modulaciÓn del jugo de toronja sobre la isoenzima...
TRANSCRIPT

1
MODULACIÓN DEL JUGO DE TORONJA SOBRE LA ISOENZIMA 1A1
RESUMEN
Las dietas humanas contienen tanto sustancias que causan el cáncer como sustancias
con acción protectora; por lo tanto, la incidencia de enfermedades crónicas que
incluyen el cáncer dependerá del consumo relativo de ambos tipos de compuestos.
Aunado a lo anterior, las interacciones que se presentan entre los alimentos y otros
carcinógenos que se encuentran en el medio ambiente son factores de riesgo para el
desarrollo de la enfermedad. En este sentido, se han publicado numerosas
investigaciones sobre la interacción del jugo de toronja (JT) con diversos fármacos, la
mayoría sustratos de CYP3A4, sin embargo, solo existen dos estudios realizados in
vitro con dos compuestos presentes en el JT (bergamotina y naringenina) quienes
inhibieron a CYP1A1 y a CYP1A2. Estas dos subfamilias de isoenzimas
biotransforman a diversos contaminantes ambientales, cuyos intermediarios
metabólicos y/o sus productos se han identificado como carcinógenos potenciales,
entre ellos el benzo (a) pireno (B[a]P). Considerando lo anterior, el objetivo de este
trabajo fue determinar el efecto del JT y del JT mas B[a]P sobre el contenido y la
actividad de la CYP1A1 hepática e intestinal in vivo. Se ocuparon 4 lotes de 6 ratones
cada uno. La administración por grupo fue la siguiente: aceite de maíz (testigo
negativo), B[a]P (100 mg/kg) como inductor enzimático, JT (41.6 μL/g), y B[a]P (100
mg/kg) en una sola administración vía i.p., 16 horas después se administró el JT v.o.
(41.6 μL/g), al completarse las 24 horas de inducción, los animales se sacrificaron y se
les extrajo el hígado e intestino delgado para obtener las fracciones S9 y microsomal
de cada órgano, las cuales se dividieron en alícuotas y se conservaron a -70°C.
Posteriormente se realizó la cuantificación de proteínas totales, la
inmunoelectrotrasferencia y la prueba de o-desetilación de la 7-etoxiresorufina para
evaluar la actividad enzimática de la CYP1A1. Los resultados muestran claramente
que: 1- El JT inhibió, tanto la cantidad como la actividad de CYP1A1; 2-El B[a]P
funcionó como inductor de la enzima; 3-Existe interacción entre el JT y B[a]P,
afectando la cantidad y la actividad de la isoforma.
INTRODUCCION
La Toxicología Genética estudia los efectos dañinos de los agentes químicos, físicos y
biológicos sobre el ADN y en los procesos genéticos de las células vivas, agentes que
son conocidos como genotóxicos y pueden ser de origen endógeno o exógeno. Dentro
de los endógenos, se encuentran por ejemplo, los radicales libres producidos durante
la respuesta inmunológica ante una infección o simplemente los producidos en el

2
metabolismo basal. Entre los exógenos, se han identificado infinidad de sustancias
químicas productos de la contaminación ambiental, incluyendo metales pesados,
productos de la combustión del petróleo o sus derivados, además de algunos virus y
radiaciones de alta energía como los rayos UV. Aunado a los genotóxicos, existen
muchos factores individuales o ambientales que pueden contribuir o aumentar el daño,
tales como: la edad, el sexo, la alimentación, hábitos o costumbres personales, nichos
ocasionales o laborales y el hábitat. De esta forma, que al identificar y caracterizar a
los agentes genotóxicos, así como los factores que los favorecen, se pueden
implementar estrategias para evitar o disminuir el daño genotóxico, lo cual se conoce
como antimutagénesis.
Las sustancias que reducen o evitan el daño al ADN, se denominan antimutágenos o
antigenotóxicos. Una gran mayoría de estos agentes se han encontrado en de
productos naturales como los alimentos, en especial las frutas y verduras. En este
contexto existen datos epidemiológicos que asocian bajos niveles en la incidencia de
algunos tipos de cáncer, especialmente aquellos encontrados en el tracto
gastrointestinal y respiratorio, con un buen suministro de frutas y verduras en la dieta
diaria.
Por otro lado, el organismo posee múltiples mecanismos de defensa para evitar que el
ADN sea alterado por los genotóxicos o, en su defecto, mecanismos de reparación
que actúan cuando el daño ya está hecho; así pues, existe cierto equilibrio para se
evitar que los genotóxicos nos dañen, impidiendo que se eleve la frecuencia de
mutaciones en nuestro organismo. El uso adecuado de los antimutágenos también
contribuye a disminuir el desarrollo de enfermedades crónico degenerativas.
TORONJA
La toronja es una hibridación natural que se dio en las islas Barbados en el siglo XVII,
entre un naranjo dulce y un pomelo (Citrus grandis). Su taxonomía y características
son las siguientes:
-Familia: Rutaceae
-Género: Citrus
-Especie: paradisi
-Porte: Reducido. Tronco corto y copa compacta, brotes color púrpura y escasa
espinosidad.
-Hojas: tamaño medio a grandes algo vellosas, con alas grandes y olor típico.
-Flores: grandes de color verdoso y estambres reducidos.
-Fruto: Hesperidio. Consta de: exocarpo (flavedo: presenta vesículas que contienen
aceites esenciales), mesocarpo (albedo: pomposo y de color blanco) grueso y

3
endocarpo (pulpa: presenta tricomas con jugo blanco, rosa o rojo). De tamaño grande
y forma redonda y algo aplastada. Superficie con glándulas prominentes con aceites.
Las variedades de toronja pueden clasificarse en dos grupos. En el primero se
incluyen las variedades blancas o comunes, la Duncan y Marsh son las más
importantes. En el segundo se engloba las variedades pigmentadas, como la
Burgundy, la Ruby (Ruby red, Redblush, Henninger), la Star Ruby y la Thompson o
Pink Marsh. La producción de toronja a nivel mundial supera las 3,8 millones de
toneladas siendo el principal productor de toronja en el mundo Estados Unidos
(2,500,200) seguido de Israel (347,500) y China (302,000). Cuba, Argentina, Brazil y
México son los países latinoamericanos con mayores niveles de producción.
Un vaso (250 mL) de jugo de toronja (JT) aporta aproximadamente 85 calorías de las
cuales 2 provienen de lípidos, 4 de proteínas y 79 de carbohidratos. Con respecto a su
valor nutricional, cabe mencionar que es una bebida baja en lípidos (la mayor parte
mono o polinsaturados), contiene al rededor de 1g de proteínas totales, lo que
representa 2% de la dosis diaria recomendada (DDR), incluye la presencia de 18
amino ácidos dentro de los que destacan el triptófano, fenilalanina, tirosina, treonina,
metionina, cisteína, isoleucina, leucina, lisina, ácido glutámico y aspártico. Con
respecto a los oligoelementos, el JT contiene potasio, calcio, cobre, magnesio, fósforo,
manganeso, potasio, hierro y zinc. También es una fuente importante de vitamina C ya
que sobrepasa la DDR en un 46%, contiene vitamina A (12% DDR), B6 (6% DDR),
ácido pantoténico, folato y tiamina, y en concentraciones bajas, vitaminas D, E,
riboflavina y niacina.
Por otro lado, el JT contiene sustancias que le confieren diferentes actividades
biológicas como las furanocumarinas o flavonoides, siendo la bergamotina y la
naringina (de 10 a 50 mg / 100 g de JT, los principales compuestos de estos grupos,
respectivamente.
La actividad antimicrobiana de la toronja se encuentra principalmente en los
compuestos extraídos de la cáscara. Se ha probado esta actividad principalmente
contra bacterias Gram-positivas ya que las Gram-negativas presentan más resistencia;
también el JT ejerce actividad antimicótico, ya que redujo hasta un 80% el crecimiento
de Aspergillus parasiticus, A. toxocarium, A. flavus y A.oryzae var. effusus in vitro;
además inhibió significativamente el crecimiento micelial y la germinación de esporas
de Penicilluim spp.
Como insecticida, se ha demostrado que el aceite esencial de la toronja ha presentado
un control efectivo de Callosobruchus muculatus, así como una disminución en la
producción de sus huevos. En la Musca domestica y Culex pipiens ha presentado
actividad insecticida en larvas y en adultos.

4
En el aspecto enzimático, el JT inhibe la 11-α-hidroxiesteroide deshidrogenasa, la cual
oxida al cortisol para inactivar a la cortisona, de tal manera que en dosis altas el JT,
produce un incremento en el efecto mineralocorticoide. Induce a la quinona reductasa
en células de ratón in vitro, la cual tiene actividad anticancerígena. El aceite esencial
de la toronja induce un incremento de la glutation S-transferasa administrado por vía
oral en ratón, enzima antioxidante que se relaciona también en procesos
antimutagénicos.
El JT reduce la concentración sanguínea de colesterol, así como una disminución de
aterosclerosis. Lo anterior se ha atribuido a las pectinas presentes en el jugo, las
cuales adsorben las grasas y otros nutrientes. Además, en otros estudios se ha
demostrado que la naringina también reduce las concentraciones plasmáticas de los
triglicéridos, el colesterol total y de baja densidad, sin embargo, este efecto no se
observa con el colesterol de alta densidad.
El JT contiene varias sustancias que por separado han demostrado ser antioxidantes,
tal es el caso de las vitaminas, A, C y E, ácidos hidroxicinámicos como el telúrico,
sinápico, p-cumárico y caféico, así como oligoelementos como el Fe, Zn, Cu, Se, Mn y
los propios flavonoides, como la naringina.
Por otro lado el JT y algunos de sus componentes probados por separado han
interactuado con la familia enzimática del citocromo P450 (CYP); algunas
furanocumarinas provenientes de extractos del JT inhibieron in vitro a isoenzimas
como CYP1A2, CPY2C9, CYP2C19, CYP2D6 Y CYP3A4, siendo la bergamotina el
inhibidor mas potente de las primeras cuatro isoenzimas; y el principal derivado de la
bergamotina (el monómero 6’,7’-dihidroxibergamoina DHB), y dos dímeros de éstos,
los más potentes en la inhibición de la CYP3A4.
En otros estudios, con bergamotina sobre microsomas humanos o de ratón, se
encontró la inhibición de CYP2B6 y CYP3A5, sugiriéndose como mecanismo a la
unión covalente entre la bergamotina o sus derivados con las isoenzimas. También se
observó la inhibición de la CYP1A1 y 1B1 al reducir su transcripción y actividad. Otro
compuesto que se ha estudiado es el flavonoide principal del JT, la naringina, de la
cual se ha demostrado que inhibe la CYP3A4 al interferir con el metabolismos de
algunos medicamentos que se biotransforman en esa isoenzima; aunque se ha
reportado también un efecto inductor sobre la Cyp3a4 intestinal en ratón. Además, se
ha reportado a la naringenina (principal derivado de la naringina), como inhibidor de la
CYP1A2 in vitro, sobre la 3-desmetilación de la cafeína.
El JT ha mostrado propiedades antigenotóxicas in vivo, ya que redujo la genotoxicidad
producida por agentes con diferentes mecanismos de acción. Así, se observó la
reducción de alrededor del 80% en la frecuencia de intercambio de cromátidas

5
hermanas (ICH) producida por la daunorrubicina (productor de radicales libres);
también con la técnica de ICH el JT protegió 45 % el daño producido por el mutágeno
directo, metil-metanosulfonat. Con la técnica de micronucleos en sangre periférica, el
JT disminuyó 54 y 70% la frecuencia de EPCMN originadas por la ifosfamida
(mutágeno indirecto).
Asimismo, algunos componentes del JT también han dado resultados satisfactorios en
estudios de antimutagénesis, tal es el caso de la naringina la cual al utilizarla en contra
de la inducción de MN en contra de la ifosfamida y el metil metano sulfonato, produjo
una genoprotección de 46 y 40%, respectivamente, ó al usarla contra la
daunorrubicina con el ensayo cometa, se obtuvo un 70% de protección en células
hepáticas de ratón. Por otra parte la bergamotina mostró una significativa reducción en
iniciación del cáncer de piel en ratones tratados con benzo[a]pireno (B[a]P), así como
la reducción de la cantidad de aductos formados por el derivado 7-8 B[a]P, en células
humanas MCF-7, al parecer mediante la inhibición del su biotransformación en la
CYP1A1.
BENZO(A) PIRENO
Está presente en el petróleo y sus derivados, y puede ser generado en la combustión
incompleta o pirólisis de materia orgánica; puede ser liberado al medio ambiente por:
derrames petroleros, fugas de óleo o gasoductos, el uso doméstico o industrial del
carbón, de la madera y de combustibles fósiles, en el humo del tabaco, etc. Debido a
su amplia distribución, a sus características fisicoquímicas y efectos biológicos, el
B[a]P es un contaminante ambiental que representan un importante riesgo ecológico y
para la salud humana.
Dentro de las fuentes de exposición se encuentran: la ingesta de alimentos o agua
contaminados, la forma de preparación o de cocción de algunos alimentos (productos
ahumados, al carbón o fritos); por vía tópica, al entrar en contacto con aguas o
sedimentos contaminados o bien por vía inhalatoria (gases de la combustión de
materias orgánicas, humo del cigarro, gases de desecho de motores de combustión
interna.
Por otro lado se han realizado varios ensayos desde principios de los 80’s en donde se
relaciona el B[a]P con procesos de mutagénesis, carcinogénesis (principalmente está
relacionado con el cáncer de pulmón hígado vejiga y piel), así como en cardiopatías y
con abortos.
Las vías de absorción del B[a]P son la inhalatoria, oral o dérmica. Para la vía
inhalatoria la retención y absorción del B[a]P a través del tracto respiratorio depende
del tamaño de partícula, para vías aéreas altas, pueden ser eliminadas por estornudo

6
o en el esputo, o en su defecto pasar a la vía gástrica y absorberse posteriormente,
mientras que, a nivel bronquial o alveolar se pueden absorber a través de los
macrófagos o por el epitelio. Por la vía oral el B[a]P se absorbe en el intestino del 10 al
60 % de la dosis ingerida.
El B[a]P es biotransformado principalmente por la CYP1A1 en un primer paso a los
derivados 4,5 óxido y 7,8 óxido-B[a]P, de los cuales el primero es poco tóxico en los
mamíferos, por su transformación subsecuente a 4,5 dihidrol-B[a]P realizada por la
epóxido hidrolasa, lo cual facilita su conjugación con glutatión, ácido glucurónico o por
sulfonaciones para su eliminación (reacciones de Fase II). El segundo derivado
también es sustrato de la epóxido hidrolasa formando 7,8 dihidrol-B[a]P pero éste es
metabolizado por la dihidrol deshidrogenasa para formar las estructuras quinona y
semiquinona correspondientes, la que su vez forma un equilibrio de oxido-reducción,
reacción que es productora de radicales libres. Por otro lado el derivado 7,8 dihidrol-
B[a]P puede ser metabolizado nuevamente por la CYP1A1 o por el sistema de
cooxidación por parte de la prostaglandin H sintetasa, dando como resultado el
derivado 7,8 dihidrol-9,10 epoxido-B[a]P, el cual es reconocido como el derivado
mutagénico mas potente en mamíferos por su capacidad para unirse covalentemente
con ADN, ARN y proteínas, siendo inductor de oncogenes, con lo que puede contribuir
en el proceso de la carcinogénesis.
EL CITOCROMO P450 Y LA ISOENZIMA CYP1A1
El sistema enzimático más importante que cataliza las reacciones metabólicas de la
fase I es el citocromo P450 (CYP). En 1958 se descubrió que cuando los microsomas
de hígado de cerdo o de rata se trataban con ditionito de sodio en un ambiente
saturado con monóxido de carbono, presentaban una gran absorción a 450 nm, lo cual
no era usual para los otros 10 pigmentos conocidos. El pigmento responsable de este
fenómeno se llamó P (por pigmento) 450 (por la longitud de onda).
Todas las enzimas del citocromo P450 son hemoproteínas. El fierro del grupo hemo en
el citocromo P450 usualmente se encuentra en estado férrico (Fe 3+), cuando se
reduce al estado ferroso (Fe 2+) las enzimas pueden realizar su función de
monooxigenasas o hidroxilasas, también llamadas oxigenasas de función mixta, en
términos generales el proceso es el siguiente: después de la unión del sustrato al
citocromo P450, el Fe3+ es reducido a Fe2+ gracias a un electrón donado por la
flavoproteína NADPH reductasa, de esta forma el oxígeno molecular (O2) es unido al
fierro reducido [Fe2+O2] complejo que es convertido a [Fe2+OOH] por la adición de un
protón (H+) y un segundo electrón donados por la NADPH reductasa y el citocromo b5,
a partir de este intermediario se forma H2O y el radical (FeO)3+ el cual trasfiere su

7
oxígeno al sustrato, dando como resultado el respectivo derivado y el citocromo P450
en su estado inicial con el Fe3+.
El metabolismo de las sustancias exógenas por el CYP no siempre lleva a la
inactivación de estos compuestos; en muchos casos, los productos de la oxidación o
de la reducción han mostrado la capacidad de iniciar procesos tóxicos los cuales
pueden derivar en la carcinogénesis, mutagénesis ó teratogénesis.
La familia CYP1 está constituida por dos subfamilias la CYP1A y la CYP1B, a su vez,
la subfamilia CYP1A, antiguamente llamada P448, se divide en dos miembros, la
CYP1A1 y la CYP1A2, las cuales son de los CYP mas conservados evolutivamente
entre los mamíferos. La inducción de la CYP1A1 se produce con diferentes
xenobióticos como el 3-metilclorantreno, la b-naftoflavona, el 2,3,7,8-tetraclorodibenzo-
p-dioxina (TCDD), o diferentes hidrocarburos como el B[a]P. Dicha inducción se logra
cuando cualquiera de estos xenobióticos se une al receptor de hidrocarburos arílicos
(Ahr) a nivel citoplasmático, desplazando a una proteína de choque térmico, la HSP90,
a la cual se encuentra regularmente unido el Ahr. El complejo formado por el
xenobiótico y el Ahr atraviesa la membrana nuclear y se une a secuencias específicas
del ADN actuando como potenciador de la transcripción del RNA mensajero de
diferentes enzimas, entre ellas las de la familia CYP1, incrementando su
concentración. En este contexto, el B[a]P favorece su propia biotransformación.
JUSTIFICACIÓN Y OBJETIVOS
Existen pocos estudios sobre el potencial antigenotóxico del jugo de toronja y no
existe ninguno que evalúe su efecto contra el benzo[a]pireno, el cual es un
contaminante ambiental ampliamente distribuido que puede ocasionar diversos
efectos perjudiciales al hombre y otras especies. Por otro lado, existe evidencia de
interacciones del JT ó algunos de sus componentes con la isoenzima CYP1A1, lo que
podría explicar una probable vía de acción antigenotóxica contra el hidrocarburo.
Si el jugo de toronja es capaz de inhibir la actividad de la CYP1A y por ende alterar su
capacidad de biotransformación, entonces se observará dicho fenómeno con
promutágenos tales como el B[a]P que se activan con esta isoenzima.
Debido a lo anterior, el objetivo general de este trabajo fue determinar el efecto del JT
sobre el contenido y la actividad de la CYP1A1 hepática e intestinal, mediante las
técnicas de inmunoelectrotransferencia y de la o-desetilación de la 7-etoxiresorufina.
METODOLOGIA
Se ocuparon 4 lotes de 6 animales cada uno, se les administraron los compuestos
correspondientes como se muestra en la Tabla III. Para este ensayo se utilizó al B[a]P
(100 mg/kg de peso) como inductor enzimático, en una sola administración vía i.p., 16
horas más tarde se administró el JT v.o. en la dosis más alta utilizada en el ensayo

8
antigenotóxico (41.6 B[a]P μL/g), al completarse las 24 horas de inducción, los
animales se sacrificaron y se les extrajo el hígado e intestino delgado para obtener las
fracciones S9 y microsomal de cada órgano, las cuales se dividieron en alícuotas y se
conservaron a -70°C.
Posteriormente se realizó la cuantificación de proteínas totales, la
inmunoelectrotrasferencia (IET) y la prueba de o-desetilación de la 7-etoxiresorufina
(EROD) para evaluar la actividad enzimática de la CYP1A1..
Distribución de grupos experimentales. Efecto de la CYP1A1 in vivo.
LOTE COMPUESTO 1 DOSIS COMPUESTO 2 DOSIS
1 Aceite de maíz 0.01mL/g --- ---
2 --- --- Jugo de toronja 41.6 μL/g
3 Benzo[a]pireno 100 mg/kg --- ---
4 Benzo[a]pireno 100 mg/kg Jugo de toronja 41.6 μL/g
RESULTADOS
A continuación se presentan los resultados con respecto al efecto del JT sobre la
CYP1A1 in vivo, en el siguiente orden: la determinación de proteínas totales, la
inmunoelectrotransferencia (IET) y la prueba de EROD realizada en las fracciones S9
y las microsomales, de hígado e intestino delgado.
La cuantificación de proteínas totales fue un parámetro necesario para realizar las
demás técnicas; la IET se estableció de manera especifica para observar la acción del
JT sobre la cantidad de CYP1A1, mientras que con la prueba de EROD se cuantificó el
efecto del JT sobre la actividad enzimática independientemente de la cantidad de
enzima.
En lo referente al hígado, se pueden observar en la Tabla IV las concentraciones
totales de proteínas en mg/mL de las fracciones hepáticas estudiadas; es evidente la
mayor concentración de proteínas totales en las fracciones S9 que en las fracciones
microsomales, esto es congruente con la naturaleza de las mismas fracciones, ya que
la fracción microsomal está contenida en la fracción S9, sin embargo, no existen
diferencias significativas, observándose que los intervalos de las concentraciones de
proteínas totales se encuentran entre 28.36 y 31.71 mg/mL para la fracción S9 y entre
16.31 y 19.04 mg/mL para los microsomas.

9
Tabla IV. Concentración hepática de proteínas totales.
Lotes Concentración
mg/mL
S9 de hígado tratados con aceite de maíz. (S9HA) 31.24
S9 de hígado tratados con B[a]P (S9HB) 31.71
S9 de hígado tratados con JT (S9HJT) 29.33
S9 de hígado tratados con JT y B[a]P (S9HBJT) 28.36
Microsomas de hígado tratados con a de
maíz
(MHA) 15.91
Microsomas de hígado tratados con BaP (MHB) 19.04
Microsomas de hígado tratados con JT (MHJT) 16.31
Microsomas de hígado tratados JT y BaP (MHBJT) 17.51
En la Figura 6 se muestra la fotografía electrónica de la membrana de nitrocelulosa,
resultado final del IET obtenido con el hígado, en donde se observan en el carril 1 el
corrimiento de CYP1A estándar, del 2 al 5 los correspondientes a la fracción S9 y del 6
al 9 los correspondientes a los microsomas, cada fracción está ordenada de acuerdo
al compuesto administrado, es decir, primero el del grupo TN (aceite de maíz),
después el TP (B[a]P 100 mg/kg), a continuación el administrado con JT (41.6 μL/g), y
por último el grupo combinado de B[a]P mas JT.
El primer anticuerpo que se utilizó reconoce a la CYP1A, de tal forma que se unió
tanto a la CYP1A1 como a la CYP1A2, esta última tiene un peso molecular menor, de
tal forma la banda de mas arriba corresponde a la CYP1A1 y la otra a la CYP1A2,
como se puede observar claramente en el carril 1.
Los resultados de la fracción S9 y de la microsomal son similares, de tal forma que
para el TN, (carriles 2 y 6) la expresión basal que existe es de CYP1A2 mientras que
no se observa CYP1A1. Para el TP, (carriles 3 y 7) la inducción por parte del B[a]P es
clara para las dos isoenzimas al compararlos con el TN. Con respecto al efecto del JT
se observa (carriles 4 y 8), que no difieren de su TN, es decir, que no modificó la
cantidad basal de CYP1A en el hígado, y tampoco influyó de con la inducción del
B[a]P (carriles 5 y 9); resultados que sugieren que en el hígado del JT per se no actuó
como inductor enzimático ni interfirió con la inducción producida por el B[a]P sobre la
CYP1A1.

10
1 2 3 4 5 6 7 8 9
Figura 6. Inmunoelectrotransferencia de CYP1A hepática. Efecto del JT sobre el
contenido enzimático.
(1) CYP1A control, (2) S9 de hígado tratados con aceite de maíz (S9HA), (3) S9 de
hígado tratados con B[a]P (S9HB), (4) S9 de hígado tratados con JT (S9HJT), (5) S9
de hígado tratados con JT y B[a]P (S9HBJT), (6) Microsomas de hígado tratados con
a. de maíz (MHA), (7) Microsomas de hígado tratados con B[a]P (MHB), (8)
Microsomas de hígado tratados con JT (MHJT), (9) Microsomas de hígado tratados JT
y B[a]P (MHBJT).
Con respecto a la actividad de la CYP1A1 hepática los resultados están expresados
en pmol de resorufina formada en un minuto por un miligramo de proteína total
utilizada (pmol/min/mg), en la Figura 7 se presentan los resultados referentes a la
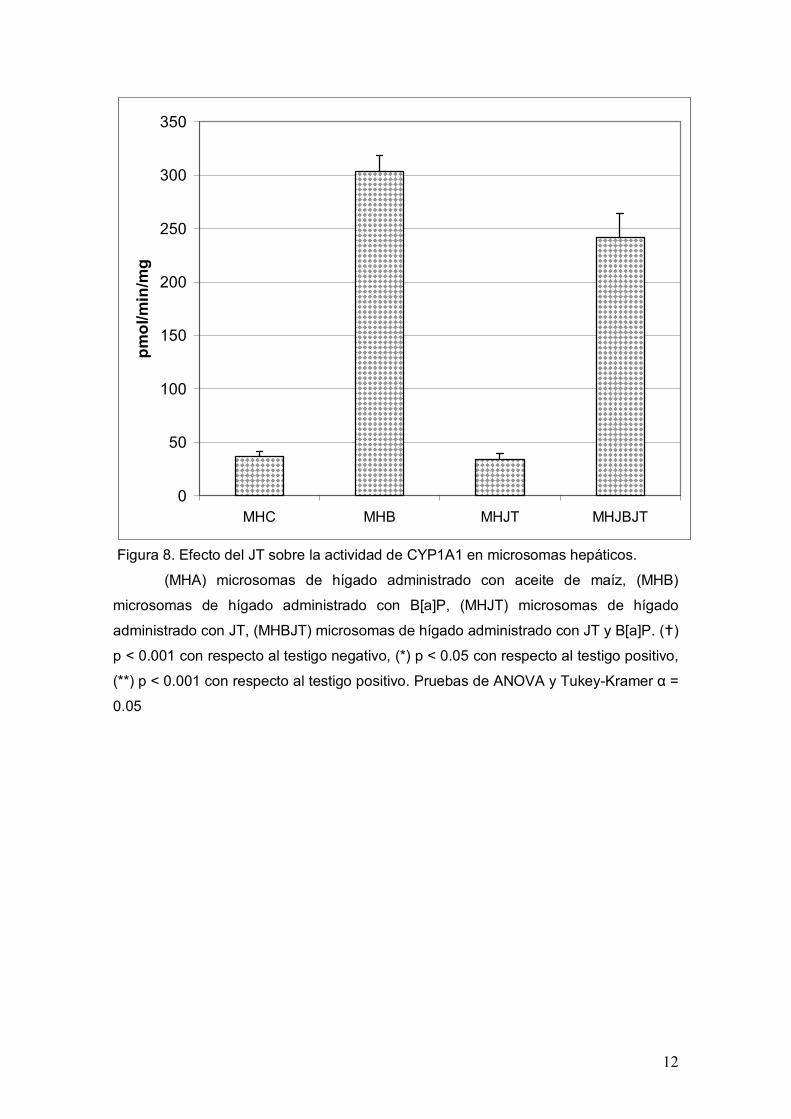
fracción S9 y en la Figura 8 los de la fracción microsomal. En ambas figuras el orden
de los lotes es el mismo, primero el TN después el TP a continuación el administrado
con JT y al último el administrado con B[a]P mas JT.
Para la fracción S9, su nivel basal de actividad promedio fue de 7.6 pmol/min/mg
(S9HA), para el TP (S9HB) la actividad aumentó 10.27 veces el nivel basal, es decir,
su valor fue de 78.5 pmol/min/mg en promedio, presentando diferencia
estadísticamente significativa al comparar ambos testigos.
En el caso del JT, se obtuvo una actividad de CYP1A1 de 8.8 pmol/min/mg similar al
del TN y no hubo diferencia significativa, lo cual sugiere que el JT por si mismo no
induce ni inhibe la actividad basal de la enzima. En el caso del lote combinado
(S9HBJT) la actividad de CYP1A1 se llegó a 63 pmol/min/mg mostrando diferencias
significativas con el TN y con el TP, lo que sugiere que el JT inhibió la actividad de la
enzima en un 22 % al compararla con el TP.
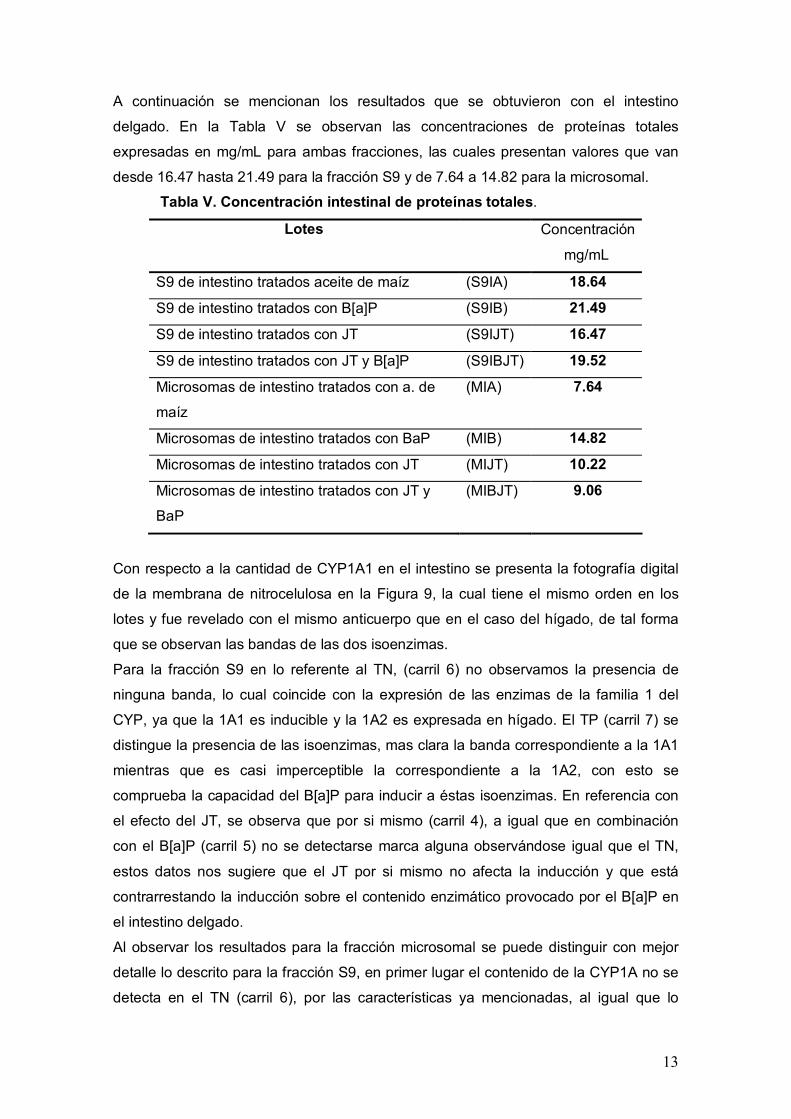
En el caso de los microsomas el comportamiento de la actividad de CYP1A1 es similar
al observado en la fracción S9. EL valor basal (MHA) fue de 37 pmol/min/mg, mientras
que para el TP (MHB) fue de 303.7 lo que representa un aumento de 8.2 veces de la
CYP1A1
CYP1A2

11
actividad de CYP1A1. En el grupo que se administro JT (MHJT) se obtuvo un valor
promedio de 33.9 pmol/min/mg y no difirió estadísticamente del TN. Respecto al grupo
combinado que recibió JT más B[a]P (MHBJT) se observa una reducción de la
actividad del 23 % al presentar un valor promedio de 242 pmol/min/mg obteniendo
diferencias significativas con el TP y con el TN. Por lo tanto, estos datos sugieren que
no se pierde la propiedad inhibitoria del JT debido al proceso de purificación de los
microsomas a partir de la fracción S9.
0
10
20
30
40
50
60
70
80
90
100
S9HC S9HB S9HJT S9HBJT
pm
ol/
min
/mg
Figura 7. Efecto del JT sobre la actividad de CYP1A1 en la fracción S9 hepática.
(S9HA) S9 de hígado administrado con aceite de maíz, (S9HB) S9 de hígado
administrado con B[a]P, (S9HJT) S9 de hígado administrado con JT, (S9HBJT) S9 de
hígado administrado con JT y B[a]P. () p < 0.001 con respecto al testigo negativo, (*)
p < 0.05 con respecto al testigo positivo, (**) p < 0.001 con respecto al testigo positivo.
Pruebas de ANOVA y Tukey-Kramer α = 0.05
*
** **
S9HA
12
0
50
100
150
200
250
300
350
MHC MHB MHJT MHJBJT
pm
ol/m
in/m
g
Figura 8. Efecto del JT sobre la actividad de CYP1A1 en microsomas hepáticos.
(MHA) microsomas de hígado administrado con aceite de maíz, (MHB)
microsomas de hígado administrado con B[a]P, (MHJT) microsomas de hígado
administrado con JT, (MHBJT) microsomas de hígado administrado con JT y B[a]P. ()
p < 0.001 con respecto al testigo negativo, (*) p < 0.05 con respecto al testigo positivo,
(**) p < 0.001 con respecto al testigo positivo. Pruebas de ANOVA y Tukey-Kramer α =
0.05
13
A continuación se mencionan los resultados que se obtuvieron con el intestino
delgado. En la Tabla V se observan las concentraciones de proteínas totales
expresadas en mg/mL para ambas fracciones, las cuales presentan valores que van
desde 16.47 hasta 21.49 para la fracción S9 y de 7.64 a 14.82 para la microsomal.
Tabla V. Concentración intestinal de proteínas totales.
Lotes Concentración
mg/mL
S9 de intestino tratados aceite de maíz (S9IA) 18.64
S9 de intestino tratados con B[a]P (S9IB) 21.49
S9 de intestino tratados con JT (S9IJT) 16.47
S9 de intestino tratados con JT y B[a]P (S9IBJT) 19.52
Microsomas de intestino tratados con a. de
maíz
(MIA) 7.64
Microsomas de intestino tratados con BaP (MIB) 14.82
Microsomas de intestino tratados con JT (MIJT) 10.22
Microsomas de intestino tratados con JT y
BaP
(MIBJT) 9.06
Con respecto a la cantidad de CYP1A1 en el intestino se presenta la fotografía digital
de la membrana de nitrocelulosa en la Figura 9, la cual tiene el mismo orden en los
lotes y fue revelado con el mismo anticuerpo que en el caso del hígado, de tal forma
que se observan las bandas de las dos isoenzimas.
Para la fracción S9 en lo referente al TN, (carril 6) no observamos la presencia de
ninguna banda, lo cual coincide con la expresión de las enzimas de la familia 1 del
CYP, ya que la 1A1 es inducible y la 1A2 es expresada en hígado. El TP (carril 7) se
distingue la presencia de las isoenzimas, mas clara la banda correspondiente a la 1A1
mientras que es casi imperceptible la correspondiente a la 1A2, con esto se
comprueba la capacidad del B[a]P para inducir a éstas isoenzimas. En referencia con
el efecto del JT, se observa que por si mismo (carril 4), a igual que en combinación
con el B[a]P (carril 5) no se detectarse marca alguna observándose igual que el TN,
estos datos nos sugiere que el JT por si mismo no afecta la inducción y que está
contrarrestando la inducción sobre el contenido enzimático provocado por el B[a]P en
el intestino delgado.
Al observar los resultados para la fracción microsomal se puede distinguir con mejor
detalle lo descrito para la fracción S9, en primer lugar el contenido de la CYP1A no se
detecta en el TN (carril 6), por las características ya mencionadas, al igual que lo

14
observado para el JT (carril 8), es decir el JT per se no interfiere con el contenido
enzimático de la CYP1A. Para el TP (carril 7) se confirma la presencia mayoritaria de
la CYP1A1 en comparación con la 1A2, la cual es apenas perceptible. Para el lote
combinado (carril 9), se observó que si existe inducción para la CYP1A, lo que no se
apreciaba en la fracción S9, sin embargo, al compararlo con el TP, se observa que el
JT si está interfiriendo con la inducción por parte del B[a]P, ya que se observa una
clara disminución en banda correspondiente a la CYP1A1 y un aumento en la
correspondiente a la 1A2, estos resultados nos sugieren que el JT esta modulando
diferencialmente la inducción sobre la CYP1A por parte del B[a]P, al reducir el
contenido de la 1A1 e incrementar la 1A2.
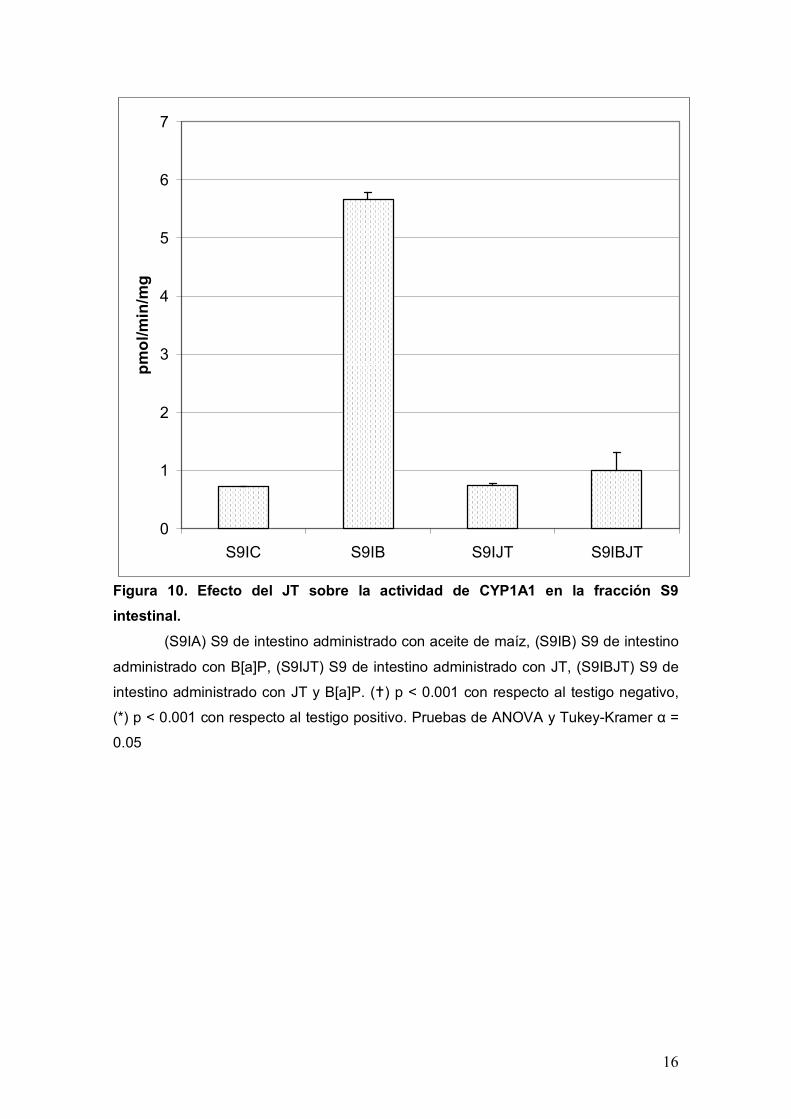
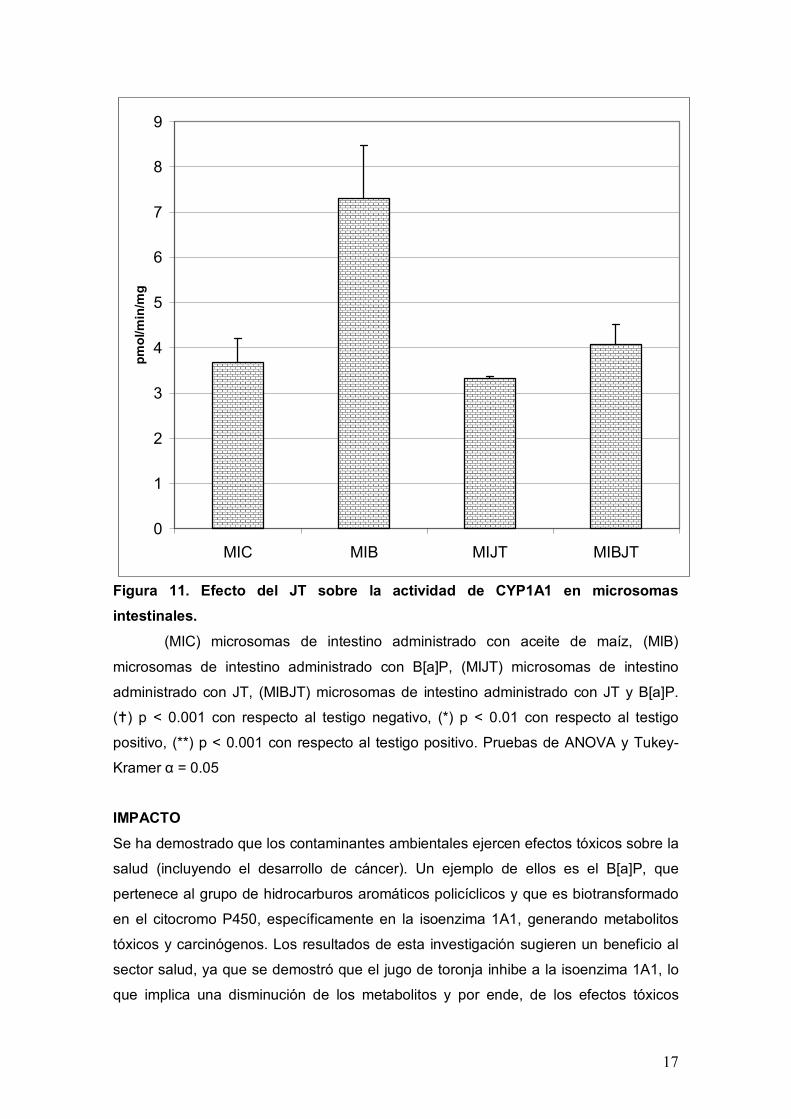
Con respecto a la actividad de CYP1A1 en intestino se presentan los datos con el
mismo orden y formato que para hígado, de tal forma que en la Figura 10 se muestran
los resultados de la fracción S9 y el la Figura 11 los correspondientes a la fracción
microsomal. Así pues, para ambas fracciones el comportamiento de la enzima es
similar.
1 2 3 4 5 6 7 8
9
Figura 9. Inmunoelectrotransferencia de CYP1A en intestino delgado. Efecto del
JT sobre el contenido enzimático.
(1) CYP1A1 control, (2) S9 de intestino tratados aceite de maíz (S9IA), (3) S9 de
intestino tratados con B[a]P (S9IB), (4) S9 de intestino tratados con JT (S9IJT), (5) S9
de intestino tratados con JT y B[a]P (S9IBJT), (6) Microsomas de intestino tratados
con a. de maíz (MIA), (7) Microsomas de intestino tratados con B[a]P (MIB), (8)
Microsomas de intestino tratados con JT (MIJT), (9) Microsomas de intestino tratados
con JT y BaP (MIBJT).
En la fracción S9 el valor basal de la actividad fue de 0.7 pmol/min/mg (S9IA), y de 5.6
pmol/min/mg para el TP (S9IB), lo que representa un incremento de 8 veces con
respecto a la basal. Para el lote administrado con JT (S9IJT) la actividad fue de 0.7
pmol/min/mg mientras que para el lote que se administro JT más B[a]P (S9IBJT) se
obtuvo un valor de 1 pmol/min/mg, ambos resultados difieren estadísticamente con el
CYP1A1
CYP1A2

15
TP, al ser similares al TN. Para los microsomas tenemos para el TN (MIA) un valor de
3.6 pmol/min/mg, y para el TP (MIB) un valor de 7.3 pmol/min/mg, lo cual representa
un aumento de 2 veces siendo estadísticamente significativo. Para grupo tratado con
JT (MIJT) y el tratado con JT más B[a]P (MIBJT), tenemos valores de 3.3 y de 4
pmol/min/mg respectivamente, los cuales no difieren estadísticamente con el TN pero
sí con el TP.
Estos resultados sugieren que en el intestino delgado: el TN presentó actividad basal
de EROD debido a niveles no detectados de CYP1A en la IET. En el TP la inducción
del B[a]P en la cantidad de CYP1A1 observada en la IET, se vio reflejada en la
actividad de ésta al incrementar la producción de resorufina. En lo referente al JT se
observó que por si mismo no afecta ni la cantidad ni la actividad de la CYP1A1 al
mantenerse igual que el TN, sin embargo, el JT interfiere con la inducción por parte del
B[a]P, inhibiendo la actividad de EROD llevándola hasta niveles basales, esto debido
en parte a la disminución que causo sobre la cantidad de la CYP1A1.
16
0
1
2
3
4
5
6
7
S9IC S9IB S9IJT S9IBJT
pm
ol/m
in/m
g
Figura 10. Efecto del JT sobre la actividad de CYP1A1 en la fracción S9
intestinal.
(S9IA) S9 de intestino administrado con aceite de maíz, (S9IB) S9 de intestino
administrado con B[a]P, (S9IJT) S9 de intestino administrado con JT, (S9IBJT) S9 de
intestino administrado con JT y B[a]P. () p < 0.001 con respecto al testigo negativo,
(*) p < 0.001 con respecto al testigo positivo. Pruebas de ANOVA y Tukey-Kramer α =
0.05
17
0
1
2
3
4
5
6
7
8
9
MIC MIB MIJT MIBJT
pm
ol/m
in/m
g
Figura 11. Efecto del JT sobre la actividad de CYP1A1 en microsomas
intestinales.
(MIC) microsomas de intestino administrado con aceite de maíz, (MIB)
microsomas de intestino administrado con B[a]P, (MIJT) microsomas de intestino
administrado con JT, (MIBJT) microsomas de intestino administrado con JT y B[a]P.
() p < 0.001 con respecto al testigo negativo, (*) p < 0.01 con respecto al testigo
positivo, (**) p < 0.001 con respecto al testigo positivo. Pruebas de ANOVA y Tukey-
Kramer α = 0.05
IMPACTO
Se ha demostrado que los contaminantes ambientales ejercen efectos tóxicos sobre la
salud (incluyendo el desarrollo de cáncer). Un ejemplo de ellos es el B[a]P, que
pertenece al grupo de hidrocarburos aromáticos policíclicos y que es biotransformado
en el citocromo P450, específicamente en la isoenzima 1A1, generando metabolitos
tóxicos y carcinógenos. Los resultados de esta investigación sugieren un beneficio al
sector salud, ya que se demostró que el jugo de toronja inhibe a la isoenzima 1A1, lo
que implica una disminución de los metabolitos y por ende, de los efectos tóxicos

18
producidos por el B[a]P. Es probable que ocurra lo mismo con otros contaminantes
ambientales derivados de la combustión del petróleo. De tal manera que con este
proyecto se establecen las bases para continuar estudiando las interacciones entre un
alimento de fácil acceso y las enzimas involucradas en la biotransformación de
xenobióticos tóxicos; con la finalidad de reducir la incidencia de cáncer en nuestro
país.