las especies de brúquidos (gorgojos de las leguminosas) de ... · forma de casquete esférico por...
TRANSCRIPT

«Las especies de Brúquidos (gorgojos de las leguminosas) de interés agrícolay fitosanitario (Col. Bruchidae)»
I: Caracteres generales
R. Yus RAMOS
En el presente trabajo, el autor hace una revisión de los caracteres generales,morfológicos y biológicos, de los coleópteros pertenecientes a la familia Bruchidae,con lo que pretende introducir el estudio sistemático de las especies de interés agrí-cola y fitosanitario, como base para posteriores observaciones de inmediata aplica-ción práctica.
R. Yus RAMOS.—Centro Experimental «La Mayora» (C.S.I.C.). Algarrobo-Costa.Málaga.
INTRODUCCIÓN
La familia Bruchidae es un grupo homogé-neo de insectos pertenecientes al orden Co-leóptera, filogenéticamente comprendido en-tre otros Phytophaga tales como Chrysomeli-dae y Curculionidae, y cuya principal carac-terística biológica es la de presentar un régi-men alimenticio de tipo cletrofágico, esto es,devorador de semillas, en su mayor parte deespecies de Papilionáceas, entre las cualesquedan comprendidas numerosas especies deleguminosas comestibles, por lo cual estosinsectos revisten una gran importancia eco-nómica.
Los Brúquidos de interés agrícola y fitosa-nitario han sido objeto de numerosos estudiosencaminados al conocimiento de su biología,para posteriores ensayos destinados a su con-trol y lucha. En Estados Unidos (BRIDWELL,
1932; LARSON, 1924 y 1927), en Alemania
(ZACHER, 1930), en Inglaterra (HERFORD,
1935; SOUTHGATE, 1957), en Francia (LE-
PESME, 1945; VAYSSIERE, 1944), en Argelia(DE LUCA, 1956), etc.
A pesar de su gran importancia, en Españacarecemos actualmente de precedentes, tantoen lo que se refiere a los estudios sistemáti-cos como a los biológicos y faunísticos, y tansólo hemos encontrado algunos datos en lasComunicaciones del Boletín Informativo delServicio de Defensa contra Plagas e Inspec-ción Fitopatológica.
A la vista de la escasez de referencias bási-cas en nuestro país, pretendemos dar a cono-cer el «status» actual de estos insectos, paralo cual hemos dividido el trabajo en dos par-tes: una destinada a un conocimiento generale introductorio y otra encaminada al conoci-miento sistemático y biológico de las especiesperjudiciales a nuestros cultivos y almacenesde leguminosas.

La morfología externa de los distintos es-tados es la siguiente:
Huevo
No presenta caracteres especiales. Carecede micropilo. De tamaño variable, general-mente alrededor de 0,5 mm. de longitud y0,2 mm. de anchura, aunque hay especies (Bru-chidius) que presentan una longitud muchomenor: 0,2 mm. y otras (Pachymerus) muchomayor: 1 mm. Forma generalmente ovoide,unas veces casi subcilíndricos por ser másalargados (Acanthoseelides) y otras veces enforma de casquete esférico por ser más acor-tados {Callosobruchus). Suele presentar unacara dorsal más convexa (Fig. 14), de coriónmás endurecido y una cara ventral más plana,de corion más blando y por donde se fija alsustrato. Al principio el corion es hialino,translúcido, de superficie lisa y en el trans-curso de su evolución se torna más oscuro,de amarillo pálido a beige, opaco y de superfi-cie rugosa. En la mayor parte de las especiespuede apreciarse un extremo más ancho yabombado que corresponde con la posiciónde la cabeza del embrión.
Larva
En este estado, la morfología larvaria secorresponde con una etología diversa, dandolugar a un fenómeno de evolución larvariaformada por varios estadios claramente dife-renciables y que en conjunto recibe el nom-bre de hipermetamorfosis. En una hiperme-tamorfosis completa podemos distinguir dosestadios, de los cuales el primero presentauna larva primaria o crisomeloide y el se-gundo presenta una larva secundaria o melo-lontoide. Sin embargo, no todas las especiespresentan esta evolución y en determinados
casos se produce una aceleración de la onto-genia de tal modo que la larva neonata ya esde tipo melolontoide sin pasar por la fase delarva crisomeloide.
a) Larva primaria: llamada también neo-nata o crisomeloide, se trata de unalarva pequeña, generalmente de 0,6-1 mm. de longitud, arqueada, de cuerpodorsalmente convexo e inferiormentecasi plano, teniendo su máxima an-chura hacia la mitad anterior, estre-chándose hacia atrás. Cuerpo blanconiveo a excepción de la cabeza y placapronotal que son parduzcas y provistode sedas de diversos tamaños reparti-dos regularmente en cada segmento(Fig. 1).
Cabeza ovoide, subglobulosa, bastantegrande, un poco aplastada y ligera-mente quitinizada, parcialmente embe-bida en el protórax. Cráneo con unasutura epicraneal bien evidente, líneapor donde se realizará la emergencia dela larva durante ía muda, que junto conlas dos suturas frontales forman untriángulo frontal (Fig. 1), con sedasdispuestas a cada lado y poros senso-riales situados cerca de las suturas. An-tenas muy cortas, de 2 artejos, pococoloreadas, situadas en una escotaduracircular del epístoma, cerca de las fo-sas mandibulares (Fig. 1). Presenta unartejo basal redondeado, aplastadodorsalmente y del que sobresale un ar-tejo apical cilindrico, generalmenteterminado en una o varias sedas, au-mentando el número de éstas en cadamuda y cuya disposición y desarrolloson característicos en cada especie.Clípeo trapezoidal, más ancho que lar-go, coloreado. Labro cuadrangular, li-geramente convexo y provisto de sedasde disposición característica en cada
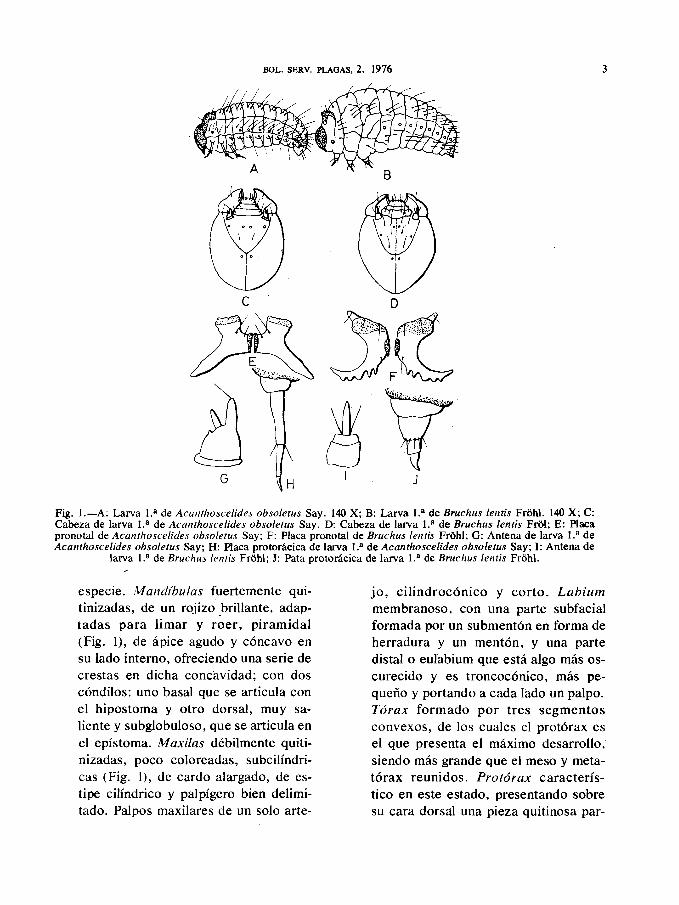
Fig. 1.—A: Larva 1.a de Acanthoscelides obsoletus Say. 140 X; B: Larva 1.a de Bruchus lentis Fróhl. 140 X; C:Cabeza de larva 1.a de Acanthoscelides obsoletus Say. D: Cabeza de larva 1.a de Bruchus lentis Fról; E: Placapronotal de Acanthoscelides obsoletus Say; F: Placa pronotal de Bruchus lentis Fróhl; G: Antena de larva 1.a deAcanthoscelides obsoletus Say; H: Placa protorácica de larva 1.a de Acanthoscelides obsoletus Say; I: Antena de
larva 1.a de Bruchus lentis Fróhl; J: Pata protorácica de larva 1.a de Bruchus lentis Fróhl.
especie. Mandíbulas fuertemente qui-tinizadas, de un rojizo brillante, adap-tadas para limar y roer, piramidal(Fig. 1), de ápice agudo y cóncavo ensu lado interno, ofreciendo una serie decrestas en dicha concavidad; con doscóndilos: uno basal que se articula conel hipostoma y otro dorsal, muy sa-liente y subglobuloso, que se articula enel epístoma. Maxilas débilmente quiti-nizadas, poco coloreadas, subcilíndri-cas (Fig. 1), de cardo alargado, de es-tipe cilindrico y palpígero bien delimi-tado. Palpos maxilares de un solo arte-
jo, cilindrocónico y corto. Labiummembranoso, con una parte subfacialformada por un submentón en forma deherradura y un mentón, y una partedistal o eulabium que está algo más os-curecido y es troncocónico, más pe-queño y portando a cada lado un palpo.Tórax formado por tres segmentosconvexos, de los cuales el protórax esel que presenta el máximo desarrollo,siendo más grande que el meso y meta-tórax reunidos. Protórax caracterís-tico en este estado, presentando sobresu cara dorsal una pieza quitinosa par-

duzca llamada placa pronotal o «ruptorovis» (Fig. 1) que consiste en una placamóvil formada por dos piezas quitino-sas con una rama dorsal más corta yotra ventral más larga y divergentes,unidas simétricamente en una líneamediana y formando una H o una X, deconformación característica en cadaespecie, presentando dos ramas supe-riores lisas y dos posteriores provistasde sedas, crestas y dientes de disposi-ción y número característicos en cadaespecie.
Mesotórax más estrecho que el metató-rax dorsalmente, e inferiormente, cadaterguito presenta dos regiones denomi-nadas pre-escutum y escuto-escutellumdivididos por un pliegue. Pleuritos bas-tante convexos, escindidos en dos ló-bulos que se prolongan hacia el abdo-men. Esternitos presentando un lóbulocoxal a cada lado, punto de inserciónde las patas, siendo el protorácico elmenos desarrollado. Patas pequeñas,delgadas, formadas por 4 artejos (Figura1), las protorácicas dirigidas hacia de-lante y menos desarrolladas y el restohacia atrás y más desarrolladas, sobretodo las metatorácicas. Presentan unanca globulosa asentada sobre el lóbulocoxal, un fémur alargado, troncocónicoy en cuyo extremo presenta general-mente una corona variable de sedas,tibia más corta y cilindrica y tarso máscorto aún y formado por una uña úni-ca. Estigmas protorácicos formadospor un par, del tipo uniforados, de pa-redes lisas, de boca rojiza y oval, si-tuados cerca de las mesopleuras.
Abdomen enteramente blanco, carno-so, formado por 9 segmentos cuya an-chura va disminuyendo progresiva-mente hacia atrás. Los 8 primeros
segmentos están ocupados dorsalmen-te por su pre-escutum y escuto-escu-tellum, que están bien delimitados;el 9.° está poco diferenciado y presen-ta dos lóbulos que son utilizados paraayudar a la marcha; pleuritos con dosborlas longitudinales, poco evidentesen los dos últimos segmentos. Escle-ritos ligeramente deprimidos y dismi-nuyendo progresivamente de tamañohacia atrás. Todos los segmentos pre-sentan una quetotaxia característica encada especie y, a excepción del 9.°, to-dos llevan un par de estigmas abdomi-nales uniforados, de orificio oval y pos-teriormente se hacen circulares, el másimportante es el primer par, pues estáformado por varios sacos aeríferos yel resto va disminuyendo progresiva-mente de importancia.
b) Larva secundaria: llamada tambiénmelolontoide o curculionoide. En esteestadio se producen profundos cambiosque afectan fundamentalmente a la ca-beza (se hace más voluminosa, pero re-lativamente más pequeña, quedandoescondida dentro del protórax), pro-noto (desaparece la placa pronotal), an-tenas (aparecen más quetas) y patas(van perdiendo su función y el desarro-llo de los lóbulos coxales acaban poresconderlas, quedando aparentementeo totalmente ápodas) (Fig. 2).
Cabeza más clara, rosácea, práctica-mente escondida dentro del protórax,algo más grande, subhemiesférica, consuturas borradas progresivamente, sinporos sensoriales. Antenas salientes,con dos artejos subcilíndricos casi igua-les y coronados por quetas, sedas oapéndices ampuliformes. Clípeo curvi-línea y trapezoidal, transverso. Labrotransverso (Fig. 2), aplastado, con una

Fig. 2.—A: Larva 2.a de Acanthoscelides obsoletas Say. 35 X; B: Larva 2.a de Bruchus lentis Fróhl. 35 X; C:Complejo clípeo-labral de larva 2.a de Acanthoscelides obsoletus Say; D: Complejo clípeo-labral de larva 2.a deBruchus lentis Fróhl; E: Maxila derecha de larva 2.a de Acanthoscelides obsoletus Say; F: Maxila derecha de
larva 2.a de Bruchus lentis Fróhl.
quetotaxia aumentada, dispuesta en fi-las simétricas. Mandíbulas muy desa-rrolladas, tan anchas como largas, conuna concavidad interna más larga yprovista de sedas, con cóndilo basalmuy desarrollado. Maxilas de cardo yestipe alargados (Fig. 2), de secciónrectangular, con sedas subparalelas enla base y luego divergentes. Palpígeroprovisto de algunos poros sensoriales.Palpos con un solo artejo, provisto ensu ápice de varias protuberancias papi-liformes. Labium más desarrollado,con predominante desarrollo del eula-bium, con estipe y palpos aislados enprogresivas mudas.
Tórax ocupando la mitad del cuerpo enlas primeras mudas, hasta hacerse rela-tivamente más pequeño y ocupando 1/3del cuerpo. Protórax mucho más desa-rrollado que los restantes segmentos,sin placa protorácica. Pleuritos conpliegues transversales más evidentes.Patas rechonchas por un aumento dedesarrollo de los lóbulos coxales, conancas mal delimitadas, en algunas es-pecies llegan a desaparecer, apare-ciendo en su lugar un lóbulo coronadopor una serie de quetas diminutas.Conforme avanzan las mudas, las patasmesotorácicas se dirigen hacia delante,quedando dirigidas hacia atrás tan sólo

las metatorácicas. La sutura que se-para el pre-escutum y el escutellummesotorácicos se confunde con la su-tura posterior del protórax. El númerode sedas disminuye.Abdomen con segmentos más volumi-nosos, sobre todo dorsalmente, adop-tando en conjunto una disposición ar-queada, comprimidos unos con otros,con pleuras más sobresalientes. Pre-senta una sección pleuro-tergal semi-circular y una sección pleuro-esternalpoligonal curvilínea. Cada terguitotiene dos regiones: escutum y escute-llum. El esternito no presenta división.Los dos últimos segmentos son máspequeños e indiferenciados. Los es-tigmas están presentes en cada seg-mento a excepción del último; estánmuy desarrollados, insertos en la mitadde una placa dorsoventral o alar, conexcrecencias quitinosas, con boca circu-lar e internamente con mayor abun-dancia de sacos aeríferos.
Ninfa
Al término de una serie de mudas, cuyonúmero oscila entre 3-6, la larva adulta, sinpasar por una muda se distiende y acusa unaserie de cambios que en conjunto recibe elnombre de preninfa, antesala de la verdaderaninfa.
a) Preninfa: de forma oval-alargada,adelgazada anteriormente y presen-tando su máxima anchura hacia el 6.°segmento abdominal (Fig. 3). Desapa-recen las patas (si las hubiera) y en sulugar quedan los lóbulos coxales pro-vistos de abundantes quetas en el ápi-ce. Todos los segmentos (torácicos yabdominales) son muy evidentes y la
cabeza queda replegada sobre el protó-rax.
b) Neoninfa: al término de la muda de lapreninfa aparece una forma incompletay esbozada de ninfa, caracterizada poruna disminución de longitud debido aun retraimiento de los segmentos y unabatimiento de la cabeza y protóraxsobre la parte ventral (Fig. 3). Elcuerpo es globuloso, glabro y blanconiveo, casi transparente. Aparece unacabeza voluminosa, con un esbozo deojos compuestos, rudimentos de palposlabiales y maxilares. Las antenas apa-recen ya con un número de artejos pe-queño y con un desarrollo de los arte-jos apicales bruscamente superior, loscuales se apoyan en la articulacióntibio-femoral de las patas anteriores. Elprotórax aparece claramente delimi-tado pero muy voluminoso. Las patasson muy gruesas, sin articulacionesevidentes, con un gran desarrollo delos futuros tarsos y dispuestos de unaforma peculiar, siempre constante: fé-mur y tibia plegados, con tarsos dirigi-dos hacia abajo. También puede apre-ciarse una región elitral con algunas es-trías esbozadas y un abdomen consegmentos no delimitados, a excepcióndel último.
c) Ninfa definitiva: de forma poco dife-rente a la neoninfa, pero con órganosmás delimitados y más perfectamenteformados (Fig. 3). La cabeza se hacemás pequeña, apareciendo unos ojoscompuestos reniformes más perfectosy un rostro más estrechado y prolon-gado. Las antenas se han estilizado sinaumentar de longitud y sus artejos sonmás semejantes entre sí. Protórax,asimismo más empequeñecido. Patasmás estilizadas, con articulaciones evi-

Fig. 3.—A: Preninfa de Bruchus lentis Fróhl. 30 X; B: Neoninfa de Bruchus lentis Frohl. 30 X; C: Ninfa deBruchus lentis Frohl. 30 X.(inspirado en Y. DE LUCA)
dentes. Alas sobresaliendo por debajode los élitros, aún no replegadas. Ab-domen voluminoso, con segmentosbien evidentes, pudiéndose distinguirya el sexo, pues el 9.° segmento apa-rece escotado en los machos y recto enlas hembras.
Imago
La familia Bruchidae agrupa un conjuntode especies que presentan una gran homoge-neidad de caracteres en el adulto. El tamañovaría en un corto intervalo comprendido en-tre 1 mm. (Br. perparvulus Boh.) a 8 mm.(Pachymerus difformis OÍ.), aunque por tér-mino medio no suelen pasar de 2-4 mm. delongitud. Su forma es oval, unas veces acor-
tada y tomando aspecto rechoncho, y otrasalargada, llegando a ser en ocasiones extre-madamente delgados (Br. cinerascens Gyll.).
La pubescencia es un carácter importanteen esta familia, aunque no tan decisivamentea nivel de especie, por cuanto se trata de unacualidad de estado, esto es, susceptible devariación según la planta huésped o clima eincluso hasta por la propia variabilidad del in-secto.
Está formada por pelos de disposición ge-neralmente tendida, normalmente bastantecortos, de apariencia escamosa y cuya densi-dad es variable, en muchos casos llegando aocultar los tegumentos. En general podemosafirmar que la pubescencia se hace más densaen el lóbulo basal del pronoto, en el escudetey en la sutura, apareciendo manchas triangu-lares muy características. En ocasiones, la

pubescencia es clara: blanca a amarillo-pajizay otras veces aparece más oscura: rojiza,ocre, gris, negra. En conjunto adoptan dispo-siciones características formando dibujos enel pronoto y los élitros.
Los tegumentos son en general de tonali-dad oscura, variando del negro al parduzco,con un brillo más o menos intenso, normal-
mente ocultado por la pubescencia. En oca-siones aparecen zonas de color rojizo quecontrastan con el resto que es oscuro (Br. bi-guttatus OÍ.), o bien este color se extiendeampliamente (Caryoborus pallidus OL). Lasuperficie rara vez es lisa, en virtud de la pre-sencia de una puntuación de densidad y con-formación variables y en ocasiones por la
Fig. 4. A: Cara dorsal de imago hipotético de Brúquido B: Cara ventral de imago hipotético de Brúquido
1.— Clípeo2.—Frente3.—Pata protorácica4.—Pronoto5.—Escudete6.—Calus humeral7.—Pata me so torácica8.—Sutura elitral9.—Interestría elitral
10.—Estrías elitrales11. —Pata metatorácica12.—Pigidio
1.—Palpos maxilares2.—Mentón3.—Pregula4.—CueUo5.—Epímera y episterna proto6.—Prosternón7.—Ancas pro torácicas8.—Epímera mesotorácico9.—Espiterna mesotorácico
10.—Mesosternón11. —Epipleura elitral12.—Espisterna metatorácico13. —Metas ternón14.—1.° segmento abdominal15.—2.° segmento abdominal16.—3.° segmento abdominal17.—4.° segmento abdominal18.—5.° segmento abdominal19.—Pigidio

presencia de estrías cuya presencia es siem-pre constante en los élitros. Otras veces en-contramos granulaciones a modo de tubércu-los en disposición lineal (Br. tuberculatusHochh.) y en ciertas especies en la base de la5.a interestría elitral (Br. biguttatus OÍ.). Porúltimo, hay especies que presentan surcos,generalmente localizados en el pronoto (Br.holosericeus Schón.), rara vez en la cabeza(B. laticollis Boh.) donde es más frecuente lapresencia de una pequeña cresta longitudinalmediana situada en la frente.
La cabeza es de apariencia trapezoidal,globosa en la mitad basal, ensanchándose enla mitad por la presencia de los ojos y estre-chándose hacia la extremidad en forma derostro aplastado, ligeramente inclinado haciadelante y susceptible de abatirse sobre la
parte anterior del prosternen (Fig. 5). Vistadorsalmente, encontramos en la base el cue-llo (Fig. 5) más o menos globoso, al que siguela frente, que ocupa el espacio interocular,luego el epístoma o clípeo que soporta ante-riormente el labro y lateralmente las mandí-bulas. Ventralmente encontramos en la base lacavidad occipital en posición inclinada, a laque sigue más anteriormente la gula y a con-tinuación la pregula que soporta las piezas la-biales.
Ojos globosos, a veces bastante salientes,escotados anteriormente y adoptando un as-pecto reniforme, con la rama ventral más an-cha.
Antenas insertas delante de la escotaduraocular, de longitud y conformación variables,compuestas de 11 artejos, de los cuales el 1.°
Fig. 5.—A: Cara dorsal de cabeza de Bruchus pisorum L; B: Cara ventral de cabeza de Bruchus pisorum L; C:Cara lateral de cabeza de Bruchus pisorum L; D: Mandíbula izquierda de Bruchus pisorum L; E: Maxila derecha
de Bruchus pisorum L; F: Labium de Bruchus pisorum L.

y el 11.° son oblongos y el resto varía segúnlas especies y el sexo. En general, a mayorlongitud de las antenas corresponden artejosmás dentados, por lo que encontramos ante-nas cortas y subfiliformes y antenas largas,pasando de la base al pronoto, y aserradas(Fig. 6).
Mandíbulas triangulares, aplastadas dorso-ventralmente. Presentan una base plana condos engrasamientos o cóndilos, puntos de in-serción de los músculos masticadores y unápice puntiagudo, curvado hacia el interior enforma de diente (Fig. 5). Mola aplastada ycortante, con una expansión laminar más omenos cortante.
Maxilas falciformes, con cardo alargado,sinuoso y estipe comprimido dorsoventral-mente (Fig. 5); gálea y lacinia anchas, provis-tas de pelos sobre su margen interno y máslargos sobre el ápice; palpígero reducido y enocasiones ausente, con palpos maxilares lar-gos, compuestos de tres artejos, aunque enocasiones (Acanthoscelides) presenta cuatroartejos.
Labium transverso, generalmente lobuladoanterolateralmente. En su mitad anterior sesitúa la lengüeta labial, más o menos circular(Fig. 5), provista de poros sensoriales en sucara dorsal y una serie de sedas en su margenanterior. Naciendo de la mitad de ésta se en-
Fig. 6.—A: Antena izquierda de Caryopemon Llostet Pie; B: Antena izquierda de Bruchidius jocosus Schónh; C:Antena izquierda de Pachymerus accaciae Gyll; D: Antena izquierda de Bruchidius cinerascens Gyll; E: Antenaizquierda de Bruchidius stylophorus K. Dan; F: Antena izquierda de Bruchidius biguttatus 01; G: Antena iz-
quierda de Bruchidius seminarius L; H: Antena izquierda de Bruchus laticollis Boh.

E F RY76
Fig. 7.—A: Pronoto de Bruchus perezi Kraatz; B: Pronoto de Bruchidius cinerascens Gyll; C: Pronoto de Callo-sobruchus chinensis L; D: Pronoto de Caryoborus pallidus 01; E: Cara ventral de protórax de Bruchus pisorum L;
F: Cara lateral de protórax de Bruchus pisorum.
cuentran los palpos labiales, que son cortos ycompuestos de tres artejos.
El protórax es de forma variable (Fig. 7),generalmente cónico, con la parte anteriormás estrecha, otras veces más ancho quelargo o transverso. Con escleritos soldados,formando un armazón sólido. Episterna yepímera soldados. Pronoto algo convexo, conborde anterior algo redondeado y ángulos an-teriores atenuados, obtusos. Lados rectos oredondeados, en ocasiones con un diente oescotadura situada más o menos hacia la mi-tad (Bruchus). Borde posterior bisinuado,formando un lóbulo basal más o menos acen-tuado y ángulos posteriores agudos. Proster-nón reducido por la proximidad de las cavi-dades coxales y con el borde anterior esco-tado para dar alojamiento al cuello\
En relación con el mesotórax, ésteespoco de-sarrollado (Fig. 8). Consta dorsalmente del es-cudete o mesonoto de forma variable (Figura8), que queda soldado a las restantes piezasdel mesoterguito. Mesosternón corto, conpre-escutum y post-escutellum soldados yformando una pieza subtriangular; mesoepi-meron estrecho, reducido a una fina bandatriangular que toca o no a las cavidades coxa-les (Fig. 8).
El metatórax consta dorsalmente del meta-noto, de naturaleza membranosa, no visibleexteriormente por quedar oculto por los éli-tros (Fig. 8). Ventralmente consta del metas-ternón que se prolonga anteriormente enforma de apófisis que avanza entre las ancasmesotorácicas, soldándose lateralmente conla episterna metatorácica y posteriormente

^ u RVR Y76
Fig. 8.—A-l: Escudete de Bruchus lentis Fróhl; A-2: Escudete de Bruchus rufimanus Boh; A-3: Escudete deAcanthoscelides obsoletus Say; A-4: Escudete de Pachymerus accaciae Gyll; A-5: Escudete de Bruchidius poupi-llieri All; A-6: Escudete de Callosobruchus maculatus F; A-7: Pseudopachymerus lallemantii Mars; A-8: Escu-dete de Spermophagus sericeus Geoffr; B: Metanoto de Bruchus pisorum L; C: Cara ventral de meso-metatórax
de Bruchus pisorum L; D: Cara ventral de meso-metatórax de Zabrotes subfasciatus Boh.
formando el reborde de las cavidades coxalesmetatorácicas que a su vez están limitadas in-teriormente por la apófisis del 4.° esternitoabdominal (Fig. 8).
Las patas son órganos muy importantes enla ordenación sistemática de las especies. Sudesarrollo va aumentando desde las patasanteriores a las posteriores. Coxa-oval-subglobulosa, trocánter pequeño y ocu-pando tan sólo la parte postero-basal del fé-mur (Fig. 9). Fémur oblongo-alargado o fusi-forme, alcanzando su máximo desarrollo enlas patas posteriores, carácter que, aunqueevidente en las especies paleárticas, se vemucho más acentuado en los géneros tropica-les (Pachymerinae). Ventralmente presenta
dos costillas más o menos desarrolladas deli-mitando un canal donde se aloja el margen in-terno de la tibia cuando ésta se abate; nor-malmente simple en los dos primeros paresde patas y con escotaduras o dientes en elmargen apical interno de las patas posterio-res. Estos son de conformación variable, en-contrándose unas veces un diente fuerte yagudo que nace de la costilla externa yadopta forma de escotadura (Bruchus), otrasveces es más pequeño o ausente (Bruchidius)y otras forma un peine compuesto de diminu-tos dientes (Pachymerus, Caryoborus) (Fi-gura 9).
Tibia de desarrollo acorde con la del fémurcorrespondiente, generalmente las anteriores

Fig. 9.—A-l: Fémur metatorácico deBruchus lentis Fróhl; A-2: Fémur metatorácico de Callosobruchus chinensisL; A-3: Fémur metatorácico de Pseudopachymerus lallemantii Mars; A-4: Fémur metatorácico de Bruchidiuscaninus Kraatz; A-5: Fémur metatorácico de Acanthoscelides obsoletus Say; A-6: Fémur metatorácico de Pa-chymerus accaciae Gyll; B: Pata metatorácica de Bruchuspisorum L; C: Detalle de los tarsos deBruchus pisorum
L; D: Detalle de las uñas de los tarsos de Bruchus pisorum L.
e intermedias son cilindricas y las posteriorespiramidales. En las especies de Bruchus losmachos presentan un desarrollo de las tibiassuperior al de las hembras, frecuentementelas anteriores son más ensanchadas y aplas-tadas dorsoventralmente y las intermediasson arqueadas, más robustas y provistas dedientes o espolones en el margen apical in-terno.
Tarsos parecidos en los tres pares de patas.Presentan cuatro artejos visibles, por lo quefueron considerados tetrámeros (4-4-4), aun-que un estudio más detallado demuestra quepresentan un quinto artejo de desarrollo ín-fimo y que se encuentra ocluido en el tercerartejo, que siempre es bilobulado, por lo queen realidad son pentámeros (5-5-5) o más co-
rrectamente, criptopentámeros. El 5.° artejopresenta un par de uñas simples, no hendi-das, algo engrosadas en la base (Fig. 9).
Los élitros son generalmente ligeros, dequitinización no muy pronunciada, en reposocubriendo las piezas dorsales del meso y me-tatórax, así como los terguitos abdominales, aexcepción del último o pigidio. Ligeramenteconvexo en el disco, con el borde anterior si-nuoso. Calus humeral bien aparente. Superfi-cie elitral surcada por 10 estrías longitudina-les, de las cuales las centrales están más omenos acortadas (Fig. 10). Interestrías algoconvexas, generalmente punteadas y rara veztuberculadas (Br. tuberculatus).
Todas las especies conocidas presentan unpar de alas membranosas metatorácicas, aun-

que hay especies que presentan mutacionesápteras (Cali, maculatus F.). La nerviaciónpuede clasificarse como del tipo adephaga,es decir, con nervaduras transversales y laM-l muy separada apicalmente de la M-2. Depigmentación escasa o nula, predominandouna tonalidad clara y translúcida; con unaorla de pequeños pelos desde el ápice hasta ellóbulo anal.
La nervadura costal (CO) y subcostal (SC)forman un tronco quitinizado del que arrancala nervadura radial (R-l) de desarrollo varia-ble (Fig. 10). La R-2 aparece en la línea apicalmediana y la mediana (M-l) corta oblicua-mente el primer tercio apical del ala libre, de-teniéndose en el borde donde forma un vérti-ce. La M-2 arranca del tronco subcostal yestá bien desarrollada. Las cubitales (C-l,
C-2) se reúnen distalmente y por último,puede aparecer una tenue nervadura anal (A)en el lóbulo de su nombre, que en ocasionespuede llegar a desaparecer.
Abdomen
Formado por 9 segmentos compuestos poruna parte superior o tergo de naturalezamembranosa, una parte lateral o pleura for-mada por dos zonas: una cercana a la suturapleural o epipleura y la siguiente a ésta o hi-popleura, e inferiormente el esternón de natu-raleza más quitinizada. De los 9 terguitos sóloson visibles exteriormente el 9.° o pigidio. Delos 9 esternitos sólo son visibles cinco deellos, ya que los tres primeros son de natura-
Fig. 10.—A: Élitro izquierdo de Bruchus pisorum L; B: Ala derecha de Acaníhoscelides obsoletus Say; C: Caradorsal de abdomen de Bruchus pisorum L; D: Cara ventral de abdomen de Bruchus pisorum L.

leza membranosa, con suturas poco aparen-tes. Le siguen el 4.° y 5.° que están soldadosformando un gran esternito con un procesointercoxal mediano (Fig. 10). Los segmentos6.° a 9.° son estrechos y el 9.° puede esco-tarse para dar alojamiento a la punta del pigi-dio, carácter que aparece sólo en los machos.
Órgano genital del macho
Presentan 4 testículos reunidos en dos pa-res separados. Cada testículo está formadopor 5 glándulas testiculares que se reúnenunas junto a otras como las hojas de un libro(Fig. 11). Cada testículo presenta un corto
Fig. 11.—A: Complejo visceral de genitalia de macho deBruchus rufipes Herbst.1.—Testículos2.—Canal deferente3.—Glándulas accesorias4.—Vesícula seminal5.—Canal eferente6.—Ampolla eyaculadora7.—Lóbulo mediano8.—Espícula gastral9.—Saco interno
10.—Tegmen
B: Segmento genital de Bruchus rufipes Herbst.
1.—Terguito2.—Esternito
C: Espícula gastral de Bruchus rufipes Herbst.D: Exofalo de Bruchus rufipes Herbst.
1.—Orificio apical2.—Lóbulos laterales3.—Vaina4.—Columna tegminal5.—Conducto eyaculador
E: Endofalo de Bruchus rufipes Herbst.
F: Tipos de escleritos de endofalo
a.—espinab.—espículac.—dentículosd.—serrata

canal que desemboca, junto con el otro delmismo par, en un canal seminal o deferenteque en su trayecto tiene un par de glándulasaccesorias que desembocan en un ensacha-miento del canal deferente llamado vesículaseminal. Ambos canales deferentes desembo-can en un único canal, denominado canaleyaculador, que presenta un ensanchamiento oampolla eyaculadora, al que sigue un cortotrayecto que se introduce en la armadura ge-nital por el orificio basal e internamente seensancha formando el endofalo o saco in-terno.
La armadura genital constituye una trans-formación de los segmentos X y XI del abdo-men que tras un proceso de invaginación on-togénica, quedaron en posición interna, acu-sando profundas modificaciones. Consta dedos partes:
a) Segmento genital: producto de latransformación del X segmento abdo-minal. Constituye una pieza membra-nosa que rodea al órgano copulador ensu parte distal, bajo el pigidio. En elborde posterior del órgano copuladorse encuentra una pieza quitinosa enforma de horquilla denominada espí-cula gastral, producto, al parecer de latransformación del XI urosternito, ycuya misión es la de sostener al órganocopulador (Fig. 11).
b) Órgano copulador: formado a su vezpor dos partes: una externa, más o me-nos quitinizada o exofalo y una interna,de naturaleza blanda llamada endofaloo saco interno.
1. El exofalo: es del tipo «cucujoi-deo» y similar al de otros Phyto-phaga. Es de morfología variable,presentando características propiasen cada especie. Es de naturalezamembranosa, poco quitinizado ycasi nulamente pigmentado. Está
formado por varias partes cuya de-nominación genérica, promulgadapor SNODGRASS, ha sido estable-cida para el caso especial de estafamilia otra denominación análogapor KINGSOLVER que establece dos
partes fundamentales: tegmen y ló-bulo mediano.
El tegmen está formado por tresregiones fundidas en una sola pieza(Fig. 11): los lóbulos laterales (pa-rameros o estilos) que están ligadosventralmente y rodeando al lóbulomediano en forma de vaina y antesde llegar al ápice se bifurcan en doslóbulos simétricos de forma aplas-tada o filiforme. En el ápice se si-túan sedas o papilas de disposiciónvariada y cuya misión es de tiposensorial. La vaina producto de lasoldadura de los dos lóbulos latera-les puede progresar excepcional-mente hacia el ápice formando unapieza única e inferiormente vuelvea bifurcarse en dos ramas for-mando un anillo denominado piezabasal, la cual se prolonga poste-riormente en una sola pieza situadapor encima del lóbulo mediano yque recibe el nombre de columnategminal, que junto con la piezabasal forman importantes puntosde inserción de los músculos re-tractores y protractores del lóbulomediano.
El lóbulo mediano es una estruc-tura tubular que sirve de protec-ción y alojamiento del endofalo ymúsculos motores del mismo du-rante la cópula. Presenta un orifi-cio basal de entrada del canal eya-culador (Fig. 11). En los génerosprimitivos (Pachymerus, Caryobo-

rus) esta pieza basal es una piezasimple, similar al de otros Phyto-phaga, pero en la mayor parte de losBrúquidos, el ápice del lóbulo me-diano lleva uno o dos escleritos amodo de valvas protectoras deno-minadas valva dorsal y ventral. Aexcepción de algunos géneros, laválvula dorsal está ausente y en sulugar hay frecuentemente una es-tructura membranosa a modo devaina que sobresale del orificioapical. La vaina basal se conectacon la columna tegminal medianteunas fibras musculares, formandocon ella una bomba auxiliar cuyamisión es la de distender el saco in-terno por contracciones de dichabomba e insuflación de hemolinfa.
2. El endofalo o saco interno consti-tuye la única estructura que pene-tra en el conducto vaginal de lahembra durante la cópula (Fig. 11).Una vez evertido, puede apreciarseuna superficie externa de aparien-cia rugosa, debido a la disposiciónde numerosos escleritos de diver-sos tipos y cuya misión es, al pare-cer, la de retención del saco du-rante la cópula. Esta gran variedadde tipos de escleritos, ha permitidodilucidar algunos problemas taxo-nómicos, ya que su disposición yestructura es característica de cadaespecie. Pueden ser espinas, den-tículos, espículas, etc.
Órgano genital de la hembra
Las hembras presentan dos ovarios dis-puestos uno a cada lado del tramo final delabdomen. Cada ovario presenta 6 ovariolas oramas ovígeras terminadas en un filamento(Fig. 12). Todas las ramas desembocan en un
canal u oviducto que, junto con el otro par,se reúnen en la vagina, sobre la cual se sitúael saco glandular y la espermateca, comuni-cando con la vagina por un tubo que en suparte distal queda protegido por la armaduragenital.
La armadura genital es un tubo constituidopor el X segmento abdominal. Este presentauna parte ventral o urosternito a modo devalvas cuyo ápice se adelgaza en dos estilosprovistos de sedas sensoriales (Fig. 12) y unaparte dorsal o uroterguito de naturalezamembranosa y provisto también en su ápicede sedas de misión sensorial. Al igual que enlos machos, el XI urosternito se encuentramodificado en forma de espícula gastral cuyamisión sea similar a la del macho, esto es, lade anclaje de la armadura.
Dimorfismo sexual
Aparentemente, la mayor parte de los Brú-quidos no presentan un dimorfismo sexualacusado, pero un examen más detallado nosrevela caracteres sexuales secundarios en lossiguientes órganos:
a) Antenas: generalmente más desarrolla-das en los ó ó, por cuanto respecta a lalongitud y desarrollo de los artejos(Fig. 13).
b) Patas: donde se aprecian diferenciasque afectan al desarrollo de las tibias,donde aparecen espolones y espinas dediversas conformaciones, así como di-lataciones, etc., en los <3ó (Fig. 13).
c) Élitros: donde aparecen diferencias encuanto a la forma. Así, en los Só ellado externo aparece bruscamente es-trechado en su mitad posterior, debajodel calus humeral, mientras en las 9 9no se aprecia este cambio (Fig. 13).
d) Ojos: más desarrollados en los ó ó, porlo que el espacio interocular aparecemás estrecho que en las 9 9 (Fig- 13).

Fig. 12.—A: Complejo de genitalia de hembra deBruchus pisorum L.
1.—Rama ovígera2.—Ovario3.—Oviducto4.—Canal deferente5. —Espermateca6.—Saco glandular
7.—Vagina8.—Armadura genital9.—Espícula gastral
B: Armadura genital de hembra de Bruchus pisorum L.
1.—Estilos internos del X uroterguito2.—X urosternito3.—X uroterguito
e) Pigidio y IX urosternito: en los óó elpigidio es más convexo y de ápice pun-tiagudo que en las 99- P° r o t r a parteel IX urosternito aparece escotado ensu margen inferior mediano en los ó ó,mientras que en las 9 9 n o aparece y esrecto (Fig. 13).
f) Coloración: afectando distinta colora-ción en las patas y artejos de las ante-nas. También hay diferencias en el di-bujo elitral que forma la pubescencia,etcétera.
Régimen alimenticio
Los Brúquidos constituyen una familia derégimen alimenticio fitófago predominante-mente cletrofágico, esto es, devorador desemillas. Sin embargo, este carácter, por serpropio de la larva, nos conduce a distinguirdos categorías alimenticias a lo largo de lavida del insecto, ligadas a su función bioló-gica:
a) Alimentación de la larva: ligada prefe-rentemente a la conservación del indi-

Fig. 13.—A: Antenas izquierdas de Bruchidius jocosus Schónh; B: Patas mesotorácicas de Bruchus atomarius L;C: Abdomen de Bruchidius perparvulus Boh; D: Élitros de Callo sobruchus chinensis L; E: Cabezas de Bruchi-dius tuberculatus Hochh.
viduo, esto es, a asegurar la evoluciónhasta el estado adulto. En este estadoel insecto se alimenta principalmentede semillas y en raras excepciones sealimenta de otra parte de la planta,
b) Alimentación del adulto: ligada prefe-rentemente a la conservación de la es-pecie, esto es, asegurar la reproduc-ción. A este respecto, los trabajos deY. DE LUCA (1952) demuestran que elgrado de madurez de las gónadas de-pende del grado de ingesta de granosde polen. En este estado el animaltiene una longevidad corta, dirigida tansólo al acoplamiento y la reproducción,alimentándose de granos de polen, néc-tar de la flor y tan solo ocasionalmente,
devora las partes tiernas de la plantahuésped de la larva (parénquima foliar,sépalos).
Según los datos de que se dispone hoy día,los Brúquidos han sido señalados en 52 fami-lias de vegetales, entendiéndose que de éstassólo unas pocas son huéspedes de las larvas yel resto han sido señaladas bien en las flores obien en la corteza y frondosidad de las plan-tas donde el adulto busca el abrigo contra lasinclemencias del invierno.
De las 800 especies de Brúquidos que seconocen, 300 han sido señalados sobre Papi-lionáceas, 200 sobre otras familias de plantas,principalmente Dicotiledóneas (95 %) y elresto (300) se desconoce la planta que fre-cuenta o parásita.

Los trabajos de Y. DE LUCA (1952) orien-tados a buscar algún patrón común que liguelas plantas ante la selección de sus parásitos,fueron infructuosos, llegando a la conclusiónde que, dada la escasez de datos bioquímicosde la gran mayoría de las especies huéspedes,hoy día es imposible determinar conclusionessobre la selección de la planta huésped. Noobstante, parece ser que hay sustancias comoel Pinitol que se encuentra en algunas plantasde abrigo (Gymnospermas: Coniferas, Cupre-sáceas, y Angiospermas: Fagáceas, Salicá-ceas) y que también está presente en la fami-lia electiva de estos insectos, es decir, las Pa-pilionáceas, pero el hecho de que esta sustan-cia no se encuentre en otras plantas electivas,descarta este factor como determinante gene-ral de la selección de la planta huésped. Sinembargo, sí parecen ser determinantes las va-riaciones en la tasa de lípidos de las semillas,y la coincidencia de la época de floración delas plantas, así como el valor cuantitativo ycualitativo del grano de polen.
En general, podemos considerar que losBrúquidos presentan un régimen alimenticiooligofágico, es decir, con un espectro alimen-ticio que abarca algunas especies, general-mente de un mismo género perteneciente a lafamilia Papilionaceae. No obstante, existencasos de monofagia, como B. pisorum L., queataca casi exclusivamente a los guisantes, ytambién existen especies polífagas que abar-can un número considerable de géneros deLeguminosas y en determinados casos puedenllegar a invadir otras semillas de distintogrupo botánico, como Gramíneas. Entre estasespecies encontramos las más perjudiciales ala agricultura: Cali, chinensis L., B. rufima-nus Boh., etc. Otras especies varían su es-pectro alimenticio según las circunstanciasambientales, tal es el caso dzAcanth. obsole-tas Say. cuyo comportamiento alimentario seve bastante afectado en circunstancias expe-
rimentales, de tal modo que de su régimennatural de tipo monofágico pasa a una polifa-gia de amplio espectro.
Excepcionalmente encontramos especiescuya especialización no ha llegado a la elec-ción de la familia Papilionaeceae como plantahuésped. Tales son los casos de especies deSpermophagus que se desarrollan en semillasde Convolvulaceae; en Brasil, Caryoborusbractis se desarrolla en Palmáceas como Co-pernica cerífera y en nuestra región encon-tramos especies como Br. biguttatus 01. y Br.cinerascens Gyll. que se desarrollan en Um-belíferas.
Ciclo biológico
Los Brúquidos, merced a una diferencia-ción del comportamiento en sus diversos es-tadios larvarios, presentan un desarrollo en elcual acusan diferencias morfológicas que enconjunto se denomina hipermetamorfosis, fe-nómeno que encontramos en otros grupos deColeópteros tales como Carábidos, Staphilí-nidos, Meloideos, etc. Básicamente presen-tan dos tipos de larva a lo largo de su desarro-llo ontogénico:
a) Larva primaria: llamada también neo-nata o crisomeloide, representaría eleslabón que los une a los Crisomélidos,por cuanto se refiere a la presencia depatas ante la exigencia del desplaza-miento para la búsqueda del alimento.
b) Larva secundaria: llamada tambiénmelolontoide o curculionoide, repre-sentaría el eslabón que filogenética-mente los une a los Curculiónidos porcuanto se refiere a la ausencia de patasfuncionales, ante una vida endofítica,fundamentalmente cletrofágica, dondeel movimiento es casi nulo por tener elalimento en su inmediato alcance.

La hipermetamorfosis, si bien es común amuchas especies de Brúquidos, está más bienpresente en aquellas especies filogenética-mente más próximas a los Crisomélidos; perohay otras especies que, merced a su cursoevolutivo, han sufrido un proceso de acelera-ción de la ontogenia o taquigénesis, por lacual la larva neonata es ya ápoda, ya que enrealidad las patas son órganos en regresión enesta familia pues carecen de función. Talesson los casos de las especies de Spermopha-gus, B. signaticornis, Br. perparvulus, etc.
Desde el punto de vista del ciclo biológico,encontramos dos grandes grupos en estos in-sectos:
a) Monovoltinos: especies que presentanuna sola generación al año, muy rara-mente dos generaciones. Sus larvasson incapaces de desarrollarse en gra-nos secos y por lo general sólo se de-sarrolla una larva por semilla. Lapuesta se realiza exclusivamente en elcampo y el ciclo dura por término me-dio unos 50 días, cifra que varía segúnla especie y las condiciones ecológicas.El adulto pasa por un largo período deinactividad durante el invierno. Son loscasos de la mayor parte de las especiesde nuestra región y muchas de ampliadispersión, tales como B. pisorum L.,B. rufimanus Boh., B. brachialisFahrs., B. signaticornis Gyll., B. lentisFról., etc.
b) Polivoltinos: especies que presentanmás de dos generaciones al año, gene-ralmente de tres a cuatro generacionesen circunstancias ecológicas propicias,cifra que varía según la temperatura yhumedad del granero, así como la tasade agua de las semillas. A este grupopertenecen especies de origen exótico,frecuentemente tropical, como Acanth.obsoletas Say., Call, chinensis, L.,
Cali, maculatus F., Zabr. subfasciatusBoh., etc. La puesta se realiza primeroen el campo y luego, al poder desarro-llarse la larva en semillas secas alma-cenadas, la realizan sobre los granossecos. Estas generaciones se sucedenininterrumpidamente, por lo cual, eladulto, al abrigo de los graneros, notiene que invernar. A este grupo perte-necen pues, las especies más perjudicia-les para nuestra agricultura.
Podemos abordar la etología de estos insec-tos en los siguientes apartados:
1.—Puesta
Al llegar la estación propicia, generalmenteprimavera avanzada en las especiesmonovolti-nas, una vez verificado el acoplamiento, lahembra busca la planta huésped para realizarla puesta. El período de puesta depende fun-damentalmente del clima y la fenología de laplanta huésped. En las especies polivoltinas,existe una generación que realiza la puesta enla naturaleza, en los cultivos, pero en sucesi-vas generaciones la realizará en los graneros,directamente sobre los granos almacenados.Durante la época de puesta existe un máximode actividad de ovoposición que se mantienedurante pocos días y luego baja rápidamentemanteniéndose durante pocos días a un nivelconstante hasta desaparecer. Esta actividadpuede ser interrumpida por factores externos,principalmente de tipo ecológico, lo que haceque la eclosión ño sea sincrónica en la mayorparte de las veces.
Durante la puesta, la hembra se dirige a losfrutos tempranos de la planta huésped, si-tuándose cerca del cáliz y explorando, me-diante sucesivos contactos de antenas y pal-pos, la superficie del fruto. Esta exploraciónva encaminada a la búsqueda del lugar propi-cio para la puesta, lo suficientemente cercanoal lugar donde se encuentra la semilla y, por

otro lado, para comprobar si el fruto es vir-gen, es decir, si no hay efectuada ya unapuesta. Caso de haber una puesta, general-mente abandonan la vaina y se dirigen a otrofruto donde no haya ninguna puesta. El lugarde la puesta varía según el estado fenológicode la planta, así encontramos que en los fru-tos tempranos se realiza cerca del cáliz, pró-ximo al lugar de inserción de la semilla y enfrutos maduros y dehiscentes, directamentesobre la semilla a través de la hendidura dedehiscencia. Las especies polivoltinas hacenuna puesta con menos cuidados, frecuente-mente poniendo paquetes de 2-30 huevos, di-rectamente sobre los granos almacenados obien en las paredes del recipiente, próximos aéstos. Hay especies monovoltinas, como B.pisorum L., que hacen una puesta numerosaen cada fruto, pero como veremos más ade-lante, sólo se desarrollará una larva por gra-no. Este fenómeno puede ocurrir cuando sonescasos los frutos ya que la hembra suele op-tar por una puesta numerosa antes que dejarlos huevos sobre un soporte cuya distanciarespecto de la semilla impedirá el alcance dela larva neonata. En condiciones experimen-tales, donde no hay frutos para la puesta, éstase realiza sobre diversos soportes tales comotallos, hojas, paredes del recipiente, etcétera,pero preferentemente sobre superficies rugo-sas, ya que en superficies lisas el huevo nopuede adherirse bien.
En el momento de la puesta, la hembradeja caer una gota de líquido hialino, viscoso,que al contacto con el aire se seca rápidamen-te. Inmediatamente después de dejar caer di-cho líquido, pone un huevo con una orienta-ción (ya realizada en los conductos genitales)constante: la cara más plana sobre el sustra-to. Es importante para el insecto que elhuevo esté bien adherido por dos razones: deuna parte, se impide que los agentes exterio-res, como el viento y la lluvia, lo desprendan y
por tanto lo alejen de la planta huésped y porotra parte se asegura la penetración de lalarva neonata en el fruto.
Al parecer, en la elección de la plantahuésped intervienen tanto factores externoscomo factores propios de la especie, funda-mentalmente ciertos tropismos. Así se haconstatado que, entre diversas variedades deuna misma planta y en un mismo cultivo, lahembra suele elegir una variedad determina-da, posiblemente en función de un quimiotro-pismo más o menos selectivo, según las espe-cies. Tal es así, que hay variedades de gui-santes denominados «resistentes» que apenasconsiguen el reclamo de B. pisorum L. Porotra parte, la hembra suele hacer la puesta enaquellas variedades de estado fonológico másavanzado, donde la semilla es más visiblemerced a la dehiscencia.
2.—Evolución del huevo y eclosión
La hembra de los Brúquidos suele haceruna puesta numerosa, generalmente de varioscentenares de huevos. Una de las posiblescausas de tan elevado número de huevesdebe ser su alto índice de mortalidad. Enefecto, un gran número de huevos no termi-nan su evolución, ya sea por factores exter-nos: roces de mamíferos con las plantas, pre-dación, parasitismo, etc., y otros propios dela puesta: el huevo queda poco adherido alsustrato e impide la emergencia de la larva alperder el apoyo del corion.
El período de evolución del huevo o incu-bación es bastante rápido en condiciones cli-máticas óptimas, por término medio dura unadecena de días. Sin embargo, esta cifra, ca-racterística de cada especie, puede alterarsevariando desde muy pocos días, merced a unincremento de humedad y temperatura am-bientales, a más de 15 días en condicionesmuy frías o secas.
Se puede seguir a «grosso modo» la evolu-

ción del huevo exteriormente. El primer díael huevo aparece como una bolsa hialina ytransparente, luego pasa a ser translúcido yno transparente y ya en los últimos díaspuede apreciarse por transparencia el em-brión, sobre todo la cabeza, que aparece "másoscura por su quitinización.
En su último estadio embrionario, la lár-vula se encuentra con el dorso en contactocon la parte más plana del huevo, esto es, encontacto con el sustrato (Fig. 14), con la ca-beza dirigida hacia la parte más abombada delhuevo, ligeramente retraída sobre el protó-rax, dejando vacía aproximadamente 1/5parte del huevo, precisamente detrás de lacabeza. En esta posición las piezas bucales
no están en contacto con el sustrato, por loque a continuación la lárvula va a efectuar ungiro lateral de 180° que la pondrá ventral-mente en contacto con dicho sustrato y en-tonces estará en condiciones de penetrar através del fruto y llegar a la semilla. Para ello,contrae y luego extiende el cuerpo, con loque consigue que la placa pronotal tome an-claje en las paredes del corion, al cual sueledañar en su parte más delgada. A continua-ción, ayudándose de las patas, labios anales yespinas abdominales, gira poco a poco hastaquedar en posición lateral (Fig. 14), que-dando inmóvil en esta posición durante algúntiempo. Por último, y merced a los anclajes ypalancas efectuadas sobre las paredes del co-
Fig. 14.—A-F: Diversas fases del proceso de penetración de la larva neonata de Bruchus ulicis Muís.

non mediante mandíbulas, placa pronotal, pa-tas, etc., y contracciones del cuerpo, consiguepor fin llegar a la posición correcta, es decir,ventralmente, en contacto con el sustrato(Fig. 14).
En esta posición, la lárvula inicia la pene-tración a través de las paredes del corion yfruto para llegar a la semilla. Para esta opera-ción, la lárvula toma apoyo en el polo poste-rior del abdomen; luego, apoyándose con pa-tas y mandíbulas, se contrae vigorosamente,quedando la placa pronotal con la pared dor-sal del corion y las mandíbulas perpendicu-larmente a la pared inferior o delgada delmismo (Fig. 14). En esta postura, la larvacomienza a taladrar el corion, consiguiendopracticar un agujero por donde meterá pri-mero la cabeza (Fig. 14) y luego, gracias alanclaje de la placa pronotal sobre el borde dedicho agujero, consigue meter el tórax trashaber conseguido profundizar el taladraje. Deaquí se desprende la necesidad de que elhuevo esté bien adherido al sustrato, pues delo contrario, aunque se verifique la eclosión yla lárvula se desplaze hasta la semilla, al ca-recer de punto de apoyo, muere en pocas ho-ras. En algunas especies polivoltinas comoAcanth. obseletus Say., la larva neonata,muy móvil, soslaya este problema al encon-trarse con los granos directamente en el gra-nero y tener opción y elegir aquellos que yatengan algún agujero o hendidura provocadospor sus congéneres o bien accidentalmente.Durante el taladraje, las limaduras son dirigi-das hacia atrás y son diminutas esférulas quepueden ser confundidas con excretas del in-secto. Estas limaduras pasan primero por laparte ventral, movidas por las patas, luego,mediante contracciones del cuerpo sobre lasparedes de la galería, pasan por los lados ypor último salen al exterior dorsalmente que-dando amontonados en la parte posterior delhuevo (Fig. 14).
3.—Biología de la larva
La duración del desarrollo larvario varíasegún la especie y las condiciones ecológicas,tanto del propio sustrato como ambientales.Como se ha mencionado en las especies mo-novoltinas, al no poder desarrollarse en gra-nos secos de los almacenes, han de hacerfrente a unas condiciones menos fluctuantesque en las especies polivoltinas que están amerced de las condiciones ambientales delgranero. Así, se ha visto que para la especiepolivoltina Cali, maculatus F. la duraciónmedia del desarrollo larvario es de 17-22 días,pero esta cifra varía grandemente según lascircunstancias desde 9 días a 8 meses. Sinembargo, la variabilidad del período de desa-rrollo larvario es más pequeña en las especiesmonovoltinas y por regla general la duraciónes de 15-30 días.
Durante este período, la larva sufre una se-rie de mudas cuyo número depende de la es-pecie, si presenta o no hipermetamorfosis yque por término medio suelen ser 3 mudas.Según Y. DE LUCA (1952) en B. lentis Fról.existen 5 mudas. Las especies monovoltinasque atacan a semillas que luego se almacena-rán, no pueden continuar el ciclo ya que elgrano se seca y en estas condiciones la larvaes incapaz de proseguir el desarrollo.
Una vez conseguida la penetración total dela larva en la semilla, ésta sufre una mudaoriginando una larva secundaria con regre-sión de los caracteres crisomeloides (patas,placa pronotal). Esta larva continúa la pene-tración en la semilla a continuación de la pe-queña galería, que va aumentando de diáme-tro conforme ésta aumenta de tamaño, en lassucesivas mudas. Esta progresión siempre eshacia el interior, y en ningún caso la larva re-trocede o abandona la galería.
La galería de entrada suele ser corta, noafectando a los cotiledones, cilindrica y algo

arqueada, bordeada de detritos. La muda severifica del siguiente modo: aparece una inci-sión dorsal mediana longitudinal. Esta inci-sión se va abriendo por la presión del cuerpode la nueva larva. A esta incisión le sucedeotra en la cápsula cefálica, la cual se abre porla frente a lo largo de las suturas en tres par-tes. La larva se contorsiona y va saliendoapicalmente, replegando la exuvia haciaatrás, quedando al final pegada al extremodorsal posterior del abdomen. Tras la muda, lalarva permanece un corto tiempo inmoviliza-da. Luego continúa su actividad y sigue li-mando las paredes del albumen lateralmente,de lado a lado, al tiempo que compacta lasparedes con una secreción bucal. En este es-tado, la larva empieza a devorar los cotiledo-nes y la cámara va ensanchándose. Al finaldel desarrollo de la larva, nos encontramostantas celdas como mudas haya tenido la lar-va. Estas celdas van aumentando de tamañoprogresivamente y la última celda suele abar-car a una o dos precedentes. Poco antes deentrar en la ninfosis, la larva prepara la celdaninfal, para lo cual alisa las paredes de la úl-tima cámara y lima la pared dorsal de lamisma hasta llegar a la cutícula de la semilla,la cual respeta y deja a modo de opérculo pordonde saldrá el imago sin esfuerzo alguno,con tan solo una ligera presión. Hay especiestropicales como Pachym. accaciae Byll. cuyalarya adulta sale de la semilla y a continua-ción entra en ninfosis.
Un hecho importante desde el punto devista ecológico, por cuanto se refiere a lacompetencia intraespecífica, consiste en elnúmero de larvas que atacan a una mismasemilla. Como habíamos apuntado anterior-mente, en ciertas condiciones, la hembrapuede poner numerosos huevos en unamisma vaina. Sin embargo, a la postre nosencontramos que en la mayor parte de las es-pecies monovoltinas tan sólo se desarrolla
una larva por cada semilla. Este fenómeno sedebe a una competencia trófica ligada al ta-maño de la semilla. En efecto, puede ocurrirque en una misma semilla entren varias larvas,pero al llegar a la 2.a muda la larva que hayamudado antes, se encontrará que su galeríainvade a otras practicadas por otras larvasque aún no han mudado. Entonces, en suprogresión las mata, prevaleciendo al términodel desarrollo larvario nada más que una lar-va. En cambio, tanto en las especies polivol-tinas como en las monovoltinas que atacan asemillas de gran tamaño (Ej. B. rufimanusBoh. en habas), se pueden desarrollar unamedia de 2-3 larvas por semilla y en ciertasespecies este número puede llegar a ser con-siderable, como ocurre con Acanth. obsoletasSay., cuyas larvas llegan a invadir en bloquela semilla hasta agotar por completo todo elalbumen de la misma.
La ninfosis aparece en el último estadiolarvario en que la larva se extiende en lacelda ninfal y adopta una conformación depreninfa, quedando inmóvil, y en el curso deuna media de 10 días, la larva va aumentandode tamaño, sufriendo una serie de transfor-maciones que en última instancia darán lugaral imago. En algunas especies monovoltinas,el imago saldrá al exterior y tras cortos reco-rridos buscará un refugio para pasar el in-vierno, pero en otras especies el imago quedaen el interior de la celda ninfal y entra en dia-pausia invernal, no saliendo del mismo hastala primavera siguiente. Esto sucede cuandolas vainas son indehiscentes.
4 Biología del imago
Una vez formado el imago, el opérculocoincide estar suficientemente reseco, por loque es bastante fácil empujarlo con la cabezay abrirlo. La salida del imago está favorecida

por el incremento de la temperatura y el des-censo de la humedad ambiental, así como elhecho de que las vainas sean dehiscentes,pues caso de no serlo el adulto permaneceráen el interior hasta la primavera siguiente.
Una vez fuera, el imago es guiado por unaserie de tropismos que le conducen bien abuscar un refugio para pasar el invierno obien, caso de llegar la primavera, a buscar laplanta huésped, acoplarse y hacer la puesta.La longevidad suele ser corta, pero este pe-ríodo se modifica en función de los factoresclimatológicos y la alimentación.
Experimentalmente, hemos podido com-probar que el imago presenta un fototropismopositivo y un geotropismo negativo, por loscuales el insecto se dirige hacia la proceden-cia de la luz y siempre hacia arriba. Pensa-mos que estos tropismos favorecen la salidadel insecto de la vaina y le mueven a dirigirsehacia partes aéreas de la planta. Asimismo,favorece la salida del adulto su manifiestotermotropismo por el cual, al aumentar latemperatura, no sólo eclosionan más adultos,sino que éstos manifiestan una mayor activi-dad, que se traduce también en una puestamás abundante y un desplazamiento más con-tinuado. A este respecto, cada especie tieneunos límites de actividad en función de latemperatura, teniendo un óptimo por el cualdesarrollan su máxima actividad. Ligado altermotropismo, se encuentra un higrotropis-mo, por el cual un ambiente muy húmedo,como puede ser el resultante de una prima-vera muy lluviosa, favorece la emergenciamasiva de estos adultos.
También hemos podido comprobar expe-rimentalmente que el imago de los Brúquidospresenta un claro tigmo-tropismo, por el cualsuelen preferir las superficies rugosas. Estopuede estar en función de la búsqueda de re-fugios y abrigos invernales. Resulta curiosoobservar que nada más salir de la celda nin-
fal, los adultos permanecen inmóviles y agru-pados unos junto a otros en las proximidadesdel opérculo.
No podemos pasar por alto el manifiestoquimiotropismo de estos insectos, uno de loscaracteres más importantes para conseguir eléxito de la reproducción en la identificaciónde la planta huésped, carácter común a otrosfitófagos. Este quimiotropismo es altamenteselectivo para algunas especies, sobre todopara las especies monófagas y oligófagas. Apesar de la importancia de este carácter en laselección de la planta huésped, son numero-sas las lagunas que poseemos en el estado ac-tual de nuestros conocimientos. Se sabe quemuchas especies tienen apetencia por el pini-tol, alcaloide que se encuentra en las Legu-minosas y en ciertas plantas de abrigo, peroresulta insuficiente para dilucidar los factoresquímicos que atraen a estos insectos.
Otro fenómeno común a sus parientes fitó-fagos es el de simulación del estado demuerte o tanatosis cuando es estimulado bienpor cambios bruscos de luz o bien por unavibración poco corriente. Inmediatamentedespués de haber sido estimulado y cuandoeste estímulo es débil, el insecto suele cam-biar de postura y se esconde detrás de lasramas. Si este estímulo persiste o es másfuerte, el insecto se retrae y cae libremente alsuelo, en ocasiones queda retenido por el fo-llaje, donde permanecen en una postura in-móvil que recuerda a la del estado de muerte.Este queda generalmente reposando sobre sudorso, con la cabeza replegada sobre el tóraxy con las antenas reposando sobre la articula-ción tibio-femoral de las patas anteriorescuando éstas son cortas y a lo largo del vien-tre cuando son largas; el tórax también sedobla hacia adelante, el 1.° y 2.° par de patasreplegadas, con tibias y fémures juntos y tar-sos dirigidos hacia abajo, divergentes y repo-sando sobre el fémur siguiente; 3.er par de pa-

tas con fémur y tibia algo separados y tarsosdirigidos hacia abajo convergentes. Es decir,una postura muy parecida a la de la ninfa,pero diferenciándose del verdadero estadomortal en que tanto cabeza como tórax no es-tán tan replegados y las patas tienen una dis-posición irregular, a menudo dirigidas haciaarriba, con los tarsos entrecruzados. Estainmovilidad es poco duradera y la estimula-ción de ciertos órganos, como antenas, pal-pos y abdomen, devuelven la actividad alanimal. Por otra parte, el haber repetido mu-chas veces la tanatosis puede impedir, por unfenómeno de aprendizaje, que el animalvuelva a acusar dicho estado.
Durante el invierno, el imago permaneceinmóvil, según algunos autores en diapausiainvernal en sus refugios o abrigos. Al llegarlos primeros días soleados del año, el animalacusa algún movimiento pero no se decide asalir hasta llegada la primavera, de tal modoque conforme va aumentando la temperatura,el número de adultos va aumentando en elcampo.
Durante el invierno, en su inmovilidad, sealimenta de las reservas energéticas y algúnque otro alimento ocasional que ingieren enlos días soleados. A la llegada de la primave-ra, al incrementar su actividad y estar agota-das sus reservas, necesitan alimentarse. Estaalimentación es fundamentalmente a base degranos de polen, lo cual favorece la madura-ción de las gónadas, y en ocasiones mordis-quean otros órganos de la planta, fundamen-talmente el parénquima foliar.
Llegada la maduración de las gónadas, severifica el acoplamiento. Este suele durar al-rededor de 10-15 minutos, y para ello el ma-cho se sitúa sobre el dorso de la hembra sinmás ceremonias, a continuación palpa consus antenas la frente de la hembra, al tiempoque con las patas anteriores mantiene reple-gadas las antenas de la hembra. Al parecer
esto provoca una disminución considerablede la agresividad de la hembra y es curiosoque en estos contactos no se verifica la tana-tosis por parte de la hembra. Mediante el 2.°y 3.° par de patas, el macho se agarra a laspleuras de la hembra. En nuestra opinión, esmuy probable que la existencia de espolonesy horquillas en la extremidad de las tibiasmesotorácicas en muchos machos en Bruchuspuede estar en relación con esa función copu-ladora. En esta posición el macho saca los es-tilos del tegmen y pene con los cuales esti-mula a la hembra en la abertura anal-genitalhasta provocar la apertura del pigidio porparte de la hembra. Por último, tras ponerseen contacto sendas armaduras genitales, elendofalo es evaginado merced a la presión dela hemolinfa y se introduce en los conductosgenitales de la hembra, tras lo cual inyectalos espermatozoides, que quedan almacena-dos en las espermateca de la hembra. Tras lacópula, la hembra aguarda un período nece-sario para la fecundación y comienza a reali-zar la puesta.
Consideraciones sobre la Ecología
a) Influencia del clima
El factor climático afecta tanto directacomo indirectamente a la dinámica de pobla-ciones de los brúquidos. En efecto, tanto elviento como la temperatura y humedad de laregión influyen en el desarrollo de la plantahuésped y, por tanto, también influye en elporcentaje de eclosión de brúquidos. Todaespecie tiene una temperatura óptima dondese desarrolla su máxima actividad, que estáincluida en un más o menos amplio margende temperatura favorable y unos límites tantopor debajo como por encima de esta zona fa-vorable, a partir de las cuales su actividad va

disminuyendo considerablemente hasta pro-vocarle la muerte. La temperatura y humedadambientales influyen en la duración del ciclo,en el vuelo o dispersión de los adultos, en laabundancia de la puesta, en el índice de mor-talidad, en la longevidad del adulto, en laeclosión, en la duración de la ninfosis, etc. Aeste respecto, se sabe que en ambientes muysecos el ciclo se alarga considerablemente, elvuelo de los adultos es más pesado llegando aser casi nulo, la puesta es escasa, el adultopuede llegar a morir antes de la reproducción,los huevos no eclosionan o se secan, etc.
b) Influencia del terreno
Se ha podido comprobar que la textura ycomposición de la tierra, al influir en la cali-dad y desarrollo de la planta, influye tambiénen el porcentaje de brúquidos que atacan a laplanta. En efecto, los terrenos de grano blan-do, de naturaleza calcárea, al guardar más lahumedad, dan plantas de mayor desarrollo,cosa que no ocurre con los terrenos duros, denaturaleza silícea, que guardan menos la hu-medad.
c) Influencia de la situación de la planta
Está directamente relacionada con la in-fluencia del clima. En efecto, las plantas ex-puestas hacia el sur tienen una precocidad ydesarrollo mayores y el insecto tiende a para-sitar las plantas más desarrolladas. Asimismola altitud también influye en la dinámica depoblaciones de estos insectos, puesto que lasplantas situadas más altamente tienen unafloración más temprana, anticipándose a laemergencia de adultos, por lo que el ataquede éstos se reduce considerablemente.
Relaciones interespecíficas
En este capítulo vamos a estudiar princi-palmente a los fenómenos de parasitismo ydepredación dirigidos contra las poblacionesde Brúquidos.
a) Parasitismo
Se puede decir que todos los Brúquidospresentan una o varias especies de parásitos.Estos son himenópteros endoparásitos ento-mófagos, pertenecientes a los subórdenesChalcidoidea e Ichneumonoidea, preferente-mente las familias Chalcididae y Braconidaerespectivamente. Han sido señalados multi-tud de parásitos de Brúquidos, y algunos au-tores como BRIDWELL e Y. DE LUCA hanefectuado una gran labor recopiladora de to-dos estos datos.
Hay especies e incluso géneros de acciónespecífica sobre Brúquidos. Sin embargo, hansido señalados multitud de parásitos polífagosy otros que, con toda seguridad, no son pará-sitos específicos de Brúquidos, sino de Mi-cro lepidópteros, Curculiónidos, Cynípedos yCecidómidos, lo cual puede ser debido a quefueron obtenidos en frascos de cría proceden-tes de otros fitófagos de legumbres.
El parasitismo comienza con una explora-ción de la hembra del parásito en las vainasde leguminosas, donde comienza a palpar conlas antenas la superficie y en cuanto detectala presencia de un huevo, la hembra sevuelve e introduce su oviscapto a través delcorion y deposita un huevo. La larva del pa-rásito se desarrolla en el interior del cuerpodel embrión y se alimenta posteriormente delos tejidos de la larva y la ninfa hasta provo-carle la muerte en el último estadio larvario obien en la ninfa. El adulto ya evolucionado,sale del cuerpo de la ninfa y abandona la

celda ninfal por el opérculo. Otras veces hade atravesar las paredes de la cámara ninfalpor no haber sido preparado aún el opérculo.Para que el parásito salga al exterior es pre-ciso que la vaina sea dehiscente, aunque enocasiones puede llegar a atravesar las paredesde la vaina, pero en caso de que las vainas nose abran, generalmente" no podrán salir ymueren dentro.
Estos parásitos aparecen en un porcentajemuy pequeño en la población de Brúquidos,en parte debido a la tasa de mortalidad a queéstos están sometidos, ya que unas vecesmueren al no poder salir de los frutos y otrasveces son a su vez parasitados (fenómenodenominado hiperparasitismo) por otros hi-menópteros, sobre todo pertenecientes al su-borden Chalcidoideos, preferentemente lasfamilias Eupelmidae y Eurytomidae, que ha-cen la puesta posteriormente a la del parásito,de forma tal que su larva se desarrolla a ex-pensas de las otras dos.
b) Depredación
Además del parasitismo, los Brúquidos es-tán sometidos a la presión de depredadoreshimenópteros pertenecientes a los subórde-nes Vespoidea, preferentemente la familiaBethylidae, y Sphecoidea, preferentemente lafamilia Cerceridae. Estos insectos atacan alas larvas de Brúquidos, devorando sus hue-vos o la larva neonata. También verifican unaimportante acción depredadora ácaros perte-necientes a las familias Anystidae y Pyemoti-dae que devoran los huevos y hasta incluso lalarva neonata.
Importancia económica
Como hemos hecho constatar a través delpresente trabajo, los Brúquidos presentan un
régimen alimenticio cletrofágico, esto es, de-vorador de semillas, fundamentalmente deLeguminosas.
Las Leguminosas constituyen uno de loscultivos hortícolas más importantes para lamayoría de los países, sobre todo para aque-llos que están en vías de desarrollo. Una granproporción de estos cultivos van dirigidos a laalimentación humana (judías, garbanzos, len-tejas, habichuelas, guisantes, etc.) y ciertoporcentaje va dirigido a la industria química yfarmacéutica. Una de las propiedades de lasLeguminosas es que pueden utilizarse variosórganos (hojas, raíz, semillas) sin necesidadde un tratamiento industrial, tienen un altonivel energético y cierto contenido vitamíni-co. Hoy día, los países más avanzados tien-den a sustituir el cultivo de variedades muyenergéticas y de bajo contenido vitamínicopor aquellas de alto contenido en vitaminaspero de bajo nivel energético. Su cultivo, sibien puede adaptarse hasta ciertas altitudes,necesita bastante agua y sol, por lo que suelehacerse en países de clima templado o cálido.Una proporción del producto local va dirigidaal autoconsumo y otra va dirigida a su co-mercialización. En este último caso, los cul-tivos pueden ser extensivos para especies queno necesitan de grandes cuidados, e intensi-vos, en los que ha de invertirse buena canti-dad de fertilizantes y antiparasitarios. Contodo, su rendimiento es elevado, pudiéndoseconseguir varias cosechas al año, y hoy día lamecanización y la mejora técnica de los culti-vos ha provocado una disminución del costoy un aumento del consumo.
Las Leguminosas constituyen uno de losprincipales productos mercantiles de muchospaíses. El intercambio comercial (importa-ción y exportación) es vital para muchos deellos. Sin embargo, frecuentemente el rendi-miento se ve disminuido por acción de lasplagas de Leguminosas, entre las que desta-

can las provocadas por los «gorgojos» o Brú-quidos, deteriorando la mercancía y ocasio-nando pérdidas que han llegado a millones dedólares para algunos países. Según el «Reportof the Pest Infestation Laboratory» de 1962,la extensión de las pérdidas llegó a un 70 %en Leguminosas y la causa principal se debióa la invasión de Brúquidos. Actualmenteexiste una copiosa bibliografía que recoge da-tos sobre continuas catástrofes en el productonacional bruto de algunos países por la inva-sión de estos insectos, ya sea en el campo oen los graneros. Así, según FLETSCHER, tansolo en la provincia de Ontario (Canadá)existe una pérdida de un millón de dólares alaño por la invasión de B. pisorum L. SegúnY. DE LUCA, en Argelia se pierde el 10 % dela cosecha de lentejas por la invasión de B.lentis Fról. Según KlEFFER, las pérdidas oca-sionadas por Acanth. obsoletus Say. y Call,maculatus F. en 1926 fueron cifradas en unmillón de chelines al año tan solo en algunospuntos.
Sin embargo, la cuestión del «brucage» deleguminosas ha sido tratada de una forma se-cundaria, cuando es sabida la importanciaque revisten las leguminosas en los países envías de desarrollo. Ello se debe al abismo queexiste entre las normas alimenticias de lospaíses en vías de desarrollo y los denomina-dos «países pilotos». Estos últimos consumenmuchas menos leguminosas, viniendo a de-cirse que constituyen «un plato pobre». Sinembargo, para otros países es primordial: enFrancia, el pedido de Leguminosas a U.S.A.aumentó en 9 millones desde 1959 a 1964. EnArgelia hubo en 1956 una producción de200.000 quintales, lo que produjo un beneficiode 400 a 1.600 millones de francos.
En España, la comercialización de legumi-nosas reviste bastante importancia. Según el«Anuario Estadístico de Producciones Agrí-colas», en el año 1943 la superficie de legu-
minosas cultivadas se estimó en 1.446.000hectáreas, con una cosecha media anual de7.071.000 quintales. Sin embargo, hubo unapérdida de 707.100 quintales, valorada en98.432.600 pesetas, debido al ataque de insec-tos, entre los cuales figuran con especial im-portancia los Brúquidos (Cuadro 1). Entre losproductos de mayor exportación predominanlas judías verdes: así, en 1975 la superficiecultivada fue de 22.000 hectáreas, lo que su-pone una producción estimada en 188.000 to-neladas, de las cuales la mayor parte se des-tina al autoconsumo y otra parte a la exporta-ción, siendo rechazada una mercancía esti-mada en 12.648 toneladas (Cuadro 5). La im-portación va acrecentándose de unos años aotros; así, entre 1973 y 1974 se vio aumentadaen un 5,2 % (Cuadro 4). La exportación tam-bién va acusando un notable ascenso, pero noobstante, la mercancía que España exportapresenta un estado fitosanitario de mayor«brucage» que la que importa de otros países(Cuadro 3).
Por otra parte, la noción de «valor alimenticio» de estos productos ha evolucionado detal forma, que la exigencia del consumidor, y,por tanto, la del comerciante, viene a recha-zar toda mercancía que presente un solo pa-rásito, exagerando en el pedido diciendo:«exento de todo ataque de insectos». Estasexageraciones son comprensibles en paísescon un período de superproducción y un tipode alimento muy evolucionado, lo que haconducido a un rechazo de un estudio pri-mordial al problema del estado fitosanitario,tratándose muy secundariamente, y en oca-siones hasta se minusvalora, el cultivo de es-tas plantas, en lugar de abordar el problemade raíz. A este respecto, Y. DE LUCA (1966)estima necesario fijar unas tasas de insecta-ción lo suficientemente objetivas para que,independientemente de la mejora de cultivos,se pueda valorar el estado sanitario real de la


Judías verdes
Superficiecultivada
(Has.)
22.000
Producciónestimada
(Tms.)
188.000
Exportación(Tms.)
15.543
Rechazado(Tms.)
12.648
Autoconsumo)(Tms.)
72.457
mercancía y de esta forma tener un índiceorientador para llegar a la mejora de la cali-dad de los productos, y de este modo noguiarse por rigores excesivos de apreciacióntotal, cuyo valor es:
Mediante esta tasa se podrá valorar si elataque se debe a especies monovoltinas o es-pecies polivoltinas, lo cual es importante, yaque en este último caso el ataque reviste ma-yor gravedd.
Medios de lucha
Hoy día existe abundante bibliografía refe-rente a este capítulo, por lo cual se sabe quese han utilizado multitud de métodos de lu-cha. Sin embargo, muchas tentativas dirigidascontra estos insectos han dado poco resul-tado y ello se debe en gran parte al escasoconocimiento de la biología de las especies.Nos limitaremos a preconizar, en base anuestro estudio, los medios de lucha suscep-tibles de llevarse a la práctica, a la vista desus resultados.
a) Medios preventivos
Quizá sean los más importantes, dado quepara ello no se necesita por lo general de
grandes gastos en aparatos y productos, y encambio se consigue una cosecha menos «tra-tada» y, por tanto, más natural. Por otra par-te, de la importancia que se le dé a estos mé-todos depende en gran parte la intensidad deposteriores métodos y su éxito.
La recolecta de frutos ha de ser total, estoes, con la mínima pérdida de semillas o frutosen el campo. Conviene no amontonarlos du-rante mucho tiempo en el campo, ya no sólopor el recalentamiento que provoca en losfrutos, sino porque, en estas condiciones, losadultos emergen con mayor facilidad y esca-parían a los abrigos invernales para atacarmasivamente las siguientes plantaciones. Porotra parte, han de utilizarse para la siembrasemillas sanas, pues las invadidas dan plantasdébiles y su rendimiento es mínimo y ademásliberarían al insecto alojado en ellas.
Otros medios preventivos consisten en rea-lizar la siembra (sobre todo en países cálidos)en una fecha tal que la maduración de los fru-tos se anticipe o pase de la época de madura-ción de las gónadas de los adultos, lo que in-cidirá sobre la eficacia de puesta y la intro-ducción de la larva en la semilla. General-mente se aconseja sembrar al suroeste mez-clando estas plantas con maíz que actúacomo tutor. Por otra parte conviene recogerlos frutos frescos de la periferia y dejar ma-durar los del centro, ya que el ataque co-mienza por la periferia.

b) Tratamiento en el granero
Una vez efectuada la recolecta y el desgra-ne, las semillas son introducidas en grandesalmacenes, en almacenes estancos con turbo-ventilador o en autoclaves de desinsectación.En esta fase, se detecta la invasión de brú-quidos por la emergencia de adultos y apari-ción de semillas opereuladas. En este mo-mento conviene distinguir si el ataque se debea una especie monovoltina o polivoltina. Enel primer caso, los daños no aumentarán du-rante el período de conservación y tan solo esurgente el tratamiento en el segundo caso.
La fumigación se realiza en depósitos es-tancos, mediante la técnica de proyección porla parte superior de los montones. Otras ve-ces se realiza en el mismo puerto, bajo lonas.Se ha utilizado mucho el sulfuro de carbono,pero hoy día se utilizan mejor (por ser másinofensivos para el hombre) productos talescomo el tetracloruro de carbono (40 gr./hcl.),aunque retrase algo la germinación, y sobretodo el lindano y bromuro de metilo. Se hande tapar todos los resquicios del almacén conmallas muy finas y ha de hacerse actuar alinsecticida durante 48 horas a menos de 20°C.En las autoclaves de desinsectación se haceun vacío parcial de 650-700 mm. y luego sehace entrar al insecticida con algo de CO2; acontinuación se vuelve a hacer el vacío y porúltimo se deja entrar el aire. Este métodosuele ser de los más efectivos.
Los tratamientos físicos son de ejecuciónlarga y aleatoria. Estos consisten en someteral granero a un calentamiento seco de 55-60°C. y generalmente matan a los insectos en30 minutos. Tienen la ventaja de que, almismo tiempo que matan al insecto, preser-van a la semilla contra posteriores ataques(polivoltinos), ya que al disminuir el conte-nido de agua de las semillas, se impide el ata-que de los insectos que requieren cierta tasa
de humedad para la penetración. Sin embar-go, esto requiere ciertas precauciones pues elpoder germinativo de la semilla disminuye li-geramente, así como su peso como mercan-cía.
También el frío actúa sobre los insectos yes sabido que a menos de 10°C. se hace im-posible la reproducción y a 0°C. no sobreviveninguno.
Otros procedimientos físicos han sido en-sayados, tales como el vacío, los rayos infra-rrojos, campo electrostático, etc., sin éxitosmuy aparentes.
A la hora de comercializar la mercancía seimpone un método eficaz para separar lassemillas atacadas de las sanas. Se han ensa-yado varios métodos pero sin resultado satis-factorio, ya que la selección se efectúa sobresemillas operculadas, es decir, aquellas quehan sido abandonadas por el adulto, lo que noes suficiente desde el punto de vista sanitario.
c) Tratamiento en el campo
Siempre ha sido bastante difícil, pues exigeun perfecto conocimiento de la biología delparásito y el huésped, para determinar conexactitud el período de susceptibilidad másfavorable. Para llevar a cabo un tratamientoeficaz, se precisa seguir los siguientes pasos:
1. Saber qué estado se va a destruir2. Escoger el producto adecuado3. Determinar el momento favorable de
luchaPara la destrucción de imagos hoy día se
han impuesto los insecticidas fosfóricos, porsu acción sistémica y gran poder de penetra-ción, actuando sobre el adulto y matándoloen 24 horas y, al mismo tiempo, merced a suacción ovo-larvicida, que impide la penetra-ción de la larva en el fruto. El tratamiento hade iniciarse cuando aparecen los adultos en

los cultivos, ya que la puesta depende más dela maduración de las gónadas que del estadofenológico de la planta huésped. Para elegir elmomento de la lucha ha de conocerse bien nosólo la biología del insecto, sino la fenologíade la planta huésped y las variaciones de éstasegún circunstancias climáticas y según la va-riedad.
La lucha biológica dirigida contra estos in-sectos es un método que se ha ensayado pocoy de momento no han habido grandes éxitos.A este respecto, BOGDANOV y KATKOV pien-san que ciertas variedades de plantas seránmás susceptibles que otras de recibir puestas;posiblemente se trate de plantas relativa-mente resistentes. Así ocurre con variedadestardías para parásitos precoces o variedadestempranas para parásitos tardíos. Otras va-riedades presentan diferencias en la tasa deagua y lípidos, lo que de alguna forma debeinfluir en el comportamiento de la hembra du-rante la puesta y supervivencia de la larva.
En cuanto a la utilización de parásitos, los
resultados han sido insignificantes, cuando nonulos. En U.S.A. se hizo un gran esfuerzopara aclimatar el parásito Triaspis thorácicacon el fin de combatir varios Brúquidos,como B. pisorum L., B. rufimanus Boh. y B.brachialis Fahr., y para ello se soltaron143.400 especímenes durante 4 años, sin con-seguir aclimatarlos. Por otra parte, no consi-deramos muy aconsejable la utilización deparásitos dado que éstos presentan un altoíndice de mortandad, la mayor parte de lasveces por no poder salir del fruto y otras porestar a su vez parasitados por otros himenóp-teros, por lo que aparecen en una proporciónmuy pequeña en las poblaciones de Brúqui-dos y, por otra parte, su acción no impide eldeterioro de la semilla, por cuanto el parásitomata al gorgojo cuando éste se encuentra yaen fase de larva adulta o ninfa, cuando elgrano ha sido ya deteriorado. Su utilizaciónrequiere un profundo conocimiento de la di-námica de poblaciones de huésped y parásito,cosa que, de momento, es escaso.
ABSTRACT
Yus RAMOS, R., 1976.—Las especies de Brúquidos (gorgojos de las Leguminosas) deinterés agrícola y fitosanitario (Col. Bruchidae). I: Caracteres generales. Bol. Serv.Plagas, 2: 1-35.
In this paper, the author makes a review of the general morfologic and biologiccharacters of the beetles relates to familly Bruchidae, with the purpose to introducethe systematic study of the species of agronomic and fitosanitary importance, as no-tion for ulterior observations of inmediate practic aplication.
REFERENCIAS
BALACHOWSKY, A. y MESNIL, L. 1936: Les insectes nui-sibles aux plantes cultivées. P. Lechevalier, Paris;t. II; 1921 págs.
CHITTENDEN, F. H. 1912: The cowpea weevil, U.S.Dept. Agrie, B. 96-part. VI; págs. 83-94.
DAVIAULT, L. 1928: Sur le développement post-embryonnaire de la Bruche du Haricot, Acanthosce-lides obtectus Say. Ann. Soc. Ent. Fr., t. XCVII,págs. 105-132.
DE LA PUERTA CASTELLÓ, L. 1973: Estado fitosanitario
de los productos importados durante el año 1972 porla zona de Valencia. Bol. lnfor. Plag., Minist. Agrie,n.° 105, págs. 75-80.
FRANSSEN, C. J. 1962: Lebenswijze en de bestrijding vande bonekever (Acanthoscelides obsoletus Say.).Versl. Landbourk. Ondrz-, t. 67; 88 págs.
HERFORD, G. M. 1935: A key to the members of the fa-mily of economic importance in Europe. Trans. Soc.Brit. Ent., t. II, págs. 1-32.
HOFFMANN, A. 1945: Coléoptéres Bruchides et Anthribi-

des. Faune de France: 44: Ed. P. Lechevalier, Paris;184 págs.
HORBER, E. 1950: Beitrag zur biologie und bekámfungdes species enbohnenkáfers (Acanthoscelides obtec-tus) (Col. Bruchidae). Mitt. Schweiz. Ent. Ges., t. 23;págs. 233-244.
KINGSOLVER, J. M. 1970: A study of male genitalia inBruchidae (Coleóptera). Proc. Ent. Soc. Washington,t. 72-n.° 3; págs. 370-386.
LABEYRIE, M. V. 1957: Influence des techniques de ré-colte des haricots sees sur l'intensité des attaques dela Bruche (Acanthoscelides obsoletos Say.).
LARSON, A. O. 1924: The effect of weevily seed beansupon the bean crop and upon the dissemination ofweevils. Bruchus obtectus Say. and Bruchus quadri-maculatus F.
LARSON, A. O. 1927: The host-selection as applied toBruchus quadrimaculatus F. Ann. Ent. Soc. Amer.,t. XX, págs. 37-53.
LEONIDE, A. 1960: Notes sur les zoocénosis des goussesde certaines Légumineuses. B. Soc. Entom. Franq.t. V, págs. 274-281.
LEPESME, P. 1941: A propos des Bruches et de la désin-sectation des legumes sees. Bull. Se. Pharm., t.XL-VIII, págs. 9-10.
LEPESME, P. 1942: Sur l'éclosion et le comportement dela larve néonate á'Acanthoscelides obtectus Say.(Col. Bruchidae). Rev. Fr. Ent. Fr., t. XLVII, págs.7-15.
LESPESME, P. 1945: Les coléptéres des denrées alimentaireset des produits industriéis entreposés. Ed. P. Lé-chevalier, Paris; 335 págs.
LUCA Y. DE 1956: Contributions á l'étude morphologi-que et biologique de Bruchus lentis Fróh. Essais delutte. Ann. Inst. Agrie. Algérie, t. X, fase. 1; 94 págs.
LUCA, Y. DE 1965: Catalogue des Métazoaires parasiteset prédateurs de Bruchidés (Col.) J. Stored Product.Res., t. I, págs. 51-98.
LUCA, Y. DE 1966: Alimentation imaginale des Bruchi-dés (Col.) Parasítica, t. 22-n.° 1; págs. 26-54.
LUCA, Y. DE 1966: Considerations sur le spectre vege-tal d'étalement des Bruchidés (Col.) Parasítica, t.23-n.° 2; págs. 56-65.
LUCA, Y. DE 1969: Au sujet des caractéristiques d'unlot de graines dépreciées para les Bruchidés (Col.).Ann. Soc. Hort. Hist. Nat. Hérault, t. 109 (2); págs.92-101.
MIRANDA NIEVES, R. 1969: Plagas y enfermedades másfrecuentes en los productos agrícolas de exportación.Bol. Infor. Plag. Minist. Agrie, Madrid; n.° 66, págs.1-44.
MUKERJEE, S., CHATTERJEE, S. N. 1951: Morphology ofthe genital structure of some of the Bruchidae (Larii-dae) of India and Ceylan and their taxonomic impor-tance. Jnd. Journ. Entom., t. XIII, part. 1, págs.1-28.
PARKER, H. L. 1957: Notes sur quelques Bruches et leursparasites eleves des graines de Légumineuses (Col.).Bull. Soc. Ent. Fr., t. 62; pág. 168-179.
SCHVESTER, D. 1957: Sur le dévelopement de la Bruche(Acanthoscelides obsoletus Say.) chez diverses varié-tés de Haricots en stock. Rev. Zool. Agrie. Appl.,págs. 120-125.
SHARP, D. y MUIR, F. 1912: The comparative anatomy ofthe male genital tube in Coleóptera. Trans. Ent. Soc,págs. 557-558.
SHOMAR, N. F. 1963: A monographic revision of the5r«-chidae of Egypt (UAR) (Coleopteres). Bull. Soc. Ent.Egypte, t. XLVII, págs. 141-196.
SKAIFE, S. H. 1926: The bionomics of the Bruchidae.South. Afr. J. Sc. Pretoria; t. 23, págs. 575-588.
TERAN, A. L. 1969: Observaciones sobre las estructurasgenitales de los machos de diversos géneros de Bru-chidae (Col.). Acta Zool. Lillneana, t. 22; págs. 307-336.
UTIDA, S. 1942: Host parasite interaction in the experi-mental population of the azuki bean weevil C. chi-nensis. The effect of density of parasite populationon the growth of the host population and also of theparasite population. Oyo-Kontyu, t. 4 (2), págs. 117-128.
ZACHER, F. 1930: Untersuchungen zur Morphologie undBiologie der Samenkafer (Bruchidae-Lariidae). Arb.Biol. Reischs. Land. Forstw., t. XVIII, págs. 223-230.