mg+2 aminoácido+arnt+atp...
TRANSCRIPT
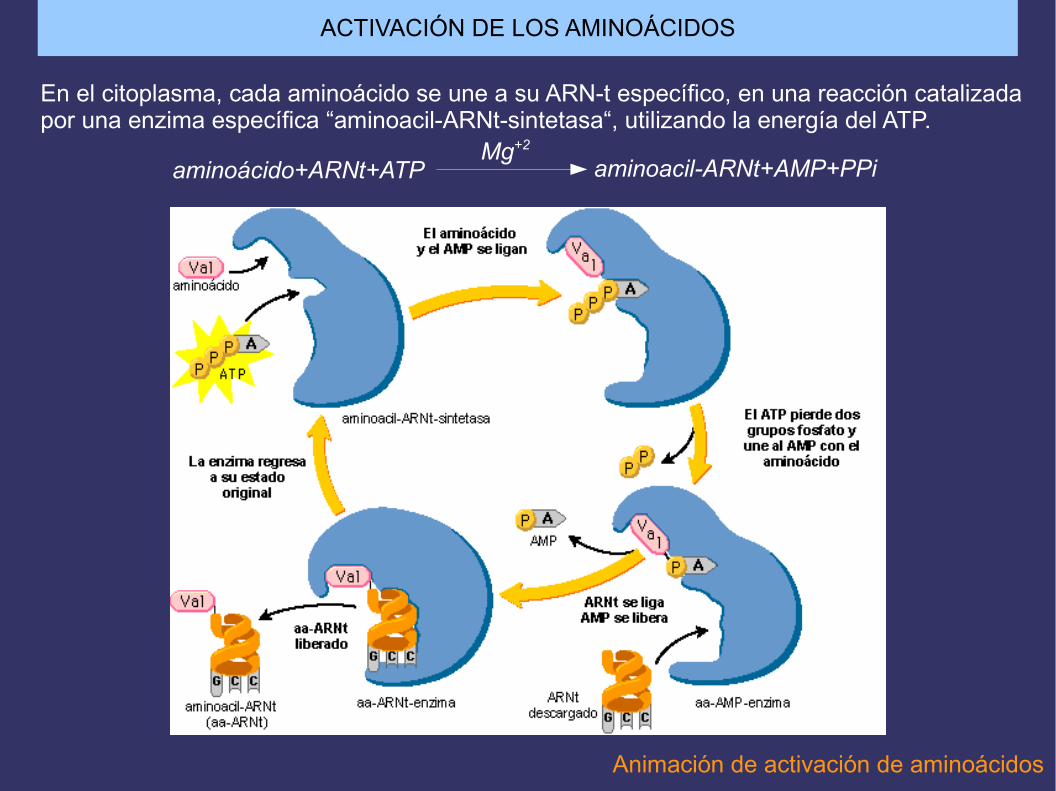
Animación de activación de aminoácidos
En el citoplasma, cada aminoácido se une a su ARN-t específico, en una reacción catalizadapor una enzima específica “aminoacil-ARNt-sintetasa“, utilizando la energía del ATP.
aminoácido+ARNt+ATP aminoacil-ARNt+AMP+PPiMg+2
ACTIVACIÓN DE LOS AMINOÁCIDOS
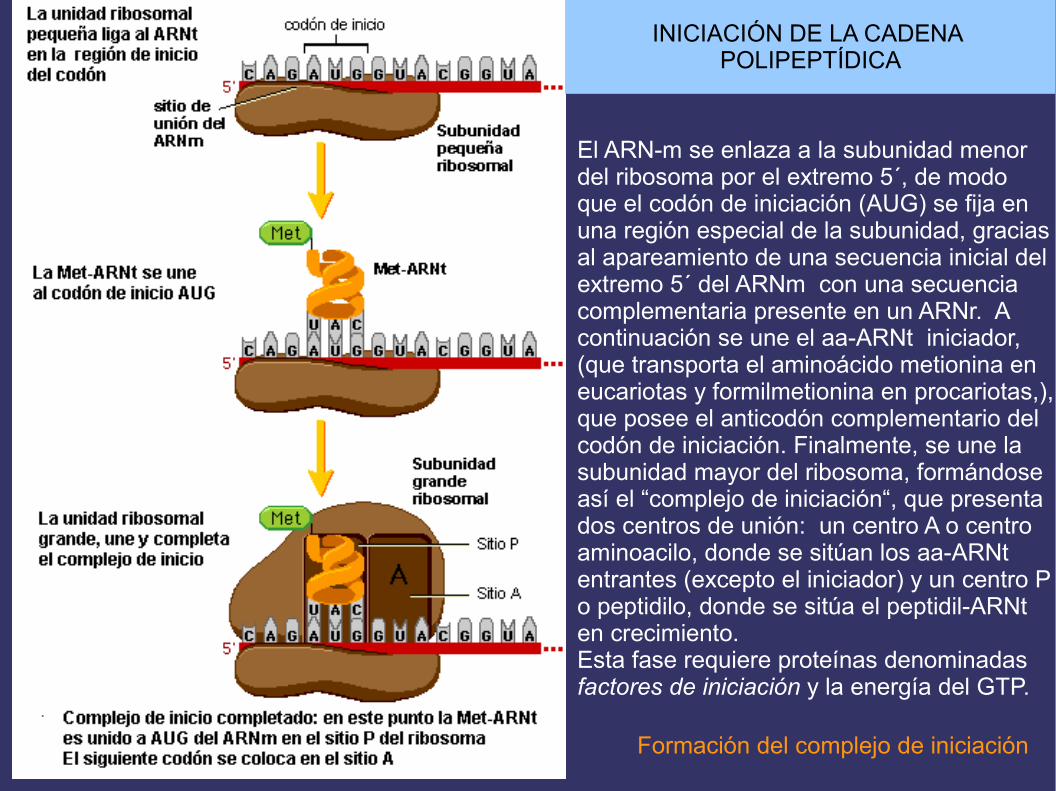
Formación del complejo de iniciación
INICIACIÓN DE LA CADENA POLIPEPTÍDICA
El ARN-m se enlaza a la subunidad menordel ribosoma por el extremo 5´, de modo que el codón de iniciación (AUG) se fija enuna región especial de la subunidad, gracias al apareamiento de una secuencia inicial del extremo 5´ del ARNm con una secuencia complementaria presente en un ARNr. A continuación se une el aa-ARNt iniciador, (que transporta el aminoácido metionina en eucariotas y formilmetionina en procariotas,),que posee el anticodón complementario delcodón de iniciación. Finalmente, se une lasubunidad mayor del ribosoma, formándoseasí el “complejo de iniciación“, que presenta dos centros de unión: un centro A o centro aminoacilo, donde se sitúan los aa-ARNt entrantes (excepto el iniciador) y un centro Po peptidilo, donde se sitúa el peptidil-ARNt en crecimiento.Esta fase requiere proteínas denominadasfactores de iniciación y la energía del GTP.

Elongación de la cadena polipeptídica
ELONGACIÓN DE LA CADENA POLIPÉPTIDICA
El crecimiento de la cadena polipeptídica tiene lugar mediante la formación de enlaces péptídicos entre los aminoácidos sucesivos. Esta fase requiere también factores proteicos de prolongación y la energía del GTP. El aa-ARNt correspondiente al siguiente codón del ARNm entra en el centro A y aparea su triplete anticodón, mediante puentes de hidrógeno, con el codón del ARNm. Una enzima del ribosoma, la peptidil-transferasa, cataliza la formación del enlace peptídico entre el grupo carboxilo del primer aminoácido (que se desprende así de su ARNt) y el grupo amino del aminoácido recién llegado, formándose un dipeptidil-ARNt en el centro A, mientras el ARNt descargado sigue en el centro P. El ribosoma se desplaza sobre el ARN-mla distancia de un codón hacia el extremo 3´, lo queprovoca la salida del ARN-t descargado y el desplazamiento del dipeptidil-ARN-t al centro P, dejando libre el centro A. Ahora, con la entrada delsiguiente aminoacil-ARN-t se repite el ciclo. En cada ciclo se añade un aminoácido a la cadena peptídica,que va creciendo desde el extremo amino al carboxilo, a medida que el ribosoma avanza desde el extremo 5´ al 3´ del ARN mensajero

Terminación de la síntesis proteica
TERMINACIÓN DE LA TRADUCCIÓN
La cadena polipeptídica está finalizada y unida al último ARN transferente en forma de peptidil-ARN-t en el centro P. En el centro A libre hay un codón de fin de síntesis para el que ningún ARN transferente tiene un anticodón complementario. Este codón de stop es reconocido por factores proteicos de liberación, lo que provoca la separacióndel polipéptido, que se libera del último ARN-t,la disociación del ribosoma en sus dos subunidades y la liberación del ARN-m, que será degradado.

Animación de la traducción
Animación Síntesis de proteínas (mcgraw-hill)
A medida que el ribosoma se desplaza a lo largo del ARN-m desde el extremo 5´ al 3´, va sintetizando una cadena polipeptídica del extremo amino al carboxilo.
Animación con formación de enlaces
A continuación tienes tres animaciones del proceso de traducción:

Animación de polirribosoma
POLIRRIBOSOMAS O POLISOMAS
Un ARN-m puede ser traducido, simultáneamente, por varios ribosomas que constituyen, en conjunto, un polirribosoma o polisoma. Cada ribosoma sintetiza una cadena polipeptídica y, por tanto, se sintetizarán tantas copias de la cadena peptídica como ribosomas hayan traducido al ARN-m.
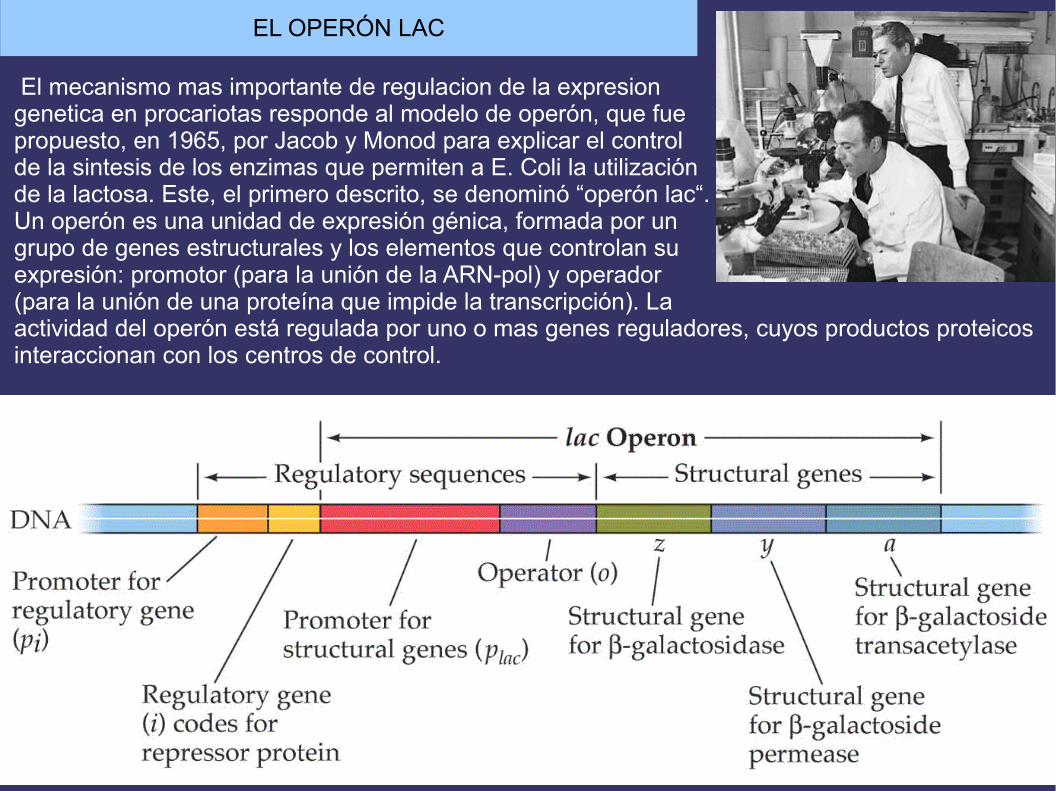
EL OPERÓN LAC
El mecanismo mas importante de regulacion de la expresion genetica en procariotas responde al modelo de operón, que fue propuesto, en 1965, por Jacob y Monod para explicar el control de la sintesis de los enzimas que permiten a E. Coli la utilización de la lactosa. Este, el primero descrito, se denominó “operón lac“. Un operón es una unidad de expresión génica, formada por ungrupo de genes estructurales y los elementos que controlan suexpresión: promotor (para la unión de la ARN-pol) y operador (para la unión de una proteína que impide la transcripción). Laactividad del operón está regulada por uno o mas genes reguladores, cuyos productos proteicosinteraccionan con los centros de control.

Inductorpresente Inductor
ausente
CONTROL DE LA TRANSCRIPCIÓN DE LOS GENES DEL OPERÓN LAC
Animación de regulación del operon lac
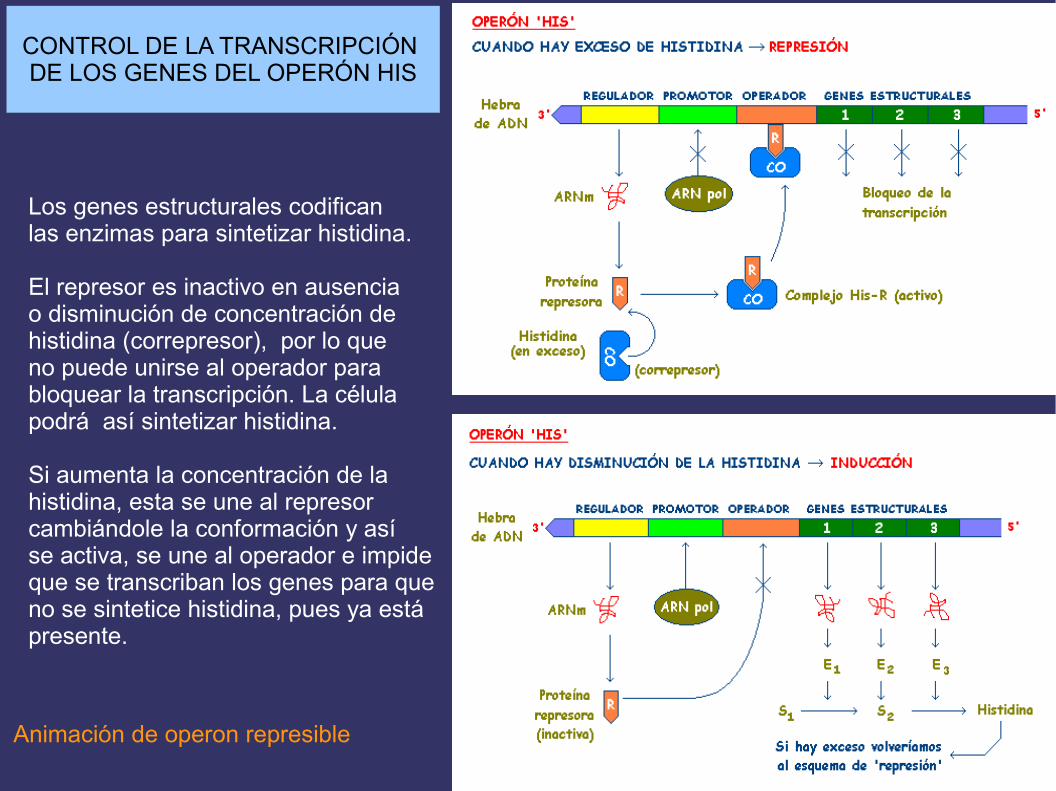
CONTROL DE LA TRANSCRIPCIÓN DE LOS GENES DEL OPERÓN HIS
Los genes estructurales codificanlas enzimas para sintetizar histidina.
El represor es inactivo en ausenciao disminución de concentración dehistidina (correpresor), por lo queno puede unirse al operador para bloquear la transcripción. La célulapodrá así sintetizar histidina.
Si aumenta la concentración de lahistidina, esta se une al represorcambiándole la conformación y asíse activa, se une al operador e impideque se transcriban los genes para queno se sintetice histidina, pues ya estápresente.
Animación de operon represible
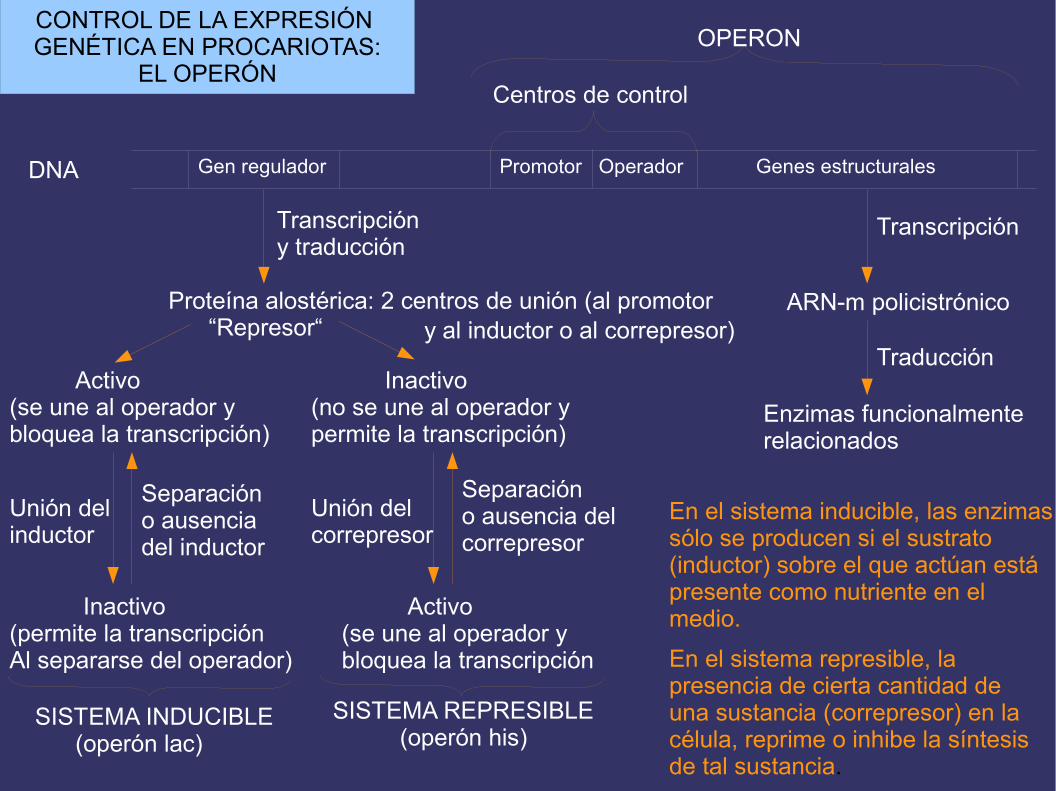
Gen regulador Promotor Operador Genes estructuralesDNA
Centros de control
OPERON
ARN-m policistrónico
Enzimas funcionalmenterelacionados
Transcripción
Traducción
Transcripción y traducción
Proteína alostérica: 2 centros de unión (al promotor “Represor“
Activo(se une al operador y bloquea la transcripción)
Inactivo(no se une al operador ypermite la transcripción)
Inactivo(permite la transcripción Al separarse del operador)
Unión delinductor
Separacióno ausencia del inductor
Activo(se une al operador y bloquea la transcripción
Unión delcorrepresor
Separacióno ausencia del correpresor
SISTEMA INDUCIBLE (operón lac)
SISTEMA REPRESIBLE (operón his)
y al inductor o al correpresor)
CONTROL DE LA EXPRESIÓN GENÉTICA EN PROCARIOTAS:
EL OPERÓN
En el sistema inducible, las enzimas sólo se producen si el sustrato (inductor) sobre el que actúan está presente como nutriente en el medio.
En el sistema represible, la presencia de cierta cantidad de una sustancia (correpresor) en la célula, reprime o inhibe la síntesis de tal sustancia.

CONTROL DE LA EXPRESIÓN GÉNICA EN EUCARIOTAS (I)
En eucariotas, la regulación de la expresión génica es mucho mas compleja que en procariotas.Aunque cada una de las células contiene el genoma completo del organismo, en cada una deellas solo se expresa una fracción de sus genes estructurales. Los diferentes tipos celularestienen estructuras y funciones distintas debido a la presencia de grupos diferentes de proteínasespecializadas, aunque todas poseen el conjunto básico de enzimas para la catálisis de lasrutas metabólicas centrales. Además, durante el desarrollo embrionario de los organismos superiores, la síntesis de las distintas clases de proteínas especializadas ha de estar tambiénexactamente programada con respecto al tiempo y a su secuencia de aparición.
En diferentes células del organismo,o en la misma célula en diferentescircunstancias, se expresan diferentes genes. Al igual que las procariotas, lascélulas eucariotas contienen proteínasreguladoras de genes que se unen asecuencias específicas del ADN, parafacilitar o inhibir la transcripción. Losdistintos tipos celulares del organismopluricelular contienen, pues, conjuntosde proteínas reguladoras de genes distintos y, debido a ello, cada tipocelular transcribe grupos de genesdistintos.

CONTROL DE LA EXPRESIÓN GÉNICA EN EUCARIOTAS (II)
El control de la expresión génica en eucariotas puede realizarse a distintos niveles:.
- Factores de transcripción- Grado de condensación de la cromatina- Grado de metilación de los genes
-Empalme alternativode exones
-Mecanismos que determinan la supervivencia del ARNm en el citosolo si el ARNm es o no traducido
-Mecanismos que determinan la activación o desactivación de una proteína o el tiempo de supervivencia de la misma
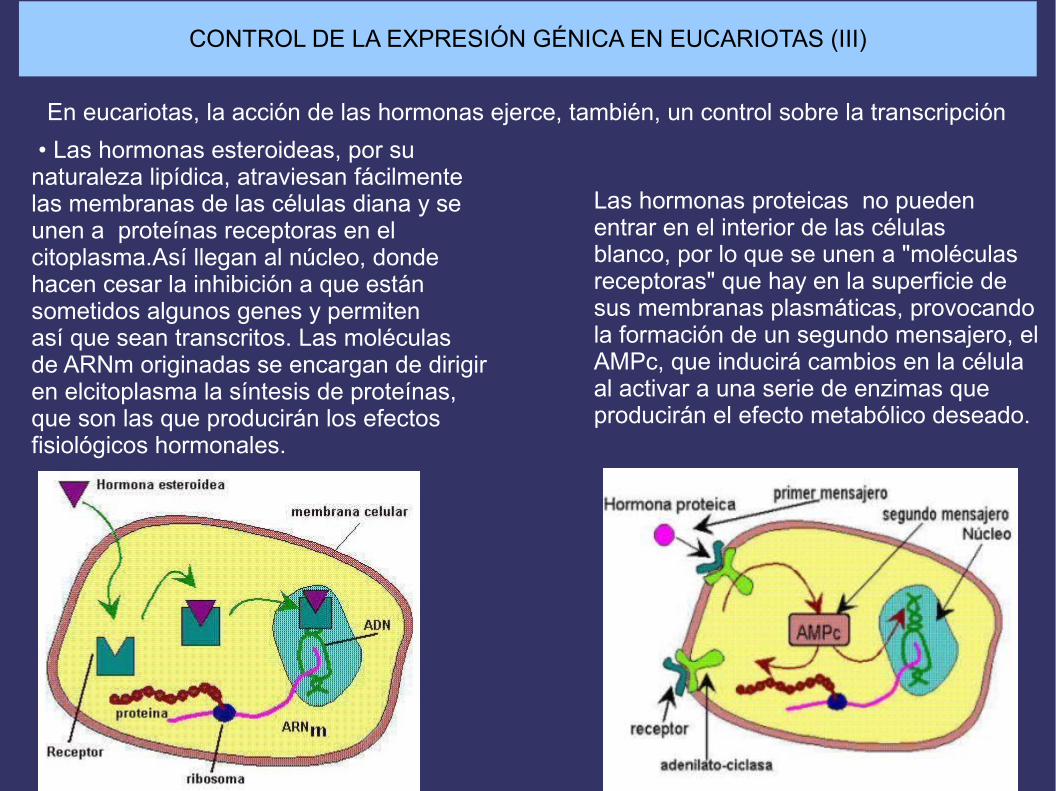
CONTROL DE LA EXPRESIÓN GÉNICA EN EUCARIOTAS (III)
En eucariotas, la acción de las hormonas ejerce, también, un control sobre la transcripción • Las hormonas esteroideas, por su naturaleza lipídica, atraviesan fácilmente las membranas de las células diana y se unen a proteínas receptoras en el citoplasma.Así llegan al núcleo, donde hacen cesar la inhibición a que están sometidos algunos genes y permiten así que sean transcritos. Las moléculas de ARNm originadas se encargan de dirigir en elcitoplasma la síntesis de proteínas, que son las que producirán los efectos fisiológicos hormonales.
Las hormonas proteicas no pueden entrar en el interior de las células blanco, por lo que se unen a "moléculas receptoras" que hay en la superficie de sus membranas plasmáticas, provocando la formación de un segundo mensajero, el AMPc, que inducirá cambios en la célula al activar a una serie de enzimas que producirán el efecto metabólico deseado.
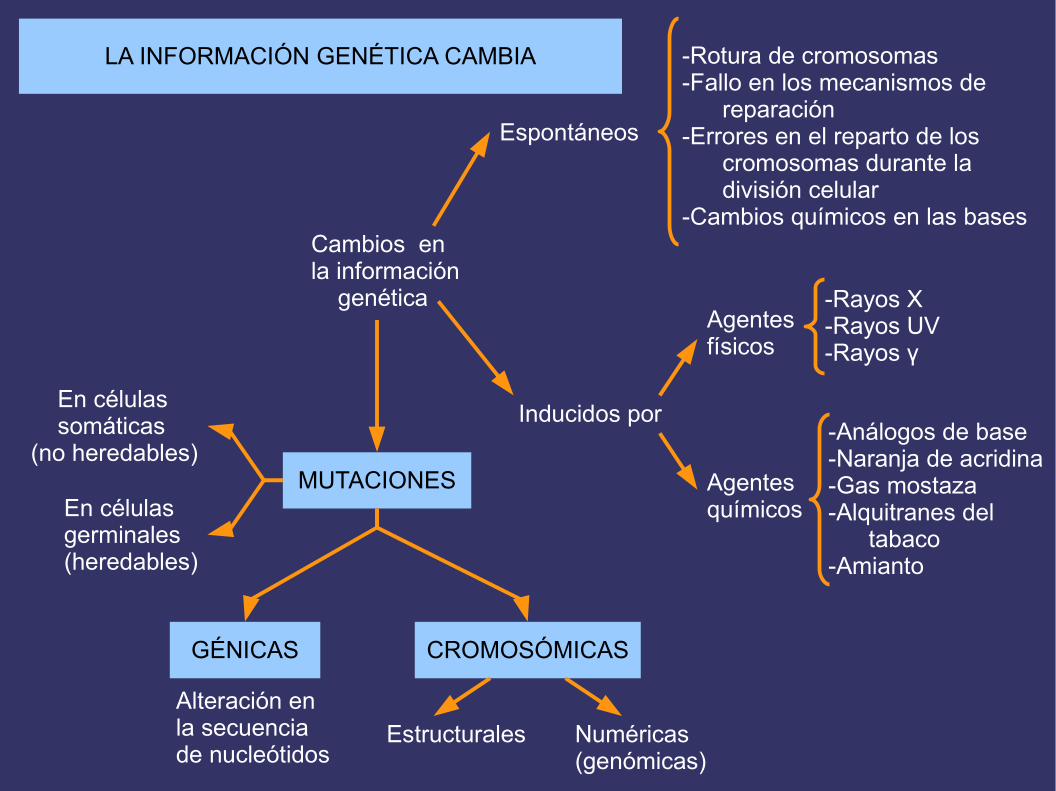
Cambios en la información genética
Espontáneos
Inducidos por
-Rotura de cromosomas-Fallo en los mecanismos de reparación-Errores en el reparto de los cromosomas durante la división celular-Cambios químicos en las bases
Agentes físicos
-Rayos X-Rayos UV-Rayos γ
Agentesquímicos
-Análogos de base-Naranja de acridina-Gas mostaza-Alquitranes del tabaco-Amianto
MUTACIONES
En células somáticas(no heredables)
En célulasgerminales (heredables)
GÉNICAS CROMOSÓMICAS
Alteración enla secuencia de nucleótidos
Estructurales Numéricas(genómicas)
LA INFORMACIÓN GENÉTICA CAMBIA

MUTACIONES GÉNICAS
Sustituciones Cambio de una base por otra
Inserciones o AdicionesGanancia de uno o masnucleótidos
Deleciones Pérdida de uno o mas nucleótidos
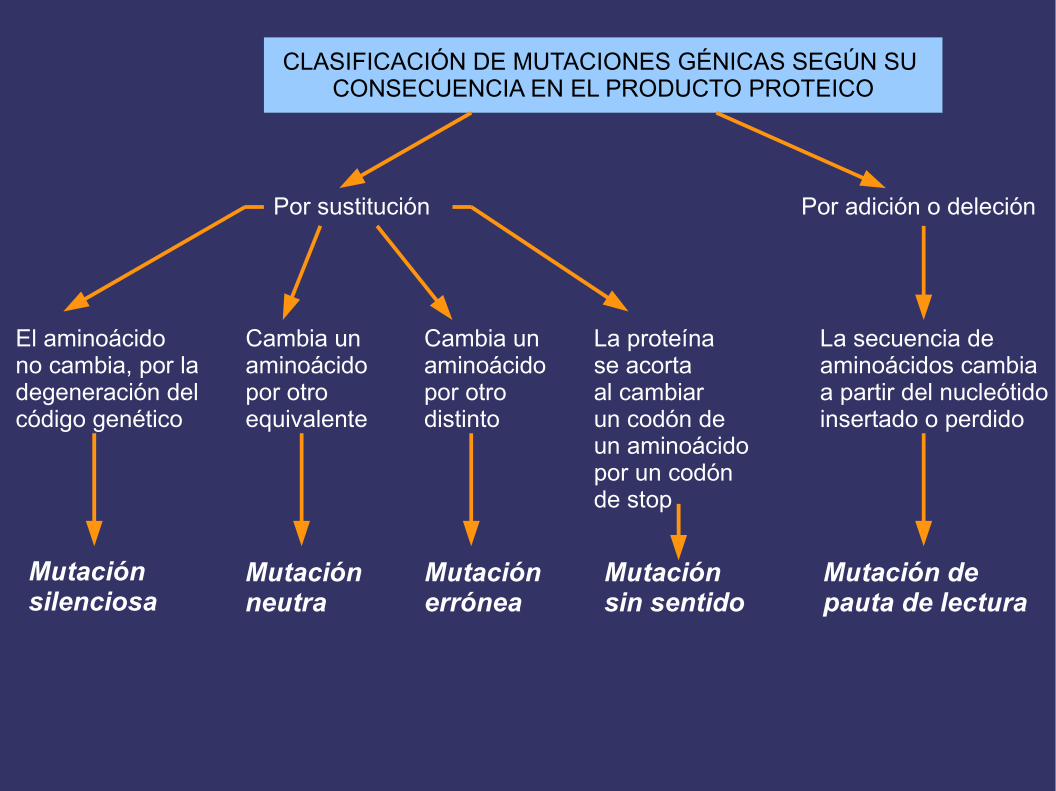
CLASIFICACIÓN DE MUTACIONES GÉNICAS SEGÚN SU CONSECUENCIA EN EL PRODUCTO PROTEICO
Por sustitución
El aminoácido no cambia, por la degeneración del código genético
Cambia unaminoácidopor otroequivalente
Cambia unaminoácidopor otrodistinto
La proteínase acortaal cambiar un codón de un aminoácido por un codón de stop
Por adición o deleción
La secuencia de aminoácidos cambiaa partir del nucleótidoinsertado o perdido
Mutaciónsilenciosa
Mutación neutra
Mutaciónerrónea
Mutación sin sentido
Mutación depauta de lectura


UNA MUTACIÓN GÉNICA POR SUSTITUCIÓN ES LA CAUSA DE LA ANEMIA FALCIFORME
La mutación provoca el cambio del aminoácido enposición 6 de la cadena beta de la hemoglobina,que se traduce en una hemoglobina que formapolímeros, produciendo glóbulos rojos en forma de hoz.

MUTACIONES CROMOSÓMICAS
DELECIONES DUPLICACIONES INVERSIONES TRANSLOCACIONES

Las características fenotípicas son microcefalia, hipertelorismo, pliegues epicánticos, orejas de implantación baja, a veces con apéndices preauriculares y micrognatia. Otros problemas son el retraso mental grave y las malformaciones cardíacas
Síndrome del maullido de gato o cri du chat: deleción del brazo corto del cromosoma 5
Vídeo
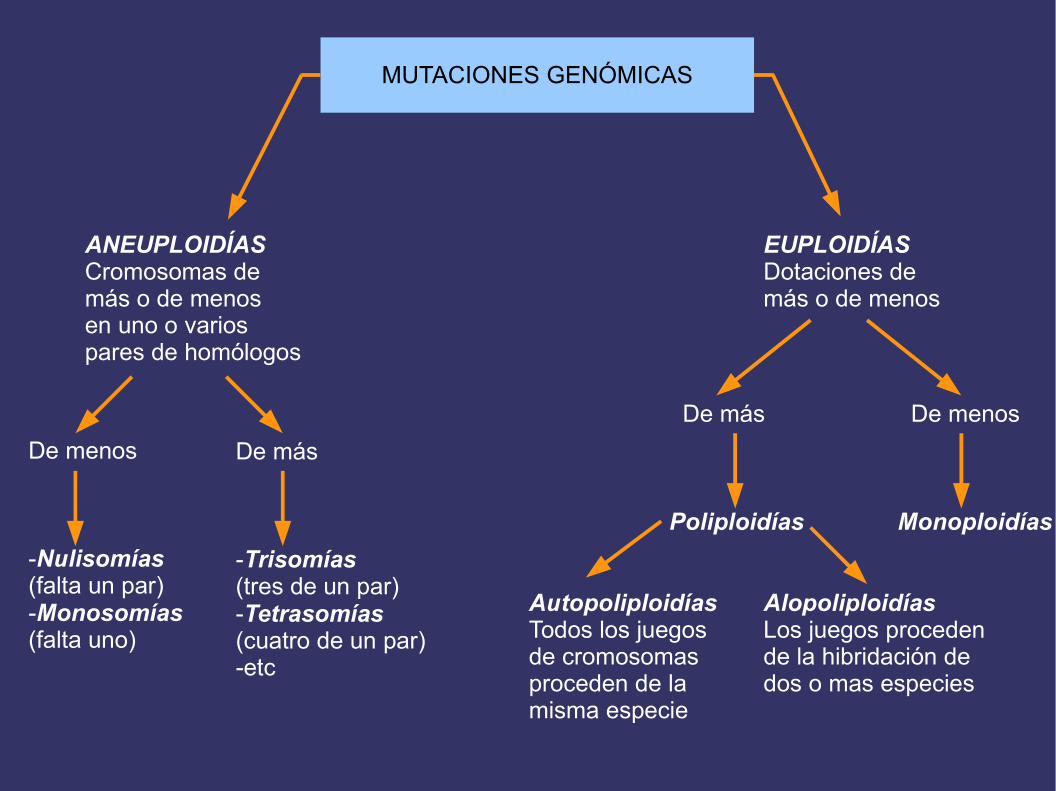
MUTACIONES GENÓMICAS
ANEUPLOIDÍASCromosomas de más o de menosen uno o variospares de homólogos
EUPLOIDÍASDotaciones de más o de menos
De menos
-Nulisomías(falta un par)-Monosomías(falta uno)
De más
-Trisomías(tres de un par)-Tetrasomías(cuatro de un par)-etc
De más
Poliploidías
AutopoliploidíasTodos los juegosde cromosomasproceden de lamisma especie
AlopoliploidíasLos juegos procedende la hibridación dedos o mas especies
De menos
Monoploidías

LA NO DISYUNCIÓN MEIÓTICA PUEDE SER CAUSA DE LAS ANEUPLOIDÍAS
Durante la meiosis, para la formación de gametos, puede haber fallos en la separación de los cromosomas homólogos durante la primera división meiótica, o en la separación de lascromátidas hermanas en la segunda. En cualquiera de estos casos los gametos no recibenel mismo número de cromosomas, es decir, se formarán gametos con cromosomas de maso de menos y, si esos gametos intervienen en la fecundación, originarán zigotos con unnúmero anormal de cromosomas, dando lugar a individuos monosómicos o trisómicos.La no disyunción o no separación de cromosomas puede afectar a cualquier cromosoma, autosómico o sexual. No es un fenómeno exclusivo de la meiosis, sino que puede ocurrirtambién durante la mitosis. Cuando esto ocurre en una célula embrionaria da lugar almosaicismo o individuos llamados mosaicos, que poseen células normales y otras célulascon el número anormal de cromosomas, y pueden presentar algunos o muchos caracteresde un determinado síndrome.

Síndrome de Down: trisomía 21
En el 95% de las personas con este síndrome, el cromosoma extra aparece debido a la no disyunción durante la primera división meiótica, llamándose a esta variante, “trisomía libre”.
En un 3% de los casos, estesíndrome se debe a unatranslocación. El cromosoma 21 extra o un fragmento del mismo se encuentra pegadoa otro cromosoma, que sueleser el 14. A efectos deinformación genética siguetratándose de una trisomía 21 ya que se duplica la dotación genética de ese cromosoma.
En un 2% de los casos la trisomía 21 se produce tras la concepción, por lo que no aparece en todas las células del individuo sino sólo en las que proceden de la primera célula mutada. El porcentaje de células afectadas puede abarcar desde unas pocas a casi todas, según el momento en que se haya producido la segregación anómala de los cromosomas homólogos. Estos individuos se denominan “mosaicos”.
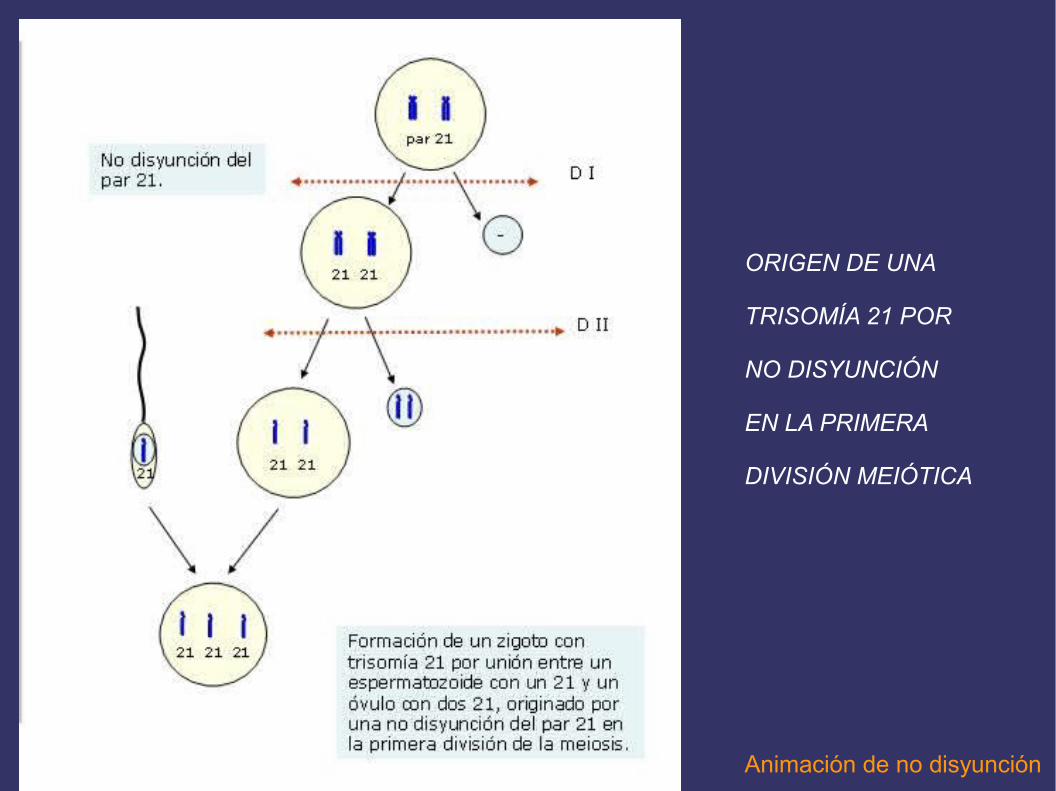
Animación de no disyunción
ORIGEN DE UNA
TRISOMÍA 21 POR
NO DISYUNCIÓN
EN LA PRIMERA
DIVISIÓN MEIÓTICA
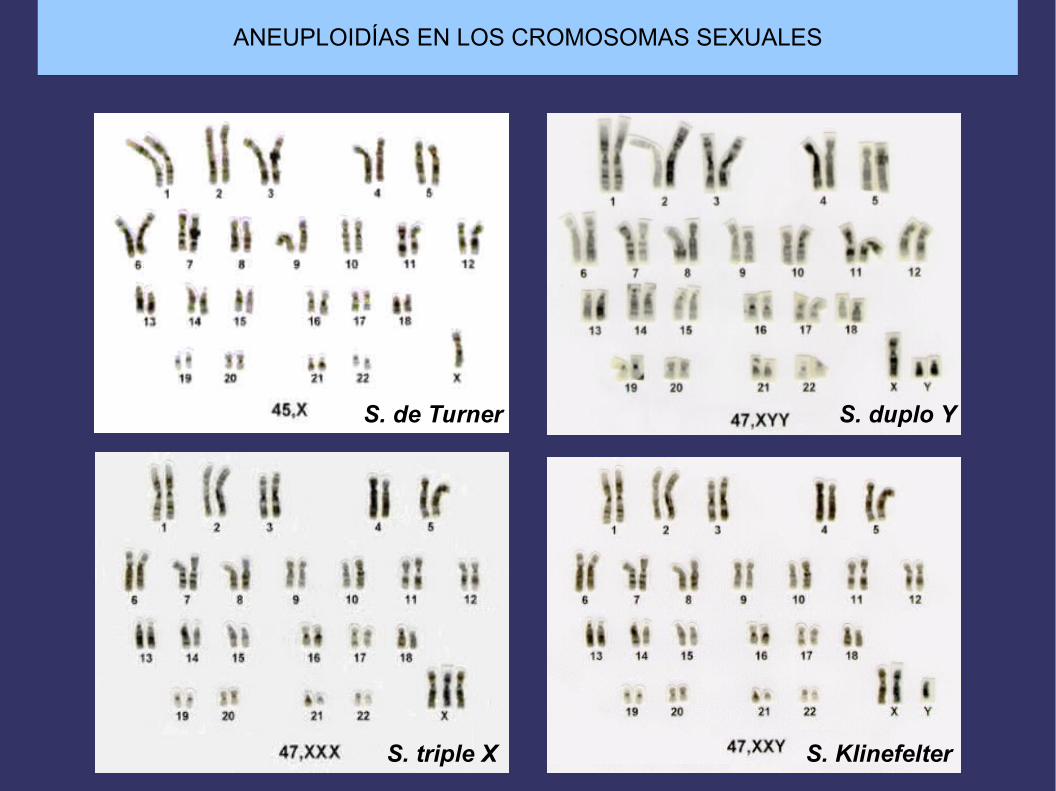
ANEUPLOIDÍAS EN LOS CROMOSOMAS SEXUALES
S. de Turner S. duplo Y
S. triple X S. Klinefelter

Síndrome de Klinefelter
Sindrome de Turner

Un organismo que contiene en sus células somáticas la mitad de los cromosomas pertenecientes a su especie se llama monoploide y si su dotacion cromosómica está compuesta por mas de dos genomas o juegos completos de cromosomas se denomina poliploide (triploide, tetraploide, etc). Si los genomas son iguales, el poliploide es un autopoliploide y si no son iguales, pues proceden de dos o mas especies,entonces se llama alopoliploide. La poliploidía es muy común en plantas pero no en animales, debido a que las plantas toleran mejor que los animales los desbalances genéticos y se pueden propagar vegetativamente. Actualmente, se piensa que entre el 30 y el 70% de las Angiospermas son poliploides. Avena, trigo, cacahuete, manzana, plátano, patata, algodón, caña de azúcar, etc son poliploides. La poliploidía puede producirse espontáneamente, como consecuencia de fallos en la división nuclear, o experimentalmente con compuestos como la colchicina. Las células poliploides son frecuentemente mayores, y en consecuencia también lo son las plantas, frutos, etc. Esto tiene importancia comercial y por eso se utilizan mucho las plantas poliploides en agricultura.
EUPLOIDÍAS
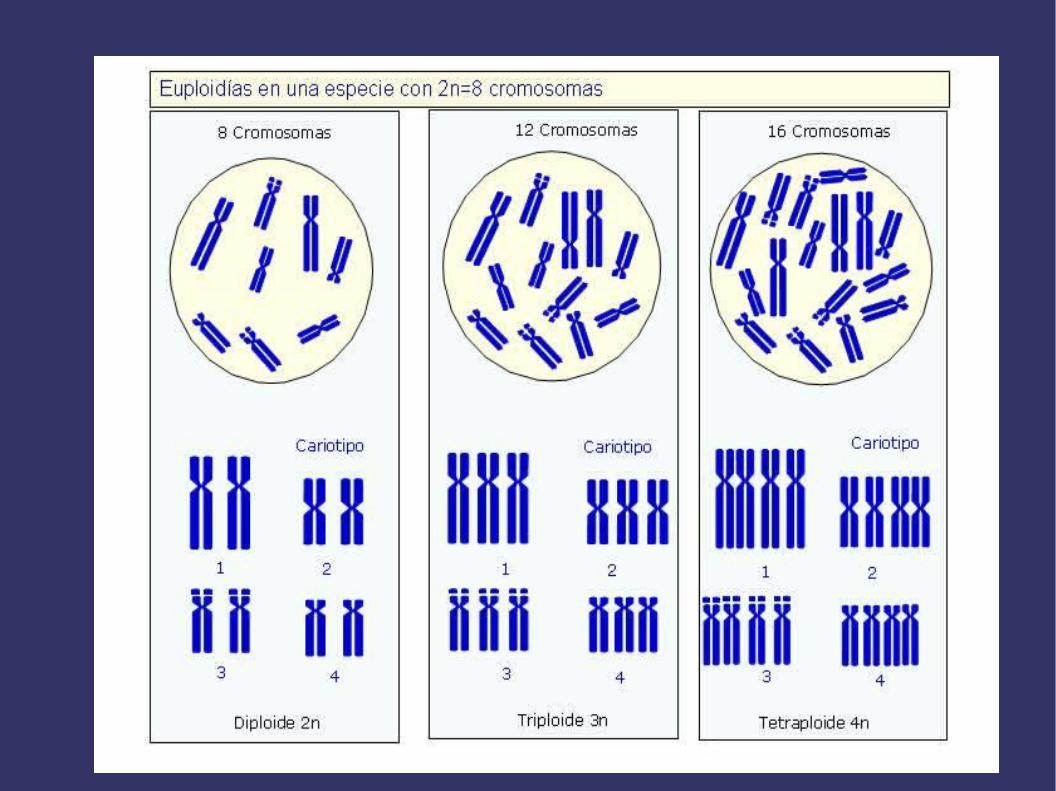


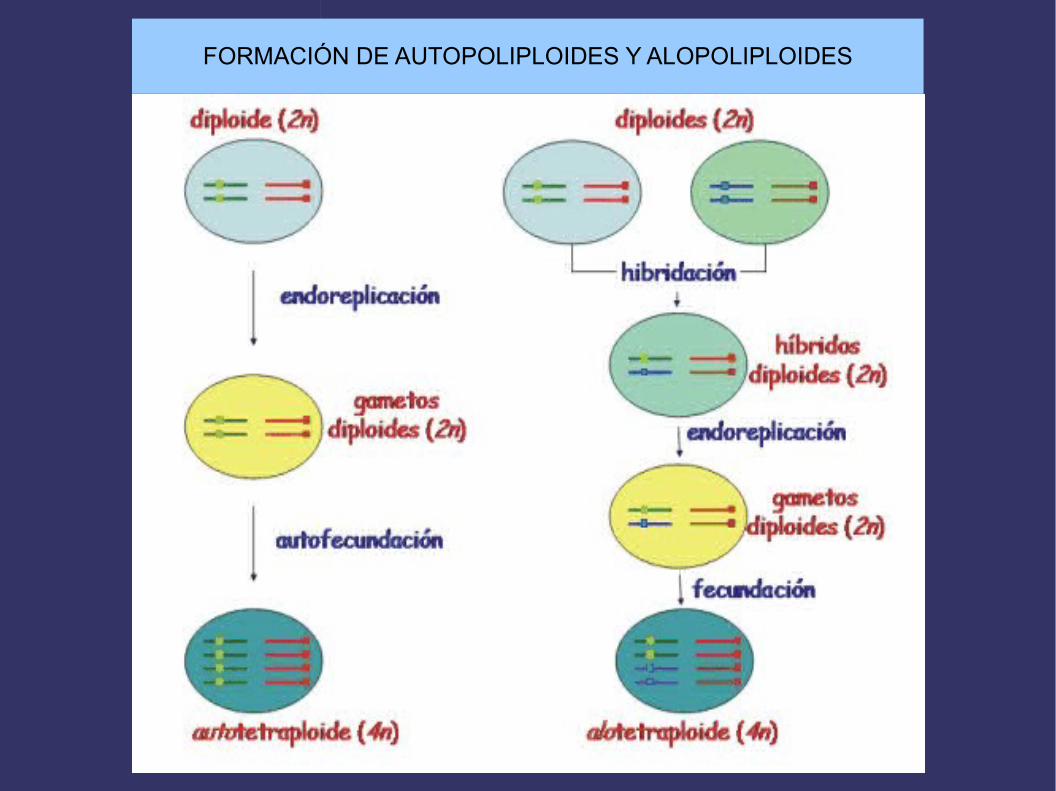
FORMACIÓN DE AUTOPOLIPLOIDES Y ALOPOLIPLOIDES

EVOLUCIÓN DEL TRIGO:El genoma del trigo está formado por varios genomas diferentes

Son aleatorias, no se producencomo respuestaa una necesidad
Produce diferencias anatómicasy funcionales entre los individuos de la población,perjudiciales o beneficiosaspara la adaptación al medio
Las variaciones beneficiosastienden a permanecer en lapoblación, pues aumentan latasa de supervivencia de losindividuos que las poseen, quedejan mas descendientes.Las variaciones perjudicialestienden a desaparecer.
Generan
de la población al ambiente.La población va cambiandoconforme cambian lascondiciones ambientales.
Las especies cambian yse convierten en otras
MUTACIONES
VARIABILIDAD GENÉTICA
SELECCIÓN NATURAL
ADAPTACIÓN
EVOLUCIÓN
RELACIÓN DE LAS MUTACIONES CON LA EVOLUCIÓN:La mutación genera los cambios sobre los que puede operar
la Selección

Darwin postuló que los individuos de una misma especie no son iguales entre sí, sino que presentan variaciones que se pueden transmitir a la descendencia. Estas variaciones hacen que cada uno tenga distintas capacidades para adaptarse al medio natural, reproducirse exitosamente y transmitir sus rasgos a su descendencia. El crecimiento de las poblaciones es exponencial y los recursos son limitados. En la competencia por la supervivencia tiene lugar una selección natural que favorecerá a los individuos mejor adaptados, que tendrán así mas oportunidades de reproducirse y transmitir sus caracteres a la siguiente generación, mientras que los peor adaptados serán eliminados. Con el tiempo, los rasgos de los individuos que mejor se adaptan a las condiciones naturales se vuelven más comunes y la población evoluciona. La acumulación gradual de caracteres ventajosos para distintos ambientes daría lugar, con el tiempo, a la aparición de nuevas especies.Darwin murió sin conocer las causas de la variabilidad en las poblaciones, ya que los resultados de los experimentos de Mendel son posteriores (1866) y apenas tuvieron difusión hasta 1900.
A lo largo de cinco años —entre 1831 y 1836— viajando a bordo del Beagle, Charles Darwin, recogió datos botánicos, zoológicos y geológicos que le permitirían formular, durante los siguientes veinte años, una explicación coherente sobre la diversidad de la vida. En 1858, Darwin se vio obligado a presentar sus trabajos, cuando recibió el manuscrito de un joven naturalista, A. R. Wallace, que había llegado de manera independiente a sus mismas conclusiones , es decir, a la idea de la evolución por medio de la selección natural.Darwin publicó su obra “Sobre el origen de las especies por medio de la selección natural“ en 1859, en la que introduce los conceptos de variabilidad de la descendencia y selección natural.
CHARLES DARWIN Y LA SELECCIÓN NATURAL

MELANISMO INDUSTRIAL: UN EJEMPLO DE EVOLUCIÓN POR SELECCIÓN NATURAL
Biston betularia, la mariposa del abedul, es una mariposanocturna, de la que existen dos variedades: una gris claroy moteada y otra melánica, gris oscura, resultante de una mutación. En Inglaterra, a mediados del siglo XIX, la forma clara era la predominante y la melánica no representaba mas del 1% de las capturas. La contaminación provocadapor la revolución industrial mató los líquenes de los troncosde los bosques y los ennegreció. La consecuencia fue un progresivo aumento de la variedad melánica, que remplazócasi completamente a la variedadad clara en las zonasindustrializadas. El aumento se debía a que las formasclaras destacaban durante el día sobre el tronco negruzcode los árboles, siendo presa fácil de los pájaros que se alimentaban de ellas, mientras que las formas oscurasquedaban ahora camufladas. La selección natural, que en esta caso actúa mediante la contaminación y la depredación,eliminó en mayor medida las formas claras y aumentó la frecuencia de las oscuras, que tienen mayor probabilidad desupervivencia y, por lo tanto, de reproducción, seleccionándose el genotipo mutante. Hoy día,gracias a las medidas anticontaminantes, el proceso se ha invertido y está aumentando lafrecuencia de las formas claras. Este fenómeno, que se ha observado en otras mariposassimilares, tanto americanas como europeas, se denomina melanismo industrial.

Mutaciones en genes específicos del ADN de las células
Reparación o muerte celular
No hay reparación
Células alteradas
Proliferación rápida e incontrolada
TUMOR Formación de una masa celular
METÁSTASISMigración a otros tejidos, a través de la sangre o lalinfa, donde crecen y reemplazan al tejido normal
Distintas proteínas de membranaCambios en la estructura del citosqueleto...
Agentes cancerígenos: físicos, químicos, biológicos
MUTACIONES Y CÁNCER

Autosuficiencia en el crecimiento
Evasión de la Apoptosis
Angiogénesis sostenida
Invasión yMetástasis
Resistencia a estímulos antiproliferativos
Inmortalidad
Cada uno de estos cambios en la fisiología celular, adquiridos durante el desarrollo de los tumores, representa una ventaja en la lucha contra los complejos sistemas de defensa con que cuentan las células paramantener estable la armonía funcional de los tejidos.
Las capacidades que adquieren las células para el desarrollode la enfermedad (Hanahan y Weinberg,2000)
Vídeo de Angiogénesis
Vídeo de Metástasis

Genes reguladores del ciclo celular
PROTOONCOGENES GENES SUPRESORES DE TUMORES
Activan los procesosde crecimiento y proliferación celular
Inhiben los procesosde crecimiento yproliferación celular
Activación permanentepor mutación
ONCOGENESGanancia de función(efecto dominante)
Inactivación por mutación
Pérdida de función(efecto recesivo)Transformación
de una célula normal en maligna
Para que un cáncer pueda progresar y desarrollarse, deben producirse al menos media docena de mutaciones que afecten a varios genes reguladores. Sin embargo, otros tipos de genes también pueden participar en la malignidad, facilitando la capacidad invasiva del tumor (por ejemplo, mutaciones en las proteínas del citoesqueleto que favorecen la motilidad celular).
genes supr. tumores oncogenes

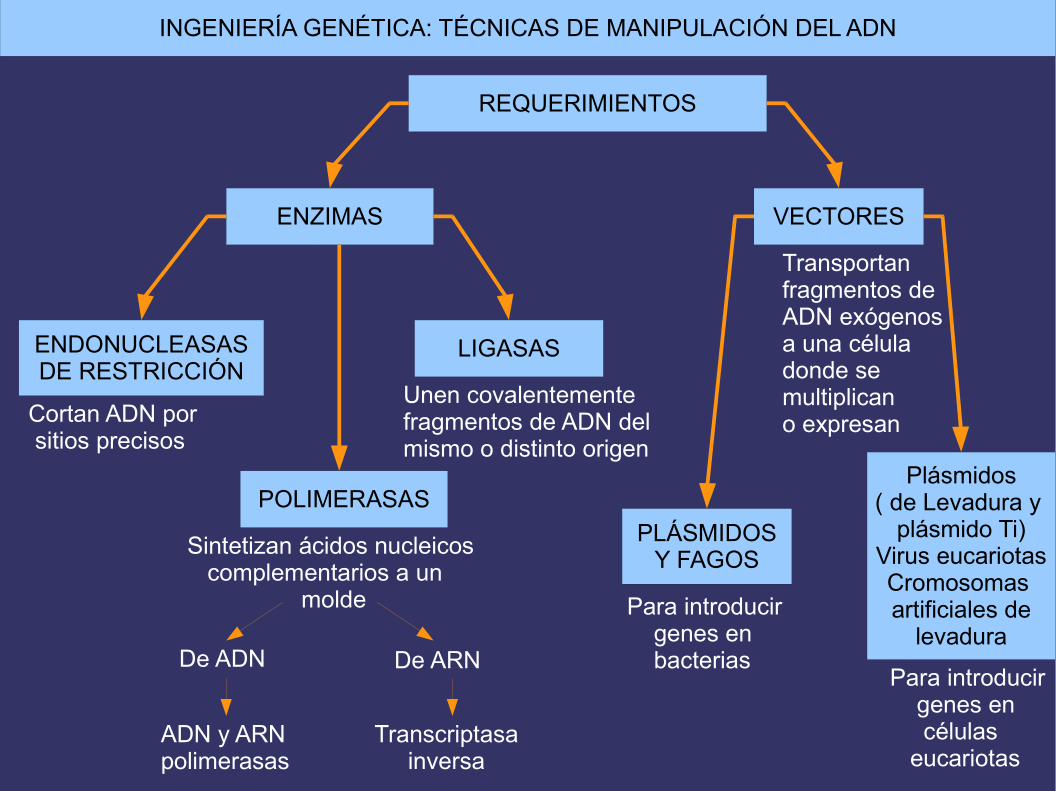
REQUERIMIENTOS
ENZIMAS VECTORES
Transportan fragmentos de ADN exógenos a una céluladonde se multiplican o expresan
ENDONUCLEASASDE RESTRICCIÓN
POLIMERASAS
LIGASAS
Cortan ADN por sitios precisos
Sintetizan ácidos nucleicos complementarios a un molde
Unen covalentementefragmentos de ADN delmismo o distinto origen
De ADN De ARN
ADN y ARN polimerasas
Transcriptasa inversa
Para introducir genes en bacterias
Para introducir genes en células eucariotas
PLÁSMIDOSY FAGOS
Plásmidos( de Levadura y
plásmido Ti)Virus eucariotasCromosomas artificiales de
levadura
INGENIERÍA GENÉTICA: TÉCNICAS DE MANIPULACIÓN DEL ADN
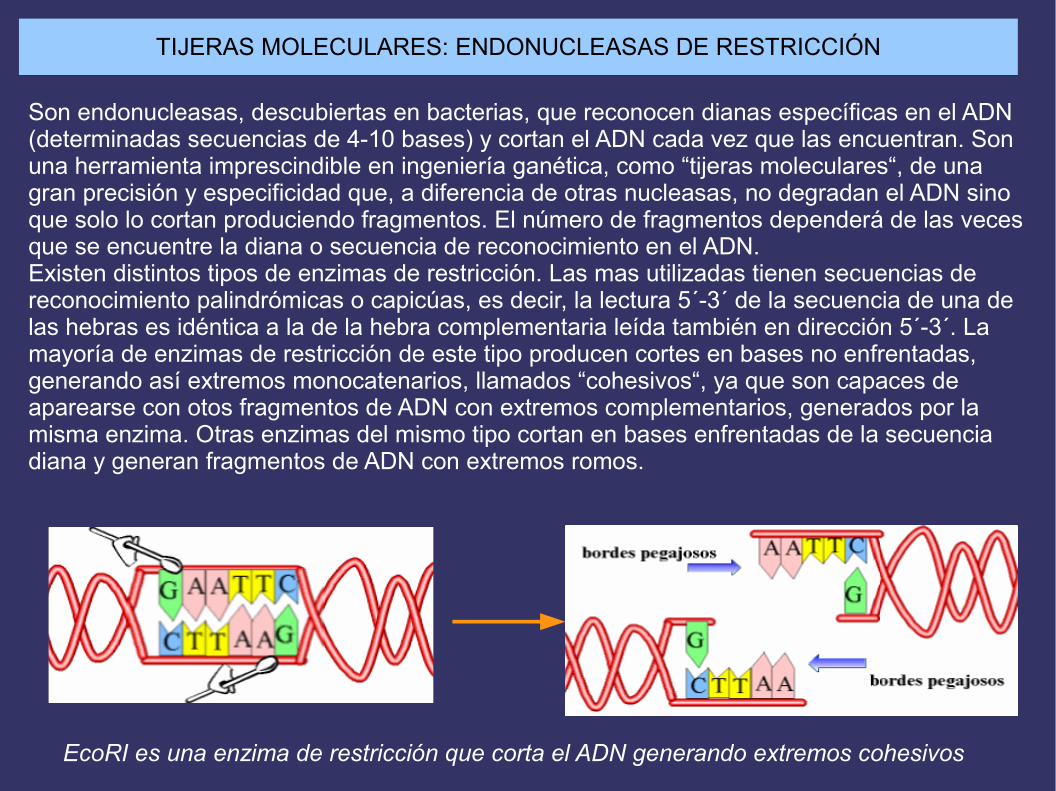
TIJERAS MOLECULARES: ENDONUCLEASAS DE RESTRICCIÓN
Son endonucleasas, descubiertas en bacterias, que reconocen dianas específicas en el ADN (determinadas secuencias de 4-10 bases) y cortan el ADN cada vez que las encuentran. Son una herramienta imprescindible en ingeniería ganética, como “tijeras moleculares“, de una gran precisión y especificidad que, a diferencia de otras nucleasas, no degradan el ADN sino que solo lo cortan produciendo fragmentos. El número de fragmentos dependerá de las vecesque se encuentre la diana o secuencia de reconocimiento en el ADN.Existen distintos tipos de enzimas de restricción. Las mas utilizadas tienen secuencias de reconocimiento palindrómicas o capicúas, es decir, la lectura 5´-3´ de la secuencia de una delas hebras es idéntica a la de la hebra complementaria leída también en dirección 5´-3´. La mayoría de enzimas de restricción de este tipo producen cortes en bases no enfrentadas, generando así extremos monocatenarios, llamados “cohesivos“, ya que son capaces de aparearse con otos fragmentos de ADN con extremos complementarios, generados por la misma enzima. Otras enzimas del mismo tipo cortan en bases enfrentadas de la secuencia diana y generan fragmentos de ADN con extremos romos.
EcoRI es una enzima de restricción que corta el ADN generando extremos cohesivos

SECUENCIAS DIANA DE DOS ENZIMAS DE RESTRICCIÓN QUE PRODUCEN FRAGMENTOS CON DOS TIPOS DE EXTREMOS: COHESIVOS Y ROMOS
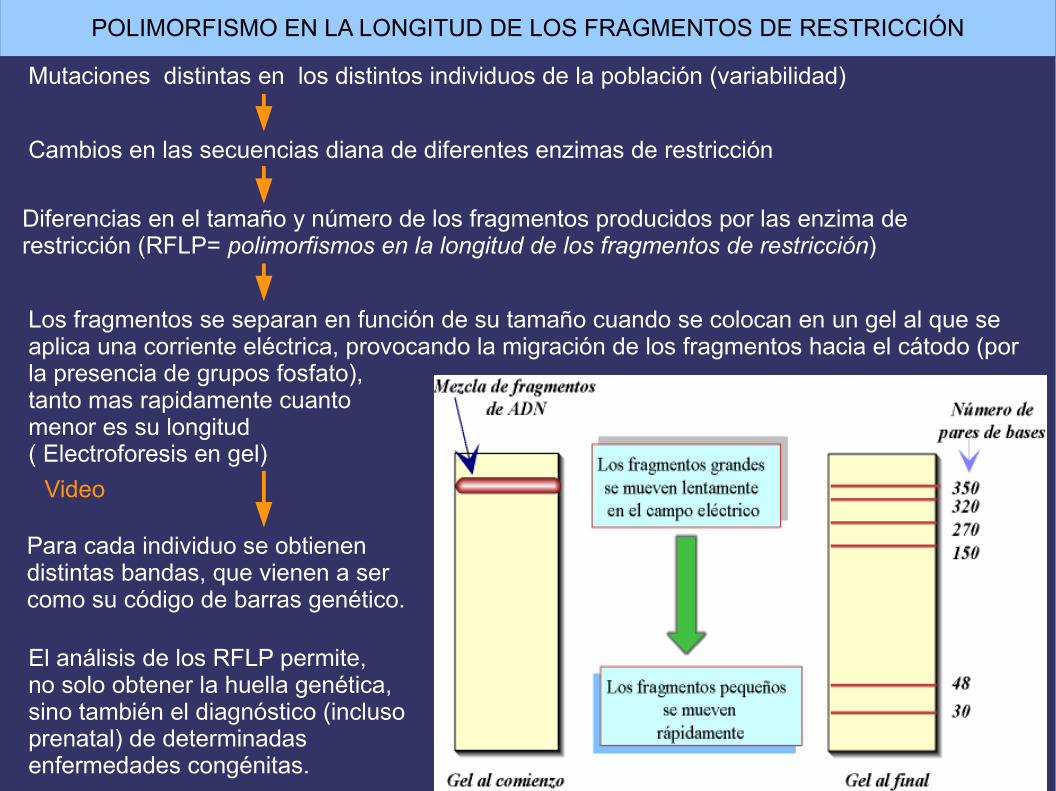
Mutaciones distintas en los distintos individuos de la población (variabilidad)
Cambios en las secuencias diana de diferentes enzimas de restricción
Diferencias en el tamaño y número de los fragmentos producidos por las enzima de restricción (RFLP= polimorfismos en la longitud de los fragmentos de restricción)
Los fragmentos se separan en función de su tamaño cuando se colocan en un gel al que se aplica una corriente eléctrica, provocando la migración de los fragmentos hacia el cátodo (por la presencia de grupos fosfato), tanto mas rapidamente cuanto menor es su longitud( Electroforesis en gel)
Para cada individuo se obtienendistintas bandas, que vienen a sercomo su código de barras genético.
El análisis de los RFLP permite,no solo obtener la huella genética,sino también el diagnóstico (inclusoprenatal) de determinadas enfermedades congénitas.
POLIMORFISMO EN LA LONGITUD DE LOS FRAGMENTOS DE RESTRICCIÓN
Video

CLONACIÓN= OBTENCIÓN DE COPIAS IDÉNTICAS
De un gen o un fragmento de ADN
De células Del genoma de un ser vivo
In vivo
Dentro deuna célulahospedadora
In vitro
Mediantela PCR
Se aísla unacélula y se permite sureproducciónpara generarmuchas célulasidénticas (un clon de células)
Introduciendo todo el genoma en una célulareproductora (óvulo)previamente enucleada(transferencia nuclear)
Se induce el desarrollo embrionario
Implantación en el útero de una madreportadora donde se completará el desarrollo embrionario
Utilización delblastocisto para obtención decélulas madrede las que sepueden obtenerdistintos tejidos
Clonación reproductiva
Clonación terapeútica
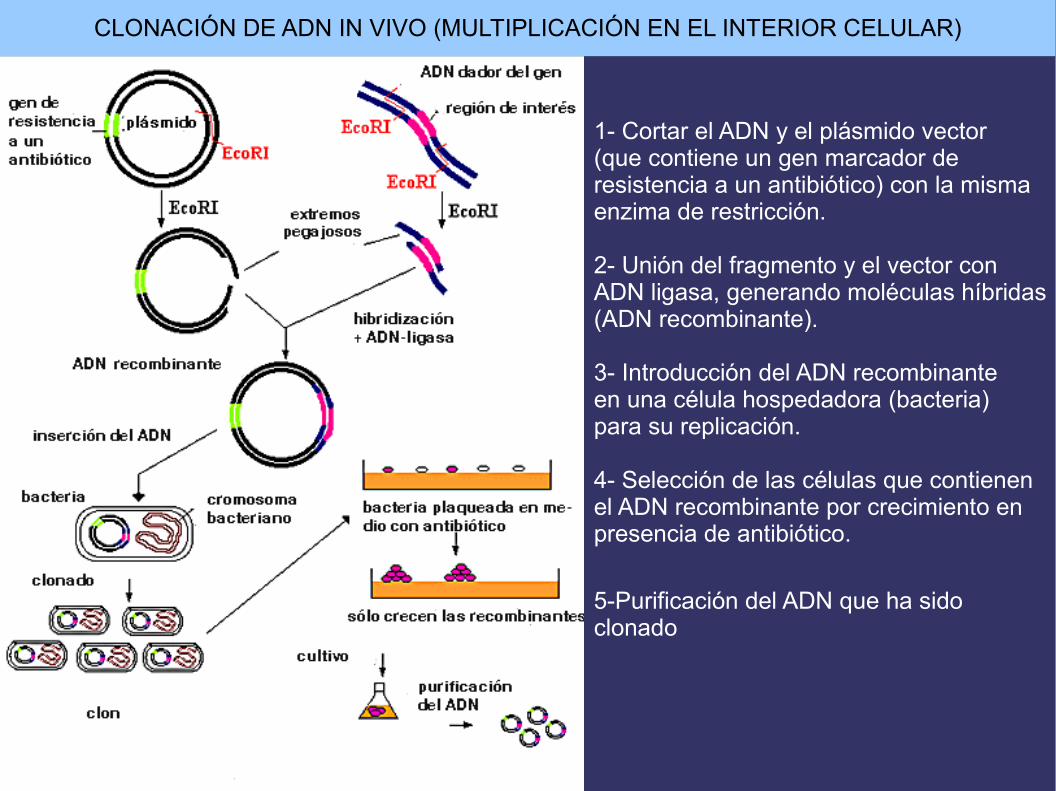
1- Cortar el ADN y el plásmido vector(que contiene un gen marcador deresistencia a un antibiótico) con la misma enzima de restricción.
2- Unión del fragmento y el vector conADN ligasa, generando moléculas híbridas(ADN recombinante).
3- Introducción del ADN recombinante en una célula hospedadora (bacteria)para su replicación.
4- Selección de las células que contienenel ADN recombinante por crecimiento enpresencia de antibiótico.
5-Purificación del ADN que ha sido clonado
CLONACIÓN DE ADN IN VIVO (MULTIPLICACIÓN EN EL INTERIOR CELULAR)

CLONACIÓN TERAPEÚTICA
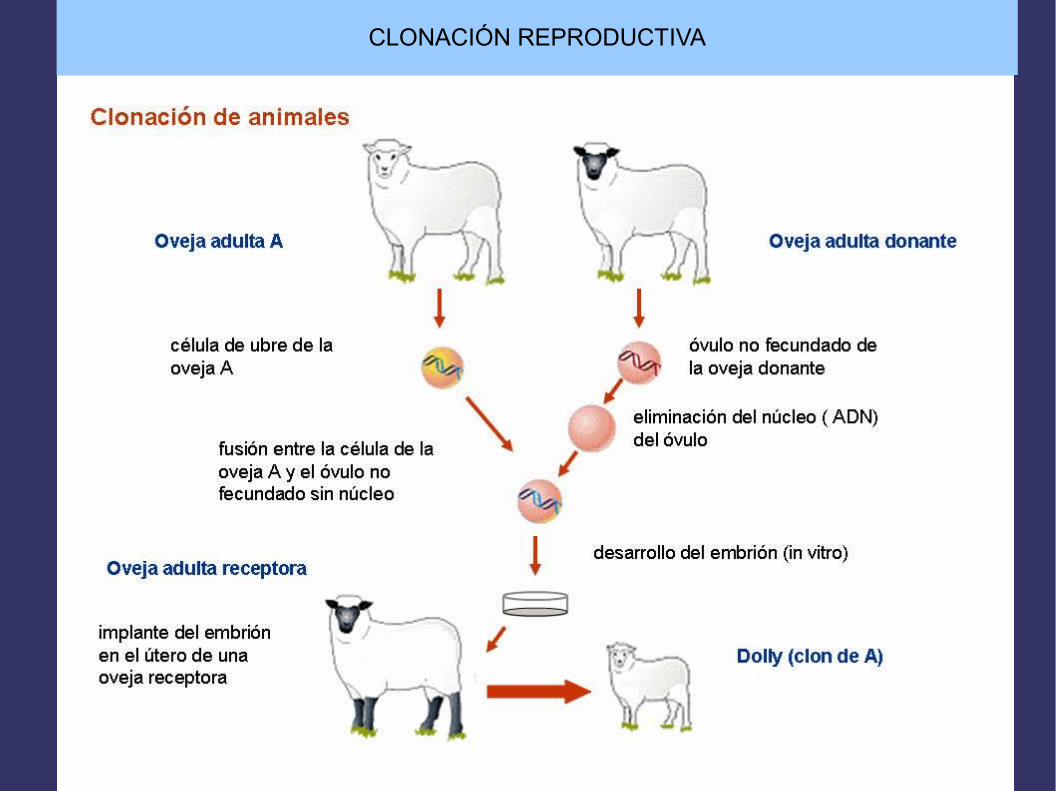
CLONACIÓN REPRODUCTIVA

Obtención de proteínas de interés mediante ingeniería genética
Actualmente se obtienen por ingeniería genética el factor VIII de coagulación, la
hormona del crecimiento, la insulina, el interferón y algunas vacunas antivirales como
las de la hepatitis A y B.

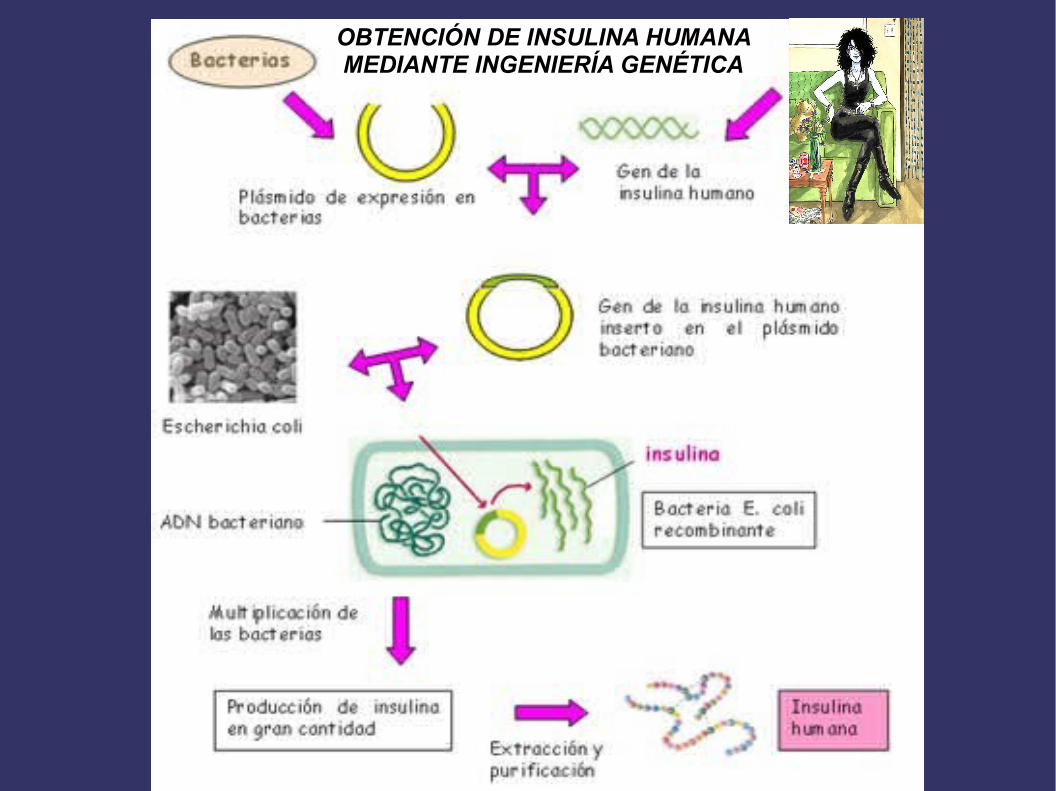
OBTENCIÓN DE INSULINA HUMANA MEDIANTE INGENIERÍA GENÉTICA
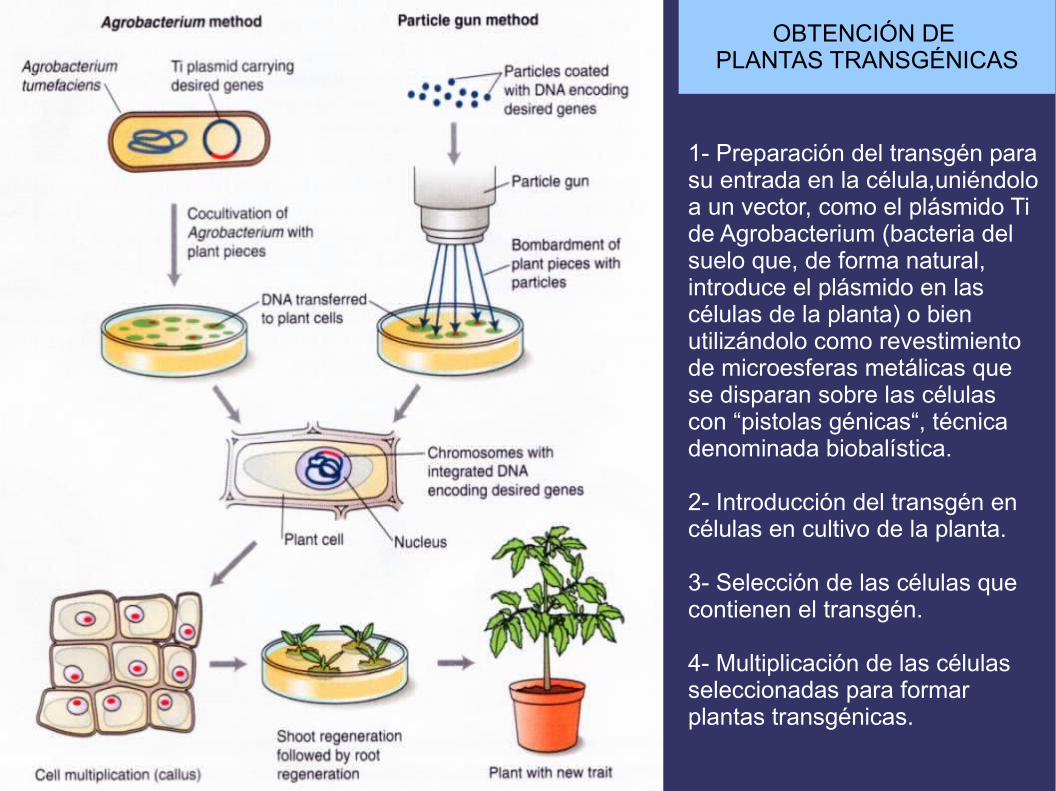
1- Preparación del transgén para su entrada en la célula,uniéndolo a un vector, como el plásmido Tide Agrobacterium (bacteria delsuelo que, de forma natural,introduce el plásmido en las células de la planta) o bien utilizándolo como revestimiento de microesferas metálicas que se disparan sobre las células con “pistolas génicas“, técnicadenominada biobalística.
2- Introducción del transgén en células en cultivo de la planta.
3- Selección de las células que contienen el transgén.
4- Multiplicación de las célulasseleccionadas para formar plantas transgénicas.
OBTENCIÓN DE PLANTAS TRANSGÉNICAS

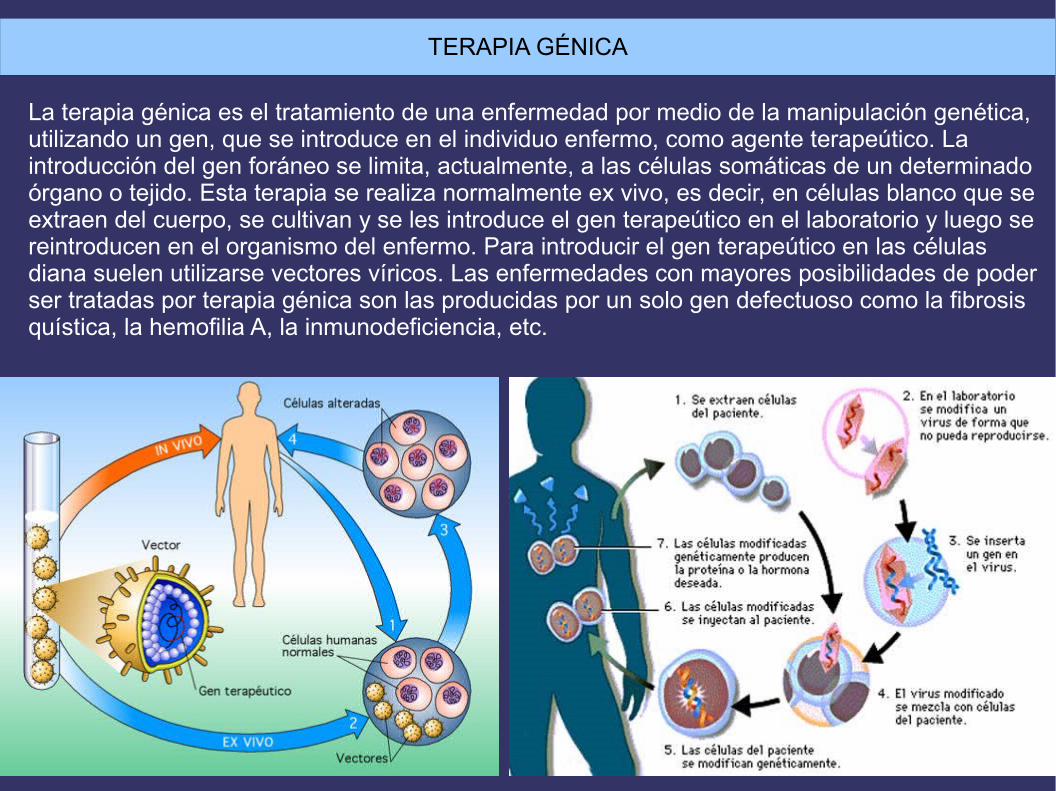
TERAPIA GÉNICA
La terapia génica es el tratamiento de una enfermedad por medio de la manipulación genética,utilizando un gen, que se introduce en el individuo enfermo, como agente terapeútico. Laintroducción del gen foráneo se limita, actualmente, a las células somáticas de un determinadoórgano o tejido. Esta terapia se realiza normalmente ex vivo, es decir, en células blanco que seextraen del cuerpo, se cultivan y se les introduce el gen terapeútico en el laboratorio y luego sereintroducen en el organismo del enfermo. Para introducir el gen terapeútico en las células diana suelen utilizarse vectores víricos. Las enfermedades con mayores posibilidades de poderser tratadas por terapia génica son las producidas por un solo gen defectuoso como la fibrosisquística, la hemofilia A, la inmunodeficiencia, etc.

APLICACIONES EN MEDICINA ▪ Producción de proteínas para tratar o prevenir enfermedades (hormonas, factores de coagulación, interferón, vacunas...). ▪ Terapia génica. APLICACIONES EN AGRICULTURA Y GANADERÍA. Obtención de organismos transgénicos. ♦ Plantas transgénicas. Las aplicaciones agrícolas tienen como objetivo: ▪ Conseguir plantas resistentes a herbicidas, al ataque de insectos o a enfermedades víricas. ▪ Mejora del producto: aumento del valor nutritivo. ▪ Conseguir plantas fijadoras de nitrógeno atmosférico, lo que evitaría la utilización de elevadas cantidades de fertilizantes químicos. ▪ Maduración controlada de frutos y flores. ▪ Tolerancia a condiciones ambientales extremas de temperatura, salinidad... ♦ Animales transgénicos. Las aplicaciones son múltiples, como la mejora en la producción ( los mejores resultados se han obtenido con peces, como el salmón, la carpa y la lubina. A individuos de estas especies se les ha añadido el gen de la hormona del crecimiento, lo que produce un aumento de tamaño del pez en muy poco tiempo. En el salmón se ha introducido otro gen, "el anticongelante", que permite su cría en aguas muy frías) o su utilización en campos distintos como el conocimiento del papel de un gen determinado (su regulación y la función de la proteína) o el estudio de una enfermedad humana de origen genético, la producción de proteínas de interés (humanas o de otro tipo), o la obtención de órganos para xenotransplantes (por ej. de cerdos a los que se les eliminan los genes que codifican las proteínas responsables del rechazo), etc.
APLICACIONES DE LA INGENIERÍA GENÉTICA