memorias de avances de investigaciÓn 2019
TRANSCRIPT

0

0
MEMORIAS DE AVANCES DE INVESTIGACIÓN
POSGRADO EN FITOSANIDAD 2019
COLEGIO DE POSTGRADUADOS
CAMPUS MONTECILLO
EDITORES
OBDULIA LOURDES SEGURA LEON
REMIGIO ANASTACIO GUZMÁN PLAZOLA
MARCO ANTONIO MAGALLANES TAPIA
GONZALO ESPINOSA VÁSQUEZ
COMITÉ ORGANIZADOR
REMIGIO ANASTACIO GUZMÁN PLAZOLA
OBDULIA LOURDES SEGURA LEÓN
MARCO ANTONIO MAGALLANES TAPIA
GONZALO ESPINOSA VÁSQUEZ
COORDINACIÓN DE DIFUSIÓN
CLAUDIA CONTRERAS ORTÍZ
DISEÑO DE PORTADA
JORGE VALDEZ CARRASCO
Montecillo, Texcoco, México, 15 de noviembre de 2019

1
AVANCES DE INVESTIGACIÓN - POSGRADO EN FITOSANIDAD 2019
PRESENTACIÓN
El evento de Avances de Investigación, tuvo su origen en 1990, como una iniciativa dentro
del entonces Instituto de Fitosanidad, ahora Posgrado en Fitosanidad. En 2019, se cumplen
28 años de su inicio y por primera vez; estos se unen al festejo del 60 aniversario de la
creación de los Programas que le dieron su origen: Fitopatología y Entomología, en 1959.
El evento de presentación de los avances de investigación del Posgrado en Fitosanidad
tiene diferentes objetivos académicos, entre estos: a) que los estudiantes de maestría y
doctorado próximos a graduarse como maestros o doctores en ciencias expongan, de forma
oral, los trabajos de investigación que desarrollan; b) que obtengan experiencia para
presentase ante un público académico experto en el área; c) favorecer la interacción
académica entre las diferentes áreas de la Fitosanidad y del Colegio de Postgraduados y d)
conocer los avances científicos y tecnológicos que se desarrollan en favor de la protección
vegetal.
Los trabajos que se presentan son elaborados por estudiantes y miembros de su Consejo
Particular, con el objetivo de garantizar la calidad. En este año, la memoria incluye 42
trabajos de investigación en diferentes áreas de protección vegetal, en las que se incluyen:
cultivos básicos e industriales, hortalizas, frutales, forestales, taxonomía, manejo, análisis de
riesgo, biotecnología y genómica, y salud humana, en donde las diferentes disciplinas se
unen para responder a problemas nacionales con herramientas de vanguardia, en las dos
líneas de investigación del Posgrado registradas ante el CONACYT: 1) Diagnóstico,
Ecología y Manejo Integrado de Plagas y 2) Biotecnología, Inocuidad y Bioseguridad.
COMITÉ ORGANIZADOR

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
2
CONTENIDO
BACTERIAS ASOCIADAS AL PANEL DE PICA EN EL CULTIVO DE HULE (HEVEA
BRASILIENSIS MUELL. ARG.) EN VERACRUZ, MÉXICO 6
DIVERSIDAD BACTERIANA ASOCIADA AL INTESTINO DE TRES DEPREDADORES DE
DACTYLOPIUS OPUNTIAE (HEMIPTERA: DACTYLOPIIDAE) 11
EVALUACIÓN DE ATRAYENTES PARA LA CAPTURA DE ANTHONOMUS EUGENII
(COLEOPTERA: CURCULIONIDAE) EN CAMPO 16
IDENTIFICACIÓN DE PROTEÍNAS DE UNIÓN A ODORANTES (OBPS) DEL
DESCORTEZADOR DE LAS ALTURAS DENDROCTONUS ADJUNCTUS BLANDFORD, 1897 22
INTROGRESIÓN DE LOS GENES DE RESISTENCIA A ROYA AMARILLA YR5 Y YR15 EN LA
VARIEDAD DE TRIGO BORLAUG 100 F2014 27
DETECCIÓN DE PEPPER MILD MOTTLE VIRUS (VIRGAVIRIDAE: TOBAMOVIRUS) POR
MEDIO DE VARIOS JUEGOS DE INICIADORES ESPECIFICOS EN RT-PCR MULTIPLEX 33
EVALUACIÓN DE LA ACTIVIDAD ANTAGÓNICA DE BACILLUS CONTRA CORYNESPORA
CASSIICOLA 38
CARACTERIZACIÓN POLIFÁSICA DE PECTOBACTERIUM Y DICKEYA SPP. INDUCTORAS DE
PUDRICIONES BLANDAS EN MÉXICO 43
IDENTIFICACIÓN Y REGIONALIDAD DE ESPECIES DE FUSARIUM ASOCIADAS A
MARCHITEZ Y PUDRICIÓN SECA DEL COGOLLO EN AGAVE TEQUILANA WEBER VAR.
AZUL 48
ESTIMACIÓN DE CARGA DE INÓCULO IN SITU DE FUSARIUM SPP. EN HIJUELOS DE
AGAVE TEQUILANA A DIFERENTES NIVELES DE INDUCTIVIDAD EPIDÉMICA REGIONAL 54
DENSIDAD DE TRAMPAS CON FEROMONA PARA EL MONITOREO DE STENOMA
CATENIFER (LEPIDOPTERA: ELACHISTIDAE) EN HUERTOS DE AGUACATE HASS 59

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
3
CARACTERIZACIÓN Y DISTRIBUCIÓN DE COLLETOTRICHUM SPP. CAUSANTES DE
ANTRACNOSIS EN FRUTOS DE AGUACATE EN MÉXICO, INCLUYENDO TRES NUEVAS
ESPECIES 65
RESPUESTA A LA INFECCIÓN POR FUSARIUM EUWALLACEAE (HYPOCREALES:
NECTRIACEAE) EN CUATRO VARIEDADES DE AGUACATE EN MÉXICO 71
PATOGENICIDAD DE HONGOS ASOCIADOS A EUWALLACEA NR. FORNICATUS EN
AGUACATE (PERSEA AMERICANA MILL.) CV. HASS 77
ESCAMAS ARMADAS (HEMIPTERA: DIASPIDIDAE) Y SUS PARASITOIDES EN AGUACATE
(PERSEA AMERICANA MILL.) EN CUATRO ENTIDADES DE LA REGIÓN CENTRO
OCCIDENTE DE MÉXICO 83
ESCAMAS (HEMIPTERA: COCCOIDEA) ASOCIADAS AL CULTIVO DE YACA (ARTOCARPUS
HETEROPHYLLUS LAM.) EN NAYARIT 89
POTENCIAL Y ESTABILIDAD DE LA RESISTENCIA DE DIAPHORINA CITRI (HEMIPTERA:
LIVIIDAE) A INSECTICIDAS 95
CICLO DE VIDA Y PARÁMETROS BIOLÓGICOS DE B. CALIFORNICUS Y B. YOTHERSI
(ACARI: TENUIPALPIDAE) EN NARANJA CITRUS SINENSIS 99
IDENTIFICACIÓN DE PLANTAS HERBÁCEAS, ORNAMENTALES Y MALEZAS PORTADORAS
DE CITRUS LEPROSIS VIRUS EN VERACRUZ, MÉXICO 103
CONSUMO DEL ESTADO LARVAL DE TRES ESPECIES DE CRISOPAS ALIMENTADAS
CON MELANAPHIS SACCHARI 110
EVALUACIÓN EN JAULAS DE CAMPO DE DOS PARASITOIDES DE HUEVO DE SPODOPTERA
FRUGIPERDA (LEPIDOPTERA: NOCTUIDAE) 115
PARÁMETROS DEMOGRÁFICOS DE LAETILIA COCCIDIVORA (LEPIDOPTERA: PYRALIDAE)
ALIMENTADO DE COCHINILLA SILVESTRE Y GRANA COCHINILLA 121

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
4
PARÁMETROS DEMOGRÁFICOS DE TRICHOPRIA DROSOPHILAE (HYMENOPTERA:
DIAPRIIDAE) SOBRE TRES HUÉSPEDES DIFERENTES 127
CALIDAD DE LAS FEROMONAS SINTÉTICAS UTILIZADAS EN EL TRAMPEO DE
LEPIDÓPTEROS DE IMPORTANCIA CUARENTENARIA 133
PREFERENCIA DE UN COCCINÉLIDO Y UN DÍPTERO POR LOS DIFERENTES ESTADOS DE
DESARROLLO DE DACTYLOPIUS OPUNTIAE (HEMIPTERA: DACTYLOPIIDAE) 139
PLANTAS CON FLORES COMO ALIMENTO DE PARASITOIDES EN EL CONTROL
BIOLÓGICO DE LA PALOMILLA DORSO DIAMANTE 145
ANÁLISIS DE RIESGO DE PLAGAS PARA PREVENIR LA INTRODUCCIÓN DE ACIDOVORAX
CITRULLI A MÉXICO 150
MODELO GEOESPACIAL DE RIESGO FITOSANITARIO: XYLELLA FASTIDIOSA SUBSP.
FASTIDIOSA EN MÉXICO 155
IDENTIFICACIÓN, HOSPEDANTES Y DAÑOS DE DELIA SPP. (DIPTERA: ANTHOMYIIDAE)
EN CULTIVOS DE CRUCÍFERAS EN MÉXICO 159
REVISIÓN DE LA ESPERMATECA DE LAS ESPECIES DE LA TRIBU XYLEBORINI
(COLEOPTERA: SCOLYTIDAE) PRESENTES EN MÉXICO 165
REVISIÓN DE LA TRIBU PACHYMERINI (COLEOPTERA: BRUCHIDAE: PACHYMERINAE)
PARA MÉXICO Y FILOGENIA DEL GÉNERO CARYOBRUCHUS 170
IDENTIFICACIÓN MORFOLÓGICA DE SIETE ESPECIES DEL GÉNERO CUSCUTA
(CONVOLVULACEAE) DE IMPORTANCIA AGRÍCOLA Y FORESTAL 175
FACTORES BIÓTICOS Y ABIÓTICOS QUE FAVORECEN LA INCIDENCIA Y DESARROLLO
DE LASIODIPLODIA THEOBROMAE EN PINO CARIBEÑO (PINUS CARIBAEA VAR.
HONDURENSIS) 181

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
5
HONGOS ENDÓFITOS: UNA ALTERNATIVA BIOLÓGICA PARA EL MANEJO DE
NEMATODOS AGALLADORES 187
PAQUETE TECNOLÓGICO PARA EL MANEJO DE LOS NEMATODOS NACOBBUS ABERRANS
Y MELOIDOGYNE INCOGNITA EN TOMATE EN INVERNADERO 193
GANANCIAS EN CICLOS DE SELECCIÓN S1 RECURRENTE DE MAÍZ (ZEA MAYS L.) PARA
RENDIMIENTO Y RESISTENCIA AL CARBÓN DE LA ESPIGA (SPORISORIUM REILIANUM
F.SP. ZEAE) 199
¿PUEDE AEDES AEGYPTI L. ADAPATRSE A LA ZONA DEL LAGO DE TEXCOCO Y
AMENAZAR A MÁS DE 36 MILLONES DE PERSONAS? 204
PROPENSIÓN A RESISTENCIA DE AEDES AEGYPTI (DIPTERA: CULICIDAE) AL
INSECTICIDAS BOTÁNICO GREEN CONTROL ULV 209
HONGOS ASOCIADOS AL ESCARABAJO AMBROSIAL XYLOSANDRUS CURTULUS
(EICHHOFF) (COLEOPTERA: SCOLYTIDAE) EN ZIRACUARETIRO, MICHOACÁN, MÉXICO
215
ASOCIACIÓN DE XYLELLA FASTIDIOSA CON DIFERENTES ESPECIES DE PLANTAS
CULTIVADAS Y SILVESTRES EN PARRAS DE LA FUENTE, COAHUILA 220
DIVERSIDAD Y FLUCTUACION POBLACIONAL DE LA ACAROFAUNA ASOCIADA A
ZARZAMORA (RUBUS ULMIFOLIUS) Y PRIMER REPORTE DE DIPTACUS SP. 226
SILICIO COMO POTENCIALIZADOR DE FUNGICIDAS CONTRA BOTRYTIS CINEREA EN
FRUTOS DE ZARZAMORA 231

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
6
BACTERIAS ASOCIADAS AL PANEL DE PICA EN EL CULTIVO
DE HULE (Hevea brasiliensis Muell. Arg.) EN VERACRUZ, MÉXICO
Iris Marley Pérez Gálvez1, Sergio Aranda Ocampo1*, Adriana Rosalía Gijón Hernandez2, Victoria
Ayala Escobar1, Elías Ortíz Cervantes3
1 Posgrado en Fitosanidad-Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco,
Estado de México, México. 2 Cenid Comef, Instituto Nacional de Investigaciones Forestales, Agrícola y Pecuarias. Av. Progreso 5, Santa
Catarina, Coyoacán, CDMX, México. 3Campo Experimental El Palmar, CIR-Golfo Centro, Instituto Nacional de Investigaciones
Forestales, Agrícola y Pecuarias. Km. 16 Carretera Tezonapa-El Palmar, Tezonapa, Veracruz, México. *Autor de correspondencia:
INTRODUCCIÓN
El hule natural se obtiene del látex que producen aproximadamente 895 especies vegetales;
sin embargo, en la actualidad, Hevea brasiliensis Muell Arg. es la única de importancia a
nivel comercial por su alto nivel de producción y calidad del látex, el cual constituye la
materia prima en la industria para la fabricación de una gran cantidad de artículos plásticos
de diversos usos (Picón et al., 1997).
El árbol del hule (H. brasiliensis Muell. Arg.) es originario de la cuenca baja del Río
Amazonas en Brasil y su cultivo inició a finales del siglo XIX (Compagnon, 1998). En
México, este cultivo representa un producto estratégico para el desarrollo de las regiones del
trópico húmedo (Rojo et al., 2005). Los estados con mayor producción de este cultivo son
Veracruz, Chiapas, Tabasco, Oaxaca y Puebla. Veracruz ocupa el primer lugar con una
superficie sembrada de 13 719 ha y una producción de 75 922.65 t (SIAP, 2018).
El árbol de hule es susceptible a diversas enfermedades bióticas producidas por bacterias,
hongos, virus y nematodos, que afectan la raíz, el panel de pica, tallo, ramas y hojas
(ANACAFÉ, 2004). Por lo anterior, las enfermedades se han convertido en la principal
limitante para el óptimo desarrollo del cultivo de hule en América. En México, no existen
investigaciones que hayan abordado el estudio de las enfermedades causadas por bacterias
en este cultivo. Por lo anterior, el objetivo de esta investigación fue caracterizar e identificar,
mediante un enfoque polifásico, las bacterias asociadas al cultivo de hule en el estado de
Veracruz, México.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
7
MATERIALES Y MÉTODOS
Muestreo. Se realizaron muestreos del cultivo de hule del panel de pica y del síntoma
“parche gangrenoso” en Tezonapa, Veracruz, durante los meses de agosto y diciembre de
2018. El material vegetal se depositó en papel estraza y bolsas de plástico para su transporte
y análisis al Colegio de Posgraduados, Campus Montecillo.
Aislamiento de bacterias. Se aislaron los microorganismos asociados al panel de pica y
parche gangrenoso en H. brasiliensis. De cada muestra se tomaron trozos de tejido (0.5 cm)
y se desinfestaron con hipoclorito de sodio (NaOH) al 1.5 % durante 1 min, seguido de tres
lavados con agua destilada estéril. Los trozos de tejido desinfestado se sembraron en agar
nutritivo (AN) y se incubaron a 28 ± 1 °C durante 72 h. Asimismo, trozos del material vegetal
se colocaron en tubos de agua destilada estéril y se realizaron diluciones en serie (10 × 100 -
10 × 10-4). De la suspensión se sembraron 50 µL, por estría cruzada, en medio agar nutritivo
(AN). Las colonias del crecimiento observado se purificaron y transfirieron a los medios B
de King (BK) y AN. Las colonias bacterianas puras se preservaron en tubos criogénicos (2
mL) en agua destilada estéril y en glicerol al 40 %.
Reacción de hipersensibilidad en tabaco y pudrición del tubérculo de papa de
aislamientos bacterianos. Los aislamientos se sometieron a una prueba de reacción de
hipersensibilidad, la cual consistió en infiltrar en el envés de hojas de tabaco (Nicotiana
tabacum), una suspensión bacteriana de 10 × 108 UFC/mL. Para la prueba de pudrición de
papa, se cortaron rebanadas de tubérculo de aproximadamente 0.5 cm y se colocaron en una
charola que contenía sanitas humedecidas con agua destilada estéril. A cada una de las
rebanadas se les hizo una incisión superficial, colocando crecimiento de cada uno de los
aislamientos bacterianos y se incubaron a 28 °C ±1 durante 24 h. Todos los aislamientos que
indujeron una reacción de hipersensibilidad o pudrición blanda en las rodajas de papa, se
seleccionaron para su posterior caracterización e identificación.
Caracterización genética. La identificación molecular de los aislamientos con el método
de Reacción en Cadena de la Polimerasa (PCR), se llevó a cabo con los oligonucleótidos
universales 8F (5´- AGAGTTTGATCCTGGCTCAG) y 1492R (5´-

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
8
GGTTACCTTGTTACGACTT) para la amplificación del gen 16S rADN. Los fragmentos
amplificados se analizaron en geles de agarosa al 1 %, con 0.4 μL de bromuro de etidio y se
visualizaron en un fotodocumentador Infinitity-ST5 Vilber Lourmat. El tamaño del amplicón
se cortó para su purificación y se envió a secuenciar a Macrogen Korea
(http://www.macrogen.com/eng/) en Seúl, República de Corea.
Secuenciación y edición de secuencias. Las secuencias obtenidas se editaron con el
programa BioEdit Sequence Alignment Editor. La secuencia consenso se sometió a un
análisis de alineamiento y homología con la herramienta bioinformática BLAST (Basic Local
Alignment Search Tool) en GenBank del National Center of Biotechnology Information
(NCBI) (https://www.ncbi.nlm.nih.gov/genbank).
RESULTADOS Y DISCUSIÓN
Se obtuvieron 32 aislamientos bacterianos asociados al panel de pica y al parche
gangrenoso que fueron positivos, ya sea a la reacción de hipersensibilidad en tabaco,
pudrición del tubérculo de papa o a ambas pruebas. Los estudios moleculares mostraron que
el tamaño del amplicón fue de aproximadamente 1,450 pb usando los oligonucleótidos 8F y
1492R.
Los alineamientos en el GenBank, de las secuencias obtenidas de la amplificación del gen
16S rADN, identificaron a bacterias como Pseudomonas parafulva con un 99.93 % de
identidad, P. flavescens con 99.42 % y P. putida con 100 % (Cuadro 1), esta última tiene
potencial para el control biológico de Xanthomonas cucurbitae, Acidovorax avenae subsp.
citrulli y Fusarium oxysporum (Montejo et al., 2014).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
9
Cuadro 1. Caracterización de cepas asociadas al panel de pica y parche gangrenoso en
hule.
Número de cepa
Pudrición blanda en tubérculo de papa
Hipersensibilidad en tabaco Fluorescencia Oxidasa Gram RYU Resultado BLAST % de
identidad
1 + + - - - Enterobacteriaceae 99.70 2 + + - - - Pseudomonas parafulva 99.93
3 + + - - - Stenotrophomonas maltophilia 99.93
4 + + + + + Bacillus safensis 99.86 5 + + + + - - Pseudomonas flavescens 99.42 6 + + - - Pseudomonas putida 100 7 + - - - Stenotrophomonas sp. 99.93 8 + - - - Pantoea sp. 98.65 9 + + + + Bacillus paramycoides 100
10 + - - - Enterobacter cloacae 99.93 11 + - - - Stenotrophomonas sp. 99.78
12 + - - - Stenotrophomonas maltophilia 100
13 + + - - Pseudomonas putida 99.85 +: reacción positiva; -: reacción negativa.
En México no existen investigaciones sobre las enfermedades causadas por bacterias en
el cultivo de hule; a nivel mundial, solo se han identificado bacterias del género Pseudomonas
que afecta el follaje, en India (Izquierdo, 2008) y X. campestris causante del tizón en hojas
(Ariharan et al., 2015). En esta investigación, se propone realizar estudios de patogenicidad
de estos aislamientos asociados al panel de pica y parche gangrenoso en los diferentes
materiales genéticos de hule utilizados comercialmente en México.
AGRADECIMIENTOS
Agradecemos al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la
beca otorgada en la maestría para el primer autor. Al Colegio de Postgraduados Campus
Montecillo y CENID-COMEF, INIFAP por el equipo y las instalaciones donde se realizaron
los experimentos.
LITERATURA CITADA
ANACAFÉ. Asociación Nacional del Café. 2004. Cultivo de Hule. Programa de diversificación de ingresos de la empresa cafetalera, Guatemala. 23 p.
Ariharan, V.N., Meena Devi2, V.N., Parameswaran, N.K. y Nagendra Prasad, P. 2015. Occurrence of Bacterial Blight Disease in Rubber Tree (Hevea brasiliensis). Int. J. Pharm. Sci. Rev. Res., 35(2). Article No. 13, Pp. 67-69.
Compagnon, P. 1998. El caucho natural: biología, cultivo producción. Consejo mexicano del hule, A. C. y CIRAD. México. 701 p.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
10
Izquierdo, B. H. 2008. Diagnóstico del manejo de cosecha y aplicación de estimulantes en plantaciones de hule Hevea basiliensis Muell Arg. en Tabasco. Colegio de Postgraduados. 83p.
Montejo, J.G., Mena, C.J., y Núñez, R. Y. 2014. Control biológico de enfermedades mediante el tratamiento de semillas de cucurbitáceas con rizobacterias promotoras del crecimiento de las plantas (PGPR). Agrisost. 20. 12-27.
Picón, R.L., Ortiz, C.E., Hernández, C.J.M. 1997. Manual para el cultivo del hule (Hevea brasiliensis Mull Arg). INIFAP. México. Folleto técnico núm. 18. 128p.
Rojo M., G. E., Jasso M. J. y Vargas H. J. J., Palma, L. D. J. y Velázquez M. A. 2005. Biomasa aérea en Plantaciones comerciales de Hule (Hevea brasiliensis Mull. Arg.) en el Estado de Oaxaca, México. Agrociencia. Vol. 39, numero 004. Colegio de Postgraduados Texcoco, México. Pp. 449-456.
SIAP. Servicio de Información Agroalimentaria y Pesquera. 2018. Estados productores y superficies cultivadas de Hule, Producción agrícola 2018, ciclo: cíclicos y perenes, modalidad: riego + temporal. Disponible en internet: https://nube.siap.gob.mx/cierreagricola/https://nube.siap.gob.mx/cierreagricola/ (Consultado: octubre, 2019).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
11
DIVERSIDAD BACTERIANA ASOCIADA AL INTESTINO DE
TRES DEPREDADORES DE Dactylopius opuntiae (HEMIPTERA:
DACTYLOPIIDAE)
Susana Martínez Martínez1, Esteban Rodríguez-Leyva1*, W. Ariel Guzmán Franco1, Ma. Teresa
Santillán Galicia1, Antonio Hernández López2
1 Posgrado en Fitosanidad-Entomología y Acarología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230
Texcoco, Estado de México, México. 2 ENES-Unidad León, Ciencias Agrogenómicas. Genómica poblacional y evolutiva. Universidad
Nacional Autónoma de México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
Dactylopius opuntiae (Cockerell) (Hemiptera: Dactylopiidae), conocida como cochinilla
silvestre del nopal, es una de las plagas de mayor importancia en el cultivo de nopal [Opuntia
ficus-indica (L.) Miller] en México y varias partes del mundo (Vanegas-Rico et al., 2010;
Mazzeo et al., 2019). Debido a ello se han empleado varios métodos para reducir sus daños,
dentro de los cuales, el biológico se propone como una de las tácticas dentro del manejo
integrado de plagas. En los últimos años se ha reportado que ésta especie de Dactylopius
tiene depredadores en su hábitat natural, entre los más importantes se encuentran el
coccinélido Hyperaspis trifurcata, el díptero Leucopis bellula y el lepidóptero Laetilia
coccidivora (Portillo y Vigueras, 1998; Rodríguez et al., 2010; Vanegas-Rico et al., 2016).
La relación que se establece entre los enemigos naturales (depredadores) y D. opuntiae
resulta interesante, debido a que se ha argumentado que la producción de ácido carmínico
(uno de los distintivos más conocidos de especies del género Dactylopius spp.) disminuye la
depredación. Gran parte de las investigaciones han resaltado la importancia del ácido
carmínico como un mecanismo de defensa en el sistema inmune de D. opuntiae (Hernández-
Hernández et al., 2003), aunque de manera interesante la repelencia de la depredación sólo
se ha demostrado en los mismos depredadores de D. opuntiae que usan el ácido carmínico
para repeler hormigas depredadoras (Eisner et al. 1980, 1994; Stanley, 2006). Aun así, se
desconoce cómo se especializaron esos enemigos naturales para consumir dactilópidos,
debido a que se alimentan de presas que tienen concentraciones de 2-5% de ácido carmínico
de manera rutinaria, lo más interesante que se ha encontrado es cómo L. coccidivora tiene la

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
12
capacidad de alimentarse tanto de D. opuntiae como de D. coccus (8-25% de ácido
carmínico). Varios trabajos respaldan que las comunidades microbianas, que se encuentran
en el intestino de los insectos tienen influencia sobre la alimentación (Chaves et al., 2009;
Nicholson et al., 2012), debido a que los simbiontes microbianos pueden producir enzimas
que les ayudan a degradar las moléculas más grandes y hacerlas más asimilables, además de
la capacidad para depredar cierta especie de insecto (Flórez et al., 2015). Actualmente varias
técnicas moleculares han facilitado la identificación del tipo de simbiontes presentes en el
intestino de los insectos, y las funciones que estos tienen (Chaves et al., 2009). Bajo este
contexto, el objetivo principal de esta investigación es determinar la diversidad bacteriana
presente en los intestinos de tres depredadores de la cochinilla del nopal (coccinélido,
lepidóptero y un díptero) y su posible relación con la capacidad de depredación.
MATERIALES Y MÉTODOS
Material biológico. Los tres depredadores se obtuvieron de las crías que se tienen
establecidas en el laboratorio de control biológico del Posgrado en Fitosanidad del Colegio
de Posgraduados. Se tomaron aproximadamente 10 larvas de los depredadores Leucopis
bellula (L3) y Laetilia coccidivora (L5), y en el caso de Hyperaspis trifurcata (L4) se
utilizaron larvas y adultos. Cada estado de desarrollo de cada especie se colocaba en una
bolsa con CO2 durante 20 minutos, después fueron puestas en una cámara con acetato de etilo
(15 minutos), transcurrido este lapso, se desinfestaron con hipoclorito de sodio al 1% durante
10 minutos, se enjuagaron varias veces con agua destilada estéril y se colocaron en papel
absorbente estéril. Después de esta preparación se procedió a extraer los intestinos de las
larvas y adultos usando unas pinzas entomológicas (no.5); para realizar este proceso se hizo
un corte longitudinal hasta exponer por completo el intestino, el contenido de los 3 segmentos
(estomodeo, mesenteron y proctodeo) se colocó en 100 ml de medio líquido Luria-Bertaini
el cual se mantuvo en agitación constante durante 48 y 72 h.
Aislamiento de bacterias. Se realizaron diluciones seriadas de la 10-1 a la 10-6, se tomaron
100 ml de cada dilución y se colocaron por triplicado en cajas Petri con medio de cultivo
sólido R2A y Luria Bertaini, se distribuyeron con asa Digralsky. Las cajas Petri se colocaron

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
13
en una incubadora a 30 °C hasta que se observó crecimiento de bacterias, posteriormente se
aislaron y purificaron mediante la técnica de estría cruzada (Ramírez-Gama, 2016).
Caracterización micro y macroscópica de las bacterias. La caracterización
microscópica se realizó por medio de la tinción de Gram, ésta es la primera herramienta para
determinar el grupo (Gram positivo o negativo) y su morfología (cocos, bacilos, cocobacilos,
espirales, etc). En la caracterización macroscópica se observaron características como borde
o margen, color, forma, elevación y luz reflejada de la colonia (Ramírez-Gamma, 2016).
Extracción de ADN y determinación de especies bacterianas y amplificación del gen
ribosomal. La extracción de DNA se realizó con el Kit DNeasy Plant Mini kit (Qiagen),
siguiendo el protocolo de extracción de acuerdo al fabricante. Para la determinación de
especies bacterianas de los intestinos de los depredadores se analizarán las muestras de ADN
de cada especie, para ello, se usará el ADN extraído en las reacciones de PCR. El ADN
bacteriano se determinará molecularmente y se utilizarán los iniciadores universales 8F (5´-
AGAGTTTGATCCTGGCTCAG-3´) y 1492R (5´- GGTTACCTTGTTACGACTT- 3´), las
condiciones térmicas serán las siguientes: 1 ciclo de 95°C por 15 min; 30 ciclos de 95°C por
1 min, 54°C por 1 min, y 72°C por 2 min; y finalmente una extensión a 72°C por 10 min
(Galkiewicz y Kellogg, 2008). Los productos de PCR se enviarán a la compañía Macrogen
(Corea) para su secuenciación directa.
RESULTADOS
De manera general, se obtuvieron 53 cepas bacterianas de los 3 depredadores, de las cuales
29 se aislaron del medio de cultivo Luria Bertaini y 24 del medio R2A. El número de bacterias
que se aislaron por especie de depredador fue variable: L. bellula (7), L. coccidivora (7), H.
trifurcata larva (24), adulto (15). La caracterización micro y macroscópica de las bacterias
que se obtuvieron en este trabajo se emplearán para continuar el desarrollo de la
investigación. Aquí se proporciona sólo una muestra de esa caracterización de un depredador
(Cuadro 1).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
14
Cuadro 1. Características micro y macroscópicas de las bacterias aisladas del intestino de
Laetilia coccidivora.
Extracción de ADN. Para conocer las cantidades de ADN extraído de las diferentes cepas
bacterianas se empleó el NanoDrop, el cual determinó la concentración de éste ácido nucleico
en un microlitro de la muestra (Figura 1), así mismo el ADN de las cepas bacterianas se
corrió en un gel de agarosa al 1.2% y se le encontró con un peso molecular de 1 kb, tal como
se observa en la Figura antes mencionada.
Figura 1. Presencia de ADN de las cepas bacterianas de los depredadores Leucopis bellula
y Laetilia coccidivora.
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la beca completa
de doctorado otorgada al primer autor.
Cepa Tinción Forma Borde Color Forma Elevación Luz reflejada Textura B1LcL + Bacilos en cadena Ondulado Crema rizoide Convexa Si ViscozaB2LcL + Bacilos en cadena Ondulado Crema rizoide Convexa Si ViscozaB3LcL + Cocos Entero Amarilla Circular Convexa Si ViscozaB4LcL + Cocos en racimo Entero Amarilla Circular Convexa Si ViscozaB5LcL + Bacilos en cadena Entero Amarilla Circular Convexa Si ViscozaB1LcR + Bacilos en cadena Filamentoso Crema rizoide Plana Si ViscozaB2LcR + Bacilos en cadena Filamentoso Crema rizoide Plana Si Viscoza
Características microscópicas Características macroscópicas

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
15
LITERATURA CITADA
Chaves S., Neto M., Tenreiro R. 2009. Insect-symbiont system: From complex relationships to biotechnological applications. Biotechnology Journal, Wiley-VCH Verlag. 4: 1753.
Eisner T, Nowicki S, Goetz M, Meiwald J.1980. Red cochineal dye (carminic acid): its role in nature. Science 208: 1039-1042.
Eisner T, Ziegler R, McCormick JL, Eisner M, Hoebeke ER, and Meinwald J. 1994. Defensive use of an acquired substance (carminic acid) by predaceous insect larvae. Experientia 50: 610-615
Flórez LV., Biedermann P.H.W., Engl T., Kaltenpoth M. 2015. Defensive symbioses of animals with prokaryotic and eukaryotic microorganisms. Nat Prod Rep 32: 904–936.
Galkiewicz J P. y Kellogg, C. A. 2008. Cross-Kingdom Amplification Using Bacteria-Specific Primers: Complications for Studies of Coral Microbial Ecology. Applied and Environmental Microbiology. 74 (24). 7828–7831.
Hernández‐Hernández FC, García-Gil de Muñoz F, Rojas‐Martínez A, Hernández‐Martínez S, and Lanz‐Mendoza H. 2003. Carminic acid dye from the homopteran Dactylopius coccus hemolymph is consumed during treatment with different microbial elicitors. Arch Insect Biochem Physiol 54: 37-45.
Nicholson J., Holmes E., Kinross J., Burcelin R., Gibson G., Jia W., Pettersson S. 2012. Host-gut microbiota metabolic interactions. Science 336: 1262-1267.
Mazzeo G, Nucifora S, Russo A, Suma P. 2019. Dactylopius opuntiae, a new prickly pear cactus pest in the Mediterranean: an overview. Entomol Exp Appl 167: 59-72.
Ramírez-Gama R. M., Luna M. B., Mejía C. A., Velázquez M. O., Tsuzuki R. G., Vierna G. L., Hernández G. L., Müggenburg R. I. 2016. Manual de prácticas de Microbiología general. UNAM. 297 pp.
Rodríguez-Leyva E., Lomelí-Flores J R., Vanegas-Rico J. M. 2010. Enemigos naturales de la grana cochinilla del nopal Dactylopius coccus Costa (Hemiptera: Dactylopiidae). 1-12. In: L. Portillo y A. L. Vigueras (Coord.). Conocimiento y aprovechamiento de la grana cochinilla. Colegio de Postgraduados.
Stanley D. 2006. Prostaglandins and other eicosanoids in insects: biological significance. Ann Rev Entomol 51: 25-44
Vanegas-Rico J. M., Lomeli-Flores J. R., Rodríguez-Leyva E., Mora-Aguilera G, Valdez J. M. 2010. Enemigos naturales de Dactylopius opuntiae (cockerell) en opuntia ficus-indica(l.) Miller en el Centro de México. Acta Zoológica Mexicana. 26: 415-433.
Vanegas-Rico J. M., Rodríguez-Leyva E., Lomeli-Flores J. R., González-Hernández H., Pérez-Panduro A., Mora-Aguilera G. 2016. Biology and life history of Hyperaspis trifurcata feeding on Dactylopius opuntiae. Biocontrol 61: 691-701.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
16
EVALUACIÓN DE ATRAYENTES PARA LA CAPTURA DE
Anthonomus eugenii (COLEOPTERA: CURCULIONIDAE) EN CAMPO
Carlos Fernando Bautista-Hernández1*, Juan Cibrián-Tovar1, Julio César Velázquez-González2,
María del Pilar Rodríguez-Guzmán1
1 Posgrado en Fitosanidad-Entomología y Acarología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230
Texcoco, Estado de México, México. 2 Koppert de México, Circuito el Márquez norte No. 82. Parque Industrial el Márquez, C.P. 76246,
Querétaro, Querétaro, México. * Autor de correspondencia: [email protected]
INTRODUCCIÓN
En México, se producen alrededor de 157, 540 ha de chile por año, distribuidos en todo el
territorio nacional, con una producción de 3, 239, 318 ton (SIAP, 2018). El picudo del chile,
Anthonomus eugenii es la plaga de mayor importancia en el chile, se alimenta de botones
florales, flores y frutos en desarrollo, ocasionando pérdidas de hasta 100 % si no se
implementan medidas de acción oportunas. Los productos químicos son la principal forma
de control; sin embargo, su utilización produce graves daños secundarios para otros
organismos y el ambiente (Avendaño-Meza, 2017). Para su monitoreo, se emplean trampas
amarillas junto con la feromona de agregación; desafortunadamente, su efectividad
disminuye después de la inducción floral de la planta debido a la competencia con los
volátiles desprendidos durante estas etapas. En otros curculionidos se ha reportado un
sinergismo entre la feromona de agregación y los volátiles de las plantas hospederas, en el
caso de A. eugenii, los compuestos volátiles del chile se han identificado y evaluado en
experimentos de laboratorio, no obstante, para emplear estas sustancias en el monitoreo del
insecto, es necesario realizar pruebas en campo. En este trabajo se investigó el rango de
atracción de compuestos sintéticos derivados del chile y la feromona de agregación con la
liberación y recaptura de adultos de A. eugenii a 5, 10, 15, 30 y 60 m de distancia.
MATERIALES Y MÉTODOS
Atrayentes. En la mezcla se utilizó (E)-β-ocimeno (52.7 %) [CHEMOS®], (Z)-β-
ocimeno (6.0 %), 2-Isobutil-3-metoxipirazina (2.6 %), (Z)-3-hexenil acetato (25.3 %) y
terpinoleno (13.1 %) [Sigma Aldrich®]. Se colocaron 500 mg de la mezcla de los

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
17
compuestos, diluidos en aceite mineral en tubos de microcentrífuga con un volumen final de
1 ml. La feromona de agregación de A. eugenii (Z)-2-(3,3-dimetilciclohexilideno) etanol (48
%), (E)-2-(3, dimetilciclohexilideno) etanol (32 %), (Z)-(3,3-dimetilciclohexilideno)
acetaldehído (3 %), (E)-3,3-dimetilciclohexilideno) acetaldehído (2 %), (E)-3,7-dimetil-2,6-
octadien-1-ol (2 %) y (E)-ácido 3,7-dimetil-2,6-octadienoico (13 %) fueron adquiridos en
ferommis®. El aceite esencial se extrajo mediante arrastre de vapor con 100 g de botones
florales de chile cv. poblano utilizando la metodología descrita por Zheljazkov et al., (2013),
con algunas modificaciones.
Insectos. Se colectaron frutos infestados con picudos en el municipio de Villa de
Guadalupe, San Luis Potosí en cultivares de chile serrano durante agosto de 2018 y se
trasladaron al Colegio de Postgraduados. Los adultos se depositaron en recipientes de 3 L en
condiciones controladas de temperatura (26±2 º C) y fotoperiodo (13:11 h luz: oscuridad).
Cada tercer día se retiraron los adultos emergidos y se trasladaron a recipientes nuevos, donde
permanecieron hasta su uso, alimentados continuamente con frutos en desarrollo de chile
jalapeño (≤ 30 mm). Para los experimentos en campo se utilizaron adultos con más de 10
días de edad, separados por sexo de acuerdo a las características descritas por Eller et al.,
(1995). Los insectos se dejaron 12 horas sin alimento y agua antes de las pruebas en campo.
Experimentos en campo. Durante agosto-diciembre del 2018 se estableció el
experimento en un área ubicada en el Colegio de Postgraduados (19.468861 º N, -98.898833
º W), en el municipio de Texcoco, México. El terreno tenía una topografía plana, sin
presencia de cultivo de chile para evitar interferencia de picudos provenientes de campo. Se
utilizaron trampas amarillas (30.5 cm x 15 cm), impregnadas con pegamento (Adhequim®),
donde se colocaron los tratamientos; estos se instalaron un día antes de iniciar el experimento
a una altura de 0.30 m en dirección a los vientos predominantes. Se evaluó una mezcla de
compuestos sintéticos del chile (MS), aceite esencial de botones florales del chile (AE),
feromona de agregación (FA), feromona de agregación: mezcla sintética (FA:MS), feromona
de agregación: aceite esencial (FA:AE) y un testigo (TA). Los tratamientos se distribuyeron
en un diseño bloques completamente al azar, con tres repeticiones por tratamiento con una
separación de 100 m entre cada repetición y tratamiento.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
18
Se liberaron adultos de A. eugenii en los cuatro puntos cardinales de cada tratamiento
durante las 13: 00 a 17: 00 horas de la tarde, de acuerdo al periodo de mayor actividad de
adultos de A. eugenii (Muñiz-Merino et al., 2014). En el primer (5 m) y segundo experimento
(10 m) se liberaron 10 picudos (5 hembras/ 5 machos), en el tercer experimento (15 m) se
liberaron 12 adultos (6 hembras/ 6 machos), para el cuarto experimento (30 m) se liberaron
14 picudos (7 hembras/ 7 machos) y en el quinto experimento (60 m) se liberaron 18 picudos
(9 hembras/ 9 machos). Después de cada evaluación se realizó el monitoreo con la feromona
comercial durante una semana para asegurar que no existían picudos y se dejaron dos
semanas para que el atrayente se disipara. Las trampas se revisaron una vez por semana,
durante 15 días que duró cada experimento en cada distancia evaluada. Los insectos
recapturados se colocaron en hexano para quitar el pegamento y se almacenaron en alcohol
al 70 % hasta la identificación del sexo.
Análisis estadístico. Los datos del experimento se analizaron mediante modelos lineales
mixtos con el programa SPSS v. 25.0 (IBM Corp. 2019), utilizando los tratamientos como
factor fijo y las repeticiones anidadas en semanas como factor aleatorio. Para averiguar si
existía asociación entre las capturas y los diferentes tratamientos se utilizó la prueba de
ANOVA.
RESULTADOS Y DISCUSIÓN
Las trampas ubicadas a 5 m de distancia con FA, FA: MS y FA: AE recapturaron
significativamente más picudos que las trampas con MS, AE y TA (F=31.558; df= 5, 48; P
< 0.05). Las trampas con FA registraron 81 % de adultos, seguida por las trampas con FA:
MS y FA: AE con 72 % de recaptura respectivamente. Similarmente, Figueroa-Castro et al.,
2016 reportaron mayores capturas de S. acupunctatus en trampas con feromona ubicadas a
distancias menores del punto de liberación. En contraste, las trampas con MS y AE
presentaron 20 y 22 % de picudos, en comparación con el control (TA) donde se obtuvo 17
%. Posiblemente estos resultados tengan que ver con el tipo de liberador utilizado, ya que
en ambos casos, los tubos de microcentrífuga se emplearon como dispersores.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
19
Figura 1. Porcentaje de adultos de A. eugenii recapturados a 5 m de distancia. Mezcla
sintética (MS), aceite esencial (AE), feromona de agregación (FA), feromona de agregación:
mezcla sintética (FA: MS), feromona de agregación: aceite esencial (FA: AE) y testigo (TA).
Letras diferentes indican que las medias de los tratamientos son significativamente diferentes
(Tukey, P<0.05).
A 10 m de distancia del punto de liberación, las trampas con FA, FA: MS y FA: AE
presentaron significativamente mayores recapturas en comparación con las trampas con MS,
AE y TA (F=52.509; df= 5, 40; P < 0.05). Las trampas con FA, FA: MS y FA: AE registraron
75 %, 61 % y 63 % de recaptura. Por su parte, las trampas con MS presentaron 16 %, AE con
13 %, mientras que TA registró 8 %. La mezcla sintética utilizada ha mostrado atracción en
pruebas de laboratorio (Muñiz-Merino et al., 2014; Bautista-San Juan et al., 2019), no
obstante, en este experimento, estos solo fueron eficientes a esta distancia. Posiblemente la
adición de otros compuestos liberados durante la alimentación de A. eugenii a la mezcla
evaluada podría mejorar la respuesta del insecto.
Las trampas con FA, FA: MS y FA: AE ubicadas a 15 m de distancia también registraron
significativamente mayores recapturas (F=124.310; df= 5, 40; P < 0.05), donde se obtuvieron
56 %, 44 % y 49 % de recaptura respectivamente. En el cuarto experimento, las trampas
ubicadas a 30 m de distancia las trampas con FA recapturaron 39 % de insectos, mientras que
las trampas con FA: MS y FA: AE registraron 33 y 32 % de adultos. La combinación entre
la feromona y los compuestos provenientes del hospedero no aumentaron el número de
recapturas, como se ha reportado en otros trabajos (Wibe et al., 2014). Posiblemente las
neuronas receptoras de A. eugenii para volátiles vegetales tengan mayor sensibilidad que las
a a
b b b
a
020406080
100
MS AE FA FA: MS FA: AE TA
Picu
dos
reca
ptur
ados
(P
rom
edio
±EE
Tratamiento
AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
20
neuronas receptoras de feromonas como ocurre con A. grandis (Magalhães et al., 2018), por
lo que el insecto necesita cantidades específicas del compuesto para poder detectarla.
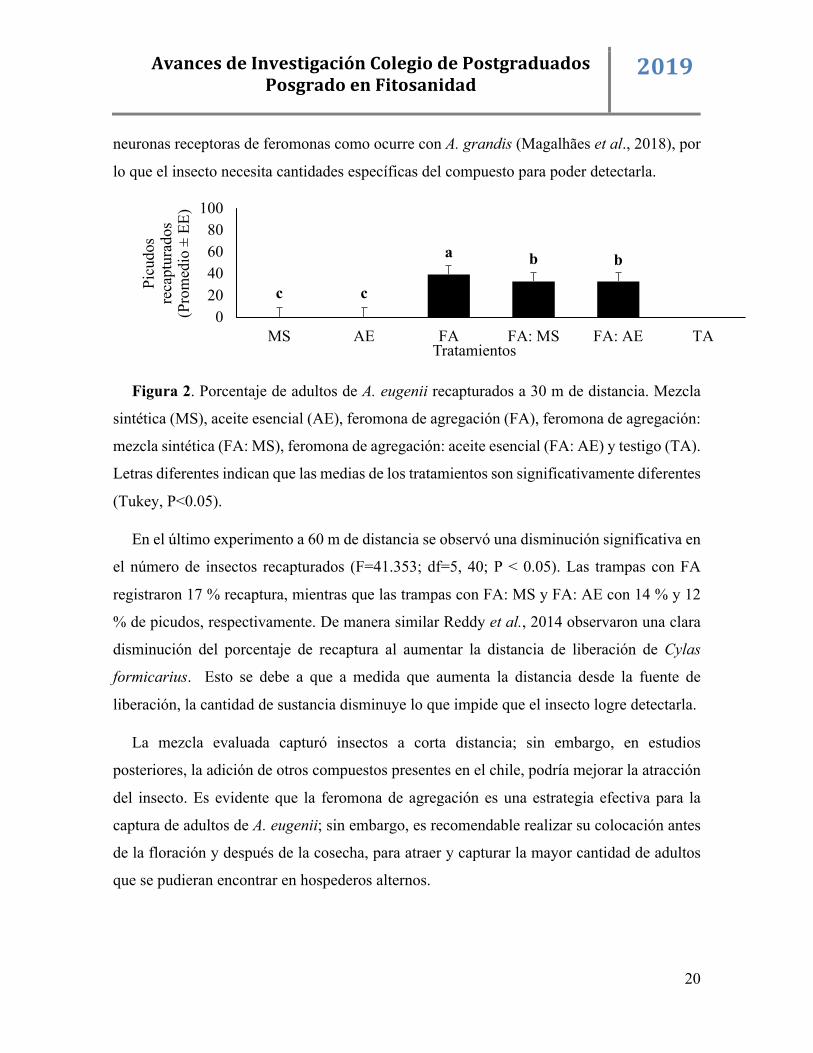
Figura 2. Porcentaje de adultos de A. eugenii recapturados a 30 m de distancia. Mezcla
sintética (MS), aceite esencial (AE), feromona de agregación (FA), feromona de agregación:
mezcla sintética (FA: MS), feromona de agregación: aceite esencial (FA: AE) y testigo (TA).
Letras diferentes indican que las medias de los tratamientos son significativamente diferentes
(Tukey, P<0.05).
En el último experimento a 60 m de distancia se observó una disminución significativa en
el número de insectos recapturados (F=41.353; df=5, 40; P < 0.05). Las trampas con FA
registraron 17 % recaptura, mientras que las trampas con FA: MS y FA: AE con 14 % y 12
% de picudos, respectivamente. De manera similar Reddy et al., 2014 observaron una clara
disminución del porcentaje de recaptura al aumentar la distancia de liberación de Cylas
formicarius. Esto se debe a que a medida que aumenta la distancia desde la fuente de
liberación, la cantidad de sustancia disminuye lo que impide que el insecto logre detectarla.
La mezcla evaluada capturó insectos a corta distancia; sin embargo, en estudios
posteriores, la adición de otros compuestos presentes en el chile, podría mejorar la atracción
del insecto. Es evidente que la feromona de agregación es una estrategia efectiva para la
captura de adultos de A. eugenii; sin embargo, es recomendable realizar su colocación antes
de la floración y después de la cosecha, para atraer y capturar la mayor cantidad de adultos
que se pudieran encontrar en hospederos alternos.
c c
a b b
020406080
100
MS AE FA FA: MS FA: AE TA
Picu
dos
reca
ptur
ados
(P
rom
edio
±EE
)
Tratamientos

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
21
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca otorgada al primer
autor para realizar los estudios de maestría; al Colegio de Postgraduados, Campus Montecillo
por las facilidades otorgadas para realizar el experimento, así como a la Junta Local de
Sanidad Vegetal del Altiplano Centro de San Luis Potosí (JLSVAC) por el apoyo en la
recolección de muestras de chiles infestados.
LITERATURA CITADA
Avendaño-Meza, F. 2017. Efectividad biológica de insecticidas para el manejo de la resistencia del picudo del chile. Revista Iberoamericana de las Ciencias Biológicas y Agropecuarias 6 (11): 23-38.
Bautista-San Juan, A., J. Cibrián-Tovar., R. M. López-Romero, N. Martínez-Bautista. y N S. Gómez-Domínguez. 2019. Atracción de adultos de Anthonomus eugenii (Cano) a mezclas de compuestos volátiles sintéticos. Southwestern Entomologist 44 (3): 743-754.
Eller, F. J. 1995. A previously unknown sexual character for the pepper weevil (Coleoptera: Curculionidae). Florida Entomologist 78 (1): 180-183.
Figueroa-Castro, P., H. Rodríguez-Rebollar., H. González-Hernández., J. F. Solís-Aguilar., J. I. Del Real-Laborde., J. L. Carrillo-Sánchez & J. C. Rojas. 2016. Attraction range and inter-trap distance of pheromone baited traps for monitoring Scyphophorus acupunctatus (Coleoptera: Dryophthoridae) on blue agave. Florida Entomologist 99 (1): 94-99.
Muñiz-Merino, M., J. Cibrián-Tovar, C. Hidalgo-Moreno, N. Bautista-Martínez, H. Vaquera-Huerta y C. Aldama-Aguilera. 2014. Compuestos volátiles atraen al picudo (Anthonomus eugenii Cano) del chile (Capsicum spp.) y presentan sinergia con su feromona de agregación. Agrociencia 48 (8): 819-832.
Magalhães, D. M., M. Borges., R. A. Laumann., C. M. Woodcock., D. M. Withall., J. A. Pickett., M. A. Birkett., M. C. Blassioli-Moraes. 2018. Identification of volatile compounds involved in host location by Anthonomus grandis (Coleoptera: Curculionidae). Frontiers in Ecology and Evolution. 6 (98): 1-12.
Reddy, G. V. P., S. Wu., R. C. Mendi. & R. H. Miller. 2014. Efficacy of pheromone trapping of the Sweetpotato weevil (Coleoptera: Brentidae): based on dose, septum age, attractive radius, and mass trapping. Environmental Entomology 43 (3): 767-773.
Wibe, A., A. K. Borg-Karlon., J. Cross., H. Bichao., M. Fountain., I. Liblikas & L. Sigsgaard. 2014. Combining 1, 4-dimethoxybenzene, the major flower volatile of wild strawberry Fragaria vesca, with the aggregation pheromone of the strawberry blossom weevil Anthonomus rubi improves attraction. Crop Protection 64: 122-128.
Servicio de Información Agroalimentaria y Pesquera, México (SIAP). 2018. Anuario estadístico de la producción agrícola. Recuperado el 19 de junio de 2019 de www.siap.gob.mx/cierre-de-la-produccion-agricola-por-estado.
Zheljazkov, V. D., C. L. Cantrell., T. Astatkie & E. Jeliazkova. 2013. Distillation time effect on lavender essential oil yield and composition. Journal of Oleo Science. 62 (4): 195-199.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
22
IDENTIFICACIÓN DE PROTEÍNAS DE UNIÓN A ODORANTES
(OBPs) DEL DESCORTEZADOR DE LAS ALTURAS Dendroctonus
adjunctus Blandford, 1897
Brenda Torres Huerta1, Obdulia Lourdes Segura León1*, Adriana Rosalía Gijón Hernández2,
Héctor González Hernández1 1 Posgrado en Fitosanidad-Entomología y Acarología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230
Texcoco, Estado de México, México. 2 Centro Nacional de Investigación Disciplinaria en Conservación y Mejoramiento de Ecosistemas
Forestales, Instituto Nacional de Investigaciones Forestales Agrícolas y Pecuarias, Av. Progreso 96, Santa Catarina, 04010, Ciudad de
México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
Los escarabajos descortezadores son un componente integral de los ecosistemas forestales,
sin embargo, existen especies que bajo ciertos factores sus poblaciones se tornan en
epidemias y afectan grandes extensiones de arbolado. La mayoría pertenecen al género
Dendroctonus Erichson (Curculionidae: Scolytinae) el cual contiene los descortezadores de
mayor importancia ecológica y económica en Norteamérica y México, la especie más
importante en alturas elevadas (>2,800 msnm) es D. adjunctus, considerado plaga primaria,
sus infestaciones requieren de la aplicación de campañas emergentes de saneamiento (Hlásny
et al., 2019).
Estos insectos tienen un complejo sistema de comunicación guiado por sustancias
químicas conductuales (semioquímicos) que dirigen comportamientos importantes como la
dispersión, búsqueda, localización de huéspedes y apareamiento. Dado el impacto que
ocasiona a nivel mundial, en los últimos años se ha incrementado la investigación de los
genes que codifican las proteínas involucradas en la percepción de semioquímicos y en el
procesamiento de señales (Shepherd y Sullivan 2013; Fettig, 2013).
Estudios recientes han establecido un punto de partida para el uso potencial de las
proteínas de unión a odorantes (OBPs), consideradas el primer filtro en la detección y
liberación de estímulos químicos del medio externo para la activación de las neuronas
olfativas. Entre las diferentes aplicaciones biotecnológicas para estas proteínas destacan la

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
23
creación de biosensores, la interferencia en los mecanismos de ubicación a los hospederos y
los comportamientos de apareamiento y la supresión del efecto estimulante de sabores
atractivos, por lo que podrían desempeñar un papel importante como parte de las estrategias
en el manejo integrado de plagas (Zhou et al., 2010; Venthur y Zhou, 2018).
Por lo anterior, el objetivo del presente estudio fue analizar el transcriptoma basado en la
secuenciación de siguiente generación RNA-Seq de cabezas de D. adjunctus para identificar
el repertorio de genes olfativos de OBPs involucrados en el procesamiento del olor, debido
a que la identificación de los genes quimiosensoriales y su entendimiento a nivel molecular
tienen potencial para el desarrollo de herramientas que pueden aplicarse en el manejo
integrado de plagas.
MATERIALES Y MÉTODOS
Colecta del material biológico. Los descortezadores se colectaron de forma directa en la
Zona Militar 31 Rancho Nuevo, ubicado en San Cristóbal de las Casas, Chiapas en el 2018,
para ello se seleccionaron tres árboles de diámetros menores a 20 cm y alturas abajo de los
30 metros, con síntomas de brotes activos por descortezador. Se derribaron y con ayuda de
un hacha y pinzas entomológicas se realizó la colecta manual de adultos maduros, los
escarabajos se preservaron en tubos de microcentrífuga con 80 µl de RNAlater™ Storage
Solution, Thermo Fisher Scientific con el fin de mantener integro el RNA. Posteriormente
los tubos se colocaron durante una noche en refrigeración a -4°C y después a -20°C, donde
permanecieron hasta su procesamiento.
Extracción de ARN total, construcción de librerías de cDNA y secuenciación. Se
contabilizaron los descortezadores colectados y se determinaron a especie con la ayuda de
claves taxonómicas (Wood, 1985 y Cibrián et. Al, 1995). Se sexaron por medio del tubérculo
frontal y genitalia, se disectaron 60 cabezas de cada sexo, se separaron en viales con 30µl de
RNAlater™ y se almacenaron a -20°C. La extracción del ARN total se realizó en tres pools
de 40 cabezas (20 hembras y 20 machos) con el kit SV Total RNA Isolation System,
Promega, la calidad y concentración del ARN se midió en un Thermo Scientific™ NanoDrop
2000. Una vez que se cumplió con la calidad requerida, el material genético se envió a BGI

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
24
para la construcción de tres librerías de cDNA y su secuenciación con la plataforma
BGISEQ-500, a Hong Kong, China.
Limpieza y ensamble de secuencias. Se verificó la calidad de las tres librerías con el
programa FastQC v0.10. (Andrews, 2010), la limpieza de adaptadores y reads de baja calidad
no fue necesaria. Se realizó un ensamble de novo con las tres librerías con el programa Trinity
v 2.0.6 (Haas et al., 2013) contenido en el Docker trinityrnaseq con el valor default de
kmer=25. Se verificó la calidad del ensamble completo con el programa rnaQUAST v.0.3.0
(Bushmanova et al., 2016) y se limpió de contaminantes y vectores con las bases de datos
EmVec y UniVec con BLASTn (Camacho et al., 2009) Se obtuvieron los contigs generados
con Trinity y se registró el total de contigs generados, longitud N50, longitud media y el
GC%.
Anotación de Ontología de Genes (GO) y análisis de homología. Para mapear y obtener
información de la función molecular, componente celular y proceso biológico u ontología de
genes (GO) asociados a los unigenes, se utilizó la rutina de mapeo del programa
HMMER2GO-version 0.17.9 (Zenodo, 2019) contenido en el Docker sestaton/HMMER2GO
contra una base de datos de Coleóptera personalizada HMM de Pfam. Su anotación se realizó
con un valor de E de 1e-6, peso GO=5 y cutoff=55 y los resultados de las anotaciones GO se
visualizaron y graficaron en WEGO 2.0 (Ye et al., 2018). Por otro lado, para el análisis de
homología se realizó un BLASTx (Camacho et al., 2009) contra un conjunto de datos que se
construyó con secuencias de proteínas de Insecta de la base de datos UniProtKB y un umbral
de valor E de 1e-5. Los resultados que se obtuvieron se formatearon y se importaron a
Blast2GO (Conesa et al., 2005) para obtener la distribución de especies encontradas en los
hits. Además, se seleccionaron aquellos transcritos cuya descripción y términos GO
asignados, correspondieran con proteínas involucradas en la recepción de olores, en
específico de proteínas de unión a odorantes para su posterior análisis y se reasignaron
nombres en base a la descripción de la anotación de BLASTx.
RESULTADOS Y DISCUSIÓN
Se obtuvieron un total de 198 descortezadores (Curculionidae: Scolytinae) de los cuales
142 se identificaron como Dendroctonus adjunctus y 56 Ips spp.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
25
Limpieza y ensamble de secuencias. Con la plataforma BGISEQ-500 se obtuvo en
promedio 96,631,854 reads (3.5 GB) con una longitud total de 100 pb a partir de tres librerías
de cabezas de machos y hembras de Dendroctonus adjuntus, con un Q20 de 98.13% y GC(%)
de 36.89. El ensamble de las tres librerías resultó en 71,157 transcritos con una longitud
media de 574 pb y una longitud N50 de 3,126 pb con un GC% de 41.2. Posterior a la
agrupación y el filtrado de redundancia se obtuvo un total de 44, 420 unigenes.
Anotación de Ontología de Genes (GO) y análisis de homología. En cuanto a la
anotación funcional, de los 44, 420 unigenes el 32% fueron anotados con base en la
homología de la secuencia. De estos el 19.97% correspondieron a procesos biológicos,
mientras que el 26% y 58.21% a componentes celulares y función molecular
respectivamente. En este ultimó las funciones fueron: de unión (51%) y actividad catalítica
(39%), que según Gu et al., 2014 refleja potencialmente los procesos metabólicos del tejido
antenal. Por otra parte, procesos moleculares (41%) y metabólicos (44%), fueron las
principales subcategorías representadas de procesos biológicos, y en la categoría de
componentes celulares, las partes celulares (22%) se encuentran entre las subcategorías con
más transcritos anotados.
Distribución de especies. Se registraron un total de 142,616 hits al realizar la búsqueda
de homología BlastX con entradas en la base de datos de insecta de UniprotKB con un evalue
de E10-5. En cuanto a la distribución de las especies, el 53.05% de los registros cotejados
tuvieron una compatibilidad con genes de Dendroctonus ponderosae Hopkins, 1902, el
6.46% con Photinus pyralis Linnaeus, 1767 y el 5.84% con Tribolium castaneum Herbst,
1797, todos ellos insectos pertenecientes al orden Coleoptera, el primero y el que obtuvo la
mayor similitud de los transcritos, es su congénere, pertenece a la misma subfamilia
(Scolytinae) de Curculionidae.
Identificación de genes olfativos putativos. Los análisis del transcriptoma de D.
adjunctus se enfocaron a las familias de genes que están asociadas al procesamiento de
olores. Mediante el análisis homológico se identificaron un total de 204 genes, con funciones
de procesamiento olfativo, de los cuales se identificaron 35 genes putativos de proteínas de
unión a olores (OBPs), 12 proteínas chemosensoriales (CSPs), 8 proteínas de membrana de

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
26
neurona sensorial (SNMP), 77 receptores de olor (Ors), 31 receptores ionotropicos (Irs), 20
receptores gustatorios (Grs), 4 receptores ionotropicos glutamicos (IgluRs) y 8 enzimas
putativas de degradación xenobiótica/olorante.
AGRADECIMIENTOS
Agradecemos al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la
beca otorgada en la maestría para el primer autor. Al Colegio de Postgraduados, Campus
Montecillo, por el equipo y las instalaciones donde se realizaron los experimentos y la
asesoría de sus Profesores. Finalmente, al Centro Nacional de Investigación Disciplinaria en
Conservación y Mejoramiento de Ecosistemas Forestales INIFAP por el apoyo y
financiamiento para la colecta del material biológico.
LITERATURA CITADA
Fettig J., C. 2013. Chemical Ecology and Management of bark beetles in Western coniferous forests. Biofertilizers & Biopesticides. 4: 1-2.
Hlásny, T., P. Krokene, A. Liebhold, C. Montagné-Huck, J. Müller, H. Qin, K. Raffa, M-J. Schelhaas, R. Seidl, M. Svoboda y H. Viiri. 2019. Living with bark beetles: impacts, outlook and management options. From Science to Policy. European Forest Institute.
Shepherd P. W., y B. Sullivan T. 2013. Southern Pine Beetle, Dendroctonus frontalis, Antennal and Behavioral Responses to Nonhost Leaf and Bark Volatiles. J Chem Ecol. 39: 481-493.
Venthur, H. y J-J. Zhou Z. 2018. Odorant Receptors and Odorant-Binding Proteins as Insect Pest Control Targets: A Comparative Analysis. Chemosensory Targets for Insect Control. 9: 1-16.
Zhou, Z. J-J., L. Field M. y X. Li. 2010. Insect odorant-binding proteins: do they offer an alternative pest control strategy? Outlooks on Pest Management. 21: 31-34.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
27
INTROGRESIÓN DE LOS GENES DE RESISTENCIA A ROYA
AMARILLA Yr5 Y Yr15 EN LA VARIEDAD DE TRIGO BORLAUG
100 F2014
Yerica Renata Valdez-Rodríguez1, José Sergio Sandoval-Islas1, Julio Huerta-Espino2, Héctor
Eduardo Villaseñor-Mir2, Olga Gómez-Rodríguez1
1Posgrado en Fitosanidad-Fotopatología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco,
Estado de México, México. 2Instittuto Nacional de Investigaciones Forestales Agrícolas y Pecuarias, Campo Experimental Valle de México,
Carretera los Reyes-Texcoco km.13.5, Coatlinchan, Texcoco, Estado de México, México. *Autor de correspondencia:
INTRODUCCIÓN
El trigo (Triticum aestivum L.) es el segundo cereal más cultivado después del maíz. En 2017,
la producción mundial de trigo fue de 771 718 579 t de las cuales México aportó 3 503 521
t (FAOSTAT, 2019). En las zonas productoras de trigo de México existen las condiciones
climatológicas para el desarrollo de las tres royas que lo atacan: la roya del tallo, causada por
el hongo Puccinia graminis f. sp. tritici; la roya de la hoja causada por P. triticina y la roya
lineal o amarilla causada por P. striiformis f. sp. tritici (Huerta-Espino et al., 2014).
La roya amarilla es una enfermedad importante del trigo en Estados Unidos y México; en
este último se han registrado pérdidas de rendimiento del 60-80% (Díaz et al., 2018). El uso
de cultivares resistentes es la mejor estrategia de lucha contra las royas del trigo, ya que es la
vía más económica para controlar y minimizar las pérdidas, porque no ocasiona un costo
adicional para el agricultor y es segura para el medio ambiente (Singh et al., 2002). Sin
embargo, la aparición continua de nuevas razas de roya o el incremento de las poblaciones
de razas antiguas hace necesaria la liberación de variedades nuevas con resistencia a las razas
comunes en las áreas productoras (Solís et al., 2016). Un estudio realizado por Ali et al.,
(2017) en el que se analizaron 887 aislados correspondientes a 79 razas provenientes de 35
países diferentes durante 2009-2015, mostró que no existen reportes de virulencia a los genes
Yr5 y Yr15 que confieren resistencia al hongo causante de la roya amarilla.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
28
Debido a la importancia de estos genes, el presente trabajo tuvo como objetivo transferir
los genes Yr5 y Yr15 a la variedad de trigo harinero Borlaug 100 F2014 y obtener familias
resistentes con los genes Yr5 y Yr15 y analizar sus características.
MATERIALES Y MÉTODOS
La introgresión de los genes se realizó mediante retrocruzas utilizando como progenitor
recurrente a la variedad Borlaug 100 y como progenitor donador a la variedad Blanca Grande
515. Se obtuvieron poblaciones a partir de dos (Bor100*2/BGde515) y tres
(Bor100*3/BGde515) retrocruzas, una vez realizada cada retrocruza se utilizaron marcadores
moleculares para detectar la presencia de los genes Yr5 y Yr15 en diferentes individuos en
las poblaciones segregantes; además se llevó a cabo selección por criterios agronómicos
tomando en cuenta altura y porte de la planta. Se obtuvieron 390 líneas, mismas que fueron
evaluadas en etapa de planta adulta en Chapingo, México, en el Campo Experimental Valle
de México del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias
(INIFAP-CEVAMEX), bajo incidencia natural de roya amarilla. La severidad se evaluó de
acuerdo a la escala de 0-100 (Roelfs et al., 1992).
La evaluación de las líneas a nivel de plántula se realizó en el Laboratorio Nacional de
Royas y otras Enfermedades de Cereales (LANAREC) del INIFAP- CEVAMEX. El material
se sembró en charolas de plástico con sustrato; se evaluaron 25 genotipos por charola. Dos
semanas después de la siembra las plántulas se inocularon con una suspensión de
urediniosporas en aceite mineral (Soltrol® 170) con un atomizador conectado a un compresor
eléctrico utilizando la raza CMEX14.25. Las plántulas inoculadas se colocaron en una
cámara de incubación con temperaturas de 4 - 7 °C por 24 h y rocío al 100% de humedad
relativa por 12 h. Posteriormente se mantuvieron en invernadero y se registró su reacción a
la roya amarilla (tipo de infección) 15 días después de la siembra, usando la escala del 0 - 9
(Roelfs et al.,1992), donde los valores 0, 1, 2, 3, 4, 5 y 6 se consideran como reacciones de
resistencia y 7, 8 y 9, reacciones de susceptibilidad.
El análisis molecular se llevó a cabo en el laboratorio de Biotecnología del Programa de
Mejoramiento Molecular del Trigo en el Centro Internacional de Mejoramiento de Maíz y

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
29
Trigo (CIMMYT). La extracción, cuantificación y valoración de la calidad del ADN se
realizó de acuerdo con los protocolos de laboratorio y aplicaciones para trigo descritos por
Dreisigacker et al. (2016). Los 390 genotipos se evaluaron con los marcadores tipo SNP
(Single Nucleotide Polymorphism) Y5/SPKASP3 y Yr5New para el gen Yr5 (Marchal et al.,
2018), Yr15-R5 y Yr15-R8 para el gen Yr15 (Ramírez-González et al., 2015).
Adicionalmente se utilizaron los marcadores Yr17 (STS) y VPM_SNP (Helguera et al.,
2003) para observar la frecuencia del gen Yr17. En el caso de los marcadores SNP se utilizó
la tecnología de genotipificación Kompetitive Allele Specific PCR genotyping system. Se
utilizaron muestras control FAM y VIC correspondientes a los alelos positivo y negativo,
HET (heterocigotos) y muestra blanco (agua); se ejecutó con la temperatura de amplificación
más favorable de acuerdo con las condiciones de cada marcador (Dreisigacker et al., 2016).
La lectura de las placas del producto de PCR se realizó en lector de placas fluorescentes
BMG Pherastar Plus y para la visualización geográfica de datos genotípicos se utilizó el
Software KlusterCallerTM. Se consideran como positivos los datos genotípicos que
presentaron el alelo favorable para resistencia a roya y como negativos a los datos genotípicos
que no portan el alelo. En el caso del marcador STS para el gen Yr17, las condiciones para
la amplificación fueron de acuerdo Dreisigacker et al. (2016) y el producto de PCR se
visualizó en gel de agarosa al 1.5 %.
RESULTADOS Y DISCUSIÓN
En la evaluación en plántula, 88 líneas fueron susceptibles a la raza CMEX14.25, es decir,
mostraron un tipo de infección de 7- 9 en la escala de Roelfs et al. (1992). Del total de líneas
susceptibles en estado de plántula doce de ellas no amplificaron para ninguno de los tres
genes evaluados, aunque el resto de ellas fue positiva para el gen Yr17; se sabe que la raza
inoculada es virulenta a este gen (Huerta-Espino et al., 2015). Al realizar la evaluación en
plántula se observan tipos de infección de 0-1, mismas que se esperan en presencia de los
genes Yr5 y Yr15 (Roelfs et al., 1992); sin embargo, dado que no hay virulencia para ninguno
de ellos, mediante la evaluación en plántula no es posible determinar si están presentes en
conjunto o por separado.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
30
La evaluación molecular mostró que del total de 390 líneas únicamente cuatro de ellas
fueron positivas a los genes Yr5, Yr15 y Yr17, todas generadas a partir de dos retrocruzas. El
número de líneas positivas a los genes Yr5 y Yr15 de forma aislada es muy bajo en
comparación de aquellas positivas a Yr17. Lo anterior se debe a que el progenitor recurrente
(Borlaug 100) posee el gen Yr17 y conforme se avanza en el número de retrocruzas se va
recuperando un mayor porcentaje del genoma de dicho progenitor. A pesar de que se ha
detectado virulencia hacia Yr17 en varias partes del mundo (Robert et al., 2000), este gen
aún puede proporcionar resistencia contra varias razas de roya si se piramida con otros genes
de resistencia a la roya a amarilla (Iqbal et al., 2016).
Los resultados obtenidos entre los diferentes marcadores para cada uno de los tres genes
evaluados no presentaron diferencias importantes, para el gen Yr5 el 97.9 % de las muestras
coincidieron para ambos marcadores, 93.1 % para Yr15 y 96.9 % en el caso de Yr17.
Cuadro 1. Número de líneas positivas a los genes evaluados de acuerdo al número de retrocruzas y su porcentaje.
Gen BOR*3/BG515 % BOR*2/BG515 %
Ninguno 11 4.4 12 8.6
Yr5 1 0.4 0 0.0
Yr15 2 0.8 4 2.9
Yr17 164 65.6 43 30.7
Yr5+Yr15 0 0.0 1 0.7
Yr5+Yr17 8 3.2 30 21.4
Yr15+Yr17 45 18.0 26 18.6
Yr5+Yr15+Yr17 0 0.0 4 2.9
Total 250
140
Los resultados muestran que dos retrocruzas son suficientes para conservar el genoma del
progenitor recurrente sin perder los genes introducidos, así mismo el uso de marcadores

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
31
moleculares adecuados para cada gen permite mejorar la eficiencia y la velocidad de los
programas de selección de plantas.
AGRADECIMIENTOS
Agradecemos al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la
beca otorgada en la maestría para el primer autor. Al Colegio de Postgraduados, Campus
Montecillo, por las facilidades y la asesoría de sus Profesores. Al laboratorio de
Biotecnología del programa de Mejoramiento Molecular del Trigo en el CIMMYT y al
LANAREC del INIFAP- CEVAMEX por el equipo e instalaciones facilitadas para la
presente investigación.
LITERATURA CITADA
Ali, S., J. Rodriguez-Algaba, T. Thach, C.K. Sørensen, J.G. Hansen, P. Lassen, K. Nazari, D.P. Hodson, A.F. Justesen and M.S. Hovmøller. 2017. Yellow Rust Epidemics Worldwide Were Caused by Pathogen Races from Divergent Genetic Lineages. Front. Plant Sci. 8:1057.
Díaz-Ceniceros H. L, S. G. Leyva-Mir, H. E. Villaseñor-Mir, M. Vargas-Hernández, R. Hortelano-Santa Rosa, Y. R. Valdez-Rodríguez y E. Martínez-Cruz. 2018. Control químico de la roya lineal en diferentes etapas de desarrollo del trigo en Terrenate, Tlaxcala. Revista Mexicana de Ciencias Agrícolas, 9(5), 1067-1074.
Dreisigacker S., D. Sehgal, A. E. Reyes J., B. Luna G., S. Muñoz Z., C. Núñez R., J. Mollins and S. Mall. 2016. CIMMYT Wheat Molecular Genetics: Laboratory Protocols and Applications to Wheat Breeding. CIMMYT. Mexico, D. F. 142 p.
Food and Agriculture Organization of the United Nations. 2019. FAOSTAT Database. Rome, Italy: FAO. Consultado Octubre, 2019. Disponible en: http://faostat3.fao.org/home/E
Helguera, M., I. A. Khan, J. Kolmer, D. Lijavetzky, L. Zhong-Qi, and J. Dubcovsky. 2003. PCR assays for the Lr37-Yr17-Sr38 cluster of rust resistance genes and their use to develop isogenic hard red spring wheat lines. Crop Science, 43(5), 1839-1847.
Huerta-Espino J., M.F. Rodríguez-García, H.E. Villaseñor-Mir, Singh, R.P., E. Martínez C., R. Hortelano S.R., E. Espitia R. 2014. Descripción de las Royas del Trigo. Folleto Técnico Núm. 64. INIFAP-CIRCE-CEVAMEX. pp 32.
Huerta-Espino J., Villaseñor-Mir H. E., Rodriguez-Garcia M. F., Singh R. P. 2015. Emerging new virulence gene combinations in the Mexican Pst population, in Proceedings of the BGRI 2015 Technical Workshop 17-20 September Sydney, NSW.
Iqbal, M., M. Ejaz, I. Ahmed, A. Shahzad, and G. M. Ali. 2016. Molecular Genetic Variation for Stripe Rust Resistance in Spring Wheat. Pak. J. Agri. Sci, 53(1), 143-150.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
32
Marchal C., J Zhang, P. Zhang, P. Fenwick, B. Steuernagel, N.M. Adamski, L. Boyd, R. McIntosh, B.B.H. Wulff, S. Berry. 2018. BED-domaincontaining immune receptors confer diverse resistance spectra to yellow rust. Nat Plants 4: 662–668.
Ramírez-González RH, V. Segovia, N. Bird, P Fenwick, S. Holdgate, S. Berry, P. Jack, M. Caccamo, C. Uauy. 2015. RNA-seq bulked segregant analysis enables the identification of high-resolution genetic markers for breeding in hexaploid wheat. Plant Biotechnol J. 3:613–624
Robert O., F. Dedryver, M. Leconte, B. Rolland and C.D. Vallavieille‐Pope. 2000. Combination of resistance tests and molecular tests to postulate the yellow rust resistance gene Yr17 in bread wheat lines. Plant Breed. 119: 467-472.
Roelfs A. P., R. P. Singh y E. E. Saari. 1992. Las Royas del Trigo: Conceptos y Métodos para el Manejo de esas Enfermedades. CIMMYT. México, D.F. 81 p.
Singh R. P., J. Huerta-Espino and A. P. Roelfs. 2002. The wheat rusts In: Curtis, B. C., Rajaram, S., Gomez, S., and MacPherson, L. (Eds.). Bread wheat: improvement and production. Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO). 227-249 p
Solís-Moya E., J. Huerta-Espino, P. Pérez-Herrera, H. E. Villaseñor-Mir, A. Ramírez-Ramírez y M. de L. Cruz-González. 2016. Alondra F2014, nueva variedad de trigo harinero para el Bajío, México. Revista Mexicana de Ciencias Agrícolas, 7(5), 1225-1229.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
33
DETECCIÓN DE Pepper mild mottle virus (VIRGAVIRIDAE:
Tobamovirus) POR MEDIO DE VARIOS JUEGOS DE INICIADORES
ESPECIFICOS EN RT-PCR MULTIPLEX
Carlos Omar Medina-Molina2, Ana María Hernández-Annguiano1*, Johan Rodríguez-Mendoza2,
Daniel Leobardo Ochoa-Martínez1.
1Posgrado en Fitosanidad-Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco,
Estado de México, México. 2 SENASICA-DGSV-Centro Nacional de Referencia Fitosanitaria, Km 37.5 carretera Federal México-Pachuca,
Tecámac, Estado de México, México. *Autora de correspondencia: [email protected]
INTRODUCCIÓN
Pepper mild mottle virus (PMMoV) perteneciente al género Tobamovirus fue caracterizado
inicialmente como una cepa latente del virus mosaico del tabaco (TMV). Actualmente existen
dudas sobre la identidad y, por lo tanto, sobre la distribución geográfica de los Tobamovirus
que afectan al cultivo de chile (Capsicum annuum) (Wetter et al., 1984). El PMMoV ha sido
reportado en algunos países de América del Norte y Central, África, Australia, Asia y Europa.
(Lamb, 2001).
El genoma de PMMoV está constituido por un RNA de cadena sencilla con 6,357
nucleótidos, organizados en cuatro fragmentos de lectura abierta (ORF, por sus siglas en
inglés, Open Reading Frame) que codifican cuatro proteínas diferentes. El primer ORF
comienza en el nucleótido 70 y se extiende hasta el nucleótido 3,423, codifica una proteína
de 1,117 aminoácidos con un peso molecular estimado de 126,304 Da, denominada 126 K.
La supresión de un codón ámbar (UAG), posibilita la lectura hasta el nucleótido 4,908 y
codifica una proteína de 1,612 aminoácidos con un peso molecular calculado de 183,340 Da,
denominada 183 k; ambas proteínas poseen alta homologías de secuencias con las proteínas
126 k y 183 k del TMV, las cuales están implicadas en el proceso de replicación viral (Quadt
y Jaspars, 1989), por lo que probablemente también están involucradas en la replicación de
PMMoV. El tercer ORF comienza en el nucleótido 4,909 y termina en el 5,682 y codifica
una proteína de 257 aminoácidos con un peso molecular estimado de 28,347 Da que posee
una alta homología de secuencia con la proteína 30 K de TMV que interviene en el
movimiento célula a célula del virus a través de los plasmodesmos (Deom et al., 1987; Meshi

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
34
et. al., 1987). Por último, el cuarto ORF que se extiende entre los nucleótidos 5,685 y el 6,158
codifica a una proteína de 156 aminoácidos con un peso molecular de 17,110 Da, denominada
17.5 K y que es la proteína que forma los capsómeros de la cápside del virus. En todos los
Tobamovirus en los que ha sido estudiada la estrategia de replicación viral, las proteínas
126K y 183K se traducen directamente en RNA viral, mientras que las proteínas 30k y 17.5k
lo hacen a partir de dos RNAm subgenómicos 3´coterminales (Palukaitis et al., 1983).
El diagnóstico molecular de PMMoV se basa en el método de la retrotranscripción
acoplada a la Reacción en Cadena de la Polimerasa, conocida como RT-PCR. Esta técnica
permite la detección y amplificación de regiones específicas del genoma del virus, mediante
una copia de cDNA a partir de una región del genoma de RNA del virus y una reacción
subsecuente de PCR. La PCR multiplex es una variante de la PCR tradicional mediante la
cual se pueden detectar 2 o más fragmentos distintos de DNA diana en la misma reacción.
Es importante diagnosticar al virus con varios juegos de iniciadores, ya que actualmente
existe la probabilidad de que el virus pueda mutar, y los iniciadores flanqueen otros
Tobamovirus. Por lo anterior el objetivo de esta investigación es diseñar varios juegos de
iniciadores inéditos de tal manera que hibriden exclusivamente con el cDNA diana y puedan
trabajar en conjunto en una RT-PCR multiplex dando mayor especificidad.
MATERIALES Y MÉTODOS
Diseño de primers. Para el diseño de primers se descargaron las secuencias de PMMoV
y de todos los Tobamovirus que se encuentran en la base de datos del NCBI. Se realizaron
secuencias consenso de cada Tobamovirus incluyendo el virus de interés. Derivado de que el
virus cuenta con cuatro marcos de lectura abierta, se separaron las regiones de nucleótidos y
mediante el software Expasy, se hizo la traducción de estas secuencias a aminoácidos.
La secuencia de aminoácidos resultantes se sometió al software InterProScan, el cual nos
ayudó a determinar el tipo de proteína resultante, así como su dominio y sitio catalítico.
Posteriormente se determinó la posición del sitio catalítico en la secuencia proteica
mediante un Blast proteico ayudando a seleccionar los aminoácidos que codifican para este
lugar. Al tener los aminoácidos se realizó una traducción inversa para que a su vez se

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
35
pudieran determinar los nucleótidos y su posición involucrados para la síntesis de este sitio
en la proteína.
Teniendo la posición de los nucleótidos codificantes al sitio de la proteína y las secuencias
consenso de Tobamovirus y PMMoV, se establecieron los diferentes oligonucleotidos que se
utilizaron en esta investigación (Figura. 1).
Figura 1. Región de los primers diseñados en el genoma de PMMoV
Una vez seleccionadas las regiones que fungirán como iniciadores, se les realizó un
análisis informático mediante el software OligoAnalyzer, el cual nos arrojó información
como: la temperatura de fusión, datos de complementariedad, homodímeros, heterodímeros,
los valores de ΔG, imágenes de las posibles estructuras secundarias, entre otras. Con este
software se precisaron las temperaturas de fusión de cada juego de primers diseñados, esto
con la finalidad de usarlos en conjunto en una RT-PCR multiplex. Los diferentes juegos de
primers se probaron in silico, lo cual consiste en hacer un Blast en el software de la NCBI.
Una vez hecho esto se decidió que los primers eran funcionales y se mandaron a sintetizar.
RESULTADOS Y DISCUSIÓN
De acuerdo a los ORF presentes en el genoma del virus, se desarrollaron cinco pares de
iniciadores, cada par flanqueando el sitio catalítico de la proteína codificante, en donde se
cuidó la temperatura de fusión, porcentaje de G/C, la formación de estructuras secundarias y
el tamaño del amplicón (Cuadro 1).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
36
Cuadro 1. Iniciadores diseñados.
Nombre del oligo
Secuencia (5´- 3´)
Tm (˚C)
G/C (%)
Tamaño del amplicón
Posición en el genoma
A GCAGAACGCTGTGCACAGTCTCGC 64.6 62.5 1113
297
A´ GCCTTATCCACATCCCACTCTGAGCGC 64.5 59.3 1410
B GTAGAGTCGCAGTGAGCTCCGAATCTTTGGTG 64.2 53.1 1021
2481
B´ GCATGTCAGAAACATCTCCTGTTTTTGGTGCTGC 64 47.1 3502
C GCCTACACAGATCGAAGAATTTTTCTCAGATCTGGACTCTAATGTTCC 64.5 41.7 417
4158
C´ CCAGAGAAGGTTTGCGCCCTGTTGAATATCGG 64.9 53.1 4575
D CGCTGTCGGTGAGGTTATTAAAACCGCACC 64.1 53.3
679
4800
D´ CTGTAAGTTCAACGGGTCCTCCTTCTGACACAC 63.9 51.5 5479
E GGACGCCGAGTCTTCTTCGTTTTAACTATGGCTTACAC 64.4 47.4 509
5658
E´ CCATCATGTTTAAGGAGTTGTAGCCCAGGTGAGTCC 64.7 50 6167
Al probar los iniciadores diseñados con muestras reales se obtuvieron los dos amplicones
esperados para los pares de oligos C y E. Estos oligos registraron una temperatura de
alineamiento de 64 °C indicando que tienen potencial para ser usados en una RT-PCR
Multiplex (Figura 2). Con los restantes tres juegos de primers se continúa estandarizando la
PCR para determinar la temperatura de alineamiento adecuada.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
37
Figura 2. Amplicones obtenidos de acuerdo a los diferentes juegos de primers
AGRADECIMIENTOS
Agradecemos al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la
beca otorgada en la maestría para el primer autor y al Colegio de Postgraduados, Campus
Montecillo, por las facilidades y la asesoría de sus Profesores.
LITERATURA CITADA
Deom, C. M., Oliver, M. J. y Beachy, R. N. (1987). The 30-kilodalton gene product of tobacco mosaic virus potentiates virus movement. Science. 237, 389-394
Lamb, E. M. 2001. Pepper Mild Mottle Virus. Plant pathology Fact Shhet. University of Florida. 55p
Meshi, T., Watanabe, Y., Saito, T., Sugimoto, A., Maeda, T. y Okada, Y. (1987). Function of the 30 kd protein of tobacco mosaic virus: involvement in cell-to-cell movement and dispensability for replication. EMBO J. 6, 2557-2563
Palukaitis, P., García-Arenal, F., Sulzinski, M. A. y Zaitlin, M. (1983). Replication of tobacco mosaic virus. VII. Further characterization of singleand double-stranded virus-related RNAs from TMV-infected plants. Virology. 131, 533-545
Quadt, R. y Jaspars, E. M. J. (1989). RNA polymerases of plus-strand RNA viruses of plants. Mol plant-Microbe Interact. 2, 219-223
Wetter C., Conti M., Altschuh D., Tabillion R. y Regenmortel M. H. V., (1984). Pepper mild mottle virus, un Tobamovirus que infecta variedades de pimiento en Sicilia. Fitopatología. 74(4): 405-410

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
38
EVALUACIÓN DE LA ACTIVIDAD ANTAGÓNICA DE Bacillus
CONTRA Corynespora cassiicola
Honelver Santiago-Santiago1, Javier Hernández-Morales1*, Sergio Aranda-Ocampo1, Ciro Velasco-
Cruz1
1Colegio de Postgraduados, Fitosanidad-Fitopatología, Carretera México-Texcoco km 36.5 Montecillo, Texcoco, CP. 65230, Estado de
México, México. *Autor de correspondencia: Javier Hernández Morales ([email protected]).
INTRODUCCIÓN
En México se cultivan más de 20,000 ha de jamaica (Hibiscus sabdariffa L.); el estado de
Guerrero, ocupa el primer lugar como productor, con más del 70 % de la superficie (SIAP,
2017). Los cálices de jamaica se utilizan en la elaboración de vinos, dulces, infusiones y
refrescos, así como en la medicina tradicional, por sus propiedades antioxidantes y
antimicrobianas (Patel, 2014).
En Guerrero, desde hace varios años, se presentó la enfermedad denominada “manchado
de hojas y cálices de la jamaica”, inducida por Corynespora cassiicola (Berk. & M.A. Curtis)
C.T. Wei. En casos severos, la incidencia llega a ser del 100 %, con pérdidas totales en las
plantaciones (Ortega-Acosta et al., 2015). El manejo a esta enfermedad ha sido con
fungicidas convencionales (Ortega-Acosta et al., 2019); sin embargo, conociendo los efectos
adversos de los agroquímicos, se inclina cada vez más a un control biológico.
El género Bacillus, dadas sus potencialidades en inhibición de fitopatógenos y promoción
de crecimiento de las plantas (Izzeddin y Medina, 2011), puede ser una alternativa para el
control de C. cassiicola. Bacillus, considerada una bacteria promotora del crecimiento
vegetal, aporta beneficios a las plantas, por diversos mecanismos, que incluyen la producción
de fitohormonas, la solubilización de fosfatos, la fijación de nitrógeno y el control biológico
de patógenos (Lugtenberg and Kamilova, 2009). Por lo anterior el presente estudio tuvo como
objetivo obtener cepas de Bacillus antagónicas para el manejo de Corynespora cassiicola en
el cultivo de la jamaica, además de evaluar su efecto promotor de crecimiento vegetal.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
39
MATERIALES Y MÉTODOS
Material biológico. C. cassiicola, se aisló e identificó como agente causal del “manchado
de hojas y cálices de la jamaica” (Ortega-Acosta et al., 2016), las muestras se donaron de la
colección del Laboratorio de Patología de Cultivos Tropicales del Colegio de Postgraduados.
Las cepas de Bacillus, previamente caracterizadas como antagonistas a C. cassiicola
(Rendón-Espíritu, 2014), se obtuvieron de la colección del Laboratorio de Bacteriología del
Colegio de Postgraduados. Se utilizó la variedad criolla del cultivar de jamaica y el desarrollo
del experimento se estableció bajo condiciones de invernadero, con un promedio de
temperatura y humedad relativa de 25 °C y 74 %, respectivamente.
Diseño experimental. Se confrontaron tres cepas de Bacillus y un testigo (T1, B. subtilis;
T2, B. amyloliquefaciens; T3, producto comercial; T4, testigo negativo) con el patógeno C.
cassiicola. El diseño experimental fue completamente aleatorio (DECA) con cinco
repeticiones. Las unidades experimentales consistieron en macetas con plantas del mismo
cultivar de jamaica (variedad criolla).
Inoculación de las cepas bacterianas. Se utilizó el método Biopriming, el cual consistió
en el recubrimiento de las semillas con el agente bacteriano de biocontrol (Bacillus), junto a
un proceso de hidratación, antes de la siembra. El tratamiento a la semilla funge como un
medio importante al manejo de enfermedades en suelo y trasmitidas por semillas (Rao et al.,
2009).
Inoculación del hongo fitopatógeno. Se asperjó 5 mL de suspensión, a una concentración
de 2 × 105 esporas/mL (Kwon y Park, 2003), con un atomizador manual, de forma
independiente en las plantas. Los testigos negativos se asperjaron con agua destilada estéril.
El experimento se mantuvo bajo condiciones de invernadero.
Se evaluaron las variables porcentaje de germinación, longitud de raíz, materia seca en
plantas y raíz, incidencia y severidad. Esta última se midió en intervalos de tres días, con
ayuda de las escalas logarítmicas diagramáticas propuestas por Ortega-Acosta et al., (2016).
Las mediciones permitieron la estimación del área bajo la curva del progreso de la
enfermedad (ABCPE), variable que servirá para determinar significancia en los tratamientos.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
40
RESULTADOS Y DISCUSIÓN
Germinación. Las primeras semillas en emerger fueron las tratadas con B. subtilis (T1),
B. amyloliquefaciens (T2) y el producto comercial (T3), y al cuarto día alcanzaron el 80 %.
El testigo negativo (T4), al día seis, no había iniciado la germinación, resaltando que el
inóculo bacteriano aceleró el desarrollo de las semillas, obteniendo entonces una ventaja en
el inicio de la germinación en comparación con las semillas tratadas sin la bacteria. El T1 fue
el mejor con 95.63 % de semillas germinadas, seguido del T2 con 94.38 %, T3 con 93.75 %
y T4 con 80 %., Lo anterior coincide con lo reportado en diversos estudios, donde los
resultados demuestran un efecto positivo de germinación en las semillas tratadas con
bacterias del género Bacillus (Mojica-Marin et al., 2009).
Promotoras de crecimiento. Se demostró que el T1, T2 y T3, son responsables del
aumento en la longitud de raíz; de igual manera, la materia seca en plantas y raíz mostraron
una diferencia significativa entre los tres tratamientos y el T4. Estudios anteriores han
propuesto al género Bacillus como bacterias benéficas para mejorar la capacidad de
promoción de crecimiento de las plantas (Rojas-Solís et al., 2016).
Antagonismo. Se ha reportado que, en condiciones de invernadero, las plantas de jamaica
inoculadas con C. cassiicola manifiestan síntomas a los 12 días después de la inoculación
(Ortega-Acosta et al., 2016). En la presente investigación, a los siete días postinoculación, el
T4 manifestó síntomas con 100 % de incidencia; mientras que después de los siete días, el
T1 registró 60 %, seguido del T2 y T3 con 80 %.
La variable severidad sirvió para estimar el ABCPE, dónde se apreció una diferencia
significativa en al menos uno de los tratamientos, esto corresponde a lo escrito por Tejera et
al. (2012), quienes mencionan al género Bacillus como un gran antifúngico sobre hongos
fitopatógenos, debido a su velocidad de crecimiento, lo que le permite alcanzar con facilidad
su estado estacionario y comenzar la producción de metabolitos secundarios con actividad
antifúngica, y finalmente comenzar una colonización en la planta.
Trabajos recientes sobre el efecto antagónico de microorganismos y el desarrollo del
Biopriming, como mecanismos en la utilización de bacterias promotoras del crecimiento

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
41
vegetal, han descrito a cepas de Bacillus con la capacidad de producir auxinas estimuladoras
del crecimiento vegetal y solubilizar fosfatos (Tejera et al, 2012); lo cual, conjuntamente con
los resultados de este trabajo, se puede sustentar una interacción planta-bacteria que permite
mejorar los estados de desarrollo de la planta, que al final impactará en mayores
rendimientos.
AGRADECIMIENTOS
Agradecemos al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la
beca otorgada en la maestría para el primer autor. Al Colegio de Postgraduados, Campus
Montecillo, por el equipo y las instalaciones donde se realizaron los experimentos y la
asesoría de sus profesores.
LITERATURA CITADA
Izzeddin, A. N. and Medina, T. L. (2011). Biological control effect by antagonist on plant pathogens in vegetables for human consumption. Salus. 15(3): 8-12.
Kwon, J. H. Park, C. S. (2003). Leaf spot of cotton rose caused by corynespora cassiicola in Korea. Mycobiology. 31(1):57-59.
Lugtenberg, B. and Kamilova, F. (2009). Plant-Growth-Promoting Rhizobacteria. Annu Rev Microbiol. 63:541-556.
Mojica-Marín, V. Luna-Olvera, H. A. Sandoval-Coronado, C. F. Pereyra-Alférez, B. Morales-Ramos, L. H. González-Aguilar, N. A. Hernández-Luna, C. E. Alvarado-Gómez, O. G. (2009). Control biológico de la marchitez del chile (Capsicum annunm L.) por Bacillus thuringensis. Revista Phyton. 78:105-110.
Ortega-Acosta, S. A. Hernández-Morales, J. Ochoa-Martínez, D. L. Ayala-Escobar, V. (2015). First report of Corynespora cassiicola causing leaf and calyx spot on roselle in Mexico. Plant Disease, Saint Paul. 99(7): 1041.
Ortega-Acosta, S. A. Ochoa-Martínez, D. L. Leyva-Mir, S.G. Velasco-Cruz, C. Mora-Aguilera, J. A. Hernández-Morales, J. (2019). Control químico del manchado de hojas y cálices de jamaica en Guerrero, México. Summa Phytopathologica, 45: 38-43.
Ortega-Acosta, S.A. Velasco-Cruz, C. Hernández-Morales, J. Ochoa-Martínez, D.L. and Hernández-Ruiz, J. (2016). Diagrammatic logarithmic scales for assess the severity of spotted leaves and calyces of roselle. Revista Mexicana de Fitopatología. 34: 270-285.
Patel, S. (2014). Hibiscus sabdariffa: An ideal yet under-exploited candidate for nutraceutical applications. Biomedicine & Preventive Nutrition. 4: 23-27.
Rao, M. S. L. Kulkarni, S. Lingaraju, S. and Nadaf. H. L. (2009). Bio-priming of seeds: a potential tool in the integrated managment of Alternaria blight of sunflower. HELIA. 32: 107-114.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
42
Rendón-Espíritu, B. (2014). Calidad sanitaria y microorganismos antagónicos a Corynespora cassiicola (Berk. & Curt.) Wei en cálices frescos de jamaica (Hibiscus sabdariffa L.). Tesis de maestría. Colegio de Postgraduados. Edo. de México.
Rojas-Solís, D. Hernández-Pacheco, C. E., & Santoyo, G. (2016). Evaluation of Bacillus and Pseudomonas to colonize the rhizosphere and their effect on growth promotion in tomato (Physalis ixocarpa Brot. ex Horm.). Revista Chapingo Serie Horticultura. 22(1): 45-57.
SIAP. Servicio de Información Agroalimentaria y Pesquera. México. 2017. Disponible en: http://www.siap.gob.mx/index.php?option=com_wrapper&view=wrapper&Itemid=351.
Tejera, B. Heydrich, M. & Rojas, M. M. (2012). Antagonismo de Bacillus spp. frente a hongos fitopatógenos del cultivo del arroz (Oryza sativa L.). Revista de Protección Vegetal. 27(2): 117-122.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
43
CARACTERIZACIÓN POLIFÁSICA DE Pectobacterium Y Dickeya
spp. INDUCTORAS DE PUDRICIONES BLANDAS EN MÉXICO
Moisés García-García1, Sergio Aranda-Ocampo1*, Reyna I. Rojas-Martínez1, Daniel L. Ochoa-
Martínez1, J. Alfredo Carrillo-Salazar2.
1Postgrado en Fitosanidad-Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco Km. 36.5, Montecillo, 56230 Texcoco,
Estado de México. 2Postgrado en Recursos Genéticos y Productividad, Colegio de Postgraduados, Carretera México-Texcoco Km. 36.5,
Montecillo, 56230 Texcoco, Estado de México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
Los géneros Pectobacterium y Dickeya pertenecen a las enterobacterias asociadas a
pudriciones blandas; son fitopatógenos que causan pérdidas económicamente importantes en
una amplia gama de cultivos herbáceos, ornamentales y hortícolas por lo que son
considerados como uno de los diez agentes patógenos de plantas más importantes (Buttimer
et al., 2018).
Anteriormente se consideraba a Pectobacterium y Dickeya como patógenos de gran
importancia en el cultivo de papa ya que inducían dos enfermedades distintas: la pudrición
blanda en tubérculos y pierna negra en tallo (Toth and Birch 2005); sin embargo, en la
actualidad diversas poblaciones genéticas de estos dos géneros causan pudriciones blandas
en un amplio rango de hospedantes y cada vez se incluyen nuevas familias y especies de
plantas no reportadas con anterioridad (Baghaee-Ravari et al., 2010).
En México no existen investigaciones que hayan abordado el estudio y caracterización de
especies de Pectobacterium y Dickeya que ayuden a diferenciar entre especies aisladas en
México; Asimismo, se desconoce si estos géneros bacterianos han ampliado su rango de
hospedantes por la activación diferencial de nuevos factores virulencia, o se trate de especies
aun no descritas en el país; por lo anterior, el objetivo de esta investigación fue caracterizar
el perfil bioquímico, fisiológico, patogénico y molecular de especies de Pectobacterium y
Dickeya aislados de diferentes localidades y hospedantes en México.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
44
MATERIALES Y MÉTODOS
Se aislaron ocho cepas bacterianas a partir de cultivos de coliflor (Brassica oleracea var.
botrytis), papa (Solanum tuberosum L.), ajo (Allium sativum), tetetzo (Neobuxbaumia
tetetzo), nopal (Opuntia ficus-indica) y sábila (Aloe vera) con síntomas de pudrición blanda
en los estados de Oaxaca, Puebla y Estado de México. Los aislados fueron identificados
inicialmente como P. atrosepticum (Cp1), P. wasabiae (Cp2), P. atrosepticum (Cp3), P.
brasiliensis (Cp4), P. carotovorum (Cp5) , Dickeya chrysanthemi (Cp7) y P. cacticida (Cp11
y Cp12) respectivamente.
Para todos los aislamientos bacterianos se realizó una prueba de pudrición en tubérculos
de papa (Solanum tuberosum L.) realizando modificaciones a lo descrito por Maisuria y
Nerurkar (2013), que consistió en colocar dos discos de papa en una caja Petri con papel
absorbente previamente humedecido con agua destilada estéril. Uno de los dos discos de papa
fue inoculado con cada cepa bacteriana mientras que el otro fue inoculado con H2O estéril
como testigo. También se incluyó una reacción de hipersensibilidad en hojas de tabaco
(Nicotiana tabacum) infiltrando 1 mL de una suspensión bacteriana a una concentración de
108 UFC/mL con una aguja hipodérmica según lo establecido por Lozano y Sequeira (1970);
Moleleki et al. 2013; Soto et al., 2011).
Para medir la virulencia (actividad pectinolítica) de las cepas en estudio se obtuvo la
cantidad relativa de la actividad enzimática, para lo cual se utilizó la propuesta de Kunstmann
et al. (2006) y Franco et al. (2007) con modificaciones. La metodología empleada consistió
en preparar una suspensión bacteriana de cada cepa a una concentración de 108 UFC/mL.
Posteriormente se desinfectaron, cortaron y pesaron tubérculos de papa (cuatro mitades por
cepa más un testigo). Cada mitad de tubérculo fue inoculada con 100 μL de suspensión
bacteriana y colocada en bolsas herméticas para generar un ambiente de alta humedad e
incubadas por 24 y 48 horas a 28º C. Al cabo de los periodos de tiempo se retiró el tejido
macerado de los tubérculos. Los datos obtenidos de la diferencia de peso antes y después de
la inoculación se analizaron con el modelo Yij = µ + τi + εij para obtener una medida
directamente proporcional de la actividad pectinolítica de las enzimas que pudieran estar
siendo secretadas por cada especie bacteriana (Kunstmann et al., 2006; Franco et al., 2007).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
45
Como parte de las pruebas fisiológicas se realizó una prueba de tolerancia salina saturando
agar nutritivo con NaCl a concentraciones del 1 al 10%. Las ocho cepas fueron sembradas
en las 10 placas e incubadas por 24 horas a 28º C para observar la presencia o ausencia de
crecimiento bacteriano sobre el medio. Además, se contrastó con la propuesta de Schaad et
al. (2001) con modificaciones, que consistió en incubar un tubo con 5 mL de caldo nutritivo
a 37º C por 24 horas, posteriormente se inocularon con 5 μL de una suspensión bacteriana a
una concentración de 108 UFC/mL y se incubó otras 24 horas a 37 ºC.
Las cepas se caracterizaron bioquímicamente por tinción de Gram, metabolismo
oxidativo/fermentativo (Hugh y Leifson 1953), la actividad de la oxidasa y la catalasa
(Baghaee-Ravari et al., 2011; Ma et al. 2007 y Pitman et al. 2010), producción de fosfatasa
(Schaad et al. 2001) y prueba de Ryu (Cerny, 1976), así como la asimilación de diferentes
fuentes de carbono en medio basal como lo describe Ayers et al. (1919). Los hidratos de
carbono usados fueron: celobiosa, α-metil-glucósido, lactosa, palatinosa, trehalosa, sucrosa,
maltosa y xilosa (Xiu-Fang et. al., 2008). También se incluyeron pruebas API®20E
(BioMérieux) y un perfil de antibióticos que incluyeron: ampicilina, oxitetraciclina,
levofloxacina y eritromicina.
La fase molecular consistió en la identificación genética, para lo cual se usó la
metodología Wizard® Genomic DNA Purification Kit para la extracción de DNA total para
después amplificar el gen 16S por PCR usando la metodología GoTaq® Green Master Mix
con los primers universales 8F y 1492R para su secuenciación. La última parte de esta fase
consistirá en buscar secuencias de nucleótidos de enzimas pectinolíticas reportadas en la base
de datos del National Center for Biotechnology Information (NCBI) para saber si la
diferencia en el grado de virulencia entre las cepas se debe por la secreción de enzimas
pectinolíticas específicas.
RESULTADOS Y DISCUSIÓN
Los resultados de la prueba de pudrición de tubérculo fueron positivas para todas las
cepas, lo que indica que todas secretan enzimas pectinolíticas y tienen la facultad de macerar
tejido vegetal (Corzo-López y Quiñones-Pantoja 2017; Ngadze et al., 2010). En la prueba de

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
46
hipersensibilidad, siete de las ocho cepas no indujeron esta reacción, lo cual concuerda para
el perfil de estas bacterias, sin embargo, la cepa Cp4 dio una respuesta positiva a tal reacción
(Moleleki at al., 2013). Los resultados de la prueba de virulencia se muestran en el cuadro 1.
La prueba de comparación de medias por el método de Tukey indica que a las 24 horas la
Cp7 (D. chrysanthemi) macera una mayor cantidad de tejido seguido de la Cp11 (P.
cacticida). La tendencia a las 48 horas no cambia mucho con respecto a las 24 horas; esto
indica que las bacterias del género Dickeya son más virulentas en comparación del género
Pectobacterium, aunque entre espacies también hay diferencias no significativas, lo que
podría indicar que la virulencia entre especies de este género puede deberse a otros factores
(Kunstman et al., 2006; Franco et al., 2007).
Cuadro 1. Prueba de comparación de medias por el método de Tukey donde muestra la
generación de grupos con base en la cantidad de tejido macerado en tubérculos de papa.
24 horas 48 horas
Cepa Respuesta Grupos Cepa Respuesta Grupos
Cp7
Cp11
Cp4
Cp5
Cp2
Cp1
Cp12
Cp3
15.900
10.375
9.700
8.600
8.075
7.525
7.150
6.125
a
ab
b
b
b
b
b
b
Cp7
Cp11
Cp12
Cp4
Cp5
Cp1
Cp2
Cp3
47.750
37.525
27.975
27.100
23.725
21.525
21.175
17.725
a
ab
bc
bc
c
c
c
c
En la prueba de tolerancia salina, ninguna cepa fue capaz de crecer a concentraciones
mayores a 5% de NaCl; las cepas Cp2, Cp3, Cp11 y Cp12 solo toleraron una concentración
máxima de 4% y Cp7 hasta 2%. Los resultados de las pruebas API®20E no muestran ninguna

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
47
variación bioquímica entre especies por lo que estos marcadores no ayudan a generar
información que pueda diferenciarlos por su actividad bioquímica. Por otro lado, las pruebas
de antibiosis tampoco muestran marcadores distintivos entre especies con respecto a la
oxitetraciclina y levofloxacina ya que todas las cepas mostraron sensibilidad. Para el caso de
la eritromicina, solo Cp7 mostró sensibilidad y todas las demás fueron resistentes. Para la
ampicilina, Cp1, Cp3 y Cp5 mostraron resistencia, mientras que las otras cepas fueron
sensibles. Los resultados de las pruebas bioquímicas convencionales tampoco muestran
resultados consistentes con lo reportado en la literatura con respecto a estos patógenos, por
lo que esto refuerza lo descrito con anterioridad en el sentido que los marcadores bioquímicos
ya no son la mejor opción para una diferenciación entre especies del mismo género, por lo
que se espera que la fase molecular se obtengan mejores resultados y confiables.
AGRADECIMIENTOS
Agradecemos al Concejo Nacional de Ciencia y Tecnología (CONACYT, México) por la
beca otorgada en la maestría para el primer autor. Al Colegio de Postgraduados, Campus
Montecillo, por los equipos e instalaciones donde se realizaron los experimentos y la asesoría
de sus profesores.
LITERATURA CITADA
Baghaee-Ravari, S., Rahimian, H., Shams-Bakhsh, M., López-Solanilla, E., Lamas-Antúnez, M., and Rodríguez-Palenzuela, P. (2010). Characterization of Pectobacterium species from Iran using biochemical and molecular methods. European Journal of Plant Pathology. 129:413-425
Buttimer, C., Hendrix, H., Lucid, A., Neve, H., Noben, J-P., Franz, C., O´Mahony., Lavigne, R., Coffey, A. (2018). Novel N4-like Bacteriophages of Pecrobacterium atrosepticum. Pharmaceuticals 11:45.
Toth, I. k., and Birch, P. R. (2005). Rotting softly stealthily. Current Opinion in Plan Biology. 8: 424 - 429.
Ma, B., Hibbing, M. E., Kim, H. S., Reedy, R. M., Yedidia, I., Breuer, J., Glasner, J. D., Perna, N. T., Kelman, A., Charkowski, A. O. (2007). Host range and molecular phylogenies of the rot enterobacterial genera Pectobacterium and Dyckeya. The American Phytopathological Society. 97:1150-1163.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
48
IDENTIFICACIÓN Y REGIONALIDAD DE ESPECIES DE
Fusarium ASOCIADAS A MARCHITEZ Y PUDRICIÓN SECA DEL
COGOLLO EN Agave tequilana Weber var. azul
Viridiana López-Bautista1, Gustavo Mora-Aguilera1, Ma. Alejandra Gutiérrez-Espinosa2, Lourdes
Obdulia Segura-León3, Verónica I. Martínez-Bustamante4, Juan J. Coria-Contreras4, Coral Mendoza-
Ramos2, Baldemar Santana-Peñaloza4 1Posgrado en Fitosanidad-Fitopatología. 2Recursos Genéticos y Productividad-Fruticultura. 3Fitosanidad-Entomología y Acarología.
Campus Montecillo, Colegio de Postgraduados, Texcoco, México, C.P. 56230. 4Laboratorio de Análisis de Riesgo Epidemiológico
Fitosanitario (LANREF-CP). *Autor de correspondencia: [email protected]
INTRODUCCIÓN
La marchitez y pudrición seca del cogollo del agave, es el principal factor fitosanitario
restrictivo de producción de Agave tequilana en los últimos quince años. Se han reportado
organismos asociados a la enfermedad como Fusarium oxysporum y F. solani (Ávila et al.,
2010; Vega et al., 2013; Ramírez et al., 2017). Sin embargo, se carece de un estudio
exhaustivo, con enfoque epidemiológico a nivel regional, que considere la interacción de
diversas condiciones agroclimáticas, productivas y estacionales para dilucidar el organismo
(s) más prevalente y asociado a esta enfermedad.
El objetivo de este trabajo fue determinar la identidad y prevalencia regional de Fusarium
spp. asociado a la marchitez y pudrición seca del cogollo, bajo escenarios epidémicos
contrastantes, inferir potenciales relaciones filogenéticas y diversidad genética, con el fin de
dilucidar posibles mecanismos de evolución patogénica aplicables a estrategias de control
efectivas y sustentables.
MATERIALES Y MÉTODOS
Selección de sitios y colecta de muestras. Se colectaron muestras de suelo-raíz de 86
plantaciones comerciales de 3-4 años de edad, ubicadas en las regiones Altos, Valles, Sur de
Jalisco y Romita, Guanajuato, y seleccionadas por criterios de inductividad epidémica de
REG-N (Acevedo-Sánchez et al., 2017; Coria-Contreras et al., 2019). Los sitios se
clasificaron de acuerdo a su condición fitosanitaria predominante de marchitez (M),

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
49
pudrición seca de cogollo (P), aparentemente sanos (AS), improductivos (IP) y potencial
productivo, nunca cultivado con agave (PT). Se integraron cinco submuestras obtenidas de
foco y perifoco de la enfermedad por plantación. Así mismo, se seleccionaron 37
plantaciones comerciales para colecta de 221 muestras de tejido aéreo (hojas de cogollo, piña
y cuello) en 138 plantas, mediante un muestreo dirigido a síntomas consistentes en pudrición
seca del cogollo. Las colectas de suelo-raíz y tejido, se realizaron en mayo-junio de 2016 y
enero-febrero de 2017, respectivamente, con un total de 307 muestras.
Extracción de ADN, ensayos de PCR y secuenciación. A partir de 458 aislados
monospóricos de Fusarium spp., se realizó una selección de 113 con base en características
culturales in vitro, morfología general, fuente de aislamiento, prevalencia e inductividad
regional de la enfermedad. El ADN genómico se extrajo por el método modificado de AP
(Sambrook y Russell, 2001) y su concentración y calidad se cuantificó con NanoDrop 2000
(Thermo Fisher Scientific, USA). El ADN templado (40 ng/μL) se amplificó por PCR
convencional, usando los cebadores EF1/EF2 para el gen factor de elongación EF-1a
(O'Donnell et al., 1998), 5f2/7cr y 7cf/11ar para la segunda subunidad mayor de la ARN
polimerasa II RPB2 (O'Donnell et al., 2008 ) y NL11/CNS1 para la región espaciadora
intergénica IGS (O’Donnell et al., 2009). Se enviaron a secuenciar 399 reacciones a
Macrogen Inc. (Korea).
Alineamiento de secuencias y análisis bioinformáticos. Las 399 secuencias de ADN se
editaron y ensamblaron utilizando SeqAssem, se compararon con las bases de datos
Fusarium-MLST y GenBank mediante BLASTn y alinearon con secuencias de referencia
utilizando el algoritmo MUSCLE implementado en MEGA7. Las alineaciones consistieron
en 267 sitios parsimoniosos informativos de 794 pb para EF-1a, 493/2027 pb para RPB2 y
841/2826 pb para IGS. Los análisis filogenéticos se ejecutaron con el método Máxima
Verosimilitud (ML) en MEGA7 con 1,000 repeticiones Bootstrap. El alineamiento obtenido
de cada gen se importó a DNA Sequence Polymorphism v6.11.01 (DnaSP6), donde se generó
la lista de haplotipos y se exportó a PopART v1.7. La red de haplotipos se construyó usando
el método Minimum Spanning Network.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
50
RESULTADOS Y DISCUSIÓN
Identificación y prevalencia regional de Fusarium spp. Los 113 aislados se
identificaron como especies correspondientes a cuatro complejos de Fusarium. Un total de
39 aislados (34.51 %) correspondieron a F. fujikuroi (FFSC), 37 (32.7 %) a F. solani (FSSC),
30 (26.54 %) a F. oxysporum (FOSC) y 7 (6.19 %) a F. incarnatum-equiseti (FIESC). Las
especies que se encontraron asociadas a marchitez y pudrición seca del cogollo incluyen a F.
oxysporum (FOSC), F. solani, F. falciforme, F. neocosmosporiellum (FSSC), F.
verticillioides, F. proliferatum, F. lactis, F. andiyazi, F. nygamai, F. subglutinans (FFSC),
F. incarnatum (cepas tipo FIESC28, FIESC38 y FIESC25) y F. equiseti (FIESC35 y
FIESC3), al menos una cepa podría representar una especie filogenética nueva dentro del
FSSC, ya que formó un linaje único en las filogenias.
F. oxysporum (30/113), F. falciforme (26/113) y F. verticillioides (23/113) dominaron la
colección y representaron el 70 % (79/113) de los aislados. Las tres especies se encontraron
distribuidas en Altos, Valles y Sur. Sin embargo, F. oxysporum, fue más prevalente en Altos,
aislado principalmente de raíz y suelo en plantaciones con marchitez (15/30) y pudrición
(11/30). Interesantemente, esta especie no se aisló de los sitios IP y PT pero sí de AS, lo que
presupone dependencia del cultivo. En plantaciones AS, factores de supresividad pueden
estar limitando la expresión de la enfermedad, o la presencia de cepas con baja o nula
capacidad patogénica. La especie F. falciforme, aislada de suelo, raíz y cuello, fue prevalente
en Valles, en parcelas P (8/26), PT (7/26), M (5/26) e IP (3/26), lo que indica asociación con
ambas enfermedades y un indicador de riesgo al encontrarse en sitios potencialmente
productivos y ser la única especie aislada de sitios improductivos. F. verticillioides fue la
especie prevalente en Valles, aislada de tejido aéreo y con mayor frecuencia de la parte
superior del cogollo (18/23). Estos resultados evidencian por primera vez la asociación entre
especie y nicho-enfermedad, e indican la especialización patogénica de Fusarium.
Resultados similares se han reportado con otras especies de Fusarium asociadas a la
malformación del mango (Liew et al., 2016).
Inferencia filogenética. La filogenia entre los complejos de Fusarium, con ambos genes,
fue topológicamente concordante, permitiendo identificar cuatro complejos de especies de

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
51
Fusarium. Todos formaron grupos monofiléticos consistentes (98-100 %). Se ha reportado
que FOSC y FFSC se encuentran estrechamente relacionados, debido a que el genoma de F.
oxysporum se asemeja a especies heterotálicas, a pesar de que no se encontró un estado sexual
(O’Donnell et al., 2004; Fourie et al., 2009)
Estructura poblacional. La diversidad genética entre los 113 aislados, determinada con
los genes EF-1a y RPB2, indicaron que la región analizada de EF-1a presentó menor cantidad
de sitios polimórficos y número de mutaciones (212 y 297) en contraste con RPB2 (505 y
653); sin embargo, el índice de mutación fue mayor en el primero, considerando el número
de nucleótidos analizados (Cuadro 1). Los haplotipos obtenidos con cada gen fueron de 59 y
52 para EF-1a y RPB2, respectivamente.
Cuadro 1. Polimorfismo del ADN para 113 aislados de Fusarium spp., con base en EF-
1a y comparado con RPB2.
Polimorfismo del ADN EF-1a RPB2 Número de secuencias 113 113 Región analizada 565 1695 Número de sitios polimórficos, S 212 505 Número de mutaciones, Eta 297 653 Número de haplotipos, h 59 52 Diversidad de haplotipos, Hd 0.972 0.957 Diversidad de nucleótidos, p 0.101 0.100 Índice de mutación 0.52 0.38 Índice de sitios conservados 0.62 0.70 Índice de variación 0.52 0.46 q (S) 0.089 0.067 q (P) 0.116 0.115 Número de nucleótidos
diferentes, k 56.789 169.428
El patrón de las redes de haplotipos mostró 39/59 únicos con EF-1a. En el centro de la red
se encontró a FFSC, a partir del cual divergen los demás. Con RPB2 se obtuvieron 35/52
únicos, pero no se identificó un haplotipo o complejo central discernible de divergencia. En
ambas redes, el mayor número de sitios de cambio se dio entre miembros de FSSC y FIESC
con 81/71 y 344/54 sitios de cambio en EF-1a y RPB2, respectivamente. La distribución

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
52
geográfica demostró correlación entre diversidad de haplotipos e inductividad de
enfermedad. La región Altos, considerada la zona más inductiva, registró el mayor número
de haplotipos, seguido de Valles, Sur y Romita. Lo anterior sugiere que la región Altos puede
considerarse el centro de diversificación y dispersión de Fusarium o donde operan factores
de variación o mutagénicos de mayor intensidad. Por ejemplo, esta región tiene en promedio
un pH ácido más bajo (5.6), en comparación con Valles (6.5) y Sur (7.1), considerado
favorable a Fusarium y donde se cultiva el 51 % del agave de Jalisco.
Este estudio mostró por primera vez que existe una amplia diversidad de especies en
complejos de Fusarium, asociadas a la marchitez y pudrición seca del agave. En el presente
trabajo se reportan 10 especies clasificadas taxonómicamente con binomio latino y nueve
especies filogenéticas. En total, 19 especies asociadas a la enfermedad, 16 de estas se reportan
por primera vez y dos pendientes de clasificar.
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca otorgada para la
realización de mis estudios de maestría. A SENASICA-DGSV y CRT, por el apoyo logístico
y financiero para la ejecución del proyecto SIVEA. Así como a los miembros del Laboratorio
de Análisis de Riesgo Epidemiológico Fitosanitario (LANREF-CP).
LITERATURA CITADA
Acevedo-Sánchez, G., B. Santana-Peñaloza, V. López-Bautista, V. Martínez-Bustamante, J. Coria-Contreras, C. Mendoza-Ramos, L. Jiménez-González y G. Mora-Aguilera. 2017. Criterios epidemiológicos para implementar una red de monitoreo de enfermedades causadas por Fusarium spp. en regiones agaveras de Jalisco. Rev. Mex. Fitopatol. 35(S): 176-177.
Ávila-Miranda, M.E., J.G. Zazueta-López, C. Arias-Castro, J.J. Peña-Cabriales. 2010. Vascular wilt caused by Fusarium oxysporum in agave (Agave tequilana Weber var. azul). J. Prof. Assoc. Cactus 12: 166-180.
Coria-Contreras, J.J., G. Mora-Aguilera, M.J. Yañez-Morales, G. Acevedo-Sánchez, B. Santana-Peñaloza, C. Mendoza-Ramos, L. Jiménez-González, V.I. Martínez-Bustamante, D.C. García-Martínez and R. Rubio-Cortés. 2018. Applied regional epidemiology to inductive characterization and forecasting of blue agave gray spot (Cercospora agavicola) in Jalisco, Mexico. Rev. Mex. Fitopatol. 37(1): 71-94.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
53
Fourie, G., E.T. Steenkamp, T.R. Gordon, A. Viljoen. 2009. Evolutionary relationships among the Fusarium oxysporum f. sp. cubense vegetative compatibility groups. Appl. Environ. Microbiol. 75(14):4770-781.
Liew, E.C.Y., M.H. Laurence, C.A. Pearce, R.G. Shivas, G.I. Johnson, Y.P. Tan, J. Edwards, S. Perry, A.W. Cooke, B.A. Summerell. 2016. Review of Fusarium species isolated in association with mango malformation in Australia. Australas. Plant Pathol. 45:547-559.
O’ Donnell, K., E. Cigelnik, H.I. Nirenberg. 1998. Molecular systematics and phylogeography of the Gibberella fujikuroi species complex. Mycologia. 90:465-493.
O’ Donnell, K., D.A. Sutton, A. Fothergill, D. McCarthy, M.G. Rinaldi, M.E. Brandt, N. Zhang, D.M. Geiser. 2008. Molecular Phylogenetic Diversity, Multilocus Haplotype Nomenclature, and In Vitro Antifungal Resistance within the Fusarium solani Species Complex. 46(8): 2477-2490.
O'Donnell, K., D.A. Sutton, M.G. Rinaldi, K.C. Magnon, P.A. Cox, S.G. Revankar, S. Sanche, D.M. Geiser, J.H. Juba, J-A.H. van Burik, A. Padhye, E.J. Anaissie, A. Francesconi, T.J. Walsh, J.S. Robinson. 2004. Genetic diversity of human pathogenic members of the Fusarium oxysporum complex inferred from multilocus DNA sequence data and amplified fragment length polymorphism analyses. J. Clin. Microbiol. 42(11):5109-120.
Ramírez-Ramírez, M. J., N.A. Mancilla-Margalli, L. Meza-Álvarez, R. Turincio-Tadeo, D.G.D. Pena and M.E. Ávila-Miranda. 2017. Epidemiology of Fusarium Agave Wilt in Agave tequilana Weber var. azul. Plant Prote. Sci. 53(3).
Sambrook, J. And R.W. Russell 2001. Molecular cloning: A laboratory manual, 3rd edition. 1:1.32-1.34 Cold spring harbor laboratory press. Cold spring harbor, N.Y.
Vega-Ramos, K.L., J.X. Uvalle-Bueno, and J.F. Gómez-Leyva. 2013. Molecular variability among isolates of Fusarium oxysporum associated with root rot disease of Agave tequilana. Biochem. Genet. 51(3-4): 243-255.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
54
ESTIMACIÓN DE CARGA DE INÓCULO IN SITU DE Fusarium
spp. EN HIJUELOS DE Agave tequilana A DIFERENTES NIVELES DE
INDUCTIVIDAD EPIDÉMICA REGIONAL
Coral Mendoza-Ramos1, Juan José Coria-Contreras2, Verónica Martínez-Bustamante2, Eduardo
Guzmán-Hernández2, Baldemar Santana-Peñaloza2, Gerardo Acevedo-Sánchez2, Gustavo Mora-
Aguilera1
1 Posgrado en Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco, Estado de
México, México. 2 CP-LANREF, Programa de posgrado en Fitosanidad, Colegio de Postgraduados, Carretera México-Texcoco km 36.5,
Montecillo, 56230 Texcoco, Estado de México, México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
La producción de tequila está regulada por la NOM-006-SCFI-2012 “Denominación de
Origen del Tequila (DOT)”, la cual delimita las características y especificaciones para su
elaboración. La DOT contempla el uso particular de Agave tequilana Weber variedad azul,
así como su plantación en estados y municipios específicos de Jalisco (125), Michoacán (30),
Tamaulipas (11), Nayarit (8) y Guanajuato (7). Jalisco es el principal productor con 69 mil
hectáreas plantadas (SIAP, 2018), con un promedio anual de plantación de 120 millones de
hijuelos.
Fusarium spp. es uno de los patógenos más importantes reportados para Agave tequilana,
asociado a enfermedades de marchitez, pudrición seca del cogollo y muerte de plantas
(Castañeda, 2002; Rubio, 2007; García, et. al., 2017). Considerando los diferentes niveles de
prevalencia de Fusarium spp. en Jalisco y el movimiento no regulado de hijuelos para el
establecimiento de nuevas plantaciones, existe un alto riesgo de dispersión del patógeno a
áreas de baja o nula incidencia, comprometiendo la productividad del agave. Éste trabajo se
planteó con el objetivo de determinar la asociación de Fusarium spp. en hijuelos de Agave
tequilana y estimar la carga de inóculo, con el fin de establecer estrategias preventivas y
desarrollar algoritmos de riesgo aplicables al Sistema Integral de Protección y Vigilancia
Epidemiológica del Agave (SIVEA) como estrategia regional de manejo fitosanitario del
agave.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
55
MATERIALES Y MÉTODOS
Los experimentos de campo se realizaron de marzo a mayo de 2018 y 2019,
correspondiendo con el arranque de hijuelos para el establecimiento comercial de nuevas
plantaciones de agave azul. En 2018, la asociación y carga de inóculo externo (CIE) a nivel
de cuello y base de hojas en hijuelos se analizó en nueve plantaciones comerciales, tres por
cada región agavera de Jalisco: Altos (A), Valles (V) y Sur (S). La selección de plantaciones
tuvo criterios de prevalencia regional de marchitez-pudrición del cogollo (alta, moderada y
baja), producción de hijuelos con calibres requeridos (tamaño “lima”, “naranja” y “toronja”)
y sincronización con arranque de hijuelos por vía del productor. En cada plantación se
seleccionaron un total de 405 hijuelos, distribuidos en tres bloques de arranque, de cada uno
se seleccionaron nueve submuestras, tres por calibre, de 15 hijuelos cada una. En total se
obtuvieron 243 muestras compuestas para CIE, 27 por plantación. Adicionalmente, en cada
plantación se evaluaron 63 plantas, 21 por bloque, utilizando las escalas de severidad de
marchitez y pudrición seca del cogollo en planta de SIVEA (Jimenez, et. al., 2017). Los
hijuelos de cada muestra se lavaron in situ en 1 L de agua desmineralizada estéril. El cuello
y la base de las hojas fue la zona de lavado y el agua-suelo recuperado se guardó en tubos
Falcon de 50ml.
Para conocer la internalización de Fusarium spp. en tejido (IFT) se analizaron 30 hijuelos
provenientes de una plantación de alta inductividad, 9 de calibre lima, 12 naranja y 9 toronja
seleccionados proporcionalmente de tres bloques de la plantación. En laboratorio, por
hijuelo, se analizaron 6 tipos de tejido: cuello, piña, base de las hojas, base del cogollo y
cogollo alto.
En 2019 se repitieron los experimentos CIE y IFT. En CIE se seleccionaron 12
plantaciones distribuidas en la región de A (6) y S (6), con los mismos criterios que 2018. Se
seleccionaron 270 hijuelos totales por cada plantación, distribuidos en tres bloques de
arranque, de cada uno se seleccionaron seis submuestras, dos por calibre, de 15 hijuelos. Los
hijuelos se procesaron como se describió previamente, obteniendo 216 muestras totales para
CIE. La evaluación de enfermedad se realizó como previamente se indicó, en 200 plantas de
cinco surcos distribuidos en una hectárea. Adicionalmente, por cada plantación se extrajo

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
56
una muestra compuesta de 300 g de tierra, conformada por 6 submuestras distribuidas
equitativamente en los bloques de selección de hijuelos.
El experimento IFT se realizó en siete de las 12 plantaciones empleadas para CIE, seis en
la región Altos, por ser la más inductiva, y uno de la región Sur, de baja inductividad. Se
seleccionó un total de 140 hijuelos, 20 de tamaño naranja por cada plantación. La
internalización, identificación de Fusarium spp. y la carga de inóculo interna (CII) total de
hijuelo, se analizó en cuello, piña, base de las hojas y base del cogollo. En CII se pesó el
hijuelo para conocer la proporción de agua a utilizar (2:1, agua:tejido) y se maceró por 30
segundos a 3 HP (licuadora Blendtec classic 575).
En laboratorio, las siembras microbiológicas se realizaron in vitro en medio modificado
de Komada. En CIE se utilizó la dilución 1x10-1/2. En IFT se empleó tejido desinfestado con
cloro (2% por 2min) y alcohol (70% por 2min). En CII se sembraron 100 μl del sobrenadante
de maceración. En todos los cultivos se contabilizó el número de hongos totales y colonias
de Fusarium spp. Se realizó reaislamiento, purificación, monospórico, conservación en arena
y PDA para su procesamiento taxonómico convencional y molecular.
RESULTADOS Y DISCUSIÓN
En 2018, los datos del experimento CIE reportaron al tamaño lima con el índice más alto
en la relación Fusarium spp./hongos totales, seguido de naranja y toronja (Cuadro 1). Los
datos del 2019 presentan la misma tendencia, pero con índices más altos. Esto podría deberse
a mecanismos de resistencia que transfiere la planta madre. Los índices F/Ht en 2018
presentan una relación directa con la inductividad, mientras que en 2019 los mayores índices
se encuentran en las plantaciones de moderada inductividad. Por región, S tiene los mayores
índices en los dos años, pese a ser considerada como una región de aparente baja
inductividad.
En IFT del 2018 se aislaron 16 cepas de Fusarium spp., 9 (56%) en la base de las hojas, 3
de cuello, 2 de piña, 1 en base del cogollo y cogollo alto. Se secuenciaron por factor de
elongación y se encontraron las especies de F. oxysporum, F. solani y F. verticillioides. En
los 140 hijuelos del 209 se encontraron 276 colonias de Fusarium spp. con proporciones

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
57
similares al año anterior, donde la base de hojas representa el 76% del total (210). Existió
una relación similar a nivel de inductividad que el experimento CIE, las plantaciones
moderadas presentaron los mayores índices de Fusarium spp. en tejidos.
En CII se contabilizaron 1 128 colonias de Fusarium spp en los 140 hijuelos. En la región
A las dos plantaciones de moderada inductividad representan el 36.5% del total, seguido alta
(32.4%) y baja prevalencia (14.3%). La única plantación de la región S equivale al 16.6%.
En promedio un hijuelo de la región A, en zona de alta inductividad tiene 9 colonias de
Fusarium spp. (min= 0, max=51) colonias, hijuelos en condición moderada 10 (min= 0,
max=42) y de baja 5 (min= 0, max=12) colonia, mientras que los hijuelos de la región S, de
aparente baja inductividad, tiene 9 (min= 0, max=37) colonias promedio. En la región Sur
las condiciones climáticas, edáficas o de competencia microbiana pueden estar suprimiendo
la patogenicidad de las cepas de Fusarium spp. debido a la elevada población de hongos
totales encontrados. Sin embargo, la alta cantidad de colonias de Fusarium spp. (5-10) en
tejido interno, por hijuelo, implica un riesgo fitosanitario en la movilización de material
propagativo a nivel regional, además, marca nuevos enfoques para el manejo y control en
hijuelos.
Cuadro 1. Resultados de los experimentos CIE, IFT y CII, en 2018 y 2019.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
58
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la beca otorgada.
Al Colegio de Postgraduados, Campus Montecillo por la oportunidad para desarrollar mi
posgrado. A las empresas Sauza, Tapatío y productores de agave que proporcionaron hijuelos
para realizar esta investigación. Al equipo de laboratorio CP-LANREF por el apoyo logístico.
LITERATURA CITADA
Castañeda V. H. 2002. Aislamiento e identificación de microorganismos responsables de la marchitez del agave tequilero. p. 21-24. In: Flores L. H. E. (ed). Análisis agroecológico del Agave tequilana Weber var. azul con énfasis en problemas fitosanitarios en Jalisco. CESAVEJAL, CIRPAC, INIFAP.
García M. C., López B. V., Bustamante M. V., Gutiérrez E. M. A., Sánchez A. G., Mora A. G. 2017. Distribución y prevalencia regional de Fusarium spp. asociadas a marchitez y pudrición seca del cogollo del agave azul (Agave tequilana) en Jalisco. Revista Mexicana de Fitopatología. 35: 177
Jiménez G. L. R., Mendoza R. C., Santana P. B., Coria C. J. J., Delgado M. F., Acevedo S. G., Guzmán H. E. y Mora A. G. 2017. Escala logarítmica diagramática de severidad para medición de pudrición seca del cogollo, marchitez y mancha gris del agave azul. Revista Mexicana de Fitopatología. 35: 177
Rubio C. R. 2007. Enfermedades del cultivo de agave. p. 169-195. In Rulfo V., F. O. et al. (ed.). Conocimiento y prácticas agronómicas para la producción de Agave tequilana Weber en la zona de denominación de origen del tequila. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro de Investigación Regional del Pacífico Centro.
Servicio de Información Agroalimentaria y Pesquera (SIAP). 2018. Consultado el 05-10-2019 en https://www.gob.mx/siap/acciones-y-programas/produccion-agricola-33119

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
59
DENSIDAD DE TRAMPAS CON FEROMONA PARA EL
MONITOREO DE Stenoma catenifer (LEPIDOPTERA:
ELACHISTIDAE) EN HUERTOS DE AGUACATE HASS
Guadalupe del Carmen Velázquez Martínez1*, Héctor González-Hernández1, Armando Equihua-
Martínez1, J. Refugio Lomeli1, Julio Cesar Rojas2, José López-Collado3
1Fitosanidad-Entomología y Acarología, Colegio de Postgraduados, Carretera México-Texcoco, Km 36.5, CP 56230, Montecillo, Texcoco,
Edo. de México, México; 2Grupo Ecología y Manejo de Artrópodos, El Colegio de la Frontera Sur, Carretera Antiguo Aeropuerto Km 2.5,
CP 30700, Tapachula, Chiapas, México; 3Colegio de Postgraduados, Campus Veracruz. Carretera federal Veracruz–Xalapa km 88.5, CP
91690, Veracruz, México. Correo: [email protected]
INTRODUCCIÓN
Las larvas de la palomilla barrenadora de la semilla del aguacate Stenoma catenifer
Walsingham perforan la pulpa del fruto para después alimentarse de la semilla, aunque
también pueden atacar y secar ramas terminales (Wolfenbarger y Colburn, 1979). En México,
S. catenifer se encuentra bajo control oficial y está distribuida en los estados de Colima,
Guerrero, Jalisco y Querétaro (SENASICA, 2018), donde en ausencia de acciones de manejo,
puede provocar pérdidas cercanas al 80% en huertos comerciales de aguacate Hass, además
de restringir la comercialización nacional e internacional de esta fruta. De esta forma, los
programas de trampeo diseñados para la detección de plagas, tienen el objetivo de prevenir
la dispersión de una especie indeseable, hacia nuevas áreas geográficas, así como monitorear
la dispersión de especies exóticas cuando se establecen en nuevas áreas (Buntin, 1994). Para
el caso de S. catenifer, el sistema de trampeo con feromona sexual sintética solo se ha
implementado para detectar la presencia y conocer la dinámica poblacional de esta plaga,
pero sin la determinación de la densidad de trampas por superficie. Por lo que, el objetivo de
la presente investigación fue determinar el número óptimo de trampas con feromona sexual
por unidad de superficie (una hectárea) para la detección o monitoreo de S. catenifer.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
60
MATERIALES Y MÉTODOS
Sistema de trampeo. El estudio se realizó durante los meses de enero a mayo de 2018,
en huertos comerciales de aguacate Hass, en los municipios de Comala y Cuauhtémoc,
Colima, México (Cuadro 1).
Cuadro 1. Ubicación de las parcelas experimentales
En el experimento se evaluaron cuatro densidades de trampeo: 1) 1 trampa/ 2 ha; 2) 1
trampa/ha, 3) 2 trampas/ha y 4) 3 trampas/ha. La trampa usada fue del tipo ala de cartón
PHEROCON® 1C (Trécé Inc., Adair, OK, EUA), con la feromona sexual sintética para S.
catenifer (septos de caucho) (Pherocon® Septa lures, Trécé Inc., Adair, OK, EUA). El
experimento se estableció con un diseño en bloques completos al azar, donde cada
tratamiento incluyó cuatro repeticiones (huertos). La distancia entre trampas fue de al menos
50 m. Las trampas se revisaban quincenalmente (del 03 enero a 09 de mayo de 2018). Las
bases de las trampas con pegamento y los septos con la feromona se cambiaban cada mes.
La variable respuesta fue el número de machos de la palomilla barrenadora de la semilla del
aguacate capturadas por trampa en cada uno de los tratamientos.
Análisis de datos. Se usó un modelo lineal generalizado mixto, que sigue una distribución
binomial negativa (BN). Este modelo empleado en la notación del lenguaje R fue:
PALOMILLAS~TRATAMIENTO + FECHA (1|HUERTA/TRAMPA), el cual representa
un modelo aditivo lineal en los conteos de los machos adultos de la PALOMILLA, de los
factores a la derecha de la tilde (~), es decir, en función del TRATAMIENTO, las FECHAS
de muestreos. Se consideró que la HUERTA es una muestra aleatoria de las posibles parcelas
en la zona, lo mismo que la TRAMPA que se encuentran anidadas dentro de las huertas. El
Municipio Huerto Ubicación Altitud (msnm)
Superficie (ha)
Edad de los arboles (años)
Comala Agosto 19°23’47”N 103°43’55”
O
1102 4.1 13
Rancho Alto
19°25´8”N; 103°42’19”O
1338 9.9 48
Piedra Rajada
19°24’ 50” N 103°44’ 8” O
1199 7.1 48
Cuauhtémoc La Calma 19°25’28”N 103°35’42”
O
1523 6.9 5

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
61
efecto de los tratamientos se obtuvo mediante una prueba de F y la comparación de medias
se hizo con la prueba de Tukey con un nivel de significancia de 0.05, con los datos
transformados de acuerdo al modelo BN, los resultados se presentan en las unidades
originales. Adicionalmente se hicieron 1000 corridas con bootstrap no paramétrico para
visualizar la variabilidad en las estimaciones de los totales (Thai et al., 2013). Los intervalos
de confianza (95%) se obtuvieron calculando los cuartiles (0.025, 0.975). Todos los análisis
estadísticos fueron realizados con el programa R v. 3.4.0 (R CoreTeam, 2019).
RESULTADOS Y DISCUSIÓN
Durante el periodo del 03 enero a 09 de mayo de 2018, se capturaron en total 2,321
palomillas macho de S. catenifer. En el huerto Piedra Rajada, se obtuvieron las mayores
capturas de palomillas en trampas durante todo el periodo de experimento, con un total de
1,004 adultos machos; le siguió el huerto Rancho Alto, con 769; después el Huerto Agosto,
con 276 y el huerto La Calma, con 272 adultos. Para los tiempos de muestreo, se observa que
las mayores capturas totales de palomillas fueron en los días julianos 102 (12 de abril) con
48 palomillas para el huerto Rancho Alto; 74 (15 de marzo), con 37 palomillas para el huerto
Piedra Rajada; 129 (09 de mayo) para el huerto La Calma, con 29 palomillas y 18 (18 de
enero), con 12 palomillas para el huerto El Agosto (Figura 1).
Figura 1. Número total de palomillas del gusano barrenador de la semilla del Stenoma catenifer, capturadas en trampas con feromona sexual sintética, a diferentes densidades de trampeo y en los diferentes huertos de aguacate Hass, en los municipios de Comala y Cuauhtémoc, Colima, México. 2018. Las columnas corresponden a tratamientos y las hileras a los huertos experimentales.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
62
El tratamiento con 3 trampas/ha presentó el mayor número de palomillas capturadas, tanto
en los cuatro huertos, como en las diferentes fechas de muestreo, con un promedio de 0.84
palomillas/trampa/noche. Las capturas de palomillas se ven afectadas por los tratamientos a
un IC 95. El tratamiento de 1 trampa/ha, tuvo una captura promedio de 0.75
palomillas/trampa/noche y los tratamientos con 1 trampa/2 ha y 2 trampas/ha tuvieron
capturas de palomillas muy similares, con promedios de 0.43 y 0.44
palomillas/trampa/noche, respectivamente. Los tratamientos con 1 trampa/2 ha, 1 y 2
trampas/ ha no fueron significativamente diferentes entre ellos (Figura 2). Además, con el
muestreo bootstrap no paramétrico, se observa que en los Huertos Rancho Alto, La Calma y
Piedra Rajada los tratamientos 1 trampa/2 ha, 1 y 2 trampas/ha tienen capturas similares y el
tratamiento de 3 trampas/ha capturó más palomillas. De esta manera se confirma que el
tratamiento 3 trampas/ha es el que capturó más palomillas.
Figura 2. Promedio acumulado de palomillas del gusano barrenador de la semilla del aguacate (IC95) capturadas en las trampas con feromona sexual sintética a diferentes densidades de trampeo, en huertos de aguacate Hass, Colima, México. 2018. Barras con las letras similares no son significativamente diferentes según prueba de Tukey (α= 0.05).
Las diferencias en el número de capturas de las palomillas en el tiempo, pueden deberse
a diversos factores, sin embargo, esta investigación se diseñó con el objetivo de tener un dato
más concreto del número de trampas/ha que se necesitan para monitorear de forma más
eficiente a la palomilla S. catenifer. Hoddle et al. (2011), para la detección de S. catenifer en
huertos de exportación, con superficie de hasta 2 ha, recomiendan colocar de 2 a 5 trampas,
distribuidas al azar; mientras que, para huertos de cerca de 76 ha, colocar de 10-13 trampas,
también distribuidas al azar. En Chiapas, en huertos comerciales de aguacate Hass y de
árboles mexicanos, así como en árboles de traspatio de aguacates mexicanos, usando trampas
con la feromona sexual sintética, capturaron a las palomillas S. catenifer y al también

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
63
Elachistidae Antaeotricha nictitans (Zeller). En esa investigación, se usaron cinco trampas,
en una superficie de 5 ha (Castillo et al., 2012). El SENASICA (2017), a través del Manual
Técnico para la implementación del plan de emergencia ante la detección de barrenadores de
la semilla del aguacate en zonas libres, describe que el trampeo para S. catenifer se debe
efectuar, tan pronto se detecte un foco de infestación, para lo cual se deben instalan 17
trampas en media hectárea, 1 en el árbol positivo y las restantes en cada punto cardinal cada
130 m, que permanecerán fijas durante los seis meses del manejo del foco de infestación;
muy similar a las recomendaciones de Hoddle et al. (2011) para la detección de al menos un
macho de S. catenifer. Finalmente, nuestros resultados indican que la cantidad de trampas
influye en la cantidad de insectos capturados de S. catenifer, ya que en el presente
experimento se encontró que, si se establece 1 trampa/2 has, 1 o 2 trampas/ha, no hay
diferencia significativa en la captura de palomillas, por lo que sugerimos utilizar 1 trampa/ha
para monitorear adecuadamente a la palomilla barrenadora de la semilla del aguacate ya que
fue la trampa segunda en eficiencias de capturas y es la económicamente más viable para el
productor.
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACyT), por la beca de doctorado
otorgada a la primera autora y por el financiamiento del Proyecto Control y Manejo de Plagas
Reglamentadas del Aguacatero, Clave: SAGARPA-2016-1-277881. SAGARPA-
CONACYT. Al Colegio de Postgraduados por la oportunidad de realizar mis estudios de
Doctorado. Al Centro Nacional de Referencia de Control Biológico por las facilidades
otorgadas en la logística de los muestreos. Al Comité Estatal de Sanidad Vegetal de Colima
(Programa de Plagas Reglamentadas del Aguacatero), por el apoyo para conseguir los huertos
experimentales.
LITERATURA CITADA
Buntin G.D. 1994. Developing a primary sampling program. En: L.P. Pedigo and G.D. Buntin, Handbook of sampling methods for arthropods in agriculture. CRC Press, Boca Raton, pp. 99-115.
Castillo, A., Cruz-López, L., and Gómez, J. 2012. Moth species captured with sex pheromone of Stenoma catenifer (Lepidoptera: Elachistidae) in avocado plantations of Southern Mexico. Fla. Entomol. 95: 1111-1116.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
64
Hoddle, M.S., Millar, J.G., Hoddle, C.D., Zou, Y., McElfresh, J.S., Lesch, S. M. 2011. Field optimization of the sex pheromone of Stenoma catenifer (Lepidoptera: Elachistidae): evaluation of lure types, trap height, male flight distances, and number of traps needed per avocado orchard for detection. Bulletin of Entomological Research, 101:145-152.
RStudio Team. 2015. RStudio: Integrated Development for R. RStudio, Inc., Boston, MA URL. En línea: https://www.rstudio.com/
SENASICA. 2017. Manual Técnico para la Implementación del Plan de Emergencia ante la Detección de Barrenadores del Hueso del Aguacate en Zonas Libres. Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria (SENASICA). Dirección General de Sanidad Vegetal. 31 p. En línea: https://www.gob.mx/cms/uploads/attachment/file/238756/Manual_T_cnico_Plan_de_Emergencia_ZL_FINAL_CIRCULAR_No__040.pdf. Fecha de consulta: 20 de junio de 2019
SENASICA.2018. Distribución nacional de la palomilla barrenadora del hueso del aguacate (Stenoma catenifer) en México. Mapa dinámico. Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria (SENASICA). En línea:
Thai, H.T., Mentre, F., Holford, H.G., Veyrart-Follet, C., Comets, E., 2013. A comparison of bootstrap approaches for estimating uncertainty of parameters in linear mixed effects models. Pharm. Stat. 12, 129–140.
Wolfenbarger D., O., and Colburn, B. 1979. The Stenoma catenifer, a serious avocado pest. Proc. Fla. State Hort. Soc. 92: 275.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
65
CARACTERIZACIÓN Y DISTRIBUCIÓN DE Colletotrichum spp.
CAUSANTES DE ANTRACNOSIS EN FRUTOS DE AGUACATE EN
MÉXICO, INCLUYENDO TRES NUEVAS ESPECIES
Dionicio Fuentes-Aragón1, Hilda Victoria Silva-Rojas2, Vladimiro Guarnaccia3, José Antonio Mora-
Aguilera1, Sergio Aranda-Ocampo1, Nestor Bautista-Martínez1, Daniel Téliz-Ortíz1*
1Postgrado en Fitosanidad, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco, Estado de México,
México. 2Postgrado en Recursos Genéticos, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco,
Estado de México, México. 3Universidad de Torino, Torino, Italia. *Autor para correspondencia: [email protected]
INTRODUCCIÓN
México es el principal productor y exportador de aguacate (Persea americana Mill) en el
mundo. En el 2017, se cosecharon más de 2 millones t en México, de las cuales el estado de
Michoacán produjo el 85 % (1 700 000 t) y se exportaron más de 800 000 t, principalmente
a Estados Unidos (FAOSTAT, 2019). México se considera el centro de origen del aguacate,
debido a la primera evidencia de domesticación y consumo datado en 7000-8000 años a. C.
en el valle de Tehuacán, Puebla (Smith, 1996).
El árbol de aguacate y sus frutos son afectados por diversos hongos como Colletotrichum,
Armillaria, Pseudocercospora, Dothiorella, Botryopshaeria spp., Neofusicoccum spp.,
Raffaelea lauricola y Sphaceloma perseae (Dann et al., 2013). Entre estos, Colletotrichum
se consideró en octavo lugar en importancia económica y científica (Dean et al., 2012).
Recientemente este género se reclasificó en complejos de especies utilizado análisis
filogenéticos multilocus (Weir et al. 2012).
El conocimiento de la diversidad de especies de Colletotrichum asociado a un hospedante
en particular es muy importante para la implementación de estrategias de control eficientes
de la antracnosis. Sin embargo, la diversidad y distribución de las especies de Colletotrichum
es muy pobre y desconocida en México. El objetivo de este estudio fue caracterizar e
identificar las especies de Colletotrichum asociadas a la antracnosis en frutos de aguacate en
México, mediante un análisis polifásico.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
66
MATERIALES Y MÉTODOS
Muestreo y aislamiento. Se muestrearon frutos con síntomas de antracnosis en
Michoacán, Morelos, Puebla, Nayarit, Guerrero y Oaxaca. Porciones de exocarpio se
colocaron en medio papa-dextrosa agar y se incubaron a 25 °C durante 5 días. Todos los
hongos culturalmente similares a Colletotrichum, se purificaron y se conservaron en glicerol
al 30% a -80 °C, para su uso posterior.
Caracterización morfológica. Aislamientos representativos de cada especie se
describieron cultural y morfológicamente. Se tomaron fotografías de conidios, setas, ascas,
ascosporas, conidióforos, acérvulos y apresorios utilizando un microscopio compuesto.
Posteriormente se midieron las estructuras usando el software ImageJ. Además, se describió
el color y se registró la tasa de crecimiento de las especies.
Análisis filogenético. La extracción de DNA se llevó a cabo mediante el método de
CTAB 2% (Doyle y Doyle, 1990). Posteriormente se amplificó el gen gliceraldehído-3-
fosfato dehidrogenasa como primer filtro de la diversidad genética. Posteriormente, se
seleccionaron diferentes haplotipos y se amplificaron la región ITS y los genes actina, quitina
sintetasa, tubulina-2, histona-3, glutamina sintetasa y la región intergénica entre los genes
Apn2. Se reconstruyó la filogenia mediante los métodos de Máxima Verosimilitud (MV) e
Inferencia Bayesiana (IB). Para MV se utilizó el software raxmlGUI (Silvestro y Michalak,
2012), donde se implementaron 1000 repeticiones bootstrap mediante un análisis rápido, y el
modelo evolutivo GTR + G (General Time Reversible más sitios con distribución gamma).
Para la IB, los análisis se llevaron a cabo en el software MrBayes v3.2 (Ronquist et al., 2012).
Se llevaron a cabo dos corridas independientes con cuatro cadenas de Markov Monte Carlo
cada una. Los análisis se detuvieron hasta que la desviación estándar de los ‘splits’ alcanzó
un valor menor a 0.01. Los primeros 25% de los datos generados se descartaron debido a la
fase de “calentamiento”. Los árboles generados se visualizaron y editaron usando el software
FigTree v1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/).
Pruebas de patogenicidad. Frutos de aguacate sin síntomas aparentes se desinfestaron y
se inocularon con 7 µL de una suspensión de conidios de 1x10-6. Previamente, se realizó una
lesión en el exocarpio con una aguja de disección. Los frutos se colocaron en cámaras

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
67
húmedas y se incubaron a 25 °C en oscuridad durante 7 días. Frutos inoculados con agua
destilada estéril se usaron como controles. Los síntomas se evaluaron de acuerdo con la escala
visual de antracnosis en frutos de aguacate (Sharma et al. 2017).
RESULTADOS Y DISCUSIÓN
Se obtuvieron 236 aislamientos, de los cuales se secuenció el gen gpadh de 106
aislamientos representativos, seleccionados con base en características culturales y
morfológicas similares. De estos, se seleccionaron 36 cepas para el análisis filogenético
multilocus. Los árboles filogenéticos generados en ambos análisis (MV y IB) mostraron
topologías similares. Se identificaron 12 especies correspondientes a cuatro complejos de
especies asociadas a la antracnosis en frutos de aguacate en México. Es decir, C.
chrysophilum, C. siamense, C. tropicale, C. jiangxiense y C. kahawae subsp. ciggaro en el
complejo C. gloeosporioides; C. godetiae, C. fioriniae y C. nymphaeae en el complejo C.
acutatum; C. kasrtii y C. morelense sp. nov. en el complejo C. boninense; C. pueblense sp.
nov. en el complejo C. magnum; y C. michoacanense sp. nov. como especie simple. Las
nuevas especies y las reportadas por primera vez en el mundo, asociadas a la antracnosis en
aguacate, presentaron muy baja frecuencia de aislamientos, además de encontrarse en una
sola región productora. En México, solamente se han reportado a C. godetiae, C. karstii y C.
fructicola como causantes de antracnosis en aguacate (Velázquez-del Valle et al., 2016;
Fuentes-Aragón et al., 2018). Aunque, Silva-Rojas y Ávila-Quezada (2011) reportaron a C.
gloeosporioides, C. boninense y C. acutatum como causantes de antracnosis en aguacate,
solo han utilizado la región ITS para la reconstrucción filogenética, la cual se ha demostrado
que no posee la suficiente resolución para discriminar las especies dentro de los complejos
(Liu et al., 2015).
Colletotrichum siamense fue la más abundante en frecuencia de aislamientos (35.56%),
seguido por C. kahawae subsp. ciggaro (20.08%), C. karstii (10.87%) y C. godetiae
(10.04%). La mayor diversidad genética se encontró en la región de Puebla con siete especies
presentes (C. kahawae subsp. ciggaro, C. godetiae, C. karstii, C. fioriniae, C. chrysophilum,
C. nymphaeae and C. pueblense sp. nov.). La alta diversidad en esta región puede deberse a
dos razones; la primera, debido a que el centro de origen del aguacate se ubicó en esta región

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
68
y ha permitido la coevolución con Colletotrichum; segunda, la capacidad de patogenicidad
cruzada de las especies de Colletototrichum que afectan a otros cultivos frutícolas como pera,
manzana y durazno. Por ejemplo, se han asociado con alta frecuencia de aislamiento a C.
fioriniae, C. nymphaeae, C. godetiae, C. karstii y C. siamense, causantes de antracnosis en
pera, manzana y durazno en diversas partes del mundo (Bragança et al. 2016; Grammen et
al. 2018).
La morfología de las especies fue similar con las cepas tipo o ex-tipo, en cuanto a tamaño
y forma de conidio. Características sobresalientes de las especies nuevas en medio PDA
fueron: C. michoacanense sp. nov. presentó, conidios cilíndricos, ligeramente agudos en uno
de los extremos, de 16.7 × 5.0 µm; setas que variaron de 67 a 102 µm de longitud. La colonia
presentó apariencia algodonosa con micelio aéreo, presencia abundante de masas de conidios
en la parte central. Colletotrichum morelense sp. nov. presentó conidios cilíndricos, de 14.3
× 6.1 µm, conidióforos muy largos que variaron de 53.7 a 94.7 µm en longitud y setas de 120
µm. La colonia presentó poco micelio, con abundantes masas de conidios dispersos en la caja
Petri. Colletotrcichum pueblense sp. nov presentó conidios cilíndricos, algunos ligeramente
fusiformes, de 16.1 × 4.7 µm, y abundante presencia de setas. La colonia presentó micelio
superficial, de color café-claro, que se torna café-oscuro con el tiempo; ausencia aparente de
masas de conidios.
Todas las especies evaluadas en las pruebas de patogenicidad sobre frutos de aguacate
fueron patogénicas. C. siamense y C. tropicale fueron las más virulentas (categoría 8 y 9).
Síntomas típicos de antracnosis se observaron a los 7 días después de la inoculación, mientras
que el control no presentó síntomas. Las cepas inoculadas se reaislaron presentando la misma
morfología y secuencia en gapdh, completando los postulados de Koch. En estudios previos,
el método de inoculación sin herida no mostró síntomas aparentes a los 7 días después de la
inoculación, para ninguna de las especies. Resultados similares se encontraron para nueve
especies causantes de antracnosis en aguacate en Israel, donde no causaron síntomas en cv.
Hass con el método de inoculación sin herida (Sharma et al. 2017).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
69
Conocer las especies causantes de antracnosis ayudará a mejorar las estrategias de control
para mejorar la calidad de los frutos, así como evitar la pérdida de rendimiento en presencia
de la enfermedad en etapas tempranas de desarrollo del fruto.
AGRADECIMIENTOS
Al CONACyT por la beca otorgada al primer autor para la realización de sus estudios de
Doctorado. Al Colegio de Postgraduados por las facilidades proporcionadas para la
realización de la investigación.
LITERATURA CITADA
Bragança, C. A. D., U. Damm, R. Baroncelli, N. S. M. Junior, P. W. Crous. 2016. Species of Colletotrichum acutatum complex associated with anthracnose diseases of fruit in Brazil. Fungal Biol. 120: 547-561.
Dann EK, R.C. Ploetz, L. M. Coates, and K. G. Pegg. 2013. Foliar, fruit and soilborne diseases. In: The Avocado. Botany, Production and Uses. B. Schaffer, B. N. Wolstenholme, A. W. Whiley (Eds.). Second Edition. CAB International. Oxfordshire, UK. Pp:380-422.
Dean R., J. L. Kann, Z. A. Pretorius, et al. 2012. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 13:414-430.
FAOSTAT, 2019. Producción y exportación de aguacate. http://www.fao.org/faostat/es/?#data/QC/visualize (Consulta: Octubre 2019).
Fuentes-Aragón D, S. B. Juárez-Vázquez, M. Vargas-Hernández, and H. V. Silva-Rojas. 2018. Colletotrichum fructicola, a member of Colletotrichum gloeosporioides sensu lato is the causal agent of anthracnose and soft rot in avocado fruits cv.‘Hass’. Mycobiology. 46: 92-100.
Grammen, A., M. Wenneker, J. V. Campenhout, K. T. K. Pham, W. V. Hemelrijck, D. Bylemans, A. Geeraerd, and W. Keulemans. 2019. Identification and pathogenicity assessment of Colletotrichum isolates causing bitter rot of apple fruit in Belgium. Eur. J. Plant Pathol. 153: 47-63.
Liu F, B. S. Weir, U. Damm, P. W. Crous, Y. Wang, B. Liu, M. Wang, M. Zhang, and L. Cai. 2015. Unravelling Colletotrichum species associated with Camellia: employing ApMat and GS loci to resolve species in the C. gloeosporioides complex. Persoonia. 35: 63-86
Silvestro, D., and I. Michalak. 2012. raxmlGUI: a graphical front-end for RAxML. Org. Divers. Evol. 12: 335-337.
Sharma G, M. Maymon, and S. Freeman. 2017. Epidemiology, pathology and identification of Colletotrichum including a novel species associated with avocado (Persea americana) anthracnose in Israel. Sci. Rep. 7: 15839.
Ronquist, F., N. Teslenko, P. van der Mark, D. L. Ayres, A. Darling, S. Höhna, B. Larget, L. Liu, M. A. Suchard, and J. P. Huelsenbeck. 2012. MrBayes 3.2: efficient Bayesian

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
70
phylogenetic inference and model choice across a large model space. Syst Biol. 61:539-542.
Silva-Rojas HV, and G. D. Ávila-Quezada GD. Phylogenetic and morphological identification of Colletotrichum boninense: a novel causal agent of anthracnose in avocado. Plant Pathol. 60: 899-908.
Smith C. E. J. 1966. Archeological evidence for selection in avocado. Econ. Bot. 20:169-175.
Velázquez-del Valle MG, A. Campos-Martínez, H. E. Flores-Moctezuma, R. Suárez-Rodríguez, J. A. Ramírez-Trujillo, A. N. Hernández-Lauzardo. 2016. First report of avocado anthracnose caused by Colletotrichum karstii in Mexico. Plant Dis. 100:534.
Weir B. S., P. R. Johnston, and U. Damm. 2012. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 73: 115-180.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
71
RESPUESTA A LA INFECCIÓN POR Fusarium euwallaceae
(HYPOCREALES: NECTRIACEAE) EN CUATRO VARIEDADES DE
AGUACATE EN MÉXICO
Otilio Cruz-Martínez1, Daniel Teliz-Ortíz1*, Clemente de Jesús García-Ávila2, Dionicio Alvarado-
Rosales1
1Posgrado en Fitosanidad-Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco,
Estado de México, México.2Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria, Km. 37.5 Carretera Federal México-
Pachuca, 55740 Tecámac, Edo. de México, México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
México es el principal productor y exportador de aguacate en el mundo (FAOSTAT, 2019,
TRADEMAP, 2019), cuenta con una superficie sembrada de 218 492.93 ha (SIAP, 2019) y
permite la generación de 400,000 empleos directos e indirectos, además de emplear más de
9 millones de jornales al año. La actividad productiva genera importantes divisas e ingresos
que contribuyen a mejorar la calidad de vida de los más de 23 300 productores que participan
en cada uno de los eslabones de la cadena de valor. Los principales estados productores del
cultivo son: Michoacán, Jalisco, Edo. de México, Colima y Nayarit (APEAM, 2018;
APEAJAL, 2018, y SIAP, 2019, con datos de cierre 2017).
La producción de aguacate y otros frutales se ve amenazada por el hongo Fusarium
euwallaceae el cual fue detectado en el Municipio de Tijuana, Baja California, México
(NAPO, 2016). Este hongo (Figura 1) afecta los haces vasculares de las plantas hospedantes,
lo cual impide el transporte de agua y nutrientes, ocasionando el marchitamiento de ramas y
eventualmente la muerte de la planta (Freeman, 2013). Por lo anterior, el presente trabajo
tuvo el objetivo de evaluar la respuesta de cuatro variedades de aguacate cultivadas en
México, a la infección por F. euwallaceae, a fin de proporcionar a las autoridades
fitosanitarias elementos técnicos y científicos que permitan fortalecer las estrategias de
vigilancia y manejo de la enfermedad.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
72
Figura 1. Fusarium euwallaceae; a) Crecimiento micelial en forma radial en medio de
cultivo Papa-dextrosa-Agar (PDA); b) Conidios multiseptados, con la parte media superior
más ancha, célula apical redondeada y papilada; c) Corte longitudinal de tallo de aguacate
inoculado con F. euwallaceae.
MATERIALES Y MÉTODOS
El experimento se realizó bajo condiciones de invernadero, considerados como de
bioseguridad nivel 2, ubicado en la Unidad Integral de Servicios, Diagnóstico y Constatación
(UISDC), del Servicio Nacional de Sanidad Inocuidad y Calidad Agroalimentaria
(SENASICA), en Tecámac, Estado de México. El inoculo de Fusarium euwallaceae se
obtuvo de tejido vegetal enfermo colectado en Tijuana, Baja California, México. Las cepas
del hongo fueron identificadas y caracterizadas morfológicamente y confirmadas mediante
la extracción de DNA y la técnica de PCR. Las cuatro variedades de aguacate evaluadas
fueron 1) Hass flor de María, 2) Hass Méndez, 3) Hass y 4) Criollo Mexicano, de 2 años de
edad, con una altura promedio de 177 cm y 1.33 cm de diámetro del tallo.
El experimento se llevó a cabo bajo un dieseño completamente al azar, con dos factores
de estudio, A=Variedad de la planta de aguacate y B= El efecto del hongo Fusarium
euwallaceae; el factor A consideró cuatro niveles: 1= Flor de María, 2= Méndez, 3=Hass y
4= Criollo; el factor B consideró dos niveles: 1 = Plantas Inoculadas con F. euwallaceae y 2
= Plantas inoculadas con agua estéril. Al realizar todas las combinaciones entre factores y
niveles se obtuvieron 8 tratamientos. Cada tratamiento contó con cinco repeticiones.
La inoculación se realizó a través de la inyección de 20 µl de una suspensión con 3.6
millones de conidios/ml en tres orificios del tallo principal de la planta provocados con un
a
)
b
)
c
)

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
73
taladro y una broca de 1 mm. El primer orificio se realizó a 40 cm, el segundo a 60 cm y el
tercero a 80 cm de la parte basal del suelo de la maceta. Posteriormente se colocó una rodaja
de 1 cm2 de PDA estéril, con la finalidad de proporcionar alimento al hongo en los primeros
días de inoculación. Por otra parte, a las plantas usadas como testigo se inyectaron con 20 µl
de agua destilada estéril. Finalmente, se selló la herida con vaselina y Parafilm®.
Para determinar el efecto de la infección por F. euwallaceae, se consideró la toma de
variables respuesta a partir del 12 de diciembre de 2018 hasta el 12 de abril de 2019. El
experimento se realizó en dos fases, en la primera se tomaron medidas repetidas con
intervalos de 30 días, las variables medidas fueron: 1) Altura de la planta (A), (midiendo con
flexómetro de 3 m) desde el piso hasta el ápice de la planta. 2) Diámetro de tallo (DT)
(midiendo con vernier (de 16 cm) en los tres puntos donde se realizó la inoculación. 3)
Número de ramas (NR) (se contaron todas las ramas presentes en árbol). La segunda fase se
realizó al final del experimento (a los 120 días después de la inoculación), en donde se midió
la longitud de la necrosis en la base del tallo (LNBT), longitud de la necrosis en la mitad del
tallo (LNMT) y longitud de la necrosis en el ápice del tallo (LNAT), derivado de la
inoculación de F. euwallaceae, con una navaja de disección se hicieron cortes longitudinales
y con una regla se midió el tejido con necrosis visible.
Para confirmar la patogenicidad de F. euwallaceae se tomaron muestras de las plantas
inoculadas con el hongo y testigos. Posteriormente se realizó el aislamiento e identificación
a través de morfología.
Finalmente, se realizó el análisis utilizando el paquete estadístico SAS, con los datos
mensuales de longitud, diámetro, número de ramas y longitud de la necrosis. El modelo
utilizado fue: yij=μ+ai+bj+abij+εij. Donde: Yij= Variable respuesta; µ= Media general; ai= el
efecto de la i-ésima variedad; bj= Efecto del j-ésimo tipo de inoculación; abij= el efecto
conjunto de la variedad y el tipo de inoculación; y εij= el error experimental.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
74
RESULTADOS Y DISCUSIÓN
Las variables morfológicas, altura de la planta, diámetro basal, medio y apical, así como
el número de ramas en las cuatro variedades de estudio no fueron afectadas
significativamente (p≤ 0.05) al ser inoculadas con F. euwallaceae. Esto probablemente se
debe a que en un ambiente natural las plantas son atacadas por el complejo simbiótico
Euwallacea sp. – Fusarium euwallaceae, donde los escarabajos ambrosiales construyen
galerías en el xilema, mientras que la propagación de su simbionte fúngico puede obstruir el
transporte de agua y minerales, dando como resultado el marchitamiento y la muerte del árbol
hospedante (Freeman et al., 2013), mientras que en el presente estudio se analizó únicamente
el efecto del hongo en las cuatro variedades de aguacate, sin estar presente el vector.
Por otra parte, la inoculación en la zona apical del tallo causó diferencias altamente
significativas (p≤ 0.05) [Cuadro 1]. La variedad que presentó el mayor daño por necrosis
(LN) del tejido fue el criollo mexicano, con valores medios de 11.8 cm de longitud, seguido
de la variedad Hass y la variedad Flor de María con 7.6 cm y 7.3 cm de longitud,
respectivamente. Mientras que la variedad Méndez tuvo el menor daño, con valores medios
de 3.34 cm de longitud de la necrosis del tejido. En contraste, las plantas testigo, inoculadas
con agua destilada estéril, expresaron una necrosis propia del proceso de cicatrización de la
herida, cuyos valores medios fueron de 0.38 a 0.66 cm, los cuales se obtuvieron en las
variedades Flor de María y Méndez; mientras que en la variedad Hass y Criollo Mexicano se
tuvieron valores medios superiores, 2.22 cm y 1.52 cm ,respectivamente; esto probablemente
se debió a que durante la inoculación o posterior a ella, tres plantas de la variedad Hass se
infectaron con otros organismos, entre los que se identificaron los hongos Acremonium sp.,
Alternaria alternata y Aspergillus niger. Mientras que para las plantas de aguacate criollo se
encontraron dos plantas con Alternaria alternata y Aspergillus niger. Es importante
mencionar que en todas las plantas usadas como testigo no se aisló el hongo Fusarium
euwallaceae.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
75
Cuadro 1. Efecto de la inoculación de Fusarium euwallaceae en el nivel de daño LN de cuatro variedades de aguacate
Tratamiento Variedad LNBT LNMT LNAT
Inoculadas con F. euwallaceae
Flor de María 9.2 NS 10.1 NS 7.3 **
Méndez 6.4 NS 6.12 NS 3.34 **
Hass 8.3 NS 9.9 NS 7.6 **
Criollo 8.55 NS 9.38 NS 11.88 **
Agua destilada estéril
Flor de María 1.01 NS 1.02 NS 0.66 **
Méndez 0.61 NS 1.52 NS 0.38 **
Hass 2.79 NS 3.05 NS 2.22 **
Criollo 0.336 NS 0.36 NS 1.524 **
** Altamente significativo (significancia a una probabilidad de p≤ 0.05).
NS: El efecto del tratamiento sobre el nivel de daño LN no es estadísticamente significativo.
Con base en los resultados obtenidos en el presente estudio se concluye de forma
preliminar que a pesar de existir muerte celular del tejido en los sitios con inoculación de
Fusarium euwallaceae, éste no es capaz de afectar por sí solo las variables morfológicas
evaluadas durante los 120 después de la inoculación, por lo que su virulencia está
determinada por la presencia del insecto vector, el cual se encarga de diseminarlo en
diferentes partes del árbol. Esto sugiere que, en caso de dispersarse y establecerse en áreas
de cultivo, una de las estrategias de manejo podría enfocarse al control del vector.
AGRADECIMIENTOS
Agradecemos al Colegio de Postgraduados, Campus Montecillo, por el acompañamiento
y la asesoría de sus Profesores. Al Servicio Nacional de Sanidad, Inocuidad y Calidad
Agroalimentaria por el apoyo con las instalaciones y equipos para el desarrollo de la
investigación.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
76
LITERATURA CITADA
APEAJAL, 2018. Asociación de Productores Exportadores de Aguacate de Jalisco, A.C. http://www.congresoaguacate.com/apeajal-1. Fecha de consulta 25 de Julio de 2018.
APEAM, A.C., 2018. Comunicado de prensa. Por atentar y poner en riesgo a la industria del aguacate de México, APEAM realiza proceso de expulsión conforme a sus estatutos. Asociación de Productores y Empacadores Exportadores de Aguacate de México. En línea: http://www.apeamac.com/archivos/prensa/comunicado-de-prensa-8-feb-2017.pdf.
FAOSTAT. 2019. “Statistics of crops production by country” Food and Agriculture Organization. En línea: http://www.fao.org/faostat/es/#rankings/countries_by_commodity. fecha de consulta: mayo de 2019.
Freeman, Stanley & Sharon, Michal & Maymon, M & Mendel, Zvi & Protasov, Alex & Aoki, Takayuki & Eskalen, Akif & O'Donnell, K. (2013). Fusarium euwallaceae sp nov.-a symbiotic fungus of Euwallacea sp., an invasive ambrosia beetle in Israel and California. Mycologia. 105. 10.3852/13-066.
NAPPO. Sistema de Alerta Fitosanitaria – Notificación oficial de plagas. 2016. Detección del escarabajo barrenador Euwallacea sp. y su hongo simbionte Fusarium euwallaceae, en zonas urbanas de Tijuana, Baja California, México. en Línea: https://www.pestalerts.org/espanol/oprDetail.cfm?oprID=680. Fecha de consulta: mayo de 2019.
SIAP. 2019. Cierre de producción agrícola por cultivo. Ciclo agrícola 2017. Servicio de Información Agroalimentaria y Pesquera. En línea: http://www.siap.gob.mx/cierre-de-la-produccion-agricola-por-cultivo/. Fecha de consulta: abril de 2019.
TRADEMAP, 2019. Estadísticas del comercio para el desarrollo internacional de las empresas. Datos comerciales mensuales, trimestrales y anuales. Valores de importación y exportación, volúmenes, tasas de crecimiento, cuotas de mercado, etc. En línea: http://www.trademap.org/Index.aspx. Fecha de consulta: 30 de mayo de 2019.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
77
PATOGENICIDAD DE HONGOS ASOCIADOS A Euwallacea nr.
fornicatus EN AGUACATE (Persea americana Mill.) cv. HASS
Nayeli Carrillo-Ortiz1,2, Cristian Nava-Díaz1*, Victoria Ayala-Escobar1, Magnolia Moreno-
Velázquez2
1Posgrado en Fitosanidad - Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, 56230 Texcoco, Estado de
México, México. 2Centro Nacional de Referencia Fitosanitaria, Dirección General de Sanidad Vegetal, Carretera Federal México- Pachuca
km 37.5, Centenario de la Educación, Col. Santa Ana, 5740 Tecámac, Estado de México. *Autor de correspondencia: [email protected].
INTRODUCCIÓN
El daño de muerte regresiva en los árboles de aguacate (Persea americana Miller) es causado
por especies simbióticas que pertenecen al Clado 3 del complejo Fusarium solani, de los
cuales su vector es un escarabajo ambrosial (Mendel et al., 2012). Adicionalmente, en las
galerías se desarrollan un conjunto de hongos, levaduras y bacterias, denominados complejos
multiespecíficos (Norris, 1965; citado por Mueller et al., 2005). El escarabajo ambrosial
identificado morfológicamente como Euwallacea fornicatus consiste de varias especies
cripticas con diferencias en sus secuencias de DNA. Stouthamer et al. (2017) identificó tres
grandes linajes (clados) con filogeografía distinta. Una de estas especies, Kuroshio Shot Hole
Borer (KSHB) se reportó por primera vez en Tijuana, Baja California, México en 2016
(García et al. 2016). El hongo asociado al escarabajo KSHB fue identificado como Fusarium
kuroshio, posteriormente se asoció a Graphium kuroshio sp nv. como parte de un complejo
de hongos en simbiosis (Na et al. 2017). Esta asociación representa un serio riesgo para la
producción de aguacate y otras especies de importancia agrícola y forestal en el país. Por
ello, el objetivo del presente trabajo es determinar el grado de patogenicidad de los hongos
asociados al escarabajo ambrosial en aguacate cultivar Hass, inoculados de manera individual
y en combinaciones entre ellos. Así también, corroborar mediante pruebas moleculares la
identidad de E.nr fornicatus y el diagnóstico de las especies de los hongos asociados a este.
MATERIALES Y MÉTODOS
Colecta de material. Se colecto tejido vegetal e insectos de las galerías en el xilema de
árboles (Liquidambar sp., Populus sp., y Erythrina sp.) con síntomas y signos característicos
de la muerte regresiva por Fusarium en los municipios de Tijuana y Rosarito, Baja California.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
78
Los especímenes se colocaron en viales con agua destilada estéril, adecuadamente
etiquetados y sellados.
Identificación de insecto. La identificación morfológica del género de los escarabajos
ambrosiales se llevó a cabo con la clave de Wood (1982) y Rabaglia et al. (2006). Se tomaron
fotografías con microscopio electrónico de barrido JEOL GSM6390 en el Colegio de
Postgraduados. Para la identificación molecular, se hizo la extracción de DNA total de los
insectos según el protocolo de diagnóstico fitosanitario “Amplificación de la Región mtDNA
CO-I, para la identificación de insectos por PCR punto final” (CNRF-SENASICA, 2018).
Los amplicones obtenidos con los oligonucleótidos LCO01490 y HC02198 se enviaron a
secuenciar a Macrogen, Inc. Las secuencias se compararon en la base de datos de secuencias
génicas NCBI (National Center for Biotechnology Information). Con la ayuda del algoritmo
BLAST (Basic Local Alignment Search Tool) se observó la identidad arrojada. Se realizó un
alineamiento múltiple de secuencias y construcción de árbol filogenético con el Software Bio
Edit Sequence Alignment Editor v7.2.5 y MEGA7.
Aislamiento de hongos a partir de insectos. Se desinfestaron usando la metodología de
You et al. (2015). Se sembró por estriado 50 μL del agua de los enjuagues en medio Papa
Dextrosa Agar (PDA) con la finalidad de descartar crecimientos de hongos no objetivo. La
cabeza y cuerpo de los insectos se maceraron por separado en 200 μL de agua destilada
estéril, se tomaron 100 μL de la suspensión y se sembró por estriado en medio PDA e
incubaron a 25 °C durante 5 días bajo oscuridad. Para obtener cultivos puros, por medio de
las técnicas punta de hifa y monospóricos se aislaron los diferentes crecimientos.
Aislamiento de hongos a partir de tejido. Se tomaron pequeños trozos del xilema con
coloraciones café marrón a café oscuro alrededor de las galerías. Para desinfestarlos, se
utilizó la metodología propuesta por Freeman et al. (2013) y se sembraron en cajas Petri con
medio PDA e incubaron a 25°C bajo oscuridad durante cinco días. Los cultivos
permanecieron en revisión constante durante 10, 15 y 30 días.
Identificación. A partir de preparaciones permanentes y caracteres morfológicos se
identificaron a género. Para la identificación molecular se realizó extracción de las cepas
puras con el método de propuesto por Cenis (1992). Se utilizaron los primers ITS 1 e ITS 4
propuestos por White et al. (1990) para las cepas de Graphium sp., los primers EF-1 y EF-2

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
79
(O´Donnell et al., 1998), los primers F5 y R8 (RPB1) (O´Donnell et al., 2010) y los primers
5F2/7CR (RPB2) (O´Donnell et al., 2007) para las cepas de Fusarium sp. La mezcla de
reacción fue preparada a una concentración final de Buffer 1X, 1.5 mM de MgCl2, 0.2 mM
de DNTP´s, 0.5µM de cada oligonucleótido, 2.5 U de TaqPolymerasa y 2 ng/µL de DNA,
dando un volumen final de 25 µL, los productos fueron amplificados en un termociclador
T100TMBioRad y un TechneFlexigene utilizando los parámetros de ciclaje modificados de
White et al. (1990), O´Donnell et al. (1998), O´Donnell et al. (2007) y O´Donnell et al.
(2010). Los amplicones obtenidos se secuenciaron en el laboratorio de biología molecular
del CNRF, en un secuenciador Aplied Biosystems 3230® de cuatro capilares. Las secuencias
obtenidas se compararon en BLAST a través del NCBI. Además el análisis múltiple de
secuencias y la construcción de árboles filogenéticos se realizaron con los Softwares Bio Edit
Sequence Alignment Editor v7.2.5 y MEGA7.
Pruebas de patogenicidad. Se inocularon árboles de aguacate cv. Hass de
aproximadamente un año de edad. Los tratamientos consistieron en:
Cuadro 1. Descripción de los tratamientos a empleados en el experimento
Tratamientos Composición Repeticiones
T I Testigo 5
T II F. kuroshium 5
T III F. solani 5
T IV Grahiumsp. 5
T V F. kuroshium + F. solani 5
T VI F. kuroshium + Graphiumsp. 5
T VII F. solani + Graphiumsp. 5
T VIII F. kuroshium + F. solani + Graphiumsp. 5
Se realizaron evaluaciones mensuales, tomando en cuenta la altura y el grosor de cada
planta. Los datos se analizaron con ANOVA utilizando el software SAS Versión 9.0. Para la
comparación de medias se usó la prueba Tukey (α=0,05).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
80
RESULTADOS Y DISCUSIÓN
El escarabajo ambrosial fue identificado por morfología como Euwallacea sp., y se
ratificó molecularmente como Euwallacea sp. # 5(KSHB) con una cobertura e identidad de
100% en GenBank, dando el mismo resultado con análisis filogenético. Los hongos con
mayor presencia se identificaron por morfología como Graphium sp., Fusarium euwallaceae
y F. solani. Para las cepas del genero Fusarium el análisis in silico a través de BLAST de los
oligonucleótidos EF-1α y RPB1 en GenBank mostraron diferencias en similitud (99% a 98%)
con secuencias de Fusarium sp., F. solani y F. euwallaceae principalmente. Mientras que el
análisis de secuencias con Bio Edit Editor v7.2.5 y MEGA7, se identificaron las especies
como F. kuroshium y F. solani. Para el género Graphium, las secuencias con los
oligonucleótidos ITS14 tuvieron una similitud de 99% con las secuencias de Graphium
jumulu, G. Penicillioides, y G. basintruncatum. Debido a que no se obtuvo la identidad exacta
con ITS mediante análisis de secuencias, se requiere la amplificación de los oligonucleótidos
EF1F (GCGGTGGTATCGACAAGCGT) y EF2R (AGCATGTTGTCGCCGTTGAAG)
como lo sugiere Lynch et al. (2017) y Na et al. (2017), para la determinación de la especie
de este último. Las pruebas de patogenicidad no han mostrado síntomas ni diferencias
estadísticamente significativas. Na et al. (2017) mostraron la patogenicidad en aguacates
jóvenes de F.kuroshium y G. kuroshium en menor grado. Mientras que Freeman et al. (2018)
menciona que estos no actúan como patógenos bajo infestación natural en el xilema.
AGRADECIMIENTOS
Mi más sincero agradecimiento al Colegio de Postgraduados, Campus Montecillo por
permitirme estudiar una maestría y a los Profesores que me han enseñado y asesorado en este
trayecto.
LITERATURA CITADA
Cenis, L. J. (1992). Rapid extraction of fungal DNA for PCR amplification. NucleicAcidsResearch, 20, 2380. doi: 10.1093/nar/20.9.2380
Freeman, S., A. Protasov, M. Sharon, K. Mohotti, M. Eliyahu, N. Okon, M. Maymon, and Z. Mendel. 2013 Fusarium euwallaceaesp. nov. - a symbioticfungus of Euwallaceasp., aninvasive ambrosia beetle in Israel and California. Mycologia. 105 (6): 1595 – 1606.doi: 10.1007/s13199-013-0222-6

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
81
García, C., Trujillo, F.J., López, J.A., González, R., Carrillo, D., Cruz, L., Ruiz, I., Quezada, A. y, N. Acevedo. 2016. Firstreport of Euwallaceaenr. fornicatus (Coleptera: Curculionidae) in México. Florida. EntomologicalSociety 555-556
Mendel, Z., Protasov, A., Sharon, M., Zveibil, A., Yehuda, S.B., y Freeman, S. (2012). AnAsian ambrosia beetleEuwallaceafornicates and its novel symbioticfungus Fusarium sp., pose a seriousthreat to theIsraeli avocado industry. Phytoparasitica, 40, 1-4. doi: 10.1007/s12600-012-0223-7
Mueller, U.G., Gerardo, N.M, Aenen, D.K., Six, D.L. &Schultz, T.R. (2005) Theevolution of agricultura in insects. Rev. Ecol. Syst., 36: 536-95. doi: 0.1146/annurev.ecolsys.36.102003.152626
SENASICA (2018). Amplificación de la Región mtDNA CO-I, para la identificación de insectos por PCR punto final. Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria. México D.F. Protocolo de Diagnóstico. 30 p
Stouthamer, R., Rugman-Jones, P., Thu, P.Q., Eskalen, A., Thibault, T., Hulcr, J., Wang, L-J., Jordal,B.H., Chen, C-Y., Cooperband, M., Lin,C-S., Kamata, N. Lu. S-S., Masuya, H., Mendel, Z., Rabaglia, R., Sanguansub, S., Shih, S.S. Sittichaya, W. y Zong, S. (2017). Tracingtheorigin of a crypticinvader: phylogeography of theEuwallaceafornicatus (Coleoptera: Curculionidae: Scolytinae) speciescomplex. Agricultural and ForestEntomology, 1-10. doi: 10.1111/afe.12215
Na, F., Carrillo, J.D., Mayorquin, J.S., Ndinga-Muniania, C., Stajich, J.E., Stouthamer,R., Huang, Y.T., Lin, Y.T., Chen, C.Y. y Eskalen, A. (2017). Two novel fungalsymbionts Fusarium kuroshiumsp., nov. andGraphiumkuroshiumsp. nov. ofKuroshioShotHoleBorer (Euwallaceaesp. nr. fornicatus) cause Fusarium Diebackonwoody host species in California. PlantDisease. 102, 1-11. doi: 10.1094/PDIS-07-17-1042-RE
O´Donell, K., Cigelnik, E., &Nirenberg, H.I. (1998). Molecular systematics and phylogeography of theGibberellafujikuroispeciescomplex. Mycologia. 90 (3), 465-493.
Rabaglia, R. J, S.A. Dole, A. Cognato. 2006. Review of American Xyleborina (Coleoptera: Curculionidae: Scolytinae) occurringnorth of Mexico, withanillustratedkey. Annals of theEntomologicalSociety of America. 99: 1034–1055.
O’Donnell, K., B. A. J. Sarver, M. Brandt, D. C. Chang, J. Noble-Wang, B. J.Park, D. A. Sutton, L. Benjamin, M. Lindsley, A. Padhye, D. M. Geiser, y T. J. Ward. 2007. Phylogeneticdiversity and microspherearray-basedgenotypingof human pathogenicfusaria, includingisolatesfromthemultistatecontactlens-associated U.S. keratitisoutbreaks of 2005 and 2006. J. Clin.Microbiol. 45:2235–2248.
O´Donell, K., Sutton, D.A., Rinaldi, M.G., Sarver, B.A.J., Balajee, S.A., Schroers, H-J., Summerbell, R.C., Robert, V., Crous, P.W., Zhang, N., Aoki, T., Jung, K., Park, J., Lee, Y.H., Kang., S., Park, B., y Geiser D.M. (2010). Internet-Accessible DNA SequenceDatabaseforIdentifyingFusariafrom Human and Animal Infections. J. Clin. Microbiol. 3708-3718 p.
You, L., D.R. Simmons, C. Bateman, D. Short, T. Kasson, and R. Rabaglia. 2015. 1 New Fungus-InsectSymbiosis: Culturing, Molecular, and HistologicalMethods Determine 2 SaprophyticPolyporalesMutualists of Ambrosiodmus Ambrosia Beetles. PlosOne. 13 p

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
82
White, J. T., Bruns, T., Lee, S., Taylor, J. (1990). Amplification and directsequencing of fungalribosomal RNA genes forphylogenetics. PCR Protocols: A guide to methods and applications. AcademicPress. 315-322
Wood, S. L. 1982. Thebark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae), a taxonomicmonograph. Great BasinNaturalistMemoirs 6: 1– 5 356.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
83
ESCAMAS ARMADAS (Hemiptera: Diaspididae) Y SUS
PARASITOIDES EN AGUACATE (Persea americana Mill.) EN
CUATRO ENTIDADES DE LA REGIÓN CENTRO OCCIDENTE DE
MÉXICO
Lázaro-Castellanos Carlos1*, Héctor González Hernández1, Jesús Romero Nápoles1, Laura Delia
Ortega Arenas1, Armando Equihua Martínez1, Salvador Ochoa Ascencio2
1 Fitosanidad-Entomología y Acarología, Colegio de Postgraduados, Campus Montecillo. Km 36.5 Carretera Federal México-Texcoco.
C.P. 56230. Montecillo, Texcoco, Edo. De México.2 Facultad de Agrobiología, Universidad Michoacana de San Nicolás de Hidalgo. Paseo
Lázaro Cárdenas Esquina Berlín s/n, Colonia Viveros, Uruapan, Michoacán, México. *Autor para correspondencia:
INTRODUCCIÓN
El estudio de las especies de escamas armadas sobre aguacate en México es limitado,
principalmente porque son consideradas como plagas secundarias en el cultivo, esto se debe
a que sus daños no son considerados de importancia económica, salvo cuando sus
poblaciones aumentan y llegan a estar presentes en altas densidades en frutos restándoles
calidad estética, e incluso, por su alimentación en ramas y hojas, también pueden ocasionar
la muerte de plantas jóvenes o adultas estresadas (Lázaro-Castellanos et al., 2012). En otros
casos, la identificación de nuevas especies de estos insectos puede tener implicaciones de
tipo cuarentenario para la comercialización de frutos de aguacate en el mercado internacional
(Evans et al., 2009; Morse et al., 2009). El manejo de esta familia de insectos escama suele
hacerse con la aplicación de insecticidas que resultan ineficientes por la presencia de la
cubierta cerosa que poseen estos insectos sobre sus cuerpos; sin embargo, suelen ser
altamente susceptibles al ataque de enemigos naturales, principalmente de avispas
parasíticas; las cuales, en el aguacatero, pueden ser afectadas por las aplicaciones de
insecticidas dirigidas a otras plagas (Rosen, 1973; Lázaro-Castellanos et al., 2012). El
conocimiento de los enemigos naturales también es escaso en nuestro país. Los estudios
realizados por Lázaro-Castellanos et al. (2012) en Michoacán y Lázaro-Castellanos et al.
(2018) en el Estado de México, Morelos y Puebla, han registrado en total a 18 especies de

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
84
avispas parasíticas de las familias Aphelinidae, Encyrtidae y Signiphoridae, además de un
hiperparasitoide del género Marietta y un Coccinellidae depredador. Por lo anterior, el
presente trabajo tiene como objetivo identificar las especies de escamas armadas y sus
enemigos naturales en huertos de aguacate comerciales y de traspatio en los estados de
Colima, Jalisco, Nayarit y Querétaro.
MATERIALES Y MÉTODOS
Se realizaron colectas de ramitas, hojas y frutos de aguacate infestados con escamas
armadas en huertos comerciales y de traspatio en los municipios de Comala, Cuauhtemoc
(Colima), Concepción de Buenos Aires, San Gabriel, Sayula, Tolimán, Zapotlán el Grande,
Zapotiltic (Jalisco), Tepic, Xalisco (Nayarit), Cadereyta, Peñamiller y Tolimán (Querétaro).
En el laboratorio se revisaron las muestras con microscopio estereoscópico y se obtuvieron
ejemplares de escamas armadas adultas para su posterior montaje, las escamas parasitadas se
colocaron en cápsulas de gelatina transparente para la obtención de parasitoides adultos,
también una parte del material vegetal no revisado se colocó en cajas Petri para la emergencia
de parasitoides a temperatura de 20±4 °C y HR de 60±10%; todos los especímenes obtenidos
se conservaron en alcohol al 80%. El montaje de escamas armadas se realizó con la técnica
de Kostarab (1963) y para parasitoides se utilizó la de Noyes con ligeras modificaciones de
Myartseva (Lázaro-Castellanos et al., 2012) para cada montaje en laminillas con bálsamo de
Canadá. Para la identificación de escamas armadas se utilizaron las claves de Evans et al.
(2009); para los enemigos naturales, las claves de Myartseva y Ruiz-Cancino (2001) para
Marietta; Myartseva y Evans (2008) para Encarsia; Myartseva et al. (2010) para Aphytis;
Ramírez-Ahuja et al. (2015) para Signiphoridae; y la identificación de Encyrtidae fue
realizada por la Dra. Beatriz Rodríguez Vélez del Centro Nacional de Referencia de Control
biológico (DGSV-SENASICA-SADER).
RESULTADOS Y DISCUSIÓN
Se obtuvieron un total de 35 muestras en 33 huertos, se identificaron a 10 especies de
escamas armadas y 21 especies de parasitoides pertenecientes a las familias Aphelinidae (13),
Encyrtidae (5) y Signiphoridae (3) (Cuadro 1). Las especies de escamas armadas

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
85
identificadas fueron: Acutaspis albopicta (Cockerell), A. scutiformis (Cockerell),
Chrysomphalus dictyospermi (Morgan), Diaspis c.a. coccois (Lichtenstein), Hemiberlesia
cyanophylli (Signoret), H. diffinis (Newstead), H. lataniae (Signoret), Howardia biclavis
(Comstock), Melanaspis deklei (Deitz & Davidson) y Pseudoparlatoria parlatorioides
(Comstock). De los parasitoides, las especie identificadas fueron: Aphytis diaspidis
(Howard), A. holoxanthus (DeBach), A. lepidosaphes (Compere), A. lingnanesis (Compere),
A. melinus (DeBach), A. proclia (Walker), Encarsia aurantii (Howard), E. citrina (Craw), E.
juanae (Myartseva & Evans), E. perniciosi (Tower), E. subelongata (Myartseva & Evans),
E. tarsalis (Myartseva), Marietta montana (Myartseva & Ruiz-Cancino), Archinus sp.,
Metaphycus sp. 2, Metaphycus sp. 4, Plagiomerus diaspidis (Crawford), Pseudhomalopoda
prima (Girault), Chartocerus sp., Signiphora flavella (Girault) y S. perpauca (Girault).
Cuadro 1. Escamas armadas, abundancia, distribución y sus enemigos naturales en cuatro
estados de la región Centro Occidente de México.
Escama armada Estados (# huertos) Parasitoides
A. albopicta Jalisco (4) Aphytis holoxanthus, A. lepidosaphes, A. lingnanensis, A. melinus, S. flavella, Ps. prima.
A. scutiformis Colima (3), Nayarit (1) A. holoxanthus.
Chry. dictyospermi Colima (2), Nayarit (6) A. holoxanthus, A. melinus, E. juanae, E. perniciosi.
D. c.a. coccois Jalisco (1), Querétaro (2) A. holoxanthus, A. lepidosaphes, S. flavella.
H. cyanophylli Colima (1), Jalisco (1), Nayarit (2),
Querétaro (2)
A. holoxanthus, A. melinus, Metaphycus sp. 4.
H. diffinis Colima (3), Jalisco (5), Nayarit (7),
Querétaro (3)
A. holoxanthus, A. lepidosaphes, A. lingnanensis, A.
melinus, Archinus sp., Chartocerus sp., E. juanae, E.
subelongata, E. tarsalis, Metaphycus sp. 2, Metaphycus
sp. 4, P. diaspidis, S. flavella, S. perpauca.
H. lataniae Colima (2), Jalisco (10), Nayarit
(7), Querétaro (5)
A. diaspidis, A. holoxanthus, A. lepidosaphes, A.
lingnanensis, A. melinus, A. proclia, E. aurantii, E. citrina,
E. juanae, M. montana, Metaphycus sp. 4, S. flavella, S.
perpauca.
Ho. biclavis Colima (1), Nayarit (3) A. holoxanthus, S. perpauca.
M. deklei Nayarit (3)
Pseu.
parlatorioides
Nayarit (2), Querétaro (1) A. lepidosaphes, A. lingnanensis, A. melinus.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
86
De las especies de escamas armadas, H. lataniae y H. diffinis fueron las especies con
mayor abundancia y distribución en los cuatro estados muestreados ya que se encontraron en
24 y 18 huertos, respectivamente; mientras que Chr. dictyospermi estuvo presente en ocho
huertos de dos estados, en el caso de H. cyanophylli, esta escama estuvo presente en seis
huertos, pero distribuido en los cuatro estados. Las cuatro escamas mencionadas tienen un
amplio rango de hospedantes que incluyen a aguacate, en lo que se refiere a su distribución,
con excepción de H. diffinis (que sólo se ha encontrado en países del continente americano),
se consideran que son especies cosmopolitas (García-Morales et al., 2016), lo que explica su
alta presencia en los huertos de aguacate. De las especies restantes de escamas armadas, M.
deklei se registra por primera vez sobre aguacate en México, mientras que las otras cinco ya
han sido reportadas en algún momento asociadas al cultivo en Michoacán u otros estados
(González et al., 2008; Morse et al., 2009; Lázaro-Castellanos et al., 2018). De los
parasitoides, todas las especies de Aphytis presentan nuevos registros de escamas
hospedantes y distribución para el país; en Encarsia, con excepción de E. citrina, todas las
especies presentan nuevos registros de escamas hospedantes y distribución para México
(Myartseva y Evans, 2008; Lázaro-Castellanos et al., 2012; Lázaro-Castellanos et al., 2018);
en Signiphora, ambas especies presentan también nuevos registros de distribución y escamas
hospedantes para el país (Ramírez-Ahuja et al., 2015; Lázaro-Castellanos et al., 2018); en
Encyrtidae, P. diaspidis presenta un nuevo registro de distribución, mientras que Ps. prima
se registra por primera vez en el cultivo de aguacate atacando un nuevo hospedante, además
de una nueva distribución para México (Trjapitzin et al., 2008). Las especies de parasitoides
S. flavella fue la especie con mayor abundancia y distribución, ya que se obtuvieron 78
individuos distribuidos en los cuatro estados; le siguieron E. juanae con 54 individuos
distribuidos en dos estados (Jalisco y Nayarit); A. holoxanthus con 42 individuos distribuidos
en tres estados (Jalisco, Nayarit y Querétaro); y A. lepidosaphes con 41 individuos
distribuidos en los mismos estados que la especie anterior.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
87
AGRADECIMIENTOS
Al CONACyT por la beca otorgada al primer autor en sus estudios de doctorado, al
Colegio de Postgraduados por los recursos económicos aportados para la realización del
presente estudio, a los Comités Estatales de Sanidad Vegetal de los estados de Colima, Jalisco
y Querétaro, al Dr. Agustín Robles Bermudes de la Universidad Autónoma de Nayarit y al
Centro Nacional de Referencia de Control Biológico, así como a los productores de aguacate
en los cuatro estados visitados, por las facilidades brindadas en los muestreos a los huertos
de aguacate.
LITERATURA CITADA
Evans, G. A., G. W. Watson, and D. R. Miller. 2009. A new species of armored scale (Hemiptera: Coccoidea: Diaspididae) found on avocado fruit from Mexico and a key to the species of armored scales found on avocado worldwide. Zootaxa, 1991: 57-68.
García-Morales, M., B. D. Denno, D. R. Miller, G. L. Miller, Y. Ben-Dov, & N. B. Hardy. 2016. ScaleNet: A literature-based model of scale insect biology and systematics. Database. doi: 10.1093/database/bav118. http://scalenet.info
González H., H., L. D. Ortega A., N. Villegas J., M. F. López D. y P. Rojas R. 2008. Escamas armadas (Hemiptera: Diaspididae) del aguacate Persea americana var. Hass en huertos certificados para exportación del estado de Michoacán. Sin publicar.
Kostarab, M. 1963. The armored scale insect on Ohio, (Homoptera: Coccoidea: Diaspididae). Bulletin of the Ohio Biology Survey. Ohio State, 2, 120 p.
Lázaro-Castellanos, C., H. González.Hernández, J. R. Lomelí-Flores, S. N. Myartseva, L. D. Ortega-Arenas y S. Ochoa-Ascencio. 2012. Enemigos naturales de escamas armadas (Hemiptera: Diaspididae) en aguacate Hass en Michoacán, México. Revista Colombiana de Entomología, 38(1): 6-13.
Lázaro-Castellanos, C., H. González-Hernández, J. Romero-Nápoles, L. D. Ortega-Arenas, A. Equihua-Martínez y S. Ochoa-Ascencio, S. 2018. Diversidad de especies de escamas armadas (Diaspididae) y sus parasitoides en aguacate (Persea americana Mill.) en tres estados del centro de México. Memoria Avances de Investigación de Fitosanidad 2018.
Morse, J. G., P. F. Rugman-Jones, G. W. Watson, L. J. Robinson, J. L. Bi, and R. Stouthamer. 2009. High levels of exotic armored scales on imported avocado raise concerns regarding USDA-APHIS’ Phytosanitary Risk Assessment. J. Econ. Entomol., 102(3): 855-867.
Myartseva, S. N. and G. Evans. 2008. Genus Encarsia Foster of Mexico (Hymenoptera: Chalcidoidea: Aphelinidae) A revisión, key and description of new species. Primera edición. Departamento de Fomento Editorial, UAT. Ciudad Victoria, Tamaulipas, México. 320 p.
Myartseva, S. N. y E. Ruiz-Cancino. 2001. Mexican species of parasitoid wasps of the genus Marietta (Hymenoptera: Aphelinidae). Florida Entomologist, 84(2): 293-297.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
88
Myartseva, S. N., E. Ruíz-Cancino, J. M. Coronado-Blanco. 2010. El género Aphytis Howard (Hymenoptera: Chalcidoidea: Aphelinidae) en México, clave de especies y descripción de una especie nueva. Dugesiana, 17(1): 81-94.
Ramirez-Ahuja, M. L., A. Dal Molin, A. González-Hernández, y J. B. Woolley. 2015. Sinopsis y clave para la identificación de las especies de Signiphora (Hymenoptera: Sigiphoridae) de México, con notas sobre biología y distribución. Revista Mexicana de Biodiversidad, 86: 337-347.
Rosen, D. 1973. Methodology for biological control of armored scale insects. Phytoparasitica, 1(1): 47-54.
Trjapitzin, V. A., S. N. Myartseva, E. Ruíz-Cancino y J. M. Coronado-Blanco. 2008. Clave de Géneros de Encyrtidae (Hymenoptera: Chalcidoidea) de México y un catálogo de las especies. Serie Avispas parasíticas de plagas y otros insectos. Primera edición. Departamento de Fomento Editorial, UAT. Ciudad Victoria, Tamaulipas, México. 264 p.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
89
ESCAMAS (HEMIPTERA: COCCOIDEA) ASOCIADAS AL
CULTIVO DE YACA (Artocarpus heterophyllus Lam.) EN NAYARIT
Saúl Arias-Corpuz1, Jesús Romero-Nápoles1*, Luis Martín Hernández-Fuentes2, Héctor González-
Hernández1, Carlos Patricio Illescas-Riquelme3
1 Posgrado en Fitosanidad-Entomología y Acarología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230
Texcoco, Estado de México, México. 2 Campo Experimental Santiago Ixcuintla-INIFAP. Carretera Internacional México-Nogales Km 6,
Santiago Ixcuintla, CP. 63300, Nayarit, México. 3 Centro de Investigación en Química Aplicada. Blvd. Enrique Reyna Hermosillo No.140,
CP: 25294, Saltillo, Coahuila, México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
En México, el cultivo de Yaca cuenta con una superficie sembrada de 1,275 ha, con una
producción anual de 18,610 ton; los principales estados productores son Nayarit, Colima,
Veracruz, Jalisco, Sinaloa, Hidalgo y Michoacán. Particularmente, Nayarit cuenta con más
del 90% de la producción nacional con 17,053 ton distribuidas en 1,150 ha (SIAP 2016). Este
frutal es considerado como un cultivo rentable debido a que la mayoría de su producción es
destinada a exportación, principalmente a Estados Unidos de América (Luna et al, 2013). Sin
embargo, este cultivo al igual que otros presentan problemas fitosanitarios que limitan su
producción, entre estos destacan las plagas insectiles tales como escamas, cochinillas y
pulgones, las cuales afectan la calidad y cantidad de producción (Nova y Arrambide, 1994;
Crane y Balerdi, 2000).
Se reportan varias especies de escamas (Hemiptera: Coccoidea) que atacan a hojas y
frutos, como Pinnaspis strachani, Aspidiotus destructor, Protopulvinaria mangiferae,
Protopulvinaria pyriformis en Florida (Crane y Balerdi 2000) y Ferrisia virgata,
Nipaecoccus viridis, Pseudococcus corymbatus en las Islas del Pacífico (Elevitch y Manner
2006). Particularmente, para México se registran tres especies de cocoideos, Coccus viridis,
Aulacaspis tubercularis y Maconellicoccus hirsutus, las cuales podrían afectar el desarrollo
y producción del cultivo (Rodríguez-Palomera et al, 2017). Debido a que no existe un manejo
adecuado de las plagas, por la falta de información de las especies de importancia económica
que afectan a dicho cultivo, el presente estudio tuvo como objetivo identificar las especies de
cocoideos asociados al cultivo de yaca en el estado de Nayarit.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
90
MATERIALES Y MÉTODOS
La recolecta de insectos escama (Hemiptera: Coccoidea) asociados al cultivo de yaca, se
realizó cada 15 a 21 días en huertos de las principales zonas productoras de Nayarit. Se
eligieron huertos a diferente grado altitudinal entre los 2 metros sobre el nivel del mar hasta
el punto más alto donde se cultive la yaca (Cuadro 1).
Se realizaron muestreos directos sobre el follaje, frutos y ramas para recolectar los insectos
escama. De las muestras recolectadas, una parte se conservó en tubos (Eppendorf) con
capacidad de 1.5 ml y con alcohol etílico al 96%; otra parte de las muestras consistió en tejido
vegetal infestado depositados en bolsas (Ziploc). Tanto las muestras en alcohol como las del
tejido vegetal fueron trasladadas al laboratorio. Se depositaron las muestras infestadas en
cajas Petri con el objetivo continuar con su desarrollo y obtención de los insectos adultos o
emergencia de parasitoides de los insectos escama.
Se procesaron las muestras en alcohol para la identificación taxonómica, de acuerdo con
la técnica propuesta por Koztarab (1963) para insectos escama, la cual consiste en macerar
en KOH 10% todos los tejidos en un termoblok (Falc) a una temperatura de 80ºC por 10-15
min y dejar solo el exoesqueleto del insecto escama. Bajo el microscopio estereoscópico se
presionó al ejemplar con la ayuda de una aguja para sacar los tejidos macerados. El material
ya transparente se pasó a alcohol ácido acético por 10 min; después se hizo una tinción con
fucsina ácida por 10 min para resaltar las estructuras esclerosadas; posteriormente se
deshidrató el ejemplar pasándolo por una serie de alcoholes en porcentajes crecientes (80, 96
y 100%), por 10 min en cada paso; finalmente se aclararon los ejemplares en aceite de clavo
mínimo 20 min, para después montarlos permanentemente en bálsamo de Canadá en porta y
cubre objetos. Para la determinación específica de los ejemplares por taxonomía morfológica,
se usarán claves especializadas (Ferris, 1942; Hamon & Willimas, 1984; Willimas & Granara
de Willink, 1992; Rámos-Portilla y Caballero, 2017).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
91
Cuadro 1.Huertos considerados para la recolecta de insectos escama en los estados de Nayarit y Colima, 2018.
FECHA DE COLECTA LOCALIDAD ALTITU
D (MSNM) COORDENADAS
05-JUN-18 Las Varas, Compostela, Nay. 34 21.071159 N; -105.102304 O
07-JUN-18 Las Varas, Compostela, Nay. 34 21.071159 N; -105.102304 O
25-JUL-18 Jalcocotán, San Blas, Nay. 545 21.2939 N; 105.459 O
25-JUL-18 Tecuitata, San Blas, Nay. 243 21.2628 N; 105.858 O
25-JUL-18 Tecuitata, San Blas, Nay. 243 21.2628 N; 105.858 O
25-JUL-18 El Llano, San Blas, Nay. 30 21.2516 N; 105.1049 O
25-JUL-18 Las Varas, Compostela, Nay. 34 21.1024 N; 105.1027 W
27-JUL-18 Santa Rita, Manzanillo, Col. 2 18.5942 N; 104.436 W
27-JUL-18 Ej. Venustiano Carranza, Manzanillo, Col. 26 19.0126 N; 104.549 W
09-OCT-18 Altavista, Compostela, Nay. 103 21.061314 N; 105.103529 O
10-OCT-18 El Ahuehuete, Tepic, Nay. 860 21.3110 N; 104.5614 O
RESULTADOS Y DISCUSIÓN
Se identificaron cuatro familias y cinco especies de insectos escama: escamas suaves
Coccidae (2), escama armada Diaspididae (1), insectos perla Monophlebidae (1) y piojos
harinosos Pseudococcidae (1) (Cuadro 2).
De estas especies, la escama de nieve Pinnaspis strachani (Cooley) (Disapididae),
presentó la mayor distribución; es polífaga, de distribución cosmopolita, presente en 114
países y se alimenta aproximadamente de 244 géneros de 74 familias botánicas (García-
Morales, 2016). En México se ha detectado a P. strachani en cítricos, aunque no como una
plaga importante (González y Atkinson, 1984). En la India, se ha reportado a esta escama
como plaga importante de la yaca (Rajkumar et al., 2018).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
92
Cuadro 2. Insectos escama asociados a huertos de yaca en Nayarit y Colima, 2018.
FAMILIA ESPECIE DE ESCAMA
ESTRUCTURA ATACADA
LOCALIDAD ALTITUD
(MSNM)
DIASPIDIDAE Pinnaspis strachani (Cooley)
Frutos Las Varas, Compostela, Nay.
34
DIASPIDIDAE Pinnaspis strachani (Cooley)
Frutos Las Varas, Compostela, Nay.
34
DIASPIDIDAE Pinnaspis strachani (Cooley)
Frutos Jalcocotán, San Blas, Nay.
545
DIASPIDIDAE Pinnaspis strachani (Cooley)
Tronco, ramas y follaje Tecuitata, San Blas, Nay.
243
COCCIDAE Milviscutulus manguiferae (Green)
Hojas Tecuitata, San Blas, Nay.
243
DIASPIDIDAE Pinnaspis strachani (Cooley)
Frutos El Llano, San Blas, Nay.
30
Ramas El Llano, San Blas, Nay.
30
DIASPIDIDAE Pinnaspis strachani (Cooley)
Frutos El Llano, San Blas, Nay.
30
DIASPIDIDAE Pinnaspis strachani (Cooley)
Tallo Las Varas, Compostela, Nay.
34
DIASPIDIDAE Pinnaspis strachani (Cooley)
Frutos Las Varas, Compostela, Nay.
34
MONOPHLEBIDAE
Icerya sp. Ramas Santa Rita, Manzanillo, Col.
2
PSEUDOCOCCIDAE
Pseudococcus calceolariae Maskell
Ramas Ej. Venustiano Carranza, Manzanillo, Col.
26
COCCIDAE Coccus nr. pseudomagnoliarum
Ramas El Ahuehuete, Tepic, Nay.
860
COCCIDAE Milviscutulus manguiferae (Green)
Hoja Altavista, Compostela, Nay.
103
Las escamas Milviscutulus manguiferae (Green) y Coccus nr. pseudomagnoliarum
(Kuwana), sólo se han observado atacando en forma esporádica a follaje y ramas de los
árboles de yaca.
Por otro lado, la escama acanalada Icerya sp. 1. (Monophlebidae) detectada en el huerto
de Santa Rita en Manzanillo, Colima, posiblemente sea un nuevo reporte de la especie para
México y para el cultivo, por lo cual, se enviará para su revisión al especialista de insectos
escamas el Dr. Kondo Takumasa de la Corporación Colombiana de Investigación
Agropecuaria (CARPOICA), en Palmira, Valle, Colombia. Por su parte, el piojo harinoso

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
93
Pseudococcus calceolariae Maskell (Pseudococcidae) se ha detectado en México en forma
aislada en naranjo y en obelisco Hibiscus rosa-sinensis (Malvaceae) (Arriola-Padilla, 2009).
De manera particular, India reporta varias especies de insectos escama, tales como la
escama Drosicha manguiferae (Monophlebidae); la escama armada Aspidiotus destructor
(Disapididae); las escamas suaves Milviscutulus manguiferae y Ceroplastes rubens
(Coccidae) así como a los piojos harinosos Nipaecoccus viridis y Planococcus lilacinus
(Pseudococcidae), de distribución cosmopolita y especies polífagas (Rajkumar et al., 2018).
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca otorgada. Al
Colegio de Postgraduados, Campus Montecillo, por el equipo y las instalaciones donde se
realizó la investigación y la asesoría de sus Profesores.
LITERATURA CITADA
Arriola-Padilla, V.J. 2009. La familia Pseudococcidae (Hemiptera: Coccoidea) en México. Tesis Doctor en Ciencias. Colegio de Postgraduados. Campus Montecillo, Texcoco, Edo. de México. 228 p.
Crane J.H. y Balerdi C.F. 2000. La Jaca (Artocarpus heterophyllus Lam.) en Florida. Consultada el 30 de octubre de 2018. Disponible en: http://hammock.ifas.ufl.edu
Elevitch C.R., and Manner H.I. 2006. Artocarpus heterophyllus (jackfruit). 17 p. In: Elevitch C.R. (ed.). Species profiles for pacific island agroforestry. Permanent Agriculture Resources (PAR), Holualoa Hawai. Consultada el 30 de octubre de 2017. Disponible en: https://retirenicaragua.files.wordpress.com/2012/05/a-heterophyllus-jackfruit1.pdf
Ferris, G.F. 1942. Atlas of the Scale Insects of North America. Series IV. The Diaspididae. Stanford University Press, Stanford, Ca. p: 385-448.
García Morales M, Denno BD, Miller DR, Miller GL, Ben-Dov Y and Hardy, NB. 2016. ScaleNet: A literature-based model of scale insect biology and systematics. Database. In: http://scalenet.info. doi: 10.1093/database/bav118.
González, H.H. y T.H. Atkinson. 1984. Coccoideos (Hom: Coccoidea) asociados a árboles frutales de la región Central de México. Agrociencia 54: 207-225.
Hamon, A.B., and M.L. Willimas. Th escale insects of Florida (Homoptera: Coccoidea). Florida Department of Agiculture &v Cosctumer Services. Div. Plant Ind. Vol 11.
Kosztarab M. 1963. The armored scale insect of Ohio (Homoptera: Coccoidea: Diaspididae). Bulletin of the Ohio Biological Survey 2: 1-120.
Luna-Esquivel E. G., Alejo-Santiago G., Ramírez-Guerrero L.G., Arévalo-Galarza M.L. 2013. La yaca (Artocarpus heterophyllus Lam.) un fruto de exportación. Agro Productividad, 6(5): 65-70. Rajkumar, M.B, B. Gundappa, M.M. Tripathi, and S. Rajan.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
94
2018. Pest of Jackfruit, chapter 18. Pp: 587-602. In: Pest and Their Management. Omkar (Editor). Springer Nature Singapore Pte Ltd. ISBN 978-981-10-8687-8 (eBook).
Ramos-Portilla, A.A., and A. Caballero. 2017. Diaspididae on Citrus spp. (Rutaceae) from Colombia: New records and a taxonomic key to their identification. Rev. Fac. Nac. Agron. 70(2): 8139-8154.
Rodríguez-Palomera M., Cambero-Campos J., Luna-Esquivel G., Robles-Bermúdez A., Cambero-Nava K.G. 2017. Entomofauna asociada al cultivo de yaca (Artocarpus heterophyllus lam.) en Nayarit, México. Entomología mexicana, 4: 220−225.
Williams D.J., y M.C. Granara de Willink. 1992. Mealybugs of Central and South America. C. A. B. International Institute of Entomology. London. 635 p.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
95
POTENCIAL Y ESTABILIDAD DE LA RESISTENCIA DE
Diaphorina citri (HEMIPTERA: LIVIIDAE) A INSECTICIDAS
Víctor Hugo García Méndez1, Laura Delia Ortega Arenas1*, Ángel Lagunes Tejeda1, Hussein
Sánchez-Arroyo1, Juan Antonio Villanueva-Jiménez2, Joel Lara-Reyna3
1Colegio de Postgraduados Campus Montecillo, Km 36.5 Carretera Federal México-Texcoco, Texcoco, Estado de México, México. 2Colegio de Postgraduados, Campus Veracruz, Km 88.5 Carretera Federal Xalapa-Veracruz, Manlio Fabio Altamirano, Veracruz, México, 3Colegio de Postgraduados Campus Campeche, Km 17.5 Carretera Haltunchén-Edzná, Champotón, Campeche, México. *Autor de
correspondencia: [email protected]
INTRODUCCIÓN
Ante la detección del Huanglongbing (HLB) - Diaphorina citri (Hemiptera: Liviidae), en
México, se estableció la estrategia de manejo que se basa en tres aspectos fundamentales; la
detección y eliminación oportuna de árboles enfermos, el empleo de plantas producidas en
viveros certificados y el control del insecto vector en Áreas Regionales de Control (ARCOs)
(Robles-García, 2012). En este último se contempla la aplicación regional de insecticidas en
periodos cortos definidos, con base en épocas biológicamente justificadas con el objetivo de
evitar la dispersión de la bacteria Candidatus Liberibacter asiaticus (CLas) (SENASICA,
2018).
Sin embargo, está documentado que poblaciones de D. citri de huertas comerciales de
Colima, Jalisco, Michoacán, Nayarit y Veracruz son capaces de resistir el efecto letal de
diferentes insecticidas (Vázquez-García et al., 2013; García-Méndez et al., 2016), esto ha
sido un obstáculo importante para el manejo del HLB-D. citri (Coy et al., 2016) y evidencia
la necesidad de comprender el fenómeno de la resistencia de D. citri a insecticidas para
definir adecuadamente los productos a utilizar para su control (Tian et al., 2019).
Por lo anterior el presente estudio tuvo como objetivos el conocer la estabilidad que tiene
la resistencia de D. citri a clorpirifos en ausencia de presión de selección, y determinar el
potencial de la resistencia de D. citri a clorpirifos e imidacloprid bajo presión de selección.
Todo con el fin de aportar más información acerca del fenómeno, que permita retrasar lo más
posible su desarrollo en campo.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
96
MATERIALES Y MÉTODOS
Para cumplir con los objetivos se utilizaron dos colonias de D. citri. La primera,
susceptible ubicada en el Colegio de Postgraduados, Campus Montecillo, la cual se ha
mantenido durante 10 años en cría constante y sin exposición a plaguicidas. La segunda fue
una población que mostró ser, en estudios previos, 80 veces resistente a clorpirifos, originaria
de una huerta comercial de lima persa de Martínez de la Torre, Veracruz. Ambas colonias se
mantuvieron separadas y en condiciones controladas de invernadero (25 ± 5ºC, 60±5 % de
humedad relativa y fotoperiodo natural).
La colonia de Martínez de la Torre, se subdividió en tres para realizar diferentes
experimentos. En cada generación se determinó la dosis letal mediana (CL50) de clorpirifos
(Tridente, 94% de pureza) e imidacloprid (Bayer, 98% de pureza), así como la dosis letal
80% (CL80) de cada uno. Para su estimación se realizaron bioensayos con una torre de
aspersión Burgerjon con el método de Colín et al. (1994) con ligeras modificaciones.
Dos colonias se expusieron, de forma independiente, cada dos generaciones con la CL80
de cada insecticida, estimada previamente; después, la CL50 de la generación tratada se
dividió entre la estimada en la generación anterior y con el radio de resistencia (RR)
resultante se determinó el cambio de la resistencia al producto en el experimento de potencial.
Una tercera colonia se mantuvo sin presión de selección y también con el RR, entre una
generación y la anterior, se estableció el cambio de la resistencia en el experimento de
estabilidad. El mismo experimento de potencial se realizó en dos colonias, de la población
susceptible, con clorpirifos e imidacloprid, respectivamente.
RESULTADOS Y DISCUSIÓN
Los resultados mostraron que la población de Martínez de la Torre tuvo potencial positivo
de desarrollo de la resistencia por la presión de selección con los insecticidas utilizados. El
RR para clorpirifos aumentó de 29.6X a 89.5X; mientras que el RR para imidacloprid
aumentó de 1.42X a 19.42X. Por otro lado, en la población susceptible lo mismo se observó
para imidacloprid, que alcanzó el mismo potencial de resistencia (RR= 18X) en cinco

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
97
generaciones, no así para clorpirifos debido a que el desarrollo en la colonia fue menor, con
un máximo de 4.5X.
El potencial de resistencia a clorpirifos en la población de Martínez de la Torre puede
relacionarse con el historial de aplicación en la zona, que ha propiciado la selección de
fenotipos con enzimas capaces de soportar dosis altas a dicho insecticida (Tiwari et al., 2012)
y que han sido reportadas en estudios previos con estudios de susceptibilidad en la zona
(García-Méndez et al., 2016). Lo cual indica que, bajo presión de selección, la población
puede mantener un potencial de resistencia en aumento positivo.
Lo mismo sucedió para imidacloprid, pero a niveles más bajos, debido a que aún no se
encontraban presentes los mecanismos metabólicos para soportar el efecto tóxico del
neonicotinoide (Tian et al., 2019), es decir, se encontraba en un estado susceptible, motivo
por el cual los valores de potencial son muy similares a los mostrados en población
susceptible. Sin embargo, no se debe desestimar la capacidad de D. citri para sobrevivir a
imidacloprid pues en una población de invernadero mostró la capacidad de ser 4265 veces
resistente a este producto (Vázquez-García et al., 2013).
La disminución de la resistencia a clorpirifos observada en la colonia de Martínez de la
Torre mostró una clara inestabilidad que ha fluctuado desde los 55.5X hasta los 18.8X. Dicha
inestabilidad no ha sido reportada para D. citri y puede tener impacto en las prácticas de
manejo de esta plaga debido a que los resultados indican que en ausencia de presión de
selección con este organofosforado es posible retornar a un estado de susceptibilidad.
Los resultados mostraron que el desarrollo de la resistencia de D. citri a clorpirifos e
imidacloprid fue lento e inestable en un principio en una población con genes de
susceptibilidad, pero en una población con fenotipos resistentes y con presión de selección
constante con clorpirifos la resistencia tuvo una tendencia a aumentar, lo cual se pudo
contrarrestar de forma rápida una vez que hubo ausencia del tóxico en la población. Esta
información puede ser de utilidad en el establecimiento de una estrategia de manejo de
insecticidas para el combate de |D. citri en el programa ARCOs.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
98
AGRADECIMIENTOS
Al CONACyT por la beca otorgada al estudiante y a los proyectos que financiaron la
investigación, así como al Colegio de Postgraduados.
LITERATURA CITADA
Coy, M. R., L. Bin and L. L. Stelinski. 2016. Reversal of insecticide resistance in Florida populations of Diaphorina citri (Hemiptera: Liviidae). Fla Entomol 99:26-32.
Colin, M.E., F. Ciavarella, G. Otero-Colina, and L. P. Belzunces. 1994. A method for characterizing the biological activity of essential oils against Varroa jacobsoni. In New perspectives on Varroa. Ed. A. Matheson IBRA, Cardiff, U. K.:109-114.
García-Méndez, V. H., L. D. Ortega-Arenas, J. A. Villanueva-Jiménez and H. Sánchez-Arroyo. 2016. Susceptibilidad de Diaphorina citri Kuwayama (Hemiptera: Liviidae) a insecticidas en Veracruz, México. Agrociencia 50:355-365.
Robles-García, P. 2012. Protocolo para establecer Áreas Regionales de Control del Huanglongbing y el Psílido Asiático de los Cítricos (ARCOs). mx/?doc=9364 [Consulta: 8 de julio de 2018].
Servicio Nacional de Sanidad Inocuidad y Calidad Agroalimentaria (SENASICA). 2018 https://www.gob.mx/cms/uploads/attachment/file/302344/Estrategia_HLB_2018_ver15Feb018_ConAnexos.pdf [Consultado: 1 abril 2018].
Tian, F., C. Li, Z. Wang, J. Liu J and X. Zeng. 2019. Identification of detoxification genes in imidacloprid-resistant Asian citrus psyllid (Hemiptera: Liviidae) and their expression patterns under stress of eight insecticides. Pest Manag Sci 75: 1400-1410.
Tiwari, S., L. L. Stelinski and M. E. Rogers. 2012. Biochemical Basis of Organophosphate and Carbamate Resistance in Asian Citrus Psyllid. J Econ Entomol 105: 540-548.
Vázquez-García, M., J. Velázquez-Monreal, V. M. Medina-Urrutia, C. D. J. Cruz-Vargas, M. Sandoval-Salazar, G. Virgen-Calleros, and J. P. Torres Morán. 2013. Insecticide resistance in adults Diaphorina citri Kuwayama from lime orchards in Central West Mexico. Southwest Entomol. 38: 579–596.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
99
CICLO DE VIDA Y PARÁMETROS BIOLÓGICOS DE B.
californicus Y B. yothersi (ACARI: TENUIPALPIDAE) EN NARANJA
Citrus sinensis
Castro-Resendiz Carmen Asunción1, Otero Colina Gabriel1, Quijano Carranza Juan Ángel2,
Martínez Meyer Enrique3, González Hernández Héctor1
1 Postgrado en Fitosanidad - Entomología y Acarología. Colegio de Postgraduados (CP). Carretera México-Texcoco km 36.5, estado de
México, México. C. P. 56230; [email protected] 2 Campo Experimental Bajío-Instituto Nacional de Investigaciones
Forestales,Agrícolas y Pecuarias (INIFAP). Celaya Guanajuato, México. Apdo. Postal 112. C.P. 38000, 3 Departamento de Zoología,
Instituto de Biología, Universidad Nacional Autónoma de México (UNAM). Ciudad de México, México. Apdo. Postal 70-153. C.P. 04510.
INTRODUCCIÓN
Kitajima et al. (2003) observaron que la leprosis de tipo citoplásmico (CiLV-C) prevalece
preferentemente en sitios de baja altitud y de clima cálido, mientras que la leprosis de tipo
nuclear (CiLV-N y OFV-citrus) prevalece en sitios de mayor altitud y de clima templado. De
manera similar, Roy et al. (2015) observaron que B. yothersi habita en México sobre todo en
sitios bajos de clima cálido; esto es, a lo largo de las planicies costeras, mientras que B.
californicus habita en sitios altos, de clima templado; esto es, en el Altiplano Mexicano y en
las Sierras Madres. Lo anterior llevó a dichos autores a suponer que el principal vector del
CiLV-C es B. yothersi, y que el principal vector del OFV-citrus es B. californicus. Esta
suposición fue plenamente confirmada por García-Escamilla et al. (2017), quienes
concluyeron que existe una especificidad total entre los virus nuclear y citoplásmico y sus
respectivos vectores. De lo anterior se desprende que la distribución de los virus causantes
de leprosis citoplásmica y nuclear corresponde a la de sus respectivos vectores, los que tienen
distribución alopátrica, por lo menos en México. Dado que tanto el CiLV-C como el OFV-
citrus al parecer se encuentran en un proceso de expansión, el conocimiento de las
preferencias climáticas de sus vectores ofrece elementos para determinar su potencial de
expansión y, por consiguiente, ofrece elementos para contener su avance. En este contexto,
el objetivo de este trabajo es determinar la biología de B. californicus y B. yothersi (Acari:
Tenuipalpidae) en condiciones controladas (15 a 30°C, humedad relativa de 60 ± 5 %)
confinados en naranja (Citrus sinensis), mediante la construcción de tablas de vida.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
100
MATERIALES Y MÉTODOS
Los ácaros. En este estudio se utilizaron ácaros procedentes de las colonias uniparentales
o isolíneas (formadas a partir de una sola hembra) con algunas modificaciones del método
descrito por Franco et al. (2007) de B. californicus y B. yothersi, establecidas en el laboratorio
de Acarología del Postgrado en Entomología y Acarología, en Montecillo, municipio de
Texcoco, estado de México. Estas crías permanentes están establecidas en frutos de naranja
agria (Citrus aurantium) a 25°C y humedad relativa de 60 ± 5 %.
Se confirmó la identificación de las especies mediante análisis moleculares, microscopía
electrónica y de contraste de fases.
El hospedero. Todos los frutos sobre los cuales se dio seguimiento al desarrollo y
supervivencia de B. californicus y B. yothersi se colectaron en el predio “La fortuna” en la
localidad Novara del municipio de Martínez de la Torre, Veracruz. Longitud: O 97° 4' 48" y
Latitud: N 20° 3' 5" ubicado a una altitud de 100 m.
Diseño experimental. Se colectaron naranjas (Citrus sinensis) con edad intermedia de
cosecha (color verde del fruto), se lavaron y se les sumergió hasta la mitad en parafina
fundida para que duraran más tiempo antes de marchitarse. La parte no cubierta con parafina
de cada naranja fue dividida en cuatro partes con adhesivo atrapamoscas (TAnglefoot ®)
para impedir el paso de ácaros de una a otra de las divisiones de la naranja. Se realizó el
seguimiento del desarrollo, supervivencia y fecundidad de B. californicus y B. yothersi, para
lo cual se formaron cohortes de un mínimo de 50 ejemplares de cada especie, iniciadas con
huevos de edad conocida, un huevo en cada división de las naranjas. Las cohortes obtenidas
se incubaron a temperaturas constantes de 15.0, 20.0, 22.5, 25.0 y 30.0 ± 2°C, humedad
relativa de 65-75% y fotoperiodo de 14-10 horas (luz-oscuridad, respectivamente). Se
observó la duración de cada etapa de desarrollo, así como la supervivencia de todos los
ejemplares de cada cohorte, mediante registro cada 24 horas, hasta la muerte del último
ejemplar

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
101
RESULTADOS
La duración de los diferentes estadios para B. californicus se presenta en la figura 1A y la
información correspondiente para B. yothersi se presenta en la figura 1B. El número de
repeticiones (ejemplares) a 20.0, 22.5, 25.0 y 30.0 °C fue de 135, 74, 51, 72 respectivamente
para B. californicus y para B. yothersi a 20.0, 22.5, 25.0 y 30.0 °C fue de 130, 72, 56 y 72
respectivamente. La temperatura de 20° C se evaluó en un mayor número de ocasiones para
ambas especies por que no se registró respuesta posterior al estadio de protoninfa.
A) B)
Figura 1. Duración en días de los diferentes estadios de desarrollo de A) B. californicus y de B) B. yothersi
en naranja (Citrus sinensis) a temperaturas constantes de 20.0, 22.5, 25.0 y 30.0 °C, humedad relativa de 60 ±
5 %, fotoperiodo de 14:10 h, luz y oscuridad, respectivamente.
De manera consistente se observó que al incremento en la temperatura de incubación
correspondió una reducción en el tiempo de desarrollo. Las gráficas de la Figura 1 muestran
un tiempo aparentemente más corto a 20°C (B. yothersi) o 22.5° (B. californicus), pero esto
más bien se debió a que un reducido número de ejemplares completaron el estadio de
desarrollo. Brevipalpus yothersi tuvo tiempos de desarrollo significativamente más cortos
que B. californicus, lo que explicaría por qué este ácaro es predominante en sitios bajos, de
clima cálido; sin embargo, no explica por qué B. californicus es predominante en sitios altos,
de clima templado. Se postula que esta última especie es más resistente a heladas, que se
presentan en los sitios altos, pero está pendiente el desarrollo de estudios para confirmar esta
hipótesis.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
102
AGRADECIMIENTOS
Agradecemos al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la
beca de maestría otorgada al primer autor. Al Servicio Nacional de Sanidad, Inocuidad y
Calidad Agroalimentaria (SENASICA) por el financiamiento de esta investigación.
Finalmente, al Colegio de Postgraduados, Campus Montecillo, por las instalaciones y la
asesoría de sus profesores.
LITERATURA CITADA
Franco A., Rebelles R., Zacarias S., Falqueto A. (2007) Predation potential of three species of phytoseiids on Oligonychus ilicis (McGREGOR, 1917) (Acari: Tetranychidae). Coffee Science. 2. 175.
García-Escamilla P., Duran-Trujillo Y., Otero-Colina G., Valdovinos-Ponce G., Santillán-Galicia M., Ortiz-García C., Velázquez-Monreal J. y Sánchez-Soto S. (2017) Transmission of viruses associated with cytoplasmic and nuclear leprosis symptoms by Brevipalpus yothersi and B. californicus. Trop. plant pathol.
Kitajima, E., W., Chagas C., M., Rodrigues J., C.V. (2003) Brevipalpus-transmitted plant virus and virus-like diseases: cytopathology and some recent cases. Experimental and Applied Acarology. 30(1-3): 135-160.Experimental and Applied Acarology 30 (1-3): 161-179.
Roy, A., Hartung, J. S., Schneider, W. L., Shao, J., León, M. G., Melzer, M. J., Beard, J. J., Otero-Colina, G., Bauchan, G. R., Ochoa, R., Brlansky, R. H. (2015) Role bending: complex relationships between viruses, hosts and vectors related to Citrus leprosis, an emerging disease. Phytopathology. 105: 1013-1025.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
103
IDENTIFICACIÓN DE PLANTAS HERBÁCEAS,
ORNAMENTALES Y MALEZAS PORTADORAS DE Citrus leprosis
virus EN VERACRUZ, MÉXICO
María de los Ángeles Cruz López1*, Ochoa Martínez Daniel Leobardo1, Cristian Nava Díaz1, Andrés
Quesada Salinas2, Jessica Berenice Valencia Luna2 y José Abel López Buenfil2
1Posgrado en Fitosanidad-Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco, Km 36.5, Montecillo, Edo. de México, CP
56230. 3Centro Nacional de Referencia Fitosanitaria-DGSV, Km 37.5, Carretera Federal México-Pachuca. Edo. de México, C.P. 55740.
*Autor de correspondencia: [email protected]
INTRODUCCIÓN
La leprosis de los cítricos es considerada como la enfermedad viral más importante en la
industria citrícola. En Brasil, cada año se invierten 80 millones de dólares para el control del
ácaro vector Brevipalpus sp. (Bastianel et al., 2010). Esta enfermedad ha incrementado su
importancia económica en América debido a su rápida dispersión por varios países del Centro
y Sur, causando pérdidas económicas considerables durante 15 años en Argentina, Uruguay,
Venezuela, Costa Rica, Panamá, Honduras, Guatemala y recientes en Perú, Bolivia y
Colombia. Los últimos países donde se ha reportado el virus son México y Belice (CABI,
2016). En México esta enfermedad representa una amenaza para la citricultura nacional, la
cual actualmente se distribuye en 28 estados, con una superficie sembrada de 563 679
hectáreas, producción superior a los 7.6 millones de toneladas y un valor de producción de
22 490 millones de pesos para el año 2017 (SIAP, 2017).
El virus CiLV-C afecta varias especies de cítricos, principalmente naranjos (Citrus
cinensis) y mandarinos (Citrus reticulata) (Bastianel et al., 2006; Bastianel et al., 2007). En
México se identificó como presente en varios huertos comerciales de naranja dulce Valencia
y Hamlin (C. sinensis) en Huimanguillo y Cunduacán, Tabasco (Izquierdo et al., 2011). En
Querétaro se reportaron siete especies del genero Citrus spp. (pomelo, limón, lima,
mandarina, naranja agria, lima dulce y naranja dulce) con detecciones de CiLV-N asociados
a síntomas específicos de leprosis. Hasta 2012, CiLV-N solo se había encontrado en los
estados de São Paulo, Rio Grande do Sul y Minas Gerais en Brasil y Boquete en Panamá
(Roy et al., 2015). En el año 2014, en Jalisco y Chiapas se reportó una nueva variante del

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
104
virus leprosis en naranja agria (C. aurantium), denominada como Citrus chlorotic spot virus
(CiCSV) que mostró una gran similitud con el virus Orchid Fleck Virus (OFV), dando una
homología global entre CiCSV y OFV / CiLV-N cerca del 90%. Estos virus pueden ser
considerados variantes o especies estrechamente relacionadas, todos pertenecen a la rama
dichorhavirus.
Inicialmente, las únicas plantas reconocidas como hospedantes naturales de CiLV eran
rutáceas del género Citrus (Rodrigues et al., 2003). Sin embargo, Colariccio et al. (1995)
lograron transmitir mecánicamente el virus CiLV-C en hospedantes herbáceos como
Chenopodium amaranticolor, C. quinoa (Chenopodiaceae) y Glomphrena globosa
(Amaranthaceae), respondiendo a la exposición con lesiones localizadas. Lovisolo et al.
(2000), ampliaron el registro de hospedantes alternos del virus a 13 especies, pertenecientes
a las familias Amaranthaceae, Tetragoniaceae y Chenopodiaceae. Nunes et al. (2012),
realizaron un experimento de infección de CiLV-C a través de ácaros vectores encontrando
plantas susceptibles al virus tales como Hibiscus rosa-sinensis, Malvaviscus arboreus
(Malvaceae), Grevilea robusta (Proteaceae) y Bixa Orellana (Bixaceae), plantas utilizadas
como cortavientos en huertos. En Colombia, el virus se encontró afectando en forma natural
las hojas de Swinglea gluinosa, especie ampliamente utilizada en el país como cerca viva
(León et al., 2008). Una observación serendipítica reveló que el frijol común (Phaseolus
vulgaris L.) es susceptible a la infección del ácaro por CiLV-C, revelando ser una excelente
planta indicadora, produciendo lesiones localizadas necróticas cinco días después de la
inoculación mediante ácaros (Groot et al., 2006; Garita et al., 2013).
Por lo anterior, el objetivo de la presente investigación fue identificar plantas herbáceas,
ornamentales y malezas portadoras de Citrus leprosis virus (CiLV-C2 y CiLV-N), Citrus
Citrus chlorotic spot virus y Orchid fleck virus mediante tres métodos de extracción en
muestras procedentes de los municipios de Papantla, Tihuatlan y Cazones, Veracruz.
MATERIALES Y MÉTODOS
Colecta de plantas. Se realizaron recorridos de campo en huertas abandonadas,
comerciales y de traspatio en los municipios de Papantla, Cazones de herrera y Tihuatlan

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
105
Veracruz. Se colectaron plantas con síntomas característicos en hojas y tallos, estos
consistieron en manchas circulares cloróticas pequeñas (1-2 mm), manchas con centro
necrótico y halo amarillo, manchas anilladas de color verde a café amarillentas (2-3 cm), que
en ocasiones estas manchas se tornan rugosas de apariencia corchosa de color café obscuro
con agrietamientos, conforme a los síntomas que se presentan en cítricos (Rodrigues, 2000;
Kitajima et al., 2003; Bastianiel et al., 2006, SENASICA, 2017), así como en las familias
que se han reportado como hipersensitivas al CiLV-C y CiLV-N en Brasil. Se recolectaron
20 plantas en el primer muestreo: 10 plantas de la especie Syngonium neglectum, 5 de
Vainilla planifolia y 5 Piper nigrum; en el segundo muestreo se recolectaron 55 plantas: 15
plantas de la especie Syngonium neglectum, 15 de Vainilla planifolia, 5 de Tabernaemontana
alba, 5 de Tabernaemontana amygdalfolia, 5 de Ipomoea sp., 5 de Momordica charantia y
5 de Piper nigrum; cada muestra fue de 5 plantas por especie. A tres plantas se les quitaron
hojas con los síntomas antes indicados para la detección de Citrus leprosis virus por cada
muestra de planta se obtuvieron dos repeticiones para su análisis y las dos restantes se dejaron
intactas para su identificación taxonómica con la ayuda de claves taxonómicas y comparación
de especímenes.
Las muestras fueron procesadas en los laboratorios de Virología y Malezas del Centro
Nacional de Referencia Fitosanitaria en Tecámac, México
Preparación del tejido vegetal. Se lavaron las muestras con agua destilada y se secaron.
Se cortaron las partes de avance de las lesiones características del virus y se pesó 0.1 g de
tejido vegetal colocándolas en viales para extracción.
Métodos de extracción de ARN. Se realizó la extracción de ARN por tres diferentes
métodos: Dodecil Sulfato de Sodio (SDS) y Acetato de potasio (KC2H3O2) por Dellaporta
(1983), bromuro de hexadecil trimetil amonio (CTAB) por Zamboni, et al. (2018), y
mediante el kit Promega (2012) que utiliza Tiocianato de guanidina (GTC). Todas las
extracciones se realizaron en un volumen de 50 µL de agua destilada estéril y almacenada a
-20 °C. Se evaluaron 45 muestras de hojas por cada método de extracción.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
106
Síntesis de cDNA. Se utilizó un volumen total de reacción de 20 µL, que contenía 5µL
de RNA, 2 µL de buffer PCR 10X (Invitrogen™), 2 µL de MgCl2 [(50 mM)] (Invitrogen™,),
2 µL de dNTP (10 mM), 0.25 µL de Oligo (dT)12-18 [50 µM] (Invitrogen™), 0.5 µL de
RNase out (40 U/µL) (Invitrogen™), 0.25 µL de MMLV 200 U/µL (Invitrogen™) y 8.0 µL
de agua grado PCR. Las condiciones de retrotranscripción fueron las siguientes: 42°C por 30
minutos, 99°C por 5 minutos y 5°C por 5
PCR. Se realizó la PCR de la síntesis de cDNA, utilizando iniciadores específicos para
CiLV-C (Locali et al., 2003), CiLV-N (Roy et al., 2014), CiCSV (Cruz et al., 2014) y OFV
(Ali et al, 2014). Se preparó un volumen final de 25 µL con 2.5 µL de buffer PCR 10 X,
1.25 µL de MgCl2 (50 mM) (Invitrogen™), 0.625 µL de dNTP (10 mM), 1µL de cada
iniciador (10 mM), 0.5 µL de Taq polimerasa (5 U/µL) (Invitrogen™), 3 µL de cDNA y
15.125 µL de agua grado PCR. Como control endógeno de la reacción se amplificó un
fragmento del gen 18S ribosomal con los iniciadores Forward (5’-
ACGGATCGCACGGCCTTCGTG-3’) y Reverse (5’-ACCAGACTTGCCCTCCAATGG-
3’) que amplifican un fragmento de 300 pb siguiendo el protocolo descrito por Zamboni et
al., (2008). Los productos de PCR fueron sometidos a electroforesis en gel de agarosa al 2%,
a 90 voltios durante 80 minutos para ser visualizados en un fotodocumentador.
RESULTADOS
Se realizaron dos muestreos en los municipios de Papantla, Tihuatlan y Cazones de
Herrera, Veracruz, en los meses de julio y noviembre del año 2018. Se colectaron plantas de
6 familias como Orchidaceae, Araceae, Convolvulaceae, Piperaceae, Apocynaceae y
Cucurbitaceae que presentaban síntomas sospechosos de las variantes de leprosis. Las
especies que se colectaron fueron: Syngonium neglectum, Vainilla planifolia, Piper nigrum,
Tabernaemontana alba, Ipomoea sp, Momordica charantia y Tabernaemontana
amygdalfolia, que presentaron síntomas de manchas circulares cloróticas, manchas con
centro café amarillentas, manchas necróticas de aspecto rugoso a corchosos y con halo
amarillo.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
107
RT-PCR con primers específicos de Citrus leprosis virus (CiLV-C2 y CiLV-N), Citrus
necrotic spot virus y Orchid fleck virus. Se realizó la RT-PCR para obtener el cDNA de las
extracciones de ARN por los tres métodos analizados. En la primera colecta, se analizaron
las especies Syngonium neglectum, Vainilla planifolia, y Piper nigrum con los primers
específicos de Citrus leprosis virus (CiLV-C y CiLV-N), Citrus chlorotic spot virus y Orchid
fleck virus; así mismo, se realizó la PCR para el control endógeno 18S ribosomal para
determinar la integridad de la extracción de ARN. Se corrieron en geles de electroforesis al
2% y se le dieron lectura en el Imager Gel DocTM EZ. Finalmente no se obtuvieron bandas
de amplificación para Citrus leprosis virus (CiLV-C y CiLV-N), Citrus chlorotic spot virus y
Orchid fleck virus.
En la segunda colecta se analizaron las especies S. neglectum, V. planifolia,
Tabernaemontana alba, Tabernaemontana amygdalfolia, Ipomoea sp., P. nigrum,
Momordica charantia con los primers específicos de Citrus leprosis virus (CiLV-C2 y CiLV-
N), Citrus chlorotic spot virus y Orchid fleck virus; asimismo, se realizó la PCR del gen
endógeno 18S para ver la integridad de las extracciones. En la segunda colecta se tuvieron
resultados negativos. Derivado de ello se realizó una tercera colecta para hacer pruebas
finales.
DISCUSIÓN Y CONCLUSIONES
Se determinó que las especies Syngonium neglectum, Vainilla planifolia, Piper nigrum,
Tabernaemontana alba, Ipomoea sp, Momordica charantia y Tabernaemontana
amygdalfolia, que presentaron síntomas de manchas circulares cloróticas, manchas con
centro café amarillentas, manchas necróticas de aspecto rugoso a corchosos y con halo
amarillo son negativas a Citrus leprosis virus (CiLV-C2 y CiLV-N), Citrus chlorotic spot
virus y Orchid fleck virus.
LITERATURA CITADA
Bastianel, M., Freitas-Astúa, J., Kitajima, E. W., and Machado, M. A. 2006. The citrus leprosis pathosystem. Summa Phytopathologica, 32(3): 211-220.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
108
Bastianel, M., Nicolini, F., Rodrigues, V., Medina, C., Segatti, N., Freitas-Astúa, J., Novelli, V. M., e Machado, M. A. 2007. Incidência de leprose em variedades de tangerinas e híbridos. Fi- topatologia Brasileira 32:293.
Bastianel, M., Novelli, V., Kitajima, E. W., Kubo, K. S, Bassanezi, R. B., Machado, M. A and Freitas-Astúa, J. 2010. Citrus leprosis Centennial of an unusual mite-virus pathosystem. Plant Disease 94(3): 284-292.
CABI. 2016. Crop Protection Compendium Global Module. CAB International. UK. Consultado en línea: http://www.cabi.org. Fecha de consulta abril de 2016
CABI. 2016. Crop Protection Compendium Global Module. CAB International. UK. Consultado en línea: http://www.cabi.org. Fecha de consulta septiembre de 2017.
Colariccio, A., Lovisolo, O., Chagas, C. M., Galletti, S. R, Rosseti, V e Kitajima, E. W. 1995. Mechanical transmission and ultrastructural aspects of citrus leprosis disease. Fitopatologia Brasileira (20): 208-213.
Garita LC, Tassi AD, Calegario RF, Kitajima EW, Carbonell SAM, Freitas-Astúa J (2013) Common bean (Phaseolus vulgaris L.): Experimental local lesion host for Citrus leprosis virus C (CiLV-C) and some other cytoplasmic-type Brevipalpus-transmitted viruses. Plant Disease (in press).
Groot TVM, Freitas-Astúa J, Kitajima EW (2006) Brevipalpus phoenicis transmits citrus leprosis virus, cytoplasmic type (CiLV-C) to common bean (Phaseolus vulgaris) under experimental conditions. Virus Review and Research 11 (Suppl.):67-68.
Izquierdo C, I., Zermeño D, L. F., Mendez, W., Otero-Colina, G., Freitas-Astúa, J., Locali-Fabris, E. C., De Moraes, G. J., Faier C, R., Tassi, A. D., and Kitajima, E. W. 2011. Confirmation of the presence of the Citrus leprosis virus C (CiLV-C) in Southern Mexico. Phytopathology, 105: 564-575.
Kitajima E. W., Rezede, J. A. M., and Rodrigues, J. C. V. 2003. Passion fruit green spot virus vectored by Brevipalpus phoenicis (Acari:Tenuipalpidae) on passion fruit in Brazil. Experimental and Applied Acarology, (30):225-231.
León, M. G., Becerra, C. H., Freitas-Astúa, J., Salaroli, R. B., and Kitajima, E. W. 2008. Natural infection of Swinglea glutinosa by the cytoplasmic type (CiLV-C) in Colombia. Plant Dis. 92:1364-1364.
Locali E.C., Freitas A. J., Souza A.A., Takita M.A., Astúa M. G., Antonioli R., Kitajima E.W. y Machado M.A. 2003. Development of a molecular tool for the diagnosis of leprosis, a major threat to citrus production in the Americas. Plant Disease 87:1317-1321.
Lovisolo O, Colariccio A, Masenga V. 2000. New Experimental Hosts and Further Information on Citrus Leprosis Virus. Fourteenth IOCV Conference, 2000—Other Viruses. 164-173.
Rodrigues, J. C. V. 2000. Relacao patogeno-vetor-planta no sistema leprose dos citros. Tesis Doc. USP Piracicaba, SP. 168p.
Rodrigues, J.C., E.W. Kitajima y C.M. Chagas. 2003. Citrus leprosis virus vectored by Bre-vipalpus phoenicis (Acari: Tenuipalpidae) on citrus in Brazil. Experimental and Applied Acarology 30: 161-179. Kluwer Academic Publishers 161-179 p.
Roy, A., León, M.G., Stone, A. L., Schneider, W. L., Hartung, J. S., and Brlansky, R. H. 2014. First Report of Citrus leprosis virus Nuclear Type in Sweet Orange in Colombia. Abstract. Plant Disease. 98(8) 1162.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
109
Roy, A., Stone, A. L., Shao, J., Otero-Colina, G., Wei, G., Choudhary, N., Achor, D., Levy, L., Nakhla, M, K., Hartung, J. S., Schneider, W. L., and Brlansky, R. H. 2015. Identification and molecular characterization of nuclear Citrus leprosis virus, a member of the proposed Dichorhavirus genus infecting multiple Citrus species in Mexico.
SENASICA. 2017. Leprosis de los cítricos (Citrus leprosis virus). Dirección General de Sanidad Vegetal. Programa de Vigilancia Epidemiológica Fitosanitaria. Con la colaboración del Laboratorio Nacional de Referencia Fitosanitaria (LaNREF). Ciudad de México. Ficha técnica No. 35. 24p.
SIAP (Servicio de Información Agroalimentaria y Pesquera). 2017. Anuario Estadístico de la Producción Agrícola. En línea: http://infosiap.siap.gob.mx/aagricola_siap_gb/icultivo/index.jsp. Fecha de consulta: 23 de junio de 2017.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
110
CONSUMO DEL ESTADO LARVAL DE TRES ESPECIES DE
CRISOPAS ALIMENTADAS CON Melanaphis sacchari
Gonzalo Espinosa-Vásquez1*, Héctor González-Hernández1, J. Refugio Lomeli-Flores1, Raquel
Alatorre-Rosas 1, L. Delia Ortega-Arenas 1, J. Luís Carrillo-Sánchez 1, J. Fernando Solís-Aguilar2
1 Posgrado en Fitosanidad, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco, Estado de México,
México. 2 Departamento de Parasitología Agrícola, Universidad Autónoma Chapingo. Km. 38.5 Carr. México-Texcoco, 56230 Texcoco,
Estado de México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
En México, los principales estados productores de sorgo han sido Tamaulipas,
Guanajuato, Sinaloa y Michoacán y en 2014 se sembraban alrededor de 2 millones de ha, con
una producción de más de 8 millones de ton; sin embargo, en 2018 la superficie del cultivo
disminuyó hasta llegar a 1.3 millones de ha, con una producción de 4.5 millones de toneladas
(SIAP, 2015, 2019), muy posiblemente debido a la llegada en 2013 del pulgón amarillo del
sorgo Melanaphis sacchari (Zehntner) (Hemiptera: Aphididae), el cual, llegó a ser la plaga
más importante en diversas regiones productoras de sorgo del país (SENASICA, 2015;
Rodríguez-del-Bosque y Terán, 2015). A nivel mundial, se registran al menos 47 especies de
enemigos naturales asociados con M. sacchari, entre estas se encuentran las crisopas. Los
adultos de estos insectos se alimentan regularmente de polen, néctar y mielecilla, mientras
que las larvas son depredadoras muy voraces de un amplio rango de insectos, principalmente
sobre aquellos de cuerpo blando (New, 2001; Athhan et al., 2004; Mansoor et al., 2013).
Aunque se han reportado varias especies alimentándose del M. sacchari, se desconocen
algunos aspectos de su biología, uno de estos es el consumo de los ínstares larvales de los
crisópidos, el cual es importante evaluar antes de su incorporación a un programa de manejo
integral de plagas (Salamanca-Bastidas et al., 2010; Paredes et al., 2013), por tal razón, el
objetivo del presente estudio fue comparar la capacidad de consumo del estado larval de
Chrysoperla carnea, C. comanche y C. externa, alimentadas con el pulgón amarillo del sorgo
M. sacchari en condiciones de laboratorio.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
111
MATERIALES Y MÉTODOS
Condiciones generales del experimento. Este experimento se realizó en el Laboratorio
de Control Biológico del Posgrado en Fitosanidad, Entomología y Acarología, del Colegio
de Postgraduados, Texcoco, Estado de México. El experimento se desarrolló bajo
condiciones controladas de 25 ± 2 ºC, 60 ± 10% de HR y fotoperiodo 12:12 (L:O). La arena
experimental consistió de una caja Petri de plástico de 4.0 cm de diámetro y 1.5 cm de altura,
con la tapa perforada (Ø= 2.0 cm) y cubierta con tela organza para permitir la ventilación.
En la base de la caja Petri se vertieron 2.0 mL de agar-agua y sobre esta capa se colocó un
disco de hoja de sorgo del híbrido UPM-219. Las arenas se remplazaron cada tercer día hasta
culminar el experimento.
Cría de insectos. Las ninfas de tercer y cuarto estadio de M. sacchari se obtuvieron a
partir de una colonia establecida en invernadero del Colegio de Postgraduados, Campus
Montecillo, Texcoco, Estado de México. Mientras que los huevos de C. carnea, C. comanche
y C. externa se obtuvieron a partir de una colonia establecida en el laboratorio anteriormente
mencionado. Los adultos fueron alimentados con una dieta elaborada por levadura de cerveza
y miel (1:1) que se ofrecía en algodón humedecido y que se cambiaba cada 2 días.
Desarrollo del experimento. De cada especie de Chrysoperla se usaron huevos de ≤ 6 h de
edad; cada huevo se colocó en una arena experimental para evitar el canibalismo entre larvas
al eclosionar. Como presas, se utilizaron ninfas de tercer y cuarto estadio de M. sacchari.
Cada 24 h se les ofreció 50, 70 y 100 presas a las larvas de primer, segundo y tercer ínstar
larval de crisópidos, respectivamente. El experimento se estableció en un diseño
completamente al azar, con 20 repeticiones para cada tratamiento bajo condiciones
controladas. Cada 24 h se registró el número de ninfas de M. sacchari consumidas.
Análisis de datos. Los datos de presas consumidas por las larvas de crisopas se sometieron
a un análisis de varianza y de ser necesario se aplicó una prueba de separación de medias con
Tukey (α=0,05), usando para ambos casos el programa SAS (2002).
RESULTADOS Y DISCUSIÓN
El rango de consumo del 1er ínstar larval de Chrysoperla fue de 107.60±0.86 a
119.85±0.80 pulgones, sin observar diferencia significativa entre especies. En el segundo

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
112
ínstar larval, C. comanche alcanzó un consumo de 79.05±1.52 pulgones, mientras que, C.
carnea y C. externa consumieron mayor e igual número de pulgones. El tercer ínstar larval
de las especies de Chrysoperla tuvieron un mayor consumo de ninfas de M. sacchari que los
otros ínstares, con un rango de consumo de 212.94±1.48 a 250.70±1.29 pulgones
consumidos; sin embargo, no se encontró diferencia significativa (Cuadro 1).
Cuadro 1. Consumo promedio de cada ínstar larval de tres especies de Chrysoperla,
alimentadas con ninfas de Melanaphis sacchari.
Tratamientos Ínstares larvales
L1 L2 L3 Total
C. carnea 119.85±0.80a 116.15±1.20ª 250.70±1.29a 486.70a
C. comanche 107.60±0.86a 79.05±1.52b 212.94±1.48a 400.82b
C. externa 116.05±0.89a 122.94±1.34a 246.39±1.50a 474.17a
P 0.1029 <0.0001 0.1785 0.0013
Letras diferentes en la misma columna indican diferencias significativas con Tukey
(α=0.05).
Los resultados indican que las tres especies de Chrysoperla registraron alta capacidad de
consumo (>400 ninfas), tan sólo el tercer ínstar larval consumió más que los ínstares uno y
dos juntos. Esta misma tendencia se registra en diversas investigaciones con larvas de tercer
ínstar de C. carnea sobre Aphis gossypii, Ropalosiphum maidis, Bemisia tabaci, huevos de
Sitotroga cerealella y Ephestia kuehniella, Amrasca devastans y Phenacoccus solenopsis
(Solangi et al., 2013; Saeed and Razaq, 2015; Saleh et al., 2017; Kabir et al., 2018). Sin
embargo, Panth et al. (2017) encontraron que la larva de tercer ínstar de C. carnea consumió
menor número de Brevicoryne brassicae, A. craccivora y A. fabae. Mientras que, Kabir et
al. (2018) encontraron resultados similares sobre A. gossypii y R. maidis. De acuerdo a los
resultados registrados en este trabajo, el último ínstar de las tres especies de Chrysoperla
presentó mayor tasa de consumo que el primer y segundo, esto podría deberse a su tamaño y
a los mayores requerimientos dietéticos para continuar con las siguientes fases biológicas
(Atthan et al., 2004; Ulhaq et al., 2006; Sattar et al., 2007).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
113
El consumo promedio durante todo el estado larval mostró diferencia significativa, C.
comanche consumió 400.82 ninfas de M. sacchari, mientras que, las otras dos especies de
Chrysoperla mostraron mayor capacidad de consumo, pero sin deferencia significativa entre
ellas (C. carnea consumió 486.70 y C. externa 474.17 ninfas del pulgón). La variación en la
tasa de consumo podría deberse al estadio ninfal de la presa sobre la cual se lleva a cabo el
estudio, es decir entre menor sea el tamaño de la presa, mayor sería la tasa de consumo del
depredador y viceversa, esto lo constatan los resultados obtenidos por Rashid et al. (2012),
Saeed and Razaq (2015), Saleh et al. (2017) y Manjunatha et al. (2018).
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca otorgada al primer
autor, al Colegio de Postgraduados por las facilidades para el desarrollo del programa de
posgrado. A la Fundación Guanajuato Produce A.C., por financiar el proyecto “Evaluación
de control biológico del PAS en Guanajuato y su compatibilidad con el control químico” en
el año 2017, del cual forma parte esta investigación.
LITERATURA CITADA
Athhan, R., B. Kaydan, and M. S. Ozogokce. 2004. Feeding activity and life history characteristics of the generalist predator, Chrysoperla carnea (Neuroptera: Chrysopidae) at different prey densities. J. Pest. Sci. 77: 17-2.
Kabir, K. K., H. Mannan S., S. Mureed M., Z. Rajput, M. Irfan J., P. Mureed M., and A. Ali K. 2018. Some parameters of Chrysoperla Carnea on floral feeds of castor and feeding potential on selective aphid species. J. of Entomol. and Zool. Studies 6: 860-863.
Manjunatha, D. K., S., E. Sowmya, and M. Maruthi. 2018. Feeding potential of Chrysoperla carnea (Steph) on different host. J. of Entomol. and Zool. Studies 6: 1028-1030.
Mansoor, M.M., N. Abbas, A. Shad S., K. Pathan A., and M. Razaq. 2013. Increased fitness and realized heritability in emamectin benzoate resistant Chrysoperla carnea (Neuroptera: Chrysopidae). Ecotoxicology 22: 1232-1240
New, T. R. 2001. Introduction to the systematics and distribution of Coniopterygidae, Hemerobiidae, and Chrysopidae used in pest management. In: McEwen, P., R. New, T., E. Whittington A. (Eds.), Lacewings in the crop environment. Cambridge University, UK, pp. 6-28.
Panth, M., G. Sarala and K. Kapil. 2017. Biology and predatory potential of Chrysoperla carnea S. (Neuroptera: Chrysopidae) reared on different species of aphid under laboratory condition. Int. J. of Advanced Research in Science, Engineering and Technology 4: 3847-3851.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
114
Paredes, D., M. Campos y L. Cayuela. 2013. El control biológico de plagas de artrópodos por conservación: técnicas y estado del arte. Ecosistemas 22: 56-61.
Rashid, M. U., M. Khattak, K. Abdullah, M. Amir, M. Tariq, and S. Nawaz. 2012. Feeding potential of Chrysoperla carnea and Cryptolaemus montrouzieri on cotton mealybug, Phenacoccus solenopsis. The J. of Animal & Plant Sci. 22: 639-643.
Rodríguez-del-Bosque, L.A. y A. Terán. 2015. Melanaphis sacchari (Hemiptera: Aphididae). A new sorghum insect pest in México. Southwest. Entomol. 40: 433-434.
Saeed, R., and R. Muhammad. 2015. Effect of prey resource on the fitness of the predator, Chrysoperla carnea (Neuroptera: Chrysopidae). Pakistan J. Zool. 47: 103-109.
Salamanca-Bastidas, J., E. Varón-Devia, and O. Santos-Amaya. 2010. Cría y evaluación de la capacidad de depredación de Chrysoperla externa sobre Neohydatothrips signifer, trips plaga del cultivo maracuyá. Corpoica Cienc. Tecnol. Agropec. 11: 31-40.
Saleh, A.A., H. El-Sharkawy, F. El-Santel, and A. El-Salam. 2017. Studies on the Predator Chrysoperla carnea (Stephens) in Egypt. Int. J. Environ. 6: 70-77.
Sattar, M., B. Fatima, N. Ahmed, and H. Abro. 2007. Development of larval artificial diet of Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae). Pakistan J. Zool. 39: 103-107.
SENASICA, 2015. Programa de trabajo de la campaña contra el pulgón amarillo a operar con recursos del componente de Sanidad Vegetal del Programa de Sanidad e Inocuidad Agroalimentaria 2015 en el estado de Nayarit. Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria (SENASICA). Oficio 03553. Ciudad de México, México. 13 p.
SIAP. 2015. Anuario estadístico de la producción agrícola. Consultado en octubre de 2019 en https://nube.siap.gob.mx/cierreagricola/
SIAP. 2019. Anuario estadístico de la producción agrícola. Consultado en octubre de 2019 en https://nube.siap.gob.mx/cierreagricola/
Solangi, A.W., G. Lanjar, N. Baloch, M. Rais, and A. Khuhro. 2013. Population, host preference and feeding potential of Chrysoperla carnea (Stephens) on different insect hosts in cotton and mustard crops. Sindh Univ. Res. J. (Sci. Ser.) 45: 213-218.
Ulhaq, M.M., A. Sattar, Z. Salihah, A. Farid, A. Usman, and U. Khattak. 2006. Effect of different artificial diets on the biology of adult green lacewing (Chrysoperla carnea Stephens.). Songklanakarin J. Sci. Technol. 28: 1-8.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
115
EVALUACIÓN EN JAULAS DE CAMPO DE DOS PARASITOIDES
DE HUEVO DE Spodoptera frugiperda (LEPIDOPTERA: NOCTUIDAE)
Jannet Jaraleño Teniente1, J. Refugio Lomelí Flores1, Esteban Rodríguez Leyva1, Rafael Bujanos-
Muñiz2
1Posgrado en Fitosanidad, Entomología y Acarología. Colegio de Postgraduados Carretera México-Texcoco km 36.5, Montecillo, 56230
Texcoco, Estado de México. 2INIFAP. Campo Experimental Bajío. Carretera Celaya – San Miguel de Allende km. 6.5, Celaya, 38010
Celaya, Guanajuato, México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
En México el maíz es uno de los cultivos más importantes y el gusano cogollero, Spodoptera
frugiperda Smith (Lepidoptera: Noctuidae) representa el principal problema fitosanitario de
este cultivo. Se estima que durante un ciclo agrícola se realizan de dos a tres aplicaciones de
insecticidas para combatir esta plaga (Blanco et al., 2014). El uso de enemigos naturales que
parasitan huevos de esta plaga, como Trichogramma spp., con liberaciones inundativas ha
sido el método de control biológico más empleado para su combate (Bahena y Cortez, 2015).
Sin embargo, otros parasitoides de huevo que atacan a S. frugiperda podrían proporcionar
mejores niveles de parasitismo que las especies de Trichogramma (Cave, 2000), dentro de
estos se ha señalado a Telenomus remus Nixon (Orden: Familia). Este es un parasitoide de
huevos de varias especies de lepidópteros, especialmente del género Spodoptera (Wojcik et
al., 1976). La ventaja de T. remus como agente de control biológico de S. frugiperda reside
en que puede parasitar masas de huevos superpuestas y con presencia de escamas (Gerling
and Schwartz, 1974), y los niveles de parasitismo pueden ser de 80 a 100% (Wojcik et al.,
1976). Por otro lado, Trichogramma pretiosum Riley, el parasitoide que comercialmente se
libera contra plagas de lepidópteros en México ha proporcionado resultados variables, y no
existen registros tan elevados de parasitismo (Murillo, 1977; Peralta, 1980). Debido a las
características biológicas de T. remus, tales como tamaño, especificidad y capacidad para
parasitar masas de huevos cubiertas por escamas, el objetivo de este trabajo fue comparar el
parasitismo de T. remus contra el parasitismo de T. pretiosum sobre huevos de S. frugiperda
en condiciones de jaulas de campo.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
116
MATERIALES Y MÉTODOS
Cría de insectos. T. remus se recuperó con centinelas (huevos de S. frugiperda) expuestos
en campos de maíz en el municipio de Tapachula, Chiapas, en otoño de 2018, y se reprodujo
en laboratorio sobre huevos de S. frugiperda. Los huevos se obtenían de una cría de dicha
especie que se mantuvo con la dieta propuesta por Poitut y Bues (1974) con modificaciones
de Alejandro Pérez-Panduro (Colegio de Postgraduados). Los parasitoides adultos se
alimentaban con una solución de agua azucarada al 10 %, y se les proporcionaron diariamente
huevos de S. frugiperda, de menos de 24 h de edad. Las crías de S. frugiperda y T. remus se
mantuvieron a 25±2°C, 75±5% HR y 12:12 h L:O. Por otra parte, T. pretiosum se obtuvo de
una cría comercial de la empresa Organismos Benéficos para la Agricultura S.A. de C.V.
(OBASA), Jalisco, México.
Parasitismo en condiciones jaulas de campo. Se evaluaron dos especies de parasitoides
(Te. remus y T. pretiosum) y tres dosis de liberación; las dosis consistieron en la proporción
de uno, dos y tres adultos del parasitoide por un huevo del huésped. Se utilizó un diseño
factorial (3x2) y un testigo (sin liberación de parasitoides). La unidad experimental consistió
de una jaula de tela de organza (3 x 3 x 1.8 m) donde se aislaron 3 surcos de maíz híbrido,
cada uno con 10 plantas, en donde se colocaron las masas de huevos (centinelas). Los
tratamientos se distribuyeron en un diseño experimental completamente al azar y se
realizaron cuatro repeticiones por tratamiento. En campo primero se establecieron las jaulas
para aislar los grupos de plantas. Luego, con una aspiradora motorizada se eliminaron los
organismos no deseados (plagas y entomófagos) de todas las plantas dentro de las jaulas.
Posteriormente, dentro de cada jaula, se colocaron aleatoriamente cinco masas de huevos de
S. frugiperda en el envés de las hojas, siguiendo la técnica de DaSilva et al. (2015);
inmediatamente después se liberaron los parasitoides. Las masas de huevos estuvieron
expuestas 48 horas. Después de este periodo los huevos se retiraron y se mantuvieron a
25±2°C y 75±5% HR.
El parasitismo se evaluó en cada masa de huevos; primero se fotografiaron las masas y
posteriormente se analizó cada imagen con el programa Image J. Los datos se analizaron

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
117
mediante regresión logística (p ≤ 0.05), y la comparación de medias se realizó mediante el
método de contrastes. Los análisis se desarrollaron con el programa R versión 3.6.
RESULTADOS Y DISCUSIÓN
El parasitismo de T. remus fue superior al de T. pretiosum en todas las dosis (p≤0.001).
En el caso de T. remus el parasitismo varió poco entre dosis y el porcentaje más alto (29.9%)
correspondió a la dosis de un parasitoide por un huevo de la plaga. En masas donde se
presentó parasitismo de T. remus se observó que este parasitaba más del 50% de la masa,
llegando en ocasiones a 93% de huevos parasitados sobre una masa. En cuanto a T.
pretiosum, el porcentaje de parasitismo más alto fue 4.19%. Este nivel de parasitismo sucedió
a pesar de que a las masas expuestas eran de una sola capa de huevos y sin escamas (Figura
1).
Figura 1. Porcentajes de parasitismo de Trichogramma pretiosum y Telenomus remus sobre huevos de
Spodoptera frugiperda en jaulas de campo (p≤0.001).
En general, las dosis de liberación de uno y dos parasitoides por huevo de S. frugiperda
fueron las mejores, y no fueron estadísticamente diferentes entre sí. Si se considera el costo
de producción, podría pensarse que la menor dosis puede ser valiosa para desarrollar ensayos
de campo. Debido a que la proporción sexual de la especie en condiciones de cría es de 1:1,
la dosis puede expresarse también como 0.5 hembras de T. remus por huevo de S. frugiperda.
En otros trabajos se indicó que dosis menores de liberación podrían alcanzar porcentajes
05
10152025303540
1 2 3
PORC
ENTA
JE D
E PA
RASI
TISM
O
±EE
DOSIS
Tr. pretiosum Te. remus
a a b
cd
d

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
118
elevados de control. Por ejemplo, Pomari et al. (2013) reportaron que en condiciones de
invernadero 0.165 hembras de T. remus por huevo de S. frugiperda proporcionó 70% de
parasitismo; Queiroz et al. (2017) con 0.13 a 0.15 hembras del parasitoide por huevo de la
plaga obtuvieron 71% de parasitismo. El mejor nivel de parasitismo en este trabajo se logró
con 13.8 hembras por m2 con parasitismo promedio de 29.9%. Figueiredo et al. (2002)
reportaron resultados similares a este trabajo, con 12 hembras/ m2 obtuvieron 88.3% de
parasitismo. De las 80 masas expuestas, T. remus parasitó 42.5% mientras que T. pretiosum
sólo lo hizo en 10%. No se descarta que los bajos niveles de parasitismo en este trabajo se
hayan debido a la falta de estímulos emitidos por las masas de huevos, esto pudiera estar
relacionado con el manipuleo de las masas de huevos de S. frugiperda, ya que a las masas se
les removieron las capas superiores de huevos, así como las escamas de la hembra para
estandarizar el tamaño de los centinelas. Se sabe que parasitoides de huevo son influenciados
por kairomonas emitidas por la palomilla, que utilizan para localizar al huésped (Fatouros et
al., 2008). Se ha comprobado que los compuestos (Z)-9-tetradeceno-1-ol acetato y (Z)-9-
dodeceno-1-ol acetato, componentes de la feromona sexual de S. frugiperda aumentan las
tasas de parasitismo de T. remus (Nordlund et al., 1983), y probablemente un manipuleo
excesivo de las masas de huevos, o la remoción de escamas influenció los resultados de este
trabajo. En trabajos futuros se recomienda considerar esta variable.
T. remus es un parasitoide que se utiliza con éxito en siete países latinoamericanos, y se
comercializa y recomienda contra plagas de lepidópteros (Cave, 2000). A pesar de esto,
prácticamente sólo existe un trabajo que evaluó a T. remus en México, y no hay ningún
programa de seguimiento. Aparentemente existe un potencial muy grande de T. remus para
usarse en México; sin embargo, hacen falta programas de evaluación y seguimiento en campo
para determinar cuál enemigo natural es más eficiente para mejorar el control de S.
frugiperda.
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la beca completa
de maestría para la primera autora. Al Dr. Iván Ramírez Ramírez del Posgrado de Genética

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
119
que permitió el desarrollo de los experimentos en campos de maíz en el Colegio de
Postgraduados.
LITERATURA CITADA
Bahena Juárez, F., Cortez Mondaca, E., 2015. Gusano cogollero del maíz, Spodoptera frugiperda (Lepidopera: Noctuidae), pp. 181–250. En: Casos de Control Biológico en México 2. Arredondo B. H. C., Rodríguez B. L. A. Eds. Biblioteca básica de agricultura, Jalisco México.
Blanco, C.A., Pellegaud, J.G., Nava-Camberos, U., Lugo-Barrera, D., Vega-Aquino, P., Coello, J., Terán-Vargas, A.P., Vargas-Camplis, J., 2014. Maize pests in Mexico and challenges for the adoption of integrated pest management programs. J. Integr. Pest Manag. 5: 1–9.
Cave, R.D., 2000. Biology, ecology and use in pest management of Telenomus remus. BioControl 21: 21–26.
DaSilva, C.S.B., Vieira, J.M., Loiácono, M., Margaría, C., Parra, J.R.P., 2015. Evidence of exploitative competition among egg parasitoids of Spodoptera frugiperda (Lepidoptera: Noctuidae) in maize. Rev. Colomb. Entomol. 41: 184–186.
Fatouros, N.E., Dicke, M., Mumm, R., Meiners, T., Hilker, M., 2008. Foraging behavior of egg parasitoids exploiting chemical information. Behav. Ecol. 19: 677–689.
Ferrer, F., 2001. Biological control of agricultural insect pests in Venezuela; advances, achievements, and future perspectives. BioControl 22: 67–74.
Figueiredo, M.L.C., Lucia, T.M.C.D., Cruz, I., 2002. Effect of Telenomus remus Nixon (Hymenoptera: Scelionidae) density on control of Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) egg masses upon release in a maize field. Rev. Bras. Milho E Sorgo 1: 12–19.
Gerling, D., Schwartz, A., 1974. Host selection by Telenomus remus, a parasite of Spodoptera littoralis eggs. Entomol. Exp. Appl. 17: 391–396.
Murillo Cruz, J.A., 1977. Resumen del avance de los programas de control biológico en la región central de la república mexicana. pp. 46–54. En: Memorias de la V Reunión Nacional de Control Biológico y Sector Agropecuario Organizado, Dirección General de Sanidad Vegetal. Departamento de Control Biológico, Cd. Victoria, Tamaulipas,
Nordlund, A., Lewis, W.J., Gueldner, C., 1983. Kairomones and their use for management of entomophagous insects. J. Chem. Ecol. 9: 695–701.
Peralta, F.G. 1980. Incidencia de insectos benéficos y plaga en maíz y sorgo. 159-168, En: Memorias de la VIII Reunión Nacional de Control Biológico y Sector Agropecuario Organizado. Manzanillo, Colima, México, 22-25 Abril 1980. Dirección General de Sanidad Vegetal. Departamento de Control Biológico. Manzanillo, Colima, México.
Poitout, S.; Bues, R. Elevage de chenilles de vingt-huit espèces de Lépidoptères Noctuidae et de deux espèces d'Arctiidae sur milieu artificiel simple. Particularités de l'élevage selon les espèces. Ann. Zool. Ecol. Anim. 1974. 6: 431-441.
Pomari, A.F., Bueno, A. de F., Bueno, R.C.O. de F., Menezes Junior, A. de O., Fonseca, A.C.P.F., 2013. Releasing number of Telenomus remus (Nixon) (Hymenoptera:

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
120
Platygastridae) against Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) in corn, cotton and soybean. Ciênc. Rural 43: 377–382.
Queiroz, A.P. de, Bueno, A. de F., Pomari-Fernandes, A., Bortolotto, O.C., Mikami, A.Y., Olive, L., 2017. Influence of host preference, mating, and release density on the parasitism of Telenomus remus (Nixon) (Hymenoptera: Platygastridae). Rev. Bras. Entomol. 61: 86–90.
Wojcik, B., Whitcomb, W.H., Habeck, D.H., 1976. Host range testing of Telenomus remus (Hymenoptera: Scelionidae). Fla. Entomol. 59:195–198.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
121
PARÁMETROS DEMOGRÁFICOS DE Laetilia coccidivora
(LEPIDOPTERA: PYRALIDAE) ALIMENTADO DE COCHINILLA
SILVESTRE Y GRANA COCHINILLA
Oscar A. Barreto-García1, Esteban Rodríguez-Leyva1, J. Refugio Lomeli-Flores1, Juan M. Vanegas-
Rico1, Ana Lilia Vigueras2, and Liberato Portillo2
1Posgrado en Fitosanidad, Entomología y Acarología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco,
Estado de México. 2Departamento de Botánica y Zoología (CUCBA), Universidad de Guadalajara, Nextipac, 45200 Zapopan, Jalisco, México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
Dactylopius opuntiae Cockerell (Hemiptera: Dactylopiidae) se considera plaga primaria de nopal
[Opuntia ficus-indica (L.) Miller (Caryophyllales: Cactaceae)] en México (Griffith 2004), también
se reconoció como plaga en Brasil y en al menos diez países de la cuenca del Mediterráneo
(Mazzeo et al. 2019) y es plaga introducida. En Etiopía D. coccus Costa; especie aprovechable
para extraer ácido, en 2009 se reportó como plaga invasora (Belay 2015).
Ambas especies de dactilópidos se alimentan directamente del floema de los cladodios, y
altas infestaciones de D. opuntiae (> 75% en el área del cladodio) ocasionan la muerte de las
plantas (Vanegas-Rico et al. 2010). No obstante, en ningún país se había presentado una plaga de
D. coccus como está sucediendo en Etiopía (Belay 2015). El daño por estas especies de
Dactylopius es relevante, ya que O. ficus-indica se considera la cactácea cultivada de mayor
importancia en el mundo, extendiéndose a lo largo de 4.5 millones de hectáreas en cuatro
continentes (Mazzeo et al. 2019).
Para el manejo de ambas especies de dactilópidos se ha propuesto como alternativa el
control biológico (Vanegas-Rico et al. 2010; Vigueras y Portillo et al. 2014). Existen 13 enemigos
naturales de estas especies de Dactylopius, todos son depredadores (Vanegas–Rico et al. 2010).
No obstante, se desconocen los parámetros demográficos de la mayor parte del complejo de éstos.
L. coccidivora se considera el enemigo natural más voraz de D. coccus en las áreas productoras de
esta especie en invernaderos comerciales y D. opuntiae en campo en México (Vigueras y Portillo
2014; Vanegas-Rico et al. 2010, 2017 se desconoce si el porcentaje de ácido carmínico impacta
en la biología de cada especie ya que el ácido carmínico se considera un mecanismo de defensa,

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
122
disuasivo de la depredación, presente en el género Dactylopius spp. (Eisner et al. 1994). El objetivo
de este trabajo fue determinar los parámetros demográficos de L. coccidivora alimentado de D.
opuntiae y D. coccus.
MATERIALES Y MÉTODOS
Éxito reproductivo de L. coccidivora. Se eligieron 50 pupas de L. coccidivora para cada
presa, Se formaron 20 parejas, y cada pareja se colocó en una jaula de alambre (60 x 35x 60 cm)
cubiertas de tela organza; en caso de la muerte de un macho éste se sustituyó de forma inmediata
por otro. Cada jaula tenía, en la parte superior, un tubo de PVC (Ø = 2) que sirvió para colgar un
cladodio infestado con la presa respectiva (50% de infestación). Cada cladodio se removía cada
24 h y se contaban los huevos por hembra por día. Para fertilidad se aislaron 80 huevos de manera
individual de cada tratamiento, y se observó porcentaje de emergencia de larvas de primer ínstar;
este ensayo se desarrolló en condiciones de invernadero (25 ± 15°C, 70 ± 20% HR), y fotoperiodo
natural de primavera en la región de Guadalajara, Jalisco. Cada hembra se consideró una
repetición, se realizaron nueve repeticiones por tratamiento, y el experimento completo se repitió
dos veces con desfase de dos semanas (en total 18 repeticiones por presa).
Ácido carmínico por especie de Dactylopius. El contenido de ácido carmínico por cada
especie de Dactylopius se determinó por el método propuesto por Méndez et al., (2004). La
determinación del contenido de ácido carmínico fue sobre estados de desarrollo de ninfa I, y II,
hembra juvenil (sin presencia de huevos), hembra grávida.
Análisis de datos. Los valores de fecundidad, fertilidad y tiempo de desarrollo se utilizaron
para elaborar una tabla de vida y fertilidad. La tasa neta de reproducción (R0), el tiempo de
generación (T), la tasa intrínseca de incremento natural (rm) y la tasa finita de incremento (ʎ) se
obtuvieron mediante el programa de Maia et al., (2000) en Lifetable SAS (SAS Institute, 2000.
Además, se realizó pruebas de t de Student´s para grupos apareados.
RESULTADOS
Éxito reproductivo de L. coccidivora. El periodo de preoviposición fue de 3 días sobre
ambas presas (Figura 1). El 65% de las hembras de L. coccidivora colocaron grupos de 3 a 7
huevos, y el 35% restante los colocaron de forma individual. Los sitios de oviposición más
comunes fueron las areolas o gloquidos, además de hendiduras o pequeñas grietas en la epidermis

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
123
del cladodio. De los huevos colocados de manera individual prácticamente el 90% se localizaron
sobre los capullos de machos de los dactilópidos de ambas presas, el 10% de los huevos restantes
se localizaron debajo de hembras juveniles o grávidas. Las hembras que se alimentaron de D.
opuntiae colocaron 38.30 ± 5.65 huevos versus los 18.65 ± 3.57 de hembras alimentadas con D.
coccus, y esa diferencia fue significativa (t= 0.198, df=86, p= 0.021). La fertilidad también tuvo
diferencias, 92.68 ± 0.8 % versus 58.88 ± 1.27%, para huevos que provinieron de hembras
alimentadas con D. opuntiae o D. coccus, respectivamente (Figura 1).
Figura 1. Fecundidad de Laetilia coccidivora criada en dos especies de Dactylopius (25±15°C, 70±20% HR).
La alimentación sobre ambas especies de Dactylopius proporcionó la nutrición para que en
ambos casos se obtuvieran parámetros favorables para el crecimiento poblacional de L.
coccidivora (Cuadro 1). El tiempo de generación de L. coccidivora fue 42.37 y 36.6 días cuando
se alimentó de D. opuntiae o D. coccus, respectivamente, y se encontraron diferencias entre
tratamientos. La (R0) no fue diferente entre tratamientos, sin embargo los parámetros (rm), (λ), y
(DT) fueron favorables a las hembras que se alimentaron con D. opuntiae (Cuadro 1).
Cuadro 1. Parámetros demográficos (medias y niveles de confianza 95%) para Laetilia coccidivora
alimentada con dos especies de Dactylopius.
Dieta mixta
Tiempo de generación (T)
(días) Tasa neta de
reproducción (R0) Tasa intrínseca de
crecimiento natural (rm) Tasa finita de
crecimiento (λ) Tiempo de
duplicación (DT)
D. coccus 42.37
(42.00-42.73) 5.82
(4.85-6.79) 0.041
(0.037-0.045) 1.042
(1.038-1.046) 16.60
(15.08-18.12)
D. opuntiae 36.6 *
(34.97-38.28) 10.11
(4.5-15.68) 0.064 *
(0.048-0.080) 1.066 *
(1.049-1.083) 10.49 *
(7.25-13.73) Las medias dentro de una columna seguidas del asterisco son significativamente diferentes (t
de Student, p >0.05).
-113579
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
Fecu
ndid
ad
Edad en días
D. coccus D. opuntiae

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
124
Concentración de ácido carmínico. A excepción del primer ínstar ninfal, la concentración
de ácido carmínico fue diferente entre especies (t= 0.019, df = 3, p = 0.039), y entre estados de
desarrollo; en el segundo ínstar y el estado adulto la concentración fue mayor en D. coccus que D.
opuntiae (Cuadro 2).
Cuadro 2. Porcentajes de ácido carmínico en diferentes estados de desarrollo de Dactylopius
spp.
Estado de desarrollo D. opuntiaeiae D. coccus Ninfa I 1.77% 1.96% Ninfa II 2.01% 8.95%*
Adultas juveniles 5.32% 25.23%* Adultas grávidas 3.11% 23.58%*
*En cada fila los valores seguidos con el asterisco son diferentes entre sí (t de Student´s, p>
0.05).
DISCUSIÓN
Laetilia coccidivora es una especie nativa de Norteamérica y depreda insectos sésiles de las
familias Coccidae, Pseudococcidae, Eriococcidae, y Dactylopiidae (Scholtens y Solis 2015).
Además, es uno de los enemigos naturales más frecuentes y abundantes de D. opuntiae en áreas
naturales y cultivadas de Opuntia spp. en el sur de Estados Unidos de América y México; y
también es frecuente sobre D. coccus en las explotaciones comerciales en invernadero en México,
donde se cultiva para la obtención del ácido carmínico (Vigueras y Portillo 2014; Vanegas-Rico
et al. 2010, 2017, Cruz-Rodríguez et al. 2016).
Los resultados de este trabajo son de interés en al menos dos vertientes; en la primera se
demostró que el ácido carmínico, o precursores de éste, que producen D. opuntiae y D. coccus
como sustancias de novo afecta el desempeño de sus insectos depredadores, alargando el tiempo
de desarrollo y disminuyendo la supervivencia y fecundidad, como ya se conocía y había reportado
con sustancias que adquieren o secuestran los fitófagos de las plantas que se alimentan (Duffey
1980). La segunda tiene que ver con la selección natural; L. coccidivora ha estado expuesta a una
presión de selección para consumir presas con una concentración de ácido carmínico, o precursores
en la ruta metabólica de éste, 500% mayor a la que se presenta de manera natural en D. opuntiae,

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
125
y ha logrado adaptarse. Tanto así que L. coccidivora se considera el enemigo natural más voraz de
D. coccus en las áreas productoras de esta especie en invernaderos comerciales en México
(Vigueras y Portillo 2014; Vanegas-Rico et al. 2010, 2017). Con esta evidencia, es recomendable
realizar trabajos para estimar la voracidad de este depredador en ambas especies de presas, para
entender su papel como regulador natural de poblaciones de Dactylopius en cultivos de O. ficus
indica en las regiones donde estas especies de insectos son problema como plaga en este cultivo
de importancia mundial.
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la beca otorgada
durante los estudios de maestría para el primer autor.
LITERATURA CITADA
Belay, T. 2015. Carmine cochineal: fortune wasted in northern Ethiopia. Journal of the Professional Association for Cactus Development 17: 61-80.
Cruz-Rodríguez, J. A., González-Machorro E., Villegas González A. A., Rodríguez Ramírez M. L., Mejía Lara F. 2016. Autonomous biological control of Dactylopius opuntiae (Hemiptera: Dactylopiidae) in a prickly pear plantation with ecological management. Environmental Entomology 45: 642-648.
Duffey, S. S. 1980. Sequestration of plant natural products by insects. Annual Review of Entomology 25: 447-477.
Eisner, T., Ziegler R., McCormick J. L., Eisner M., Hoebeke E. R., and Meinwald J. 1994. Defensive use of an acquired substance (carminic acid) by predaceous insect larvae. Experientia 50: 610-615.
Griffith, M. P. 2004. The origins of an important cactus crop, Opuntia ficus‐indica (Cactaceae): new molecular evidence. American Journal of Botany 91: 1915-1921.
Mazzeo, G., Nucifora S., Russo A., Suma P. 2019. Dactylopius opuntiae, a new prickly pear cactus pest in the Mediterranean: an overview. Entomologia Experimentalis et Applicata 167: 59-72.
Méndez, J., González, M., Lobo M. G., Carnero A. 2004. Color quality of pigments in cochineals (Dactylopius coccus Costa). Geographical origin characterization using multivariate statistical analysis. Journal of Agricultural and Food Chemistry 52: 1331-1337.
Scholtens, B. G., Solis M. A. 2015. Annotated check list of the Pyraloidea (Lepidoptera) of America North of Mexico. ZooKeys 535: 1–136.
Vanegas-Rico J. M., Lomeli-Flores J. R., Rodríguez-Leyva E., Mora-Aguilera G., Valdez J. M. 2010. Enemigos naturales of Dactylopius opuntiae (Cockerell) en Opuntia ficus-indica (L.) Miller en el centro de México. Acta Zoológica Mexicana (n. s.) 26: 415-433.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
126
Vanegas-Rico, J. M., Pérez-Panduro, A., Lomeli-Flores, J. R., Rodríguez-Leyva, E., Valdez-Carrasco, J. M., Mora-Aguilera, G. 2017. Dactylopius opuntiae (Hemiptera: Dactylopiidae) population fluctuations and predators in Tlalnepantla, Morelos. Folia Entomológica Mexicana 3: 23-31.
Vigueras, A. L., & Portillo, L. (2014). Control de cochinilla silvestre y cría de grana cochinilla. Consejo Estatal de Ciencia y Tecnología 1: 1-66.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
127
PARÁMETROS DEMOGRÁFICOS DE Trichopria drosophilae
(HYMENOPTERA: DIAPRIIDAE) SOBRE TRES HUÉSPEDES
DIFERENTES
José Mauricio Esteban-Santiago1*, Esteban Rodríguez-Leyva1, J. Refugio Lomeli-Flores1 y Jaime
González-Cabrera2
1Colegio de Postgraduados, Posgrado en Fitosanidad, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco, Estado de México,
México.2Centro Nacional de Referencia de Control Biológico, CNRF-DGSV, SENASICA. 28110 Tecomán, Colima, México.
INTRODUCCIÓN
La mosca del vinagre de las alas manchadas, Drosophila suzukii Matsumura (Diptera:
Drosophilidae), es nativa del sureste asiático y en México se reportó por primera vez en
noviembre de 2011 (Senasica 2013). D. suzukii tiene un ovipositor aserrado y un
comportamiento diferente a otros drosofílidos, lo que le confiere la capacidad de atacar frutos
inmaduros, sus larvas se alimentan de la pulpa causando pérdida de turgencia de los frutos y
reduciendo su valor comercial (Rota-Stabelli et al. 2013).
El control biológico puede ser una alternativa para reducir las poblaciones de esta plaga,
sin causar impacto ambiental desfavorable, sobre todo en las áreas aledañas a los cultivos o
aquellas que no tienen manejo del problema. El endoparasitoide pupal Trichopria
drosophilae Perkins (Hymenoptera: Diapriidae) recibió atención porque tiene alta eficacia
de parasitismo sobre D. suzukii (Schlenke et al. 2007; Wang et al. 2016; Rossi Stacconi et
al. 2015) y otras especies de drosofílidos (Chabert et al. 2012). Se asume que el tamaño del
parasitoide está directamente relacionado con el del huésped (Mackay, 2010). Asimismo, un
mayor tamaño en el parasitoide, le puede conferir mejor aptitud física y biológica (Schlenke
et al. 2007; Liu et al. 2011). Lampson et al. (1996) reportaron que el tamaño de machos y
hembras de parasitoides aumentaban conforme aumentaba el tamaño del hospedero, y que el
tamaño de los parasitoides tiene efectos en varias características biológicas como la
longevidad, competitividad y aptitud reproductiva.
Trichopria drosophilae es uno de los candidatos para usarse como agente de control de
D. suzukii. No obstante, es importante conocer los factores que pueden incidir en su cría y

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
128
reproducción masiva. De acuerdo con los datos reportados en diferentes trabajos se puede
deducir que un huésped de mayor tamaño para T. drosophilae le puede conferir ventajas en
varios aspectos biológicos. Por tal motivo, este trabajo tuvo como objetivos determinar el
tiempo de desarrollo, fecundidad y parámetros poblacionales de T. drosophilae criado en
diferentes huéspedes.
MATERIALES Y MÉTODOS
Cría de los insectos. Se utilizaron tres especies de drosofílidos, D. suzukii, D.
melanogaster y D. immigrans. La cría de D. suzukii se inició con individuos del Centro
Nacional de Referencia de Control Biológico (CNRCB) de Tecomán, Colima; la cría de D.
melanogaster con individuos del Posgrado en Recursos Genéticos y Productividad-Genética
del Colegio de Postgraduados, Campus Montecillo, y la cría de D. immigrans con individuos
del laboratorio de genética y toxicología ambiental-Banco de Moscas, de la Facultad de
Ciencias de la Universidad Nacional Autónoma de México (UNAM). La colonia de T.
drosophilae la proporcionó el CNRCB. Para la reproducción y mantenimiento de D. suzukii
se utilizó plátano fresco (González-Cabrera et al. 2018), y para D. melanogaster se utilizó
una dieta estándar. Referente a la reproducción y mantenimiento de D. immigrans se utilizó
una dieta artificial (medio banana-Opuntia) recomendada por la Dra. Patricia Ramos de FC-
UNAM. Las crías, y los ensayos experimentales se mantuvieron en condiciones de
laboratorio a 25 ± 1°C, 60-70% HR y fotoperiodo 10:14 L:O. Para cada experimento se
usaron individuos de T. drosophilae como mínimo de la segunda generación proveniente de
cada especie de huésped.
Tiempo de desarrollo de T. drosophilae. En recipientes de plástico de 29.5 ml se
colocaron 40 pupas de cada especie de huésped de 16 a 24 h de edad y se expusieron durante
6 h a cuatro hembras apareadas de T. drosophilae de 4 d de edad (edad a la que tienen la
mayor carga de huevos). Después del tiempo de exposición las pupas se mantuvieron hasta
la emergencia de adultos de avispas o moscas y las observaciones se realizaron cada 12 h.
Este experimento se repitió 10 veces.
Fecundidad de T. drosophilae. Este experimento se inició con hembras y machos de ≤
24 h de edad. La fecundidad de T. drosophilae se evaluó en 15 parejas por cada especie de

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
129
huésped. Cada hembra estuvo con un macho, y en el caso de muerte de un macho se sustituyó
por otro. Cada pareja de T. drosophilae se colocó en recipientes de plástico de 59 ml. Cada
tapa del recipiente tenía un orificio, sellado con tela de organza, para permitir ventilación.
Asimismo, se incluyó una tira de papel con unas líneas de miel de abeja con polen. Cada 24
h se exponían 20 pupas del huésped de ≤ 24 h de edad de acuerdo a cada tratamiento; después
de ese tiempo, las pupas se reemplazaban por otras. Posterior a eso, se esperó hasta la
emergencia de adultos de T. drosophilae o de moscas. Este procedimiento se continuó hasta
la muerte de la última hembra del parasitoide. Una vez que las moscas o parasitoides dejaron
de emerger se estimó la fecundidad y la supervivencia. Al final, también se estimó la
proporción sexual. Para mayor comodidad los parasitoides emergidos de pupas de D.
melanogaster se denominarán Tdmelanogaster, los de D. suzukii como Tdsuzukii y a los
emergidos de D. immigrans como Tdimmigrans.
Análisis. El tiempo de desarrollo de huevo a adulto se analizó mediante las pruebas Log-
rank del Proc LIFETEST con ajuste de Bonferroni. La prueba de Log-rank otorga el mismo
peso a cada medición durante todo el período experimental.
RESULTADOS Y DISCUSIÓN
Tiempo de desarrollo de T. drosophilae. El tiempo de desarrollo del parasitoide T.
drosophilae varió de un mínimo de 15.74 días para los machos de Tdmelanogaster y un
máximo de 19.7 días para las hembras de Tdimmigrans. El tiempo de desarrollo de machos
o hembras del parasitoide que emergieron de D. melanogaster no fue diferente (P=0.255 >
0.05), pero si se presentó diferencia en el tiempo de desarrollo entre sexos (P<0.0001) cuando
el parasitoide se desarrolló en D. suzukii y D. immigrans (Cuadro 1).
Cuadro 1. Tiempo de desarrollo en días de Trichopria drosophilae desarrollado en diferentes huéspedes.
Tratamiento Machos (media ± EE) Hembras (media ± EE) D. melanogaster 15.74 ± 0.735 16.79 ± 0.569 D. suzukii 16.74 ± 0.749 18.70 ± 0.516 * D. immigrans 17.75 ± 0714 19.7 ± 0.516 *
Valores entre filas seguidos del asterisco son significativamente diferentes (prueba Log-Rank, p< 0.001).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
130
Fecundidad de T. drosophilae. Los adultos de T. drosophilae provenientes de los tres
huéspedes se aparearon y comenzaron la oviposición en las primeras 24 h de edad y las
hembras colocaron la mayoría de los huevos durante los primeros 5 d de vida (Figura 1).
La longevidad de hembras de Tdmelanogaster fue de mínimamente 10 d, y el promedio
de 12.93 d. Para las hembras de Tdsuzukii la mínima fue 17 d y el promedio de 18.6 d;
respecto a las hembras de Tdimmigrans, la mínima fue 17 d, y el promedio de 19.6 d.
Figura 1. Fecundidad diaria de Trichopria drosophilae (=Td) sobre diferente huésped. D. melanogaster (m) D. suzukii (s) D. immigrans (i).
La fecundidad promedio fue mayor para Tdimmigrans con un valor de 46.3, mientras que
para Tdsuzukii fue de 44.8 y el menor promedio de fecundidad correspondió a
Tdmelanogaster con 36.6. Referente a la proporción sexual ésta fue similar con valores de
0.572, 0.579, 0.53 para Tdimmigrans, Tdsuzukii y Tdmelanogaster respectivamente.
(Cuadro 2).
Cuadro 2. Proporción sexual y fecundidad promedio de Trichopria drosophilae emergidos de los diferentes huéspedes
Tratamiento Fecundidad promedio Tratamiento Proporción sexual promedio
Tdimmigrans 46.33 a Tdsuzukii 0.5793 a Tdsuzukii 44.86 a Tdimmigrans 0.5726 a Tdmelanogaster 36.66 b Tdmelanogaster 0.5358 b
Valores entre columnas seguidos por la misma letra no muestran diferencia significativa (α=0.05)

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
131
Con este trabajo se corroboró que los huéspedes influyen el tiempo de desarrollo de los
parasitoides, y es probable que ese tiempo de desarrollo también se relacione con la cantidad
de recursos (diferente tamaño) que ofrecía cada huésped.
Los parasitoides criados en D. immigrans y D. suzukii tuvieron valores más altos de
fecundidad comparados con los emergidos de D. melanogaster, esto sugiere que las
diferencias de tamaño en el huésped influyen en las características reproductivas del
parasitoide. Estos resultados son similares a los reportados por Chen et al. (2018) quienes
indicaron que el tamaño del huésped le confirió ventajas en fecundidad a T. drosophilae.
Aunque no fueron conclusivos los resultados de proporción sexual, también se notó una
tendencia hacia la proporción de hembras dependiendo del tipo de huésped, estos resultados
los reportaron Boycheva et al. (2019), y se sugiere seguir evaluando los huéspedes de este
trabajo para concluir al respecto. Los hallazgos de este trabajo invitan a continuar
investigando los factores que pueden influir en la eficacia de T. drosophilae para mejorar la
crías y evaluar su potencial para suprimir poblaciones de D. suzukii.
AGRADECIMIENTOS
A la Dra. P. Ramos de la FC-UNAM por la disponibilidad de compartir material de su
banco de drosofílidos. Al Colegio de Postgraduados y al Centro Nacional de Referencia de
Control Biológico por la oportunidad para desarrollar la parte científica y experimental y al
CONACYT por la beca otorgada.
LITERATURA CITADA
Boycheva, W., Romeis, J., Collatz J. 2019. Influence of the rearing host on biological parameters of Trichopria drosophilae, a potential biological control agent of Drosophila suzukii. Insects 10: 183.
Chabert, S., Allemand, R., Poyet, M., Eslin, P., Gibert, P. 2012. Ability of European parasitoids (Hymenoptera) to control a new invasive Asiatic pest. Drosophila suzukii. Biological Control 63: 40–47.
Chen, J., S. Zhou, Y. Wang, M. Shi, X. Chen, J. Huang. 2018. Biocontrol characteristics of the fruit fly pupal parasitoid Trichopria drosophilae (Hymenoptera: Diapriidae) emerging from different hosts. Scientific Reports. 8: 13323.
González-Cabrera, J., Garcia-Cancino, M.D., Moreno-Carrillo, G., Sanchez-Gonzalez, J.A., Arredondo-Bernal, H. 2018. Fresh banana as an alternative host for mass rearing Drosophila suzukii. Bulletin of Insectology 71: 65-70.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
132
Lampson, L.J., Morse, J.G., Luck, R.F. 1996. Host selection, sex allocation, and host feeding by Metaphycus helvolus (Hymenoptera: Encyrtidae) on Saissetia oleae (Homoptera: Coccidae) and its effect on parasitoid size, sex, and quality. Environmental Entomology 25: 283–294.
Liu, Z.D., Xu B.B., Li L., Sun, J.H. 2011. Host-size mediated trade-off in a parasitoid Sclerodermus harmandi. Plos One 6: e23260.
Mackay, T.F.C. 2010. Mutations and quantitative genetic variation: lessons from Drosophila. Philos. Trans. R. Soc. Lond B Biol. Sci. 365: 1229–1239.
Rota-Stabelli, O., Blaxter, M., Anfora, G. 2013. Quick Guide: Drosophila suzukii. Current Biology 23: 8-9.
Schlenke, T.A., Morales, J., Govind, S., Clark, A. G. 2007. Contrasting infection strategies in Generalist and specialist wasp parasitoids of Drosophila melanogaster. PLoS Path. 3: 1486–1501.
Stacconi, R., Buffington, M., Daane, K.M, Dalton, D.T., Grassi, A., Kaçar, G., Miller, B., Miller, J.C., Baser, N., Ioriatti, C., Walton, V.M., Wiman, N.G, Xingeng Wang, Anfora G. 2015. Host stage preference, efficacy and fecundity of parasitoids attacking Drosophila suzukii in newly invaded areas. Biological Control 84: 28–35.
Wang, X.G., Kacar, G., Biondi, A., Daane, K.M. 2016. Life-history and host preference of Trichopria drosophilae, a pupal parasitoid of spotted wing drosophila. BioControl 61: 387–397.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
133
CALIDAD DE LAS FEROMONAS SINTÉTICAS UTILIZADAS EN
EL TRAMPEO DE LEPIDÓPTEROS DE IMPORTANCIA
CUARENTENARIA
José Manuel Montiel-Castelán1, Juan Cibrian-Tovar1*, Nestor Bautista-Martinez1, José Abel
López-Buenfil2
1 Posgrado en Fitosanidad-Entomología y Acarología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230
Texcoco, Estado de México, México. 2 Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria, Boulevar Adolfo Ruiz
Cortinez, No. 5010, Colonia Insurentes Cuicuilco, Coyoacan, Ciudad de Mexico. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
En México el Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria
(SENASICA) realiza desde el año 2010, la Vigilancia Epidemiológica Fitosanitaria de plagas
de importancia cuarentenaria, de las cuales de un listado de plagas de mas de 1,000, se
seleccionan 35 plagas, considerando factores como el potencial de introducción, potencial de
establecimiento y consecuencias económicas y sociales (SAGARPA-SENASICA, 2019).
Para el año 2019 se monitorean 14 especies de insectos a través de estrategias como el
trampeo, utilizando feromonas, de las cuales no es posible conocer su calidad y eficiencia,
debido a que se trata de plagas ausentes, sin embargo, se han detectado en México por primera
vez a Grapholitha molesta, Grapholita prunivora, Grapholita packardi, Euwallacea sp. y
Argirotaenia franciscana (SAGARPA- SENASICA, 2019).
Haciendo referencia a los lepidópteros plaga que están bajo un programa de detección por
ser considerados de importancia cuarentenaria, (10 especies) son: Epiphyas postvittana,
Cactoblastis cactorum, Helicoverpa armigera, Spodoptera litura, Lobesia botrana,
Grapholita molesta, Argyrotaenia franciscana, Tuta absoluta, Lymantria dispar y
Thaumatotibia leucotreta, para lo cual se utilizan feromonas sexuales, mismas que proceden
de distintos formuladores y proveedores (SAGARPA- SENASICA, 2019). Los sistemas de
trampeo de lepidópteros considerados plagas ausentes de un área, están influenciados por
diferentes factores, la trampa, el clima, la fenología de los cultivos, la biología de la plaga y
el atrayente (Barrera et al., 2006).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
134
Para el caso de México, el sistema de trampeo de plagas de importancia cuarentenaria, a
base de feromonas sexuales, es necesario que se verifique que los compuestos que contiene
la feromona se encuentren en las proporciones adecuadas para que cumpla con la función de
capturar las especies que se buscan, además no existen medidas regulatorias que garanticen
que los productos (atrayentes) utilizados son los adecuados.
Por lo anterior, el sistema de trampeo que se utiliza en México para la Vigilancia de Plagas
Cuarentenarias, requiere que aplique un control de calidad de los semioquímicos utilizados,
toda vez que de esto depende su capacidad de atracción y como resultado una eficiencia en
las capturas. Por tal motivo, en esta investigación se plantea como objetivo determinar la
calidad de los semioquímicos utilizados en México para la detección de lepidópteros de
importancia cuarentenaria, lo que involucra evaluar contenido y concentración, según lo
indicado por la marca comercial y la literatura. En segunda instancia, se realiza un estudio
para evaluar la tasa de liberación de las feromonas de trampas colocadas en campo de durante
0,10,20 y 30 días.
MATERIALES Y MÉTODOS
Análisis de la composición y proporción de compuestos que tienen feromonas
comerciales de 10 especies de lepidópteros cuarentenados. Se realizó la selección del
disolvente adecuado que nos permitiera extraer los analitos de interés sin sustraer otras
sustancias que no son objeto de este estudio, de los disolventes hexano, diclorometano,
acetato de etilo, acetona, etanol y cloroformo, utilizando septos nuevos en siete mililitros de
los disolventes con agitación manual por 7 minutos.
Se realizó la elección del método de extracción de los analitos mediante extracción en fase
solida, extracción con aireación dinámica y extracción con disolventes por inmersión. Esta
última fue la técnica seleccionada, para ello se colocó un septo comercial con feromona (se
usó Pherocon 3416 de H. armigera), y se añadieron siete mililitros de acetato de etilo (AcEt)
en un vial de 20 mL. Se tapó y se empezó a medir el tiempo para siete minutos, agitando
manualmente los primeros dos minutos, dejando reposar tres minutos y agitando

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
135
manualmente los últimos dos minutos. Después se extrae el septo y se tapa el vial
guardándolo hasta su uso en congelación a -20 °C.
Las feromonas extraidas de los septos comerciales de 10 especies de lepidópteros de dos
empresas, se analizaron mediante cromatografía de gases (CG) en un cromatógrafo Agilent
7890A (Agilent Technologies®, USA), acoplado a un detector de masas (DM) Agilent
5975C. La columna fue una HP 5-ms 30 m × 0.250 mm y 0.25 µm de grosor de fase
estacionaria. El gas acarreador fue helio con grado de pureza 99.9 % con flujo en columna
de un mililitro por minuto, en modo splitless. El horno se inició en 50 °C y se incrementó en
15 °C/minuto hasta alcanzar la temperatura final de 280 °C, la corrida se hizo por 25 minutos.
Se inyectó un microlitro de cada uno de los extractos (a menos que se indiquen
modificaciones). El MSD se operó en modo scan, en un rango de 30-550 uma. La interface
se mantuvo a 280 °C, el detector y cuadrupolo a 150 y 230 °C, respectivamente.
La adquisición de datos fue en modo impacto electrónico, a 70 eV (Velázquez et al. 2104).
Los compuestos del cromatograma y los espectros respectivos se corroboraron con los
contenidos de la biblioteca NIST (2014) y con los estándares auténticos, entre ellos: Z-8-
dodecen-1-ol acetato, Z-9-tetradecen-1-ol y (Z,E)9,12-tetradecen-1-il acetato. En todos los
casos se estimó el error estándar para los picos de interés y abundancia de cada uno de ellos
en los cromatogramas.
Cambios en la tasa de liberación de tres feromonas sexuales de tres especies de
lepidópteros cuarentenados. Se colocarón en campo trampas con feromonas de tres
especies (Helicoverpa armigera, Tuta absoluta y Spodoptera litura), con 4 repeticiones, se
colectaron los septos expuestos durante 0, 10, 20 y 30 días, se realizó la extracción de los
analitos a través de la metodología de inmersión y a través del análisis cromatográfico
mencionado anteriormente.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
136
RESULTADOS Y DISCUSIÓN
Cuadro 1. Resultados del análisis cromatográfico de feromonas sintéticas de 10 especies de lepidóteros de varias marcas comerciales en México. Se indican las referencias respectivas
Especie Marca 1 Marca 2 Referencia
Argyrotaenia franciscana
Cumple con la identidad adecuada, y la proporción se acerca a la esperada de 1:15
Cumple con la identidad adecuada, y la proporción se acerca a la esperada de 1:15
Hills, A.S., R.T. Cardé, H. Kido and W.L. Roelofs. 1975
Cactoblastis cactorum Cumple con la identidad adecuada y la proporción (mayor area y duración) respectiva.
Cumple con la identidad adecuada y la proporción respectiva
Cibrián-Tovar, J., J. Carpenter, S. Hight, T. Potter, G. Logarzo and J.C. Velázquez- González.
2017.
Epiphyas postvittana Cumple con los estándares de identidad química, cantidad (0.8 mg) y proporción de los compuestos (20:1)
Cumple con los estándares de identidad química, cantidad (0.8 mg) y proporción de los compuestos (20:1)
Bellas, T.E., R.J. Bartell and A. Hill. 1983
Grapholita molesta Cumple con los compuestos y la proporción adecuada
Solo se logro detectar solo un compuesto
Bisotto-de-Oliveira, R., R. M. Morais, I. Roggia, S. J. N. Silva, J. Santana and C. N. Pereira. 2015.
Helicoverpa armigera Cumple con los compuestos y la proporción adecuada 1.2 mg
Solo se logro detectar un compuesto el mayoritario Chumakov M.A., and
Kuznetsova T.L. 2009
Lobesia botrana Cumple con los compuestos y la proporción adecuada
Cumple con los compuestos y la proporción adecuada
Ideses, R., J.T. Klug, A.Shani, S. Gothilf and E. Gurevitz. 1982.
Lymantria dispar Cumple con los compuestos y la proporción adecuada
Cumple con los compuestos y la proporción adecuada (no se detectaron antioxidantes)
Charlton R.E. and R. T Carde. 1982
Spodoptera litura Solo se puedo detectar el compuesto mayoritario (solo un antioxidante)
Cumple con los compuestos y la proporción adecuada (0.9 mg) Youshima T, Tamaki Y,
Kamano S, Oyama M, 1974.
Thaumatotibia leucotreta
Solo se detectó uno de los componentes y un antioxidante
Solo se detectó uno de los componentes y un antioxidante
Persoons, C. J., F. J. Ritter, and W. J. Nooijen. 1977.
Tuta absoluta Cumple con los componentes solo que no con la proporción 1:3
Cumple con los compuestos y la proporción adecuada
Attygalle, A.B., Jham, G.N., Svatos, A., Frighetto, R.T.S., Ferrara, F.A., Vilela, E.F., Uchoa- Fernandes, M.A., and Meinwald, J. 1996.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
137
En la mayoría de las feromonas se observe que cumplen con la identidad y proporción
esperada y solo en algunos casos no se detectó uno de los compuestos y la proporción fue
variable.
Cambios en la tasa de liberación de tres feromonas sexuales de tres especies de
lepidópteros cuarentenados. En la Helicoverpa armigera los compuestos identificados
fueron (z)-11-hexadecenal (98) y (z)-9-hexadecenal (2), lo cual es es coincidente con
(Chumakov M.A., and Kuznetsova T.L. 2009), ambos compuestos comparados con los
tiempos de retención de los estándares inyectados, el compuesto que de acuerdo a los autores
anteriores y Zhang, JP 2012, es mas abundante (z)-11 hexadecenal se presentó en este estudio
con una abundancia mayor y constante durante los 30 días.
En la feromona de Spodoptera litura los compuestos identificados fueron (Z,E)-9,11-
Tetredecatrienyl acetato (90) y (Z,E)-9,12-Tetradecadienyl acetato (10), lo cual es
coincidente con Sun, F.3003, se midió la tasa de liberación de la feromona, la cual se mantuvo
constante durante los 30 días, ya que el Z,E-9,12-Tetradecadienyl acetato se mantuvo en una
concentración promedio de 4.48 ng por cada uL.
Para Tuta absoluta uno de los compuestos que reporta Ferrara, 2001, (E,Z,Z)-3,8,11
Tetradecadienyl acetato se identificó durante el estudio, iniciando con una concentración de
49.16 ng por uL a los 0 días, 38.65 a los 10 días, 36.26 a los 20 días y alos 30 dias 21.52 ng
por uL.
Con los datos anteriores, se puede inferir que las feromonas comerciales de Helicoverpa
armigera, Spodoptera litura y Tuta absoluta, cumplen con la expectativa de duración de 30
días, periodo establecido en los programas fitosanitarios de detección.
AGRADECIMIENTOS
Agradecemos al Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria por
la oportunidad y tiempo para realizar esta investigación.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
138
LITERATURA CITADA
Cibrián-Tovar, J., J. Carpenter, S. Hight, T. Potter, G. Logarzo and J.C. Velázquez- González. 2017. Reinvestigation of Cactobastis cactorum (Lepidoptera: Pyralidae) Sex pheromone for improved attractiveness and greater specificity. En: V. D.C. Shields, ed. Biological control of pest and vector insects. INTECH, Ed. P. 119-131.
El-Sayed A.M. 2019. The pherobase. Database of pheromones and semiochemicals. <http://www.pherobase.com>©2003-2019.
Heath, R. R., P.E A. Teal, N.D. Epsky, B. Dueben, S. D. Hight, S. Bloem, J.E. Carpenter; T. J. Weissling; P. E. Kendra and J. Cibrián-Tovar. 2006. Pheromone-based attractant for males of Cactoblastis cactorum (Lepidoptera: Pyralidae). Environ. Entomol. 35 (6): 1469 – 1476.
Hills, A.S., R.T. Cardé, H. Kido and W.L. Roelofs. 1975. Sex pheromone of the orange tortrix moth, Argyrotaenia citrana (Lepidoptera: Tortricidae). Journal of Chemical Ecology 1(2):215-224.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
139
PREFERENCIA DE UN COCCINÉLIDO Y UN DÍPTERO POR LOS
DIFERENTES ESTADOS DE DESARROLLO DE Dactylopius opuntiae
(HEMIPTERA: DACTYLOPIIDAE)
Raquel Salas-Monzón, *Esteban Rodríguez-Leyva, J. Refugio Lomeli-Flores, Juan M. Vanegas-Rico
Posgrado en Fitosanidad, Entomología y Acarología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230
Texcoco, Estado de México, México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
En varias especies de depredadores, las larvas y los adultos prefieren alimentarse de
diferentes estados de desarrollo de la misma presa. Esta capacidad de elección puede estar
asociada a su calidad es decir a los nutrientes que pueden obtener de ella (Vanegas-Rico et
al. 2016), pero al mismo tiempo al trabajo energético que puede tomarles manejar la presa y
su ganancia neta, y a los riesgos que representa alimentarse de presas grandes que pueden
causar daño por el contra ataque; a decir verdad, generalmente la elección se debe a la
combinación de todos esos factores (Rodríguez-Leyva et al. 2019). En este sentido, la
preferencia de la presa en enemigos naturales tiene un papel importante no sólo en el
aprovechamiento de los recursos presa, sino también en la supresión de especies fitófagas
(Xu y Enkegaard, 2010).
En México se han reportado a los depredadores Hyperaspis trifurcata Schaeffer,
(Coleoptera: Coccinellidae) y Leucopis bellula Williston, (Diptera: Chamaemyiidae)
alimentándose de la cochinilla silvestre del nopal, Dactylopius opuntiae (Cockerell)
(Hemiptera: Dactylopiidae), ésta último es la plaga más importante del género Opuntia spp.
(Caryophyllales: Cactaceae), y plaga clave en Opuntia ficus-indica, que es la cactácea más
cultivada en todo el mundo (Mann, 1969; Vanegas-Rico et al., 2010).
Los depredadores H. trifurcata y L. bellula son de los más frecuentes y abundantes en
campo, además de tener una aparente relación denso-dependiente con D. opuntiae (Vanegas-
Rico et al. 2010; 2016; 2017). La utilización de estos enemigos naturales como un método
de control biológico, puede ser una alternativa para el manejo de la cochinilla silvestre del
nopal, pero se desconocen varias características de su biología y comportamiento. Hasta el

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
140
momento se sabe que H. trifurcata puede completar su ciclo cuando se alimenta de ninfas
del primer instar; sin embargo, cuando se alimentó de una colonia mixta de D. opuntiae, sus
parámetros poblaciones aumentaron (Vanegas-Rico et al. 2016). Por otro lado, no hay
información de L. bellula y se podría suponer que por su comportamiento de vivir
prácticamente debajo de colonias maduras de la presa prefiere alimentarse en mayor
proporción de estados inmaduras de D. opuntiae en lugar que de adultas (Vanegas-Rico et
al., 2010) y eso podría influenciar la manera que ambos depredadores forrajean a la presa en
campo, y podrían influir en su regulación El objetivo de este trabajo fue conocer la
preferencia de estos depredadores sobre los distintos estados de desarrollo de D. opuntiae.
MATERIALES Y MÉTODOS
Cría de Dactylopius opuntiae. La colonia de D. opuntiae se obtuvo de una cría
establecida en el Colegio de Postgraduados (CP), Texcoco, Estado de México, en condiciones
de invernadero (21 ± 10°C, 70 ± 10% HR). El aumento de la cría se realizó sobre cladodios
de nopal, Opuntia ficus-indica, de un año de edad, y se infestaron mediante la técnica
propuesta por Vanegas-Rico et al. (2016).
Cría de depredadores. Los depredadores Hyperaspis trifurcata y Leucopis bellula se
obtuvieron de crías establecidas en el laboratorio de control biológico del Posgrado en
Fitosanidad, en el Colegio de Postgraduados. Las larvas de ambas especies se mantuvieron
en cajas de Petri (Ø =9 x 1.5 cm de altura) y se alimentaron con inmaduros y adultas de D.
opuntiae. Los adultos de H. trifurcata y L. bellula se mantuvieron en jaulas entomológicas
(21 x 31 x 20 cm) cubiertas con tela organza; en la jaula se les proporcionaban gotas de miel
sobre la tela y agua en un recipiente de vidrio (50 mL). A los adultos se les ofreció cada tercer
día una colonia de todos los estados de desarrollo de D. opuntiae para favorecer su
oviposición (Vanegas-Rico et al. 2016). Las crías de los depredadores y todos los
experimentos se establecieron en cámara de cría a 25±1˚ C, 60±10% HR y 12:12 h L: O.
Pruebas de elección de L. bellula e H. trifurcata sobre hembras de D. Opuntiae. La
arena experimental para los ensayos fue de una caja de Petri (Ø =2.5 x 1.5 cm de altura). A
cada unidad se le colocó dos hembras adultas grávidas (huevos y ninfas de primer instar (N1).
Posteriormente, se introdujo de manera individual una larva del tercer instar de L. bellula o

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
141
H. trifurcata con previo ayuno (24 h) y se le permitió forrajear durante 24 h. Los ensayos se
distribuyeron empleando un diseño completamente al azar, con 20 repeticiones por
depredador. Como todas las etapas de desarrollo de D. opuntiae se incluyeron en la misma
arena experimental, cada arena se consideró como un bloque en un diseño de bloques
completamente al azar. El componente del bloque aleatorio se contabilizó mediante el análisis
de datos en un modelo lineal mixto generalizado (GLMM), con mortalidad en cada etapa de
desarrollo de D. opuntiae como efecto principal fijo, y se replicó (en ocasiones) como un
efecto de bloque aleatorio, con distribución binomial y función de enlace logit. Todos los
análisis se realizaron utilizando GenStat v. 8.0 (Payne et al., 2005).
RESULTADOS Y DISCUSIÓN
Hyperaspis trifurcata y L. bellula son enemigos naturales que se consideran agentes
potenciales de control biológico de D. opuntiae por su frecuencia, abundancia y adaptabilidad
a una gran cantidad de ambientes (Vanegas-Rico et al. 2010, 2016, 2017; Mendel et al. 2019).
No obstante, aunque se conocía que ambas especies se pueden alimentar de todos los estados
de desarrollo de la presa (Vanegas-Rico et al. 2010, 2016), se desconocían aspectos básicos
de comportamiento y preferencia sobre esos estados de desarrollo.
Leucopis bellula consumió todas las etapas de desarrollo de D. opuntiae (Cuadro 1), pero
presentó mayor proporción de consumo por hembras grávidas en comparación con huevos y
N1 de D. opuntiae (F2, 47=25.94, P=0.001). Esta preferencia por insectos adultos, quizá se
deba a que las larvas de L. bellula tienen poca movilidad y no presentan un comportamiento
activo en la búsqueda de sus presas, además esto se relaciona con la presa, que es sésil y sin
ninguna actividad defensiva (Barriault et al., 2019, Rodríguez-Leyva et al., 2019). Por otro
lado, se ha observado que las larvas de L. bellula se localizan frecuentemente en la parte
ventral de la hembra, donde también se encuentran huevos y ninfas (Vanegas-Rico et al.
2010) teniendo la oportunidad de consumir huevos y adultos (Kimberling y Price, 1996) así
como sucede en otras especies de Leucopis sp. (Barriault et al., 2019). También puede
sugerirse que probablemente la preferencia de L. bellula hacia el consumo de hembras
grávidas le ayude a obtener los nutrientes que necesita para su desarrollo de manera más
eficiente (Vanegas-Rico et al. 2016), porque hubo la disponibilidad de huevos y ninfas de

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
142
primer instar, y consumió de todos los estados de desarrollo, pero prefirió adultas (Cuadro
1).
Cuadro 1. Proporción (media ± error estándar) de huevo; ninfas 1 (N1) y hembras grávidas
de Dactylopius opuntiae consumidos por tercer instar de H. trifurcata y Leucopis bellula en
condiciones controladas 25±1˚ C, 60±10% HR y 12:12 h L: O). Proporción de estados de desarrollo de D. opuntiae consumidos
Depredador Huevo N1 Hembra grávida
Leucopis bellula 0.14±0.05 0.10±0.05 0.42±0.05*
Hyperaspis trifurcata 0.15±0.05A 0.10±0.05A 0.13±0.05A Los asteriscos indican diferencias significativas entre columnas para L. bellula. Entre columnas con la
misma letra no difieren estadísticamente entre sí para H. trifurcata.
El depredador H. trifurcata al igual que L. bellula consumió todos los estados de desarrollo
de D. opuntiae. Sin embargo, en esta especie no se encontró preferencia por alguno de ellos
(F2,54=0.56, P=0.758) (Cuadro 1). Esta diferencia en la biología y comportamiento de los dos
depredadores de la misma presa tiene implicaciones científicas y prácticas, que necesitaran
de más experimentos para responderlas. Lo primero es que aparentemente H. trifurcata es
más activo que L. bellula en campo y laboratorio (Vanegas-Rico et al. 2010, 2016, 2017)
pero consumiendo una proporción similar de los tres estados de desarrollo de la presa logra
obtener la nutrición necesaria (Cuadro 1). Esto es valioso porque el mismo depredador tuvo
mejores parámetros demográficos cuando se alimentó de una dieta mixta, que sólo de primer
instar de la misma presa (Vanegas-Rico et al., 2016), lo que podría confirmar que prefiere
comer de los tres estados de desarrollo de la presa a la vez, aparentemente sin preferencia por
un estado en particular, y así puede alcanzar su máximo potencial reproductivo (Vanegas-
Rico et al., 2016). Este comportamiento de alimentarse de todos los estados de desarrollo de
la presa también lo presenta H. polita, aunque éste si presentó preferencia por los huevos de
su presa (Farhadi et al., 2017). Por el contrario H. campestris presentó una alimentación
similar entre huevos y ninfas I de D. opuntiae (El Aalaoui et al. 2019). Se recomienda realizar
observaciones de campo para confirmar si este comportamiento en la explotación diferencial

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
143
de las presas de ambos depredadores se mantiene para entender los procesos de regulación
de D. opuntiae.
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la beca otorgada
en la maestría para la primera autora.
LITERATURA CITADA
El Aalaoui, M., R. Bouharroud, M. Sbaghi, M. El Bouhssini, y L. Hilali. 2019. Predatory potential of eleven native Moroccan adult ladybird species on different stages of Dactylopius opuntiae (Cockerell) (Hemiptera: Dactylopiidae). Boletín EPPO. 0:1-6
Barriault, S., M. Fournier, A. O. Soares, and E. Lucas. 2019. Leucopis glyphinivora, a potential aphidophagous biocontrol agent? Predation and comparison with the commercial agent Aphidoletes aphidimyza. BioControl. 64:21-31.
Farhadi Z, M. Esfandiari, M. S. Mossadegh, and P. Shishehbo. 2017. Prey stage preference and functional response of the coccinellid Hyperaspis polita Weise by feeding on the mealybug Nipaecoccus viridis (Newstead). Plant Pest Res. 7:63–78.
Kimberling, D. N., and P. W. Price. 1996. Competition, leaf morphology, and host clone effects on eeaf–galling grape Phylloxera (Homoptera: Phylloxeridae). Environmental Entomology. 25, 1147-1153.
Mann, J. 1969. Cactus-feeding insects and mites. Smithsonian Institution Press, Washington, US.
Mendel, Z., A. Protasov, J. M. Vanegas-Rico, J. R. Lomeli-Flores, P. Suma, E. Rodríguez-Leyva. 2019. Classical and fortuitous biological control of the prickly pear cochineal Dactylopius opuntiae in Israel. Biological Control, BICO-19-00. Submitted. 10-06-2019.
Payne, R.W., D. A. Murray, S. A. Harding, D. B. Baird, D. M. Soutar. 2005. GenStat for Windows Introduction. Hemel Hempstead, United Kingdom.
Rodríguez-Leyva, E., J. R. Lomeli-Flores y N. S. Gómez-Domínguez. 2019. Biología, ecología y comportamiento de insectos depredadores. Biblioteca Básica en Agricultura (BBA), en prensa.
Vanegas-Rico, J. M., E. Rodríguez-Leyva, J. R. Lomeli-Flores, H. González-Hernández, A. Pérez-Panduro, G. Mora-Aguilera. 2016. Biology and life history of Hyperaspis trifurcata feeding on Dactylopius opuntiae. BioControl. 61: 691-701.
Vanegas-Rico, J. M., J. R. Lomeli-Flores, E. Rodríguez-Leyva, G. Mora-Aguilera, J. M. Valdez. 2010. Natural enemies of Dactylopius opuntiae (Cockerell) on Opuntia ficus-indica (L.) Miller in Central México. Acta Zool. Mex. (n.s.) 26: 415-433.
Vanegas-Rico, J. M., A.Pérez-Panduro, J. R. Lomeli-Flores, E. Rodríguez-Leyva, J. M. Valdez-Carrasco y G. Mora-Aguilera. 2017. Dactylopius opuntiae (Cockerell) (Hemiptera: Dactylopiidae) population fluctuations and predators in Tlalnepantla, Morelos, Mexico. Folia Entomol. Mex. (n. s.) 3: 23−31.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
144
Xu X and A. Enkegaard. 2010. Prey preference of the predatory mite, Amblyseius swirskii between first instar western flower thrips Frankliniella occidentalis and nymphs of the two-spotted spider mite Tetranychus urticae. J. Insect Sci. 10:149

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
145
PLANTAS CON FLORES COMO ALIMENTO DE PARASITOIDES
EN EL CONTROL BIOLÓGICO DE LA PALOMILLA DORSO
DIAMANTE
Luis Felipe San Marino Cid-Aguilar1, J. Refugio Lomeli-Flores1*, Esteban Rodríguez-Leyva1
1Colegio de Postgraduados, Posgrado en Fitosanidad, Entomología y Acarología, Carretera México-Texcoco km. 36.5, Montecillo, 56230
Texcoco, Estado de México, México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
El brócoli es una de las hortalizas más importantes en Guanajuato, durante 2017 se
sembraron 23,060 ha y se produjeron 364,658 ton; esta cantidad representó el 63.4% de la
producción nacional y más del 50% se destinó a la exportación (SIAP, 2018). Plutella
xylostella Linnaeus, 1758 (Lepidoptera: Plutellidae), o palomilla dorso de diamante (PDD),
se considera la principal plaga de brócoli ya que su presencia en el producto comercial es
motivo de una disminución del valor y de rechazo de un cargamento (Bujanos et al., 2013).
Diadegma insulare (Cresson, 1865) (Hymenoptera: Ichneumonidae) es el parasitoide más
eficiente para el control biológico de P. xylostella, y el mejor distribuido y adaptado en
América (Bolter y Laing, 1983). Al igual que otros parasitoides, D. insulare prefiere
permanecer en ambientes con fuentes de alimento (flores con néctar y polen) y refugios
abundantes (Idris y Grafius, 2001), participando en el control de plagas cuando las
densidades son bajas. Un ejemplo de esto es el uso de flores de panalillo (Lobularia maritima
(L.) Desv. (Brassicaceae) en el cultivo de brócoli que, aunado a una reducción del uso de
insecticida, ha demostrado un aumento del parasitismo natural sobre P. xylostella (Tovar et
al., 2007). Considerando lo anterior, en Guanajuato se ha propuesto explorar el control
biológico por conservación como alternativa de combate de la PDD. Se piensa que la
incorporación de plantas de alysium (Lobularia maritima) pudiese favorecer ese tipo de
control biológico. Por lo anterior se planteó el presente trabajo con el objetivo de evaluar el
efecto de asociar plantas de alysium con brócoli comercial y comparar la respuesta de la
fauna benéfica y la PDD.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
146
MATERIALES Y MÉTODOS
El presente estudio se realizó en un predio del poblado “El Varal” en el municipio de
Abasolo, Guanajuato en los meses de enero a marzo de 2019. En el predio se estableció
control biológico por conservación, para ellos se asociaron plantas con flores (alysium) al
brócoli el mismo día del trasplante. Se establecieron tres tratamientos, cada tratamiento de
0.5 ha: 1) Alysium intercalado al 1% en brócoli; 2) Alysium intercalado al 3% en brócoli; y
3) Testigo regional (manejo tradicional del productor, aplicación de insecticidas de acuerdo
a recomendaciones de sus técnicos). Las plántulas de alysium las proporcionó la empresa
Gigante Verde (B&G Foods Manufacturing México S de RL DECV).
Para cada tratamiento se realizó un muestreo semanal en cinco de oros evaluando en cada
punto 10 plantas al azar, se contabilizó, recolectó y registró todos los individuos de la PDD.
Adicionalmente, se realizaron 20 redeos por tratamiento. Los insectos colectados se
colocaron en bolsas de plástico herméticas y se agregó etanol 70% para fijarlos.
Las larvas de la PDD recolectadas de campo se llevaron al laboratorio, allí se colocaron
en cajas Petri y se alimentaban con follaje inmaduro de brócoli cada 2 días. Los especímenes
emergidos se separaron utilizando un microscopio estereoscópico y se registró el número de
especímenes por especie de cada muestra. Los parasitoides se procesaron e identificaron
mediante claves específicas de cada grupo. Con los valores de número de larvas por planta
se describió la fluctuación poblacional, también se registró a los enemigos naturales
presentes.
Análisis estadístico. Para observar si había algún efecto de los tratamientos en la densidad
poblacional de la PDD, se compararon los números totales de los estados inmaduros por cada
10 plantas muestreadas. Además, se calculó el porcentaje de parasitismo entre los
tratamientos por fecha de muestreo, y se compararon mediante un análisis de varianza;
cuando se encontraron diferencias entre tratamientos se aplicó una prueba de separación de
medias LSD (p ≤ 0.05). Para cada uno de estos análisis estadísticos se usó el paquete
estadístico Statistix 8.1.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
147
RESULTADOS Y DISCUSIÓN
Las poblaciones de la PDD no alcanzaron el umbral de acción fijado para la zona (0.5
larvas/planta) en las primeras siete semanas del cultivo. Las primeras poblaciones de la plaga
se presentaron a niveles muy bajos en la tercera semana después del trasplante. Aunque se
presentó un pico de la PDD en la tercera semana de febrero en el lote con 3% de plantas, la
cual fue superior a lo encontrado en los otros tratamientos, estas poblaciones bajaron
significativamente la semana siguiente. A pesar de los niveles bajos de la PDD, el total de
inmaduros recolectados fue casi tres veces mayor en el lote del testigo regional en
comparación al que contenía 1% de flores en la última semana de febrero y las primeras dos
de marzo. Finalmente, el productor decidió realizar acciones de control la segunda semana
de marzo mediante la aplicación de un producto químico lo que provocó que los niveles de
plaga disminuyeran a casi cero en los siguientes dos muestreos.
El único parasitoide de la PDD recolectadas en los lotes experimentales fue Diadegma
insulare, con porcentajes de parasitismo entre 0 y 100% sin observarse una tendencia clara
entre los tratamientos. No obstante, se puede decir que el porcentaje de parasitismo fue
ligeramente mayor en parcelas con 1% de Alysium en la primera y segunda semana de marzo.
Considerando los tres tratamientos y todas las fechas de colecta se recuperaron en total 100
larvas y/o pupas de la PDD y de ellas el 18% estaban parasitadas. La presencia de plantas de
Alysium dentro de los tratamientos ocasionó una tendencia en la densidad poblacional de la
PDD. El total de inmaduros recolectados de la PDD fue hasta tres veces mayor en el lote sin
presencia de flores (52 inmaduros de la PDD) en comparación al lote con 1% de Alysium
(14 inmaduros), esta diferencia se registró en tres semanas. A pesar de esas diferencias en
densidad población en dichas semanas, sólo se detectó una pequeña tendencia a
incrementarse el parasitismo en las dos primeras semanas de marzo. Ese parasitismo no
superó el 29% en los lotes con Alysium. Ese porcentaje de parasitismo fue semejante a lo
reportado en la región en una temporada similar sin plantas de alysium (McCully y Salas-
Araiza, 1990), pero aparentemente superior al parasitismo en el tratamiento sin presencia de
Alysium.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
148
Con las limitaciones de haber hecho el estudio en condiciones no aptas (baja incidencia
de PDD) y sin repeticiones, se pudiera sugerir que la inclusión de plantas con flores pudiera
favorecer la presencia de parasitoides y/o depredadores de la PDD, como se ha señalado en
otros trabajos (Olson et al., 2005; Wade et al., 2008); sin embargo, al utilizar plantas con
flores de la misma familia botánica del cultivo a proteger, en este caso Alysium en brócoli,
pudiera traer consecuencias negativas. Un indicio de que esto pudo haber ocurrido en los
lotes con 3% de Alysium, donde se presentó una ligera tendencia a mayores densidades de la
plaga, aunque estadísticamente no hubo diferencia con los lotes con de 1% de Alysium.
El Alysium se ha usado experimentalmente como fuente de néctar y polen en diversos
programas de control biológico por conservación de la PDD y su parasitoide D. insulare
(Johanowicz y Mitchell, 2000; Lavandero et al., 2005), Estos autores mencionan la
importancia de esa planta como fuente de néctar y polen para enemigos naturales, lo que
incrementó su permanencia en campo y los niveles de control. Sin embargo, la utilización de
Alysium en brócoli debe tomarse con cautela (Winkler et al., 2010). La PDD se pudiera
reproducir en ella, o los adultos de esta plaga pudieran obtener alimento de esta planta con lo
que incrementaría su potencial biótico. Se debe continuar con la investigación confirme si la
PDD realmente prefiere poner huevos en Alysium comparado con brócoli (lo que podría
convertirla en una planta trampa), y que disminuye la sobrevivencia de las larvas que se
alimentan en Alysium. Esto depende de las variedades de Alysium y de brócoli, pero es algo
que se tiene que responder antes de poder recomendar la asociación de esas plantas con
intención de mejorar el control biológico por conservación.
A manera de conclusión, se puede decir que, si bien la inclusión de plantas de Alysium en
el cultivo de brócoli puede tener un efecto positivo en la permanencia y parasitismo de D.
insulare sobre la PDD, se deben realizar más estudios para determinar cuál es la variedad
más conveniente y la densidad óptima de plantas con flores asociadas en el brócoli para
favorecer la presencia de enemigos naturales, y en qué periodo puede ser útil esa propuesta
de manejo en el cultivo.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
149
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la beca otorgada
en la maestría para el primer autor.
LITERATURA CITADA
Bolter C. J., Laing J. E.1983. Competition between Diadegma insulare (Hymenoptera: Ichneumonidae) and Microplitis plutellae (Hymenoptera: Braconidae) for larvae of Diamondback moth Plutella xylostella (L.) (Lepidoptera: Plutellidae). Proceedings of the Entomological Society of. Ontario.114:1-10.
Bujanos R., Marín Jarillo A., Díaz Espino L. F., Gámez Vázquez A. J., Ávila Perches M. A., Herrera Vega R., Dorantes González R. A. y Gámez Vázquez F., P. 2013. Manejo integrado de la palomilla dorso de diamante Plutella xylostella (L.) en la región del Bajío, México. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Folleto Técnico Núm. 27. 40 p.
Idris, A. B., and E. Grafius. 2001. Evidence of Diadegma insulare (Cresson), a parasitoid of diamondback moth, Plutella xylostella (L.), present in various habitats. Pakistan Journal of Biological Sciences 1: 742-743.
Johanowicz, D. L., and E. R. Mitchell. 2000. Effects of sweet alyssum flowers on the longevity of the parasitoid wasps Cotesia marginiventris (Hymenoptera: Braconidae) and Diadegma insulare (Hymenoptera: Ichneumonidae). Florida Entomologist 83: 41-47.
Lavandero, B., S. Wratten, P. Shishehbor, and S. Worner. 2005. Enhancing the effectiveness of the parasitoid Diadegma semiclausum (Helen): movement after use of nectar in the field. Biological Control 34: 152-158.
Olson, D., K. Takasu, and W. J. Lewis. 2005. Food needs of adult parasitoids: Behavioral adaptations and consequences. Pp. 137-147. In: F. L. Wackers, P. C. J. van Rijn and J. Bruin (Eds.). Plant-provided food for carnivorous insects: a protective mutualism and its applications. Cambridge University Press.
SIAP, 2018. Resumen de producción agrícola por estado: Brócoli, 2017. Sistema de Información Agroalimentaria y Pesquera (SIAP). SAGARPA. Fuente: http://infosiap.siap.gob.mx:8080/agricola_siap_gobmx/AvanceNacionalCultivo.doF fecha de consulta: 10 de octubre de 2019.
Tovar Hernández, H., N. Bautista Martínez, J. Vera Graziano, A. D. Suárez Vargas, y S. Ramírez Alarcón. 2007. Fluctuación poblacional y parasitismo de larvas de Copitarsia decolora Guenée, Plutella xylostella L. y Trichoplusia ni Hübner (Lepidoptera) en Brassica oleracea L. Acta Zoológica Mexicana 23: 183-196.
Wade, M. R., M. P. Zalucki, S. D. Wratten, K. A. Robinson. 2008. Conservation biological control of arthropods using artificial food sprays: Current status and future challenges. Biological Control 45: 185–199.
Winkler, K., F.L. Wackers, A.J. Termorshuizen and J. C. van Lenteren. 2010. Assessing risks and benefits of floral supplements in conservation biological control. BioControl 55: 719–27.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
150
ANÁLISIS DE RIESGO DE PLAGAS PARA PREVENIR LA
INTRODUCCIÓN DE Acidovorax citrulli A MÉXICO
Vícente Rosas Medina1*, Ana María Hernández Anguiano1, Cristian Nava Díaz1, José Gustavo
Torres Martínez2
1 Posgrado en Fitosanidad-Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco,
Estado de México, México. 2 Dirección del Cemtro Nacional de Referncia Fitosanitaria, de la Dirección General de Sanidad Vegetal del
SENASICA, Km 37.5 Carretera Federal México-Pachuca, Tecamác, Edo. de México. C.P. 554074. *Autor de correspondencia:
INTRODUCCIÓN
El Análisis de Riesgo de Plagas (ARP) de este trabajo tiene por objeto determinar el riesgo
fitosanitario que representa la bacteria Acidovorax citrulli en la importación de material
vegetal propagativo (semilla botánica y plántulas) para siembra de especies de hospedantes
primarios: sandía (Citrullus lanatus) y melón (Cucumis melo), así como de hospedantes
secundarios: pepino (Cucumis sativus) y calabacita (Cucurbita pepo), originarias de países
para los cuales se tienen establecidas medidas fitosanitarias aplicables para su importación a
México.
Se realizó la búsqueda, categorización, análisis y síntesis de información técnico-científica
en bases de datos, normatividad nacional e internacional, para documentar y hacer el ARP
con base a lo establecido en la Norma Internacional sobre Medidas Fitosanitarias NIMF No.
11 Análisis de Riesgo de Plagas para Plagas Cuarentenarias, de la Convención Internacional
de Protección Fitosanitaria (IPPC, por sus siglas en inglés). Derivado del proceso de ARP,
se determinó que Acidovorax citrulli es una plaga cuarentenaria para México y que se
encuentra asociada a la vía de importación de material vegetal propagativo (semillas y
plántulas) de especies hospederas de la bacteria.
MATERIALES Y MÉTODOS
El Análisis de Riesgo de Plagas se realizó conforme lo establece la normatividad
internacional, tomando como referencia el Acuerdo sobre la Aplicación de Medidas
Sanitarias y Fitosanitarias (AMSF) de la OMC y la Norma Internacional para Medidas

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
151
Fitosanitarias NIMF No. 11.- Análisis de riesgo de plagas para plagas cuarentenarias, la
cual ofrece el proceso para determinar si una plaga es de interés cuarentenario para un área
o un país. El ARP consideró el desarrollo de las etapas marcadas en la citada NIMF 11:
Etapa 1 (inicio del proceso) consiste en la identificación de plagas y las vías que suscitan
preocupación, en relación con el área de ARP identificada.
Etapa 2 (evaluación del riesgo) comienza con la categorización de las plagas individuales
para determinar si se cumplen los criterios para incluirlas entre las plagas cuarentenarias. La
evaluación del riesgo continúa con una valoración de la probabilidad de entrada,
establecimiento y dispersión de la plaga y de sus consecuencias económicas potenciales.
Etapa 3 (manejo del riesgo) consiste en determinar opciones con respecto al manejo para
reducir los riesgos identificados en la etapa 2. Esas opciones se evalúan en función de su
eficacia, viabilidad y repercusiones con el fin de seleccionar las que son apropiadas.
RESULTADOS Y DISCUSIÓN
Acivodorax citrulli no se encuentra establecido en México. No se encontraron
documentos con calidad de registros que refieran la presencia de la bacteria en México.
Identificación de hospedantes primarios y secundarios de Acivodorax citrulli.
Hospederos primarios: Sandía (Citrullus lanatus) y melón (Cucumis melo). Hospederos
secundarios: Pepino (Cucumis sativus), calabaza moschata (Cucurbita moschata) y
calabacita (Cucurbita pepo) (Isakeit et al., 1997; O'Brien y Martin, 1999; Walcott et al.,
2000; Liu et al., 2012; Horuz et al., 2014; Li et al., 2014; Langston et al., 1999; Martin et
al., 1999).
Asociación con la vía de importación. Se identifica la asociación de la bacteria con
semillas y material vegetal propagativo (plantas) (Elizalde et al., 2011; CAB International, 2018;
Walcott, 2008; Kucharek et al, 1993; Hopkins et al., 1996).
Distribución Mundial de Acidovorax citrulli. Los países en los cuales se tiene detectada
la presencia de la bacteria Acivodorax citrulli son: Asia: China (Wang et al., 2009; Liu et al.,

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
152
2012), Israel, Japón, República de Corea, Malasia, Taiwán, Tailandia (CAB International,
2018) y Turquía (Mirik et al., 2006; Horuz et al., 2014). Norteamérica: Canadá (CAB
International, 2018) y Estados Unidos de Norteamérica (Isakeit et al., 1997; Walcott et al.,
2000, CAB International, 2018). Centroamérica y el Caribe: Costa Rica y Trinidad y Tobago
(CAB International, 2018). Suramérica: Brasil (Marques et al., 2015; CAB International,
2018)). Europa: Grecia (CAB International, 2018), Hungría (Palkovics et al., 2008), Italia y
Serbia (Popović e Ivanović, 2015; CAB International, 2018). Oceanía: Australia (Martin et
al., 1999; O'Brien y Martin, 1999), Guam e Islas Marianas del Norte (CAB International,
2018).
Material vegetal propagativo de importación. En el Modulo de Requisitos
Fitosanitarios para la Importación, existen claves de combinación para el ingreso de semillas
y plántulas de especies hospederas de Acidovorax citrulli, procedentes de países donde está
presente la bacteria, en las cuales actualmente no se tienen establecidas medidas fitosanitarias
aplicables para prevenir el ingreso de esta bacteria (SENASICA, 2018).
Medidas Fitosanitarias para el ingreso de material vegetal propagativo. Se determina
la necesidad de que las Autoridades Fitosanitarias de México (SENASICA) lleven a cabo
una actualización de las medidas fitosanitarias aplicables para el ingreso de material vegetal
propagativo (semillas y plántulas), provenientes de países donde se identificó la presencia de
Acidovorax citrulli.
Identificación de medidas fitosanitarias para el manejo del riesgo de Acidovorax
citrulli. Se proponen medidas fitosanitarias de mitigación del riesgo que pueden ser aplicadas
al material vegetal propagativo (semillas y plántulas) de especies hospederas de Acidovorax
citrulli, cuando provengan de países con presencia de la bacteria.
AGRADECIMIENTOS
Al Colegio de Postgraduados, Campus Montecillo, y la Dirección General de Sanidad
Vegetal del SENASICA, por el apoyo brindado con equipo e instalaciones para la realización

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
153
del trabajo, así como la coordinación y asesoría de sus Profesores y personal técnico
especializado.
LITERATURA CITADA
CAB International. 2018. Crop Protection Compendium. Wallingford, UK: CAB International. En línea: http://www.cabi.org/cpc/
Elizalde J., N. A., Hernandez M., J., Leyva M., S. G., Nava D., C., Sequeria, R. A., Fowler, G., Magarey, R. 2011. Evaluación del riesgo de Acidovorax avenae subsp. citrulli asociadas a semilla de sandía de importación a México. Revista Mexicana de Fitopatologia, 29(2): 133.145.
Hopkins, D., Stall, B., Kucharek, T., Gay, D., Gitaitis, R., Cook, W., Keinath, A., and Latin, R. 2000. Bacterial fruit blotch of watermelon. Special Interstate Cooperative Publication SICP-1. En línea: http://plantpath.ifas.ufl.edu/media/plantpathifasufledu/factsheets/sicp1.pdf
Horuz, S., Cetinkaya-Yildiz, R., Mirik M., and Aysan, Y. 2014. Ocurrence, isolation, and identification of Acidovorax citrulli from melon in Turkey. Plant Protection Science, 50(4):179-183. En línea: http://www.agriculturejournals.cz/publicFiles/136824.pdf
Isakeit, T., Black, M. C., Barnes, L. W., Jones, J. B. 1997. First report of infection of honeydew with Acidovorax avenae subsp. citrulli. Disease Notes. Plant Disease, 81(6):694. En línea: http://apsjournals.apsnet.org/doi/abs/10.1094/PDIS.1997.81.6.694C
IPPC. 2017. Norma Internacinal para Medias Fitosanitarias (ISPM) 11 Análisis de Riesgo de Plagas para Plagas Cuarentenarias. International Plant Protection Convention (IPPC). En línea: https://www.ippc.int/es/publications/639/
Kucharek T, Perez Y, Hodge C, 1993. Transmission of the watermelon blotch bacterium from infested seed to seedlings. Phytopathology, 83:467.
Langston, D. B., Walcott, R. D. Jr., Gitaitis, R. D., and Sanders, F. H. Jr. 1999. First report of a fruit rot of pumpkin caused by Acidovorax avenae subsp. citrulli in Georgia. Disease Notes. Plant Disease, 83(2):199. En línea: http://apsjournals.apsnet.org/doi/abs/10.1094/PDIS.1999.83.2.199B
Liu, J., Luo, S. Z., Zhang, Q., Wang, Q. H., Chen, J. F., Guo, A. G., and Shan, W. X. 2012. Tn5 transposon mutagenesis in Acidovorax citrulli for identification of genes required for pathogenicity on cucumber. Plant Pathology, 61:364-374. En línea: http://onlinelibrary.wiley.com/doi/10.1111/j.1365-3059.2011.02519.x/epdf
Li, J-F., Lin, T., and Ma, G-B. 2014. Effects of bacterial fruit blotch pathogen on treated melon seed. 7-9 pp. In: Cucurbitaceae 2014 Proceedings. Havey, M., Weng, Y., Day, B., and Grumet, R. (eds.). Bay Harbor, Michigan, USA. 204 p. En línea: http://cuke.hort.ncsu.edu/cgc/conferences/cuc2014proceedings.pdf
Marques S., K. M., Silva J., A., Siqueira G., M. A., Bernardi L., N., Pereira L., M. do C. C., Ramos M., R. L., Barbosa S., E. 2015. Polyphasic analysis of Acidovorax citrulli strains from northeastern Brazil. Scientia Agricola, 73(3):252-259. En línea: http://www.scielo.br/pdf/sa/v73n3/0103-9016-sa-73-3-0252.pdf

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
154
Martin, H. L. O’Brien, R. G., and Abbott D. V. 1999. First report of Acidovorax avenae subsp. citrulli as a pathogen of cucumber. Disease Notes. Plant Disease, 83(10):965. En línea: http://apsjournals.apsnet.org/doi/abs/10.1094/PDIS.1999.83.10.965D
Mirik, M., Aysan, Y., and Sahin, F. 2006. Ocurrence of bacterial fruit blotch of watermelon caused by Acidovorax avenae subsp. citrulli in the Eastern Mediterranean región of Turkey. Disease Notes. Plant Disease, 90(6):829. En línea: http://apsjournals.apsnet.org/doi/abs/10.1094/PD-90-0829C
O'Brien, R. G., and Martin, H. L. 1999. Bacterial blotch of melons caused by strains of Acidovorax avenae subsp. citrulli. Australian Journal of Experimental Agriculture, 39(4):479-485. Abstract. En línea: http://www.cabi.org/cpc/abstract/19991006151
Palkovics, L., Petróczy, M., Kertész, B., Németh, J., Bársony, Cs., Mike, Zs., and Hevesi, M. 2008. First report of bacterial fruit blotch of watermelon caused by Acidovorax avenae subsp. citrulli in Hungary. Disease Notes. Plant Disease, 92(5):834. En línea: http://apsjournals.apsnet.org/doi/abs/10.1094/PDIS-92-5-0834C
Popović, T., and Ivanović, Ž. 2015. Ocurrence of Acidovorax citrulli causing bacterial fruit blotch of watermelon in Serbia. Disease Notes. Plant Disease, 99(6):886. En línea: https://apsjournals.apsnet.org/doi/full/10.1094/PDIS-12-14-1276-PDN
SENASICA. 2018. Módulo de consulta de requisitos fitosanitarios para la importación de mercancías de origen vegetal. Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria (SENASICA). En línea: http://sistemas.senasica.gob.mx/mcrfi/
Walcott, R. R., Langston D. B. Jr., Sanders, F. H. Jr., and Gitaitis, R. D. 2000. Investigating intraspecific variation of Acidovorax avenae subsp. citrulli using DNA fingerprinting and whole cell fatty acid analysis. Phytopathology, 90(2):191-196. En línea: http://apsjournals.apsnet.org/doi/pdf/10.1094/PHYTO.2000.90.2.191
Wang, X., Li, G., Jiang, D., and Huang, H-C. 2009. Screening of plant epiphytic yeasts for biocontrol of bacterial fruit blotch (Acidovorax avenae subsp. citrulli) of hami melón. Biological Control, 50:164-171.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
155
MODELO GEOESPACIAL DE RIESGO FITOSANITARIO: Xylella
fastidiosa subsp. fastidiosa EN MÉXICO
Maria Margarita Oliva-Hurtado1, Daniel Téliz-Ortiz1*, Laura Delia Ortega-Arenas1, Andrés
Quezada-Salinas2
1Posgrado en Fitosanidad-Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco,
Estado de México, México. 2SENASICA- Centro Nacional de Referencia Fitosanitaria, Carretera Federal México-Pachuca Km 37.5,
Técamac, Estado de México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
En México se destinan 36 654 ha para el cultivo de uva. Baja California es el tercer estado
productor de uva de mesa y el primer estado con mayor producción de uva industrial (SIAP,
2019). El municipio de Ensenada cuenta con 166 de las 195 unidades de producción de vid
de la entidad; la producción de vino es de suma importancia para la economía del estado, esta
posee una calidad internacional reconocida con más de 400 premios y reconocimientos
internacionales (Consejo Mexicano Vitivinícola, 2010). En 1995 el INIFAP detectó por
primera vez en Ensenada la presencia de Xylella fastidiosa en plantas de Vid. Los principales
hospedantes de Xylella fasdtidiosa subsp. fastidiosa reportados por EFSA (2018) son: vid
(Vitis vinífera L.), alfalfa (Medicago sativa L.), maple (Acer spp.), almendro (Prunus dulcis
Mill., L.), café (Coffea arabiga), durazno (Prunus persica), y naranja (Citrus cinensis). Esta
enfermedad ha causado pérdidas de rendimiento del 20-40% en almendros de California,
EUA (Sisterson et al., 2012), del cual el 9 % de los árboles infectados murieron en el periodo
de ~7 años. En el cultivo de café en Brasil Rocha et al. (2006) indica que un aumento de 1 %
en la incidencia de la enfermedad causa una pérdida de hasta 1.28 bolsas (60 kg) de café por
hectárea.
Entre los principales vectores de la Enfermedad de pierce se encuentra Homalodisca
liturata y Xyphon fulgidum, presentes en el estado de Baja California. A partir del 2003 el
Comité Estatal de Sanidad Vegetal de Baja California implementó la campaña contra la
Enfermedad de Pierce, con actividades encaminadas a reducir el riesgo que representa la
enfermedad para la actividad vitivinícola en todas las zonas productoras de Vid en Baja
California (CESAVEBC, 2015). El presente estudio tiene como objetivo desarrollar un

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
156
modelo geoespacial fitosanitario de Xylella fastidiosa subsp. fastidiosa en Baja California,
México para identificar el patrón de dispersión de los cicadelidos transmisores de la bacteria
y poder identificar las zonas vitícolas de bajo riesgo de establecimiento de la enfermedad.
MATERIALES Y MÉTODOS
Material. Se documentó la biología del patógeno y sus dos vectores principales
reportados en Baja California: Homalodisca liturata y Xyphon fulgidum de artículos
científicos internacionales y nacionales, así como documentos emitidos por diversas
instancias gubernamentales de los países con presencia de la bacteria. A partir de esto se
generaron capas de la información (shapefiles) de temperatura, superficie sembrada de
hospedantes, detecciones de la bacteria y captura de vectores (registros de campaña de la
DGSV) y un modelo biológico MAXENT de acuerdo a los parámetros de reproducción y
supervivencia documentados.
Al generar las capas de información georreferenciada se elaboraron diferentes modelos:
establecimiento; hospedantes: agrícolas, forestales y silvestres; distribución potencial (índice
de similitud climática MAXENT); condiciones climatológicas óptimas para la bacteria y
hospedantes silvestres de los vectores. Y el modelo de dispersión local: mediante la
abundancia estacional de los vectores.
RESULTADOS Y DISCUSIÓN
A nivel nacional se cuantificó un total de 3,932,247.04 hectáreas de cultivos susceptibles
a Xylella fastidiosa subsp. fastidiosa: vid (Vitis vinífera L.), alfalfa (Medicago sativa L.),
café (Coffea arabiga), durazno (Prunus persica), naranja (Citrus cinensis), romero
(Rosmarinus officinalis L.). Así también especies forestales como maple (Acer spp.),
almendro (Prunus dulcis Mill., L.), nogal (Juglans regia L.), magnolia (Magnolia
grandiflora L.), rosa laural (Nerium oleander L.), sauco (Sambucus sp.), Spartium junceum
L., y un total de 26 especies silvestres distribuidas en 7,695 sitios. A nivel estatal en Baja
California se cuantificaron 34 271 hectáreas de cultivos susceptibles de vid (Vitis vinífera
L.), alfalfa (Medicago sativa L.), durazno (Prunus persica), naranja (Citrus cinensis) y un

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
157
total de 19 especies silvestres distribuidas en 189 sitios. Cabe destacar que el municipio de
Ensenadas es uno de los principales municipios con mayor densidad de sitios con
hospedantes silvestres, al igual que Baja California Sur, Sonora, Chihuahua, Durango,
Sinaloa, Querétaro, Michoacán, Estado de México y Campeche (SIAP, 2019; Instituto de
Biología, 2019; Villaseñor y Espinoza, 1998; CONABIO, 2019; Vibrans, 2012; Rzedowski
y Rzedowski, 2004 y Herbanwmex, 2019.
Se corrió el modelo de distribución potencial (MAXENT) para Xylella fastidiosa subsp.
fastidiosa a nivel nacional; este arrojó que el 70% del territorio nacional cuenta con altas
condiciones similares de temperatura y precipitación con los sitios reportados con presencia
de la bacteria a nivel mundial (GBIF, 2019; IVIA, 2017; SENASICA, 2019). El estado de
Baja California presenta un alto índice de similitud de 0.5 a 0.9 en la mayor parte del estado,
así mismo los estados que presentan un alto nivel de similitud son Sonora, Sinaloa, Baja
California Sur, Coahuila, Veracruz, Oaxaca, Campeche, Yucatán, Durango, Nayarit y
Zacatecas; el resto de los estados presentan una menor similitud climática.
Aunado a esto se creó el modelo de condiciones climáticas óptimas para el desarrollo de
la bacteria, lo que arrojó que la parte centro y norte del estado de Baja California cuenta con
temperaturas máximas normal óptimas para su desarrollo; asimismo el 80% del territorio
nacional presenta esta condición, a su vez, en condiciones de temperatura mínima normal la
mayor parte del estado de Ensenada presentan condiciones mínimas óptimas; el 70% del
territorio nacional presenta esta misma condición, exceptuando la parte centro y norte de
México (Sierras y Llanuras del Norte) que presentan condiciones por debajo de las óptimas
mínimas. En menor proporción del territorio nacional, el 20%, presenta temperaturas
superiores al umbral máximo de desarrollo de la bacteria, entre 35°C y 43°C. Los estados
con estas condiciones son: Sonora, Baja California Sur, Sinaloa, Chihuahua, Coahuila,
Nuevo León, Michoacán, Campeche y Yucatán.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
158
AGRADECIMIENTOS
Agradecemos al Colegio de Postgraduados, Campus Montecillo y a la DGSV del
SENASICA por el equipo y las instalaciones donde se realizó la investigación y la asesoría
de sus Profesores.
LITERATURA CITADA
CESAVEBC, 2015. Comité Estatal de Sanidad Vegetal de Baja California. Campaña contra la Enfermedad de Pierce. Informe de actividades correspondientes al mes de marzo. En línea: http://www.cesavebc.com/CAMPANAS/INFORMES%20MENSUALES/PIERCE/ MARZO.pdf
CONABIO, 2019. Especies. En línea: http://enciclovida.mx/explora-por-region Consejo Mexicano Vitivinícola, 2010. Uva y vino. En línea: https://uvayvino.org.mx/ EFSA, 2018. European Food Safety Authority (EFSA). Update of the Xylella spp. host plant
database. En linea: www.efsa.europa.eu/efsajournal GBIF, 2019. Xylella fastidiosa. En línea: https://www.gbif.org/species/3222355 Herbanwmex, 2019. Red de Herbarios del Noroeste de México. En linea:
http://herbanwmex.net/portal/taxa/index.php?taxon=Conium+maculatum&formsubmit=Search+Terms
Instituto de Biología (IBUNAM). 2019. Plantas Vasculares. En Portal de Datos Abiertos UNAM, México, en linea: http://datosabiertos.unam.mx/biodiversidad/
Rocha, J. G.; Zambolim, F.; Vale, F. X. R.; Zambolim, E. M.; Bergamin Filho, A.; Jesus Júnior, W. C. y Hau, B. (2006): Quantificação dos danos causados por Xylella fastidiosa em cafeeiro; Summa Phytophatologica (32 S); pp. 90-91.
Rzedowski, G. C. y Rzedowski, J. 2004. Manual de malezas de la región de Salvatierra, Guanajuato. 315 p. In: Flora del Bajío y Regiones Adyacentes. Rzedowski, G. C. y Rzedowski, J (eds). Instituto de Ecología. Fascículo XX. México.
SENASICA, 2019. Estrategia operativa de la campaña contra Enfermedad de Pierce (Xylella fastidiosa subsp. fastidiosa).
SIAP, 2019. Cierre de la producción agrícola. En línea: https://nube.siap.gob.mx/cierreagricola/
Sisterson, M. S.; Ledbetter, C. A.; Chen, J. C.; Higbee, B. S.; Groves, R. L. y Daane, K. M. 2012): Management of almond leaf scorch disease: long-term data on yield, tree vitality, and disease progress; Plant Disease (96); pp. 1037-1044.
Vibrans, H. 2012. Malezas de México. CONABIO. En línea: http://www.conabio.gob.mx/malezasdemexico/2inicio/home-malezas-mexico.htm
Villaseñor R., J. L: y Espinosa G., F. J. 1998. Catálogo de malezas de México. 1ª ed. UNAM, CONACOFI y Fondo de Cultura Económica. México. 449 p.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
159
IDENTIFICACIÓN, HOSPEDANTES Y DAÑOS DE Delia spp.
(DIPTERA: ANTHOMYIIDAE) EN CULTIVOS DE CRUCÍFERAS EN
MÉXICO
Ricardo Meraz Álvarez1; Néstor Bautista Martínez1*; Carlos Patricio Illescas Riquelme2; Héctor
González Hernández1; Ángel Lagunes Tejeda1 y Rafael Bujanos Muñiz3
1Colegio de Postgraduados, Posgrado en Fitosanidad en Entomología y Acarología, Carretera México-Texcoco Km. 36.5, Montecillo,
Texcoco 56230, Estado de México; 2Centro de Investigación en Química Aplicada, Blvd. Enrique Reyna Hermosillo No.140 C.P. 25294
Saltillo, Coahuila; 3Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Carretera Celaya-San Miguel de Allende,
Celaya, Guanajuato. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
El cultivo de crucíferas en México es altamente rentable, ya que en algunas regiones como
el Bajío, éstas se consideran las hortalizas de mayor importancia socioeconómica, debido a
la superficie sembrada; por ejemplo, tan solo en Guanajuato con 24, 885.61 ha de brócoli y
1, 226.38 ha de coliflor, de acuerdo con el SIAP, (2018), así como por los ingresos que se
obtienen de su comercialización y por la gran cantidad de fuentes de trabajo que generan
directamente en las labores propias del cultivo en el campo, en particular de los cultivos de
brócoli y coliflor, con alrededor de 110 jornales de trabajo por hectárea al año, e
indirectamente, por la cantidad de personal que se ocupa durante el proceso, empaque y
transporte. Adicionalmente, el producto que se cosecha del cultivo de crucíferas se destina
principalmente al mercado de exportación, lo que representa una significativa fuente de
divisas para el país (Agenda Técnica Agrícola Guanajuato, 2017).
En los últimos años, en distintas regiones del país se han presentados brotes importantes
de la mosca de la raíz de las crucíferas del género Delia; sin embargo, la información que
existe sobre la identidad de este insecto no está suficientemente respaldada y solo se ha hecho
a nivel de género. Al respecto, existen reportes del estado de Guanajuato en donde se
menciona que moscas el género Delia se encuentran atacando cultivos como maíz y frijol
(Marín, 2001). En la región de Acatzingo, Puebla, en el ciclo primavera-verano del año 2000,
se detectó a esta plaga en el cultivo de col y se identificó como Hylemya (= Delia) sp. (Barrios
et al., 2004).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
160
En virtud de que en el manejo integrado de cualquier plaga es fundamental el diagnóstico
confiable y dada la importancia económica de este grupo de insectos, los objetivos de la
presente investigación fueron: identificar las especies de Delia en cultivos de brócoli
(Brassica oleracea var. italica), col (B. oleracea var. capitata) y coliflor (B. oleracea var.
botrytis) en México; así como determinar los tipos de daños que ocasionan y su rango de
hospedantes.
MATERIALES Y MÉTODOS
Las colectas para determinar presencia de Delia en crucíferas cultivadas y silvestres, se
realizaron de junio 2017 a diciembre de 2018, en los estados de Guanajuato, Puebla y Estado
de México. Los cultivos incluidos en las colectas fueron brócoli (Brassica oleracea L. var.
italica), col (B. oleracea var. capitata) y coliflor (B. oleracea var. botrytis); así como, nabo
(Brassica napus L.), rábano (Raphanus sativus L.) y otras crucíferas silvestres como rábano
silvestre (Raphanus raphanistrum L.), nabo de campo (Brassica campestris L.) y mostacilla
(Sisymbrium irio L.). En el caso de las crucíferas cultivadas, en cada predio se seleccionaron
entre 10 y 15 plantas con síntomas de marchitamiento, plantas contiguas con menos
desarrollo y algunas con apariencia sana. Para el caso de crucíferas silvestres, las plantas se
seleccionaron al azar dentro y en los márgenes externos del cultivo comercial, éstas
generalmente no presentaban síntomas de marchitamiento, por lo que el tamaño de muestra
varió desde 5 hasta 20 plantas, dependiendo de su abundancia dentro del cultivo, como
consecuencia del control de la maleza. Las plantas inspeccionadas en las que se detectó la
presencia de larvas de Delia se extrajeron completamente junto con el suelo adherido a las
raíces. Posteriormente, todas las plantas colectadas en un mismo predio, se conservaron en
botes de plástico de 2 a 3 L de capacidad y se etiquetaron por localidad, fecha y hospedante,
tratando de separar entre hospedante cultivado y silvestre. El material se trasladó al
Laboratorio de Entomología Agrícola del Colegio de Postgraduados, Campus Montecillo,
Texcoco, Estado de México, para su confinamiento. Los botes con las muestras se
mantuvieron en una cámara de cría con una temperatura de 26 ± 2°C, humedad relativa de
60 ± 20% y un fotoperiodo de 12:12 (luz: oscuridad) hasta la emergencia de los adultos. Los
adultos emergidos se contabilizaron y se separaron por sexo para cada muestra.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
161
Identificación de especies. La identificación se realizó en el Laboratorio de Entomología
Agrícola del Colegio de Postgraduados, Campus Montecillo; sobre machos adultos y se
tomaron como referencia las claves de Darvas y Szappanos (2003) y Savage et al. (2016).
Para soportar la identificación de las especies, se extrajo la genitalia de machos y se
fotografiaron con un Photomicroscope III Carl ZeissÒ (Carl Zeiss, Alemania). También para
corroborar la identidad de las especies se enviaron ejemplares adultos a la Dra. Jade Savage,
de Bishop´s University en Quebec, Canadá, especialista en este grupo de insectos. También,
se proporcionaron ejemplares adultos (hembras y machos) y algunos estados inmaduros de
ambas especies al Centro Nacional de Referencia Fitosanitaria, de la Dirección General de
Sanidad Vegetal, SENASICA, Tecámac Estado de México. De igual manera, se depositaron
ejemplares adultos y larvas en la Colección Entomológica del Colegio de Postgraduados,
Campus Montecillo.
RESULTADOS Y DISCUSIÓN
Las especies identificadas fueron Delia planipalpis Stein y Delia platura Meigen (Figura
1, A y B). Las genitalias extraídas de los machos de D. planipalpis y D. platura (Figura 2)
son similares a las ilustradas por Wang et al (2014) y Savage (2016). La genitalia del macho
es una herramienta fundamental para la identificación de especies de la familia
Anthomyiidae, por lo que la identificación de estas especies basada solo en la quetotaxia,
generalmente no resulta suficiente y ha causado confusiones en la literatura (Darvas y
Szappano 2003).
Figura 1. A) Macho de Delia planipalpis; B) Macho de Delia platura.
Daños. De acuerdo con las observaciones de campo y de la colecta de plantas con
síntomas y daños, las hembras de Delia spp., ovipositan en la base del tallo de la planta y en
A

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
162
el suelo cercano a la planta y una vez emergida la larva, se empieza a alimentar del tejido
externo del tallo y se introduce en este o en las hojas basales.
Figura 2. Genitalia de los machos. V esternito abdominal de D. planipalpis (A) y de D.
platura (B). Terminalia de D. planipalpis (C) y de D. platura (D).
El nivel de daño que ocasionan las larvas de Delia spp., está en función de la edad de la
planta, ya que entre los 10 a 30 días después del trasplante, las plantas dañadas presentan
síntomas parecidos a los que ocasiona la falta de agua. Las larvas de Delia spp., en los
cultivos de B. oleracea (var italica, capitata y botrytis), generalmente se encuentran en la
base de la planta alimentándose de la corona de la raíz, por lo que al dañar el sistema radicular
la planta muere o retrasa su crecimiento notablemente. Además, se pueden ver galerías en el
tallo principal por el efecto de su alimentación, así como los orificios de salida de las larvas
de tercer instar que se salen para pupar en el suelo. Las pupas se pueden observar en el sitio
donde se extrae la planta y en el sustrato adherido a las raíces de ésta. En plantas con más de
30 días después del trasplante, se observa que éstas pueden tolerar el daño ocasionado por la
alimentación de las larvas, sin embargo, dichas plantas serán de menor tamaño. Durante los
recorridos de campo se pudo constatar la presencia de adultos de D. planipalpis y D. platura
en las orillas de los cultivos referidos; sin embargo, esto no es un indicativo de que haya
A B
D C
E

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
163
larvas ocasionando daños significativos al cultivo, lo cual coincide con Savage et al. (2016),
quienes mencionan que algunas especies de Delia pueden ser muy abundantes como adultos,
pero que raramente están involucradas en los daños en el cultivo. Incluso, en cultivos de col
próximos a la cosecha, se observaron adultos en la periferia del cultivo que probablemente
vienen de cultivos aledaños y hospedantes silvestres. Estas observaciones coinciden con lo
observado por Hawkes (1972), quien señala que los adultos de Erioischia brassicae (=Delia
radicum), una vez que localizan el cultivo, permanecen en las orillas durante la mañana y por
la tarde se mueven dentro del cultivo para ovipositar y finalmente regresan a las orillas del
cultivo al atardecer.
AGRADECIMIENTOS
Se agradece al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca
otorgada al primer autor para sus estudios de doctorado. Al Colegio de Postgraduados,
Campus Montecillo, por el equipo y las instalaciones donde se realizaron la identificación y
toma de fotografías. También se reconoce y agradece la colaboración de la especialista en
Anthomyiidae, Dra. Jade Savage, quién corroboró la identidad Delia planipalpis y Delia
platura en Bishop´s University, Quebec, Canadá.
LITERATURA CITADA
Barrios DB, Alatorre RR, Bautista M N, Calyecac CHG. 2004. Identificación y fluctuación poblacional de plagas de col (Brassica oleracea var Capitata) y sus enemigos naturales en Acatzingo, Puebla. Agrociencia. 38 (2) 239-248.
Darvas B, Szappanos A. 2003. Male and female morphology of some central european Delia (Anthomyiidae) pests. Acta Zoologica Academiae Scientiarum Hungaricae. 49(2): 87-101.
Malloch JR. 1920. Descriptions of Diptera of the Families Anthomyidae and Scatophagidae (Part XII of Series). The Ohio Journal of Science. 20(7): 267-291.
Hawkes C. 1972.The diurnal periodicity and cycle of behaviour of the adult cabbage root Fly (Erioischia brassicae). Annals of Applied Biology. 70: 109-118.
Marín JA. 2001. Insectos plagas de maíz. Guía para su identificación. SAGARPA. INIFAP. Campo experimental Bajío. Folleto técnico No. 1. Celaya, Guanajuato, México. 29 p.
Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. Coordinación de Fundaciones Produce. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. 2017. Agenda Técnica Agrícola Guanajuato. 274 p.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
164
Savage J, Fortier AM, Fournier F, Bellavance V. 2016. Identification of Delia pest species (Diptera: Anthomyiidae) in cultivated crucifers and other vegetable crops in Canada. Canadian Journal of Arthropod Identification No. 29. 40 p.
SIAP, 2018. https://www.gob.mx/siap/acciones-y-programas/produccion-agricola-33119. Servicio de Información Agroalimentaria y Pesquera. Consultado el 19 de julio de 2019.
Wang QK, Yang YZ, Liu MQ, Zhang D. 2014. Fine Structure of Delia platura (Meigen) (Diptera: Anthomyiidae) Revealed by Scanning Electron Microscopy. Microscopy Research and Technique. 77: 619-630.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
165
REVISIÓN DE LA ESPERMATECA DE LAS ESPECIES DE LA
TRIBU XYLEBORINI (COLEOPTERA: SCOLYTIDAE) PRESENTES
EN MÉXICO
Mauricio Pérez Silva1, Armando Equihua Martínez1*, Thomas H. Atkinson2, Jesús Romero
Nápoles1, Obdulia L. Segura León1 y José Abel López Buenfil3
1Instituto de Fitosanidad, Colegio de Postgraduados, Km. 36.5 Carretera México-Texcoco, Texcoco, Estado de México 56230, México. 2University of Texas Insect Collection, 3001 Lake Austin Blvd. Austin, Texas 78703, Estados Unidos 3Servicio Nacional de Sanidad,
Inocuidad y Calidad Agroalimentaria. Unidad Integral de Diagnóstico, Servicios y Constatación, Tecámac, Estado de México 55740,
México. *Autor de Correspondencia: equihuaa@colpos. mx
INTRODUCCIÓN
La tribu xyleborini pertenece a un grupo de insectos que comúnmente son conocidos como
escarabajos ambrosiales, algunas de las especies incluidas en este grupo se consideran plagas
importantes tanto a nivel forestal como agrícola; en los últimos años han cobrado mucha
atención debido a la introducción de especies en Norteamérica que han impactado en gran
medida especies de árboles nativos y representa un riego potencial considerable (Wood,
1982; Hulcr et al. 2007; Rabaglia et al., 2006). Esto ha generado que en los últimos años se
dé mayor atención al grupo en diferentes áreas de estudio.
La espermateca es una estructura esclerosada ubicada dentro de la cavidad vaginal y su
principal función es el almacenamiento del esperma (Lindroth y Palmen, 1970);
frecuentemente se ha hecho referencia a esta estructura por su gran valor taxonómico para
especies cercanas. Recientemente Pérez et al (2018) resaltaron su valor para especies de
Xyleborus. Los elementos más importantes que componen a la espermateca son la cápsula
espermática (cornu y nodulus), glándula espermática y el conducto espermático (Ferronato,
2000; De Marzo, 2008). Si bien la función de almacenamiento se lleva a cabo en la cápsula
espermática, la glándula provee de los materiales esenciales para su correcto almacenamiento
y el conducto espermático conecta a la espermateca con la cavidad vaginal (Pérez et al, 2018).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
166
Tomando en cuenta esta información el objetivo de trabajo es registrar la variación
morfológica de la espermateca entre las diferentes especies de la tribu Xileborini presentes
en México.
MATERIALES Y MÉTODOS
Para la extracción de la espermateca se separó el abdomen del resto del cuerpo del insecto,
el cual se colocó en KOH al 10% a 80° C, entre 20 y 30 minutos; posteriormente se
enjuagaron con agua y después de un minuto se colocaron en ácido acético glacial al 5%,
posteriormente se pasaron a alcohol al 70%, luego 5 minutos en Alcohol al 100% y
finalmente se colocaron en xilol para montarlos en bálsamo de Canadá. Las estructuras se
observaron a través de un microscopio y se tomaron fotografías.
Se realizó la disección de un total de 68 ejemplares (Cuadro 1) pertenecientes a los 10
géneros y a 33 especies de las 55 especies de Xyleborini presentes en México; se
complementó el presente análisis con las espermatecas de 15 especies del género Xyleborus
mexicanos previamente analizados por Pérez et al. (2018). Las especies que no se incluyeron
en el presente trabajo fueron: Ambrosiodmus sp. Coptoborus catulus, Dryocoetoides
asperulus, Dryocoetoides sp., Theoborus coartatus, T. micarius y Xyleborus vismiae debido
a la falta de ejemplares.
Cuadro 1. Especies de Xyleborini usados para la extracción de la espermateca. Especie N* Colecta
Ambrosiodmus coffeiceus 2 MÉXICO: Puebla, Xicotepec; 02-V-2002; Col. J. Acuña- Soto;
CEAM (1). Veracruz, Jalapa; 12-IX-1983; CEAM1 (1).
Ambrosiodmus ferus 2 MÉXICO: Chiapas, Huixtan; 11-V-2017; DGSV2 (1). Veracruz, Jalapa; 1-IV-1984; Col. F. A. Noguera; CEAM (1).
Ambrosiodmus hagerdoni 2 MÉXICO: Chiapas, Tapachula; 13-XII-2017; Col. J. Barrera (1).
Chiapas, Tapachula; 11-X-2017; Col. J. Barrera (1)
Ambrosiodmus obliquus 3
MÉXICO: Oaxaca, Valle Nacional; 1-XII-2017; Col. M. Pérez-Silva (2). Veracruz, Uxpanapa; 10-XII-1982; Col. T. H. Atkinson;
CEAM (1) Ambrosiodmus rubricollis 2 ESTADOS UNIDOS DE AMÉRICA: Tennessee, Davidson; 00-I-
1994; Col. B. Kaufman; CEAM (2). Ambrosiodmus rugicollis 2 MÉXICO: Veracruz, Jalapa; 16-VIII-1983; Col. F. A. Noguera;
CEAM (2).
Ambrosiodmus rusticus 3 MÉXICO: Michoacán, Uruapán; 01-IX-2017; Col. M. O. Lazaro-Dzul (1). Morelos, Huitzilac; 2-VI-1984; Col. E. Martínez y E.
Saucedo; CEAM (2).
Ambrosiodmus scalaris 1 MÉXICO: Veracruz, Los Tuxtlas; 18-sep-1985; Col. L. Cervantes, CNIN3.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
167
Coptoborus pseudotenuis 6
MÉXICO: Chiapas, Tapachula; 23 al 27-I-2001, 21 al 28-II-2001 y 9 al 16-III-2001; (4). Veracruz, Coatepec; 09-V-2019; Cols. M.
Ochoa y J. Ríos (2). Coptoborus silviasalasi 2 MÉXICO: Oaxaca; 24-VI-2009; Col. T. H. Atkinson; CEAM (2).
Coptoborus tolimanus 1 MÉXICO: Veracruz, Uxpanapa; 27-IV-1982; Col. T. H. Atkinson; CEAM (1).
Coptoborus vepatoriorus 1 MÉXICO: Veracruz, Minatitlán; 28-IV-1982; Col. T. H. Atkinson;
CEAM (1).
Dryocoetoides capuccinus 8
MÉXICO: Guerrero, Petatlán; 16 al 21-XII-2016; Col. A. Hernández (5). Michoacán, Zumpimito; 00-VIII-2017; Col. A.
Equihua M. (3). Euwallacea kuroshio 2 MÉXICO: Baja California, Tijuana; 5-VI-2017; DGSV (2).
Euwallacea posticus 1 MÉXICO: Oaxaca, Valle Nacional; 1-XII-2017; Col. M. Pérez Silva (1).
Sampsonius dampfi 1 MÉXICO: Chiapas, Tapachula; 00-VIII-2018; Col. M. Pérez Silva (1).
Sampsonius mexicanus 1 MÉXICO: Tabasco, Huimanguillo; 13-XII-2012; Col. M. Pérez De la Cruz; UJAT (1).
Sampsonius reticulatus 1 MÉXICO: Veracruz, Los Tuxtlas; 4-IV-2014; Col. M. Pérez Silva (1).
Taurodemus flavipes 1 MÉXICO: Oaxaca, Uxpanapa; 24-V-1981; Col. A. Equihua M.; CEAM (1).
Taurodemus sharpi 1 MÉXICO: Veracruz, Los Tuxtlas; 4-IV-2014; Col. M. Pérez Silva (1).
Theoborus incultus 1 MÉXICO: Campeche, Escarcega; 3-II-1984; Col. A. Estrada V.; CEAM (1)
Theoborus ricini 5 MÉXICO: Chiapas, Tapachula; 13-XII-2017; Col. J. Barrera (1). Oaxaca, Valle Nacional; 2-XII- 2019; Col. M. Pérez-Silva (1);
Tabasco, Macuspana; 25-X-2016; Col. V. Arriola (3).
Theoborus theobromae 1 MÉXICO: Veracruz, Uxpanapa; 22-V-1981; Col. A. Equihua; CEAM (1)
Xyleborinus gracilis 3 MÉXICO: Guerrero, Petatlán; 16 al 21-XII-2016 y 21 al 28-IV-
2017; Col. A. Hernández (2). San Luis Potosí, Axtla de Terrazas; 11 al 15-IV-2017; Col. M. Pérez-Silva y I. Ríos-Ibarra (1).
Xyleborinus intersetosus 3
MÉXICO: Puebla, Zihuateutla; 3-IV-2002; Col. J. Acuña-Soto (2). Veracruz, Los Tuxtlas; 29 al 31-VIII-2017; Cols. M. Pérez-Silva y I.
Ríos- Ibarra (1). Xyleborinus saxeseni 2 MÉXICO: Jalisco, La primavera; 2-III-2011 (1).
Xyleborinus tribuloides 1 MÉXICO: Oaxaca, P. Blanca; 26-X-1986; Col. A. Burgos; CEAM (1).
Xyleborus pubescens 1 MÉXICO: Chiapas, Altamirano; 16-V-2011; Col. R. Altuzar-Menda; LARSF4 (1).
Xyleborus sparsipilosus 3
MÉXICO: Veracruz, Uxpanapa; 10-XII-1983; Col. T. H. Atkinson; CEAM (2). Veracruz, Uxpanapa; 31-I-1984; Col. T. H.
Atkinson; CEAM (1).
Xyleborus titubanter 1 MÉXICO: Chiapas, Las Margaritas; 11-VI-2016; Col R. González; ECO-TAP-E5 (1).
Xylosandrus compactus 3 ESTADOS UNIDOS DE AMÉRICA: Sin datos (3). Xylosandrus curtulus 1 MÉXICO: Nayarit; 2018; Col. L. Domínguez (1).
Xylosandrus morigerus 2 MÉXICO: Chiapas, Tapachula; 14-II-2018; Col. J. Barrera (2). *Número de ejemplares analizados. 1Colección entomológica del Colegio de Postgraduados, Montecillo. 2Colección de Insectos de la Dirección General de
Sanidad Vegetal. 3Colección Nacional de Insectos. 4Colección de Insectos del Laboratorio de Análisis y Referencia de Sanidad Forestal. 5 Colección entomológica del Colegio de la Frontera Sur, Tapachula.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
168
RESULTADOS Y DISCUSIÓN
La variación morfológica de la espermateca tanto a nivel de género como de especie fue
muy amplia, lo cual dificultó generalizar en un solo esquema la información de dicha
estructura para todas las especies de Xyleborini, sin embargo, se tomó en cuenta la
nomenclatura propuesta por Pérez et al. (2018), quienes profundizaron en el análisis de la
espermateca del género Xyleborus, ya que la mayoría de las especies se ajustaron a su
propuesta, además de que su esquema incluye los componentes básicos de la espermateca.
Es importante reconocer que existen cuatro áreas que en todas las especies son consistentes
y que ayudaron a la caracterización tanto a nivel de especies como de géneros: la glándula
espermática, el nodulus, el cornu y la unión entre la cápsula y el conducto espermático a
través del collum o unidos directamente al nodulus (Figura 1).
Figura 1. Esquema general de la espermateca del género Xyleborus. a=apodema; c=cornu; co=collum; dv=valvula del conducto; mc=constricción media; n=nodulus; sd=conducto espermático; sg=glándula espermática; sgd= conducto de la glándula espermática (tomada de Pérez et al. 2018).
En México existen reportados 10 géneros de Xyleborini (Atkinson 2019) de los cuales
Euwallacea, Xyleborinus, Xyleborus y Xylosandrus presentan gran similitud y se ajustan al
esquema presentado anteriormente, mientras que los géneros Ambrosiodmus, Coptoborus,
Dryocoetoides, Sampsonius, Taurodemus y Theoborus presentan características muy
particulares y no se ajustan al esquema propuesto, además de que pueden representar un
carácter de gran valor para separar géneros.
Hasta el momento se ha analizado la espermateca del 87.3% de las especies de la tribu
Xyleborini, es decir 48 de las 55 especies registradas para México, esta estructura resultó ser

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
169
un carácter importante en el reconocimiento, tanto de las especies como de los géneros que
constituyen la tribu para México.
AGRADECIMIENTOS
Agradecemos en primer lugar a CONACyT por la beca otorgada y al Colegio de
Postgraduados; a la M. C. Cristina Mayorga, al Dr. Armando Burgos, M. C. Leonardo
Delgado, al Dr. Manuel Pérez y a la Dra. Lourdes Chamorro y todas aquellas personas
(Investigadores, técnicos y estudiantes) que nos apoyaron y permitieron revisar sus
colecciones y a la Biol. R. Ivette Ríos Ibarra, al Biol. Manuel Ochoa, al M. C. Eduardo
Chamé, al Dr. Artemio Hernández, Dr. Jesús A. Acuña-Soto y al Dr. Juan M. Vanegas por
su colaboración en las colectas y la donación de ejemplares.
LITERATURA CITADA
Atkinson, T. H. 2019. Bark and ambrosia beetles. Disponible en: www.barkbeetles.info/about.php. (October 2019).
De Marzo, L. 2008. Biodiversitá della spermateca nei coleotteri. Atti Accademia Nazionale Italiana di Entomologia Anno LVI: 69–96.
Ferronato, E. M. O. 2000. A cápsula da espermateca dos Eumolpinae (Coleoptera: Chrysomelidae) asociados a Theobroma cacao L. (Sterculiaceae) do sul da Bahia. Anais da Sociedade Entomológica do Brasil 29: 57–63.
Hulcr, J., S. A. Dole, R. A. Beaver y A. I. Cognato. 2007. Cladistic review of generic taxonomic
characters in Xyleborina (Coleoptera: Curculionidae: Scolytinae). Systematic Entomology 32: 568–584
Lindroth, C. H., y E. Palmén. 1970. Coleoptera (pp.80–87). In: Taxonomist’s Glossary of Genitalia in Insects. 2° edition (S. L. Tuxen, editor). Munksgaard, Copenhagen, Denmark.
Pérez S. M., A. Equihua M., J. M. Valdez C y E. G. Estrada V. 2018. Spermathecae of the Mexican Species of Xyleborus Eichhoff (Coleoptera: Curculionidae: Scolytinae). Coleopterist bulletin 72 (3): 616-624.
Rabaglia R. J., S. A. Dole, y A. I. Cognato. 2006. Review of American Xyleborina (Coleoptera: Curculionidae: Scolytinae) occurring North of México, with an illustrated key. Annals of the Entomological Society of America 99:1034–1056
Wood, S. L. 1982. The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytinae), a taxonomic monograph. Great Basin Naturalis Memoirs 6: 1–1356.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
170
REVISIÓN DE LA TRIBU PACHYMERINI (COLEOPTERA:
BRUCHIDAE: PACHYMERINAE) PARA MÉXICO Y FILOGENIA
DEL GÉNERO Caryobruchus
Sergio Godínez Cortés1*, Jesús Romero-Nápoles1, Obdulia Lourdes Segura León1, Armando
Equihua Martínez1* y Julián Bueno Villegas2
1Laboratorio de Taxonomía de Insectos, Posgrado en Fitosanidad-Entomología y Acarología, Instituto de Fitosanidad, Colegio de
Postgraduados (Campus Montecillo). Carretera México-Texcoco, Km 36.5, Código Postal 56230; Montecillo, Texcoco de Mora, Estado
de México, México. 2Laboratorio de Sistemática Animal, Centro de Investigaciones Biológicas, Universidad Autónoma del Estado de
Hidalgo, C. P. 42184, Mineral de la Reforma, Hidalgo, México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
Las especies de la tribu Pachymerini Bridwell 1929, corresponden a coleópteros
pertenecientes a la familia Bruchidae, caracterizados como fitófagos obligados de semillas
durante su estado inmaduro (Kingsolver, 2002; Yus et al., 2007). La tribu está conformada
por cinco géneros actuales y uno extinto: Butiobruchus Prevett 1966, Caryoborus Schöenherr
1833, Caryobruchus Bridwell 1929, Mesopachymerus (†) Poinar 2005, Pachymerus
Thunberg 1805 y Speciomerus Nilsson 1993 (Romero, 2019).
Los Pachymerini presentan una distribución exclusiva en el Nuevo Mundo y casi
restringida a la región Neotropical (Borowiec, 1987); aunque su origen es Neártico, su mayor
irradiación y diversificación proviene de Sudamérica (Poinar, 2005). Los Pachymerini se
caracterizan por consumir exclusivamente semillas de palmas de la familia Arecaceae; y
morfológicamente por presentar un proceso prosternal bien marcado, siempre separando
completamente las procoxas, nunca agudo; el primer segmento protarsal y metatarsal
fuertemente expandido triangularmente en el ápice y una longitud corporal entre 8-20 mm
(Nilsson y Johnson, 1993).
Para México, se conocen exclusivamente la presencia de tres géneros: Caryobruchus,
rebasando la distribución Neotropical a la Neártica; Speciomerus, con una distribución
limitada hasta el sur de México, en Chiapas; y Pachymerus, con una distribución dudosa e
incierta en territorio mexicano (Romero, 2019). El escaso número de especies y su casi nulo
conocimiento sobre la asociación con sus plantas hospederas, a pesar de que muchas de ellas

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
171
tienen interés comercial masivo y artesanal (Pavón et al., 2006), es una razón para
profundizar en su estudio.
Debido a la aparición de nuevos datos de colecta y de distribución, de la asociación con
sus hospederos, y a la aparición de nuevos taxa dentro del género Caryobruchus, se realiza
la revisión de la tribu Pachymerini para México. Con lo que se busca obtener la siguiente
información: literatura sinonímica, diagnosis, dimorfismo sexual, distribución, hospederos,
material examinado, ilustraciones, mapas de distribución, claves para géneros y especies; y
el esclarecimiento de las relaciones filogenéticas de las especies dentro del género
Caryobruchus, mediante un análisis cladístico por Parsimonia con datos morfológicos.
MATERIALES Y MÉTODOS
Colecciones entomológicas. Se solicitó a préstamo y se revisó todo el material referente
a Pachymerini para México depositado en las colecciones de: BYUC (Brigham Young
University Collection. Brigham Young University, Utha), CEAM (Colección Entomológica
del Colegio de Postgraduados, Colegio de Postgraduados, Estado de México), CNIN
(Colección Nacional de Insectos, Universidad Nacional Autónoma de México, Ciudad de
México), TMMC (Austin Insect Collection. University of Texas, Austin) y USNM (United
States National Museum. Smithsonian Institution, Washington DC).
Base de datos y literatura especializada. Se consultaron y cotejaron los registros
conocidos e inéditos alojados previamente en la base de datos BRUCOL (2019) y de distintas
fuentes de literatura especializada; así como de la compilación de datos inéditos de etiquetas
de los ejemplares de museo en préstamo; vaciando la información a la misma BRUCOL
(2019), implementada en un programa de diseño de base de datos (Paradox® Versión X6,
Corel Corporation).
Digitalización de ejemplares. Se realizó la digitalización y esquematizaron cada una de
las especies de Pachymerini en México. Para adultos, se utilizó un microscopio estereoscopio
(Olympus® SZX7), una fuente de iluminación (Fiber-Lite® MI-150), una cámara digital
(Olympus® E-620) y el software para manejo de la cámara (Olympus Studio® 2.22). Para
las genitalias, se utilizó una cámara digital (AmScope® 5 mm, modelo MW 1000), adaptada

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
172
a un microscopio óptico (Zeiss® Axiostar), y el software para toma de fotografía
(AmScope® versión X86, 3.7.3980). Todas las imágenes fueron procesadas y editadas por
medio de softwares libres (Gimp 2, CombineZP e ImageJ).
Clave taxonómica. Se elaboró una clave dicotómica para identificación de géneros y
especies de Pachymerini en México, siguiendo el formato de Kingsolver (2004), la
terminología de Nilsson y Johnson (1993), complementándola con información de Godínez-
Cortés (2017).
Mapas de distribución. Actualmente se está eligiendo el mejor software para documentar
la distribución de las especies, con utilidad para poderle dar una interpretación biogeográfica;
principalmente en el flujo génico y de dispersión de las especies de Caryobruchus, en el
continente americano.
Análisis filogenético. Se trabaja en la elaboración de una matriz de aproximadamente 30
caracteres morfológicos para realizar un análisis cladístico mediante Parsimonia, con lo cual
se espera establecer el origen y las relaciones filogenéticas entre de las especies del género
Caryobruchus. El estudio se conforma con nueve taxa terminales, siete para el grupo interno
(Caryobruchus spp.), y dos para el grupo externo (Epitrix cucumeris (Harris, 1851) y
Diabrotica speciosa (Germar, 1824). Las búsquedas de los árboles más parsimoniosos se
realizarán utilizando el algoritmo heurístico de búsqueda tradicional, se elegirá el árbol más
parsimonioso solo, o por consenso estricto, aplicando ponderación contra la homoplasia
(Goloboff y Farris, 2001; Goloboff et al., 2008). Los caracteres binarios o multiestado, serán
tratados como no aditivos y con pesos iguales. El análisis será implementado en el programa
TNT (Tree analysis using New Technology) v 1.1, actualizado hasta el día de hoy (Goloboff,
Farris y Nixon ® 2019).
RESULTADOS Y DISCUSIÓN
Se logró identificar a nivel de especie todos los ejemplares provenientes de los préstamos
de las distintas colecciones y museos, sumado a su curación, mediante el montaje, etiquetado
y sexado. Las especies de la tribu Pachymerini presentes en México resultaron ser: P. cardo
(Fåhraeus, 1839), S. giganteus (Chevrolat, 1877), C. curvipes (Latreille, 1811), C. gleditsiae
(Johansson and Linnaeus, 1763), C. maya Nilsson, 1993, C. rubidus (Chevrolat, 1877) y C.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
173
veseyi (Horn, 1873); así como el surgimiento de una nueva especie, Caryobruchus mayledi
Godínez-Cortés y Romero, 2020.
Derivado de los datos inéditos de BRUCOL (2019), de la literatura consultada y de los
datos de etiquetas observadas, fue posible recopilar nuevos registros de distribución y
asociación de hospederos. Para cada una de dichas especies, se estableció de manera
detallada: su literatura taxonómica y sinonímica, una diagnosis mediante la distinción de sus
rasgos más representativos, el señalamiento puntual del dimorfismo sexual, su distribución,
las plantas hospederas asociadas, registros previamente publicadas y otras inéditas,
ilustraciones de machos y hembras adultos y sus respectivas genitalias, así como una clave
para la identificación de géneros y especies.
Algunos datos puntuales y relevantes de la investigación son: Se confirma la especie P.
cardo en territorio mexicano, con datos específicos en el estado de Jalisco, con una
distribución que rebasa los límites Neotropicales, así como C. gleditsiae, como la especie
más polífaga y con la mayor presencia, con un rango de distribución muy amplio, bien
establecida en México.
Por otro lado, si se logra implementar el análisis cladístico por Parsimonia, se logrará
llegar al esclarecimiento de las relaciones filogenéticas de las especies dentro del género
Caryobruchus, dilucidando su parentesco, distribución y predecir su origen, además de poder
abordar una posible hipótesis sobre su tipo de especiación, que servirá como antecedente para
entender la diversificación de la tribu en América y su exclusividad en el nuevo mundo.
De manera más clara y útil, al lograr documentar mapas de distribución, imágenes
digitalizadas de las especies y clave para su identificación, se logrará obtener una herramienta
sólida para su conocimiento, identificación y tratamiento, disponible para todo mundo. Por
lo que los resultados finales de esta investigación, formarán un cúmulo completo de los
aspectos sistemáticos básicos, que nos permitirán abordar a las Pachymerini en México, en
cualquier perspectiva o campo de la biología.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
174
AGRADECIMEINTOS
Completa gratitud a CONACYT por la beca "Inversión en el Conocimiento" que han
financiado mis estudios doctorales, así como al Colegio de Postgraduados por brindarme las
facilidades para desarrollar dicho proyecto.
LITERATURA CITADA
Borowiec, L. 1987. The genera of seed-beetles (Coleoptera, Bruchidae). Polskie Pismo Entomologiczne 57: 3-207.
Godínez-Cortés, S. 2017. Biodiversidad de la familia Bruchidae (Insecta: Coleoptera) en el estado de Hidalgo, México. Tesis de Maestría, Colegio de Postgraduados, Estado de México.
Goloboff, P. A., y J. S. Farris. 2001. Methods for quick consensus estimation. Cladistics 17: 26-34.
Goloboff, P. A., J. M. Carpenter, J. S. Arias, y D. R. Miranda Esquivel. 2008. Weighting against homoplasy improves phylogenetic analysis of morphological data sets. Cladistics 24: 1-6.
Kingsolver, J. M. 2002. Bruchidae. In: Arnett, J. R. H., Thomas, M. C., Skelley, P. E. y J. H. Frank (eds.). American beetles. Polyphaga: Scarabaeoidea through Curculionoidea, volume 2. CRC Press, New York. pp. 602-608.
Godínez-Cortés, S. 2015. Brúquidos: Algunos ejemplos de la importancia cultural de los escarabajos de las semillas. In: Castaño-Meneses, G. y J. L. Navarrete-Heredia (eds.). Avances en la Entomología Cultural Mexicana. Universidad Nacional Autónoma de México y Universidad de Guadalajara, Jalisco. pp. 77-82.
Kingsolver, J. M. 2004. Handbook of the Bruchidae of the United States and Canada (Insecta, Coleoptera), Volume I. Department of Agriculture, Technical Bulletin No.1912, Washington, D. C., Unites States of America.
Nilsson, J. A., y C. D. Johnson. 1993. A Taxonomic Revision of the Palm Bruchids (Pachymerini) and a Description of the World Genera of the Pachymerinae. Memoirs of the American Entomological Society 41: 1-104.
Poinar, G. 2005. A Cretaceous Palm Bruchid, Mesopachymerus antiqua, n. gen., n. sp. (Coleoptera: Bruchidae: Pachymerini) and biogeographical implications. Proceedings of Entomological Society of Washington 107: 392-397.
Pavón, N., R. Escobar, y R. Ortíz-Pulido. 2006. Extracción de hojas de la palma Brahea dulcis en una comunidad otomí en Hidalgo, México: efecto sobre algunos parámetros poblacionales. Interciencia 31:1-6.
Romero N., J. 2019. BRUCOL Database (actualizada). Programa de Entomología y Acarología, Instituto de Fitosanidad, Colegio de Postgraduados, Estado de México, México. (consultada el: 12-X-2019).
Yus R., R., J. M. Kingsolver, y J. Romero N. 2007. Sobre el estatus taxonómico actual de los brúquidos (Coleoptera: Bruchidae) en los Chrysomeloidea. Dugesiana 14: 1-21.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
175
IDENTIFICACIÓN MORFOLÓGICA DE SIETE ESPECIES DEL
GÉNERO Cuscuta (Convolvulaceae) DE IMPORTANCIA AGRÍCOLA
Y FORESTAL
Sonia Monroy-Martínez1, Clemente de Jesús García Ávila2, Obdulia Lourdes Segura León1,
Dionicio Alvarado Rosales1*
1 Posgrado en Fitosanidad-Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco,
Estado de México, México. 2 Servicio Nacional de Sanidad Inocuidad y Calidad Agroalimentaria. Centro Nacional de Referencia
Fitosanitaria. Km 37.5, Carretera Federal México-Pachuca, 55740 Tecámac, Estado de México, México. *Autor de correspondencia:
INTRODUCCIÓN
Las plantas parásitas, como Cuscuta, se asocian con la reducción o modificación de
estructuras vegetativas, las hojas se reducen a escamas y las raíces desaparecen y se
reemplazan por haustorios; estás se encuentran en al menos 13 familias, con
aproximadamente 4000 especies (1 % de plantas angiospermas) (Stefanović et al., 2007). El
término parásita, se refiere a la alimentación de los hospederos (del griego para = al lado;
sitos = alimento), manteniéndolos vivos hasta que han completado su ciclo de vida. La mayor
diversidad de especies se encuentra en América (Yuncker, 1932), principalmente en regiones
con clima cálido y húmedo; parasitan principalmente plantas dicotiledóneas y algunas
monocotiledóneas (Dawson et al., 1994); algunas especies pueden ser muy específicas con
su hospedante, mientras que otras tienen un amplio rango.
La identificación de Cuscuta sp., se realiza con claves taxonómicas con base en
características morfológicas como: tamaño, forma, color, tipos de hojas, flores, frutos,
semillas, entre otras (Cullen, 2006), características que generalmente se presentan en la etapa
de floración y que son útiles para su identificación morfológica, así como como, para que a
partir de estas se genere un perfil genético con base en marcadores moleculares que permita
más adelante su identificación en cualquier estado fenológico. Por lo anterior y la necesidad
de materiales didácticos que permitan la identificación in situ; además, de conocer la
distribución de algunas especies que se encuentran en el país, el objetivo del presente trabajo
fue elaborar una guía ilustrada que incluya características que apoyen a la identificación de
siete especies del género Cuscuta, con reporte de presencia en México, que sirva de base para

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
176
generar el perfil genético de estas en apoyo al proceso de identificación y diagnostico de
plagas de importancia económica.
MATERIALES Y MÉTODOS
El presente trabajo se realizó en las instalaciones del Centro Nacional de Referencia
Fitosanitaria (CNRF) de la Dirección General de Sanidad Vegetal, en el Laboratorio de
Roedores, Aves y Malezas. Se identificaron 56 ejemplares depositados en el herbario del
laboratorio y se realizaron tres recolectas de ejemplares con presencia de estructuras florales
para su identificación, que se encontraban parasitando cultivos de importancia agrícola o
forestal, en regiones del estado de México, Morelos y Sonora. Los ejemplares recolectados
se fotografiaron in situ, con una cámara Canon, EOS 1000D (Canon Mexicana, S. de R.L. de
C.V., México).
La identificación de cada uno de los especímenes se realizó por medio de las estructuras
florales, siguiendo la metodología de Wright (2011). De cada uno de los ejemplares se
removieron algunas flores y se colocaron en un vaso de precipitado de 50 mL, se agregó
etanol al 50 % para rehidratarlas. Posteriormente, se colocó el vaso sobre una plancha de
calentamiento con agitación hasta alcanzar el punto de ebullición. Después de enfriarse, las
flores se tomaron con una pinza de relojero, se depositaron en una caja Petri, se observaron
en un microscopio estereoscopio Stemi 2000-C, (Carl Zeiss de México, S. A. de C. V.,
México); posteriormente se cortaron, para observar los caracteres taxonómicos de cada flor:
cáliz, corola, escamas infraestaminales, estilos, estigmas, cápsula y semillas.
Con la cámara fotográfica AxioCam ERc5s y el software ZEN 2012 blue edition (Carl
Zeiss de México, S. A. de C. V., México), se tomaron fotografías de estas características,
utilizando diferentes aumentos, según el tamaño de las estructuras. Las fotografías de las
estructuras de la flor, fueron ensambladas mediante el programa CombineZP 1.0 (CombineZ-
Pyramid por sus siglas en inglés) (Hadley, 2006), con la finalidad de obtener una sola
fotografía de cada estructura. Se usó el programa GIM 2.10.0 (GNU Image Manipulation
Program, por sus siglas en inglés) para editar las fotografías y agregar la escala milimétrica.
Se realizaron mediciones de las características florales con el programa ZEN 2012 blue
edition (Carl Zeiss de México, S. A. de C. V., México), las cuales se utilizaron para hacer la

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
177
comparación de medidas con lo reportado por Riviere et al. (2013), Welsh et al. (2010) y
Wright et al. (2012); además, de servir como apoyo en la identificación junto con las claves
taxonómicas y descripciones de acuerdo a Yuncker (1932; 1965), Costea y Tardif (2006),
Costea et al. (2012, 2013). Asimismo, se consultaron ejemplares de colección determinados
por el Dr. Mihai Costea especialista de este género, del Departamento de Biología,
Universidad Wilfrid Laurier, Canadá; así como, la observación de ejemplares escaneados de
los herbarios MEXU del Instituto de Biología de la UNAM (IBUMAN) y el New York
Botanical Garden Steere Herbarium (NYBG).
RESULTADOS Y DISCUSIÓN
Se revisaron 59 ejemplares del género Cuscuta, con ayuda de las claves taxonómicas,
mediciones, fotografías obtenidas y comparación de ejemplares, se identificaron siete
especies.
Los cortes que se realizaron a las flores permitieron observar y medir las estructuras para
identificar las especies. Se tomaron fotografías de algunas estructuras como corte del cáliz,
corte de la corola y escamas infraestaminales (Figura 1), para que se pudieran observar las
características y diferencias de cada una de estas especies.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
178
Figura 1. Escamas infraestaminales. A. C. campestris Yunck. B. C. chinensis Lam. var. applanata (Engelm.)
Costea y Stefanov. C. C. corymbosa Ruiz y Pav. var. stylosa Engelm. D. C. indecora Choisy E. C. costaricensis
Yunck. F. C. mitriformis Engelm. Ex Hemsl. G. C. tinctoria Mart. ex Engelm. A-G = 1 mm
De acuerdo a lo mencionado por Costea y Tardif (2006), las flores de C. campestris llegan
a medir entre 1.5 a 2 mm de largo, mientras que los tubos de la corola de las especies
recolectadas midieron 1.1 mm. En este trabajo se observó una disminución de 0.4 mm, la
diferencia de tamaño puede deberse a la plasticidad fenotípica y la variabilidad genética que
puede influir en los caracteres para identificación, como indica Palacio-López y Rodríguez-
López 2007, donde las plantas se adaptan y pueden producir diferentes respuestas a las
condiciones ambientales en las que se están desarrollando.
En el caso de C. corymbosa, la corola midió 3.4 mm, según lo que indica Carranza (2008),
las flores pueden llegar a medir entre 4 mm. De acuerdo a las mediciones tomadas el tamaño
no concuerda, pero para diferenciar la variedad con la que se trabajó, se revisó la descripción
de Costea et al. (2012), para C. corymbosa var. grandiflora, donde indica que las flores llegan
a medir 4 mm de longitud y el cáliz llega a la mitad de estas. Según lo reportado para C.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
179
corymbosa var. stylosa, las flores miden aproximadamente 3 mm de longitud y el cáliz es
más corto que la mitad del tubo de la corola, y las escamas infraestaminales llegan a la mitad
del tubo de la corola (Carranza, 2008), por lo que de acuerdo a esta descripción la especie
que se identificó fue C. corymbosa var. stylosa.
La especie de C. costaricensis se encuentra dentro de las medidas reportadas por Carranza
(2008) y Costea et al. (2011), para la longitud de flor, longitud de escamas y fimbrias, pero
existe una diferencia en la longitud del cáliz de 0.7 mm, ya que lo reportado es de 3.2-3-5
mm y la medida que se tomo fue de 2.5 mm, causado quizás por causas ambientales del
medio donde se desarrolló.
Por otro lado C. chinensis var. applanata no muestra cambios significativos, el tamaño es
similar a lo reportado por Costea (2011). El tamaño de C. indecora corresponde a lo reportado
por Costea et al. (2006) y Yuncker (1932). No existió diferencia en las medidas que se
tomaron en C. mitriformis y lo reportado por Carranza (2008); asimismo, las medidas de C.
tinctoria, se encuentran dentro del rango que indican Yuncker (1932) y Costea et al. (2013).
Las medidas que se obtuvieron de la corola, cáliz y escamas infraestaminales, no
concordaron al 100 % con las que indicaronWelsh et al. (2010), Wright et al. (2012) y Riviere
et al. (2013); sin embargo, las fotografías obtenidas y las demás caracteristicas que se
mencionan en las claves taxonómicas ayudaron a identificar a estas especies; asimismo se
pudo elaborar una guía con las principales caracteristicas de estas especies.
AGRADECIMIENTOS
Al Colegio de Postgraduados, Campus Montecillo por las facilidades brindadas y la
asesoría de sus profesores.
LITERATURA CITADA Carranza, E. 2008. Flora del bajío y de regiones adyacentes. Convolvulaceae II. Fascículo
155. Instituto de Ecología A. C. Costea, M., I. R. Garcia y S. Stefanović. 2011. ‘Horned’ dodders: phylogenetic relationships
and two new species within Cuscuta chapalana complex (Convolvulaceae). Botany 89: 715–730.
Costea, M., R. S. Felger, D. F. Austin, T. R. Van Devender y J. J. Sánchez-Escalante. 2012. Convolvulaceae of Sonora, México. II: Cuscuta. Journal of the Botanical Research Institute of Texas 6(2): 529-550.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
180
Costea, M., I. R. García, K. Dockstader y S. Stefanović. 2013. More problems despite bigger flowers: Systematics of Cuscuta tinctoria clade (subgenus Grammica, Convolvulaceae) with description of six new species. Systematic Botany 38(4): 1160-1187. DOI: https://doi.org/10.1600/036364413X674887.
Cullen, J. 2006. Practical plant identification. Cambridge University. 357. Dawson J. H, L. J Musselman, P. Wolswinkel e I. Dorr. 1994. Biology and control of
Cuscuta. Reviews of Weed Science 6: 265–317. Hadley, A. 2006. CombineZM and CZBatch - Free image stacking software for depth of field
correction. http://www.hadleyweb.pwp.blueyonder.co.uk/ Palacio-López, K. y N. Rodríguez-López. 2007. Phenotypic Plasticity in Lippia alba
(Verbenaceae) in Response to Water Availability in Two Light Environments. Acta Biológica Colombiana, 12: 187-198.
Riviere S., C. Clayson, K. Dockstader, M. A. R. Wright y M. Costea. 2013. To attract or to repel? Diversity, evolution and role of the ‘‘most peculiar organ’’ in the Cuscuta flower (dodder, Convolvulaceae) the infrastaminal scales. Plant Systematics and Evolution 24 p. DOI: https://doi.org/10.1007/s00606-012-0741-2
Stefanović, S., M. Kuzmina y M. Costea. 2007. Delimitation of major lineages within Cuscuta subgenus Grammica (Convolvulaceae) using plastid and nuclear DNA sequences. American Journal of Botany 94(4): 568-589.
Welsh, M., Stefanović, S. y M. Costea. 2010. Pollen evolution and its taxonomic significance in Cuscuta (dodders, Convolvulaceae). Plant Systematics and Evolution 285: 83-101. DOI: https://doi.org/10.1007/s00606-009-0259-4
Wright, M. 2011. The evolution of sexual reproduction in Cuscuta (Convolvulaceae). Theses and dissertations. 1039.
Wright, M. A. R., M. D. Ianni y M. Costea. 2012. Diversity and evolution of pollen-ovule production in Cuscuta (dodders, Convolvulaceae) in relation to floral morphology. Plant Systematics and Evolution 298: 369-389. DOI: https://doi.org/10.1007/s00606-011-0550-z
Yuncker T. G. 1932. The genus Cuscuta. Memoirs of the Torrey Botanical Club 18, 113–331.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
181
FACTORES BIÓTICOS Y ABIÓTICOS QUE FAVORECEN LA
INCIDENCIA Y DESARROLLO DE Lasiodiplodia theobromae EN PINO
CARIBEÑO (Pinus caribaea var. hondurensis)
Laura Rosney Jiménez-González1, Dionicio Alvarado-Rosales1*, Victoria Ayala-Escobar, Armando
Gómez-Guerrero2, Sergio Hernández Pablo3
1 Posgrado en Fitosanidad-Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco,
Estado de México, México. 2 Posgrado en Ciencias Forestales, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo,
56230 Texcoco, Estado de México, México. 3 Agroforestal UUMBAL, Las Choapas, Veracruz. *Autor de correspondencia:
INTRODUCCIÓN
El Pino caribeño o amarillo (Pinus caribaea var. hondurensis (Séneclauze) W.H.G. Barret &
Golfari) es el pino tropical con mayor distribución geográfica, encontrado en forma natural
en Belice, Guatemala, Honduras, Nicaragua y sur de México, actualmente se ha distribuido
en otros estados con fines comerciales e industriales (Perry, 1991; CONABIO, 1962). Uno
de los problemas de importancia que se ha reportado en esta especie es Lasiodiplodia
theobromae, patógeno endófito que se manifiesta en árboles bajo estrés fisiológico y posee
la cualidad de estar asociado desde semilla, afectando así a nivel de plántulas (Gonzatto, et
al., 2015; Cilliers et al., 1993). En árboles adultos este patógeno ocasiona el manchado azul
de la madera reduciendo la calidad de la misma (Mohali, 1993).
Estudios realizados en Campeche por Tucuch et al. (2005) menciona que la intensidad de
la enfermedad se ve favorecida cuando la humedad relativa es mayor a 80% con temperaturas
entre 26 - 32°C. Debido a la importancia que posee la especie en el sector industrial, el
presente estudio tiene como objetivo determinar los principales factores involucrados en
favorecer la presencia de Lasiodiplodia teobromae en plantaciones de Pino tropical
localizados en Sayula y Las Choapas, Veracruz así como evaluar la patogenicidad de las
cepas obtenidas en cada región.
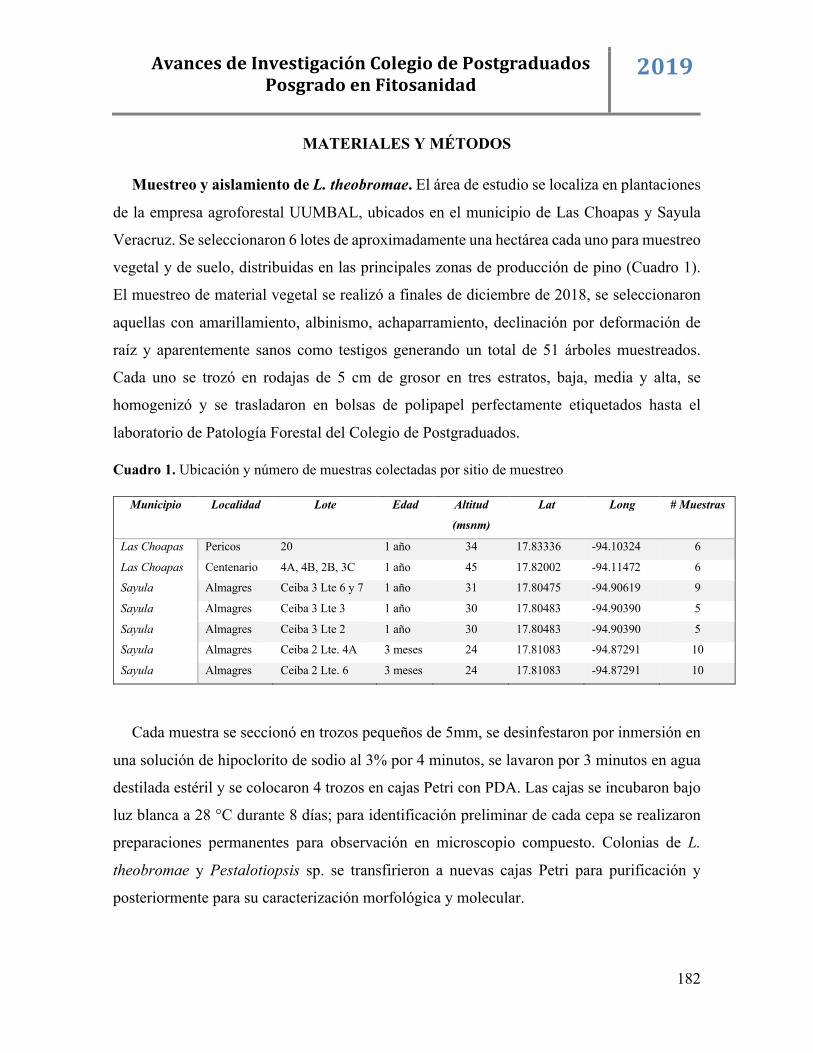
AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
182
MATERIALES Y MÉTODOS
Muestreo y aislamiento de L. theobromae. El área de estudio se localiza en plantaciones
de la empresa agroforestal UUMBAL, ubicados en el municipio de Las Choapas y Sayula
Veracruz. Se seleccionaron 6 lotes de aproximadamente una hectárea cada uno para muestreo
vegetal y de suelo, distribuidas en las principales zonas de producción de pino (Cuadro 1).
El muestreo de material vegetal se realizó a finales de diciembre de 2018, se seleccionaron
aquellas con amarillamiento, albinismo, achaparramiento, declinación por deformación de
raíz y aparentemente sanos como testigos generando un total de 51 árboles muestreados.
Cada uno se trozó en rodajas de 5 cm de grosor en tres estratos, baja, media y alta, se
homogenizó y se trasladaron en bolsas de polipapel perfectamente etiquetados hasta el
laboratorio de Patología Forestal del Colegio de Postgraduados.
Cuadro 1. Ubicación y número de muestras colectadas por sitio de muestreo
Municipio Localidad Lote Edad Altitud
(msnm)
Lat Long # Muestras
Las Choapas Pericos 20 1 año 34 17.83336 -94.10324 6
Las Choapas Centenario 4A, 4B, 2B, 3C 1 año 45 17.82002 -94.11472 6
Sayula Almagres Ceiba 3 Lte 6 y 7 1 año 31 17.80475 -94.90619 9
Sayula Almagres Ceiba 3 Lte 3 1 año 30 17.80483 -94.90390 5
Sayula Almagres Ceiba 3 Lte 2 1 año 30 17.80483 -94.90390 5
Sayula Almagres Ceiba 2 Lte. 4A 3 meses 24 17.81083 -94.87291 10
Sayula Almagres Ceiba 2 Lte. 6 3 meses 24 17.81083 -94.87291 10
Cada muestra se seccionó en trozos pequeños de 5mm, se desinfestaron por inmersión en
una solución de hipoclorito de sodio al 3% por 4 minutos, se lavaron por 3 minutos en agua
destilada estéril y se colocaron 4 trozos en cajas Petri con PDA. Las cajas se incubaron bajo
luz blanca a 28 °C durante 8 días; para identificación preliminar de cada cepa se realizaron
preparaciones permanentes para observación en microscopio compuesto. Colonias de L.
theobromae y Pestalotiopsis sp. se transfirieron a nuevas cajas Petri para purificación y
posteriormente para su caracterización morfológica y molecular.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
183
Pruebas de Patogenicidad. Para verificar la patogenicidad de los aislamientos de
Lasiodiplodia y Pestalotiopsis se realizó inoculaciones en plantas de 4.5 meses de edad. Se
utilizaron dos cepas de Lasiodiplodia seleccionados de acuerdo a la zona de estudio y uno de
Pestalotiopsis sp. Se cultivaron en PDA a 29 °C durante 14 días hasta tener presencia de
picnidios con conidios maduros y conidios de Pestalotiopsis sp., con el fin de asegurar la
infección.
La cantidad de inóculo usado fueron discos de PDA de 3mm de diámetro con micelio y
pequeños picnidios y conidios caso Pestalotiopsis. Cada cepa incluyendo el testigo discos de
PDA puro, se inocularon en 20 plantas, 15 establecidos en ambiente controlados con 12 horas
luz y 12 de oscuridad a un rango de temperatura 29-31 °C; en condiciones de campo se
establecieron 5 plantas con rango de temperatura 25-43 °C. Para la inoculación, se realizó
una herida de 5mm en forma de U invertida desprendiendo la corteza.
En la herida se colocó en contacto directo con el cambium expuesto, el disco de PDA, se
cubrió con la corteza extraída mas una capa de algodón estéril humedecida con agua destilada
estéril. Posteriormente se cubrió con cinta parafilm para proteger de la desecación y otros
contaminantes.
Análisis de micoflora. La colecta de muestras de suelo para análisis de micoflora se
dividió en 2 fases, con el fin de realizar comparaciones de dos épocas diferentes, la primera
correspondió a la época de sequía en mayo de 2019 y la segunda correspondió al de lluvia
que fue la segunda semana de agosto de 2019, se realizó de forma dirigida en árboles
sospechosos a Lasiodiplodia más un aparentemente sano. Se colectaron 400g de suelo a una
profundidad de 30cm dejando un espaciamiento de 15 cm de la raíz principal. Posteriormente
cada muestra se etiquetó para su traslado al laboratorio de la empresa UUMBAL y
posteriormente al laboratorio de Patología Forestal del Colegio de Postgraduados.
Para estimar la población de micoflora presente se utilizó la técnica de dilución de suelo
PDA-TS (Papa – Dextrosa – Agar- Tergitol – Estreptomicina) de Steiner y Watson (1965).
De cada muestra se pesaron 10g de suelo y se colocó dentro de un matraz Erlenmeyer con 90
mL de agua destilada estéril (un matraz por muestra) como solución madre; de esta solución

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
184
(10-1), se tomó 1 mL y se adicionó a un tubo de ensaye con 9 mL de agua destilada estéril
(10-2), la dilución se repitió y se obtuvo la dilución 10-3. De esta suspensión se tomó 1mL, el
cual se colocó 0.05 mL por caja en 10 repeticiones. Posteriormente, a cada caja se le adicionó
medio PDA-TS tibio, se agitó ligeramente para obtener una dispersión homogénea;
finalmente las cajas se mantuvieron a temperatura ambiente bajo luz blanca. El conteo de los
propágulos se realizó 6 días después de la siembra.
RESULTADOS Y DISCUSIÓN
De las 51 muestras vegetales colectadas y sembradas en medio PDA, se obtuvo presencia
de Lasiodiplodia theobromae en 34.5% y Pestalotiopsis sp. en 63.5%, siendo Sayula con el
mayor número de plantas enfermas. No se encontró una asociación directa con árboles que
presentaron síntomas de amarillamiento debido a que en árboles aparentemente sanos se
obtuvieron los mismos resultados. Con estos datos se corrobora que Lasiodiplodia causante
de la muerte descendente es un patógeno endófito y que cuando las coníferas u otras especies
están en condiciones de estrés se vuelven vulnerables al ataque del mismo (Gonzatto, et al.,
2015).
En cuanto a las pruebas de patogenicidad, el 80% de las plantas inoculadas mostraron
síntomas de muerte descendente a los 10 días después de la inoculación, con desarrollo de
picnidios alrededor de la zona de la herida. La manifestación rápida de los síntomas
probablemente se debe a las heridas profundas realizadas en la base del tallo de las plántulas,
facilitando la penetración del patógeno al tejido vascular (Ploetz et al., 1996; Pavlic et al.,
2004) así como al microambiente (>30°C y 100% de humedad relativa) generado por la
cubierta plástica y el algodón humedecido.
Para el análisis de micoflora, los hongos encontrados e identificados en el primer periodo
(mayo 2019) correspondieron a 2 géneros principalmente que son Aspergillus y Penicillium,
con un total de 6640 aislados, donde el 90% correspondió al género Aspergillus,
principalmente la especie A. niger. En el segundo periodo, correspondiente a época de lluvias
se encontraron 6371 aislados, al igual que en el periodo de sequía el género predominante
fue Aspergillus con un 80%. La predominancia de estos géneros se puede usar como
indicadores de baja fertilidad del suelo debido a que la diversidad y actividad fúngica

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
185
depende directamente de varios factores del suelo, como el pH, materia orgánica, el nivel de
humedad, aireación, temperatura, estación del año y composición de la vegetación
(Alexander, 1987). Por ello la baja fertilidad del suelo se puede considerar como un factor
que favorece la presencia de Lasiodiplodia, debido a que, al no haber suficiente
disponibilidad de nutrientes para los árboles, estos tienden a presentar estrés fisiológico.
AGRADECIMIENTOS
Se agradece al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la
beca otorgada en la maestría para el primer autor. Al Colegio de Postgraduados, Campus
Montecillo, por el equipo y las instalaciones donde se realizaron las actividades y la asesoría
de sus Profesores. A la Empresa UUMBAL por las instalaciones y al personal que nos
apoyaron durante los recorridos.
LITERATURA CITADA
Alexander, M. 1987. Introducción a la Microbiología del suelo. AGT editor, S.A. México. 491 p.
Cilliers, A. J., Swart, W. J., Wingfield, M. J. 1993. A review of Lasiodiplodia theobromae with particular reference to its occurrence on coniferous seeds. Suid -Afrikaaense Bosboutydskrif 166: 47-52
CONABIO. 1962. Comisión Nacional Para el Conocimiento y Uso de la Biodiversidad.http://www.conabio.gob.mx/conocimiento/info_especies
Gonzatto, M. C., Briao, M. M., Mezzomo R., and R.L. Silveira. 2015. Lasiodiplodia theobromae associated with seeds of Pinus spp. originated from the northwest of Rio Grande do Sul, Brazil. Forest Sciences, Piracicaba, v. 43, n. 107, p. 639-646.
Mohali S, Burgess T.I, and M.J. Wingfield 2005. Diversity and host association of the tropical tree endophyte Lasiodiplodia theobromae revealed using simple sequence repeat markers. Forest Pathology 35:385-396.
Pavlic, D., B. Slippers, T. A. Coutinho, M. Gryenhout, and M. J. Wingfield. 2004. Lasiodiplodia gonubiensis sp. nov., a new Botryosphaeria anamorph from native Syzygium cordatum in South Africa. Studies in Mycol. 50: 313-322.
Perry, J.P. 1991. The Pines of Mexico and Central America. Portland, Oregon. Tember Press. 231 p.
Ploetz, R. C., D. Benscher, A. Vazquez, A. Colls, J. Nagel, and B. Schaffer. 1996. A reexamination of mango decline in Florida. Plant Dis. 80: 664-668.
Steiner, G. W. y R.D. Watson. 1965. Use of surfactants in the soil dilution and plate count method. Phytopathology. 55: 728-730.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
186
Tucuch C.F.M., Palacios P.A., Ku N.R. y E.C. Guzmán. 2005. Manejo del cultivo de mango en el estado de Campeche. Campo Experimental Edzna, INIFAP. Folleto Técnico. Campeche, Camp., México. 33-34p.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
187
HONGOS ENDÓFITOS: UNA ALTERNATIVA BIOLÓGICA PARA
EL MANEJO DE NEMATODOS AGALLADORES
Manuel Silva-Valenzuela1, Emma Zavaleta-Mejía1, Rosa Helena Manzanilla-López3, Reyna Isabel
Rojas-Martínez1, Martha Lydia Macías Ruvalcaba2, Sergio Aranda-Ocampo1
1Fitosanidad- Fitopatología, Colegio de Postgraduados, 2Instituto de Química, Universidad Nacional Autónoma de México 3Centro de
Desarrollo de Productos Bióticos de México, Instituto Politécnico Nacional. [email protected]
INTRODUCCIÓN
En México Meloidogyne incognita, M. enterolobii y Nacobbus aberrans provocan severas
pérdidas económicas en diferentes cultivos. Su control se realiza con plaguicidas, lo que
representa un riesgo para la salud (Degenkolb y Vilcinskas, 2016). Por tal razón, la tendencia
internacional está enfocada en generar alternativas biológicas de manejo de nematodos
fitopatógenos, como el uso de hongos endófitos benéficos (HEB) (Compant et al., 2016). La
protección que los HEB proveen a la planta contra nematodos se explica por mecanismos de
acción como: parasitismo en huevos, juveniles de segundo estadio (J2) y hembras;
producción de compuestos nematostáticos y nematicidas; promoción del crecimiento de la
planta e inducción de resistencia (Romera et al., 2019; Schouten, 2016). Con base en estos
antecedentes se llevó a cabo el aislamiento de hongos endófitos de plantas de chile y tomate
con síntomas de agallamiento, con la finalidad de conocer su potencial como agentes de
control biológico de M. enterolobii, M. incognita y N. aberrans. Con 14 de los aislamientos
obtenidos se realizaron ensayos para determinar su capacidad parasítica en huevos y J2 de
los tres nematodos; el efecto nematostático y nematicida de sus filtrados sobre J2, y su efecto
promotor de crecimiento en plantas de chile y tomate. Los mejores resultados se obtuvieron
con los aislamientos S1JH2, S3R3B y XRC3, que fueron evaluados con el objetivo de
conocer su capacidad de inducir resistencia y su nivel de protección en chile (Capsicum
annuum var. Yolo Wonder) y tomate (Solanum lycopersicum var. Rio Grande) contra M.
enterolobii, M. incognita y N. aberrans.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
188
MATERIALES Y MÉTODOS
Material biológico. De raíces agalladas con M. enterolobii (Me), M. incognita (Mi) y N.
aberrans (Na), se extrajeron huevos y J2 de cada nematodo siguiendo el método descrito por
Vrain, (1977) y Villar-Luna et al. (2009). Los aislamientos endófitos S1JH2, S3R3B y XRC3
fueron reactivados en placas de papa-dextrosa-agar y el inóculo se obtuvo a partir de su
crecimiento en medio líquido papa-dextrosa. Las plántulas de chile var. Yolo Wonder y de
tomate var. Rio Grande, se obtuvieron de semillas desinfestadas con hipoclorito de sodio al
1% y germinadas bajo condiciones asépticas.
Combinaciones hospedante-aislamiento-nematodo evaluadas. Para los experimentos
de inducción de resistencia y protección contra la infección por los nematodos se evaluaron
las siguientes combinaciones: en chile S1JH2 contra Me y Na, S3R3B contra Mi y Na, y
XRC3 contra Me y Na; y en tomate S3R3B contra Mi y Na, y XRC3 contra Me y Na.
Ensayo inducción de resistencia. Plántulas de chile y tomate con el primer par de hojas
fueron trasplantadas en macetas conteniendo 40 g de arena. Para obtener plantas con doble
sistema radical, a las 2 semanas, la raíz de cada planta se separó en dos partes colocando cada
una de ellas en una maceta conteniendo 80 g de arena estéril. Enseguida, una de las macetas
se etiquetó como lado inductor (I) y la otra como lado respuesta (R) (Figura 1). A la siguiente
semana, el lado I se inoculó con 4 mL de una suspensión de 1 x 106 propágulos (fragmentos
de micelio) /mL del hongo a evaluar. La inoculación se repitió a los cinco y 10 días
posteriores. En el lado I de plantas testigo solo se aplicó agua estéril. Al décimo día, en el
lado R se inocularon 2,000 J2 de cada nematodo a evaluar. El experimento para cada
combinación (hospedante-hongo-nematodo) se realizó bajo un diseño completo al azar con
dos tratamientos y cuatro repeticiones. Las plantas se mantuvieron en una cámara de
crecimiento a 27±1 °C y con un fotoperiodo de 12 h luz y 12 h oscuridad. Para M. enterolobii
y M. incognita el ensayo se evaluó a las 5 semanas posteriores a la inoculación (spi) con el
nematodo y para N. aberrans a las 7 spi, y se registró el índice de agallamiento y la cantidad
de J2 por gramo de raíz. El ensayo se repitió una vez más.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
189
Figura 1. Plantas con doble sistema radical.
Ensayo de protección. Para cada combinación se estableció un experimento bajo un
diseño completo al azar con 10 repeticiones por tratamiento, los cuales fueron: Testigo (T),
Hongo endófito (H), Nematodo (N) y Hongo + Nematodo (H+N). Plántulas de chile y tomate
con el primer par de hojas se trasplantaron a macetas conteniendo 80 g de arena estéril.
Transcurrida una semana, los tratamientos H y H+N se inocularon con 4 mL de una
suspensión de 1 x 106 propágulos/mL del hongo a evaluar, la inoculación se repitió a los cinco
y 10 días posteriores al trasplante. En las plantas de los tratamientos T y N solo se aplicó
agua estéril. Al décimo día, las plantas de los tratamientos N y H+N fueron inoculadas con
2000 J2 de cada nematodo a evaluar. Las plantas se mantuvieron en un cuarto de crecimiento
a 27±1 °C y con un fotoperiodo de 12 h luz y 12 h oscuridad. Trascurridas 6 spi con el
nematodo se evaluó el experimento registrando el índice de agallamiento, peso seco de raíz
y de la parte aérea. Cada experimento se repitió una vez más.
Análisis estadístico. Previo al análisis de varianza (ANOVA) se determinó si los datos
obtenidos cumplían los supuestos estadísticos. La comparación de medias de los tratamientos
se realizó con la prueba de Tukey (P≤0.05). Se utilizó el programa “R Project for Statistical
Computing” versión 3.3.3.
RESULTADOS Y DISCUSIÓN
En chile los aislamientos endófitos que indujeron resistencia al ataque de N. aberrans
fueron S1JH2 y XRC3, dado que el índice de agallamiento y la cantidad de J2/g de raíz fue
significativamente menor a la observada en plantas no tratadas (Figura 2). Asimismo, en
tomate el aislamiento S3R3B indujo resistencia al ataque de M. incognita y de N. aberrans y

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
190
XRC3 solo para N. aberrans (Figura 3). Para M. enterolobii ninguno de los aislamientos
evaluados redujo el ataque de este nematodo (datos no mostrados).
Figura 2. Efecto de la inducción de resistencia por S3R3B, XRC3 y S1JH2 en jitomate y chile sobre N. aberrans. IA: Índice de
agallamiento, J2/g: Juveniles de segundo estadio por gramo de raíz, Na/Agua: lado inductor sin tratar /lado respuesta inoculado con N.
aberrans; Na/S3R3B: lado inductor inoculado con S3R3B/lado respuesta inoculado con N. aberrans, Na/XRC3: lado inductor inoculado
con XRC3/lado respuesta inoculado con N. aberrans y Na/S1JH2 lado inductor inoculado conS1JH2 /lado respuesta inoculado con N.
aberrans. La línea en cada barra corresponde a la desviación estándar. Cada barra representa el promedio de 8 repeticiones correspondiente
a dos ensayos y barras con la misma letra no son significativamente diferentes (Tukey, p˂0.05).
Figura 3. Efecto de la inducción de resistencia por S3R3B en jitomate y chile sobre M. incognita. IA: Índice de agallamiento, J2/g: Juveniles
de segundo estadio por gramo de raíz, Mi/Agua: lado inductor sin tratar/lado respuesta inoculado con M. incognita; Mi/S3R3B: lado
inductor inoculado con S3R3B/lado respuesta inoculado con M. incognita. La línea en cada barra corresponde a la desviación estándar.
Cada barra representa el promedio de 8 repeticiones correspondiente a dos ensayos y barras con la misma letra no son significativamente
diferentes (Tukey, p˂0.05).
En el ensayo de protección, plantas de chile inoculadas con S1JH2 y XRC3 redujeron
significativamente el índice de agallamiento y su presencia permitió acumulación de biomasa
(Cuadro 1 y 2).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
191
Cuadro 1. Efecto del hongo endófito S1JH2 en el agallamiento radical inducido por Nacobbus aberrans en chile (Capsicum annuum var. Yolo Wonder)
Tratamiento Nacobbus aberrans
Chile IA NR PSR PSF
T 0±0 c 1±0 b 0.07±0.01 b 0.21±0.03 a H 0±0 c 1±0 b 0.09±0.02 b 0.23±0.03 a N 87±6.32 a 2.5±0.97 a 0.06±0.02 b 0.09±0.01 c
H+N 42.5±12.07 b 1±0 b 0.13±0.01 a 0.14±0.01 b T: Testigo; H: Hongo endófito; N: Nematodo; H+N: Hongo endófito + Nematodo; IA: índice de agallamiento; NR: Necrosis radical; PSR: peso seco raíz; PSF: peso seco follaje. Cada media representa el dato de 20 repeticiones provenientes de dos ensayos, medias con la misma letra en cada columna no son significativamente diferentes (Tukey, P˂0.05).
En tomate se observó que los aislamientos XRC3 y S3R3B protegieron a la planta del
ataque de N. aberrans y M. incognita (Cuadros 2 y 3, respectivamente). Cuadro 2. Efecto del hongo endófito XRC3 en el agallamiento radical inducido por Nacobbus aberrans en chile (Capsicum annuum var. Yolo Wonder) y tomate (Solanum lycopersicum var. Rio Grande)
Tratamiento Nacobbus aberrans
Chile IA NR PSR PSF
T 0±0 c 1±0 a 0.05±0.01 b 0.17±0.03b H 0±0 c 1±0 a 0.10±0.01 a 0.23±0.02 a N 87.0±6.32 a 1.3±0.48 a 0.11±0.01 a 0.08±.0.01 d
H+N 72.5±7.9 b 1.5±0.84 a 0.10±0.02 a 0.12±.0.01 c
Tratamiento Tomate IA NR PSR PSF
T 0±0 c 1±0 b 0.07±0.00 c 0.27±0.03 b H 0±0 c 1±0 b 0.12±0.01 b 0.31±0.01 a N 78.0±6.32 a 1.7±0.48 a 0.14±0.02 ab 0.17±0.01 c
H+N 55.0±10.54 b 1.2±0.42 b 0.17±0.02 a 0.24±0.03 b T: Testigo; H: Hongo endófito; N: Nematodo; H+N: Hongo endófito + Nematodo; IA: índice de agallamiento; NR: Necrosis radical; PSR: peso seco raíz; PSF: peso seco follaje. Cada media representa el dato de 20 repeticiones provenientes de dos ensayos, medias con la misma letra en cada columna no son significativamente diferentes (Tukey, P˂0.05).
En contraste, ningún aislamiento evaluado en este ensayo protegió a las plantas del ataque
de M. enterolobii (datos no mostrados) confirmando su alta virulencia y agresividad (Rutter
et al., 2019). Cuadro 3. Efecto del hongo endófito S3R3B en el agallamiento radical inducido por Meloidogyne incognita en tomate (Solanum lycopersicum var. Rio Grande)
Tratamiento Meloidogyne incognita
Tomate IA NR PSR PSF
T 0±0 c 1±0 b 0.06±0.02 b 0.24±0.05 a H 0±0 c 1±0 b 0.09±0.03 b 0.24±0.06 a N 99±3.16 a 2.1±1.87 a 0.06±0.03 b 0.06±0.02 c
H+N 82±9.48 b 1.3±0.9 ab 0.15±0.03 a 0.14±0.02 b T: Testigo; H: Hongo endófito; N: Nematodo; H+N: Hongo endófito + Nematodo; IA: índice de agallamiento; NR: Necrosis radical; PSR: peso seco raíz; PSF: peso seco follaje. Cada media representa el dato de 20 repeticiones provenientes de dos ensayos, medias con la misma letra en cada columna no son significativamente diferentes (Tukey, P˂0.05).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
192
Las combinaciones S1JH2-chile-Na, S3R3B-tomate-Mi, XRC3-chile-Na y XRC3-
tomate-Na presentaron resultados similares a los obtenidos en los ensayos de inducción de
resistencia; lo que sugiere que uno de los posibles mecanismos de acción involucrados en la
protección sea la inducción de resistencia. Una respuesta similar se reporta para M. incognita
en plantas de tomate inducido por Fusarium oxysporum 162 y para Trichoderma harzianum
T-78 (Romera et al., 2019; Dababat y Sikora, 2007). La detección y cuantificación de los
niveles de expresión de genes relacionados con la defensa en presencia y ausencia del hongo
y del nematodo contribuirían a tener un mejor entendimiento de este mecanismo de
protección.
Los resultados obtenidos indican que las combinaciones promisorias para llevar a cabo
evaluaciones en invernadero son: S1JH2-chile-Na, S3R3B-tomate-Mi, XRC3-chile-Na y
XRC3-tomate-Na.
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por la beca otorgada para la
realización de mis estudios de doctorado (CVU 484085). Al consejo particular por el
seguimiento, orientación y facilidades proporcionadas en la realización de esta investigación.
LITERATURA CITADA
Compant S., Saikkonen K., Mitter B., Campisano A., Mercado-Blanco J. 2016. Editorial special issue: soil, plants and endophytes. Plant Soil 405: 1 – 11.
Dababat, A. E. A., Sikora R. A. 2007. Induced resistance by the mutualistic endophyte, Fusarium oxysporum strain 162, toward Meloidogyne incognita on tomato. Biocontrol Science and Technology 17: 969-975.
Degenkolb, T., Vilcinskas A. 2015. Metabolites from nematophagous fungi and nematicidal natural products from fungi as an alternative for biological control. Part I: metabolites from nematophagous ascomycetes. Applied Microbiology and Biotechnology 100: 3813-3824.
Romera, F. J., García, M. J., Lucena C., Martínez-Medina A., Aparicio M. A., Ramos J., Alcántara E., Angulo M., Pérez-Vicente R. 2019. Induced Systemic Resistance (ISR) and Fe Deficiency Responses in Dicot Plants. Frontiers in Plant Science 10: 1-17.
Schouten A. 2016. Mechanisms involved in nematode control by endophytic fungi. Annual Review of Phytopathology 54: 121–142.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
193
PAQUETE TECNOLÓGICO PARA EL MANEJO DE LOS
NEMATODOS Nacobbus aberrans y Meloidogyne incognita EN
TOMATE EN INVERNADERO
Marco Antonio Magallanes-Tapia1, Cristian Nava-Díaz1, Howard Ferris2, Daniel L. Ochoa-
Martínez1, Juan Manuel González-Camacho1, Ignacio Cid del Prado-Vera1*
1 Posgrado en Fitosanidad-Fitopatología, Colegio de Postgraduados, Campus Montecillo, Carretera México-Texcoco km 36.5, Montecillo,
56230, Texcoco, México. 2 Department of Entomology and Nematology, University of California, One Shields Avenue, Davis, CA 95616,
USA. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
En México, el tomate (Solanum lycopersicon L.) representa 37 % del valor total de las
exportaciones hortícolas y 16 % de las agropecuarias, convirtiéndolo en el principal producto
hortícola de exportación (SIAP-SAGARPA, 2015). Los nematodos agalladores, Nacobbus
aberrans y Meloidogyne sp., afectan a este cultivo y causan perdidas económicas
considerables, al reducir su producción entre 7 y 70 % (Cid del Prado et al., 1997; 2001;
Moens et al., 2009). Las estrategias de control de nematodos incluyen la aplicación de
nematicidas por su rápido efecto; sin embargo, el costo económico y repercusión ambiental
es muy elevado. En este sentido, es importante la aplicación de un control integrado con
técnicas que complementen el manejo de los nematodos agalladores en tomate y otros
cultivos. El uso de microorganismos antagonistas, enmiendas orgánicas, biofumigación,
solarización del suelo y acolchado, han mostrado eficacia en el manejo de nematodos
agalladores; no obstante, su empleo ha sido aislado o poco integrado (Pérez-Rodríguez et al.,
2007; Herrera-Parra et al., 2009; Cid del Prado et al., 2010). Por lo anterior, la siguiente
investigación tuvo como objetivo evaluar la combinación de la biofumigación, agentes de
control biológico y nematicidas químicos, para el manejo integrado de los nematodos
Nacobbus aberrans y Meloidogyne incognita en tomate, bajo condiciones de invernadero.
MATERIALES Y MÉTODOS
La investigación se llevó a cabo en un invernadero comercial de tomate, variedad Condor
(BHN Seeds), de 475 m2, con antecedentes de infestación por N. aberrans y M. incognita,

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
194
ubicado en San Diego, Texcoco, México. La estrategia de manejo comprendió la
combinación de la biofumigación [brócoli + estiércol de borrego], agentes de control
biológico y nematicidas químicos para el manejo de las especies de nematodos mencionadas.
El experimento fue un diseño de bloques completos al azar generalizados (DEBAG), con seis
tratamientos [T1, Purpureocillium lilacinum (producto comercial®); T2, Pochonia
chlamydosporia + Fluopiram (producto comercial®); T3, P. lilacinum (producto comercial®)
+ Fluopiram (producto comercial®); T4, Fluopiram (producto comercial®); T5, Avermectina
(producto comercial®); T6, testigo absoluto) y cuatro repeticiones. Las unidades
experimentales (UE) consistieron de 12 plantas en un área de 2.4 m2. Las variables a evaluar
incluyeron la población de nematodos agalladores en el suelo, el índice de agallamiento de
las raíces (IAR) y el rendimiento de tomate. El análisis del patrón de distribución espacial,
de dichas evaluaciones, se efectuó con el software Surfer (v15).
La densidad de población inicial y la fluctuación de N. aberrans y M. incognita, se estimó
en cinco colectas de suelo a una profundidad de 20 cm, antes del trasplante de plántulas y
posterior al trasplante, cada 30 días. La extracción de nematodos se llevó a cabo por la técnica
de tamizado (Cobb, 1918) y centrifugado-flotación (Jenkins, 1964; Ayoub, 1977). El IAR se
llevó a cabo en seis evaluaciones no destructivas, iniciando 30 días después del trasplante
(ddt) y una cada mes. Se evaluó una planta por tratamiento, tomando como base el porcentaje
de raíz agallada propuesto por Bridge y Page (1980), que incluye los niveles 1 al 10. El
rendimiento de tomate se estimó con la cosecha de los frutos de cada tratamiento por UE.
Con los datos obtenidos se realizó un análisis de varianza (ANOVA) y comparación múltiple
de medias utilizando la prueba de Tukey (P > 0.05) en el paquete estadístico R para Mac OS
X (versión 3.4.3).
RESULTADOS Y DISCUSIÓN
La densidad total promedio de N. aberrans varió entre 28 y 81 especímenes en 200 g de
suelo. El T3 y el T5 registraron los promedios totales bajo y alto, respectivamente, en todo
el ciclo del cultivo. La población más baja de este nematodo agallador se presentó a los 30
ddt, durante la etapa de floración del cultivo; mientras que la más alta a los 60 ddt

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
195
(fructificación y formación del fruto). En las evaluaciones realizadas predominaron los J2,
con un bajo porcentaje de hembras inmaduras y machos. El ANOVA no mostró diferencias
significativas entre los tratamientos con respecto a la población acumulada de N. aberrans
en cinco colectas; por lo tanto, no hay evidencias para indicar cuál fue el mejor tratamiento,
bajo estas condiciones de experimentación. Se registraron diferencias entre bloques (B) y la
comparación de medias muestra que el B4 y el B1 presentan la mayor y menor población del
fitoparásito, respectivamente. Es importante señalar que la población de M. incognita fue
muy baja en todas las colectas ejecutadas, con un máximo de tres organismos registrados (J2)
durante el experimento.
Por otro lado, referente al IAR promedio de las plantas evaluadas, los niveles fluctuaron
entre 3 y 5 durante el desarrollo del cultivo, en los distintos tratamientos, destacando el testigo
absoluto (T6), donde su valor se mantuvo en cuatro, en las seis evaluaciones realizadas;
mientras que T2, T4 y T5 registraron el nivel más bajo (3). El ANOVA no indicó diferencias
significativas entre los tratamientos, lo que indica que estos producen el mismo efecto en la
planta durante el cultivo. En cambio, no todos los bloques producen el mismo efecto, la
comparación de medias muestra que el B4 es significativamente diferente, con un mayor IAR
promedio (3.8) mayor con respecto al resto de los bloques. Es importante mencionar que el
síntoma de “rosario”, ocasionado por N. aberrans, predominó en todas las evaluaciones del
IAR. Se ha registrado que la aplicación de residuos de brasicas, como biofumigantes,
disminuye el IAR, ya que contienen glucosinolatos que se hidrolizan por la enzima
mirosinasa y producen epthionitriles, isotiocianatos y tiocianatos, que son compuestos con
propiedades nematicidas (Kirkegaard et al., 1993). Con relación a ello, en un estudio
realizado en tomate, infectado por M. incognita, se disminuyó el IAR en 50 % por medio de
la aplicación de brócoli (Kaskavalci et al., 2009). Asimismo, se ha reportado que la
incorporación al suelo de residuos de crucíferas y sorgo, en combinación con P.
chlamydosporia, disminuye las poblaciones de Meloidogyne sp. en invernadero (Cid del
Prado et al., 2010). Además, Bello et al. (2004) recomiendan una dosis general de 50 t/ha de
brasicas; la eficacia de la biofumigación puede variar según las propiedades bioquímicas del
suelo y el método de aplicación de las plantas. En el presente estudio, se aplicaron 60 t/ha de

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
196
brócoli triturada e incorporada a 30 cm de profundidad en el suelo para el proceso de
biofumigación, durante 30 días.
Con relación al rendimiento total acumulado de tomate, en 14 cosechas, el T2 registró la
mayor producción con 685 t/ha, en tanto que el T3, la menor (620 t/ha). No obstante, el
análisis estadístico no indica diferencias significativas entre ellos. Por el contrario, se
registraron diferencias significativas entre los bloques, donde la comparación de medias
señala que el B4 y el B1 presentaron el mayor y menor rendimiento, respectivamente. Al
respecto, se reportó que los tratamientos con nematicidas químicos no tuvieron una influencia
significativa en el rendimiento de pepino afectado por altas densidades de M. incognita
(Hajihassani et al., 2019).
Se ha documentado la eficacia de agentes de biocontrol (P. lilacinum y P.
chlamydosporia) contra N. aberrans y M. incognita (Brand et al., 2004; Flores-Camacho et
al., 2007); a pesar de ello, en el presente estudio no se evidenció el manejo efectivo de ambas
especies de nematodos agalladores mediante los tratamientos estudiados. Este resultado
coincide con lo reportado por Gortari y Hours (2016), quienes mencionan que no se observó
una actividad antagónica concluyente de P. lilacinum sobre N. aberrans en tomate. Así
mismo, cabe señalar que en el presente trabajo se evaluaron dos combinaciones de manejo
(T2 y T3), los cuales no fueron significativamente diferentes con el resto de los tratamientos.
Esta observación coincide con un estudio donde se documentó el efecto de nematicidas
químicos y P. lilacinum sobre N. aberrans, reportándose que el nematicida afectó
negativamente al hongo nematófago (Eguiguren-Carrión, 1995). Es importante mencionar
que Fluopiram (Pyridinyl-ethyl-benzamide) inicialmente se desarrolló como un fungicida, de
modo que la molécula podría estar afectando el desarrollo de las especies de hongos en
mención (T2 y T3) (Labourdette et al., 2011).
Por otro lado, se observó que el patrón de distribución de N. aberrans en el suelo, el IAR
y el rendimiento de tomate, fueron más evidentes en el B4 del sitio experimental. Este hecho
se puede asociar al declive que tiene el terreno, área en la que se acumuló mayor humedad.
Los principales grupos tróficos registrados durante la colecta de muestras incluye a

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
197
fitoparásitos (Tylenchorhynchus sp., Helicotylenchus sp., Tylenchus sp., Hoplolaimus sp.,
Criconematidos), Micófagos (Acrobleles sp. y Aphelenchus sp.) y bacteriófagos
(Rhabditidos).
AGRADECIMIENTOS
Agradecemos al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la
beca otorgada para los estudios de doctorado del primer autor; al Colegio de Postgraduados
Campus Montecillo por el equipo y las instalaciones donde se realizaron los experimentos y
la asesoría de sus Profesores.
LITERATURA CITADA
Ayoub, M.S. 1977. Plant nematology: An agricultural training aid. 157 p. State of California, Department of Food and Agriculture, Division of Plant Industry, Laboratory Services Nematology, Sacramento, California, USA.
Bello, A., J.A. López-Pérez, A. Garcia-Alvarez, R. Sanz, A. Lacasa. 2004. Biofumigation and nematode control in the Mediterranean region, in: R.C. Cook, D.J. Hunt (Eds.), Proceedings, IV International Congress of Nematology 133-149.
Brand, D., S. Roussos, A. Pandey, P.C. Zilioli, J. Pohl and C.R. Soccol. 2004. Development of a bionematicide with Paecilomyces lilacinus to control Meloidogyne incognita. App. Biochem. Biotech. 118: 81-88.
Bridge, J. and S. Page. 1980. Estimation of root-knot nematode infestation levels on roots using a rating chart. Trop. Pest Manag. 26: 296-298.
Cid del Prado, V. I., M.A. Lucero-Pallares and I. Pérez-Rodríguez. 2010. Biofumigation a very efficient alternative for the control of root-knot nematode Meloidogyne spp. in vegetables pro- duced in house shadows. Nematropica 40: 128.
Cid del Prado, V.I., A.J. Cristóbal, A.G.E. Franco y L.R.H. Manzanilla. 1997. Manejo de poblaciones de Nacobbus aberrans en el cultivo de jitomate (Lycopersicon sculentum Mill.). En Avances en la investigación (pp. 171–173). Montecillo: Instituto de Fitosanidad. Colegio de Postgraduados.
Cid del Prado, V.I., S.A. Tovar, y J.A. Hernández. 2001. Distribución de especies y razas de Meloidogyne en México. Rev. Mex. Fitopatol. 19: 32-39.
Cobb, N.A. 1918. Estimating the nema population of the soil. Agrc. Tech. Circ. Bur. Pl. Ind. U.S. Dep. Agric, N.1.
Eguiguren-Carrion, R. 1995. Control of Meloidogyne incognita and Nacobbus sp. with Paecilomyces lilacinus in the green house and effect of nematicides on the fungus, Biocont. 1: 41-49.
Flores-Camacho, R., R.H. Manzanilla López, I. Cid del Prado-Vera and A. Martínez-Garza. 2007. Control de Nacobbus aberrans (Thorne) Thorne y Allen con Pochonia chlamydosporia (Goddard) Gams y Zare. Rev. Mex. Fitopatol. 25: 25-34.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
198
Gortari, M.C., R.A. Hours. 2016. Purpureocillium lilacinum LPSC # 876: Producción de conidias en cultivos sobre sustratos sólidos y evaluación de su actividad sobre Nacobbus aberrans en plantas de tomate. Rev. Fac. Agron. 115: 239-249.
Hajihassani, A., R.F. Davis and P. Timper. 2019. Evaluation of selected nonfumigant nematicides on increasing inoculation densities of Meloidogyne incognita on cucumber. Plant Dis. https://doi.org/10.1094/PDIS-04-19-0836-RE
Herrera-Parra, E., J. Cristóbal-Alejo and J.M. Tún-Suárez. 2009. Water extracts of Calea urtici-folia Mill. for the control of Meloidogyne incognita. Nematropica 39: 289-296.
Jenkins, W.R. 1964. A Rapid Centrifugal-Flotation Technique for Separating Nematodes from Soil. Plant Disease Report 48: 692.
Kaskavalci, G., Y. Tuzel, O. Dura, G.B. Oztekin. 2009. Effects of alternative control methods against Meloidogyne incognita in organic tomato production. Ekoloji 18: 23-31.
Kirkegaard, J.A., P.A. Gardner, J.M. Desmarchelier and J.F. Angus. 1993. Biofumigationusin Brassica species to control pests and diseases in horticulture and agriculture. In: Wratten, M., Mailer, R.J. (Eds.), 9th Australian Research Assembly on Brassicas. British Society for Plant Pathology, Agricultural Research Institute, Wagga Wagga, pp. 77-82.
Labourdette, G., H. Lachaise, H. Rieck and D. Steiger. 2010. Fluopyram: efficacy and beyond on problematic diseases. In: Modern Fungicides and Antifungal Compounds VI. 16th International Reinhardsbrunn Symposium, Friedrichroda, Germany.
Moens, M., R.N. Perry, J.L. Starr. 2009. Meloidogyne species: a diverse group of novel and important plant parasites. In: Perry, R.N., M. Moens, J.L. Starr (eds). Root-knot nematodes. CABI Publishing, Wallingford, pp 1-17.
Pérez-Rodríguez, I., A. Doroteo-Mendoza and F. Franco-Navarro. 2007. Isolates of Pochonia chlamydosporia var. chlamydosporia from Mexico as potencial biological control agents of Nacobbus aberrans. Nematropica 37: 127-134.
SIAP (Servicio de Información Agroalimentaria y Pesquera) SAGARPA (Secretaría de Agricultura Ganadería Desarrollo Rural Pesca y Alimentación). 2015. Producción agrícola. http://www.siap.gob.mx/cierre-de-la-produccion-agricola-por-estado/

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
199
GANANCIAS EN CICLOS DE SELECCIÓN S1 RECURRENTE DE
MAÍZ (Zea mays L.) PARA RENDIMIENTO Y RESISTENCIA AL
CARBÓN DE LA ESPIGA (Sporisorium reilianum f.sp. zeae)
Gelasino Díaz Ramírez1, María Del C. Castillo1, Carlos De León1*, Daniel Nieto Ángel1
1 Fitosanidad-Fitopatología. Colegio de Postgraduados. Km. 36.5 Carretera México-Texcoco, Montecillo, Texcoco 56230, Estado de
México. * Autor de correspondencia: [email protected]
INTRODUCCIÓN
El cultivo de maíz ocupa el primer lugar mundial dentro de los cereales, con una producción
de 1 134 Mt que superan al trigo y arroz (FAOSTAT, 2017). México produce un total de 21
185 003 t con un rendimiento promedio 3.9 t ha-1 (SIAP, 2018). Las enfermedades son la
principal causa que limita la producción. El carbón de la espiga causado por Sporisorium
reilianum f. sp. zeae causa pérdidas considerables, ya que el porcentaje de incidencia está
correlacionado directamente con el porcentaje de pérdida en producción (Fullerton et al.,
1974). Esta enfermedad se encuentra distribuida en los estados de Guanajuato, Jalisco,
Michoacán, Tamaulipas, Hidalgo y el Estado de México (Sánchez, 1988; CESAVEM 2006).
Debido a esto, en 2006 y 2007 el Colegio de Postgraduados (CP), el Servicio Nacional de
Sanidad, Inocuidad y Calidad Alimentaria (SENASICA) y la Dirección General de Sanidad
Vegetal crearon el proyecto PM0531 y PM0542 que contempla el mejoramiento de caracteres
agronómicos de interés y resistencia al carbón de la espiga en dos poblaciones de maíz con
amplia base genética, una con endospermo blanco y otra con endospermo amarillo. En esta
investigación se evaluaron seis ciclos de selección (C0-C5) de la población blanca y cinco
ciclos de selección (C0-C4) de la amarilla, los cuales se obtuvieron por selección recurrente
intrapoblacional de familias S1. Se evaluaron las ganancias genéticas por ciclos de selección
para rendimiento, incidencia del carbón de la espiga y otros caracteres agronómicos. El
objetivo del proyecto es generar variedades sintéticas con mayor rendimiento, resistentes al
carbón de la espiga, con bajo costo de la semilla, que el agricultor puede sembrar hasta por
cinco ciclos sin disminuir el rendimiento, ya que la variedad sintética se mantiene estable.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
200
MATERIALES Y MÉTODOS
Material genético: En al año 2006 se colectaron entre 35 y 45 variedades locales e
híbridos que se cultivan en la región del altiplano para generar una amplia base genética, que
fue el punto de partida para generar los diferentes ciclos de selección de poblaciones con
endospermo blanco y amarillo, respectivamente. Se recombinaron por dos años por el método
modificado de medios hermanos y posteriormente se siguió el programa de mejoramiento
por selección recurrente intrapoblacional de familias S1.
Siembra de los diferentes ciclos de selección: La siembra de los ciclos de selección se
realizó en Mixquiahuala, Hidalgo en el ciclo de primavera-verano de 2019. Se evaluaron seis
ciclos de la población con endospermo blanco (C0-C5), cinco ciclos de la población con
endospermo amarillo (C0-C4) y se incluyó un híbrido comercial como testigo (BG1384W).
Se utilizó un diseño de bloques completos al azar con tres repeticiones; se sembraron 39
parcelas, cada una en surcos de 3 m de largo con 22 plantas por surco, 0.16 m de distancia
entre plantas y 0.8 m entre surcos, con una densidad de población de 80 000 plantas ha-1.
Variables a estudiar: Se analizaron las ganancias genéticas de cada ciclo de selección
para rendimiento, biomasa seca, número de hojas, altura de planta y mazorca, días a floración
masculina y femenina, aspecto de planta y mazorca, incidencia de roya y carbón de la espiga.
Se realizó un análisis de varianza y comparación de medias con la prueba de Diferencia
Mínima Significativa (DMS) al nivel de 0.05 de probabilidad. Posteriormente se realizó un
análisis de regresión lineal para los caracteres agronómicos evaluados, con en el fin de
determinar el progreso del mejoramiento genético entre los diferentes ciclos de selección.
RESULTADOS Y DISCUSIÓN
En los Cuadros 1 y 2, se presentan los promedios de los caracteres evaluados para las
poblaciones con endospermo blanco y amarillo. No se contempló el análisis de varianza ni
la regresión lineal de los ciclos 2 de ambas poblaciones porque se obtuvieron pocas plantas
debido a problemas de germinación. Para la mayoría de los caracteres evaluados no hubo
diferencias estadísticas con excepción al número de hojas totales de la población blanca que
mostró diferencia significativa al 0.001. Sin embargo, el análisis de regresión lineal para las
poblaciones blanca y amarilla mostraron ganancias para cada ciclo evaluado en los caracteres

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
201
agronómicos (Cuadro 3), excepto para aspecto de mazorca que aumento en un 10 % en la
población blanca y 1.8% en la población amarilla. La incidencia de carbón para la población
amarilla mostró un incremento de 12.5% por ciclo, cuando se esperaba una disminución en
ambas variables. Este comportamiento se debe a que las semillas no se inocularon y a la
distribución irregular del inóculo en campo. No obstante, se obtuvo un progreso genético en
los diferentes ciclos de las dos poblaciones evaluadas. Los resultados obtenidos para las
variables de interés son similares con lo reportado por Mendoza et al. (2000), quienes
evaluaron cuatro ciclos de selección y reportaron una ganancia genética por ciclo de 3.09 %
para rendimiento. Maya et al. (2002) evaluaron dos poblaciones de maíz y obtuvieron una
ganancia genética por ciclo de 8.3% para la variable rendimiento, una reducción en número
de días del -2.3 % para floración femenina y aumento del 8.5 % en sanidad de mazorca por
ciclo. Shah et al. (2007), reportaron una ganancia genética del 19 % para rendimiento y una
reducción en severidad de -26 % por ciclo para Bipolaris maydis. Ruiz de Galarreta y Álvarez
(2000) encontraron una respuesta positiva para rendimiento, acame de plantas y humedad de
grano, pero los días a floración femenina aumentaron conforme avanzaban los ciclos de
selección. La variabilidad en la respuesta de las poblaciones evaluadas se debe a diversos
factores como la heredabilidad de los caracteres evaluados, el genotipo, la interacción
genotipo-ambiente, así como posibles errores de muestreo. En este trabajo se encontró que
la población de endospermo blanco mostró mejor respuesta a la selección recurrente de
familias S1 con respecto a la población amarilla.
AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
202
Cuadro 1. Medias de los caracteres agronómicos evaluados de la población de endospermo blanco.
Ciclos Hojas Indice alt
Aspecto Enfermedades Rend tot ASI Maz Pta Carbón Roya t ha-1
C0 19.8 a 0.6 a 3.6 ab 1.1 a 3 a 10 a 16.8 a 11.1 a C1 17.5 b 0.5 ab 3.6 ab 1.3 a 2.5 ab 4.2 a 7.2b 11.3 a C3 16.1 c 0.5 ab 4.6 a 1.8 a 2 ab 2.4 a 5.9b 12.1 a C4 17.5 b 0.5 ab 2.6 c 1.6 a 3 a 1.3 a 6.4b 11.5 a C5 17.1 bc 0.6 a 3.6 ab 1.5 a 1.6 c 2.7 a 7.3b 12.4a
TEST 16.8 bc 0.5 c 2.3 c 1.6 a 2.3 ab 1.4 a 8 b 11.3 a
DMS 1.3 0.09 1.6 0.7 1.2 7.7 7 4.1
CV 4.3 9.7 27.7 26.8 29 25.1 42 21.8 Cuadro 2. Medias de los caracteres agronómicos evaluados de la población de endospermo amarillo.
Ciclos Hojas
Indice alt
Aspecto Enfermedades Rend tot ASI Maz Pta Carbón Roya t ha-1
C0 19.5 a 0.6 a 4 a 1.1 b 3 a 6.9 a 15.2a 15.4 a C1 18.7 a 0.6 a 4.3 a 1.3 ab 2 b 10.4 a 7.7 b 12.3 ab C3 19 a 0.6 a 2.3 a 1.5 ab 3 a 8.4 a 5.7 b 9.7 ab C4 19.3 a 0.5 a 2.6 a 1.1 b 2.6 ab 10.5 a 6.6 b 16.6 a
TEST 18.8 a 0.5 a 2.6 a 2 a 3 a 1.4 a 7.2 b 12.1 ab
DMS 1.4 0.09 2.3 0.7 0.9 9.3 6.8 7.6 CV 4.2 9.5 42.8 28.2 19.4 68 39 36

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
203
Cuadro 3. Ganancias genéticas promedio por ciclos de los caracteres agronómicos evaluados en la población blanco y amarillo.
Carácter ∆G ciclo-1 (%) ∆G ciclo-1 (%) población blanco población amarillo
Índice de altura -0.5 -5 ASI (Anthesis-Silking Interval) -2.7 -16.3 Aspecto de planta -4.9 -0.6 Aspecto de mazorca 10 1.8 Número de hojas totales -2.7 -0.15 Materia seca 6.9 12.1 Incidencia del carbón -17.5 12.5 Incidencia de roya -11.7 -18.3 Incidencia pudrición de mazorca -7.9 -16.8 Rendimiento 2.5 -0.6
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por el financiamiento otorgado
para la realización de estudios de maestría.
LITERATURA CITADA
CESAVEM. 2006. Carbón de la espiga del maíz (Sphacelotheca reiliana). Comité Estatal de Sanidad Vegetal del Estado de México. Folleto para productores. Toluca, edo. Méx., México. 8p.
FAOSTAT (Food and Agriculture Organization). © FAO Statistics Division. 2017. Fullerton R., A., D. Scott J., G. Graham J. 1974. Effects of fungicides on the control of head
smut of maize, and the relationship between infection level and crop yield. New Zealand J. Extl Agri. 2:177-179.
Mendoza M., A. Oyervides., L. Latournerie., H. de León. 2000. Selección recurrente en maíz para el trópico húmedo. Agron. Mesoam. 11(1):89-93.
Maya L., J., y D. Ramirez J. 2002. Selección recurrente en tres poblaciones de maíz para el subtrópico de México. Rev. Fitotec. Mex. 25(2):201-207.
Ruiz de Galarreta J., y I. Álvarez A. 2000. Six cycles of S1 recurrent selection in two Spanish maize synthetics. Span. J. Agric. Res. 5:193–198
Sánchez, H. 1988. Enfermedades del maíz. pp.1-11. In: Teliz, D. (ed.). Enfermedades del maíz, frijol, trigo y papa. Colegio de Postgraduados. Montecillo, Méx., México. 84p.
Shah S., S., H. Rahman., I. Khalil H., and M. Iqbal. 2007. Recurrent selection for maydis leaf blight resistance and grain yield improvement in maize. Pak. J. Biol. Sci. 10(20): 3632-3637.
SIAP (Servicio de Información Agroalimentaria y Pesquera). Anuario Estadístico de la Producción Agrícola 2018. http://nube.siap.gob.mx/cierre_agricola/.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
204
¿PUEDE Aedes aegypti L. ADAPATRSE A LA ZONA DEL LAGO DE
TEXCOCO Y AMENAZAR A MÁS DE 36 MILLONES DE
PERSONAS?
Selene Bello-Rivera1, J. Concepcion Rodriguez-Maciel1*
1 Posgrado en Fitosanidad-Entomología y Acarología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230
Texcoco, Estado de México, México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
El mosquito, Aedes aegypti L. (Diptera:Culicidae) es la especie de insecto más peligrosa
para el ser humano pues transmite enfermedades severas como dengue hemorrágico, dengue
clásico, chikungunya, virus del Mayaro, zika y fiebre amarilla (Rigau-Pérez et al., 1998).
Su distribución geográfica se sigue ampliando debido al aumento de la temperatura derivada
del calentamiento global (Marquez et al., 2018) y a la capacidad de la larva para vivir en
aguas contaminadas (McNaughton et al., 2018). Kuri-Morales et al., (2017) detectó
individuos autóctonos de Ae. Aegypti en la Ciudad de México, a no más de 20 km de esta
zona lacustre del Lago de Texcoco (LT). Este lugar se localiza aproximadamente a 2 km de
la Ciudad de México y tiene una superficie de 2,252 ha; alberga cuerpos de agua salina con
valores de pH que van de 8-11 (CONAGUA, 2014; Guzmán-Quintero et al. 2007). En
consecuencia, es cuestión de tiempo para que este vector pueda tener las condiciones de
temperatura para adaptarse a vivir en esta amplia zona, a menos de que el factor agua sea
una limitante. Por tanto, el objetivo del presente estudio consistió en determinar la influencia
del tipo de agua de la zona lacustre del Lago de Texcoco (LT) sobre el desarrollo de larvas
de Ae. Aegypti.
MATERIALES Y MÉTODOS
Población de insectos. Se utilizó la raza New Orleans de A. aegypti susceptible a
insecticidas, proporcionada por La Facultad de Ciencias Biológicas de la Universidad
Autónoma de Nuevo León, México.
Tratamientos. Se utilizaron cinco muestras de agua: a) agua de la llave del Colegio de
Postgraduados, mismo que se encuentra a 10 km del Lago de Texcoco, b) agua de lluvia

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
205
recolectada de la zona del LT, antes de hacer contacto con el suelo. En el caso del Lago de
Texcoco se hizo una estratificación de los cuerpos de agua presentes: c) Muestra 1: Agua del
lago de Texcoco (Coordenadas: 19.4553-98.9867) representativa de los charcos formados en
epoca de lluvias; con 15 cm de profundidad, d) Muestra 2: Agua del Lago de Texcoco
(Coordenadas: 19.45.28-98.9765) representativa de la orilla (20 cm de profundidad) de los
lagos presentes; y e) Muestra 3: Agua del lago de Texcoco (Coordenadas: 19.4612-98.9886)
representativa del vaso de regulación que corre a 4 metros del lago Nabor carrillo, 27 de
profundidad. Se realizó un análisis a cada muestra de agua.
Tiempo de desarrollo. Se vertieron 3 litros de agua potable en un recipiente de plástico
de 30 x 20 x 30 cm, al cual se le adicionaron papeletas con huevos de la raza New Orleans,
se introdujo a la cámara de cría y 4 horas después se extrajeron las larvas para colocarlas
individualmente en tubos falcon de 50 mL PD1001B (crm globe, USA). Se utilizó huevo de
48 horas de edad (CENAPRECE, 2015). Con una cohorte de 400 individuos en estado L1 se
depositaron en cada tratamiento con una densidad de 1 larva cm-3 (40 larvas por tubo/10
tubos*tratamiento) y se agregó a diario en cada tubo, 12 mg (0.3 mg/individuo) de alimento
molido para Hamster “RedKite 450 G” (Grupo Lomas, Ciudad de México, México).
Las observaciones se realizaron diariamente hasta que éstas llegaron al estado de pupa.
Posteriormente, las pupas retiraron con la ayuda de una pipeta de transferencia. En cada
evaluación se determinaron el número de larvas vivas y el instar. El medio acuoso donde se
desarrollaron las larvas se cambió cada dos dias (Quispe-Pretel et. al. 2015) para evitar
formacion de hongos por el alimento.
Se prepararon tubos de microtitulación (tapa perforada) con 1 mL de agua de cada
tratamiento correspondiente. Cuando se observó en los tubos falcon la presencia de pupas,
estas se transfirieron a tubos de microtitulación 7-72077 (Cientifica Senna, CDMX, Mexico)
para la emergencia de adultos e iniciar el sexado y así obtener machos y hembras virgenes,
mismos que se utilizaron en las evaluaciones de fecundidad y fertilidad.
Una vez emergidos los adultos vírgenes, se determinó el dimorfismo sexual mediante las
antenas (Ebeling, 1975), y se colocaron 10 parejas en jaulas entomológicas (30x30x30 cm).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
206
Para su alimentación se colocó algodón embebido en agua azucarada al 1%. Como fuente de
proteína para las hembras se colocó diariamente un recipiente de polipropileno VP4CH
(REYMA, Leon GTO. MEXICO.) con 10 mL de sangre fresca de cerdo (Sus scrofa spp.) al
cual se le adicionó el anticoagulante INHEPAR 1000 UI/mL (PISA Farmaceutica,
Guadalajara Jalisco, MEXICO.) a la dosis de 7 mL/L sangre, y se puso en baño María a 37°
C durante 5 min. Posteriormente dicho recipiente se tapó con una de membrana Parafilm®
M PM-992 (Bemis, USA.). Para hacer este alimento más apetecible a las hembras, se
impregnó de sudor humano y se colocó invertido en la parte superior de la jaula
entomológica.
Para determinar el tiempo de desarrollo se hicieron, para todos los estados biológicos,
observaciones diarias. Los tiempos totales de desarrollo de huevo a adulto de Ae. aegypti, se
compararon utilizando una prueba No pareada (a= 0.05%) de t de student, y ANOVA (p ≤
0.05%). La longevidad de adultos se comparó (entre tratamientos) con una prueba de
ANOVA (p ≤ 0.05%). Los análisis se desarrollaron con el software Statistix 8.1.
RESULTADOS Y DISCUSIÓN
Los rangos de pH del lago de Texcoco son altos (8.7-9.7) y el sodio se encuentra presente
desde 15 hasta 126 mg/L. Las tres muestras del lago presentan altos valores de aniones (Cl=
1.09-172, SO42= 166-266, F= 0.06-0.38), cationes mg/L (Ca= 1.4-2.2, Mg= 0.9-1.6, K= 3-
14), y alta conductividad electrica (2.85-17.82 dSm-1), debido al alto contenido de sales
disueltas. Los tratamientos con agua potable y agua de lluvia se utilizaron testigos.
La prueba de eclosión indica que los huevos viables reaccionan al estimulo del agua en
los diferentes tratamientos, a solo horas después de su contacto. Los huevos presentes en
agua de la llave comienzan a eclosionar más rápido (0.23 días ± 0.02 a); aquellos expuestos
al agua de lluvia tardaron más tiempo a eclosionar (0.72 días± 0.05 a). Los huevos expuestos
a los diferentes tratamientos de agua provenientes del Lago de Texcoco, eclosionaron a los
0.35-0.42 días.
El tiempo total de desarrollo de A. aegypti en todos los tratamientos fue mayor a 11 días.
El tiempo mas largo de desarrollo ocurrió en los individuos cuyas larvas estuvieron expuestas

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
207
al agua de la llave (11.72 d), y el tiempo más breve ocurrió cuando las larvas estuvieron
expuestas al agua del Lago de Texcoco (muestra 3; en la muestra 3; 11.0 d). El porcentaje de
mortalidad de larvas expuestas en el agua del lago de Texcoco fueron los siguientes: muestra
1 (47%), muestra 2 (61%) y muestra 3 (57%). En el estado de pupa también hay mortalidad
(< 12%); en este caso la mortalidad mas elevada se observó en larvas presentes en el agua de
lluvia con 12% y menor mortalidad en el agua de la llave y la muestra 3 del Lago de Texcoco
con valor de 2%.
La curva de sobrevivencia muestra que los adultos de A. aegypti cuya larva estuvo
expuesta al agua potable, fueron más longevos; casi el 50% de los individuos vivieron 30 d,
y los menos longevos fueron aquellos casos donde la respectiva larva estuvo en contacto con
agua del Lago de Texcoco (muestras 2 y 3). En todos los tratamientos, los primeros adultos
que emergieron fueron las hembras y también éstas fueron las más longevas. Las hembras
provenientes de larvas que se desarrollaron en agua potable llegaron a vivir hasta 55.7 días.
Las hembras que murieron primero fueron las de la muestra 3 del Lago de Texcoco. En todos
los tratamientos, los machos comienzan a morir a partir del día 31, y no llegan con vida al
día 40.
Los diferentes tipos de agua presentes en la zona lacustre del Lago de Texcoco no
representan obstáculo para la eclosión de huevo, desarrollo tanto de larvas como de pupas de
A. aegypti. Por tanto, se considera que, si la temperatura de esta zona sube a un nivel
favorable para esta especie, la invasión ocurrirá y amenazará a un núcleo poblacional de más
de 36 millones de personas.
AGRADECIMIENTOS
Agradezco al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la
beca otorgada para realizar los estudios de posgrado del primer autor. Al Colegio de
Postgraduados, Campus Montecillo, por el equipo y las instalaciones donde se realizaron los
experimentos y la asesoría de sus Profesores.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
208
LITERATURA CITADA
CENAPRECE Centro Nacional de Programas Preventivos y Control de Enfermedades. 2015. GUÍA PARA LA INSTALACIÓN Y MANTENIMIENTO DEL INSECTARIO DE Aedes aegypti (DIPTERA: CULICIDAE). http://www.cenaprece.salud.gob.mx/programas/interior/vectores/descargas/pdf/GuiaInstalacionMantenimientoInsectarioAedesAegypti.pdf. Fecha de consulta 18 de agosto 2019
CONAGUA, 2014. PROYECTO HIDRÁULICO DEL LAGO DE TEXCOCO. MEXICO. Ebeling, W. 1975. Urban Entomology. University of California. Division of Agricultural
Sciences. Guzmán-Quintero, A., Palacios-Vélez, O. L., Chávez-Morales, J., Nikolskii-Gavrilov, L.
2007. LA CONTAMINACIÓN DEL AGUA SUPERFICIAL EN LA CUENCA DEL RÍO TEXCOCO, MÉXICO. Agrociencia 41 (4): 385-393
Kuri-Morales, P., Correa-Morales, F., González-Acosta, C., Sánchez-Tejeda, G., Dávalos-Becerril, E., Fernanda Juárez-Franco, M., Díaz-Quiñonez, A., Huerta-Jimenéz, H., Mejía-Guevara, M.D., Moreno-García, M., González-Roldán, J.F., 2017. First report of Stegomyia aegypti (= Aedes aegypti) in Mexico City, Mexico. Med. Vet. Entomol. 31: 240–242.
Márquez, B. Y., Monroy, C. K. J., Martínez, M. E. G., Peña G. V. H., Monroy, D. A. L. 2019. Infuencia de la temperatura ambiental en el mosquito Aedes spp y la transmisión del virus del dengue. Rev CES Med; 33(1): 42-50.
McNaughton, D., Miller, E. R., & Tsourtos, G. (2018). The Importance of Water Typologies in Lay Entomologies of Aedes aegypti Habitat, Breeding and Dengue Risk: A Study from Northern Australia. Tropical medicine and infectious disease, 3(2), 67. doi:10.3390/tropicalmed3020067
Rigau-Pérez, J.G., Clark, G.G., Gubler, D.J., Reiter, P., Sanders, E.J., Vorndam, A. V, 1998. Dengue and dengue haemorrhagic fever. Lancet 352: 971–7.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
209
PROPENSIÓN A RESISTENCIA DE Aedes aegypti (DIPTERA:
CULICIDAE) AL INSECTICIDAS BOTÁNICO GREEN CONTROL
ULV
Filemón Morales Hernández1, J. Concepción Rodríguez Maciel1*, Ángel Lagunes Tejeda1, Celina
Llanderal Cázares1, Gustavo Ramírez Valverde2, Gonzalo Iván Silva Aguayo3.
1 Posgrado en Fitosanidad-Entomología y Acarología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, CP
56230, Texcoco, Estado de México, México. 2Socioeconomía, Estadística e Informática, Colegio de Postgraduados, Carretera México-
Texcoco km 36.5, Montecillo, CP 56230, Texcoco, Estado de México, México. 3Departamento de Producción Vegetal, Facultad de
Agronomía, Universidad de Concepción, Chillán, Chile. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
El mosquito Aedes aegypti L. se considera la especie de insecto más importante como vector
de enfermedades al ser humano, ya que transmite patógenos arbovirales que causan
enfermedades como zika, dengue, dengue hemorrágico, virus del Mayaro, fiebre amarilla y
chikungunya (Leta et al., 2018). En México, desde 1950 se han utilizado insecticidas para
combatir mosquitos vectores (García et al., 2009). Entre 1981 y 1999, el insecticida
organofosforado malatión se utilizó ampliamente en aplicaciones espaciales como adulticida
contra A. aegypti y actualmente el organofosforado temefos se sigue utilizando como
larvicida; entre 2000 y 2011, los programas de control de vectores en México se basaron en
el uso del insecticida permetrina (García et al., 2009). Este uso desmedido de insecticidas ha
llevado al desarrollo acelerado de la resistencia a insecticidas en A. aegypti.
El Centro Nacional de Programas Preventivos en México y Control de Enfermedades
(CENAPRECE) (2018) documenta resistencia generalizada a piretroides en México, por lo
que este fenómeno representa una amenaza al combate de dicha especie. Actualmente, los
insecticidas botánicos han estado remplazando gradualmente a los insecticidas sintéticos
tradicionales contra una amplia variedad de plagas. Este tipo de insecticidas generalmente se
desarrollan a partir de metabolitos secundarios de plantas y tienen toxicidad en insectos a
través de múltiples mecanismos de acción, por lo que su propensión a desarrollar resistencia
en la mayoría de los casos es baja (Cui et al., 2016; Pavela and Benelli, 2016). En México,
en el año 2017, por primera vez la Secretaría de Salud a través del CENAPRECE (2017)

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
210
incluyó dentro de los insecticidas recomendados para el combate de A. aegypti al insecticida
comercial GREEN CONTROL ULV, formulado a base de extractos de piretrinas I y II
(Chrysanthemum cinerariaefolium (Trevir.) Vis.), aceite extracto de canela (Cinnamomum
zeylanicum Blume) y aceite extracto de neem (Azadirachta indica A. Juss). No se tienen
antecedentes del grado de propensión a resistencia a este producto y por ende, se desconoce
el grado de restricción que debe tener su uso en campo, para aumentar su vida útil. Por tanto,
el objetivo de esta investigación fue determinar la propensión del mosquito Ae. aegypti a
desarrollar resistencia al insecticida GREEN CONTROL ULV.
MATERIALES Y MÉTODOS
Ubicación de los experimentos. Esta investigación se llevó acabo de junio de 2017 a
agosto de 2019 en el laboratorio de Toxicología del Postgrado de Entomología y Acarología
del Colegio de Posgraduados, Campus Montecillo, Texcoco, Estado de México, México.
Población de Aedes aegypti. Se recolectó huevo de siete poblaciones de campo de
acuerdo a la metodología del CENAPRECE (2015). Dichas poblaciones son originarias de
La Paz, Baja california Sur; Culiacán, Sinaloa; Guadalajara, Jalisco; Ciudad Valles, San Luis
Potosí; Tejupilco, Estado de México; Cárdenas, Tabasco y Campeche, Campeche. Se
tomaron al azar de 300 a 400 larvas de tercer instar de cada población de campo y se
mezclaron para formar una población compuesta (Compuesta S0), la cual se crió por tres
generaciones sin presión de selección para favorecer el intercambio y la recombinación
genética. La crianza de las poblaciones de A. aegypti se llevó a cabo de acuerdo a la
metodología establecida por la Organización Mundial de la Salud (World Health
Organization, 2005).
Insecticida. Se utilizó el insecticida GREEN CONTROL ULV® (Extracto de piretrinas
naturales I y II al 17.5 %, aceite extracto de canela 9.35 % y aceite extracto de neem 9.35 %,
solución oleosa, Formulab de México, S. de R.L. de C.V., México) y las concentraciones
utilizadas fueron preparadas con acetona de grado técnico.
Procedimiento de selección. El procedimiento de selección de la población Compuesta
S0 consistió en la exposición durante 15 minutos, de mosquitos adultos en botellas Wheaton

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
211
de 250 ml impregnadas con el insecticida GREEN CONTROL ULV®. Se expusieron por
botella de 150 a 200 adultos, sin diferenciar sexo, recién emergidos y sin alimentarse. En
total, por generación se seleccionaron entre 4,000 y 5,000 adultos durante 13 generaciones.
Las botellas Wheaton se impregnaron con insecticida a la concentración de 0.4 mg de
ingrediente activo/botella para las primeras cuatro generaciones, 0.8 mg/L de la generación
S5 a la S8 y 1 mg/L de la generación S9 a la S13, de acuerdo a la metodología establecida
por los Centers for Disease Control and Prevention (CDC) (CDC, 2012). Después de la
selección con insecticida, los mosquitos supervivientes se introdujeron en una jaula
entomológica (30 x 30 x 30 cm) y se alimentaron con una solución acuosa azucarada al 10
%.
Procedimiento de bioensayo. Se monitoreó la respuesta de las poblaciones en mosquitos
adultos mediante bioensayos de acuerdo al procedimiento estandarizado por los CDC (2012).
Después de 24 horas de recuperación posteriores a la exposición al insecticida, se registró el
porcentaje de mortalidad y se consideraron adultos muertos aquellos que no presentaron
movimientos.
RESULTADOS Y DISCUSIÓN
Se observó una reducción en la susceptibilidad de acuerdo a las estimaciones de las
Concentraciones Letales a nivel del 50% y 95% de mortalidad (CL50 y CL95) (Cuadro 2).
Aunque existen diferencias significativas en la respuesta de la población Compuesta antes y
después de la presión de selección, se considera que estos cambios no son significativos
debido a que los valores de RR son bajos para considerar que se desarrolló resistencia, de
acuerdo a los criterios de Georghiou, (1990) y Naqqash et al., (2016).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
212
Cuadro 2. Respuesta de hembras adultas de Aedes aegypti L. a presión de selección con el insecticida botánico GREEN CONTROL ULV.
G. † n¶ gl§ b±EE Þ
CL50¤ IC†† 95%
mg/botella
CL95 ¤IC†† 95%
mg/botella Pr >
χ²¶¶ RR50
§§ RR95§§
S0 600 4 3.9±0.37 0.051 0.045 - 0.060
0.13 0.10 - 0.19 0.095
S2 700 5 3.3±0.22 0.095 0.087 - 0.103
0.29 0.25 - 0.34 0.3 1.9 2.2
S5 600 4 5.0±0.33 0.122 0.115 - 0.129
0.25 0.23 - 0.29 0.12 2.4 1.9
S8 800 6 6.6±0.4 0.31 0.3 - 0.33
0.55 0.52 -0.61 0.57 6.1 4.2
S10 875 5 5.9±0.33 0.2 0.19 - 0.21
0.38 0.35 - 0.41 0.26 3.9 2.9
S12 875 5 4.9±0.28 0.27 0.26 - 0.28
0.59 0.54 - 0.67 0.33 5.3 4.5
S13 1125 7 3.84±0.2 0.159 0.151 - 0.168
0.42 0.38 - 0.47 0.84 3.1 3.2
†Generación de selección; ¶Total de larvas tratadas; §Grados de libertad; Þ Pendiente ± error estándar; ¤Concentración estimada que provoca 50% (95 %) de mortalidad; ††Intervalo de confianza al 95%; ¶¶Probabilidad mayor a χ²= prueba de ajuste a una línea recta; §§Respuesta relativa = CL50(95) población de campo/ CL50(95) población susceptible.
El insecticida GREEN CONTROL ULV está formulado a base de piretrinas, aceite de
neem (mezcla de aleloquímicos como la azadiractina, nimbina y salaniana), así como aceite
extracto de canela que tiene como componente principal el cinamaldehido. Estos
componentes presentan elevada actividad insecticida y varios modos de acción, lo que
confiere un bajo riesgo de desarrollar resistencia en poblaciones de insectos (Con et al., 2009;
Dua et al., 2009; Georghiou, 1990; IRAC, 2012; Siegwart et al., 2015).
AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) y al Colegio de
Postgraduados por el financiamiento y apoyo brindado a esta investigación.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
213
LITERATURA CITADA
CENAPRECE, 2018. Resultados Preliminares Estudios de Adulticidas. Centro Nacional de Programas Preventivos en México. Secretaría de Salud. México. https://www.gob.mx/cms/uploads/attachment/file/466748/Resultados_Preliminares_Adulticidas_2018_Publicacio_n.pdf (Consultado: 11/10/2019)
CENAPRECE. 2017. Productos recomendados por el CENAPRECE para el combate de insectos vectores de enfermedades a partir de 2017. Centro Nacional de Programas Preventivos en México. Secretaría de Salud. México. https://www.gob.mx/cms/uploads/attachment/file/236439/Lista_actualizada_de_productos_recomendados_por_Cenaprece2017.pdf (Consultado: 11/10/2019)
CENAPRECE. 2015. Guía Metodológica para la Vigilancia Entomológica con Ovitrampas. Centro Nacional de Programas Preventivos en México. Secretaría de Salud. México. https://www.gob.mx/cms/uploads/attachment/file/37865/guia_vigilancia_entomologica_ovitrampas.pdf (Consultado: 11/10/2019)
Porras, Gina, Calvo M., Esquivel M., Sibaja M., Madrigal-carballo Sergio. 2009. Quitosano n-acilado con cinamaldehído: un potencial bioplaguicida contra agentes patógenos en el campo agrícola. Revista Iberoamericana de Polímeros. 10(3), 197–206.
Cui, S., Wang L., Ma L., Geng X. 2016. P450-mediated detoxification of botanicals in insects. Phytoparasitica. 44, 585–599. https://doi.org/10.1007/s12600-016-0550-1
Dua, V.K., Pandey A.C., Raghavendra K., Gupta A., Sharma T., Dash A.P. 2009. Larvicidal activity of neem oil (Azadirachta indica) formulation against mosquitoes. Malar. J. 8, 2–7. https://doi.org/10.1186/1475-2875-8-124
García, G.P., Flores A.E., Fernández-Salas I., Saavedra-Rodríguez K., Reyes-Solis G., Lozano-Fuentes S., Bond J.G., Casas-Martínez M., Ramsey J.M., García-Rejón J., Domínguez-Galera M., Ranson H., Hemingway J., Eisen L., Black IV W.C. 2009. Recent rapid rise of a permethrin knock down resistance allele in Aedes aegypti in México. PLoS Negl. Trop. Dis. 3. https://doi.org/10.1371/journal.pntd.0000531
Georghiou, G. P.. 1990. Overview of Insecticide Resistance. Managing Resistance to Agrochemicals. p 18–41. https://doi.org/10.1021/bk-1990-0421.ch002
IRAC, 2012. IRAC Mode of Action Classification Scheme. Insecticide Resistance Action Commettee. https://www.irac-online.org/modes-of-action/ (Consultado: 11/10/2019)
Leta, S., Beyene T.J., De Clercq E.M., Amenu K., Kraemer M.U.G., Revie C.W. 2018. Global risk mapping for major diseases transmitted by Aedes aegypti and Aedes albopictus. Int. J. Infect. Dis. 67, 25–35. https://doi.org/10.1016/j.ijid.2017.11.026
Naqqash, M.N., Gökçe A., Bakhsh A., Salim M. 2016. Insecticide resistance and its molecular basis in urban insect pests. Parasitol. Res. 115, 1363–1373. https://doi.org/10.1007/s00436-015-4898-9
Pavela, R., Benelli G., 2016. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends Plant Sci. 21, 1000–1007. https://doi.org/10.1016/j.tplants.2016.10.005
CDC. 2012. Guideline for Evaluating Insecticide Resistance in Vectors Using the CDC Bottle Bioassay. Centers for Disease Control and Prevention.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
214
https://www.cdc.gov/malaria/resources/ pdf/fsp/ir_manual/ir_cdc_bioassay_en.pdf (Consultado: 11/10/2019)
Robertson, J.L., Jones M.M., Olguin E., Alberts B. 2017. Bioassays with Arthropods, Third Edit. ed. CRC Press, Boca Raton, FL.
Siegwart, M., Graillot B., Lopez C.B., Besse S., Bardin M., Nicot P.C., Lopez-Ferber M. 2015. Resistance to bio-insecticides or how to enhance their sustainability: A review. Front. Plant Sci. 6, 1–19. https://doi.org/10.3389/fpls.2015.00381
World Health Organization, 2005. Guidelines for laboratory and field testing of mosquito larvicides. https://apps.who.int/iris/bitstream/handle/10665/69101/WHO_CDS_WHOPES _GCDPP_2005.13.pdf?sequence=1&isAllowed=y (Consultado: 11/10/2019).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
215
HONGOS ASOCIADOS AL ESCARABAJO AMBROSIAL
Xylosandrus curtulus (Eichhoff) (COLEOPTERA: SCOLYTIDAE) EN
ZIRACUARETIRO, MICHOACÁN, MÉXICO
Francisca De la Cruz-Martínez1, Jesús Romero Nápoles1*, Armando Equihua Martínez1, José Abel
López Buenfil2, Santos Gerardo Leyva-Mir3
1 Posgrado en Fitosanidad-Entomología y Acarología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230
Texcoco, Estado de México, México. 2 Centro Nacional de Referencia Fitosanitaria, km 37.5 Carretera Federal México-Pachuca, Tecámac,
Estado de México, 55740. 3 Universidad Autónoma Chapingo, Departamento de Parasitología Agrícola, km 38.5 Carretera México-
Texcoco, Chapingo, Estado de México, 56230. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
La relación simbiótica entre escarabajos descortezadores y sus hongos asociados ha sido
ampliamente estudiada por la importancia mundial en áreas forestales, sin embargo,
recientemente ha despertado un interés muy especial los escarabajos ambrosiales y sus
hongos ambrosiales, enfocados principalmente a dos géneros, Xyleborus spp. y Euwallacea
spp. En ese sentido, en la Unión Europea se realizó el estado del arte de los escarabajos
ambrosiales del género Xylosandrus, por considerarlos un riesgo en viveros forestales y en
el comercio de plantas, de 2014 a 2017, se incluyeron sitios de inspección en España, Francia
e Italia (IUFRO, 2018); asimismo, en Estados Unidos las investigaciones están enfocadas a
tópicos de evolución ecológica, patogenicidad y manejo, incluyendo a Xylosandrus
amputatus (Blandorf, 1894), X. compactus (Eichhoff, 1876), X. crassiusculus (Motschulsky,
1866), X. discolor (Blandford, 1898) y X. germanus (Blandorf, 1894).
En México, el escarabajo ambrosial Xylosandrus curtulus (Eichhoff) se encuentra
distribuido en Campeche, Chiapas, Colima, Hidalgo, Jalisco, Morelos, Nayarit, Oaxaca, San
Luis Potosí, Tabasco, Tamaulipas, Veracruz y Yucatán (Atkinson, 2012); posteriormente
Lázaro (2019) registra a Xylosandrus curtulus, como primer registro para Michoacán
asociado al cultivo de aguacate.
Estos escarabajos han desarrollado una estructura especializada similar a un saco
(micangio) para la transmisión del simbionte fúngico y la dispersión en árboles hospedantes

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
216
(Batra, 1963), también pueden introducir hongos fitopatógenos en ambientes naturales y
áreas cultivadas, por ejemplo, Raffaelea lauricola asociada a Xyleborus glabratus
(O’Donnell et al., 2015), ambos amenazando el cultivo de aguacate en la costa este de
Estados Unidos de América. Algunos hongos asociados a Xylosandrus spp. son patogénicos,
por ende, si se establecieran en áreas naturales, los daños podrían ser catastróficos (Vannini
et al., 2018).
Los objetivos planteados en este trabajo se indican a continuación: 1) aislar los hongos
asociados al escarabajo ambrosial Xylosandrus curtulus Eichhoff, proveniente de un huerto
comercial de aguacate en Ziracuaretiro, Michoacán, México y 2) tipificar el micangio
mesonotal en X. curtulus.
MATERIALES Y MÉTODOS
El estudio se realizó en escarabajos ambrosiales colectados en el huerto comercial de
aguacate Hass “La Ziranda”, en Ziracuaretiro, Michoacán (1,304 msnm, 19.2400º N,
101.5456º W) del 2 al 8 de junio de 2019. Para la captura se utilizaron ocho trampas con
etanol como atrayente y agua destilada como medio de colecta. Se utilizaron 40 individuos a
las 72 horas de haberlos colectado, para esto cada individuo fue seccionado en dos partes, la
primera considerando cabeza y protórax con el primer par de patas y la segunda porción
estuvo integrada por mesotórax, metatórax, abdomen y patas correspondientes. Dos insectos
(cuatro porciones) se colocaron por caja Petri, en total se prepararon 20 cajas en Cámara
Húmeda y 20 en medio Agar Spezieller Nährstoffarmer (SNA). Aproximadamente al quinto
día de sembrado se aislaron y purificaron hongos y bacterias, utilizando punta de hifa,
cultivos monospóricos, así como Sacabocados® de 0.5 mm, en diversos medios de cultivo,
tales como Papa Dextrosa Agar (PDA), PDA con antibiótico y Papa Dextrosa Agar
BDBioxon®. Las cepas se mantuvieron bajo condiciones de luz natural y temperaturas que
oscilan entre 25 a 27 grados. Todo el procedimiento se llevó acabo en el laboratorio de
Micología del Depto. de Parasitología Agrícola de la Universidad Autónoma Chapingo.
Una vez que se obtuvieron las cepas de hongos y bacteria purificadas, se ingresaron al
Laboratorio de Diagnóstico Integral Fitosanitario del Colegio de Postgraduados (LADIFIT-

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
217
COLPOS) para extracción de ADN, amplificación, secuenciación e identificación
correspondiente.
Con respecto a la tipificación del micangio, se tomaron ocho especímenes almacenados
bajo condiciones de -5 ºC, se descongelaron uno a uno, durante 5 minutos y mediante
disección manual, con pinzas entomológicas no. 5 y pincel no. 000, utilizando agua o
glicerina como medio de disección, se eliminaron las partes del cuerpo, quedando sólo la
parte dorsal del mesotórax, éste se colocó en portaobjetos con glicerina en posición ventral
para su observación en microscopio compuesto.
Para la identificación de la especie de Scolytidae se utilizaron las claves de Wood (1982),
además también se contó con una identificación molecular para lo cual se siguió el protocolo
de diagnóstico fitosanitario denominado “Amplificación de la región mtDNA CO-I, para la
identificación de insectos por PCR punto final LC01490/HC02198-SENASICA” (2017), en
este procedimiento se incluye a detalle la extracción, amplificación y secuenciación de DNA.
En lo que respecta al análisis de datos, se aplicó el siguiente Modelo Lineal Generalizado con objeto de identificar diferencias significativas entre medios de cultivo, regiones e interacción entre éstos:
Yijk = µ + Mi + Rj + M*Rij + eijk
Donde,
Yijk = Número de patógenos en el i – ésimo medio y el j – ésima región de la k – ésima repetición; donde i = 1, 2; j= 1, 2; k = 1, …, 20.
µ = La media general de patógenos.
Mi = El medio de cultivo, donde i = 1 cámara húmeda; i = 2 SNA.
Rj = La región del cuerpo del insecto, donde j = 1 abdomen; j = 2 cabeza
eijk = Error experimental asociado al i – ésimo medio y j – ésima región de la k – ésima repetición.
RESULTADOS Y DISCUSIÓN
Referente a la colecta del escolítido en las trampas repartidas en el huerto, el cual contaba
con una superficie de 3.5 ha, se colectó un total de 76 especímenes.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
218
Con respecto a la identidad de la especie, mediante las claves y por comparación con
material determinado en la Colección de Insectos del Colegio de Postgraduados (CEAM),
ésta se identificó como Xylosandrus curtulus (Eichhoff, 1869). Referente a la identificación
molecular, se llevó a cabo la limpieza y ensamble de las secuencias asimismo la búsqueda
por homología y los análisis filogenéticos están en proceso.
Se purificaron siete cepas de hongos y mediante preparaciones temporales con cinta
adhesiva se identificaron cuatro géneros: Colletotrichum spp., Cladosporium spp.,
Trichoderma spp. y Penicillium spp.; aún estamos en espera de los resultados específicos de
la identidad de cada aislamiento para realizar la discusión correspondiente. En el mismo
estatus se encuentra la cepa bacteriana.
Se encuentra en proceso la aplicación del Modelo Lineal Generalizado para determinar si
existen diferencias significativas de los medios de cultivo, las regiones del insecto y su
interacción.
Referente al micangio, se corrobora su ubicación en el mesotórax y consiste de dos sacos
oblongos, hialinos y membranosos, con medidas de 0.19 mm de largo por 0.19 de ancho,
embebidos en la cavidad del mesotórax y adheridos a éste por un extremo.
Francisca De la Cruz-Martínez,
2019

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
219
AGRADECIMIENTOS
Agradecemos al Consejo Nacional de Ciencia y Tecnología (CONACYT, México) por la beca otorgada en la maestría para el primer autor; al Colegio de Postgraduados, Campus Montecillo y al Departamento de Parasitología Agrícola de la Universidad Autónoma Chapingo por el equipo, materiales y las instalaciones donde se realizaron los experimentos; a los integrantes del Comité y al Dr. Santos Gerardo Leyva Mir por su apoyo, asesoría y enseñanzas en el desarrollo de la investigación; al Sr. Israel Solís Velázquez, propietario del huerto “La Ziranda” por las facilidades brindadas durante las actividades de colecta. Finalmente, a todos los amigos y colegas que me apoyaron incondicionalmente para concluir con éxito esta investigación.
LITERATURA CITADA
Atkinson, T. H., 2012. Estado de conocimiento de la taxonomía de los escarabajos descortezadores y ambrosiales de México (Coleoptera: Curculionidae: Scolytinae). In: Memoria XVI Simposio Parasitología Forestal. Consejo Nacional Forestal. 13-27.
Batra, L. R. 1963. Ecology of ambrosia fungi and their dissemination by beetles. Transactions of the Kansas Academy of Science (1903), 66(2), 213-236.
Doyle, J. J. and Doyle, J. L. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin, 19, 11-15.
Lázaro-Dzul, M.O. 2019. Monitoreo de Scolytinae (COLEOPTERA: CURCULIONIDAE) con atrayentes químicos en huertos de aguacate en Michoacán. Montecillo, Estado de México. 28-31p.
O’Donnell, K., S. Sink, R. Libeskind-Hadas, R. C. Ploetz, Konkol, J. N. Ploetz, D. Carrillo, A. Campbell, R.E. Duncan, M.T. Kasson, P. N. H. Liyanage, A. Eskalen, D. M. Geiser, J. Hulcr, J., C. Bateman, S. Freeman, Z. Mendel, P. R. Campbell, A. D. W. Geering, T. Aoki, A. A. Cossé and A. P. Rooney, (2015) Cophylogenetic analysis of the Fusarium–Euwallacea (COLEOPTERA: SCOLYTINAE) mutualism suggests their discordant phylogenies are due to repeated host shifts. Fungal Genetics and Biology, 82, 277–290.
SENASICA. 2017. Protocolo de diagnóstico fitosanitario “Amplificación de la región mtDNA CO-I, para la identificación de insectos por PCR punto final LC01490/HC02198-SENASICA”. Centro Nacional de Referencia Fitosanitaria, Dirección General de Sanidad Vegetal, SENASICA.
Vannini, A., S. Speranza, M. Contarini, I. Sferrazza, F. Michelangeli and Carmen Morales Rodriguez. 2018. State of the Art: The Ambrosia beetle Xylosandrus compactus and the symbiotic fungi: a new risk for European native flora associated to nurseries and trading of living plants. Diseases and insect forest nurseries working party meeting. International Union of Forest Research Organizations (IUFRO). Kuşadasi, Turquía. Wood, S. L. 1982. The Bark and Ambrosia Beetles of North and Central America
(Coleoptera: Scolytidae), a taxonomic monograph. Great Basin Nat. Mem. No. 6.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
220
ASOCIACIÓN DE Xylella fastidiosa CON DIFERENTES ESPECIES
DE PLANTAS CULTIVADAS Y SILVESTRES EN PARRAS DE LA
FUENTE, COAHUILA
Sandra Lourdes Moya-Hernández1, Ana María Hernández-Anguiano1*, Sergio Aranda-Ocampo1,
José Abel López-Buenfil2
1 Posgrado en Fitosanidad-Fitopatología, Colegio de Postgraduados, Carretera México-Texcoco km 36.5, Montecillo, 56230 Texcoco,
Estado de México, México. 2 SENASICA-SADER. Centro Nacional de Referencia Fitosanitaria. Carretera Federal México-Pachuca Km
37.5, Tecámac, CP. 55740, Estado de México. *Autor de correspondencia: [email protected]
INTRODUCCIÓN
El municipio de Parras de la Fuente Coahuila, México es una importante zona productora de
uva de mesa, con aproximadamente 450 hectáreas y una producción promedio de 2 352.1
toneladas valorada en cerca de 45 millones de pesos (SIAP, 2019). Esta producción se
encuentra amenazada por la enfermedad de Pierce, causada por Xylella fastidiosa, bacteria
que no solo ataca a la vid (Vitis vinifera L.) sino también a un gran número de especies de
plantas herbáceas (malezas o vegetación silvestre) de plantas arbustivas o leñosas, cultivadas
o adventicias, de uso agrícola u ornamental (Baldi y La Porta, 2017; EFSA, 2018; Marco-
Noales et al., 2017). Algunas de las especies vegetales no desarrollan síntomas de
enfermedad, pero todas son de importancia epidemiológica (Janse y Obradovic, 2010;
Welington et al., 2002). Por lo anterior, se planteó esta investigación con el objetivo de
determinar la asociación de X. fastidiosa con las especies de plantas cultivadas y silvestres
aledañas a los viñedos de esta importante zona productora de uva de mesa.
MATERIALES Y MÉTODOS
Recolecta. En los meses de junio, agosto, septiembre y octubre de 2018 se recolectó
material vegetal en los predios la Encantada y San Lorenzo del grupo comercial Casa
Madero; también, en agosto, se hizo una captura con trampa amarilla de insectos en la
vegetación silvestre aledaña a los viñedos. Las muestras recolectadas correspondieron a: 1)
plantas de vid con síntomas similares a la enfermedad de Pierce; 2) árboles frutales y
forestales y plantas ornamentales; 3) plantas silvestres o malezas (tres ejemplares de cada

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
221
una), con síntomas o asintomáticas (EFSA, 2018). Un ejemplar de cada planta silvestre fue
prensado en campo de acuerdo al Manual de recolecta, preservación y envío de ejemplares
de malezas (SENASICA, s. f.). Las muestras se trasladaron al Centro Nacional de Referencia
Fitosanitaria (CNRF) de la Dirección General de Sanidad Vegetal en Tecámac, Estado de
México para su análisis. La identificación taxonómica de las plantas silvestres y de los
insectos estuvo a cargo de personal técnico de los laboratorios de Malezas y Entomología y
Acaralogía del CNRF, respectivamente.
Detección. La detección de X. fastidiosa se llevó a cabo mediante las variantes punto final
y tiempo real de la Reacción en Cadena de la Polimerasa (PCR, por sus siglas en inglés). La
extracción de DNA de peciolos y nervadura central de hojas (0.5 g de tejido) y de las cabezas
de insectos se hizo por el método de CTAB 2% (Loconsole et al., 2014) con algunas
modificaciones. Se verificó la pureza y cantidad del DNA por espectrofotometría (Nanodrop
2000®).
La reacción PCR punto final se estableció con los pares de iniciadores específicos:
XYgyr499-F/XYgyr907-R, HL5/HL6 y RST31/RST33 los cuales amplifican los genes gyrB,
HL y rpoD, respectivamente (Rodrigues et al., 2003; Francis et al., 2006; Minsavage et al.,
1994). Los productos de PCR se enviaron a secuenciar al laboratorio de Biología Molecular
del CNRF (ABI PRISM 3130 de Applied Biosystems). Las secuencias sentido y antisentido
de cada gen se editaron con el programa BioEdit® v 7.0.9.0 (Hall, 1999) para obtener las
secuencias consenso las cuales se sometieron a comparación de similitud contra las
secuencias de X. fastidiosa depositadas en el GenBank NCBI (National Center for
Biotechnology Information), con el programa bioinformático BLASTn (Basic Local
Alignment Search Tool nucleotide) para comprobar la identidad de la bacteria. Mientras que
la reacción PCR tiempo real se estableció con los iniciadores XF-F/XF-R y la sonda XF-P
Taqman (Harper et al., 2010).
RESULTADOS Y DISCUSIÓN
De 83 muestras de especies de plantas analizadas, X. fastidiosa se detectó en cinco
muestras de vid, cuatro especies de árboles frutales (Carya sp., Ficus carica., Olea europea

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
222
y Cydonia sp.) y seis especies de plantas silvestres (Salsola kali., Celtis pallida, Phragmites
sp., un pasto de la familia Poaceae, Rubus sp. y Tribullus terrestris) (Figura 1). Las especies
de plantas positivas a X. fastidiosa pertenecen a las familias: Vitaceae, Juglandaceae,
Moraceae, Oleaceae, Rosaceae, Amaranthaceae, Cannabaceae, Poaceae y Zygophyllaceae.
Con excepción de la familia Zygophyllaceae, especies de estas familias se encuentran
reportadas en la lista de hospedantes de X. fastidiosa de la Autoridad Europea de Seguridad
Alimentaria (EFSA, 2018). En este estudio Tribullus terrestris (maleza) y Cydonia sp (árbol
frutal conocido como membrillo) de la familia Zygophyllaceae se proponen como nuevos
hospedantes o reservorios de X. fastidiosa en Parras de la Fuente, Coahuila.
Figura 1. Plantas silvestres positivas a X. fastidiosa. A y B) Tallos y hojas de Salsola kali
(rodadera). C y D) ramas y hojas de Celtis pallida (acebuche). E) hojas con escaldadura de
Phragmites sp. (carrizo gigante). F) frutos y flores de Rubus sp. (mora silvestre).
Actualmente se reportan 563 especies de plantas asociadas con X. fastidiosa (EFSA,
2018), la mayoría de ellas son asintomáticas y pueden actuar como reservorios contribuyendo
al establecimiento, mantenimiento y dispersión de la bacteria (Landa et al., 2017). En esta
investigación, las plantas silvestres que resultaron positivas a X. fastidiosa fueron
C
A
F E
D
B

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
223
asintomáticas. Presumiblemente estos reservorios encontrados en la vegetación adyacente a
los viñedos contribuyen a la trasmisión de la bacteria a las plantaciones de vid. En las plantas
de vid se registró escaldadura en los márgenes de las hojas, “islas verdes” y momificación de
frutos, síntomas característicos de la enfermedad de Pierce. De seis ejemplares de insectos
solo uno resultó positivo a X. fastidiosa el cual se identifica como Homalodisca sp. Las
plantas de vid que resultaron positivas, así como la trampa amarilla en la cual se capturó a
los ejemplares de insectos, se ubicaron en el borde de las plantaciones de vid, lo que coincide
con lo reportado respecto a que la vegetación natural riparia es refugio natural de insectos
vectores y que la mayor incidencia de la enfermedad se presenta en los bordes del viñedo
(Morente y Fereres, 2017; Purcell, 1974). La vegetación silvestre adyacente a los viñedos
suele albergar especies de plantas que son el alimento principal de insectos vectores los
cuales se trasladan a las plantaciones de vid en donde trasmiten a la bacteria causante de la
Enfermedad de Pierce. Es por eso que las plantas silvestres son importantes en la
epidemiología de la enfermedad de Pierce y en el mantenimiento de la diversidad genética
de X. fastidiosa (Sicard et al., 2018).
Los resultados obtenidos en este estudio indican que X. fastidiosa se encuentra asociada a
diversas especies de plantas silvestres (rodadera, acebuche, carrizo gigante y mora silvestre)
y cultivadas (vid, olivo, nogal y membrillo) en Parras de la Fuente, Coahuila.
AGRADECIMIENTOS
Al Servicio Nacional de Sanidad Inocuidad y Calidad Agroalimentaria, al Centro Nacional
de Referencia Fitosanitaria y al Laboratorio de Bacteriología, ambos del CNRF, por las
facilidades otorgadas a la primera autora para cursar sus estudios de posgrado y llevar a cabo
su trabajo de investigación. Al personal académico del Colegio de Postgraduados, Campus
Montecillo, por la asesoría y enseñanzas proporcionadas.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
224
LITERATURA CITADA
Baldi P, and La Porta N. 2017. Xylella fastidiosa: Host Range and Advance in Molecular Identification Techniques. Front. Plant Sci. 8:944. doi: 10.3389/fpls.2017.00944.
EFSA (European Food Safety Authority) 2018. Scientific report on the update of the Xylella spp. host plant database. EFSA Journal 2018;16(9):5408 87 pp. https://doi.org/10.2903/j.efsa. 2018.5408.
Francis MH, Lin J, Cabrera-La Rosa, Doddapaneni H and Civerolo EL. 2006. Genome-based PCR primers for specific and sensitive detection and quantification of Xylella fastidiosa. European Journal of Plant Pathology 115: 203-213. doi: 10.1007/s10658-006-9009-4.
Hall, T. A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 41:95-98.
Harper SJ, Ward LI and Clover GR. 2010. Development of LAMP and real-time PCR methods for the rapid detection of Xylella fastidiosa for quarantine and field applications. Phytopathology 100: 1282-1288. https://doi.org/10.1094/PHYTO-06-10-0168.
Janse JD and Obradovich A. 2010. Xylella fastidiosa: its biology, diagnosis, control and risks. Journal of Plant Pathology 92, 35-48. Disponible en línea: file:///C:/Users/Sandy/Downloads/2504-1747-1-PB.pdf
Landa B, Navas JA, Montes M. 2017. Enfermedades causadas por la bacteria Xylella fastidiosa Capítulo 2. Características generales de Xylella fastidiosa. Monografías 32. Cajamar caja Rural. pp:47-59. Disponible en línea: https://www.publicacionescajamar.es/pdf/series-tematicas/informes-coyuntura-monografias/enfermedades-causadas-por-la-bacteria.pdf
Loconsole G., Saponari M., Potere O., Palmisano F y Boscia D. Current protocols for the detection of Xylella fastidiosa in host plants and vectors. Workshop Manual. International symposium on the European outbreak of Xylella fastidiosa in olive. Gallipoli, Locorotondo, Italy. 21-24 october, 2014.
Marco-Noales E, Landa B, López MM. 2017. Enfermedades causadas por la bacteria Xylella fastidiosa. Capítulo 1. Monografías 32. Cajamar caja Rural. pp:19-45. Disponible en línea: https://www.publicacionescajamar.es/pdf/series-tematicas/informes-coyuntura-monografias/enfermedades-causadas-por-la-bacteria.pdf
Minsavage GV, Thompson CM, Hopkins DL, Leite RM and Stall RE. 1994. Development of a polymerase chain reaction protocol for detection of Xylella fastidiosa in plant tissue. Phytopathology 84: 446-461. Disponible en línea: https://www.apsnet.org/publications/phytopathology/backissues/Documents/1994Articles/Phyto84n05_456.PDF
Morente M and Fereres A. 2017.Enfermedades causadas por la bacteria Xylella fastidiosa. Capítulo 4 Vectores de Xylella fastidiosa. Monografías 32. Cajamar caja Rural. pp:73-93. Disponible en línea: https://www.publicacionescajamar.es/pdf/series-tematicas/informes-coyuntura-monografias/enfermedades-causadas-por-la-bacteria.pdf
Rodrigues ML, Silva-Stenico ME, Gomes J, Lopes JS and Tsai SM. 2003. Detection and Diversity Assessment of Xylella fastidiosa in Field-Collected Plant and Insect Samples by Using 16S rRNA and gyrB Sequences. Applied and environmental microbiology, July 2003, p. 4249–4255. doi: 10.1128/AEM.69.7.4249-4255.2003.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
225
SIAP. 2019. Servicio Nacional de Información Agroalimentaria y Pesquera. Anuario Estadístico de Producción Agrícola.
Sicard A, Zeilinger AR, Vanhove M, Schartel TE, Beal DJ, Daugherty MP and Almeida R. 2018. Xylella fastidiosa: Insights in to an Emerging Plant Pathogen. Annu. Rev. Phytopathol. 2018. 56:181–202. https://doi.org/10.1146/annurev-phyto-080417-045849.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
226
DIVERSIDAD Y FLUCTUACION POBLACIONAL DE LA
ACAROFAUNA ASOCIADA A ZARZAMORA (Rubus ulmifolius) Y
PRIMER REPORTE DE Diptacus sp.
Sandra Gonzalez-Domínguez1, Ma. Teresa Santillán-Galicia1*, Ariel Guzmán-Franco1, Clemente de
Jesús Avila-García2, José Abel López-Buenfil 2, Felipe Romero-Rosales1
1Postgrado en Fitosanidad-Entomología y Acarología. Colegio de Postgraduados, Km 36.5 Carretera México-Texcoco, Montecillo,
Texcoco, Edo. de Mexico, 56230. 2 Centro Nacional de Referencia Fitosanitaria. Km 37.5, Carretera Federal México-Pachuca, Tecámac,
Edo. de México, C.P. 55740. *Corresponding autor: [email protected]
RESUMEN
Se estudió la diversidad de especies y la fluctuación poblacional de ácaros en huertas de
producción orgánica y convencional de zarzamora var. Tupy, en Michoacán, México. El
periodo de muestreo se realizó en el ciclo productivo 2018. Los ácaros identificados fueron
Tetranychus urticae, Brevipalpus yothersi, Brevipalpus californicus, Diptacus sp.,
Tarsonemus sp. Polyphagotarsonemus latus, Brachytedeus mexicana, Tydeus kochi
Amblyseius andersoni. Se determinó la fluctuación poblacional de las familias con mayor
abundancia: Tetranychidae, Diptilomiopidae y Phytoseiidae, en las cuales se encontraron
diferencias estadísticas durante el periodo de estudio. La mayor diversidad de ácaros se
encontró en el sistema de producción orgánica, además, las etapas fenológicas con mayor
presencia fueron las “vegetativas” y en la “inducción a la producción”. Se realizó por primera
vez en México la descripción morfometría de una especie de vida libre de Diptilomiopidae,
Diptacus sp., en hojas de Rubus ulmifolius, la descripción se basa en hembras, machos y
estados inmaduros.
MATERIALES Y METODOS
Se realizó la toma de muestra de material vegetal de zarzamora (Rubus ulmifolius) var.
Tupi, en seis huertas de los municipios de los Reyes de Salgado y Peribán, Michoacán. Tres
huertas con manejo orgánico y tres convencional, en el ciclo productivo de 2018. En cada
huerta se tomaron cuarenta muestras compuestas (yemas axilares vegetativas, fructíferas,
hojas completamente desarrolladas, flores, frutos en desarrollo y completamente

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
227
desarrollados), en cada estrato de la planta y en cada una de las etapas fenológicas del cultivo.
Se eligieron 10 surcos distribuidos equitativamente a lo largo y ancho de la huerta, y en cada
uno de ellos se tomaron cuatro muestras compuestas.
Procesamiento de las muestras vegetales. Para la obtención de los ácaros, las estructuras
vegetales recolectadas en cada punto de muestreo de cada huerta se sumergieron en un
recipiente de 1L con una solución de agua jabonosa. La solución, junto con las estructuras
vegetativas se agitaron vigorosamente durante un minuto para desprender a los ácaros. En el
caso de las yemas y los frutos, estos se cortaron en partes pequeñas, antes de colocarlos en la
solución jabonosa. Posteriormente, el contenido se vertió en una torre de tamices de 200μ,
400 μ y 500 μ, colocados en orden descendente.
Diversidad de especies. Para determinar la diversidad de especies de ácaros en el cultivo
de zarzamora, todos los individuos recolectados en cada muestreo fueron montados entre
porta y cubreobjetos con liquido de Hoyer (Krantz y Walter, 2009) y medio de montaje Keifer
F modificado (Amrine y Manson, 1996). Una vez realizadas las preparaciones fueron
mantenidas a una temperatura de 50°C durante 15 días. Posteriormente se limpiaron y
sellaron con pintura anticorrosiva (Comex®). La identificación de las especies se realizó con
claves taxonómicas dicotómicas específicas para las familias y especies encontradas, usando
un microscopio de contraste de fases marca Leica.
Fluctuación poblacional. La fluctuación poblacional se determinó de los ácaros
recolectados en todas las huertas durante los muestreos realizados. Los datos de temperatura
y humedad fueron obtenidos de la estación meteorológica Los Reyes, 19° 35.93' N | 102°
29.44' W GlobalMet, 2016. Antes del análisis de datos, los números de ácaros se
transformaron utilizando Box Cox (λ=0.1), posteriormente se analizaron utilizando los
modelos lineales mixtos, que incluyeron un modelo de covariancia, efectos de tiempo (e
interacciones) y los componentes de varianza estimado utilizando la probabilidad máxima
restringida en el paquete de Software estadístico GenStat v.8.0 (Payne et al., 2005).

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
228
Descripción de Diptacus sp. Debido a la alta incidencia observada de Diptacus sp. en
todos los muestreos y que es una especie no reportada en zarzamora en México, se procedió
a realizar una descripción. Los especímenes fueron observados en un microscopio de
contraste de fases Olympus BX41 ®, con la adaptación de una cámara digital Olympus U-
TV1X-2®, vinculada al programa InfarView V. Las estructuras relevantes para la taxonomía,
se observaron con objetivos de 10 x, 40 x y 100 x. Se digitalizaron utilizando el programa
Image J V.4.3.0.
Para la observación y descripción de los empodios y la genitalia, se utilizaron imágenes
obtenidas de Microscopia Electrónica de Barrido, de la UME, del Colegio de Postgraduados,
Campus Montecillo.
RESULTADOS Y DISCUSIÓN
Diversidad de especies. Se determinó la asociación de Tetranychus urticae, Brevipalpus
yothersi, B. californicus, Diptacus sp., Tarsonemus sp., Polyphagotarsonemus latus, Tydeus
kochi, Brachytideus mexicana. Además, los depredadores Ambyseius andersoni, y la especie
micófaga Tyrophagus sp. La diversidad de los ácaros encontrados fue similar en los cultivos
con manejo convencional y orgánico; sin embargo, la proporción de individuos que se
encontraron en cada grupo fue mayor en el manejo orgánico. Los ácaros de las familias
Tarsonemidae y Tenuipalpidae, se presentaron en mayor proporción en el manejo
convencional. En cuanto a ácaros agentes de control biológico de la familia Phytoseiidae, se
presentaron en mayor proporción en huertas con manejo orgánico.
Fluctuación poblacional. En general, la distribución de ácaros fue diferente en cada etapa
fenológica del cultivo en los dos sistemas de producción orgánico y convencional. Las
especies con mayor abundancia fueron T. urticae y Diptacus sp. En el manejo convencional
Diptacus sp., tuvo mayor población en la etapa “vegetativa” y de “inducción a la
producción”, al igual que T. urticae. En el manejo orgánico, las mayores poblaciones se
presentaron en las etapas de “inducción a la producción”, “inducción a la floración” y en
“Flor y fruto”. El manejo agronómico determina la presencia o ausencia de follaje y otras
estructuras vegetales. T. urticae y Diptacus sp., fueron las especies con mayor abundancia.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
229
Con base en los resultados obtenidos, las etapas fenológicas en las que se encontró mayor
presencia de ácaros se asocian principalmente a temperaturas cálidas; el clima seco, la baja
humedad y las temperaturas superiores a los 30 °C favorecen el crecimiento de ácaros
(Dhooria 2016; Akyyazi et al., 2016). T. urticae fue la plaga más abundante en ambos
sistemas productivos, no obstante, las poblaciones cambiaron en las fechas de muestreo,
como en el cultivo de fresa, donde las poblaciones de ácaros fueron diferentes debido a las
diferencias de temperatura y disponibilidad de alimento (Nyoike y Liburd 2013); esto puede
deberse a su comportamiento y patrón de abundancia estacional ya que son distintos en cada
cultivo (Farazmand y Amir-Maafi, 2018). T. urticae, se presentó principalmente cuando las
temperaturas eran cálidas, por lo que los factores ambientales tienen un impacto significativo
en el comportamiento biológico (Fazlul et al., 2010). Las poblaciones de Brevipalpus
yothersi y B. californicus, tuvieron baja densidad, quienes se ven afectadas por factores
bióticos y abióticos (De Carvalho et al., 2008; Akyyazi et al., 2016), tales como la
precipitación sobre las principales estructuras vegetales, época del año y la planta hospedera
(Salinas-Vargas et al., 2013).
Descripción de Diptacus sp. Diptacus sp., fue la especie más abundante. Se han descrito
siete especies del género en Rubus sp. La especie encontrada en México comparte
características con D. rubuscolum sp. nov., reportada en Brasil, se distingue principalmente
por el tamaño diminuto de las sedas sc, la ornamentación del escudo prodorsal en forma de
red, con líneas medias incompletas, líneas admedias conectadas con líneas transversales,
formando una especie de celdas. Sin embargo, se puede llegar a confundir con D.
gigantoruba, D. caesius, D. rubi y D. glaber. La especie Diptacus sp., encontrada en México,
es más larga, que las anteriores. En comparación con D. rubuscolum sp.nov., Diptacus sp.
(México), las sedas 2ª, d, e y f son más cortas. El empodio dividido cuanta con 15 (2) radios,
no así D. rubuscolum que tiene 4 radios empodiales. Además, la placa genital con micro-
tubérculos desordenadas.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
230
LITERATURA CITADA
Akyasi R., Uerckermann EA., Soysal M., Akyol D. (2016). Population dynamics of mites (Acari) on Diospyros kaki Thunb. and Diospyros lotus L. (Ebenaceae) res in Ordu, Turkey. Systematic & Applied Acarology 21(10): 1334–1345.
De Carvalho MJL, Sato ME, Raga A, Arthur V. (2008). Population dynamics of phytophagous and predaceous mites on coffee in Brazil, with emphasis on Brevipalpus phoenicis (Acari: Tenuipalpidae). Exp Appl Acarol 44:277–29.
Dhooria MS. 2016. Management of mites under organic farming. In: Fundamentals of Applied Acarology, Springer, Singapore, 317-326.
Farazmand A., Amir-Maafi M. (2018). A population growth model of Tetranychus urticae Koch (Acari: Tetranychidae). Persian Journal of Acarology, 7(2): 193-201.
Fazlul HM., Khaleqzzaman M., Islam W. (2010). Population dynamics of Tetranychus urticae Koch and Phytoseiulus persimilis Athias-Henriot on three host plants. Pakistan Entomologist, 32(1): 6-11.
Nyoike TW., Liburd OE. (2013). Effect of Tetranychus urticae (Tetranychidae), on marketable yields of field-grown strawberries in North-Central Florida. Journal of Economic Entomology, 106(4):1757-1766.
Payne RW, Harding SA, Murray DA, Soutar DM, Baird DB, Welham SJ, Kane AF, Gilmour AR, Thompson R, Webster R, Tunnicliffe WG (2005) GenStat Release 8 reference manual. VSN International, Hemel Hempstead.
Salinas-Vargas D., Santillán-Galicia MT., Valdez-Carrasco J., Mora-Aguilera G., Abb tanacio-Serrano Y., Romero-Pescador P. (2013). Species composition and abundance of Brevipalpus spp., on different citrus species in mexican orchards. Neotrop Entomol, 42:419–425

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
231
SILICIO COMO POTENCIALIZADOR DE FUNGICIDAS CONTRA
Botrytis cinerea EN FRUTOS DE ZARZAMORA
José Terrones-Salgado1*, Daniel Nieto-Ángell, Cristian Nava-Diaz1, Daniel Téliz-Ortiz1, Prometeo
Sánchez García2, Rómulo García Velasco3, Moisés Roberto Vallejo Pérez4
1Programa de Fitosanidad-Fitopatología. 2Edafología. Colegio de Postgraduados, Campus Montecillo. Km 36.5 Carr. México-Texcoco,
México: 56230. 3UAEM. Tenancingo Edo. de México. 4CONACyt-UASLP. Álvaro Obregón No. 64, Col. Centro, San Luis Potosí, S.L.P.
C.P. 78000. *Correspondencia: [email protected].
INTRODUCCIÓN
La zarzamora es un cultivo frutícola económicamente importante que se cultiva en México y
en todo el mundo (Li et al., 2012). En 2018 en México se obtuvieron 258 141 t, el principal
productor es Michoacán (SAGARPA, 2018); la producción está amenazada por
enfermedades, donde el moho gris causado por Botrytis cinerea es la enfermedad más
importante (Li et al., 2012). En México se ha reportado en diferentes regiones productoras
de dicho cultivo (Terrones, et al., 2019) causando pudrición suave de color marrón, donde se
forman conidióforos y conidios de color gris que le da a la enfermedad su nombre de “moho
gris” (Fillinger y Walker, 2016).
Si la enfermedad no se controla puede ocasionar importantes pérdidas. El control químico
es el principal medio eficiente y confiable contra el moho gris y representa el 8% del mercado
mundial de fungicidas (Phillips y McDougall 2012). Existen siete modos de acción de
fungicidas unisitio específicos y multisitio para el control de B. cinerea autorizados para
aplicarse en zarzamora en México en cumplimiento con USA y E.U., los cuales son
fluazinam, fenhexamid, tiofanato metil, captan, pirimethanil, fludioxonil, boscalid,
azoxistrobin e iprodiona (ANEBERRIES, 2019), estos se aplican a intervalos de 7-14 días,
lo cual incrementa los costos de producción y pueden generar resistencia (Phillips y
McDougall 2012). Para minimizar el uso de estos se puede utilizar el Silicio el cual modifica
las propiedades de la pared celular inhibiendo la penetración de los hongos y activa la
expresión de genes de resistencia de las plantas (Fauteux et al., 2005). Existen muchos
estudios sobre la disminución de enfermedades con Si en diferentes patógenos, sin embargo,
no existe un reporte formal sobre la aplicación de Si en zarzamora para el control del moho

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
232
gris. Por lo que el objetivo de este estudio fue realizar control químico de nueve botryticidas
de diferentes familias químicas solos y en combinación con silicato de potasio para
potencializar su efecto contra B. cinerea en condiciones de campo.
MATERIALES Y MÉTODOS
Tratamientos. En plantas de zarzamora var. Tupi, en campo, se realizaron dos
experimentos: en el primero se evaluaron los fungicidas fluazinam, fenhexamid, tiofanato
metil, captan, pirimethanil, fludioxonil, boscalid, azoxistrobin e iprodiona más un testigo,
solos y en combinación con silicato de potasio a dosis recomendada en la etiqueta del
producto. En el segundo experimento se evaluaron tres dosis, baja, media y alta de
pirimethanil y azoxistrobin en combinación con silicato de potasio.
Diseño y unidad experimental. Los experimentos se condujeron bajo un diseño factorial,
el primero 10×2, fueron 20 tratamientos con cuatro repeticiones y se utilizaron 80 unidades
experimentales, el segundo fue 2×3, fueron 6 tratamientos con cuatro repeticiones y se
utilizaron 24 unidades experimentales; cada una consistió de una parcela de 7 m de largo por
2 m de ancho con 14 plantas de zarzamora distribuidas en hilera. Ambos experimentos se
repitieron en el tiempo.
Aplicación de tratamientos. Se realizaron a intervalos de 15 días hasta llegar al final de
la cosecha. En total se realizaron seis aplicaciones, con una aspersora motorizada
(Maruyama, MS072H) con boquilla de cono hueco realizando una cobertura total del follaje
(gasto 800 l/ha).
Variables. Al momento de la cosecha se seleccionaron 100 frutos cada unidad
experimental para la evaluación de incidencia, severidad, ° Brix, rendimiento y concentración
de Silicio en la planta.
Análisis de datos. Los datos de las variables fueron sometidos a un análisis de varianza y
comparación de medias (Diferencia Mínima Significativa) utilizando el programa SAS
versión 9.3.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
233
RESULTADOS Y DISCUSIÓN
Evaluación de incidencia de B. cinerea. En el primer experimento el ABCPE varió
significativamente en ambos ensayos (F = 540.5, p = ˃0.0001) (F = 376.27, p = ˃0.0001), las
plantas tratadas con Azoxistrobin más silicato de potasio presentaron menor ABCPE
significativamente diferentes al resto de los tratamientos, con valores de 1320 y 1099 en el
primer y segundo ensayo. En el segundo experimento en ambos ensayos, se identificaron
diferencias significativas (F = 495.59, p = ˃0.0001) (F = 645.87, p = ˃0.0001) en el ABCPE.
Las plantas tratadas con dosis media y alta de azoxistrobin más silicato de potasio presentaron
menor ABCPE significativamente diferentes al resto de los demás tratamientos, con valores
de 671.3 (dosis media) y 532.5 (dosis alta) en el primer ensayo y 607.5 (dosis media) y 463.13
(dosis alta) en el segundo.
Evaluación severidad de B. cinerea. En el primer experimento, en el primer ensayo, en
el ABCPE hubo diferencias significativas (F = 502.96, p = ˃0.0001). Los mejores
tratamientos iprodiona, iprodiona más silicato de potasio y azoxistrobin mas silicato de
potasio presentaron menor ABCPE y fueron significativamente diferentes al resto de los
tratamientos, con valores de 355.8, 338.2 y 298.5 respectivamente. En el segundo ensayo,
hubo diferencias significativas (F = 373.06, p = ˃0.0001) en el ABCPE. Todos los
tratamientos fueron diferentes al testigo, siendo la combinación de azoxistrobin más silicato
de potasio el tratamiento que más redujo la severidad de la enfermedad con un valor de 214.5.
En el segundo experimento en ambos ensayos se observaron diferencias significativas en el
ABCPE (F = 613.39, p = ˃0.0001) (F = 795.13, p = ˃0.0001); las plantas tratadas con dosis
media y alta de azoxistrobin más silicato de potasio presentaron menor ABCPE el cual fue
significativamente diferente a los demás tratamientos con valores de 277.05 y 247.13,
respectivamente, en el primer ensayo y 265.72 y 233.50, respectivamente, en el segundo
ensayo.
Sólidos solubles totales. En el primer experimento, en los °Brix en ambos ensayos se
identificaron diferencias significativas (F = 611.47, p = ˃0.0001) (F = 347.31, p = ˃0.0001).
En las plantas que se trataron con iprodiona más silicato de potasio y azoxistrobin solo, se
encontró la mayor cantidad de solidos solubles con valores de 7.49 y 7.45, respectivamente,

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
234
para el primer ensayo y 7.55 y 7.50 para el segundo. En el segundo experimento en los °Brix
en los dos ensayos, se identificaron diferencias significativas (F = 527.56, p = ˃0.0001) (F =
2234.48, p = ˃0.0001). En las plantas que se trataron con la dosis alta de azoxistrobin más
silicato de potasio se encontró la mayor cantidad de solidos solubles con valores de 7.61 y
7.70 para el primer y segundo ensayo.
Concentración de silicio. En el primer experimento, en ambos ensayos, se identificaron
diferencias significativas (F = 661.90, p = ˃0.0001) (F = 661.28, p = ˃0.0001). Las plantas
donde se aplicó azoxistrobin más silicato de potasio fueron significativamente diferentes; se
encontró mayor concentración de Si en ambos ensayos con valores de 50.79 y 53.75 ppm
para el primero y segundo ensayo. En el segundo experimento, en el primer ensayo se
identificaron diferencias significativas (F = 563.40, p = ˃0.0001). Las plantas tratadas con
dosis alta de azoxistrobin más silicato de potasio fueron significativamente diferentes ya que
hubo mayor concentración de Si con valores de 51.41. En el segundo ensayo hubo diferencias
significativas (F = 1430.31, p = ˃0.0001). Las plantas que se trataron con dosis media y alta
de azoxistrobin más silicato de potasio fueron significativamente diferentes con valores de
52.02 y 53.01 ppm respectivamente.
Rendimiento. En el primer experimento, en ambos ensayos, se encontraron diferencias
significativas (F = 429.24, p = ˃0.0001) (F = 3065.12, p = ˃0.0001). Las plantas asperjadas
con el fungicida azoxistrobin en combinación con silicato de potasio fueron
significativamente diferentes en ambos ensayos, presentando el mayor rendimiento de 12.44
y 13.62 t ha-1 en el primer y segundo ensayo. En el segundo experimento, en los dos ensayos,
se encontraron diferencias significativas (F = 1116.69, p = ˃0.0001) (F = 3269.08, p =
˃0.0001). Las plantas tratadas con dosis alta de azoxistrobin más silicato de potasio fueron
significativamente diferentes, presentando el mayor rendimiento de 13.22 y 13.99 t ha-1 para
el primer y segundo ensayo, respectivamente.
Con la adición de silicato de potasio en todos los fungicidas se obtuvo una reducción de
la incidencia y severidad del moho gris, lo cual se vio reflejado en el ABCPE; esto se debe a
que el Si modifica las propiedades de la pared celular, la estructura de la planta y crea una
capa externa dura y activa las defensas naturales de las plantas, principalmente en la

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
235
expresión de genes (Fauteux et al., 2005). Con base en los conceptos generales de Jennings
(2007) sobre nutrición y crecimiento micelial de hongos, al adherirse el Si a la pared celular,
las enzimas de las hifas no actúan eficientemente en desdoblar la celulosa a glucosa y la
nutrición del hongo es afectada negativamente por lo que la enfermedad se reduce (Datnoff
et al., 2011). El azoxistrobin inhibe la oxidación de NADH y la síntesis de ATP (Fillinger et
al., 2016); es un fungicida muy eficiente por actuar en la respiración fúngica y producción
de energía (Ishii et al. 2009); cuando se aplicó solo presentó buen control de la enfermedad
en las primeras tres evaluaciones; a partir de la cuarta se observó que no controlaba de la
misma manera por lo cual se identificó que el patógeno pudo haber desarrollado resistencia;
esto no se observó cuando el azoxistrobin se combinó con silicato de potasio; este efecto se
debe a las propiedades que presenta el Si (Rodrigues et al., 2015) por tal motivo la
combinación de silicato de potasio con botryticidas en el control del moho gris causado por
B. cinérea disminuye eficientemente la incidencia y la severidad así como evita que el
patógeno desarrolle resistencia a fungicidas.
AGRADECIMIENTOS
Al CONACYT por la beca No. 429090 otorgada al primer autor para realizar sus estudios
de doctorado y al Colegio de Postgraduados por el apoyo en mi formación académica.
LITERATURA CITADA
ANEBERRIES, 2019. Internet. http://www.aneberries.mx/lista-de-plaguicidas-autorizados-aneberries-2019 fecha de consulta. 18/06/19.
Datnoff L., E., H Snyder G., and H Korndörfer G. 2011. Silicon in Agriculture. Studies in Plant Science, 8. Florida, USA. 403p. http://dx.doi.org/10.1016/S0928-3420(01)80001-X
Fauteux F., W Rémus B. and G Menzies J. 2005. Silicon and plant disease resistance against pathogenic fungi. FEMS Microb Lett 249:1–6. doi: 10.1016/j.femsle.2005.06.034
Fillinger and Walker. 2016. Chemical Control and Resistance Management of Botrytis Diseases. In S. Fillinger, Y. Elad (eds.), Botrytis – the Fungus, the Pathogen and its Management in Agricultural Systems, DOI 10.1007/978-3-319-23371-0_10
Jennings D., H. 2007. The Physiology of Fungal Nutrition. Cambridge University Press. New York, USA. 622p. DOI:10.1007/BF02908824
Li X., D Fernández O., W Chai, F. Wang and G Schnabel. 2012. Identification and prevalence of Botrytis spp. from blackberry and strawberry fields of the Carolinas. Plant Disease 96(11): 1634–1637. https://doi.org/10.1094/PDIS-02-12-0128-RE
Phillips M., W., A. and J McDougall. 2012. Crop protection market trends and opportunities for new active ingredients. Abstracts of Papers of the American Chemical Society 244.

AvancesdeInvestigaciónColegiodePostgraduadosPosgradoenFitosanidad
2019
236
Rodrigues E., J. 2015. Silicon Control of Foliar Diseases in Monocots and Dicots. In: F.A. Rodrigues, L.E. Datnoff (eds.), Silicon and Plant Diseases, DOI 10.1007/978-3-319-22930-0_4
SAGARPA, Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación. Servicio de información agroalimentaria y pesquera. 2018. https://www.gob.mx/siap/acciones-y-programas/produccion-agricola-33119. (Consulta, mayo de 2018).
Terrones S., J., D., Nieto A., C Nava D., D Téliz O., R García V., M R Vallejo P and P Sánchez G. 2019. Botrytis cinerea causing gray mold in blackberry fruit in Mexico. Mexican Journal of Phytopathology 37(3). DOI: 10.18781/R.MEX.FIT.1906-1