la singularidad de la especie humana: de la … · evolución tiene de hecho trascendentales...
TRANSCRIPT

Carlos Beorlegui
La singularidad de la especie humana
De la hominización a la humanización
DeustoDigitalPublicaciones


© Universidad de Deusto - ISBN 978-84-9830-383-4
La singularidad de la especie humana
De la hominizacion a la humanización


© Universidad de Deusto - ISBN 978-84-9830-383-4
Carlos Beorlegui
La singularidad de la especie humana
De la hominizacion a la humanización
2011Universidad de Deusto
Bilbao

© Universidad de Deusto - ISBN 978-84-9830-383-4
Serie Filosofía, vol. 38
Aprobado para su publicación por la Comisión de Publicaciones de la UD, tras dictamen fa-vorable emitido en un proceso de evaluación por pares externos a la Universidad de Deusto.
Cualquier forma de reproducción, distribución, comunicación pública o transformación de esta obra sólo puede ser realizada con la autorización de sus titulares, salvo excepción prevista por la ley. Diríjase a CEDRO (Centro Español de Derechos Reprográficos, www.cedro.org) si necesita fotocopiar o esca-near algún fragmento de esta obra.
Ilustración de portada: Diseño realizado a partir de un con-cepto expositivo mostrado en el Museo de la Evolución Humana (Burgos, 2011).© LIT Images
© Publicaciones de la Universidad de DeustoApartado 1 - 48080 Bilbaoe-mail: [email protected]
ISBN: 978-84-9830-383-4Depósito legal: BI - 571-2011
Impreso en España/Printed in Spain

© Universidad de Deusto - ISBN 978-84-9830-383-4
A la memoria de mis hermanos, Juan Clemente, Mercedes y Francisco Javier.
En recuerdo de Txema Mardones y Txema Fínez, amigos y compañeros de inquietudes intelectuales y vocacionales.

© Universidad de Deusto - ISBN 978-84-9830-383-4

© Universidad de Deusto - ISBN 978-84-9830-383-4
«Es indiscutible que los grandes temas —la existencia de Dios, nuestra visión de la naturaleza humana y nuestra relación con otras criaturas— se ven afectados por las ideas acerca de la evolución.
¿Por qué a nosotros (que no somos biólogos) debería importar-nos la evolución? La respuesta que ofrezco en este libro es que la evolución tiene de hecho trascendentales consecuencias para nuestra visión de nosotros mismos y de nuestro lugar en el universo, pero que no tiene en realidad la clase de consecuencias más ampliamente difundidas en la actualidad. En particular, tiene una utilidad limitada en lo que se refiere al esclarecimiento de la naturaleza humana».
Jean DUPRÉ, El legado de Darwin. Qué significa hoy la evolu-ción, Buenos Aires, Katz Editores, 2006, pp. 9 y 12.
«En el reino animal, el hombre es el único ser que puede ir más allá de sí mismo, que puede fijarse ideales o, dicho en otras pala-bras, que puede reconocer un sentido a su existencia. Y este sentido no es otro que poder vivir su vida como si debiera ser juzgada, como si la vida debiera responder a una llamada, a una exigencia, a una esperanza que trasciende la animalidad del hombre y que funda su humanidad —entendemos por esto último su capacidad de ser algo distinto que una bestia—.»
Jean GRONDIN, Del sentido de la vida. Un ensayo filosófico, Bar-celona, Herder, 2005, p. 24.

© Universidad de Deusto - ISBN 978-84-9830-383-4



© Universidad de Deusto - ISBN 978-84-9830-383-4
Índice
Introducción . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19
Capítulo 1. El entronque de la especie humana en la biosfera . . . . . . . . . . 31
1. Los avatares de la autocomprensión antropológica. . . . . . . . . . . . . . . . . . 312. La biología pre-darwiniana . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 363. La revolución de Charles Darwin (1809-1882). . . . . . . . . . . . . . . . . . . . . 39
3.1. El largo camino hacia la teoría de la selección natural . . . . . . . . . . . 39
a) Los primeros pasos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40b) La teoría de la selección natural . . . . . . . . . . . . . . . . . . . . . . . . . . 41
3.2. El origen del hombre y las discrepancias con R. Wallace . . . . . . . . . 433.3. El impacto del darwinismo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45
3.3.1. El darwinismo frente a la visión tradicional del hombre, de la realidad y de Dios . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
a) Nueva visión del hombre y de la realidad . . . . . . . . . . . . . 46b) El enfrentamiento religioso . . . . . . . . . . . . . . . . . . . . . . . . 48
1. La teoría tradicional del diseño . . . . . . . . . . . . . . . . . . . 482. William Paley (1743-1865) y su teoría del diseño. . . . . 493. La selección natural de Ch. Darwin. . . . . . . . . . . . . . . . 51
3.3.2. Oposición desde la filosofía idealista . . . . . . . . . . . . . . . . . . . 563.3.3. Impacto del darwinismo en el ámbito sociológico y económico 593.3.4. El impacto de la teoría de Darwin en la Antropología socio-
cultural . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61
4. G. Mendel (1822-1884) y el desarrollo de la genética . . . . . . . . . . . . . . . 62
4.1. G. Mendel y los inicios de la genética. . . . . . . . . . . . . . . . . . . . . . . . 62

12
© Universidad de Deusto - ISBN 978-84-9830-383-4
4.2. El redescubrimiento de la genética mendeliana. . . . . . . . . . . . . . . . . 644.3. Genética versus selección natural . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
a) Las teorías saltacionistas o mutacionismo . . . . . . . . . . . . . . . . . . 66b) Las teorías neolamarckianas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 69c) Teorías ortogenéticas. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 70d) El neodarwinismo de Auguste Weismann. . . . . . . . . . . . . . . . . . . 70
5. La teoría sintética de la evolución. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71
5.1. Los pasos hacia la primera síntesis . . . . . . . . . . . . . . . . . . . . . . . . . . 715.2. La teoría sintética. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 735.3. Las teorías alternativas a la teoría sintética . . . . . . . . . . . . . . . . . . . . 75
5.3.1. Teorías neolamarckianas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 765.3.2. Teorías neutralistas o estocásticas . . . . . . . . . . . . . . . . . . . . . . 78
5.4. Unas cuantas evidencias constatables . . . . . . . . . . . . . . . . . . . . . . . . 80
Capítulo 2. El darwinismo cuestionado . . . . . . . . . . . . . . . . . . . . . . . . . . . . 83
1. Los puntos débiles y las críticas al darwinismo en la actualidad . . . . . . . 832. La condición científica de la teoría de la selección natural. . . . . . . . . . . . 853. Las diversas propuestas sobre el ritmo del proceso evolutivo . . . . . . . . . 874. Críticas al darwinismo desde los fundamentalismos religiosos . . . . . . . . 90
4.1. El conflicto entre evolución y religión . . . . . . . . . . . . . . . . . . . . . . . 904.2. Precisiones epistemológicas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 934.3. La problemática relación entre ciencia y religión. Cuatro posturas
diferentes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99
a) Conflicto . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 100b) Independencia. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 102c) Diálogo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103d) Integración . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103
4.4. Las críticas de los creacionismos fundamentalistas. . . . . . . . . . . . . . 1044.5. La teoría del Diseño Inteligente (D.I.). . . . . . . . . . . . . . . . . . . . . . . . 110
a) Los argumentos y las debilidades del Diseño Inteligente. . . . . . . 112b) La teoría del DI no es una teoría científica y es una mala teología 116
4.6. Evolución darwinista y fe cristiana . . . . . . . . . . . . . . . . . . . . . . . . . . 1194.7. El diálogo con la ciencia: oportunidad para la fe y la teología . . . . . 1254.8. Ciencia y fe como aliadas y complementarias. . . . . . . . . . . . . . . . . . 127
5. El darwinismo y la actual comprensión del ser humano. . . . . . . . . . . . . . 128
Capítulo 3. La antropogénesis. La dimensión filogenética y la cuestión del poligenismo/monogenismo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 131
1. La microevolución y la macroevolución. . . . . . . . . . . . . . . . . . . . . . . . . . 131
1.1. La microevolución . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 132

13
© Universidad de Deusto - ISBN 978-84-9830-383-4
1.2. La macroevolución o tipogénesis . . . . . . . . . . . . . . . . . . . . . . . . . . . 1331.3. El paso de la macro a la microevolución . . . . . . . . . . . . . . . . . . . . . . 136
2. La dimensión filogenética del origen del hombre. . . . . . . . . . . . . . . . . . . 137
2.1. Las diversas fases de la aparición de la especie humana. . . . . . . . . . 138
2.1.1. El punto de vista de la Paleoantropología . . . . . . . . . . . . . . . . 140
a) La línea de los hominoideos o antropomorfos . . . . . . . . . . 143b) Los homínidos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 145
b.1) Los Orrorin tugeniensis . . . . . . . . . . . . . . . . . . . . . . . 147b.2) Los Ardipithecos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 148
1. Ardipithecus kadabba . . . . . . . . . . . . . . . . . . . . . . 1482. Ardipithecus ramidus . . . . . . . . . . . . . . . . . . . . . . . 148
b.3) Los Australopithecos . . . . . . . . . . . . . . . . . . . . . . . . . 149
1. Australopithecus anamensis . . . . . . . . . . . . . . . . . 1502. Australopithecus afarensis. . . . . . . . . . . . . . . . . . . 1513. Australopithecus africanus . . . . . . . . . . . . . . . . . . 1524. Australopithecus garhi. . . . . . . . . . . . . . . . . . . . . . 153
b.4) Los Paranthropos . . . . . . . . . . . . . . . . . . . . . . . . . . . . 154
c) Los humanos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 155
c.1) El homo habilis. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 156c.2) El «grado erectus» . . . . . . . . . . . . . . . . . . . . . . . . . . . 158
1. Homo ergaster . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1592. Homo erectus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1603. El homo antecesor . . . . . . . . . . . . . . . . . . . . . . . . . 1624. ¿Homo floresiensis?. . . . . . . . . . . . . . . . . . . . . . . . 164
c.3) Los neandertales y el homo sapiens moderno . . . . . . 165
1. Los Neanderthales . . . . . . . . . . . . . . . . . . . . . . . . . 1662. Los Cromañones. El homo sapiens . . . . . . . . . . . . 168
2.1.2. El punto de vista de la biología molecular . . . . . . . . . . . . . . . 171
a) A la búsqueda del «reloj molecular» . . . . . . . . . . . . . . . . . 172b) Implicaciones respecto al origen del hombre moderno. . . 177
3. La disputa entre el poligenismo y el monogenismo . . . . . . . . . . . . . . . . . 181
3.1. ¿Polifiletismo o monofiletismo? . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1823.2. ¿Poligenismo o monogenismo? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 187
Capítulo 4. Las transformaciones genéticas y morfológicas y la cuestión del sentido (azar o finalidad) de la evolución . . . . . . . . . . . . . . . . . . . . . . . 197
1. Las transformaciones genéticas y morfológicas . . . . . . . . . . . . . . . . . . . . 197

14
© Universidad de Deusto - ISBN 978-84-9830-383-4
2. Las dos fronteras de la hominización . . . . . . . . . . . . . . . . . . . . . . . . . . . . 200
2.1. El salto mutacional . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2002.2. La encefalización . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 208
3. Los rasgos morfológicos de la hominización . . . . . . . . . . . . . . . . . . . . . . 210
3.1. La bipedestación o postura erguida . . . . . . . . . . . . . . . . . . . . . . . . . . 2113.2. El desarrollo del encéfalo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2193.3. La mano prensil e inteligente. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2283.4. La transformación de los rasgos faciales . . . . . . . . . . . . . . . . . . . . . . 230
4. ¿Azar o finalidad en la evolución y en el universo? . . . . . . . . . . . . . . . . . 233
4.1. La cuestión acerca del azar o la finalidad en la evolución . . . . . . . . 2334.2. La recuperación del azar y de la probabilidad . . . . . . . . . . . . . . . . . . 2354.3. La finalidad teleonómica en el mundo de la biología . . . . . . . . . . . . 2394.4. ¿Finalidad o azar en la historia del universo? . . . . . . . . . . . . . . . . . . 246
4.4.1. ¿Cosmocentrismo o antropocentrismo? . . . . . . . . . . . . . . . . . 247
a) Razones del anti-antropocentrismo o de las tesis del cosmo-centrismo. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 248
b) Razones a favor del antropocentrismo . . . . . . . . . . . . . . . . 249
4.4.2. Entre el principio cosmológico o copernicano y el principio antrópico . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 250
a) El principio cosmológico o copernicano de la tesis cosmo-céntrica . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 250
b) El principio antrópico . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 252
1. La versión débil del principio antrópico (Weak Anthropic Principle: W.A.P.) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 253
2. Versión fuerte (Strong Anthropic Principle: S.A.P.). . . . 2543. El principio antrópico participativo (P.A.P.) de J. Wheeler 255
c) Valoración glogal de los dos principios . . . . . . . . . . . . . . . 258
4.4.3. Entre el azar y la finalidad: una cuestión de opción filosófica 259
Capítulo 5. El proceso de humanización. La dimensión conductual . . . . . . 263
1. La humanización como complemento de la hominización . . . . . . . . . . . . 2632. Los principales rasgos culturales y conductuales . . . . . . . . . . . . . . . . . . . 265
2.1. La conciencia refleja o autoconciencia . . . . . . . . . . . . . . . . . . . . . . . 2652.2. La imaginación y la capacidad simbolizadora. . . . . . . . . . . . . . . . . . 2742.3. El lenguaje . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2762.4. La creación y uso de herramientas y armas . . . . . . . . . . . . . . . . . . . . 2912.5. Dominio y uso del fuego . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3042.6. Los usos alimenticios. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3132.7. El hábitat humano. La construcción de refugios y casas . . . . . . . . . . 322

15
© Universidad de Deusto - ISBN 978-84-9830-383-4
2.8. La socialidad: de la familia a los primeros grupos sociales . . . . . . 327
2.8.1. La sexualidad humana y las primeras células familiares . . . 3282.8.2. El salto de la naturaleza a la cultura: el tabú del incesto . . . 3332.8.3. La formación de sociedades complejas. . . . . . . . . . . . . . . . . 338
2.9. La aparición de la capacidad estética y de las obras de arte . . . . . . 342
2.9.1. Estética animal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3432.9.2. Los inicios de los objetos de arte . . . . . . . . . . . . . . . . . . . . . 346
a) Las tesis gradualistas frente a las rupturistas . . . . . . . . . . 347
a.1) Las tesis gradualistas . . . . . . . . . . . . . . . . . . . . . . . . 348a.2) La tesis rupturista . . . . . . . . . . . . . . . . . . . . . . . . . . . 349
b) Una visión gradual más completa . . . . . . . . . . . . . . . . . . 351
2.10. Los primeros vestigios de la religión y de la moral . . . . . . . . . . . . 353
a) Los primeros enterramientos . . . . . . . . . . . . . . . . . . . . . . . . . . . 354b) Las primeras creencias (¿La primera «metafísica»?). . . . . . . . . 359
3. Concluyendo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 361
Capítulo 6. La antropogénesis. El punto de vista ontogenético. . . . . . . . . . 363
1. La problemática relación entre filogénesis y ontogénesis: una visión evo-lutiva de la embriología. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 363
2. Las diferentes fases del proceso ontogenético . . . . . . . . . . . . . . . . . . . . . 369
2.1. Fase previa: de los gametos a la célula germinal (pre-ontogénesis). . 3702.2. Fase zigótica: de la célula germinal o zigoto a la anidación o implan-
tación . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3712.3. Fase embrionaria o de gastrulación . . . . . . . . . . . . . . . . . . . . . . . . . . 3742.4. Fase fetal . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3752.5. Fase de oriogénesis y de gerontogénesis . . . . . . . . . . . . . . . . . . . . . . 377
3. La ontogénesis humana vista desde la embriología comparada . . . . . . . . 3784. El especial puesto de la especie humana en el proceso ontogenético . . . . 3815. Cuestiones filosóficas planteadas en el terreno de la ontogénesis . . . . . . 386
5.1. ¿La célula germinal es un ser humano?. . . . . . . . . . . . . . . . . . . . . . . 3865.2. ¿Cuándo se llega a ser persona? . . . . . . . . . . . . . . . . . . . . . . . . . . . . 388
a) La célula germinal es ya una persona. . . . . . . . . . . . . . . . . . . . . . 389b) Persona: entre la anidación y la formación del sistema inmuno-
lógico . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 392c) La postura emotivista y utilitarista . . . . . . . . . . . . . . . . . . . . . . . . 397d) La persona como entidad relacional: a partir del nacimiento. . . . 404
5.3. ¿Cómo se llega a ser persona? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 408

16
© Universidad de Deusto - ISBN 978-84-9830-383-4
Capítulo 7. La singularidad de los humanos: entre el antropocentrismo y el reduccionismo biológico . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 415
1. El antropocentrismo en cuestión . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4152. La comparación hombre-animal: dos enfoques complementarios (cientí-
fico y filosófico) y cuatro ámbitos de comparación . . . . . . . . . . . . . . . . . 419
2.1. Ciencia y filosofía: dos modos complementarios de acercarse a la realidad y al ser humano . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 419
2.2. Cuatro ámbitos de comparación . . . . . . . . . . . . . . . . . . . . . . . . . . . . 421
2.2.1. El ámbito bioquímico y genético . . . . . . . . . . . . . . . . . . . . . . 4212.2.2. El punto de vista anatómico y morfológico . . . . . . . . . . . . . . 4272.2.3. La comparación desde el proceso embriológico . . . . . . . . . . . 4292.2.4. Comparación desde la estructura comportamental . . . . . . . . . 433
2.2.4.1. Las tesis deterministas . . . . . . . . . . . . . . . . . . . . . . . . 435
a) El determinismo genético de la sociobiología . . . 435b) El determinismo ambiental del conductismo . . . . 436
2.2.4.2. Entre la genética y el ambiente. . . . . . . . . . . . . . . . . . 437
3. La singular estructura comportamental de los seres humanos . . . . . . . . . 4394. ¿Ampliación de la «humanidad» a los grandes simios? . . . . . . . . . . . . . . 4425. La específica y singular estructura esencial de la especie humana . . . . . . 446
Capítulo 8. Entre la biología y la cultura . . . . . . . . . . . . . . . . . . . . . . . . . . . 453
1. La emergencia de la cultura desde la biología . . . . . . . . . . . . . . . . . . . . . 453
1.1. El ser humano, fruto de dos herencias . . . . . . . . . . . . . . . . . . . . . . . . 4541.2. ¿Cultura animal? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4561.3. El salto de la biología a la cultura . . . . . . . . . . . . . . . . . . . . . . . . . . . 462
2. La conjugación de la cultura con la biología. . . . . . . . . . . . . . . . . . . . . . . 467
2.1. El reduccionismo biológico . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 469
a) Biologismo extremo o naturalismo biológico . . . . . . . . . . . . . . . 469b) Biologismo o naturalismo moderado . . . . . . . . . . . . . . . . . . . . . . 470
2.2. El biologismo funcionalista o subordinacionista. . . . . . . . . . . . . . . . 4742.3. El culturalismo dualista . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4762.4. El culturalismo estructurista . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 479
3. La relación de la biología con la ética. . . . . . . . . . . . . . . . . . . . . . . . . . . . 4814. El ser humano, unidad bio-cultural . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 491
Capítulo 9. Continuidad y ruptura: la especie singular . . . . . . . . . . . . . . . . 493
1. Una visión bio-filosófica de lo humano . . . . . . . . . . . . . . . . . . . . . . . . . . 4932. El reconocimiento de nuestra condición animal . . . . . . . . . . . . . . . . . . . . 494

17
© Universidad de Deusto - ISBN 978-84-9830-383-4
3. El hombre, fruto de un largo proceso filogenético y ontogenético . . . . . . 4974. Continuidad y emergencia cualitativa . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4995. El animal bio-cultural . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5046. La legitimidad del humanismo antropocéntrico . . . . . . . . . . . . . . . . . . . . 5067. ¿Somos el resultado del puro azar, o podemos apostar por un fundamento
último y providente del universo?. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 5088. Concluyendo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 510
Bibliografía . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 511


19
© Universidad de Deusto - ISBN 978-84-9830-383-4
Introducción
El ser humano tiene como elemento constitutivo de su condición la capacidad y la necesidad de tener una imagen de sí mismo. Ningún otro ser vivo posee esta capacidad, ni tampoco siente necesidad de ello. Nues-tra extraordinaria inteligencia y el poder convertirnos a nosotros mismos en objeto de reflexión nos permite hacernos preguntas sobre todo, tam-bién sobre nuestra propia realidad. Esta cualidad no es meramente un lujo, algo que realizamos en momentos de ocio y de descanso, sino que repre-senta un ejercicio vital para nuestra supervivencia, para realizarnos y estar a la altura de nuestra específica condición. La naturaleza no nos ha dotado de unos recursos y procedimientos automáticos para sobrevivir, sino que la deficiencia biológica con la que nacemos, consecuencia de nuestra pre-maturidad, nos obliga a poner a prueba nuestra capacidad intelectual y to-dos los medios que tenemos a nuestro alcance para superar la prueba de la selección natural.
Pero no se trata sólo de no tener la existencia asegurada. Al estar do-tados de una naturaleza abierta, necesitamos construir permanentemente nuestra peculiar forma de existir. Nacemos inacabamos y tenemos que ir realizándonos, sin tener asegurada la forma de hacerlo y el éxito final. Por eso, el ser humano necesita tener una imagen de sí mismo que se convier-ta en meta de su propia autorrealización. De ahí que la exigencia de esta imagen de sí no sea un dato superficial, sino que constituye una necesidad perentoria para dirigir hacia ella todos sus esfuerzos hacia la realización de sí mismo.
Cada individuo ha tenido que construir su propia imagen, así como cada cultura y cada época histórica. Por eso, no puede extrañarnos la am-plia proliferación de imágenes de lo humano con que nos encontramos a

20
© Universidad de Deusto - ISBN 978-84-9830-383-4
lo largo de todas las épocas históricas, así como la pluralidad de propues-tas antropológicas que vemos hoy día en nuestro entorno. Y ello es así porque a la cuestión sobre la naturaleza y el ser de lo humano se respon-de y se puede responder de múltiples formas, tantas como seres humanos, culturas y cosmovisiones existen. Cada uno de nosotros proyecta sobre el horizonte una imagen ideal y utópica a la que tratamos de dar alcance, aunque de hecho nunca lo consigamos.
Si a lo largo de la mayoría de la historia humana ha predominado la tendencia a entenderse el ser humano en referencia a lo sobrenatu-ral, como criatura de Dios, situando lo esencial de su condición en un peldaño por encima de todo lo creado, sin sentirse en continuidad con el resto de las cosas, y considerando el mundo sólo como el entorno mate-rial donde tiene que sobrevivir, desde mediados del siglo XIX el ser hu-mano ha ido descubriendo, con sorpresa y cierta resistencia al principio, y con admiración después, su común origen con el resto de las especies que forman la biosfera. Con la propuesta de Ch. Darwin sobre la selec-ción natural se produce un cambio revolucionario de autocomprensión del ser humano, que le lleva a entenderse más cercano al resto de los se-res vivos, y, por ello, a tener que admitir la centralidad de lo corpóreo en la constitución de su ser, produciéndose una radical biologización de la Antropología.
El ser humano toma conciencia de que no es tanto una res cogitans in-dependiente de lo corpóreo, sino que tal capacidad intelectiva es resulta-do de un proceso evolutivo, que le ha dotado de unos rasgos específicos, entre ellos un cerebro con especial complejidad, que le permite pensar y actuar al estilo propio de los humanos. Por tanto, no sólo no tenemos que menospreciar lo corpóreo como algo accidental, o como un estorbo que es-tamos obligados a aguantar, sino que tenemos que admitir que somos ra-dicalmente cuerpo, racionalidad encarnada.
Es evidente que este cambio de óptica ha tenido importantes conse-cuencias para el estudio de nuestra especie, de tal modo que parecía que la filosofía ya no tenía casi razón de ser en el ámbito antropológico, sien-do reemplazada por las diferentes Antropologías científicas, con el peligro de querer reducir lo humano a un capítulo más de la zoología, la etología, o de cualquier otra rama científica que estudia la composición de los de-más seres vivos y sus complejos comportamientos. Todo ello era conse-cuencia, por un lado, de las fructíferas perspectivas que para el estudio científico de los humanos había abierto el evolucionismo y la teoría de la selección natural de Darwin, así como, por otro, la pretensión de muchos intérpretes de la teoría evolutiva de que con sus planteamientos se llegaba a demostrar de forma contundente que no tenía sentido seguir defendiendo la centralidad de nuestra especie y menos aún su dimensión trascendente,

21
© Universidad de Deusto - ISBN 978-84-9830-383-4
teniéndose que admitir su reducción a una especie animal más, todo lo ex-traordinaria que se quiera considerar.
Estas pretensiones biologistas y reduccionistas representaban un de-safío demasiado fuerte como para que se pudiera pasar por alto. Se ponía en cuestión no sólo la legitimidad de la dimensión trascendente de nues-tra especie, sino también cualquier tesis antropocéntrica que exigiera de forma inevitable la pertinencia del enfoque filosófico en el estudio del hombre. Se nos planteaban como consecuencia una serie de cuestiones fundamentales por resolver. ¿Son suficientes las aportaciones de las di-ferentes ciencias de lo humano para dar cuenta total de su peculiaridad y su especificidad? ¿No parece que de ese modo sólo nos quedamos con un amplio abanico de datos sobre las diversas dimensiones que conforman su enorme riqueza de perspectivas? ¿No parece que la propia naturaleza de la pregunta que más nos interesa, el ser y el sentido de lo humano, como mirada unitaria y totalizante que supera lo meramente fáctico, escapa a las pretensiones y posibilidades de lo científico?
El contraataque de la filosofía desde los inicios del siglo XX, con las propuestas de renovar la Antropología filosófica de la mano de Max Scheler, Helmut Plessner y sus seguidores, orientó la pregunta acerca de lo específico de la especie humana hacia un diálogo necesario y fructífero entre las diversas ciencias humanas y la filosofía, para intentar desentra-ñar la complejidad de lo humano, conjugando las aportaciones científicas y la crítica filosófica. Pero no constituyó una línea de reflexión valorada unánimemente, no sólo en la medida en que desde la filosofía no todos aceptaron esta propuesta concreta de Antropología filosófica, sino porque sigue persistiendo en muchos científicos el convencimiento de que no se necesita la filosofía para dar cuenta de la especificidad del ser humano, entendiendo que las ciencias son suficientes para realizar con éxito esta función.
Como puede comprenderse, esta cuestión no es baladí, ni represen-ta un tema que sólo interesa a un grupo de académicos que se entretienen en temas demasiado abstractos y al margen de los intereses concretos de los ciudadanos de a pie, sino que constituye un conjunto de cuestiones de gran calado y consistencia (aunque su concreción teórica resulta a veces difícil de entender y valorar), y de consecuencias muy serias y determi-nantes en el ámbito de la ética y de nuestro modo de vivir y de organizar la sociedad.
Y son temas tanto más importantes y determinantes por cuanto en la actualidad nos hallamos inundados de publicaciones de todo tipo y de no-ticias periodísticas de gran impacto entre el público, y en las que se pro-ponen y se presentan noticias y avances científicos cuya interpretación distorsiona en ocasiones la imagen que se tiene del ser humano. Esto lleva

22
© Universidad de Deusto - ISBN 978-84-9830-383-4
a la necesidad de analizar el alcance de esas afirmaciones científicas sobre lo humano, su pertinencia y sus limitaciones, así como la obligación de conjugar sus aportaciones con un enfoque filosófico crítico. Estas son al-gunas de las motivaciones que se hallan al fondo de este libro.
Cuando publiqué hace algunos años Antropología filosófica. Noso-tros: urdimbre solidaria y responsable (Bilbao, Universidad de Deusto, 1999), indicaba en el prólogo que era la primera parte de un proyecto antropológico que exigía una continuación. Desde entonces llevo reco-pilando materiales para escribir la segunda parte del libro, pero otras muchas tareas (académicas y extra-académicas) me han impedido reali-zarlo en un tiempo razonable, tal y como lo tenía programado. Y cuando me puse ya en serio a realizarlo, al comenzar el primer capítulo sobre la condición bio-cultural del ser humano, me fui dando cuenta de que la amplitud e importancia del tema hacía que se fuera convirtiendo en un libro autónomo.
Un programa completo de Antropología filosófica debiera compren-der tres partes fundamentales: una primera, dedicada a labores de funda-mentación teórica y encargada de delimitar lo que se suele denominar el estatuto epistemológico de la materia en cuestión; una segunda, de tipo histórico, encaminada a presentar el surgimiento de esta disciplina en su etapa moderna, de la mano de quien se suele considerar el iniciador de la moderna Antropología filosófica, Max Scheler, junto con otros autores de su misma línea, como Plessner y Gehlen, empeñados todos ellos en estudiar de un modo renovado la singularidad y especificidad de lo hu-mano, distinguiendo entre el modo de hablar del ser humano propio, por un lado, de la filosofía del hombre y de las antropologías científicas, y el de la Antropología filosófica, por otro; y la tercera parte se tendría que centrar en el estudio de las múltiples dimensiones existenciales de la rea-lidad humana.
Las dos primeras partes de este amplio programa estaban ya presen-tadas en mi libro Antropología filosófica. Nosotros: urdimbre solidaria y responsable. Quedaba pendiente, por tanto, para un segundo volumen todo el desarrollo de la tercera parte, que comprendería el estudio, como he apuntado, de las denominadas dimensiones existenciales de la realidad humana, o simplemente de las diferentes facetas que configuran nuestra realidad, como son la condición bio-cultural del ser humano, su unidad psico-somática, la individualidad o subjetividad, la libertad, la interper-sonalidad y socialidad del hombre, la historicidad, el problema del mal, el fracaso y la muerte, la condición elpídica (la esperanza) y utópica de la realidad humana, y, para concluir, la apertura del ser humano a la pre-gunta por el fundamento de la realidad, es decir, la referencia del ser hu-mano a lo divino.

23
© Universidad de Deusto - ISBN 978-84-9830-383-4
Como he dicho más arriba, sólo el estudio de la primera de las facetas de la tercera parte del programa, la condición bio-cultural del ser humano, constituye una temática tan amplia, tan profundamente estudiada en casi todas sus parcelas, que resulta más que suficiente como para convertir su estudio en un libro autónomo y de gran amplitud. Basta echar un vistazo a lo aportado recientemente por disciplinas como la genética, la biología molecular, la paleoantropología, la embriología, la psicología del desarro-llo, la etología, la psicología evolucionista, etc., para advertir que preten-der lograr una síntesis adecuada de todo ese conjunto de disciplinas sobre el ser humano, y proponer desde ahí la imagen específica de lo humano que de tales investigaciones se desprende, casi constituye una pretensión prometeica. De ahí que lo presentado en estas páginas no puede tener más pretensiones que un acercamiento modesto a los aspectos más rele-vantes que sobre nuestra condición humana nos aportan este abanico de ciencias genéticas, biológicas y del comportamiento, para proponer, tras un diálogo crítico con ellas, la pertinencia renovada de una imagen huma-nista del ser humano y una visión antropocéntrica de nuestro mundo.
Desde que se fue abriendo paso el paradigma de la evolución a lo lar-do del siglo XIX, y Darwin planteó su teoría de la selección natural como el motor de dicho proceso evolutivo, completado más adelante con las propuestas de la genética, iniciadas con los primeros descubrimientos de sus leyes básicas por G. Mendel y continuadas con su redescubrimiento en 1900 por tres científicos a la vez (caso extraordinario, aunque no úni-co, en la historia de la ciencia), los desarrollos posteriores de la propia genética, la biología molecular, la paleoantropología, la embriología, la etología y demás ciencias del comportamiento, han ido transformando de forma fundamental la imagen que tenemos de nosotros mismos, haciéndo-nos ver la radical familiaridad que la especie humana posee con el resto de las especies vivas, con las consiguientes consecuencias que esto ha su-puesto para nuestra relación con el entorno de la biosfera y del universo, y para ir transformando el ámbito de la ética, de las relaciones sociales y políticas, así como nuestras concepciones religiosas. Podríamos decir que nada de nuestra concepción anterior del hombre y de la realidad ha queda-do indiferente y sin transformar.
Si el ser humano antes de las teorías evolucionistas se comprendía como un ser singular y al margen del conjunto de los demás seres vivos, interpretándose desde el horizonte de una cosmovisión religiosa (habla-mos sobre todo del ámbito occidental cristiano) como creado directamen-te por Dios del barro de la tierra, y a su imagen y semejanza, apoyando dicha teorización antropológica en una interpretación casi literal y poco crítica de la Biblia judeocristiana, las aportaciones de la biología evolu-cionista y demás ciencias afines parecen haber hecho tambalearse todas

24
© Universidad de Deusto - ISBN 978-84-9830-383-4
estas creencias y convicciones. De tal modo que, a partir de las aportacio-nes de Darwin, el ser humano se descubre anclado y situado como una es-pecie más dentro de la biosfera, de la que ha surgido, a través del común proceso evolutivo, a partir de las especies que nos preceden (los australo-pithecos). La tarea que tiene por delante, ante este panorama, un estudio de Antropología filosófica (en la línea que le corresponde a una reflexión radical sobre lo específico de lo humano) no puede ser otra que acoger y examinar las diversas aportaciones que las múltiples ciencias (naturales y humanas) nos van ofreciendo sobre la condición humana, para después reflexionar sobre ellas desde un enfoque filosófico crítico.
Las dos cuestiones o tareas más acuciantes que este cambio de pers-pectiva antropológica nos dejan pendientes, desde nuestro punto de vista, son, en primer lugar, analizar y separar del conjunto de aportaciones cien-tíficas sobre lo humano lo correspondiente al ámbito de la ciencia y lo si-tuado en el terreno de las interpretaciones filosóficas e ideológicas. Y, en segundo lugar, plantearnos la cuestión de si tras la teoría de la selección natural no tenemos más remedio que abandonar las pretensiones humanis-tas de seguir defendiendo una diferencia cualitativa entre el ser humano y el resto de las especies animales. Ambas cuestiones están íntimamente re-lacionadas, puesto que si distinguimos y diferenciamos adecuadamente el nivel científico y el filosófico, tendremos que reconocer que las aportacio-nes científicas no pueden llegar más que a constatar datos y hechos sobre la dimensión fáctica de lo humano, sin corresponderle realizar afirmacio-nes de valor y de sentido sobre el ser humano. Por tanto, la cuestión de si es pertinente o no seguir defendiendo las tesis humanistas, no es tarea de la ciencia sino de las interpretaciones filosóficas. Y dentro del nivel filo-sófico nos encontraremos siempre con una gran variedad y amplitud de propuestas e imágenes sobre lo humano, ninguna de las cuales podrá arro-garse la capacidad de considerarse la única verdadera, sea que invoque para ello la autoridad de la ciencia o la inspiración divina. Un modelo an-tropológico, para pretender ser tomado en serio, no podrá oponerse a los datos probados de las ciencias, y tener una coherencia interna, pero no po-drá arrogarse la pretensión dogmática de ser el único verdadero, ni aducir, como hemos señalado, datos científicos para demostrar su certeza.
Uno de los errores más extendidos en muchas publicaciones sobre es-tos temas consiste precisamente en no hacer una buena distinción entre el nivel científico y el filosófico, e intentar sacar conclusiones incorrectas y radicales tanto de la teoría de la selección natural en su conjunto, como de determinadas aportaciones parciales de una ciencia determinada. Pero tan ilegítimo es, como tendremos ocasión de ver, el cientifismo naturalista,que niega la pertinencia de la filosofía en beneficio exclusivo de la cien-cia, sin advertir que esas mismas afirmaciones son filosóficas y no cientí-

25
© Universidad de Deusto - ISBN 978-84-9830-383-4
ficas, como el fundamentalismo religioso, incapaz también de distinguir entre ciencia y filosofía/teología, descartando dogmáticamente cualquier afirmación de la ciencia cuando no concuerdan con una verdad religiosa deducida de una lectura acrítica de su libro sagrado.
El título del libro por el que al final nos hemos decantado, tras no po-cas vacilaciones, La singularidad de la especie humana. De la hominiza-ción a la humanización, hace referencia a la radical condición bio-cultural de nuestra especie. Lo que nos ha hecho humanos es tanto un proceso de evolución que nos entronca con el resto de las especies vivas (hominiza-ción), como también el salto al mundo de la cultura, como consecuencia de la emergencia de un sistema cualitativamente nuevo de vivir, el espe-cífico de la especie humana, que nos ha permitido desprendernos de los automatismos biológicos para poder hacernos cargo de nuestra vida e ir conformándola a través de nuestras decisiones libres (humanización). Por eso, la tesis humanista que queremos defender se encamina a entender al ser humano en radical continuidad con el mundo de la biosfera, al mismo tiempo que situado en un nivel singular que lo distingue cualitativamente del resto. La especie humana ha ido configurando el continente de la cul-tura, atisbado y anticipado de alguna manera por diversas especies ani-males anteriores, para irla ensanchando, colonizando y completando, y en la medida en que la humanización no ha terminado, la antropogénesis se halla a mitad de camino, hallándose el ser humano en permanente proceso de realización.
En el primer capítulo presentamos el profundo cambio que se produce en la imagen de los humanos como consecuencia de la teoría de la selec-ción natural de Darwin, siguiendo al naturalista inglés en la evolución de su pensamiento, su relación con R. Wallace y el conjunto de reacciones que se fueron produciendo ante sus propuestas. Igualmente hacemos re-ferencia breve y suficiente a las investigaciones punteras de Mendel en el mundo de la genética y el redescubrimiento de sus teorías años después, para más adelante confluir con la teoría de la selección natural en la de-nominada teoría sintética de la evolución. Con motivo del doscientos ani-versario del nacimiento de Darwin y de los ciento cincuenta años de la publicación de El origen de las especies, ha visto la luz una cantidad de bibliografía que resulta imposible controlar. La pretensión de mi breve síntesis sobre estos temas se reduce a encuadrar la problemática de la teo-ría de la evolución, de su síntesis posterior con los avances de la genética, y servir de acercamiento básico a la reflexión sobre estos puntos.
Este primer capítulo se completa con un segundo en el que recojo y completo algunas de las críticas que sobre la teoría de la selección natural se han ido vertiendo, desde su condición de teoría científica, las polémicas sobre el ritmo de la evolución, y las críticas a la teoría evolucionista desde

26
© Universidad de Deusto - ISBN 978-84-9830-383-4
los creacionismos fundamentalistas y la más moderna teoría del Diseño In-teligente. Como toda gran teoría científica, la teoría de la selección natural de Darwin ha tenido sus defensores y detractores, convirtiéndose en mu-chos momentos las discusiones a favor y en contra en una defensa a ultran-za o en un ataque descalificador, sin medias tintas. No cabe duda de que estas discusiones de todo o nada dificultan el avance de la ciencia, que se ha ido conformando por sucesivos avances y retrocesos, y sucesivas mejo-ras. La teoría de la selección natural se halla en la actualidad aceptada por la inmensa mayoría del mundo científico, pero eso no quita que muchos de sus aspectos secundarios y periféricos tengan que ser contrastados y veri-ficados, pudiendo resultar algunos de ellos fallidos o incompletos, sin que eso suponga poner en cuestión la centralidad de la teoría.
Tras presentar en sus líneas generales la teoría evolucionista, nos de-tenemos en el capítulo tercero en exponer el proceso de conformación de la especie humana desde el punto de vista filogenético, echando mano de las aportaciones de la paleoantropología y de la biología molecular. Como se puede imaginar, al igual que en relación con las investigaciones sobre Darwin y las teorías rivales, los avances y publicaciones sobre los oríge-nes de la especie humana se producen con tal rapidez y aportan tal canti-dad de publicaciones al respecto, que resulta imposible estar a la última. Además, no puede ser esa nuestra pretensión, en la medida en que nos interesa resaltar los resultados más importantes y básicos de este cam-po para poder defender una interpretación de lo humano que se apoye y respete las investigaciones que las ciencias correspondientes nos están aportando. Completamos el capítulo acercándonos a lo que los saberes científicos nos dicen en la actualidad sobre la ya antigua disputa entre el poligenismo y el monogenismo, con las consecuencias que estas eviden-cias suponen para el ámbito filosófico y teológico.
El capítulo cuarto complementa el punto de vista filogenético sobre el origen del ser humano, describiendo las progresivas y profundas transfor-maciones genéticas y morfológicas que se producen en el género humano desde que se atraviesan las denominadas dos fronteras de la hominización, la cromosómica y la cerebral hasta la aparición del género humano. Tras si-tuar a la especie humana en el proceso diacrónico de la evolución, se hace necesario completar el proceso presentando el conjunto de cambios que nos han llevado a ser como somos. La frontera clave es la genética, esto es, el salto mutacional de veinticuatro pares de cromosomas que caracte-riza a los póngidos, a veintitrés pares que constituye la dotación cromosó-mica humana. Y la consecuencia de ello es el desarrollo espectacular del cerebro, la postura erguida, la mano prensil y las trasformaciones faciales. Todas estas metamorfosis forman un proceso coevolutivo, que comienza con el bipedismo y sigue con el resto de los demás cambios, y en el que un

27
© Universidad de Deusto - ISBN 978-84-9830-383-4
pequeño avance en una de ellas produce otros posteriores en el resto, ge-nerando así un dinamismo de interacción que parece no se detiene. Hemos introducido como complemento de este capítulo, y desde la perspectiva del conjunto del proceso filogenético humano, la cuestión tan importante de si se puede concluir del proceso evolutivo una visión teleológica o azaro-sa dentro del mismo, de igual modo que en el conjunto de la historia del universo, haciendo referencia al denominado principio antrópico, en sus diferentes versiones (débil y fuerte) e interpretaciones. Como en otras dis-cusiones similares, advertimos también aquí la importancia decisiva de distinguir entre el punto de vista científico y el filosófico y teológico.
El proceso de hominización se completa y se enriquece en el ser hu-mano con el proceso de humanización, sobre el que nos detenemos en el capítulo quinto. El momento en el que el género humano se desgaja de los australopitecos está definido por la emergencia de una serie de cua-lidades mentales, como la autoconciencia, la capacidad simbólica e ima-ginativa, al mismo tiempo que un aumento considerable de la capacidad de construir y manejar herramientas, con lo que emerge de ese modo el mundo de la cultura. A partir de ese momento, el ser humano empie-za de forma progresiva a tomar las riendas de su propia humanización, quedando en cierta medida la evolución en manos del ser humano. Cada uno de los rasgos que componen la dimensión humanizadora del género humano se hallan ya apuntados en las especies animales cercanas, pu-diéndose poner en duda la diferencia cualitativa entre hombres y anima-les, pero si la comparación se realiza desde una visión conjunta, enten-demos que no queda ninguna duda de que con el género humano se ha producido la emergencia de una forma de vida cualitativamente distinta a las anteriores, conformada por una síntesis de biología y cultura que no se daba hasta entonces.
El capítulo sexto completa la visión de la antropogénesis con la di-mensión ontogenética y el punto de vista de la embriología. La mirada a la ontogénesis resulta cada vez más necesaria y significativa por el hecho de que la filogénesis, que durante mucho tiempo se ha desarrollado de es-paldas y al margen de la faceta ontogenética, está corrigiendo actualmente su autosuficiencia y sus carencias, contribuyendo a descubrir la necesidad de que vayan de la mano ambos enfoques. Pero también es fundamental el enfoque ontogenético por las decisivas cuestiones morales, legales y reli-giosas que en su ámbito se hallan planteadas. En la actualidad, todos sabe-mos la centralidad de las discusiones sobre el estatuto ontológico y ético del embrión humano, cuestión no sólo relevante en el entorno académico, sino también en el entorno legal y social. Un enfoque adecuado de estos temas representa una ayuda fundamental para abordar de forma adecuada los problemas éticos, con el apoyo de la antropología.

28
© Universidad de Deusto - ISBN 978-84-9830-383-4
Ya hemos indicado que el enfoque fundamental de todo el libro está encaminado a mostrar las características genéticas, morfológicas y cul-turales de la especie humana con el objetivo de mostrar su especificidad frente al conjunto de las demás especies vivas que conforman el conjunto de la biosfera. En el capítulo séptimo nos detenemos directamente a dilu-cidar esta cuestión, realizando un ejercicio comparativo entre el ser huma-no y el resto de las especies animales, en cuatro momentos centrales: ge-nética, morfología, embriología y estructura comportamental, para llegar a la conclusión que estamos defendiendo en el conjunto del libro: el géne-ro humano se halla en continuidad evolutiva con el resto de las especies, al mismo tiempo que se sitúa en un nivel nuevo, emergido en el mismo proceso evolutivo, que le dota de una diferencia cualitativa respecto a sus predecesores animales, diferencia que se apoya en último término en la capacidad de autoconciencia y de libertad.
En el capítulo octavo acometemos el estudio de la conjunción en el ser humano de sus dos dimensiones esenciales, la biológica y la cultural, presentando las cuatro posturas básicas que sobre esta temática se dan en la actualidad. La postura que consideramos más consistente es la que en-tiende que biología y cultura no son dos aspectos extrínsecos, sino que se hallan conformando en el ser humano una estructura única, en la que la cultura se une con la biología, pero reordenándola y absorbiéndola desde el nivel de lo psíquico. Por tanto, ni la cultura está subordinada a la biología, ni constituye una dimensión tan independiente y autónoma de ella como defiende un cierto tipo de culturalismo dualista. La especie humana consti-tuye una estructura bio-cultural, que se manifiesta en su ser y en su actuar, siendo nuestra sensibilidad inteligente y nuestra intelección sentiente.
El capítulo noveno y último viene a ser como un resumen conclusivo, donde exponemos las conclusiones que pretendíamos extraer del conjun-to de nuestras reflexiones, enunciadas ya en cierto modo en esta introduc-ción. Los rasgos biológicos y comportamentales que las ciencias nos apor-tan sobre la especie humana, nos dan suficiente apoyo argumental como para defender la singularidad del ser humano dentro del proceso evoluti-vo. Frente a quienes concluyen que las tesis darvinianas habrían demos-trado que las tesis humanistas y antropocéntricas han quedado obsoletas, entendemos que los datos científicos, si se interpretan adecuadamente, constituyen un apoyo necesario y suficiente para seguir manteniendo la diferencia cualitativa del ser humano frente al resto de las especies vivas. Diferencia que, como hemos repetido continuamente, no implica separa-ción. La diferencia ha de conjugarse con la continuidad, puesto que nues-tra singularidad no consiste en no ser ya animales, sino en serlo de otra manera, al estar dotados con una específica estructura bio-cultural en la que ambos elementos se necesitan y se potencian.

29
© Universidad de Deusto - ISBN 978-84-9830-383-4
Como hemos indicado al comienzo, un estudio tan amplio como el presente no puede aspirar a ser ni original, ni contener hasta el último dato de cada una de las disciplinas antropológicas sobre las que reflexionamos. Hemos tratado únicamente de aportar los elementos fundamentales para realizar una síntesis suficiente sustentadora de la tesis filosófica que es-tamos defendiendo sobre la específica y singular constitución esencial de la especie humana. Entendemos que puede ser una aportación valiosa al servicio de quienes, sin ser especialistas en estos campos, y posean un interés por estar al tanto de las aportaciones más recientes de las ciencias antropológicas, desean adquirir una visión realista y crítica sobre lo espe-cífico de la realidad humana. Pensamos que la aportación más provechosa de este libro está, más que en exponer novedades punteras en el terreno de las ciencias antropológicas, en acercarse de forma crítica a los datos que ellas nos presentan, de tal manera que se consiga analizar y superar las afirmaciones tendenciosas tanto de los cientifismos reduccionistas como de los creacionismos fundamentalistas. Porque si algo puede aportar una reflexión filosófica crítica sobre el ser humano (es la meta perseguida desde siempre por la Antropología filosófica, con mejor o peor acierto) es precisamente hacernos conscientes de la insuficiencia de las aportacio-nes científicas para dar cuenta de la realidad humana, y de la necesidad de conjugar las evidencias empíricas con las interpretaciones críticas de la filosofía, siempre plurales y necesitadas de contrastación. En esa contras-tación y discusión crítica entre las diferentes propuestas antropológicas es donde situamos nuestra propuesta de seguir defendiendo una visión huma-nista y antropocéntrica de la condición humana, sin necesidad de diluirla en la biología ni elevarla a una condición angélica insostenible.
En la medida en que, como hemos dicho, las aportaciones científicas sobre los diferentes aspectos de las realidad humana y animal resultan im-posibles de dominar y abarcar en su totalidad, aportamos al final del libro una amplia bibliografía para quien quiera completar aspectos que no se han tocado aquí con la suficiente amplitud, bibliografía que como es lógi-co no pretende ser exhaustiva, entre otras cosas porque, además de ser un imposible lograrlo dada la amplitud de publicaciones existentes sobre to-dos estos temas, ocuparía un espacio demasiado amplio para poder abar-carlo. Por ello, me he limitado casi en exclusiva a presentar simplemente la bibliografía que he utilizado en los diferentes capítulos del libro.
Los diversos materiales que constituyen estos nueve capítulos han sido utilizados en gran medida en varios de los cursos de filosofía impartidos tanto en la Universidad de Deusto como en la Universidad Centroameri-cana (UCA) José Simeón Cañas de San Salvador (El Salvador) a lo largo de estos últimos años, siendo corregidos y mejorados como consecuencia del diálogo con los alumnos de los diferentes cursos, a quienes expreso mi

30
© Universidad de Deusto - ISBN 978-84-9830-383-4
agradecimiento. Entre las personas a quienes quiero agradecer sus comen-tarios y aportaciones se encuentran Leandro Sequeiros, Roberto Aretxaga, Ángel García Larraz y Lorena Zuchel. Todos ellos han contribuido a me-jorar con sus críticas y aportaciones el texto inicial, muy distinto del que aparece publicado. Junto a ellos, han contribuido de forma muy especial a esta mejora, y les expreso mis agradecimientos más sinceros, Manuel Ca-bada y Sixto J. Castro, quienes desde su función de lectores críticos pro-puestos por la Universidad de Deusto, leyeron con detenimiento y empa-tía el borrador inicial y presentaron con sinceridad y acierto los cambios y las mejoras que resultaba necesario acometer. Junto a ellos, y de una forma especial, Manuel J. Heredia Noriega ha contribuido con su lectura meticulosa a superar en el texto defectos de estilo, repeticiones y errores tipográficos. Es de justicia agradecerle el tiempo dedicado a esta enojosa y necesaria tarea. Los errores e insuficiencias que el texto sigue teniendo, son únicamente responsabilidad mía.
Escribir un libro tan complejo como éste representa un esfuerzo impor-tante, realizado a lo largo de varios años de reflexión continuada. Por eso, aun conscientes de las limitaciones que contiene, nos daríamos por satisfe-chos si sirviera para un mejor conocimiento de la especie humana como realidad singular, y para una defensa inteligente de la misma frente a quienes pretenden diluirla como una cosa más entre el resto de las realida-des de nuestro mundo. Y esta pretensión es importante no sólo desde el punto de vista meramente teórico y académico, sino también por las con-secuencias de tipo ético y social que se hallan en juego, puesto que sólo desde la defensa de la singularidad ontológica y ética de nuestra especie parece posible seguir luchando por la dignidad de todos los seres huma-nos, en especial y sobre todo por aquellos situados socialmente por debajo de las capas más influyentes de la sociedad. En muchas ocasiones corren el peligro de que sus derechos estén situados por detrás (si no olvidados) de las mascotas de quienes están sobrados de recursos económicos y cul-turales. Nuestro proceso de humanización irá acercándose a unas metas dignas del ser humano cuando vayamos conformando una sociedad en la que todos los seres humanos tengamos igualdad real de derechos, sin dis-tinción de sexos, clases sociales, culturas y religiones, conseguido todo, claro está, sin olvidarnos del respeto a las condiciones de vida de los ani-males y del cuidado y sostenimiento del entorno ecológico, que nos con-tiene y sin el que nada somos ni podríamos existir.

31
© Universidad de Deusto - ISBN 978-84-9830-383-4
Capítulo 1
El entronque de la especie humana en la biosfera
1. LOS AVATARES DE LA AUTOCOMPRENSIÓN ANTROPOLÓGICA
Los avances tan espectaculares en el ámbito de las ciencias en ge-neral, y de las ciencias humanas en particular, en los dos últimos siglos sobre todo, han producido un vuelco significativo en la autocompren-sión del ser humano. De entenderse como una especie única y salida di-rectamente de las manos de Dios, hemos pasado a reconocernos como una especie más emergida del proceso evolutivo. Podemos afirmar que desde mitad del siglo XIX, con la aparición de la teoría de la selección de Darwin y los avances en el campo de la genética, tras G. Mendel y sus continuadores, se ha producido lo que podemos denominar un fuerte pro-ceso de biologización de la Antropología. De tal modo que en la actuali-dad se está extendiendo un modo de pensar que tiende a diluir la frontera entro lo humano y el resto de los animales. Se pretende quebrar con ello la común e inveterada tradición antropocéntrica del humanismo filosófi-co y religioso que ha dominado la totalidad de las culturas humanas. Los argumentos que se presentan no son triviales, y cobran una mayor fuer-za al calor de las exageraciones que la cosmovisión antropocéntrica ha mantenido sobre la superioridad de la especie humana y su pretendida condición de rey absoluto de la naturaleza. Pero este modo de pensar ma-terialista y reduccionista parece olvidar, o tal vez menospreciar, muchos aspectos o cualidades que conforman la especie humana, y que abogan claramente por una diferencia esencial entre ella y el resto de los seres integrantes de la biosfera.

32
© Universidad de Deusto - ISBN 978-84-9830-383-4
Por de pronto, el ser humano es el único ser vivo que se hace proble-ma de sí mismo, que ha tomado conciencia de su condición existencial, y se muestra crítico consigo mismo y con el resto de lo que existe. Esta capacidad la ha ejercido, de modo más o menos explícito y consciente, desde siempre, desde que empezó a ser homo, representando esa capa-cidad inquiriente el salto de lo no-humano a lo humano y constituyendo tal cualidad una de los distintivos clave de su especie. Es el salto de la naturaleza a la cultura, del determinismo biológico a la indeterminación de lo cultural. También es importante señalar que tal cualidad comporta-mental no constituye un lujo sino una necesidad. En la medida en que la biología no nos ha dotado de estructuras comportamentales fijas e ins-tintivas, no tenemos mas remedio que pensar cómo tenemos que actuar, qué alternativas entre las que se nos presentan tenemos que elegir, y, en definitiva, qué modelo de autorrealización queremos perseguir. Así, como dice Michael Landmann, cada anthropos se convierte necesaria-mente en un anthropo-logos, en un ser consciente y rector de su propia realización1.
Pero desde esos orígenes, los seres humanos han ido afinando su pro-pio retrato, pasando de una cripto-antropología a una antropología cons-ciente y explícita2, pero también sometida a continuos cambios y revisio-nes. Esos cambios podríamos sintetizarlos en los siguientes:
1) El paso de verse como una realidad más dentro del cosmos, a la necesidad de hacer de sí un objeto específico y autónomo de reflexión, así como de conformar una disciplina específica, la Antropología, que tenga por objeto propio y exclusivo la reali-dad humana. Durante la época griega y medieval, el ser humano constituía una parte más del conjunto de los saberes, constituyén-dose durante la Edad Media el tratado De Anima para el estudio del ser humano, junto a otros capítulos del mismo tratado dedi-cados a Dios, a los animales, y al resto de las demás creaturas naturales. De hecho, el nombre de Antropología no aparece has-ta el s. XVI (O. Casmann)3. Y no será más que a partir de I. Kant cuando se constituye la reflexión filosófica acerca del ser huma-no en un tema sustantivo y con pretensiones de ser el centro de la filosofía. Para Kant, las tres preguntas en las que sintetiza el sa-
1 Cfr. Landmann, Michael, Antropología filosófica. Autointerpretación del hombre en la historia y en el presente, México, UTEHA, 1961, p. 8.
2 Cfr. ibídem, pp. 10 y ss.3 El humanista protestante O. Casmann publicó en 1596 un libro que tenía por primera
vez el título de Antropología: cfr. Landmann, M., o.c., p. 1.

33
© Universidad de Deusto - ISBN 978-84-9830-383-4
ber filosófico, se resumen en una cuarta: ¿qué es el hombre? De ahí también que M. Foucault, cuando en su libro Las palabras y las cosas4 analice críticamente las posibilidades epistemológicas de una antropología filosófica desde la óptica del presente, pueda afirmar que el hombre es un invento reciente, así como decretar, desde la perspectiva de una nueva episteme estructuralista, que el hombre ha muerto.
2) Un nuevo horizonte de reflexión sobre lo humano ha consisti-do en pasar de una concepción temática de sí a otra problemáti-ca5. El ser humano comienza percibiéndose como un tema de re-flexión, con muchas lagunas desconocidas, pero seguro de poseer ya las bases fundamentales de su condición y de su existencia desde las raíces esenciales de la cosmovisión en la que se susten-ta. En cambio, a medida que ha ido estudiando y comprendiendo más hondamente los diversos aspectos de su realidad, se ha ido comprendiendo más hondamente como problema. Ahora bien, la problematicidad de la condición humana, como indica M. Buber, no es sólo una cualidad de la antropología contemporánea, sino que ha sido un rasgo que ha ido emergiendo de modo intermiten-te a lo largo de la historia de la filosofía occidental6. Pero es en el momento presente cuando más conscientemente habría asumido el ser humano su radical condición problemática, produciéndose la aparente contradicción entre el hecho de que nunca como hasta ahora el ser humano ha conocido más cosas acerca de sí mismo, al mismo tiempo de que nunca como hasta ahora se ha sentido tan problemático. Como indican tanto Heidegger como M. Scheler7, el ser humano no sabe lo que es y sabe que no lo sabe. Hemos pasa-do, pues, de entendernos desde una esencia ontológica ya sabida y constatada, a entendernos como una realidad abierta, en continuo proceso de realización y construcción desde nuestras propias de-cisiones libres.
3) Se ha producido también el paso de un estudio del ser humano desde el enfoque de la filosofía especulativa, a otro en el que co-bra cada vez más preponderancia el enfoque científico. Es el paso
4 Madrid, Siglo XXI, 1968.5 Cfr. García Bacca, J. D., Antropología filosófica contemporánea, Caracas, UCV,
1947, cap. 2.º; Morey, M., El hombre como argumento, Barcelona, Anthropos, 1987, p. 9; Beorlegui, C., Antropología filosófica. Nosotros: urdimbre solidaria y responsable, Bilbao, Universidad de Deusto, 2004 (2.ª ed.), pp. 45-49.
6 Cfr. Buber, M., ¿Qué es el hombre?, México, FCE, 1949. 7 Cfr. Heidegger, M., Kant y el problema de la metafísica, México, FCE, 1954; Scheler, M.,
El puesto del hombre en el cosmos, Argentina, Losada, 1938.

34
© Universidad de Deusto - ISBN 978-84-9830-383-4
de lo que se ha denominado la filosofía del hombre a las antropo-logías científicas, ya en pleno siglo XIX. También es verdad que todos los filósofos que han teorizado sobre el ser humano, han tenido en cuenta los datos y aportaciones de las ciencias de su tiempo, pero aparte de que el nivel técnico que las ciencias de esas épocas poseían era muy endeble, las aportaciones de las ciencias poseían un carácter más bien secundario y periférico. Y no podía ser de otra forma, puesto que, como vamos a ver en el apartado si-guiente, los rasgos que definían al ser humano eran de tipo espiri-tual y racional, terreno específico de la filosofía, y no tanto de tipo material y corpóreo, ámbito de las ciencias.
4) Un cuarto rasgo consiste en pasar de una concepción antropológi-ca basada en los aspectos racionales y espirituales (el ser humano es definido como animal racional, animal que tiene alma) a otra en la que se tienen más en cuenta, y en algunos casos predomina, los aspectos materiales y corpóreos. Se partía de una concepción dualista, en la que lo anímico se imponía de modo rotundo sobre lo corpóreo, que aparecía como algo a la postre complementario, y más todavía como un estorbo a controlar. No es que no se tuvie-ra en cuenta anteriormente lo corpóreo, sino que se consideraba como algo accidental y secundario. Ello suponía una concepción dualista de la condición humana, conformada por un alma y su cuerpo como dos sustancias diferentes y autónomas, aunque no todos los modelos antropológicos de esas épocas defendieran pos-turas dualistas. En el entorno de la cultura homérica, así como en la literatura bíblica de los semitas, el ser humano era concebido desde una radical unidad que se desplegaba en una pluralidad de perspectivas, que no suponían aceptar el dualismo ontológico8. De igual manera, el hilemorfismo de Aristóteles y de Santo Tomás entendía la realidad humana como compuesta por dos principios ontológicos inseparables, frente al dualismo de sustancias defen-dido por Platón y posteriormente por Descartes y sus seguidores. Para el filósofo francés, lo que define la esencia de lo humano es el alma en cuanto conciencia, definida como «una sustancia cuya única esencia y naturaleza toda es pensar», y «no necesita, para ser, de lugar alguno, ni depende de cosa alguna material; de suer-te que este yo, es decir, el alma por la cual yo soy lo que soy, es enteramente distinta del cuerpo y hasta más fácil de conocer que éste, y, aunque el cuerpo no fuese, el alma no dejaría de ser cuan-
8 Cfr. Gevaert, J., El problema del hombre, Salamanca, Sígueme, 1976.

35
© Universidad de Deusto - ISBN 978-84-9830-383-4
to es»9. Pero lo que constituye un giro esencial en la concepción actual del ser humano es la superación de la visión dualista acer-ca del mismo y la correspondiente acentuación de la importancia del cuerpo en la definición de lo humano. Con toda tranquilidad podemos decir ahora que el hombre es su cuerpo, aunque tal afir-mación se entienda de muy diferente manera desde un monismo fisicalista que desde una postura funcionalista, o desde el emer-gentismo. No es momento de detenernos en presentar las diver-sas teorías actuales sobre el clásico problema de las relaciones alma-cuerpo, o mente-cuerpo10, pero sí es importante señalar que las posturas más aceptadas en la actualidad desechan una idea de alma separada del cuerpo para aceptar (excepto los monismos re-duccionistas) una idea de mente o psique como estructura o modo específico de estar sistematizado el cerebro o el cuerpo humano.
Este conjunto de virajes o de giros profundos en la concepción del ser humano, especialmente el último descrito, se va produciendo como consecuencia del espectacular avance que experimentan las ciencias na-turales desde el Renacimiento, sobre todo la fisiología, la zoología, la medicina, la morfología comparada y el resto de las ciencias dedicadas al estudio de los rasgos físicos y de los comportamientos de los animales. Todos estos avances van mostrando la similitud física del cuerpo humano con el resto de las especies animales, y desembocando en la evidencia de la común naturaleza de las diferentes especies vivas, sin que pueda esca-par a esta común esencia biológica la especie humana. Todo iba apuntan-do hacia una concepción evolutiva de la biosfera, e incluso del conjun-to de la realidad cósmica, en la que todo conforma un conjunto unitario tanto en su composición material última como en su concatenación his-tórica. En la unificación de este modo de ver las cosas, y sobre todo en el cambio tan drástico que supuso para la concepción del ser humano, la teoría de la selección natural de Ch. Darwin ocupa un lugar preferencial y definitivo.
9 Descartes, R., De método, A. T., VI, 558 (tr. cast., Discurso del método, Madrid, Es-pasa-Calpe, 1937/1970 (12.ª ed.), p. 50; cfr. Meditaciones IX, 63 (tr. cast.: Meditacionesmetafísicas, Madrid, Espasa-Calpe, 1937/1970, 12.ª ed.).
10 Cfr. Gardner, H., La nueva ciencia de la mente (Paidós, Barcelona,1987; 2.ª ed.: 2000); Martínez-Freire, P. F., La nueva filosofía de la mente (Gedisa, Barcelona, 1995); Liz, M., Perspectivas actuales en filosofía de la mente (Gobierno de Canarias, Tenerife, 2001); Broncano, F. (ed.), La mente humana (Trotta, Madrid, 1995); Moya, C., Filosofíade la mente (Universitat de València, Valencia, 2004); Martínez-Freire, P. F. (ed.), Filoso-fía actual de la mente (Contrastes, Suplemento 6, Valencia, 2001). Cfr. Churchland, P. M., Materia y conciencia. Introducción contemporánea a la filosofía de la mente, Barcelona, Gedisa, 1999.

36
© Universidad de Deusto - ISBN 978-84-9830-383-4
2. LA BIOLOGÍA PRE-DARWINIANA
Aunque la biología se ha hecho popular y ha incidido de forma impor-tante en el ámbito del pensamiento a raíz de los avances y propuestas de Ch. Darwin, sin embargo tampoco sus teorías hubieran sido posibles sin las aportaciones anteriores11. Ya los presocráticos y los más importantes filósofos griegos, como Aristóteles, dedicaron su atención al estudio de los animales, pero desde una óptica estática. Aun así, sus trabajos fueron sentando las bases de las futuras clasificaciones de los seres vivos. Duran-te la Edad Media, se sigue con la misma concepción fixista de las especies vivas, interpretando la biosfera desde la óptica de la Biblia, entendida de modo literal.
Con el Renacimiento se da comienzo a las ciencias empíricas, produ-ciéndose grandes avances en el campo de la anatomía de la mano de Ve-salio. Desde la óptica religiosa predominante en la Europa cristiana, se consideraba al cuerpo humano como algo único, como parte de una per-sona humana, prohibiéndose cualquier tipo de investigación sobre el mis-mo. Pero poco a poco se fueron superando estas barreras, avanzando los estudios comparativos sobre las formas y estructuras corpóreas entre los diversos animales y el ser humano. Estos estudios van a despertar la idea de la variación de estas formas anatómicas en el tiempo, quedando en en-tredicho la noción fixista de especie. A esto se añadieron los importantes descubrimientos geográficos realizados en esa época, que traerían al mis-mo tiempo noticias de nuevas especies de plantas y de animales, así como el descubrimiento de los primeros fósiles. Las cuestiones que se plantea-ban los estudiosos se originaban del hecho de por qué estas desconocidas especies vivas se daban sólo en esos lugares y no en otros.
A partir del s. XVII es cuando propiamente se comienza la historia de la biología en sentido estricto y empírico, de la mano de figuras como el ita-liano Francesco Redi, que realiza la primera demostración de la no existen-cia de la generación espontánea (aunque la demostración definitiva la rea-lizará más adelante L. Pasteur), y el holandés Antony Van Leeuwenhoek, quien provisto de una rudimentaria tecnología microscópica descubrirá los glóbulos de la sangre, los infusorios y las bacterias. Ambos científicos plantearán la cuestión central sobre la relación entre la materia y la vida,
11 Cfr. Rostand, J., Introducción a la historia de la biología, Barcelona, Planeta, 1985; Radl, E. M., Historia de las teorías biológicas, Madrid, Alianza, 2 vols., 1988; Grasa Her-nández, R., El evolucionismo: de Darwin a la sociobiología, Madrid, Cincel, 1986; Ba-rahona, A./Ayala, Fr. J., El siglo de los genes. Patrones de explicación en genética, Madrid, Alianza, 2009; Buskes, Chris, La herencia de Darwin. La evolución en nuestra visión del mundo, Barcelona, Herder, 2009, cap. 1.º.

37
© Universidad de Deusto - ISBN 978-84-9830-383-4
produciéndose dos tipos de respuestas muy diferentes: la de los epigenéti-cos o transformistas y la de los preformacionistas o fijistas.
a) La postura fijista es la que predominará hasta el siglo XIX, defen-diendo la invariabilidad de las especies, de tal modo que, siguien-do la mentalidad bíblica, cada especie viva habría sido creada di-rectamente por Dios.
b) Los transformistas defendían, en cambio, que unas especies prove-nían de otras, negando la total independencia de unas especies res-pecto a las demás.
A pesar de sus contrapuestos planteamientos, ambas posturas reali-zaron aportaciones importantes a la historia de la biología, porque ambas tenían parte de verdad. Así, Linneo era creacionista y fijista, pero no le impidió realizar sus excelentes clasificaciones taxonómicas sin las que el transformacionismo no habría progresado. Y Buffon, defensor de la pos-tura transformacionista, aplica sus planteamientos sólo a las especies infe-riores, no a las superiores, ya que el transformismo era considerado como una degradación de las especies, no como progreso12.
Pero ya a partir del siglo XVIII se va generalizando la idea de progreso,como consecuencia del influjo del tiempo y de la historia en el ámbito del ser humano y de la realidad, aunque la sensibilidad por lo histórico como elemento constitutivo de la realidad no se impondrá hasta el siglo XIX. Por otro lado, el estudio de los diversos fósiles encontrados va produciendo en los científicos el convencimiento de la ampliación de la edad de la tierra, contraviniendo las deducciones bíblicas. Buffon llega a proponer la cifra de 70.000 años para la historia de la tierra, organizados esos años en siete etapas, hasta llegar a la aparición del hombre.
El primer científico que se puede considerar ya evolucionista fue J. Bta. Lamarck13. Es el primero que propone un sistema evolucionista más o menos coherente y completo, en obras como Historia de los anima-les sin vértebras (1815-1822), amplia obra de siete volúmenes precedidos por una introducción, y Filosofía zoológica (1809), donde desarrolla la primera teoría positiva de la evolución de los seres vivos14. Lamarck par-tía de dos convicciones de partida. La primera consistía en defender que los seres vivos se hallan distribuidos en una escala que va de los más sim-ples a los más complejos. Y la segunda idea consideraba que esta escala
12 Sobre Linneo y Buffon, cfr. Schwoerbel, W., Evolución, Barcelona, Salvat, 1986, pp. 56-60; Radl, E. M., o.c., vol. 2.º, pp. 1-6.
13 Cfr. Rostand, J., o.c., 79 y ss.; Grasa Hernández, R., o.c., 34 y ss.; Schwoerbel, W.,o.c., 60 y ss.; Radl, E. M., o.c., pp. 7 y ss.
14 Cfr. Rostand, J., o.c., 81 y ss.; Radl, E- M., o.c., pp. 11 y ss.

38
© Universidad de Deusto - ISBN 978-84-9830-383-4
no era completamente regular sino imperfecta. Y este conjunto de cam-bios que conforman esa escala de especies vivas, era consecuencia y fruto de la evolución.
Lamarck consideraba que la evolución era consecuencia de varios factores, el primero de los cuales era la tendencia de lo viviente a com-plejificarse, debido a una fuerza interna que tiende de forma constante a complicar la organización. Junto a esta fuerza complejificadora, actúa una segunda, que empuja a los seres vivos a la adaptación a su entorno, a las circunstancias. Para Lamarck, el medio actúa sobre los organismos creando necesidades, y los organismos se modifican para adaptarse al medio. De ahí que la vida no sea perfecta, sino que depende de la fuerza adaptadora a las circunstancias del medio. Por eso que J. Ruffié ha de-nominado a su planteamiento necesidad sin azar15. El tercer elemento de su teoría, por el que Lamarck es un referente en las teorías evolucio-nistas, es la transmisión a los descendientes de los caracteres adquiridos. De todos modos, esta idea no es original de Lamarck sino al parecer un tópico de la época, y tampoco la defendió con demasiada radicalidad. Sabemos, además, que el propio Darwin la defendió sin problemas en algunos momentos de su vida. El cuarto factor de la teoría lamarckiana es la generación espontánea, entendiendo que las especies se van rege-nerando permanentemente, renovándose continuamente los organismos más pequeños.
Lamarck también se ocupó de situar a los seres humanos en el proce-so evolutivo, no vacilando en colocarlos en parentesco con los animales. Para Lamarck, el hombre desciende de los animales en el ámbito de lo corpóreo, pero también defiende que el origen de la especie humana tiene que haberse producido por un sistema original y diferente al del resto de las especies animales.
A pesar de su importancia en la historia de la biología, sus ideas no tu-vieron demasiada acogida, por una cierta ingenuidad a la hora de explicar sus teorías, no probando nada de lo que defendía de forma seria, además de seguir todavía defendiendo teorías como la del flogisto. De ahí que en sus polémicas con el fijista Cuvier16, de más seriedad científica y de mayor prestigio, saliera siempre derrotado. Por otro lado, es posible que el mo-mento en el que Lamarck exponía sus ideas fuera todavía pronto para que la sociedad aceptase las ideas evolucionistas. De todos modos, la historia posterior no ha olvidado sus aportaciones y considera a Lamarck como un antecedente necesario y fundamental de las teorías de Ch. Darwin.
15 Cfr. Ruffié, J., De la naturaleza a la cultura, Barcelona, Muchnik Editores, 1982. 16 Cfr. Schwoerbel, W., o.c., 65-68.

39
© Universidad de Deusto - ISBN 978-84-9830-383-4
3. LA REVOLUCIÓN DE CHARLES DARWIN (1809-1882)
Lamarck había centrado su interés por la evolución en su aspecto o di-mensión temporal, es decir, se centró en la evolución vertical. En cambio, Darwin centró su interés en el problema de la diversidad, esto es, en el ori-gen de las especies, su diversificación en una dimensión geográfica. Se tra-taba, pues, de la evolución horizontal. Como ocurre en todo autor, las ideas de Darwin fueron fruto de múltiples influencias de otros autores, así como de circunstancias y experiencias de su propia vida. Estaba claro que la idea de la evolución estaba flotando en el ambiente cultural desde hacía algún tiempo, y Darwin había leído a los diversos autores evolucionistas, como Lamarck y su abuelo Erasmus Darwin, entre otros. Igualmente Darwin fue muy influenciado por autores como el geólogo Lyell, y por sociólo-gos como Malthus y Spencer. Todas estas influencias es lo que ha hecho a veces afirmar a determinados estudiosos que la originalidad de Darwin era poca o ninguna. Pero no cabe duda de que esa opinión no tiene nin-gún sentido, en la medida en que Darwin, como todo autor original, recoge elementos de otros y los sintetiza y estructura de forma nueva, y esa es su aportación específica. No en vano su teoría también fue consecuencia de sus experiencias directas, como las observaciones sobre la selección artifi-cial, y sobre todo las que fue recogiendo en su viaje en el Beagle.
3.1. El largo camino hacia la teoría de la selección natural
Sobre Ch. Darwin hay una bibliografía ya muy numerosa, tanto so-bre su vida como el desarrollo histórico de sus aportaciones17. Por eso,
17 Cfr. Darwin, Charles, Autobiografía, Barcelona, Edit. Alta Fulla, 1987; Id, Autobio-grafía, Pamplona, Editorial Laetoli, 2008; Bowler, Peter J., Charles Darwin. El hombre y su influencia, Madrid, Alianza, 1995; Burkhardt, F. (ed.), Cartas de Darwin (1825-1859); Des-mond, A./Moore, J., Darwin, N.York/London, W. W. Norton and Company, 1991; Tort, P. (dir.), Dictionnaire du darwinisme et de l’évolution, 3 vols., París, PUF, 1996; Tort, P., Para leer a Darwin, Madrid, Alianza, 2000; Mayr, E., Una larga controversia: Darwin y el darwi-nismo, Barcelona, Crítica, 1992; Hemleben, Johannes, Darwin, Madrid, Alianza, 1980 (2.ª ed.); Gould, S. J., The Structure of Evolutionary Theory, Cambridge (Mas.)/London, Harvard Uni-versity Press, 2002 (trad. cast.: La estructura de la evolución, Barcelona, Tusquets, 2004);Soler, Manuel, Evolución. La base de la biología, Granada, Proyecto Sur Ediciones, 2002; Young, David, El descubrimiento de la evolución, Barcelona, Edic. del Serbal, 1998; VV.AA., Evolución hoy, Barcelona, Tusquets, 2001; Chauvin, Rémy, Darwinismo. El fin de un mito, Madrid, Espasa-Calpe, 2001; Vallejo, F., La tautología darwinista y otros ensayos de biología, Madrid, Taurus, 2002; Sampedro, Javier, Deconstruyendo a Darwin, Barcelona, Crítica, 2002; Ruse, M., Charles Darwin, Buenos Aires, Katz Editores, 2008; Eldredge, Niles, Darwin. El descubrimiento del árbol de la vida, Buenos Aires, Katz Editores, 2009.

40
© Universidad de Deusto - ISBN 978-84-9830-383-4
no nos vamos a extender mucho exponiendo los detalles del proceso histórico de sus descubrimientos, sino sólo en los aspectos más signifi-cativos.
a) Los primeros pasos18
Darwin era hijo de un médico y quiso en principio seguir la profe-sión de su padre, pero sus desmayos al contemplar operaciones le obli-garon a cambiar de profesión y matricularse en teología, sirviendo esa circunstancia para entrar en contacto con el libro de W. Paley en el que presentaba la influyente teoría sobre el diseño, aceptada en principio por el joven Darwin. Pero tampoco el oficio de pastor anglicano le con-venció, dedicándose a la investigación de la naturaleza, su pasión desde niño. Esa afición es la que le llevó a presentarse a la plaza de naturalista que se ofertaba en la expedición cartográfica del bergantín Beagle, que la marina británica había preparado para cartografiar las costas de Suda-mérica. A lo largo de su viaje en el Beagle, que duró cinco años (1831-1836), Darwin fue observando la enorme diversidad de flora y de fauna de los lugares que iban visitando, y fue descubriendo también multitud de fósiles de animales desaparecidos, preguntándose por las causas de tal desaparición. Un ámbito ecológico clave fue el de las islas Galápagos, donde observó los parecidos entre tipos de tortugas, pinzones y otros mu-chos animales, dándole que pensar las diferencias de estas especies en las diferentes islas.
Pero Darwin volvió de su periplo marítimo sin tener todavía una ex-plicación y una síntesis de sus descubrimientos y teorías. Poco a poco, y bajo la influencia de diversos autores, irá progresivamente madurando su forma de pensar. Siguiendo las teorías geológicas de Lyell, se desmarca de los planteamientos catastrofistas, que sostienen que habrían desaparecido especies enteras en el pasado como efecto de catástrofes geológicas. Fren-te a ese modo de pensar, Darwin propugna más bien procesos gradualis-tas, evoluciones progresivas, tanto en la aparición como en la desaparición de las especies, de tal forma que considera que la evolución se va confor-mando como la suma de pequeños cambios, pequeñas causas pasadas que producen grandes efectos posteriores. De este modo, más que seguir un proceso de inducción, como acumulación de múltiples pruebas empíricas, para desembocar en su teoría de la selección natural, lo que hace Darwin
18 Cfr. Ayala, Fr. J., Darwin y el Diseño Inteligente, Madrid, Alianza, 2007, cap. 2.º; Sampedro, J. L., Deconstruyendo a Darwin, Barcelona, Crítica, 2002, cap. 1.º y 2.º; Tort, P., Para leer a Darwin, Madrid, Alianza, 2001.

41
© Universidad de Deusto - ISBN 978-84-9830-383-4
es ir poniendo a prueba y desechando (ensayo y error) diversas teorías y pistas faltas, hasta llegar a su posición definitiva19.
En su Autobiografía nos indica también Darwin que, en esa madura-ción de su teoría, le iluminó grandemente la lectura del libro de Malthus On Population. Pero todavía le faltaban varios datos para que se com-pusiera y encajara toda su teoría. Otros autores que le influyeron fueron A. Comte (Cours de Philosophie Positive) y sus ideas sobre el progreso en la evolución y en la historia; Adam Smith y sus teorías económicas; el estadístico Quetelet, ocupado con las ideas de Malthus sobre el creci-miento geométrico de la población frente al crecimiento aritmético de los recursos económicos. Todos estos autores fueron fundamentales para que Darwin llegara a conformar su teoría de la selección natural como motor del proceso evolutivo, de tal forma que «la extensión de la economía del “laissez faire” a la biología origina la teoría de la evolución mediante la selección natural»20. El orden armónico y estable, según A. Smith, surgi-rá de la criba y eliminación de los individuos. Lucha y competencia, que-dando los mejores. Así se hace posible el progreso.
b) La teoría de la selección natural
Lo novedoso del planteamiento de Darwin está en su renuncia a con-troles exteriores a la naturaleza, a leyes perfeccionadoras internas o a re-gularidades que operan sobre la totalidad. De este modo, Darwin enuncia una teoría de la adaptación local. Es decir, para él el orden de la natura-leza es un resultado accidental o consecuente con la lucha de los indi-viduos. Hacia 1839 Darwin tenía ya la base fundamental de su teoría21.En 1842 escribe ya un primer resumen breve, de unas treinta y cinco pá-ginas, que se convertirán dos años más tarde en un amplio volumen. En 1856, a sugerencias de Lyell, Darwin se decide a realizar una redacción más extensa. Pero lo que va a espolear a Darwin a escribir definitivamen-te sus planteamientos de cara hacerlos públicos es la lectura, en 1848, del escrito de Russell Wallace dando cuenta del descubrimiento por su parte
19 Cfr. sobre el método científico de Darwin: Ruse, M., Charles Darwin, Buenos Aires, Katz Editores, 2008.
20 Grasa Hernández, R., o.c., 43.21 Para la cronología completa de la evolución del pensamiento de Darwin, cfr. Tort,
Patrick, Para leer a Darwin, Madrid, Alianza, 2001, pp. 11-27. Cfr. Ayala, F., Darwin y el Diseño Inteligente, Madrid, Alianza, 2007, pp. 47 y ss.; Ruse, M., Charles Darwin, o.c.;Eldredge, N., Darwin. El descubrimiento del árbol de la vida, o.c.; Sampedro, J., Decons-truyendo a Darwin. Barcelona, Crítica, 2002, cap. 1.º; Fernández-Rañada, A., Los científi-cos y Dios, Madrid, Trotta, 2008, pp. 118 y ss.

42
© Universidad de Deusto - ISBN 978-84-9830-383-4
de la teoría de la selección natural. A la vista de ello, y con objeto de de-jar clara la paternidad de este descubrimiento en años anteriores, Darwin se decidirá a escribir pormenorizadamente sus ideas y la historia de sus descubrimientos, dando lugar a lo que será su famoso libro El origen de las especies (1859)22.
Al recibir la carta de Wallace dándole indicaciones de que tam-bién había descubierto por su cuenta la teoría de la selección natural que él había ya concebido desde años antes, pero que por prudencia no la había presentado públicamente, se pone en contacto con Wallace y acuerdan presentar al mismo tiempo sus investigaciones en la Linnean Society de Londres. Ambos autores son modelo de honestidad en el res-peto a la verdad y a los acontecimientos, puesto que también Wallace reconoció en esta comparecencia pública la primacía de Ch. Darwin en el descubrimiento de la teoría23. Y ahora es cuando Darwin se dispone a expresar abierta y explícitamente sus pensamientos, redactando en el transcurso de trece meses El origen de las especies, que aparecerá pu-blicado en 1859.
En resumen, el planteamiento de Darwin estaba articulado por una percepción dinámica del mundo, dentro de la cual las especies están cam-biando continuamente, mediando un proceso de transformación gradual y continuo, sin saltos bruscos (pero que el propio Darwin no sabe explicar). Entre las diferentes especies se da una comunidad de descendencia, a di-ferencia de los planteamientos de Lamarck, que defendía líneas evolutivas independientes. Para Darwin, los organismos y especies que mostraban semejanzas se debía a que estaban emparentados, en la medida en que te-nían antepasados comunes. Y el hombre también posee igual comunidad de descendencia que el resto de los animales, no representando ningún tipo de singularidad dentro del proceso evolutivo. De este modo, aunque el cuadro taxonómico de Linneo se mantenía en sus grandes líneas, el sen-tido del cuadro cambiaba totalmente, puesto que se pasaba de una figura estática a otra dinámica y evolutiva.
Ahora bien, este proceso evolutivo necesitaba un mecanismo que lo moviera y explicara, pues el cambio, según Darwin, no se da por simple azar ni tampoco por un impulso interno de tipo larmarckiano. Este fue el descubrimiento original de Darwin: el cambio evolutivo se debe a la se-lección natural, inspirada en las teorías de Malthus: frente a la sobreabun-dancia de vida y la escasez de recursos, sobreviven los más aptos. Pero
22 Cfr. Darwin, Ch., El origen de las especies por medio de la selección natural, Ma-drid, Alianza Editorial, 2009 (prólogo de Fr. J. Ayala, e introducción de Diego Núñez).
23 Para las relaciones entre Darwin y Wallace, cfr. Haken, H., Fórmulas de éxito en la naturaleza, Madrid, Salvat, 1986, cap, 7.º, 64 y ss.

43
© Universidad de Deusto - ISBN 978-84-9830-383-4
esta selección se produce por etapas, no por mero azar, de tal modo que se empieza produciendo una enorme cantidad de variabilidad de formas de vida, de diferencias intra-específicas, y sobre ellas actúa la selección, haciendo que sobrevivan los organismos y las especies más aptas. Así, los mejor dotados, en la medida en que presentan las variaciones más venta-josas, resultan favorecidos en el proceso de la lucha por la supervivencia en el proceso de adaptación al medio ambiente. De este modo, la evolu-ción es consecuencia tanto del azar como de la necesidad, de variaciones fortuitas y de la selección natural.
La principal laguna de la teoría darwiniana estaba en el desconoci-miento por parte de Darwin de los mecanismos del proceso genético de la diversificación de los rasgos que se transmiten por herencia. Como se sabe, esta era la aportación genial de las investigaciones de G. Mendel, que quedaron sin ser conocidas por la comunidad científica, y solamente medio siglo más tarde se redescubrirían y se darían a conocer, para con-formar, junto con la teoría de la selección natural, a mitades del siglo XX
la denominada teoría sintética de la evolución. Pero no adelantemos acontecimientos. Este desconocimiento de los avances de Mendel es lo que explica que Darwin pudiera defender en varios momentos, sobre todo al final de su vida, la teoría lamarckiana de la herencia de los caracteres adquiridos, entendiendo que había subestimado la importancia del medio en el proceso evolutivo, defendiendo la teoría de la pangénesis. Aunque también es cierto que esta teoría ocupa un papel más bien secundario den-tro de su hipótesis general.
3.2. El origen del hombre y las discrepancias con R. Wallace
Si en un principio las coincidencias fueron totales entre los dos co-descubridores de la teoría de la selección natural, posteriormente se irían mostrando las profundas discrepancias entre ambos. Los ámbitos de dis-crepancia se centraban en la selección sexual, la evolución de la especie humana y las teorías de Wallace (1823-1913) sobre el hiperseleccionismo.
Como se sabe, Darwin defendió la selección sexual como un elemen-to complementario de la selección natural, aunque no coincidente. La se-lección sexual orientaba la presencia de determinados rasgos en machos y hembras, que les daban ventaja de cara al apareamiento sexual y a la transmisión de sus rasgos genéticos en la generación siguiente. Pero la di-námica de esta selección podía originar una deriva que repercutía en ca-racteres perjudiciales para la supervivencia de tales individuos. Se produ-ce esta acentuación de rasgos tanto en los machos como en las hembras, aunque en cada uno de los dos sexos la lógica era diferente.

44
© Universidad de Deusto - ISBN 978-84-9830-383-4
Wallace se opuso a estos planteamientos, porque, según él, compro-metía la teoría general de la lucha por la vida, además de que subrayaba en demasía la intervención de los animales, en concreto de las hembras. Por lo visto, Wallace aceptaba los planteamientos de Darwin referidos a los machos, pero no en el caso de las hembras.
Pero la principal discrepancia se situaba sobre todo en la diferente for-ma de entender la incidencia del proceso evolutivo en la especie humana. Estas diferencias se refieren a las ideas expresadas por Darwin en sus li-bros El origen del hombre (1871) y La expresión de las emociones (1872). Wallace se negó siempre a admitir que el origen de la especie humana se debiera solamente al mecanismo de la selección natural, y más en concre-to la parte más elevada del ser humano, el cerebro. Entendía que los ele-mentos más específicos del ser humano, como son la capacidad intelec-tual y los demás elementos de la cultura, como la moralidad y la religión, y el cerebro como base fisiológica de los mismos, no podía venir sólo del proceso mecánico de la evolución sino de Dios.
Darwin argumentaba de forma contraria. Dada la capacidad de ex-plicación total de la selección natural, pensaba que había que defender el materialismo filosófico, a pesar de la prudencia con la que se refirió siem-pre a los temas religiosos. Para Darwin, el hombre procede de animales inferiores a él, y las capacidades intelectuales y morales también proce-den por evolución desde las mismas capacidades de los animales. En con-creto, las capacidades intelectuales provienen, según Darwin, del desarro-llo de la capacidad lingüística, y las capacidades morales se han originado de los instintos sociales y familiares. La naturaleza moral del hombre se explica como efecto del progreso de las facultades del raciocinio, la flexi-bilización de los afectos y simpatías, la educación, el ejemplo, la costum-bre y la reflexión, junto al instinto social adquirido por selección natural. Los planteamientos de Wallace le preservan de caer en el racismo, en el que Darwin está más propenso a caer, en la medida en que creía que todos los grupos humanos tienen las mismas capacidades intelectuales inna tas, de modo que el primitivismo se debe a la no utilización de cier-tas capacidades, pero no a su ausencia. De ahí que, para Wallace, todas las culturas humanas son iguales, pues su cerebro, que proviene de Dios, es en todos igual.
El tercer aspecto de discrepancia se refiere al hiperseleccionismo de-fendido por Wallace, según el cual cada parte del cuerpo serviría para una sola actividad y sería el resultado de un proceso selectivo específico. Darwin era en esto más flexible, en la medida en que consideraba que la selección natural modela órganos para una función o grupo de funciones, pero con un amplio abanico de posibilidades, y con unos límites que per-miten funciones polivalentes y abundantes interacciones. Estaba claro

45
© Universidad de Deusto - ISBN 978-84-9830-383-4
que, según la visión de Darwin, la naturaleza no funciona como un inge-niero que planifica con antelación y con lógica finalística sus obras, sino que procede más bien de forma imperfecta y «chapucera», con proyectos aproximados a partir de un número limitado de piezas24. La vida genera cambios al azar, y esas novedades sobreviven si son capaces de adaptar-se posteriormente al medio ambiente. En caso contrario, desaparecen. En este modo de entender las cosas radica la superioridad del planteamiento de Darwin. La selección natural, tal y como él la entendía, se opone a todo tipo de finalismo, entendido éste como la aceptación de una fuerza exte-rior y consciente que dirija la evolución.
Este rechazo del finalismo aleja también a Darwin de Lamarck, quien defendía la transmisión de los caracteres adquiridos, apoyándose en una teo-ría de la variación dirigida, lógica que sí se da en el ámbito de la cultura hu-mana, pero no en el de la genética. Darwin, como ya lo hemos señalado, ar-ticula su teoría basada en dos momentos: las variaciones al azar, aleatorias, y la selección del medio que opera sobre esa variación, permaneciendo los más aptos. Se dan, pues, dos lógicas: la darwiniana, que opera en el ámbi-to de la genética, y la lamarckiana, que funciona en el terreno de la cultura, donde ya se dan mentes conscientes que funcionan con lógica finalística. Si Lamarck tuviera razón, el ritmo de la evolución se acortaría notablemente. Pero las evidencias científicas no han logrado probar de momento los plan-teamientos lamarckianos, sino más bien lo contrario, aunque se han dado en varios momentos falsos anuncios en sentido contrario.
Nos hallamos, pues, ante dos lógicas para interpretar la evolución, cada una de ellas con distinto trasfondo filosófico: la lógica finalística, defensora de una racionalidad que podríamos llamar prospectiva, frente a otra lógica meramente retrospectiva, que funciona relacionando el azar y la selección natural. La primera funciona en el mundo de la cultura, pero no en el de la evolución biológica, en la que se da solamente la segunda.
3.3. El impacto del darwinismo
El impacto de las teorías darvinianas en su entorno social fue, como se sabe, muy fuerte y determinante, a la vez que muy variado. Mientras un grupo numeroso e importante de científicos e intelectuales aceptaron entu-siasmados sus conclusiones, otros presentaron diversas críticas a elementos parciales de la teoría, que Darwin tuvo en cuenta y con el tiempo corrigió y
24 Este concepto de selección natural desde la óptica del amateurismo chapuceroes el que resaltará en nuestro tiempo Fr. Jacob, en su Lógica de lo viviente, Barcelona, Laia, 1977.

46
© Universidad de Deusto - ISBN 978-84-9830-383-4
completó; y hubo también un tercer grupo, que se opuso radicalmente a la selección natural desde posturas filosóficas y teológicas aprióricas25.
La verdad es que los críticos tenían sobrados aspectos para indicar nu-merosas debilidades a la teoría de Darwin, puesto que se trataba de una hi-pótesis que, si bien contaba con diversas evidencias y una gran coherencia a su favor, también presentaba diversos flancos débiles. Entre ellos se adu-cían la falta de pruebas directas de la eficacia de la selección natural y del proceso de especiación; la falta de evidencia de especies transicionales entre otras dos conocidas; el problema del desarrollo por etapas, y de su utilidad en cada una de ellas, de órganos complejos como el ojo de los vertebrados; el no haber pruebas de que la edad de la tierra fuera tan larga como para ha-ber posibilitado la lentitud de la selección natural; la relación entre leyes de la herencia y la selección natural; la inexistencia de la herencia de los carac-teres adquiridos; etc. Como hemos indicado, Darwin salió al paso paciente-mente de estas objeciones, con la ayuda de Wallace y de T. Huxley, sirvien-do esta confrontación para limar y completar su teoría26.
Pero no nos vamos a detener en la discusión sobre estos detalles con-cretos que ponen en evidencia las insuficiencias y limitaciones de la teoría de la evolución y de la selección natural darviniana, que eran consecuen-cia de las insuficiencias de los avances científicos de esa época. Años des-pués, con el avance de los descubrimientos en geología, biología, genética y demás, se irían solventando varias de esas dificultades y conformada la denominada teoría sintética de la evolución, como veremos más adelante.
Más nos interesa aquí centrarnos en la oposición que tuvo Darwin desde planteamientos filosóficos y teológicos, y que mostraban las pro-fundas implicaciones y consecuencias cosmovisionales que la selección natural tenía para la comprensión de la imagen del hombre y del mundo que hasta entonces se tenía.
3.3.1. El darwinismo frente a la visión tradicional del hombre, de la realidad y de Dios
a) Nueva visión del hombre y de la realidad
Los avances de los saberes científicos en el campo de la zoología, la biología, la genética, etc., a partir del siglo XIX, en especial desde las
25 Cfr. Hemleben, Darwin, Madrid, Alianza, 1980 (2.ª ed.); Leith, B., El legado de Darwin, Barcelona, Salvat, 1986.
26 Sobre el diálogo de Darwin con sus críticos, en las ediciones posteriores de El origen de las especies, cfr. Tort, P., o.c., p. 42 y ss.

47
© Universidad de Deusto - ISBN 978-84-9830-383-4
aportaciones de Ch. Darwin, han ido mostrando de modo progresivo una evidencia hoy día incontestable, pero que con anterioridad ni siquiera se sospechaba: la procedencia por evolución de la especie humana desde el ámbito de la biosfera, y, por tanto, la familiaridad y comunidad del ser hu-mano con el mundo del resto de los animales.
Esta progresiva evidencia supuso, como es lógico, un cambio radical en el enfoque de los estudios sobre el ser humano. Hasta Darwin, el hom-bre se comprendía como un ser o una especie creada directamente por Dios, formada del barro de la tierra y animada por el aliento (espiritus)insuflado por Dios sobre él, según la interpretación literal del pasaje del Génesis (2,7). De ninguna manera se llegaba a sospechar, por tanto, que el ser humano constituía un eslabón más del proceso evolutivo, el último escalón aparecido en el continente animal. Hasta ese momento, el imagi-nario cultural humano se asentaba en una cosmovisión estática y religiosa, desde el supuesto de que «la vida fue plasmada concedida inmediatamen-te por Dios. Las diversas clases de plantas y animales existen según un plan divino. Por eso tienen una subsistencia que se halla por encima del tiempo y de la historia de los seres vivos individuales»27.
Pero a lo largo de s. XIX, tanto la geología como la morfología com-parada van aportando pruebas cada vez más evidentes de la unidad evolu-tiva del mundo de la vida, y, por tanto, el parentesco entre las diferentes especies vivas, imponiéndose poco a poco el paradigma de la evolución, aunque no siempre se entendiera de la misma manera28. La aportación de Darwin al proceso de explicación del proceso evolutiva con la teoría de la selección natural constituyó la pieza clave para la interpretación del ori-gen de los seres vivos, convirtiéndose la fecha de su teorización en un an-tes y un después en la historia de la Antropología físico-biológica y en la comprensión del ser humano en general.
La teoría de la selección natural de Darwin vino, pues, a significar un cambio drástico en el modo de entender la realidad humana, porque no
27 Häring, H., «La teoría de la evolución como megateoría del pensamiento occiden-tal», Concilium, 2000, n.º 284, 27-40; 27.
28 Cfr. Bowler, Peter J., Charles Darwin. El hombre y su influencia, Madrid, Alian-za, 1995; Tort, P. (dir.), Dictionnaire du darwinisme et de l’évolution, 3 vols., París, PUF, 1996; Mayr, E., Una larga controversia: Darwin y el darwinismo, Barcelona, Crítica, 1992; Gould, S. J., The Structure of Evolutionary Theory, Cambridge (Mas.)/London, Harvard University Press, 2002 (trad. cast.: La estructura de la evolución, Barcelona, Tus-quets, 2004); Soler, Manuel, Evolución. La base de la biología, Granada, Proyecto Sur Ediciones, 2002; Young, David, El descubrimiento de la evolución, Barcelona, Edic. del Serbal, 1998; VV.AA., Evolución hoy, Barcelona, Tusquets, 2001; Chauvin, Rémy, Darwi-nismo. El fin de un mito, Madrid, Espasa-Calpe, 2001; Vallejo, F., La tautología darwinista y otros ensayos de biología, Madrid, Taurus, 2002.

48
© Universidad de Deusto - ISBN 978-84-9830-383-4
sólo implicaba un cambio de modelo antropológico, sino también una re-volución paradigmática en lo cosmovisional, con fuertes repercusiones, en consecuencia, en el campo de la teología, de la geología, de la sociología, de la biología y de casi todos los ámbitos del saber. En el terreno de la an-tropología, suponía pasar de entender al ser humano como una estructura estática, salida directamente de Dios, al igual que el resto de las demás especies vivas, como queda plasmado en el modelo clasificatorio del na-turalista sueco Linneo, a entenderlo como el resultado de un proceso evo-lutivo surgido del conjunto de la biosfera, a través de mecanismos seme-jantes a los que han seguido el surgimiento en ella de las demás especies animales. En el campo de la geología sucedía algo similar, puesto que se va pasando (antes que en el campo antropológico) de una concepción es-tática de la Tierra y de todo el universo a una visión dinámica, en la que se entiende la realidad material como atravesada por un proceso dinámico del que, en un momento determinado, surgirá la vida en toda su paulatina riqueza y complejidad. Por tanto, cae por los suelos la máxima medieval «natura non facit saltus», para dar paso a una cosmovisión unitaria y di-námica, que concibe al universo como un paulatino dinamismo que abar-ca todo el cosmos, tanto la realidad material como los seres vivos, dina-mismo del que no queda excluido el ser humano.
b) El enfrentamiento religioso
Las tesis evolutivas de Darwin venían a quebrar y dejar como obsole-ta la teoría predominante, basada en lo que W. Paley había denominado la teoría del diseño. El conjunto de la biosfera no estaba ordenada según los planes de Dios, sino siguiendo el incontrolable cúmulo de circunstancias selectivas ambientales.
1. La teoría tradicional del diseño
Las grandes religiones han tratado siempre de explicar el origen del universo, de la tierra y de la vida dentro de ella, así como el origen de los humanos, como obra de un Dios omnipotente y omnisapiente. Desde los años posteriores a la época del destierro en la historia veterotestamentaria, así como desde los orígenes del cristianismo, se veía el ordenado diseño del universo como una prueba de la existencia de Dios y de su poder creador de todo lo que hay. Así lo expresan los relatos del Génesis, en el Antiguo Tes-tamento, como también los escritores cristianos, entre los que cabe destacar a San Agustín y a Santo Tomás y sus vías para acercarse a la existencia de Dios, siguiendo los pasos de Aristóteles. Su quinta vía es un claro ejem-

49
© Universidad de Deusto - ISBN 978-84-9830-383-4
plo del argumento a partir de la teoría del diseño. Este argumento se repitió de forma normal y reiterativa durante toda la Edad Media.
La denominada teología natural, basada en partir del orden y rasgos de la naturaleza mundana para desde ahí llegar hasta Dios, entendiendo estas reflexiones como pruebas demostrativas, fue rechazada ya por la Re-forma protestante, en la medida en que los seguidores de Lutero y Calvino consideraban que la naturaleza humana estaba corrompida, y, por tanto, era incapaz de alcanzar a través de ella y de sus leyes un conocimiento de Dios y de sus atributos. Una expresión moderna de este modo de ver las cosas es la denominada teología dialéctica del teólogo protestante Karl Barth, que advierte entre el mundo y Dios más un salto cualitativo y una ruptura que una continuidad y una cierta analogía, que pueda ser aprove-chada por la inteligencia humana para el acercamiento a Dios.
Pero, aun así, se seguía manteniendo la idea del diseño como la prue-ba más acabada y convincente de la acción creadora y ordenadora de Dios sobre el mundo, en la línea del deísmo. La primera formulación de la teo-ría del diseño, antes del siglo XVIII, se debe al clérigo naturalista inglés, John Ray (1627-1705), en su obra The Wisdom of God Manifested in the Works of Creation (La sabiduría de Dios manifestada en las obras de la creación), de 1691. Incluso Voltaire y el resto de los filósofos y científicos ilustrados aceptaron una concepción del mundo apoyada en el argumento de la teoría del diseño. Consideraban que así como un reloj, como expre-sión de una realidad compleja, ha necesitado ser hecho por un relojero, así también el mundo exigía necesariamente, para explicar su existencia y su forma de ser, referirse a la acción previsora y creadora de Dios, como in-teligencia suprema29.
2. William Paley (1743-1865) y su teoría del diseño
Pero el autor que ha quedado como referente de esta teoría es, sin duda, W. Paley, autor de Teología natural (1802), texto donde defendía ampliamente la pertinencia de este modo de ver las cosas como muestra inequívoca de la existencia de Dios. Tras una primera época en la que su interés se centró en la lucha por la abolición del comercio de esclavos, se dedicó posteriormente a los temas científicos y teológicos. Su obra central, Teología natural, o Evidencia de la Existencia y Atributos de la Divinidad, influyó fuertemente, como ya sabemos, en el joven Charles Darwin. El argumento central se apoyaba en la idea razonable de que «no puede haber diseño sin diseñador».
29 Cfr. Ayala, F. J., o.c., p. 32.

50
© Universidad de Deusto - ISBN 978-84-9830-383-4
W. Paley deducía la existencia de Dios a partir del obvio diseño de los seres humanos y de sus diferentes órganos. Su argumentación tenía dos partes: los organismos dan prueba de que son diseñados, y sólo un Dios omnipotente podría explicar la perfección, multitud y diversidad de los diseños. Para esta reflexión se fijaba sobre todo en la compleji-dad del ojo humano, en el que veía una gran similitud con el mecanismo del telescopio. En su libro va analizando las diferentes partes del ojo y las ve tan complejas y tan bien diseñadas, que deduce que no pueden ser una consecuencia de un mero azar, sino de un diseño previsto por un diseña-dor inteligente.
La fuerza del argumento de Paley contra el azar se basa, como indica Ayala30, en la idea de lo que llama relación. Es decir, en palabras de Paley, «cuando varias partes contribuyen a un efecto único o, lo que es lo mis-mo, cuando un efecto es producido por la acción conjunta de diversos ins-trumentos, el ajuste de dichas partes o instrumentos entre sí con objetivo de producir, por su acción conjunta, dicho efecto, es lo que yo llamo rela-ción; y donde esto se observa en las obras de la naturaleza o del hombre, me parece que conlleva una prueba decisiva de juicio, intención, arte»31.Por tanto, cuando se advierte ese ajuste relacional, es cuando tenemos que concluir que se da una intencionalidad, propia de una mente diseñado-ra32. Los productos del azar, por el contrario, no muestran relación entre sus partes, y no constituyen, por tanto, una complejidad organizada. Paley concluye de ello que la complejidad organizada y la función útil para la que se supone han sido construidas las realidades complejas evidencian y exigen un diseñador inteligente. Y la diversidad, la riqueza y la universali-dad de los diseños indican que sólo Dios, Creador omnipotente, podría ser ese Diseñador Inteligente.
De todos modos, la honradez intelectual de Paley encuentra dificul-tades para explicar las imperfecciones, los defectos y el dolor y crueldad tan abundantes en la creación, y hacerlas compatibles y coherentes con la noción de un creador del mundo. Por un lado, es evidente para Paley que ingenios tales como el ojo y el riñón no pueden ser resultado de principios o procesos naturales, al estilo de las leyes de la mecánica de Newton con las que explicaba los movimientos de los planetas. Esa realidad capaz de
30 Cfr. Ayala, F. J., o.c., pp. 35 y ss. 31 Paley, W., Natural Theology, Nueva York, American Tract Society, 1802, pp. 175-176.
Cita tomada de F. J. Ayala, o.c., pp. 36-37.32 Este argumento es similar al que recientemente denomina uno de los defensores ac-
tuales del Diseño Inteligente, M. Behe, complejidad irreductible, en su libro The Darwin’s black box, New York, The Free Press, 1996 (trad. cast.: La caja negra de Darwin, Madrid, Edit. Andrés Bello, 2000).

51
© Universidad de Deusto - ISBN 978-84-9830-383-4
diseñar, Dios, tienen que ser una persona (poseedora de conciencia y de pensamiento). Pero, por otro lado, también se le presentaban serias difi-cultades para explicar el sentido de determinados órganos o partes de los seres vivos que parecen innecesarios o superfluos, o incluso la presencia de órganos imperfectos y disfuncionales. Por tanto, Paley advierte que el estudio de la creación lleva tanto a apreciar los grandes aciertos y maravi-llas, como también errores e imperfecciones, como el punto ciego del ojo, o la abundancia de crueldad y sadismo entre los animales. A pesar de es-tas reflexiones, Paley no llega nunca a sacar la conclusión de que, si atri-buimos a Dios como Diseñador los aciertos de la creación, también ten-dríamos que atribuirle los defectos y errores.
Además de la obra de Paley, y centrando las reflexiones más allá del mundo de la biología, también por esa época se publicaron diversas obras sobre la presencia de la acción de Dios en un mundo estructura-do por las diversas leyes naturales. Una primera obra es un amplio tex-to, publicado como ocho tratados (1833-40), patrocinados por el octavo conde de Bridgwater, y dedicada a mostrar el poder, la sabiduría y la bondad de Dios tal y como se manifiesta en la creación. El geólogo Hugh Miller, en su obra The Testimony of the Rocks (El testimonio de las rocas),utilizó también el argumento del diseño, a partir de la belleza de la crea-ción, dato que manifiestaría claramente la intervención del Creador. Simi-lares reflexiones y argumentaciones utilizaba Charles Bell, en su tratado The Hand, Its Mechanisms and Vital Endowments As Evincing Design (La mano, sus mecanismos y dotes vitales como designio evidente), basándose en la admiración de la mano y del brazo humanos, así como de las alas de los pájaros, para llegar a la conclusión de que reflejan innegablemente la sabiduría y el poder de Dios33.
Charles Darwin dice en sus escritos autobiográficos que leyó de joven a Paley, y que le gustó y convenció. Pero, tras su viaje de cinco años en el Beagle, abandonó su teoría al descubrir una explicación científica alterna-tiva para el diseño de los organismos. Con esta explicación, que Darwin denominó selección natural, la ciencia dio un salto cualitativo.
3. La selección natural de Ch. Darwin34
El empeño central de la obra de Darwin, El origen de las especies,consistió en tratar de resolver el problema que representaba para Paley ex-plicar de manera científica el diseño de los organismos. Para Darwin, la
33 Cfr. Ayala, F. J., o.c., pp. 40-41. 34 Cfr. Ayala, F. J., Darwin y el Diseño Inteligente, o.c., cap. 2.º; Sampedro, J. L., De-
construyendo a Darwin, o.c. cap. 1.º.

52
© Universidad de Deusto - ISBN 978-84-9830-383-4
complejidad, diversidad y la existencia de estos maravillosos ingenios no son resultado de un diseñador inteligente, sino más bien de procesos na-turales.
La evidencia de la evolución surgía como consecuencia necesaria de la teoría de la selección natural tal y como la entendía Darwin. En los pri-meros capítulos de su obra principal, Darwin presenta pruebas de la selec-ción artificial de animales y plantas. La naturaleza aporta variaciones, y los cuidadores humanos seleccionan los mejores ejemplares. Pues lo mis-mo ocurre con el ambiente natural de modo espontáneo, seleccionando los rasgos de lo que Weismann denominaba el plasma germinal. Por tanto, son rasgos adaptados que proceden de un mecanismo que funciona como un diseño. Pero no es un diseño, puesto que para que sea tal se necesita una inteligencia que, desde un propósito previo, orienta la acción creado-ra; pero en el caso de la naturaleza, se trata de un proceso espontáneo, sin inteligencia, y cuya racionalidad se ve sólo a posteriori.
Darwin acepta que los seres vivos y sus órganos están diseñados para ciertos cometidos, para determinadas funciones, y, por ello, están adapta-dos para determinadas formas de vida. Pero tanto esas formas con sus res-pectivas funciones no las ha dibujado ningún diseñador inteligente, sino que tienen su explicación de forma natural, desde una racionalidad retros-pectiva, no prospectiva (García Bacca). Y este modo de entender el pro-ceso evolutivo no lo reducía Darwin al mundo de los animales, sino que lo extendía también al del género humano, a diferencia de Alfred Wallace (1823-1913), el otro descubridor de la teoría de la selección natural, con el que discrepaba acerca de la inserción de la especie humana en el proce-so evolutivo.
No es, pues, de extrañar que este cambio de paradigma cosmovisional, fuera visto como una seria amenaza a la cosmovisión religiosa judeocris-tiana, puesto que suponía romper con la hasta entonces imperante concep-ción de una relación milagrera de Dios respecto al mundo, y propugnar una consecuente autonomía de las leyes del universo, no necesitado, por tanto, de la intervención directa y permanente de Dios para ir configuran-do el mundo en su admirable pluriformidad y orden. No es, por tanto, ex-traño que el planteamiento darwiniano tuviera en sus comienzos una opo-sición dura, visceral y sistemática, desde múltiples ámbitos de la sociedad, en especial desde el estamento eclesial35, siendo paradigmático el enfren-
35 La bibliografía sobre el rechazo desde ámbitos religiosos de la evolución y de la se-lección natural es muy amplia. Para una visión global del problema, cfr. Barbour, I. G., Re-ligión y ciencia, Madrid, Trotta, 2004; Id., Problemas de religión y ciencia, Santander, Sal Terrae, 1972; Evolución y fe, Concilium, 2000, n.º 284 (número monográfico); Núñez, Die-go, «La religión y la ciencia (Historia de las controversias entre ambas)», Mundo Científi-

53
© Universidad de Deusto - ISBN 978-84-9830-383-4
tamiento verbal tenido en 1860 (al año siguiente de la publicación del Ori-gen de las especies de Ch. Darwin), en la reunión de la Asociación Britá-nica para el Progreso de la Ciencia, entre el obispo Samuel Wilberforce y el amigo de Darwin, Thomas Huxley36.
En realidad, las reacciones de los creyentes ante la teoría de la se-lección natural de Darwin fueron muy variadas. Es cierto que los sec-tores más tradicionales se opusieron frontalmente por las razones que vamos a indicar, pero también es cierto que los sectores más abiertos y liberales defendieron desde el principio la compatibilididad de la fe cris-tiana y la teoría darwiniana, indicando que el cristianismo no implica defender la creación directa por parte de Dios de cada especie separada, sino que la afirmación central de la cosmovisión cristiana es que todo lo que hay debe su existencia a la acción creadora de Dios, sea cual sea el modo como entendamos el proceso de creación. Incluso algunos teólo-gos protestantes consideraban que el proceso evolutivo es el modo como Dios se ha servido para crear las diversas especies vivas. Y lo mismo que no nos impide creer en un Dios creador y ordenador del universo la de-fensa de la autonomía del movimiento de los planetas siguiendo las le-yes descubiertas por los cosmólogos, tras el paso del geocentrismo al heliocentrismo, de igual modo cabe compaginar la acción de Dios y las autonomía natural de la biosfera. En esta línea se entienden las afirma-ciones, en 1885, de A. S. Strong, presidente del Rochester Theological Seminar del estado de Nueva York: «Aceptamos el principio de la evo-lución pero la consideramos sólo como el método elegido por la inteli-gencia divina»37.
De todos modos, es razonable entender la oposición de los sectores más conservadores, en la medida en que, como hemos visto, la teoría de la selección natural suponía un golpe muy importante a los principios bá-sicos de la religiosidad tradicional, basada en una concepción de la acción de Dios sobre el mundo de forma directa y milagrera. La teoría darwi-niana atacaba esa forma tradicional de entender la acción de Dios en tres puntos fundamentales. El primero se refiere a la función del azar en el proceso de la transmisión de los rasgos genéticos, en contraste con la vi-sión cristiana de un mundo ordenado por Dios desde un plan sabio y pro-
co, 1996, n.º 166, 247-256; Gutiérrez Lombardo, Raúl, «La teoría de la evolución y la Igle-sia Católica», Ludus Vitalis, X (2002), n.º 17, 111-122; Riaza, José María, La Iglesia en la historia de la ciencia, Madrid, BAC, 1999, Nicolau Pous, Fr., Iglesia y ciencia a lo largo de la historia, Barcelona, Ed. Claret, 2002 (Ed. SCIRE, 2003).
36 Cfr. Polkinghorne, John, Ciencia y Teología. Una introducción, Santander, Sal Terrae, 2000, pp. 19-21; Fernández-Rañada, A., o.c., 121 y ss.
37 Cita tomada de ibídem, p. 122.

54
© Universidad de Deusto - ISBN 978-84-9830-383-4
videncial. Aunque Darwin no conocía el modo azaroso como se producen las combinaciones genéticas (puesto que no conoció las aportaciones de Mendel, y menos aún los descubrimientos posteriores de la genética sobre las mutaciones), consideraba que la selección natural actuaba sobre la re-producción de los caracteres biológicos transmitidos de una generación a otra a través de un proceso azaroso.
El segundo de los motivos de rechazo se refiere a la negación que la selección natural realiza sobre la teoría del diseño. Para Darwin, como ya lo hemos visto, el proceso evolutivo no se debe a un diseño salido de una mente inteligente y consciente, sino a una racionalidad a posteriori depen-diente de las circunstancias del ambiente. La naturaleza parece que tiene la capacidad de simular un diseño, pero no lo es tal, al menos consciente y a priori. Simplemente la estructura genética produce de forma azarosa nuevas formas de vida, y el ambiente las selecciona, perviviendo las más aptas a las circunstancias de ese ambiente. Este modo de actuar parece si-mular un propósito, pero no es tal. Como indica Fernández-Rañada, «la naturaleza sería como una persona que se pierde al avanzar con los ojos vendados por un bosque espeso, cambiando constantemente de rumbo por tropezar contra los árboles. Si al final llega a un destino, eso no significa necesariamente que hubiese pretendido llegar allí»38. En este modo de ver las cosas, parece que la acción de Dios está de sobra, y la naturaleza se nos aparece como totalmente autosuficiente.
Y una tercera razón del rechazo se debe a las teorías de Darwin sobre el origen de las leyes morales, origen que para él había que buscar no en algo distinto al proceso evolutivo, en contra de las tesis de Wallace y de otros autores39. Pero Darwin no defendió el relativismo moral, como algu-nos han sostenido, sino que consideraba que los valores morales van apa-reciendo a lo largo del desarrollo evolutivo de los instintos sociales. Pero, como indica Fernández-Rañada, «esta opinión era muy difícilmente acep-table para muchos sectores religiosos porque implicaba la posibilidad de una ética independiente de la teología»40.
Estas razones son las que explican que se extendiera pronto la con-vicción, alentada tanto por los creyentes conservadores como también por los intérpretes más radicales del darwinismo, de que el cristianismo y el darwinismo son incompatibles, situando el punto de mayor conflicto e in-compatibilidad en la antítesis entre el diseño y el azar, con las consecuen-
38 Ibídem, p. 123.39 Cfr. Darwin, Ch., The Expression of the Emotions in Man and Animals, London,
John Murray, 1872 (trad. cast.: La expresión de las emociones en los animales y en el hom-bre, Madrid, Alianza, 1998).
40 O.c., p. 124.

55
© Universidad de Deusto - ISBN 978-84-9830-383-4
cias que llevaba sobre la falta de destino y la imposibilidad de configurar una teoría sobre el sufrimiento y el mal, en la medida en que surgirían por puro azar y por la voluntad de los seres humanos, y no por voluntad divi-na41. Lo curioso es que Darwin no se consideró nunca ateo, sino agnósti-co, y siempre fue muy cuidadoso en sus afirmaciones sobre las implica-ciones de su teoría sobre las verdades religiosas. Todo parece indicar que la muerte temprana de su hija mayor Annie, a los diez años, fue un factor que le alejó de la religión, haciéndole perder la fe en un Dios bueno que cuida providentemente de sus criaturas42. A pesar de ello, en carta a su amigo Asa Gray, reflexionando sobre el bien y el mal en el mundo, afir-maba que «no puedo mirar a este universo maravilloso, especialmente a la naturaleza humana, y concluir que todo es producto de la fuerza bruta. Me inclino a pensar que todo resulta de leyes diseñadas, con los detalles, buenos o malos, dejados a la suerte de lo que podemos llamar azar. Siento muy profundamente que estas cosas son demasiado difíciles para la inteli-gencia humana»43.
Aquí puede verse la honestidad con la que Darwin reflexionaba so-bre estas hondas cuestiones acerca del sentido de la existencia humana, no considerando incompatibles la evolución y la fe religiosa, como se dedu-ce de estas palabras escritas por él tres meses antes de morir: «Me parece absurdo dudar que un hombre pueda ser, a la vez, un teísta ardiente y un evolucionista (…). Contestando a su pregunta le diré que mi opinión fluc-túa a menudo. En las fluctuaciones más extremadas, no he llegado nunca a ser un ateo, en el sentido de negar la existencia de un Dios. Creo que en general (más y más según me hago viejo), aunque no siempre, la descrip-ción más correcta de mi postura es la de agnóstico»44.
En la actualidad, la conciliación entre la teoría de la evolución y la fe cristiana es un hecho generalizado, excepto entre los defensores del crea-cionismo fundamentalista45. Si para muchos científicos el hecho de la evo-
41 Para un estudio panorámico y amplio sobre el conflicto entre darwinismo y cris-tianismo, cfr. Ruse, M, ¿Puede un darwinista ser cristiano?, Madrid, Siglo XXI, 2007; Gould, S. J., Ciencia versus religión, Barcelona, Crítica, 2007; Barbour, I. G., El encuentro entre ciencia y religión. ¿Rivales, desconocidas o compañeras de viaje?, Santander, Sal Terrae, 2004; Id., Religión y ciencia, Madrid, Trotta, 2004.
42 Cfr. Gould, S. J., Ciencia versus religión, o.c., pp. 33 y ss. 43 Cita tomada de Fernández-Rañana, A., o.c., p. 184. 44 Cita tomada de ibídem, p. 184. 45 Cfr. Davies, M. W., Darwin y el fundamentalismo, Barcelona, Gedisa, 2004; Drees,
Willem B., «Creacionismo y evolución», Concilium, 2000, n.º 284, 55-64; Sequeiros, L., Raíces de la humanidad. ¿Evolución o creación?, Santander/Madrid, Sal Terrae/Fe y Se-cularidad, 1992; Gomis, A./Josa, J., «Creacionismo y evolución. Imágenes de la polémica darwinista en España», Mundo Científico, 2002, n.º 233, pp. 20-29.

56
© Universidad de Deusto - ISBN 978-84-9830-383-4
lución suponía prescindir de toda referencia a instancias divinas, para los creyentes suponía simplemente la necesidad de cambiar el modo de enten-der la realidad de Dios y su relación con el mundo. Tal es el caso de ecle-siásticos contemporáneos de Darwin, como Frederick Temple y Charles Kingsley, para quienes era evidente que había que aceptar las afirmacio-nes confirmadas por las ciencias, pero entendiendo que «la selección na-tural estaba relacionada con el “cómo” de la acción creadora de Dios», y defendiendo que «la evolución reemplazaba el concepto de acción instan-tánea del Creador por la más sutil y satisfactoria idea de una creación que, una vez implantada en el ser, era capaz de ir haciéndose a sí misma»46.Hace unos pocos años, el Papa Juan Pablo II afirmaba oficialmente que el evolucionismo es conciliable con el dogma católico, dos puntos de vista «que parecían irreconciliables»47. En realidad, esta declaración del Papa venía simplemente a confirmar lo que, desde hace tiempo, constituye una doctrina teológica normal entre los teólogos oficiales de las más impor-tantes confesiones cristianas.
3.3.2. Oposición desde la filosofía idealista
Pero no sólo la oposición al darwinismo, por lo que suponía una re-volución cosmovisional, provenía del campo religioso, sino también del campo filosófico idealista. El pensamiento filosófico occidental, cuya máxima figura había sido Hegel, completando sus planteamientos con las teorías historicistas, estaban basadas en un teleología cósmica, en una ra-zón universal que suponía una evolución dirigida conscientemente y no sometida a la mera selección natural por parte del ambiente. Del mismo modo, se habían impuesto en esa época las teorías ortogenéticas, que de-fendían la existencia de la evolución de líneas filéticas, siguiendo una ruta rectilínea predeterminada, cuya dirección no podía estar orientada por la selección natural.
Por tanto, en el ámbito filosófico y científico el oponente teórico fun-damental era el esencialismo, presente en el pensamiento occidental desde el presocrático Parménides, aunque ya estaba decayendo desde la segunda mitad del s. XVIII con el auge de los historicismos y de las diversas filoso-fías de la historia. Este esencialismo daba por hecho que las especies te-nían ya una estructura fija, inamovible, y que se transmitía de generación
46 Ibídem, p. 20. Las comillas son del autor. 47 Juan Pablo II, «Mensaje a los miembros de la Academia pontificia de ciencias, reu-
nidos en Roma para su asamblea plenaria (22-10-96)», Ecclesia, 16 de noviembre de 1996, n.º 2.815, pp. 25-26.

57
© Universidad de Deusto - ISBN 978-84-9830-383-4
en generación desde sus orígenes; y esas esencias fijas que caracterizaban a cada especie, provenían directamente de la creación divina, o bien de un mundo platónico compuesto por formas puras.
Los planteamientos de Darwin suponían una visión muy diferente, entendiendo que los organismos vivos se caracterizan por su singulari-dad, y dentro del conjunto poblacional que forma una especie, los valores medios y comunes sólo son abstracciones. Además, si unas especies pro-ceden de otras, a través de lo que la genética de poblaciones ha denomi-nado deriva genética, en los primeros momentos de la conformación de una nueva especie, a partir de otra, la diferencia entre los individuos que inician la separación de las dos especies es pequeña e inapreciable. Así, pues, el concepto de especie no obedece a una noción de esencia estática, sino que es un proceso evolutivo en permanente transformación. La disputaentre saltacionistas y gradualistas (postura defendida por Darwin) estaba mezclada con la discusión sobre el esencialismo, de tal forma que el sal-tacionismo venía a ser una nueva versión del esencialismo. Veremos más adelante que esta discusión no está cerrada del todo.
Por tanto, admitir los planteamientos de Darwin suponía al pare-cer admitir el reduccionismo materialista, y hacer desaparecer a Dios del ámbito de lo mundano, en la medida en que el proceso evolutivo, y el origen y la transformación de las diversas formas de vida, se debía ex-clusivamente a mecanismos inmanentes a la propia naturaleza. No es de extrañar que el darwinismo fuera favorable y fervientemente acogido por filósofos importantes de la época como Marx, Engels, Nietzsche, Kro-potkin, Bergson, Unamuno y otros muchos. Marx y Engels expresaron en diversas cartas intercambiadas entre sí que el darwinismo era un «golpe de gracia a la teleología», y Marx intentó dedicar el volumen I de El Ca-pital a Darwin. Esta orientación materialista influyó posteriormente de tal modo en el avance de la biología que, como afirma Kolakowski, se tendió a partir de entonces a «consolidar una imagen del mundo en que todas las situaciones de la vida humana podían reducirse a situaciones biológicas»48.
La influencia en Nietzsche se dio a través del socialdarwinista Lange, quien popularizó los conceptos lucha por la vida y supervivencia del más apto. Estos planteamientos le ayudaron a Nietzsche a oponerse a la ética de la compasión de Schopenhauer y a sustituirla por el derecho de los fuertes y la primacía del superhombre. Igualmente tuvo un fuerte im-pacto en el pensador ruso Kropotkin (1842-1921), pensador anarquista opuesto al darwinismo social, quien en su libro Ayuda mutua: un factor
48 Cita tomada de Grasa Hernández, R., o.c., 67.

58
© Universidad de Deusto - ISBN 978-84-9830-383-4
de la evolución49 expuso una visión crítica y corregida de la interpreta-ción tradicional de la selección natural en clave de violencia individualista. Para Kropotkin, la versión individualista y agresiva de la selección natural no es más que el lastre ideológico que se ha colgado a la teoría de Darwin como consecuencia de la influencia del darwinismo social de Spencer. La experiencia muestra, según el escritor ruso, que el más apto en el proceso de adaptación al medio no es siempre el más agresivo sino el más coope-rativo. Por tanto, la ayuda mutua es un factor de adaptación tan eficaz o más que el egoísmo agresivo. «Ayuda mutua, justicia, moralidad: tales son las etapas subsiguientes que observamos al estudiar el mundo animal y el hombre. Constituyen una necesidad orgánica que lleva su justificación en sí misma, que vemos confirmada en todo el reino animal (…). Los senti-mientos de ayuda mutua y de justicia y de moralidad están arraigadas hon-damente en el hombre con toda la fuerza de los instintos. El primero de ellos —el instinto de Ayuda Mutua— aparece como el más fuerte»50.
También el filósofo francés Henri Bergson creó una versión original del darwinismo, rechazando el mecanicismo y admitiendo un finalismo parcial, en su conocida obra La evolución creadora51. Admite Bergson el origen común de todas las variedades de vida, incluida la humana, como una granada que se abre y estalla en fragmentos. Pero entiende que el hom-bre es el término y la finalidad de la creación. Para Bergson, la fuerza de la evolución lleva dos caminos: el del instinto, cuya mayor expresión se da en los insectos, y la de la inteligencia, cuya cumbre está en el hombre. En los demás caminos, la fuerza de la evolución o èlan vital parece que termina en un callejón sin salida. El resto del mundo es el humus sobre el que de-bía nacer el ser humano. Por tanto, para Bergson, la evolución se nos pre-senta como la historia de las victorias de la vida sobre la materia.
En el ámbito de la intelectualidad española también tuvo Darwin un gran impacto, sobre todo en los intelectuales de la denominada generación del 98, con Unamuno a la cabeza52.
49 Kropotkin, Peter, Mutual Aid: A factor on evolution, Londres, 1902 (Trad. cast. de Carlos Díaz, El apoyo mutuo. Un factor de la evolución, Bilbao, Edit. Zero ZYX, 1970 (2.ª ed.: 1978).
50 Kropotkin, P., La moral anarquista, Madrid, 1977, 73-74. Cfr. Montagu, A., ¿Qué es el hombre?, Buenos Aires, Paidós. 1966.
51 Madrid, Edic. Austral, 1912.52 Cfr. Girón Sierra, A., Evolucionismo y anarquismo en España. 1882-1914, Madrid,
CSIC, 1996; Id., En la mesa con Darwin. Evolución y revolución en el movimiento li-bertario en España (1869-1914), Madrid, CSIC, 2005; Núñez, Diego, Introducción a Ch. Darwin, El origen de las especies por medio de la selección natural, Madrid, Alianza, 2009, pp. 26-33; Gomis, Alberto/Josa, Jaume, Bibliografía crítica ilustrada de las obras de Darwin en España (1857-2008), Madrid, CSIC, 2009.

59
© Universidad de Deusto - ISBN 978-84-9830-383-4
3.3.3. Impacto del darwinismo en el ámbito sociológico y económico
Pero no sólo la teoría de la evolución darwiniana tuvo un fuerte im-pacto en el terreno religioso y filosófico, sino que también lo tuvo en el ámbito sociológico y económico. El denominado darwinismo social gozó de una gran influencia a finales del siglo XIX y comienzos del XX, no sólo por su pretensión de fundamentar biológicamente las ciencias sociales, sino sobre todo porque, en su versión conservadora, representaba la legiti-mación del liberalismo económico y el primitivo y más feroz capitalismo industrial.
El autor más representativo de esta corriente fue H. Spencer. Antes de conocer El origen de las especies de Darwin, tenía ya elaborada su teoría social en su libro Social Statics (1850), donde acuña la expresión lucha por la existencia, expresión que, como se sabe, no es de Darwin ni nunca la aceptó53. Desde posturas ultraliberales, H. Spencer defendía que la mecánica de la selección natural se aplicaba también en el terre-no de la cultura humana, pudiéndose aplicar a ella la «lógica» del mundo de la biología. De ahí que se haya aplicado a sus teorías sociológicas el apelativo de «darwinismo social». La verdad es que Spencer corrigió en parte la teoría darwiniana, puesto que agudiza dentro de la lógica de la selección natural la «supervivencia del más fuerte». En realidad, hay que aclarar que la teoría spenceriana no se origina en Darwin, sino que su concepción evolutiva de la historia humana y de toda la realidad es fruto de la cosmovisión evolucionista imperante en aquella época en Europa occidental, sirviéndole la teoría de la selección de Darwin, interpretada de forma muy personal, como útil apoyo teórico, escorando su interpre-tación de modo indebido desde parámetros exclusivamente competitivos, cuando se ha observado que no siempre es más eficaz para la superviven-cia la competencia a muerte sino la cooperación, como hemos visto más arriba de la mano de Kropotkin. De ahí que haya quien considere que más lógico que denominar la teoría spenceriana de darwinismo social,habría que denominarla spencerismo social, y a la de Darwin, de spence-rismo biológico54.
53 Cfr. sobre la relación entre el darwnismo social de Spencer y Darwin, Tort, P., Paraleer a Darwin, o.c., 63 y ss. Cfr. Gutiérrez, A./Ortega, R., «Ciencia e ideología en una po-sible antropología darviniana: una crítica a Ptrick Tort» (manuscrito).
54 Cfr. Lewontin, R. C./Rose, S./Kamin, L. J., No está en los genes. Racismo, genética e ideología, Barcelona, Crítica, 1987; Kropotkin, P., Mutual Aid, Boston, Porter Sargent, 1955 (trad. cast.: El apoyo mutuo. Un factor de la evolución, Bilbao, Zero ZYX, 1970); Beorlegui, C., Antropología filosófica. Nosotros: urdimbre solidaria y responsable, Bilbao, Universidad de Deusto, 1999, 2004 (2.ª), cap. 3, pp. 144-191.

60
© Universidad de Deusto - ISBN 978-84-9830-383-4
Spencer, como defensor del liberalismo, era partidario de la propie-dad privada y de la libre empresa, y contrario a la intervención estatal en el ámbito económico para corregir las desigualdades, así como crítico del socialismo y del comunismo. Refutando a Malthus, entiende que el exce-so de población y la falta de recursos se puede superar con el desarrollo de la inteligencia. Fue quien acuñó la expresión superviviencia del más apto, dando por hecho que el más apto será siempre el que busca su pro-pio interés y no duda de poner todas sus fuerzas al servicio de ese objeti-vo, sin importarle la moralidad o no del procedimiento. Defendía también una visión lamarckiana de la transformación de las especies, así como la selección social y el determinismo racial.
Hizo también una biologización de la ética, cayendo en la denomina-da falacia naturalista, en la medida en que infería de las leyes naturales conclusiones morales. En la medida en que el proceso evolutivo se orien-ta hacia la supervivencia de los más aptos, de los más fuertes, las espe-cies y los individuos que han sobrevivido son los mejores, no sólo desde el punto de vista fáctico sino también moral. Por tanto, la evolución su-pone progreso, en la medida en que iría de lo peor y más imperfecto ha-cia lo mejor y más perfecto. El mal es, por tanto, una inadaptación, que, al igual que la pobreza, tenderá a desaparecer por sí sola, puesto que la tendencia a la perfección humana es inevitable. Por ello, Spencer con-sidera que no hace falta la intervención del Estado ni dictar demasiadas leyes. Basta reforzar la libertad individual, y en ese poner en práctica las cualidades de cada uno, los más aptos sobreviven y los demás pere-cen. Quien no resiste en esta lucha, debe morir, puesto que es un estorbo para el perfeccionamiento de la especie. Claro está que estas teorías pre-suponían implícitamente el racismo y la sobrevaloración de los factores hereditarios, en cuanto determinismo conductual, así como la justifica-ción del imperialismo y del colonialismo, y la justificación de las guerras como instrumento de evolución55.
De este modo, aunque el darwinismo social no es de directa filiación darviniana, funcionó (y en algunos autores todavía sigue funcionando) como justificación del capitalismo basado en el individualismo posesivo frente a las reivindicaciones sociales. La verdad es que se puede discutir si Darwin se opuso o no claramente a estos planteamientos, pero lo que está claro es que no necesariamente se tiene que interpretar la teoría de la selec-ción natural en los términos en que lo hace el darwinismo social, pudién-
55 La influencia de estas teorías de H. Spencer, más o menos corregidas, fue muy gran-de a lo largo de la segunda mitad del s. XIX en todas las naciones latinoamericanas: cfr. Beorlegui, C., Historia del pensamiento filosófico latinoamericano, Bilbao, Universidad de Deusto, 2004 (2.ª ed.: 2006), pp. 265 y ss.

61
© Universidad de Deusto - ISBN 978-84-9830-383-4
dose entender, como veíamos en Kropotkin y otros, en clave de coopera-ción y ayuda mutua56.
3.3.4. El impacto de la teoría de Darwin en la Antropología socio-cultural
Para ver el impacto del darwinismo en el ámbito de la Antropología sociocultural, hay que ser conscientes de que la construcción de la teoría evolucionista se debió en gran medida a las convicciones evolucionistas de las disciplinas histórico-sociales. Aunque es cierto también que la cris-talización del darwinismo sirvió para dar nuevos impulsos a esas discipli-nas, y en concreto a la Antropología socio-cultural.
Ya sabemos que la primera gran escuela que emerge en la historia de la moderna Antropología socio-cultural se denominó precisamente evolu-cionista57. Los primeros antropólogos culturales, como Bachoffen o Mai-ne, no debían nada a Darwin en su evolucionismo. Pero la influencia es evidente en los autores posteriores más importantes, como Taylor, Morgan, Frazer, Spencer, Wake, Starcke y otros más.
Esta escuela evolucionista partía del a priori metafísico de que to-das las culturas tenían una esencia común, que se expresaba de forma unitaria en tres fases de evolución: salvajismo, barbarie y civilización.La cultura europea occidental, como consecuencia de un claro etnocen-trismo cultural, era la modélica y más desarrollada, y el resto se hallaba en alguna de las dos fases anteriores, tarea que perseguía determinar el método comparativo que utilizaban los antropólogos. Así mismo, todas las situaciones culturales se enfocaban desde el seleccionismo evolutivo. Así, por ejemplo, el tabú del incesto y la ley de la exogamia se entendían como mecanismos de supervivencia y de progreso, en la medida en que los grupos exogámicos tienen mejores posibilidades de adaptación y de supervivencia.
Pero estos planteamientos evolucionistas fueron criticados posterior-mente por Franz Boas y su escuela, el particularismo cultural, así como por la escuela difusionista. Boas criticó y puso en cuestión los aprioris
56 Cfr. Kropotkin, Piotr, El apoyo mutuo. Un factor de la evolución, Madrid, Ed, Zero Zyx, 1970; 2.ª ed.: 1978 (introducción de Carlos Díaz); Id., La selección natural y el apo-yo mutuo, Madrid, CSIC, 2009 (Introducción de A. Girón Sierra); Lewontin, R./Kamin, S./Rose, L. J., No está es los genes, o.c.; Gutiérrez, A./Ortega, R., «Ciencia e ideología en una posible antropología darwiniana: una crítica a Patrick Tort», o.c.
57 Cfr. Harris, M., El desarrollo de la teoría antropológica. Una historia de las teo-rías de la cultura, Madrid, Siglo XXI, 1978; caps. 6.º y 7.º; Bohannan, Paul/Glazer, Mark (eds.), Antropología. Lecturas, Madrid, McGraw-Hill, 1992 (2.ª ed.); Beorlegui, C., Antro-pología filosófica. Nosotros: urdimbre solidaria y responsable, o.c., pp. 208-209.

62
© Universidad de Deusto - ISBN 978-84-9830-383-4
metafísicos y no demostrados empíricamente desde los que partía el evo-lucionismo cultural, proponiendo contra ello una mayor fidelidad a los datos empíricos en el trabajo de campo. Y el difusionismo del P. Schmidt y sus seguidores proponía otro método diferente para acercarse a la ex-plicación de las similitudes entre culturas diferentes: no tanto como con-secuencia de hallarse en similar fase de desarrollo cultural, sino como consecuencia de los diversos intercambios comerciales entre los centros culturales promotores de cultura58.
A pesar de estas críticas y del desprestigio consecuente del primer evolucionismo cultural, en mitad del siglo XX se han vuelto a reprodu-cir diferentes corrientes neo-evolucionistas, como las de Leslie Whithe y Gordon Childe, y el materialismo cultural de Marvin Harris, que también se considera neo-evolucionista59.
4. G. MENDEL (1822-1884) Y EL DESARROLLO DE LA GENÉTICA
4.1. G. Mendel y los inicios de la genética60
Las aportaciones de G. Mendel a la genética fueron claves para com-pletar el conocimiento del proceso evolutivo. Mendel había nacido en 1822, en un pueblecito de Moravia (posteriormente Checoslovaquia, y actualmente Chequia). En su convento de Brünn (agustinos) se dedicó a realizar sus experimentos con guisantes, y a través de sus experimentos y observaciones, llegó a descubrir por su cuenta las leyes fundamentales de la herencia genética. Sus experimentos estaban guiados por dos leyes centrales:
a) la ley de la disyunción de los caracteres en las células reproducto-ras del híbrido, esto es, los caracteres de los padres se separan para conformar las células reproductivas o gametos;
58 Sobre estas dos escuelas, cfr. Harris, M., o.c., cap. 9.º y 14.º; Beorlegui, C., o.c., pp. 209-210.
59 Cfr. Harris, M., El materialismo cultural, Madrid, Alianza, 1987 (2.ª ed.). 60 Cfr. Rostand, J., o.c., cap. 18.º, 169 y ss.; Castrodeza, Carlos, «G. J. Mendel (1822-1884)»,
Revista de Occidente, 1984, n.º 32, 7-26; Cordón, Faustino, «La genética, a los cien años de Mendel (1822-1884)», El País, 6 de enero de 1984, 22-23; Fox Keller, Evelyn, El siglo del gen. Cein años de pensamiento genético, Barcelona, Península, 2002; Barahona, A./Ayala, Fr. J., El siglo de los genes. Patrones de explicación en genética, Madrid, Alianza, 2009, cap. 2.º; Grasa Hernández, R., El evolucionismo: de Darwin a la Sociobiología, Ma-drid, Cincel, 1986, pp. 76-89.

63
© Universidad de Deusto - ISBN 978-84-9830-383-4
b) y la ley de la independencia de los caracteres. Es decir, contra la opinión de muchos pensadores, Mendel defendía que el patrimonio hereditario estaba formado por unidades separadas, que en el futu-ro serían denominados genes.
Los experimentos realizados con guisantes confirmaron de modo em-pírico sus presupuestos, y escribió una memoria de sus trabajos (Versuche über Planzen-Hybriden: Investigación sobre Plantas híbridas), presen-tada en 1865, a la Sociedad de Historia Natural de Brünn. Esta memoria fue leída por Mendel dos veces públicamente y se publicó en el volumen anual de los Informes de la citada Sociedad. También envió copias a va-rios científicos notables, entre ellos a Carl Wilhelm von Nägeli, célebre botánico austríaco, pero ni él ni el resto de los científicos que recibieron la memoria la valoraron como se merecía.
Años después, Mendel fue nombrado obispo, lo que, unido al escaso eco de sus investigaciones, supuso el abandono de su vocación científica. Jean Rostand señala la maravilla que supone que un solo hombre cons-truyera de golpe una nueva ciencia, la genética, cuya base está magistral-mente descrita en su memoria de 1865, de forma madura y acabada61. No se conoce otro caso tan extraordinario en la historia de la ciencia, salvo quizás el de L. Pasteur y sus trabajos sobre los fermentos y microbios.
El éxito de Mendel en sus investigaciones se debió, sin duda, a dos cosas:
— a su genio intuitivo que le llevó a defender la unidad del carácter genético y la separación entre unos genéticos y otros;
— y a la suerte de experimentar con guisantes, planta en la que los he-chos de la herencia se dan con gran nitidez.
Lo que resulta más extraño es la poca atención que se dio a sus des-cubrimientos expresados en su memoria. Las razones de esta escasa difu-sión y poco éxito en el entorno del mundo científico parece que se debió a la conjunción de varias circunstancias: la poca difusión de la revista en la que se publicó la memoria; la lejanía de Moravia de los centros de difu-sión científica y cultural; el, en opinión muy acertada de algunos, carácter insólito de la metodología empleada: el carácter estadístico de los resulta-dos. No podían probar que la herencia no fuera intermedia, excepto para algunos casos especiales, poco relacionados con el modelo darwiniano. Es decir, variaciones importantes parecían ser, todavía, más continuas que discontinuas. Y sobre todo, parece que el motivo más determinante de la falta de atención a los descubrimientos de Mendel se debía a la excesiva
61 Cfr. Rostand, J., o.c., p. 174.

64
© Universidad de Deusto - ISBN 978-84-9830-383-4
originalidad de los descubrimientos. Podríamos decir que Mendel llegaba demasiado pronto para su época, por lo que no nos puede extrañar que no pudiera ser comprendido. Por ello, Medel, en opinión de algunos, más que estar aislado geográficamente, estaba aislado culturalmente, es decir, tenía poca conexión con las preocupaciones de la época.
4.2. El redescubrimiento de la genética mendeliana
Si resulta extraño y curioso el asilamiento sufrido por la publicación de la memoria de los descubrimientos de Medel, más curioso resulta el redescubrimiento de su obra por parte de tres científicos a la vez.
A finales de marzo de 1900, el botánico holandés Hugo de Vries pu-blicó dos notas, en Francia y Alemania, expresando sus descubrimientos en cruces de plantas. Había descubierto la ley de la disyunción, es decir, los caracteres de los padres se separan en las células reproductoras del hí-brido. Pero, al mismo tiempo que da a conocer su descubrimiento, recono-ce que esta ley ya la había descubierto treinta y cinco años antes G. Men-del, un monje agustino. Esto lo llegó a saber a través de la lectura de una obra de Focke62, donde se daba cuenta de esta noticia. Por tanto, de Vries había descubierto esta ley sin tener ninguna noticia de que ya lo había he-cho con tantos años de anticipación G. Mendel, y sin ni siquiera conocer la obra y la importancia de sus investigaciones.
Lo curioso es que un mes más tarde (24 de abril de 1900), Carl Correns,botánico de la Universidad de Tübingen (Alemania), presentó resultados similares en sus investigaciones, realizados con guisantes y maíz. Y el mes de junio, el botánico vienés, Erich Tschermak, presentaba las mismas conclusiones tras haber realizado experimentos con guisantes. Como es lógico, estos dos últimos científicos quedaron estupefactos ante la refe-rencia a Mendel realizada por de Vries en su escrito63.
A partir de este momento, las leyes de la hibridación se aceptaron ple-namente, y la biología dio un paso fundamental de cara a su historia pos-terior. En realidad, las teorías de Weismann habían preparado el camino, y ya no resultaban estos descubrimientos una novedad tan grande como habían supuesto en la época de Mendel. Con gran acierto, dice Rostand al respecto: «Sin disminuir en un ápice el mérito de los tres redescubri-
62 Planzen mischlingen, 1881. 63 Cfr. Rostand, J., o.c., cap. 19.º; Barahona, Ana/Ayala, Fr. J., o.c.; Judson, H. F., El
octavo día de la creación, México, Ediciones Castel Mexicana, S. A. y Conacyt, 1987; Su-sanne, Ch./Chiarelli, Br./Salces, I., «Bases genéticas», en Rebato, E. y otros (eds.), Paracomprender la Antropología biológica, Estella, Verbo Divino, 2005, cap. 2.º, pp. 33-83.

65
© Universidad de Deusto - ISBN 978-84-9830-383-4
dores, puede decirse que encontraron lo que era normal que hallasen en su época. Son de su tiempo y su obra es hija del estado de la ciencia que les rodea. Pero, los 35 años de adelanto que Mendel tiene respecto a ellos, miden el grado de la potencia anticipadora del genio. Con el tranquilo aplomo de su pensamiento, un hombre solo se ha adelantado a la ciencia; un hombre que era apenas un sabio, supo otear lo que sólo verán los sa-bios 35 años después»64.
4.3. Genética versus selección natural
Pero seguimos con las paradojas y curiosidades, puesto que, desde la perspectiva de lo que más adelante se denominó la teoría sintética de la evolución, los avances de la genética ayudan a comprender y a confirmar las afirmaciones de Darwin sobre la selección natural, pareció en estos primeros momentos del redescubrimiento de las leyes de la genética que las cosas podían ser al revés, es decir, se llegó a pensar que estos descu-brimientos ponían en cuestión las tesis de Darwin. En palabras de R. Gra-sa Hernández, «el redescubrimiento de las leyes de Mendel no sólo no reforzaron la teoría darwiniana de la selección natural, sino que contribu-yeron decisivamente a que perdiera prestigio entre los biólogos»65.
La razón de ello la justifica el gran genetista Th. Dobzhansky con varios argumentos. En primer lugar, estos genetistas no eran naturalistas; es decir, estaban metidos en el exclusivo círculo de la genética, y les faltaba conoci-mientos sobre los patrones de variación en poblaciones naturales, así como las interacciones entre organismos y medio. Sólo tenían experiencia de cul-tivos en jardín de semillas importadas, y de la Drosophila en el laboratorio. Pero también, en segundo lugar, estos científicos eran tipologistas, es decir, les parecía más importante el individuo mutante que la población a que per-tenecían. Y en tercer lugar, como misioneros de una teoría nueva, quitaban importancia a todo fenómeno que pudiera restarle importancia.
Todo esto llevó a que, desde estos comienzos del redescubrimiento de las leyes de la genética, los más importantes genetistas combatieron las teorías darwinianas, al tiempo que iban haciendo importantes aportacio-nes a la historia de la genética, como vamos a ver. Al calor de este desper-tar de la genética, fueron apareciendo tres teorías que se oponían, de dife-rente modo, a la selección natural del darwinismo: la teoría saltacionista, el neolamarkismo y la ortogenética.
64 O.c., p. 176. 65 O.c., p. 77.

66
© Universidad de Deusto - ISBN 978-84-9830-383-4
a) Las teorías saltacionistas o mutacionismo66
Estas teorías ya existían en tiempos de Darwin, pero se convierten en predominantes a partir de 189467, con el descubrimiento del mutacionis-mo por Hugo de Vries. Los primeros saltacionistas fueron T. H. Husley y William Bateson, pero el mayor impacto y auge que recibió esta postura vino de parte del genetista de Vries con su obra publicada en 1901-1903, Die Mutationstheorie (La teoría de las mutaciones).
De Vries descubrió dos tipos de variaciones en la herencia: la variabi-lidad individual (que supone cambios en algunos rasgos de los individuos, cambios que pueden ser unas veces mejoras y otras no) y la variación dis-continua, que lleva hacia la especiación (o cladogénesis). La primera es la que origina las variaciones pequeñas que tiene un individuo a lo largo de su vida. Estas variaciones no hacen que nazcan nuevas especies. Sólo sir-ven para que mejore la especie en ciertos rasgos, poco a poco. En cambio, sí se produce la aparición de nuevas especies con las variaciones disconti-nuas, que, según de Vries, suceden de forma brusca, por macromutaciones.Para de Vries, por tanto, la especiación se debe al origen espontáneo y sú-bito de una variación discontinua68. Así, pues, no se da ni transición ni pre-paración visible69.
Utilizando cultivos de plantas, como la onótera de grandes flores, u onótera de Lamarck, procedente de América, de Vries observa que, de entre miles de plantas normales, aparecen en cada generación unas po-cas diferentes, aberrantes. Y saca de ello la conclusión de que se trata del punto de partida de verdaderas especies elementales, aparecidas por un proceso de variación brusca o mutación. Es decir, la nueva especie nace repentinamente, con sus caracteres plenamente desarrollados, sin pasar por ninguna etapa progresiva o transitoria. Estas mutaciones aparecen de forma esporádica, y sólo en pequeñas cantidades (menos del 1 por mil, en los experimentos de de Vries).
La mutación sólo puede venir de un cambio en el propio germen. Y es un cambio que se produce en el mecanismo interno de bipartición de las células sexuales de los progenitores, y sin ninguna relación con las cir-cunstancias externas del medio, ni las cualidades adquiridas de los padres. Estas mutaciones sólo se forman en ciertas épocas de la vida de la espe-
66 Cfr. Leith, B., El legado de Darwin, Barcelona, Salvat, 1986.67 Esto se debió a la nueva teoría de de Vries sobre la evolución, apoyada en pacientes
investigaciones emprendidas desde el año 1886, con su obra Die Mutationstheorie, publi-cada entre 19001 y 1903.
68 Cfr. Ruffié, J., De la biología a la cultura, Barcelona, Muchnik Ediciones, 1982.69 Cfr. Rostand, J., o.c., cap. 19.º, p. 177.

67
© Universidad de Deusto - ISBN 978-84-9830-383-4
cie, épocas definidas y probablemente de corta duración. Así, para encon-trarse con ellas, hay que tener suerte, la que tuvo de Vries con la onótera.
Conviene tener en cuenta, de todas formas, que las variaciones de de Vries difieren radicalmente de las variaciones de Lamarck. Además, difie-ren de la denominada variación darwiniana o weismanniana, por el hecho de que la mutación de de Vries crea directamente la nueva especie elemental; mientras que, para Darwin y Weismann, la especie sólo se crea poco a poco, mediante una lenta y radical selección de variantes. De Vries hace intervenir también a la selección natural, pero de muy diferente forma a como la enten-día Darwin, puesto que no todas las mutaciones son positivas o viables. Sólo salen adelante algunas, y la mayoría perecen, debido a que la selección natu-ral actúa como criba. Ahora bien, Darwin plantea la lucha por la existencia en el interior de la especie, y de Vries sólo entre las especies elementales.
Así, como indica J. Rostand, «ya que la nueva especie nace de un solo individuo mutante y no, como admite Darwin, en lo selecto de cada genera-ción, de Vries prefería el término de elección, que se aplica a la de un indi-viduo más que a la de un grupo, al término de selección»70. Igualmente, la diferencia con Darwin y Weismann está en que la mutación es una modifi-cación repentina de uno de los elementos, y no una lenta e incesante modi-ficación de los elementos del plasma germinativo. Es decir, para de Vries, el plasma germinativo es mucho más estable que para los otros dos científicos; entre una mutación y la siguiente, permanece perfectamente fijo.
Los descubrimientos de de Vries crearon un gran revuelo y grandes polémicas, y se convirtieron en las ideas dominantes entre 1900 y 1910, teniendo más repercusión que el mismo redescubrimiento de Mendel. Además, se hizo tal hincapié en el aspecto genético que se pensó que con-tradecía y echaba por tierra el planteamiento de Darwin. De Vries llegó a afirmar que la lucha por la existencia y la selección natural no sólo no ori-ginaban las especies, sino que las exterminaban71. Algunos vieron en el mutacionismo la negación de la idea de transformismo, y la vuelta a las concepciones rupturistas o catastrofistas. Por el momento, los experimen-tos de de Vries se impusieron, y se completaron, llegándose a quedar claro una serie de realidades:
— el patrimonio hereditario es completamente estable, excepto cuan-do se dan mutaciones. J. Monod dirá más tarde que la base de la re-producción consiste en combinar el azar y la necesidad72;
70 Ibídem, 180. La cursiva es del autor. 71 Cfr. Grasa Hernández, R., o.c., p. 79. 72 Cfr. Monod, J., El azar y la necesidad. Ensayo sobre la filosofía natural de la filoso-
fía moderna, Barcelona, Barral, 1971 (9.ª ed. 1977).

68
© Universidad de Deusto - ISBN 978-84-9830-383-4
— y que el único modo de variación que se da en la naturaleza vivien-te es el modo brusco y esporádico73. La cuestión se centra ahora en saber si el procedimiento actual de formación de especies es el mis-mo que originó los cambios evolutivos en épocas pasadas.
Los experimentos genéticos continuaron con más vigor todavía y se centraron en hallar la base celular de los caracteres hereditarios. Ya a fina-les del siglo XIX (en 1880), los citólogos aceptaban la hipótesis de que la sustancia celular se hallaba en los cromosomas. Se trataba de unir el men-delismo con la hipótesis cromosómica. Es decir, ver si las unidades here-ditarias de Mendel residían realmente en los cromosomas.
El éxito de este trabajo le correspondió a Thomas Hunt Morgan y a su escuela74. Sus investigaciones las realizó con la Drosophila melanogaster(también denominada mosca del vinagre), animal muy adecuado para este tipo de investigaciones75. A cada unidad cromosomática o gen (según el nombre dado por Johannsen) le correspondía una característica en el fe-notipo. Morgan defendió y distinguió el orden de los genes en los cromo-somas, presentándolos como verdaderos planos de los cromosomas de la drosophila. Como puede imaginarse, el escepticismo sobre esas propues-tas fue muy grande al principio, pero Morgan seguía aportando pruebas, a través de cruces de razas, y por medio de observación al microscopio (bandas de alineamientos, posibles genes). En la actualidad, como conse-cuencia de la continuación de estas investigaciones, y acumulando nuevos descubrimientos, se ha constituido una nueva e importantísima ciencia, la genética.
Con estos nuevos descubrimientos se ha dejado claro el problema de las mutaciones. La biología ha dado un salto espectacular en este punto, y de ser la célula la unidad de acción y de investigación, se ha pasado a serlo más bien los genes, en la actualidad como un fragmento del largo filamento de la doble hélice del ADN76. De todos modos, en ese estado de las investigaciones, la evolución parecía reducirse a un problema de genes, que se transmiten invariablemente o con mutacio-nes esporádicas.
73 Cfr. Ruffié, J., De la biología a la cultura, o.c. 74 Morgan nació en 1866, y fue profesor del Instituto Tecnológico de Pasadena (Califor-
nia). Cfr. Barahona, A./Ayala, Fr. J., El siglo de los genes, o.c., pp. 59 y ss. Cfr. Schoerbel, W., Evolución, Barcelona, Salvat, 1938, cap. 1.º.
75 Cfr. Rostand, J., o.c., p. 184, nota 3. 76 Para seguir sobre el tema de los genes, cfr. J. Rostand, o.c., pp. 186-88; Ayala, Ori-
gen y evolución del hombre, Madrid, Alianza, 1980, cap. 2.º; Barahona, A./Ayala, Fr. J., Elsiglo de los genes, o.c., cap. 5.º; Monod, J., El azar y la necesidad, o.c.; Fox Keller, E., El siglo del gen. Cien años de pensamiento genético, o.c.

69
© Universidad de Deusto - ISBN 978-84-9830-383-4
Pero los avances posteriores han demostrado que de Vries, al lado de sus grandes aportaciones en el ámbito de la genética, estaba equivocado en dos puntos:
— la mutación sólo aporta una variación ligera, y, en general, sólo así sucede. Son necesarias numerosas mutaciones para constituir una nueva especie;
— además, no se dan períodos concretos de mutabilidad, sino que en cualquier momento y en cualquier especie, se puede mutar la he-rencia. Se sigue aquí la impredictibilidad cuántica, el puro azar.
Y lo que está claro es que la mutación parece ser independiente de las condiciones externas del ambiente. Aunque pueden provocarse muta-ciones sometiendo a los genes a la acción de factores como los rayos X, o los rayos de radium. Pero esto es un tema muy distinto al problema del lamarckismo.
b) Las teorías neolamarckianas
Siempre se han dado ciertos restos de lamarckismo, aunque de dife-rente tipo. Pero no siempre presuponían y se añadían a sus teorías el fi-nalismo, que viene a ser el principal rasgo de la teoría de Lamarck. In-cluían una teoría de la herencia y una de la evolución, pero cada una de ellas eran muy diferentes entre sí. Entre esas teorías se dan varias, como, en primer lugar, el geoffrismo, consistente en defender que el cambio evo-lutivo se debe a la influencia directa del entorno. Esta teoría fue bastante popular en los comienzos del siglo XX. Pero también, en segundo lugar, teorías que defendían la influencia de las fuerzas mentales, de la concien-cia, en el proceso evolutivo. Así, por ejemplo, lo defendía Cape, y el psi-colarmarckismo de Pauli77.
Todas estas teorías neolamarckianas defienden la herencia de los ca-racteres adquiridos, por lo que aceptaban más la adaptación que la va-riación por azar. De modo que, con los descubrimientos de De Vries y Morgan, se avinieron a la evidencia. Pero posteriormente se ha vuelto a reaceptar el lamarckismo y a defender la influencia de las proteínas sobre los ácidos nucleicos (ADN y ARN)78.
77 Cfr. Jacob, Fr., La lógica de lo viviente, Barcelona, Edit. Laia, p. 10.: referencia al pasaje bíblico del génesis entre Jacob y su suegro Laván, en el que las ovejas se van apa-reando ante ramas descortezadas por franjas, defendiéndose la influencia del entorno físico en el apareamiento y el tipo de descendientes.
78 Cfr. Leith, B., El legado de Darwin, Barcelona, Salvat, …

70
© Universidad de Deusto - ISBN 978-84-9830-383-4
c) Teorías ortogenéticas
Ya hemos hecho referencia a ellas varias veces con antelación. El fi-nalismo biológico era doctrina común en el siglo XIX, consistente en de-fender, por un lado, que el principio de perfección es inmanente a toda forma de vida orgánica; y, por otro lado, que la evolución sólo puede ope-rar en una sola dirección más o menos rectilínea, y además, en un proceso ascendente. A este principio lo denominó Eimer ortogénesis. Algo pare-cido a esto podemos considerar las tesis de Teilhard de Chardin sobre su principio Omega, o también la tesis de Berg sobre la nomogénesis.
Pero resulta difícil explicar la causa de este principio ortogenético:unos lo atribuyen a unos principios teleológicos inmanentes; otros, a mis-teriosos procesos internos, de modo que se caía de nuevo en el principio embriológico de la preformación en la evolución.
Ahora bien, a pesar de que la ciencia biológico-genética echa por tierra la ortogénesis, se sigue añorando la aceptación de un finalismo en la evolución, como defienden hoy en día la teoría del Diseño Inteligente, so-bre la que hablaremos en el siguiente capítulo.
d) El neodarwinismo de Auguste Weismann79
Ya en 1903, Weismann postuló la existencia de genes, y, sin saber-lo, aplicó en su teoría de la evolución la mecánica mendeliana al darwi-nismo. Para Weismann, la selección se hace sobre unas partículas que llama determinantes, que transmiten los caracteres hereditarios. Pero la selección fundamentalmente se realiza a través de las células sexuales, puesto que son las únicas que transmiten los caracteres genéticos a los descendientes.
Habría, pues, en cada organismo dos partes: el soma, conjunto no re-productor, y el germen, contenido en las células sexuales. De ahí la afir-mación de uno de los discípulos de Weismann, S. Butler, indicando que la gallina es un medio imaginado por un huevo para lograr que ponga otro huevo. Esta misma idea la repetirá, referida a los genes, el fundador de la sociobiología, E. O. Wilson80.
Weismann rechazaba claramente la unión de la teoría de Darwin con la herencia de los caracteres adquiridos. La variabilidad genética, factor
79 Aunque se ha solido denominar también neodarwinismo a la teoría sintética de la evolución, parece más adecuado reservar el nombre de neodarwinismo para la teoría de Weismann.
80 Cfr. el prólogo de su Sociobiología. La nueva síntesis, Barcelona, Omega, 1975.

71
© Universidad de Deusto - ISBN 978-84-9830-383-4
necesario para la selección, la explicaba desde la comprensión del ex-traordinario poder de la recombinación sexual para crear la variabilidad genética (por el procedimiento del crossing over). Más tarde admitió el principio de selección germinal, postulando la posibilidad de variación dirigida, causada y guiada por las condiciones de vida de los organismos. Negaba todo principio ortogenético, pero aceptaba que la selección de ciertos rasgos (plumas largas, por ejemplo) favorecía simultáneamente a aquellos genotipos que tenían tendencia a variar el tamaño de las plumas de la cola.
Pensaba de este modo porque la variación al azar y la selección no pa-recían factores suficientes para explicar fenómenos tan curiosos como la reducción gradual de los órganos rudimentarios o inútiles (los ojos de los animales cavernícolas).
Esta teoría es el equivalente de las teorías modernas acerca de que el genotipo establece limitaciones definidas sobre la variación genética, y marca las posibilidades del fenotipo, lo que unido a la selección, fa-vorable o desfavorable de ciertos genes reguladores, puede explicar los supuestos rasgos ortogenéticos observados81. Como puede entenderse, el impacto de Weismann en la biología evolutiva ha sido fuerte y perma-nente. En realidad, la mayoría de los problemas debatidos en los últimos ochenta años del siglo XIX, los planteó él, y así preparó el terreno para el redescubrimiento de Mendel, como ya lo hemos apuntado, al obligar a los lamarckianos a probar y encontrar una buena base a su teoría. También es cierto que el rechazo de sus teorías por humanistas, científicos y filósofos fue total, o casi total.
5. LA TEORÍA SINTÉTICA DE LA EVOLUCIÓN82
5.1. Los pasos hacia la primera síntesis
Ya hemos visto cómo los primeros pasos de la genética, tras las apor-taciones de de Vries, se alejaban de la selección natural. Pero otros teó-ricos fueron avanzando hasta darse cuenta de que podían perfectamente compatibilizarse los dos elementos, la evidencia genética y la selección
81 Cfr. Grasa Hernández, R., o.c., 83; Monod, J., El azar y la necesidad, o.c., últimoscapítulos.
82 Cfr. Dobzhansky, Th./Ayala, Fr. J./Stebbins, G. L./Valentine, J.W., Evolución, Barce-lona, Omega, 1983; Schoerbel, W., Evolución, Barcelona, Salvat, 1986, cap. 4.º.

72
© Universidad de Deusto - ISBN 978-84-9830-383-4
natural, así como el carácter gradual de ésta. Porque ya hemos visto cómo se fue refutando poco a poco tanto el mutacionismo como el salta-cionismo83.
Podríamos decir que los investigadores del ámbito biológico se di-vidieron en dos campos de investigación: los interesados en la mecánica de la herencia, y los estudiosos de los aspectos evolutivos. Los primeros, aunque comenzaron siendo antidarwinistas, poco a poco fueron advirtien-do que la evolución no es un fenómeno que sucede sólo en el individuo, en sus mutaciones y traspasos de herencias, sino más bien un fenómeno poblacional. Este convencimiento es el que hizo que apareciera una nueva rama de la genética, la genética de poblaciones, que resultó a la postre de-cisiva para la recuperación del darwinismo.
Bajo la denominación de genética de poblaciones se suele situar, como indica M. Ruse, «a los estudios que conciernen esencialmente a la herencia de grupo, contrapuestos al estudio del individuo, y so-breentienden que se trata de una porción del campo global de la genéti-ca mendeliana»84. Dentro de estos estudios se advierten dos tendencias: la genética de poblaciones matemática, en la que se sitúan R. A. Fischer, J. B. S. Haldane y S. Wright; y la genética de poblaciones ecológica, que estudiaba poblaciones reales de organismos vivos en el laboratorio y en su medio ambiente, y entre sus investigadores se hallaban, entre otros, Chetverikov y Th. Dobzhanski. Ambas corrientes tuvieron el mérito de acabar con el pensamiento tipológico, influido por el esencialismo. La importancia de esto se halla en el hecho de que toda la historia del pensa-miento evolutivo está marcada por dos tipos de pensamiento: el popula-cionista (genética de poblaciones) y el tipológico.
El pensamiento populacionista enfatiza la singularidad de cada cosa. Los colectivos son sólo aglomeraciones de individuos, y sólo pueden des-cubrirse colectivamente en términos estadísticos. La única realidad exis-tente propiamente tal son los individuos de las poblaciones. Se pueden establecer promedios estadísticos y seguir su variación. Así, el tipo o pro-medio es una abstracción y lo único real es la variación.
El pensamiento tipológico considera, por el contrario, que el tipo (al estilo del eidos platónico) es lo real, y la variación sería sólo una ilusión.
Los estudios de genética de poblaciones tuvieron la virtud de ampliar los puntos de mira de los estudios genéticos iniciados por de Vries, y abrir-se a la introducción sintética de la selección natural, que dieron como re-
83 Cfr. Grasa Hernández, R., o.c., p. 84 y ss. 84 Ruse, M., Filosofía de la biología, Madrid, Alianza, p. 38. El cap. 3.º lo dedica a la
genética de poblaciones y su validez científica: pp. 38 y ss.

73
© Universidad de Deusto - ISBN 978-84-9830-383-4
sultado la teoría sintética. Todos los cultivadores de esta disciplina, en sus dos líneas de trabajo, insisten en la importancia de la selección natural y en la inexistencia de la presión puntuacionista y de la herencia de los caracteres adquiridos. Por tanto, para ellos existiría solamente una evolución gradual. Además, no hay conflicto para ellos entre la discontinuidad de los genes y la continuidad de la variación individual (contra De Vries y la presión de la mutación).
Con todos estos avances, se mostró que las especies no son entidades esencialistas sino agregados de poblaciones naturales, reproductivamente aislados entre sí, y que ocupan nichos específicos en la naturaleza. La pa-labra especie es un término relacional; esto es, los genes de un individuo son componentes del mismo pool genético85.
5.2. La teoría sintética
Se formuló por primera vez hacia finales de los años treinta y comien-zos de los cuarenta, y se reformula a partir de 1950, con el nacimiento de la genética molecular86. Entre 1940 y 1960, la aceptación fue casi total. Desde 1970, con motivo de la genética molecular, se ha vuelto a poner en cuestión esta teoría sintética.
En la actualidad, frente a los defensores de la síntesis, se sitúan tres grupos contrarios: en primer lugar está la línea de quienes, fieles a Darwin en general, cuestionan la síntesis por creer que ha expandido demasiado sus dominios y que los últimos niveles de cambio evolutivo responden a causas diferentes. Por tanto, les acusan de querer abarcar demasiado con pocos datos87; otra línea considera que los hallazgos recientes refutan el darwinismo, y proponen teorías alternativas, como el neutralismo y la teo-ría estocástica, y el neolamarckismo; y una tercera línea contraria está for-mada por los sociobiólogos, que propugna su propia síntesis, más omnia-barcadora que la de la teoría sintética.
La teoría sintética persigue conjugar complementariamente las ideas básicas de Darwin con la genética de poblaciones, las observaciones ex-perimentales sobre la variabilidad de las poblaciones naturales, y la teo-ría cromosómica de la herencia. Los autores que la llevaron a cabo fue-
85 Cfr. Ayala, Fr. J., Origen y evolución del hombre, Madrid, Alianza, 1980, cap. 4.º, pp. 86 y ss.
86 Cfr. Ruse, M., Filosofía de la biología, o.c., cap. 10.º; Barahona, A./Ayala, Fr. J., Elsiglo de los genes, o.c., cap. 5.º.
87 Leith, Brian, El legado de Darwin, Barcelona, Salvat, 1986, cap. 11.º.

74
© Universidad de Deusto - ISBN 978-84-9830-383-4
ron, sobre todo, Th. Dobzhanky, Julian Huxley, E. Mayr, G. Simpson, B. Rensch y G. L. Stebbins88.
Lo central de su teoría se encaminaba a la unión de la selección na-tural de Darwin con las leyes de Mendel y demás avances genéticos. Sus presupuestos de partida fueron los siguientes:
1) las interacciones entre los organismos y sus entornos varían mucho geográfica e históricamente;
2) existe una continuidad de la herencia (y de la tradición cultural); 3) las regularidades se alteran esporádicamente y al azar.
Lo que une la variabilidad y la evolución es la selección natural, ya que toda población tiene siete características: variabilidad genética, re-combinación genética (por la reproducción sexual), continuidad heredi-taria, capacidad de mutación, capacidad excesiva de reproducción, inte-gración del genotipo (todos los genes de un organismo), y las limitaciones del acervo genético (pool genético).
El proceso selectivo depende de las interacciones entre los organis-mos y su medio ambiente, y éstas pueden ser muy diversas, implicando o no lucha por la existencia. La variabilidad genética en una población es enorme; y los más aptos en la reproducción aportan a la generación si-guiente mayor proporción de genes o alelos. Así, la dirección evolutiva de la población dependerá del medio ambiente y del contenido genético de la población.
Por tanto, los procesos básicos tenidos en cuenta por la teoría sintética son estos tres: la mutación, la recombinación genética, y la selección na-tural. Pero sigue habiendo discrepancias sobre el grado de importancia de cada uno de ellos, en particular en relación al azar. En síntesis, las afirma-ciones de la teoría sintética se concentran en estas tres:
— la unidad de transmisión, conservación y mutación es la misma: el gen;— la unidad de selección es (como sostuvo Darwin) el individuo, no
el gen (como defienden los sociobiólogos); pero el proceso es más complejo que la simple interpretación de la lucha por la supervi-vencia. El problema se centra en la supervivencia de qué tipo de genes de un individuo se hallan o no presentes en la generación siguiente. No es central que el individuo sobreviva, sino que deje descendencia con posibilidad de sobrevivir; esto es lo que se deno-mina eficacia reproductiva (fitness);
— la unidad de evolución no es ni el gen ni el individuo, sino la pobla-ción. Y más en concreto, la población que comparte determinado
88 Cfr. Dobzhansky, Th., y otros, Evolución, o.c.

75
© Universidad de Deusto - ISBN 978-84-9830-383-4
genoma. Es decir, lo que evoluciona es una población mendeliana con recombinación genética a causa de la reproducción sexual, una población que produce constantemente nuevos genomas, a causa de la recombinación.
5.3. Las teorías alternativas a la teoría sintética
En el ámbito de la genética, el dogma o planteamiento general que se da por aceptado e indiscutido defiende la relación unidireccional del genotipo al fenotipo, es decir: ADN-ARN-Proteìnas, no produciéndose en ningún caso la relación inversa: la transmisión de los caracteres ad-quiridos. El fenotipo no puede influir en el genotipo. Pero este asunto es más complejo. En un individuo, el genotipo no cambia, pero el fenotipo sí, a lo largo de su vida. La razón es muy sencilla y fundamental: el ge-notipo no especifica directamente el fenotipo, sino que sólo apunta a y especifica un marco fenotípico, por así decirlo. El genotipo establece una gama de posibilidades más amplio de lo que de hecho se concretará y se realizará. Por tanto, hay un margen de acción a través del medio (la so-ciobiología ha tenido bien en cuenta todo esto, con sus teorías sobre la epigenética)89.
Los genes (segmentos de ADN) se suelen dividir en dos tipos diferen-tes: estructurales y reguladores. Los estructurales sintetizan proteínas, a partir de aminoácidos. Suelen ser los genes más estudiados, y son inflexi-bles a las variaciones ambientales. En cambio, los genes reguladores se en-cargan de regular el tiempo, la actuación y desactivización de los estructu-rales, y parecen más flexibles. De este modo, se abre la posibilidad de que sean sensibles a la presión del medio sobre el fenotipo. Si esto fuera así, y se demostrara esta influencia, se agrietaría la teoría sintética, y la socio-biología recibiría un buen apoyo (en algunas de sus tesis menos extremas). La polémica está servida, pero no está todavía nada claro. Por otro lado, también es cierto que la teoría sintética está seriamente cuestionada por un conjunto de teorías rivales. Como dice S. J. Gould, «muchos evolucionis-tas empezamos a cuestionar esta síntesis y a respaldar la perspectiva jerár-quica de que los distintos niveles de cambio evolutivo a menudo reflejen diferentes tipos de causas»90.
Todo esto hace que tomen fuerza teorías alternativas, como la estocás-tica y la neolamarckiana, que renacen periódicamente como el ave fénix.
89 Cfr. Lumsdem, Ch. J./Wilson, E. O., El fuego de Prometeo, México, FCE, 1985; Beorlegui, C., Antropología filosófica, o.c., pp. 189 y ss.
90 Cita tomada de Grasa Hernández, R., o.c., p. 105.

76
© Universidad de Deusto - ISBN 978-84-9830-383-4
La primera concede la importancia fundamental al azar, y la segunda, a la necesidad, a la constancia de la base genética. Veamos lo central de ambas teorías empezando por la segunda.
5.3.1. Teorías neolamarckianas91
Estas teorías han sido defendidas, en esta nueva etapa, por autores como P. Grassé, A. Vandel y P. Winttrebert. Para Grassé, lo viviente lleva en sí mismo su poder evolutivo, de tal modo que la evolución se desarro-lló por un programa inmutable y necesario. Según él, esto se ve claro si se examinan los fósiles. Wintrebert considera la adaptación como una reac-ción orgánica frente a exigencias exteriores: un conjunto de respuestas activas frente a estímulos ecológicos.
Lo común a estos autores consiste en negar el azar y quedarse con la necesidad. Se suelen llamar neolamarckianos a todos los paleontólogos y zoólogos que niegan el papel de la mutación y del azar en la evolución. La evolución se explicaría como consecuencia de una necesidad que to-davía desconocemos. Sus argumentaciones se basan en cinco puntos, que nos vamos a limitar a enunciar:
— El fenómeno evolutivo es irregular. — El proceso evolutivo es anterior a la selección. — El fenómeno adaptativo puede exagerarse hasta convertirse en una
desventaja.— Tendencias evolutivas parecidas corresponden a presiones selecti-
vas distintas. — Y por sí solas las mutaciones no justifican la evolución. En re-
lación a este aspecto, los neolamarckianos esgrimen dos argu-mentos: en primer lugar, los organismos dotados de la más alta capacidad de mutación no siempre son los de mayor dinámica evolutiva. Por ejemplo, las bacterias y las drosophilas experimen-tan muchas mutaciones, pero apenas han evolucionado desde hace casi mil millones de años. En cambio, poblaciones de homínidos han experimentado múltiples cambios de especies en poco tiem-po. Y, en segundo lugar, en los metazoos, la aparición de nuevos órganos y funciones no se puede considerar que se deban a una mutación única, ni a una serie de mutaciones simples. Se trata de fenómenos complejos en los que intervienen muchos elementos ajustados entre sí.
91 Cfr. Ruffié, J., o.c., 123 y ss, y Grasa Hernández, R., o.c., 107 y ss.

77
© Universidad de Deusto - ISBN 978-84-9830-383-4
En definitiva, para los lamarckianos, las mutaciones, sin más, no pueden generar más que razas, pero no especies. Es lo que ocurre con la domesticación. Y se trata de un argumento de peso, aunque se puede contrarrestar diciendo que las domesticaciones afectan a períodos cor-tos. Con todo, Dobzhansky, investigando con las drosophilas, sí que creó especies nuevas. Pero muchos zoólogos aceptan que el esquema darwi-niano sirve para explicar la microevolución (evolución diversificadora), pero no lo consideran capaz de explicar la tipogénesis (formación de nuevas especies).
Para el neolamarckismo el origen de nuevas especies se debe buscar en otros lados. Implica la aparición de nuevos genes, que se forman de manera preferencial (por fenómenos de convergencia). No surgen, pues, al azar, sino como consecuencia de un estímulo externo. Con ello se plan-tea la cuestión de la ascensión de la información genética. La ascensiónes el fenómeno contrario de la descensión, esto es, la tendencia de lo que se suele denominar, como ya dijimos, el dogma central de la genética: ADN-ARN-PROTEINAS. Se sabe que, por una transcriptasa inversa,una porción del ARN puede provocar la síntesis del segmento de ADN correspondiente. Hay, pues, una cierta ascensión en la información. Pero, de momento, no hay pruebas de que las enzimas sean capaces de provocar la síntesis del ADN que las codifica.
Pero sí hay influencia del ambiente en variaciones enzimáticas (eco-proteínas: su síntesis está inducida por el medio). Se trata, pues, de enzi-mas de adaptación. Por ejemplo, hay enzimas digestivas modificadas se-gún el tipo de alimentos de ratones y cobayas. No se sabe si es hereditario (o puede serlo), o si es sólo adaptación de un individuo concreto.
De todos modos, hay autores que, como J. Ruffié, piensan que el pro-blema de la discusión entre darwinismo y lamarckismo está superado y es un falso problema. Ruffié piensa que la cuestión está en que, dada la enor-me amplitud de la información genética de un grupo, cualquier dificultad ecológica nueva tiene la práctica seguridad de recibir una respuesta ade-cuada. Por eso, es ocioso contraponer un modelo instructivo (lamarckiano) a un modelo selectivo (darwiniano). La realidad se desprende de ambos. Un ejemplo claro es el esquema o sistema inmunitario92. A un antígeno extraño se opone un anticuerpo. Visto así, parece un esquema instructivo. Pero se ha descubierto que preexisten múltiples informaciones en la línealinfocitoria, que son capaces de sintetizar múltiples anticuerpos, antes del contacto con los antígenos. Se tiene, pues, una gama enorme de informa-ción. Así, pues, el sistema inmunitario construye muchos antígenos como
92 Cfr. Ruffié, J., o.c., p. 133.

78
© Universidad de Deusto - ISBN 978-84-9830-383-4
prueba, y luego se queda con el que sirve en cada caso concreto. Según el tipo de antígenos, el sistema inmunitario hace ponerse en acción parte de la enorme herencia genética que está en letargo. Por consiguiente, se trata de un modelo en parte instructivo (según la dificultad), y en parte también selectivo (información).
Si la herencia es muy rica, los dos polos coinciden. Y esto vale para otros muchos más casos. Por tanto, tras vilipendiar a Larmarck y a los lamarckianos durante un siglo, el evolucionismo tiene hoy posturas más matizadas. Parece hoy tan inverosímil la teoría de la mutación correcta y singular como el papel del uso y desuso del desarrollo de los órganos.
5.3.2. Teorías neutralistas o estocásticas93
El máximo exponente de esta teoría es el científico japonés, M. Ki-mura, quien presentó ya sus planteamientos en 1968, con aportaciones de Crow, Jamasaki, Jacquard, Feldman y otros. Para Kimura, el papel central de la evolución está en el azar. Las mutaciones no tienen ningún valor se-lectivo; el azar constituye el papel esencial en el reparto y en el futuro de los genes.
Para apuntalar estos planteamientos, se fundamenta en dos datos:
— Los cálculos de Haldane demuestran la lentitud de fijación de una mutación aislada, incluso cuando es favorable. Es tal la lentitud, que una especie nueva (que necesita una serie de mutaciones) no habría tenido posibilidad de aparecer jamás.
— El extremo polimorfismo de lo viviente (Lewontin y otros), mostra-do en la electrofóresis de las enzimas, polimorfismo que afecta a todoslos loci (lugares o fragmentos de la herencia genética presente en el ADN). Esto sólo se explica si admitimos la ausencia de todo valor selectivo para la mayoría de los genes. En el caso contrario, el po-limorfismo genético es tal, que casi todos los individuos serían via-bles. Un polimorfismo tan amplio sólo es concebible si afecta a ge-nes rigurosamente neutros.
En conclusión, para Kimura la mayoría de las mutaciones son neutras.No responden ni a una ventaja ni a una desventaja clara. Es un fenómeno aleatorio. Así, la evolución es un proceso estocástico, guiado sólo por el azar. De ahí que estos autores abandonen la teoría darwiniana, puesto que niegan todo proceso selectivo.
93 Sigo a Ruffié, J., o.c., pp. 135 y ss.

79
© Universidad de Deusto - ISBN 978-84-9830-383-4
El modelo neutralista se puede resumir así: aunque sea desfavorable, una mutación tiene una cierta probabilidad de difundirse e incluso de sus-tituir a los demás alelos, a condición de que surja una población de efecti-vos reducidos. La inmensa mayoría de las mutaciones se pierden; sólo un pequeño porcentaje se fija (2 ó 3% para mutaciones con una ventaja se-lectiva del 1% en una población numéricamente baja). En este caso, sería mucho mayor el papel del azar que el de la presión selectiva, en el proce-so evolutivo.
Para Kimura, el papel de la selección sería esencialmente negativo, y se limitaría de forma más o menos rápida a las mutaciones más deletéreas (sobre todo, las que ponen en peligro una función biológica fundamental). Así, en conclusión, la selección actuaría más como un guardián, un pro-tector de las mutaciones demasiado nocivas, que como factor de progre-so, capaz de retener sin cesar los genotipos más favorables por su mejor adaptación. El proceso selectivo sería debido sobre todo al azar, y no a la selección natural.
Pero, para Ruffié, a pesar de que el modelo matemático de la teoría estocástica es perfecto, no cuadra con los hechos experimentales, con los experimentos realizados por científicos como Lewontin, Prakash, Ayala y otros. Tales experimentos llevan a las siguientes constataciones:
a) En poblaciones que viven en la misma área ecológica, la frecuen-cia de un alelo, en un momento dado, es la misma. Si sólo intervi-niera el azar (y fueran poblaciones aisladas), se observarían varia-ciones considerables de un grupo a otro. De hecho, se observa lo que cabría prever desde la óptica darwinista: el alelo más frecuen-te, o el más persistente, debe corresponder a la mutación que con-fiera el máximo de ventajas selectivas.
b) El esquema kimuriano no puede explicar la evolución estacional de las frecuencias génicas (estudiadas por Dobzhansky y Proesin-ger). Se separan dos poblaciones, y, al cabo de un tiempo, las fre-cuencias génicas tienden a igualarse, acercándose a la frecuencia media, de equilibrio, propio de la población salvaje. La evolución sigue siempre una dirección privilegiada: la que permite una ma-yor eficacia respecto a unas condiciones ambientales dadas.
Se puede admitir que el proceso kimuriano intervenga en genes indi-ferentes (neutros), o dotados de un poder selectivo débil. Pero tales genes no tendrían ninguna influencia sobre la selección. Además, la genética mendeliana considera que los genes verdaderamente neutros son excep-cionales. La hipótesis neutralista ha sido aceptada por los matemáticos y bioquímicos, porque se adapta a su modo de razonar. Pero no lo aceptan los zoólogos, enfrentados a la complejidad de la materia viva.

80
© Universidad de Deusto - ISBN 978-84-9830-383-4
Habría, de todos modos, una posibilidad de conciliar el modelo selec-tivo y el neutralista. Lewontin propuso que la unidad de selección sería no el gen, sino un segmento cromosómico (o todo el genoma). Hasta ahora, la unidad genómica era el cistrón (un trozo de cromosoma). Este gen, aisla-damente, no tendría valor selectivo. En cambio, el conjunto funcional en el que se incluye el gen, no es neutro. Llega así Lewontin al concepto de «seg-mento cromosómico característico», unidad de evolución. Se trataría de una especie de supergen. Quizás por ahí pueda llegarse a una cierta solución.
Partiendo del azar, que preside la aparición del polimorfismo genéti-co, la evolución es el fruto de necesidades ecológicas. La adaptación es una respuesta a las dificultades. El modelo estocástico no puede explicar ni las radiaciones adaptativas, ni tampoco los fenómenos de convergencia entre grupos muy alejados. La solución de Lewontin puede haber ayudado a un cierto acercamiento.
5.4. Unas cuantas evidencias constatables94
Según señala J. Ruffié, cualquiera que sea la hipótesis sobre la evolu-ción que consideremos más verdadera (seleccionista, mutacionista, esto-cástica), parece que se imponen las siguientes constataciones generales:
a) El primum movens del fenómeno evolutivo se sitúa en el nivel mo-lecular (que, de hecho, es el nivel macromolecular). Aquí es donde se sitúa el lugar de la aparición y de las transformaciones del ser vivo. Y esto se debe a las mismas propiedades de las macromo-léculas. Por debajo de un cierto nivel de complejidad, no podemos hablar ni de viviente ni de previviente. La existencia de especies y su evolución supone un mecanismo lo bastante rígido como para asegurar su transmisión invariante (necesidad), pero suficiente-mente flexible como para dar entrada a los errores y aportar nue-vas ventajas (azar). El privilegio del nivel macromolecular se debe a las propiedades de las mismas macromoléculas: son las primeras unidades que contienen información para poder asegurar la exis-tencia de seres vivos.
b) Pero no hay que menospreciar el resto de los niveles de integra-ción. El ser vivo es el conjunto de diversos niveles, que se integran y se interpenetran. No todo se reduce al nivel molecular, genético. A. Vandel95 habla de la formación y funcionamiento de un ser vivo
94 Cfr. Ruffié, J., o.c., pp. 158 y ss. 95 Cfr. Ruffié, J., o.c., pp. 143 y ss.

81
© Universidad de Deusto - ISBN 978-84-9830-383-4
como fruto de la integración de varios niveles. El conjunto total es la megagénesis. La evolución, en su conjunto, es una serie de nive-les de integración cada vez más complejos96. Y esos niveles son: el nivel molecular (aquí se halla el problema del origen de la vida), el nivel celular (aquí se hallan los ácidos nucleicos, proteínas y de-más), el nivel pluricelular (metazoos), y el nivel social. La socie-dad constituye un superindividuo formado por sujetos que pueden consagrarse cada uno a una actividad preferencial. Los procesos de integración en las sociedades de insectos son esencialmente bioló-gicos (innatos y adquiridos, en gradiente). En los vertebrados, los procesos son a la vez biológicos y psicológicos, y en los hombres, fundamentalmente psicológicos y culturales. Así, se da en la con-ducta humana mucha mayor flexibilidad que en las especies ani-males anteriores.
Por consiguiente, según este punto de vista organicista,
— la evolución está constituida por varios niveles de integración. En cada nivel, se siguen manteniendo las unidades del nivel an-terior, pero agregados en super-individuos.
— Por otro lado, las unidades que forman un nuevo nivel, no difie-ren fundamentalmente de las unidades aisladas del nivel prece-dente. Pero se especializan y presentan fenómenos de jerarquías, necesarias para funcionar con orden y eficacia.
— En cada nivel, surgen caracteres nuevos, desconocidos en el nivel precedente, pero que no parecen ligados a una remodelación fun-damental de la información genética, puesto que no hay ninguna modificación en la estructura de los componentes. Se trata más bien del establecimiento de nuevas relaciones que, por una parte, modifican quizás el funcionamiento del genoma (al especializar-se una célula, se inhibe parte de su material genético), y, por otra parte, revelan cualidades desconocidas hasta el momento, pero presentes en estado potencial en los niveles precedentes. Es el fe-nómeno denominado epigénesis. El organismo permite concebir la evolución como una verdadera ontogénesis, marcada por un cierto número de etapas que corresponden a los niveles de integración.
c) Cada plano de organización tiene su importancia. Y el conocimien-to de cada nivel no puede reducirse sólo al anterior, sino al conoci-miento directo de las cualidades de ese nivel, pues cada nivel tiene
96 Cfr. Luisi, Pier Luigi, La vida emergente. De los orígenes químicos a la biología sin-tética, Barcelona, Tusquets, 2010.

82
© Universidad de Deusto - ISBN 978-84-9830-383-4
sus cualidades específicas, irreductibles a las de los niveles ante-riores. En cada nivel aparecen cualidades nuevas, que no pueden descubrirse a priori en el nivel anterior. Pero, al mismo tiempo, por más nuevo que parezca lo que ocurre en un nivel, siempre está condicionado, en última instancia, por el nivel precedente. Así, cuanto más inferior sea un nivel, más explicativo será. En conse-cuencia, este modo organicista de ver las cosas nos indica que se da una autonomía dentro de cada nivel, pero una determinación o condicionamiento de unos niveles sobre otros.
d) El nuevo viviente es, pues, un edificio construido por la sucesión de niveles de integración. Y está conformado por dos movimientos en sentido opuesto:
— ascendente, contenido en la información genética. Del nivel mo-lecular se llega al social. Cualquier modificación aleatoria del molecular tiene su traducción en el nivel superior. El azar es quien guía esta vía ascendente;
— descendente, constituido por las fuerzas de la selección, que se ejercen desde los niveles superiores a los inferiores. La po-blación es donde se produce el primer impacto de la selección, antes que en el individuo, y a través de él, en el genoma y en el gen.
Del encuentro de estas dos fuerzas nace el fenómeno de la evolución. Pero estos dos movimientos todavía no están del todo elucidados. Pero se ha adelantado mucho, y con el tiempo irán quedando claros muchos me-canismos que hoy día están todavía oscuros.

83
© Universidad de Deusto - ISBN 978-84-9830-383-4
Capítulo 2
El darwinismo cuestionado
1. LOS PUNTOS DÉBILES Y LAS CRÍTICAS AL DARWINISMO EN LA ACTUALIDAD1
En la actualidad, la teoría de la evolución, aunque tenga todavía al-gunos puntos débiles y requiera ser complementada en algunos aspectos periféricos, es aceptada por la mayoría de la comunidad científica como fundamentalmente verdadera2. Dentro de esta teoría hay que distinguir, como señala Fco. J. Ayala, entre el hecho de la evolución, la historia de la evolución y las causas de la evolución de los organismos3. «El origen evolutivo de los organismos, dice Ayala, es hoy una conclusión científica establecida con un grado de certeza comparable a otros conceptos científi-cos ciertos, tales como la redondez de la tierra, la rotación de los planetas alrededor del sol o la composición molecular de la materia. Este grado de certeza, que va más allá de toda duda razonable, es lo que señalan los bió-logos cuando afirman que la evolución es un “hecho”»4.
1 Cfr. Sampedro, Javier, Deconstruyendo a Darwin, Barcelona, Crítica, 2002; Tort, Patrick,Para leer a Darwin, Madrid, Alianza, 2001; Dupré, John, El legado de Darwin. Qué signi-fica hoy la evolución, Buenos Aires, Katz Editores, 2006; Toro, M:A./López-Fanjul, C., «In-derminaciones del neodarwinismo», Teorema, XII/3 (1982), 251-274.
2 Mayr, E., Una larga controversia: Darwin y el darwinismo, Barcelona, Crítica, 1992; Bowler, P. J., The New-Darwinian Revolution (Reinterpreting a historical Myth), Baltimore,The Johns Hopkins University Press, 1988; Sampedro, J., o.c.; Vallejo, F., La tautología darwinista y otros ensayos de biología, Madrid, Taurus, 2002; VV.AA., Evolución hoy,Barcelona, Tusquets, 2001.
3 Cfr. Ayala, Fco. J., La teoría de la evolución. De Darwin a los últimos avances de la genética, Madrid, Temas de Hoy, 1999, pp. 17 y ss.
4 Ibídem, 17.

84
© Universidad de Deusto - ISBN 978-84-9830-383-4
Demostrada la verdad de este hecho, los evolucionistas se han esfor-zado en mostrar la estructura completa de este proceso evolutivo, concate-nando las diversas especies de seres vivos en una historia evolutiva en la que van quedando fijados y señalados los ritmos de la evolución, la apari-ción de las diversas especies, la desaparición de muchas de ellas, el modo como han ido colonizando las diferentes regiones de nuestro planeta, des-de que la vida surgió en él hace unos 3.500 millones de años hasta hoy, y otras muchas cuestiones similares.
Las aportaciones de Darwin al esclarecimiento del proceso evoluti-vo se orientan a mostrar evidencias sobre el hecho en sí de la evolución, pero su aportación más fundamental se refiere a dilucidar las causas de la evolución, al motor que empuja y orienta todo este impresionante proce-so. Para Darwin, ese motor no es otro que la selección natural, diferen-ciándose de las tesis de Lamarck, que, como ya ha quedado claro, defen-día la herencia de los caracteres adquiridos.
A pesar de este amplio consenso entre los científicos sobre el hecho y el motor clave de la evolución, no significa que la teoría de la selec-ción natural lo explique todo y disponga de todas las evidencias necesa-rias para dejar totalmente demostrado el paradigma de la evolución. En realidad, rara es la teoría que posee tales evidencias completas. Siguiendo los planteamientos popperianos, ocurre más bien que una teoría científi-ca es tal cuando su propuesta puede ser contrastada empíricamente, y si-gue siendo aceptada y considerada provisionalmente verdadera hasta que no se presenten contrapruebas empíricas o de coherencia interna que de-muestren su falsedad o sus limitaciones. Y esto se aplica ejemplarmente al darwinismo.
Recogeremos a continuación las críticas al darwinismo desde tres puntos de vista. La de quienes ponen en duda su carácter de teoría cien-tífica, considerando que a lo más representa una comprobación a pos-teriori del proceso evolutivo, pero dudando que tenga capacidad, como se supone tienen que tener todas las teorías científicas, para predecir los acontecimientos. Por otro lado, están los que ponen en cuestión la pro-puesta gradualista del ritmo evolutivo, proponiendo en contraposición otros modelos alternativos, como son el rupturismo o la teoría de equi-librios puntuados. Y en tercer lugar, recogeremos las críticas a la teoría darwiniana desde los creacionismos fundamentalistas y la teoría del di-seño inteligente (DI), posturas que consideran que la selección natural no puede ser una teoría verdadera, en la medida en que contradice las afirmaciones de la Biblia (es el caso de los creacionismos fundamenta-listas), o bien niegan que la teoría de la selección natural aporte pruebas suficientes para ser considerada una teoría científica aceptable, consi-derándola más bien una hipótesis teórica similar y alternativa a la que

85
© Universidad de Deusto - ISBN 978-84-9830-383-4
ellos proponen (teoría del Diseño Inteligente), defendiendo, por ello, una igualdad de trato en los programas de estudio en la enseñanza se-cundaria.
2. LA CONDICIÓN CIENTÍFICA DE LA TEORÍA DE LA SELECCIÓN NATURAL
Posiblemente haya sido Popper el filósofo que más ha puesto en tela de juicio la condición de teoría científica predictiva del darwinismo, sien-do también Popper el filósofo de la ciencia que más se ha ocupado de la teoría neodarwinista. Popper califica a esta teoría de «programa metafí-sico de investigación por atribuirle una incapacidad predictiva en lo que respecta a las consecuencias esenciales del proceso evolutivo: la adapta-ción y la especiación»5.
Es bastante común y repetida la acusación de que la propuesta darwi-niana sobre la supervivencia del más apto es una simple tautología. La se-lección natural indica que sobreviven los más aptos, y cuando se pregunta quiénes son los más aptos, la respuesta es los que sobreviven. Expuesto así, el cariz tautológico es evidente. Pero, como indican Toro y López-Fanjul, es posible desligar los conceptos de adaptación y de eficacia bio-lógica, definiéndolos independientemente6. Un modo de adaptación es la solución concreta que un organismo vivo construye de cara a camuflarse en su entorno ambiental, y el valor de tal estrategia se puede valorar en términos de calidad de tal proyecto. Las críticas de Popper al evolucio-nismo en este punto adolecen de historicismo, puesto que parece que lo examina en términos de lo que ha ocurrido en el pasado, y en ese punto parece evidente que el neodarwinismo queda circunscrito a la descripción y explicación de los modos posibles en que un determinado proceso de especiación se ha dado.
5 Toro, M. A./López-Fanjul, C., o.c., p. 263. Cfr. Popper, K. R., «Darwinism as a meta-physical research programme», sección 37 de la obra de K. R. Popper Unended Quest, ci-tada en la nota 14, «Evolution and the tree of knowledge», sección 7 de la obra de Popper Objective Knowledge. An Evolutionary approach (Oxford University Press, edición revi-sada, 1974; traducción española: Conocimiento objetivo, Madrid, Tecnos, 1974); Vallejo, Fernando, La tautología darvinista y otros ensayos de biología, Madrid, Taurus, 2002.
6 Esta idea ya ha sido defendida por Fco. J. Ayala en el último capítulo de la obra Evolution,de Th. Dobzhansky, Fco. J. Ayala, G. L. Stebbins y J. W. Valentine, W. H. Freeman and Company, San Francisco, 1977, pp. 504-505, y también por R. C. Lewontin, «Adaptation», Scientific American, 239: 157169, 1978 (traducción española en Investigación y Ciencia,26: 138-149, 1978).

86
© Universidad de Deusto - ISBN 978-84-9830-383-4
Pero la evolución sigue su marcha y está funcionando en el presente y de cara al futuro. Y el darwinismo tiene la pretensión de tener capaci-dad de predicción de los fenómenos adaptativos. Es evidente que la direc-ción que siga el proceso selectivo depende del modo como cambien las circunstancias del entorno, y estos cambios son difíciles de predecir, pero cada vez es esto más factible. Por otra parte, «aunque cualquier suceso de especiación es único, ello no imposibilita a priori su predicción, puesto que ésta se hace en función de unas fuerzas generales que actúan esencial-mente de la misma manera en cualquier comunidad de organismos»7. Los cambios ambientales que se están sucediendo en la actualidad son debidos a la actuación humana, y estos cambios son previsibles, por desgracia, y de ellos depende en lo fundamental la presión que actualmente ejerce la selección natural sobre la mayoría de las especies vivas.
Otra dificultad que Popper propone a la capacidad predictiva del darwinismo hace referencia a lo que denomina el «dualismo genético». Popper supone que el neodarwinismo predice cambios cuya dirección es aleatoria, en la medida en que la mutación se produce al azar. Pero, en opinión de Toro y López-Fanjul, «es la variación del medio, a través del mecanismo de selección natural, la que determina la dirección del cam-bio evolutivo y aquella puede ocurrir, y de hecho muchas veces ocurre, en una determinada dirección»8. La hipótesis popperiana del «dualismo ge-nético» distingue dos tipos de mutaciones: las anatómicas y las del com-portamiento, y da por hecho que ambas aparecen independientemente, siendo las segundas menos perjudiciales que las primeras. Según eso, las segundas son las que primero se incorporan al acervo genético y dirigen, en cierta medida, la evolución. Popper pone como ejemplo la extensión del cuello de las jirafas, y se pregunta si primero es el hábito de comer ho-jas altas seguido de un cuello más largo, o al revés, si primero es tener el cuello largo y luego la aparición de un hábito alimenticio propiciado por ese rasgo anatómico. Los críticos de Popper, Toro y López-Fanjul, entien-den que estos planteamientos popperianos son bastante confusos, aparte de que «no está claro en qué se diferencian del razonamiento darwinista tradicional»; y, por otro lado, «no parece que haya razón alguna a favor de los mutantes etológicos como menos nocivos que los morfológicos, o vi-ceversa»; y además, en tercer lugar, «no hay por qué suponer que las mu-taciones etológicas se incorporen antes que las morfológicas»9.
Lo que Toro y López-Fanjul deducen es que Popper está dando por supuesto en sus planteamientos la existencia de macromutaciones de
7 Toro, M. A./López-Fanjul, C., o.c., p. 265. 8 Ibídem, 266. 9 Ibídem, 266-267.

87
© Universidad de Deusto - ISBN 978-84-9830-383-4
efectos catastrofistas o drásticos, cosa que, desde los planteamientos del darwinismo ortodoxo, no se acepta como un hecho comprobado, sino que más bien los diversos rasgos biológicos o comportamentales suelen ser resultado de la acumulación gradual de pequeñas mutaciones, sobre las que a continuación actúa la selección natural. «Las mutaciones “mons-truosas” aparecidas durante ochenta años de investigación genética, no pasan de ser formas aberrantes que no cabe confundir con novedades evolutivas»10.
3. LAS DIVERSAS PROPUESTAS SOBRE EL RITMO DEL PROCESO EVOLUTIVO
Aceptar el evolucionismo no supone estar de acuerdo sobre cómo se produce el ritmo evolutivo que lleva a la generación de nuevas especies. Este problema ya estuvo presente en los tiempos de Darwin, pero en la actualidad se ha renovado con especial profundidad y virulencia. Ya vi-mos que Darwin fue siempre partidario de la postura que se denomina gradualista, consistente en defender que los cambios en el proceso evo-lutivo se producen de forma continuada y gradual, sin grandes saltos o rupturas.
En cambio, el genetista Hugo de Vries defendió una postura rupturis-ta, apoyándose en la existencia de macromutaciones, capaces de aportar cambios profundos en ciertos individuos y de formar especies nuevas en un lapso breve de tiempo. Esta postura rupturista o saltacionista, opues-ta al gradualismo, fue defendida tempranamente por R. B. Goldschmidt, quien consideraba que las nuevas especies surgen de modo brusco y re-pentino como consecuencia de la aparición de un tipo de individuos, de-nominados monstruos esperanzadores, portadores de una macromutación de gran efecto, o bien afectados por una reordenación genética global, lle-gando de este modo a convertirse en iniciadores de nuevos linajes y espe-cies evolutivas11.
Esta teoría rupturista o saltacionista ha representado una alternativa al gradualismo de Darwin desde que éste propuso su teoría selectiva. La defendió nada menos que el amigo y principal defensor de Darwin, T. H. Huxley, quien escribía al propio Darwin diciéndole que su teoría «está innecesariamente sobrecargada con la suposición de que Natura
10 Ibídem, 267. 11 Cfr. Golschmidt, R. B., The Material Basic of Evolution, Yale University Press,
New Haven, 1940 (traducción española en Madrid, Espasa-Calpe, S.A., 1943).

88
© Universidad de Deusto - ISBN 978-84-9830-383-4
non facit saltus»12, esto es, que en la naturaleza no hay discontinuidades. Por el contrario, como ya vimos en su momento, los genéticos más impor-tantes del primer tercio de este siglo (Bateson, de Vries, Morgan Johanssen,etc.) eran totalmente partidarios de teorías evolutivas saltacionistas.
Estos planteamientos saltacionistas de los primeros genetistas fueron continuados por Goldschmidt, y, en parte, hoy en día está siendo asumida por la tercera de las teorías que veremos enseguida, la de los equilibriospuntuados, o equilibrios interrumpidos. Normalmente los defensores de la teoría saltacionista suelen considerar al individuo como la unidad de la evo-lución, a diferencia de los neodarwinistas, para quienes este lugar lo ocu-pa la población. Esta es la razón de por qué a veces las teorías saltacionis-tas pueden resultar, según Toro y López-Fanjul, más atractivas, «puesto que introducen una discontinuidad radical entre el hombre y los restantes seres vivos»13. Pero entendemos que la diferencia cualitativa entre hom-bre y demás animales se puede defender tanto desde el gradualismo, como desde el saltacionismo, como con los equilibrios puntuados.
La teoría de los equilibrios puntuados, o equilibrios interrumpidos (punctuated equilibria)14, fue planteada recientemente por primera vez por S. J. Gould (Universidad de Harvard, USA) y por N. Eldredge (Mu-seo Americano de Historia Natural, Nueva York). Pretende situarse en medio, de forma sintética, de las dos teorías anteriores, la gradualista y la saltacionista. Defiende que en la historia evolutiva se dan largos períodos de normalidad, es decir, de acumulación progresiva y gradual de cambios y mutaciones (mejoras en las especies, anagénesis), junto a momentos en
12 Cita tomada de Toro, M. A./López-Fanjul, C., o.c., 273, nota 39. 13 Ibídem, p. 273, nota 39. 14 La traducción al castellano de esta teoría no está del todo consensuada. Aunque la
mayoría de los autores la traducen como equilibrios puntuados, otros, como Toro y López-Fanjul, prefieren denominarla equilibrios interrumpidos: o.c., p. 271, nota 30. Estos auto-res entienden que la expresión inglesa, punctuated equilibria, «alude a las interrupciones bruscas de largos períodos de estabilidad, por cuanto el participio punctuated se refiere al modo de separar unas fases de otras mediante pautas o puntos. Las traducciones literales puntuado o pausado, ni transmiten fácilmente esa imagen de intermisión ni es ese su sig-nificado más común. Los adjetivos puntual o pautado nada tienen que ver con lo que se quiere representar; el primero de ellos se ha usado en la jerga de laboratorio (como tam-bién el barbarismo puntuacional) por su semejanza fonética con la palabra inglesa. Por ultimo, el calificativo intermitente sugiere una tendencia cíclica que no es del caso. En nuestra opinión, los términos interrumpido, discontinuo, discreto, reflejan más claramente la idea contenida en la expresión original, aunque los dos últimos sólo lo hacen en su acep-ción matemática menos general. Por estas razones, nos parece que la traducción más aceptable de punctuated equilibria es equilibrios interrumpidos y así la utilizaremos aquí» (o.c., pp. 272-273, nota 30). De todos modos, aunque los argumentos de estos autores son de gran peso, el nombre que en el entorno científico español se está imponiendo es el de equilibrios puntuados, y es el que seguiremos aquí.

89
© Universidad de Deusto - ISBN 978-84-9830-383-4
que se producen rápidamente mutaciones importantes que da lugar a la aparición de especies nuevas (cladogénesis)15. Claro que la rapidez de que estamos hablando es relativa y comparativa, puesto que hablamos de pe-ríodos que abarcan miles de años. La primera vez que defendieron esta teoría fue en 1972, y la completaron en 199316. Según estos autores, en el proceso de formación de las especies no sólo se da un proceso gradual, evolutivo, sino que se producen conjuntamente revoluciones, saltos, ex-plosiones repentinas de cambio rápido. De modo que la evolución es una síntesis o conjunto de largos períodos de equilibrio estático con momentos de rápidos cambios, en los que aparecen nuevas especies17.
El darwinismo ortodoxo, defensor de la teoría sintética, desde la óp-tica de la genética de poblaciones, apoya el gradualismo. Pero, a pesar de ello, tampoco es necesario para el neodarwinismo defender que el cambio evolutivo tenga que darse necesariamente de forma gradual, pudiéndose producir determinados cambios de forma rápida, relativamente. Por eso que hay quienes desde el darwinismo ortodoxo y gradualista consideran que la teoría de los equilibrios puntuados no se diferencia demasiado de la teoría gradualista, entendida de forma flexible.
De todos modos, los defensores de esta nueva teoría la consideran ori-ginal y diferente al gradualismo, apoyándose en dos características de los registros fósiles:
1) «la morfología de las especies fósiles suelen mantenerse con muy poca variación durante largos períodos de tiempo, del orden de va-rios millones de años (estatismo)»;
2) «los cambios morfológicos experimentados por un determinado li-naje fósil ocurren durante lapsos relativamente cortos (discontinui-dades del registro), del orden de cincuenta a cien mil años, y están comúnmente asociados con fenómenos de especiación»18.
Estas evidencias fósiles estarían mostrando, según estos autores, que no se da en la historia de la evolución esa supuesta gradualidad en el pro-ceso evolutivo. A lo más, se daría tal gradualidad en el ámbito de la mi-croevolución, pero no en el de la macroevolución, esto es en el proceso de formación de especies y linajes nuevos. Y, de hecho, si se observa la
15 Cfr. Sampedro, Javier, o.c., cap. 5.º. 16 Cfr. Nature, 18 de noviembre de 1993. El texto donde de modo más claro y amplio
se explicita esta teoría es el artículo de S. J. Gould, «El equilibrio puntuado y el enfoque jerárquico de la macroevolución», Revista de Occidente, 1982, n.º 18-19, 121-148.
17 Cfr. Michael Gross, «El doble juego de una proteína», Mundo Científico, 1999, n.º 204, 28-31; Henry Gree, «Explosiones repentinas de cambios. La nueva visión, 21 años después», El País, 8-XII-1993, Futuro, p. 3.
18 Toro, M. A./López-Fanjul, C., o.c., 262.

90
© Universidad de Deusto - ISBN 978-84-9830-383-4
historia de las diferentes especies animales en el ámbito de nuestro pla-neta, su aparición histórica no se ha dado de forma escalonada y gradual, sino que parece que la aparición más numerosa se dio en el momento que se ha denominado la explosión cámbrica19. Este fenómeno consiste en el curioso hecho de que la mayoría de los linajes zoológicos existentes en la actualidad aparecieron súbitamente en el período geológico denominado Cámbrico. El fenómeno es tanto más curioso por cuanto, hasta esa época geológica, las especies vivas de que se tenía noticias eran tan sólo fósiles de microbios unicelulares. Y a partir de ese momento, unos 530 millo-nes de años, se produce la gran explosión de la mayoría de los antepasados animales que existen en la actualidad. De tal modo que «la aparición relati-vamente súbita, hacia el principio del Cámbrico —hace unos 530 millones de años—, de casi todos los bauplanes (planes generales de diseño animal) que existen o han existido sobre la Tierra sigue siendo un profundo miste-rio, por más que el planeta estuviera hiperpoblado de microbios durante al menos los 3.000 millones de años anteriores. El fenómeno es tan extraor-dinario que hasta en la más sobria literatura técnica recibe un nombre que parece ideado por un creativo publicitario: la explosión cámbrica»20.
Todas estas evidencias son las que hacen que la teoría de los equili-brios puntuados sea cada vez más aceptada y parece la teoría que se está imponiendo en la actualidad, aunque todavía tenga que ser probada y ma-tizada en muchos aspectos, y no deje de tener enemigos radicales.
Como puede verse, la teoría darwinista de la selección natural sigue siendo una teoría aceptada mayoritariamente como un hecho científico, pero que dista mucho de poder explicar todos los fenómenos que se pro-ducen en el proceso evolutivo. Y es bueno que no se la considere como un dogma inamovible, sino como una teoría que tiene que estar continua-mente falsándose y poniéndose a prueba.
4. CRÍTICAS AL DARWINISMO DESDE LOS FUNDAMENTALISMOS RELIGIOSOS
4.1. El conflicto entre evolución y religión
Ya vimos en el capítulo anterior que, desde la aparición de El origen de las especies, se produjo un fuerte rechazo de las teorías darwinistas de parte de la facción más conservadora del cristianismo. Esta oposición se
19 Cfr. Sampedro, Javier, Deconstruyendo a Darwin, o.c., caps. 6.º y 8.º. 20 Ibídem, p. 78.

91
© Universidad de Deusto - ISBN 978-84-9830-383-4
ha ido manteniendo desde entonces, agudizándose progresivamente en los entornos de diversas confesiones evangélicas, que desde una postura reli-giosa fundamentalista sostienen no haber más verdad que la literalidad bí-blica, por lo que resulta errónea cualquier teoría que se le oponga. De ese modo, surge la cuestión de si hay compatibilidad entre darwinismo y fe cristiana, y en general entre ciencia y fe21.
Las posturas más beligerantes contra el darwinismo se sitúan en las con-fesiones evangélicas de algunos estados de los USA, los denominados gru-pos creacionistas fundamentalistas, empeñados en conseguir leyes civiles que prohíban la inclusión del darwinismo en los temas de enseñanza de las escuelas, o que admitan al mismo tiempo que el darwinismo el fundamenta-lismo bíblico o la teoría creacionista22. Más recientemente, este fundamen-talismo radical ha dado paso a una postura más moderada y pretendidamen-te asumible, que recibe el nombre de teoría del Diseño Inteligente23.
Estas teorías conservadoras han recibido críticas desde muchos pun-tos de vista, pero sobre todo desde un cientifismo extremo, liderado por el sociobiólogo inglés Richard Dawkins, junto al filósofo norteamericano Daniel Dennett y otros más24. Sus críticas no se han limitado al ámbito de
21 Cfr. Ruse, M., ¿Puede un darvinista ser cristiano? La relación entre ciencia y re-ligión, Madrid, Siglo XXI, 2007; Sequeiros, L., ¿Puede un cristiano ser evolucionista?,Madrid, PPC, 2009; Ayala, Fr. J., Darwin y el Diseño Inteligente. Creacionismo, cristianis-mo y evolución, Madrid, Alianza, 2007; Küng, H., El principio de todas las cosas. Ciencia religión, Madrid, Trotta, 2007; Gould, S. J., Ciencia vs. Religión. Un falso conflicto, Bar-celona, Crítica, 2000; Fernández Rañada, A., Los científicos y Dios, Madrid, Trotta, 2008 (nueva edición); Buskes, Chris, La herencia de Darwin. La evolución en nuestra visión del mundo, Barcelona, Herder, 2009.
22 Cfr. Ayala, Fco. J., Darwin y el Diseño Inteligente. Creacionismo, cristianismo y evolución, o.c., cap. 8.º, «Creacionismo y fundamentalismo en Estados Unidos»; Sequei-ros, L., o.c., cap. 4.º, «La resistencia al evolucionismo: las marejadas creacionistas»; Davies, M. W., Darwin y el fundamentalismo, Barcelona, Gedisa, 2004. De todos modos, la palabra creacionismo tiene también que ser matizada, porque, frente a un creacionismo fundamentalista, hay que tener en cuenta lo que podríamos llamar un creacionismo ilustra-do, que defiende la fe en la creación del mundo por Dios, pero no al modo fundamentalista, sino compatibilizando el nivel científico con el teológico.
23 Sobre la teoría del Diseño Inteligente, cfr. Ayala, Fco. J., o.c. Los tres más impor-tantes teóricos del Diseño Inteligente son Ph. E. Johnson (Juicio a Darwin, Madrid, Homo Legens, 2007), M. Behe (Darwin’s Box. The Biochemical Challange to Evolution, N. York, The Free Press, 1996), y W. A. Dembski (Diseño Inteligente. Respuesta a las cuestiones más espinosas del diseño inteligente, Madrid, Homo Legens, 2006).
24 Cfr. Eagleton, T., Reason, Faith, and Revolution: Reflections on the God Debate,New Haven, Yale University Press, 2009; Armstrom, K., The Case for God, Nueva York, Knopf, 2009 (trad. cast.: En defensa de Dios. El sentido de la religión, Barcelona, Paidós, 2009); McGrath, A. E./McGrath, J. C., The Dawkins Delusion? Atheist Fundamentalism and the Denial of the Divine, Londres, SPCK, 2007; McGrath, Alister E., «Los ateísmos de superventas. El nuevo cientifismo, Concilium, 2010, n.º 337, 555-567.

92
© Universidad de Deusto - ISBN 978-84-9830-383-4
las publicaciones escritas, sino que han originado también agresivas cam-pañas publicitarias, consistentes en colocar carteles propagandísticos en autobuses urbanos de diversas ciudades europeas, con consignas como «There is no God» (Dios no existe), con la consiguiente contraofensiva de algunos sectores cristianos más conservadores.
Estamos, por tanto, ante un tema candente y de gran actualidad, pero también de enorme complejidad25. Las relaciones entre la ciencia y la re-ligión ha sido estudiada con suficiente amplitud y profundidad como para que nos tengamos que detener aquí demasiado en este punto. Simplemen-te voy a limitarme a presentar de modo esquemático las posturas más im-portantes defendidas en este debate entre la ciencia y la fe, y en situar el esquema básico desde el que hay que entenderlo.
Considero que el marco de reflexión más interesante y adecuado para abordar la relación entre ciencia y fe es el marco epistemológico, esto es, presentar claramente la distinción entre estos dos modos de saber y de dar cuenta de la realidad que constituyen la ciencia, por un lado, y la fe y la teología, por otro, y reflexionar a continuación, desde esos parámetros, a cerca de si la teoría científica de la selección natural puede ser compatible o no con una mirada de fe y su reflexión racional correspondiente, como es la teología26. Entiendo que este punto es el aspecto clave y fundamental a la hora de plantear y de tratar de resolver la cuestión de si son compati-bles o no, o de cómo conjugar la teoría de la evolución de Darwin y la fe cristiana.
La tesis que considero más acertada es la que defiende la compatibi-lidad ente darvinismo y cristianismo, porque ambos planteamientos co-rresponden a planos epistemológicos diferentes, que se pueden y se deben complementar e integrar. La condición más adecuada para una integra-ción correcta consiste en evitar las dos posturas extremas: el materialismo cientifista y reduccionista, consistentes en afirmar que no hay más reali-
25 La bibliografía sobre la historia de la relación entre el evolucionismo y religión es muy amplia. Para una visión global del problema, cfr. Barbour, I. G., Religión y ciencia,Madrid, Trotta, 2004; Id., Problemas de religión y ciencia, Santander, Sal Terrae, 1972; Evolución y fe, Concilium, 2000, n.º 284 (número monográfico); Núñez, Diego, «La re-ligión y la ciencia (Historia de las controversias entre ambas)», Mundo Científico, 1996, n.º 166, 247-256; Gutiérrez Lombardo, Raúl, «La teoría de la evolución y la Iglesia Cató-lica», Ludus Vitalis, X (2002), n.º 17, 111-122; Riaza, José María, La Iglesia en la historia de la ciencia, Madrid, BAC, 1999, Nicolau Pous, Fr., Iglesia y ciencia a lo largo de la his-toria, Barcelona, Ed. Claret, 2002 (Ed. SCIRE, 2003); Sequeiros, L., o.c., caps. 1.º y 2.º; Núñez, Diego, «La religión y la ciencia (Historia de la controversia entre ambas)», MundoCientífico, 1996, marzo, n.º 166, 247-256; Haught, John F., Cristianismo y ciencia. Hacia una teología de la naturaleza, Santander, Sal Terrae, 2009; Udías Vallina, Agustín, Cienciay religión. Dos visiones del mundo, Santander, Sal Terrae, 2010.
26 Cfr. Udías Vallina, A., o.c., cap. 2.º.

93
© Universidad de Deusto - ISBN 978-84-9830-383-4
dad que la realidad material o física, y tampoco ningún otro saber legíti-mo y verdadero que el que se basa en el método de las ciencias naturales, esto es, el método empírico; y el fundamentalista religioso, también deno-minado creacionismo, defensor de una interpretación literal de la Biblia y de una intervención milagrera de Dios en los acontecimientos del mundo.
En consecuencia, la postura de integración entre ciencia y fe que de-fiendo aquí, supone dejar claro, por el contrario, que la ciencia no es más que un tipo de saber junto a otros (filosofía, teología, estética, etc.), a quien le corresponde dar cuenta de la realidad en un nivel determinado de la misma (en concreto, la dimensión fáctica), por lo que no le corresponde dirimir cuestiones a acerca del sentido de la realidad (tarea propia de la fi-losofía y de la religión); e igualmente, supone superar, desde el campo de la opción religiosa, una concepción literal y fundamentalista de los conte-nidos de la Biblia y replantear de otro modo la idea de Dios y su interven-ción en el mundo.
4.2. Precisiones epistemológicas
Como ya hemos avanzado, considero que el problema de la correcta conjugación entre ciencia y fe/teología pasa por un correcto entendimien-to de los diferentes niveles epistemológicos de la realidad.
Para entender esta afirmación, podríamos referirnos a un ejemplo sen-cillo. Si nos ponemos a contemplar, por ejemplo, un árbol (un manzano, o un cerezo), advertimos sus múltiples caras o enfoques desde los que pode-mos y solemos contemplarlo: la cara estética, económica, biológica, filo-sófica, teológica, etc. Desde cada uno de esos enfoques, podemos descri-bir el árbol de forma diferente.
Si nos preguntan cuál es el enfoque más verdadero, nos quedaremos en principio extrañados por la pregunta, puesto que la respuesta evidente es que todos son igualmente verdaderos, cada uno en su nivel. Y el con-junto de los diversos puntos de vista es lo que constituye ese árbol del que estamos hablando.
En vez de centrarnos en un objeto particular del mundo, podemos re-ferirnos al conjunto de la realidad. La ciencia nos puede decir cómo es la realidad y cómo funciona. Por ejemplo, cuál es la velocidad de la luz, cómo es la estructura del átomo, cuántos cromosomas tiene el cariotipo de una especie viva, etc., pero no podrá responder, porque no le correspon-de, por qué esas realidades son así y no de otra manera; y, en definitiva, no podrá responder por qué la realidad existe en vez de no existir, por qué hay ser y no nada. Esa es precisamente la pregunta filosófica por excelen-cia, según Leibniz, Heidegger y otros filósofos. En este tipo de cuestio-

94
© Universidad de Deusto - ISBN 978-84-9830-383-4
nes es donde experimentamos la pluralidad de niveles de la realidad, y la complejidad del esfuerzo intelectivo del ser humano por entenderla y captarla.
En realidad, la razón humana, esto es, el modo como tratamos de cap-tar la realidad y dar cuenta de la misma, es así de compleja. En sus inicios, comenzó siendo una visión racional única y global, con el nombre de filo-sofía (amor a la sabiduría), y más adelante se fue desgajando y explicitan-do en diversos ámbitos del saber, que no siempre se han interrelacionado y complementado de un modo pacífico y fructífero. La epistemología se-ría la parte de la racionalidad o del saber que intenta delimitar las diferen-tes parcelas de los saberes, las normas adecuadas para no interferir entre sus fronteras, así como las reglas de complementariedad entre los diversos niveles de la racionalidad.
Con este telón de fondo de la epistemología es como tenemos que plantearnos la cuestión de la relación entre darwinismo y cristianismo, puesto que en el fondo se trata de cómo conjugar una teoría científica con otra filosófica o teológica. Dentro de lo que se lleva denominando el re-torno de lo sagrado, cada vez cobra más actualidad la discusión sobre la relación entre las teorías científicas y las filosóficas y teológicas, sobre todo en los ámbitos más problemáticos, como son el origen, la estructura y la historia del universo, el origen de la vida, su evolución y complejifi-cación, el origen del ser humano, la naturaleza de la mente y su conjuga-ción con lo somático, los múltiples problemas que se plantean en el ám-bito de la bioética, la eutanasia, etc. Las mayores dificultades a la hora de concordar posiciones no provienen de los contenidos específicos de las diferentes posturas cosmovisionales desde las que cada interlocutor habla e interpreta la realidad, sino sobre todo del diferente modo como se en-tiende correctamente la distinción y conjugación de los planos o enfoques específicos de la ciencia, la filosofía y la teología27. Aunque, en el fondo, los dos aspectos del problema (contenidos cosmovisionales y distinción epistemológica) están interrelacionados y se influyen mutuamente.
Por eso, en la clarificación de las diferencias entre las diversas pro-puestas que hoy día contienden sobre la comprensión de la evolución y el lugar del ser humano en ella, es fundamental y básico hacer una buena la-bor de clarificación sobre la distinción de planos epistemológicos. Es muy probable que si llegamos a entendernos en este punto, resulte mucho más fácil clarificar las diferencias e iniciar el camino que nos lleve a la posible superación de las mismas.
27 Para quien quiera añadir a la discusión la relación con la estética, puede ver Capelle, Ph.,«La relation philosophie-théologie: histoire tourmentée, géographie nouvelle», Esprit, 117 (2007), n.º 171, 1-8.

95
© Universidad de Deusto - ISBN 978-84-9830-383-4
Considero que una correcta mirada epistemológica parte de las si-guientes precisiones:
a) la ciencia se limita a, o le corresponde solamente dar cuenta de la realidad fáctica, de la constatación de los hechos que pueden ser observados y comprobados empíricamente por cualquier observa-dor humano, así como las leyes que rigen la interrelación entre los diversos hechos de la realidad. Dejemos de lado, de momento, la distinción y diferenciación entre los diversos tipos de ciencias, las naturales y las humanas, porque nos complejificaría demasiado el tema y no contribuiría a aclararlo;
b) en cambio, la filosofía se mueve en el terreno de las interpretacio-nes de tales hechos; lo que se denomina el ámbito del sentido. Ello hace que las exigencias de validación de sus afirmaciones sea de diferente género que las verdades científicas, en la medida en que la evidencia de su verdad no pertenece al campo de la constata-ción empírica o sensible, sino más bien a su coherencia interna y a su plausibilidad con el funcionamiento de los hechos que trata de interpretar desde una teoría sistémica más abarcadora. Pero, así como en el campo de las afirmaciones científicas cabe una demos-tración empírica casi definitiva (aunque también en el ámbito cien-tífico se halla el ser humano en búsqueda constante), y los científi-cos llegan en su mayoría a una constatación objetiva (la estructura del ADN, la del átomo de Hidrógeno, la raíz cuadrada de 25, etc.), no ocurre lo mismo en el terreno de la filosofía, donde se pueden dar, y se dan, sobre un mismo hecho varias interpretaciones. Y ello no debe ser considerado (como a veces se hace desde determina-das posturas científicas, o más bien filosóficas, reduccionistas, ma-terialistas) como una limitación e insuficiencia de la filosofía, sino más bien como una consecuencia de su específico modo de dar cuenta de la realidad. El dato específico del saber filosófico está en que interpreta la realidad desde un a priori cosmovisional en el que se apoya y desde el que da sentido a todos los datos de la rea-lidad. Y en la medida en que tal cosmovisión no parte de la mera experiencia empírica, sino que tiene otros muchos componentes que la sostienen (educación cultural, creencia religiosa, opciones y experiencias personales, etc.), y desde el cual cada sujeto huma-no termina de construir un mundo que se le presenta abierto y, en cierto modo, sin terminar, hay una parte de opción, de creación y de libertad inevitable en toda interpretación filosófica del mundo. Por eso, no debe extrañar que la filosofía esté constituida por una inevitable y legítima pluralidad de puntos de vista, a la que se opo-

96
© Universidad de Deusto - ISBN 978-84-9830-383-4
nen los diferentes dogmatismos y fundamentalismos, y pluralidad que, por otro lado, no hay que confundir con una postura relativis-ta disolvente;
c) por su parte, la teología se mueve, al igual que la filosofía, en el ámbito de las interpretaciones, partiendo también y apoyándose en opciones aprióricas, cosmovisionales de la realidad, diferenciándo-se en este punto de la filosofía en que forma parte de dichas cos-movisiones la fe en Dios y en su acción sobre la realidad, apelan-do para estas afirmaciones a fuentes reveladas, aunque también (al menos en el cristianismo) mediadas por la racionalidad filosófica, que entiende que tales afirmaciones teológicas, aunque no son de-mostrables sí son razonables, esto es, coherentes con la racionali-dad humana (fides quaerens intelectum).
Estas distinciones, a pesar de su aparente coherencia y plausibilidad, están muy lejos de ser reconocidas y aceptadas por todos. Podríamos co-legir que hay varias razones, sobre todo dos, que explican en gran me-dida estas discrepancias. La primera tiene que ver con la extraordinaria labilidad de las fronteras entre cada mirada epistemológica. No todos aceptan la distinción entre hechos e interpretaciones, hechos y valores, o entienden tal distinción de maneras muy diversas y confusas. De he-cho, cuando se quiere hacer ver que la ciencia no puede responder a las cuestiones sobre por qué hay realidad en vez de la nada, o por qué la rea-lidad es así y no de otra manera, suelen algunos considerar que son cues-tiones no resueltas todavía por la ciencia, pero que quizás en el futuro se podrán responder desde la misma ciencia, no siendo conscientes de que tales cuestiones pertenecen a niveles epistemológicos diferentes, que no hay que mezclar. Y la segunda consiste en entender que la propia dife-renciación de planos epistemológicos supone ya de entrada un modo es-pecífico de resolver o de entender el conjunto del problema y una visión cosmovisional y totalizante de la realidad. Es decir, tiene el riesgo de ser una forma entre otras (como toda propuesta filosófica) de entender y de resolver el problema, con lo cual parecería ser a la vez juez y parte de la distinción.
Entre las posturas más reacias a la aceptación de esta distinción de planos se hallan dos planteamientos extremos ya indicados antes: el na-turalismo filosófico y los creacionismos fundamentalistas. El primero no admite la distinción de planos de la realidad, y da por hecho que no existe más realidad que la que se puede demostrar con el método de las ciencias naturales. Y consideran los defensores de esta postura que al querer pre-sentarles, o hacerles advertir, la existencia del nivel del sentido, les esta-mos metiendo de rondón un ámbito imaginario, y por tanto no existente,

97
© Universidad de Deusto - ISBN 978-84-9830-383-4
como es el de la racionalidad filosófica o metafísica, donde se sitúa tam-bién el ámbito de las religiones. Para ellos se trata, sin más, de propuestas metafísicas sin sentido. Por tanto, para esa postura no habría más raciona-lidad que la propia de las ciencias, quedando el ámbito de los valores pos-tergado a la región de los gustos o preferencias irracionales.
Las limitaciones de esta postura son evidentes: la primera consis-te en no advertir que las bases teóricas de su planteamiento no se pue-den demostrar empíricamente, son propuestas filosóficas que escapan a la verificación científica; y además, la evidente y necesaria distinción entre ciencia y filosofía se advierte precisamente cuando se plantean las preguntas sobre los límites de la ciencia. Está claro que las diferen-tes ciencias pueden llegar a descubrir todas las leyes del universo y de sus diferentes partes, no sólo en el ámbito físico, cosmológico, quími-co, biológico, sino también psicológico e histórico-social. En todos es-tos campos, y en el conjunto total del universo, no podrá ir nunca más allá de constatar las leyes fácticas que rigen su funcionamiento, pero no puede evitar que el ser humano se plantee preguntas más radicales: ¿por qué existe el universo en vez de no existir? ¿por qué es así y no de otra manera? ¿qué sentido tiene nuestra existencia, a dónde vamos, de dónde venimos, para qué todo, por qué existe el mal y el dolor, etc.? Estas cues-tiones escapan a la mera reflexión científica, aunque no a los científicos, que como seres humanos pueden y deben planteárselas, debiendo ser conscientes de que el ámbito de las preguntas y de las respuestas escapa a los dominios de la ciencia, y se sitúan en el terreno de las propuestas de sentido filosóficas y teológicas.
Por otro lado, los creacionismos fundamentalistas, aunque no nieguen en principio la distinción de planos, consideran que la única verdad se ha-lla en el nivel religioso, presente en los libros sagrados de turno, interpre-tados de modo literal y acrítico. De modo que si existe discrepancias entre ciencia, filosofía y libros sagrados, son éstos (interpretados dogmática-mente por la autoridad religiosa de turno) los únicos jueces desde los que dirimir el conflicto28.
Ambas posturas no aciertan a entender la realidad en su justa comple-jidad e interrelación. Según las precisiones epistemológicas que hemos presentado, a la hora de resolver la cuestión que nos ocupa, la posibilidad o no de integrar y hacer compatibles el evolucionismo científico y el cris-tianismo, tenemos que distinguir dos momentos o planos de reflexión:
28 Para un estudio exhaustivo y sistemático de las diferentes posturas sobre la relación entre ciencia y fe, cfr. I. Barbour, Problemas de religión y ciencia, Salamanca, Sígueme, 1966; Id., Religión y ciencia, Madrid, Trotta, 2004; Id., El encuentro entre ciencia y reli-gión. ¿Rivales, desconocidas o compañeras de viaje?, Santander, Sal Terrae, 2000.

98
© Universidad de Deusto - ISBN 978-84-9830-383-4
a) el primero tiene que centrarse en saber aceptar y distinguir adecua-damente entre los dos niveles epistemológicos indicados, el cientí-fico y el filosófico. Este es un paso fundamental, sin el cual no es posible entenderse ni resolver adecuadamente no sólo la cuestión que nos ocupa, sino cualquier conflicto entre una teoría científica y otra filosófica o teológica;
b) y dentro ya del nivel filosófico, como ámbito del sentido, en el que convergen las diversas interpretaciones, nos encontraríamos con una amplia pluralidad de cosmovisiones filosóficas, y también teo-lógicas, que tratan de dar sentido a la realidad, tal y como nos la presentan las teorías científicas.
En el nivel primero, se sitúa la contrastación y la reflexión sobre los datos científicos, así como la constatación de que la ciencia no es sufi-ciente para dar cuenta de la totalidad de la realidad.
Y en el segundo, la inevitable necesidad de admitir una pluralidad de interpretaciones, entre las cuales estaría la religiosa, pero también la atea o agnóstica. Según esto, las conclusiones a las que llegamos son varias:
a) a la hora de dar sentido al hecho de la evolución, tan legítima es la postura religiosa como la atea o agnóstica, porque, ante la opa-cidad inevitable de la realidad, cabe igualmente concluir que todo el proceso evolutivo, así como toda la historia del universo, es re-sultado del azar o de cualquier otro mecanismo impersonal, como también optar por la postura religiosa;
b) las dos no pueden ser igualmente verdaderas, puesto que son con-tradictorias, pero desde nuestra mirada intrahistórica no tenemos posibilidades de dirimir entre la verdad de una u otra de las in-terpretaciones; por eso, la única salida es la opción por una de las dos, la fe, o la no fe; por tanto, ambas opciones pertenecen al mis-mo nivel epistemológico: el de la fe y la opción personal;
c) en conclusión, son compatibles en principio la teoría darwinista y la cosmovisión cristiana sobre el mundo y la historia del universo y el desarrollo de la biosfera, puesto que ambas pertenecen a nive-les epistemológicos diferentes, y pueden complementarse.
Por consiguiente, también parece claro que, en virtud de esta distin-ción de planos epistemológicos, no resulta sostenible la postura del re-duccionismo materialista, o naturalismo cientifista, por insuficiente y contradictoria:
a) por insuficiente, puesto que las ciencias no lo abarcan todo, sino el nivel de las descripciones fácticas; más allá tienen que ceder el puesto a la filosofía y a la teología; el materialismo reduccionista

99
© Universidad de Deusto - ISBN 978-84-9830-383-4
pretende ser con frecuencia sólo ciencia, pero es más bien, y con toda legitimidad, una propuesta filosófica;
b) y por contradictoria: afirmar que la ciencia es suficiente para dar cuenta de la realidad, no pertenece al ámbito científico sino al fi-losófico; por lo tanto, el reduccionismo materialista, que pretende ser una teoría científica, constituye de hecho una tesis filosófica, que además consideramos falsa. Esto es, se está intentando pasar como afirmaciones y conclusiones científicas lo que no pasan de ser propuestas filosóficas, que, por definición, no son demostra-bles empíricamente, y de ahí que puedan presentarse y ser acepta-das varias teorías rivales al mismo tiempo, con la misma preten-sión de verdad.
4.3. La problemática relación entre ciencia y religión. Cuatro posturas diferentes
La relación entre ciencia y teología, fe o religión, ha sido siempre com-pleja. Cuando no se sabe bien esta historia, se proyectan sobre ella nuestros prejuicios e intereses, pensando que las cosas han sido en el pasado igual que en el presente, y no es del todo cierto. Por tanto, hay que desmontar muchos prejuicios sobre la forma en que a veces se entiende y se afirma que han sido las relaciones en el pasado entre ciencia y fe. Un prejuicio primero y fundamental, que quienes investigan la historia de esta relación señalan como bastante frecuente, es la incorrecta teoría de que la ciencia ha estado siempre en oposición a la religión, como si el trabajo científico fuera totalmente incompatible con los planteamientos religiosos.
Nada más lejos de la realidad, porque el saber ha estado siempre interrelacionado desde el comienzo de la historia humana, formando un todo desde el que se han ido desgajando las diferentes parcelas y especia-lidades, y al inicio no había conflictos entre las diferentes miradas sobre la realidad, sea la mirada científica, literaria, histórica, filosófica, teológi-ca o mística. Además, es una verdad incontestable que el saber filosófico y científico han nacido al amparo de la religión; aunque no sin conflicto. De hecho, el judeocristianismo, con su esencial y radical sesgo desmitifi-cador y secularizador de la realidad (en el relato del Génesis es esto lo que nos viene a decir: Dios es trascendente al mundo, y el mundo tiene sus le-yes autosuficientes; por tanto, el mundo no es Dios; el hombre no tiene que adorarlo, sino que puede conocerlo, darle nombre y dominarlo, aun-que no de una forma despótica) ha contribuido de forma determinante en los orígenes y en los avances de la ciencia moderna. No en vano la ciencia moderna se ha desarrollado en el entorno de la Europa cristiana, aunque

100
© Universidad de Deusto - ISBN 978-84-9830-383-4
también en el contexto de la sociedad islámica. Y la mayoría de los gran-des científicos modernos han sido cristianos, e incluso en la mayoría de los casos también muy religiosos. Bastaría hacer un recorrido por la histo-ria de la ciencia para comprobarlo29.
Bien es cierto que la mirada científica y la mirada religiosa tienen ele-mentos específicos y más que suficientes como para entrar en conflicto en determinadas ocasiones. Pero esta conflictividad se ha dado más que por la incompatibilidad de ambos enfoques, por las insuficiencias de los propios intelectuales a la hora de hacer ciencia o interpretar la religión. El científico se centra en describir los fenómenos naturales tal y como aparecen a nuestra observación directa, pero a veces se tiende, por parte de algunos, a absolu-tizar este punto de vista y a considerar que todo lo que hay se reduce a esa única dimensión. En cambio, la teología o la fe se centran en el nivel del sentido, aunque también hay ciertos planteamientos que tienden a no acep-tar del todo la autonomía de lo mundano, y, en consecuencia, a defender que la acción de Dios sobre el mundo se confunde y se mezcla con los factores materiales que originan los diversos acontecimientos naturales.
Por eso que, más que hablar de conflicto entre ciencia y fe, hay que hablar de conflicto entre determinados científicos y determinados cre-yentes o teólogos. Pero purificados de estos errores y derivas incorrectas, creemos que lo más acertado es defender que son perfectamente compati-bles la visión científica y la religiosa, entendidas ambas de forma adecua-da, aunque eso no quita que se mantengan de vez en cuando, o incluso en no pocos momentos, diversas dificultades a la hora de conjugarlas en rela-ción a diversos temas puntuales.
Los diversos modos como se suelen interrelacionar y conjugar la cien-cia y fe pueden resumirse en cuatro posturas30:
a) Conflicto
Para esta postura, no hay posibilidad de conjugar fe y ciencia, puesto que se trataría de dos dimensiones contrapuestas de mirar la realidad. O se es científico o se es creyente, pero no se puede ser las dos cosas al mismo
29 Cfr. Sequeiros, L., o.c., p. 182; Fernández Rañada, A., o.c., p. 288; Riaza, J. M., LaIglesia en la historia de la ciencia, Madrid, BAC, 1999; Nicolaus Pous, Fr., Iglesia y cien-cia a lo largo de la historia, Barcelona, Ed. Claret, 2002; Udías Vallina, A., Ciencia y reli-gión. Dos visiones del mundo, Santander, Sal Terrae, 2010.
30 Cfr. Barbour, Ian G., textos citados en la nota 28; 2004; Polkibghorne, John, Cienciay Teología Una introducción, Santander, Sal Terrae, 2000; Id. (ed.), La obra del amor. La creación como kénosis, Estella (Navarra), Verbo Divino, 2008.

101
© Universidad de Deusto - ISBN 978-84-9830-383-4
tiempo. El error básico de esta postura es no aceptar el pluralismo episte-mológico, esto es, la complejidad de la realidad, sobre la que cabe hacer constataciones de hecho, ciencia, y preguntas por el sentido, filosofía y teología. Por tanto, como sólo se puede ver la realidad, según los defenso-res de estos planteamientos, desde un único punto de vista, se entiende y se vive la ciencia y la fe como dos teorías sobre la realidad que se sitúan en el mismo y único nivel, y por tanto se hallan en conflicto y resultan ex-cluyentes.
No es extraño, por ello, constatar que dentro de esta postura se si-túan tanto científicos como creyentes, enfrentándose los planteamien-tos de los reduccionistas científicos y de los creyentes fundamentalistas. Para los primeros, no hay más forma de dar cuenta de la realidad que a través del método científico, entendido bajo el método positivista. Entre los defensores de estos planteamientos están sobre todo intelec-tuales como R. Dawkins y D. Dennet31. En cambio, para los fundamen-talistas cristianos, la verdad sobre el conjunto del universo se halla en la Biblia, entendida al pie de la letra. De modo que, cuando hay una discre-pancia entre los científicos y el libro sagrado, no hay duda de quién tiene la verdad y quién se equivoca. Aquí se sitúan los fundamentalistas clá-sicos de ámbito norteamericano, sobre todo, y en parte también los de-fensores de la teoría denominada del Diseño Inteligente32.
El error del cientifismo naturalista consiste en convertirse y actuar como una religión, puesto que tiene la pretensión de que una teoría cien-tífica es también una cosmovisión, una propuesta de sentido33. El funda-mentalismo religioso absolutiza también la mirada religiosa, atribuyéndo-le la tarea de dar cuenta de la realidad material, cosa que le corresponde a la ciencia. De ahí que ambos planteamientos no puedan por menos que chocar, porque pretenden ocupar todo el espectro del saber, y se miden como dos propuestas excluyentes y alternativas. La ciencia quiere conver-
31 Cfr. Dawkins, R., El espejismo de Dios, Madrid, Espasa, 2007; Id., El capellán del diablo, Barcelona, Gedisa, 2005; Dennet, D., Breaking the Spell. Religión as a natural Phenomenon, Londres, Viking, 2006 (trad. española: Romper el hechizo. La religión como fenómeno natural, Buenos Aires, Katz Editores, 2006).
32 Cfr. los textos sobre los fundamentalismos clásicos. Cfr. Ayala, Fco. J., Darwin y el Diseño Inteligente, o.c., cap. 8.º, «Creacionismo y fundamentalismo en Estados Unidos», pp. 163 y ss. Sobre la teoría del Diseño Inteligente, cfr. Johnson, Ph. E., Juicio a Darwin,Madrid, Homo Legens, 2007; Dembski, W. A., Diseño Inteligente. Respuestas a las cues-tiones más espinosas del diseño inteligente, Madrid, Homo Legens, 2006; Behe, M., Darwin’s Black Box: The Biochemical Challenge to Evolution, Nueva York, The Free Press, 1996.
33 Cfr. McGrath, Alister, «Los ateísmos de superventas: el nuevo cientifismo», Conci-lium, 2010, n.º 337, 555-567.

102
© Universidad de Deusto - ISBN 978-84-9830-383-4
tirse en una religión más, y la religión quiere ocuparse de ámbitos del sa-ber que no le corresponde.
El enfrentamiento de estas dos posturas extremas suele ser lo que sal-ta a la prensa y lo que suele llegar al gran público, con el correspondiente desenfoque teórico que puede advertirse, con lo cual se hace un flaco fa-vor a una reflexión seria de este problema fundamental. Advertimos con ello un dicho popular: los extremos se tocan y se alimentan mutuamente. Al cientifismo naturalista le conviene tener delante a un fundamentalismo primitivo y extremo, porque con él se siente a gusto para vencerlo fácil-mente; y lo mismo le ocurre al fundamentalismo, cuando se encuentra con una postura tan extrema y radical como la suya, y tan poco dada a precisar y a matizar, sino a salir a la búsqueda del hereje y a llevarlo a la hoguera.
b) Independencia
Los que se sitúan en esta postura consideran que no hay, ni puede ha-ber, conflicto entre el punto de vista de la ciencia y el de la fe, porque cada uno de los dos se mueve en niveles de reflexión diferentes. La cien-cia se centra en decirnos cómo es la realidad material, y la religión, en cómo debemos vivir, proponiéndonos diversas orientaciones de sentido y salvación34.
El acierto de esta postura está en distinguir estos dos planos epistemo-lógicos sobre los que se mueven la ciencia y la fe, o la teología. Pero su insuficiencia consiste en considerar que la separación de planos hace que parezca que ciencia y fe no tienen nada que decirse ni dialogar. De ese modo, esta postura se apoya en un fondo teórico dualista, en el que por un lado va la ciencia y los aspectos empíricos, y, por otra, la filosofía y la re-ligión, condenando al ser humano a vivir un mundo escindido en dos par-tes que no se relacionan ni tienen nada que decirse, cosa que es manifies-tamente inexacta.
De ese modo, dentro ya de la teología, resulta difícil conjugar la rea-lidad fáctica de lo mundano con su sentido y significado, abriéndose un abismo insalvable entre el Dios de la creación y el Dios de la salvación.
En conclusión, la ventaja de evitar los conflictos se salda con el in-conveniente y el error de escindir de forma definitiva nuestra realidad mundana, que, por el contrario, se nos muestra como una realidad única aunque compleja.
34 En esta postura se sitúa Stephen Jay Gould, Ciencia versus religión. Un falso con-flicto, Barcelona, Crítica, 2000.

103
© Universidad de Deusto - ISBN 978-84-9830-383-4
c) Diálogo
Esta postura trata de comparar los métodos que emplean tanto la ciencia como la teología, advirtiendo sus semejanzas y sus diferencias entre ellos. En concreto, estudian los modelos y analogías que utilizan la ciencia y la teología para tratar de comprender e imaginar lo que no puede ser directamente observado (por ej., la estructura y los componen-tes del átomo, por los científicos, o la existencia y realidad de Dios, por la teología).
Otro punto de diálogo se centra en advertir los diferentes momentos en los que la ciencia llega en sus investigaciones a cuestiones límite, en los que advierte que ya no puede responder con sólo el enfoque científico (Ej.: la creación del mundo, o el hecho de que el mundo se nos aparece como un todo ordenado e inteligible, ante lo cual surgen las cuestiones post-científicas de por qué es así y no de otra forma).
También constituyen ocasiones de diálogo al utilizar algunos concep-tos científicos como analogías para hablar de cómo se puede entender la relación de Dios con el mundo. Por ejemplo, la comunicación de infor-mación, o entender a Dios como el determinador de las indeterminacio-nes cuánticas que deja abierta la física cuántica, proceso que atribuir a la acción de Dios no supondría violar ninguna ley de la física actual.
Podríamos seguir poniendo múltiples ejemplos, pero siempre llegaría-mos a la conclusión de que, aunque este modo de proceder es muy fructí-fero y necesario, en la medida en que advertimos un importante y clarifica-dor acercamiento entre ambos enfoques, vemos también su insuficiencia, pudiéndose profundizar más en el acercamiento entre ciencia y fe, como propone la postura siguiente.
d) Integración
Esta postura va más allá de las dos anteriores, y trata no sólo de ad-vertir la diferencia de niveles epistemológicos en los que se mueven la ciencia y la teología, o establecer diálogos entre las semejanzas estructu-rales entre los dos métodos, el científico y el teológico, sino de llegar a in-tegrar más estrechamente ambas disciplinas.
Esta integración, desde el punto de vista de la teología, se puede hacer de diferentes formas, siguiendo a I. G. Barbour:
— La que realiza la denominada teología natural: trata de buscar en la naturaleza y en los avances de la ciencia pruebas para demostrar la existencia de Dios y las afirmaciones de la teología.

104
© Universidad de Deusto - ISBN 978-84-9830-383-4
— La defendida por la teología de la naturaleza: no busca pruebas demostrativas de las verdades que defiende la fe y la teología, sino simplemente mostrar que los avances de la ciencia no son contra-rios a la teología, sino más bien lo contrario: le sirven de apoyo y hacen razonables sus afirmaciones.
La primera postura trata de reavivar los planteamientos de la ya supe-rada (aunque no por todos) teología natural, existente ya desde la Edad Media, y consistente en defender que las verdades teológicas, entre las que estaría la existencia de Dios, se pueden demostrar, basándose también en la teoría de las dos vías de la única verdad.
En cambio, lo que Barbour denomina teología de la naturaleza con-sidera que las verdades de la fe no pueden ser demostradas, pero sí son razonables, considerándose una opción más al lado de otras opciones filo-sóficas y cosmovisionales de entender la realidad. De este modo, sostiene que la fe (la teología), la filosofía y la ciencia puede conjugarse de modo adecuado para sostener que se puede ser creyente y admitir los datos con-solidados de la ciencia, en este caso la teoría de la selección natural pro-puesta por Darwin.
De cara al tema que nos ocupa, las dos posturas que vamos a tener en cuenta, y que suelen ser las que salen más al gran público y ayudan a pre-sentar de un modo más claro el problema, son la primera y la tercera, en este caso en la versión de lo que hemos denominado teología de la natu-raleza.
4.4. Las críticas de los creacionismos fundamentalistas35
Dentro de los diferentes grupos e iglesias cristianas, siempre se han dado grupos numerosos que se han opuesto a la evolución y a la selección darviniana, desde el convencimiento de que la única verdad sobre el hom-bre y sobre el mundo no puede provenir más que de Dios y de su palabra,
35 Cfr. Scott, E. C., Evolution vs. Creationism. An Introduction, Westport, CT, GreenwoodPress, 2004; Numbers, R. L., Darwinism comes to America, Cambridge, MA, Harvard Uni-versity Press, 1998; Lindberg, D. L./Numbers, R. L. (eds.), God and Nature. Historical Essays on the Encounter between Christianity and Science, Berkeley, CA, University of California Press, 1986; Davies, M. W., Darwin y el fundamentalismo, Barcelona, Gedisa, 2004; Ayala, F. J., Darwin y el Diseño Inteligente. Creacionismo, cristianismo y evolución,Madrid, Alianza, 2007, cap. 8, «Creacionismo y fundamentalismo en Estados Unidos», pp. 163 y ss.; Ruse, M., ¿Puede un darwinista ser cristiano? La relación entre Ciencia y Religón, Madrid, Siglo XXI, 2004; Sequeiros, Leandro, ¿Puede un cristiano ser evolu-cionista?, Madrid, PPC, 2009.

105
© Universidad de Deusto - ISBN 978-84-9830-383-4
la Biblia. En la medida en que esa palabra tiene que ser definitiva e infali-ble, la Biblia como palabra de Dios tendría que ser entendida y manejada de forma literal y directa. A los defensores de estos planteamientos se les ha denominado creacionistas, y creacionismo a su doctrina, situándose es-tos grupos en la actualidad sobre todo en el ámbito norteamericano.
En lo que resta de este capítulo nos vamos a detener en presentar, de forma sintética, los planteamientos del creacionismo fundamentalista,para en el apartado siguiente dedicarnos a la versión más seria, y con pre-tensiones científicas, de esta tendencia creacionista, denominada teoría del Diseño Inteligente.
Pero antes es importante hacer una observación sobre el nombre crea-cionismo, en la medida en que este apelativo puede referirse simplemente a la fe en Dios como creador del mundo, pero también a la teoría funda-mentalista defensora de una interpretación literal del texto bíblico. El pri-mer creacionismo, que podemos denominarlo creacionismo ilustrado, es la postura de la mayoría de los creyentes que intentan compatibilizar la fe en la creación del mundo por Dios con una visión autónoma del mundo, objeto de la investigación científica, no viendo oposición con lo que afir-ma la teoría evolucionista. En cambio, el creacionismo fundamentalista es anti-evolucionista, y entiende la creación del mundo y la acción de Dios sobre él de forma milagrera, negando la autonomía de lo mundano respec-to a la acción de Dios en el discurrir de su historia, sometida a sus diferen-tes leyes internas. Aquí nos vamos a detener en este creacionismo funda-mentalista, especialmente virulento y aceptado en el entorno de diferentes grupos evangélicos norteamericanos. De ahí que algunos estudiosos le de-nominen fundamentalismo protestante36.
Dentro de este amplio y heterogéneo movimiento, se advierten dife-rentes formas o escuelas: el creacionismo de la Tierra Reciente, el crea-cionismo de la Tierra Plana, el Geocentrismo, y el creacionismo de la Tierra Antigua37. A todos ellos hay que añadir, como ya dijimos más arri-ba, la versión más reciente y modernizada del creacionismo del DiseñoInteligente. Veamos los aspectos más importantes de cada una de estas versiones del creacionismo fundamentalista.
El creacionismo de la Tierra Reciente es el que interpreta de modo más literal todos los relatos del Génesis referentes a la creación de las dis-tintas partes del universo y de los seres vivos en la Tierra. Basándose en una interpretación literal del proceso bíblico de la creación, defienden que el mundo tiene una antigüedad de 6.000 años (se originó 4.000 años antes
36 Cfr. Küng, Hans, El principio de todas las cosas. Ciencia y religión, Madrid, Trotta, 2007, pp. 99 y ss.
37 Cfr. Ayala, F. J., Darwin y el Diseño Inteligente, o.c., pp. 166-167.

106
© Universidad de Deusto - ISBN 978-84-9830-383-4
de la era cristiana), cálculo que se saca de sumar las edades de los patriar-cas y el conjunto de la genealogía presentada en los evangelios de Mateo (1,1-17) y Lucas (3,23-38). Su interpretación literal de la Biblia les lleva también a defender la realidad histórica del diluvio universal. Este movi-miento no está formado por gente inculta y sin prestigio social. Prueba de ello es que «el Institute for Creation Research en San Diego, California, es un centro notable del creacionismo de la Tierra Reciente. Posee un mu-seo, publicaciones y actividades académicas»38.
El creacionismo de la Tierra Plana es una versión del grupo anterior, y, como su nombre indica, sostiene que la Tierra es plana y tiene la for-ma de una moneda con bordes redondeados, y no como una esfera, tal y como es evidente por las muestras científicas. Tampoco este movimiento es despreciable por su composición e influencia, puesto que tiene unos 3.500 miembros y están organizados en la International Flat Earth Researche Society of Lancaster39.
El creacionismo denominado Geocentrismo acepta la forma esféri-ca de la Tierra, pero sigue defendiendo que constituye el centro del sis-tema solar, en vez de reducirse a ser un simple planeta del sistema solar. Y una prueba de que su peso cultural es importante lo constituye el he-cho de que «una sesión plenaria en la conferencia de 1985 de la Bible-Science Association consistió en un debate entre dos geocentristas y dos heliocentristas»40.
El creacionismo de la Tierra Antigua acepta los datos y pruebas de las diferentes ciencias sobre la edad de la Tierra, y reconoce su antigüedad, pero hay diferentes versiones de este tipo de creacionismo, y cada uno de ellos pone diversos matices a la aceptación de los datos científicos. Según el de-nominado Creacionismo Día-Era, en su empeño por seguir apostando por la literalidad de la interpretación bíblica, defiende que los días a los que se re-fiere la Biblia hay que interpretarlos no tanto como días solares, sino como largos períodos de tiempo o eras. Otros creacionistas de esta corriente sostie-nen «la creencia en sucesivas intervenciones de Dios, quien creó el mundo así como todas las criaturas vivas de forma separada, por etapas»41.
Como indica L. Sequeiros42, este amplio movimiento fundamentalista ha seguido una estrategia anti-evolucionista a lo largo de un amplio perío-
38 Ibídem, p. 166. 39 Cfr. ibídem, p. 166. 40 Cfr. Scott, E. C., o.c., p. 58. Tomo el dato de Ayala, F. J., o.c., p. 166. 41 Ayala, F. J., o.c., pp. 167. 42 Cfr. Sequeiros, L., «Dioses, monos y humanos: La respuesta de la Ciencia y de la
Biblia a los orígenes de la humanidad», en VV.AA., Biblia y Cultura, Bilbao, Universidad de Deusto, 2008, pp. 97-115. Cfr. también Id., ¿Puede un cristiano ser evolucionista?, o.c., pp. 93 y ss.

107
© Universidad de Deusto - ISBN 978-84-9830-383-4
do de tiempo que se puede dividir en tres etapas: en la primera, entre 1910 y 1940, «los antievolucionistas intentaron y lograron en algunos casos que se aprobara una legislación que eliminase la evolución en las escuelas y en los libros de texto»43. Pero estos triunfos parciales no duraron mucho, y al ser derogadas las leyes que habían conseguido en algunos estados, se inicia el segundo período, entre los años de 1960 hasta casi la actuali-dad, basado en una versión más científica del creacionismo. La actualidad creacionista está marcada, en un tercer estadio, con la aparición de la teo-ría denominada Diseño Inteligente44.
La primera etapa del creacionismo fundamentalista estuvo centra-da en contrarrestar la progresiva introducción, en los programas escola-res de la secundaria, de la enseñanza de las teorías evolucionistas, unido también al hecho de asociar a las tesis científicas del evolucionismo las tesis políticas e ideológicas del darwinismo social. Estos factores lleva-ron a los fundamentalistas a impulsar la aprobación de leyes en diversos Estados para proteger a los jóvenes de las para ellos peligrosas teorías evolucionistas. En la disputa entre la verdad bíblica y la científica, tra-taron de demostrar que las afirmaciones bíblicas tenían más consistencia que las científicas. Así es como aparece ya en 1909 una nueva versión de la Biblia, llevada a cabo por Cyrus I. Scofield (1843-1847), y en la que defendía la existencia de un importante intervalo de tiempo entre la creación del universo, según el primer versículo del Génesis, y el res-to de la creación, narrado a partir del versículo segundo, tesis que ya había defendido anteriormente Thomas Chalmers (1780-1847). Algunos años más tarde, G. McCready Price (1870-1963), perteneciente a los Adven-tistas del Séptimo Día, comenzó a defender la historicidad del diluvio universal, fenómeno que, según él, explicaría la formación de las rocas y de los fósiles descubiertos y estudiados por los paleontólogos. De este modo, atribuía intencionalidad científica a los relatos bíblicos y negaba las tesis evolucionistas.
La movilización social y política de los grupos fundamentalistas con-siguió que para antes de 1925 se hubieran aprobado en treinta y siete Esta-dos norteamericanos diversas leyes para prohibir la enseñanza de las tesis evolucionistas en las escuelas públicas, penalizando y castigando a los pro-fesores que las incumplieran. Se hizo mundialmente famoso el juicio con-tra el profesor de secundaria de Tenesse, J. Th. Scopes (1900-1970), quienen 1925 fue condenado por enseñar el evolucionismo en sus clases. Du-rante esos años triunfó y se extendió notablemente la tendencia anti-evo-
43 Sequeiros, L., «Dioses, monos y humanos», o.c., p. 105. 44 Sigo en la descripción de las dos etapas primeras del fundamentalismo a L. Sequei-
ros, ¿Puede un cristiano ser evolucionista?, o.c., pp. 98 y ss.

108
© Universidad de Deusto - ISBN 978-84-9830-383-4
lucionista, llegándose a fundar en 1935 la Religión and Science Associa-tion, cuyo presidente fue L. Allen Highley, profesor de la Universidad de Chicago. Las desavenencias entre sus socios hizo que algunos de ellos se escindieran para fundar otra asociación, liderados por Price y otros Ad-ventistas, asociación que muy pronto sufrió también escisiones, que oca-sionó su desaparecición en 1948.
Al mismo tiempo, en 1941 fue fundada por un grupo de evangélicos The american scientific affiliation, constituida por científicos cristianos y encaminada a extender la necesidad de que las verdades bíblicas y cientí-ficas concordaran, en vez de oponerse. La condición para ello estaba en que las tesis científicas tenían que someterse a las verdades de la Biblia, leída de modo literal, ya que la Biblia por principio no podía equivocarse. En este movimiento la figura clave fue el geoquímico J. L. Kulp, conven-cido de la necesidad de no mezclar la geología con la evolución de cara a la concordancia entre ciencia y Biblia.
La segunda etapa del anti-evolucionismo se inicia con los años 60, momento en el que los Estados Unidos están volcados en un ambicioso programa científico de exploración del espacio, para responder al reto so-viético. Ese programa incluía la ampliación de la enseñanza de las mate-rias científicas desde la secundaria, entre cuyos contenidos estaba la teoría evolucionista, como se indica en una ley publicada en 1963. Como puede imaginarse, esta tendencia crea un gran malestar entre los ámbitos conser-vadores, a pesar de que en la enseñanza se distinguía claramente entre el hecho de la evolución y las teorías filosóficas que la interpretaban. Pero las familias conservadoras entendían que ese era un camino peligroso para socavar la fe religiosa de sus hijos.
En estos años, a diferencia de la fase anterior, la estrategia de los fun-damentalistas irá en la línea de presentar razones científicas para oponerse al evolucionismo. Sus argumentos intentaban mostrar que la tesis bíblica era una teoría tan científica como la teoría evolucionista, y que obligar a enseñarse ésta en las escuelas iba en contra de la libertad religiosa, en la medida en que sería obligar a estudiar una teoría que no está probada en absoluto. En realidad, la estrategia que se siguió comprendía tanto la pre-tendida categoría científica de las teorías bíblicas como también el negar la firmeza científica de las tesis evolucionistas, en la medida en que el evolucionismo sólo sería una simple hipótesis de trabajo, muy lejos de es-tar demostrada por los parámetros científicos.
Una figura clave de este momento es H. M. Morris (1918-2006), in-geniero hidráulico, considerado como el padre del creacionismo cientí-fico, defensor de estos planteamientos desde muy joven, a través de sus tempranos libros That you might believe (Lo que usted debe creer, 1946) y The Bible and modern science (La Biblia y la ciencia moderna, 1951),

109
© Universidad de Deusto - ISBN 978-84-9830-383-4
donde «defendía la realidad histórica del hecho de la creación de la nada por Dios, e intentaba demostrar científicamente la existencia histórica del diluvio universal»45. Y para que sus afirmaciones tuvieran peso, se esforzaba en aportar pruebas científicas. Pero el creacionismo científi-co cobró más peso con las aportaciones del profesor de Antiguo Testa-mento, John C. Whitcomb (n. 1924), quien publicó con Morris un libro que tuvo una gran influencia entre los creacionistas, The Genesis flood(El Diluvio del Génesis). Whitcomb creó en Michigan un centro de in-vestigación, The Creation Research Society (CRS, Sociedad para la in-vestigación de la creación), exponiendo sus tesis más rígidas sobre este punto a través de la revista Creation research Society Quaterly (CRSQ, Revista de la Sociedad para la investigación de la creación), publicada a partir de 1964.
La estrategia de los creacionistas fundamentalistas se orientó a conse-guir mayorías políticas que aprobaran leyes contra la enseñanza del evolu-cionismo en las escuelas. Lo consiguieron en el Estado de Arkansas. Pero en 1968 el Tribunal Supremo de los Estados Unidos publicó una ley por la que consideraba contraria a la Constitución cualquier ley que prohibie-ra la enseñanza de la evolución en las escuelas públicas. Los creacionistas fundamentalistas siguieron batallando en el terreno de las ideas, publicán-dose en 1970 por la citada The creation research Society, el libro Biology:a search for order in complexity (La Biología: un búsqueda de un orden en la complejidad), donde se advierte ya en el título la orientación de pre-tender ver en el ámbito de la lógica de la vida la presencia de un diseño, pudiéndose así demostrar en él la acción de Dios sobre el origen y el de-sarrollo de la biosfera.
Morris seguía empeñado en defender la historicidad del diluvio universal, y publicó en 1974 un nuevo libro sobre el tema, con el títu-lo genérico Sientific creationism (Creacionismo científico). Así mismo, Morris siguió peleando para que las tesis creacionistas fueran intro-ducidas en los programas escolares de estudio en igualdad de trato con el evolucionismo, consiguiendo durante la etapa de la presidencia de Reagan que se aprobasen en varios Estados, de tal forma que, al ini-cio de los años 80, se había conseguido este objetivo nada menos que en veintisiete de ellos.
Pero la movilización de científicos evolucionistas llevó a la derrota de estos planteamientos, en sendos juicios públicos, en los estados de Arkansas(el caso denominado McLean vs. Arkansas) y en Louisisna (caso Edwards vs. Aguillard). Los argumentos jurídicos se apoyaban en el hecho de que
45 Ibídem, p. 103.

110
© Universidad de Deusto - ISBN 978-84-9830-383-4
situar las teorías creacionistas al mismo nivel que el evolucionismo supo-nía confundir una teoría científica con otra religiosa, siendo contrario a la libertad de conciencia y de religión enseñar en las escuelas estatales una creencia religiosa concreta46.
Aunque las teorías del creacionismo fundamentalista no están exclu-sivamente presentes en los Estados Unidos sino que también se han de-fendido en Europa, los representantes europeos no han conseguido el peso social ni tampoco representantes intelectuales de peso. Eso hace que sus seguidores se limiten a traducir y repetir lo que escriben los representan-tes estadounidenses47.
4.5. La teoría del Diseño Inteligente (DI)
En el juicio de Luisiana de 1987, en el que el Tribunal Supremo de Estados Unidos suprimió la ley del Estado sureño sobre la igualdad de trato entre el creacionismo y el evolucionismo en las escuelas públicas, empiezan a aparecer y a concretarse los argumentos que serán utilizados por la corriente más renovada del creacionismo fundamentalista: el Di-seño Inteligente (DI). La disputa legal no se terminó con la resolución sobre Louisiana, sino que se continuó con otro nuevo capítulo en Dover (Pennsylvania), en 2004. La Junta Directiva Escolar de la Región de Do-ver, adoptó en octubre de 2004 la siguiente resolución: «A los estudiantes se les pondrá al corriente de los vacíos/problemas de la teoría de Darwin y de que hay otras teorías de la evolución, entre ellos el diseño inteligente, pero sin limitarse a él»48.
A raíz de ello, se inició una fuerte confrontación ante los tribunales de los Estados Unidos, en la medida en que el Distrito Escolar de la Re-gión de Dover reforzó los planteamientos de la Junta Directiva Escolar de la Región, obligando a todos sus profesores en sus aulas a la lectura de un declaración con afirmaciones como éstas: «Debido a que la Teoría de Darwin es una teoría, sigue siendo puesta a prueba mientras se descubren nuevas evidencias. Una Teoría no es un hecho… El Diseño Inteligente
46 Cfr. ibídem, 102-106; Ayala, F. J., Darwin y el Diseño Inteligegnte, o.c., pp. 168 y ss.
47 Cfr. Sequeiros, L, o.c., pp. 106-107. Sobre la incidencia de estas corrientes en Es-paña, cfr. Molina, E., «Los argumentos geológicos y paleontológicos de los creacionistas “científicos”: ignorancia y pseudociencia», en Molina, E./Carreras, A./Puertas, J. (eds.), Evolucionismo y racionalismo, Zaragoza, Universidad de Zaragoza, 1998; Gomis, A./Josa, J.,«Imágenes de la polémica darwinista en España. Creacionismo y evolucionismo», MundoCientífico, 2002, n.º 233, pp. 20-29.
48 Cfr. Ayala, F. J., o.c., p. 169.

111
© Universidad de Deusto - ISBN 978-84-9830-383-4
es una explicación del origen de la vida que difiere del punto de vista de Darwin. El libro de referencia Of Pandas and People (La gente y los pan-das), está al alcance de los estudiantes que puedan estar interesados en comprender lo que realmente implica el Diseño Inteligente»49.
Tras recurrirse esta orden ante la Corte Federal del Distrito Central de Pensylvania por once padres, se celebró el juicio en otoño de 2005, haciéndose famoso el veredicto de 139 páginas emitido por el juez fe-deral John E. Jones III, en el que falla contra las tesis creacionistas con los siguientes argumentos: «La Política de los defensores del DI viola la Clásula que establece el Estado Laico en la Primera Enmienda de la Constitución de los Estados Unidos», ordenándose también que a los «Defensores se les impone de forma permanente que no mantengan la Política del DI en ninguna escuela dentro del Distrito Escolar de la Re-gión de Dover»50.
En el juicio se pidieron las opiniones de expertos de primera línea y sirvieron para que el juez Jones III presentara una sentencia en la que hace interesantes y acertadas afirmaciones sobre la naturaleza de las teo-rías creacionistas (teoría filosófico-teológica, pero no ciencia) y sobre la relaciones correctas entre la ciencia y las interpretaciones filosóficas y teológicas sobre ella. De ahí que afirme en su sentencia: «La abrumadora evidencia aportada en el juicio estableció que el DI es una opinión reli-giosa, tan sólo una nueva etiqueta del creacionismo, y no una teoría cien-tífica» (p. 43). «El DI no está apoyado por ninguna investigación, datos o publicaciones científicamente contrastables» (p. 87). Todo esto hace con-cluir que «presenta a los estudiantes una alternativa religiosa disfrazada de teoría científica, (…) y enseña a los estudiantes a renunciar a la inves-tigación científica» (p. 49)51.
Es posible que los intentos de los teóricos del DI por ser homologada su teoría como una hipótesis científica, sigan adelante recurriendo estas sentencias judiciales. Lo que sí está claro es que las publicaciones que de-fienden estos planteamientos siguen apareciendo y extendiéndose tanto en los Estados Unidos como en Europa, y en el resto del mundo, tratando de ir ganando adeptos.
Veamos a continuación los autores más significativos de esta corrien-te, sus obras e ideas más significativas, así como un análisis crítico sobre su incorrecta pretensión de considerarse una teoría científica, así como una valoración sobre su desacertada teología de fondo52.
49 Texto tomado de Ayala, F. J., o.c., pp. 169-170. 50 Texto tomado de Id., p. 171. 51 Textos tomados de Id., p. 170. 52 Cfr. Ayala, F. J., Darwin y el Diseño Inteligente, o.c., cap. 7.º, pp. 143 y ss.

112
© Universidad de Deusto - ISBN 978-84-9830-383-4
a) Los argumentos y las debilidades del Diseño Inteligente53
Como ya quedó indicado, la teoría del Diseño Inteligente (DI) cons-tituye la tercera etapa del creacionismo fundamentalista, aunque algunos de sus autores más significativos intentan desmarcarse hasta cierto pun-to de este movimiento tan conservador. Se trata del empeño por situarse frente a las tesis del evolucionismo, pero no tanto desde posturas explí-citamente teológicas, sino presentando argumentos que pretenden con-siderar su postura como una teoría científica más, rival de la selección natural de Darwin.
Los líderes ideológicos del DI son sobre todo tres: Phillip Johnson, Michael Behe y William Demski. Ph. Johnson se hizo conocido por su libro Juicio a Darwin54, en el que toda su argumentación se centra en re-saltar los vacíos explicativos del darwinismo así como sus supuestas con-tradicciones. Johnson no es científico de profesión, sino abogado, y por eso, como él mismo lo indica en su libro, sus argumentaciones van en la línea de descubrir las supuestas flaquezas del darwinismo, para dejar bien sentado que no se trata de una teoría científica comprobada y demostra-da, sino una simple hipótesis de trabajo que se halla muy lejos de resultar convincente.
Michael Behe es un bioquímico que sitúa sus argumentaciones en el terreno de las pruebas científicas, centrando toda su argumentación, en su libro básico Darwin’s Black Box: The Biochemical Challange to Evo-lution (La caja negra de Darwin. El reto bioquímico a la evolución)55,en mostrar que el mundo de los seres vivos está lleno de mecanismos tan complejamente construidos (complejidad irreductible), que se ne-cesita necesariamente echar mano de un diseñador para explicarlos, siendo insuficiente la solución de la selección natural que propone el darwinismo.
William Dembski completa el argumento de Behe desde plantea-mientos matemáticos y estadísticos, defendiendo que esos mecanismos complejos que constituyen los diversos órganos de los seres vivos, así como el origen de las diversas especies, son de tan baja probabilidad (complejidad específica), que necesariamente tenemos que echar mano de un diseñador consciente e inteligente para atribuirle la autoría de ta-
53 Cfr. Johnson, Ph. E., Juicio a Darwin, Madrid, Homo Legens, 2007; Dembski, W. A., Diseño inteligente. Repuestas a las cuestiones más espinosas del diseño inteligente, Ma-drid, Homo Legens, 2006.
54 O.c. Cfr. Id., The Wedge of Truth, Downers Grove, II: Inter Varsity Press, 2000. 55 Nueva York, The Free Press, 1996. Trad. cast.: La caja negra de Darwin. El reto de
la bioquímica a la evolución, Madrid, Edit. Andrés Bello, 2000.

113
© Universidad de Deusto - ISBN 978-84-9830-383-4
les hechos y realidades. Tales son los argumentos básicos de su libro The Desing Inference56.
Los argumentos centrales de los defensores del DI son, si nos atenemos a lo fundamental, dos. Por un lado, «que los organismos son tan complejos que ningún proceso natural podría explicar su origen: más bien los orga-nismos y sus partes evidencian que han sido diseñados para cumplir ciertas funciones. Sostienen que la selección natural no puede explicar el diseño de los mamíferos, o los flagelos de las bacterias»57. Y, en segundo lugar, que la teoría de la selección natural no es una teoría científica firmemente asentada y comprobada, sino que se limita a ser una simple teoría, esto es, una hipó-tesis entre otras, ante la cual se pueden presentar legítimamente otras hi-pótesis alternativas, una de las cuales es la del DI que ellos defienden.
Los defensores del DI presentan su teoría como un paradigma cien-tífico similar a cualquier otro, y, por ello, defienden la legitimidad de competir con otros paradigmas científicos en vigor, como el evolucionis-mo darwinista. En ese sentido, su interés se centra en desmarcarse de los diversos creacionismos fundamentalistas58, y, por otro lado, de quienes niegan a su teoría el estatus de teoría científica. Por tanto, «el diseño inte-ligente no es creacionismo ni naturalismo. Tampoco es un compromiso o síntesis de estas posiciones. Este nuevo enfoque se limita a seguir simple-mente la evidencia empírica de diseño hasta donde ésta conduzca. El dise-ño inteligente es una tercera vía»59.
De entrada, Dembski define el DI como «la ciencia que estudia los signos de la inteligencia»60. Pero es importante advertir que la noción de ciencia que tienen estos autores es muy problemática61, porque, por un lado, parece que no acaban de aceptar la distinción entre ciencia e inter-pretación filosófica, esto es, entre hechos e interpretaciones, que vendría hecha ya desde Kant, porque, según ellos, habría tenido la dañina conse-cuencia de relegar a Dios, la religión y la moral, al mero plano privado, ya que no tendrían nada que decir en el ámbito de la creación, ni tampo-co a la hora de formular las leyes morales y del comportamiento62; pero,
56 Cfr. Dembski, W. A., The Design Inference. Eliminating Chance through Small Probabilities, Cambridge, Cambridge University Press, 1995 (Trad. esp.: Diseño Inteli-gente, Madrid, Homo Legens, 2006).
57 Ayala, F. J., o.c., p. 144. 58 Cfr. Dembski, W. A., Diseño Inteligente, o.c., p. 24. 59 Dembski, W. A., o.c., 24.60 O.c., 18; cfr. 31-36. 61 Cfr. Dembski, W. A., parte seis de su obra; y en el libro de Ph. Johnson hay capítulos
finales que ponen en cuestión la idea general de ciencia para proponer otra. 62 Cfr. Colson, Ch. W., en la «Presentación» del libro de W. A. Dembski ya citado,
pp. 11-12

114
© Universidad de Deusto - ISBN 978-84-9830-383-4
por otra parte, parecen aceptarla, jugando a ser su doctrina una auténtica teoría científica, por lo demás auténticamente revolucionaria63, pero dan-do por hecho que hay que desconfiar e incluso modificar la definición de ciencia64.
El centro de la argumentación de Dembski se orienta a demostrar que hay argumentos suficientes y contundentes como para afirmar que las te-sis del darwinismo naturalista, que afirma que todo se debe a leyes me-cánicas en las que todo está regido por el azar y la necesidad, son insufi-cientes, teniéndose que aceptar que hay muchos ámbitos del mundo de la biología en que hay que echar mano del diseño inteligente. En definitiva, según Dembski, la afirmación central de las tesis del DI es que «hay sis-temas naturales que no pueden ser adecuadamente explicados en términos de fuerzas naturales carentes de dirección y que exhiben rasgos que en cualquier otra circunstancia atribuiríamos a la inteligencia»65.
Claro que estas afirmaciones hay que probarlas, y a ello se dedica De-mbski en su texto con ahínco y perseverancia, argumentando en una doble dirección: construir un sistema teórico desde el que definir una estructura de diseño inteligente, y mostrar también estructuras biológicas en las que se daría tal DI, no pudiéndose en ningún caso explicarse por meras causas naturales de azar-necesidad. La primera parte de la estrategia se centra en explicar el concepto de complejidad especificada (Dembski)66, cercano y similar al de complejidad irreducible (Behe)67. Para convertirse el paradig-ma del DI en alternativa al evolucionismo darwinista, Dembski considera que tiene que ser capaz de realizar cinco tareas: establecer criterios fiables para detectar inteligencia; aplicar estos criterios a los sistemas biológicos (aquí es donde se sitúan los conceptos ya indicados de complejidad espe-cificada o irreducible: «si éste consta de varias partes relacionadas entre sí de modo tal que la eliminación de una de ellas destruye la función en-tera del sistema»68); mostrar que estos criterios eliminan efectivamente los mecanismos materiales; reconceptualizar la biología evolutiva dentro
63 Cfr. Dembski, W., o.c., pp. 15 y ss. 64 Cfr. Johnson, Ph. E., o.c., cap. 9.º, «Las reglas de la ciencia» (132-145). Las críticas
de Johnson a la definición académica de ciencia se encaminan sobre todo a mostrar que los científicos darwinistas mezclan en sus argumentaciones afirmaciones científicas (criterios em-piristas) y filosóficas (tesis naturalistas): «el naturalismo da por supuesto que todo el rei-no natural constituye un sistema cerrado de causas y efectos, que no puede ser influido por nada de su “exterior”» (p. 136).
65 O.c., 25.66 O.c., pp. 87 y ss; 307 y ss.67 Cfr. Behe, Michael J, Darwin´s Black Box: The Biochemical Challenge to Evolution,
Nueva York, Free Press, 1996. 68 Dembski, W. A., o.c., 307.

115
© Universidad de Deusto - ISBN 978-84-9830-383-4
de un marco teórico de diseño; e inspirar un programa fértil de investiga-ción biológica que sea exclusivamente teórico-del-diseño69. De estos cin-co puntos, los dos últimos considera Dembski que son de momento una «cuestión prometedora», pero los tres primeros, en cambio, constituyen una palpable realidad.
Los casos concretos que los teóricos del DI presentan como prueba de la existencia de diseño inteligente son varios, como son la conformación de los ojos, las extremidades, las alas y otros órganos complejos, pero el caso más típico es la propuesta de Behe sobre el flagellum bacterial,como prueba de un sistema bioquímico irreductiblemente complejo70. Se trataría de una complejidad tal que no se habría podido dar una situación intermedia desde un sistema biológico anterior a él, que habría evolucio-nado, como dicen los darwinistas ortodoxos, a partir de un sistema menos complejo, por acumulación de cambios y novedades parciales. Lo correc-to, según la teoría del DI, es pensar que ha emergido de golpe, como un sistema diseñado por una inteligencia, resultando imposible entenderlo de otro modo.
En cambio, los evolucionistas darwinianos presentan multitud de pruebas para mostrar que los argumentos de Behe son palmariamente equivocados, como muestra detenidamente Ayala respecto al caso del ojo y del flagelo bacteriano, así como en el caso del sistema de coagulación de la sangre, el sistema inmunológico y otros fenómenos similares71. Las evidencias científicas apuntan a que la formación de estos sistemas com-plejos no se han dado a partir de saltos del tipo «todo o nada», sino que se explican todos a partir de un proceso gradual de mejora desde un sistema imperfecto a sistemas progresivamente mejorados, en los que se da, como en todo proceso emergente, tanto una continuidad como una mejora cua-litativa. En todos los ejemplos citados puede explicar el avance a través de la lógica evolutiva que defiende la teoría de la selección natural. Es-tas evidencias científicas contra la argumentación de Behe hicieron que el juez Jones, en la sentencia ya indicada del 2005 concluyera para este pun-to: «Por tanto hallamos que la declaración del profesor Behe a favor de la complejidad irreductible ha sido refutada en artículos de investigación académica y ha sido rechazada por la comunidad científica en general»72.
Como hemos indicado ya, estos teóricos del DI afirman que, en prin-cipio, no pretenden deducir de sus argumentaciones ninguna tesis religio-sa. Esa sería más bien la pretensión de la teoría antigua que se denominó
69 Cfr. ibídem, pp. 305-311. 70 Cfr. Behe, M. J., o.c.71 Cfr. Ayala, F. J., o.c., pp. 149-157. 72 Cita tomada de ibídem, p. 156.

116
© Universidad de Deusto - ISBN 978-84-9830-383-4
del diseño, presente en la historia del pensamiento desde épocas antiguas y defendida en la época de Darwin por Paley73. El DI se limita a mostrar que hay estructuras biológicas que sólo se pueden explicar desde un di-seño previo, dejando de lado las cuestiones acerca de la naturaleza del supuesto diseñador74. Por lo tanto, según ellos, no se trataría de una pro-puesta de teología natural75, ya que en ese caso no pasaría de ser, en el fondo, una propuesta teológica disfrazada.
Además, le resulta curioso y extraño a Dembski que las propuestas del DI hayan sido contestadas tanto por los científicos naturalistas como también por diversos teólogos. Pero no podía ser menos, puesto que desde ambos lados la propuesta presenta muchas debilidades. No es posible detenernos aquí en una crítica pormenorizada de las deficiencias del DI. Para ello hay que remitirse a libros que ya lo han hecho de un modo más o menos completo y profundo76. Nos vamos a limitar aquí a indicar y ana-lizar las dos limitaciones que considero más significativas.
b) La teoría del DI no es una teoría científica y es una mala teología
Coincido con Fco. J. Ayala77 en afirmar que el DI es un mal paradig-ma tanto en lo científico como en lo teológico. Es decir, su propuesta no es un paradigma científico sino una (mala) teoría filosófica, y, al mismo tiempo, se apoyan en un trasfondo teológico que no está en sintonía con el Dios del Evangelio de Jesús (o de la teología cristiana). Vamos a intentar explicitar esta doble crítica.
La pretensión de querer mostrar que existen estructuras biológicas tan complejas (complejidad especificada o irreducible) que sólo se pueden explicar desde la hipótesis de un diseño inteligente, es inconsistente. Por un lado, ya hemos indicado que la ciencia aporta suficientes pruebas como para demostrar que tales complejidades no son fruto de un salto de la nada al todo, sino fenómenos que se han ido produciendo por una acumulación gradual de mejoras y pequeños cambios. Pero, por otro lado, defender las tesis del DI supone apelar a un sujeto que ha diseñado tales estructuras de una complejidad irreductible. Es verdad que se apresuran a decir que ellos
73 Cfr. Dembski, W. A., o.c., pp. 67 y ss.; Ayala, Fco. J., o.c., pp. 31 y ss. 74 Cfr. Dembski, W. A., pp. 21 y 22. 75 Así lo entiende I. Barbour, en su intervención en la American Academy of Religión:
cfr. Dembski, W. A., o.c., 45-46.76 Cfr. Ayala, Fco. J., o.c.; Ruse, M., o.c.; y Barbour, I., El encuentro entre ciencia y re-
ligión, o.c., pp. 142-147; Küng, H., o.c. 77 O.c., pp. 21-22.

117
© Universidad de Deusto - ISBN 978-84-9830-383-4
no quieren detenerse en analizar la naturaleza de tal sujeto o inteligencia, sino a mostrar los efectos: una estructura diseñada de tal modo que no puede atribuirse a sólo causas materiales azarosas. Pero es fundamental tener que referirnos a esa inteligencia o sujeto para explicitar qué concep-ción de Dios y de la realidad tienen. No caben más alternativas, respecto a la autoría de tal supuesto diseño, que Dios, o algún espíritu invisible, un demiurgo (ángel o demonio). Querer implicar a Dios supondría acep-tar que realiza continuamente acciones milagrosas y que actúa como una causa segunda más, lo cual supone una concepción de Dios pre-ilustrada y milagrera. Por otro lado, admitir la existencia de númenes espirituales infra-divinos como sujetos a los que hay que atribuir tales diseños, supone también una concepción de la realidad milagrera y mágica.
Una idea ilustrada de Dios no puede situarlo al nivel de las causas se-gundas, sino más bien como fundamento de toda la realidad78. Una con-cepción de Dios como fundamento de la realidad supone entenderlo como soporte metafísico del mundo, que lo crea y lo mantiene en la existencia, pero, al mismo tiempo, el hecho de la creación supone también la total autonomía del mundo en su funcionamiento interno. De ahí que la exis-tencia de Dios y su interacción con el mundo no pueden ser demostradas, sino sólo postuladas. Dios no es demostrable, aunque su existencia es ra-zonable, deducible al menos desde la contingencia del universo79.
De ahí que, en nuestro acercamiento a la realidad mundana, tengamos que distinguir entre la descripción fáctica de su funcionamiento, tarea que le corresponde a la ciencia, y la interpretación del sentido de las grandes cuestiones que se plantean más allá o en el límite de la reflexión científi-ca. Así, en el estudio de las estructuras biológicas, donde los defensores del DI quieren ver un diseño inteligente, tienen que tenerse en cuenta dos niveles o facetas: la mera descripción del funcionamiento material de tales estructuras (tarea del científico, en cuyo análisis tiene que mantenerse en el puro nivel de la concatenación de causas materiales, y ahí es donde tie-ne la total legitimidad para defender un materialismo reduccionista en el nivel metodológico), y el nivel de la interpretación, pudiéndose proponer diferentes propuestas alternativas, entre las cuales se dan la atea materia-lista y la que advierte tal complejidad en la estructura de un ojo, un ala, un flagelo bacteriano, o cualquier otro organismo vivo, que en esa compleji-
78 Cfr. Zubiri, X., El hombre y Dios, Madrid, Alianza/Sociedad de Estudios y Publica-ciones, 1986.
79 Sobre la noción teológica de creación, cfr. Küng, H., o.c. pp. 93 y ss.; Edwards, Denis, El Dios de la evolución. Una teología trinitaria, Santander, Sal Terrae, 2006; Haught, John F., Cristianismo y ciencia. Hacia una teología de la naturaleza, Santander, Sal Terrae, 2009.

118
© Universidad de Deusto - ISBN 978-84-9830-383-4
dad «ve», «cree» que hay que postular en ella una actuación teleológica, procedente de Dios o de un ente divino. Pero querer empeñarse en saltar del nivel interpretativo al mero nivel científico, para defender que la pro-puesta de advertir un diseño inteligente en una estructura biológica com-pleja es una tesis científica, es mezclar ciencia con filosofía o teología, y es mostrar que la idea que se tiene de Dios se acerca mucho al Dios «tapa-agujeros», o al Dios milagrero, difícil de aceptar por creyentes ilustrados y que han superado una visión del mundo encantado y propio de una vi-sión animista.
Por tanto, en su idea de Dios y de su actuación en el mundo, el DI muestra también una mala teología, porque, además de las razones ya expuestas, supone no advertir que si se quiere ver en toda realidad com-pleja la presencia y la actuación de un diseñador inteligente, que en últi-mo término sería Dios, no sólo hay que atribuirle las cosas buenas y bien diseñadas, sino también los fallos y males del mundo y de la biosfera. La tendencia de los creacionistas es interpretar el conjunto del universo como un cosmos bien diseñado y maravillosamente bien organizado, cuando es evidente que, junto a las maravillas y perfecciones, el universo y el mun-do de la vida nos muestran también sus numerosos fallos y defectos. Vi-mos que W. Paley ya lo tenía en cuenta, y no sabía responder de un modo convincente a las serias cuestiones que la existencia del mal plantea a la fe en un Dios creador, bueno y providente, atribuyéndolas a causas que desconocemos, pero no a Dios.
Ayala, en su crítica a este aspecto del DI, hace ver que, si miramos el modo como está construido el ojo humano, por poner el ejemplo más so-corrido que se suele presentar por los defensores de la teoría del diseño y del DI, vemos que su estructura no tiene nada de perfecta, en la medida en que el nervio óptico, que conecta el ojo con el cerebro, está situado en el ojo huma-no de tal forma que produce un punto ciego en la retina, cosa que no ocurre, por ejemplo, en los calamares ni en los pulpos. Por tanto, si lo hubiera hecho un diseñador consciente, resulta lógico pensar que lo habría hecho mejor y sin ese fallo. El modo de resolver el problema de estos fallos es en los teóri-cos del DI similar a Paley, como se ve en Behe, quien responde a la cuestión de por qué se hizo de esa forma defectuosa en vez de hacerse de otra manera más acertada, apelando a que «las razones para que un diseñador haga o no haga cualquier cosa son prácticamente imposibles de saber a no ser que el di-señador te diga específicamente cuáles son sus razones»80.
Este modo de responder puede ser suficiente dentro del nivel teológi-co, pero no lo es como respuesta científica, puesto que se trata de una ar-
80 Behe, W., o.c., p. 223. Cita tomada de Ayala, F. J., o.c., p. 157.

119
© Universidad de Deusto - ISBN 978-84-9830-383-4
gumentación que no se puede comprobar de modo empírico, requisito ne-cesario para la ciencia. Además, dentro ya de la respuesta teológica, situar la acción de Dios en el ámbito del diseño directo de las estructuras vivas y de las leyes específicas del universo, supone una mala teodicea, un resol-ver de modo insuficiente el problema del mal, puesto que vemos no sólo que los organismos no son tan perfectos como se puede creer de forma in-genua, «sino también que las deficiencias y las disfunciones son ubicuas, lo que evidencia un diseño más “incompetente” que “inteligente”»81.
De este modo, como advierte Ayala, la teoría de la selección natural explica mejor que la teoría del DI estas imperfecciones de la naturaleza, sin necesidad de atribuírselas a Dios. Eso le lleva a concluir que la teoría de la selección natural ha sido «el regalo de Darwin a la teología»82, por-que la selección natural actúa seleccionando a posteriori las formas más adecuadas a cada ambiente de forma aproximada, no de modo perfecto, como lo haría un diseñador inteligente que actuara siguiendo un diseño, al estilo de una racionalidad prospectiva. Es la lógica aproximada y chapu-cera con la que, según nos dice F. Jacob, actúa la evolución por selección natural83, más acorde con la idea de un Dios creador que se autolimita en su relación con el mundo, y deja que actúen las leyes de la naturaleza, con sus limitaciones e imperfecciones.
Y de este modo, más que ser la teoría de la selección natural y las teorías sobre la historia del universo y la mecánica cuántica, basadas en las leyes probabilísticas y azarosas, frente a las propuestas determinísticas del pasado, un obstáculo para la fe en Dios, constituyen más bien una ocasión para re-novar la teología y la idea de Dios y su actuación sobre el mundo, así como para renovar la idea de nuestra libertad, del mal y de las grandes cuestiones que inquietan al ser humano acerca del sentido del mundo y de la vida84.
4.6. Evolución darwinista y fe cristiana
La opinión de que darwinismo y fe cristiana son incompatibles apa-rece con cierta frecuencia en autores y libros importantes, en los que se advierte la incorrección epistemológica de ciertas afirmaciones básicas.
81 Ayala, F. J., o.c., p. 158. Las comillas son del autor. 82 Cfr. ibídem, p. 158. 83 Cfr. Jacob, Fr., La lógica de lo viviente. Una historia de lo viviente, Barcelona, Edit.
Laia, 1977. 84 Cfr. Barbour, I., El encuentro entre ciencia y religión. ¿Rivales, desconocidas o com-
pañeras de viaje?, Santander, Sal Terrae, 2004, cap. 6.; Beorlegui, C., «Evolución y reli-gión, ¿Incompatibilidad o integración?» (texto a publicar en Realidad (UCA, El Salvador), 2010).

120
© Universidad de Deusto - ISBN 978-84-9830-383-4
Como muestra podemos ver dos ejemplos. El primero está sacado del li-bro de Javier Sampedro, Deconstruyendo a Darwin85, un libro excelen-te desde el punto de vista científico, pero, a mi entender, con errores de bulto desde el punto de vista filosófico. En una de sus páginas dice lo si-guiente: «Darwin había descubierto por fin una alternativa creíble al crea-cionismo, a la perogrullada que todo el mundo había dado por sentada hasta entonces, y que formulaba —o mejor, que ni formulaba— que las cosas de diseño inteligente, como los relojes y los seres vivos tenían for-zosamente que haber sido diseñadas por una inteligencia, como un reloje-ro o un dios. Fue la teoría de la selección natural la que refutó el famoso argumento teológico del diseño, tan pía y meticulosamente ensam-blado por el reverendo Paley. Si quieren loar a la persona que mató a Dios, no busquen en el entorno de Nietzsche. Pidan la lista de tripulantes del H. M. S. Beagle. La selección natural, es decir, la muerte de Dios, es la razón de la celebridad de Darwin fuera del ámbito de la biología»86.
Como puede verse, para J. Sampedro, de una teoría científica se pue-de concluir, de modo contundente y demostrativo, una afirmación filo-sófica y teológica: la supuesta muerte o no existencia de Dios. Está visto que a muchos científicos no les vendría mal algunas clases de epistemolo-gía. Porque no se trata de considerar ilegítima su postura atea o agnóstica (cada uno es libre de optar por el tipo de respuesta que considere más ade-cuada a la pregunta sobre el sentido último de la realidad), sino simple-mente considerar ilegítima su pretensión de que el ateísmo o agnosticismo se deduzca directa y necesariamente (como prueba concluyente) de una afirmación o teoría científica, en este caso la selección natural.
Un segundo ejemplo lo tomo del también excelente libro de Steven Mithem, Arqueología de la mente: orígenes del arte, de la religión y de la ciencia87. Mithem no se refiere al proceso evolutivo, en general, sino al modo como hay que entender el surgimiento de la mente humana. De sus reflexiones extraigo dos párrafos en los que se puede ver la in-correcta deducción filosófico-teológica de unas bases científicas. «Los creacionistas creen que la mente surgió de repente y ya completamen-te formada. De acuerdo a su visión de las cosas, fue un producto de la creación divina. Están equivocados: la mente tiene una larga historia evolutiva, y puede explicarse sin recurrir a poderes sobrenaturales»88.«La mente humana es un producto de la evolución, no una creación so-brenatural. He puesto claramente sobre la mesa la evidencia al respecto.
85 Barcelona, Crítica, 2002.86 Ibídem, p. 23. Las cursivas de la frase final son nuestras. 87 Barcelona, Crítica, 1998.88 P. 14.

121
© Universidad de Deusto - ISBN 978-84-9830-383-4
He especificado los “qués”, los “cuándos” y los “porqués” de la evolu-ción de la mente»89.
Para Mithem, desde los presupuestos del reduccionismo materialista, que dan por hecho un monismo ontológico y epistemológico, la realidad no tiene más que un solo plano, objeto de la ciencia, donde no cabe en ab-soluto la posibilidad de conjugar el nivel fáctico (en el que la emergencia de la vida, de la mente y de cualquier otro fenómeno biológico y mental, tiene que ser explicada por las ciencias correspondientes), con el nivel del sentido, donde cabe tanto interpretar todos esos fenómenos desde el pun-to de vista del azar, como también entenderlos desde una intervención de Dios, respetando por supuesto el ámbito de las denominadas causas se-gundas. Por eso, entendemos que cabe conjugar y complementar una teo-ría de la mente con un creacionismo ilustrado.
Pero veamos un poco más detenidamente estos puntos. Siguiendo lo que hemos indicado más arriba sobre la necesidad de distinguir el pla-no científico, el filosófico y el teológico, hay que reconocer que la teoría de la selección natural es una teoría científica, y su aceptación y rechazo tiene que realizarse en el mismo nivel, es decir, con argumentos de tipo científico.
Y, a partir de esos hechos científicos, cabe preguntarse por el sentidodel proceso evolutivo, haciéndonos preguntas tales como si en la evolu-ción hay azar, necesidad, o finalismo; o si cabe descubrir en ese mismo proceso evolutivo un mero resultado de proceso azarosos y aleatorios, o bien puede caber también una interpretación religiosa. Tanto si respon-demos con una interpretación atea o bien con otra postura religiosa, hay que aceptar que ambas respuestas son legítimas y razonables. Son legí-timas, porque la mera constatación de hechos no es suficiente para dar cuenta de la realidad, y la inteligencia humana, en cuanto razón (siguien-do la distinción de Zubiri: aprehensión primordial de realidad, logos y razón), está empujada a preguntarse por el sentido y fundamento profun-do de la realidad90. Y cualquier propuesta de sentido en el ámbito de la razón exige algún tipo de prueba racional, pero no es demostrable empí-ricamente, puesto que constituye una propuesta de sentido que no puede ser demostrada con el mismo tipo de requisitos empíricos que nos exigen las ciencias naturales.
Y las dos propuestas, la creyente y la no creyente, son razonables, en cuanto poseen una coherencia interna y no son contradictorias, y no se oponen a los datos científicos. Sólo se requiere de una propuesta filosófica
89 P. 230. 90 Cfr. Zubiri, X, Inteligencia sentiente, 3 vols., Madrid, Alianza/Sociedad de Estudios
y Publicaciones, 1980-1983.

122
© Universidad de Deusto - ISBN 978-84-9830-383-4
para ser razonable que sea coherente internamente y que no se oponga a los datos científicos debidamente contrastados. Por tanto, la teoría cientí-fica de la selección natural no se opone a la existencia de Dios, ni tampo-co demuestra que existe. En cuanto teoría científica es totalmente neutra respecto a cualquier propuesta de sentido. Ahora bien, esta teoría científi-ca no acepta cualquier tipo de idea de Dios; por ejemplo, no es consistente ni compatible con una idea de un Dios milagrero (el dios tapa-agujeros),que actúa directamente en los fenómenos naturales como si fuera una cau-sa segunda más, en expresión de la teología medieval.
Por consiguiente, es totalmente legítimo que cualquier científico, o un pensador cualquiera, diga que, según la teoría darvinista, le pare-ce más razonable la no existencia de Dios y opta por un sentido ateo del universo. Pero esa deducción no es una conclusión científica, sino filosó-fica o cosmovisional, que tiene el estatus de opción de fe, de opción vital sobre su vida y el universo. Lo mismo que hace el creyente: desde los da-tos que le aporta la experiencia del modo de ser del mundo, que la cien-cia la va completando y corrigiendo históricamente: cree y deduce que todo tiene sentido desde Dios, aunque también está claro que el progresi-vo conocimiento de la realidad le va haciendo comprender que la acción de Dios sobre el mundo no puede ser ni situarse al mismo nivel que las demás realidades mundanas. Si Dios existe, no puede ser una cosa u ob-jeto más del mundo, sino el fundamento de todo, sea lo que sea lo que entendamos por ello.
Por otro lado, como ya dijimos en el capítulo anterior, si nos atene-mos a lo que sabemos de la vida del propio Ch. Darwin, su postura res-pecto al alcance metafísico y religioso de su teoría era menos extrema y antirreligiosa que muchos de sus seguidores, y tampoco era tan clara-mente atea como algunos quieren suponer. Lo cierto es que Darwin no se consideró nunca ateo, sino agnóstico, y siempre fue muy cuidadoso en sus afirmaciones sobre las implicaciones de su teoría sobre las verda-des religiosas.
Por tanto, parece razonable concluir que se pueden compatibilizar los datos de las ciencias respecto a los grandes momentos de la historia del universo y de la tierra con las verdades de fe del cristianismo, como son el origen del universo y la creación de Dios, el origen de la vida, la apari-ción de las diferentes especies vivas y la aparición del género humano con la afirmación de la acción de Dios en cada uno de estos momentos, acción que no hay que considerarla milagrosa, al estilo de lo que defiende la teo-ría del Diseño Inteligente, sino una acción como fundamento de la reali-dad en todos sus ámbitos y en todos sus momentos históricos.
De hecho, la Iglesia Católica ha aceptado oficialmente la compatibili-dad entre el evolucionismo y la fe cristiana. El Papa Juan Pablo II, en un

123
© Universidad de Deusto - ISBN 978-84-9830-383-4
mensaje dirigido a los miembros de la Academia pontificia de ciencias91,reunidos en Roma para su asamblea plenaria del 22 de octubre de 1996, expresó pública y oficialmente la compatibilidad entre el evolucionismo darvinista y el cristianismo92. Lo que resultó sorprendente es que muchos comentaristas de medios de comunicación relevantes consideraran que se trataba de una gran novedad en la Iglesia Católica, cuando, como indicaba A. Matabosch, «en realidad se trata de una expresión pública y de magis-terio ordinario de lo que hace más de treinta años se enseña en todas las facultades de Teología y hasta se expone en los catecismos para niños»93.Es importante también matizar que Darwin nunca fue condenado por la Iglesia94, a diferencia de Galileo. La disputa propiamente se produjo, en-tre las diferentes iglesias, sólo con la Iglesia Católica, cuando el darwinis-mo se convirtió en una tesis filosófica materialista, de la mano sobre todo del biólogo alemán Haeckel, discípulo de Darwin, con la que pretendía demostrar que del darwinismo se deducía el ateísmo, la no existencia de Dios, y la aparente contradicción entre evolución y los relatos del Géne-sis. El Papa hace en su mensaje varias afirmaciones de interés. La prime-ra, que la evolución está ya consolidada como teoría científica, esto es, no es una mera hipótesis («la teoría de la evolución es más que una hi-pótesis»); por otro lado, también considera que es necesaria una interpre-tación rigurosa, no simplemente literal, de la palabra de Dios; afirma, en tercer lugar, que hay que distinguir entre el hecho de la evolución, de los mecanismos que la provocan (la mayor o menor importancia de la selec-ción natural, de las mutaciones al azar, etc.), y, de hecho, aunque la teoría evolucionista es ya reconocida por la mayoría de los científicos como una teoría consistente, advierte también que «más que de la teoría de la evolu-ción, conviene hablar de las teorías de la evolución»; y, por último, indica
91 Esta Academia tuvo sus inicios ya en el siglo XVI, aunque fue refundada por Pío XI hace más de setenta años, y está integrada por unos setenta científicos de prestigio de todo el mundo: cfr. Gomis, J., «Sobre el evolucionismo y la evolución de la Iglesia», IglesiaViva, 1996, n.º 186, pp. 555-560; 556.
92 La Iglesia Católica aceptó oficialmente la compatibilidad del evolucionismo y la fe cristiana en un mensaje del Papa Juan Pablo II a los miembros de la Academia pontificia de ciencias, con motivo de su reunión en Roma para su asamblea plenaria del 22 de octu-bre de 1996. Ecclesia, 1996, n.º 2.815, 16 de noviembre, pp. 25-26.
93 Matabosch, A., «El evolucionismo y la Iglesia», La Vanguardia, 1996, 25 de noviem-bre. De todos modos, hay otros comentaristas que, sin ser conscientes de esta realidad, in-cidieron más en añadir al asunto del evolucionismo al resto de enfrentamientos de la Iglesia Católica con la ciencia: José Manuel Vidal, «Darwin, perdónalos. 2000 años de errores de la Iglesia», El Mundo, 1996, 27 de octubre.
94 En realidad, como indica J. I. González Faus en La autoridad de la verdad, Santan-der, Sal Térrea, 2006, al año de la publicación del Origen de las especies, fue condenado por un sínodo episcopal de Colonia.

124
© Universidad de Deusto - ISBN 978-84-9830-383-4
que hay que distinguir entre el hecho y la teoría científica de la evolución y las teorías filosóficas que la interpretan, descartando entre ellas los re-duccionismos materialistas95.
Como ya se ha dicho, la inmensa mayoría de teólogos católicos expli-can en sus cátedras teológicas la compatibilidad del evolucionismo darvi-niano con la teología católica, diferenciando por un lado la teoría científi-ca de la evolución de sus interpretaciones materialistas reduccionistas, y, desde la teología, haciendo hincapié en la necesidad de una interpretación correcta de la Biblia, aplicando para ello los estudios sobre los géneros li-terarios y demás avances en el estudio de la interpretación bíblica96. Por eso que resultó extraño que varios comentaristas de periódicos europeos de prestigio, al glosar la declaración del Papa de 1996, además de conside-rarla una gran novedad (cosa que, como ya hemos dicho, no es cierto) la consideraron como un síntoma de resquebrajamiento del sistema doctrinal dogmático de la Iglesia. Es sintomático de una interpretación de este estilo, el editorial del prestigioso periódico parisino Le Monde, que afirmaba: «El distanciamiento así tomado con la lectura fundamental de la Biblia amena-za con afectar las estructuras de todo el edificio dogmático cristiano, fun-dado sobre el pecado original y la existencia del mal… La Iglesia se desdi-ce de sus previas posturas creando una vertiginosa fosa a sus pies»97.
El desfase que se advierte entre esta forma de pensar de ciertos comenta-ristas, supuestamente inteligentes y enterados, y el estado ordinario de la teo-logía eclesial así como de la práctica de las catequesis al interior de la Iglesia, hacen sospechar que la tarea de explicar al exterior de la Iglesia los cambios y modos de pensar y de evolucionar el pensamiento teológico se está llevan-do de un modo deficiente, porque la distancia no puede ser mayor98.
En conclusión, la opinión de muchos teólogos es que, lejos de ser in-compatible y negativa la relación y conjugación de la ciencia y la teolo-gía, para estos teólogos y teóricos evolucionistas creyentes99, la teoría de
95 Cfr. Matabosch, A., o.c.96 Cfr. Gomis, Joaquim, «Sobre el evolucionismo y la evolución de la Iglesia», Iglesia
Viva, 1996, n.º 186, 555-560; ISSAR, Aire S., «¿Se llevan bien la Biblia y la ciencia? Las plagas de Egipto y del Exodo pasadas por el tamiz de la hidrogeología», Mundo Científico,1996, n.º 166, 240 y ss.; Gutiérrez Lombardo, Raúl, «La teoría de la evolución y la Iglesia Católica», Ludus Vitalis, X(2002), n.º 17, 111-122; Alemañ Berenguer, Rafael, Evolucióny creación. Entre la ciencia y la creencia, Barcelona, Ariel, 1996; Núñez, Diego, «La re-ligión y la ciencia (Historia de las controversias entre ambas)», Mundo Científico, 1996, marzo, n.º 166, 247-256.
97 Cfr. Gomis, J., o.c., 557.98 Cfr. ibídem, 557. 99 Cfr. Edwards, Denis, El Dios de la evolución. Una teología trinitaria, Santander, Sal
Térrea, 2006; Barbour, I.G., o.c.; Ayala, Fco. J., Darwin y el Diseño Inteligente, o.c.; Küng, Hans, El principio de todas las cosas, o.c.

125
© Universidad de Deusto - ISBN 978-84-9830-383-4
la selección natural en lugar de ser un obstáculo para la fe, es una ocasión propicia para purificar la teología cristiana y entender del modo más ade-cuado la acción sobre el mundo, y purificando las superadas teodiceas y teorías acerca del mal, como vamos a ver en el último apartado de esta charla o escrito100.
4.7. El diálogo con la ciencia: oportunidad para la fe y la teología
De las cuatro posturas en las que resumía I. G. Barbour la contro-versia entre ciencia y teología, ya indicamos que la que nos resultaba más convincente era la cuarta, la integradora, pero en la línea de propo-ner una teología de la naturaleza, orientada a presentar la razonabilidad de la fe desde los datos más actuales de las diferentes ciencias. No se trata, ya lo decíamos, de resucitar o mantener la ya periclitada teolo-gía natural y sus pretensiones de demostrar la existencia de Dios y los praeambula fidei con la ayuda exclusiva de la razón, con el peligro de mantener un concepto de dios tapa-agujeros, que es permanentemen-te devaluado y tiene que retroceder a medida de que una nueva teoría científica halla una explicación natural a lo que hasta ese momento se consideraba un misterio inexplicable sin echar mano de Dios. Se trata, más bien, de seguir los planteamientos de una teología de la naturaleza, que respeta y se apoya en los datos científicos, no tanto para probar y demostrar las verdades de fe, sino para algo más modesto como es ha-cerlas plausibles y razonables, es decir, mostrar la no contradicción y la perfecta compatibilidad entre las verdades de la ciencia con las de la teología.
La postura de diálogo, por tanto, parte de la demarcación entre la mirada científica y filosófico-teológica, considerando ambas como le-gítimas y complementarias, en la medida en que la primera se mueve en el nivel de la constatación de datos, del funcionamiento de la reali-dad, mientras que la filosófico-teológica lo hace en el nivel de las in-terpretaciones, teniendo cada nivel diferentes modos de validación y demostración. Esto supone deslegitimar las pretensiones del naturalis-mo materialista de mezclar, a veces inconscientemente, ambos niveles, deduciendo de los datos científicos su postura filosófica reduccionista. Pero también se deslegitima desde aquí las pretensiones de los creacio-nistas y posturas similares de presentar un Dios milagrero, que se con-
100 Fr. J. Ayala, en su libro Darwin y el diseño inteligente, varias veces citado, titula el apartado donde reflexiona sobre este punto de este modo tan significativo: «El regalo de Darwin a la teología», p. 158.

126
© Universidad de Deusto - ISBN 978-84-9830-383-4
funde en su relación con el mundo con el resto de causas segundas, po-niendo en entredicho tanto la trascendencia de Dios como la autonomía del mundo. Claro que el gran reto de la teoría de las posturas del diálo-go e integración consiste en intentar proponer modelos de la actuación de Dios sobre el mundo que salve tanto la autonomía del mundo como la trascendencia de Dios.
El empeño de esta postura se orientará precisamente a mostrar que este modo de entender a Dios no sólo no supone un empobrecimiento de la imagen que la teología nos muestra de Dios, sino más bien una ex-celente oportunidad para renovar, purificar y enriquecer dicha imagen. Es verdad que la ciencia actual ha planteado serios reparos a diversos aspectos de la noción tradicional de Dios; pero al mismo tiempo le pre-senta un apoyo inestimable para proponer y hacer plausible otras imá-genes más adecuadas de Dios101. Los aspectos teológicos más renova-dos serían, en primer lugar, la imagen de Dios creador de un mundo en evolución, frente a un mundo estático102; una mejor conjugación entre ley y azar, determinismo y probabilidad en la naturaleza; una más ade-cuada comprensión de la libertad humana, frente a la tradicional teoría sobre la presciencia y preordenación divina, así como del problema del mal y del sufrimiento103; la superación de una imagen patriarcalista y androcéntrica de Dios, desde una nueva imagen de Dios Padre-Madre, recuperando para esa imagen de Dios muchos atributos femeninos que han sido tradicionalmente postergados y minusvalorados; y, por último, la superación de la intolerancia religiosa desde un diálogo interreligio-so abierto.
En resumen, la evolución y avance de las diferentes ciencias, tanto na-turales como humanas, constituye un acicate y una oportunidad para reali-zar un cambio en el modo de entender a Dios y su actuación en el mundo y entre los seres humanos. Pero, como puede verse, no se trata más que de intentos y barruntos de acercarse a la realidad de Dios, que siempre se nos aparecerá como un gran misterio. Pues, como decía San Agustín, «si lo comprendemos, entonces es que no es Dios».
101 Cfr. Barbour, I., El encuentro entre ciencia y religión, o.c., cap. 6.º.102 Cfr. Schmitz-Moormann, K. y Salmon, J. F., Teología de la creación de un mun-
do en evolución, Estella, Verbo Divino, 2005; Edwards, D., El Dios de la evolución, o.c.,cap. 3.º.
103 Cfr. Estrada, J. A., La imposible teodicea. La crisis de la fe en Dios, Madrid, Trotta, 1997; Ricoeur, Paul, El mal. Un desafío a la filosofía y a la teología, Buenos Aires, Amo-rrortu, 2006; Albertos, Jesús E., El mal en la filosofía de la voluntad de Paul Ricoeur,Pamplona, EUNSA, 2008; Greshake, Gisbert, ¿Por qué el Dios del amor permite que su-framos?, Salamanca, Sígueme, 2008; Gebara, Ivonne, El rostro oculto del mal, Madrid, Trotta, 2002.

127
© Universidad de Deusto - ISBN 978-84-9830-383-4
4.8. Ciencia y fe como aliadas y complementarias
En resumen, como hemos podido ir viendo, en este tema tan actual y recurrente de la relación entre ciencia y fe, se ha mostrado que no sólo no son dos saberes contrarios e incompatibles, sino más bien complementa-rios y llamados a integrarse adecuadamente.
Pero, para llegar a estas conclusiones, es necesario comenzar situan-do a cada tipo de saber en el nivel epistemológico que le corresponde. Los saberes científicos se ocupan de decirnos cómo es la realidad en sus diferentes facetas y aspectos, pero no les corresponde adentrarse en el ámbito del sentido o del significado cosmovisional, tarea propia de la fi-losofía y de la teología.
Y ya dentro del nivel del sentido, nos hallamos siempre con una plu-ralidad de planteamientos cosmovisionales y religiosos, la mayoría de los cuales son compatibles con los datos de las ciencias, aunque no todos lo son de la misma forma. De ahí que, en la medida en que la realidad no es transparente sino opaca a nuestras ansias de sentido, tenemos que deducir que tan legítima y razonable es la opción por la no fe como por la fe en Dios. Aunque no cualquier imagen de Dios es adecuada y compatible con la realidad que nos descubre la ciencia104.
Por tanto, si Dios es el creador del mundo, su realidad no puede ser incompatible con la obra que ha realizado. Pero Dios no es un objeto más del mundo, sino su fundamento. De ahí que no podamos pretender demos-trarlo y describirlo con los mismos métodos que utilizamos para describir y conocer los objetos del mundo, sino solamente postularlo como una hi-pótesis razonable.
Así, pues, son perfectamente compatibles la convivencia de opciones existenciales diferentes, como son la fe y el ateísmo, o el agnosticismo, aunque no todas ellas pueden ser igualmente verdaderas. Pero todas esas posturas son, en principio, compatibles con las verdades científicas, en la medida en que pertenecen a niveles diferentes de realidad, que se nos pre-senta como opaca y necesitada de sentido. La ciencia es el primer acerca-miento a la realidad, pero no el último; y la filosofía y la teología preten-den apuntar a la verdad definitiva sobre el mundo y la existencia humana, pero no pueden hacerlo sin la mediación de las ciencias. Las ciencias nos dan respuestas exactas sobre verdades penúltimas, decía P. Laín Entral-go, mientras que la filosofía y la teología, respuestas imprecisas sobre las cuestiones últimas, que son las que más nos interesan105. Ante el caso fa-
104 Cfr. Küng, Hans, Ser cristiano, Madrid, Cristiandad, 1977, pp. 62-104.105 Cfr. Laín Entralgo, P., Qué es el hombre. Evolución y sentido de la vida, Oviedo,
Ediciones Nobel, 1999.

128
© Universidad de Deusto - ISBN 978-84-9830-383-4
tídico de la muerte de una persona caída desde la ventana de un edificio, ¿qué nos importa más saber: la velocidad con la que ha caído, o cualquier otro tipo de datos que la ciencia puede resolver, o responder al dilema de si se ha caído, le han empujado o se ha tirado, y por qué? ¿Diríamos que estas últimas cuestiones no son importantes, o lo son menos, porque no se pueden responder con la exactitud del método científico? Sería absurdo dar una respuesta de ese estilo, cuando son precisamente esas cuestiones las que verdaderamente nos importan.
Podemos concluir, por tanto, con estas iluminadoras palabras de la carta que Juan Pablo II dirigió, en 1988, al jesuita P. Georges Coyne, di-rector en esos años del Observatorio Vaticano, escrita con motivo del cen-tenario de los Principia de Newton, con las que indicaba que, en vez de considerar incompatibles y enemigas a la ciencia y la reflexión teológi-ca, ambas pueden salir favorecidas en sus diálogos y encuentros. Decía el Papa: «La ciencia puede purificar a la religión del error y de la supers-tición; la religión puede purificar a la ciencia de idolatría y falsos absolu-tos. Cada una puede traer a la otra hacia un mundo más amplio, en el que ambas pueden florecer»106.
5. EL DARWINISMO Y LA ACTUAL COMPRENSIÓN DEL SER HUMANO
En resumen, aunque, como ya ha quedado señalado, la teoría de la evolución ha sido completada y perfeccionada, configurándose lo que se llama la teoría sintética de la evolución107, en la que se suman y com-plementan las tesis de Darwin con la dimensión genética de las leyes mendelianas y avances posteriores, entre los que se sitúa la genética de poblaciones, el estudio del ser humano no puede ser acometido con se-riedad y profundidad sin hacerlo desde el horizonte de la teoría de la evolución. Por tanto, cuando se trata de preguntarnos qué es el ser hu-mano, cómo se define frente a sí mismo y frente al resto de la realidad, y cuál es su puesto en el conjunto del cosmos, no podemos dejar de lado el estudio del proceso evolutivo en el que la ciencia nos lo muestra in-sertado. Este estudio nos proporciona datos fundamentales para acertar a describir de modo adecuado la realidad humana, puesto que, como seña-la Fco. J. Ayala, «no cabe duda de que una comprensión apropiada de la
106 Cita tomada de L. Sequeiros, ¿Puede un cristiano ser evolucionista?, o.c., p. 17. 107 Cfr. Ayala, Fco. J., o.c., p. 37-39.

129
© Universidad de Deusto - ISBN 978-84-9830-383-4
naturaleza humana requiere el conocimiento de que desciende de los an-tropoides, y, a través de ellos, de organismos simples a través de miles de millones de años de cambio gradual pero cumulativamente enorme. Es cierto que el descubrimiento del origen evolutivo de la humanidad no es suficiente para darnos a entender qué es el hombre, pero proporciona el punto de partida para ello»108.
Ahora bien, a pesar de que sin la teoría de la evolución y la selección natural no entendemos nada de lo humano, también es cierto que, a dife-rencia de determinados planteamientos reduccionistas y biologistas, tam-poco es suficiente la visión biologista del ser humano para dar cuenta de su auténtica naturaleza. Si como indica John Dupré, «es indiscutible que los grandes temas —la existencia de Dios, nuestra visión de la naturaleza humana y nuestra relación con otras creaturas— se ven afectados por las ideas acerca de la evolución»109, también son ciertas estas otras palabras suyas: «la evolución tiene de hecho trascendentales consecuencias para nuestra visión de nosotros mismos y de nuestro lugar en el universo, pero que no tiene en realidad la clase de consecuencias más ampliamente di-fundidas en la actualidad. En particular, tiene una utilidad limitada en lo que se refiere al esclarecimiento de la naturaleza humana»110. Por tanto, si la naturaleza o condición humana no se entiende sin su componente gené-tico y biológico, también hay que afirmar que la naturaleza humana no se reduce a esa única dimensión. Somos una unidad bio-cultural que no con-viene olvidar, si no queremos caer en un biologismo reduccionista y en un determinismo genético paralizante.
El estudio de los aspectos genéticos y biológicos de lo humano le corresponde en primer lugar a la Antropología físico-biológica111, cuya pregunta fundamental vendría a ser «¿quiénes son los hombres, los seres humanos?». Esto es, ¿qué rasgos son los que definen y caracterizan, en medio del conjunto de las especies vivas, a esta especie que somos noso-tros, la especie humana? Se trata, por tanto, de señalar los rasgos genéti-cos, biológicos, etológicos y comportamentales que configuran a nuestra especie.
Esto nos obliga a tener que estudiar al ser humano encuadrado en el conjunto de la biosfera, desde la comparación con el resto de las espe-cies vivas, en especial con las especies más cercanas y emparentadas a la nuestra. Por tanto, las cuestiones que tenemos que plantearnos e intentar responder son las siguientes:
108 Ayala, Fco. J., Origen y evolución del hombre, Madrid, Alianza, 1980, p. 153.109 El legado de Darwin, o.c., p. 9. 110 Ibídem, p. 12. 111 Cfr. Beorlegui, C., o.c., cap. 3, pp. 144 y ss.

130
© Universidad de Deusto - ISBN 978-84-9830-383-4
— ¿en qué se parece y en qué se diferencia la especie humana del res-to de las especies animales?
— sean cuales sean las diferencias, ¿lo son de tipo meramente cuanti-tativo o cualitativo?
— ¿lo específico de la especie humana se deduce de los parecidos con los demás animales, en especial las especies más cercanas a la nuestra (es la tesis del etólogo K. Lorenz y de los sociobiólogos), o más bien de las diferencias (como señalan las que vamos a denomi-nar antropologías humanistas);
— por tanto, ¿lo que define a la especie humana son sus características biológicas y genéticas, o más bien su condición cultural, esto es, el hecho de que sea un animal creador y creado por la cultura, aunque no sea fácil ponerse de acuerdo sobre el concepto de cultura, y so-bre si podemos aceptar algo así como una cultura animal?
— ¿cómo entender correctamente la relación que en el ser humano se da entre lo biológico y lo cultural? ¿qué diferentes modos existen de conjugar ambos ingredientes, y cuál sería el más adecuado?
— y, en definitiva, si la especie humana proviene por evolución del ám-bito de lo biológico (y nunca dejará de estar afincada en el ámbito de la biosfera) para asentarse y configurarse en lo cultural, ¿nos define más aquello de lo que venimos y procedemos, o aquello hacia lo que nos dirigimos, todo el continente de lo cultural, como continente inaca-bado y en permanente configuración y transformación?
— Otro modo de decirlo sería preguntarnos cómo se conjuga en el ser humano lo innato y lo adquirido, y cómo se compagina nuestra condición de seres libres con nuestra condición de especie animal.
Todas estas cuestiones se nos agolpan al iniciar esta temática tan apa-sionante y compleja, además de tan llena de actualidad, en virtud de los rápidos y constantes descubrimientos que se van sucediendo en el terreno de las diferentes ciencias humanas que estudian el origen del género hu-mano. En los capítulos que vienen a continuación trataremos de respon-derlas, hasta donde los descubrimientos científicos y la reflexión crítica de la filosofía nos lo permitan, comenzando por los datos sobre el origen del género humano, tanto en su dimensión filogenética como ontogenéti-ca, para después preguntarnos por los rasgos específicos que nos permitan seguir defendiendo la singularidad de la especie humana frente al resto de las demás especies animales que componen el ámbito de la biosfera.

131
© Universidad de Deusto - ISBN 978-84-9830-383-4
Capítulo 3
La antropogénesis. La dimensión filogenética y la cuestión del poligenismo/monogenismo
1. LA MICROEVOLUCIÓN Y LA MACROEVOLUCIÓN
Hemos hecho referencia ya en el capítulo anterior, aunque de forma muy superficial, a la microevolución y a la macreoevolución, como dos aspectos cercanos y relacionados, aunque muy diferenciados, del proceso evolutivo. Se trata de dos subprocesos o líneas evolutivas complementa-rias, aunque no todos los expertos están de acuerdo a la hora de estudiar sus contenidos y su incidencia en el conjunto de la evolución1. Veamos los rasgos más significativos de ambos.
a) Microevolución: se llama así al proceso de desintegración de una especie joven en diferentes grupos poblacionales, que llegarán a formar más adelante grupos múltiples, e incluso especies nue-vas. Así, de un grupo o especie arquetípica, se originan órdenes, familias o géneros nuevos. Además de microevolución, se deno-mina también a este proceso evolución especializadora o diversi-ficadora. Aparece, pues, como un movimiento descendente o di-versificador.
b) Macroevolución: es el punto de vista o proceso que nos conduce a la sucesión de arquetipos, es decir, a la aparición periódica de nue-vas formas de organización. Se trata, por tanto, de un proceso de
1 J. Ruffié, o.c., cap. 1.º, p. 26 y ss., señala que, en el fondo, no existe diferencia entre ambos procesos, dándose entre ellos a lo más una diferencia de grado.

132
© Universidad de Deusto - ISBN 978-84-9830-383-4
naturaleza ascendente y complejificadora. En relación a estos dos procesos, la pregunta básica que se suele plantear es: ¿cómo se pro-duce la especiación, esto es, la aparición de especies nuevas?
1.1. La microevolución
El proceso que se sucede en esta línea evolutiva parece sencillo: una especie joven, no especializada, con alto potencial genético y evolutivo, intenta establecerse en diversos nichos ecológicos. En cada nicho se da una determinada presión selectiva, y sobrevivirán y se difundirán los ge-nes que ayudan al grupo a su mejor adaptación. Se llega a diversificar un grupo de otro por efecto de la deriva genética, que pueden formar especies nuevas, por imposibilidad de cruzarse entre ellas. Así, de una especie indi-ferenciada llegarán a formarse varias especies diferenciadas. Cada especie se va adaptando a su nicho ecológico, produciéndose un cierto empobreci-miento del genoma del grupo, del polimorfismo genético, lo que dificulta-ría una nueva adaptación, si se produce un cambio ecológico brusco.
La microevolución es, por tanto, un proceso sin retorno, aunque siem-pre queda el acervo genético no expresado en el fenotipo, y que hace que pueda adaptarse de nuevo. Esta realidad ha permitido que desaparecieran muchas especies a lo largo de la historia de la evolución2. En cada uno de los procesos de microevolución, se produce un esquema general, que se repite con cierta constancia, y que suele constar de tres fases:
a) Fase de juventud: un grupo pequeño nuevo, no especializado, constituye el grupo pionero, la materia prima. Se trata de grupos oscuros, que no suelen dejar huellas en forma de fósiles y formas similares. Su escasa especialización les permite crear nuevos lina-jes más que prosperar en su forma inicial. Son especies-visagras(por ejemplo, las especies que se dieron entre reptiles y pájaros, o entre reptiles y mamíferos).
b) Fase de expansión o conquista: es la fase en la que el grupo pione-ro se extiende en múltiples nichos, y se adapta (si puede) a múltiples ambientes. Es la crisis evolutiva, como la denomina A. Vandel. Si se adaptan, vivirán; y si no, perecerán. Aquí se ve clara la tenden-cia a la especialización. En realidad, toda la vida surgió, al parecer, de los mares. Y en una segunda fase, saltó a tierra firme y al aire. Y
2 Cfr. Ayala, F. J., Origen y evolución del hombre, Madrid, Alianza, 1980; Id., La teoría de la evolución. De Darwin a los últimos avances de la genética, Madrid, Temas de Hoy, 1999.

133
© Universidad de Deusto - ISBN 978-84-9830-383-4
también ocurrió que algunos animales volvieron de nuevo al medio acuático, como es el caso de los insectos, ácaros y moluscos pulmo-nados, entre otros. Y se adaptan a ese nuevo medio por medio de la transformación de un órgano, que asegura la función indispensable en el nuevo ambiente. Al mismo tiempo, de forma complementaria, se van perdiendo órganos que ya no servirán para la vida en el nue-vo ámbito ecológico. Por ejemplo, las branquias cuando se sale del entorno acuático. Los órganos perdidos nunca reaparecen. Esa pa-rece ser la norma general, que se denomina regla o ley de Dollo, o también ley de la irreversibilidad de la evolución.
c) Fase de senectud: es la fase en la que el linaje entra en regresión. En general, las especies mejor adaptadas son las que desaparecen pri-mero. Al final, persisten sólo las especies denominadas reliquias o fósiles, en áreas especialmente protegidas. Antes se creía que esta regresión dependía de la competitividad natural intraespecífica. Pero hoy se sabe que el problema es más de tipo de variación ecológica,en la medida en que dichas especies no son capaces de superar los cambios bruscos en su entorno ecológico. Es decir, su especializa-ción les hace perder el poder de reconversión a nuevos entornos. A mayor especialización, mayor sensibilidad al cambio del medio. Este proceso de envejecimiento afecta a la mayoría de las especies vivas actuales. El hombre apareció tardíamente en un mundo enve-jecido, entre linajes en decadencia. Es probable que en la actualidad sólo entre los insectos se den algunos procesos de especiación.
Este proceso evolutivo ha sido denominado evolución adaptativa (an-glosajones), evolución-dispersión (Teilhard de Chardin), y evolución diver-sificadora o microevolución (A. Vandel). Pero este proceso no puede, por sí solo, abarcar todo el transformismo, en la medida en que sólo puede origi-nar múltiples variaciones a partir de un esquema de base, pero no crea nin-guno nuevo. Esto sólo lo puede hacer la macroevolución o tipogénesis.
1.2. La macroevolución o tipogénesis
Es el fenómeno por el que aparecen nuevos tipos de organización de seres vivientes. Schinden y Wolf lo denominaron tipogénesis, y A. Vandel, macroevolución. Ya no se trata aquí de una evolución especializadora, re-lacionada siempre con detalles anatómicos o fisiológicos, sino de la apa-rición de nuevos tipos, más o menos diferentes que los anteriores, y que serán, a su vez, en momentos posteriores el punto de partida de ciclos mi-croevolutivos.

134
© Universidad de Deusto - ISBN 978-84-9830-383-4
La tipogénesis recorre varios niveles, cada uno marcado por un esque-ma propio de organización, que se diferenciará en múltiples formas espe-cializadoras. Estos niveles forman los tipos y clases de la taxonomía; y en la fase de microevolución formará los géneros, especies y razas.
La macroevolución ha producido tipos muy diversos: espongiarios, celentéreos, anélidos, moluscos, artrópodos, equinodermos, procordados y vertebrados. Pero sigue, al parecer, una norma constante: la del desarrollo progresivo del sistema nervioso y de los órganos sensoriales. Este proceso posee un alto valor selectivo, como vamos a ver. Este desarrollo del siste-ma nervioso se observa en cada nivel evolutivo. En los más bajos, no hay psiquismo. En los intermedios, aparecen pequeños ganglios nerviosos, y una progresiva unión en cadena de dichos ganglios. Y en los reptiles y pe-ces, se produce ya una pequeña e incipiente cerebralización.
Y la misma tendencia se da en el conjunto de los niveles evolutivos. En cada nuevo tipo de organización, el sistema nervioso se complica y el psiquismo aumenta. En los artrópodos (insectos) y los vertebrados, extre-mos de las dos grandes ramas del reino animal, esta tendencia conduce a procesos de socialización, aunque formen modelos sociales diferentes. Pero en ambos casos se da agrupación de individuos jerarquizados. Por tanto, el desarrollo del sistema nervioso y del psiquismo parece ser la ten-dencia fundamental de la evolución.
Además, en la evolución, el sistema nervioso se comporta de forma muy distinta a los otros órganos. La hipertrofia de un aparato se traduce siempre en un cierto grado de especialización, produciendo una mayor adaptación del individuo a un medio y a un estilo de vida. Se trata de una de las bases de la microevolución. Pero el sistema nervioso es el único aparato que puede sufrir un desarrollo considerable sin que implique la especialización orgánica del individuo. El crecimiento del sistema ner-vioso y del psiquismo constituyen el fundamento mismo de la macroevo-lución. Simplificando, la macroevolución no es más que una especializa-ción hacia el psiquismo. Esto, como afirma Ruffié, no es una profesión de fe vitalista, sino un dato objetivo de la zoología, y ha de ser aceptado como tal.
La macroevolución, por tanto, sigue una dirección, que no es otra que la mayor complejidad del sistema nervioso y del psiquismo. Y esta com-plejidad coincide con un desarrollo de las facultades psíquicas, que culmi-na en los últimos estadios de los grandes phylum (phyla) del reino animal: los insectos y los primates. Tanto unos como otros presentan importantes fenómenos de socialización. Esto no es puro azar, sino explicable desde el esquema del darwinismo: según las condiciones ecológicas en que vi-vimos, el desarrollo del sistema nervioso y de sus funciones constituye la adquisición dotada de mayor poder selectivo.

135
© Universidad de Deusto - ISBN 978-84-9830-383-4
Estamos en un mundo de depredadores, en el que unos grupos viven a costa de otros. El enorme desarrollo de las plantas, con la fotosíntesis, con sistema autotrófico, parece que se tenga cortado por ahí cualquier progre-so y no se permita ya avanzar en el camino de la evolución. Pero el reino vegetal servirá para abastecer al reino animal con materia orgánica. Los animales son heterótrofos, y poseen mayor movilidad. Los primeros ani-males eran herbívoros, y después aparecieron los carnívoros, alimentán-dose a expensas de los demás. En cada nuevo estadio, se da un incremento de la eficacia, de logros del sistema nervioso y de las funciones psíquicas. El herbívoro parece no necesitar grandes capacidades psíquicas. En cam-bio, el carnívoro sí, porque al alimentarse de otros animales vivos, tiene que poder defenderse y saber atacar.
Las especies mejor informadas del mundo exterior son las más aven-tajadas, porque pueden reaccionar mejor. Con el desarrollo psíquico, se produce una diferenciación de los órganos reguladores e integradores (li-beración de los obstáculos externos). Aparece también la homeotermia,una forma de homeostasis (se da ya en los pájaros y en los mamíferos, algo que no tienen los reptiles ni los batracios, animales de sangre fría), que ayuda a soportar los cambios climáticos y a liberarse de sus incle-mencias.
Lo más importante, pues, que se produce en el terreno de la ma-croevolución es el desarrollo del psiquismo, que permite establecer fuer-tes lazos sociales e ir conformando comunidades cada vez más fuertes, complejas y estables. La creación de comunidades implica especialización entre los miembros, integración armónica de los miembros, y poseer al-gún medio de comunicación entre sí. La socialización aumenta la eficacia del individuo. Cada animal se convierte en un especialista, que realiza una función mejor que el animal solitario, ya que éste tiene que solucionar él solo todas sus necesidades. Aumenta de este modo la eficacia del conjun-to, en la medida en que cada uno trabaja para todos. La sociedad forma un todo; viene a ser como un super-individuo, más eficaz que la suma de un conjunto de individuos aislados.
La socialización constituye el estadio más avanzado de la macroevo-lución: corresponde a un cambio de niveles de integración. Se ha dicho que el sistema nervioso era el único órgano que podía complejificarse sin que el individuo emprendiera el camino de especialización orgánica. El desarrollo del sistema nervioso sí es especialización, pero en sentido diferente al normal. Esta especialización no se da respecto al medio par-ticular, sino respecto al conjunto ecológico. El animal de más elevado psi-quismo es el más competitivo. Así, el hombre es el más dotado desde este punto de vista, porque, en primer lugar, percibe mejor su entorno y lo in-tegra mejor; en segundo lugar, posee mejores medios de comunicación, lo

136
© Universidad de Deusto - ISBN 978-84-9830-383-4
que le permite socializar mejor el conocimiento; y, en tercer lugar, puede componer sociedades más amplias y complejas. Y todo esto es fruto de la presión selectiva.
1.3. El paso de la macro a la microevolución
Si sólo hubiera microevolución, el mundo de lo vivo no hubiera llega-do muy lejos. La evolución adaptativa no es capaz de crear un tipo verda-deramente nuevo. Hipertrofia o suprime, pero no innova. Es sinónimo de senectud; evolución sin esperanza.
En cambio, la creación supone una marcha atrás, un verdadero rejuve-necimiento, que libera al animal de la especialización. La desdiferencia-ción es el precio que hay que pagar para recuperar potencialidades que le permite dar un nuevo salto evolutivo, y reemprender un nuevo salto evo-lutivo. Estos rejuvenecimientos esporádicos, que permiten pasar de un ci-clo microevolutivo al siguiente, forman la trama de la macroevolución y corresponden casi siempre a la neotenia.
La neotenia es la aparición de facultades reproductivas en un orga-nismo que ha conservado sus caracteres embrionarios. Cuando la neo-tenia se hace permanente, crea una nueva especie que conserva, en el estado adulto, la mayoría de los caracteres fetales; o sea, una falta de especialización. Es el fenómeno que otros han llamado paidomorfosis o paidogénesis (W. Garstang, 1922). Esto se da, de forma especial, en la especie humana, puesto que, estudiando su proceso embriológico, el re-cién nacido nos recuerda en muchos aspectos el feto de los grandes mo-nos3. Por eso, a este fenómeno se le denomina también fetalización.
En conjunto, la evolución consistiría en una serie de ciclos especiali-zadores en el curso de los cuales las formas se multiplican, y en gran me-dida desaparecen. Pero, de vez en cuando, surgen por neotenia algunos tipos nuevos portadores de potencialidades evolutivas. Son el punto de partida de un nuevo nivel, que seguirá el proceso microevolutivo. Las for-mas de transición que han permitido el paso de un nivel a otro, y que son la trama de la macroevolución, son pocas, y normalmente poco persisten-tes, Pero sobre esto se tiene hoy todavía poca información.
La aparición periódica de nuevos tipos en grupos envejecidos cons-tituye la ley de transmisión (A. Vandel), y funciona como una serie de plataformas, en las que se diferenciarían una multiplicidad de especies
3 Cfr. Ruffié, J., o.c., p. 32; Christen, Yves, El hombre biocultural. De la molécula a la civilización, Madrid, Cátedra, 1989.

137
© Universidad de Deusto - ISBN 978-84-9830-383-4
(microevolución)4. La evolución sería como un inmenso libro en que cada capítulo narra la aventura de un tipo de organización, que resulta una aventura monótona, con idénticas fases: nacimiento, expansión y enveje-cimiento. Es el proceso que hemos denominado microevolución.
En cambio, la macroevolución conforma la sucesión de los diversos ca-pítulos del libro, y nos habla de seres cada vez más complejos. El hombre constituye el último capítulo de la historia de la vida, pero con sólo algunas páginas del primer acto. El segundo acto, la especialización, está sólo esbo-zado. El ser humano, al acceder a la cultura, ha cambiado los condiciona-mientos que dirigían un linaje hacia la evolución especializadora y hacia la senectud. De este modo, el hombre permanecerá siempre joven e indiferen-ciado. La historia natural no llegará más lejos, porque las últimas páginas no se escribirán jamás. Y es mejor que así sea, pues en la evolución biológi-ca, la marcha triunfal de la conquista es también la de la muerte.
El ser humano, pues, es una especie más que ha emergido en el proce-so evolutivo siguiendo las leyes generales que la han conformado a lo lar-go de su compleja historia. Hemos indicado los aspectos generales de este proceso evolutivo, tanto en el aspecto microevolutivo como macroevolu-tivo. A continuación nos vamos a detener en estudiar el proceso de forma-ción de la especie humana, o antropogénesis, como último eslabón surgi-do de la línea de los primates.
2. LA DIMENSIÓN FILOGENÉTICA DEL ORIGEN DEL HOMBRE
Se denomina antropogénesis al proceso completo de la formación del ser humano. Abarca tanto el proceso de formación de la especie (filogéne-sis) como el de cada individuo (ontogénesis). Estudiaremos los dos pro-cesos, empezando por la filogénesis. Y dentro de esta dimensión filogené-tica, en este capítulo nos centraremos en el proceso diacrónico, histórico, de la aparición de nuestra especie como un paso más de la evolución. En el capítulo cuarto nos dedicaremos a mostrar los cambios genéticos y mor-fológicos experimentados por el género humano a lo largo del proceso de humanización, para detenernos en el capítulo quinto en el proceso de huma-nización, es decir, en los aspectos culturales y conductuales que el ser hu-mano fue experimentando en el largo proceso de convertirse en humano.
4 Cfr. Vandel, A., L’homme et l’Evolution, 1958. Vandel presenta en este texto un diagrama filogenético, donde se advierte la correlación entre los diferentes tipos y espe-cies, viéndose cómo se conjuga la microevolución con la macroevolución: Cfr. Ruffié, J.,o.c., p. 33.

138
© Universidad de Deusto - ISBN 978-84-9830-383-4
2.1. Las diversas fases de la aparición de la especie humana
Creo que podemos afirmar sin exageración que nos hallamos en una nueva edad de oro de la paleoantropología (estudio del hombre antiguo, ciencia que se encarga de investigar todo lo referente a los orígenes del ser humano) y de otras ciencias afines y auxiliares (paleontología, zoolo-gía, biología, genética, arqueología, etc.), por lo que los científicos cada vez son más cautos a la hora de hacer afirmaciones tajantes, y son cada vez más conscientes de la provisionalidad de sus descubrimientos e inter-pretaciones. Igualmente tenemos que ser conscientes de que los métodos de medición de las edades de los fósiles y otros restos antiguos encontra-dos, se hallan en permanente proceso de perfeccionamiento, y, por ello, sus resultados son siempre aproximativos, y aunque se avanza con pasos agigantados en el empeño de conseguir métodos de medición más precisa, es importante advertir que las cifras que se manejan son siempre provisio-nales y expuestas a revisión continua.
Hasta no hace mucho tiempo, la paleoantropología era la única dis-ciplina científica que se encargaba de investigar todo lo referente al ori-gen del ser humano y sus diferentes etapas y características. Hoy día, los avances en biología molecular5 están permitiendo un acercamiento al pro-blema desde el modo como la estructura molecular, biológica, de los seres vivos se ha ido combinando y descomponiendo, asistiendo al nacimiento de lo que ya se está denominando la Antropología molecular6.
Por tanto, la paleoantropología y la biología molecular constituyen en la actualidad dos modos complementarios (aunque sean vistos por al-gunos otros autores como dos métodos enfrentados y excluyentes) de acercamiento al problema del origen del ser humano. Ambos modos de ac-ceso al problema tienen sus ventajas, pero también sus respectivos incon-venientes o limitaciones.
Las ventajas de la paleoantropología se hallan en que, cuando se des-cubren restos fósiles, parece que nos situamos ante pruebas objetivas y contundentes sobre aspectos fundamentales de nuestros diversos antepa-sados. Los inconvenientes surgen de la lentitud de los descubrimientos, en la medida en que se depende de los aleatorios hallazgos fósiles, hallaz-
5 Cfr. Ricards, Olga, y otros, «La Biología molecular», en Rebato, E., y otros (eds.), Para comprender la Antropología biológica. Evolución y biología humana, Estella, Verbo Divino, 2005, pp. 473-483; Johanson, Donald y Edey, Maitland, El primer antepasado del hombre, Barcelona, Planeta, 1987 (3.ª de.); Id., La cuestión esencial. Preguntas y respues-tas sobre la evolución, Barcelona, Planeta, 1990; Lewin, Roger, La interpretación de los fósiles, Barcelona, Planeta, 1987.
6 Cfr. Lewin, Roger, «El nacimiento de la Antropología molecular», Mundo Científico,1991, n.º 119, pp. 1222-1231.

139
© Universidad de Deusto - ISBN 978-84-9830-383-4
gos no planeados sino descubiertos muchas veces por un golpe de suerte. Además, se trata de restos incompletos, que hay que estudiar con mucho cuidado tanto en sí mismos como dentro del contexto geológico y paleon-tológico en el que hay que situarlos. En resumen, se tiene que ir compo-niendo el rompecabezas con pocas piezas y, en la mayoría de los casos, muy incompletas. Además, no todas las partes del cuerpo humano ni sus herramientas o restos culturales pueden fosilizarse; tampoco siempre se fosilizan las que son susceptibles de hacerlo. Debemos añadir a lo anterior el hecho de que los métodos de datación cronológica de los fósiles, aun-que van perfeccionándose continuamente, todavía son limitados para da-tar con precisión los hallazgos. Desde el método llamado del «carbono ca-torce» hasta los actuales del «argon-carbono» o «argon-argon», los pasos han sido espectaculares7. Las limitaciones, por tanto, son evidentes.
Las ventajas y limitaciones de la biología molecular parecen ser com-plementarias respecto a la paleoantropología, puesto que su objetivo se centra en advertir los cambios que en el proceso evolutivo se han dado den-tro de la estructura biológica y molecular de cada especie. De tal modo que cada célula de un ser vivo se convierte en una especie de caja negra don-de han quedado reflejados los cambios que se han producido a lo largo de todo el proceso evolutivo anterior. Sólo se necesita descubrir los elementos, dentro de esa estructura proteínica, que nos indiquen el ritmo y los cam-bios producidos. Es lo que se denomina el reloj molecular. De este modo, la ventaja de la vía molecular para descubrir la historia de la evolución con sus pasos correspondientes, como son, la separación y parentesco de cada una de las especies respecto a las demás, y los momentos en que tales bi-furcaciones se han realizado, es extraordinaria. Ya no se necesita esperar a los hallazgos azarosos de los restos fósiles para dibujar el proceso evoluti-vo, puesto que todo se hallaría escrito en nuestra estructura molecular y en el cariotipo. Pero las limitaciones son también grandes, puesto que, como veremos cuando más adelante nos detengamos con más detalle en el análi-sis de los descubrimientos de la biología molecular, se está todavía lejos de haber descubierto con precisión todos los mecanismos de las transformacio-nes proteínicas, así como de poder defender con certeza que esos cambios se han dado de modo uniforme, aspecto absolutamente necesario para poder ser considerado como un reloj molecular certero y fiable.
Teniendo en cuenta todas estas limitaciones, vamos, pues, a presentar a grandes rasgos lo que hoy día nos dice la paleoantropología y la biolo-gía molecular (antropología molecular) sobre nuestros orígenes.
7 Cfr. Lewin, R., La interpretación de los fósiles, o.c., y Arsuaga, J. L./ Martínez, I., Laespecie elegida, Madrid, Temas de Hoy, 1998; Cavalli-Sforza, Luca y Francesco, ¿Quiénessomos? Historia de la diversidad humana, Barcelona, Crítica, 1999.

140
© Universidad de Deusto - ISBN 978-84-9830-383-4
2.1.1. El punto de vista de la Paleoantropología8
Los saberes científicos ofrecen cifras cada vez más aquilatadas so-bre la historia del universo y del sistema solar al que pertenecemos, así como del origen de nuestro planeta y del surgimiento de la vida en ella9.A pesar de ello, las cifras que se nos dan hay que entenderlas y aceptarlas como cantidades aproximativas y susceptibles de corrección.
— Según estos datos, el Universo, según la teoría del Big-Bang, se originó en una fortísima explosión hace aproximadamente entre 15.000 y 10.000 millones de años10.
— El origen de la Tierra se sitúa hace entre unos 4.600 y 4.500 millo-nes de años11.
— La aparición de la vida en nuestro planeta se habría dado hace 3.800 millones de años12.
8 La bibliografía sobre este punto es inabarcable. Boyd, Robert/Silk, J. B., Cómo evolu-cionaron los humanos, Madrid, Barcelona, Ariel, 2001; Cela Conde, C. J./Ayala, F. J., Senderos de la evolución humana, Madrid, Alianza, 2001; Arsuaga, J. L./Martínez, I., La especie ele-gida, Madrid, Temas de Hoy, 1998; Arsuaga, J. L., El enigma de la esfinge, Barcelona, Plaza y Janés, 2001; Bermúdez de Castro, José M.ª, El chico de la Gran Dolina. En los orígenes de lo humano, Barcelona, Crítica, 2002; Carbonell i Roura, E./Mosquera Martínez, M., Las claves del pasado. La llave del futuro, Tarragona, Arola Editores, 2000; Rebato, E., y otros, Para comprender la Antropología biológica, o.c.; Ruffié, J., o.c., pp. 161 y ss.; Coppens, Y., La historia del hombre, Barcelona, Tusquets, 2009; Ayala, Fr. J., Origen y evo-lución del hombre, o.c., cap. 5.º; Ayala, Fr. J./Cela Conde, C. J., La piedra que se volvió pa-labra. Las claves evolutivas de la humanidad, Madrid, Alianza, 2006.
9 Para un acercamiento a esta datación cronológica más completa, cfr. Gribbin, John y Gribbin, Mary, Hijos del hielo. Clima y orígenes del hombre, Madrid, Pirámide, 1992; Erickson, Jon, La extinción de las especies, Madrid, McGraw-Hill, 1992, p. 22; Sagan, Carl, Los dragones del Edén, Barcelona, Crítica, 1993, pp. 22 y ss.
10 Cfr. Hawking, S., Historia del tiempo. Del bi-bang a los agujeros negros, Barcelona, Crítica, 1988; Id., El universo en una cáscara de nuez, Barcelona, Crítica/Planeta, 2002; Coles, Peter, Hawking y la mente de Dios, Barcelona, Gedisa, 2004.
11 Cfr. Luisi, Pier Luigi, La vida emergente. De los orígenes químicos a la biología sin-tética, Barcelona, Tusquets Editores, 2010, p. 71; Erickson, Jon, La extinción de las espe-cies. Evolución, causas y efectos, Madrid, McGraw-Hill, 1992, pp. 1 y ss.
12 Cfr. Oparin, A.I., Proishkhozhddenie Zhisni, Moskowski Rabocii, 1924; Id., The Origin of Live, Dover Publications, 1953 (trad. esp.: El origen de la vida, Madrid, Akal, 1994); Id., The Origin of Live on Erth, Academic Press, 1957, 3.ª ed. (trad. esp.: El origen de la vida sobre la Tierra, Madrid, Tecnos, 1979); Haldane, J. B. S., «The Origin of Live», Rationalist Annual,1929, 148, pp. 3-10; Id., «The origine of live», New Biol., 1954, 16, pp. 12-27; Luisi, Pier Luigi, La vida emergente. De los orígenes químicos a la biología sintética, Barcelona, Tusquets, 2010, cap. 3.º; Erickson, Jon, o.c., pp. 21 y ss.; Lane, Nock, Los diez grandes inventos de la evolución,Barcelona, Ariel, 2009, cap. 1.º, «El origen de la vida». No hacemos referencia aquí a los auto-res que defienden el posible origen extraterrestre de la vida. Son tesis que tienen que ser todavía contrastadas, pero que constituyen un punto de investigación central de la astrobiología actual.

141
© Universidad de Deusto - ISBN 978-84-9830-383-4
— El origen de los primates se situaría alrededor de 65/70 millones de años13.
— Si nos preguntamos por el antepasado común de los homínidos y de los simios o primates, la respuesta resulta en la actualidad todavía muy oscura, pero se calcula que hay que situarlo entre 24 y 14 mi-llones de años antes de nosotros14.
— La separación entre los póngidos (orangután, gorila y chimpancé) y los Australopitecos, se suele situar entre los 14 y 11 millones de años, con las precisiones que más adelante se harán.
— Y la aparición del primer hombre (Homo habilis) se produjo hace 2,4 millones de años.
Si se comparan estas fechas, con todo lo que tienen de provisionales y aproximativas, advertimos que el ser humano ha emergido en el proce-so evolutivo en fechas muy recientes. Algunos autores han realizado un ejercicio comparativo de estas fechas con el calendario de un año solar. De tal modo que, si consideramos que el origen del universo marca el arranque del año, la especie humana sólo habría hecho acto de presencia hacia las 22,30 de la noche del último día del año. Somos, pues, una es-pecie que acaba de estrenarse en la existencia15.
La historia de la paleoantropología está llena de interesantes episo-dios de descubrimientos apasionantes, así como de equivocaciones nota-bles y de fraudes espectaculares16. Pero aquí no vamos a detenernos en tantos detalles, porque nos extenderíamos más de lo que corresponde a
13 Cfr. Tudge, Colin/Young, Josh, El eslabón. La historia del fósil de primate más an-tiguo del mundo, Barcelona, Debate, 2010. En este libro se nos describe la historia del descubrimiento de un fósil, Ida, en la Fosa de Messel, cerca de Frankfurt, de una antigüe-dad de 47 millones de años, y que representaría el primer eslabón entre los primeros pri-mates y nosotros.
14 La dificultad de situar una fecha razonable para este antepasado común «tiene que ver con el hecho de que el registro fósil de los simios del Mioceno, con ser muy abundante, resulte parcial. En el Mioceno temprano (entre 24 y 16 m. a.), los ejemplares hallados son africanos, como, por ejemplo, el Procónsul. Del Mioceno medio (16 a 10 m. a.) contamos con los Dryopithecus europeos y los Afropithecus y Kenyapithecus africanos. Del Mioceno superior (de 10 a 5 m. a.), los fósiles disponibles son asiáticos (Sivapithecus, Lufengpithecusy Ankarapithecus, entre ellos) y europeos (con Oreopithecus y Ouranopithecus, por nom-brar sólo dos géneros que se añaden al Dryopithecus). No se han hallado fósiles de homi-noideos del Mioceno superior en África, salvo restos demasiado incompletos como para poder sacar conclusiones evolutivas de ellos»: Ayala, Fr. J., /Cela Conde, C. J., La piedra que se volvió palabra. Las claves evolutivas de la humanidad, o.c.
15 Cfr. C. Sagan, o.c., p. 23.16 Cfr. Johanson, Donald/Edey, Maitland, El primer antepasado del hombre, Barcelona,
Planeta, 1982; Lewin, Roger, La interpretación de los fósiles. Una polémica búsqueda del origen del hombre, Barcelona, Planeta, 1989.

142
© Universidad de Deusto - ISBN 978-84-9830-383-4
este texto. Solamente vamos a fijarnos los resultados más sobresalientes de las diversas investigaciones, para exponer el estado actual de lo que se sabe sobre los antecedentes de nuestros orígenes.
Desde la aparición de El origen de las especies y El origen del hom-bre, los dos textos centrales de Ch. Darwin, el entronque del ser huma-no en la biosfera se expresaba con la afirmación de que descendemos del mono, en la medida de que los monos, sobre todo el chimpancé, es la es-pecie, de entre las actualmente existentes, más cercana y parecida a la nuestra. A partir de ahí, la ciencia se ha ido esforzando en la búsqueda del denominado eslabón perdido, de la especie o especimen intermedio en-tre el chimpancé y el ser humano. Pero las evidencias fósiles que durante este siglo han ido apareciendo, nos han ido mostrando que la realidad es mucho más compleja de lo pensábamos, tanto respecto a nuestra conexión con los simios, como también respecto a la evidencia de existir múltiples eslabones intermedios.
El cuadro taxonómico del mundo de la biosfera sitúa a los seres huma-nos, la especie humana (o el conjunto de las subespecies que componen el linaje humano), dentro del orden de los primates, que, a su vez, estaría com-puestos por diversos subórdenes, superfamilias y familias, según el siguien-te cuadro17:
— Orden de los primates:
• Suborden 1. Prosimii. • Suborden 2. Tarsioidea. • Suborden 3. Antropoidea:
— Superfamilia 1. Ceboidea (monos del Nuevo Mundo). — Superfamilia 2. Cercopithecoidea (monos del Viejo Mundo). — Superfamilia 3. Hominoidea.
— Los hominoideos, a su vez, están clasificados en tres grandes familias:
1) Familia 1. Hilobatidae (gibón). 2) Familia 2. Pongidae (orangután, gorila, chimpancé). 3) Familia 3. Hominidae.
Esta clasificación corresponde a los grupos de seres que siguen vivos en la actualidad. Pero los descubrimientos paleontológicos nos indican
17 Cfr. Cela Conde, C./Ayala, F. J., Senderos de la evolución humana, o.c. (Alianza,2001), p. 84. Los autores toman el cuadro taxonómico de Simpson, G. G., «The Principles of Classification and a Classification of Mammals», Bulletin of the American Museum of Natural History, 85: 1-350; Arsuaga, J. L., El collar del neandertal, Madrid, Temas de Hoy, 1999, p. 78.

143
© Universidad de Deusto - ISBN 978-84-9830-383-4
que existieron otras especies entre el chimpancé y el hombre actual. Lo quesí está claro es que, como señala José Luis Arsuaga, «nos hemos queda-do solos. No hay ninguna especie animal que se parezca a la nuestra, ya que somos únicos. Un abismo nos separa en cuerpo y sobre todo en men-te del resto de las criaturas vivientes»18. ¿Qué ha pasado para que esto sea así? ¿Qué otras especies vivas han ido emergiendo y desapareciendo, de-jando en el camino la especie humana tal y como actualmente somos so-bre la tierra? Estas son las preguntas que se ha ido haciendo e intentando responder la paleoantropología.
a) La línea de los hominoideos o antropomorfos
Empezaremos por la familia de los hominoideos, ampliando y comple-tando en primer lugar el cuadro que más arriba acabamos de presentar19.
Superfamiliade los
HOMINOIDEOS
1. Póngidos
Gibones y siamang (simios inferiores)
Orangutanes
Gorilas
Chimpancés: Chimpancé pigmeoChimpancé común
2. Homínidos
Orrorin tugeniensisArdipithecus (Kadabba, Ramidus)
Australopithecos
A. Anamensis A. AffarensisA. Áfricanus A. Garhi
Humanos
H. HabilisH. Erectus/ergaster
H. AntecesorPre-heidelbergensis Heidelbergensis Neanderthal
H. Sapiens (Cro-magnon)
18 Arsuaga, J. L., El collar del neandertal, o.c., p. 27.19 Cfr. Arsuaga, J. L., El collar de l neandertal, o.c., p. 32; Bertranpetit, J./Junyent, Cr.,
Viaje a los orígenes, Barcelona, Península, 2000, p. 82.

144
© Universidad de Deusto - ISBN 978-84-9830-383-4
En síntesis, la especie humana actual pertenece, dentro de las siste-matizaciones de los taxonomistas, a la superfamilia de los hominoideos,que comprende a su vez dos familias: los póngidos y los homínidos20.La familia de los póngidos o simios suele ser también clasificada en dos grupos: los simios inferiores, a los que pertenecen los gibones y los sia-mang, y los simios superiores, que comprenden los orangutanes, gorilas y chimpancés. Dentro de los chimpancés, hay que distinguir las dos es-pecies existentes: el chimpancé pigmeo y el chimpancé común. Algu-nos incluso hablan de una tercera especie de chimpancés, Pan troglodyte verus. Pero no todos la admiten como especie diferenciada21. Las eviden-cias fósiles muestran que la relación entre estas diferentes especies de póngidos se configura en forma de un árbol con diferentes ramas, dando lugar a lo que los expertos denominan un dendrograma, o diagrama de árbol. En este dendrograma se advierte que la primera separación se pro-dujo entre los gibones y el resto; a continuación, se dio otra separación entre el orangután y el resto; posteriormente, un antepasado común a los chimpancés y a los homínidos se separó del gorila. La separación final se realizó, por tanto, entre los chimpancés y la especie que dio origen a los diversos homínidos22.
Los primeros hominoideos conocidos pertenecían al género Pro-consul23, cuyos restos fueron descubiertos en Uganda y Kenia, y cuya antigüedad se calcula entre 22 y 17 millones de años (época del Mioce-no Inferior). Dentro de este género se sitúan varias especies: Proconsul africanus y Rangwapithecus gordoni. La primera es la mejor conocida y de la que se tienen más datos. En el Mioceno Medio se produce la pri-mera salida de África de los hominoideos, hace aproximadamente unos trece millones de años, a través de los driopithecos, dentro de los cuales se sitúan tres ramas: los Afropithecini, Keniapithecini y Dryopithecini.Y ya en el Mioceno Superior se produjo un amplio abanico de especies de hominoideos, entre los cuales cabe señalar, en Europa, Dryopithecus,Oreopithecus24 y Ouranopithecus, así como Ankarapithecus, en Turquía, Sivapithecus/Ramapithecus, en Pakistán, Lufengpithecus, en China, y Otavipithecus, en Namibia (África).
20 Cfr. Arsuaga, J. L., o.c., pp. 31-32. Cfr. Cela Conde, C. J./Ayala, F. J., o.c., p. 87. 21 Cfr. Cela Conde, C. J./Ayala, F. J., o.c., p. 87. 22 De Bonis, Louis, «Nuestros antepasados más remotos», Investigación y Ciencia,
1999, Temas n.º 19, 4-7. 23 Cfr. Cela Conde, C. J./Ayala, F. J., o.c., pp. 91 y ss. 24 El Oreopithecus bambolii, mono europeo de hace 8 millones de años, fue descubier-
to por M. Köhler, S. Moyá, Jordi Agustí e Isabel Pellejero, quienes consideran que camina-ba completamente erguido, adelantándose de este modo la fecha del inicio de este modo de caminar. Cfr. El País, 15 de octubre de 1997, p. 28.

145
© Universidad de Deusto - ISBN 978-84-9830-383-4
Se solía considerar hasta hace no muchos años que el género Aegypto-pithecus (descubierto en el yacimiento de Fayum, Egipto), era el eslabón entre los chimpancés y los primeros hominoideos. Pero desde los años ochenta se ha desechado esta tesis, considerándose que Aegyptopithecus puede ser un ancestro común de los hominoideos y de los cercopitecos(monos del viejo mundo), con una antigüedad de unos treinta o treinta y dos millones de años. La separación entre los grandes simios y los homi-noideos se produjo en una fecha que hay que situar entre 14 y 11 millones de años, en el momento en el que se produjo un enfriamiento del planeta que contribuyó a una importante degradación de los bosques tropicales en el África oriental. Ahora bien, el paso entre los hominoideos y los prime-ros homínidos sigue siendo un enigma, pudiéndose afirmar algunas hipó-tesis. Es posible que los Ouranopithecus, que vivieron hace entre 9 y 10 millones de años, fueran los antecesores comunes de los grandes simios africanos y el ser humano. Entre hace 7 y 8 millones de años se produciría la separación de los gorilas, por un lado, y la línea de los chimpancés y los humanos, por otro. Y en un tercer momento se produjo la separación pos-terior, hace unos 6 millones de años, de los chimpancés y los homínidos25.
b) Los homínidos26
La separación entre los grandes simios y los homínidos se produjo hace unos 8 millones de años (algunos alargan la distancia y la sitúan en-tre 10 y 8 millones). Pero, ¿a qué se debió esta separación? Todo parece apuntar a la confluencia de un enfriamiento del planeta y una serie de mo-vimientos tectónicos que originaron, entre otros efectos, la formación y hundimiento del valle del Rif, falla que atraviesa el continente africano de norte a sur, en su zona oriental. Como consecuencia de ello, al oeste de la falla (entre el Rif y el Atlántico) las lluvias permanecieron con igual intensidad, pero no así en la parte este, donde las lluvias se dieron de for-ma discontinua, deteriorándose la capa forestal27. Al variar las condicio-nes ecológicas, se produce también un reacomodo de los seres vivos que predominan en ambas zonas. En la zona occidental de África siguieron habitando nuestros antepasados póngidos, adaptados a una vida arbórea, desplazándose entre las ramas y no necesitando utilizar la postura bípeda.
25 Cfr. Cela Conde, C. J./Ayala, F. J., o.c., p. 140. 26 Cfr. Cela Conde, C. J./Ayala, F. J., o.c., cap. 4.º, «La familia de los homínidos»,
pp. 137 y ss. 27 Cfr. para estos datos, Coppens, Yves, «Somos africanos y tenemos 7 millones de
años», El País, 19 diciembre de 1991, pp. 60-61.

146
© Universidad de Deusto - ISBN 978-84-9830-383-4
En cambio, en el este, el clareamiento progresivo del bosque, por el des-censo de la pluviosidad, fue facilitando la presencia de un nuevo tipo de especies, los homínidos (ardipithecos y australopithecos) irían adoptando progresivamente la postura erguida para adentrarse en un terreno de saba-na, no arbóreo. Esto es lo que llama Y. Coppens la East Side Story28. Estas conclusiones quedan avaladas por el hecho de que en la zona oriental del Rif apenas se han encontrado restos fósiles de póngidos, correspondientes a los 8 últimos millones de años, mientras que en la zona occidental están apareciendo innumerables de ellos atribuidos a las diferentes especies de los australopitecos y de los humanos29.
En esta estrecha franja de tierra es donde aparecen los primeros an-tepasados del hombre actual, los llamados homínidos. Pero en la familia de los homínidos no se encuentra sólo la estirpe humana (con las diversas especies o subespecies homo), sino que se halla compuesta por una serie de especies intermedias, que se ha denominado, hasta hace no mucho, de una forma genérica con el nombre de Australopithecos. Ahora bien, desde hace algunos años, debido a varios descubrimientos paleoantropológicos importantes, el mapa clasificatorio ha cambiado y se ha complejificado recomponiéndose de la forma que hemos presentado en el cuadro anterior. Es decir, entre los australopitecos y los chimpancés hay que situar a los Orrogin tugeniensis y a las diferentes especies de los Ardipithecus.
De este modo, aunque no todos los especialistas aceptan como sufi-cientemente probada esta nueva clasificación, la familia de los homínidosse inició con la aparición de los orrorin, para dar paso posteriormente a los ardipithecus y luego a los australopithecus. Cuando se clasifican bajo un mismo nombre diferentes especies o familias, se da por hecho que tie-ne que haber alguna característica común que los acerca y define. En este caso, la supuesta característica común de los homínidos sería su capacidad para caminar erguidos, esto es, el bipedismo30. Pero, como es lógico, esta
28 Cfr. o.c., 60.29 «En un cuarto de siglo de intensivas investigaciones, dice Y. Coppens, en África
oriental, entre los doscientos mil restos de vertebrados fósiles de los últimos 8 millones de años, no hemos encontrado nunca el más mínimo fragmento de hueso o diente de gori-la o pregorila, de chimpancé o prechimpancé, mientras que reunimos casi 2.000 de huesos de homínidos, prehumanos y humanos. En 1983, propuse, por tanto, un modelo que tiene en cuenta todos esos datos y que puede formularse así: los chimpancés, gorilas y hombres son primos, y, si jamás se los ha encontrado juntos, es simplemente porque nunca lo han estado. Su antepasado común probablemente había vivido allí, en un paisaje de bosques y sabanas arboladas, que se extendía hace aproximadamente 10 millones de años, por todo África ecuatorial, desde el Atlántico hasta el Océano Indico», o.c., 60.
30 Cfr. Leaky, Meave/Walker, Alan, «Fósiles de antiguos homínidos africanos», Inves-tigación y Ciencia, 1999, Temas n.º 19, 22-27; Arsuaga, J. L., La especie elegida, Madrid, Temas de Hoy, 1998, cap. 5.º, «El chimpancé bípedo».

147
© Universidad de Deusto - ISBN 978-84-9830-383-4
capacidad no se poseería desde el principio, sino que se empezó a utilizar de modo muy incipiente y provisional al inicio, para irse perfeccionando posteriormente. Pero se trata de una propuesta no demostrada en su totali-dad, en la medida en que no se poseen suficientes evidencias fósiles en to-das las especies homínidas como para afirmar con toda seguridad que sea un rasgo compartido por todas las especies. En su momento nos centrare-mos en las características del bipedismo de los homínidos y las diferentes teorías que pretenden explicar su origen, sus ventajas y el momento en que se inició. Por el momento nos vamos a centrar en indicar las caracte-rísticas de cada una de estas subfamilias.
b.1) Los Orrorin tugeniensis
Serían los primeros homínidos, desgajados de los chimpancés. Has-ta finales del año 2000, los restos fósiles de los homínidos más antiguos eran de 4,4 millones de años. Pero ese mismo año se descubrieron unos fósiles de mayor antigüedad que se denominaron el hombre del milenio,y sus descubridores, un equipo franco-keniata bajo la dirección de Brigitte Senut y Martin Pickford, lo denominó Orrorin Tugeniensis31. El 25 de oc-tubre de 2000, el keniata Cheboi Kiptalam descubrió un fragmento de mandíbula con algunos dientes, y posteriormente se descubrieron algu-nos dientes más junto con otros huesos. En total se calcula que pertene-cían a cinco individuos diferentes, cuatro adultos y un niño. Se calcula que podrían tener una estatura de 1,4 metros, y podrían caminar ergui-dos, dado sus potentes fémures y el importante desarrollo de la apófisis del glúteo (lugar donde se insertan los músculos en el hueso), aunque su tipo de vida estaría todavía muy ligada a un estilo de vida fundamental-mente arborícola32.
El nombre de orrorin significa hombre primitivo, en el idioma de la región keniana donde se descubrió; y tugeniensis se debe a haberse en-contrado en un lugar de los montes Tugen, situados en la región central de Kenia. Según los análisis de biología molecular realizados sobre los restos fósiles, la antigüedad que se puede atribuir a estos restos se sitúa entre 5 y 7-8 millones de años. Por otro lado, por algunas características de su es-tructura fenotípica, los restos de los orrorin tugeniensis se parecerían más al hombre moderno que los de Lucy, que vivió tres millones de años des-pués. Por ello, los especialistas consideran que quedan pendientes muchos
31 Cfr. Senut, B., et al., «First hominid from the Miocene (Lukeino Formation, Kenia)», C. R. Acad. Sci., 2001, 332: 137-144.
32 Cfr. Baur, M./Ziegler, G., La aventura del hombre. Todo empezó en África, Barcelo-na, Maeva, 2003, pp. 27-29.

148
© Universidad de Deusto - ISBN 978-84-9830-383-4
interrogantes, por lo que futuros descubrimientos llevarán a confirmar las suposiciones actuales o quizás tengan que corregirlas.
b.2) Los Ardipithecos
Pero no sólo los Orrorin se sitúan en la actualidad entre los grandes si-mios y los australopitecos, sino también otra subfamilia denominada Ardipi-thecos. Se han descubierto hasta el presente restos correspondientes a dos es-pecies de Ardipithecos: el Ardipithecus ramidus y el Ardipithecus kadabba.
1. Ardipithecus kadabba
Fue descubierto, al igual que el Ardipithecus ramidus, por el equipo de T. White y Yohannes Haile-Selassie, y presentado en marzo de 2004 en la revista Science33. Los restos fósiles que acreditan la existencia de esta especie son seis dientes, a los que se atribuye una antigüedad entre 5,2 y 5,8 millones de años, y que fueron descubiertos en la región de Asa Koma (Etiopía). Se trata de unos dientes caninos que presentan unos rasgos muy primitivos (grandes y muy afilados), y que los haría casi más cercanos a los monos que a los antiguos ancestros humanos. Como puede verse, de-fender la consistencia de esta nueva especie con el descubrimiento de tan sólo estas piezas dentales resulta un tanto arriesgado, si no fuera por el prestigio de las autoridades científicas que lo proponen.
Esta supuesta especie posee una antigüedad mayor que el Ardipithe-cus ramidus, y juntos formarían el género o subfamilia de los Ardipithecus, propuesta que está muy lejos de ser aceptada por todos y que necesitará ser confirmada por posteriores descubrimientos fósiles para poder lograr un consenso más generalizado, que en la actualidad está muy lejos de conseguir.
2. Ardipithecus ramidus
Fue descubierto por Tim White, en una zona de Etiopía, y se le atribu-ye una antigüedad de cuatro millones y medio de años. El descubrimiento se produjo en 1992, pero White y su equipo no dio a conocer los resul-tados hasta 199434. Al parecer, White descubrió numerosos fragmentos,
33 Cfr. Begun, David R., «The Earliest Hominist. Ist Less More?», Sciencie, vol. 303, n.º 5663, 5 de marzo de 2004, pp. 1478-1480.
34 Cfr. White, Tim D. /Suwa, Gen/ Asfaw, Berhane, «Australopithecus ramidus, a new species of early hominid from Aramis, Ethiopia», Nature, 371 (1994), n.º 6.495, 13 de sept., 306-312; cfr. también pp. 313-333, y 280-281.

149
© Universidad de Deusto - ISBN 978-84-9830-383-4
unos diecisiete, de esta nueva especie, y todavía sigue estudiando sus ca-racterísticas, antes de hacer públicos la conclusiones que mostrarían un in-forme amplio sobre las características de esta nueva especie. En la medi-da en que no se tiene evidencia fósil del esqueleto, no se podía saber con exactitud cuál era su forma de locomoción. Pudiera ser que representara el primer eslabón entre la locomoción cuadrúpeda y la bipédica, o también es posible que todavía no se hubiera adaptado al bipedismo.
Se trataría de una especie acostumbrada a vivir en la selva lluviosa (pluvisilva), al estilo de los gorilas y chimpancés actuales. «Por el tipo de dentición, señala J. L. Arsuaga, se ha deducido que se alimentaba de los mismos vegetales que los chimpancés, es decir, frutas, brotes, tallos tier-nos y hojas frescas»35.
Pero el Ardipithecus ramidus poseyó una característica que lo acerca a los humanos actuales, alejándolo de los chimpancés, y consiste en poseer unos caninos reducidos, signo de que su alimentación se orientaba del ré-gimen vegetariano al carnívoro. «Por este único rasgo, advierte Arsuaga, se sitúa a la especie Ardipithecus ramidus entre los homínidos», separado ya de los póngidos. De todos modos, es interesante advertir que, aunque en el informe para la revista Nature T. White colocó a esta especie entre los Australopithecus, posteriormente la denominó Ardipithecus, para sig-nificar su posición intermedia entre los Australopithecus y los chimpan-cés. Lo que parece de momento razonable afirmar es que «el Ardipithecus ramidus era un homínido muy antiguo y primitivo del este de África, y que nosotros procedemos de esta especie, o de una especie muy similar, que vivió hace unos cuatro millones y medio de años en África, proba-blemente en su región oriental»36. Pero hay muchos expertos que mues-tran claramente sus dudas a la hora de incluir o no estos restos entre los homínidos37.
b.3) Los Australopithecos38
La característica general que distingue a los australopithecos, al igual que todos los homínidos, respecto a los chimpancés es que aquéllos em-piezan a caminar erguidos de forma permanente. Pero en realidad se tra-ta de una afirmación excesivamente global y no del todo probada, puesto que no se sabe, como veremos enseguida, si los australopithecos más an-tiguos dominaban el bipedismo de forma permanente. Estas especies apa-
35 Arsuaga, J. L., o.c., p. 36.36 Arsuaga, J. L., o.c., p., 38. 37 Cfr. Cela Conde, C. J./Ayala, F. J., O.C., pp. 138 y 142. 38 Cfr. White, T. D., «Los australopitecinos», Mundo Científico, 1983, n.º 21, p. 18 y ss.

150
© Universidad de Deusto - ISBN 978-84-9830-383-4
recieron hace aproximadamente cinco millones de años, produciéndose su desaparición un millón de años después de la aparición del primer hom-bre. De modo que varios australopithecos convivieron simultáneamente con algunas especies humanas (homo habilis y homo erectus).
El primer descubrimiento de los australopithecos data del hallazgo por Raymond Dart del cráneo del denominado Niño de Taung, en 1924. El nombre se debe a su condición morfológica entre el hombre y el mono, hallados en primer lugar en la zona austral (en concreto, en Áfri-ca del Sur)39. Un momento fundamental en la configuración de los aus-tralopithecos fue el hallazgo del Australopithecus affarensis, por Donald Johanson y sus colaboradores, en la zona del Afar, Etiopía40. Pero la dé-cada de los noventa va a representar una época gloriosa para el avance en el esclarecimiento de las diversas especies de los australopithecos. De tal modo que, en la actualidad, los avances paleoantropológicos consideran que este grupo estaría formado por las especies que vamos a presentar a continuación.
En un principio se situó entre los australopitecos a la especie que lue-go su descubridor, T. White, ha redefinido como Ardipithecus ramidus.Pero en la actualidad se considera el género Australopithecus configura-da por cuatro especies: A. anamensis, A. afarensis, A. africanus y A. garhi.
1. Australopithecus anamensis41
Los restos fósiles que se tienen de esta especie fueron descubiertos por Meave Leakey, esposa de Richard Leakey, en Kanapoi (Kenia) y Allia Bay, a pocos kilómetros del lago Turkana, lugar donde han sido encontra-dos otros muchos e importantes restos fósiles. Esta especie habría vivido hace unos 4,1 millones de años (entre 4,17 y 4,07 millones de años).
Sus rasgos específicos estarían situados en su dentición y en el modo de andar. Respecto a lo primero, sus muelas son más grandes y con esmal-te más grueso que el A. ramidus, prueba de que su alimentación estaba compuesta, además de frutos tiernos, de vegetales (granos y frutos secos, además de bulbos, tubérculos, raíces, etc.) que necesitaban una mastica-ción bastante prolongada, además de llevar con ellos algunas partículas
39 Cfr. Lewin, R., La interpretación de los fósiles, Barcelona, Planeta, 1987; Johan-son, Donald/Edey, Maitland, El primer antepasado del hombre, Barcelona, Planeta, 1987 (3.ª ed.); Bacon, Anne-Marie, «Los australopitecos», Investigación y Ciencia, 1999, Temas n.º 19, 16-22.
40 Cfr. Lewin, R., o.c.; Johanson, D./Edey, M., o.c.; Cela Conde, C. J./Ayala, Fr. J., Sen-deros de la evolución, Madrid, Alianza, 2001, cap. 4.º.
41 Cfr. Leaky, Meave y Walter, Alan, «Antiguos fósiles de homínidos en África», Inves-tigación y Ciencia, 1997, n 251, 70-75.

151
© Universidad de Deusto - ISBN 978-84-9830-383-4
minerales que producían, como consecuencia, un fuerte desgaste de las muelas. En la medida en que estos productos alimenticios se hallan en bosques más secos que los que habitaba el A. ramidus, se piensa que el habitat del A. anamensis había variado sensiblemente.
En relación a su modo de caminar, el análisis de la tibia casi completa que se ha hallado nos permite afirmar que esta especie caminaba de forma bipédica de modo continuo.
2. Australopithecus afarensis
La definición y descripción de esta especie se debe sobre todo a Do-nald Johanson, el descubridor de los restos más importantes y significati-vos de la misma42. Los restos se hallaron en Tanzania y Etiopía, en la tierra de los Afar (de ahí la denominación de «afarensis»). La abundancia de fó-siles encontrados nos permiten una mayor información sobre la especie.
Por sus características dentarias se deduce que su alimentación era exclusivamente vegetariana, propia de bosque seco. Su postura era clara-mente erguida, pero seguía todavía adaptado a poder trepar y vivir en los árboles, aspecto deducido de la largura de sus brazos en comparación con las piernas. Su tamaño era más bien pequeño, comparado con nosotros, y no mucho mayor que los chimpancés actuales. Los machos llegaban a me-dir alrededor de 135 cms. de altura con unos 45 kgs. de peso; y las hem-bras, 105 cms. de altura, con unos 30 kgs. de peso. Puede verse, por tanto, que el dimorfirmo sexual (diferencia de tamaño entre ambos sexos) era bastante acentuado.
Los dos más importantes fósiles hallados de esta especie son un es-queleto femenino casi completo, que Johanson, su descubridor, denominó Lucy, y un cráneo masculino también casi entero, con una capacidad cra-neana estimada en 500 cc. Otros cráneos apuntan a una capacidad cranea-na inferior a ésta. Vemos con ello que su cerebro es sólo un poco mayor que el de los chimpancés, que se sitúa cercano a los 400 cc.
La antigüedad de esta especie se sitúa entre 3 y 4 millones de años antes de nosotros, que vivió en el centro de África, no sólo en la zona del noreste, sino también en zonas de más al centro, como El Chad, donde, como señala J. L. Arsuaga, «recientemente el paleontólogo francés Michel Brunet ha encontrado restos de Australopithecus de esta misma edad»43.Ningún fósil por sí mismo nos puede responder a la cuestión de si es un antepasado nuestro o no, pero parece bastante probable. De todos modos,
42 Cfr. Johanson, D./Edey, M., o.c. 43 O.c., p. 40.

152
© Universidad de Deusto - ISBN 978-84-9830-383-4
no todos los paleoantropológos están de acuerdo en situarlo en la línea directa de nuestra evolución. Mientras su descubridor, Donald Johanson, y una gran mayoría de paleoantropólogos lo consideran como probable, otros lo sitúan más bien en un lugar colateral, como una rama paralela a los homínidos, con un mismo antepasado común, el ya estudiado Austra-lopithecus anamensis.44 Necesitamos todavía más datos fósiles para dejar zanjada la cuestión. Pero está imponiéndose de modo más amplio la tesis de situar al Australopithecus afarensis en la línea de ascendencia de las diversas especies o subespecies homo.
3. Australopithecus africanus45
La antigüedad de los fósiles encontrados de esta especie se sitúa entre los 3 y 2,5 millones de años, descubiertos en diferentes puntos de Sudáfri-ca. Su parecido físico con los A. afarensis es importante, y no parecen te-ner un cerebro mucho más grande, puesto que se calcula que se sitúa entre 500 y 530 cc.
El descubrimiento más reciente (1998) lo realizó el conocido pa-leoantropólogo Philip Tobias, junto con su colaborador Ron Clarke, con-sistente en un esqueleto casi completo, que podría tener una antigüedad de 3,5 millones de años. Las investigaciones para estudiar las caracterís-ticas de estos restos fósiles todavía están en plena marcha, por lo que ha-brá que esperar a las conclusiones definitivas. Pero hay que señalar que se tratarán de conclusiones de gran importancia, en la medida en que, como señala J. L. Arsuaga, «si el esqueleto de verdad es contemporáneo del Australopithecus afarensis, y de una especie distinta (tal vez una forma antigua de Australopithecus africanus), podría competir seriamente por el puesto de antecesor de todos los homínidos posteriores (es decir, con me-nos de 3 millones de años); entre ellos nos encontramos nosotros»46.
Uno de los rasgos para considerar al Australopithecus africanus espe-cie diferente del Australopithecus afarensis es su dentición, formada por muelas más grandes, propias de una alimentación necesitada de una masti-cación más prolongada sobre productos más duros que los que ingerían sus antecesores. De modo que además de frutos tiernos y hojas tiernas, tenían como alimentación raíces y semillas de diversas hierbas de la sabana.
44 Cfr. Arsuaga, J. L., El collar del neandertal, o.c., p. 78. Otros muchos autores, de la talla de Ph. Tobias, Y. Coppens y R. Leakey, dudan también de que al Australopithecusaffarensis haya que situarlo en la línea directa del origen del linaje humano.
45 Cfr. Cela Conde, C. J./Ayala, Fr. J., Senderos de la evolución humana, o.c., pp. 214 y ss.
46 El collar del neandertal, o.c., p. 42.

153
© Universidad de Deusto - ISBN 978-84-9830-383-4
4. Australopithecus garhi
El acontecimiento más reciente dentro de la recomposición de la fa-milia de los australopitecos lo compone el anuncio de Tim White (Univer-sidad de Carlifornia, Berkeley) y de Berhane Asfaw, etíope, de haber des-cubierto restos fósiles (seis homínidos) de una nueva especie, denominada por él y su equipo de investigadores como Australopithecus garhi47. El lugar del descubrimiento es otra vez la región del Medio Awash (Etiopía), un desierto inhóspito donde vive el pueblo Afar, lugar donde fueron des-cubiertos los restos fósiles más importantes del A. afarensis. Su ubicación cronológica se sitúa entre el Australopithecus afarensis y el Homo habilis,con una antigüedad de 2,5 millones de años.
La altura media de esta especie sería de menos de metro y medio, y su cerebro alcanzaría un tercio de nuestra capacidad. Los rasgos morfológi-cos son una rara combinación (de ahí el nombre impuesto, gahri: sorpre-sa, en la lengua de los Afar) de rasgos simiescos (brazos largos) con otros más humanos (piernas largas y dentición cercana a la humana). Otro ele-mento de gran valor fue el hecho de haber descubierto junto a estos restos del llamado Australopithecus gahri, otros restos fósiles de animales con indicios de haber sido descarnados con instrumentos de piedra, así como huesos machacados para absorber sus tuétanos. Estos indicios vendrían a confirmar una cierta utilización de herramientas por individuos prehuma-nos. O dicho de modo complementario, nos llevaría a discutir si no nos hallamos ante una especie que, dada la utilización de instrumentos inte-ligentes, habría que situar en la línea homo. De hecho, este punto vendría a confirmar el dato de las herramientas encontradas no mucho antes en Gona (Awash Medio) de piedras talladas con una antigüedad calculada de 2,6 millones de años, es decir, anterior a los restos que hoy día conocemos del Homo habilis, supuesto primer hombre. Este dato sobre las herramien-tas indica también que el Australopithecus gahri poseería un método de aprovecharse de alimentos como la carne y el tuétano, que le proporciona-ría una base alimenticia más rica, base de una mejor adaptación al entor-no ecológico en que vivían, y la posibilidad de extender el espacio de su existencia.
Pero la capacidad craneana de esta nueva especie lo acerca mucho al Australopithecus afarensis. «El rostro se proyecta hacia adelante, dice T. White, la caja craneana es pequeña y con cresta, pero los premolares y los molares son enormes. Nunca se había visto esta combinación de ras-gos, y por eso hemos nombrado una nueva especie»48. Con los datos que
47 Cfr. Science, 23 de abril de 1999, vol. 284, n.º 5414, pp. 545-700. 48 Cfr. Rivera, Alicia, reportaje en El País, 23-abril-1999, p. 30.

154
© Universidad de Deusto - ISBN 978-84-9830-383-4
se poseen, no se sabe si nos hallamos ante el antepasado más cercano del hombre, aunque T. White lo cree muy posible. Vemos, pues, que nos ha-llamos ante avances importantes en el conocimiento de nuestros orígenes, así como ante interrogantes y problemas que, de momento, la ciencia no puede solucionar.
Podemos ver, por tanto, que el momento en que la línea Homo se se-para de los Australopithecos no está del todo aclarada, disputándose va-rias supuestas especies el puesto de último eslabón o antepasado de nues-tra línea evolutiva. Lo que sí es evidente es que no fuimos los humanos los únicos descendientes y continuadores de la línea de los australopithe-cos, puesto que hubo algunas especies contemporáneas al H. habilis y al Homo erectus. A estas especies se les ha llamado Paránthropos.
b.4) Los Paranthropos
Se trata de homínidos con una morfología muy similar a los Australo-pithecos, e incluso al Homo habilis. Pero la diferencia más notable se ha-lla en su aparato masticador, que se fue especializando en «productos ve-getales duros y fibrosos, muy abrasivos»49.
El grupo de los paranthropos constituye una línea paralela a los hu-manos, separada de ella hace unos 2,8 millones de años, representando, como señala Arsuaga, «una respuesta al mismo cambio climático de hace 2,8 millones de años que propició el origen del género Homo»50.
Dentro de los paranthropos se conocen tres especies: el Paranthropus aethiopicus, que habitó el este de África hace unos 2,6 millones de años; el Paranthropus boisei, posterior al aethiopicus y habitante de la misma región que él; y el Paranthropus robustus, que habitó las tierras sudafri-canas. Todas estas especies desaparecieron hace un millón de años; al me-nos, no se han encontrado restos fósiles en fechas posteriores.
Aunque hemos dicho que los Paranthropos son una línea paralela a los humanos, compartiendo con nosotros un antepasado común aus-tralopitheco, no está claro cuál sería esa especie en la que entroncarlos. «La mayoría de los autores la entroncan a la altura de los Australopithe-cus afarensis, que sería un antepasado común de los parántropos y de los humanos»51. Pero pudiera ser ese antepasado común el Australopithecusanamensis, especie, por lo demás, muy parecida al Australopithecus afa-rensis. Sólo investigaciones posteriores más detalladas podrán sacarnos de estas dudas.
49 Arsuaga, J. L., El collar del neandertal, o.c., p. 65.50 Ibídem, p. 65.51 O.c., 65-66.

155
© Universidad de Deusto - ISBN 978-84-9830-383-4
De todos modos, el esquema o árbol genealógico representativo de la distribución que hemos seguido aquí, estaría configurado por la propuesta de Tim White, en su trabajo de presentación del Australopithcus garhi52.Igualmente, es interesante contrastarlo con el esquema de J. L. Arsuaga, en El collar del Neanderthal53.
c) Los humanos
Así como los paleoclimatólogos (estudiosos de las variaciones y evo-luciones del clima terráqueo en las épocas antiguas) consideran los movi-mientos tectónicos como el hecho fundamental que dio origen a la agudi-zación de la falla del Rif y que llevó a la bifurcación o separación entre los póngidos y los homínidos, el fenómeno que con toda probabilidad parece explicar la separación entre los australopithecos y los humanos es el fuerte enfriamiento que sufrió toda la Tierra hace unos 3 millones de años54. Este enfriamiento duró unos 900.000 años. Fue una de las varias glaciaciones que se produjeron en la historia de nuestro planeta. La conse-cuencia de ello fue la transformación de nuevo del entorno ecológico. «Se produjo una evolución, según Y. Coppens, de una sabana muy frondosa y terrenos forestales con grandes árboles tropicales (antrocaryon) que exis-tían desde hace 3 millones de años, a una sabana llena de claros con sólo algunas zonas arboladas (ficus, myrianthoxylon) a lo largo de los ríos, hace unos 2 millones de años»55.
Ante este cambio climático y ecológico, las especies de australopithe-cos existentes adoptaron una doble estrategia, correspondiente a las dos ramas que les sucedieron: la línea prehumana, que estaba constituida por los Parántropos, y que siguió viviendo hasta cerca de un millón de años antes de la fecha actual; y la línea humana, de la que somos los seres hu-manos actuales los únicos supervivientes. Y lo que nos dio ventaja en el
52 Cfr. Science, 23 de abril de 1999, vol. 284, n.º 5414, págs. 545-700. 53 O.c., p. 78. 54 Para la influencia de la climatología en el origen y evolución de las especies, y tam-
bién en el origen de la especie humana, cfr. Gribbin, J. y M., Hijos del hielo. Clima y orí-genes del hombre, o.c.; Erickson, J., La extinción de las especies. Evolución, causas y efec-tos, o.c.
55 O.c., p. 61. El modo de mostrar la objetividad de estas afirmaciones consiste en analizar los porcentajes de polen fosilizado de los diferentes vegetales hallados entre los terrenos excavados. «La relación entre la cantidad de polen de árboles y la de polen de hierbas puede considerarse un importante punto de referencia: ¡dicha relación fue de 0,4 hace 3 millones de años, y alcanzó el 0,01 hace 2 millones de años!», Y. Coppens,o.c., p. 61.

156
© Universidad de Deusto - ISBN 978-84-9830-383-4
proceso selectivo fue el extraordinario aumento del cerebro y todos los frutos culturales que originó.
Entre las diferentes grupos o especies de la línea homo, se solían con-siderar, casi en línea recta de sucesión, el homo habilis, el homo erectus, el homo sapiens (hombre de Neanderthal) y el homo sapiens sapiens (hom-bre de Cro-Magnon). Los avances de la paleoantropología en los últimos años ha puesto en crisis esta clasificación, introduciendo serias noveda-des, como vamos a ver. Sólo nos centraremos en los aspectos imprescindi-bles para el objetivo que nos hemos propuesto aquí.
c.1) El homo habilis
Se le considera el primer grupo poblacional que inicia el linaje (mejor que especie) de los humanos56. Sus características específicas, por las que hay que considerarlo homo, y la primera especie o subespecie humana, son las herramientas que utilizaba, la dimensión de su cerebro, y la loco-moción bípeda de forma permanente.
Esta tercera característica no se inicia, como ya hemos visto en el homo habilis, sino en muchos de los australopithecos, y es posible que en todos (faltan en algunos las evidencias fósiles). Pero en el homo habi-lis esta locomoción se perfeccionó, dándole la posibilidad de usar las ma-nos para múltiples finalidades, sobre todo para el uso y construcción de herramientas57. Su tamaño era más bien pequeño (poco más de un metro de altura), con largas manos y piernas cortas, por lo que todavía poseería una cierta facilidad para trepar a los árboles y vivir en la frontera entre el bosque y la sabana, al igual que sus antecesores los australopithecos. Por eso señala Arsuaga que «desde el punto de vista de la morfología no hay muchos motivos para aceptar el Homo habilis en nuestro propio género y sería más gráfico denominarlo “Australopithecus habilis”: así el lector sa-bría mejor de qué clase de homínido estamos hablando. Sin embargo, no
56 Estamos utilizando repetidas veces el concepto de especie sin haberlo definido con antelación. Los especialistas no tienen un concepto claro, a priori, de especie, sino más bien a posteriori: forman especies diferentes aquellos grupos cuyos apareamientos sexuales no producen descendencia, o una descendencia no fértil. Pero en el caso de gru-pos poblacionales anteriores a nosotros no podemos demostrar si se da esa posibilidad o no, por lo que hablar dentro de la línea homo (al igual que dentro de los australopithcos) de especies distintas, es pura hipótesis. Tenemos que hablar, por tanto, tanto de especiescomo de subespecies. Por eso, en muchos momentos nos referiremos más bien al linaje o género humano.
57 Veremos más adelante la implicación del bipedismo en el uso inteligente de la mano, pero para ver la específica configuración del aparato locomotor en los humanos. Cfr. Ar-suaga, J. L., La especie elegida, Madrid, Temas de Hoy, 1998, cap. 5.º.

157
© Universidad de Deusto - ISBN 978-84-9830-383-4
está claro que su mente fuera la de un australopiteco, la mente de un mono en definitiva»58.
Efectivamente, el cerebro del homo habilis posee un volumen medio de alrededor de 630 cc., sensiblemente mayor que el de los australopithe-cos59. Eso le daba una importante ventaja en el proceso de adaptación al ambiente y en la lucha selectiva. Los restos fósiles encontrados (los más antiguos se deben a L. Leakey y algunos otros componentes de su misma familia: su mujer, Mary y su hijo Richard) le conceden una antigüedad entre 2,3 y 1,5 millones, y el lugar de su hallazgo se extiende dentro de una región comprendida entre Etiopía (valle del río Omo y Hadar), Kenia (lago Turkana) y Tanzania (quebrada de Olduvai). Esta referencia geográ-fica (así como la cronológica) es la que hace más plausible considerar al Australopithecus gahri como el antecesor del homo habilis.
Como hemos señalado, la mayor amplitud de cerebro le permitió el uso y la construcción de herramientas, elemento fundamental para distin-guirlo de las especies anteriores. Precisamente la razón de denominarlo homo habilis está en el descubrimiento junto a sus restos fósiles de gui-jarros retocados, constituyendo el primer invento y los primeros indicios de la cultura humana. Volvemos a repetir que no hay total evidencia de que las herramientas, en este caso los artefactos líticos (de piedra), no sean más que inventos humanos, puesto que parece seguro que un cierto tipo de herramientas habrían sido utilizados por algunas de las especies de australopitecos60.
Pero hay que distinguir claramente entre herramientas consistentes en objetos no modificados (como ramas, huesos, piedras) y objetos modifica-dos para que puedan ser utilizados mejor para una finalidad determinada. Sólo en este segundo caso parece razonable atribuir en los sujetos de tales transformaciones una inteligencia que pueda considerarse como humana. Y ese sería el paso que habría dado el homo habilis respecto a las espe-cies anteriores. La paleontología ha registrado como herramientas más antiguas las encontradas en Gona, en la región del Hadar (tierra del Affar, Etiopía), atribuyéndoles una antigüedad de hasta 2,5 millones de años, una cifra superior a los restos fósiles encontrados hasta ahora del homo habilis, que no llegan más que hasta 2,33 millones de años (en concreto,
58 El collar del Neandertal, o.c., 45-46.59 El problema respecto a delimitar el volumen del cerebro del homo habilis está en las
sensibles diferencias existentes entre los diversos cráneos fósiles encontrados, que varían desde 510 cc. hasta 752 cc. Cfr. Arsuaga, J. L., El collar del Neandertal, o.c., p. 46 y ss.
60 Tendremos ocasión de hablar más adelante sobre las herramientas y su origen. Baste decir aquí que hay muchos autores que consideran la herramienta un artefacto prehumano: cfr. Coppens, Y., La rodilla de Lucy. Los primeros pasos hacia la humanidad, Barcelona, Tusquets, 2005.

158
© Universidad de Deusto - ISBN 978-84-9830-383-4
un maxilar con el paladar y unos dientes, descubiertos por D. Johanson hace unos pocos años).
Los expertos suelen clasificar el tipo de herramientas, y de las culturas correspondientes a ellas, en función de la complejidad que suponen. Las herramientas líticas del homo habilis eran muy más elementales, puesto que consistían en cantos de piedra a los que se les había sacado algún filo al golpearlos con otra piedra. Los arqueólogos suelen denominar a esta in-dustria como Olduvayense (en referencia a Olduvai, zona donde se halló por primera vez) o Modo Técnico I61.
c.2) El «grado erectus»
En la clasificación tradicional, tras el homo habilis se situaba el homoerectus, a quien se le atribuía la primera salida de África. Pero posterior-mente se defendía la teoría de que quizás homo erectus sería el nombre bajo el que habría que situar los restos fósiles hallados en el continente asiático. En cambio, a los restos de esa misma antigüedad encontrados en África se los situaba bajo la denominación de homo ergaster. Incluso se consideraba al homo ergaster como un eslabón anterior al homo erectus.No vamos a detenernos en los detalles de la polémica, sino sólo indicar que, siguiendo el planteamiento de Cela Conde y Ayala, vamos a situar a todos los humanos de la época del Pleistoceno bajo la denominación de grado erectus, considerando al homo ergaster como un tipo de homo per-teneciente a tal grado.
El Pleistoceno se extiende entre 1,6 millones de años hasta los 100.000 años antes de nuestra época, y se suele dividir en tres etapas: el Pleistoceno inferior (1,6 m. a.-0,73 m. a.), el Pleistoceno medio (0,73 m. a.-0,13 m. a.) y el Pleistoceno superior (130.000 a.-100.000 a.)62. En el Pleistoceno su-perior existían todavía los paranthropos, mientras que en el Pleistoceno medio parece que se dio una fuerte reducción de las diversas especies ho-mínidas, de tal modo que hacia medio millón de años antes de nosotros ya no quedarían más que una o dos especies. De tal forma que en el Pleisto-ceno medio sólo quedaría el homo erectus, o las diversas especies que se
61 Cfr. Arsuaga, J. L., o.c., p. 51 y ss. «Se ha dicho, comenta Arsuaga, que los instru-mentos (cualesquiera que fuesen) producidos por el Homo habilis eran “instrumentos bio-lógicos”, es decir, que potenciaban o prolongaban la morfología del individuo. Sin embar-go, desde que los caninos empezaron a reducirse en el Ardipithecus ramidus, los homínidos no disponen de buenas herramientas naturales para cortar la piel y la carne de los animales muertos, ni tampoco tienen modo de fracturar los huesos para extraer el tuétano, por lo que los instrumentos de piedra realmente representaron la llave, o una de ellas, para abrir una nueva despensa», o.c., 51-52.
62 Cfr. Cela Conde, C. J./Ayala, F. J., o.c., pp. 294 y ss.

159
© Universidad de Deusto - ISBN 978-84-9830-383-4
encierran dentro del grado erectus. Ya hemos dicho que se ha solido dis-tinguir entre homo ergaster y el homo erectus, encerrándose bajo la pri-mera denominación a los restos fósiles encontrados en el África del Este, considerándolo el sucesor directo del homo habilis, mientras que el homoerectus sería un sucesor del homo ergaster, la primera especie humana emigrada fuera de África y situada en diversas regiones asiáticas. Pero en la actualidad se atribuyen a la denominación de homo erectus restos fó-siles hallados en África, en concreto en Olduvai, denominándose homoerectus olduvaiensis63. De todos modos, presentamos los rasgos y carac-terísticas de ambas especies, aun considerando que puedan agruparse bajo la denominación indicada: grado erectus.
1. Homo ergaster
Como he indicado, tras el Homo habilis se solía situar en la línea evo-lutiva al Homo erectus. Pero los descubrimientos más recientes ponen en entredicho esta relación, y se sitúan en las franjas cronológicas interme-dias una serie de restos fósiles que hacen pensar en la presencia de una es-pecie intermedia denominada Homo ergaster. Los fósiles pertenecientes a esta especie se encontraron en la cuenca del lago Turkana (Kenia), de una antigüedad entre 1,8 y 1,6 millones de años64. Igualmente, hay indicios de la presencia del Homo ergaster en África del Sur (por restos encontrados en Swartkrans), por lo que esta segunda especie humana se habría ya ex-tendido a diferentes partes de África, desde el lugar de origen, el noreste.
Ahora bien, el solapamiento de la datación cronológica entre restos fósiles de Homo habilis y Homo ergaster podría restar fuerza a la tesis de considerar a este segundo como una especie diferente. La verdad es que no es raro que convivan al mismo tiempo y en los mismos lugares dos especies contiguas en el árbol genealógico. Lo lógico es que las especies contiguas cronológicamente se especialicen después en ámbitos geográ-ficos diferentes. Así, según Arsuaga, el Homo habilis se habría situado en la zona del lago Turkana, y el Homo ergaster, en Olduvai. Y progresiva-mente se va imponiendo la especie nueva, desapareciendo poco a poco la anterior.
Lo central es mostrar las diferencias morfológicas y culturales de esta supuesta nueva especie. En el aspecto morfológico, Homo ergaster era
63 Cfr. ibídem, p. 295. 64 Arsuaga hace referencia al descubrimiento reciente, por un grupo italiano de la Uni-
versidad de Florencia, de un cráneo de un millón de años de antigüedad (descubierto en la depresión de Danakil, en Eritrea), que podría pertenecer a esta especie. Cfr. Arsuaga, J. L., El collar del neandertal, o.c., p. 56.

160
© Universidad de Deusto - ISBN 978-84-9830-383-4
bastante diferente al Homo habilis, puesto que poseía una talla muy pa-recida a la nuestra, como lo demuestra el hallazgo, por Richard Leackey, de un esqueleto casi completo en el entorno del lago Turkana. Así mis-mo, la capacidad craneana se había desarrollado bastante, hasta alcanzar unas dimensiones aproximadas entre 800 y 900 cc. Es verdad que estas dimensiones cerebrales tienen que relativizarse, dado que a un cuerpo más grande le corresponde siempre unas dimensiones cerebrales también más amplias, sin suponer con ello más inteligencia. Lo que mostraría la mayor capacidad intelectual del Homo ergaster sería, en cambio, la superior tec-nología manifestada en las herramientas que era capaz de construir y uti-lizar. Se le atribuye un tipo de herramientas que se denomina Achelense(por el lugar donde se hallaron por primera vez, Saint-Acheule, Francia) o Modo Técnico II65.
Con el Homo ergaster es con quien, en opinión de Arsuaga, se ma-nifiesta la especial configuración de los seres humanos, en la medida en que somos «anatómicamente sólo unos primates con postura erguida, una novedad evolutiva interesante pero en ningún sentido más extraordinaria que el vuelo de los murciélagos o la adaptación al mar de los cetáceos; y al mismo tiempo unos organismos radicalmente diferentes de todas las demás criaturas vivientes por su asombrosa inteligencia, capacidad de re-flexión y plena conciencia de sí mismos en todos sus actos»66.
Otra peculiaridad de esta especie se halla en su adaptación a un nue-vo nicho ecológico, en la medida en que habría abandonado totalmente un entorno con cierta densidad arbórea para adentrarse de lleno en entor-nos abiertos. La aridez de los entornos donde vivía había aumentado por un nuevo enfriamiento climático, por lo que la sabana estaba alcanzando mayores ámbitos de extensión. Las fuentes de alimentación eran, además de la vegetal, la procedente de carne de otros animales, no tanto cazados directamente, sino de restos de lo cazado por otras especies depredadoras. Se trata del sistema de carroñeo que ya había sido iniciado por el Homohabilis. Todos estos elementos hacen pensar que el ser humano estaba ya capacitado para extenderse fuera de África.
2. Homo erectus
No todos los paleoantropólogos aceptan la distinción entre el Homoergaster y el Homo erectus, considerando que los fósiles africanos situa-dos dentro de la especie del Homo ergaster no pasarían de ser más que
65 Cfr. Arsuaga, J. L., El collar del neandertal, o.c., pp. 62 y ss. 66 O.c., 64.

161
© Universidad de Deusto - ISBN 978-84-9830-383-4
ejemplares del Homo erectus. El Homo erectus había sido configurado como especie a partir de los fósiles encontrados en el entorno de Asia. Fue E. Dubois, en 1891, quien denominó Pithecanthropus erectus a los restos fósiles que encontró en la isla de Java. La capacidad craneal de es-tos fósiles oscila entre 800 y 1.059 cc., por lo que apenas hay diferencia con las dimensiones del Homo ergaster. «Sin embargo, desde el principio los cráneos asiáticos son más robustos (extremadamente robustos en algu-nos casos) que los africanos, y hay algunas otras diferencias en la base del cráneo que podrían justificar la distinción entre especies»67. La distinción, por tanto, es demasiado débil y lábil, y tendrá que ser probada o rechaza-da a partir de ulteriores avances paleontológicos.
Lo que sí parece un hecho aceptado por todos es que el Homo erectusrepresenta la especie primera que sale de África, presentándose sus res-tos fósiles más antiguos en Java y en diferentes partes de Asia68. Los restos más famosos de Asia son los hallados en la cueva de Zhoukoudian, cer-ca de Pekín69. La capacidad craneana de estos fósiles se sitúa entre 915 y 1.225 cc. Respecto a los utensilios que podía utilizar, no se tiene demasia-da claridad sobre ello. La verdad es que son escasos los artefactos encon-trados junto a los restos humanos fósiles. Y están muy poco elaborados, por lo que se ha pensado que el Homo erectus asiático no llegó a dominar la industria de tipo achelense, bien sea porque no la necesitó, o porque salió de África antes de que se desarrollara allí este nivel de la industria lítica. De todas formas, contra estas explicaciones se levantan ciertos des-cubrimientos de arqueólogos chinos que han encontrado herramientas que pueden considerarse del Modo Técnico II, aunque no lleguen al nivel de la industria achelense (hachas bifaces, talladas por las dos caras)70.
Se ha considerado durante mucho tiempo la posibilidad de que el Homo erectus pudiera llegar a usar y dominar el fuego, atestiguado sobre todo por los restos hallados en Zhoukoudian (Pekín), pero hoy día se duda
67 O.c., p. 68. Señala, no obstante, Arsuaga que «éstas son discusiones entre los espe-cialistas, ya que se trata en los dos casos, fósiles africanos y fósiles asiáticos, esencialmen-te del mismo tipo de homínido», p. 68-69.
68 Descubrimientos más recientes adelantan, de modo significativo, la fecha de salida de África de los primeros humanos. Se trata de dos cráneos encontrados en el yacimiento de Dmanisi (república de Georgia), datados con una antigüedad de 1,7 millones de años. Hasta ahora, los fósiles anteriores más antiguos se consideraban de un millón de años de antigüedad. El descubrimiento lo ha realizado un grupo de científicos georgianos, dirigi-dos por David Lordkipanidze, durante el verano de 1999, y han sido publicados ahora los resultados en la revista Science, del 12 de mayo de 2000. Cfr. Agustí, Jordi/Lordkipanidze, David, Del Turkana al Cáucaso. La evolución de los primeros pobladores de Europa, Bar-celona, RBA Libros, 2005.
69 Cfr. Arsuaga, J. L., o.c., pp. 67-70. 70 Cfr. Cela Conde, C. J./Ayala, F. J., Senderos de la evolución humana, o.c., pp. 327 y ss.

162
© Universidad de Deusto - ISBN 978-84-9830-383-4
seriamente de este dato, en la medida en que este yacimiento chino fue excavado con una tecnología un tanto superada, y sus restos desaparecie-ron durante un oscuro episodio de la guerra entre China y Japón. Las mis-mas dudas se extienden sobre los supuestos restos arqueológicos que ates-tiguarían la existencia de enterramientos y de lugares de culto religioso71.
Cabe preguntarse si se han encontrado restos de homo erectus en Eu-ropa, puesto que tras su salida de África podría haberse extendido tanto por Asia como por Europa. Además está el problema de por dónde salió de África, si por la parte nororiental (Egipto) o también por la norocciden-tal (estrecho de Gibraltar o por Sicilia y sur de Italia)72. Los descubrimien-tos de Atapuerca parecen avalar la tesis de un asentamiento relativamente temprano en Europa del homo ergaster, para dar lugar a la nueva especie que Arsuaga y sus colegas han denominado Homo antecesor.
3. El homo antecessor73
Tras el Homo erectus, en la paleoantropología común se solía situar el hombre de Neanderthal. Pero los descubrimientos de la cueva de Ata-puerca (Burgos), por un grupo de científicos españoles, han dado un giro importante al mapa evolutivo de los humanos. En los yacimientos de la Gran Dolina se han encontrado restos humanos de una antigüedad de 800.000 años o más. Como señala J. L. Arsuaga74, uno de sus descubrido-res, no se trata de restos que puedan situarse dentro del Homo ergaster ni del Homo erectus. Tampoco pertenecen a los neandertales, por lo que se imponía la creación de una nueva especie, por sus rasgos tan especiales y diferentes a las otras especies conocidas.
Podría pensarse que estos restos cabían dentro de la denominación de Homo heidelbergensis, pero esta denominación, piensan los científicos españoles, sirve más bien para nombrar a la especie inmediatamente an-
71 Cfr. Cavalli-Sforza, Luca y Franceso, ¿Quiénes somos? Historia de la diversidad hu-mana, Barcelona, Crítica, 1999, pp. 56-59; Arsuaga, J. L., o.c., pp. 66-70.
72 Cfr., sobre la presencia del «grado erectus» en Europa, Cela Conde, C. J./Ayala, F. J.,o.c., pp. 316 y ss.
73 Arsuaga et alli, Nature, 1994, n.º 362, p. 354; Arsuaga, J. L., Mundo Científico,1993, 143, 167; Id., «Atapuerca y el origen del hombre», Mundo Científico, 1996, n.º 170, p. 601; Id., «Homo Antecessor», Mundo Científico, 1997, n.º 181, pp. 649 y 651; Cervera, J./Arsuaga, J. L./Bermúdez, J. M.ª/Carbonell, E., Atapuerca: Un millón de años de historia,Madrid, Ed. Complutense/Plot Edic., 1999; Díez, C./Moral, S./Navazo, M., La Sierra de Atapuerca. Un viaje a nuestros orígenes, Burgos, Fundación Atapuerca, 2009; «Atapuer-ca», número monográfico de Journal of Human Evolution, 37(1999), n.º 3-4, sept.-oct.
74 Cfr. El collar del neandertal, o.c., pp. 74 y ss.; Id., «Nuestros primeros padres», ElPaís Semanal, 2003, 3 de agosto, 28-35

163
© Universidad de Deusto - ISBN 978-84-9830-383-4
terior a los neandertales. Por tanto, el Homo antecessor se situaría en un momento anterior a la separación de las dos líneas humanas que vinieron después: por un lado los neandertales, con el Homo heidelbergensis por delante, y por otro, los cromañones. Entre las herramientas que utilizaban, no aparecen las hachas bifaces, siendo más bien sus utensilios del tipo que se ha denominado Modo Técnico I, o Pre-Achelense, aunque ya en Áfri-ca se había desarrollado desde mucho antes el Modo Técnico II75.
Los yacimientos de Atapuerca nos indican aspectos interesantes so-bre el comportamiento de estos antecesores nuestros76. Muchos de los restos fósiles «muestran evidencias, dice Arsuaga, de haber sido descar-nados y troceados por otros humanos sin ningún miramiento»77, prueba evidente de usos antropofágicos dentro de esta especie. En las excavacio-nes de la Sierra de Atapuerca (en la Sima de los Huesos), también se han encontrado restos fósiles del Homo heidelbergensis, supuesto antepasado de los neandertales, como ya ha quedado dicho. La denominación de esta especie se debe a los restos encontrados (mandíbula de Mauer) cerca de Heidelberg (Alemania). La industria utilizada es claramente Achelense,Modo II, con abundante presencia de hachas bifaces.
Dentro de los humanos del Pleistoceno, se ha querido situar al deno-minado hombre de Orce, denominación dada a los restos fósiles encon-trados en 1982 en el pueblo de Orce, en la depresión de Guadrix-Baza, dentro de la provincia de Granada (España)78. El equipo de Josep Gibert, su descubridor, lo catalogó como homínido, aunque otros especialistas lo consideran más bien perteneciente a una especie équida. Mientras varios paleoantropólogos atribuyen con claridad la condición de homínidos a estos restos, otros siguen teniendo serias dudas sobre ello. Su antigüedad se situaría alrededor de 1,8 millones de años (aunque hay quienes rebajan esta cifra hasta sólo un millón de años), con lo que serían quizás los restos del europeo más antiguo.
De todas formas, independientemente de la atribución o no del hom-bre de Orce a la línea de los humanos, los restos fósiles de herramientas encontradas en la zona del sureste español, y atribuidas a la cultura o tra-dición olduvaiense, refuerzan la tesis de la posible llegada del ser humano a Europa, procedente de África, a través de la vía occidental.
75 Cfr. Carbonell, E. y otros, «Instrumentos de los homínidos de Atapuerca. Las prime-ras comunidades humanas», Mundo Científico, 1997, n.º 175, 54-59.
76 Cfr. Carbonell, E./Arsuaga, J. L. y resto del equipo, «Homo Antecessor y su medio natural. Cómo eran y cómo vivían los europeos más antiguos», Mundo Científico, 1998, n.º 192, 42-49.
77 El collar del neandertal, o.c., p. 76. Cfr. también Arsuaga, J. L., «Canibalismo prehis-tórico», El País, 11-XII-1996, p. 36.
78 Cfr. Cela Conde, C. J./Ayala, F. J., o.c., pp. 327 y ss.

164
© Universidad de Deusto - ISBN 978-84-9830-383-4
4. ¿Homo floresiensis?79
Un descubrimiento paleoantropológico reciente ha introducido en el esquema evolutivo de los humanos serios interrogantes y no pocas difi-cultades. Se trata del denominado Homo Floresiensis, Hombre de Flores,denominado así por haberse encontrado sus restos en la isla de Flores, In-donesia, por antropólogos australianos e indonesios. Sus características físicas resultan sorprendentes, puesto que poseía una estatura de tan sólo un metro de alto, unos 30 kilos de peso y una capacidad cerebral de unos 380 cm3, similar a la de un chimpancé. Lo curioso es que se han encontra-do junto a estos restos unas herramientas líticas de avanzada complejidad, similar a las del hombre moderno. En esta misma isla de Flores se habían descubierto con antelación unas herramientas de piedra de 800.000 años de antigüedad. Si a todo esto se añade la necesidad de poseer una técnica de construir balsas para poblar la isla, resulta difícil explicar esto y las herramientas halladas, si tenemos que atribuirlas a un ser con un cerebro similar al del chimpancé.
Normalmente los restos antropológicos suelen ser noticia porque mar-can hitos de antigüedad y de inicio de un nuevo tipo de humano, una nue-va especie, pero estos restos del Homo Floresiensis se convierten también en noticia porque representan el límite de supervivencia de una especie que los investigadores sitúan dentro del homo o grado erectus. El proble-ma está en que el homo erectus poseía una estatura y capacidad craneana mucho mayores. La posible explicación estaría en aceptar que fue un gru-po de homo erectus los que llegaron a la isla, y, por razones de carencia de presión evolutiva que amenazara su existencia, inició un proceso de disminución de tamaño que se da también en diversas especies animales, del que un ejemplo claro está en un tipo de elefante enano, el Stegodón,presente también en la isla de Flores. Según esta teoría, el homo floresien-sis sería una versión enana del homo erectus. Pero no es una explicación suficiente, puesto que el tamaño de su cerebro no es proporcional al del conjunto del cuerpo.
Todos estos enigmas resultan inexplicables de momento tanto para los antropólogos que intervinieron en su descubrimiento en 2003 (la publi-cación de los resultados se ha hecho en 2004, en la revista Nature), Peter Brown, M. Morwood y Bert Robert, australianos, como para el resto de
79 Cfr. Morwood, M. J. y otros, «Archaeology and Age of a New Hominin from Flores in Eastern Indonesia», Nature, vol. 431, 28 de octubre de 2004, pp. 1087-1091; Brown, P. y otros, «A New Small-Bodied Hominin from the Late Pleistocene of Flores, Indonesia», Nature, vol. 431, 28 de octubre de 2004, pp. 1055-1061; Wong, Kate, «El Hombre de Flo-res», Investigación y Ciencia, abril 2005, n.º 343, pp. 22-31; Id., «Nueva luz sobre el hom-bre de Flores», Investigación y Ciencia, 2010, enero, n.º 400, pp. 60-67.

165
© Universidad de Deusto - ISBN 978-84-9830-383-4
paleoantropólogos. Los análisis genéticos que se están intentando hacer de su ADN mitocondrial, quizás puedan aclarar algunos de estos interrogan-tes, y aportar datos más o menos seguros sobre el lugar que les correspondeen el dendograma evolutivo, así como el momento en que se distanciaron evolutivamente del hombre actual. Tampoco se descarta el descubrimien-to de cualquier nuevo dato que nos aporte nuevas luces sobre esta especie tan problemática.
En definitiva, ¿qué decir sobre el «grado erectus» y sus diferentes propuestas de clasificación de las diversas especies que lo abarcarían? Tras los dicho hasta ahora, parece que se pueden deducirse las siguientes conclusiones80:
— el grado erectus se inició en África del este, alrededor de 1,8 y 1,9 millones de años, momento en que aparecen unos nuevos seres con cambios anatómicos de la bóveda craneal y un aumento impor-tante de tamaño;
— estos primeros erectus mantienen al inicio la cultura olduvaiense de los habilis, pero las irán cambiando hacia el tipo de herramientas denominadas achelenses (bifaces, que no se hallan casi en Asia, y que en Europa aparecerán sólo en época muy posterior);
— la salida de África se produce pronto, orientándose hacia Asia y Europa, no sabiéndose todavía si el lugar de salida fue sólo Egipto o también Gibraltar, o Sicilia;
— la evolución de los diferentes tipos de homo erectus se daría en cada lugar de acomodo. Así, el homo erectus en sentido estricto se dio en Asia; en Europa occidental surgió el homo antecessor, al que sucedería el homo heidelbergensis;
— en el paso al Paleolítico superior se dan diferentes transformaciones en cada continente: en Asia permaneció el homo erectus; en Euro-pa, el homo antecessor daría lugar al homo heidelbergensis y luego al hombre de Neanderthal; y en África, el homo antecessor, o el equivalente a esta especie, daría lugar al homo sapiens.
c.3) Los neanderthales y el homo sapiens moderno
Como puede verse, el salto del grado erectus hasta el hombre moder-no no está claro, y para explicarlo se han propuesto varias hipótesis. Todas coinciden en afirmar que entre el erectus y el hombre moderno se dieron varios seres humanos intermedios, pero surgen las diferencias a la hora de denominarlos y de establecer el cuadro clasificatorio que explique las re-
80 Cfr. ibídem, pp. 337 y ss.

166
© Universidad de Deusto - ISBN 978-84-9830-383-4
laciones entre ellos. Dos son las hipótesis más importantes que se dispu-tan la hegemonía:
a) El modelo de evolución multirregional, que defiende que el origen del hombre moderno es el resultado de la mezcla de grupos dife-rentes que experimentaron el salto mutacional genético en diferen-tes partes del mundo a la vez.
b) Y el modelo que defiende una salida única desde África, que con-sidera que el origen del hombre moderno es único y se dio en Áfri-ca, desde donde salió posteriormente para ir imponiéndose como especie única en el resto de la tierra, desplazando a los neanderta-les y a los restos de homo erectus en el ámbito asiático.
Más adelante nos detendremos en los argumentos de cada una de estas dos teorías o modelos. En la medida en que nos hallamos ante un gran vacío de evidencias, los paleoantropólogos sitúan en este gran espa-cio que va entre el grado erectus y el hombre moderno diversas especies de humanos situadas en otros tantos espacios geográficos y franjas histó-ricas. En esta franja se sitúan las especies denominadas preneandertha-les, existentes tanto en África (homo heidelbergensis, homo rodhesien-sis, homo swanscombensis, etc.), como en Europa81 (homo heidelbergensis,homo steinheimensis, etc.) y Asia (en Java y en China). Pero no nos va-mos a detener en tantos detalles, sino que nos limitaremos a presentar los elementos y rasgos más significativos de los hombres de Neanderthal y los modernos o de Cro-Magnon.
1. Los Neandertales
El tiempo que va entre 127.000 y 40.000 años, es lo que se suele deno-minar «el tiempo de los neandertales»82. Posiblemente sea la especie huma-na sobre la que mejor información tengamos sobre sus características, dado el gran número de fósiles con que se cuenta. Hacia los 40.000 años, apare-cen en Europa los primeros antecesores del hombre actual, los cromañones,aunque su origen africano hay que situarlo hace alrededor de 100.000 años. De modo que entre los 40.000 y los 30.000 años convivieron en el mismo espacio europeo las dos especies, desapareciendo definitivamente los nean-dertales hacia los 30.000 años.
81 La Sima de los Huesos, en los yacimientos de la Sierra de Atapuerca, encierra la in-formación más amplia sobre esta época que va entre el grado erectus y el hombre de nean-dertal y el moderno.
82 Cfr. Arsuaga, J. L., El collar del neandertal, o.c., p. 77.

167
© Universidad de Deusto - ISBN 978-84-9830-383-4
La industria específica de los neandertales corresponde al Modo Téc-nico III, llamado también Musteriense (Le Moustier, Francia)83. Arsuaga señala que en buena lógica a los neandertales se les debiera llamar cal-penses, dado que fue en Calpe (el nombre que se daba a Gibraltar) donde se dio a conocer por primera vez un resto fósil de esta especie, con ocho años de antelación al hallazgo de la cueva Feldhofer, en el valle del río Neander (Alemania). Aunque el primer resto se localizó con todavía ma-yor antelación en la cueva de Engis (Bélgica), en 1829 o 1830. «El nom-bre científico Homo neanderthalensis fue creado por William King en una comunicación leída en la reunión de 1863 de la British Association for the Advancement of Science»84.
A pesar de que se conozcan bastante bien las características de los neanderthales, dado el gran número de fósiles que se tiene, hay algunos aspectos de gran importancia sobre los que se tienen grandes sombras. Es el caso de la cadera y de la base del cráneo, por otro lado fundamentales para el estudio del parto y del habla, respectivamente. Las características morfológicas de los neandertales se refieren sobre todo a determinados arcaísmos, rasgos propios de la línea evolutiva y que nosotros hemos per-dido. Los neandertales tenían la pigmentación de la piel blanca, mientras que los cromañones la tenían más morena, dado que éstos procedían de África mientras que los primeros se habían aclimatado al entorno euro-peo, un ámbito menos cálido. Otras diferencias importantes se refieren a la configuración del rostro, en el que sobresalía un toro supraorbitario muy desarrollado en los neandertales, pero sobre cuyas características y origen del mismo no vamos a detenernos aquí85.
Su peso corporal era superior al hombre moderno, pudiéndose con-siderar que los individuos masculinos pesarían alrededor de 80 kgrs. Si se tienen en cuenta los restos de neandertales encontrados en Atapuerca (Sima de los Huesos), se puede deducir, dada la fortaleza de los fému-res encontrados, que la estatura de los individuos masculinos pudo ser entre 170 y 180 cms., por lo que no es exagerado atribuir un peso de no menos de 90 kg.86 Estas dimensiones corporales, mayores que las de los cromañones, hace que se pueda leer de modo razonable la comparación de las dimensiones craneales entre ambas especies. La mayor capacidad craneana encontrada en un neandertal se ha dado en el yacimiento israe-lí de Amud, y arroja la cifra de 1.750 cc., cantidad superior a cualquier
83 Par una visión completa de los neandertales, cfr. Cavalli-Sforza, Luca y Francesco,¿Quiénes somos? Historia de la diversidad humana, o.c., 59-70.
84 Arsuaga, J. L., El collar del neandertal, o.c., 81.85 Cfr. Arsuaga, J. L., o.c., pp. 91-97.86 Cfr. Arsuaga, J. L., o.c., 102.

168
© Universidad de Deusto - ISBN 978-84-9830-383-4
dato encontrado incluso entre los cromañones. Pero es importante tener en cuenta que la masa encefálica es correlativa al peso global del cuer-po, denominándose a esa proporción índice de cefalización. De modo que hay que advertir que no por tener una masa encefálica mayor se po-see, sin más, mayor inteligencia. Los restos culturales de los neanderta-les nos inclinan a pensar que se trataba de una especie muy parecida a nosotros, aunque sus rasgos físicos tuvieran un acusado primitivismo, e incluso se pueda afirmar (lo que no está todavía probado) que no llega-ron a poseer la capacidad de hablar de modo articulado como nosotros87.En definitiva, como señala Arsuaga, «no tenemos por lo tanto a partir de los fósiles argumentos para suponer a los neandertales menos capaces mentalmente que nosotros; más apropiado sería decir que nosotros so-mos menos pesados»88.
Uno de los problemas que más interés despierta en la actualidad en-tre los paleoantropólogos se refiere a la convivencia y transición entre los neandertales y los cromañones89. Ya hemos señalado que los datos fósi-les muestran que hubo unos 10.000 años de convivencia entre ellos (en-tre los 40.000 y 30.000 años). Una de las investigaciones más interesan-tes en este punto se refiere a los esfuerzos por dilucidar si hubo entre los neandertales y los cromañones mezcla parental e intercambio genético. El científico sueco, Svante Pavo, investigador del Max Plank Institute de Leizig (Alemania), es quien está abanderando las investigaciones sobre el ADN de los neandertales, para ver con ello si existe en el hombre actual restos de la herencia genética de los neandertales90. De momento, las res-puestas son negativas, aunque todavía las investigaciones se hallan en sus comienzos. Pero todo parece indicar que la respuesta será más bien nega-tiva. La conclusión es que no se sabe mucho sobre las relaciones que hubo entre las dos especies, ni a qué se debió la desaparición de los neanderta-les y su sustitución total y definitiva por los cromañones.
2. Los Cromañones. El homo sapiens
La mayoría de los paleoantropólogos piensan en la actualidad que los neandertales son una especie colateral y no un antepasado genético de los
87 Cfr. B. Arensburg y A. M. Tilier, «El lenguaje del hombre de neanderthal», MundoCientífico, 1990, n.º 107, nov. o.c., cap. 16.º; cfr. Arsuaga, «La especie elegida», donde se inclina por atribuir también lenguaje articulado a los neandertales. Pero este tema lo trata-remos más adelante, con más amplitud.
88 O.c., p. 105.89 Cfr. Cavalli-Sforza, Luca y Francesco, ¿Quiénes somos?, o.c., pp. 60 y 72-73. 90 Cfr. Rivera, Alicia, Entrevista a Svante Pavo: «Buscamos las diferencias genéticas
entre los neandertales y los humanos actuales», El País, 4 de abril de 2007, p. 35.

169
© Universidad de Deusto - ISBN 978-84-9830-383-4
cromañones91. Aunque no todos están de acuerdo, la mayoría parece tam-bién inclinarse por considerar que los neandertales evolucionaron de esla-bones anteriores (H. heidelbergensis, u Homo antecessor) fuera de África. En cambio, el Hombre de Cro-Magnon tendría su origen en la zona cen-tral de África92, y, desde allí, al igual que el Homo erectus, se extendería poco a poco por el resto del planeta, saliendo de África por la zona del Mar Rojo93. De ahí que su contacto e imposición sobre los neandertalesocurrió de modo diferente y en épocas distintas según los lugares de Oriente Medio (Israel, Siria, Líbano) y de Europa a que hagamos refe-rencia.
La relación entre ambas especies, como es lógico suponer, no fue uni-forme. En unos sitios se habría dado una sustitución bélica o más agresi-va, y en otros, hay indicios de convivencia pacífica e incluso de compartir una misma cultura. Los yacimientos donde se advierte una convivencia de mayor interacción pacífica se sitúan en el entorno de Israel94. En ellos se advierte que en la época de hace unos 100.000 años, esa zona constituía, como señala el paleoantropólogo israelí Yoel Rak, el límite meridional de ocupación de los neandertales y el límite septentrional de los cromaño-nes95. Los indicios arqueológicos apuntan a una presencia alternativa de ambas especies en las mismas cuevas y hábitats. Según Y. Rak, «no hay indicios de competencia ni de lucha entre ambas especies humanas. Los neandertales eran muy especializados, eran muy dependientes del entor-no, de las condiciones climáticas, estaban muy bien adaptados al frío y cuando el clima cambió no pudieron sobrevivir. El homo sapiens es más flexible, por su capacidad cerebral, para adaptarse y solucionar los proble-mas, no mediante la biología, sino por su inventiva»96.
La convivencia pacífica de Oriente Medio se decantó por la sustitu-ción de los neandertales por los cromañones, que fueron extendiéndose poco a poco por Asia y Europa. Las evidencias fósiles de Europa defien-
91 Es interesante desde este punto de vista la polémica sobre el famoso «niño de Lape-do», fósil descubierto en Portugal, en 1998, que según sus descubridores, entre los que se encuentra Joao Zilhao, sería un híbrido genético entre neandertal y cromañón. Pero la ma-yoría de los paleoantropólogos se inclina por la hipótesis contraria. Cfr. Cela Conde, C. J./Ayala, F. J., o.c., pp. 415-417; El País, 18 de febrero de 2000, p. 40
92 Cavalli-Sforza, L. y Fr., o.c., p. 70. 93 Cuando hayamos expuesto los planteamientos de la Antropología molecular, nos re-
feriremos a la disputa entre las teorías multifocales y unifocales sobre el origen del hombre moderno.
94 Sobre las diferentes teorías y datos sobre la relación entre neandertales y hombre moderno, cfr. Cela Conde, C. J./Ayala, F. J., o.c., cap. 8.º, pp. 389 y ss.
95 Cfr. la entrevista realizada a Y. Rack por Alicia Rivera, El País, 5 de abril de 2000, p. 49.
96 Ibídem, p. 49.

170
© Universidad de Deusto - ISBN 978-84-9830-383-4
den este hecho, en la medida en que en el norte de Europa van desapare-ciendo los neandertales con más rapidez que en el sur97.
Como hemos señalado, no se sabe todavía cuál fue el factor determi-nante respecto a la imposición de los cromañones sobre los neanderta-les. Ya hemos dicho que no está nada claro que los primeros poseyeran una mayor inteligencia, a tener por su capacidad craneana, puesto que los neandertales poseían por término medio mayor capacidad, aunque revelen un menor índice cefálico. Pero los cromañones tenían un tipo de indus-tria, un tipo de organización social y cultural, posiblemente basado todo ello en el dominio del lenguaje articulado, que les dio una determinante superioridad sobre su otro contrincante humano. Las herramientas que utilizaban los cromañones se considera que pertenecían al denominado Modo Técnico IV, llamado también Auriñacense, superior al Modo III de los neandertales. Este mayor desarrollo cultural les permitió ir pasando pro-gresivamente de un estilo de vida propio de los cazadores-recolectores a un estilo de vida más sedentario, el de los agricultores y ganaderos98.
De todas maneras, nos señala Arsuaga, «tenemos aún una imagen bo-rrosa de esta fascinante etapa de la Prehistoria. ¿Qué fue lo que empujó a los cromañones hacia el sur hace 30.000 años? ¿Coincidió con un cambio climá-tico y ecológico que destruyó los encinares mediterráneos? Si fue así, ¿por qué los cromañones eran más eficaces en los paisajes desolados? ¿Hubo al-gún cruce entre unos humanos y otros, aunque fuera en algún remoto rincón? Tengan un poco de paciencia, se está investigando en el tema»99.
Lo que sí es evidente es que los cromañones se van progresivamente aposentando en toda la superficie del planeta, quedándose ya como única especie humana. Tras situarse primero, como hemos señalado en Oriente Medio, se expandieron por todo Asia. Se han encontrado restos fósiles en China con una antigüedad de 60.000 años, aunque es una datación no muy precisa. En esa misma época llegaron a Nueva Guinea y Australia100.
97 Cfr. Arsuaga, J. L., «El último neandertal», El País, 22 de diciembre de 1999, p. 47. 98 Esta orientación progresiva hacia un estilo de vida más sedentario por parte de los
cromañones, está confirmado por los análisis del ángulo formado por la cabeza del fémur, realizados por Erik Trinkaus, «El cuello del fémur: ¿un indicador social para los antropólo-gos?», Mundo Científico, 1994, n.º 151, 982-983.
99 «El último neandertal», El País, 22 de diciembre de 1999, p. 47.100 Respecto a la llegada del hombre moderno a Australia, Cavalli-Sforza señala que
«puede que llegara en embarcaciones, cruzando un brazo de mar que en aquella época no era tan ancho, pero de todos modos suponía un obstáculo que sólo se podía salvar con al-gún medio náutico, por primitivo que fuera. En Australia se han encontrado los restos fósi-les de un hombre (que según la mayoría de los paleoantropólogos es un hombre moderno) de hace 40.000 a 35.000 años, y yacimientos con abundante material arqueológico bien da-tado en torno a los 55.000-60.000 años atrás», o.c., p. 71.

171
© Universidad de Deusto - ISBN 978-84-9830-383-4
La llegada del hombre moderno a Europa fue más bien tardía, entran-do por Oriente, entre 40.000-35.000 años, como ya hemos señalado an-teriormente. Y más adelante se orientó hacia las zonas del norte de Asia, zonas más frías y desérticas. Y de allí se dirigiría a América, penetrando por el norte, a través del estrecho de Bering, en el período en que estaban unidos los dos continentes por el hielo (la última glaciación). Estamos ha-blando de fechas ya muy recientes, hace unos 15.000 años.
Con la llegada del hombre moderno, se va imponiendo unos nuevos patrones culturales. Ya hemos señalado que el tipo de industria lítica se denomina auriñacense, sustituyendo a la musteriense de los neandertales. Pero no sólo se dan innovaciones tecnológicas, sino que también aparece con el hombre moderno una gran sensibilidad artística. De esta época se van encontrando abundantes grabados y pinturas murales, sobre todo en la región del suroeste francés y del norte de España (País Vasco y Can-tabria), así como abundantes estatuillas, tanto para adorno personal como con finalidad religiosa.
Dentro ya de la historia del hombre moderno, un momento clave lo constituye la llamada revolución neolítica, realizada hace unos 10.000 años, momento que coincide también con el paso de una cultura de cazadores-recolectores a otra sedentaria, de agricultores y ganaderos101.
2.1.2. El punto de vista de la biología molecular
Señalamos en su momento que el punto de vista de la paleoantropolo-gía tenía la ventaja sobre la biología molecular de presentar evidencias di-rectas a través de los fósiles que se van encontrando. Pero estos descubri-mientos están sometidos a la buena suerte, además de que tales hallazgos son parciales y difíciles a veces de datar e interpretar. La biología molecu-lar tiene la ventaja de que no necesita esperar a la suerte de los hallazgos fósiles, sino que le basta analizar la estructura molecular de nuestras célu-las y las de las demás especies animales para deducir de ahí la historia del proceso evolutivo. También es cierto que su desventaja o sus problemas se hallan en la dificultad de determinar el ritmo de transformación de esa estructura molecular, con la que va configurando lo que los expertos de-nominan el reloj molecular.
Los avances de esta rama moderna de la ciencia le están permitiendo competir y colaborar con la paleoantropología en el empeño por desentra-ñar las preguntas referidas al origen y tiempo aproximado del primer ser
101 Todos estos temas los trataremos en el capítulo 5.º.

172
© Universidad de Deusto - ISBN 978-84-9830-383-4
humano, y al origen y fechas de la aparición del hombre moderno. Las aportaciones de la biología molecular a la resolución de estas cuestiones van siendo tan importante que está dando origen a la conformación de una nueva rama en el campo de la antropología, la Antropología molecular102.
a) A la búsqueda del «reloj molecular»
Los orígenes de esta disciplina se remontarían a 1962, cuando Emile Zuckerkandl acuñó este título en un simposio científico celebrado en Burg Wartenstein (Austria). Allí lanzó la idea de utilizar la biología molecular para el estudio de la evolución, produciendo un fuerte debate entre los es-tudiosos de la Antropología física. Ahora bien, quien practicó de forma concreta y exitosa este procedimiento fue Morris Goodman, biólogo de la Wayne State University of Detroit103, comparando «la reactividad de las proteínas de suero de monos y hombres (conocidos bajo el nombre común de hominoides), utilizando una técnica inmunológica para poner de mani-fiesto sus afinidades genéticas»104.
Goodman entendía que cuanto más antigua es la separación entre dos especies en el proceso evolutivo, más diferencias genéticas y moleculares habrá entre ellas, por lo que las reacciones que cada una de ellas desarro-llará ante un reactivo serán específicas y diferentes. Como consecuencia de sus investigaciones, Goodman concluyó que el hombre es más cercano y parecido genéticamente al chimpancé que lo que éste pueda ser respec-to al gorila y al orangután. Esto suponía dar por hecho que no siempre los parecidos y diferencias en el ámbito morfológico son conformes con las diferencias de tipo genético y molecular. Por tanto, los modelos clasifica-torios y taxonómicos de las diferentes especies evolutivas debían cambiar-se y trabajar conjuntamente el punto de vista morfológico y el molecular.
Pero nuevos y más importantes avances en el campo de la biología molecular se añadieron poco después de la mano de Allan Wilson y Vin-cent Sarich, bioquímicos de la Universidad de California, en Berkeley105.Analizando las transformaciones de la estructura molecular, que funciona-ría, según ellos, como un reloj molecular, dedujeron que el origen de los
102 Cfr. Lewin, Roger, «El nacimiento de la Antropología molecular», Mundo Científi-co, 1991, n.º 119, 1222-1231.
103 Cfr. ibídem, p. 1224.104 Lewin,R., o.c., 1224;. cfr. también el trabajo de M. Goodman, «Man’s place in the
phylogeny of the primates as reflected in serum proteins», en S. L. Washburn, Classifica-tion and Human Evolution, Metheun and Co., 1964.
105 Los resultados de su investigación los publicaron en Science, 158, 1200, 1967. Cfr. Lewin, R., La interpretación de los fósiles, o.c., 99-106.

173
© Universidad de Deusto - ISBN 978-84-9830-383-4
homínidos era más bien un fenómeno reciente, rebajando la fecha de su aparición hasta unos 7,5 millones de años. Por lo tanto, según Sarich, «ya no era posible seguir considerando un homínido a un fósil que se remon-tase a más de ocho millones de años, cualquiera que fuese su aspecto»106.Eso afectaba de lleno a la discusión, entonces de plena actualidad, sobre si el Ramapithecus era o no ascendiente directo de los humanos. Los mé-todos moleculares lo descartaban totalmente, y se necesitaron nuevos des-cubrimientos paleoantropológicos para resolver la cuestión. El descubri-miento del Sivapithecus mostró que ambos tipos de fósiles eran similares y antecesores del orangután107.
Ahora bien, donde más se ha recrudecido la polémica por las aporta-ciones de la biología molecular se debe a los datos aportados sobre el ori-gen del hombre moderno108. En los últimos tiempos se han propuesto dos hipótesis. Todos los estudiosos de este tema tan interesante y complejo parecen estar de acuerdo en que fue el Homo erectus el primero que aban-donó el continente africano, hace aproximadamente un millón de años, ocupando de modo paulatino Asia y Europa. Posteriormente, hace unos 250.000 años, aparecieron los hombres modernos. Pero aquí ya no hay acuerdo respecto a dónde se originan. El modelo multirregional defiendeque el hombre moderno es producto de mezclas genéticas entre diferen-tes grupos de hombre, desde el homo erectus y el hombre de Neanderthal,para dar lugar al Homo sapiens sapiens. Por tanto, no habría surgido en un solo lugar, sino que fue configurándose desde el intercambio y la con-fluencia de varios centros de fusión.
En contraposición a esa teoría, hay una segunda, que recibe varias de-nominaciones, como modelo del Arca de Noé, modelo Out of África, o modelo de la Eva mitocondrial109, y que defiende que el origen del hom-bre moderno es el resultado de la evolución de una sola población de hu-manos que tuvo su origen en África110. De ahí se habrían extendido por toda Eurasia y resto del mundo, sustituyendo al resto de humanos situados allí en épocas anteriores, sin haberse mezclado genéticamente con ellos. Ya vimos al dar el punto de vista de la paleoantropología, que ésta era la opinión de la mayoría de los que trabajan esa disciplina. Pero donde las
106 Cita tomada de Lewin, R., «El nacimiento de la antropología molecular», o.c., 1225.107 Para la disputa sobre el Ramapithecus y el Sivapithecus, cfr. R. Lewin, «La interpre-
tación de los fósiles», o.c., caps., 5 y 6, pp. 78 y ss.; Andrews, Peter, «Un antepasado para el orangután», Mundo Científico, 1982, n.º 20, pp. 1257-1260.
108 Cfr. Darlu, Pierre/Tassy, Pacal, «El ADN, África y el hombre», Mundo Científico,n.º 73, 1987, pp. 943-945.
109 Cfr. Lewin, R., «El nacimiento de la Antropología molecular», o.c., 1227.110 Cfr. Stringer, C. B. y Andrews, P., Science, 239, 1263, 1988.

174
© Universidad de Deusto - ISBN 978-84-9830-383-4
evidencias resultan más abrumadoras a favor de esta tesis es en el terreno de la antropología molecular111.
Los datos que pretenden confirmar las tesis del modelo unirregionalse presentaron en una publicación de Allan Wilson y sus colegas, en la que daban a conocer sus investigaciones sobre el ADN mitocondrial112. Las mitocondrias son unos pequeños orgánulos celulares, presentes en todas las células de los organismos superiores. Toda célula está compuesta de un núcleo, el citoplasma y la membrana que envuelve a la célula. Las mi-tocondrias se hallan en el citoplasma. Una mitocondria «tiene la forma de una pequeña bacteria, y probablemente lo sea: una bacteria que hace más de mil millones de años que se adaptó a la vida en simbiosis con la célula y pasó a ser una parte muy importante de ella, ya que desempeña la fun-ción de central energética»113.
Pero lo específico de las mitocondrias, para el tema que nos ocupa, con-siste en que sigue poseyendo una cierta independencia respecto a la célula, puesto que tiene en su estructura un pequeño cromosoma con su correspon-diente ADN. «El pequeño cromosoma que se encuentra en la mitocondria humana está formado por unos 15.600 nucleótidos, por lo que es mucho más corto que cualquiera de los cromosomas del núcleo celular, que constan de decenas o centenares de millones»114. Pero el ADN mitocondrial, por no pertenecer al núcleo de la célula, no está sometido a las particiones (meio-sis) y recombinaciones que experimenta el ADN nucleico, durante el pro-ceso de la formación de las células sexuales (gametos) y su posterior fusión con el gameto del otro sexo para conformar la célula germinal en el proceso de la fecundación de óvulo por el espermatozoide.
Además, otra característica del ADN mitocondrial consiste en que su transmisión se da exclusivamente por vía femenina115, ya que se sitúa en el
111 Cfr. Ayala, F. J., «¿Desde África? Una perspectiva de los elementos poblacionales en la aparición de los seres humanos modernos», Ludus Vitalis, VIII (2000), n.º 13, 135-156; Id., «El mito de Eva y la evolución humana», en Forum Deusto, Innovación y cambio. Ha-cia una nueva sociedad, Vol. II, Bilbao, Universidad de Deusto, 1997, 121-137; Moya, A., «La genética de poblaciones y la trazabilidad genética de la historia de la vida: el poder de los marcadores moleculares», Ludus Vitalis, VIII (2000), n.º 13, 157-161; Mestres, F., «Procedimientos utilizados por la evolución molecular para entender el origen del hombre moderno», Ludus Vitalis, VIII (2000), n.º 13, 163-170.
112 Cfr. Cann, R. et al., Nature, 325, 31, enero de 1987. 113 Cavalli-Sforza, Luca y Francesco, ¿Quiénes somos?, o.c., 77; cfr Ayala, Fco. J.,
La teoría de la evolución, Madrid, Temas de Hoy, 1999, cap. 12, «Evolución molecular», pp. 179 y ss.
114 Cfr. Cavalli-Sforza, L. y Fr., o.c., 80.115 Investigaciones recientes parece que ponen en entredicho esta afirmación, puesto que
se han dado muestras en algunos casos de transmisión también por vía masculina. Cfr. Strauss, Evelyn, «El tiempo variable de los relojes de DNA», Mundo Científico, 1999, n.º 204, 20-23.

175
© Universidad de Deusto - ISBN 978-84-9830-383-4
citoplasma, y la aportación masculina se centra sólo en la mitad del ADN nucleico. El ADN mitocondrial tiene de vez en cuando cambios, mutacio-nes, al igual que lo tienen otras partes de la estructura celular, molecular. Se trata de unas mutaciones que al parecer mantienen un ritmo constante en el proceso de mutación y transformación, constituyéndose en un reloj molecular.
Esa es la tesis de Alan Wilson y sus colegas en el trabajo que hicie-ron público en 1987 y que tan fuerte impacto produjo no sólo en el ámbito científico sino también en toda la opinión pública, al hacerse referencia a la Eva mitocondrial. El trabajo publicado hacía referencia al estudio del ADN mitocondrial, analizando el 9% de nucleótidos de una muestra de individuos humanos pertenecientes a Asia, Australia, Europa, Nueva Gui-nea y África, comparándola con un grupo de muestra africana, compuesto en su mayor parte por negros americanos116. Los datos obtenidos permitía situar a todos los seres humanos actualmente existentes en una relación genética configurada por un árbol genealógico constituido por dos ramas, «una sólo contiene tipos de ADN mitocondrial africano y la segunda una mezcla de todos los tipos. En otras palabras, el árbol evolutivo parece “enraizado” en África, con ramas que se han desarrollado ulteriormente en el resto del “Antiguo Mundo”»117. Eso supone que, si se da por hecho que el ritmo de las mutaciones es constante, el grupo africano es el más antiguo y los que primero han ido evolucionando.
Intentando calcular el momento a partir del cual se comenzó este pro-ceso evolutivo, los investigadores consideran que habría habido un antepa-sado común, una mujer, que habría vivido entre 140.000 y 290.000 años. Ante esto, los artículos periodísticos empezaron a hablar de una Eva afri-cana, interpretándose incluso en la línea de dar la razón a una interpreta-ción literal y fundamentalista de la Biblia118. De ahí que A. Wilson, pre-maturamente fallecido en el verano de 1991, ha tenido que salir al paso de estas inexactas y tendenciosas interpretaciones, puesto que decir que el análisis de ADN mitocondrial apunte a una mujer, situada en África hace unos 200.000 años, de la que todos los seres humanos actuales poseería-mos al menos un fragmento de su ADN mitocondrial, no implica afirmar que esa mujer sea la representante femenina de una primera pareja de hu-manos de la que descenderíamos todos, como el punto de origen del hom-bre moderno, el llamado homo sapiens sapiens.
116 Cfr. Lewin, R., «El nacimiento de la Antropología molecular», o.c., 1227.117 Ibídem, 1228. El entrecomillado es del autor. 118 Cfr. Ayala, F., «El mito de Eva y la evolución humana», o.c.; Berger, L. R./Hilton-
Barber, B., Tras las huellas de Eva, Barcelona, Ediciones B, 2001; Sykes, Bryan, Las siete hijas de Eva, Madrid, Debate, 2001.

176
© Universidad de Deusto - ISBN 978-84-9830-383-4
Las críticas de los científicos se han dirigido a varios aspectos de las conclusiones de A. Wilson. La primera se orienta a negar que se pueda ha-blar con tanta seguridad de un reloj molecular. El grupo de Wilson calcu-laba que el ritmo de mutación de los nucleótidos era del 2 al 4% por mi-llón de años. Pero, a parte de considerar algunos que se trata de un ritmo demasiado rápido y que situaba el origen del hombre moderno en una fe-cha muy reciente, se dudaba que se tuvieran datos suficientes como para hablar de un ritmo constante en la mutación de los nucleótidos, con lo que el concepto de reloj molecular sería difícil mantenerlo119. Por tanto, se consideraba que aunque los datos aportados son de gran interés, todavía tiene que avanzarse mucho en precisión para que puedan ser considerados como datos totalmente seguros.
Por otro lado, aunque A. Wilson y sus colaboradores no pensaban, como hemos dicho, en una primera mujer de la que se habría originado el hombre moderno, sí que pensaban en un cierto estrangulamiento pobla-cional como paso de una especie o subespecie a otra. Así lo indican estas palabras de Wilson: «Es tentador establecer una relación entre la apari-ción del tipo de ADN mitocondrial ancestral y una reducción drástica del tamaño de la población (estrangulamiento)»120. Pero más tarde, el mismo Wilson reconoció que era desafortunado afirmar eso, puesto que todas las evidencias científicas posteriores se inclinan a pensar que no es necesario ningún tipo de estrangulamiento para dar origen a una nueva especie, y en el caso de los humanos modernos parece que los datos apuntan a que es altamente improbable tal estrangulamiento121.
Igualmente, investigaciones más recientes indican que el ADN mito-condrial (ADNmt) no sólo se transmite por vía femenina, sino que a veces también puede ser transmitido por vía masculina122. A pesar de ello, otros investigadores están intentando encontrar relojes moleculares diferen-tes al ADNmt, que nos permitan llegar a saber algo similar en el engarce
119 Cfr. Darlu, Pierre y Tassy, Tassy, «El ADN, África y el hombre», Mundo Cien-tífico, 1987, n.º 73, 943-945; 945. Cfr. también R. J. Britten, Science, 231, 1393, 1986; Strauss, E., «El tiempo variable de los relojes de DNA», o.c.
120 Cita tomada de Lewin, R., «El nacimiento de la Antropología molecular», o.c., 1229.121 Cfr. Klein, Jan/Takahata, N./Ayala, F. J., «Polimorfismo del MHC y origen del hom-
bre», Investigación y Ciencia, 1994, n.º 215, 14-19. Cfr. Lewin, R., artíc. cit., p. 1229, nota 9, donde se hace referencia a las investigaciones de J. Klein y colegas del Max-Plank Institut für Biologie de Tubinga, sobre los genes del complejo principal de histocompatibilidad, llamado complejo HLA en los humanos, aspecto sobre el que trabajan también F. J. Ayala y colegas.
122 Cfr. Strauss, Evelyn, «El tiempo variable de los relojes de DNA», o.c.; en pp. 20-23 señala que «nuevos datos de la biología de las mitocondrias vuelven a cuestionar la preci-sió de los relojes moleculares. Estos relojes no van a la misma velocidad en todos los gru-pos de animales y en todos los períodos de la evolución. Además, es posible que no here-demos estos orgánulos sólo de la madre» (p. 20).

177
© Universidad de Deusto - ISBN 978-84-9830-383-4
masculino de nuestra evolución123. De todos modos, todas estas críticas a los datos del trabajo de Wilson no son tan demoledoras como para invali-dar el camino emprendido por la antropología molecular, sino que indican que se trata de un camino de gran interés pero todavía incompleto y nece-sitado de maduración. De hecho, determinados aspectos de la antropolo-gía molecular coinciden con los datos aportados por la paleoantropología, y en la actualidad le sirven para caminar ambas ciencias de la mano en el empeño de conseguir más precisión en muchos aspectos todavía oscuros sobre el origen del hombre moderno124.
b) Implicaciones respecto al origen del hombre moderno
Ya apuntamos un poco más arriba que en la actualidad existen dos modelos sobre el origen del hombre moderno, el modelo multirregional y el unirregional, denominado modelo del Arca de Noé, Out of África o de la Eva mitocondrial. Los defensores de la antropología molecular se incli-nan todos por este segundo modelo, en el que coinciden con muchos pa-leoantropólogos.
En realidad, cabría diferenciar en tres los modelos o teorías sobre el origen del hombre moderno, siguiendo a Ch. B. Stringer125. El modelo
123 En nov. de 1995 se produjo la noticia de que genetistas estadounidenses y británicos creían haber encontrado un Adán ancestral, que habría vivido hace unos 188.000 años, en África, constituyendo la pareja de la Eva mitocondrial de las investigaciones de A. Wilson y colegas. Estas conclusiones las basaban en el estudio de segmentos específicos de ADN del cromosoma Y (masculino). El estudio de los científicos estadounidenses, de la Univ. de Arizona, publicado en Nature, sitúa a este Adán en la fecha dada anteriormente. Pero los científicos ingleses, de la Univ. de Cambridge, también publicado en Nature, acercan la fe-cha hasta unos 49.000 años. Otro grupo de investigadores estadounidenses, esta vez de la Univ. de Yale, cuyas conclusiones han publicado en la rev. Science (nov. de 1995), sitúan la presencia de este Adán ancestral hacia unos 270.000 años. Pero el problema, como he-mos señalado antes, de estas investigaciones es que no se acaba de tener claro que el ritmo de la evolución de los nucleótidos del ADN se constante, y, por tanto, si se puede hablar o no de «reloj» molecular. Cfr. Ayala, Fco. J., «El mito de Eva y la evolución humana», en VV.AA, Innovación y cambio. Hacia una nueva sociedad, Bilbao, Universidad. de Deus-to, 1997, vol. II, 121-137, pp. 121(hace referencia a mayo de 1995, cuando hacen público el estudio los de Yale y Harvard) y 135 (el Adán ZFY). Cfr. también Ayala, F. J., «¿Desde África? Una perspectiva de los elementos poblacionales en la aparición de los seres huma-nos modernos», o.c., pp. 148-150.
124 Cfr. Sykes, Bryan, Las siete hijas de Eva, Madrid, Debate, 2001. 125 Cfr. Stringer, Ch. B., «¿Está en África nuestro origen?», Investigación y Ciencia,
1991, febrero. Después ha sido recopilado en Bertanpetit, Jaume (comp.), Orígenes del hombre moderno, Barcelona, Prensa Científica, S.A., 1993, 12-19 (las citas las hago por esta publicación).

178
© Universidad de Deusto - ISBN 978-84-9830-383-4
multirregional (también llamado por B. Vandermeersch como hipótesis del candelabro126) defiende, como ya hemos indicado, que el origen del hombre moderno no se dio en un solo lugar, sino en varios, fruto de mez-clas genéticas con especies anteriores, desde el Homo erectus, primer es-pecie humana que sale de África. Según esta teoría, los caracteres racia-les, que son antiquísimos, evolucionaron, en largos períodos de tiempo, en las mismas regiones donde se encuentran en la actualidad. De modo que estas características raciales serían descendencia de los neanderta-les en Europa, y del homo erectus en Java y regiones limítrofes. Esta teoría fue propuesta por F. Widenreich, en 1945. Entre los paleoantro-pólogos que defienden este modelo se hallan, entre otros, Alan G. Thorne y Milford H. Wolpoff127.
El segundo modelo es el de dispersión desde África (Out of África).Todos los hombres modernos, según este modo de pensar, procedemos de África, lugar desde el que se extendió por todo el mundo. Por tanto, los rasgos raciales se fueron adquiriendo como consecuencia del largo proce-so adaptativo a los diferentes ambientes ecológicos del planeta. Esta espe-cie nueva no se mezcló con los humanos de especies anteriores, sino que fue sustituyéndolos de una forma sobre cuyos detalles no se tiene un co-nocimiento completo sino hipótesis muy generales128.
Un tercer modelo se denomina de flujo genético o hibridación. Esta teoría «retrotrae las poblaciones modernas a una red de antiguos linajes, cuya contribución genética variaría de una región a otra, y cuya tasa de intercambio aumentaría quizás a medida que fueran evolucionando los hombres modernos»129.
Hemos ido viendo, al estudiar el punto de vista de la paleoantropolo-gía, que la fecha atribuida a la aparición del hombre moderno ha ido re-trocediendo desde las primeras aproximaciones, que la situaban en torno a 35/40.000 años, hasta situarla más allá de los 100.000, debido a nue-vos descubrimientos paleontológicos, a nuevos métodos de datación, y a los progresos en la genética o biología molecular. Si los descubrimien-tos fósiles de los yacimientos de Oriente Medio situaban la aparición del hombre moderno alrededor de unos 100.000 años, los datos genéticos lo
126 Cfr. «Homo sapiens sapiens: lo que dicen los fósiles», Mundo Científico, 1995, n.º 160, 710-716.
127 Cfr. Thorne, A.G./Wolpoff, M. H., «Evolución multirregional de los humanos», In-vestigación y Ciencia, junio de 1992, recopilado en Bertranpetit, Jaume (comp.), Orígenesdel hombre moderno, o.c., 26-32.
128 Cfr. Wilson, A. C./Cann, R. L., «Origen africano reciente de los humanos», Investi-gación y Ciencia, 1992, n.º 189, 8-13; Ayala, F. J., «¿Desde África? Una perspectiva de los elementos poblacionales en la aparición de los seres humanos modernos», o.c.
129 Ch. Stringer, B., o.c., p. 12.

179
© Universidad de Deusto - ISBN 978-84-9830-383-4
retrasan a unas fechas que se sitúan entre 140.000 y 290.000 años. Aun-que se trata de datos todavía muy imprecisos, que tendrán que contras-tarse a medida que avancen las investigaciones. Apoyándose en datos genéticos y paleoantropológicos, surge la teoría del Arca de Noé (porque da por hecha la desaparición de las demás poblaciones humanas), defen-dida por W.W. Howell, en 1976130. Los datos genéticos en que se apo-ya, proceden de los trabajos de Rebecca Cann, Mark Stoneking y Allan Wilson, de la Universidad de Berkeley sobre el ADNmt131, y en otros es-tudios basados en el DNA del núcleo celular (ADNn)132. Los datos apor-tados por las investigaciones moleculares pretenderían ser concluyentes sobre la inexistencia de restos de ADNmt y ADNn en la estructura ge-nética del hombre actual. Pero esto, según algunos paleoantropólogos crí-ticos, sería válido para Europa, donde la sustitución de los neandertales por los cromañones parece ser que fue total. Pero no se tiene la seguri-dad de que eso fuera lo que sucedió en Asia, donde las investigaciones están apoyadas en pocos datos.
Por eso, Bernard Vandermeersch propone un modelo sintético, o hi-pótesis de la evolución reticulada, basada en un esquema reticulado de la evolución133. Este esquema, en realidad, se acerca más al modelo multirre-gional o del candelabro, siendo casi una complejificación del mismo, de-fendiendo enlaces y mezclas genéticas entre poblaciones contiguas, hasta dar como resultado la mezcla de estructura genética y racial que constitu-ye la pluralidad de poblaciones del hombre moderno.
Como puede verse, estamos todavía muy lejos de llegar a consensos fundamentales sobre los aspectos más importantes a cerca del origen del hombre moderno, y en determinadas ocasiones da la impresión de que cada investigador y cada escuela o punto de vista de investigación se fija sólo en sus propias pruebas, pareciendo que nos hallamos ante un diálogode sordos, como indica Geoffry A. Clark134. Pero tenemos la confianza de que nuevos avances en las diferentes ciencias nos harán avanzar y se irá desvelando el misterio.
130 Cfr. Howel, W.W., Journal of Human Evolution, 5, 477, 1976; Vandermeersch, Bernard, «Homo sapiens sapiens: lo que dicen los fósiles», Mundo Científico, 1995, n.º 160, pp. 710-716.
131 Cfr. Cann, R.I. et Al., Nature, 325, 31, 1987; Wilson, Allan C. y Cann, Rebecca L., «Origen africano reciente de los humanos», Investigación y Ciencia, 1992, junio, recopila-do por Jaume Bertranpetit (comp.), o.c., 20-25.
132 Cfr. Wainscoat, J. S. et al., Nature, 319, 491, 1986.133 Cfr. el art. cit., p. 716. En la pág. 714, nos muestra con sendos esquemas las tres teo-
rías que acabamos de presentar. 134 Cfr. Clark, G. A., «Origen del hombre: un diálogo de sordos», Mundo Científico,
1994, n.º 146, 462-467.

180
© Universidad de Deusto - ISBN 978-84-9830-383-4
Lo que está claro es que la vía de la biología molecular, con todas las insuficiencias que se quiera, está suponiendo una ventaja y unos avan-ces insustituibles. Además, si se tiene en cuenta el modo como funcio-nan el punto de vista genético y el morfológico (sólo éste en algunos ca-sos queda fosilizado), pueden compaginarse las discrepancias de fechas, como advierte de forma inteligente Luca Cavalli Sforza. Así, la fecha de 190.000 años que aporta la evidencia genética y los 100.000 años que señala la datación paleoantropológica, pueden conjugarse de modo ra-zonable. En primer lugar, «conviene precisar, señala Cavalli-Sforza, que los 190.000 años de Wilson no se refieren al primer hombre moderno. Si acaso, al tratarse de mitocondrias, se refieren a la primera mujer, de ahí el nombre de Eva. En realidad, esos 190.000 años señalan un hecho muy particular: en una mujer tuvo lugar la mutación de un solo nucleótido, por lo que su hija resultó distinta de las demás. Los descendientes de aque-lla mujer han tenido destinos geográficos distintos, pero parece probable, aunque no es seguro, que la progenitora común vivió en África. Sus des-cendientes que salieron de África no se desperdigaron necesariamente jus-to después de la mutación. Al contrario, debió pasar bastante tiempo entre el origen de la mutación y la formación de un grupo de descendientes en algún punto (desconocido) del continente africano, que traspasó sus lími-tes y se adentró en Oriente Medio a través del istmo de Suez, o en Arabia a través del Mar Rojo. Puede que la fecha de la migración sólo fuera un poco anterior a la que se atribuye a los primeros cráneos hallados en Is-rael, y es muy probable que sea bastante más tardía la fecha en que tuvo lugar la mutación mitocondrial, que es el origen del árbol de Wilson. Las dos fechas, mitocondrial y arqueológica, son distintas, y la primera tiene que ser anterior a la segunda»135.
Un paso adelante para complementar la vía molecular y la paleoantro-pológica será poder extraer y analizar fragmentos del ADN de los huesos fósiles, como ya parece que se está intentando hacer con fósiles de nean-dertales136, para poder señalar de modo complementario su edad y otras características importantes. Y de ese modo, como señala R. Lewin, «los datos moleculares y los datos fósiles se habrán convertido finalmente en aliados. La antropología molecular todavía tiene prometedoras perspecti-
135 Cfr. Cavalli-Sforza, Luca y Francesco, ¿Quiénes somos? Historia de la diversidad humana, o.c., p. 83. Cfr. Cavalli-Sforza, L., «Genes, pueblos y lenguas», Investigación y Ciencia, enero 1992, recopilado en J. Bertranpetit (comp.), o.c., 49-56; Lázaro Alcay, Car-los, «Luigi Luca Cavalli-Sforza: Genealogía de la humanidad», Mundo Científico, 1994, n.º 144, 232-239.
136 Cfr. Sanpedro, Javier, «¿Tenemos sus genes?», El País, 10-12-2006 (Domingo), pp. 1-3; Id., «El proyecto genoma Neandertal arranca con un millón de “letras” de ADN de un fósil», El País, 16-11-2006, p. 40.

181
© Universidad de Deusto - ISBN 978-84-9830-383-4
vas ante ella»137. Prometedoras perspectivas y difíciles problemas que re-solver todavía.
3. LA DISPUTA ENTRE EL POLIGENISMO Y EL MONOGENISMO
En relación con la dimensión filogenética del género humano, se sus-citó en su momento la cuestión acerca del origen del primer ser humano, es decir, si provenimos de un único filum evolutivo, o de varios (polifi-letismo o monofiletismo). Y si dentro de ese único filum, provenimos de una única pareja humana, o más bien de un grupo poblacional más amplio (monogenismo o poligenismo).
Esta problemática o disputa tiene tanto una vertiente científica como también religiosa. En un principio, se suscitó la cuestión en un contexto fundamentalmente religioso, en la medida en que la discusión acerca de si el género humano proviene de un único principio genético o de varios di-ferentes, parecía poner en cuestión el relato del Génesis sobre el origen de los humanos a partir de una única pareja, Adán y Eva. Hasta la aparición del paradigma evolutivo, no había problema ni discusión sobre este punto: se daba como un hecho indiscutible que todos los humanos descendíamos de Adán y Eva. De este modo, quedaba claramente resuelto el problema teológico sobre el pecado original, de tal modo que del pecado cometido por la primera pareja hemos quedado todos contagiados por el simple he-cho de ser descendientes biológicos de ella, y, por consiguiente, todos es-tamos necesitados de la redención de Jesucristo, redención que todos reci-bimos a través del bautismo, como medio e instrumento (sacramento) de filiación divina y de aplicación personal de la redención y del perdón del pecado original.
Pero la teoría de la evolución a través de la selección natural, propues-ta por Ch. Darwin, puso en tela de juicio los planteamientos monogenistas y, como consecuencia, los cimientos de la doctrina del pecado original y sus consecuencias teóricas.
La visión científica y anti-finalista de la evolución parecía implicar que se viera el tema de la hominización como fruto y consecuencia exclu-siva de las simples causas y procesos naturales. Y se pudo llegar a admitir, por parte de algunos, la posibilidad de que los humanos actuales no proce-diéramos de una única pareja original, sino de varias, que habrían existido en momentos diferentes. Ello explicaría la existencia de múltiples razas
137 «El nacimiento de la antropología molecular», o.c., 1231.

182
© Universidad de Deusto - ISBN 978-84-9830-383-4
dentro de la especie humana. Incluso cabía plantearse la cuestión de si te-nemos que seguir hablando de una única especie humana o de varias, tan-tas como razas existan.
Por tanto, aparte de las implicaciones religiosas, esta cuestión tiene también consecuencias científicas, filosóficas y sociológicas, en la medi-da en la que se pone en cuestión la consistencia del concepto de raza, y la unidad o no de la especie humana. Frente a una concepción unitaria de la especie humana, dentro de la cual las diferencias raciales se consideran accidentales, fue emergiendo en el siglo XIX una concepción ideológica y pseudo-científica de la especie humana dentro de la cual los diferentes co-lectivos raciales se entendía que provenían de diferentes especies antece-soras, dando lugar a un determinismo genético y biológico responsable de la desigualdad de capacidades individuales así como de las desigualdades sociales, culturales.
Dentro de esta problemática, hay que distinguir dos áreas de discusión: la referente a la polémica entre el polifiletismo y el monofiletismo, y la que se da entre el poligenismo y el monogenismo. En la primera se discute si todos los grupos poblacionales humanos, o razas, procedemos de la misma línea evolutiva (filum) o de varias diferentes; mientras que la segunda hace referencia a la cuestión de si, aun procediendo todos los humanos del mis-mo filum evolutivo, provenimos de una única pareja inicial o de varias.
3.1. ¿Polifiletismo o monofiletismo?
Esta disputa se inicia mucho antes de la propuesta de la teoría de la selección natural de Darwin, situándose a la base de las teorías racis-tas, que se venían imponiendo desde el siglo XVIII138. Hasta la revolución francesa, el racismo científico fue defendido sólo por una minoría, pues-to que, aunque se veían las diferencias entre naciones, culturas y grupos humanos, se atribuían las diferencias a factores sociales y culturales, de tal modo que, a lo largo del siglo XVIII, «la balanza naturaleza-cultura se inclinó siempre y claramente hacia el lado de la cultura»139. En esa época se aceptaba sin discusión los planteamientos monogenistas/monofiletistas
138 Cfr. Harris, Marvin, El desarrollo de la teoría antropológica. Una historia de las teorías de la cultura, Madrid, Siglo XXI, 1985 (5.ª ed.), cap. 4. «Apogeo y decadencia del determinismo racial»; Ruffié, J., De la biología a la cultura, Barcelona, Muchnik Edito-res, 1982, pp. 315 y ss.; Gould, S. J., La falsa medida del hombre, Barcelona, Crítica, 2007 (edición revisada y aumentada); Susanne, Ch./Rebato, E./Deligne, J., «Razas y racismo», en Rebato, E./Susanne, Ch./Chiarelli, B. (eds.), Para comprender la Antropología biológi-ca. Evolución y biología humana, Estella, Verbo Divino, 2005, pp. 655-664.
139 Harris, M., o.c., 71.

183
© Universidad de Deusto - ISBN 978-84-9830-383-4
del Génesis, de modo que se consideraba que las diferencias posteriores entre los humanos eran producto del proceso evolutivo, en el que los fac-tores del medio ambiental eran los determinantes. Eso no evitaba defen-der claramente posturas etnocéntricas, a favor en concreto de la suprema-cía de la raza blanca, como es el caso de científicos tan destacados como J. Blumenbach en Alemania, o G. L. Leclerc, conde de Bufón, en Francia. Todos ellos aceptaban el relato bíblico del origen único del género huma-no, explicando el curso degenerativo de la aparición posterior de las dife-rentes razas como fruto de condiciones ambientales140.
A pesar de que la opinión mayoritaria entre los científicos estaba a favor del monogenismo, también había autores que defendían el polige-nismo, basándose a su vez en autores del siglo anterior, como Isaac La Peyrère, autor de Preadamitae (1655), quien «mantenía que Adán era sólo el progenitor de los judíos, mientras que otros pueblos antiguos, como los caldeos, egipcios, los chinos y los mejicanos, descendían de antepasados preadamitas»141. En un siglo como el XVIII, en el que abundaban los ata-ques racionalistas a la religión, se acogían de buen grado estos ataques a la Biblia por parte de varios de los filósofos más relevantes del siglo XVIII.Así, Voltaire se reía de que los judíos, un grupo humano sin relevancia es-pecial, pretendieran considerarse como los antepasados de toda la huma-nidad. También defendieron posturas poligenistas David Hume, Henry James, Edward Long y Charles White.
En general, los defensores de las teorías poligenistas se inclinaban por el determinismo racial. Por el contrario, los monogenistas defendían la igualdad racial, como es el caso de Hunter y J. Herder, entre otros142.Hacia finales del siglo XVIII, la teoría poligenista se complicó y se mezcló con el problema de la esclavitud. En principio, parecería que los defenso-res de la esclavitud tendrían que haber sido partidarios del poligenismo, y los antiesclavistas, del monogenismo, pero, «como el historiador William Stanton ha demostrado (1960), el poligenismo, pese a ofrecer una justi-ficación racional para tratar a ciertos grupos humanos como animales de otra especie, jamás llegó a imponerse como ideología del esclavismo»143.
Pero donde se dio una defensa fuerte del poligenismo fue en el seno de la primera escuela de antropología americana, de la mano de L. Agassiz y S. G. Morton144. Morton era médico y profesor de anatomía en la Universi-
140 Cfr. ibídem, 72-73.141 Ibídem, 75.142 Cfr. ibídem, 75-76. 143 Ibídem, 77. 144 Para un estudio detenido de las teorías de Agassiz y de Morton, cfr. Gould, S. J., La
falsa medida del hombre, o.c., pp. 86-125.

184
© Universidad de Deusto - ISBN 978-84-9830-383-4
dad de Filadelfia, y estudiando de forma científica las capacidades cranea-les de diferentes razas (los resultados los presentó en tres libros que tuvie-ron una gran acogida: Crania Americana, 1840, Crania Aegyptiaca, 1844, y el epítome de toda su colección de cráneos, 1849), mostraba de forma contundente la superior capacidad cerebral de los blancos sobre las otras razas, siendo la negra la de menor capacidad. De tal modo que esos datos le llevaban a la conclusión de que cada raza había tenido un origen evoluti-vo diferente, conformando cada raza no una degeneración posterior tras un origen único, sino cada una de ellas una especie diferente, con su filogenia separada, producida desde hace varios miles de años. Ahora bien, como ha mostrado de forma clara y contundente S. J. Gould, en La falsa medida del hombre145, los prejuicios racistas de Morton le llevaron a falsear los datos, aunque no se pueda achacar a sus estudios ningún tipo de fraude científico.
Como hemos indicado más arriba, no hay una exacta correlación en-tre los partidarios del poligenismo y los planteamientos esclavistas, pues-to que, como indica William Stanton en su estudio citado, un autor como John Bachman, contemporáneo y adversario de las teorías de Morton, era también defensor de la esclavitud. Bachman era pastor luterano en Charleston (Carolina del Sur), y, aunque defendía el monogenismo, no lo consideraba opuesto a considerar la raza negra como inferior a la blanca, basándose en el pasaje bíblico de la maldición de Noé a su hijo Cam, su-puesto padre de la raza negra (Génesis, 9, 18-27). Así, pues, como indica Marvin Harris, por lo que le interesaba a Bachman «oponerse al polige-nismo no era, desde luego, por favorecer la abolición, sino, al revés, por salvar la justificación bíblica de la esclavitud»146.
La mayoría de los libros sobre Antropología cultural, tanto en Europa como en América, entre los años 1800 y 1849, se centraban en gran me-dida en la discusión entre el poligenismo y el monogenismo, de tal modo que la aparición de El origen de las especies de Ch. Darwin fue determi-nante para aclarar esta discusión. Tal como estaban planteadas, las dos posturas estaban equivocadas, puesto que, respecto a los monogenistas, si la humanidad tenía un antepasado común, no sería Adán sino un individuo (o pareja) de alguna especie de mono anterior. Y respecto a los poligenis-tas, ya no parecía que hubiera que apelar a diversos principios para las ra-zas humanas. Para el seguidor de Darwin, Thomas Huxley, la teoría de la selección natural había conciliado lo bueno de ambas posturas. Así, «los monogenistas conservaron su común humanidad y su progresismo; los po-ligenistas se vieron apoyados en su crítica a la Biblia y su cientifismo»147.
145 O.c., pp. 102 y ss. 146 Harris, M., o.c., p. 79. 147 Ibídem, p. 80.

185
© Universidad de Deusto - ISBN 978-84-9830-383-4
Pero no era tan evidente esta conclusión, puesto que James Hunt, pre-sidente de la Sociedad Antropológica de Londres, acusó a Th. Huxley de monogenista disfrazado, puesto que, en opinión de Hunt, la teoría de Darwin apoyaba mucho mejor que hasta entonces la postura de que las ra-zas humanas pertenecían a especies diferentes, e incluso algunas de ellas desaparecerían con el tiempo, debido al empuje de las más fuertes, dentro de la dinámica de la lucha por la vida y la supervivencia que se da entre los diversos colectivos vivientes.
Con el tiempo, se llegó a aceptar que todos los grupos humanos con-temporáneos pertenecían a una misma especie, pero quedaba todavía la cuestión de por cuánto tiempo han mantenido las razas dentro de la es-pecie una filogenia separada. Las discusiones del siglo XIX sobre este as-pecto han tenido su continuación en el siglo XX en autores como Ernest Hooton y Carlton Coon, que defienden que las diferentes razas proceden de líneas evolutivas diferentes, considerando que «los diferentes tipos ra-ciales han sufrido una evolución paralela en la transición de los australo-pitecos al homo sapiens. Debe señalarse que todos estos intentos de salvar la genealogía separada de los caucasoides son pura especulación en el mo-mento en que aducen antigüedades del orden de decenas de miles de años. Los fósiles no nos dicen nada del color de la piel, la sección del cabello, el tamaño o la forma de la nariz y de los labios o los pliegues epicánticos, que son precisamente los rasgos en los que se basan los constructos racio-lógicos tradicionales»148.
A pesar de todos estos flecos del poligenismo, es evidente que la teo-ría de la selección natural de Darwin supuso un avance fundamental para reconfigurar la discusión entre poligenistas y monogenistas, situando el problema en otros parámetros más serios y científicos, y ayudando a rom-per los estrechos márgenes de discusión reducidos a aceptar o no de modo literal el relato bíblico del Génesis. A partir de Darwin y de los avances científicos en diferentes áreas de la Antropología físico-biológica, el con-cepto ideológico de raza fue sufriendo un deterioro determinante, en la medida en que, si hasta entonces los rasgos diferenciadores de cada raza eran aspectos fenotípicos y superficiales, los avances de la genética y de la biología (entre otros, los avances en la investigación sobre los grupos sanguíneos, y, sobre todo, la común estructura genética de todos los hu-manos) fueron mostrando de forma evidente el común origen genético y evolutivo de los colectivos humanos actuales, por lo que se demuestra de modo evidente que los humanos formamos una única especie, siendo las diferencias raciales sólo elementos accidentales y secundarios.
148 Ibídem, p. 81.

186
© Universidad de Deusto - ISBN 978-84-9830-383-4
Uno de los últimos reductos del polifiletismo, serían las tesis más ac-tuales del denominado policentrismo, defendido por algunos paleoantro-pólogos para explicar el origen de la pluralidad de las poblaciones huma-nas actuales. Ya vimos en otra parte anterior de este capítulo, las dos tesis contrapuestas (desde la biología molecular y la paleoantropología) sobre el origen del homo sapiens sapiens: la teoría que apuesta por una única oleada expansiva (la segunda, tras la del homo erectus) que se extendió por todo el planeta, imponiéndose sobre los restos del hombre de Nean-dertal (en Europa) y del homo erectus en Asia, frente a la teoría policén-trica o del candelabro, partidaria de considerar que el hombre actual es el resultado de la mezcla de varios grupos poblacionales, que habrían tenido diversos saltos mutacionales en distintos lugares geográficos, y se habrían ido mezclando genéticamente entre ellos, explicándose de ese modo la pluralidad de rasgos morfológicos de los hombres actuales. Pero también quedó clara la postura de los estudiosos de la biología molecular, indican-do que no parece existir ninguna evidencia de restos genéticos en las po-blaciones humanas actuales, ni de los neandertales (en esto no hay tanta claridad) ni del homo erectus. Eso no demuestra que no se hubiera dado en algún momento de la historia pasada cruces e intercambios genéticos. Pero lo que sí parece demostrarse es que, hasta el momento (y según lo que la ciencia actual sabe sobre los análisis del ADN fósil), esos posibles cruces genéticos no han dejado ninguna huella significativa en nuestra es-pecie149.
En resumen, como afirma J. Ruffié, «en el plano biológico, hoy en día no hay nada que nos autorice a descomponer la especie humana en ra-zas autónomas. Pero para muchos antropólogos es difícil renunciar a este concepto»150. El mismo Ruffié hace referencia, como ejemplo de estas reticencias, al caso de H. V. Vallois, quien en 1950 respondía a un cues-tionario de la UNESCO diciendo: «La existencia de la raza es un hecho incuestionable», aunque matizaba esta afirmación señalando la existencia de dos tipos de razas, las primarias («grandes razas o grupos raciales») y las secundarias («razas de segundo orden»). Las razas primarias, según Vallois, podrían haberse convertido en especies autónomas «si el aisla-miento que imperaba en el momento de su formación hubiera persistido… Actualmente no se puede considerar que hayan alcanzado este estadio. Dado el estado actual de las cosas se puede presumir incluso que seguirán una evolución inversa»151.
149 Cfr. Ayala, F. J./Cela Conde, C. J., La piedra que se volvió palabra. Las claves evo-lutivas de la humanidad, o.c., pp. 111 y ss.
150 O.c., p.322.151 Citas tomadas de J. Ruffié, o.c., 322-323.

187
© Universidad de Deusto - ISBN 978-84-9830-383-4
A pesar de todas las evidencias científicas, ha costado mucho aban-donar el uso y las aplicaciones del concepto de raza, advirtiéndose que «los últimos defensores de la raza fueron casi siempre los morfólogos, mientras que los genéticos siempre se han situado entre los defensores de las poblaciones. Dado el progreso de la genética humana, hoy en día ningún biólogo admite la existencia de razas en la especie humana; al-gunos aún utilizan el término, pero la mayoría lo despojan de su sentido zoológico»152. Ante la extrañeza de la persistencia del racismo y del uso de la idea de raza, sólo podemos entender esa contumacia por el peso de la costumbre y, sobre todo, por los intereses sociológicos que el mito del racismo permitía mantener, como fue el caso del nazismo. A consecuencia de los horrores nazis, la UNESCO organizó cuatro conferencias interna-cionales de antropólogos (entre 1950 y 1967), llegándose a la conclusión unánime de que el concepto de raza ya no puede ser usado para referirse a la especie humana: «en el hombre, la raza es más un mito social que un fenómeno biológico»153.
En conclusión, en la disputa entre el polifiletismo y el monofiletismo, las evidencias científicas se inclinan clara y rotundamente hacia el mono-filetismo. Los datos biológicos y genéticos nos indican que el género hu-mano, tanto si nos referimos a su origen en su primera especie de homohabilis, como a nuestra especie, homo sapiens sapiens, posee un único origen filético en ambos casos, constituyendo las tradicionales diferencias raciales meros aspectos secundarios, que no pueden servir para deducir la diversidad específica de sus diferentes grupos poblacionales, ni para con-cluir de ello diferencias en las capacidades intelectuales y culturales, ni menos aún para mantener cualquier tipo de privilegios o algún tipo de di-ferencias en derechos e igualdad de oportunidades ante las leyes sociales.
3.2. ¿Poligenismo o monogenismo?
La verdadera discusión sobre el poligenismo-monogenismo se centra en la actualidad en dilucidar si el origen de la especie humana, o mejor, del género humano se debe a una pareja humana central (monogenismo) o más bien a un colectivo humano más amplio con el que se habría dado el tránsito de la especie anterior a la humana (poligenismo). Como pue-de suponerse, las pruebas para demostrar la verdad o no de cada una de las dos posturas no pueden provenir de la paleoantropología, puesto que
152 Ruffié, J.; o.c., 323.153 Ibídem, p. 324.

188
© Universidad de Deusto - ISBN 978-84-9830-383-4
tales pruebas no dejan rastros fósiles, teniéndose que recurrir al análisis de cómo entienden los biólogos y genetistas que se produce el origen de una especie; esto es, el salto de una especie madre a otra, u otras espe-cies hijas.
La idea de especie, como quedó indicado en su momento, ha sido ob-jeto de múltiples discusiones entre los científicos, habiéndose pasado de un concepto morfológico a otro genético. Los individuos de una misma especie pueden realizar intercambios sexuales fértiles, cosa que no pue-den hacer los que pertenecen a especies diferentes. Hay casos especiales en que, aunque pueden tener descendientes, éstos son estériles (los mulos, fruto del intercambio entre caballos y asnos). Si ese es el criterio de dife-renciación, la prueba de si dos individuos pertenecen a la misma especie o no, no puede determinarse a priori, sino a posteriori. De ahí que cuando hablamos de los diferentes tipos de homo (habilis, erectus, etc.), no poda-mos saber de forma taxativa si eran especies diferentes o simples grupos poblacionales diferentes pero de la misma especie. Hablamos, por ello, unas veces de género humano (con varias especies distintas dentro), y otras, de una única especie humana.
El problema está en cómo se produjo el salto de una especie pre-huma-na (la última de las especies de los australopitecos) a la primera humana (homo habilis), o el origen del homo sapiens sapiens (hombre de Cro-mañón) a partir de la especie (o sub-especie) anterior (¿homo antecesor,homo heidelbergensis?). En cualquiera de estos casos, al igual que si nos refiriéramos al origen de cualquier especie animal que se origina por mezcla sexual, el proceso consistente en el salto de una especie a otra no parece realizarse a partir de una única pareja sexual, que experimenta el adecuado salto mutacional en su cariotipo, para transmitirlo después a sus descendientes, dando así lugar a una nueva especie.
En primer lugar, ya vimos en su momento154 que es poco probable que se den macromutaciones repentinas que den lugar, en períodos cortos de una o dos generaciones, a nuevas especies, siendo lo más normal y evi-dente el proceso gradualista de acumulación de pequeñas mutaciones, que suponen la introducción de una deriva genética dentro de un grupo pobla-cional en la especie origen, para ir poco a poco produciéndose un proceso de aislamiento, normalmente de tipo geográfico, para luego producirse un aislamiento genético. Esa deriva genética, orientada a formar una nueva especie, puede tener en algunos casos vuelta atrás, pero lo normal es que se consolide la diferencia para terminar formando una especie diferente.
154 Cfr. capítulo 1.º, apartado 4.2. El redescubrimiento de la genética mendeliana, don-de De Vries defendía la existencia de macromutaciones como causa de la aparición brusca de nuevas especies.

189
© Universidad de Deusto - ISBN 978-84-9830-383-4
Por todo esto, lo que resulta más evidente, tanto para los humanos como para cualquier otra especie animal, es que el proceso de formación de la nueva especie no se pueda producir por el sistema de un estrecho cuello de botella (situación en la que quedan de una especie unos pocos individuos, o incluso una sola pareja), ya sea que ese fenómeno lo enten-damos como consecuencia de una catástrofe natural o social, que aísle a esa pareja, o sea un fenómeno de tipo genético (producto de una macro-mutación repentina), para dar después origen al resto de la nueva especie. Las investigaciones genéticas de algunos estudiosos apuntan a considerar como totalmente necesaria la presencia de un colectivo de al menos varias decenas o centenares de individuos como condición necesaria para poder generarse una nueva especie.
Seguimos en estos planteamientos las investigaciones de F. J. Ayala, Jan Klein y Naoyuki Takahata sobre la estructura genética del sistema in-munitario humano y sus repercusiones sobre el problema del poligenis-mo/monogenismo155. Estos científicos tratan de mostrarnos que, frente a la hipótesis de que nuestra especie pueda haberse originado a partir de un grupo poblacional más bien pequeño, las evidencias científicas que ellos manejan nos sitúan en un escenario en el que se necesita un colectivo po-blacional compuesto por varias decenas de miles de personas. Sus inves-tigaciones están basadas en estudios «sobre la evolución de los genes que controlan la capacidad del sistema inmunitario para reconocer las proteí-nas foráneas»156.
Para entender esto tenemos que ver cómo funciona el sistema inmu-nitario en los seres vivos, como sistema que vigila la intromisión de elementos extraños en un sistema vivo compuesto por células que po-seen un cariotipo idéntico. Los primeros avances en esta dirección los realizó Peter A. Gorer, al descubrir en las células de todos los organismos unos marcadores moleculares por los que se distinguen unos individuos de otros dentro de una misma especie, determinando de este modo lo que se denomina compatibilidad hística (compatibilidad entre unos tejidos ce-lulares con otros), condición fundamental para poder realizarse los tras-plantes de órganos. Gorer mostró que las moléculas que desempeñan la función principal en esta compatibilidad son «un largo grupo de loci ge-néticos que ocupan una región de un cromosoma: el complejo principal de histocompatibilidad (MHC)»157. Su función consiste en iniciar un proceso de reacción contra los parásitos.
155 Cfr. Klein, J./Takahata, N./Ayala, F. J., «Polimorfismo del MHC y origen del hom-bre», Investigación y Ciencia, 1994, n.º 215, 14-19.
156 Ibídem, p. 14. 157 Ibídem, 14.

190
© Universidad de Deusto - ISBN 978-84-9830-383-4
«El MHC humano, denominado complejo antigénico linfocitario hu-mano (HLA), contiene más de 100 genes y ocupa una región cromosómica de más de 4 millones de pares de bases»158. El hecho de que se produzcan rechazos de tejidos trasplantados, lleva a concluir que cada individuo de cada especie posee moléculas MHC distintas. Y estos complejos de mo-léculas se van sucediendo y complejificando en el proceso evolutivo, de tal modo que sólo una fracción de este tipo de sistemas y combinaciones po-sibles se dan en las poblaciones humanas. Por tanto, nos encontramos ante un hecho claro: «la gran diversidad de la secuencia de nucleótidos de los diferentes alelos MHC»159. De unos genes determinados de una especie se han ido formando otros, en línea evolutiva, complejificándose el proceso y pudiéndose seguir la filogenia de los alelos de un gen presente en una espe-cie a través del procedimiento de la coalescencia. Cualquier rasgo genético actual (sea el sistema inmunológico o los grupos sanguíneos) se derivan de otros que les precedieron, «de tal forma que si rastreamos su historia hacia atrás coalescen en un alelo inicial único, el “cenancestro”. El cenancestro es el gen del que descienden todos los alelos presentes en un locus preci-so dentro de las poblaciones actuales. La existencia de un solo cenancestro para cada gen no implica que no existieran alelos diferentes de otros indi-viduos coetáneos con el cenancestral; lo que se está diciendo es que esos otros alelos no tienen descendientes en la humanidad actual. Los diversos alelos que se encuentran en los humanos de hoy día habrían surgido a tra-vés del tiempo por medio de las mutaciones. La idea tampoco implica que el individuo en el que se daba el alelo ancestral fuera el único individuo del que descienden todos los humanos actuales»160.
Ayala y Cela Conde están refiriéndose con estas apreciaciones a las equivocadas conclusiones que determinados comentaristas sacaron so-bre lo que se denominó la Eva mitocondrial o Eva africana161. Tras los análisis de la biología molecular al reconstruir la filogenia del mtDNA (ADN mitocondrial), los científicos llegaron a identificar un cenances-tro, situado en una mujer que habría vivido en África hace entre 150.000 y 200.000 años, siendo ese cenancestro común a todos los seres humanos actuales. Por ello, se recurrió al nombre de Eva, en referencia al relato del Génesis. Pero, en coherencia con lo que Ayala y Cela Conde nos indican, no se trata de que esta mujer ancestral fuera la única mujer ancestral de todos nosotros, los humanos actuales. Como ambos autores nos indican,
158 Ibídem, p. 15. 159 Ibídem, p. 15. 160 Ayala, F. J./Cela Conde, C. J., o.c., p. 114 (las cursivas son de los autores). 161 Cfr. el final del capítulo primero del presente libro, donde se habla de las investiga-
ciones de la biología molecular en relación sobre los orígenes del hombre actual.

191
© Universidad de Deusto - ISBN 978-84-9830-383-4
«el error surge de la confusión entre genealogías de genes y genealogías de individuos. La “Eva mitocondrial” de la que descienden todos los mtDNA actuales era una molécula de ADN, no una mujer»162.
Se puede, por tanto, utilizar la teoría de la coalescencia sobre otros rasgos genéticos, como es el caso del sistema inmunológico, comparando los sistemas humanos con los de los simios y otras especies vivas. Los re-sultados ilustrativos que arrojan estas investigaciones nos hacen ver que «algunos alelos humanos son más diferentes entre sí de lo que lo son los alelos de chimpancés, macacos y otros simios. Esto implica que tales ale-los existían ya mucho antes del origen de los homínidos»163.
Retomando la referencia a los estudios sobre el sistema inmunológico humano, nos decían Klein, Takahata y Ayala que el MHC humano (com-plejo antigénico linfocitario humano: HLA) contiene más de cien genes. Estos científicos se han dedicado a estudiar el gen funcional DRBI, des-cubriendo que tiene un cenancestro que vivió hace unos 35 millones de años, mucho antes de que existiera el género humano, si tenemos en cuen-ta que, como sabemos, el homo habilis apareció aproximadamente hace dos millones y medio de años.
Carecemos de información suficiente sobre el número mínimo de in-dividuos que se exige para componer el grupo fundador de una especie. «Pero el polimorfismo MHC ofrece la posibilidad de estudiar indirecta-mente la especiación: si se conociera el número de alelos MHC que una especie ha heredado de su antecesora (así como la tasa de mutación y la intensidad de la selección) podría calcularse el tamaño de la población fundadora»164.
En función de esas variables, los autores nos indican que «a partir del número conocido de alelos MHC-DRBI existentes, nosotros calcula-mos un tamaño eficaz a largo plazo de la población humana, en los úl-timos 0,5 millones de años, del orden de 100.000 individuos. Los cómpu-tos realizados por otros loci MHC humanos suelen dar valores similares. Y también se indican grandes tamaños de población para otras especies de primates cuyo polimorfismo MHC está bastante bien definido»165. Los mismos autores son conscientes de que ha podido haber circunstancias en las que, debido a catástrofes o situaciones de epidemias o hambrunas, se haya podido producir una reducción drástica de población, hasta llegar a formarse hipotéticos cuellos de botella genéticos. Incluso puede darse el caso de que un pequeño grupo de población emigre y se separe del grupo
162 Ayala, F. J./Cela Conde, C. J., o.c., p. 118163 Ibídem, pp. 118-119. 164 Klein, J./Takahata, N./Ayala, F. J., o.c., 17.165 Ibídem, p. 17.

192
© Universidad de Deusto - ISBN 978-84-9830-383-4
de origen, pudiendo desde el nuevo ámbito donde se asentara dar lugar a una nueva especie. Partiendo de esta hipótesis, los autores han desarrolla-do diversas simulaciones en el ordenador, llegando a la conclusión de que «la población fundadora de una especie no puede estar formada por me-nos de 500 individuos reproductores»166.
Estas apreciaciones se refieren a casos hipotéticos en que se darían fe-nómenos que, como decimos, se suelen denominar cuellos de botella ge-néticos. Por tanto, estos estudios contradicen las teorías que atribuyen los inicios de una especie a un grupo pequeño de fundadores. Más bien ocurre que «el polimorfismo MHC excluye rotundamente la posibilidad de que las poblaciones humanas deriven de un solo individuo, de una Eva»167.Por el contrario, las evidencias científicas se inclinan a pensar que lo nor-mal es que estos parámetro sobre el grupo originante de una especie, tam-bién se tienen que aplicar al caso humano, deduciéndose que tuvo que estar compuesto al menos por 500 individuos reproductores, y «probable-mente nunca menos de 100.000. El origen de los homínidos, como el ori-gen del Homo sapiens, no fue consecuencia de un cuello de botella extre-mo, durante el cual nuestros antepasados se redujesen a una sola pareja o a muy pocos individuos»168.
No cabe duda de que se trata de investigaciones científicas que tienen que ser contrastadas y completadas, pero todo parece apuntar a que, inde-pendientemente del número mínimo de individuos que se quiera postular o que las investigaciones científicas posteriores nos lleguen a demostrar, la tesis de una única pareja como originante del género humano o de la especie Homo sapiens no parece tener mucha cabida razonable, puesto que no tiene sentido apelar a esta tesis en lo humano cuando en los oríge-nes de las demás especies vivas las apreciaciones científicas van por otro lado. Por lo tanto, en la disputa entre el poligenismo y el monogenismo, todas las evidencias apuntan a dar la razón al poligenismo.
La cuestión que nos queda plantear es si con esto se socavan las tesis teológicas sobre la interpretación de la Biblia y sobre el pecado original. Y la respuesta es que no. Siguiendo las tesis que hemos propuesto en el capítulo anterior sobre la relación entre la dimensión científica, filosófi-ca y teológica de una teoría, tenemos que dar autonomía a cada uno de los diferentes niveles epistemológicos de reflexión, y dejar que las diferentes ciencias investiguen estos asuntos con total libertad.
Pero, junto a esto, hay que ser conscientes de que los conflictos de las tesis poligenistas con la interpretación de la Biblia y la doctrina del peca-
166 Ibídem, p. 18. 167 Ibídem, p. 18.168 Ayala, F. J./Cela Conde, C. J., o.c. p. 119.

193
© Universidad de Deusto - ISBN 978-84-9830-383-4
do original, sólo se dan desde un determinado y ya superado modo de en-tender tanto los estudios bíblicos como el teologúmeno del pecado origi-nal, como vamos a ver a continuación.
En relación a las interpretaciones bíblicas, los estudios críticos, co-menzados ya en la segunda mitad del siglo XIX, han experimentado avan-ces tan importantes como para que en la actualidad se acepte mayorita-riamente entre los estudiosos de la Biblia que no se trata de relatos con intencionalidad histórica, similares a como en la actualidad se entienden los estudios históricos, sino que se trata de textos con intencionalidad teo-lógica. Sólo unas minorías fundamentalistas se aferran en la actualidad a una interpretación literal e historicista de los textos bíblicos.
Las referencias en el Génesis a Adán y Eva proceden de dos relatos dife-rentes: uno de ellos pertenece a la tradición sacerdotal (Gen., 1, 26-30; 5, 1-5) y el otro, a la tradición yahvista (Gen., 2,4-4,2.25.26). Según algunos estu-diosos de la Biblia, el personaje Adán es más bien visto como un persona-je colectivo (como especie) en el texto de la tradición sacerdotal, mientras que en el yahvista es visto más bien como un individuo singular169.
Pero en ambos casos los personajes de Adán y Eva no son figuras his-tóricas, sino simbólicas y teológicas, no teniendo sentido discutir sobre su antigüedad o sus rasgos característicos e históricos dentro de los estudios científicos sobre el proceso evolutivo.
Claro que esta forma de pensar se ha ido imponiendo, dentro del cam-po teológico y del magisterio eclesial, muy poco a poco. El primer Papa que se mostró receptivo a estos estudios críticos, y se refirió al tema del poligenismo, fue Pío XII, quien, reflexionando sobre la conjugación de las verdades científicas y la verdad bíblica en relación con la evolución y el poligenismo, indicaba que «no se ve claro cómo tal sentencia pueda compaginarse con lo que las fuentes de la verdad revelada y los documen-tos del magisterio de la Iglesia enseñan acerca del pecado original, que procede del pecado verdaderamente cometido por un solo Adán y que, di-fundiéndose a todos los hombres por la generación, es propio de cada uno de ellos»170.
Lo que se quiere decir es que de suyo la hipótesis sobre la alternativa monogenismo/poligenismo, en cuanto se centra en tratar de explicar bio-lógicamente el origen de la especie humana, no pertenece al ámbito de la
169 Cfr. Camps, Guíu, «Los datos bíblicos y en el número de los primeros progenito-res», en Crusafont Pairo, M./Meléndez, B./Aguirre, Emiliano de (dirs.), La evolución, Ma-drid, BAC, 1986, pp. 938-957; R. Lavocat/H. Cazelles, «Polygènisme», en Suplément au Dictionnaire de la Bible, París, 1967, col. 90-110.
170 Encíclica Humani Generis, 1950, en DS, 3897 (Documentos bíblicos, n. 703): cita tomada de Camps, G., o.c., p. 944.

194
© Universidad de Deusto - ISBN 978-84-9830-383-4
fe. Pero, en cuanto pueden derivarse implicaciones importantes para la teología, es un tema pertinente para ella, puesto que se enfrenta a la ver-dad sobre el pecado original cometido por nuestros primeros padres y transmitido a todos sus descendientes humanos. Por tanto, si se viera que la tesis poligenista puede compaginarse con las verdades de fe implicadas, no habría inconveniente en que los creyentes pudieran apoyar esta hipóte-sis como compatible con las afirmaciones teológicas.
Ya hemos visto que hay que distinguir entre la alternativa entre poli-filetismo/monofiletismo y entre poligenismo/monogenismo. En relación a la primera alternativa, esto es, si los humanos provenimos de un úni-co origen filético o de varios, ya hemos visto que hoy día es aplastante la mayoría de científicos que defienden el monofiletismo, desautorizando cualquier planteamiento a favor de las teorías racistas basadas en una plu-ralidad de orígenes filéticos. Por tanto, aquí habría total sintonía entre la ciencia y la fe. Pero, en relación a la alternativa poligenismo/monogenis-mo, vimos que todas las evidencias apuntan a un origen del género huma-no en el que no tiene sentido hablar de una primera pareja humana desde la que se habrían derivado los demás de la especie o especies posteriores, sino más bien de un colectivo originante amplio.
Según estos planteamientos, y de acuerdo con la interpretación bíbli-ca encaminada a entender las figuras de Adán y Eva no tanto como per-sonajes históricos sino simbólicos, no habría problema en compaginar la verdad científica con la teológica si entendemos las figuras de Adán y Eva como una representación simbólica y colectiva de la especie humana. La dificultad estaría en cómo entender la teología del pecado original. Pero tal dificultad sólo existe si seguimos entendiendo el relato del Génesis de un modo literal e histórico.
La opinión más amplia entre los teólogos estaría reflejada en estas pa-labras de W. Pannenberg: «La tesis de la universalidad y radicalidad del pecado, que es el fundamento cristiano del pecado original, no precisa de la hipótesis auxiliar del llamado monogenismo, o sea, el origen de todos los hombres a partir de una única pareja, que, al mismo tiempo, es la au-tora del pecado humano»171. Los teólogos, animados por la jerarquía ecle-sial, buscan una reinterpretación más adecuada a la doctrina del pecado original, que supere la doctrina tradicional, nacida con San Agustín al in-terpretar algunas afirmaciones de San Pablo.
171 Cfr. Pannenberg, W., Antropología en perspectiva teológica, Salamanca, Sígueme, 1993, p. 173. Cfr. también Feiner, J., Mysterium Salutis, II (1967), pp. 562-581; 564; Seyes, J: A., Antropología del hombre caído. El pecado original, Madrid, BAC, 1991; Concilium, 2004, n.º 304 (número monográfico sobre el pecado original: «El pecado origi-nal: ¿un código de falibilidad?»).

195
© Universidad de Deusto - ISBN 978-84-9830-383-4
La teología actual reinterpreta la doctrina del pecado original172, en clave más social que individual, y dentro de una visión renovada de la realidad de Dios, del hombre y de la teología en general. González Faus resume su reinterpretación del pecado original en cuatro grandes afirma-ciones: la primera sostiene que «el ser humano está moral e interiormente deteriorado: se encuentra ante el mal en un estado que no es de sola ex-posición o vulnerabilidad, sino de debilidad e inferioridad»; en segundo lugar, «este deterioro es obra del hombre mismo: de la estructuración del pecado a todos los niveles de la vida humana»; en tercer lugar, «este dete-rioro comienza ya con el origen de la historia humana» (sea como sea el modo de entender ese origen); y en cuarto lugar, «ese deterioro contradice la voluntad de Dios sobre el hombre»173. Estas afirmaciones fundamen-tales han sido expresadas en el relato bíblico, y en los diversos escritos de la tradición eclesial, con un ropaje propio de cada época, estando impreg-nados los diversos relatos con múltiples elementos míticos que envuelven las convicciones esenciales de fondo (referencias al paraíso, a los persona-jes de Adán y Eva, al limbo, etc.), siendo necesario separar esas verdades fundamentales de los ropajes culturales y míticos propios de una mentali-dad que ya no es la nuestra, y que, por tanto, más que ayudar a entender nuestra fe, en muchos casos la estorban y nos desvían de su adecuada in-terpretación.
De este modo, vemos que los conflictos entre la ciencia y la fe, en vez de significar la ruptura entre ambas perspectivas, sirven para su mutuo enriquecimiento. De este modo, no se ve que, en principio, los avances de las diferentes ciencias sobre el origen de la humanidad puedan llevar a conflictos graves con la fe cristiana, que, guiada por la revelación bíbli-ca, nos hace ver que la Biblia, como ya indicaba San Agustín, no nos dice cómo es el cielo sino cómo tenemos que vivir para llegar a él.
172 Para una reinterpretación amplia y completa del pecado original, cfr. González Faus, J. I., Proyecto de hermano. Visión creyente del hombre, Santander, Sal Terrae, 1987, cap. 6.º «El pecado original», pp. 299-386; Ruiz de la Peña, J. L., Qué hay del pecado ori-ginal, Madrid, Fundación Santa María, 1992; Fernández, Domiciano, El problema del pe-cado original, Madrid, Nueva Utopía, 1999; Vidal, Marciano, Pecado estructural y respon-sabilidad personal, Madrid, Fundación Santa María, 1991.
173 Ibídem, pp. 365-366.


197
© Universidad de Deusto - ISBN 978-84-9830-383-4
Capítulo 4
Las transformaciones genéticas y morfológicas y la cuestión del sentido (azar o finalidad) de la evolución
1. LAS TRANSFORMACIONES GENÉTICAS Y MORFOLÓGICAS
En capítulo anterior hemos hecho referencia al proceso de hominiza-ción desde el punto de vista diacrónico, recogiendo los datos que tanto la paleoantropología como la biología molecular poseen sobre los pasos producidos en el proceso evolutivo hasta desembocar en el hombre ac-tual. Ahora nos vamos a detener en estudiar qué cambios se han ido pro-duciendo tanto en la estructura morfológica como comportamental desde el primer grupo humano hasta configurar al ser humano tal como es en la actualidad. Es decir, ser trata de preguntarnos y de ir mostrando por qué los seres humanos somos como somos, y cuáles han sido los diversos ava-tares que nos han llevado a ser así.
Aceptado como paradigma científicamente probado la teoría de la evolución, explicada desde la selección natural darwiniana, parecería que el ser humano, al igual que el resto de las especies vivas tendría que ser fruto de las mutaciones azarosas y la lógica de la selección natural, sin que el propio ser humano actúe en ningún momento como sujeto de su propia configuración. Pero vamos a ver que eso no fue así. El ser hu-mano ha sido en parte sujeto y objeto de su propio desarrollo y consti-tución. Se han imbricado en esa larga tarea dos procesos complementa-rios: el de hominización y el de humanización. Entendemos por proceso de hominización al conjunto de transformaciones que los seres humanos

198
© Universidad de Deusto - ISBN 978-84-9830-383-4
han ido experimentando desde que se dio la primera mutación genéti-ca que inició la separación de la última especie pre-humana, hasta el último paso que ha terminado por conformarnos morfológicamente tal y como somos en la actualidad los seres humanos. En cambio, podría-mos denominar proceso de humanización al conjunto de transformacio-nes comportamentales, conductuales y culturales que nos han configurado como animales racionales y culturales, única especie en su género. En la medida en que la especie humana (o las diferentes subespecies del lina-je humano) se halla dotada de consciencia, racionalidad y libertad, esto es, que no está fijada su estructura comportamental por su herencia ge-nética, no tiene más remedio que hacerse a sí mismo, en diálogo con el entorno ecológico como también con el resto de compañeros de especie. Y esa tarea comenzó ya desde el primer instante de su existencia como género humano.
Ahora bien, las ciencias que nos acercan a nuestros orígenes, nos han enseñado a darnos cuenta de que estos dos procesos, de hominiza-ción y de humanización, no se suceden separadamente el uno al otro, de modo que el segundo empieza sólo cuando el primero ha terminado su tarea, sino que se imbrican recursivamente, se implican mutuamente, de modo que un avance en uno de ellos produce como consecuencia avan-ces en el otro. Y ese proceso de progresiva influencia imbricatoria es el que nos ha ido constituyendo como seres humanos, como vamos a ir viendo. Por tanto, es fundamental, a la hora de entender el proceso que nos ha ido constituyendo como humanos, que veamos todo este aconte-cer en su conjunto, abarcando todos los elementos que configuran esta estructura antropogenética.
El conjunto de todos los elementos que han conformado el proceso antropogenético, estaría ordenado como el siguiente esquema:
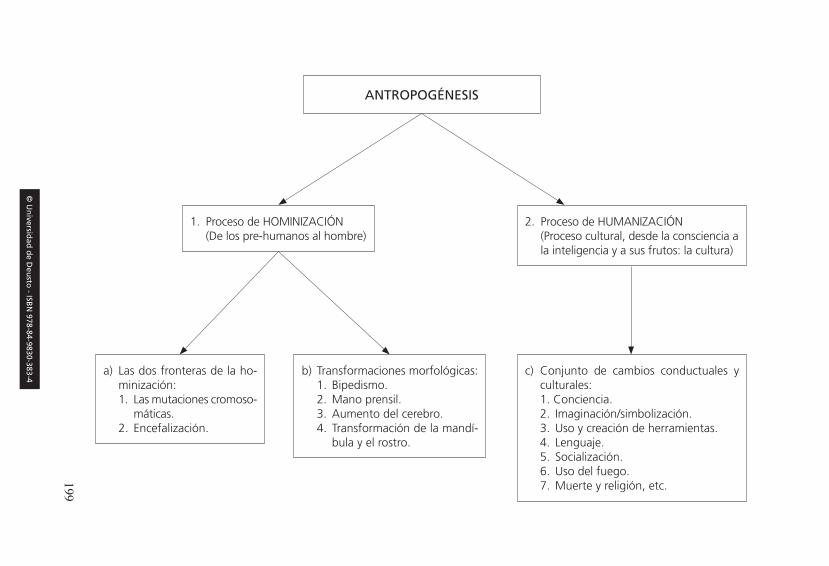
199
© U
niversid
ad d
e Deu
sto - ISB
N 978-84-9830-383-4
ANTROPOGÉNESIS
1. Proceso de HOMINIZACIÓN(De los pre-humanos al hombre)
2. Proceso de HUMANIZACIÓN(Proceso cultural, desde la consciencia a la inteligencia y a sus frutos: la cultura)
a) Las dos fronteras de la ho-minización:1. Las mutaciones cromoso-
máticas.2. Encefalización.
b) Transformaciones morfológicas:1. Bipedismo.2. Mano prensil.3. Aumento del cerebro.4. Transformación de la mandí-
bula y el rostro.
c) Conjunto de cambios conductuales y culturales:1. Conciencia.2. Imaginación/simbolización.3. Uso y creación de herramientas.4. Lenguaje.5. Socialización.6. Uso del fuego.7. Muerte y religión, etc.

200
© Universidad de Deusto - ISBN 978-84-9830-383-4
2. LAS DOS FRONTERAS DE LA HOMINIZACIÓN
La seña más explícita que distingue a los seres humanos del resto de los seres vivos es, sin duda, su inteligencia y su capacidad de ser cons-ciente de sí y de sus acciones. De ahí que sea el proceso de encefalización el aspecto central en el que hay que centrarse a la hora de entender el pro-ceso antropogenético. Pero esta progresiva encefalización está posibilita-da, como todo cambio morfológico, por nuestra base genética, nuestro ca-riotipo. Por eso, el primer paso en la hominización hay que situarlo en los primeros saltos mutacionales que empezaron a hacernos genéticamente diferentes de nuestros antecesores. Así, pues, las dos aunténticas fronteras de la hominización, como señala Jacques Ruffié1, son los saltos mutacio-nales y la encefalización.
2.1. El salto mutacional
Una nueva especie se forma a partir de otras ya existentes, a través de las mutaciones genéticas. El mecanismo biológico nos enseña que lo pro-pio de la vida es su capacidad de replicación, de formar nuevos seres vi-vos a partir de la confluencia de los gametos masculinos y femeninos (en los animales superiores, donde la reproducción se hace por unión sexual, y por la fusión de la herencia genética de los dos sexos). Lo normal es que la fase reproductiva genere miméticamente individuos semejantes a sus progenitores.
Pero en ese proceso reproductivo surgen continuos errores2, las muta-ciones. Se trata de un fenómeno totalmente aleatorio y azaroso, no previsto ni preparado por ningún mecanismo teleológico. Además, las mutaciones se están produciendo continuamente en el proceso evolutivo, pero sólo algu-nas de ellas, las que resultan viables, permanecen, desechándose las restan-tes, la mayoría, por no ser ventajosas para los nuevos organismos. La crea-ción de nuevas especies es consecuencia de este proceso aleatorio que es la mutación genética. De modo que el proceso evolutivo es una confluencia de un momento aleatorio, las mutaciones (azar), y otro de acomodación al am-biente, según la ley de la selección natural. El célebre genetista J. Monod lo sintetizó en el título de su libro, El azar y la necesidad3.
1 Cfr. Ruffié, J., De la biología a la cultura, Barcelona, Muchnik Editores, 1982.2 Cfr. Poitier, P. y otros, «La importancia de ser imperfectos», Investigación y Ciencia,
1994, n.º 217, 67-79.3 Cfr. Monod, Jacques, El azar y la necesidad, Barcelona, Barral Editor, 1977.

201
© Universidad de Deusto - ISBN 978-84-9830-383-4
Pero el proceso completo de la influencia de las mutaciones en la for-mación de las nuevas especies no se conoce en todos sus detalles, puesto que se dan mutaciones de diferentes tipos, y su influencia en la configura-ción de nuevas especies es muy diverso. De ahí que se suele hablar, en el proceso evolutivo, de dos fenómenos distintos aunque complementarios: la anagénesis y la cladogénesis4. La anagénisis, evolución anagenética o evolución lineal, es el proceso de acumulación de mutaciones ventajosas para una especie, sin que desemboque en una transformación de esa espe-cie en otra. Son mutaciones que suponen ventajas específicas para ciertos individuos de esa especie, ventajas que, puesto que son rasgos positivos, se irán imponiendo en el conjunto de la especie, produciendo una deriva genética en la dirección de tales rasgos ventajosos. Un ejemplo de anagé-nesis lo constituye el progresivo aumento de la dimensión encefálica en las diversas especies de humanos. O la reducción de cuatro a un dedo en la pata del caballo actual (desde el Hyracothrium, hace unos 50 millo-nes de años). O tambien, el crecimiento de la estatura de los homo, desde 1,20 m a 1,70 m, por término medio. Podríamos poner muchísimos ejem-plos de todas las especies.
La cladogénesis constituye el proceso mutacional que desemboca en la formación de una nueva especie (de clado, rama, en el árbol de la evo-lución). Así, de un antepasado común se originaron por un lado los chim-pancés, y por otro, los ardipithecos y australopitecos, hace varios millones de años. Este proceso cladogenético es el responsable de la formación, a lo largo de un complejo proceso de acumulación de cambios positivos, de las más de dos millones de especies vivientes, entre animales y plantas. Y lo mismo que aparecen, van desapareciendo también las especies, como consecuencia de la continua presión selectiva. Fco. J. Ayala nos hace ver que lo cálculos científicos concluyen que han desaparecido, por selección natural, el 99,9% de las especies que han existido5. La cladogénesis es, pues, el proceso por el que una especie existente se divide en dos o más. Las pruebas fósiles que encuentran los investigadores, muestran que la evolución morfológica es, en general, un proceso gradual, fruto de la acu-mulación de pequeñas mutaciones. Ahora bien, el registro fósil es discon-tinuo, puesto que no fosilizan todos los pasos del proceso, ni se encuen-tran todos los restos que han fosilizado. Por tanto, la mera referencia a los restos fósiles encontrados no sirve para mostrarnos cómo se produce este proceso, si de forma gradual o de modo discontinuo.
4 Cfr. Ayala, Fco. J., «El prodigio de la evolución», Muy Interesante, 1996, n.º 27, 16-21;cfr. pp. 19-20.
5 Cfr. ibídem, p. 19-20.

202
© Universidad de Deusto - ISBN 978-84-9830-383-4
Porque este es un problema, el ritmo de la evolución, sobre el que los especialistas no se ponen de acuerdo. De ahí que existan tres modelos o teorías que tratan de explicarlo: la teoría saltacionista o rupturista; la teo-ría gradualista y la teoría del equilibrio puntuado.
1) La teoría rupturista o saltacionista defiende que las especies se crean de modo repentino, por macromutaciones, procesos acele-rados que en poco tiempo separan a los nuevos individuos de la especie de la que proceden. Esas mutaciones introducen unas no-vedades y ventajas tan fuertes, que la ruptura se produce de modo rápido y claro, originando un aislamiento sexual casi completo y una acomodación suficiente a un nuevo nicho ecológico6.
2) La teoría gradualista propone que la formación de nuevas espe-cies es consecuencia de una larga acumulación cuantitativa de mu-taciones parciales, que, en un momento determinado (la acumula-ción cuantitativa se convierte en salto cualitativo), desencadena la formación de una nueva especie, separada ya de la especie madre por un largo profeso de diferenciación. Esta teoría la propuso ya Darwin, y en la actualidad la defienden los que podríamos deno-minar darwinistas ortodoxos, entre otros, Richard Dawkins7.
3) Hoy día, la teoría que parece se va imponiendo entre los cientí-ficos es la llamada teoría del equilibrio puntuado, propuesta por S. J. ould (Univ. de Harvard, USA) y Niles Eldredge (Museo Ame-ricano de Historia Natural, Nueva York) por primera vez en 1972, y completada en 19938. Según ellos, en el proceso de formación de las especies no sólo se da una dinámica gradual, sino que se produ-cen conjuntamente revoluciones, saltos, explosiones repentinas de cambio rápido. De modo que la evolución es una síntesis o conjun-to de largos períodos de equilibrio estático con momentos de rápi-dos cambios, en los que aparecen nuevas especies9.
6 De hecho, noticias de hace pocos años, de 1998, nos indican que el genetista Chung-I Wu y sus colegas de la Universidad de Chicago (USA) han dado a conocer, en la revista Science (20 de nov. de 1998), el resultado de sus investigaciones con moscas Drosophila:«un solo gen que muta muy deprisa, explica por sí mismo el origen de algunas especies de moscas». Cfr. El País, 25-nov.-1998, p. 33 (crónica de Javier Sampedro).
7 Cfr. El relojero ciego, Barcelona, Ed. Labor, 1988.8 Cfr. Nature, 18 de nov. de 1993. 9 Cfr. Gross, Michael, «El doble juego de una proteína», Mundo Científico, 1999,
n.º 204, 28-31. El autor nos habla de un descubrimiento, salido del campo de las proteínas antiestrés, que revelaría un mecanismo molecular que explicaría una evolución a dos velo-cidades, aportando con ello nuevos elementos en favor de la teoría de equilibrios puntea-dos. Cfr. también Henry Gee, «Explosiones repentinas de cambios. La nueva visión, 21 años después», El País, 8-XII-1993, Futuro, p. 3.

203
© Universidad de Deusto - ISBN 978-84-9830-383-4
Como ocurre en otros capítulos problemáticos del saber científico, aunque esta tercera teoría es la que resulta más plausible, por su al pare-cer mayor capacidad explicativa y su progresiva mayor aceptación por la comunidad científica, todavía tienen que aportarse nuevos datos para de-jar resuelto el problema. Pero la cuestión acerca de cómo se dio el salto de la especie anterior a la humana está llena de dificultades. Por un lado, los saltos mutacionales y la estructura genética de las especies vivas no dejan fósiles. De ahí que no podemos esperar a un hallazgo paleontológico para que la cuestión se resuelva. Por otro lado, la biología o la antropología mo-lecular sólo nos puede mostrar cómo se realiza el fenómeno reproductivo y cómo entendemos que se da el salto de una especie a otra. Pero el acerca-miento al problema del primer hombre tiene que hacerse a través de un su-puesto experimento teórico, consistente en aportar la teoría más plausible que nos permita comprender cómo pudo haber sucedido, teniendo en cuen-ta cómo funcionan los mecanismos genéticos normales.
Por de pronto, lo que hay que explicar es el salto de un cariotipo10
pre-humano (compuesto por 24 pares de cromosomas, como es el caso de la especie viva más cercana a nosotros, el chimpancé) a un cariotipo hu-mano (23 pares de cromosomas)11. El fenómeno de la especiación, esto es, la formación de especies a partir de otra anterior, aunque pueda tener momentos de transformación rápida (como indica la teoría de equilibriospuntuados), es un proceso que dura mucho tiempo, y en un primer mo-mento hay que suponer que los individuos portadores de mutaciones ge-néticas, que van a ir dando paso a una nueva especie, viven en medio de
10 Se denomina cariotipo al conjunto de cromosomas presentes en una célula. Cada cé-lula de un ser vivo contiene, por lo general, el cariotipo propio de la especie a la que perte-nece, como su signo de identidad. Se llama, por otra parte, «genotipo» al conjunto de genes presentes en un individuo de una especie viva. El genotipo está codificado en el ADN, en forma de doble hélice, en la que se encuentran los diferentes nucleótidos (adenina, guani-na, citosina y timina). El genotipo se expresa en el fenotipo, esto es, la morfología que con-forma a un ser vivo como resultado de su proceso de gestación. Pero no todo el genotipo se expresa en el fenotipo, puesto que sólo lo hacen los llamados «genes dominantes», mien-tras que los «recesivos» quedan sin expresarse en el fenotipo.
11 Sobre la genética humana en relación con el origen del ser humano, cfr. Ridley, Matt, Genoma. La autobiografía de un especie en 23 capítulos, Madrid, Taurus, 2000; Smith, John Maynard, La construcción de la vida. Genes, embriones y evolución, Barcelona, Crí-tica, 1998; Ayala, Fco. J., Origen y evolución del hombre, Madrid, Alianza, 1980; id., Lateoría de la evolución, Madrid, Temas de Hoy, 1999; Mattei, Jean-François (coord.), El ge-noma humano, Madrid, Editorial Complutense, 2002; Wright, William, Así nacemos. Ge-nes, conducta y personalidad, Madrid, Taurus, 2000. Bertranpetit, Jaume/Junyent, Cristina, Viaje a los orígenes. Una historia biológica de la especie humana, Barcelona, Península, 2000; Fox Keller, Evelyn, El siglo del gen. Cien años de pensamiento genético, Barcelona, Península, 2002.

204
© Universidad de Deusto - ISBN 978-84-9830-383-4
la población de la que han surgido, con intercambios sexuales fructíferos dentro de ella12.
Eso nos aboca a darnos cuenta de que el concepto de especie es una realidad lábil y no fácil de precisar desde posturas fixistas13. Cuando se comparan dos especies vivas existentes, al haberse separado en el proceso evolutivo hace miles de años, tienen ya una estructura genética bien de-limitada y diferenciada. Pero no ocurre lo mismo si comparamos indivi-duos que forman parte de una población en el momento en el que se está ya dando el fenómeno de separación específica. La separación genética progresiva origina al cabo de un cierto tiempo un aislamiento sexual (im-posibilidad de apareamientos fecundos, o, si se tienen descendientes, se-rán estériles).
Como puede verse, es imposible advertir experiencialmente cuán-do y en qué individuos se produjeron los primeras mutaciones genéticas para ir conformando el grupo poblacional que derivará hacia una nueva especie, en este caso la especie (o género) humana. De ahí que sea en sí misma una cuestión mal planteada (por imposible de responder) pre-tender descubrir el primer integrante de la especie humana (sea varón o hembra, o ambos). Vimos en parte este problema en el capítulo anterior al tratar la cuestión del monogenismo/poligenismo14. Las investigacio-nes más convincentes se encaminan a pensar que, frente a teorías que defienden un estrechamiento poblacional, hasta incluso reducirlo a una sola pareja, de la que se originaría la nueva especie, los planteamientos más plausibles empujan a pensar que en el origen de una especie, y tam-bién en el caso de la humana, se necesita un grupo poblacional de varios miles de individuos15.
Lo que diferencia genéticamente, como ya se sabe, a la especie hu-mana actual de los póngidos es el diferente número de pares de cromoso-mas16. Ahora bien, el que la especie humana tenga 23 pares de cromo-somas, frente a los 24 pares de los póngidos, en contra de lo que pudiera pensarse, no implica pérdida de contenido genético. De hecho, como se-ñala Fco. J. Ayala, «los cromosomas de ambos grupos (hombre y pón-
12 Sobre el fenómeno de la especiación, cfr. Ayala, Fco. J., Origen y evolución del hom-bre, o.c., pp. 95 y ss.; Id., La teoría de la evolución, o.c., pp. 135 y ss.; Ruffié, J., De la biología a la cultura, o.c., cap. IV, pp. 205 y ss.; Cavalli-Sforza, L. y Fr., ¿Quiénes somos? Historia de la diversidad humana, o.c., cap. 4.º, pp. 189 y ss
13 Cfr. Prebosti, Antonio, «Concepto de especie en el darwinismo actual», Mundo Cien-tífico, 1993, n.º 141, 1040-1049.
14 Cfr. el cap. 3.º de este libro, apartado 3.15 Cfr. Klein, Jan/Takahata, N./Ayala, F. J., «Polimorfismo del MHC y origen del hom-
bre», Investigación y Ciencia, 1994, n.º 215, 14-19. 16 Cfr. Ruffié, J., o.c., pp. 206 y ss.

205
© Universidad de Deusto - ISBN 978-84-9830-383-4
gidos) son muy semejantes entre sí, y no sólo uno a uno, sino banda a banda, excepto en lo que respecta a algunas mutaciones cromosómicas, como la fusión de dos de los cromosomas de los antropoides en uno del hombre»17. En concreto, una de las diferencias más claras es que lo que en la especie humana funciona como un único cromosoma, el n.º 2, en los chimpancés funcionan como dos diferentes. En palabras técnicas: la «fu-sión de dos acrocéntricos aún presentes en los monos antropomorfos, dio lugar a un único metacéntrico en el hombre»18.
Estas mínimas diferencias nos indican que la evolución puede produ-cir, con ligeras diferencias genéticas, importantes diferencias morfológi-cas y conductuales19, como es el caso de la especie humana. «Así, señala Ruffié, la evolución se caracteriza más por una redistribución, una nue-va disposición de las diferentes partes del material cromosomático, que por la creación de nuevos segmentos. La naturaleza innova con pocos gastos»20. Por tanto, con estos mínimos cambios, pero muy significativos, por las transformaciones y consecuencias que trajeron consigo, «se pro-dujo esta translocación fundamental que puede ser considerada como la frontera cromosómica de la hominización»21.
La dificultad está ahora en el modo como resolvemos la cuestión de cómo se fueron dando estos pequeños cambios para conformar la nueva especie, dotada de 23 pares de cromosomas. Se han propuesto diferentes teorías, intentando todas ellas presentar hipótesis plausibles de cómo pu-dieron haber sucedido las cosas, sin tener posibilidad de presentar prue-bas contundentes. Todas ellas apelan a un único individuo, hombre o mu-jer, en el que se habría producido la primera mutación, y a partir del cual se habrían propagado progresivamente esos cambios a sus descendien-tes, hasta ir conformando la nueva especie. Ese cambio mutacional, que provocaría el paso de 48 a 46 cromosomas, produciría en los individuos mutados un progresivo aislamiento sexual (no es razonable ni necesario pensar que se hubiera dado una separación geográfica), que produjera un
17 Origen y evolución del hombre, o.c., 160-161.18 Ruffié, J., o.c., p. 207. «Los dos acrocéntricos responsables, sigue diciendo Ruffié,
citando al Dr. Jean De Grouchy, corresponden a 2p y 2q del chimpancé que, por fusión ter-minal, dieron un sólo elemento metacéntrico: el cromosoma n.º 2 del hombre». Ruffié ad-vierte que algunos especialistas, como Pearson, discuten que esto esté totalmente probado. Para ver gráficamente la diferencia entre los diferentes cromosomas humanos y de los pón-gidos, cfr. Fco. J. Ayala, Origen y evolución del hombre, o.c., p. 161. Véase J. L. Arsuaga, El collar del Neandertal, o.c., p. 29, donde se nos muestra la comparación gráfica del cro-mosoma 4 del hombre, el chimpancé, el gorila y el orangután.
19 Cfr. Christen, Yves, El hombre biocultural, o.c., pp. 39-40. 20 O.c., p. 206.21 Ruffié, J., o.c., 207. El subrayado es del autor.

206
© Universidad de Deusto - ISBN 978-84-9830-383-4
aislamiento genético como consecuencia de la diferencia que se daba ya en el cariotipo.
J. Ruffié presenta dos teorías, entre otras, que pueden resultar vero-símiles22. A la primera se le denomina teoría adánica; y la segunda es la propuesta por el genetista francés J. Lejeune. Es importante indicar que es-tas dos teorías se pueden interpretar en clave gradualista y rupturista, pero parecen más adecuadas para pensarlas en clave rupturista, esto es, desde la óptica de producirse macromutaciones, que en una sola generación habrían dado paso al nuevo linaje humano. Por eso mismo, en la medida en que las teorías rupturistas son menos plausibles, estas propuestas que presentamos, son poco aceptables, al menos que se entiendan como un posible modo de entender el proceso de modo gradual y aproximativo.
1) La teoría adánica consiste en partir de una anomalía mutacional en un solo individuo (Adán; esta teoría admite la posibilidad de que este primer individuo sea también una mujer en vez de un va-rón, pero se inclinan hacia el varón por la mayor probabilidad de unión incestuosa padre-hija que madre-hijo), que situado en un grupo reducido de efectivos, con una fuerte endogamia, podría transmitir en poco tiempo (una o dos generaciones) su nueva mu-tación, pasando de 48 a 47 cromosomas en un primer momento, y posteriormente a 46. Es decir, el primer paso consistiría en la mutación experimentada en el primer individuo, en el proceso de la meiosis (formación de gametos, o células sexuales: espermato-zoides u óvulos). Un porcentaje de sus hijos/as llevaría consigo un cariotipo ya mutado, similar al de su padre, 47 cromosomas. Si se produce un cruce endogámico entre este Adán y su hija, o entre hermano-hermana con idéntico cariotipo mutado (las rela-ciones incestuosas, tanto en animales como en hombres, se da con más frecuencia entre padre-hijo, menos entre hermano-hermana, y pocas veces entre madre-hijo; de ahí que se considere como más probable la condición masculina del primer individuo mutado), una proporción de los descendientes de ambos (un 25%) tendrían ya en la siguiente generación un cariotipo de 46 cromosomas23.
Esta teoría tiene la ventaja de no presentar demasiadas rarezas ex-plicativas, apoyándose en pocos acontecimientos excepcionales: un salto mutacional producido en un único individuo, una relación endogámica, plausible dentro de la etología animal, y la lógica normal de la reproducción genética.
22 Cfr. ibídem, pp. 208-212.23 Cfr. Ruffié, J., o.c., p. 109.

207
© Universidad de Deusto - ISBN 978-84-9830-383-4
2) La teoría de J. Léjeune24 presenta un modelo parecido pero más complicado, puesto que parte de «dos gemelos monozigóticos, heterocariontes (verdaderos gemelos, surgidos de la división de un solo huevo, pero uno de los cuales ha perdido el cromosoma Y (síndrome de Turner)). En este caso habría machos normales, y hembras con un solo X (síndrome de Turner). Si éstas pudieran ser fecundadas (como en el caso del ratón), con su hermano geme-lo, formarían una pareja portadora de la misma anomalía: de ahí la formación inmediata de la nueva especie a partir de una pareja única»25.
Esta teoría, como señala Ruffié, es tan posible científicamente como la adánica, pero es más compleja, porque exige que se den al mismo tiempo dos anomalías o rarezas: la mutación cromosómica y la presencia del síndrome de Turner en uno de los gemelos. A pe-sar de esa complejidad, técnicamente es impecable, y pudo perfec-tamente haber sucedido así.
De todas formas, estas teorías parten, como ya hemos indicado más arriba, de presupuestos científicos que todavía no están del todo proba-dos, como es considerar que se puede dar un salto mutacional que supo-ne el paso en un solo individuo de la fusión de dos cromosomas en uno. Pudiera ser que esta fusión se realizara a través de un proceso que durara varias generaciones. Pero, aun así, estas teorías seguirían siendo en sus líneas maestras parecidas, sólo que necesitarían ser entendidas como un proceso que se alarga en varias generaciones de individuos, en vez de considerar que se han podido realizar en una sola generación.
Cuando Ruffié escribió De la biología a la cultura26, libro en el que nos apoyamos en gran medida en estos aspectos, señalaba que cabría, para poder contrastar la firmeza de estas teorías, hacer una prueba en dirección contraria: inseminar un óvulo de chimpancé con un espermatozoide hu-mano, para ver cómo se realizaba el proceso de pasar de 46 a 48 cromo-somas. El científico francés consideraba que era posible que alguien lo hubiera intentado con antelación, pero afirmaba que «aparte de su reproba-bilidad desde el punto de vista ético, estas experiencias, gracias a Dios, han fracasado. Efectivamente, el cariotipo humano y el del chimpancé han evo-lucionado cada uno por su lado desde hace 30 millones de años (…). … el
24 Cfr. Léjeune, J., «Adam et Eve ou le monogenisme», en Nouv. Rev. Théol., 1968, n.º 90, 191; Id., «Sur les mécanismes de la spéciation», C.R. Séances de la Soc. de Biol.,169 (4) 1975, p. 828.
25 Ruffié, J., o.c., 210-211; en p. 211 se presenta el esquema gráfico del proceso de in-tercambio genético propuesto por Léjeune.
26 El original francés es de 1976.

208
© Universidad de Deusto - ISBN 978-84-9830-383-4
cariotipo humano y el cariotipo antropomorfo actualmente están demasia-do alejados para que aún sea posible un apareamiento fuera de las condi-ciones de cultivo. E incluso en condiciones tan artificiales, la divergencia es tal, que este apareamiento sólo puede durar un cierto tiempo»27.
No muchos años después de estas afirmaciones, en mayo de 1989, saltaron a la prensa internacional unas declaraciones del científico italia-no, Bruno Chiarelli (Universidad de Florencia), al periódico L’Espresso,indicando la posibilidad de que en algún laboratorio del mundo (no espe-cificaba Chiarelli ni el lugar ni los posibles científicos autores de tal he-cho) se habría intentado crear un nuevo ser viviente, mitad hombre mitad chimpancé. Según Chiarelli, tal y como afirmaba en declaraciones poste-riores, el experimento se habría realizado en los años 60 en un laboratorio de California, sin que pudiera dar más datos. Según ello, este experimen-to, tras varios meses de realización, se habría detenido para evitar conse-cuencias indeseadas y la reacción adversa de la opinión pública.
Las reacciones que esta noticia produjo (una de las más fuertes provo-cadas en la comunidad científica en los últimos años) en todos los ámbitos del mundo, se centraron tanto en su aspecto ético como en el científico, afirmando ciertos investigadores de primera línea que tal proyecto todavía pertenecía a la ciencia ficción28.
Dejando de lado el hecho en sí y su polémica posterior, queda ahí la cuestión de cuál pudo ser el camino seguido por el proceso genético de la especiación, que explicaría la aparición de los primeros humanos.
2.2. La encefalización29
La frontera cerebral se ha considerado, con razón, como uno de los distintivos de lo humano, en la medida en que lleva aparejado a ello cuali-dades tan importantes como la conciencia y el ejercicio de la racionalidad, con todas sus consecuencias. Pero la pregunta que nos seguimos haciendo
27 J. Ruffié, o.c., p. 212. La fecha de 30 millones de separación entre el chimpancé y los humanos hoy día está, como ya quedó dicho, muy recortada, situándose entre 8 y 5 millo-nes de años.
28 Cfr. entrevista de M. P. Oliva a Bruno Chiarelli, El País, 11-XI-1987, p. 30; Arias, Juan, «Hombres mono», El País, 20-mayo-87, Futuro, pp. 1, 4 y 5. Cfr. el dosier de traba-jos sobre este asunto.
29 Cfr. Allman, J. M., El cerebro en evolución, Barcelona, Ariel, 2003; Ruffié, J., o.c., pp. 212-215; Martín, Robert D., «Capacidad cerebral y evolución humana», Investigacióny Ciencia, 1994, n.º 219, 71-77; Christen, Yves, El hombre biocultural, o.c., p. 42 y ss.; Sa-gan, Carl, Los dragones del Edén. Especulaciones sobre la evolución de la inteligencia hu-mana, Barcelona, Crítica, 1993.

209
© Universidad de Deusto - ISBN 978-84-9830-383-4
es dónde podemos situar la frontera o el límite entre un cerebro (o encéfa-lo, en general)30 humano y otro pre-humano. Es la búsqueda de lo que se ha solido denominar el Rubicón cerebral31.
El acceso al problema puede hacerse de dos modos, íntimamente rela-cionados: desde la medición del volumen y estructura cerebral, y desde el estudio de las herramientas construidas y utilizadas por los portadores de cerebros tan desarrollados. El tema de las herramientas lo trataremos de modo específico en el capítulo siguiente, por lo que nos centraremos aquí en los datos que la paleoantropología y otras ciencias afines aportan sobre el volumen y la estructura cerebral.
Algunos antropólogos llegaron a situar el Rubicón cerebral en 800 cc., de modo que cualquier cerebro de menor volumen, habría que conside-rarlo como un cerebro pre-humano. Con los avances más recientes de la arqueología y de la paleontología, este límite supondría, como ya vimos, dejar fuera de la condición de humano al Homo habilis. Pero las eviden-cias fósiles indican que el Homo habilis utilizaba ya herramientas com-plejas, indicadoras de un nivel de inteligencia que consideramos humano. Por tanto, mientras no tengamos otros criterios discriminadores, la consi-deración o no de un cerebro como humano tiene que depender no tanto de un número arbitrario de volumen cerebral, sino de su relación con la cons-trucción y uso de herramientas que denoten un nivel intelectual determi-nado, suficiente para considerarlo propio de una conducta humana.
La mejor comprensión del funcionamiento del cerebro nos lleva a pensar que las simples magnitudes de volumen no son suficientes para considerar a un cerebro como humano o pre-humano. Ya vimos en su mo-mento que los paleoantropólogos advierten una correlación entre masa encefálica y dimensión global del cuerpo, por lo que tienen que recurrir al llamado índice de cefalización. Pero es que, además, lo que caracteriza a un cerebro como humano, a diferencia de otro simiesco es sobre todo su estructura. El proceso de encefalización nos indica que el cerebro no crece de forma uniforme, como si fuera algo así como un globo esférico que se va hinchando de un modo homogéneo en todas sus direcciones, sino que posee una estructuración específica (configurada por sus diversas partes o lóbulos cerebrales), que es importante tener en cuenta.
J. Ruffié señala que se suelen comparar, para resolver este problema, dos modelos de estructuras cerebrales: la simiesca y la humana. La prime-ra está formada por «lóbulos occipitales muy desarrollados y lóbulos tem-porales y parietales más reducidos»; la segunda, por «lóbulos occipitales
30 El encéfalo se denomina al conjunto de cerebro, cerebelo y bulbo raquídeo.31 Cfr. Ruffié, J., o.c., 212-213.

210
© Universidad de Deusto - ISBN 978-84-9830-383-4
relativamente reducidos, lóbulos temporales y parietales muy desarrolla-dos, y los frontales muy desarrollados»32. La razón de ello está en que en los lóbulos frontales se sitúan las operaciones más sofisticadas de la inte-ligencia, propia de los seres humanos.
El cerebro humano se fue configurando de forma progresiva, produ-ciéndose una rápida aceleración en su crecimiento a lo largo de los últi-mos 2,5 millones de años, momento en el que surge la primera especie de la línea homo. La capacidad cerebral del Homo habilis, probada por los restos culturales (herramientas, vivienda, organización social, etc.) que fue capaz de ir produciendo, fue creciendo con una gran rapidez, hasta llegar al volumen del hombre moderno (entre 1200 cc. y 1800 cc.). Pero en sus niveles de estructura mínima, podemos considerar el cerebro y el comportamiento del Homo habilis como fundamentalmente similar al del ser humano actual. De este modo, la conclusión que hay que sacar es que resulta equivocada la noción de Rubicón cerebral referido a los primeros humanos, si la situamos en un volumen tan alto y la valoramos sólo desde el único criterio de su volumen.
En resumen, la aparición dentro del proceso evolutivo de los seres hu-manos se realizó al atravesar estas dos fronteras, la cromosómica y la ce-rebral. Ahora bien, en la medida en que el desarrollo del cerebro fue posi-bilitado por la correspondiente capacidad genética para ello, tenemos que ser conscientes, como señala Ruffié, que «la frontera de la hominización fue en primer lugar una frontera cromosómica»33.
3. LOS RASGOS MORFOLÓGICOS DE LA HOMINIZACIÓN
El traspaso de la frontera cromosómica dio lugar a una nueva configu-ración morfológica que se fue conformando durante miles de años. Ese pro-ceso, pues, no sólo trajo el aumento del cerebro sino una completa transfor-mación morfológica. Sería largo que analizáramos aquí lo específico del ser humano desde el punto de vista morfológico, y sus diferencias respecto a los antropoides. Nos vamos a limitar a seguir la costumbre de centrar esas dife-rencias morfológicas en cuatro aspectos fundamentales: la bipedestación, la mano prensil, el aumento del cerebro y la transformación de la cara.
Estos cuatro rasgos no se produjeron a la vez. Veremos cómo la bipe-destación marcó el punto de salida, originando, a su vez, la utilización de
32 O.c., 213.33 O.c., 215; Id., «Les frontières cromosomiques de l’hominisation», en C.-R. Acad.
Sc., 1973, 276, seri D, 1709.

211
© Universidad de Deusto - ISBN 978-84-9830-383-4
la mano para manipular y manejar las cosas, el aumento del cerebro y la progresiva transformación de la estructura ósea y musculosa de la cara. Pero los cuatro aspectos se fueron influenciando recíprocamente, de modo que un avance en uno de los cuatro elementos suponía a su vez un avance en los otros.
3.1. La bipedestación o postura erguida34
Como ya dijimos en su momento, el bipedismo se suele considerar en la actualidad como el rasgo distintivo de los homínidos, por lo que el ca-minar erguido no es algo que se inició con el ser humano sino mucho an-tes de la aparición del homo habilis. Las cuestiones que se plantean acerca de este rasgo conductual y morfológico son varias: cuándo se inició, qué ventajas adaptativas supuso y qué cambios morfológicos se dieron para pasar de una vida arborícola a otra en tierra.
Durante mucho tiempo, los paleoantropólogos discutieron, aportando cada uno sus peculiares razones, cuál de estos cuatro rasgos se había pro-ducido en primer lugar, convirtiéndose en detonante de los demás en el pro-ceso de transformación de la morfología de los humanos. Pero el problema quedó resuelto con el descubrimiento de que los australopitecos caminaban completamente erguidos, convirtiéndose, por tanto, el bipedismo en un ras-go pre-humano, desencadenante del aumento progresivo del cerebro, de la utilización inteligente de la mano y de las transformaciones faciales.
El descubrimiento clave lo constituyó el hallazgo de los primeros res-tos del Australopithecus affarensis por Donald Johanson y sus colabora-dores35, en especial el esqueleto casi completo de Lucy, que mostraba los rasgos óseos propios del bipedismo36. Este es un dato importantísimo, en
34 Cfr. Cela Conde, C. J. /Ayala, F. J., o.c., cap. 4.º, pp. 157 y ss.: Coppens, Yves, La rodi-lla de Lucy. Los primeros pasos hacia la humanidad, Barcelona, Tusquets, 2005: Ubeda Pur-kiss, M., «Datos para una antropología del hombre en pie», Estudios Filosóficos, XL (1991), n.º 113, 7-32; Asimov, Isaac, «Sobre sus piernas traseras», El País, 8-abril-1987, futuro, p. 2.
35 Cfr. Johanson, D./Edey, M., El primer antepasado del hombre, Barcelona, Planeta, 1982 (3.ª ed.: 1987); Id., La cuestión esencial. Preguntas y respuestas sobre la evolución,Barcelona, Planeta, 1990; Coppens, Y., La rodilla de Lucy, o.c.
36 Una aportación interesante la constituye el descubrimiento, por R. J. Clarke y Ph. V. Tobias, en el yacimiento sudafricano de Sterkfontein, de restos fósiles de un homí-nido de hace unos tres millones y medio de años, que muestran unas características inter-medias entre los humanos y los simios. Se trataría al parecer de restos de un individuo de la especie Australopithecus africanus (parántropo). Según sus descubridores, cuyos resul-tados publicaron en Science, 28-julio-1995, «tiene características humanas en la parte pos-terior del pie y sorprendentemente simiescas en la parte delantera». Por tanto, caminaría erguido pero conservaba todavía la habilidad para trepar a los árboles.

212
© Universidad de Deusto - ISBN 978-84-9830-383-4
la medida en que se partía, en algunas interpretaciones antropológicas ya superadas, de una dinámica excesivamente finalística en la propia antro-pogénesis, considerando que el primer factor que había originado todos los cambios era la dimensión cerebral, que desde una cierta conscien-cia habría desencadenado después el bipedismo, la transformación de la mano prensil y todos los demás rasgos que constituyen las características morfológicas humanas. Los datos paleontológicos que se poseen en la actualidad nos indican, como hemos dicho, que el bipedismo lo poseían los australopithecos, así como los ardipithecos, e incluso el orrogin tuge-niensis. Pero estas afirmaciones, como vimos en su momento, tienen que confirmarse y contrastarse con nuevos hallazgos fósiles. Aunque parece probado que el Australopitecus anamensis (descubierto por Maeve Leacky)poseía esta cualidad bipédica, no está claro todavía que la poseyera el Ar-dipithecus ramidus, descubierto por Tim White, puesto que no se tienen, al parecer suficientes piezas fósiles para comprobarlo. Lo que separa, lo dijimos en su momento, a los chimpancés de los homínidos australopite-cos es precisamente la capacidad de caminar erguidos y adaptarse a un en-torno ecológico de sabana, desprovisto ya de un arbolado denso.
Pero además de la cuestión sobre el origen más probable del bipe-dismo, son otros dos los aspectos que consideramos pertinente tratar: las transformaciones morfológicas que con ello experimentaron los homíni-dos, y los seres humanos con ellos, y las ventajas adaptativas que justifi-can su utilización, llevándoles a abandonar la vida arborícola anterior. Los cambios morfológicos que se produjeron a partir del inicio del caminar erguido no se hicieron de modo rápido, como ninguno de los demás cam-bios. Se trata de un largo recorrido en el que no vamos a detenernos más que para señalar los aspectos más importantes37. Uno de ellos implicó la transformación de la columna vertebral: de una forma arqueada, adaptada a la locomoción cuadrúpeda, fue adoptando la forma de «S», para soste-ner el tronco vertical, apoyado en la pelvis. Esto obliga a la pelvis a irse ensanchando, para poder sostener el conjunto del cuerpo y las vísceras ab-dominales. También se produce un alargamiento de las piernas, en compa-ración con los brazos, y un desarrollo importante de la musculatura glútea, necesaria para poder sostener todo el peso del cuerpo y dotar al individuo de la fortaleza suficiente o adecuada como para andar y correr38. Al mis-
37 Para un estudio más detallado, Arsuaga, J. L., La especie elegida, o.c., cap. 5.º (93-116); Laín Entralgo, P., El cuerpo humano. Teoría actual, Madrid, Espasa-Calpe, 1989, pp. 63-66.
38 Para más precisiones respecto a la pelvis humana, cfr. Arsuaga, J. L., El collar del Neandertal, o.c., pp. 97 y ss.; Id., La especie elegida, o.c., pp. 94 y ss.; Cela Conde, C. J./Ayala, F. J., o.c., pp. 180-185.

213
© Universidad de Deusto - ISBN 978-84-9830-383-4
mo tiempo, el pie se transforma totalmente, pasando de ser una segunda mano que se agarra a los árboles, a ser un miembro adaptado a su fun-ción sustentadora39. Otra de las consecuencias, sobre la que hablaremos más adelante, se refiere a las transformaciones producidas en la forma y estructura del cráneo, que ahora ya no cuelga de la columna, sino que se sitúa apoyado en la parte superior del tronco. Se da también un progresivo alargamiento del cuello, que «se hace esbelto, en notorio contraste con el cortísimo cuello de los antropoides»40.
Los antropólogos no se ponen de acuerdo a la hora de explicar las ra-zones y las ventajas evolutivas que llevaron a los homínidos a ir adoptan-do progresivamente la postura erguida. Es evidente que supuso una mejo-ra en el proceso adaptativo al ambiente, que debido al enfriamiento de la Tierra y a la sequía persistente, produjo una nueva retirada del arbolado y una ampliación de la sabana con grandes espacios abiertos. La postura er-guida daba a la nueva especie que comenzó a usar la bipedestación, unas ventajas claras. Evidentemente, la adopción del bipedismo tuvo que ser posibilitada, como ya se dijo al hablar de las dos fronteras de hominiza-ción, por las mutaciones genéticas que lo permitieran, puesto que los po-sibles póngidos que vivían con anterioridad en la región africana en la que aparece el ser humano fueron paulatinamente desapareciendo, porque no tenían capacidad para adaptarse al nuevo entorno ecológico. Por tanto, los australopitecos estaban genéticamente capacitados para desenvolverse en este terreno abierto, aunque es lógico pensar que el salto no se dio de for-ma repentina. Habría un período de adaptación entre la vida arborícola y la desarrollada en la sabana.
Como indica Marcel Blanc, cuando los estudiosos del bipedismo se planteaban la pregunta de qué tipo de ventajas encontraba el primer ser que empezó a caminar erguido, «la respuesta más frecuentemente usada desde el siglo XIX ha sido que la posición de pie permitía a los primeros homínidos liberar sus manos y fabricar utensilios (o armas) y de esta ma-nera asegurarse la superioridad sobre los demás animales»41. Este modo de pensar situaba el origen de los cambios morfológicos y conductuales en una especie de cierta previsión intencional de la nueva especie.
Estos planteamientos no tienen ya sentido en la actualidad, en la me-dida en que las diversas especies de homínidos caminaban erguidos a pe-sar de tener una capacidad cerebral no muy superior a los chimpancés,
39 Cfr. Deloison, Y., «El pie de los primeros homínidos», Mundo Científico, 1996, n.º 164, 20-23.
40 Laín Entralgo, P., o.c., 65.41 Blanc, Marcel, «El origen del bipedismo: ¿por motivos de sexo?», Mundo Científico,
1983, n.º 25, 526-529; 526.

214
© Universidad de Deusto - ISBN 978-84-9830-383-4
de los que se fueron separando progresivamente. La razón del inicio del bipedismo hay que buscarla en la necesidad de adaptarse al nuevo entor-no ecológico producido por la sequía, que originó el retroceso de la masa boscosa y el aumento de la sabana. Este factor, el cambio vegetacional, es el «empujón inicial» hacia los homínidos, como indican Philip Tobias42
y E. Vrba43. La tesis central de este modo de ver las cosas, defiende que hace entre seis y siete millones de años, debido a ese cambio vegetacio-nal, los primeros homínidos empezaron a abandonar los bosques tropica-les para ir aprovechando los nuevos espacios que se iban abriendo con el retroceso de los bosques empezando a adoptar la postura erguida y utili-zando diversas herramientas. Este modo de ver las cosas está unido a otra temática importante, como es el tipo de alimentación de los primeros ho-mínidos, a base de carne (por carroñeo44) o de semillas y frutos, con la re-lación directa que esto tiene con el desarrollo del cerebro.
De todos modos, la postura erguida, como puede comprenderse, no fue un modo de locomoción que se consiguió de modo rápido, sino que se trató de un proceso que duró mucho tiempo, dándose al principio un estilo de vida mixto entre la vida arborícola y la terrestre en la sabana45. Lo que parece evidente es que este nuevo estilo de vida fue ventajoso para estas nuevas especies en la medida en que les proporcionó ventajas adaptativas en ese nuevo nicho ecológico. Está claro que la postura erguida perma-nente facilitó la utilización de la mano para otros menesteres diferentes a las necesarias en un tipo de vida arborícola, y a la posibilidad de usar y construir todo tipo de herramientas. Pero no es fácil explicar de forma contundente a qué se debió esta utilización de la conducta bípeda, y prue-ba de ello es que se han presentado multitud de hipótesis, ninguna de las cuales ha conseguido una explicación total y su aceptación por todos los antropólogos. De todas formas, no deben confundirse la cuestión del por-qué de la aparición del bipedismo con la pregunta acerca de los beneficios de tal postura en su adaptación a la sabana, aunque ambas cuestiones es-tán muy unidas y se suelen presentar entremezcladas a la hora de su dis-cusión. Así, una primera hipótesis explica el bipedismo desde las ventajas que proporcionaba para el transporte de la comida y el cuidado de los hi-
42 Cfr. Tobias, P., The Brain in Hominid Evolution, Nueva York, Columbia University Press, 1971.
43 Cfr. Vrba, Elisabeth, «Evolution, Species and Fossils: How does Live Evolve?», S. Afr. J. Sci., 1980, 76 (2): 61-84; Cfr. Martínez Pulico, Carolina, El papel de la mujer en la evolución humana, Madrid, Biblioteca Nueva, 2003, cap. 5.º, «Elisabeth Vrba: El pa-leoclima que nos hizo humanos».
44 Cfr. Blumenschine, Robert J./Cavallo, John A., «Carroñeo y evolución humana», In-vestigación y Ciencia, 1999, Termas n.º 19, 76-83.
45 Cfr. Cela Conde, C. J./Ayala, F. J., o.c., pp. 166-170.

215
© Universidad de Deusto - ISBN 978-84-9830-383-4
jos pequeños; una segunda se fija en su adecuación para facilitar una con-ducta vigilante, al erguirse sobre las yerbas de la sabana; otra se refiere a la exhibición fálica de los machos para atraer a las hembras; otra defiende que favorecía el seguimiento de las piezas de caza en la sabana; otra hipó-tesis se fija en la ventaja de perder calor al quedar expuestos a la radiación solar en plena sabana; y otra considera que el bipedismo es favorable en el gasto energético cuando hay que hacer marchas largas46.
Pero existen otras hipótesis, como vamos a ver a continuación. El an-tropólogo norteamericano C. Owen Lovejoy ha propuesto que el bipedis-mo se debió al nuevo modo de vivir la sexualidad y a una nueva estrategia reproductiva del ser humano47. En la medida en que el primer homínido ca-minaba erguido y no se piensa que pudiera tener capacidad de construir he-rramientas, para Lovejoy la razón de utilizar el bipedismo se halla en que les permitió a los primeros homínidos procrear más. Considera Lovejoy que existen dos estrategias reproductivas: una primera de tipo cuantitativo (tener muchos descendientes, sin poder los progenitores atenderlos adecua-damente, de modo que el proceso de selección natural permite la supervi-vencia de un porcentaje pequeño, pero suficiente) y otra de tipo cualitativo (tener menos descendientes, pero cuidarlos mejor). La tendencia general del proceso evolutivo iría de la primera a la segunda estrategia. El punto más extremo de la estrategia cualitativa se daría, según Lovejoy, en la es-pecie humana. A cada pareja humana no le interesaría tener muchos hijos al mismo tiempo, sino sólo uno, pero tienen que cuidarlo con todo esmero para que sobreviva. Eso supone la tendencia a la conformación de parejas sexuales monogámicas y fuertemente unidas, situación a la que contribu-yó la orientación que fue adoptando la sexualidad femenina, que pasó de carecer de período estrogénico (estro) a estar permanentemente disponi-ble al contacto sexual y a la procreación. Eso le permitió sujetar de modo continuo a su pareja y, adoptando la estrategia que defiende la sociobiolo-gía respecto a las relaciones sexuales48, conseguir la hembra que el macho abandone su natural tendencia promiscua y poligámica y colabore de modo
46 Cfr. Tuttle, R. H./Webb, D. M./Tuttle, N. I., «Laetoli Footprint Trails and the Evo-lution of Hominid Bipedalism», en Coppens, Y./Senut, B. (eds.), Origine(s) de la bipedie chez les hominidés, París, Editions du CNRS, 1991, 187-198; Cela Conde, C. J./Ayala, F. J.,o.c., pp. 195-199.
47 Cfr. Lovejoy, C. Owen, Science, 211, 341, 1981. Este artículo suscitó un amplio de-bate, cuyos contenidos más importantes se publicaron en Science, 217, 295, 1982. Cfr. Lovejoy, C. O., «Evolución de la marcha humana», Investigación y Ciencia, 1999, Temas n.º 19, 66-75; Chastel, Olivier y Bonnet, Xavier, «La procreación por años sabáticos», Mundo Científico, 1996, n.º 170, 624-625.
48 Cfr. Dawkins, R., El gen egoísta, Barcelona, Labor, 1979; Ruse, M., Sociobiología,Madrid, Cátedra, 1983.

216
© Universidad de Deusto - ISBN 978-84-9830-383-4
continuo en la cría y el cuidado de la prole. Esto supone ir imponiéndose un reparto de roles sexuales y culturales, en el que el sexo masculino se de-dica a la caza y el femenino a cuidar a los hijos y a la recolección de plan-tas y posteriormente a la agricultura y la ganadería. Por tanto, la postura er-guida le permitiría a la mujer llevar en brazos al recién nacido y al varón le habría permitido utilizar las manos para manejar las armas que necesitaba para la caza y la búsqueda de alimentos para su pareja e hijos. Estas ven-tajas son las que, según Lovejoy y otros autores que defienden sus teorías, como Donald Johanson49 o Helen Fisher50, explicarían el éxito reproducti-vo de nuestra especie y la razón de que se haya mantenido viva, habiendo desaparecido todos los demás competidores cercanos, tanto los cercopite-cos (papiones, macacos, etc., que vivían en su mismo entorno) como pos-teriormente los australopitecos y los parántropos.
Esta teoría ha sido muy contestada desde diferentes puntos de vista. El antropólogo francés Marcel Blanc critica los aspectos fundamentales de la misma, empezando por las razones propuestas para la desaparición de los australopitecos. Si se dice que el bipedismo es el que asegura mejor la su-pervivencia, no se explica que hayan desaparecido los australopitecos y no los grandes simios que no tenían esa propiedad. Ello parece concluir que «este dispositivo anatómico y comportamental no asegura necesaria-mente una buena capacidad reproductiva, o sea, que el éxito reproductor no es el único factor a tener en cuenta para explicar la persistencia de una especie»51.
Tampoco es muy consistente lo que esa teoría defiende acerca del nuevo comportamiento sexual de la mujer en función de configurar fa-milias monógamas. Aparte de que hay quien señala que no es cierto la re-ceptividad sexual permanente de la mujer52, tampoco está probado total-mente, desde las investigaciones antropológicas, el papel que esta teoría atribuye a la mujer dentro de las primitivas sociedades humanas. Los roles de las mujeres en las sociedades primitivas actuales son tan variados que no inducen a pensar que se pueda aplicar la tesis de Lovejoy respecto a las mujeres de las que descendemos los humanos. Y lo mismo hay que decir
49 Cfr. Johanson, D./Edey, M., El primer antepasado del hombre, Barcelona, Planeta, 1982, pp. 273 y ss. Cfr. entrevista a Donald Johanson en El País, 1 de sept. de 1993, futu-ro, p. 3.
50 Cfr. Fisher, Helen E., El contrato sexual, Barcelona, Salvat, 1987. 51 Blanc, Marcel, o.c., 528.52 «En realidad, opina Marcel Blanc, hay en el ciclo menstrual un período coincidente
de forma aproximada con la ovulación y en el curso del cual la mujer manifiesta un com-portamiento sexual más marcado. Por otra parte, los estudios efectuados con parejas monó-gamas de otras especies (siamang, gibón) no corroboran la idea de la que la sexualidad sea el “adherente” de la pareja», o.c., 528.

217
© Universidad de Deusto - ISBN 978-84-9830-383-4
respecto al modelo de familia nuclear que la teoría de Lovejoy propone, porque si algo parece propio de las culturas humanas es la gran pluralidad de modelos de familia (y de cualquier comportamiento e institución cul-tural) que se han dado y se siguen dando53. «Todo ello prueba, concluye Marcel Blanc, que el carácter genéticamente programado de la condición de la mujer y de la familia nuclear propuesto por Lovejoy, es al menos, discutible»54.
En contrapartida, la solución que propone Marcel Blanc para el origen del bipedismo no va en la línea de un mayor éxito reproductivo (Lovejoy) sino de una mejor adaptación a un nuevo nicho ecológico, en el que se po-día alimentar mejor y no tenía ninguna especie que le pudiera hacer frente en ese nuevo entorno natural. Ese ámbito ecológico estaba representado por grandes extensiones de sabana, sin apenas arbolado, donde los homí-nidos se fueron acostumbrando a alimentarse de frutas, bayas, frutos secos de los arbustos y árboles bajos. En ese entorno, resultaba especialmente ventajoso poder ponerse de pie y caminar erguido para poder advertir con suficiente antelación la llegada de los depredadores y poder resguardarse en un árbol o en cualquier otro refugio55.
Esta adaptación supuso un progresivo perfeccionamiento de la capaci-dad manipuladora de la mano respecto a objetos pequeños, como semillas y raíces, así como una preponderancia de los molares en la masticación, aspecto que parece comprobarse por los paleoantropólogos al analizar lo gastados que aparecen las piezas dentales de estos homínidos, adaptados a una alimentación vegetariana con restos de tierra, mucho más erosionante que los frutos de los árboles56. Por tanto, «esta clase de explotación de los matorrales y los árboles bajos, en la sabana, representa la ocupación de un nicho ecológico apropiado a los primeros homínidos»57, y en él se fueron asentando, no encontrando al principio competidores serios y alejando después a cualquier otro que lo pudiera invadir.
Existen otras teorías propuestas, algunas de las cuales no dejan de ser curiosas y bastante poco creíbles, como la defendida por dos investiga-
53 De ahí que determinados autores, por ej. Rebecca Cann, le haya acusado de ser su teoría «una simple proyección en el pasado del modelo occidental de la familia nuclear y de las relaciones entre hombres y mujeres»: cfr. R. L. Cann y A. C. Wilson, Science, 217, 303, 1982. Y más en concreto, algunos autores le han acusado de representar una cobertu-ra científica a las pretensiones neoconservadoras de la época del presidente Regan en sus doctrinas sobre la familia.
54 O.c., 529.55 Cfr. Leaky, R. E. y Lewin, R., Los orígenes del hombre, Madrid, Aguilar, 1980;
Leaky, R. E., La formación de la humanidad, Barcelona, Edic. del Serbal, 1981.56 Cfr. Arsuaga, J. L., El collar del Neandertal, o.c., pp. 36 y ss. 57 Blanc, Marcel, o.c., 529.

218
© Universidad de Deusto - ISBN 978-84-9830-383-4
dores australianos, Nina G. Jablonski y George Chaplin58. Frente a otras muchas hipótesis que se han propuesto, «nosotros proponemos otras hi-pótesis: la adopción de la estación bípeda por los prehomínidos estaría relacionada con un contexto social específico, el de las agresiones en el seno del grupo y entre los grupos de primates superiores. La adopción del bipedismo, derivada de la parada de intimidación que lleva a un mono a mantenerse derecho para amenazar a su adversario, disminuyó el coste de morbilidad debida a estas agresiones, lo que representa una ventaja se-lectiva significativa. Esta ventaja se habría visto acentuada por las modi-ficaciones medioambientales, y luego aumentada aún por los beneficios derivados por los otros comportamientos engendrados por el bipedismo: comportamiento de centinela, comunicación, etc.»59. No deja de ser in-teresante la propuesta, pero, aunque quieran sus defensores hacerla creí-ble desde la importancia que tienen estos comportamientos en los prima-tes superiores, no acaba de ser convincente como explicación del nuevo modo de vida de los homínidos, puesto que no está demostrada la tan alta presencia de la agresividad entre ellos que parece suponer esta teoría, uti-lizándose esta costumbre como medio de solucionar más pacíficamente los conflictos ante la escasez de agua y alimentos60.
Posiblemente no exista una única razón o punto de vista que explique completamente las razones de la adopción del bipedismo, en vistas a sus in-contestables ventajas adaptativas. Por ello pensamos que son más acertadas las soluciones holistas, las que tienen en cuanta diferentes factores o ven-tajas: alimenticias (nicho ecológico específico), etológicas, eficacia repro-ductiva, etc. En esta línea va la propuesta de Manuel Domínguez Rodrigo, en El origen del comportamiento humano61. Tras desechar las teorías del contrato sexual, de Helen Fisher, y la del eterno femenino o hipótesis de la atracción epigámica (Lovejoy), presenta la suya, como hipótesis del con-trato social o de reforzamiento de las relaciones grupales. La novedad de este planteamiento consiste en tratar de estudiar este comportamiento homí-nido desde una visión conjunta, no reducida sólo a un aspecto concreto, por muy importante y determinante que se crea. Pero además, y esto es quizá lo más importante, propone el cambio de una visión etológica (que se que-da en la descripción de la conducta) por una mirada sinérgica (que trata de comprender la conducta, estudiando la acción en su conjunto). La adopción del bipedismo, según esta propuesta, permitió una mayor cohesión social
58 Jablonski, N. G., y Chaplin, G., «Antes de los primeros pasos: los orígenes del bipe-dismo», Mundo Científico, 1994, n.º 114, 270-271.
59 Ibídem, p. 271.60 Cfr. ibídem, p. 271.61 Madrid, Librería Tipo, 1994, cap. 3.º, pp. 98 y ss.

219
© Universidad de Deusto - ISBN 978-84-9830-383-4
del grupo, factor que, en círculo virtuoso, fue propiciado por una adaptación a un nicho ecológico mejor y un reparto mejor de roles, y, a su vez, mejoró también estos elementos. Pero lo que trajo, sobre todo, como resultado fue una progresiva cohesión grupal, que se mejoró muy sensiblemente cuando los humanos estuvieron provistos de una capacidad cerebral importante y de un modo de comunicación dotado de lenguaje articulado.
Esta adaptación al bipedismo hemos de verla dentro de un período de tiempo muy amplio, de progresiva adaptación en la que incluso los hom-bres actuales nos sentimos inmersos. En ese sentido, tenemos que recono-cer que el hombre actual se halla todavía aprendiendo a caminar erguido. Se trata, pues, de un modo de comportarse relativamente reciente, en tér-minos evolutivos (hace entre 7 y 5 millones de años), relativamente costo-so, y que no hemos aprendido del todo a utilizarlo o que estamos todavía en fase de adaptación. Prueba de ello es que no nacemos dominándola y que produce en ocasiones serias lesiones de espalda como consecuencia de lo costoso que supone su utilización permanente.
3.2. El desarrollo del encéfalo
Vimos con anterioridad que el extraordinario volumen que alcanza el cerebro en las especies de la línea homo, posibilitando su extraordinaria capacidad intelectual y su conciencia, constituye uno de los elementos di-ferenciadores de los humanos, y se convierte en la segunda frontera de la hominización. En su momento ya señalamos los aspectos fundamentales del cerebro humano; aquí vamos a completar y ampliar algunos datos que allí se apuntaron.
Dejamos ya dicho que el aumento del volumen cerebral no se produce de modo uniforme, sino que sigue una configuración estructural específi-ca, que lo distingue de un cerebro de características simiescas. De ahí que resulte equivocado situar el Rubicón cerebral en una cantidad determina-da en centímetros cúbicos de volumen. Los estudios acerca del cerebro de los humanos se han extendido de un modo tal, que resulta imposible pre-tender decirlo todo en pocas líneas. Bastarán algunos aspectos que consi-deramos más significativos para la finalidad que aquí perseguimos62.
El cerebro es una parte, la más importante, del encéfalo, que compren-de también el cerebelo y el bulbo raquídeo. La historia de la conformación del encéfalo indica que primero se formó el bulbo raquídeo (el cerebro rep-
62 Cfr. Allman, J. M., El cerebro en evolución, o.c.; Braillon, M. G., El sistema nervio-so central, Madrid, Alhambra, 1980; Allen, J. S./Bruss, J./Damasio, Hanna, «Estructura del cerebro humano», Investigación y Ciencia, 2005, n.º 340, 68-75.

220
© Universidad de Deusto - ISBN 978-84-9830-383-4
tiliano), a continuación el cerebelo, y al final, el cerebro, siendo su parte más importante la corteza cerebral. Mientras el bulbo raquídeo y el cerebe-lo realizan funciones más automáticas y vegetativas, el cerebro es la base de la racionalidad, encargándose de las funciones superiores como pensar, hablar, recordar, etc.63. Y dentro del cerebro, sus diferentes partes o lóbulos se fueron conformando desde el lóbulo occipital, pasando por los parie-tales y temporales, hasta emerger en último lugar el lóbulo frontal, como sede de las funciones superiores de la racionalidad humana.
La mayor capacidad cerebral dotó a los seres humanos de una gran ventaja adaptativa, y es la razón de habernos convertido en la especie hu-mana mejor adaptada, dominando al resto de las especies y capaz de vivir en todos los ambientes del planeta, sin casi límites de adaptación. Los da-tos que se manejan respecto a la aceleración del proceso, muestran que en comparativamente poco tiempo se produjo un enorme despegue en la lí-nea de los humanos64.
Al comparar el tamaño y el peso de los cerebros en las diferentes espe-cies animales, J. M. Allman extrae tres conclusiones: la comparación tiene que tener en cuenta sus tamaños corporales; los mayores pesos cerebrales están relacionados con sus mayores necesidades energéticas; y la expan-sión del tamaño cerebral ha tenido lugar en las diferentes clases de verte-brados; no es exclusiva de los mamíferos65. En este ejercicio comparativo, para Allman hay dos partes del cerebro que resultan especialmente ilustra-tivas: la red de neuronas serotonérgicas del tronco del encéfalo y la neocor-teza. La primera es una parte básica del cerebro, que apareció muy tempra-namente en la historia del cerebro, encontrándose ya en los vertebrados más tempranos, y se ha mantenido constante hasta el cerebro de los huma-nos66. Su nombre se debe a que sus terminales axónicos segregan el neu-rotransmisor denominado serotonina, que realiza un papel fundamental en la integración de la conducta. De tal forma que su carencia puede inducir a un tipo de conducta no adaptativa y a patologías que pueden poner en peli-
63 Cfr. Arsuaga, J. L. /Martínez, Natxo, La especie elegida, o.c., cap. 8.º, «La evolu-ción del encéfalo», pp. 151 y ss.; Goleman, D., Inteligencia emocional, Barcelona, Kairós, 1996; Damasio, Antonio R., El error de Descartes. La emoción, la razón y el cerebro hu-mano, Barcelona, Crítica, 1996; Rose, Steven, Tu cerebro mañana. Cómo será la mente del futuro, Barcelona, Paidós, 2008.
64 Cfr. Sagan, Carl, Los dragones del Edén. Especulaciones sobre la evolución de la in-teligencia humana, o.c.; Seché, Andreas, «Así sentamos cabeza», Muy Interesante, 1996, n.º 27, pp. 93 y ss.; Arsuaga, J. L./Martínez, N., o.c.; Christen, Y., El hombre biocultural,o.c., pp. 42-45.
65 Cfr. Allman, J. M., o.c., p. 29-30. 66 Dice Allman: «Esta función es tan fundamental que la arquitectura básica del sistema
serotoenérgico se ha preservado durante 500 millones de años», o.c., p. 38.

221
© Universidad de Deusto - ISBN 978-84-9830-383-4
gro la vida o una correcta integración grupal67. La segunda área fundamen-tal es el neocórtex o neocorteza, evolucionada mucho más recientemente, propia sólo de los mamíferos y muy evolucionada en los seres humanos. Se trata de una capa de unos dos milímetros de espesor y compuesta por el conjunto de interacciones y conexiones sinápticas entre las diferentes neuronas, y donde se sitúan las zonas que controlan la recepción de las ter-minaciones nerviosas que controlan las diferentes partes del cuerpo, con-virtiéndose en un auténtico mapa del cuerpo. Como ha quedado dicho, el neocórtex es uno de los rasgos distintivos de los mamíferos y «evolucionó como parte de un conjunto de adaptaciones relacionadas con la homeos-tasis térmica»68. Este dato es fundamental, en la medida en que el cerebro nació y se fue configurando para servir de amortiguador de las variaciones ambientales, sean de típico físico o de tipo sociológico.
Cuando se presentan datos comparativos entre la capacidad cerebral de los humanos y las especies que nos precedieron, los especialistas desechanla mera comparación cuantitativa, para referirse al llamado índice de ce-falización, que expresa la cantidad que resulta de la proporción entre la cantidad que cabría esperar en un avance aumentativo normal, y la can-tidad que de hecho se encuentra69. Según esto, si comparando las canti-dades globales, los hombres actuales ocupamos el lugar más alto (unos 1.450 cc. de media), frente al Australopithecus afarensis (426 cc.), el Australopithecus africanus (436 cc.), el Paranthropus o Australopithe-cus robustus (523 cc.), el Homo habilis/Homo rudolfensis (619 cc.), y el Homo erectus/ergaster (805 cc.). El Hombre de Neandertal ya vimos que presentaba cantidades iguales e incluso algo mayores que el Homo sa-piens sapiens.
Si lo vemos desde el índice de cefalización, los datos que tenemos son los siguientes: Australopithecus afarensis: 1,3; Australopithecus africanus: 1,4; Paranthropus boisei: 1,5; Homo habilis/rudolfensis: 1,8; Homo ergastes/erectus: 1,9. El hombre moderno presenta un índice ce-fálico de 2,9, lo que implica que «tenemos un encéfalo 7 veces mayor de lo esperado para un mamífero de nuestro peso corporal, pero sólo 2,9 ve-ces mayor de lo que le correspondería a un primate haplorrino de nuestro tamaño»70.
Pero si la superioridad del cerebro humano es grande con estos da-tos señalados, todavía aumenta la distancia si hacemos referencia a la su-
67 Cfr. ibídem, pp. 31-38, 68 Allman, J. M., El cerebro en evolución, o.c., p. 114. 69 Cfr. AllmanN, J. M., El cerebro en evolución, o.c., pp. 28-30; Martin, R. D., «Capa-
cidad cerebral y evolución humana», Investigación y Ciencia, 1994, n.º 219, pp. 71-77.70 Arsuaga, J. L./Martínez, N., o.c., p. 160.

222
© Universidad de Deusto - ISBN 978-84-9830-383-4
perficie de nuestro cerebro. La superficie del cerebro no es lisa, sino que presenta diversas circunvoluciones y fisuras (o cisuras), con sus nombres respectivos. Si medimos la superficie de la corteza cerebral (neocórtex,lugar donde se sitúan las terminaciones nerviosas de las neuronas), la del ser humano tiene 22.260 cm2 de extensión, mientras que la del gorila es de sólo 5.500 cm2 71. Y si comparamos también el número de neuronas que posee el cerebro humano con las del gorila, el cerebro humano posee tres veces más neuronas (de 9 a 12 miles de millones), cantidad muy su-perior igualmente a las del orangután (cerca de 3 mil millones)72.
Un cerebro grande proporciona importantes ventajas en el proce-so evolutivo, pero también supone grandes costos, tanto en inversión de energía alimenticia como en tiempo y en cuidado de los progenitores73.Hay por tanto una cierta relación entre cerebro grande y dimensión corpo-ral, en la medida en que el cerebro se encarga del control de todo el cuer-po. Pero también exige grandes costos energéticos y de cuidado de los progenitores sobre todo en los primeros momentos de la vida.
El cerebro exige para su mantenimiento un gasto enorme de energía, de tal forma que, si por un lado su capacidad intelectual supone una ri-queza enorme en el proceso adaptativo, también conlleva unos costes que no siempre se pueden compensar. De ahí que cada especie posea un tipo de encéfalo correspondiente al equilibrio entre costes y beneficios74. Una pregunta interesante es, por tanto, cómo se produjo en los humanos el acomodo energético y morfológico hacia un cerebro tan desproporciona-do. Según Leslie Aiello y Peter Wheeler, el aumento del cerebro se habría realizado a costa del aparato digestivo. «En relación con su peso, los órga-nos energéticamente más costosos del cuerpo humano son el corazón, los riñones, el cerebro y el conjunto formado por el tubo digestivo más el hí-gado; el cerebro representa el 16 por ciento de la tasa metabólica basal del organismo (el gasto energético, medido por unidad de tiempo, necesario para mantener las funciones vitales de un individuo en reposo), y el tubo digestivo un porcentaje próximo, el 15 por ciento»75.
Para Aiello y Wheeler, el aumento del cerebro en los humanos se hizo a costa del acortamiento del tubo digestivo, y ello fue permitido por el pro-
71 Cfr. Ruffié, J., o.c., p. 202; Arsuaga, J. L./Martínez, N., o.c., pp. 161 y ss. 72 Cfr. Ruffié, J., o.c., p. 202.73 Cfr. Allman, J. M., o.c., cap. 7.º, pp. 165 y ss.; Martin, R. D., «Capacidad cerebral y
evolución humana», Investigación y Ciencia, 1994, n.º 219, pp. 71-77. 74 Cfr. Martin, R. D., o.c. R. D. Martin, los cambios de mayor alcance experimentados
en la sociedad humana han ido acompañados de un descenso progresivo del tamaño de nuestro cerebro, lo que explicaría, entre otras cosas, que el cerebro de los neandertales pue-da arrojar cifras de promedio más altas que las del hombre actual.
75 Arsuaga, J. L./Martínez, N., o.c., pp. 183-184.

223
© Universidad de Deusto - ISBN 978-84-9830-383-4
gresivo cambio de un modo alimenticio exclusivamente herbívoro (nece-sitado de una digestión más larga, y un aparato intestinal también largo) a otro carnívoro, con la ventaja de la mayor facilidad que posee la carne en el proceso de asimilación digestiva76. Incluso dentro de los animales her-bívoros, se dio un salto progresivo desde los hojíveros (alimentados con hojas) a los frugívoros (alimentados con frutas, como los primates)77. Por tanto, como señalan Arsuaga y Martínez, «era necesario que nos volviéra-mos carnívoros para ser inteligentes (aunque esta es una pescadilla que se muerde la cola porque los alimentos de alta calidad requieren de mayores capacidades mentales para ser localizados)»78. Y ello nos muestra de nuevo en qué medida la configuración de los humanos (y de todas las especies) es el resultado de procesos que han durado largos períodos de tiempo, interac-tuando varios factores entre sí, en forma trenzada, para apoyarse y comple-mentarse. Es lo que se denominan procesos de coevolución.
Otro factor a tener en cuenta en la evolución del cerebro de los humanos es el factor tiempo. Parece evidente que los animales que viven más tiempo son más propicios a experimentar a lo largo de su vida más variaciones am-bientales de todo tipo. Por tanto, necesitan un cerebro más capaz para poder amortiguar esas variaciones, produciéndose una relación directa entre estas dos variables. Así se ve cuando comparamos la capacidad cerebral y la lon-gevidad entre animales que se alimentan con hojas (hojíveros), con frutos (frugívoros) y los carnívoros79. Este dato se conecta con otro fundamental: los cerebros más grandes se van formando a lo largo de períodos de tiem-po más amplios, por lo que necesitan una mayor inversión de cuidados pa-rentales, siendo el ser humano el que necesita un cuidado mayor. Además, parece que se advierte que, en la medida en que es el sexo femenino el que dedica más tiempo y esfuerzo en el cuidado de los hijos, se explicaría desde ahí la mayor longevidad de las hembras sobre los machos80.
Con esto nos introducimos en uno de los aspectos de mayor interés en relación a la formación y características del cerebro humano: la rela-ción entre cerebro y entorno social. Si la función más importante del ce-rebro es servir de amortiguador de los factores ambientales, la principal circunstancia ambiental del ser humano no es el entorno ecológico sino el social y cultural. Por tanto, la tesis que parece más razonable defender es que uno de los factores más importantes que han influido en el aumen-to de la dimensión craneal humana es la condición social de los seres hu-
76 Cfr. ibídem, p. 184.77 Cfr. Allman, J. M., o.c., p. 170. 78 Arsuaga, J. L./Martínez, N., o.c., 185.79 Cfr. Allman, J. M., o.c., pp. 174-178. 80 Cfr. ibídem, pp. 182 y ss.

224
© Universidad de Deusto - ISBN 978-84-9830-383-4
manos. Hay, pues, una directa e importante relación entre cerebro y so-cialidad, de tal modo que a mayor capacidad cerebral, esto es, a mayores capacidades intelectivas existen mayores posibilidades de constituir so-ciedades más amplias, comunicadas y cohesionadas. Y, de modo comple-mentario, sociedades con mejores niveles de comunicación y de cohesión provocan y empujan hacia una mayor cerebralización81.
Es importante advertir que lo propio de los seres vivos es interactuar con el entorno de modo inteligente, acumulando información y capaci-tándose progresivamente para actuar frente al entorno, de forma tal que la vida del individuo esté protegida. Es la ley de la selección natural. Esta dialéctica individuo-entorno, entendida ahora en términos de capacidad intelectiva para actuar frente al ambiente (Umwelt, Von Uexküll), es una ley que atraviesa toda la biosfera, como lo explicita el etólogo Konrad Lorenz, en La otra cara del espejo82. Pero, entre los humanos, como ya hemos indicado, el ambiente que más determina e influye el crecimiento intelectual no es tanto el entorno ecológico, sino el social. Este dato es el que probaría la necesidad de que los seres humanos más capacitados se-rían aquéllos que estuvieran más dotados para interpretar la conducta de los demás seres humanos. En esta línea van las tesis de N. Humphrey83,el descubrimiento de las denominadas neuronas espejo84, o las tesis de R. Byrne y A. Whiten sobre lo que han denominado la hipótesis de la inte-ligencia maquiavélica (capacidad para interpretar la conducta de los otros y anticiparse a sus movimientos)85. Ahora bien, esta idea de mente ma-quiavélica no necesariamente ha de interpretarse en clave indivualista y agresiva, sino también de modo positivo, esto es, una mostración de la ra-dical reciprocidad del ser humano, vertido a los otros y dotado de la capa-cidad de poseer una inteligencia social86. «En resumen, señalan Arsuaga y Martínez, es una teoría muy respetable la de que la expansión del cerebro y la inteligencia (o al menos una parte sustancial de la misma) representa
81 Cfr. Arsuaga, J. L./Martínez, N., o.c., pp. 216 y ss. 82 Barcelona, Plaza y Janés, 1980. 83 Cfr. Humphrey, Nicholas, Consciousness Regained, Oxford, Oxford University
Press, 1983 (trad. cast.: La reconquista de la conciencia. Desarrollo de la mente humana,México, FCE, 1987); Id., The Inner Eye, Londres, Faber and Faber,1986 (trad. cast.: Lamirada interior, Madrid, Alianza, 1993); Id., A History of the Mind, Londres, Simon and Schuster, 1992 (trad. cast.: Una historia de la mente. La evolución y el nacimiento de la conciencia, Barcelona, Gedisa, 1995).
84 Cfr. Rizzolatti, G./Sinigaglia, C., Las neuronas espejo. Los mecanismos de la empa-tía emocional, Barcelona, Paidós, 2006.
85 Cfr. Byrne, R.W., The Thinking Ape: Evolutionary Origins of Intelligence, Oxford, Oxford University Press, 1995
86 Cfr. Humphrey, N., obras citadas en nota 83.

225
© Universidad de Deusto - ISBN 978-84-9830-383-4
una adaptación a la vida social, un medio en el que uno tiene que cooperar y competir a la vez con los mismos individuos»87. Pero nos estamos aden-trando por aquí en el estudio de la socialidad humana en relación con la antropogénesis, aspecto sobre el que nos extenderemos más adelante, en un capítulo posterior.
En esta relación entre tiempo y socialidad, tenemos que hacer también referencia a otro factor fundamental: la relación existente entre cerebros grandes y el cuidado parental. El coste de sacar adelante a un individuo con cerebro grande es tarea sobre todo de sus progenitores, observándose las dos estrategias reproductivas analizadas por Lovejoy y de las que hablamos en el apartado sobre el bipedismo. La línea evolutiva de los humanos se fue especializando en un menor número de descendientes, con el progresivo au-mento de los esfuerzos de los dos padres en su cuidado. En el caso del ser humano se da además un factor esencial: somos una especie que nacemos de forma prematura (animal deficiente: A. Gehlen), con el cerebro sin ter-minar de conformarse. Se daría, pues, una tendencia evolutiva a primar se-lectivamente a las crías que estuvieran más dotadas para obedecer y apren-der de sus progenitores, con el consiguiente aumento de la dependencia de ellos. De tal modo que, comparando el desarrollo embriológico de las espe-cies más cercanas al ser humano, se advierte que en nuestra especie se ha producido un fenómeno de infantilización o paidomorfismo. El ser humano viene a ser como el feto de un mono88. El desarrollo del cerebro huma-no, si tenemos en cuenta su proceso ontogenético, no termina de desarro-llarse con el nacimiento, sino que continúa varios años después de abando-nar el seno materno, siendo fundamental y decisivo el primer año post-parto (el llamado año extra-uterino del embrión, Portmann), pero alargándose la maduración varios años más, hasta diez o más. De tal modo que, aunque el cerebro del recién nacido posea todas las neuronas que le corresponden por su condición humana89, tiene que ir formando y desarrollando posterior-mente todo el conjunto de relaciones e interconexiones neuronales que le permitirán hacerse cargo de la compleja estructura comportamental del ser humano. De tal modo que la capacidad cerebral y craneana del recién na-cido es muy inferior (aproximadamente la mitad, o menos) de la que va a tener en su adultez90, posibilitando así el parto y el paso de la cabeza por el
87 O.c., p. 217.88 Cfr. Christen, Y., El hombre biocultural, o.c., pp. 67 y ss. 89 Uno de los aspectos de investigación en el campo de la neurobiología es el de la ca-
pacidad o no de regenerarse las neuronas desgastadas. Se piensa que sólo en casos muy ex-cepcionales se consigue tal regeneración. Cfr. Dusant, I. /Rubin, B. C. /Schwah, M. E., «La regeneración de las fibras nerviosas», Mundo Científico, 1993, n.º 41, 1009-1014.
90 Cfr. Trevarthen, C., «Cerebro, desarrollo del», en Gregory, R. L. (ed.), DiccionarioOxford de la mente, Madrid, Alianza, 1995, pp. 149-161.

226
© Universidad de Deusto - ISBN 978-84-9830-383-4
canal pélvico de la madre (tendremos ocasión de completar estos aspectos más adelante). Lo que hace crecer el volumen y la capacidad cerebral no se-ría, por tanto, el crecimiento del número de neuronas, sino las interconexio-nes sinápticas que se establecen entre ellas en el ejercicio de actuar el niño con el entorno, sobre todo con otros seres humanos.
Pero estas afirmaciones no son hoy día tan evidentes, y parecen estar más bien en entredicho, debido a recientes descubrimientos en el ámbi-to de la neurología. Hasta no hace mucho tiempo, el dogma central de los estudios sobre el cerebro lo constituía la afirmación de que nacemos con un número determinado de neuronas (aproximadamente ¡unos cien mil millones!91) y, si alguna muere, no se repara ni es sustituida. Este supuesto dogma ha empezado a ser puesto en cuestión por diversos hallazgos cientí-ficos. Pero el consenso sobre este punto está todavía lejos de conseguirse. Desde el año 1884, F. Nottebohm (Universidad de Rockefeller) había ob-servado el nacimiento de nuevas neuronas en animales como el canario, con ocasión de aprender nuevas habilidades. Pero estos hallazgos no se tuvieron demasiado en cuanta hasta que en 1996 E. Gould y Ch. Gross (Universi-dad de Princeton) descubrieron que en el hipocampo, parte del cerebro muy implicada en la memoria, se encontraban neuronas que se dividían y rege-neraban. Pero no sólo parece que se da neurogénesis en el hipocampo, sino también en la región frontal (parte que interviene en la ejecución de decisio-nes, la interacción social y la memoria a corto plazo), la región temporal in-ferior (reconocimiento visual de objetos y caras) y la región parietal poste-rior (sentido del espacio y el reconocimiento de objetos). Los experimentos donde se ha visto esta regeneración neuronal se han realizado en macacos, y se considera totalmente extrapolable al ser humano92. Todo esto tendría unas consecuencias muy favorables para curar muchas enfermedades con-secuentes al deterioro del sistema nervioso del cerebro, como el parkinson y el Alzheimer, y los procesos de envejecimiento en general.
Ahora bien, estas afirmaciones han sufrido últimamente un serio re-vés como consecuencia de otros experimentos llevados a cabo por Pasko Rakic y D. R. Kornack (Universidad de Yale)93, otro de los grandes espe-cialistas cerebrales, en los que no se han confirmado las tesis de E. Gould. En los experimentos de Rakic y Kornack, se producían regeneración neu-
91 Cfr. Blond, Olivier, «Las neuronas», Mundo Científico, 1999, n.º 199, 82-85; 83. 92 Cfr. Townsend, Rosa, «La regeneración cerebral abre nuevas fronteras», El País,
23 de noviembre de 1999, p. 39; Costa-Pau, Marta, «El hallazgo de que las neuronas pue-den regenerarse es crucial» (entrevista a Paola Timiras, especialista en envejecimiento), ElPaís, 28 de septiembre de 1999, p. 47; Blakeslee, Sandra, «Sorpresas de la década del ce-rebro», El País, 12 de enero de 2000, p. 31.
93 Cfr. Science, 7 de diciembre de 2001.

227
© Universidad de Deusto - ISBN 978-84-9830-383-4
ronal en el hipocampo y el bulbo olfativo, pero no en el neocortex adulto de los primates94. Así, pues, los científicos están divididos sobre el alcan-ce del proceso de neurogénesis cerebral. Lo que sí parece evidente es que la tesis central hasta no hace mucho, que negaba cualquier regeneración neuronal, parece que está empezando a resquebrajarse.
Pero lo que está claro es que el aumento de la masa cerebral que se produce en los años sucesivos al nacimiento se debe a las múltiples in-terconexiones neuronales (sinapsis) que se van produciendo entre las di-ferentes neuronas. Una neurona puede establecer contacto con otra que puede estar a varios centenares de veces la longitud de su cuerpo celular, por medio del alargamiento de su axón, en cuyo extremo se dan los co-nos de crecimiento, provistos de los filípodos, tentáculos que buscan su camino de conexión con las dendritas de otras neuronas95. Este proceso de conexión entre diversas neuronas constituye la base cerebral donde se apoyan los procesos sensoriales de recogida de datos del exterior, así como las respuestas motoras con las que se interactúa con el entorno. De este modo, las redes neuronales, que al principio son provisionales, se van consolidando para formar redes permanentes que constituyen la base de los procesos vitales permanentes del ser vivo, convirtiéndose tales proce-sos sensoriales y motores en acciones automáticas y maduras.
Pero esto no significa que la formación de redes o mapas neuronales se deba sólo a una respuesta al ambiente, sino que también responde a una orientación innata que permite al ser vivo realizar múltiples funciones, configurándose la conducta como una síntesis de orientaciones innatas y aprendidas96. Una de las teorías que resultan más interesantes para descri-bir la conformación de la estructura cerebal a través de la interacción en-tre neuronas individuales y posteriores sistemas neuronales, es la propues-ta por Gerald Edelman y su aplicación al cerebro de la selección natural darwiniana97. Según Edelman, el cerebro se va configurando genética-mente por interconexiones neuronales que forman mapas sistémicos que interactúan con el entorno del viviente. Unos mapas están conectados con
94 Cfr. Ruiz de Elvira, Malen, «Recientes experimentos niegan la formación de neuro-nas nuevas», El País, 13 de enero de 2002, p. 27.
95 Cfr. Blond, Oliver, «Las neuronas», o.c., p. 84. 96 Kalil, Ronald E., «Formación de sinapsis durante el desarrollo del cerebro», Investi-
gación y Ciencia, 1990, n.º 161, 26-34; Kennedy, H./Dehay, C., «Córtex cerebral: el peso de los genes», Mundo Científico, 1994, n.º 146, 474-475.
97 Cfr. Edelman, G., Neuronal Darwinism: The Theory of Neuronal Group Selection,Basic Books, 1987; Topobiology: An Introduction to Molecular Embriology, Basic Books, 1988; The Remembered Present: A Biological Theory of Consciousnes, Basic Books, 1989;Bright Air, Brilliant Fire: On the Matter of the Mind, Basic Books, 1992; Edelman, G./Tononi, G., El universo de la conciencia, Barcelona, Crítica, 2002.

228
© Universidad de Deusto - ISBN 978-84-9830-383-4
otros mapas, formando el entramado total del cerebro. Pero estos mapas iniciales no son definitivos, sino que vienen a ser como propuestas varia-das para la captación de sensaciones y respuestas motoras. En la medida en que el ser vivo interactúa con el entorno, determinados mapas resultan más valiosos que otros, produciéndose una selección de unos mapas en detrimento de otros. Por tanto, el resultado de la conformación de un cere-bro maduro es la suma de orientaciones genéticas innatas y del aprendiza-je experiencial, de tal modo que, como indica J. Searle, «el cerebro anda genéticamente equipado desde el nacimiento con una sobreabundancia de grupos neuronales, y su desarrollo acontece merced a un mecanismo pa-recido al de la selección natural darwiniana: algunos grupos neuronales mueren, mientras otros sobreviven y se robustecen. En algunas partes del cerebro muere hasta un 70% de las neuronas antes de que el cerebro lle-gue a madurar. La unidad seleccionada no es la neurona individual, sino grupos neuronales de cientos y hasta millones de células. La cuestión fun-damental es que el cerebro no es un mecanismo instructivo, sino un meca-nismo selectivo; es decir, que el cerebro no se desarrolla por alteraciones en un conjunto fijo de neuronas, sino por procesos selectivos que eliminan algunos grupos neuronales y robustecen a otros»98.
En resumen, como acabamos de comprobar, la configuración del ce-rebro en los seres humanos es el resultado de un complejo número de fac-tores y elementos que lo van orientando, desde el momento en el que se produce, por mutación genética, el paso de la frontera cromosómica hasta la actual configuración del cerebro humano.
3.3. La mano prensil e inteligente99
Con el inicio de la bipedestación comenzó también la paulatina trans-formación de la mano, que pasa de ser el pie delantero (aunque no tuvo nunca la forma de un pie, puesto que su forma estaba adaptada a la fun-ción de agarrarse a las ramas de los árboles) a un instrumento que maneja a la perfección toda clase de objetos y herramientas. Junto a la mano, el conjunto del brazo también experimenta variaciones, en la media en que los brazos se van acortando, comparativamente con las piernas. Los si-
98 Cfr. Searle, John, El misterio de la conciencia, Barcelona, Paidós, 2000, cap. 3.º, «Gerald Edelman y la cartografía del reingreso», pp. 45-56; la cita es de las pp. 46-47; las cursivas son del autor.
99 Cfr. Wilson, Frank R. La mano. De cómo su uso configura el cerebro, el lenguaje y la cultura humana, Barcelona, Tusquets, 2002; Alba, D. M./Moyá Solá, A. /Köhler, Meike, «El origen de la mano humana», Investigación y Ciencia, 2005, n.º 341, 46-53.

229
© Universidad de Deusto - ISBN 978-84-9830-383-4
mios poseen las piernas más cortas y los brazos más largos, lo que les per-mite el modo de andar típico que todos conocemos, apoyándose en el sue-lo con los nudillos de las manos100.
En los humanos, los brazos ya no sirven como ayuda para andar, sino para manejar objetos y herramientas, por lo que experimentan un signifi-cativo acortamiento comparativo. Posiblemente el primer uso específico de las manos fue la manipulación de semillas, raíces y otro tipo de ali-mentación propia del nuevo nicho ecológico en el que se fueron situando los homínidos. Y esta especialización de la mano se pudo haber dado ya entre los australopitecos y ardipethecos, desde el momento en que se ori-gina en ellos la bipedestación101.
Esta progresiva utilización de la mano para la manipulación de obje-tos pequeños, que requerían una gran precisión táctil, irá convirtiendo la mano en una herramienta maravillosa, capaz de realizar mil movimientos y operaciones, que no le llega a superar ni la más precisa herramienta me-cánica que después ha ido construyendo el ser humano. De hecho, ella es la que construye y maneja todas las herramientas que luego han ido apare-ciendo en el entorno cultural humano102.
Descubrimientos recientes van también aclarándonos cual es el origen físico de los dedos de las manos y de las patas de los mamíferos. Frente a teorías anteriores que pensaban que eran una prolongación de las aletas de los peces, un equipo de científicos de la Universidad de Ginebra conside-ran que los dedos de los mamíferos no tienen ninguna homología o pro-cedencia de las aletas de los peces, sino que «los dedos son huesos comple-tamente nuevos que han sido inventados por los tetrápodos y no proceden de los peces»103. Sería muy interesante, pero nos llevaría muy lejos de nuestro propósito en estas páginas, seguir la historia de la mano desde su formación en los primeros mamíferos hasta que toma la forma humana104.
Lo que hizo humana la mano fue precisamente el ser el instrumento y prolongación de un cerebro inteligente105. De ahí que sea fundamental e
100 Cfr. Arsuaga, J. L. y otros, «Los homínidos en la hipótesis de Valverde», MundoCientífico, 2000, n.º 225, pp. 58-63.
101 Cfr. Arsuaga, J. L./Martínez, I., o.c., 182-183; Cela Conde, C. J./Ayala, F. J., o.c., pp. 185 y 349-352.
102 Cfr. Richter Sherman, Claire, Writing on Hands, Washington, The Folger Shakespeare Library, 2000.
103 Cfr. Rivera, Alicia, «Los dedos son una innovación y no derivan de las aletas de los peces», El País, 28-junio-1995, p. 29.
104 Cfr. Wilson, Frank R., La mano. De cómo su uso configura el cerebro, el lenguaje y la cultura humana, Barcelona, Tusquets, 2002.
105 Chauchard, Paul, El cerebro y la mano creadora, Madrid, Narcea, 1972; VV.AA., «De la mirada al gesto. Cómo dirige el cerebro la mano», Mundo científico, 1998, n.º 192, 25-37.

230
© Universidad de Deusto - ISBN 978-84-9830-383-4
imprescindible para entender la función de la mano situarla en su relación y diálogo con el cerebro106. El cerebro, como órgano específico de la in-teligencia, necesita para llevar a cabo sus planes de órganos mediadores, y uno de ellos, sin duda el más importante, ha sido la mano. De ahí que Pierre-Gilles de Gennes, señale que «la inteligencia surgió en el hombre porque tenía manos y podía utilizarlas para hacer cosas»107.
Hemos tendido a considerar la inteligencia y la racionalidad como una propiedad abstracta y al margen del contacto con la realidad mate-rial, cuando de hecho comprende todo lo que el ser humano ha tenido que realizar para sobrevivir. De ahí que el avance de la inteligencia y el creci-miento del cerebro se han ido configurando en relación y diálogo con la realidad material, a través de la mano y del resto del cuerpo, como media-dores. De tal forma que la inteligencia y el cerebro, en la medida en que crecen y se desarrollan, orientan con mayor precisión a la mano, y ésta, en su contacto directo con la realidad, le lleva una mejor y más precisa infor-mación al cerebro, produciéndose en este círculo virtuoso un aumento de inteligencia y de precisión manipuladora.
Y la consecuencia de ese perfeccionamiento es que la mano, guiada por el cerebro, ha rebasado sus posibilidades a través de las herramientas, que se convierten en la mano artificial, o en la prolongación de las manos, como veremos más adelante. De ahí que la técnica humana, tan sofistica-da en la actualidad, sea tanto una prolongación y materialización del cere-bro como de la mano108.
Y para completar el panorama, es importante ver en qué medida la mano fue contribuyendo a la transformación de la cabeza y la cara de los humanos.
3.4. La transformación de los rasgos faciales
Una de las partes de la morfología humana que más ha evolucionado respecto a sus antecesores es la cabeza y la cara, en particular. Basta ver las ilustraciones que los paleoantropólogos nos muestran de reconstruc-ciones de caras de australopitecos y de las primeras especies humanas,
106 Es muy sugerente a este respecto la descripción sobre el diálogo entre la mente y la mano en el trabajo de un alfarero, que José Saramago nos relata en su novela La caverna,Barcelona, Alfaguara, 2001, pp. 105 y ss.
107 Entrevista de M. Ruiz de Elvira, en El País, 22-mayo.1991, p. 33. De Gennes fue Premio Nobel de Física en 1991, pero sus investigaciones se extienden también al ámbito de la antropología.
108 Cfr. Gubern, Román, El simio informatizado, Barcelona, Fundesco, 1987.

231
© Universidad de Deusto - ISBN 978-84-9830-383-4
por no referirnos ya a los simios antropomorfos109. Las transformaciones han sido muchas y de muy diverso tipo, motivadas también por muy di-versas circunstancias.
La bipedestación y su consecuencia, la utilización de la mano para manipular alimentos y herramientas, produjeron como efecto directo que fueran realizados por las manos muchos cometidos que antes se rea-lizaban con las mandíbulas: atacar y defenderse de las agresiones de los depredadores, cortar y rasgar la carne para comerla en trozos pequeños, manipular diferentes objetos, etc. Todos estos cometidos los va a rea-lizar progresivamente la mano. Las ventajas de ello van a ser conside-rables: desde la mayor precisión en partir las cosas y los alimentos, así como en cortar objetos para manipularlos, no tener necesidad de reali-zar defensas o ataques en contacto directo tan de cuerpo a cuerpo con el enemigo, puesto que ahora el brazo y la mano pueden establecer una cierta distancia, y más aún cuando la mano añade un arma o una herra-mienta, etc.
Y como consecuencia de estos beneficios, hay que advertir los cam-bios que con ello se producen. Si el proceso evolutivo va dotando de una poderosa estructura ósea, con su correspondiente masa muscular, a las mandíbulas de los carnívoros que hacen de ellas un arma sanguina-ria, la no utilización de las mandíbulas para esos menesteres hará que se vaya aligerando tanto esa masa ósea como la muscular. De ese modo, tampoco se necesitarán potentes huesos de la estructura craneana que hagan de soporte donde los tendones de la potente musculatura se ten-gan que apoyar. Se produce con ello lo que algunos han denominado el descerrojamiento del cerebro, ya que la potente estructura ósea anterior dará lugar y espacio para el avance de la masa encefálica. Por tanto, es significativo el diferente modo de configurarse el conjunto de la cabeza y la proporción dentro de ella del encéfalo. Se daría un movimiento en el que en los humanos se adelanta la frente (para dar cabida al importan-te aumento del lóbulo frontal) y retrocede y levanta la parte posterior de la cabeza (menor proporción de los lóbulos occipitales). Eso se acompa-ña también con el retroceso de la mandíbula, que dejará de tener la for-ma del morro, propia de los carnívoros depredadores, para alcanzar la forma de la mandíbula humana.
De este modo, la cabeza del ser humano va pasando de tener la forma aplanada propia de los simios, a una más erguida y estrechada. Igualmen-te, la cara pasa de presentar una orientación inclinada, a poseer un traza-
109 Cfr. Arsuaga, J. L./Martínez, N., La especie elegida, o.c., pp. 120 y ss; Lewin, R., La interpretación de los fósiles, o.c., pp. 169 ss.; Johanson, D./Hedí, M., El primer antepa-sado del hombre, o.c., pp. 252-53 y 314 y ss.

232
© Universidad de Deusto - ISBN 978-84-9830-383-4
do vertical, por adelantamiento de la frente y retroceso de la mandíbula, como hemos señalado110.
Capítulo aparte, por su interés y su complejidad, es el referente al de-sarrollo de la dentición en los seres humanos. Las investigaciones paleoan-tropológicas le dan cada vez más importancia al análisis de la dentición fósil, porque nos revelan costumbres alimenticias, edad, enfermedades, etc.111. En la línea de la hominización, vimos en su momento que se fue pasando de una dieta alimenticia fundamentalmente hervíbora en los si-mios y australopitecos, a otra de progresivo contenido carnívoro en los hu-manos, sea procedencia del carroñeo, al principio, o de la caza directa des-pués. Este cambio alimenticio influyó en la cerebralización y en la propia morfología de la dentición112. Un elemento fundamental es el referente a la relación entre el retroceso de la mandíbula y la lengua con profundización de la laringe, fenómeno fundamental que llevó al origen de la capacidad de hablar, propia del género humano, sin que se pueda en la actualidad dis-cernir la especie humana en la que se dio por primera vez este importante avance113. Pero estos aspectos los veremos en el capítulo siguiente.
En resumen, las transformaciones faciales y de la cabeza, en su totali-dad, fueron inmensas y progresivas, llegando a lo largo de este recorrido a conformarse una cabeza como la que poseemos los humanos actuales. Vemos, por tanto, cómo la interinfluencia entre la bipedestación, la utili-zación inteligente de la mano y el crecimiento del cerebro llevaron a una fuerte metamorfosis de la cara y de toda la morfología corpórea de los se-res humanos, hasta desembocar en la forma corpórea que nos caracteriza en la actualidad como especie. Hoy nadie duda de que el desencadenante de este largo proceso de transformación morfológica es el inicio del cami-nar erguido, que sirvió de detonante y de arrastre de todos los demás ras-gos. Pero dada la señal de partida, en la carrera se fueron intercambiando los demás factores señalados para ir influyéndose de forma imbricada y conformar el resultado que somos nosotros, los humanos actuales.
110 Cfr. Arsuaga, J. L./Martínez, N., La especie elegida, o c., pp. 89, 112-113, 114, 120, 122, 136, 139, 141256 276, 306-307.
111 Cfr. Bermúdez de Castro, J. M.ª, El chico de la Gran Dolina. En los orígenes de lo humano, Barcelona, Crítica, 2002, cap. 7, «El reloj biológico de los dientes», pp. 93-109; Bermúdez de Castro, J. M.ª/Sarmiento, S./Cunha, E., «Dimorfismo sexual en dientes hu-manos», Mundo Científico, 2000, n.º 214, 17-21.
112 Para las características específicas de la dentición de los humanos, cfr. Bermúdez de Castro, José M.ª, El chico de la Gran Dolina. En los orígenes de lo humano, o.c., pp. 78-80; 93-109.
113 Cfr. Arsuaga, J. L./Martínez, N., La especie elegida, o.c., cap. 14, en especial las pp. 306-307, donde se advierte la diferencia entre el denominado tracto vocal de un chim-pancé y un humano.

233
© Universidad de Deusto - ISBN 978-84-9830-383-4
Ahora bien, en la propia transformación morfológica no intervinieron solamente esta coimplicación de rasgos y aspectos morfológicos, sino que, como ya dejamos apuntado en el comienzo de este apartado, también se mezclaron factores conductuales y culturales, que fueron surgiendo al mis-mo tiempo que las transformaciones morfológicas y producidos por ellas, pero a la vez los iban cambiando y originando profundas y nuevas transfor-maciones y progresos. De este modo, lo morfológico y lo conductual/cultu-ral son dos caras o vertientes de la misma compleja realidad de los huma-no, vertientes que fueron actuando de forma cómplice y complementaria.
4. ¿AZAR O FINALIDAD EN LA EVOLUCIÓN Y EN EL UNIVERSO?
4.1. La cuestión acerca del azar o la finalidad en la evolución
Tras haber presentado el proceso filogenético del género humano, y el conjunto de cambios genéticos y morfológicos que supuso la aparición de nuestra especie, es importante que nos centremos ahora en la cuestión de si se puede advertir en ese proceso, y en el conjunto de la evolución, así como en la historia del universo, un proceso finalístico, teleológico, o se puede por el contrario concluir que ha sido todo fruto del azar y de re-laciones causales probabilísticas.
La teoría de la selección natural no ha pretendido ser para muchos de sus seguidores sólo un modelo o paradigma científico, sino que implicaba también un modo de entender el mundo prescindiendo de toda referen-cia a un supuesto diseñador inteligente. Desde muy antiguo han estado en conflicto dos modos de entender el universo y el desarrollo de las causas naturales: el modelo teleológico, según el cual se pensaba que Dios creaba y orientaba el universo, actuando en él como una causa segunda más, y el modelo natural, que defendía que en el mundo no se daba más que la inte-rrelación de causas físicas, sin referencia a ningún tipo de causa sobrena-tural o milagrosa, puesto que escaparía a una explicación empírica, propia del método de las ciencias físico-matemáticas.
Ahora bien, entre los defensores del modelo natural, se daban a su vez dos modelos alternativos: el determinismo y el azar (el juego de pro-babilidades). El conflicto entre estos dos modelos comienza ya entre los filósofos griegos114. Mientras Parménides defendía que todo está predeter-
114 Cfr. Fernández Rañada, Antonio, Los científicos y Dios, Madrid, Trotta, 2008, cap. 4.º,«El azar y la necesidad», pp. 87 y ss.; Wagensberg, Jorge (ed.), Proceso al azar, Barcelona, Tusquets, 1986.

234
© Universidad de Deusto - ISBN 978-84-9830-383-4
minado y orientado por leyes precisas, para Heráclito todo está sometido a fluidez y fugacidad. Leucipo y Demócrito defendieron tesis intermedias, puesto que su teoría atomista suponía que las cosas están hechas de áto-mos (pequeños corpúsculos indivisibles), cuya extrema dureza explicaría la tesis de Parmánides sobre lo fijo y permanente, pero sus incesantes co-lisiones introducirían el cambio constante regido por las leyes del azar, tal y como defendía Heráclito. De ahí que Demócrito afirmara que todo se debe al azar y a la necesidad, un materialismo irreligioso que indignó a Platón, pero que anticipaba una postura muy actual. De alguna manera, la historia de la física ha sido, en gran medida, la historia del esfuerzo por determinar cuánto hay en el universo de cada uno de los aspectos, azar o necesidad.
Tras la época griega y medieval, dominada por una concepción fina-lística del universo, bien sea apoyada en los grandes filósofos griegos o en la cosmovisión cristiana, la ciencia moderna, surgida a partir del Re-nacimiento, se nos presenta como opuesta al aristotelismo y a su postura finalística, en la medida en que va triunfando el mecanicismo, basado en la hipótesis de que el funcionamiento de la naturaleza es explicable en tér-minos de los movimientos de sus corpúsculos materiales. Es la tesis me-canicista defendida por Descartes, Galileo, Pascal, Copérnico y Kepler, hasta desembocar en Newton.
Para el mecanicismo, si conociéramos la posición y la velocidad de las partículas de cualquier sistema en un momento dado, la situación del conjunto del universo quedaría claramente determinada y podríamos saber sus valores y datos pasados y futuros. Bastaría aplicar las leyes del movi-miento. Los presupuestos teóricos fundamentales que están debajo de este modo de entender el mundo son, por un lado, la idea de que el tiempo y el espacio son realidades absolutas, y, por otro, la teoría del movimiento continuo. Este determinismo físico era para Newton expresión de la orga-nización con que Dios había creado el universo, aunque también pensaba que Dios intervenía de vez en cuando en el universo para corregir mo-mentos de inevitable inestabilidad.
El paradigma perfecto de la concepción determinística lo defendió el marqués Pierre Simón de Laplace (1749-1827), yendo más allá de Newton y defendiendo que todo el universo sigue leyes totalmente esta-bles y previsibles, no necesitándose echar mano de la hipótesis de Dios. Para Laplace, todo el comportamiento del sistema solar es explicable y calculable mediante la aplicación de técnicas matemáticas de la teoría de Newton sobre la gravitación universal. El problema para Laplace es-taba, por tanto, no tanto en que las leyes del universo pudieran en algún momento fallar y necesitar de una corrección (según Newton), sino en la dificultad de poseer la inmensidad de conocimientos de todos los detalles

235
© Universidad de Deusto - ISBN 978-84-9830-383-4
sobre la situación e interacción de cada partícula del universo con las de-más. Sabiendo la situación y la fuerza de interacción de cada partícula con las demás, se podrían conocer las secuencias sucesivas de todas las piezas de esa gran máquina que constituye nuestro universo.
Así, el deísmo de los primeros autores mecanicistas, se fue superando tras Laplace, prescindiéndose totalmente de la necesidad de un Dios que pone en marcha el mecanismo del cosmos. La polémica estará ahora entre la concepción determinista que pone en cuestión la libertad humana, en-tendiéndola incluso como un engaño del ser humano, entendible como un modo de actuar dirigido por la estructura biológica y genética, o la defen-sa de la libertad dentro de un mundo totalmente conformado por leyes de-terminísticas y cerradas. Todo funciona como un reloj, cuyos engranajes y piezas son los átomos y las moléculas. Así se generaliza la idea de que el mecanicismo, con su visión materialista y atea del mundo, era la única teoría compatible con las modernas teorías científicas. Esta visión meca-nicista del mundo se unió a la generalizada creencia, propia del siglo XIX,en el continuo e inevitable progreso de la historia humana, fruto de la po-derosa y humanizadora racionalidad científica, que otorgaban a los seres humanos una ilimitada esperanza en un futuro mundo cada vez mejor y en el que se conseguirían los sueños de todos los humanos. Al menos los de todos aquellos que sobrevivieran a la implacable lucha por la superviven-cia inscrita en un mundo ferozmente competitivo.
En un primer momento, había una resistencia a incluir a los seres vivos en esta concepción mecanicista, en la medida en que se los en-tendía como realidades más complejas que los objetos inanimados, y movidas por un dinamismo autónomo, difícilmente reducibles a los modelos del mecanicismo físico. La aportación de Darwin consistió precisamente en proponer un modelo de interpretación del mecanismo de los seres vivos con el que se conseguía extender la visión naturalista y mecanicista al conjunto de la biosfera, en la que se incluía también a los seres humanos115.
4.2. La recuperación del azar y de la probabilidad116
La teoría de la selección de Darwin significaba la coronación definiti-va del mecanicismo, puesto que suponía el sometimiento al mismo del úl-timo reducto que quedaba, el ámbito de los seres vivos, descubierta ahora
115 Cfr. Ayala, Fr. J., Darwin y el Diseño Inteligente, Madrid, Alianza, 2007, cap. 1.º, «Diseño inteligente», pp. 31 y ss.
116 Cfr. Fernández-Rañada, A., o.c., pp. 96 y ss.

236
© Universidad de Deusto - ISBN 978-84-9830-383-4
su regulación por leyes autónomas y bien precisas, al margen de interven-ciones sobrenaturales. Además, significaba también la ampliación al mun-do de la vida de los planteamientos deterministas, aunque sólo sea por la imposición definitiva del ambiente sobre la pluralidad de los rasgos gené-ticos transmitidos por generación.
Pero este modo de ver las cosas empezó muy pronto a revelarse como una visión demasiado simplista del mundo. El cambio, de todas formas, cabalgaba sobre una aparente contradicción. Mientras se llega a la apo-teosis y al momento del máximo éxito de la aceptación del mecanicismo determinista, cuya cima se da en las ciencias del momento, y, en el ámbi-to de la filosofía, de la mano del ateísmo antropológico de L. Feuerbach y sus seguidores, se estaba produciendo el nacimiento de la corriente con-traria que echará abajo el simplismo de la visión determinista.
Este cambio se produjo en tres ámbitos diferentes: en primer lugar, en el descubrimiento de los sistemas complejos; en segundo lugar, con el nacimiento de la nueva física, dominada por la mecánica cuántica, y, por último, con el estudio de los sistemas caóticos (la teoría del caos y de las catástrofes), en los que el determinismo básico se advierte operativo sólo en intervalos limitados de tiempo.
Antes de referirnos brevemente a estos tres campos, tenemos que de-jar claro qué entendemos por azar. Azar es el caos, la total ausencia de normas, reglas o leyes. El azar de nuestro mundo no parece ser de este tipo tan radical. Para ello, necesitamos distinguir entre conceptos como azar, probabilidad y determinismo, como lo hace J. D. García Bacca117.El azar estaría constituido por un sistema en el que se da irracionalidad en las partes y en el todo. Esto es, en ese sistema no existe ningún por qué como respuesta a cualquiera de sus acontecimientos. Las cosas suce-den porque sí. En cambio, se da un sistema de probabilidad cuando exis-te irracionalidad en las partes pero racionalidad en el todo. Sería el caso del juego de los dados. En cada tirada del dado no sabemos nunca (a me-nos que esté trucado) qué número va a salir, pero se da una ley de proba-bilidad por la que, tras un número grande de tiradas, lo probable es que el número de veces que salga cada una de las seis caras del dado se vaya igualando. En cambio, el ámbito del determinismo se da cuando existe ra-cionalidad en las partes y en el todo. Se trata de sistemas donde todo está regulado por leyes fijas y bien especificadas.
El determinismo clásico admitía el azar subjetivo (debido a nuestra ignorancia), pero no el objetivo. Pero hoy día la ciencia admite también
117 Cfr. García Bacca, J. D., Curso sistemático de filosofía actual, Caracas, Universidad Central de Venezuela, 1969, pp. 169 y ss.

237
© Universidad de Deusto - ISBN 978-84-9830-383-4
el azar objetivo en los ámbitos donde domina la probabilidad y la esta-dística. Así lo advirtió A. Cournot (1801-1877) en el ámbito de las for-mulaciones matemáticas aplicadas a la economía y a otros campos, don-de aparece el azar como colisión de cadenas causales independientes. Así surgió la mecánica estadística de la mano de J. C. Maxwell, Bolzmann y W. Gibbs. Del determinismo simplista de Laplace se salta a un nuevo de-terminismo basado en la teoría estadística, que nos muestra que los siste-ma tienden a situarse en su estado más probable, sobre todo en el ámbito de los grandes números. Este nuevo determinismo estadístico fue puesto de moda por médicos y biólogos, siendo Claude Bernard (1813-1878) quien lo puso de moda al hablar de la aparición de enfermedades por fac-tores determinantes.
Pero el desprestigio definitivo del determinismo se produjo de la mano de la teoría cuántica, construida por Max Planck, N. Bohr, W. Heisenberg, Dirac, Schrödinger, De Brogie, M. Born y otros118. Tanto el principio de indeterminación de Heisenberg (1901-1976), como el de complementarie-dad de N. Bohr, nos indican que la realidad subatómica funciona por leyes probalísticas y no determinísticas, presentándosenos ese ámbito de la rea-lidad conformado por leyes que contradicen el mundo de nuestra percep-ción sensorial. Esto rompe el principio idealista de pensar que podemos entender y apresar con nuestra lógica la realidad. La física actual nos hace ver que lo real rebasa sobradamente nuestra razón y el sentido común de nuestras intuiciones. Por tanto, al final nos queda el interrogante de si po-demos conocer lo real en sí, y, en definitiva, en qué consiste la realidad última de lo que hay. Frente al realismo (el mundo existe por sí mismo) y al positivismo (las cosas sólo existen en cuanto son percibidas), pare-ce más adecuada una tercera postura intermedia, defendida por Bernard d’Espagnat, quien nos habla de una realidad velada y lejana, que sustitu-ye a lo real empírico, reflejando sólo débilmente lo que son las cosas en sí, aunque diciendo lo que no son. En palabras de B. d’Espagnat: «Parece muy legítimo calificarlo (lo real en sí) como inconocible y velado. Pero de las dos palabras, es la segunda la que me parece la más correcta (…). Lo real en sí, aunque no es conocible en el sentido habitual de la palabra, no es tampoco rigurosamente inconocible; está velado»119.
Einstein nunca quiso aceptar el indeterminismo cuántico, aferrán-dose a la concepción determinística del mundo, en coincidencia con los planteamientos de Spinoza y basándose en una idea cerrada del univer-
118 Cfr. Lindley, David, Incertidumbre. Einstein, Heisenberg, Bohr y la lucha por la esencia de la ciencia, Barcelona, Ariel, 2008.
119 D’Espagnat, B., Une incertaine réalité, París, Gauthier Villar, 1985, p. 269. Cita to-mada de A. Fernández Rañada, o.c., p. 105.

238
© Universidad de Deusto - ISBN 978-84-9830-383-4
so, creado y ordenado sabiamente por Dios, entendido como una inteli-gencia impersonal que lo controla todo. De ahí que Einstein se negara a aceptar la visión probabilística de la realidad presentada por la mecá-nica cuántica, con el conocido argumento de que «Dios no juega a los dados»; a lo que Bohr contestaba con ironía: «Deja de decirle a Dios lo que debe hacer»120.
A pesar de todos estos avances en física cuántica, el determinismo se-guía dominando en el ámbito de lo macroscópico y supra-atómico, porque las leyes de lo subatómico no rigen el mundo macroscópico. Pero también en este campo se van descubriendo ámbitos regidos por la probabilidad y la indeterminación, como lo han demostrado la teoría del caos y de las ca-tástrofes121.
Cuando el matemático E. N. Lorenz intentaba, en 1963, entender y desentrañar las leyes que rigen la endiablada complejidad de la meteorolo-gía, llegó a formular lo que se denominó a partir de sus trabajos el efectomariposa. Como todos sabemos, los fenómenos meteorológicos dependen de una enorme cantidad de variables. Por eso, resulta muy difícil predecir los acontecimientos meteorológicos, ya que de un cambio pequeño en el inicio, se derivan un amplio abanico de consecuencias, difíciles de pre-decir y de controlar. Así, va creciendo más rápido el número de variables que el esfuerzo matemático por controlarlo. A ese movimiento desorde-nado, en el que crecen de modo inexorable los errores, se ha denominado caos. Eso nos lleva a constatar que, como indica Ilya Prigogine122, vemos el mundo a través de una ventana temporal limitada.
Siendo así las cosas, hoy día se admite que el caos es ubicuo. Somos conscientes de que está formando parte de todos los fenómenos del mun-do. El mundo, por tanto, se nos presenta en la actualidad como un sistema abierto (Popper)123, impredecible, y la metáfora del reloj que se utilizaba durante el siglo XIX para simbolizar el supuesto perfecto funcionamiento
120 Cfr. Sampedro, J., El siglo de la ciencia. Nuestro mundo al descubierto, Barcelona, Península, 2009, 58.
121 Cfr. Thom, René, Estabilidad estructural y morfogénesis. Ensayo de una teoría ge-neral de los modelos, Barcelona, Gedisa, 1987; Espinosa, Miguel, «René Thom: de la teo-ría de las catástrofes a la metafísica», en Arana, Juan (ed.), La filosofía de los científicos,Sevilla, Thémata. Revista de Filosofía, 1995, n.º 14, pp. 321-348.
122 Cfr. Prigogine, Ilya-Stengers, I., La nueva alianza. Metamorfosis de la ciencia, Ma-drid, Alianza, 1983; Prigogine, I., ¿Tan sólo una ilusión?, Barcelona, Tusquets, 1983; Que-raltó, Ramón, «El nuevo orden del tiempo. En torno a la obra de Ilya Prigogine», en Arana, Juan (ed.), o.c., pp. 305-320.
123 Cfr. Popper, Karl, El desarrollo del conocimiento científico. Conjeturas y refuta-ciones, Barcelona, Paidós Ibérica, 1982; Id., La miseria del historicismo, Madrid, Alianza, 1973; Id., La sociedad abierta y sus enemigos, Barcelona, Paidós Ibérica, 1982.

239
© Universidad de Deusto - ISBN 978-84-9830-383-4
del universo, ya no sirve. Popper contrapone a la metáfora del reloj la de la nube. Nuestro realidad es tan proteica como una nube124.
Lo curioso de todo esto es que tanto la visión mecanicista como la pro-babilística sirven al mismo tiempo para deducir de ellas la existencia de Dios como para negarla. En el primer caso, la existencia de Dios resultaría, en cierta manera, más fácilmente deducida si nos hallamos ante un mundo organizado con un sistema de leyes cerradas y determinísticas, puesto que eso nos llevaría a postular un creador de tales leyes y de ese orden tan per-fecto. Es la postura que defendía Einstein, siguiendo a Spinoza. En cam-bio, para la otra postura, Dios sería más comprensible dentro de un mundo abierto, sin estar encerrado en leyes inmutables, porque Dios se situaría por encima de esas leyes que encorsetarían y limitarían su omnipotencia.
En cambio, los defensores del materialismo filosófico consideran que la necesidad y el azar son suficientes para entender y gobernar el mundo, y que la realidad de Dios sobra en este modo de ver las cosas. Otros, por el contrario, creen que la refutación del determinismo da paso al recono-cimiento y a la defensa del libre albedrío y a la existencia de Dios. Pero entendemos que lo más acertado es pensar que la realidad de Dios no depende, ni se demuestra, desde una teoría científica, sino que tanto un modo como el otro de ver el mundo por la ciencia son compatibles tanto con la hipótesis de Dios como con su negación.
4.3. La finalidad teleonómica en el mundo de la biología
Ya vimos más arriba cómo la teoría de la selección natural de Darwin supuso la introducción, en el ámbito de la biología, de la racionalidad científica, sometiendo el ámbito de la biosfera a las leyes de la ciencia, y desterrando de ese ámbito las referencias milagreras o sobrenaturales. Pero el reducir lo biológico a las leyes científicas, negando en el proceso evolutivo una referencia a una inteligencia consciente diseñadora del pro-ceso y de las diferentes especies, no suprime la cuestión acerca del azar o la finalidad en el mundo de la biología.
La cuestión en este caso está en si se puede reducir lo biológico a las leyes de la física, o si, por el contrario, el mundo de la vida no está con-figurado por leyes específicas, entre las cuales se halla un cierto tipo de finalismo, que, para diferenciarlo del auténtico dinamismo teleológico,se denomina teleonomía. Tal es la opinión de F. J. Ayala, entre otros, a la
124 Cfr. Popper, Karl R., El universo abierto. Un argumento en favor del indeterminis-mo, Madrid, Tecnos, 1984.

240
© Universidad de Deusto - ISBN 978-84-9830-383-4
hora de diferenciar lo biológico de lo físico, y de especificar el carácter específico del mundo de la vida125.
En el ámbito de la biología, desde los años veinte del pasado siglo, se han disputado la supremacía sobre el modo de entender y explicar la diná-mica vital tres planteamientos rivales: el vitalismo, el reduccionismo y el emergentismo. Las posturas vitalistas estaban defendidas por científicos y filósofos de la talla de Hans Driesch y de Henri Bergson, quienes propug-naban la existencia de una fuerza vital, un élan vital específico, responsa-ble de la especial forma de ser y de comportarse de los seres vivos126. Esta postura vitalista no se tomó nunca excesivamente en serio, y posterior-mente ha evolucionado hacia posturas dualistas dentro de la filosofía de la mente, como es el caso del neurofisiólogo J. Eccles, continuando las tesis dualistas de Descartes y sus seguidores127.
Mucho más influyente es la postura reduccionista, seguida en la ac-tualidad por una amplia mayoría de científicos y filósofos. Su tenden-cia a reducir a la leyes físicas el conjunto de fenómenos existentes en el ámbito de la biosfera había recibido, como nos indica G. Armengol, dos apoyos recientes de gran peso: «Primero el desarrollo de la bioquímica de los ácidos nucleicos (ADN) que han permitido entender que la vida se construye desde un mecanicismo estricto (herencia y embriogénesis). Se-gundo los formalismos computacionales que han conducido a las teorías computacionales de la vida: la complejidad observacional de ésta (muy difícil de explicar hasta ahora por el mecanicismo clásico del XIX) deri-varía de que la evolución habría diseñado los seres vivos como computa-dores biológicos»128. La tesis reduccionista sigue siendo la postura más extendida en el mundo de la biología, basándose en una idea del hombre máquina, explicitada en el s. XVIII por Lametrie, y reforzada en la actua-lidad por los avances de la genética y del funcionalismo computacional en el terreno de la filosofía de la mente.
125 Cfr. Ayala, Fco. J., La evolución de un evolucionista. Escritos seleccionados, Valen-cia, Universidad de Valencia, 2006, pp. 105-122.
126 Cfr. Bergson, Henri, La evolución creadora, Madrid, Espasa-Calpe, 1973; Ortega y Gasset, J., «Ni vitalismo ni racionalismo» (1924), Obras Completas, vol, III (Madrid, Re-vista de Occidente, 1966), pp. 270-280.
127 Cfr. Cfr. Eccles, J. C./Popper, K. R., El yo y su cerebro, Barcelona, Labor, 1980; Eccles, J. C./Zeier, H., El cerebro y la mente, Barcelona, Herder, 1984; Eccles, J. C., TheHuman Psyche, Londres, Routledge, 1992; Id., Evolution of the Brain. Creation of the Self,Londres, Routledge, 1995; Candela, J. A., «El dualismo interaccionista», en Dou, A. (ed.), Mente y cuerpo, Bilbao, Mensajero, 1986, 33-44; Gutiérrez Martínez, A., «El dualismo in-teraccionista del neurofisiólogo católico John Carew Eccles», La Ciudad de Dios, CCXIV (2001), 189-205 (1.ª parte), 295-346 (2.ª parte).
128 Armengol, G, «El emergentismo, una vía humanista de la ciencia», en http://tendencias 21/tendenciasdelasreligiones, 23-08-2007, pp. 1-4; 1.

241
© Universidad de Deusto - ISBN 978-84-9830-383-4
Pero difícilmente se puede defender que el ámbito de la biología, ca-racterizada por dinámicas teleológicas o teleonómicas, se pueda reducir en su totalidad a las leyes de la física. De ahí que surgiera con fuerza la tercera postura, la emergentista. Los defensores del emergentismo consi-deran que el primer emergentista en el terreno de la biología fue el propio Darwin, en la medida en que defendía saltos cualitativos correspondientes a la aparición de cada una de las especies vivas, como formas específicas y novedosas de organizarse la materia viva. El argumento básico para de-fender la irreductibilidad de lo biológico a lo físico se centra en el hecho de que en lo biológico se da la presencia de la teleología. Aunque hay que ser conscientes de que existen diferentes tipos de teleologías.
Ayala considera que los organismos biológicos se mueven con patro-nes de explicación teleológicos, que no existen en el ámbito de lo inani-mado, y que no pueden explicarse desde el punto de vista de las leyes fi-sicoquímicas, sin que se pierda en el camino algún importante contenido explicativo.
Para poder sostener estas afirmaciones, tenemos que definir antes el concepto de teleología. Ayala es consciente de que este término «está desacreditado» en el entorno de la ciencia moderna, siendo visto por mu-chos como «una señal de superstición», no sometible a verificación empí-rica y como un resto de «una era precientífica»129. Pero esto se debe a que se piensa que una acción teleológica está causada por un agente externo y no inmanente al propio organismo. Precisamente uno de los logros de la teoría de la selección natural de Darwin, entendida como motor de la evolución, consistió en sustituir en el ámbito de la biología una teleología teológica o dualista por otra de tipo científico. «La teleología de la natu-raleza se podría explicar ahora, al menos en principio, como resultado de leyes naturales que se manifiestan en procesos naturales, sin recurrir a un Creador externo o a fuerzas espirituales o inmateriales. En este punto la biología maduró como ciencia»130.
Por tanto, si en el ámbito de las acciones humanas advertimos una es-tructura teleológica en la que interviene un agente autoconsciente y con racionalidad prospectiva y anticipadora, no ocurre lo mismo en el ámbi-to de los fenómenos biológicos, teniendo que diferenciarse entre distintos modos de entender la teleología. Todos los movimientos teleológicos tie-
129 Ayala, F. J., «La biología como una ciencia autónoma», en Id., La evolución de un evolucionista. Escritos seleccionados, o.c., pp. 105-122. De todos modos, quizás fuera me-jor referirse a teleonomía para hacer referencia a los dinamismos biológicos, reservando teleología para las acciones humanas, en las que se da un sujeto consciente que prevé y ejecuta una acción.
130 Ibídem, p. 113.

242
© Universidad de Deusto - ISBN 978-84-9830-383-4
nen en común el ser «acciones, objetos o procesos que exhiben una orien-tación hacia cierto objetivo o estado final»131. En función de ello, es fun-damental que «el objeto o proceso contribuya a la existencia de un cierto estado o propiedad del sistema»132. Por eso, se puede hablar de teleología en el funcionamiento de un riñón, y no tanto en los movimientos de un planeta o en una reacción química.
Por eso, es fundamental, para aclarar conceptos, distinguir entre tres tipos de fenómenos teleológicos, en función, como indica Ayala, de la re-lación que se da entre la estructura o el proceso y la propiedad o estado fi-nal. Siguiendo este criterio, Ayala distingue tres tipos de teleologías133:
1) Teleología consciente: es aquella en la que «el estado final u ob-jetivo es conscientemente anticipado por el agente»134. Algunos utilizan para este tipo de fenómenos el concepto de racionalidad prospectiva135 para referirse a este tipo de teleología, que se daría exclusivamente en los seres humanos, y es posible que de modo incipiente en ciertos primates. Los negadores de la existencia de procesos teleológicos en la biología, posiblemente lo hacen debido a que reducen este concepto a las situaciones en las que se va una previsión y anticipación conscientes.
2) Teleología de sistemas autorregulados o teleonómicos, cuando se da un «mecanismo que permite al sistema alcanzar o mantener una propiedad específica a pesar de las fluctuaciones del entorno»136.Un ejemplo de estos sistemas es la capacidad homeostática de los mamíferos para mantener estable la temperatura corporal. Es in-teresante hacer ver, por lo demás, que los biólogos distinguen dos tipos de homeostasis: fisiológica y de desarrollo. Los organismos que poseen la primera, tienen capacidad de mantener estables cier-tos estados fisiológicos. En cambio, la homeostasis de desarrolloes la capacidad de «regulación de las diferentes rutas que un orga-nismo puede seguir en su progreso de zigoto a adulto»137. El proce-so de gestación de los seres vivos pertenece a este tipo de homeos-
131 Ibídem, p. 114.132 Ibídem, p. 114. 133 Cfr. ibídem, 114-116.134 Ibídem, p. 114.135 Cfr. García Bacca, J. D., Curso sistemático de filosofía actual, Universidad Central
de Venezuela, Caracas, 1969, pp. 169 y ss. García Bacca distingue entre racionalidad pros-pectiva (teleológica) y retrospectiva, a posteriori. Cfr. Beorlegui, C., García Bacca. La au-dacia de un pensar (Bilbao, Universidad de Deusto, 1988), pp. 156-161.
136 Ayala, F. J., o.c., 115.137 Ibídem, p. 115.

243
© Universidad de Deusto - ISBN 978-84-9830-383-4
tasis, conformando una secuencia bien delimitada que atraviesa diferentes etapas orientadas hacia un objetivo claro, pero sin que este dinamismo esté regido por una instancia externa consciente. También hay que situar dentro de este tipo de procesos teleonó-micos los sistemas mecánicos fabricados técnicamente por el ser humano, como un termostato, en la medida en que están consti-tuidos con un sistema de autorregulación a través de un mecanis-mo de retroalimentación de la información. Pero en estos casos, se habla más bien de teleomática, esto es, un proceso teleológico ideado e impuesto por el ser humano dentro de un artefacto o má-quina cualquiera.
3) Un tercer tipo de teleología es la propia de los órganos o partes del cuerpo de los seres vivos, como la mano, el sistema circula-torio o respiratorio, etc. Todos ellos son ejemplos de «estructu-ras anatómica y fisiológicamente constituidas para realizar una cierta función»138. Al igual que los órganos fisiológicos, también pertenecen a este tipo de teleología las herramientas y artefac-tos construidos por los seres humanos, en la medida en que están conformados de un modo determinado para realizar una función específica. Como puede verse, la distinción entre estos dos últi-mos tipos de teleología es un tanto difusa, porque también el ter-cer tipo puede y suele tener ciertas capacidades de autorregula-ción y retroalimentación.
Si en el ámbito de los organismos vivos no se da teleología conscien-te, está claro que el resto de estructuras teleológicas y de homeostasis son resultado del proceso de adaptación al ambiente (selección natural). Son acomodaciones que se justifican en función de la contribución al proceso de reproducción de la especie. Pero son procesos mecanicistas e imperso-nales, con racionalidad retrospectiva, no prospectiva. Desde este punto de vista, también se puede hablar de dos niveles de teleología: específica y genérica. El dinamismo finalístico específico es el que permite relacionar el carácter o modo de ser de un organismo, o parte del mismo, con la fun-ción que desempeña. Pero el fin genérico y último perseguido por todos los caracteres y sus funciones es el éxito reproductivo. Por tanto, desde este punto de vista, «la fuente última de explicación en biología es el prin-cipio de selección natural»139.
Pero, como Ayala precisa, la selección natural puede considerarse tanto como un proceso teleológico, como también lo contrario. Es de-
138 Ibídem, p. 116. 139 Ibídem, p. 117.

244
© Universidad de Deusto - ISBN 978-84-9830-383-4
cir, por un lado, «la selección natural puede decirse que es un fenóme-no teleológico en un sentido causal. La selección natural no es una enti-dad sino un proceso puramente mecanicista. Pero se puede decir que la selección natural es teleológica en el sentido de que produce y mantie-ne órganos y mecanismos dirigidos hacia un fin, cuando las funciones que ejercen contribuyen a la eficiencia reproductiva del organismo»140.Pero no puede decirse que la selección natural sea teleológica en el sentido de que se oriente a la producción de determinados y específicos organismos o especies. En ese sentido, la selección natural es mecani-cista y oportunista. Y el resultado final de una especie puede ser tanto la adaptación exitosa como la extinción. La tendencia de la selección natural es dotar a los organismos de mecanismos eficientes de cara a la supervivencia, pero no realiza esta tendencia de modo consciente141.Ese estado final es causal y temporalmente posterior (racionalidad re-trospectiva).
De todos modos, una prueba de la complejidad del fenómeno teleo-lógico es que se pueden hacer múltiples clasificaciones, en función de los múltiples puntos de vista desde los que se examina. Si la clasificación an-terior la hacía Ayala desde la relación entre «el objeto o mecanismo y la función o propiedad a la que sirve», él mismo realiza otra clasificación complementaria, «atendiendo al proceso o agencia que da origen al siste-ma teleológico»142. Desde este segundo punto de vista, se puede hablar de teleología interna y externa. Otro modo de nombrarlas es, respectivamen-te, teleología natural y artificial. La primera es la propia de los organis-mos vivos y sus caracteres, y la segunda, la de las herramientas y servo-mecanismos humanos. Ya hemos dicho que algunos denominan a este tipo de teleología telemática.
Llegados a este punto, tenemos que preguntarnos si estos procesos teleológicos se pueden explicar desde la base de los procesos fisicoquí-micos. Parece evidente para Ayala que, aunque «las explicaciones teleo-lógicas son totalmente compatibles con las consideraciones causales», en realidad «la explicación teleológica connota algo más que su equiva-lente no teleológica»143, de modo que no se cumplen las dos condiciones propuestas por Nagel para que se dé un proceso correcto de reducción de una ciencia a otra. Nagel propone dos condiciones necesarias y sufi-cientes para poder considerar que el proceso de reducción epistemológi-ca es correcto, y las denominó condiciones de derivabilidad y de conec-
140 Ibídem, p. 117.141 Cfr. ibídem, p. 118.142 Ibídem, p. 118.143 Ibídem, p. 119

245
© Universidad de Deusto - ISBN 978-84-9830-383-4
tibilidad144. La primera afirma que «para llevar a cabo la reducción de una rama de la ciencia a otra, debe demostrarse que todas las teorías y leyes experimentales de la primera son consecuencias lógicas de los he-chos teóricos de la segunda»145. Por su parte, el principio de conectibi-lidad indica y manda que «todos los términos técnicos de la ciencia que quieren reducirse sean definidos otra vez usando términos de la ciencia a que se reduce la primera»146.
Un proceso teleológico, por tanto, está organizado para que persiga un fin, cosa que no se da en el ámbito de las relaciones meramente causales, esto es, en el ámbito de la fisicoquímica. Además, las explicaciones teleo-lógicas muestran la presencia de funciones específicas en el sistema y de una organización directora del mismo. Así, la forma de ser del corazón está en función de bombear la sangre. Y, en último término, la relación entre finalidad y medios está al servicio de una finalidad más genérica, fruto de la selección natural, como es el éxito reproductivo.
En conclusión, «el uso de las explicaciones en biología no es sólo aceptable sino de hecho indispensable. Los organismos son sistemas orga-nizados de forma dirigida»147. Y ante la objeción de algunos autores, que no ven demasiada claridad entre sistemas teleológicos y los que no lo son, Ayala propone como distinción entre ambos procesos el criterio de utili-dad, aplicable tanto para los casos de finalidad externa como interna. En el segundo caso, un sistema será teleológico «si el carácter es de utilidad para el sistema en el que existe y si tal utilidad explica la presencia del ca-rácter en el sistema»148, contribuyendo de ese modo a su adaptabilidad y al éxito reproductivo. Si nos referimos a la finalidad externa, diremos que la utilidad se comprueba y refiere al autor del sistema, en este caso, las herramientas y artefactos creados por los seres humanos: los artefactos son como son porque sirven y son útiles para realizar determinados fines. Así, «el criterio de utilidad introduce una objetividad necesaria en la de-terminación de qué mecanismos biológicos son dirigidos a un fin»149.
Eso no significa que todos los caracteres de un organismo vivo ten-gan utilidad (como a veces se ha defendido por algunos darwinistas), pero
144 Cfr. Nagel, E., The Structure of Science, Nueva York, Harcourt, Brace and World, 1961; Ayala, F. J., «Biology as an Autonomous Science», American Scientist, 1968, n.º 56, pp. 207-221 (trad. cast., «La biología como una ciencia autónoma», en Ayala, F. J., La evo-lución de un evolucionista. Escritos seleccionados, o.c., pp. 105-122; Id., «Introducción»,o.c., p. 13.
145 Ayala, F. J., o.c., p. 13. 146 Ibídem, p. 13. 147 Ibídem, p. 120. 148 Ibídem, p. 121. 149 Ibídem, p. 121.

246
© Universidad de Deusto - ISBN 978-84-9830-383-4
también se han dado casos en que determinados caracteres que no han sido adaptativos en el origen, pueden reconvertirse posteriormente en úti-les150. Esta es la idea que defiende Fr. Jacob al decir que la evolución se parece a un sistema de bricolage, en la medida en que no actúa de modo consciente y con racionalidad prospectiva, sino de modo chapucero a pos-teriori, acomodando lo que tiene para sacar de las cosas su mejor rendi-miento151.
En conclusión, así como los sistema biológicos no pueden explicarse del todo sin la referencia a la teleología (teleonomía), y de hecho son los únicos sistemas que tienen teleología interna, no sirve por el contrario la idea de teleología en el ámbito de la física y la química. Por ello, desde este punto de vista tan fundamental, es inadecuado defender que el ámbi-to de la biología se pueda reducir a explicaciones propias de las leyes fi-sicoquímicas. Así, «las explicaciones teleológicas, por tanto, diferencian a la biología del resto de las ciencias naturales»152. La conclusión es que la biología es un nivel emergente respecto a la física y la química, y no total-mente reducible a ellas. Y lo mismo hay que decir de lo psíquico, respecto de lo biológico.
4.4. ¿Finalidad o azar en la historia del universo?
No sólo tenemos que hacernos la pregunta sobre el proceso evoluti-vo en el mundo de la biosfera, sino que también es pertinente la cuestión referida a la historia del universo. ¿Tanto el origen del universo como su historia, desde el Big Bang (dando por hecho que esta teoría cosmológi-ca es la más aceptada por los científicos actuales153) hasta el presente, es
150 Cfr. Gould, Stephen Jay, El pulgar del panda. Reflexiones sobre historia natural y evolución, Barcelona, Crítica, 1994; Gould, S. J./Lewontin, R. C., «La adaptación biológi-ca», Mundo Científico, 1983, n.º 22, pp. 214-223. Algunos autores, como Gould y Lewon-tin, denominan a este fenómeno exadaptación: readaptación de un rasgo a una finalidad nueva y diferente a la que han tenido en especies anteriores en el proceso evolutivo. Cfr. también Dupré, J., El legado de Darwin. Qué significa hoy la evolución, Buenos Aires, Katz Editores, 2006, p. 58.
151 Cf. La lógica de lo viviente, Barcelona, Laia, 1977. 152 Ayala, F. J., o.c., p. 122. 153 Sobre las diferentes teorías cosmológicas, cfr. Lane Craig, W., «Naturalismo y cos-
mología», en Soler Gil, F. J. (ed.), Dios y las cosmologías modernas, Madrid, BAC, 2005, 49-99; Hawking, S., La historia del tiempo. Del Big Bang a los agujeros negros, Barce-lona, Crítica, 1988; Id., El universo en una cáscara de nuez, Barcelona, Crítica, 2002; Brockman, John (eds.), La tercera cultura (Más allá de la revolución científica), Barcelo-na, Tusquets, 1996; Ruiz de la Peña, J. L., Teología de la creación, Santander, Sal Terrae, 1986, cap. 8.º; Davies, Paul, Dios y la nueva física, Barcelona, Salvat, 1986; Id., Otros

247
© Universidad de Deusto - ISBN 978-84-9830-383-4
resultado de un proceso azaroso, o cabe hablar en este ámbito de finali-dad? Esta cuestión está íntimamente relacionada con la pregunta sobre el puesto del ser humano en el conjunto del universo. Es decir, ¿el uni-verso es cosmocéntrico o antropocéntrico? O dicho de un modo más ex-plícito: ¿el universo posee su propia finalidad, y se orienta y desarrolla al margen de la especie humana, o el fenómeno humano es el objetivo perseguido por esa explosión inicial desde la que procede todo el uni-verso conocido?
Vamos a analizar los argumentos de cada una de estas dos posturas, en dos etapas: en la primera, mostraremos la diferencia entre ambos plantea-mientos, para, en la segunda, presentar los principios teóricos en los que cada una se apoya, respectivamente: el principio cosmológico o coperni-cano, y el principio antrópico.
4.4.1. ¿Cosmocentrismo o antropocentrismo?154
Si contemplamos el universo en relación al hombre, advertimos una gran paradoja. Por un lado, si nos fijamos en las inmensas dimensiones que posee el universo, y en el carácter periférico del planeta Tierra, así como en el carácter eventual y fortuito de la emergencia del género hu-mano dentro de nuestro planeta, parecería que tenemos que sacar la con-clusión de que el ser humano es un mero accidente dentro de este azaro-so proceso tan largo y complejo. Pero, por otro, si tenemos en cuenta la cualidad probablemente única del surgimiento de la vida en el planeta (da igual si se confirmase la hipótesis de su procedencia de otro planeta), ello apoyaría la tesis de que el ser humano tiene que ser la cúspide de la pirá-mide evolutiva, el centro ontológico (no el espacial o temporal) de la tota-lidad de lo real. Veamos, pues, de modo más extenso los dos términos de esta antinomia.
mundos, Barcelona, Salvat, 1986; Küng, Hans, El principio de todas las cosas. Ciencia y religión, Madrid, Trotta, 2007; Pineda, José Avelino de la, El baile de la ciencia y la meta-física. Respuesta a Stephen Hawking, Madrid, Biblioteca Nueva, 2008; Sanguinetti, Juan José, «Algunos aspectos de la filosofía de los cosmólogos contemporáneos», en Arana, Juan (ed.), La filosofía de los científicos, o.c., pp. 285-304.
154 Cfr. Ruiz de la Peña, J. L., Teología de la creación, Santander, Sal Terrae, o.c., pp. 241 y ss.; Torres Queiruga, Andrés, Recuperar la creación. Por una religión humani-zadora, Santander, Sal Terrae, 1996; Küng, Hans, El principio de todas las cosas. Ciencia y religión, Madrid, Trotta, 2007; Sanguinetti, J. J., «Algunos aspectos de la filosofía de los cosmólogos contemporáneos», en Arana, Juan (ed.), La filosofía de los científicos, Sevilla, Thémata (Rev. de Filosofía), 1995, n.º 22, pp. 285-304; VV.AA., «Creador del cielo y de la tierra», número monográfico de Iglesia Viva, 1996, n.º 183.

248
© Universidad de Deusto - ISBN 978-84-9830-383-4
a) Razones del anti-antropocentrismo o de las tesis del cosmocentrismo
Su argumento central se basa en las enormes magnitudes que posee el cosmos. Las escalas del espacio euclidiano y del tiempo newtoniano se han agrandado de tal forma que poseen en la actualidad cifras sobre-cogedoras. Para que nos hagamos una mínima idea de ellas, hemos de tener en cuenta que la Vía Láctea posee una extensión de unos cien mil años luz (es decir, un rayo de luz tardaría ese tiempo en recorrerla de un lado a otro). Y el tamaño del grupo de las galaxias a las que pertenece, viene a ser de unos tres millones de años luz. Por otro lado, se considera que existen centenares de miles de millones de otras galaxias en el uni-verso observable, y los científicos no hacen más que ir descubriendo de continuo nuevas galaxias, aumentándose constantemente la dimensión del universo conocido por el ser humano. Siguiendo con estos datos, la nebulosa Andrómeda, la más cercana a nosotros, está a la distancia de un millón setecientos mil años luz. Y la estrella fija más cercana a no-sotros, Alfa Centauro, se halla a unos cuatro años luz dela tierra. A cada galaxia se le atribuye una media de cien mil millones de estrellas, de tal modo que, para que nos resulte más plástico, podemos decir que cada galaxia alberga más estrellas que habitantes ha tenido la tierra desde sus orígenes.
Si nos referimos ahora a las dimensiones temporales, los científicos nos indican que la edad del universo podemos calcularla entre 15.000 y 18.000 millones de años, aunque a veces algunos científicos la rebajan hasta 13.000 millones. La edad de la Tierra se sitúa entre 4 y 5 mil millo-nes de años.
A la vista de estos datos, la conclusión que parece imponerse, en pala-bras de J. L. Ruiz de la Peña, es que «somos un planeta secundario de un sol suburbial situado en el extrarradio de una galaxia irrelevante»155. Por tanto, la cuestión que se nos plantea es interrogarnos sobre si es racional seguir pensando que lo que ocurra en este pequeño planeta tiene alguna significación cósmica, esto es, alguna significación decisiva para el con-junto del universo. Parece lo más lógico deducir, a la vista de estos datos, que todo lo sucedido hasta ahora (desde el origen del universo, pasando por el origen de la vida, hasta la emergencia de la conciencia) se habría podido repetir en otros muchos lugares del espacio, en diversos momentos de la historia del universo. Es posible y razonable pensar así, aunque de hecho no sabemos nada acerca de ello.
155 Estas palabras que recoge J. L. Ruiz de la Peña, o.c., p. 242, reflejan la opinión cos-mocéntrica de autores como E. Morin, R. Ardrey o E. Chaisso, entre otros muchos.

249
© Universidad de Deusto - ISBN 978-84-9830-383-4
b) Razones a favor del antropocentrismo
Lo curioso es que precisamente los mismos argumentos que el cosmo-centrismo presenta para apoyar su postura, se pueden utilizar en su contra, sirviendo para apoyar las tesis antropocéntricas. Ruiz de la Peña presenta dos tipos de razones para apoyar las tesis del antropocentrismo: subjetivas y objetivas.
Las razones subjetivas se apoyan en el hecho de que el ser huma-no ha ido construyendo a lo largo de su historia la ilusión de que somos el centro del universo. Curiosamente un autor como Robert Ardrey, de-fensor de las tesis cosmocéntricas, utiliza este mismo argumento: «La ilusión … de creernos en una posición central nos acucia al saltar de la cama por la mañana, afeitarnos o hacernos el maquillaje y continuar con la labor diaria. Sin ella, moriríamos como individuos, nos extinguiríamos como especie»156.
En similares términos se expresa E. Jüngel, al considerar que el ser humano, tras perder la centralidad objetiva del universo tras los grandes avances científicos del inicio de la modernidad, volvió a convertirse de modo subjetivo en el centro del universo a partir de la filosofía cartesiana, pensamiento en el que «venía dada la orientación antropológica de todos los problemas de la edad moderna»157.
Pero, frente a estos planteamientos subjetivos, son más importantes los datos objetivos consistentes en advertir la suprema improbabilidad de la emergencia de la vida, y, sobre todo, de la vida humana, en la medida en que, como defienden todos los biólogos, todas las formas vivas proce-den de un único inicio, convirtiendo ese acontecimiento en un caso único, de un índice de improbabilidad altísimo, casi irrepetible158. Y lo mismo pasa con la aparición del género humano. De tal modo que, si visto desde el punto de vista cosmocéntrico el ser humano se nos presenta como algo sin importancia aparecido por azar en un planeta suburbial, esa misma im-probabilidad lleva al planteamiento antropocéntrico a considerar al ser hu-mano como un algo único y excepcional, cuya aparición y conformación suscita la admiración y las más evidentes preguntas sobre su sentido y lu-gar en el conjunto del universo.
156 Ardrey, R., La evolución del hombre: la hipótesis del cazador, Madrid, Alianza, 1983 (3.ª ed.), 228. Tomo la cita de Ruiz de la Peña, J. L., o.c., p. 243. Las cursivas son de J. L. Ruiz de la Peña.
157 Jüngel, E., Dios como misterio del mundo, Salamanca, Sígueme, 1984, p. 34. Cita tomada de J. L. Ruiz de la Peña, o.c., p. 243.
158 Cfr. Luisi, Pier Luigi, La vida emergente. De los orígenes químicos a la biología sintética, Barcelona, Tusquets, 2010, cap. 1.º.

250
© Universidad de Deusto - ISBN 978-84-9830-383-4
De ahí que un científico de la talla de J. Monod, situado en las tesis cosmocéntricas, no dude en defender la tesis de que «el acontecimiento decisivo (la aparición de la vida) no se haya producido más que una sola vez. Lo que significa que su probabilidad a priori es casi nula… La bios-fera aparece como producto de un acontecimiento único… Nuestro nú-mero salió en el juego de Montecarlo»159. De forma similar se expresan Popper, Dobzhansky, W. Thorpe y E. Morin160. Esto no se contradice con la opinión científica de que, dada la acumulación de circunstancias mate-riales en un momento determinado y en el lugar adecuado, la aparición de la pre-vida fuera inevitable «en las condiciones físico-químicas de la tie-rra prometida», como indican J. Ruffié161 e I. Prigogine162. El problema de fondo se centra en saber cómo, por qué y con qué probabilidad pudieron darse tales condiciones que posibilitaron la aparición de la vida y del gé-nero humano. Lo único que se puede hacer aquí es referirse a considera-ciones estadísticas, pero nada más. Y teniendo en cuenta que existen unos diez mil millones de planetas semejantes al nuestro, no se puede incluso descartar por completo la posibilidad de que en algunos de ellos se repi-tiera el improbable acontecimiento del surgimiento de seres vivos. Es lo que algunos han denominado la teoría de la panspermia163.
4.4.2. Entre el principio cosmológico o copernicano y el principio antrópico
Cada una de las dos caras de esta cuestión, esto es, cada uno de los dos modos de situarse ante el interrogante sobre la centralidad o no del ser humano en el universo, se apoya en dos principios alternativos: el cosmo-lógico o copernicano y el antrópico. Vamos a examinarlos por separado.
a) El principio cosmológico o copernicano de la tesis cosmocéntrica164
El origen de este principio se remonta a Copérnico (1543), cuando empezó a defender que la Tierra no era el centro del universo, sino un planeta más. En la época más reciente, la configuración de este principio
159 Monod, J., El azar y la necesidad, Barcelona, Seix Barral, 1971 (·3.ª ed.), 158-160. 160 Cfr. Ruiz de la Peña, J. L., o.c., p. 244. 161 Ruffié, J., De la biología a la cultura, o.c. 162 Cfr. Prigogine, I.-Stengers, I., La nueva alianza. Metamorfosis de la ciencia, Ma-
drid, Alianza, 1983, pp. 185 y ss. 163 Cfr. Luisi, P. L., La vida emergente, o.c., pp. 38 y ss. 164 Cfr. Gale, George, «El principio antrópico», Investigación y Ciencia, 1982, n.º 65,
pp. 93-103.

251
© Universidad de Deusto - ISBN 978-84-9830-383-4
se debería a Hermann Bondi, de la Universidad de Cambridge, quien en 1948 defendía que la posición de los observadores humanos en el univer-so no es en modo alguno privilegiada o distinta de otras posiciones. Así, las observaciones cosmológicas son válidas no sólo para la Tierra o el sistema solar, sino para las más remotas regiones del Universo. Con esta afirmación no se hace más que ser consecuente con el principio de la teo-ría de la relatividad de A. Einstein.
De este modo, este principio cosmológico señala que no sólo es la po-sición del sistema solar la que no tiene estatuto alguno de privilegio, sino que tampoco la tiene otra posición en cualquier otra parte del universo. Esto implica que la estructura del universo es uniforme, esto es, todas las regiones del universo son exactamente iguales de cara a la observación del resto del universo. Y, de hecho, repetidos experimentos científicos confirman esta apreciación. Ahora bien, esto tampoco es demasiado con-vincente, porque las distancias en la Tierra son insuficientes para demos-trar que el espacio es homogéneo, y en consecuencia los fenómenos cós-micos funcionan siempre igual.
Por eso, H. Bondi y Thomas Gold, de la Universidad de Cornell, pro-pusieron una hipótesis aún más general, denominada principio cosmoló-gico perfecto, que dice así: el universo es uniforme en el espacio y en el tiempo. Este principio es la base del modelo cosmológico estacionario, que defiende que el universo es completamente uniforme en el espacio y en el tiempo. Pero, al hacerse evidente, años más tarde, que el universo se halla en permanente expansión, este modelo se reformula con la hipótesis de que continuamente se está creando materia.
Ya hace tiempo que se abandonó el modelo del estado-estacionario, y, con él, el principio cosmológico perfecto. La causa de ese abandono se debió al descubrimiento, en 1965, de la denominada radiación de fon-do de microondas, por unos ingenieros de la empresa Bell165. Esta radia-ción de fondo sería el remanente de un estadio del universo primitivo, en un momento en el que el universo era más cálido y denso que en la actua-lidad. Este fenómeno radiactivo excluye la uniformidad temporal del uni-verso, pero proporciona la prueba más fuerte sobre la uniformidad espa-cial a gran escala. Por tanto, suponía una prueba contundente a favor del principio cosmológico, pero no en su versión perfecta.
De todos modos, estos datos pueden conjugarse con el principio an-trópico. Todo apunta y avala la tesis del inicio del universo con el Big Bang, desde un momento de concentración máxima en el que el universo
165 Cfr. Davies, Paul, Otros mundos, Barcelona, Salvat, 1986, cap. VIII, «El principio antrópico».

252
© Universidad de Deusto - ISBN 978-84-9830-383-4
no posee dimensiones y contiene una densidad aparentemente infinita. Y desde ahí se puede calcular la edad y el momento en el que se produjo esa explosión, que los científicos, como ya se dijo, calculan entre 13.000 y 20.000 millones de años. Desde esta explosión inicial, se van originando las diversas galaxias y su progresivo alejamiento con una velocidad con-formada por la constante propuesta por Edwin P. Hubble, quien en 1923 descubrió la relación entre la distancia y la velocidad de recesión.
Dependiendo de que la fuerza y la velocidad con la que las galaxias se van expandiendo, sea una u otra, se puede también decidir o considerar si el universo es abierto o cerrado. Es decir, si la velocidad traspasa deter-minado límite, el universo seguirá expandiéndose sin ningún tipo de lími-te ni vuelta atrás. Y en el caso de ser abierto, de su velocidad se calcularía que la edad de la explosión inicial se produjo hace unos 20.000 millones de años. En cambio, si se defiende la tesis de un universo cerrado, se con-sidera que el universo cesará en algún momento de expandirse y empeza-rá a contraerse, orientándose hacia el colapso final o gran implosión (BigCrunch). Las cifras que se manejan en este caso para el momento del Big Bang son unos 13.000 millones de años. Entre ambas hipótesis, pa-rece que la apoyada mayoritariamente por los científicos actuales es la de un universo cerrado.
b) El principio antrópico166
Todo lo dicho hasta ahora puede subsumirse en una formulación más ambiciosa, y dar un paso más, que se expresa de la siguiente forma: tal como ha evolucionado, el universo es tan improbable, que parece calcula-
166 Cfr. Gale, George, «El principio antrópico», o.c., pp. 94-103; Demaret, J./Barbier, CH., «Le principe anthropique en cosmologie», Révue des questions scientiphiques, 15(1981), 181-222 y 461-509; Morren, L., «De la loi d’entropie au principe anthropique», Révue Théologique de Louvain, 1984, pp. 160-183; Alonso, J. M., Introducción al principio antrópico, Madrid, Edic. Encuentro, 1989; Wilson, P. A., «What is the Explanandum in the Anthropic Principle», Am.Philos. Quarterly, 28 (1991), n.º 3, 167-174; Lerlie, John, «Time and the Anthropic Principle», Mind, 101(1992), n.º 403, 521-540; Bradner Norris, R., «Cosmos and Anthropos: A Philosophical Intepretation of the Anthropic Cosmological Principle, and Cosmos and Theos: Ethical and Teological Implications of the Anthropic Cosmological Principle, by Errol E. Harris», Zygon, 28 (1993), n.º 4, 531-536; Pérez de Laborda, Al, «El principio antrópico», Diálogo Filosófico, 7 (1991), n.º 1, 46-57; Id., Laciencia contemporánea y sus implicaciones filosóficas, Madrid, Cincel, 1985, cap. 7.º; Davies, Paul, Dios y la nueva física, o.c., cap. 13.º; Id., Otros mundos, o.c., cap. VIII; Ruiz de la Peña, J. L., o.c., pp. 245 y ss.; Mosterion, J., «Examen del principio antrópi-co en cosmología», Diálogos, 79 (2002), n.º 79, 203-236; Luisi, P. L., La vida emergente,o.c., pp. 34 y ss.

253
© Universidad de Deusto - ISBN 978-84-9830-383-4
do a propósito para que sea factible la aparición de un observador del mis-mo, el ser humano.
El número de universos posibles es casi infinito; que se haya reali-zado precisamente el modelo en el que se cumplen todas las condiciones (sumamente restrictivas) necesarias para el surgimiento de lo humano, hace pensar en un diseño previo. Este modo de pensar se ha denomina-do principio antrópico, esto es, la hipótesis de que el origen y la evolu-ción de todo el universo estaría orientado en función del surgimiento del hombre, tal y como de hecho ha surgido y es. El primero que lo formula de forma sistemática fue Carter, en 1974167, y posteriormente fue clave la obra de J. D. Barrow y F. J. Tipler, en 1986168.
Pero hay dos versiones muy diferentes de este principio: la denomina-da versión débil, y la fuerte; y, dentro de la versión fuerte, veremos tam-bién la denominada versión fuerte participativa.
1. La versión débil del principio antrópico (Weak Anthtopic Principle: W.A.P.)
Esta versión no hace más que constatar que el hecho de que en el uni-verso se halle incluido el hombre, hace evidente que se han tenido que dar las condiciones necesarias para la evolución de la vida y de la inteligen-cia. Es, como se ve, una visión del mundo desde una racionalidad retros-pectiva, a posteriori, en la que todo el universo converge en el hombre, el último ser vivo de la cadena evolutiva.
Por tanto, se puede adoptar el finalismo antrópico como perspectiva heurística o metodológica. Pero se ha de ser consciente de que la teleolo-gía del principio antrópico, en su versión débil, se tiene que quedar en el estricto plano metodológico, o, como máximo, en el epistemológico. Es decir, es una especie de teleología de como si, que constituye un puente entre el ámbito epistemológico y el ontológico.
Pero en esta versión débil, cabe preguntarse también si se da en el universo azar o finalidad. Y a la hora de contestar, nos encontramos con varias opciones.
— La primera respuesta considera que la orientación del universo ha-cia el hombre es obra de una inteligencia inmanente al universo (como si éste supiese de antemano que iba a dar a luz al hombre).
167 Cfr. Alonso, J. M., Introducción al principio antrópico, o.c., pp. 43 y ss.168 Cfr. Barrow, J. D./Tipler, F. J., The Anthropic Cosmological Principle, Oxford,
Clarendon Press, 1986.

254
© Universidad de Deusto - ISBN 978-84-9830-383-4
Es la versión que podemos denominar animista o panteísta, defen-dida por F. Hoyle169.
— La segunda opción consistiría en defender que la historia del uni-verso es fruto de una inteligencia trascendente, que dirigiría desde fuera al universo.
— Y la tercera considera que el universo está empujado simplemente por el azar, la pura casualidad. Ahora bien, ya vimos en su momen-to que esta solución no resuelve ningún problema, puesto que nos remite a un interrogante, esto es, a la simple constatación de nues-tra ignorancia. Este remitirnos sin más al azar es cuestionado en la actualidad por reputados científicos y filósofos170.
En resumen, nos encontramos con tres hipótesis: inteligencia inma-nente, inteligencia trascendente, y el azar.
2. Versión fuerte (Strong Anthropic Principle: S.A.P)
Esta versión es la que resulta más interesante y problemática, sea o no convincente, y es la que plantea una cuestión decisiva. La versión fuer-te no se limita a constatar que el universo, de hecho y a posteriori, ha desembocado en el hombre, sino que el universo debe (en un sentido fuer-te, no sólo dado-que-el-hombre-existe, sino en sentido absoluto) tener las propiedades necesarias para que sea posible el surgimiento en él de vida inteligente171.
¿Cuál es la justificación de la visión fuerte del principio antrópico? Los que la defienden se apoyan en la mecánica cuántica y en su aplica-ción al origen y evolución del universo172. Como ya sabemos, el principio de indeterminación de W. Heisenberg y el de ambigüedad de N. Bohr, nos hacen ver que en el nivel infra-atómico impera la probabilidad, no el de-terminismo, y la presencia de un observador supone introducir cambios y perturbaciones en lo observado.
Si reflexionamos sobre el origen del universo, y aceptamos como teo-ría más válida y aceptada por los científicos la del Big Bang, hay que re-
169 Cfr. Hoyle, F., The Nature of the Universe, Oxford, Basil Blackwel, 1950; Id., Eluniverso inteligente. Un nuevo enfoque del origen y la evolución de la vida, Barcelona, Grijalbo, 1984.
170 Cfr. Wagensberg, J. (dir.), Proceso al azar, o.c.171 Una versión ponderada de esta versión, puede verse en Gale, G., «A revised Design:
Teleology and Big Questions in Contemporary Cosmology», en Biology and Philosophy, 2 (1987), 475-492 (trad. esp. en Investigación y Ciencia, 1982, n.º 65, ya citado).
172 Cfr. Davies, P., Dios y la nueva física, o.c., cap. 8.º, «El factor cuántico»; Id., Otros mundos, o.c., cap. 8.º, «El principio antrópico».

255
© Universidad de Deusto - ISBN 978-84-9830-383-4
conocer que el universo tuvo un comienzo (aunque hoy día va cobrando fuerza la teoría de que el Big Bang fuera al mismo tiempo el comienzo de un ciclo de la historia del universo, y el fin de otro ciclo anterior). Pero el problema está en que hablar de tiempo cuando se habla del origen del universo posee una dificultad, en la medida en que el tiempo es una cua-lidad consustancial y correlativa a la existencia del mundo. Sin mundo no hay tiempo, por lo que no hay tiempo antes del mundo. De ahí que en las cuestiones sobre cómo fue creado el mundo, o quién lo ha causado, se tie-ne que distinguir entre el nivel científico y el ontológico. De modo que cabe, en el nivel ontológico y religioso, hablar de creación del mundo por Dios, aceptando la infinitud temporal del mundo173. Ello se debe a que cuando se habla de la creación del mundo por Dios, se hace referencia a la dependencia ontológica del mundo respecto a Dios para existir. Dios es el fundamento último de la realidad, independientemente de cuándo haya comenzado a existir.
En el ámbito de la filosofía y de la ciencia, los conceptos de causa y efecto son conceptos temporales, y no pueden aplicarse a un estado en el que el tiempo no existe. La única solución consiste en ampliar el concepto de causa. En primer lugar, reduciendo la necesidad de que la causa prece-da siempre al efecto. La cuestión está en ver si es posible que una causa actúe hacia atrás en el tiempo para producir un efecto anterior a ella. La teoría cuántica parece que así lo confirma, habiendo casos en los que se puede producir causalidad retroactiva, fruto de unas partículas hipotéticas más rápidas que la luz, llamadas taquiones.
3. El principio antrópico participativo (P.A.P.) de J. Wheeler
Esta es la hipótesis que defiende el físico John Wheeler, con su princi-pio antrópico participativo (P.A.P.)174. Para Wheeler, el principio cuántico demuestra que, en algún sentido, lo que haga un observador en el futuro determina lo que pasó en el pasado, incluso en un pasado tan remoto que la vida no existía. Wheeler introduce la mente (el observador) de una ma-nera fundamental, como estamos obligados a hacerlo en la teoría cuántica, de tal modo que para él los observadores son necesarios para que exista el universo, de tal modo que, para Wheeler, «el observador es tan esen-cial para la creación del universo, como el universo para la creación del
173 Cfr. Küng, Hans, El principio de todas las cosas. Ciencia y religión, Madrid, Trotta, 2007; Pineda, J. A. de la, El baile de la ciencia y la metafísica, Madrid, Biblioteca Nueva, 2008.
174 Sobre las tesis de Wheeler, cfr. Alonso, J. M., Introducción al principio antrópico,o.c., p. 555 y ss.

256
© Universidad de Deusto - ISBN 978-84-9830-383-4
observador»175. De este modo, Wheeler relaciona la existencia de la men-te en un estado posterior de la evolución cósmica con la misma creación del universo. Estos planteamientos tienen cierta fuerza desde la pregun-ta de qué sentido tiene un universo sin una conciencia que lo contemple y lo entienda. Por tanto, ¿tiene que ser el propio mecanismo de la existencia del universo algo sin sentido o inviable, o ambas cosas, a menos que el universo tenga garantía de producir vida, conciencia y observación en al-guna parte y durante algún breve instante de su historia futura?
Wheeler confía en que podamos descubrir en la física un principio que permita que el universo surja por su propio impulso. Wheeler ha com-parado este universo auto-causado con un circuito auto-excitado en elec-trónica. Pero estos planteamientos, aunque resulten muy sugerentes, no resultan muy convincentes, y plantean muchos interrogantes. Paul Davies observa176 que, aunque fuera posible encontrar la causa de la creación del espacio-tiempo en alguna actividad natural posterior (sea mente o ma-teria), es difícil comprender cómo la creación a partir de la nada puede ocurrir de un modo natural. Deberíamos todavía explicar el origen de la materia prima de la mente, o lo que actúa retroactivamente.
Estas observaciones de la teoría cuántica nos empujan a comprender nuestra realidad y la del universo de modo diferente a como lo perciben nuestros sentidos y la lógica natural a que estamos acostumbrados. Pero eso ya ha pasado otras veces en la historia, y nos pasa también en di-versas percepciones sensoriales diarias, como la errónea impresión que tenemos diariamente de que el sol es el que se mueve sobre nosotros mientras la tierra está quieta; o la impresión que tenemos al estar sobre un puente fluvial y mirar a la corriente de agua que tenemos debajo: pa-rece que es el puente el que se mueve y avanza, mientras que el agua es-taría quieta.
En relación a las tesis de Wheeler sobre el principio antrópico par-ticipativo, resultan interesantes, a la vez que problemáticas, los plantea-mientos del físico Hugh Everett, de la Universidad de Princeton, sobre los universos paralelos (1957), tesis desarrollada más adelante por Pryce De Uit, de la Universidad de Austin (Texas). La mecánica cuántica afirma que todos los mundos cuánticos alternativos son posibles, en la medida en que no podemos saber la posición de un electrón, pudiendo estar en diver-sos sitios a la vez. De tal forma que esas localizaciones alternativas serían igualmente reales, pudiendo existir paralelamente múltiples mundos al-ternativos. Todos esos mundos son iguales, y todos tendrían observadores
175 Cita tomada de Alonso, J. M., o.c., 59-60.176 Cfr. Davies, P., Dios y la nueva física, o.c., p. 48. Puede verse en la p. 130 el dibujo
con el que Wheeler imagina ese mundo autoeficiente.

257
© Universidad de Deusto - ISBN 978-84-9830-383-4
humanos. Sin embargo, cada conjunto de habitantes percibe únicamente su propia rama del universo.
Esos mundos paralelos, una vez desconectados, se encuentran físi-camente aislados para todo propósito práctico. De tal modo que sólo in-virtiendo el curso del tiempo se podrían volver a unir. La distancia entre un mundo y otro no se mide en unidades espaciales, porque nos hallamos ante otro concepto de distancia. Y tampoco se puede vivir en todos esos mundos de igual manera, porque en otros mundos es posible que no haya las condiciones necesarias para la vida humana, o para mi vida concreta. Los defensores de esta teoría nos advierten de que en alguno de esos mun-dos alternativos puede que no haya existido Hitler, o no hayan asesinado a Kennedy, o Judas no traicionó a Jesús, ni César atravesó el Rubicón, etc.
La cuestión que queda pendiente en estos planteamientos consiste en responder al interrogante de por qué yo (y los demás) estamos en este mundo concreto en el que estamos, y no en otro de los alternativos. Desde el punto de vista físico, nuestra existencia es posible porque, entre muchas otras posibilidades, nuestro mundo eligió esta estructuración muy orde-nada de la materia y la energía que mantiene el cosmos lo bastante frío como para que pueda haber vida. Es un mundo con un contenido de calor primigenio próximo al mínimo (unos tres grados).
Sólo en nuestro mundo existen criaturas inteligentes que se plantean preguntas sobre la cosmología y el sentido de la existencia. El resto re-corre sus historias entre tormentas rugientes y calores tórridos que nadie percibe: son, pues, estériles, violentos, y, en apariencia, sin sentido177.Ahora bien, sin recurrir a la posibilidad de mundos alternativos, y dentro de nuestro mundo, puede haber planetas similares al nuestro, con las mis-mas condiciones alternativas ambientales como para producir vida, y vida inteligente como la nuestra. Pero no lo sabemos; nadie ha conectado con nosotros, y no sabemos si será vida como la nuestra, o diferente, y, por tanto, si podríamos interactuar de modo inteligente con ella.
De lo que sí parece que la ciencia física está cada vez más segura es de que el universo en el que estamos es un universo privilegiadísimo, puesto que, de todas las alternativas de mundos que ha hecho posible la historia de nuestro mundo, el actual es el único apropiado para la vida178.Esto le lleva a Wheeler a concluir, apoyado en la mecánica cuántica, que el observador inteligente contribuye a que la realidad es como es por el acto mismo de la observación. Y de ahí concluye también, llevando el
177 Cfr. Davies, P., Otros mundos, o.c., pp. 157-158. 178 Aunque no todos los científicos opinan en la actualidad de este modo: Cfr. Jenkins, A./
Pérez, G., Investigación y Ciencia, 2010, marzo, n.º 402, pp. 12-19; Sampedro, Javier, «Hay otros mundos posibles», El País, 17 de enero de 2010, pp. 36-37.

258
© Universidad de Deusto - ISBN 978-84-9830-383-4
razonamiento al extremo, que, para que el universo sea real, ha de evo-lucionar de tal modo que en él puedan llegar a existir observadores inte-ligentes.
La justificación que utiliza es clara: «Jamás se ha dado razón algu-na de por qué ciertas constantes y condiciones iniciales tienen los valores que tienen, excepto la de que, de no ser así, algo como nuestra capaci-dad de observadores, según la conocemos, sería imposible»179. Por tanto, no acepta Wheeler que la vida y la capacidad de observación sean meros accidentes en un universo independiente de los observadores. Por eso, dice: «La mecánica cuántica nos ha obligado a tomar en serio y a sope-sar la opinión, directamente opuesta, de que tan esencial es el observa-dor para la creación del universo como el universo para la creación del observador»180.
Ahora bien, haciendo una valoración conjunta de las posiciones de Wheeler, tenemos que reconocer que, como indica G. Gale, «ha lleva-do Wheeler el principio antrópico más allá de los dominios de la lógica de la explicación: ha cruzado el umbral de la metafísica. Pocos científicos o filósofos de la ciencia se sentirían cómodos con su visión»181. Pero lo que sí es cierto es que científicos de la talla de Hawking, Collins y otros, más propicios a defender la posición del principio cosmológico, están dando entrada en sus reflexiones, si no defendiendo, cada vez más las tesis del principio antrópico, en su versión débil182.
c) Valoración global de los dos principios
A la hora de hacer una valoración conjunta y en profundidad de las dos propuestas sobre el sentido de la historia del universo, es fundamental que volvamos de nuevo a situar las diversas afirmaciones y teorías en sus correspondientes niveles epistemológicos, el científico, el filosófico y el teológico-religioso.
La ciencia tiene que limitarse al nivel de lo fáctico, a constatar los he-chos y las leyes con las que está constituido nuestro mundo. Pero no le corresponde, como tal saber científico, entrometerse en cuestiones inter-
179 Cita tomada de Gale, George, o.c., p. 103. 180 Cfr. ibídem, p. 103. 181 Ibídem, p. 103. 182 Cfr. Hawking, S., La historia del tiempo. Del Big Bang a los agujeros negros, Bar-
celona, Crítica, 1988; Id., El universo en una cáscara de nuez, Barcelona, o.c.; Id., La teo-ría del todo. El origen y el destino del universo, Barcelona, Mondadori, 2007; Coles, Peter, Hawking y la mente de Dios, Barcelona, Gedisa, 2004; Pineda, J. A. de la, El baile de la ciencia y de la metafísica, o.c.,

259
© Universidad de Deusto - ISBN 978-84-9830-383-4
pretativas o de sentido, campo específico de la filosofía y la teología. De ahí que apostar por el principio cosmológico o antrópico no es tarea de la ciencia sino de la filosofía. La ciencia aporta los datos correspondientes, y sobre ellos el filósofo (o el científico en cuanto filosofa) advierte una mera concatenación de causas y efectos en los que puede interpretar la centralidad del ser humano (principio antrópico) o no (principio cosmoló-gico o copernicano).
Ahora bien, dentro del paradigma del principio antrópico, es muy dis-tinto el estatus de la versión débil que el de las versiones fuertes. En la versión débil, no se hace más que constatar que, dentro de las diversas alternativas que podría haber seguido la historia del universo, la que de hecho ha seguido es la única, o casi única, entre las muchas alternativas posibles, que ha permitido la existencia de la vida, y del ser humano, en este pequeño planeta de nuestro sistema solar. Esta sola constatación es susceptible por sí sola, como ya lo dejamos constatado, como para susten-tar cualquiera de las dos alternativas filosóficas y teológicas. Unos dedu-cen de ella la pura probabilidad del azar para interpretar la aparición de la vida y del ser humano (principio cosmológico), y otros leen en esa rareza y mínima probabilidad de la vida la presencia de una finalidad humanís-tica (principio antrópico), sustentada incluso por un fundamento del uni-verso, que apunta a la realidad de Dios. Pero dentro sólo de las diferentes versiones del principio antrópico, la débil se sitúa en la mera constatación de hechos, mientras que la fuerte sobrepasa lo que son los planteamientos científicos para situarse en un nivel metafísico, que puede ser aceptable y respetable, pero que ya no es el de la ciencia, nivel en el que parece evi-dente quieren situarse los que lo defienden. Por eso, los planteamientos de Wheeler y sus seguidores, pertenecen más a una propuesta metafísica, y religiosa, que a una tesis científica, por más esfuerzos que hagan por fun-darla en algunos elementos poco demostrados de la mecánica cuántica.
4.4.3. Entre el azar y la finalidad: una cuestión de opción filosófica
La cuestión acerca de si la historia del universo y del origen de la vida, así como su posterior evolución, como cualquier cuestión sobre nuestra realidad mundana, posee, por tanto, dos niveles de comprensión: el científico y el filosófico. Ya hemos indicado al inicio del capítulo que la ciencia es quien se ocupa de la dimensión fáctica, y, por tanto, es la encar-gada de descubrir cómo está constituida y cómo se comporta la realidad en el ámbito de las interacciones causales de sus diferentes elementos ma-teriales. Pero ante esas leyes que configuran lo real, sean determinísticas o probabilísticas, se nos plantea siempre la pregunta por el sentido profundo

260
© Universidad de Deusto - ISBN 978-84-9830-383-4
y último del universo, concretado en cuestiones como ¿por qué la realidad es así y no de otra forma?; o más radical, ¿por qué hay realidad en vez de nada?
Este tipo de cuestiones, que pertenecen a la mirada filosófica, son las que nos hacen ver que la pregunta acerca de si el mundo es resultado del azar o de una finalidad programada, no se resuelven desde el nivel cientí-fico, sino desde una propuesta filosófica, que siempre será una más entre otras, porque nunca podrá demostrar de forma concluyente la verdad inequí-voca de su propuesta. Por eso, la opción por la condición azarosa de lo que hay es fruto de una cosmovisión que apuesta por una visión autosu-fuciente del mundo, construido a fuerza de golpes azarosos que han ido concatenando las diferentes fuerzas que lo componen, sin necesidad de echar mano de una realidad fundante y trascendente que la haya traído a ser y la mantenga en su existir. El problema está en que la experiencia que el mundo nos transmite es su radical contingencia, su insuficiencia para existir y mantenerse en su ser.
Para una opción creyente, la existencia del universo es consecuencia del acto creador de una realidad trascendente que llamamos Dios, como fundamento último, posibilitante e impelente183. La realidad de Dios es compatible con un mundo dominado por leyes determinísticas (salvando la libertad del hombre) o azarosas y probabilísticas, puesto que el ámbito de la probabilidad en el nivel de las causas científicas, no evita que Dios actúe a través de las leyes probabiíisticas que la mecánica cuántica ad-vierte en el mundo de las partículas elementales de la materia en el nivel de lo infra-atómico184. El creyente tiene la legitimidad para creer en la in-fluencia de un Dios providente bajo las leyes empíricas de lo mundano, incluso ante un mundo dominado por las leyes probabilísticas, tanto en el mundo infra-atómico como en el sucederse de las mutaciones genéticas que permiten el surgimiento de nuevas especies, hasta llegar a las que han ido componiendo el género humano. La acción providente de Dios es su-ficientemente sabia y poderosa como para orientar, en diálogo amoroso, el proceso cosmológico y evolutivo, en un acto de autoabajamiento, confor-mado por un diálogo entre él y el cosmos, así como entre él y el ser huma-no en su libertad185.
183 Cfr. Zubiri, X., El hombre y Dios, o.c., 184 Cfr. Schmitz-Moormann, Karl (en colaboración con James F. Salmon, S. J.), Teolo-
gía de la creación de un mundo en evolución, Estella, Verbo Divino, 2005; Haught, J. F., Cristianismo y ciencia. Hacia una teología de la naturaleza, Santander, Sal Terrae, 2009; Edwards, Denis, El Dios de la evolución. Una teología unitaria, Santander, Sal Terrae, 2006.
185 Cfr. Edwards, Denis, o.c.; Barbour, I., El encuentro entre ciencia y religión, Santan-der, Sal Terrae, 2004.

261
© Universidad de Deusto - ISBN 978-84-9830-383-4
Por tanto, tan legítima y razonable (desde la mera razón humana) es la postura que no ve más que el ciego azar como respuesta última al sen-tido del universo, como la que confía en una profunda orientación inte-ligente y amorosa por parte de un Dios providente, que acompaña y dia-loga con el universo, renunciando a imponer su poder a través de leyes fijas y cerradas, así como dejando la historia humana en manos de las decisiones libres de los humanos, tanto cuando hace el bien como cuando hace el mal, destruye a sus semejantes y comete actos depredadores so-bre la naturaleza, exponiéndose a una destrucción de la especie humana y de toda la biosfera. Las dos opciones no pueden ser verdaderas al mismo tiempo, pero en nuestra situación histórica no tenemos posibilidades de demostrar la verdad de ninguna de las dos, aunque a cada uno de los de-fensores de ambas opciones les parecerá más coherente y lógica su pro-pia postura.


263
© Universidad de Deusto - ISBN 978-84-9830-383-4
Capítulo 5
El proceso de humanización. La dimensión conductual
1. LA HUMANIZACIÓN COMO COMPLEMENTO DE LA HOMINIZACIÓN
Si volvemos al cuadro presentado al inicio del capítulo anterior, se pude ver que nos referíamos a la interrelación entre tres tipos de factores: las dos fronteras de la hominización, los rasgos morfológicos que expe-rimentan los humanos en el proceso de llegar a ser una nueva especie, y un tercer conjunto de factores: los cambios culturales y conductuales que van también experimentando. Una vez presentados los dos primeros apar-tados, iniciamos ahora el tercero. Podríamos subsumir los dos primeros conjuntos de factores bajo el proceso denominado de hominización, mien-tras que el conjunto de características culturales que van desarrollando los seres humanos los denominamos el proceso de humanización. Ambos es-tán mutuamente imbricados e interrelacionados, puesto que si la especie humana llega, con el paso de las dos fronteras y los cambios morfológicos subsiguientes, a constituirse como especie humana (proceso de hominiza-ción), a través de una serie de factores y de rasgos en las que ella no inter-viene, en el proceso de humanización, por el contrario, es ya el propio ser humano el que progresivamente va tomando las riendas de su propia rea-lización.
Ahora bien, no se trata de dos niveles separados y extrínsecos, sino que están mutuamente implicados e imbricados. Esto es, el proceso de hu-manización no se inicia cuando ha terminado el de hominización, sino que empieza poco después, o casi al mismo tiempo de haberse iniciado éste.

264
© Universidad de Deusto - ISBN 978-84-9830-383-4
Al menos, en la medida en que el proceso de cerebralización ha alcanza-do el nivel suficiente como para que los primeros seres humanos (homohabilis) tuvieran el mínimo de inteligencia, con los elementos específicos de la condición humana: conciencia, intencionalidad, libertad, previsión de futuro, etc. Desde ese momento, el ser humano se convierte, en parte, en sujeto de su propia realización o antropogénesis, interviniendo también al mismo tiempo en el desarrollo y transformación de los rasgos morfológi-cos que le van haciendo crecer y avanzar desde el homo habilis hasta el homo sapiens sapiens.
Podemos, pues, decir que el proceso evolutivo nos convirtió en homí-nidos (humanos), sin que el propio ser humano interviniera en ello. Como indican los filósofos existencialistas, hemos sido arrojados a la existen-cia, pero una vez situados en ella, advertimos que nuestra naturaleza, nuestro modo de ser, no está totalmente programado ni dejado al cuidado exclusivo de las leyes biológicas internas, ni al entorno ecológico externo, sino que el ser humano va tomando las riendas de su destino, orientando su existencia al mismo tiempo que va terminando de completar su propia imagen morfológica. Somos, pues, homínidos humanizados1, homínidos a quienes se les ha encargado la tarea de hacerse, de acabar de hacerse.
El conjunto de rasgos culturales o conductuales que van emergiendo con la aparición del ser humano es muy variado y complejo. Carbonell y Sala los resumen en los siguientes: «la pinza de precisión, la fabricación y la utilización de instrumentos, la caza en grupo, la construcción de refu-gios, la capacidad de transportar la comida y las materias primas a través de grandes distancias, la producción y uso del fuego, el desarrollo del len-guaje articulado, la práctica de enterrar a los muertos, el vestido, el arte, la domesticación sistemática de animales, el descubrimiento y la fabricación de los metales, la escritura, entre otras de menor entidad»2. No vamos a ocuparnos de todos estos aspectos, sino que nos vamos a detener sólo en los más significativos. Y lo haremos de un modo esquemático, puesto que extenderse sobre cada uno de ellos nos ocuparía varios volúmenes.
Los rasgos conductuales y culturales de los que nos vamos a ocupar, como más específicos de nuestra especie, y cuya puesta en marcha le ha ido convirtiendo en humano y humanizándolo progresivamente, son las siguientes:
— La conciencia refleja autoconciencia.— La imaginación y la capacidad simbolizadora.
1 Cfr. Carbonell, Eudald/Sala, Robert, Planeta humano, Barcelona, Península, 2000, p. 18.
2 Ibídem, p. 17.

265
© Universidad de Deusto - ISBN 978-84-9830-383-4
— El origen y uso del lenguaje. — La creación y uso de herramientas y armas.— Dominio y uso del fuego.— Los usos alimenticios.— Construcción de refugios y casas.— Sexualidad, familia y primeros núcleos sociales.— La capacidad artística.— El culto a los muertos y el origen de la religión.
2. LOS PRINCIPALES RASGOS CULTURALES Y CONDUCTUALES
2.1. La conciencia refleja o autoconciencia
Parece evidente que una de las características más propias de lo hu-mano es la conciencia, o mejor, la autoconciencia. Ahora bien, a pesar de la evidencia de este rasgo humano, no resulta fácil señalar en qué consiste y en qué momento de la historia del linaje humano aparece. Son preguntas demasiado complejas sobre las que no podemos aportar pruebas objeti-vas, ni desde la paleoantropología ni desde la biología molecular. Se trata, por tanto, de un tema de gran complejidad, que a lo largo de la historia del pensamiento antropológico ha dado lugar a multitud de posturas acerca de qué entendemos por conciencia, subjetividad, alma o como queramos llamarlo, y qué solución dar al denominado problema de las relaciones alma-cuerpo, mente-cerebro o mente-cuerpo3. No es este el momento de
3 Sobre este problema, cfr. Liz, M., Perspectivas actuales en filosofía de la mente, Te-nerife, Gobierno de Canarias, 2001; Rabossi, E. (comp.), Filosofía de la mente y ciencia cognitiva, Barcelona, Paidós, 1995; Gardner, H., La nueva ciencia de la mente. Historia de la revolución cognitiva, Barcelona, Paidós, 2000 (2.ª ed.); Preist, S., Teorías y filoso-fías de la mente, Madrid, Cátedra, 1994; Bechtel, W., Filosofía de la mente, Madrid, Tec-nos, 1991; Martínez-Freire, P. E., La nueva filosofía de la mente, Barcelona, Gedisa, 1995; Broncano, F. (ed.), La mente humana, Madrid, Trotta, 1995; Dou, A. (ed.), Mente y cuer-po, Bilbao, Mensajero, 1986; Searle, J., Mente, cerebros y ciencia, Madrid, Cátedra, 1985; Id., El misterio de la conciencia, Barcelona, Paidós, 2000; Mora, Fco. (ed.), El problema cerebro-mente, Madrid, Alianza, 1995; Laín Entralgo, P., El cuerpo humano. Teoría actual,Madrid, Espasa-Calpe,1989; Id., Cuerpo y alma, Madrid, Espasa-Calpe, 1991; Ruiz de la Peña, J. L., Las nuevas antropologías, Santander, Sal Terrae, 1983. Bunge, M., El proble-ma mente-cuerpo, Madrid, Tecnos, 1985; Monserrat, J., Epistemología evolutiva y teoría de la ciencia, Madrid, Publicaciones Universidad de Comillas, 1987; Moya, C., Filoso-fía de la mente, Valencia, Publicaciones Universidad de Valencia, 2004; Hierro-Pescador, José, Filosofía de la mente y ciencia cognitiva, Madrid, Akal, 2005; Canobbio, Giacomo, Sobre el alma. Más allá de mente y cerebro, Salamanca, Sígueme, 2010.

266
© Universidad de Deusto - ISBN 978-84-9830-383-4
adentrarnos en ese complejo problema, teniendo que limitarnos aquí a di-lucidar sobre el momento en que surge en la especie humana la chispa de la conciencia, su proceso de crecimiento y consolidación, y si el hecho de la conciencia, o autoconciencia, es un elemento específico de la especie hu-mana o lo comparte también con algunas otras especies animales.
La primera dificultad la tenemos ya a la hora de designar el término a utilizar: conciencia, autoconciencia, alma, psique, mente, etc., puesto que cada uno de esos conceptos tiene de fondo unos presupuestos teóricos concretos y diferenciados. Como hemos indicado, no vamos a detenernos en el problema de las relaciones alma-cuerpo, o mente-cuerpo, sino que nos detendremos en qué podemos decir sobre la conciencia y la autocon-ciencia del ser humano, como elemento específico de su condición. Una segunda dificultad se halla en el hecho de que la conciencia no fosiliza, como puede hacerlo una parte del cuerpo humano, por lo que el modo que tienen los investigadores para acercarse a la pregunta sobre su origen y sus características es a través de caminos indirectos, estudiando sus frutos y consecuencias, así como las condiciones de posibilidad de la mente: las herramientas y otros objetos culturales, lenguaje, comportamientos, cere-bro, base genética, etc.
Lo que nos interesa aquí es estudiar el tema de la conciencia y de la autoconciencia desde el contexto del proceso evolutivo. Ahora bien, el acercamiento al esclarecimiento de este problema puede hacerse desde dos puntos de vista: el filogenético y el ontogenético. El más propio de este lugar es el primero, pero en la medida de su dificultad resulta in-teresante aprovecharse de la mirada ontogenética y extrapolarla, en la medida en que sea posible y legítimo, a la perspectiva filogenética. La perspectiva ontogenética se ayuda de las investigaciones de la Psicolo-gía evolutiva y del desarrollo4, para rastrear las diferentes fases de ma-duración de la capacidad sensitiva de un ser humano, tanto en las fases de su gestación intrauterina5 como durante los primeros años después del na-cimiento6. Aunque no podemos detenernos en recoger los muchos avan-ces realizados por estas disciplinas, es adecuado afirmar que el proceso de formación de la autoconciencia atraviesa múltiples etapas y consti-tuye un gradiente continuo en el que no parecen darse rupturas o saltos cualitativos. Por eso, resulta poco lógica la pregunta sobre el primer mo-
4 Cfr. Shaffer, D. R., Psicología del desarrollo. Infancia y adolescencia, Madrid, Inter-national Thomson Editores, 2005, 5.ª edición.
5 Cfr. Fernández Mateos, L. M., El vínculo afectivo con el niño por nacer, Salamanca, Universidad Pontificia de Salamanca, 2005.
6 Cfr. Spitz, R. A., No y Sí. Sobre la génesis de la comunicación humana, Buenos Aires, Ediciones Hormé, S.A. E./Paidós, 1972 (3.ª ed.).

267
© Universidad de Deusto - ISBN 978-84-9830-383-4
mento de autoconciencia en la vida de un niño, aunque si comparamos segmentos muy alejados en ese gradiente, podemos palpar la diferencia entre un recién nacido y un niño de siete años. Pero es inútil preguntar cuál es el momento en que se pasó de la inconsciencia a la consciencia, como si fuese un salto del todo a la nada. Se trata de una frontera difusa; aunque, por otro lado, sí que podemos afirmar que, desde otro punto de vista, se da una diferencia del todo a la nada entre la inconsciencia y la autoconciencia.
Aunque no es legítimo afirmar que se dé un total paralelismo entre el proceso ontogenético y el filogenético, sí que cabe considerar que esa misma dificultad para señalar el momento crítico del salto de la incons-ciencia a la autoconciencia se da dentro del enfoque filogenético. Con el agravante de que, en el aspecto ontogenético, la ciencia tiene campo para avanzar en sus estudios empíricos, mientras que no sucede lo mismo en el enfoque filogenético, en el que no cabe más que especular y hacer saltos teóricos de lo ontogenético a lo filogenético. De todos modos, parece ló-gico pensar que la conciencia ha emergido de modo paulatino en el proce-so de la antropogénesis porque representaba un beneficio y una cualidad positiva para sus poseedores. Es evidente también que quienes tienen con-ciencia de lo que hacen poseen más capacidad a la hora de enfrentarse con su entorno y con el resto de los seres vivos, tanto si son de su misma especie como si pertenecen a otra. Pero tenemos también que distinguir entre percibir inteligentemente el entorno natural y la capacidad de ser consciente de que estoy pensando y captando lo que sucede en mi entor-no. Sería la diferencia entre lo que algunos autores denominan percepcióny sensación7. Los seres humanos, en la medida en que estamos dotados de autoconciencia (sensación), tendemos a pensar que ambos aspectos son lo mismo, pero cabe imaginar separadas la percepción de la sensación, o mi-rada interior. De hecho, parece que ese es el caso de la mayoría, si no de todos los animales.
Por tanto, la pregunta adecuada es qué ventajas supuso la autocon-ciencia para la especie humana, puesto que el proceso evolutivo le dotó y le premió con esa cualidad. La verdad es que hay conductas complejas, por ejemplo la de ciertos animales y la de ciertos robots, que son capaces de desenvolverse realizando determinadas acciones sin tener concien-cia de lo que hacen. Los humanos tenemos conciencia, pero quizás cabría imaginar que podríamos vivir sin necesidad de ella. De ahí que algunos fi-lósofos hagan referencia a los zombis, criaturas hipotéticas semejantes en
7 Cfr. Humphrey, N., L., La mirada interior, Madrid, Alianza, 1993; Id., La reconquista de la conciencia. Desarrollo de la mente humana, México, FCE, 1987; Id., Una historia de la mente. La evolución y el nacimiento de la conciencia, Barcelona, Gedisa, 1995.

268
© Universidad de Deusto - ISBN 978-84-9830-383-4
todo a nosotros, pero sin conciencia8. La cuestión de fondo es si la con-ciencia vale para algo, o bien, por qué la evolución nos dotó de concien-cia, como elemento ventajoso. Por tanto, aunque en hipótesis podríamos pensar que la evolución habría podido generar zombis, eligió el camino de los humanos, seres autoconscientes en los que la conciencia es funda-mental y parte esencial de su condición.
Y esta conciencia no sólo se ocupa de los actos inteligentes y supe-riores, como el lenguaje y el pensamiento abstracto, sino también de las emociones y de los procesos sensoriales. De este modo, la conciencia se convierte en un elemento imprescindible para que los humanos podamos interactuar con el entorno natural y animal, así como, sobre todo, con los demás seres humanos. En el ejercicio de interactuar con el entorno ecoló-gico, la conciencia humana realiza funciones parecidas a la de los demás animales, pero lo más básico de nuestra conciencia es guiarnos en las re-laciones interhumanas. Porque es fundamental ser conscientes de que las relaciones primordiales de los seres humanos no se refieren al entorno natural sino al resto de compañeros de especie. Así, la mente humana se formó y se configuró en un ámbito interpersonal y social. Y, por tanto, la inteligencia humana es fundamentalmente inteligencia social antes, y en mayor medida, que inteligencia práctica. De tal modo que se orienta en mayor medida a entender e interpretar la interioridad y las intenciones de sus congéneres humanos antes que a solventar y superar los problemas de mera supervivencia, como el comer, vestirse, escapar de los depreda-dores, y cualquier otro problema de esta índole. En realidad, ambas di-mensiones están estrechamente relacionadas, pero en el caso de los huma-nos la relación con el entorno natural y la lucha por la supervivencia hay que enmarcarla desde y subordinarla a la relación social, y no al revés, como la mayor parte de las veces se ha entendido. De este modo, todo lo que hacemos los humanos es fruto y consecuencia de este cúmulo de inte-racciones sociales. En ese sentido, como indica N. Humphrey, esa miradainterior9 con la que estamos dotados los seres humanos de modo innato, nos convierte en psicólogos naturales, capacitados para vernos a nosotros mismos conscientes de lo que significa ser yo, un ser humano, y al mismo tiempo dotados para entender e interpretar el modo como los otros seres humanos se entienden y son capaces de ponerse en el lugar de los demás.
Si contemplamos la vida humana, descubrimos que la mayoría de nuestros actos están ya orientados y encarrilados por la rutina de las cos-tumbres sociales. Pero en realidad esas costumbres e instituciones so-
8 Cfr. Buskes, Chr., o.c., p. 236-238. 9 Cfr. Humphrey, N., La mirada interior, o.c.

269
© Universidad de Deusto - ISBN 978-84-9830-383-4
ciales, auténticos carriles de nuestra existencia, están impregnados de interacciones sociales en las que los individuos tienen que poner en per-manente funcionamiento sus habilidades de psicólogos naturales. Y esta habilidad es nuestra mayor ventaja en el proceso evolutivo y en la lucha por la supervivencia. Ahora bien, el precio que pagamos por ello es el te-ner que estar en una permanente vigilancia psicológica, dentro de la cual se establece todo el conjunto de nuestras relaciones sociales: amistad, ce-los, ayuda, competencia, cooperación, agresividad, etc. La mayor tarea de todo ser humano consiste en la capacidad de socializarse correctamente y en aprender los mecanismos psicológicos con los que estamos entrela-zados por naturaleza los componentes de nuestra especie. Esta capacidad psicológica es lo que hizo más fuerte y compacta la estructura social de nuestra especie, en la medida en que favoreció un tipo de relación inter-personal y social más fuerte, un modo de organizar los diferentes roles so-ciales que dotaron al homo sapiens sapiens de una eficacia privilegiada de cara a la relación y lucha con los depredadores y las demás especies riva-les. Capacidad que aumentó con el logro posterior del lenguaje articulado. De hecho, es la única especie que ha quedado y sobrevivido a partir de los chimpancés.
Parece que la especie humana es la única dotada de esta capacidad de autoconciencia, aunque no es exagerado atribuir a ciertos animales supe-riores un cierto grado de conciencia, a tenor de las últimas investigacio-nes con chimpancés y otros simios superiores10. Nos encontramos de este modo con el problema de dilucidar si la diferencia entre la conciencia hu-mana y la de otros animales es meramente cuantitativa o cualitativa. Sin adentrarnos en grandes precisiones, porque más adelante analizaremos con más detenimiento este tema (de la distinción entre la especie huma-na y el resto de los animales), estamos de acuerdo con la afirmación de J. Ruffié respecto a que el ser humano no innova prácticamente nada, sino
10 Premack, David, «Is language the key to human intelligencie?», Science, 2003, 303, pp. 318-320; Savage-Rumbaugh, E. Sue, «Why are we afraid of apes with language?», en Scheibel, A. B./Schopf, J. W. (eds.), The origin of evolution of intelligence, Boston, Jons and Bartlett, 2001, pp. 43-69; Id. y R. Lewin, Kanzi: The ape at the brink of the human mind, New York, Wiley, 1994; Id. and Rumbaugh, D. M., «Perspectives on conscious-ness, language, and other emergent proceses in apes and humans», en Hameroff, S. R./Kaszniac, A. W./Scott, A. (eds.), Towards a scicence of consciousness II: The second Tucson discussions and debates, Cambridge, Mass., MIT Press, 1998, pp. 533-549; Id./Shanker, S./Taylor, T. J., «Apes with language», Critical Quarterly, 1996, n.º 38, pp. 45-57; Id./Id./Id., Apes, language, ande the human mind, Nueva York, Oxford University Press, 1998; Hart, S., El lenguaje de los animales, Barcelona, Omega, 1997; Kenneally, Christine, La primera palabra. La búsqueda de los orígenes del lenguaje, Madrid, Alian-za, 2009; Roger Fischer, Steven, Breve historia del lenguaje, Madrid, Alianza, 2003, pp. 32-44.

270
© Universidad de Deusto - ISBN 978-84-9830-383-4
que conserva los rasgos que han aparecido con antelación en el proceso evolutivo, pero llevándolos hasta extremos de enorme complejidad, muy lejanos de los animales anteriores11. Esto puede aplicarse también al tema de la conciencia humana. Todo ser vivo posee un centro vital, su mismi-dad, que estructura y unifica sus diferentes partes, y dirige desde sí sus diversas relaciones con el entorno (es lo que quiere decir Zubiri cuando afirma que todo ser vivo es un autós). Pero esa capacidad de ser dueño de sí mismo se va complejificando en el proceso evolutivo hasta alcanzar un alto nivel en los animales superiores, y llegar en el ser humano al nivel de la autoconciencia. De tal modo que los animales superiores saben de la realidad y de sí mismos, pero sólo el ser humano sabe que sabe. Somos animales excéntricos (Plessner)12, al estar dotados de mismidad y suidad.
Cuando se habla de la distinción entre la conciencia animal y la au-toconciencia humana se suelen distinguir dos posturas dispares: la que tiende a ver entre ellas una mera diferencia cuantitativa, y la que entiende que la distancia es esencial y cualitativa. La primera se apoya en lo que parece más bien un error, el pensar que la autoconciencia posee grados y tamaños. Es lo que N. Humphrey denomina el error de Santa Catalina(haciendo referencia al cuento de Anatole France de la Isla del Pingüino),en la medida en que parece evidente que el bucle de la autoconciencia no admite grados: o se tiene o no se tiene, y por tanto no admite grados o pel-daños intermedios13. De hecho, aunque advertimos en los animales con-ductas de alto grado de inteligencia, no parece que posean la complejidad de nuestra vida social. Y sólo en ciertos chimpancés parece hallarse una cierta conciencia de sí. Pero la única autoconciencia que conocemos es la nuestra, la de la especie humana, y desde ella tratamos de comprender a los compañeros de especie y a las demás especies vivas.
De todos modos, al comparar la conducta animal y la humana, no se da entre ambas, como ya hemos indicado, una ruptura total, como si en el hombre se diera una autoconciencia transparente y los animales fue-ran máquinas instintivas. Se da una gradación entre los animales dotados de una estructura comportamental basada en un programa cerrado a otros dotados de conductas orientadas por programas más abiertos, en los que los organismos está obligados a tomar decisiones vitales aunque no se
11 Cfr. Ruffié, J., De la biología a la cultura, o.c. 12 Cfr. Plessner, H., Dei Stufen des Organischen und der Mensch. Einleitung in die
Philosophische Antropologie (Los estratos de lo orgánico y el hombre. Introducción a una Antropología filosófica), Berlín, 1928 (2.ª ed.: 1964), Beorlegui, C., Antropología filosó-fica. Nosotros: urdimbre solidaria y responsable, Bilbao, Universidad de Deusto, 2004, pp. 374-393.
13 Cfr. La mirada interior, o.c., pp. 78 y ss.

271
© Universidad de Deusto - ISBN 978-84-9830-383-4
pueda hablar de conciencia. Y en los primates, se advierte una cierta auto-conciencia rudimentaria, dotados incluso algunos de lo que se denomina teoría de la mente: capaces de ponerse en la mente de otro. Sólo el ser hu-mano posee autoconciencia plena, capacidad de ser consciente de sí y de ponerse en el lugar del otro reciprocante.
Para entender este proceso hacia la autoconciencia, Antonio Damasio indica tres momentos relacionados, en los que se dan saltos hacia la fase siguiente: el proto-yo, la conciencia nuclear y el yo autobiográfico14. El proto-yo se compone de procesos cerebrales no conscientes, que regulan las funciones corporales, vegetativas, controlando las funciones vitales que nos permiten seguir viviendo. La conciencia nuclear provee al orga-nismo vivo de un sentido del aquí y del ahora, dotándole de experiencias conscientes, pero de un nivel primitivo, como el dolor o el placer. El ter-cer nivel corresponde al yo autobiográfico, cuando los individuos poseen ya una identidad propia, una idea desarrollada de sí mismos, y memoria; son capaces de reflexionar y de mantener los recuerdos fundamentales de su propia vida pasada. Otros estudiosos, como el psicólogo Piet Vroon, han defendido que estos tres niveles de conciencia, estarían afincados en tres niveles y etapas de la evolución del cerebro: el cerebro reptiliano, el mamífero y el humano15.
Todo esto lo que nos indica es que la conciencia fue surgiendo de un modo gradual a lo largo del proceso evolutivo. No es algo que surgió de repente, de la nada, sino que ha tenido un largo proceso de irse encen-diendo de modo gradual. Por eso que, al hablar de la conciencia humana, como autoconciencia, tenemos que considerar, al igual que vamos defen-der en todas las demás características de lo humano, un aspecto de conti-nuidad con los animales, y otro de salto, ruptura y emergencia. Entre lo humano y el resto de los animales, se da a la vez continuidad y ruptura.
El problema que ahora tenemos que dilucidar se desdobla en dos caras: cómo y cuándo se produce el surgimiento de la autoconciencia en el ser humano, de la conciencia reflexiva o de doble nivel. En ambas preguntas la dificultad es casi insalvable, porque el proceso de la formación de la con-ciencia contiene dos aspectos que hay que saber conjugar adecuadamente: por un lado, gradualidad y continuidad, y, por otro, ruptura y salto cualita-tivo. Está claro que se trata de un proceso gradual y progresivo, como una especie de gradiente progresivo, por lo que resulta difícil indicar algo así como el primer momento en el que se da o aparece la autoconciencia en la
14 Cfr. Damasio, A., El error de Descartes, Barcelona, Crítica, 1996; Buskes, Chr., o.c.,240-242.
15 Cfr. Buskes, Chr., La herencia de Darwin. La evolución en nuestra visión del mundo, Barcelona, Herder, 2009, p. 241.

272
© Universidad de Deusto - ISBN 978-84-9830-383-4
vida de una persona, ni tampoco el surgimiento de la autoconciencia en el proceso filogenético. Como resulta evidente, en ninguno de los dos aspec-tos, ontogenético y filogenético, podemos apelar a las evidencias fósiles, ni a los análisis moleculares, cerebrales o genéticos, para resolver este pro-blema. No nos queda más solución que acercarnos por caminos indirectos. Es evidente que la autoconciencia no surge desde el primer momento de la existencia de los seres humanos, sino que va precedida de un proceso de preparación en la que el niño va tomándole el pulso a su entorno, natural y humano, y a sí mismo. La toma de conciencia de su propio yo será más bien un proceso laborioso, el punto de llegada incluso de la toma de con-ciencia de la personalidad de los otros. En ese proceso tienen una gran im-portancia los juegos, en los que los niños tratan de ponerse en el lugar de los otros, en un ejercicio de aprender y de investigar lo que significa ser otros. Algo similar parece que ocurre en los sueños, y desde esta hipótesis se aclararían bastantes de los interrogantes que los especialistas se plantean a la hora de preguntarse por qué y para qué soñamos16.
Es evidente que la capacidad de la autoconciencia, y de saber poner-nos en el lugar del otro, ha ido precedida de este largo proceso de aprendi-zaje, en el que resulta muy difícil indicar el momento preciso en el que se dio el salto emergente de lo no consciente a la autoconciencia. Pero tam-bién tenemos que insistir en lo que este proceso tiene de ruptura y de salto discontinuo. Es decir, por más que insistamos en la continuidad y la gra-dualidad, también hemos de decir que se da una ruptura o salto entre la no-conciencia, o conciencia de primer grado, y la conciencia reflexiva. Es un salto de todo o nada, no caben términos medios. Las etapas anteriores serán preparatorias y anticipadoras, pero se quedan en un nivel cualitati-vamente inferior.
Aunque no podamos indicar con exactitud el momento y el proceso causal de la emergencia de la conciencia reflexiva, sí podemos referirnos con certeza a lo que podríamos considerar los tres factores que constitu-yen la condición de posibilidad de esta esencial capacidad humana: haber traspasado lo que hemos denominado las dos fronteras de la hominiza-ción, la cromosómica y la cerebral, así como un entorno social humano. Es decir, la frontera cromosómica posibilitó la formación en nuestra es-pecie de cerebros capaces de una capacidad de autoconciencia que no pa-rece darse más que en nuestra especie. Pero esta capacidad sólo es capaz de surgir, y de llegar a funcionar al estilo de los humanos, cuando el pro-ceso de maduración infantil se da en un entorno humano en el que el niño
16 Cfr. Buskes, Chr., o.c., cap. 6.º, 7.º. Una utilidad similar tendría la costumbre humana de contar cuentos y escuchar y escribir historias, algo que gusta tanto a los niños (cap. 8.º).

273
© Universidad de Deusto - ISBN 978-84-9830-383-4
aprende a ser yo, persona, porque sólo en ese entorno interpersonal es donde es tratado como conciencia reflexiva y aprende a ver comportarse a los demás humanos con las reglas de juego propias de los psicólogos naturales que somos. Y cuando carece de tal entorno y contexto, como ocurre en los diferentes casos de los llamados «niños lobos»17, dicho pro-ceso queda en suspenso o se realiza con serias patologías.
Otro aspecto fundamental que las investigaciones sobre la mente nos están dando a conocer es lo que se denomina el carácter modular de la mente. Hemos insistido en el carácter gradual de la aparición de la mente, pero, hasta no hace mucho tiempo, se pensaba que esa maduración gradual se producía como un fenómeno unitario, sin compartimentos, cosa que en la actualidad se cuestiona, o más bien se asegura que no es así. Se habría pasado, pues, de un estudio unitario de la evolución de la mente, tal y como lo realiza el psicoanálisis freudiano y las investigaciones de la psicología genética de la escuela de Piaget, entre otros, a entender la mente con una estructura semejante al plano de una vivienda con diversas habitaciones. La mente parece tener diversos compartimentos o módulos, con cierta au-tonomía, que maduran de modo autónomo y a ritmos diferentes. El prime-ro teórico de la mente que empezó a hablar de la modularidad de lo mental fue Jerry Fodor18, pero los modos en que después se ha entendido esta mo-dularidad son muy diversos, destacando los denominados psicólogos evo-lucionistas. Sin poder detenernos demasiado en este aspecto de lo mental, que en la actualidad está cobrando una importancia relevante19, podemos considerar que la teoría de la modularidad de la mente se ha ido afianzan-do desde diferentes puntos de vista, sea desde la psicología o filosofía de la mente, como también desde la lingüística generativa o la paleoantro-pología. La tesis de Fodor distinguía un módulo de percepción o sistemas de input, junto al módulo de cognición, o sistema central. Posteriormente, Howard Gardner ha acuñado el concepto de inteligencias múltiples20. Pero
17 Cfr. Itard, J., Victor de L’Aveyron, Madrid, Alianza, 1982 (introducción y comenta-rios de R. Sánchez Ferlosio); Cavada, M., La vigencia del amor, Madrid, Edic. Paulinas, 1994; Beorlegui, C., «De al autonomía a la alteridad. Hacia una nueva racionalidad», Le-tras de Deusto, 17 (1987), n.º 38, pp. 41-61; Id., «El hombre y sus imágenes. Del narcisis-mo a la alteridad», Letras de Deusto, 22 (1992), n.º 52, pp. 81-108.
18 Cfr. Fodor, J., The Modularity of Mind, Cambridge, Mass., MIT Press, 1983. 19 Cfr. para estos temas, Mithem, Steven, Arqueología de la mente, Barcelona, Críti-
ca, 1998. Sobre la Psicología evolucionista, cfr. Buskes, Chr., o.c., cap. 5; Castro Noguei-ra, Laureano/Castro Nogueira, Luis/Castro Nogueira, M. A., ¿Quién teme a la naturaleza humana?, Madrid, Tecnos, 2008, cap. 3.º; Dupre, J., El legado de Darwin, Buenos Aires, Katz Editores, 2006, cap. 6.
20 Cfr. Gardner, H., Frames of Mind: the Theorie of Multiple Intelligences, Nueva York, Basic Books, 1983 (Inteligencias múltiples, Barcelona, Paidós, 1987).

274
© Universidad de Deusto - ISBN 978-84-9830-383-4
los psicólogos evolucionistas interpretan la figura de la mente de una forma más compleja, como una navaja suiza, dotada de diversos compartimentos autónomos: la mente que rige la relación con el entorno material, con las plantas, con los animales, con los demás seres humanos, el módulo lingüís-tico y el módulo central o del yo. Cada módulo sería autónomo y poseería su propio proceso de maduración, pero llegado un momento del proceso de maduración humana, los diversos módulos entrarían en contacto unos con otros, y todos ellos se hallarían coordinados y dominados por el módulo central, el del yo autoconsciente21. Si ya es difícil, como puede imaginar-se, establecer desde el punto de vista psicológico la distinción entre estos diversos módulos, todavía lo es más establecer hipótesis consistentes sobre los momentos en los que se originan cada uno de estos módulos y las fa-ses de su evolución en el proceso evolutivo de los humanos, y sus primeros atisbos entre los primates. Pero los psicólogos evolucionistas y los paleoan-tropólogos se hallan enfrascados en el empeño de hallar pruebas objetivas que marquen los mojones de estos procesos de evolución de nuestra mente primitiva.
Como puede verse, la mente autoconsciente, rasgo que nos caracteriza como humanos, constituye una realidad de enorme complejidad y cargada de muchísimos interrogantes, no sólo en el aspecto de su origen y evolu-ción diacrónica, sino también en el aspecto de su configuración sistémica o modular.
2.2. La imaginación y la capacidad simbolizadora
La imaginación es la capacidad de conformar un mundo mental en ausencia del objeto pensado. Esta virtualidad es consecuencia del importante poder intelectual que le ha dotado el proceso de cerebra-lización. El mundo interior abstracto que el ser humano es capaz de construir, está configurado por signos y símbolos22. Los dos aspectos, imaginación y capacidad simbolizadora, están estrechamente unidos y constituyen un avance importante en el proceso evolutivo, dotándole a la especie humana de una gran ventaja en el ámbito de la selección na-tural. Aunque sus virtualidades no hay que entenderlas sólo en el marco del proceso selectivo.
21 Cfr. Mithem, S., La arqueología de la mente, o.c.22 Cfr. Cassirer, Ernst, Filosofía de las formas simbólicas, 3 vols., México, FCE, 1973-75;
Id., Mito y lenguaje, Buenos Aires, Nueva Visión, 1959; Id., Antropología filosófica, Méxi-co, FCE, 1945; Beorlegui, C., Lecturas de Antropología filosófica, Bilbao, Desclée, 1988, cap. 8.º, «El símbolo y la utopía», pp. 245 y ss.

275
© Universidad de Deusto - ISBN 978-84-9830-383-4
Por la capacidad imaginativa, el ser humano tiene la virtualidad de analizar los sucesos del pasado, evaluarlos y advertir sus fallos y aciertos, tanto de cara a la lucha por la supervivencia como de cara a las relaciones con los demás compañeros de la especie. De ese modo, es capaz de corre-gir su modo de comportarse, aprender de sus errores y aciertos, y benefi-ciarse de las lecciones que le aporta dicho análisis del pasado. Pero también puede anticipar el porvenir, imaginando diversas alternativas y modos de actuación posibles, convirtiéndose su imaginación en un auténtico labora-torio de pruebas y de ensayos, que le evitan tener que esperar a que la rea-lidad le muestre las consecuencias de los diversos modos de actuación po-sibles ante un depredador, un compañero de especie, la utilización de una herramienta, probar una técnica o procedimiento desconocido, etc. Esta capacidad imaginadora está a la base tanto de su capacidad de usar y cons-truir herramientas, como de la capacidad y uso del lenguaje.
Como hemos dicho, el modo de encerrar el mundo y los diversos fu-turibles de actuación en su escenario imaginativo es a base de símbolos y conceptos abstractos. El ser humano es, pues, un animal symbolicum(Cassirer), vive en un mundo artificial, la cultura, que colorea y transfor-ma todo el mundo natural. De ahí que podamos decir que el ser humano ha superado la naturaleza, para afincarse en ese ámbito o mundo que de-nominamos cultura.
También en este punto resulta un tanto problemático delimitar la frontera que separa la capacidad humana y la de las demás especies animales. No es fácil aceptar que la especie humana es la única que posee capacidad simboli-zadora, en la medida en que algunos simios parecen poseer un cierto tipo de capacidad simbolizadora, necesaria para poder utilizar algún tipo de lengua-je23. Para diferenciar entre la realidad humana y la de los demás animales, en lo que a este punto se refiere, se suele diferenciar entre signo y símbolo24.En nuestro proceso de comunicación con los demás, utilizamos permanen-temente señales, entendidas, en términos generales, como cosas, gestos, so-nidos, procesos, a través de las que nos referimos a las cosas del mundo a la hora de llamar la atención sobre ellas a los copartícipes de nuestras acciones comunicativas. Pero las señales pueden ser de muy diferentes tipos, distin-guiéndose aquí entre los signos y los símbolos. Serán signos los tipos de se-ñales en los que entre la señal y lo señalado existe una relación inmediata y unívoca. Así, un signo es el humo respecto del fuego, o el dibujo de una casa
23 Cfr. Gomila, A., «Evolución y lenguaje», en Broncano, F. (ed.), La mente humana,Madrid, Trotta, 1995, pp. 273-299.
24 Cfr. Laín Entralgo, Pedro, «La psicoterapia como somatoterapia: proceso de simboli-zación», en Alemany, C./García, V. (eds.), El cuerpo vivenciado y analizado, Bilbao, Des-clée de Brouwer, 1996, pp. 45-61.

276
© Universidad de Deusto - ISBN 978-84-9830-383-4
o de un animal respecto al objeto que representan. Se trata, como puede ver-se, de una relación directa y unívoca, y, también por ello, inmediata.
En cambio, en el caso de los símbolos, la relación entre la señal y su objeto no es tan directa, unívoca e inmediata, sino que se trata de una re-lación artificial y convencional, por lo que se necesitará para establecer tal convención simbólica una serie de sujetos con una mayor capacidad inte-lectual e intencional. La vida humana está llena de símbolos, de tal forma que podemos referirnos a símbolos gestuales (dar la mano), verbales (gri-tos, palabras), objetuales (banderas, señales de tráfico), poéticos (metáfo-ras), míticos (el ámbito de las diversas mitologías), etc.
Según esto, podríamos afirmar que a lo más a que pueden llegar deter-minados animales superiores (simios) es a captar y utilizar determinados signos, pero sólo los seres humanos tenemos capacidad de utilizar además símbolos. Los llamados lenguajes animales25 son modos analógicos de comunicación entre ellos, pero difícilmente pueden ser llamados lengua-jes en el mismo sentido que los seres humanos, puesto que los animales no poseen la suficiente capacidad semántica para dotar a los signos de significados abstractos, a pesar de la admiración que nos causan los des-cubrimientos del etólogo Karl von Frisch sobre el lenguaje de las abejas,y otros investigadores sobre las capacidad que pueden llegar a tener algu-nos chimpancés con el uso de los lenguajes de sordomudos26.
La cuestión que queda pendiente es cuándo los seres humanos comen-zaron a tener esta capacidad imaginativa y simbolizadora. Para responder a esta cuestión nos encontramos con las mismas dificultades que veíamos en relación a la conciencia reflexiva. Pero no resulta descabellado presu-poner la posesión de un cierto grado de estas cualidades ya en el primer homo habilis, puesto que parece necesario suponerlo de cara a la utiliza-ción de las primeras herramientas y posterior construcción, en la medida en que tenemos que dar por hecho tal capacidad de usar y de construir he-rramientas como condición primera y básica para marcar las diferencias entre las especies homo y las de los australopitecos.
2.3. El lenguaje
El lenguaje es sin duda una de las características más específicas de la condición humana, propiedad que a primera vista más la distin-gue del resto de las especies animales, puesto que se trata de un instru-
25 Cfr. la bibliografía de las notas 10 y 23. 26 Cfr. Savage-Rumbaugh, E. S./Shanker, S./Taylor, T. J., Apes, language, and the hu-
man mind, New York, Oxford University Pres, 1998

277
© Universidad de Deusto - ISBN 978-84-9830-383-4
mento de comunicación con caracteres muy singulares, tanto fonadoras y semánticas, como sintácticas y pragmáticas27. Sin embargo, a pesar de estas singularidades definitivas, se sigue poniendo en duda que esto constituya una diferencia cualitativa, defendiendo en cambio que el len-guaje humano sólo se distingue del resto de los lenguajes animales sólo por una mayor complejidad cuantitativa, no pudiéndose hallar en él una diferencia cualitativa y ontológica. Son muy reveladoras estas pala-bras de Richard Leaky: «Aquellos que desean mantener a los humanos como algo especial darán la bienvenida a pruebas que señalen un origen abrupto y reciente del lenguaje. A aquellos que se sienten cómodos con la conexión de los humanos con el resto de la naturaleza no les conster-nará un temprano y lento desarrollo de esta capacidad quintaesencial-mente humana»28.
El lenguaje humano ha sido estudiado desde múltiples puntos de vista, y desde épocas muy tempranas. Y en la actualidad, el estudio del origen y la evolución del lenguaje está siendo objeto de múltiples inves-tigaciones y de fuertes debates29. La cuestión de la procedencia del len-guaje ha sido objeto de múltiples teorías, desde quienes defendían que provenía de los gritos involuntarios surgidos de trabajos conjuntos, o de la imitación de los sonidos de los animales, hasta quienes consideraban que provenía de los gritos instintivos de placer, alegría y dolor, o incluso aportaban teorías míticas y religiosas, situando su origen en Adán en el momento de dar nombre a los animales30. De ahí que Sociedad Lingüísti-ca de París, en 1866, prohibiera seguir especulando sobre este punto, por considerar que no había base científica para ello y para aportar algo sen-sato sobre el tema.
Pero, tras unos años de estancamiento, las investigaciones sobre el origen del lenguaje están experimentando una considerable aceleración, a través de estudios de lingüistas, neurocientíficos, biólogos evolutivos, psicólogos, y paleoantropólogos, orientando sus reflexiones tanto en su dimensión ontogenética como filogenética, cobrando así estos estudios una relevancia y profundidad inesperadas y hasta ahora desconocidas. En
27 Como indica Ch. Taylor, «la cuestión del lenguaje es en cierto modo estratégica para el problema de la naturaleza humana, y el hombre es sobre todo el animal del len-guaje», La libertad de los modernos, Buenos Aires/Madrid, Amorrotu Editores, 2005, p. 35.
28 Leaky, R., El origen de la humanidad. Cita tomada de Lorenzo, G. /Longa, V. M., Homo Loquens. Biología y evolución del lenguaje, Lugo, Edit. Tris Tram, 2003, p. 85.
29 Cfr. Kenneally, Christine, La primera palabra. La búsqueda de los orígenes del len-guaje, Madrid, Alianza, 2009; Buskes; Chris, La herencia de Darwin. La evolución en nuestra visión del mundo, Barcelona, Herder, 2009, cap. 7 (Evolución y lenguaje);
30 Cfr. Buskes, Ch., o.c., 191-192.

278
© Universidad de Deusto - ISBN 978-84-9830-383-4
concreto, tras las determinantes propuestas de la gramática generativa de N. Chomsky31, se ha abierto un espléndido campo de investigación que ha dado un fuerte impulso a la reflexión sobre el origen y las características específicas del lenguaje humano.
Durante mucho tiempo se ha tenido el convencimiento de que el len-guaje humano era un producto cultural, similar en ello al resto de habi-lidades culturales propias de nuestra especie. Somos animales deficien-tes, sin una estructura instintiva fiable a la que pueda abandonarse sin reflexión, por lo que tiene que construir inevitablemente un mundo cul-tural que supla lo que la naturaleza no le ha otorgado32. La deficiencia biológica ha puesto a prueba la supervivencia de la especie humana, que ha tenido que introducir un hiato o paréntesis de reflexión, de pruebas de ensayo y error, para poner en marcha la ley de la descarga (Gehlen) e ir construyendo el mundo cultural, sustituto de la naturaleza biológica perdida. En relación con la capacidad lingüística, se suponía que el ser humano nacía como una tabla rasa y el entorno ambiental era quien se encargaba de enseñarle a hablar, para poder ponerse en contacto con sus semejantes e introyectar de ese modo el rico bagaje cultural que los seres humanos, en su condición de herederos de las generaciones ante-riores33, van transmitiendo a las sucesivas generaciones. La cultura ge-nerada con la puesta en marcha de esa ley de la descarga de que habla Gehlen, no es algo individual, que cada individuo la tiene que reinventar y poner en marcha, sino un producto social que se va osificando en las diversas instituciones que van conformando el complejo sistema social y cultural.
Ha sido recurrente, de todos modos, la polémica entre innatistas y am-bientalistas a la hora de entender el proceso del aprendizaje lingüístico. Mientras los ambientalistas extremos defendían la tesis de la tabla rasa,siendo partidarios de un cierto determinismo ambiental, como los conduc-tistas Watson y Skinner, por el contrario, ya desde Darwin, e incluso con anterioridad con W. von Humboldt, se tiende a insistir en el aspecto innato de dicho aprendizaje. Pero no ha sido hasta los estudios de N. Chomsky y sus tesis sobre la gramática generativa cuando estas tesis innatistas se han afianzado de modo definitivo. Estudiando la manera como los niños
31 Chomsky, N. (1966), Cartesian linguistic. A chapter in the history of rationalistic thought, New York, Harper and Row (Lingüística cartesiana. Un capítulo de la historia del pensamiento racionalista, Madrid, Gredos, 1969); Chomsky, N. (1968), Language and mind, New York, Harcourt Brace Jovanovich (El lenguaje y el entendimiento, Barcelona, Seix Barral, 1971).
32 Cfr. Gehlen, A., Der Mensch, Franlfurt, Athenaion Verlag, 1940 (2.ª ed. 1974) (Elhombre. Su naturaleza y su lugar en el mundo, Salamanca, Sígueme, 1980).
33 Cfr. Ortega y Gasset, J., Historia como sistema, Madrid, Espasa-Calpe, 1935 (1971).

279
© Universidad de Deusto - ISBN 978-84-9830-383-4
aprenden a hablar, resulta evidente para Chomsky que el lenguaje no es algo que el niño aprende, sino algo que acontece en él. Según Chomsky, «el lenguaje del niño “crece en la mente” como el sistema visual desarro-lla la capacidad para la visión binocular, o como el propio niño alcanza la edad de la pubertad en un cierto momento de su crecimiento. La adquisi-ción del lenguaje es algo que le sucede al niño localizado en un cierto am-biente, no algo que el niño haga»34.
Esto no supone negar la dimensión cultural del lenguaje y de la con-dición humana, puesto que el aspecto innato no lo es todo, pero sí implica la necesidad de conjugar de otro modo ambas dimensiones. Así, insistir en la capacidad innata supone entender que un individuo dotado de mayo-res capacidades genéticas será capaz de aprovechar en mayor medida las aportaciones del entorno cultural. Y no supone tampoco defender posturas deterministas, ni defensoras de cualquier clase de racismo o sexismo, sino reconocer objetivamente la parte que corresponde a cada elemento (inna-to o adquirido) y complementarlos de la forma más adecuada. Ahora bien, sería un error entender que esta mayor referencia a lo innato implica rei-vindicar de nuevo la idea de naturaleza humana, entendida como un asu-mir la idea que lo que caracteriza y define lo humano está sobre todo pre-sente en sus dotes biológicas o genéticas. En alguna medida, por ahí van algunos autores contemporáneos, sin que se atrevan a explicitar del todo sus pretensiones35.
Por tanto, el lenguaje es algo que pertenece a la dotación genética de la especie humana, transmitida por generación a todos sus integran-tes. Todos los humanos poseemos dicha capacidad, aparecida en el pro-ceso evolutivo en fecha reciente. La pregunta que se hacen hoy día los especialistas es cuándo se produjo su aparición o emergencia, y, sobre todo, por qué, siendo más importante que el momento cronológico el descubrir cuál habría sido el mecanismo y la lógica que marca la apa-rición de la capacidad lingüística tanto en la dimensión ontogenética como filogenética.
Pero entre los innatistas, y a la hora de plantearse la cuestión del ori-gen del lenguaje, los especialistas se dividen en dos líneas diferentes: los que entienden este proceso como consecuencia de la lógica de la presión selectiva, defendiendo la postura gradualista en el proceso evolutivo, y
34 Chomsky, N., Language and thought, Wkafield, Moyer Bell, 1993, 29; Lorenzo, G./Longa, V. M., o.c., 45.
35 Cfr. Pinker, S., La tabla rasa. La negación moderna de la naturaleza humana, Ma-drid, Alianza, 2004; Mosterin, J., La naturaleza humana, Madrid, Espasa-Calpe, 2006; Beorlegui, C., «A vueltas con la naturaleza humana», en Arregui, J. V. (ed.), Debate sobre las antropologías, Thémata. Revista de Filosofía (Sevilla), 2005, n.º 35, 139-150.

280
© Universidad de Deusto - ISBN 978-84-9830-383-4
los que consideran que la emergencia del lenguaje se debió a un proceso de ruptura y emergencia, coincidiendo de ese modo con la postura rup-turista o de equilibrio puntuado. Entre los primeros se hallan S. Pinker, P. Bloom, y Ph. Lieberman entre otros36. Para S. Pinker, el lenguaje hu-mano funciona como un instinto37. En cambio, para N. Chomsky y S. Jay Gould, el lenguaje no habría sido resultado de la evolución, sino que ha-bría surgido de forma repentina como resultado de un proceso de acomo-do de la masa encefálica dentro de la cavidad craneana. Así, la capacidad lingüística no habría tenido nada que ver con la biología evolutiva, sino que sería un fenómeno que S- Jay Gould denomina de exadaptación, o exaptación. Vendría a ser como las pechinas, espacios triangulares situa-dos entre las columnas y la bóveda de las catedrales38.
Ahora bien, esta separación de posturas no es tan rígida, puesto que autores como Pinker y otros, se declaran gradualistas respecto a la di-mensión filogenética, aunque defienden una postura rupturista respecto a la ontogénesis. Estas dos posturas suponen defender también, y de modo respectivo, dos modos diferentes de entender la diferencia, cualitativa o meramente cuantitativa, entre el lenguaje humano y el del resto de las es-pecies animales39.
A la hora de defender la especificidad o no del lenguaje humano, nos tenemos que preguntar si las demás especies animales poseen la capaci-dad del lenguaje, y en qué se diferencia del lenguaje humano. Para ello, tenemos que comenzar poniéndonos de acuerdo en la definición de len-guaje. Siguiendo a A. Gomila, «el lenguaje consiste en un medio de em-parejar significados (o mensajes) con sonidos, y viceversa (ya que cubre tanto los aspectos de producción como de comprensión)»40. En esta defi-nición, se considera el lenguaje humano articulado como instrumento es-pecífico y radicalmente diferente de los demás medios comunicativos de otras especies animales.
36 Cfr. Kenneally, Chr., o.c., pp. 77-125.37 Cfr. Pinker, S., The language instinct, Nueva York, William Morrow, 1994 (El instin-
to del lenguaje, Madrid, Alianza, 2001); Id., El mundo de las palabras, Barcelona, Paidós, 2007.
38 Cfr. Gould, S. J./Lewontin, R., «The spandrels of San Marcos and the Panglossian paradigm: a critique of the adapatationist programme», Proceedings of the Royal Society of London, 1979. B 205, 581-598; Gould, S. J./Vrba, E., «Exaptation: a missing term in the science of form», Palebiology, 1981, vol. 8, 4-15; Gould, S. J./Lewontin, R. C., «La adap-tación biológica», Mundo Científico, 1983, n.º 22, 214-223.
39 Cfr. Buskes, Chr., o.c., 194-202; Beorlegui, C., «La capacidad lingüística del ser hu-mano: una diferencia cualitativa», Thémata (Sevilla), 2006, n.º 37, 139-168.
40 «Evolución y lenguaje», en Broncano, F. (ed.), La mente humana, Madrid, Trotta, 1995, 273-288; 276.

281
© Universidad de Deusto - ISBN 978-84-9830-383-4
Dejando de lado las polémicas acerca de cómo entender las experien-cias de los investigadores que trabajan sobre lenguajes animales41, es evi-dente que ningún animal alcanza la capacidad fonética que posee nuestro lenguaje. Pero también se diferencia del lenguaje humano en los aspectos semántico, sintáctico y pragmático. De ahí que, sin negar la existencia de la capacidad comunicativa que poseen las diferentes especies animales para relacionarse entre sí42, la aparición del lenguaje humano articulado supuso un salto cualitativo incontestable, teniendo que distinguir clara-mente entre los lenguajes animales (todo tipo de expresiones faciales, gestos, posturas, silbidos, signos manuales, etc.) y el lenguaje humano, dotado de unas características fonéticas, semánticas, sintácticas y pragmá-ticas a las que los sistemas comunicativos animales sólo se acercan de un modo muy rudimentario. Sería como comparar, según Chomsky, y consi-derar analógico un salto del ser humano con la capacidad de volar de las aves. No se trata, si comparamos esas dos habilidades, de una mera dife-rencia cuantitativa, sino de realidades cualitativamente distintas.
En definitiva, aunque nos puedan admirar las capacidades comunica-tivas de determinadas especies de animales, no llegan más que hasta un nivel rudimentario si las comparamos con las de los seres humanos. En-tre ambos niveles se da tanto una continuidad como una ruptura. Hay una continuidad en el aspecto cognitivo, en la medida en que la conducta ani-mal y su capacidad para transmitir señales a sus congéneres están basa-das en su específico nivel cognitivo, situándose de todos modos el ser hu-mano en estos campos a una distancia ciertamente apreciable. Pero se da también una ruptura y un salto cualitativo desde el momento en el que la especie humana accede a su específica capacidad lingüística, en la medida en que el lenguaje hablado que poseemos los humanos representa una dis-tancia cualitativa tanto en el aspecto fonético, semántico, sintáctico como pragmático43.
Esto nos hace ver, como indica Bickerton, que el lenguaje humano ha aparecido recientemente en el proceso evolutivo, en fechas posterio-
41 Cfr. Premack, D., «Minds with and wihtouth language», en Weiskrantz, L. (ed.), Thought without language, Clarendon, 1988, 46-65; Savage-Rumbaugh, E. Sue, Kanzi: The ape at the brink the human mind, Nueva York, Wiley, 1994; Savage-Rumbaugh, E. S./Shanker, S./Taylor, T. J., Apes, language, and the human mind, Nueva York, Oxford Uni-versity Press, 1998; Kenneally, Christine, La primera palabra. La búsqueda de los oríge-nes del lenguaje, Madrid, Alianza, 2009.
42 Fischer, S. R., A history of language, London, Reaktino Books, 1999 (Breve histo-ria del lenguaje, Madrid, Alianza, 2003); Hart, S., El lenguaje de los animales, Barcelona, Omega, 1997.
43 Cfr. Bickerton, D., Language and species, Chicago, Chicago University Press, 1990 (Lenguaje y especie, Madrid, Alianza, 1994); Gomila, A., o.c.

282
© Universidad de Deusto - ISBN 978-84-9830-383-4
res a la separación del género humano respecto a nuestros antecesores los chimpancés. La pregunta es cuándo, y si todas las especies o subespecies humanas (desde el homo habilis hasta el homo sapiens) estaban dotadas de esta capacidad innata. Los paleoantropólogos tienen sobre este punto diferentes opiniones, apoyadas en argumentos más o menos serios44, pero no nos vamos a detener en este punto, puesto que la cuestión más impor-tante, como hemos indicado ya, no es tanto cuándo sino cómo se produjo la aparición de esta capacidad, esto es, la lógica del proceso de adquisi-ción, tanto en el aspecto filogenético como ontogenético.
Las diferentes posturas sobre el cómo surge la capacidad lingüística en la especie humana están íntimamente relacionadas con el modo como se entiende el proceso evolutivo. Mientras la postura gradualista, presen-tada ya por el propio Darwin, sostiene que todas las características propias de la especie humana tienen antecedentes en especies animales anteriores, y, por tanto, sería su aparición en los humanos consecuencia de un proceso gradual de acumulación progresiva de múltiples y pequeñas mutaciones, la postura rupturista o saltacionista considera que el proceso evolutivo, sobre todo en el momento de la aparición de nuevas especies, se debe al surgimiento de macromutaciones. La postura que hoy día va teniendo más aceptación es la que se denomina de equilibrios puntuados, o puntuacio-nismo45. Según esta tercera teoría, el proceso evolutivo es el resultado de la conjunción de momentos de avance gradual junto a otros momentos en los que se producen rupturas y emergencias macromutacionales rápidas, que originan la aparición de nuevas especies (cladogénesis) o la emergencia de características nuevas e importantes dentro de una misma especie.
Para los defensores del planteamiento gradualista, las mejoras de las diversas formas morfológicas de una especie y la función que dichas for-mas desempeñan en el proceso de la lucha por la supervivencia son con-secuencia de un continuo y prolongado proceso de mejoras mutacionales graduales, siguiendo una especie de racionalidad o proyecto inteligente que dirige esta tendencia gradual. De este modo pretenden superar cual-quier propuesta teleológica, supliéndolo por un modelo evolutivo de di-
44 Cfr. Lieberman, Ph. (1973), «On the evolution of language: A unified view», Cogni-tion, 2 («Un enfoque unitario de la evolución humana», en V. Sánchez de Zabala (ed.), So-bre el lenguaje de los antropoides, Madrid, Siglo XXI, 1976, 147-203); Arsuaga, J. L., Elcollar del Neandertal. En busca de los primeros pensadores, Madrid, Temas de Hoy, 1999; Leaky, R., La formación de la humanidad, Barcelona, Ediciones del Serbal, 1981, cap. 8, 128-143.
45 Eldredge, N./Gould, S. J. (1972), «Punctuated equilibria: An alternative to phyletic gradualism», en T. Schopf (ed.), Models in Paleobiology, San Francisco, Freeman, 82-115; Gould, S. J./Eldredge, N. (1977), «Punctuated equilibria: The tempo and the mode of evolution reconsidered», Paleobiology, 3, 115-151. Cfr. Cap. 2.º de este libro, apartado 3.

283
© Universidad de Deusto - ISBN 978-84-9830-383-4
seño óptimo que se va produciendo de forma gradual, combinando las leyes de la genética con la selección natural. Es la mejor forma, según esta teoría, de entender la formación de formas y órganos tan complejos como es un ojo humano. No lo proyecta una inteligencia consciente, sino el proceso evolutivo que funciona de modo similar a un relojero ciego46.Por tanto, la línea gradualista se apoya en dos principios básicos: formaly funcional. El funcional consiste en entender que toda modificación en un organismo vivo se justifica en función de una mejora adaptativa. De tal modo que la nueva forma orgánica que se produce en función de una lógica que funciona hacia atrás, con una cierta ingeniería inversa, produ-ciendo lo que Dennett denomina objetos diseñoides, a través, como ya se ha indicado, de la acumulación de pequeños cambios que siguen no una orientación con racionalidad prospectiva, sino más bien un principio de estupidez subyacente47.
Para la teoría puntuacionista, esta pretensión de explicar todos los cambios desde la funcionalidad adaptativa tiene el peligro de caer en el vicio del panglosismo, y, hasta cierto punto, en una explicación similar al teleologismo preconsciente que pretendían combatir. Resulta más con-vincente la propuesta de F. Jacob de ver el proceso evolutivo como una operación de bricolage, similar a una adaptación chapucera y aproxima-tiva48. La evolución sería, pues, empujada por momentos de avance gra-dualista junto a otros momentos de ruptura o saltos emergentes, en los que se darían aceleraciones rápidas y apariciones de especies nuevas. Se producirían en esos momentos críticos lo que los teóricos de las ciencias de la complejidad denominan transiciones de fase49. Además, la postura puntuacionista quiere resaltar que estos cambios rápidos (en una rapidez relativa, puesto que hablamos de amplios períodos de tiempo) pueden ser (y, de hecho, lo son en muchas ocasiones) consecuencia de una reestruc-turación formal realizada al interior de los organismos vivos, y no tanto el resultado de un proceso de presión selectiva proveniente del entorno am-biental. Desde esta lógica evolutiva es como explica Chomsky la emer-gencia de la capacidad lingüística en el ser humano.
46 Dawkins, R. (1986), The blind watchmaker, London, Longman (El relojero ciego,Barcelona, Labor, 1988); Dawkins, R. (1995), River out of Eden. A darwinian view of life,New York, Basic Books (El río del Edén, Madrid, Debate, 2000).
47 Dennett, D. (1995), Darwin’s dangerous idea, New York, Simon and Schuster (Lapeligrosa idea de Darwin. Evolución y significado de la vida, Barcelona, Galaxa Guten-berg/Círculo de Lectores, 1999).
48 Jacob, Fr. (1970), La logique du vivant. Une histoire de l’hérédité, París, Gallimard (La lógica de lo viviente. Una historia de la herencia, Barcelona, Tusquets, 1999).
49 Kauffman, S. (1995), At home in the universe. The search for the laws of self-organi-zation and complexity, New York, Oxford University Press.

284
© Universidad de Deusto - ISBN 978-84-9830-383-4
Cada una de las dos teorías evolutivas tiene su propuesta en este punto. Para los gradualistas, el origen del lenguaje humano se debe a una presión selectiva del entorno humano en el que habrían intervenido una serie muy compleja de factores, desde genéticos y morfológicos hasta culturales. En cambio, para los puntuacionistas, siguiendo la propuesta de N. Chomsky, se debería a un proceso de configuración interna de la estructura cerebral. De todos modos, hay quienes defienden una tesis rupturista o emergentista en el ámbito ontogenético, entendiendo que en la dimensión filogenética se habría dado más bien un proceso gradualista50.
Los gradualistas consideran que la forma más plausible de entender el proceso, dentro de la ortodoxia evolucionista, consiste en advertir la emergencia del lenguaje como consecuencia de un proceso de coevolu-ción de diversos factores: la base genética correspondiente51, el aumen-to y sistematización progresivos del cerebro, y los cambios en la cavidad supraglótica que llevaron a un reacomodo de la lengua y a un retroceso y abajamiento de la laringe52, así como un conjunto de factores culturales: cambios en los hábitos alimenticios y en las estrategias de caza y recolec-ción de alimentos, en el aumento y complejidad de los grupos sociales, etc.53. Todos estos elementos habrían realizado una fuerte presión selec-tiva, en la línea de pasar de una fase previa al lenguaje como capacidad innata hasta dar el salto a convertirse en un rasgo genético, que se trans-mitiría posteriormente como un rasgo innato propio de toda la especie, ex-plicándose este proceso por medio del denominado efecto Baldwin54.
Para Chomsky y sus seguidores, el origen del lenguaje no se produ-jo tanto por efecto de una presión selectiva de ese conjunto de factores externos, sino que fue sobre todo consecuencia de un nuevo acomodo y sistematización de la masa encefálica dentro de un entorno craneano es-trecho y difícil55. Esto no significa que Chomsky considere que el lengua-je ha quedado al margen de la lógica evolucionista, sino que sólo reduce
50 Pinker, S./Bloom, P. (1990), «Natural language and natural selection», Behavioraland Brain Science, 13, 707-727; Pinker, S. (1994), The language instinct. How the mind creates language, New York, W. W. Norton (El instinto del lenguaje. Cómo crea el lengua-je la mente, Madrid, Alianza, 1995); Gomila, A., o.c.
51 Holden, C. (2005), «Sur les traces du premier langage», Les Dossier de la Recherche,19, 88-92.
52 Lieberman, Ph. (1973), o.c. 53 Gomila, A., o.c.54 Richards, R. J., Darwin and the emergence of evolutionary theorie of mind and
behavior, Chicago, University of Chicago Press, 1987, 483-486.55 Chomsky, N. (1968), o.c.; Id. (1986), Knowledge of language: Its nature, origins
and use, New York, Praeger (El conocimiento del lenguaje. Su naturaleza, origen y uso,Madrid, Alianza, 1989).

285
© Universidad de Deusto - ISBN 978-84-9830-383-4
dentro de este proceso el peso de la presión selectiva del entorno. Así, «la emergencia del lenguaje podría proceder no (o al menos no directamente) de las presiones selectivas externamente ejercidas por el medio ambiente sobre los homínidos, sino de las presiones internas del propio organismo ante la necesidad de dar un acomodo apropiado a una masa encefálica en constante aumento»56.
Por tanto, este modo de explicar el proceso indica que no hay una ló-gica finalística, desde la presión selectiva, que oriente el origen de la capa-cidad lingüística hacia su conversión en un rasgo innato, aunque su adqui-sición supuso a posteriori una mejora adaptativa indudable. Así, pues, su incorporación en la lógica evolutiva es a posteriori, no a priori, de la mis-ma manera a como explica S. J. Gould la utilidad del pulgar del panda57.
Chomsky ha insistido en sus investigaciones más en el aspecto on-togenético que en el filogenético, pero defiende posturas rupturistas en ambos puntos de vista. Sus argumentaciones han ido dirigidas a mostrar la fuerte dimensión innata presente en el proceso de maduración de la ca-pacidad lingüística de los niños. Como hemos indicado ya, para Chomsky no se trata propiamente de un proceso de aprendizaje, sino de un dinamis-mo que acontece en el propio niño, de naturaleza innata. Si el lenguaje es «una particular puesta en relación entre el sonido y el sentido»58, esto es, entre la capacidad cognitiva del cerebro y el aparato fonador en que se ha convertido nuestra peculiar laringe, las investigaciones sobre el origen del lenguaje tienen que orientarse, por tanto, a tratar de explicar cómo se pu-sieron en contacto estas dos capacidades, hablar y pensar, emitir sonidos y atribuirles sentido, capacidades que podían con anterioridad funcionar separadas. De hecho, determinados chimpancés poseen una importante capacidad cognitiva que les permite interpretar la conducta ajena, pero no alcanzan a dominar la capacidad de relacionar sonidos guturales con con-tenidos semánticos.
56 Lorenzo, G./Longa, V. M., Homo loquens. Biología y evolución del lenguaje, Lugo, Ed. Tris Tram, 2003, 137; Mithen, S. (1996), The prehistory of the mind. A search for the origin of art, religion and science, London, Thames and Hudson Ltd. (Arqueología de la mente. Orígenes de la religión, el arte y la ciencia, Barcelona, Crítica, 1998); Chomsky, N. (1994), «Language from an internalist perspective», Conferencia dictada en el King’s College (Londres), 24 de mayo («El lenguaje desde una perspectiva internista», en Unaaproximación naturalista a la mente y al lenguaje, Barcelona, Prensa Ibérica, 1998, 193-230);Bickerton, D. (1990), Language and species, Chicago, Chicago University Press (Lenguaje y especie, Madrid, Alianza, 1994).
57 Cfr. Gould, S. J. (1990), The panda’s thumb: more reflexions in natural history, Lon-don, Penguin (El pulgar del panda. Reflexiones sobre historia natural y evolución, Barce-lona, Crítica, 1994); Gould, S. J./Lewontin, R. C., o.c.
58 Chomsky, N. (1968), o.c.

286
© Universidad de Deusto - ISBN 978-84-9830-383-4
El momento de la aparición de la capacidad lingüística en el ser hu-mano se produciría, según Chomsky, en el momento en que, estando ya en posesión de la capacidad mental (teoría de la mente o lenguaje del pensa-miento, según Fodor)59, se dio un reacomodo del cerebro, produciéndose una nueva sistematización del mismo, poniendo en contacto y haciendo trabajar de forma coordinada estructuras cerebrales que antes funciona-ban separadas. Se trata, por tanto, de una ruptura evolutiva, un auténtico salto emergente, que dotó al cerebro de propiedades nuevas que hasta ese momento no poseía, y que se convirtió a partir de entonces en dotación esencial de la especie humana. En concreto, se daría el salto de un proto-lenguaje, en la línea de Bickerton, para pasar a una fase posterior en la que conseguimos poseer nuestro lenguaje actual, con toda su compleji-dad sintáctica, a través de una transición de fase. De este modo, todos los individuos pertenecientes a la especie humana nacemos dotados de una disposición innata para aprender un lenguaje, una Gramática Universalque se halla, en un primer momento, en un estado cero (Eo), para alcanzar posteriormente, con la influencia externa del ambiente cultural correspon-diente, un estado final estable (Ee). Así, el aprendizaje de una lengua es la confluencia de una dotación genética con la influencia externa, sirviendo ésta de detonante y de concreción de la lengua materna cuya competencia concreta consigue el hablante.
Chomsky está echando mano, a la hora de plantear esta hipótesis, de conceptos propios de la ciencias de la complejidad como es el de auto-organi-zación, y que hacen referencia, en palabras de Goodwin, a «la capacidad de generar patrones de forma espontánea, sin instrucciones específicas que dicten lo que hay que hacer, como haría un programa genético. Es-tos sistemas sacan algo de nada. Por “nada” se entiende en este contex-to la ausencia de plan, de proyecto, de instrucciones sobre el patrón que surge»60.
En resumen, la adquisición de la capacidad lingüística, aunque a poste-riori se inscribe en el proceso evolutivo, dotando a sus poseedores de unas enormes ventajas selectivas y sirviendo de catalizador de otros muchos as-pectos fisiológicos y culturales, no tiene por qué entenderse desde la óptica de la aplicación del efecto Baldwin, tal y como lo entienden los evolucio-nistas gradualistas. Resulta más plausible la tesis de Chomsky, aunque se trata de un planteamiento que tiene que probarse científicamente.
59 Fodor, J. (1975), The language of thought, New York, Thomas Y. Crowell (El len-guaje del pensamiento, Madrid, Alianza, 1984).
60 Goodwin, B. (1994), How the leopard changed its sopots. The evolution of com-plexity, New York, Charles Schribner’s Son (Las manchas del leopardo. La evolución de la complejidad, Barcelona, Tusquets, 1998).

287
© Universidad de Deusto - ISBN 978-84-9830-383-4
Tras el breve recorrido que acabamos de hacer de la mano de las dife-rentes teorías sobre el origen y especificidad del lenguaje humano, se pue-de concluir que el lenguaje constituye uno de los elementos que permiten defender la singularidad y la diferencia cualitativa y ontológica del ser humano. El lenguaje humano se diferencia cualitativamente de las capaci-dades comunicativas de los demás animales tanto desde el punto de vista fonético, como también semántico, sintáctico y pragmático. La habilidad de conjuntar la capacidad cognitiva cerebral con los sonidos de la larin-ge representa un rasgo tardío en el proceso evolutivo, puesto que sólo el ser humano la posee. La diferencia entre las habilidades comunicativas de ciertos animales y el lenguaje humano se puede advertir, como lo hemos ya indicado, echando mano de la gran diferencia existente entre el nivel sintáctico a que pueden llegar determinados chimpancés muy entrenados (un cierto proto-lenguaje, como máximo, apelando a Bickerton) y la com-pleja capacidad recursiva de la sintaxis propia de las lenguas humanas.
Esto nos lleva a la conclusión de que la relación entre la especie humana y el resto de las especies animales es una conjunción dialéctica entre conti-nuidad y ruptura. La especie humana es una especie más del mundo de la biosfera, y, por tanto, procede de las especies anteriores a través de un proce-so gradual de complejificación, en el que han participado los mismos meca-nismos que en la aparición del resto de las especies vivas (genética y presión selectiva del entorno ambiental). Pero también se han dado en este mismo proceso evolutivo saltos y rupturas, que propiciaron la aparición y emer-gencia de especies nuevas, debido al surgimiento de mutaciones genéticas (cladogénesis). Las teorías emergentistas advierten, frente a las tesis gradua-listas, que el proceso evolutivo está lleno de casos en los que una progresiva sucesión de cambios gradualistas dan lugar, en un momento determinado, a un salto cualitativo (transición de fase) que genera una nueva estructuración de la realidad, una nueva sistematización. En la naturaleza advertimos múlti-ples situaciones de este tipo. Es el caso de cambios de estado en una realidad física (paso del estado líquido al gaseoso en el agua, a los 100º C), o el salto emergente de la vida desde la materia no viva, o la aparición de cada una de las especies vivas61. Es la tesis de los emergentismos sistémicos de cara a la explicación de la emergencia de la mente humana62.
Ese mismo salto emergente se habría producido en el ser humano, al haber atravesado las dos fronteras de la hominización: el paso de 24 pa-
61 Cfr. Luisi, Pier Luigi, La vida emergente. De los orígenes químicos a la biología sin-tética, Barcelona, Busquets, 2010.
62 Beorlegui, C. (2006), «Los emergentismos sistémicos: un modelo fructífero para el problema mente-cuerpo», Pensamiento, 62 (2006), n.º 234, 391-439; Id., Emergentismos,en vías de publicación en Pensamiento, 2010.

288
© Universidad de Deusto - ISBN 978-84-9830-383-4
res de cromosomas a 23 pares, y el proceso acelerado de la cerebraliza-ción (en poco tiempo, el cerebro del chimpancé, de 400 cm3, se convertirá en 630 cm3 en el homo habilis, y entre 1200 y 1800 en el hombre actual, homo sapiens sapiens). Este salto cualitativo del cerebro, dotado de tan extraordinarias capacidades conductuales, es el que nos permite defen-der la diferencia cualitativa en lo ontológico y conductual entre hombres y demás animales. Y en este desarrollo evolutivo, con sus diversas fases de reacomodo de sus 100 mil millones de neuronas en la estrecha cavidad craneana, es como se habría producido, según Chomsky, el salto cualita-tivo de la aparición de la capacidad innata del lenguaje en la especie hu-mana. Y con el lenguaje, se habría complejificado de modo también ex-traordinario su capacidad congnitiva y su autoconciencia, tal y como las poseemos en la actualidad los seres humanos.
Por tanto, si reduciéndonos a la dimensión puramente fáctica, esto es, al conjunto de datos científicos que sobre la especie humana nos apor-tan las diferentes disciplinas científicas (y que se remiten a mostrarnos el funcionamiento de la realidad humana), podríamos tener tal vez razones para defender la idea de que entre hombres y animales no hay más que una diferencia cuantitativa, advirtiendo, por el contrario, la ruptura emer-gentista que supone la aparición de la mente humana, con sus extraordi-narias capacidades (lenguaje articulado, pensamiento abstracto, libertad y capacidad ética, complejidad de convivencia social, apertura a la pre-gunta por el sentido de la realidad y por su fundamento último, etc.), te-nemos argumentos más que suficientes para defender con total plausibi-lidad la tesis de la diferencia cualitativa. A partir de diferencias más bien pequeñas, en el terreno genético y morfológico, la naturaleza (el creyen-te podrá ver en ello la presencia de Dios como fundamento de todo) ha generado una especie, la humana, que se sitúa, tanto en el terreno de lo ontológico como conductual y ético, en una galaxia cualitativamente su-perior. Pero esto no supone concluir de esta tesis humanista y antropo-céntrica posturas anti-ecológicas o no respetuosas con las tesis de los que propugnan un mayor respeto hacia las condiciones de vida de los gran-des simios y del conjunto de los animales, en general63. Ahora bien, otra cosa muy distinta sería defender los derechos de los animales (en sentido estricto), y atribuir a ciertas especies de simios la categoría de persona,sólo atribuible a los individuos dotados de autoconciencia, autonomía, libertad y capacidad ética. Y estos rasgos son, a todas luces, patrimonio exclusivo de la especie humana.
63 Beorlegui, C. (2005b), «Ética animal e idea de persona», en Guibert, J. M./Orella, J. L. (eds.), Preguntas éticas en cuestiones disputadas hoy, San Sebastián, Universidad de Deusto, 317-335.

289
© Universidad de Deusto - ISBN 978-84-9830-383-4
La cuestión que queda pendiente es en qué momento de la aparición del género Homo aparecería el lenguaje. Algunos consideran al homo ha-bilis como poseedor de la capacidad de hablar, en la medida en que do-minaba ya unas ciertas formas de cultura y tecnología, aunque todavía en grado muy rudimentario. Para construir las herramientas que utilizaba y poder vivir en sociedades de cierta complejidad como eran las suyas, se supone que tendría que poseer un cierto tipo de lenguaje. Con más razón habría que atribuirle el lenguaje al homo erectus, capaz de construir herramientas y de vivir en sociedades aún más complejas. Dereck Bicker-ton considera probable que el homo erectus poseyera ya un cierto proto-lenguaje64. De la misma opinión es Alan Walker, uno de los descubridores de restos de homo ergaster, coetáneo del homo erectus. Según Walker, el homo ergaster no podría hablar como nosotros, puesto que no tendría to-davía el control sutil de la respiración necesaria para hablar, dado el tipo de columna vertebral que poseía el homo ergaster65. Hay otros autores, como el psicólogo M. Corbalis, que defienden la posibilidad de que el homo erectus pudiera hablar con un lenguaje de gestos muy refinado, en la medida en que, según él, el origen del lenguaje no está en la producción de sonidos sino en el uso de gesticulaciones y expresiones faciales66.
Más fuertes son las evidencias sobre la capacidad de hablar del homoheidelbergensis, si nos basamos en los restos de esta especie hallados por los investigadores españoles de Atapuerca. Las características de su con-ducto auditivo y de su oído medio muestran su sensibilidad auditiva en un nivel fundamental para poder entender el habla, algo de lo que carecen los chimpancés67. Según estas teorías, el lenguaje humano tendría más de me-dio millón de años de antigüedad.
Pero otras investigaciones más meticulosas retrasan la aparición del lenguaje a una época entre los 150.000 y 50.000 años, momento en que surgen y viven los primeros homo sapiens sapiens. Es muy probable que también poseyera esa capacidad el hombre de Neandertal, puesto que en yacimientos israelíes se habrían encontrado restos fosilizados del hueso hioides (situado en la base de la lengua), y una laringe, pertenecientes a un ejemplar de la especie neandertal68.
64 Cfr. Bickerton, D., Language and species, Chicago, University of Chicago Press, 1990; Id., Language and human behaviour, London, Taylor and Francis, 1998.
65 Cfr. Walker, A./Shipman, P., The wisdom of bones: in search of human origins, Lon-dres, Weidenfeld and Nicolson, 1996.
66 Cfr. Corballis, M. C., From hand to mouth: the origins of language, Princeton, Prince-ton University Press, 2002.
67 Cfr. Arsuaga, J. L./Martínez, I., La especie elegida, Madrid, Temas de Hoy, 1998, cap. 16.
68 Cfr. ibídem, pp. 314-318.

290
© Universidad de Deusto - ISBN 978-84-9830-383-4
Hay quien ha defendido que la razón más convincente de la desapari-ción de los neandertales ante el impulso de los cromañones fue el no tener todavía la capacidad de hablar. Pero ya hemos visto que las evidencias fó-siles apuntan a un origen más antiguo del lenguaje. De todos modos, a pe-sar de estos resultados, la mayoría de los especialistas se inclinan a pensar que el leguaje sólo se desarrolló de forma completa con el Homo sapiens sapiens69. Los esfuerzos en aportar bases empíricas al origen del lenguaje se orientan tanto hacia el estudio del cerebro y las diversas partes cerebra-les encargadas del control del mismo (áreas de Broca y de Wernicke)70,como, en segundo lugar, hacia las transformaciones laríngeas y del apa-rato respiratorio71, y, por último, la raíz genética del habla, supuestamen-te encontrada en algunos tramos de nuestro genotipo, como el gen foxp2,según el biólogo molecular alemán Wolfgang Enard. Según él, aunque ese gen ya se halla presente en los chimpancés, en el ser humano se habría dado una mutación del mismo, razón del prendimiento de la chispa del lenguaje entre los humanos, suceso que se habría producido en los últi-mos 200.000 años72.
Como puede verse, estamos muy lejos de resolver muchos de los enigmas y misterios que rodean al extraordinario fenómeno del lenguaje, quedando todavía muchas dudas a cerca del momento en que surge esta capacidad, así como el conjunto de elementos que lo hicieron posible. Queda también por resolver la cuestión de si el lenguaje fue un efecto del proceso evolutivo, o un efecto secundario de la acomodación de la masa encefálica en la cavidad craneana, como defiende Chomsky. Lo que resul-ta totalmente evidente es que, sea una u otra la tesis verdadera, la posesión del lenguaje dotó a la especie humana de una capacidad de supervivencia y de comunicación extraordinarias: nos dotó de capacidad de interactuar entre nosotros, de tomar conciencia de nosotros mismos, y de transmitir a las generaciones siguientes la rica herencia cultural acumulada durante siglos. El lenguaje completó y multiplicó de forma considerable las capa-cidades de poseer teoría de la mente, la posibilidad de ponerse en el lugar del interlocutor, permitiéndonos de ese modo compadecernos del otro, aunque también mentirle y manipularle maquiavélicamente.
69 Para un estudio del desarrollo del lenguaje en las diversas especies humanas, cfr. Ar-suaga, J. L./Martínez, La especie elegida, o.c., cap. 16; Ayala, Fr. J./Cela Conde, C., Sende-ros de la evolución humana, Madrid, Alianza, 2001, cap. 10; Carbonell, E./Sala, R., Plane-ta humano, o.c., 11; Kenneally, Chr., La primera palabra, o.c.
70 Cfr. Buskes, Cht., o.c., 201-202.71 Cfr. Lieberman, Ph., Toward an evolutionary biology of language, Cambridge,
Mass., Belknap Press Of Harvard University Press, 2006; Laitman, J. T., «El origen del lenguaje articulado», Mundo Científico, 1986, n.º 64, 1182-1191.
72 Cfr. Buskes, Chr., o.c., 211.

291
© Universidad de Deusto - ISBN 978-84-9830-383-4
2.4. La creación y uso de herramientas y armas
Hasta hace no muchas décadas, se consideraba que las herramien-tas eran uno de los distintivos de la especie humana. De hecho, cuando L. Leaky en 1964 denominó a la primera especie humana como homo ha-bilis, lo hizo entendiendo que la habilidad de construir y usar herramientas era su característica principal y determinante, como expresión de un nivel intelectual que no poseían las especies anteriores. Pero también en esta supuesta característica humana se han producido muchos cambios de opi-nión. El criterio que ha utilizado normalmente la paleoantropología para diferenciar los fósiles humanos de los de nuestros antepasados más cerca-nos, ha sido precisamente la atribución a los supuestos fósiles humanos la autoría de construcción y uso de herramientas inteligentes. Es la razón de considerar humano al homo habilis, como ya vimos en el capítulo segun-do. Así, se consideraba a los humanos como los únicos constructores de herramientas. Como mucho, a un cierto grupo de animales más inteligen-tes se les consideraba tools users (usuarios de herramientas), mientras que se reservaba a los humanos la posibilidad de ser tools makers (constructo-res de herramientas).
La transformación de un objeto para convertirlo en herramienta lo más adecuada a la finalidad para la que se construye, se ha solido consi-derar un patrimonio exclusivo de la especie humana. Y no es de extrañar, puesto que para ello se necesita poseer un proyecto teórico, en el que se prevé la estructura de la herramienta con sus diferentes cualidades y capa-cidades, con vistas a conseguir el fin para el que se construye. Sólo quien posee una mente potente puede tener este tipo de estados mentales, capa-ces de anticiparse al tiempo, concebir propósitos, establecer relaciones causales entre objetos físicos y entre la mano y las herramientas, caracte-rísticas que hemos analizado al hablar de la capacidad de imaginación, de analizar el pasado y de prever el futuro. Esto es una de las características específicas del ser humano, al menos en su dimensión desarrollada y per-fecta, pero eso no quita que se les pueda atribuir también, en cierto grado, a determinados individuos de algunas especies animales73.
Se suele citar el caso de los pájaros carboneros en Inglaterra, capa-ces de aprender a abrir botellas de leche depositadas ante las puertas de
73 Cfr. Fossey, D., Gorilas en la niebla, Barcelona, Salvat, 1985; Goodall, J., «Nest Building Behaviour in the Free Ranging Chimpanzees», Annuary of the New York Academy of Sciences, 1962, 102, pp. 455-467; Id, En la senda del hombre, Barcelona, Salvat, 1988; Id., A través de la ventana, Barcelona, Salvar, 1994; Sabater Pi, J., Etología de la vivien-da humana. De los nidos de gorilas y chimpancés a la viviencia humana, Barcelona, La-bor, 1985.

292
© Universidad de Deusto - ISBN 978-84-9830-383-4
las viviendas humanas. Se trata de un comportamiento que es una mezcla de innato y de aprendido. Otros ejemplos más complejos se refieren a los chimpancés que son capaces de utilizar determinados objetos, una rama de árbol, cortarla para que tome la dimensión adecuada, quitarle la cor-teza, impregnarla de saliva para que resulte pegajosa e introducirla en un hormiguero con objeto de cazar hormigas de las que alimentarse.
Pero, al igual que hemos ido viendo, y tendremos ocasión de compro-bar también más adelante, en relación a otros aspectos supuestamente es-pecíficos de la especie humana (desde la conciencia, el lenguaje, el fuego y demás elementos integrantes de la cultura), las diferencias entre los hu-manos y el resto de las especies animales se va acortando y, en opinión de algunos, diluyendo y desapareciendo, a medida de que avanzan las inves-tigaciones sobre los comportamientos animales. En el caso de las herra-mientas, es fundamental considerarlas como una mediación entre la men-te y el entorno físico, configurada estructuralmente con una lógica bien precisa, continuación y ampliación de la relación del propio cuerpo con el entorno74. Esa mediación comienza, por tanto, por el propio cuerpo y con-tinúa en todo el amplísimo abanico de las herramientas. Por eso, a la hora de clasificar los diversos tipos de herramientas, se suele distinguir entre las denominadas herramientas biológicas (partes del propio cuerpo), ade-más de las herramientas consistentes en diversos objetos sin apenas trans-formar, y, por último, las herramientas propiamente dichas, consistentes en objetos transformados y trabajados.
Es evidente que la mayoría de los animales, por no decir todos, uti-lizan partes de su cuerpo como herramientas manipuladoras y transfor-madoras de su entorno, y para interactuar con sus congéneres, desde los picos hasta las pezuñas, garras, patas excavadores, etc. Son lo que hemos denominado herramientas biológicas.
Pero también hay muchas especies animales que utilizan diversos ob-jetos externos a su cuerpo como herramientas para aumentar su eficacia transformadora. Es el caso de ramas, palos, guijarros, esponjas, etc. En lí-neas generales, parece que aunque la mayoría de los primates pueden ma-nejar algunas herramientas, estos animales actúan con un comportamiento muy poco flexible; es decir, excepto en casos muy especiales, parece que utilizan esas herramientas de un modo innato, puesto que no escogen, ni modifican, ni mejoran la herramienta. Y tampoco parece que tienen capa-cidad para prever la posibilidad de utilizar estas mismas herramientas en el futuro, en situaciones similares. De ahí que una vez que las han utiliza-
74 Cfr. Leroi-Gourhan, A., El hombre y la materia (vol. I. Evolución y técnica; vol. II. Medio y técnica), Madrid, Taurus, 1988/1989; Breton, S., «La technique entre nature et cul-ture», Esprit, 1997, n.º 11, 159-180; Bourg, D., L’Homme artífice, París, Gallimard, 1996.

293
© Universidad de Deusto - ISBN 978-84-9830-383-4
do, las abandonan. Es decir, no tiene apenas capacidad para racionalizar la acción que realizan, sino que el recurso a la herramienta es una carac-terística de la especie, al estilo de como posee y utiliza otros caracteres biológicos. Las herramientas de este estilo son bastante frecuentes entre los artrópodos y en ciertos mamíferos; y está muy extendida entre los primates75.
Pero en el caso de los chimpancés, las investigaciones de las últimas décadas están aportando evidencias de un uso y fabricación de herramien-tas bastante sofisticado. Como indican Chr. Boesch y H. Boesch-Acher-mann, dos de los más importantes investigadores de la utilización de herramientas por los chimpancés, «hay que reconocer que, aparte de los chimpancés, la rareza de las utilizaciones de herramientas en los simios parece confirmar la idea preconcebida de que la herramienta es exclusi-va de los humanos»76. Estos mismos investigadores indican que, después de muchos años de estudios, «únicamente algunos babuinos, macacos u orangutanes en cautividad han sido observados utilizando, y de manera repetida, herramientas, aunque esto se debe principalmente a estimulacio-nes presentadas por los científicos a animales cautivos»77. Esto parece un tanto sorprendente, por cuanto en muchas publicaciones se indica que tan-to los gorilas como los orangutanes habían sido capaces de manifestar, en ámbitos de laboratorio, suficiente capacidad intelectual como para utilizar símbolos y manejar el lenguaje gestual de los sordomudos78. Pero parece lógico deducir que, viviendo en libertad en medio de la naturaleza, la pre-sión ambiental la resuelven sin necesidad de echar mano de herramientas. Además, investigaciones hechas en cautividad parece que indican que el nivel intelectual de estos animales es insuficiente para hacerse una repre-sentación de los rasgos físicos de las herramientas que tendrían que utili-zar, ni tampoco tiene la inteligencia suficiente como para imitar a un con-génere más hábil79.
En el caso de los chimpancés, como hemos ya apuntado, la situación es diferente. Son muy conocidas las investigaciones del psicólogo ale-mán, W. Köhler, en las primeras décadas del siglo XX en Canarias, sobre
75 Cfr. Beck, B. B., Animal tool behaviour: The use and manufacture of tools by animals,Garland Publishing, 1980; Visalberghi, E., «Los simios y la herramienta: ¿una cuestión de inteligencia?», Mundo Científico, 1989, n.º 97, 1264-1267.
76 «Los chimpancés y la herramienta», Mundo Científico, 1991, n.º 116, 842-849; 842. 77 Ibídem, 842. 78 Cfr. Chevalier-Skolnikoff, S./Poirier, F. E, Primate biosocial development: Biological,
social and ecological determinants, Garland Press, 1977; Chevalier-Skolnikoff, S., Behavioraland Brain Sciences, 12, 561, 1988; Savage-Rumbaugh et alii, «Linguistically mediated tool use ad exchange by chimpanzees», Behavioral and Brain Sciences, 1978, 1 (4), 539-545.
79 Cfr. Visalberghi, E., o.c.

294
© Universidad de Deusto - ISBN 978-84-9830-383-4
los comportamientos complejos e inteligentes de los chimpancés. En la década de los sesenta, Jane Goodall estudió los comportamientos de los chimpancés en libertad en Tanzania y descubría la fabricación de útiles para cazar termitas80. Poco después, el japonés T. Nishida confirmó las investigaciones de J. Goodall en un territorio cercano también en Tan-zania81. Y algunos años más tarde, Chr. Boesch y H. Boesch-Achermann realizaron investigaciones similares con chimpancés en Costa de Marfil (Parque Nacional de Tai), descubriendo una gran variedad de utilización de herramientas, especialmente herramientas de madera para el consu-mo de alimentos. Los Boesch llegaron a descubrir hasta veintiocho técni-cas diferentes y cuatro tipos de movimientos de la mano para manipular la herramienta82.
Construir una herramienta, aunque sea sencilla, presupone tener una cierta idea del resultado final y del fin para el que se la quiere utilizar. Esto parecería que sobrepasa la capacidad mental de los primates, cosa que sí se ha comprobado en el caso de los simios capuchinos83. Pero no es el caso de los chimpancés, que parecen poseer una cierta planificación a la hora de construir sus útiles. Esto hace que algunos investigadores, a la vista de las distintas técnicas que utilizan los diversos chimpancés, quie-ran ver en esas variaciones diferencias culturales, en la medida en que también se ha experimentado entre ellos intentos de transmitir a sus des-cendientes la técnica de la fabricación y uso de estas herramientas.
No cabe duda de que los chimpancés, en la medida de que constituyen la especie más cercana a los humanos, poseen atisbos de inteligencia y de capacidad técnica de fabricar herramientas que anticipan las capacidades humanas, pero sus habilidades tienen también muchas limitaciones, simi-lares a las que pueden tener los niños humanos de pocos años. De todas maneras, como indican E. Carbonell y R. Sala, en estos casos de manipu-lación de objetos para convertirlos en herramientas parece haber un límite, puesto que «los objetos naturales pueden, como mucho, sufrir modifica-ciones de medida pero jamás de forma, lo que significa que sus propieda-des naturales no son alteradas»84. Otros chimpancés han sido capaces de
80 Cfr. Goodall, J., In the shadow of man, Boston, Houghton Mifflin, 1971 (En la senda del hombre, Barcelona, Salvat, 1988); Id., The Chimpanzees of Gombe: Patterns of Behavior,Cambridge, Mass., Belknap Press, Harvard University Press., 1986; Through a Window, Bos-ton, Houghton Mifflin (A través de la ventana, Barcelona, Salvat, 1994).
81 Nishida, T., The social group of chimpanzees in the Mahale Mountains, Primates,1968, 9, 167-224.
82 Cfr. Boesch, Chr./Boesch-Achermann, H., Los chimpancés y la herramienta, o.c., 845-846.
83 Cfr. Visalberghi, E., «Los simios y la herramienta: ¿una cuestión de inteligencia?», o.c.84 Planeta humano, o.c., p. 30.

295
© Universidad de Deusto - ISBN 978-84-9830-383-4
recolectar un puñado de hojas secas para convertirlas en una esponja que, empapada de agua, les sirve para saciar la sed. Estas innovaciones crea-tivas suelen después contagiarse a otros integrantes de la especie, siendo normalmente los jóvenes los que innovan y contagian a sus compañeros, resultando difícil que los de más edad aprendan o acepten innovaciones que provienen de los más jóvenes. Igualmente, es frecuente ver a monos de la sabana seleccionar ramas secas como armas para ejercer acciones posteriores de caza.
También parece cierto que los chimpancés, al igual que otras especies, innovan menos en situación salvaje que en cautividad, dado que mientras vive en su habitat natural recibe menor presión del medio que en situación de cautividad y sometido a las pruebas de los investigadores. El salvaje se enfrenta siempre a los mismos problemas, resultándole suficiente para acomodarse a su entorno con sus estilos de vida innatos. En cambio, en cautividad recibe una presión mayor, presión que le lleva a aprender com-portamientos mucho más complejos. Pero aun así, su estructura compor-tamental tiene sus límites que no puede superar. Su facultad de previsión sigue siendo limitada. Como ya hemos indicado, tras alcanzar su objetivo con la ayuda de una cierta herramienta, la abandona. Si se encuentra en próximas ocasiones en situaciones similares, volverá a utilizar o a compo-ner herramientas similares. Vive, pues, al día.
En resumen, aunque entre el comportamiento animal y el humano, en relación a las herramientas, hay diferencias cualitativas claras, los com-portamientos de los primates se acercan a los humanos en una serie de rasgos dignos de señalarse, de la mano de E. Carbonell y R. Sala: «la utili-zación de objetos con transformaciones mínimas, que indica la existencia de habilidades mentales claras, aunque rudimentarias; la forma mediante la cual estas habilidades se aprenden por ensayo-error»85.
Haciendo un balance comparativo entre el comportamiento de los chimpancés y el de los humanos en relación a las herramientas, Carbonell y Sala ven al menos dos diferencias importantes. La primera se refiere a que, en los chimpancés, «la transformación de la morfología original del objeto es mínima y siempre se realiza sin la intervención de otros obje-tos construidos o adaptados: se rompen bastones con las manos o con los pies, se impregnan tallos de hierba con saliva… Finalmente, el uso al que se destina es siempre inmediatamente posterior a una adaptación o elabo-ración muy somera. Por tanto, la cadena elaboración-uso es muy corta en el espacio y en el tiempo. El lapso temporal transcurrido entre la aparición del estímulo, la elaboración y el uso es breve, sin que haya lugar en él
85 Ibídem, p. 30.

296
© Universidad de Deusto - ISBN 978-84-9830-383-4
para la previsión. La materia prima es la propia del medio en el que se de-sarrolla la vida de los chimpancés y bonobos y de aquél en el que se usa-rá el nuevo objeto, sin que exista transporte. No se encadenarán acciones conducentes a una gran transformación»86.
La segunda gran diferencia se refiere al hecho de que «el instrumen-tal y la capacidad operativa de los primates no humanos es unidireccional: no son capaces de presentar una diversidad de respuestas “técnicas” a un mismo estímulo que consistan en la producción de objetos diferentes o de cadenas de elaboración distintas, sino que siempre realizan de la misma manera los procesos en los que usan objetos. Por el contrario, los huma-nos podemos ofrecer respuestas diferentes a un mismo estímulo y vicever-sa, somos capaces de generalizar y de dar una misma respuesta a proble-mas diferentes. Nuestra capacidad es multidireccional»87.
Por tanto, en la comparación entre los comportamientos de los prima-tes y de los humanos, volvemos a encontrarnos con la misma dialéctica de continuidad y ruptura con la que nos vamos encontrando siempre. Las es-pecies animales más cercanas a la humana poseen atisbos de nuestra ma-nera de comportarnos y nos muestran la continuidad de los humanos con el proceso evolutivo, a la vez que el salto cualitativo del que procedemos. La capacidad de idear y construir herramientas que posee la especie hu-mana, aprendida y ejercida a lo largo de los dos millones y medio de exis-tencia, es de una complejidad extraordinaria88.
En primer lugar, implica la posesión de una mente enormemente compleja, capaz de prever todo tipo de situaciones y problemas, con las respuestas y soluciones de lo más variadas y acertadas. Esa capacidad anticipadora es la que le permite prever y preparar herramientas cuya uti-lización no es inmediata, sino que será posible utilizar en situaciones fu-turas. Así, antes de sentir la necesidad de las mismas, es capaz de prever su uso. Y tras utilizarlas en la realización de un determinado trabajo, las guarda para situaciones similares en el futuro. Esto le permite ir reali-zando mejoras progresivas, o inventar mejoras cualitativas. Este ha sido el procedimiento normal y continuo que ha utilizado el ser humano en el desarrollo de herramientas, utensilios y armas a lo largo de la historia, en sus diferentes culturas, llegando a dominar la naturaleza de la forma que conocemos, y representando el mayor depredador y transformador del en-torno ecológico, con peligro incluso de romper los equilibrios ecológicos y poner en riesgo la supervivencia del planeta entero y su propia supervi-vencia con él.
86 Planeta humano, o.c., 61.87 Ibídem, 61-62. 88 Cfr. Gubern, Roman, El simio informatizado, Madrid, FUNDESCO, 1987.

297
© Universidad de Deusto - ISBN 978-84-9830-383-4
Esta capacidad creativa la va realizando a través de sucesivos ejerci-cios de ensayo-error, acumulando experiencias y mejoras, y transmitién-dolas a las generaciones siguientes. Es la cultura, en su sentido más am-plio, como patrimonio de la especie, y como bagaje que se va agrandando en cada generación y transmitiendo a las siguientes.
Según lo ya indicado, la pregunta por el origen de las primeras herra-mientas en la especie humana no puede obviar el hecho de la existencia de proto-herramientas en las especies animales cercanas a la nuestra. Pero la definición del homo habilis como primera especie humana desde el cri-terio de utilización de herramientas inteligentes, especiales, conlleva la distinción o la ruptura de complejidad entre sus herramientas y las de los simios y también la de los australopitecos. De todas formas, ya vimos en su momento las discusiones sobre la evidencia científica o no de hacer tal separación, habiendo autores que se inclinan a situar al homo habilis entre los australopitecos (australopitecus habilis), aunque la mayoría de los pa-leoantropólogos siguen considerando tal especie como humana.
En el problema del origen de las herramientas tenemos que tener en cuenta dos aspectos complementarios: el relacionado con lo que podría-mos denominar las condiciones de posibilidad del uso y construcción de las herramientas inteligentes, y la pregunta sobre el momento histórico de su aparición, según los datos y pruebas empíricas manejados por la pa-leoantropología. En relación a las condiciones de posibilidad del uso de herramientas inteligentes, nos tenemos que referir a la capacidad mental e intelectual necesaria para idear la construcción de herramientas, así como al bipedismo y a la posesión de unas manos hábiles capaces de manejar-las. Respecto a la capacidad mental y cerebral, poco podemos decir. Ya veíamos que los investigadores de los chimpancés les atribuían una capa-cidad intelectual suficiente como manejar útiles complejos, mientras que no veían tal capacidad en los capuchinos, monos de América Central y del Sur. El extraordinario aumento del cerebro realizado en los homínidos y en el homo habilis, permitió que se produjera un salto cualitativo a la hora de la construcción y uso de herramientas más complejas, auténticamente humanas. En este sentido, las herramientas vienen a estar dotadas, en re-lación con la mente, de una estructura lógica y funcional semejante al len-guaje, puesto que, al igual que las palabras, representan un instrumento de mediación entre la mente y la realidad, al mismo tiempo que un medio de transformación del entorno que nos rodea, tanto humano como ecológico. Por eso que, como decíamos al hablar del primer humano, el homo habi-lis, la autoría de herramientas inteligentes es lo que adjudica la condición de humano a los cerebros de los sujetos creadores de tales herramientas.
Por otro lado, la aparición de las herramientas fue propiciada por la adopción del bipedismo y la liberación de la mano para manipular ob-

298
© Universidad de Deusto - ISBN 978-84-9830-383-4
jetos. Estos dos elementos que aparecieron ya con los australopitecos,dotaron a estas nuevas especies de unas ventajas adaptativas muy im-portantes, como ya vimos en capítulos anteriores, pero al mismo tiempo les permitieron fabricar utensilios y herramientas cada vez más eficaces e inteligentes. De este modo, se produce una fundamental y extraordi-naria colaboración entre el cerebro y la mano. «Entre las adquisiciones básicas de los homínidos humanos, la prensión de precisión puede ser la más importante y con mayor potencial transformador de todas: el poder formar una pinza con el pulgar nos permite controlar hasta extremos im-pensables el objeto que estamos manipulando, a la vez que nos permite asirlo cualquiera que sea su superficie. La combinación de las manos y el cerebro revoluciona el comportamiento de los homínidos de tal forma que, por primera vez, biología y cultura empiezan a compenetrarse»89.La complicidad entre el cerebro y la mano inteligente y hábil es lo que hizo que los humanos pudieran construir, utilizar y perfeccionar las más diversas y complejas herramientas, pudiendo de este modo defenderse y defender al grupo de cualquier ataque externo, defender el territorio donde vivían, explorar nuevos nichos ecológicos y ámbitos donde po-der cazar y buscar alimentos, competir con ventaja con depredadores por el mismo nicho ecológico, competir también con especies de homínidos menos dotados, así como con otros grupos de humanos por el territorio y las fuentes de supervivencia.
En relación a la segunda cuestión, está la pregunta sobre el origen de las primeras herramientas. Ya hemos dicho en repetidas ocasiones que la tesis tradicional consistía en situar ese momento originario en el homo ha-bilis. Pero ese convencimiento está de algún modo superado, siendo pro-bable que las primeras herramientas fueran construidas y utilizadas por algunas especies de australopitecos. Hay quienes defienden que el primer constructor de herramientas sería el australopithecus garhi, apoyándo-se en el descubrimiento, por parte del equipo de Tim White (Berkeley) y de B. Asfaw (Museo de Addis Abeba), en el valle del Awash (Etiopía), de unas marcas de talla en huesos de antílope. Según estos paleoantropólo-gos, estas marcas serían indicios claros de ser este homínido el primer co-medor de carne de la historia de la evolución humana. El tipo de marcas encontradas en dichos huesos no tienen la forma de «U», como sería lógi-co en caso de pertenecer a huellas de dientes de depredadores, sino en for-ma de «V», huella específica de las herramientas. El problema está en que no se han encontrado restos fósiles de tales herramientas. Aunque, dado que esa zona geográfica no contiene materia prima para hacer herramien-
89 Carbonell, E./Sala, R., Planeta humano, o.c., 63.

299
© Universidad de Deusto - ISBN 978-84-9830-383-4
ta, podría ser defendible que estos homínidos utilizaran y llevaran consigo herramientas que las habían construido en otras zonas cercanas90.
Hay otros autores que atribuyen las primeras herramientas a otras fami-lias de asutralopitecos o a ciertos paránthropos, contemporáneos del homo habilis. Si esta primera especie humana apareció hace 2,4 millones de años, según datan los fósiles más antiguos descubiertos, resultan muy significa-tivos los hallazgos de herramientas de fechas algo anteriores a la aparición del homo habilis (al menos, 2,6 millones de años), realizados en 1974 por Helène Roche y Jean-Jacques Tiercelin (del CNRS francés)91. En esa misma región, algunos años después, el equipo de Sileshi Sumaw (Universidad de Rutgers) ha encontrado más de un millar de objetos tallados92.
La cuestión es saber quiénes fueron los sujetos de estos útiles, que marcan la frontera entre lo humano y lo pre-humano. Ya hemos dicho que los chimpancés son capaces de utilizar piedras a modo de yunque y mar-tillo para romper cáscaras, pero no son capaces de tallar estos instrumen-tos de manera espontánea. «Por consiguiente, la talla sería específica de la humanidad»93. A pesar de ello, hay estudiosos, como Randall Susman, que han demostrado que la estructura de los australopitecos estaría dotada para poder manipular y fabricar los útiles de modo casi tan hábil como los hu-manos. Si a esto se une que las fechas de los primeros útiles son anteriores al homo habilis, nos encontraríamos con fuertes pruebas para atribuir esas herramientas a los australopitecos94. Claro que se podría defender la tesis de que el homo habilis es anterior a la fecha que se le atribuye (2,4 mi-llones de años), aunque no se han encontrado fósiles de tanta antigüedad. Pero si se encontraran, ellos podrían ser los autores de tales herramientas.
Otra tesis es atribuirla a los australopitecos, considerando la posibi-lidad de que entre las fechas de 2,5 millones y 1 millón de años convi-vieron en África varias especies de homínidos. Si las herramientas más antiguas datan de hace 2,6 millones de años, puede perfectamente haber sido construidas por dos especies de homínidos que vivieron entre hace 2,5 y 2,0 millones de años: el Paranthropus, una forma de australopitecorobusto, y el Australopithecus africanus, sucesores ambos del Australo-pithecus afarensis grácil95. Esta tesis se apoyaría en el hecho de que mu-
90 Cfr. Asfaw, B./White, T. et al., «Australopithecus garhi: A new Species of Early Ho-minid from Ethiopia», Science, 1999, 284, 629-635; Carbonell, E./Sala, R., o.c., 64-65.
91 Roche, H./Tiercelin, J. J., Acad. Sc. Paris, serie D, 1871, 1977.92 Sumaw, S. et al., Nature, 1997, 385, 333. 93 Kempf, Heervé, «El nacimiento de los útiles. Peleontólogos, etólogos y psicólogos
unen sus esfuerzos», Mundo Científico, 1997, 523. 94 Susman, R./McGraw, S. A., «¿Quién fabricó las primeras herramientas?», Mundo
Científico, 1995, n.º 159, 682-683. 95 Cfr. ibídem, p. 682.

300
© Universidad de Deusto - ISBN 978-84-9830-383-4
chas de las herramientas encontradas se hallan en los mismos yacimien-tos en que se han encontrado restos de Paranthropus robustus. Algunos análisis comparativos de la estructura de la mano humana y la de los si-mios, realizadas en concreto por John Napier, experto en anatomía fun-cional de los primates96, le han llevado a concluir que los primates tenían manos bastante adecuadas para manipular ciertas herramientas. Otros es-tudiosos se muestran más bien escépticos respecto a estas conclusiones tan optimistas.
En cambio, otros estudiosos, como Glynn Isaac, defienden que esas primeras herramientas podrían muy bien ser atribuidas a los Parantho-pos, debido precisamente a la estructura ósea y muscular de sus manos, muy parecidas a las del homo habilis, y debido también a su mayor ca-pacidad craneana97. De esta misma opinión son también R. L. Susman y W. S. McGraw, defendiendo las siguientes conclusiones: «El Homo no era el único que podía fabricar herramientas hace unos 2 millones de años. No es tampoco el primer artesano de la Historia, ya que las herra-mientas más antiguas son anteriores a su aparición. Las manos de los primeros homínidos, como los Australopithecus afarensis no eran aptas para la fabricación y uso de herramientas. En cambio, hace unos 2,5 mi-llones de años, los primeros australopitecos robustos las fabricaban en cantidad. Y para explicar la desaparición del Patanthropus, sustituido hace un millón de años por homínidos con el cerebro más desarrollado, habrá que buscar explicaciones más sutiles que su capacidad craneana limitada y las imposibilidad de fabricarse herramientas»98.
En resumen, los datos que en la actualidad se tiene sobre la antigüe-dad y la atribución de las primeras herramientas, permiten poder defen-derse hasta cuatro hipótesis, como indica Henry Gee: la primera postura las atribuye a los Paranthopus (boisei); la segunda considera como po-sible que ni el homo habilis ni el Panranthropus (boisei) fueran los auto-res, sino una nueva especie anterior, de la que todavía no tenemos rastro corpóreo fosilizado; una tercera tesis se inclina porque los fabricantes pri-meros de herramientas fueran o el Paranthropus (boisei) o algún otro ho-mínido; y una cuarta se inclinaría por atribuirlas exclusivamente al homo habilis99. H. Gee descarta esta última postura y apuesta por atribuirlas
96 Cfr. Napier, J., Scientific American, 1962, 205, 2. 97 Cfr. Isaac, G.I., en Wendorf, F./Close, A. (eds.), Advances in World Archaeology, vol. 3,
Academic Press, 1984; Susman, R./McGraw, S. A., o.c., 683.98 Susman, R./McGraw, S. A., o.c., 683.99 Cfr. Gee, H., «¿Es la fabricación de utensilios un rasgo exclusivo de los humanos?»,
El País, 12 de mayo de 1999, p. 43; Dufour, Jean-Paul, «Las primeras herramientas pudie-ron ser obras de homínidos», El País, 12 de mayo de 1999, p. 43.

301
© Universidad de Deusto - ISBN 978-84-9830-383-4
a algunas de las especies de australopitecos, de tal modo que considera que a la pregunta: «¿hay humanidad implícita en la fabricación de herra-mientas?», hay que responder que no. Ocurre, por tanto, en el ámbito de las herramientas lo que hemos indicado también sobre el bipedismo: no es un comportamiento orientado y originado finalísticamente desde un cerebro humano, sino fruto de condicionantes evolutivos anteriores a la aparición de los humanos. Y de este modo, en opinión de H. Gee, este descentramiento antropológico de las herramientas constituiría un nue-vo golpe al narcisismo y antropocentrismo tradicional. De todos modos, entendemos que estas afirmaciones no dejan de ser una prueba más de la tendencia actual más dominante de socavar a toda costa la hasta ahora in-contestable visión antropocéntrica del ser humano. De todos modos, en-tendemos que se puede defender la primera autoría de herramientas por parte de los homínidos, sin necesidad de desprenderse de la visión antro-pocéntrica tradicional, entendida adecuadamente como estamos defen-diendo en este libro.
Pero lo que sí aparece claro es que en el ámbito de la construcción y uso de las herramientas, al igual que en lo referente a la aparición de la conciencia, del lenguaje y de la capacidad simbolizadora (y de otros as-pectos culturales que veremos más adelante), la frontera entre lo humano y lo pre-humano es cada más vez más difícil de establecer, y no existe una línea totalmente precisa y definida. Pero lo que está claro es que, aunque tengamos que atribuir a los homínidos la construcción y el uso de herra-mientas inteligentes, las herramientas de los humanos representan un salto de complejidad indiscutible, iniciándose con el homo habilis una comple-ja y rica historia de perfeccionamiento de las herramientas y útiles.
Los estudiosos de las herramientas humanas suelen dividir en tres eta-pas la historia de su uso y construcción, atendiendo a su complejidad. La primera etapa, denominada Modo I, o también etapa Olduvaiense (en re-ferencia a Olduvai, lugar de Tanzania donde fueron hallados restos fósiles por primera vez), se extiende entre los 2,5 y 1,6 millones de años. «Estos artefactos, caracterizados por su escasa configuración y por la búsqueda de filos diédricos, se obtenían a través de métodos de talla aleatorios y con secuencias arquetípicas elementales. La variabilidad de rocas utilizadas es amplia y está sujeta sólo a su disponibilidad, resistencia y buena esfolia-ción. Estos instrumentos serían desechados poco después de su utilización, dada la facilidad y rapidez de la fabricación de nuevos útiles»100.
100 Carbonell, E. et alii, «Instrumentos de los homínidos en Atapuerca», Mundo Cien-tífico, 1997, n.º 175, 54-59; 55; Carbonell, E./Sala, R., Planeta humano, o.c., pp. 117-125; Arsuaga, J. L./Martínez, I., La especie elegida, Madrid, Temas de Hoy, 1998, cap, 7, 129-149;Ayala, Fco. J./Cela Conde, C. J., Senderos de la evolución humana, Madrid, Alianza, 2001,

302
© Universidad de Deusto - ISBN 978-84-9830-383-4
El Modo II, o Achelense (por haberse hallado en Saint Acheul, Francia), abarca la época que va desde 1,5 millones de años hasta los 300.000 años. La herramienta más típica de esta época es el hacha deno-minada bifaz, junto con hendedores, picos e instrumentos de gran formato, que manifiestan un alto grado de elaboración. Algunos incluso quieren ver en ellos los primeros síntomas de una visión estética de la herramienta, de-bido a la evidente constitución simétrica de dichos artefactos, sobre todo las bifaces, así como incluso una dimensión simbólica101. De todos modos, a pesar del avance que supone este tipo de herramientas respecto a las de la fase anterior, resulta impresionante ver la lentitud de tales avances, en la medida en que la amplitud de los períodos en los que se sigue constru-yendo de un modo repetitivo e invariable estas herramientas, nos hace ver la dificultad enorme que supone introducir novedades y avances en esta tecnología tan elemental, realizada por humanos dotados de una capaci-dad craneana todavía lejana a los niveles del homo sapiens. Es igualmente digno de notarse «el retraso aparente entre la aparición de estos modos en África y su presencia en el resto de los continentes»102, representando este dato una prueba más del origen africano de los seres humanos.
La etapa denominada Modo III, o Musteriense (Le Moustier, en la Dordoña francesa), se extiende desde los 300.000 años hasta la actualidad. Lo característico de esta etapa es la gran diversidad y complejidad de las herramientas que se fabrican, produciéndose ya en esta época de forma es-tandarizada. Entre los años 200.000 y los 40.000, «se extiende como una mancha de aceite la denominada “Middle Stone Age”, o Modo 3 africano, compuesta por utensilios de pequeño formato altamente elaborado»103.
No hemos hecho referencia hasta ahora a la cuestión de cuáles fueron los primeros utensilios que se fabricaron: los de piedra, o los de huesos, de madera, o de otro tipo de materia. Una primera impresión intuitiva defendería la preeminencia de la madera, por ser más fácil de manejar. Pero, en la medida en que no es fácil que deje restos fósiles, estas hipó-tesis solían ser rechazadas. Con todo, poco a poco se han ido encontran-do cada vez más restos evidentes de la antigüedad de las herramientas de madera. Así, Lawrence Leaky encontró en Koobi Fora evidencias de ob-jetos líticos, con una antigüedad de 1,5 millones de años, que trabajaron
372-374; Clarke, Roberet, El nacimiento del hombre, Barcelona, Ediciones Juan Granica, 1983, cap. 2.º; Vinagre, Fco./Vinagre Arias, A., Materiales y tecnología en las primeras ci-vilizaciones, Calamonte (Badajoz), Edit. Filarias, 2002.
101 Cfr. Arsuaga, J. L., «Un objeto simbólico», El País, 8 de enero de 2003, p. 26; Rive-ra, A, «Un hacha hallada en Atapuerca indica que ya había ritos funerarios hace 400.000 años»,El País, 8 de enero de 2003, p. 26.
102 Ibídem, p. 56. 103 Ibídem, p. 56.

303
© Universidad de Deusto - ISBN 978-84-9830-383-4
sobre madera104. Descubrimientos posteriores confirman la existencia desde antiguo de diversas herramientas de madera, utilizadas para diver-sas funciones: desde cavar para arrancar raíces y tubérculos, hasta como lanzas, jabalinas y otro tipo de armas.
Las pruebas definitivas aparecieron en diversos yacimientos ale-manes en la década de los noventa del siglo pasado. En un yacimiento situado en la localidad de Schöningen se han encontrado desde obje-tos lanzadores y otro tipo de armas para acciones de proximidad, hasta mangos para útiles cortantes. Incluso se halló un cráneo de caballo con un orificio causado por un arma puntiaguda, complemento de los ob-jetos lanzadores encontrados también en el yacimiento. Las fechas de tales herramientas son de más de 400.000 años, época del Pleistoceno Medio, y utilizados seguramente por el homo heidelbergensis, anterior al hombre de Neandertal105. También en Atapuerca, en los niveles del Pleistoceno Inferior, donde se han hallado restos del homo antecesor, con más de 800.000 años de antigüedad, se han encontrado trabajos de la madera, esto es, un tipo de trabajo técnico destinado a la producción de este tipo de instrumentos106.
La utilización de la madera como herramientas no sólo se daba en el ámbito de las armas, sino también para usos domésticos, aunque todavía «no hemos podido averiguar qué instrumentos se usaban cotidianamente en las actividades domésticas relacionadas con la alimentación, la prepa-ración de alimentos o su conservación»107. De todos modos, aunque no haya todavía evidencias de todo esto en épocas muy tempranas a la apari-ción de los primeros humanos, entre otras razones por la dificultad de que la madera fosilice con la abundancia que lo ha podido hacer la piedra, no resulta difícil imaginar que la madera fuera uno de los primeros recursos utilizados como herramientas, en la medida en que abundaba en el entorno de los primeros humanos, nacidos en los espacios entre el bosque y la sa-bana. De ahí que, como indican Carbonell y Sala, «no parece descamina-do que la madera fuera el primer material usado por nuestros antepasados, igual que vemos entre los chimpancés. La historia de los descubrimientos más recientes así lo confirma. La madera es, con toda probabilidad, la ma-teria prima más ancestral de la humanización, aunque las posibilidades que se desprenden de este hecho sólo hayan sido aceptadas de manera amplia muy recientemente»108.
104 Cfr. Carbonell, E./Sala, R., Planeta humano, o.c., 71 y ss. 105 Cfr. ibídem, pp. 72 y ss. 106 Cfr. ibídem, 76.107 Ibídem, 78. 108 Ibídem, 78-79.

304
© Universidad de Deusto - ISBN 978-84-9830-383-4
2.5. Dominio y uso del fuego
El dominio y el uso del fuego es una de los logros más importantes que ha conseguido la especie humana y que más han contribuido en su proceso de humanización. Y es, al mismo tiempo, una característica ex-clusivamente humana, en la medida en que sólo la especie humana ha conseguido perderle el miedo, controlarlo, producirlo y sacar de él todas sus virtualidades. Como en el resto de las características culturales que es-tamos analizando, vamos a describir sus rasgos y ventajas, aportando los datos de la paleoantropología sobre los primeros momentos en que los hu-manos dominaron el fuego.
Las ventajas que el ser humano ha conseguido del fuego son múlti-ples, alcanzadas no desde el principio, sino a través de un largo proceso de contacto permanente con el fuego109 La primera de las ventajas es la de iluminar. Antes de dominar el fuego, los ciclos culturales que marcaban el día y la noche era el sol. A partir de que pudo tener hogueras encendidas, pudo alargar el trabajo del día, reposar al final del mismo, reunirse toda la comunidad alrededor de él, intercambiar las diversas experiencias y co-nocimientos unos a otros, y transmitirlas a las generaciones más jóvenes, contar historias y experimentar los gozos de la vida grupal y social. No hace falta ser muy imaginativo para poder afirmar que el fuego ha sido testigo de la mayor parte de los elementos que han ido conformando los procesos de humanización a lo largo del millón y medio de años aproxi-madamente en que los humanos lo dominaron. El largo camino hacia la socialización de los humanos y de conformación de sus cosmovisiones, así como el proceso de simbolización se fue realizando en gran medida al-rededor del fuego.
Una segunda ventaja o función del fuego fue producir calor, regulan-do su intensidad añadiéndole nuevos trozos de madera para que siguiera vivo y con suficiente temperatura. De este modo, el fuego permitió al gé-nero humano habitar regiones del planeta que sin él no podría haber soña-do habitar110. Se supone, como veremos luego, que fue el homo erectus, la primera especie humana que sale de África, la que conseguiría dominar el fuego y lo utilizaría para adentrarse en regiones frías, imposibles de ha-
109 Cfr. Carbonell, E./Sala, R., Planeta humano, o.c., 129 y ss; Clarke, Robert, El naci-miento del hombre, o.c., cap. 5; Lorite Mena, José, El animal paradójico. Fundamentos de Antropología filosófica, Madrid, Alianza, 1982, pp. 375-380.
110 Cfr. Arsuaga, J. L., Los aborígenes. La alimentación en la evolución humana, Bar-celona, RBA Libros, 2002, pp. 115 y ss. El autor considera que quizás el ser humano se adentró en regiones frías antes de dominar el fuego, a base de vestidos de pieles. Esa sería quizás una de las razones de que no se hayan encontrado restos de fuego en épocas anterio-res a los 500.000 años.

305
© Universidad de Deusto - ISBN 978-84-9830-383-4
bitar con los recursos naturales, y su propia desnudez, que le permitieron hasta ese momento al homo habilis habitar sin grandes dificultades las tierras cálidas del centro de África. La función de preservar al ser huma-no del frío sería realizada antes por el inicio de construir vestidos, que se supone tuvo que ser una técnica anterior a dominar el fuego. Fue capaz de adentrarse en zonas frías, o incluso fue sorprendido, en zonas habitual-mente no tan frías, por momentos de glaciaciones y de bajadas fuertes de temperatura. Eran momentos en que todavía no había aprendido a domi-nar y aprovechar las virtualidades del fuego, pero sí pudo hacerse los pri-meros vestidos a base de pieles de los animales que cazaba. Al principio, como es lógico, utilizaba simplemente las pieles alrededor del cuerpo, atadas alrededor de la cintura, puesto que no había inventado todavía las agujas de hueso o utensilios similares111.
Al mismo tiempo que calentaba, el fuego hacía huir a los animales, de modo que resultaba un instrumento útil colocar en un rellano una bue-na hoguera encendida, dormir alrededor de ella, o ponerla en la puerta de alguna gruta, con la seguridad de que ningún animal peligroso se acer-caría a molestar, en la medida en que ninguna especie animal parece que muestra evidencias de haberle perdido el miedo al fuego, como ocurre con los humanos, conocedores de su naturaleza, sus leyes y su modo de dominarlo.
Unida a la seguridad frente a los depredadores, una función y ventaja que proporcionó el fuego a los hombres es permitirles dormir y descan-sar con seguridad. No todos los estudiosos suelen hacer hincapié en esta ventaja, que por otro lado tampoco arroja datos empíricamente muy ve-rificables. Sigue siendo una incógnita desentrañar los interrogantes que nos presentan los sueños. No sabemos por qué ni para qué soñamos, aun-que existen múltiples hipótesis para contestar esos interrogantes. Si tra-dicionalmente se han dado respuestas de tipo místico y mítico, pensando que los sueños eran el modo que tenían los dioses de ponerse en con-tacto con los humanos, o el modo como éstos se ponían en contacto con el mundo del más allá, tal y como se advierte en la Biblia y en el resto de los libros sagrados de las religiones o en los libros de las epopeyas lite-rarias más diversas, en épocas más recientes se ha tratado de acercarse a sus misterios de modo más científico. Así, el psicoanálisis lo estudia des-de dentro, analizando la naturaleza simbólica de los sueños, proponiendo hipótesis que relacionaban sus contenidos con el inconsciente o con los deseos y temores más hondos de sus protagonistas. Pero ese camino de
111 Para un acercamiento a los orígenes de los vestidos primitivos, cfr. Carbonell, E./Sala, R., Aún no somos humanos. Propuestas de humanización para el tercer milenio, Bar-celona, Península, 2002, cap. 9.º.

306
© Universidad de Deusto - ISBN 978-84-9830-383-4
reflexión, sin menospreciarlo ni considerar que es totalmente inservible, está expuesto a múltiples teorías arbitrarias y poco contrastables con una base más objetiva y razonable. Por eso, la línea más científica de acceso a nuestra mente y a los sueños ha ido extendiéndose cada vez más, a través de técnicas de medición de las diferentes ondas que los pacientes o volun-tarios emiten cuando se les aplican electrodos conectados a monitores que reflejan los electroencefalogramas112.
Se advierte con estos procedimientos científicos que el sujeto huma-no, cuando cierra los ojos emite un tipo de ondas, denominadas ondas alfau ondas de la meditación. A medida de que va entrando en un sueño más profundo, el ser humano emite otro tipo de ondas, que reflejan las diferen-tes fases por las que va a travesando su período de sueño113. Se advierte que tanto el hombre como los animales atraviesan durante la noche por núcleos de hasta cinco fases de sueño o más, núcleos que se repiten varias veces a lo largo de una noche, llegando en cada una de esas fases a un pe-ríodo especial y profundo, denominado sueño REM (movimiento rápido de ojos, en ingles: Rapid Eyes Mouvement)114, momento en el que al pa-recer se producen los sueños. Cuando se comparan los comportamientos durante el sueño y la vigilia entre los humanos y los primates, se advier-ten, como no podía ser menos, parecidos y diferencias115. El animal tiene que estar en permanente vigilancia ante la posibilidad del ataque de los depredadores, no pudiéndose entregar nunca de un modo seguro a la pla-cidez del sueño. En los animales, no aparecen las ondas alfa o de medi-tación, sino que poseen ondas transitorias, como reacción de sorpresa ante la sospecha del peligro y el mantenimiento de una permanente situación de alerta, y que producen reacciones musculares y neurovegetativas para poner al organismo en disposición de huir o de repeler cualquier agresión. En el ser humano también se dan estas ondas y reacciones consecuentes, pero puede racionalizar intelectualmente la situación de peligro. Por eso es capaz de controlar los ruidos sospechosos, controlarlos y ver cuándo carecen de importancia. De ahí que en el ser humano estas ondas desapa-recen enseguida, al comprobar que no hay peligro. Y situaciones similares posteriores ya no producen tal reacción de alerta. Se trata del fenómeno
112 Cfr. Zimmer, Dieter E., Dormir y soñar. La vida nocturna de nuestras vidas, Barce-lona, Salvat, 1986.
113 Cfr. ibídem. 114 Este fenómeno fue descubierto por casualidad por un doctorando norteamericano,
cuando investigaba los comportamientos de los voluntarios sometidos a pruebas para in-vestigar el comportamiento de los humanos al dormir, en una universidad norteamericana de California.
115 Desportes, J. P., «El comportamiento de vigilancia en los animales», Mundo Cientí-fico, 1987, n.º 67, 238-245.

307
© Universidad de Deusto - ISBN 978-84-9830-383-4
denominado de habituación. Esta habituación también se puede produ-cir en los chimpancés, pero es mucho más lenta. En otros animales infe-riores, casi no consiguen habituarse a determinados ruidos y perderles el miedo. Y en los animales más elementales, la vigilia es constante, porque el momento del sueño es cuando un ser vivo está en situación de máxima indefensión. Sólo el ser humano posee instrumentos y organización social suficiente para entregarse sin miedo al descanso reparador y al sueño, ha-biendo sido el fuego uno de los elementos que contribuyó a poder adquirir tal tranquilidad.
Cuando se alcanza la fase de sueño REM (también denominado sueñoparadójico), es cuando se da una completa relajación y una caída fuerte del tono muscular, al mismo tiempo que una disminución de la excitabi-lidad en los receptores sensoriales (ruptura con el medio y descenso de vigilancia). Lo curioso es que este desenganche muscular y sensorial con-trasta con la actividad cerebral, que parece ligada a un tratamiento de la información adquirida durante la vigilia, y a su memorización. Así, pues, parece existir una relación entre el proceso de memorización e integración de las experiencias, por un lado, y el sueño paradójico, por otro. A mayor duración y cantidad de fases de sueño paradójico, se produciría más me-morización. Si esto fuera cierto, en la medida en que el ser humano puede permitirse dormir con más tranquilidad y acceder a numerosas fases de sue-ño paradójico, poseería un mecanismo de memorización y de incremento de la capacidad cerebral e intelectual, que no poseen los demás animales. Por tanto, el uso y dominio del fuego, con las consecuencias tan impor-tantes que estamos viendo, pudo ser también uno de los factores clave en el proceso de hominización y humanización.
Además, el ser humano es capaz, como ya lo dijimos al referirnos al surgimiento y desarrollo de la conciencia, de soñar no sólo cuando duer-me, sino también estando despierto, simulando en sus sueños situaciones imaginarias e irreales para ejercitarse en esas historias y situaciones hi-potéticas. El ser humano es el primate que puede reflexionar profunda-mente, porque es capaz de aislarse suficientemente del entorno exterior, y liberar totalmente sus funciones psíquicas. Y esto es posible porque tiene los medios para preservar su seguridad. El hombre, al franquear el pasode reflexión, se encamina a través de un ciclo sin fin de acciones y retro-acciones entre su mente y el entorno y los demás humanos. Así, las reali-zaciones culturales desarrollan la inteligencia, y ésta aumenta los logros culturales.
Nos hemos extendido quizás demasiado en esta función, un ventaja aportada a los humanos por el uso del fuego, pero está justificado por la importancia que ha tenido en la propia conformación de nuestro modo de ser. Aunque hay que reconocer también que muchas de estas afirmaciones

308
© Universidad de Deusto - ISBN 978-84-9830-383-4
son meras hipótesis, que tienen que ser validadas y demostradas con el avance de las investigaciones.
Una función importantísima del fuego consistió en que su uso per-mitió la cocción de los alimentos, con lo cual consiguió enriquecer de un modo considerable su dieta y asimilar mejor los nutrientes que comía116.Ya vimos, al hablar del fenómeno de la cerebralización, las tesis de que la ingesta de proteínas animales le permitió aumentar de modo acelerado su capacidad cerebral. Los alimentos vegetales son más difíciles de asimilar y, por tanto, necesitan un proceso digestivo más largo. Ahorrando en apa-rato digestivo el ser humano pudo invertir en cerebro. Pero con el fuego, pudo cocer los alimentos vegetales, asimilar también los nutrientes nece-sarios, y ahorrar tiempo de hacer la digestión en situación de indefensión y duermevela, momento que, como acabamos de ver, es una fase muy pe-ligrosa de la existencia. Por tanto, también desde este punto de vista, des-de haber permitido al ser humano convertirse en cocinero, el fuego ha re-presentado un elemento clave en el proceso de humanización117. El fuego mejoró la dieta del ser humano y le permitió fortalecerse para subsistir y desarrollar la caza y el resto de conductas encaminadas a la supervivencia.
Los cambios de los hábitos alimenticios tuvieron también repercu-siones tanto en la musculación como en las dimensiones de los dientes y maxilares, «causada, entre otros factores, por la adaptación del cráneo y de las cavidades bucal y nasal al crecimiento cerebral… El estudio de la den-tición de los individuos de las especies heidelbergensis y neanderthalen-sis ha permitido ver que entre los primeros los alimentos consumidos eran muy abrasivos y entre los segundos, muy poco. Esto se ha podido saber por el desgaste y las estrías de los dientes, indicando un cambio notable en la dieta por la introducción del fuego, que reduce la capacidad abrasiva de los vegetales»118.
También sirvió el fuego a los primeros seres humanos para conservar los alimentos, permitiéndole guardar para tiempos de mayor escasez re-servas sobrantes en momentos determinados de buena cosecha o caza. El ser humano parece que aprendió enseguida a darse cuenta de que los
116 Cfr. Arsuaga, J. L., Los aborígenes. La alimentación en la evolución humana, Bar-celona, RBA Libros, 2002, pp. 125 y ss. Arsuaga, de todos modos, hace interesantes re-flexiones sobre el hecho de que no siempre la cocción de los alimentos los hace más enriquecidos al alimentarnos. Cfr. también Cordón, Faustino, Cocinar hizo al hombre, Bar-celona, Tusquets, 1980 (2009, 7.ª ed.).
117 Cfr. Cordón, F., o.c.; Arranz, M., «¿Un nuevo mecanismo de especiación? En diá-logo con Faustino Cordón», Cuadernos Salmantinos de Filosofía, X (1983), 283-292; Ca-rrasco i Pons, Silvia, Antropología i alimentació. Una proposta per a l’estudi de la cultura alimentaria, Barcelona, Universidad Autónoma, 1992.
118 Carbonell, E./Sala, R., o.c., 130.

309
© Universidad de Deusto - ISBN 978-84-9830-383-4
alimentos cocinados duraban más que si se mantenían crudos. Los estu-diosos descubren que los primeros humanos conservaban los alimentos bien sea ahumados o en salazón. A medida en que los humanos se organi-zaban en grupos numerosos, tenían más capacidad para conseguir piezas grandes y abundantes, lo que suponía que tenían capacidad para conseguir alimentos en excedencia, y necesitó hallar técnicas para la conservación de excedentes, tecnología que fue perfeccionando a medida que iba avan-zando en el dominio del fuego.
Hay otra función interesante desempeñada por el fuego y consiste en la limpieza del espacio. Esta función, que como indican Carbonell y Sala está poco documentada en la bibliografía, nos indica y muestra «una complejidad del comportamiento, de la estructuración del espacio y del concepto de espacio doméstico que por sí sola conduce a conclusiones obvias»119. Se ha visto en muchos yacimientos humanos que el fuego ser-vía para echar en él todo tipo de desechos, sirviendo así para la limpieza del espacio doméstico, arrojando en el fuego todos los restos no consu-mibles de comidas, sobre todo los huesos. De ese modo, el fuego va sir-viendo al ser humano para configurar un tipo nuevo de relación con el entorno, sobre todo en el ámbito doméstico, convirtiéndose la hoguera, el fuego controlado, el centro del espacio familiar y de la convivencia120.
También le sirvió el fuego al hombre para manipular otras materias primas y hacerlas maleables para otros fines. Se ve claro, en primer lu-gar, en relación con la madera: ya vimos que se han encontrado restos que muestran la técnica de quemar las puntas para hacerlas más duras y resis-tentes al convertirlas en lanzas, jabalinas o flechas. Pero también utilizaba el fuego para manipular los objetos de sílex: las piedras expuestas al calor son más maleables y se podían tallar u obtener mejores herramientas de un modo más fácil. Más adelante fue capaz de modelar con el fuego me-tales como el bronce, el cobre o el hierro, para hacer armas, herramientas y útiles culturales de todo tipo, así como metales preciosos como el oro y la plata121.
Un aspecto importante de la utilización del fuego se refiere a la ma-nipulación con él de la madera para la construcción de arcos de tejo para que la madera adquiriera más flexibilidad y tomara la forma deseada que le permitiera convertirse en un arma mortal, manejada por guerreros ex-pertos. Pero no sólo servía el fuego para la moldeabilidad de la madera en la fabricación de armas, sino que también servía para hacer múltiples objetos de cestería de mimbres, y otro tipo de objetos de mimbres y mate-
119 Ibídem, 131. 120 Cfr. Lorite Meta, J., o.c., pp. 377-379. 121 Cfr. Cfr. Carbonell, E./Sala, R., o.c., pp. 131-133.

310
© Universidad de Deusto - ISBN 978-84-9830-383-4
riales semejantes a los que se les aplicaba una capa de betún, material que sólo ha podido ser moldeado gracias al fuego122.
La pregunta que nos queda pendiente por resolver es cuándo comen-zó el ser humano a dominar el fuego y a saber provocarlo. En la medida en que no tenemos pruebas para responder estas cuestiones desde los pri-meros momentos, tenemos que echar mano de la imaginación. Es muy razonable pensar que los primeros humanos, al igual que el resto de los animales, tuvieran pavor al fuego y a sus consecuencias. Tendría expe-riencia de terribles incendios provocados por una tormenta, por erupcio-nes volcánicas, por desprendimiento de una ladera pedregosa de montaña, o por cualquier otro fenómeno natural. Con el tiempo, le iría perdiendo el miedo, llegando a acercarse y a mantenerlo encendido por períodos más o menos largos123. Así, al principio lo podría utilizar y mantener, sacándole el mayor rendimiento posible, pero no sabría todavía producirlo. El poder dominar el fuego, aunque fuera de modo precario, concedería a la tribu que lo tuviera un enorme poder, así como también sería objeto de envidias y de deseos de robárselo por los grupos humanos rivales. De ahí nacería también la necesidad perentoria de mantener encendido el fuego, dedicán-dose a ello, según parece, las mujeres124, los niños y los ancianos, mien-tras los guerreros fuertes y las mujeres capaces se dedicaban a la caza, los primeros, y a otras labores de agricultura y ganadería incipientes, las se-gundas. Así mismo, es comprensible que se proyectase sobre el fuego po-deres y cualidades divinas, fundándose en varias culturas, como entre los romanos, el grupo institucional de las vestales, encargadas de mantener el fuego sagrado, como símbolo ya en este caso de la custodia para garanti-zar la bendición y la presencia de los dioses a favor del pueblo. Todas es-tas vicisitudes y proyecciones alrededor del fuego, se han popularizado a través de la película En busca del fuego (1981), de Jean-Jacques Annaud, película que sigue la novela del mismo nombre de J. H. Rosny125.
Posteriormente, los humanos consiguieron inventar diversas técni-cas para encender fuego, sin necesidad de estar pendientes de su con-
122 Cfr. ibídem, p. 133.123 Cfr. Collina-Girad, Jacques, Le feu avant les allumettes, París, Editorial de la mai-
son des sciences de l’homme, 1998. 124 Cfr. Clarke, R., El nacimiento del hombre, Barcelona, Ediciones Juan Granica,
1983, pp. 95-98. 125 J. H. Rosny es el seudónimo común que utilizaron para escribir sus numerosas no-
veles sobre temas prehistóricos los hermanos de nacionalidad belga Joseph-Henry-Honoré Boex (1856-1940) y Seraphin-Justin-Francois Boex (1859-1948). Para un mejor conoci-miento de los dos autores y su obra: véase la edición reciente de En busca del fuego, El león de las cavernas, y Vamireh, Madrid, Valdemar, 2001, con la Presentación de Alfredo Lara López.

311
© Universidad de Deusto - ISBN 978-84-9830-383-4
servación. En los primeros momentos históricos los humanos tenían dos técnicas de producción del fuego: frotando un vara de madera con una plataforma del mismo material. La rotación repetida de la vara sobre la plataforma va produciendo calor, hasta desprender una pequeña com-bustión en la plataforma y el desprendimiento de algunas chispas, que tie-nen que ser aprovechadas añadiendo algunos materiales muy inflamables, como excrementos secos, setas o yescas. Este modo de producir fuego te-nía múltiples variables al realizarlo, pudiéndose perfeccionar el sistema para hacerlo más fácil y eficaz. Un segundo sistema o técnica consistía en aprovechar las propiedades de ciertos minerales productores de chispas al rozarse, como la pirita, rozándolo con una roca dura de cuarzo o sílex, que tenía que utilizarse al lado de materiales muy inflamables, al estilo de la técnica anterior126.
Veamos ahora qué nos dicen los hallazgos paleontológicos sobre las fechas más antiguas de aparición del fuego en los yacimientos utilizados por los humanos. Los datos más razonables apuntan a que no se utilizó el fuego en fechas anteriores al millón de años. Se supone que lo utilizaría por primera vez el homo erectus, como instrumento imprescindible para, tras su salida de África, poder adentrase en regiones frías. Pero no debió ser en los primeros años de este primer éxodo humano, porque no se han encontrado restos de fuego en yacimientos antiguos como los de Ubeidiya (Israel) y Dmanisi (Georgia)127, ni tampoco acompañando a los restos de homo erectus hallados en Java. Tampoco en Atapuerca se han encontrado restos de utilización de fuego en ninguno de sus yacimientos128. Durante bastante tiempo se consideró un yacimiento clave para afirmar el uso del fuego por el homo erectus el de Zhoukoudian, situado cerca de Pekín (Chi-na), y descubierto a principios del s. XX. Sus descubridores hacían referen-cia a hogueras con vestigios de huesos quemados y varias capas de ceni-zas. Pero este yacimiento sufrió las consecuencias de la guerra entre chinos y japoneses, antes de la segunda guerra mundial, y han desaparecido los rastros arqueológicos allí encontrados. Estudios posteriores, con metodo-logía más rigurosa, llevados a cabo por Ofer Bar-Josef, de la Universidad de Harvard, han llegado a la conclusión de que, dada la situación actual del yacimiento, no se pueden constatar ningún tipo de vestigio de cenizas ni de carbones vegetales. Pero sí han podido encontrarse restos de huesos que-mados, consecuencia tanto de fuegos naturales como producidos por ho-
126 Cfr. Carbonell, E./Sala, R., o.c., 134-135; Clarke, R., o.c., 96-97. 127 Sobre este importante yacimiento de Georgia, cfr. Agustí, Jordi/Lordkipanidze, Da-
vid, Del Turkana al Cáucaso. La evolución de los primeros pobladores de Europa, Barce-lona, RBA Libros, 2005.
128 Cfr. Carbonell, E./Sala, R., o.c., p. 136.

312
© Universidad de Deusto - ISBN 978-84-9830-383-4
gueras humanas. Pero, dada la situación del yacimiento, no parecen ser por desgracia pruebas concluyentes. Así, como indican Carbonell y Sala, «el yacimiento que presentaba el registro más antiguo de fuego antrópico que-da descartado como tal y, de paso, ha caído un mito. Es posible que existie-ran hogueras en Zhoukoudian, pero en cualquier caso no eran tan extensas como se creía. A pesar de todo, nuestra experiencia en Abric Romaní nos impulsa a no descartar definitivamente su existencia»129.
En Europa se encuentran los primeros restos de fuego en fechas an-teriores a los 400.000 años, en diversos yacimientos de Francia (Menez Dregan, Terra Amata) y Alemania (Bilzingsleben). De este modo, se su-pone que tanto el homo erectus, como sobre todo el homo heidelbergensis,hacían uso de hogueras con diversos fines domésticos, durante la época central del Pleistoceno Medio. Al principio el sistema de fuego era el de-nominado fuego plano, consistente en encenderlo directamente sobre el suelo, sin estructura externa que lo delimite. Más tarde, se utilizan fuegos con una cubeta formada en la tierra. De este modo, se evita que cualquier eventualidad no controlada pueda extender el fuego, aparte de que se con-sigue de ese modo aprovecharlo mejor, consiguiéndose temperaturas más altas. Y un tercer tipo de fuegos es el de hogueras bordeadas por piedras y materiales de todo tipo, y con diverso tipo de complejidad. Las formas más elementales son las que se encuentran en pocos sitios y en épocas más antiguas. Y cien mil años después, el uso del fuego se extendió de forma casi generalizada. Como dijimos anteriormente, el paso de usar el fuego a conseguir la técnica de producirlo, se daría en un sitio concreto, guardándolo como un tesoro de vital importancia, y dotándolo incluso de valores míticos y religiosos, aunque posteriormente se extendería rápida-mente por todos los grupos humanos.
En resumen, el fuego parece que fue dominado por los primeros hu-manos (homo erectus) algún tiempo antes de los 400.000 años, pasándose de perderle el miedo a su contagio devastador a saber dominarlo130, utili-zándolo progresivamente para las diversas funciones de las que ya hemos hablado: cocción y conservación de los alimentos, defensa de los depre-dadores, dormir mejor, manipulación de herramientas, realización de di-versos actos de socialización alrededor de las hogueras, desarrollo del len-
129 O.c., 137.130 «El dominio y la posterior utilización de esa cosa ardiente, terrorífica, viva y preca-
ria que es la llama adquirió, seguramente, innumerables tentativas, desarrolladas a lo lar-go de decenas de millares de años. No podemos reconstruirlas. Nunca sabremos cuántos intentos, cuántas esperanzas defraudadas, ni cuántas tragedias causadas por los incendios, fueron necesarios, antes de que el hombre llegara a domesticar el fuego»: Clarke, R., Elnacimiento del hombre, o.c., p. 94.

313
© Universidad de Deusto - ISBN 978-84-9830-383-4
guaje, etc. Por tanto, alrededor del fuego se produjo un espacio domésti-co y cultural de primer orden, escenario privilegiado sin ninguna duda del largo proceso de humanización.
2.6. Los usos alimenticios
Para J. L. Arsuaga, en la historia de la humanidad se han producido dos acontecimientos importantes en lo referente a su forma de alimen-tarse. El primero marca el origen del ser humano, y consiste en «la in-corporación, en cantidad importante, de productos de origen animal a la dieta»131. El segundo se produjo en fecha más reciente, y sucedió cuando «se pasó de extraer el alimento de la naturaleza a producirlo directamente por medio de la agricultura y de la ganadería. Es lo que se conoce como la Revolución Neolítica, y nosotros somos sus hijos»132.
El ser humano es una animal omnívoro, en la medida en que nos ali-mentamos tanto de vegetales y frutas, como de carne y pescado de todo tipo. Pero, como indica Arsuaga, la alimentación carnívora fue clave en el desarrollo de nuestra especie, sobre todo en relación al desarrollo del cerebro. Vimos ya, al hablar del fenómeno de la cerebralización, que la evolución de la alimentación se ha ido dando desde los animales herbívo-ros, pasando por los frugívoros, para desembocar en los carnívoros. Los primeros animales se alimentaban de plantas, teniendo que desarrollar un amplio aparato digestivo para poder asimilarlas, siendo los mamíferos ru-miantes el prototipo de aparato digestivo plenamente desarrollado para estos menesteres. Posteriormente, los primates son las especies animales especializadas en alimentarse de frutas, con la ventaja de tener una die-ta más rica en azúcares, así como representar una alimentación más fácil de asimilar. Para ello necesitaban un tipo de vista más compleja, como es el paso de una visión en blanco y negro a otra que permita percibir los colores, en especial el color de la fruta madura133. Pero lo que permitió a los humanos poseer de forma progresiva y rápida un cerebro más de-sarrollado fue el salto de la dieta herbívora y frugívora a la carnívora. Ese salto fue posible ahorrando en aparato digestivo para invertirlo en neuro-nas cerebrales, como muestran los estudios de Aiello y Séller, ya citados
131 Los aborígenes. La alimentación en la evolución humana, Barcelona, RBA Libros, 2002, p. 13; Cfr. Clarke, R., o.c., p. 56.
132 Ibídem, 14-15. 133 Cfr. Alman, J. M., El cerebro en evolución, Barcelona, Ariel, 2003, cap. 6. «El cere-
bro de los primates»; Jacobs, Gerald H./Natham, Jeremy, «La evolución de la visión de los colores en los primates», Investigación y Ciencia, 2009, n.º 393, 42-49.

314
© Universidad de Deusto - ISBN 978-84-9830-383-4
en su momento. Por tanto, lo que nos hizo inteligentes fue convertirnos en carnívoros, por la necesidad de tener una alimentación rica en proteí-nas animales, y la mayor facilidad en asimilarlas, sin tener que poseer un grande y poderoso aparato digestivo134. El cambio de dieta no sólo reper-cutió en el cambio del estómago a favor del cerebro, sino también en todo el aparato masticador y en la forma de los dientes, que se fueron haciendo más finos, añadido también esto al uso progresivo de las manos y las he-rramientas para cortar alimentos, cosa que los otros anímales carnívoros realizaban con sus potentes mandíbulas y afilados incisivos. La transfor-mación de la dentición y de la forma de la mandíbula y del aparato mas-ticador, ya vimos que había sido uno de los elementos morfológicos más representativos del proceso de hominización.
De todos modos, es importante advertir que la introducción de la dieta carnívora no significó dejar de lado la alimentación de vegetales, de raí-ces, semillas y toda clase de frutas, al mismo tiempo que la alimentación de carne le supuso a los primeros seres humanos un largo y costoso ejer-cicio de aprendizaje de la caza. Deteniéndonos en el primer momento o acontecimiento de la historia humana a los que hacía referencia Arsuaga más arriba, es importante imaginar el paso de la dieta alimenticia de los chimpancés a la de los primeros humanos, pasando por la de las diferen-tes especies de homínidos, los australopitecos. Si los chimpancés se ali-mentan de vegetales y de frutas, es de imaginar que los australopitecosiniciaran una ampliación de su dieta a base de frutos secos, semillas, raí-ces y algo de carne. Los granos secos y las semillas duras poseen una gran riqueza alimenticia, que los australopitecos empezaron a utilizar debido a poseer ya una importante habilidad para manipularlas con las manos, li-beradas tras haber empezado ya a caminar erguidos. Además, su dentición se fue adaptando a este tipo de alimentación, disminuyendo los dientes caninos y aumentando los molares. Lo mismo diríamos de las nueces, al-mendras y otros frutos secos similares, siendo capaces de utilizar piedras para partirlas y apropiarse de su rico contenido alimenticio. Por otro lado, otra fuente de alimentos sería la alimentación con bulbos, tubérculos, ri-zomas y otro tipo de raíces gruesas, que serían capaces de desenterrar con la ayuda de palos elementales usados como herramientas inteligentes135.También se alimentarían con diversos insectos, como termitas y hormigas de todo tipo, al estilo de lo que ya hacen los chimpancés136.
134 Cfr. Arsuaga, J. L./Martínez, I., La especie elegida, Madrid, Temas de Hoy, 1998, cap. 9.º.
135 Cfr. Arsuaga, J. L., Los aborígenes, o.c., pp. 55 y ss. 136 Cfr. Joulian, Frédéric, «Origen del hombre, termitas y chimpancés (¿Eran insectívo-
ros los primeros hombres?)», Mundo Científico, 2001, n.º 230, 14-15.

315
© Universidad de Deusto - ISBN 978-84-9830-383-4
Pero es también muy posible que ya los australopitecos iniciaran un tipo de alimentación a base de grasa animal, comenzando quizás por el tuétano de los huesos largos de animales grandes, tras haber aprendido a romperlos con piedras para extraer su rico contenido alimenticio. E igual-mente, aprenderían a cazar pequeñas presas animales, así como es posible que comenzaran a utilizar el sistema del carroñeo, que con toda probabi-lidad siguieron utilizando también los primeros humanos137. Aunque no tengamos del todo pruebas contundentes de estos comportamientos ali-menticios, son muy probables que existieran, anticipando los de los hu-manos posteriores.
Con la aparición del homo habilis, la alimentación fue similar en parte a lo que hemos indicado de los australopitecos. Tenían la ventaja de po-der vivir en terrenos amplios de sabana, donde podían recoger semillas, frutos secos, raíces, y aprovechar los tuétanos de los huesos de presas ma-yores matadas por los grandes depredadores, así como ejercer el carroñeo, disputándose el acceso a grandes presas matadas por los grandes felinos y dejadas todavía con cantidades notables de carne tras haber saciado su hambre. El carroñeo es un sistema de alimentación en el que concurren muchas especies de animales, y los primeros humanos competirían en el acceso a los restos dejados por los grandes felinos como las hienas, bui-tres y otras especies similares.
También es importante hacer referencia a la primera alimentación bá-sica del ser humano, y de los demás mamíferos: la leche materna. Este es un producto alimenticio básico para los primeros momentos de la exis-tencia, que se prescinde de él cuando la cría se independiza del cuidado directo de la madre. Pero es curioso que la especie humana es la única que sigue bebiendo leche a lo largo de la vida138. Aunque hoy día se dis-cute si es bueno tal hábito, o si tiene el riesgo de generar consecuencias negativas139.
Durante mucho tiempo se dio por sentado que los primeros huma-nos fueron expertos cazadores, que realizaban sus campañas de caza bien organizados y provistos de temibles, aunque todavía elementales, herra-mientas. Pero esta tesis clásica de la caza como el modo primero de pro-
137 Cfr. Arsuaga, J. L., Los aborígenes, o.c., pp. 33 y ss. 138 Cfr. Wade, Nicholas, «La tolerancia a la lactosa en África indica una reciente evolu-
ción de la especie humana», The New York Times (reproducido en El País, 27 de diciembre de 2006, p. 37).
139 Cfr. sobre este problema, y sobre si se ha dado en algunas culturas la mutación que permite asimilar la leche, que en situaciones normales la rechazaríamos, así como sus po-sibles producciones de cánceres: Boto, Angela, «¿Necesitan tomar leche los adultos? Con-troversia sobre el consumo de lácteos en la edad adulta por su posible relación con ciertos tumores», El País, 2004, 21 de diciembre, p. 37.

316
© Universidad de Deusto - ISBN 978-84-9830-383-4
veerse de proteínas animales, propuesta por Washburn y Lancaster, ade-más de la denominada por R. Ardrey la hipótesis del cazador140, se ha re-chazado en la actualidad con diferentes argumentaciones. En primer lugar, esta tesis no ha sido corroborada por el registro fósil141. Además, frente a la visión del hombre primitivo como cazador agresivo y sanguinario en la sabana abierta, las evidencias fósiles muestran más bien un tipo de caza-dor que cooperaba en mayor o menor medida para conseguir los alimen-tos. Parece más razonable pensar que los primeros humanos no pudieron al principio más que utilizar el carroñeo, para más adelante avanzar hacia técnicas de caza mucho más eficaces, dada su capacidad de formar equi-pos de cooperación compleja y disponer progresivamente de herramientas cada vez más eficaces142.
En el habitat de la sabana, los humanos tenían la ventaja de ca-minar erguido y no tener pelo y tener capacidad de sudar, lo que les permitía de forma ventajosa frente a sus competidores de ese nicho ecológico aguantar los calores del día, además de utilizar diversas herra-mientas para poder cavar y desenterrar las raíces y recoger semillas y frutos secos143.
Al principio, el ser humano sería muy poco capaz de cazar más que animales pequeños, y pescar de modo elemental. Pero, al estar dotado de un cerebro de considerable tamaño respecto a chimpancés y austra-lopitecos, pronto fue capaz de construir herramientas de caza, así como de conformar grupos unidos que sumaban sus fuerzas ante sus presas. Desde el principio el ser humano se fue haciendo cazador, llevado por la presión selectiva a necesitar carne para comer y no tener más remedio que unirse con otros de su especie para sumar esfuerzos para cazar pie-zas cada vez mayores y más alimenticias. Además, la caza no trajo sólo ventajas de cara al aporte de alimentos, sino que puede quizás decirse que «fue también la primera actividad humana socializada»144. Se tra-ta de uno de los muchos elementos comportamentales de la especie hu-mana en la que se da un círculo virtuoso, necesitándose para realizarlo una serie de condiciones previas, y enriqueciéndose tales condiciones a medida que se ejercita. En concreto, para poder conformar grupos efica-ces de cazadores, se necesitaba una mente capaz de comunicarse con los
140 Cfr. Ardrey, R., La evolución del hombre: la hipótesis del cazador, Madrid, Alianza, 1978; Cfr. Carbonell, E./Sala, R., o.c., pp. 97-102.
141 Cfr. Blumenschine, R. J./Caballo, J. A., «Carroñeo y evolución humana», Investiga-ción y Ciencia, 1992, 195, 70-77.
142 Cfr. Ayala, Fco. J./Cela Conde, C. J., Senderos de la evolución humana, Madrid, Alianza, 2001, pp. 361 y ss.
143 Cfr. ibídem, pp. 45 y ss. 144 Clarke, R., o.c., p. 56.

317
© Universidad de Deusto - ISBN 978-84-9830-383-4
otros, ponerse en el lugar del otro, designar liderazgos que impusieran conductas comunes y dirigidas, y después un reparto adecuado del bo-tín que evitara conflictos y produjera una mayor cohesión de los compo-nentes del grupo. Así mismo, suponía una evolución de la tecnología de las herramientas y de los sistemas de caza.
A lo largo de los estudios paleoantropológicos, así como de la An-tropología cultural, se ha dado por hecho que en el largo período de tiempo denominado Paleolítico, esto es, desde el origen de los prime-ros humanos hasta unos 10.000 años a. C., momento en que se inicia el gran salto del Neolítico, la forma de vida humana se centraba en el sis-tema de cazadores y recolectores, en el que se daba un reparto de roles entre los varones, cazadores, y las mujeres, recolectoras, y encargadas también de las labores domésticas y el cuidado de los hijos. En este re-parto de tareas, siempre se da por sentado que el papel predominante en el aporte alimenticio era debido al varón, quedando la mujer en un se-gundo lugar. Estudios más recientes, así como el acceso de las mujeres a las investigaciones paleoantropológicas, están derribando estos tópi-cos tan poco fundamentados e inexactos145. El reparto de roles fue un hecho evidente, en función de las diferentes características biológicas, que obligaba a la mujer a estar más dependiente de los hijos y a la ma-yor fuerza física del varón (aunque el dimorfismo sexual en el género humano es mucho menor que en otras especies de primates), pero pare-ce ser que las aportaciones alimenticias de ambos sexos eran similares, si no mayores en el caso de las mujeres. Los varones podían aportar en la caza piezas mayores, pero las aportaciones femeninas eran más constantes, como hacen ver las investigaciones de la australiana Rebeca Bliege Bird146.
Es posible que hasta la aparición del homo erectus y del homo er-gaster, los humanos no tuvieran envergadura, ni capacidad mental ni herramientas capaces como para atreverse a cobrar piezas de grandes dimensiones. «Con Homo ergaster aparece, muy probablemente, una nueva estrategia, ya que la necesidad de aumentar la aportación proteí-nica seguramente provocó un cambio en las estrategias alimentarias y en las técnicas de caza. El entorno en el que habitaban, las enormes sa-banas africanas repletas de animales de grandes dimensiones, debió de
145 Cfr. Arsuaga, J. L., Los aborígenes, o.c., pp.129 y ss. Para una visión femenina de la hominización y la humanización, cfr. Martínez Pulido, Carolina, El papel de la mujer en la evolución humana, Madrid, Biblioteca Nueva, 2003; Id., La presencia femenina en el pen-samiento biológico, Madrid, Minerva Ediciones, 2006; Nussbaum, Martha C., Las mujeres y el desarrollo humano, Barcelona, Herder, 2002.
146 Cfr. Arsuaga, J. L., Los aborígenes, o.c., pp. 132 y ss.

318
© Universidad de Deusto - ISBN 978-84-9830-383-4
facilitarlo aún más»147. En ese momento se iría dando un salto hacia el arte de cazar animales más grandes. Para ello se necesitaban varias con-diciones, como, en primer lugar, un tipo más estricto de organización, en la que toda la comunidad tenía que sentirse implicada, y que suponía un fuerte liderazgo y una importante capacidad de comunicación lingüísti-ca. Y, por otro lado, se imponía inventar nuevas formas de transporte de las grandes piezas cobradas para llevarlas a los campamentos más esta-bles, para proceder allí al descuartizamiento, aprovechamiento y almace-naje de carne, pieles y huesos. Además, se requería, como ya lo hemos repetido, la confección de herramientas de madera muy específicas para la caza, como son lanzas, jabalinas con sus lanzaderas y flechas, junto con otras herramientas líticas características del denominado Modo 2, o achelense. Estos beneficios tan grandes que produce la caza y la pro-gresiva dependencia de la misma, es lo que explica también el inicio de una mitologización de los animales148, convirtiéndolos en totems tribales y en personificación de las fuerzas divinas de la naturaleza, así como su aparición en el primitivo arte prehistórico, tanto en las cuevas como pin-turas rupestres, como también en las posteriores primeras estatuillas de piedra. El alimento y los usos alimenticios en las comidas rituales, siem-pre ha tenido en casi todas las religiones de la historia de la humanidad un alto valor religioso, dictándose leyes sobre alimentos puros e impuros y sobre su uso y modo de comerlos, estableciéndose toda una legislación ritual sobre los alimentos149.
El éxito progresivo de este tipo de caza supuso también unos medios sostenidos de alimentación rica en calorías animales, que llevó a un au-mento del cerebro, así como a un considerable incremento de la pobla-ción, produciéndose, a su vez, una mayor capacidad de formar grupos cazadores cada vez más dotados y expertos. «La caza de los grandes ani-males es una actividad propia de una buena adaptación a las sabanas y podría haber guardado relación con el desplazamiento de los homínidos a lo largo de África centroriental. Ante la competencia de los grandes depredadores, el homínido hallará una nueva forma de adaptación hasta entonces desconocida y, por consiguiente, nacerá un nuevo período en lo que respecta al desarrollo de las diferentes especies de homínidos»150.
147 Carbonell, E./Sala, R., Planeta humano, o.c., 93. 148 Cfr. Clarke, R., o.c., 58-60.149 Cfr. Soler, Jean, «Las razones de la Biblia: reglas alimenticias hebreas», en Flandrin,
J-L/Montanari, M. (dirs.), Historia de la alimentación, Gijón, Edic. Trea S. L, 2004, 81-94; Cruz Cruz, J., Alimentación y cultura. Antropología de la conducta alimentaria, Pamplona, EUNSA, 1991; Harris, M., Bueno para comer, Madrid, Alianza, 1989.
150 Clarke, R., o.c., p. 94.

319
© Universidad de Deusto - ISBN 978-84-9830-383-4
Por tanto, el ejercicio progresivo de la caza en grupo constituyó un comportamiento cultural que tuvo repercusiones tanto en el aumento del cerebro, con sus correspondientes consecuencias conductuales, así como con el avance en la técnica de las herramientas y en el crecimien-to de la cooperación, de la comunicación y de la organización social151.Un elemento fundamental en la relación entre caza y organización so-cial se refiere al reparto de los alimentos cazados. La mayoría de los animales depredadores se alimenta de sus presas en el mismo lugar donde las caza. El ser humano haría quizá lo mismo al principio. Pero bien pronto se advierten restos arqueológicos de utilización de campa-mentos de concentración y acampada más permanente a donde se lle-vaba la caza, se repartía entre los diversos grupos, se conservaban los excedentes, y se realizaban otro tipo de actividades encaminadas a la caza, como construir nuevas y mejores herramientas. Este modo de re-partir la caza es un elemento que caracteriza al ser humano, a diferencia de la mayoría de los animales152. Y supone también una alta organiza-ción social, que no dejaría de tener conflictos y ser ocasión incluso de división y ruptura de ciertos grupos. No vamos a repetir lo dicho más arriba respecto a la necesidad de superar la idea de un estricto reparto de roles sexuales de cara al aprovisionamiento de alimento, reservando a los varones la caza de piezas grandes y a las mujeres el cuidado de los hijos y del campamento. La situación sería sin duda mucho más com-pleja y variada.
No podemos detenernos en hacer una descripción de los momentos más importantes de la historia de la caza y de los progresos en la misma, con los que supone de avance en la construcción de herramientas y en la complejidad de la organización social, entre otras cosas porque respecto a las herramientas ya nos hemos extendido en páginas anteriores, y sobre la organización social nos detendremos más adelante. Como resumen, nos
151 Para la relación entre caza y organización social, cfr. ibídem, 57-58 y 95-97. 152 Cfr. Carbonell, E./Sala, R., o.c., pp. 96-97: «Otra dimensión social de la caza es
la que implica un comportamiento solidario gracias al cual se tiende a compartir la car-ne, un tipo de alimento que se presta más a desarrollar actitudes sociales de esta natura-leza. Los frutos y vegetales a menudo se consumen a medida que se recogen; en cambio, cuando se caza un gran mamífero, el gran volumen del alimento conseguido permite que pueda ser compartido por los miembros del grupo y, aún más, el hecho de compar-tir se convierte en algo necesario tanto para la cohesión interna como para aprovechar al máximo todos los recursos. Esta es la hipótesis formulada por G. Isaac en 1978: que el compartir el alimento estuvo en la base de la estructuración social y del crecimiento y la especialización del lenguaje de nuestro cerebro. En definitiva, la relación entre la caza y el consumo de carne con la complejidad social parecen muy claros, aunque nos falta es-tablecer desde cuándo es así».

320
© Universidad de Deusto - ISBN 978-84-9830-383-4
basta decir, con Carbonell y Sala, que en el caso de los humanos «par-tiendo de una situación de una inferioridad e indefinición con aprovecha-mientos secundarios, se pasa por las fases de inicio y desarrollo masivo de la caza activa, a más tardar en el Pleistoceno medio, y se llega a la fase final de aprovechamiento intensivo y máximo a partir de la segunda mitad del Pleistoceno Medio y, especialmente, en el Pleistoceno Superior y el Holoceno»153.
Y se llega en esta historia a un punto clave, que constituye el se-gundo momento decisivo del que hablaba Arsuaga: el paso al Neolíti-co, hace entre unos 15.000 y 10.000 años antes de nuestra era154. Es el momento en que se da el paso decisivo, a la vez que traumático, de un estilo de vida trashumante, propio de cazadores-recolectores, al seden-tario, con una economía basada en la agricultura y la ganadería. Pero hay que precisar también que el paso de la trashumancia a la sedenta-rización no coincidió con el descubrimiento de la agricultura, sino que se produjo antes. En estos momentos, aprovechando el buen clima, el hombre viviría al abrigo de salientes rocosos o en cavernas, producien-do los que se ha denominado la cultura natufiense155, investigada en el Oriente Próximo. En esas regiones geográficas, fueron apareciendo ce-reales salvajes (almidonero, cebada, escandia, antepasado del trigo), de los que los seres humanos aprendieron a alimentarse. Según algu-nos paleoantropólogos, los primeros pasos hacia la agricultura se da-rían, por tanto, aprovechando la recolección de estos cereales salvajes, antes de que aprendieran los humanos a cultivarlos por su cuenta. Esta tesis parece probada, al mostrarse la mayor antigüedad de las primeras aldeas y construcciones de casas que los restos de agricultura. Por tan-to, el paso de la trashumancia a la sedentarización se realizó cuando los humanos experimentan que no necesitan andar buscando comida de un sitio a otro, teniéndola en abundancia en un mismo lugar, sin necesidad todavía de generarla por los métodos agrícolas. Por tanto, la sedentari-zación precedió al dominio de la agricultura. Y lo mismo hay que decir respecto a la comida de carne: «Vivían en un medio que les proporcio-naba todo lo que les hacía falta: peces, crustáceos, moluscos, aves, pe-queños mamíferos fáciles de cazar, y cereales al alcance de la mano. Por ello, ya no tenían necesidad de ir a conquistar nuevos cazaderos»156. De ese modo, dejaron sus habitáculos al abrigo de las rocas y las cavernas, y empezaron a construir sus pequeñas y primitivas viviendas junto a los
153 O.c., p. 102. 154 Cfr. Arsuaga, J. L., Los aborígenes, o.c., pp. 99 y ss. 155 Cfr. Clarke, R., El nacimiento del hombre, o.c., pp. 183 y ss. 156 Ibídem, p. 184.

321
© Universidad de Deusto - ISBN 978-84-9830-383-4
sitios donde tenían la comida. Y así se inicia la época fundamental de la vida sedentaria, y el comienzo del Neolítico.
En esos momentos, el paso de la recolección de semillas, frutos secos y raíces a la producción de los primeros cereales, permite un cambio fun-damental en la humanidad157. Al parecer se produce este salto tanto en el ámbito del Mediterráneo, el creciente fértil, con el cultivo del trigo, como también en Asia, con el arroz, en África, con el sorgo y el mijo, y en Amé-rica con el maíz.
Igualmente, ya no se dedica el ser humano a cazar, sino que ha apren-dido a domesticar algunos animales, y, en vez de perseguirlos para cazar-los, los encierra y engorda en sus corrales para alimentarse de ellos cuan-do lo desee. No se tienen evidencias de cuándo y dónde se produjeron estas primeras prácticas de domesticación de los animales y el inicio de las consecuentes prácticas ganaderas158. Pero parece que los primeros ani-males domesticados fueron el perro159, como ayudante de caza, y luego el toro y la vaca160. Respecto al caballo todo parece apuntar a que se domes-ticó en las extensas estepas asiáticas, que van desde Ucrania hasta el Asia Central, siendo importante tenerse en cuenta de que el caballo no sólo ha sido utilizado como montura, tanto para la carga como para la guerra, sino también como alimento, hasta que su utilización estratégica para la guerra se impuso económicamente sobre su utilización como alimento, al menos en Europa, tal y como lo defiende Marvin Harris161.
Con la implantación de la ganadería y la agricultura, el ser humano experimenta un cambio drástico en su estilo de vida, pasando de un tener que estar dependiendo de perseguir sus presas y recolectar vegetales, a tener a su disposición de modo fácil todo tipo de alimentación, tanto ve-getal como cárnica. Eso supuso también la posibilidad de un aumento ex-ponencial de la demografía, así como la construcción de aglomeraciones
157 Cfr. Clarke, R., El nacimiento del hombre, o.c., cap. 10.º, «Los primeros campe-sinos».
158 Sobre la domesticación de los primeros animales, cfr. Clarke, R., o.c., cap. 13, pp. 217 y ss., donde se nos habla del perro, caballo, el toro y demás.
159 Cfr. Allman, J. M., El cerebro en evolución, o.c., pp. 211-217 («Bailando con lo-bos»).
160 «En el caso de la vaca doméstica, los estudios genéticos indican que las razas euro-peas y africanas derivan del uro (Bos taurus), mientras que las de la India proceden de otra especie de toro salvaje (Bos indicus). Los estudios de diversidad indican que nuestras va-cas domésticas vienen de uros del Próximo Oriente, que es donde la diversidad genética es máxima, mientras que el centro de origen de las razas africanas podría ser independiente y situarse en Egipto, donde hay unas dataciones de presuntas vacas domésticas de hace más de 9.000 años», Arsuaga, J. L., Los aborígenes, o.c., p. 154.
161 Cfr. Harris, M., Vacas, cerdos, guerras y brujas. Los enigmas de la cultura, Madrid, Alianza, 1980.

322
© Universidad de Deusto - ISBN 978-84-9830-383-4
urbanísticas, la conformación de alianzas de ciudades y la aparición de los primeros estados, hasta la superpoblación y la complejidad social de nuestro tiempo.
2.7. El hábitat humano. La construcción de refugios y casas
Como en el resto de los demás capítulos de la primera cultura huma-na, en lo referente a los primeros habitáculos artificiales no se tienen mu-chas evidencias claras. Y es lógico que así sea, como ocurre con las herra-mientas. Se supone que los primeros humanos vivían en climas cálidos, y estarían por ello menos necesitados de refugios fuertes para defenderse de las inclemencias de climas fríos y tormentosos. Así mismo, las prime-ras casas estarían construidas con materiales débiles, difíciles de fosilizar. Aún así, la paleoantropología y la arqueología van encontrando algunas evidencias como para poder hablar de algunos indicios objetivos de estos primeros momentos.
De todos modos, es importante que se supere la visión de tiempos atrás de creer que los primeros humanos vivían de una forma generaliza-da en cuevas, siendo casi el ser cavernícola sinónimo de hombre antiguo. Esta tendencia se debe a que hasta hace no mucho, la arqueología apenas si había encontrado restos de viviendas de nuestros antepasados fuera de estos ámbitos cavernícolas. Pero es normal que así sea, porque el resto de habitáculos en campo abierto ha sido más difícil de producir restos y de ser encontrados por los investigadores. Pero desde finales del si-glo XX las cosas van cambiando. Así, como dicen Carbonell y Sala, hoy día se da por sentado que «la vida cavernícola de los homínidos fue muy esporádica y limitada y que sólo sirvió para complementar la vida en campamentos al aire libre»162. El ser humano no vivió más que de forma muy limitada en territorios cársticos, y en esas circunstancias aprovecha-ría las posibilidades de vivir en cuevas, pero en la inmensa mayoría de los casos, vivía en terrenos abiertos de sabana, y allí tuvo que inventar e idear refugios adecuados.
En esas situaciones, los seres humanos aprenderían a levantar cons-trucciones de protección, al estilo de lo que ya hacen determinados animales con sus nidos y sus viviendas. Al principio serían muy ele-mentales, pero más adelante construyeron habitáculos más complejos y sofisticados. El habitáculo humano más antiguo fue descubierto en África por los Leaky, en la localidad de Bed I en Olduvai (Tanzania).
162 Planeta humano, o.c., 83.

323
© Universidad de Deusto - ISBN 978-84-9830-383-4
En una de sus excavaciones, descubrieron unos bloques de piedra apila-dos de forma circular, con una extensión aproximada de cinco metros de diámetro. Fuera de ese entorno, estaba lleno de restos de herramientas y huesos, pero no había nada dentro del recinto circular. Aunque no se pueda asegurar de modo definitivo que se trate de una vivienda elemen-tal, todo parece indicar que es de origen antrópico, y que no es una mera acumulación de bloques de piedra para sacar de ellos herramientas líti-cas163. Es de suponer, por su antigüedad, que esta vivienda o simple re-fugio vendría a ser «la primera prueba de organización del espacio o una construcción artificial. La acumulación intencionada podría responder a la modificación del espacio para destinarlo a algún tipo especial de acti-vidad o para rodear un espacio humano protegido»164. Dada la antigüe-dad de esta construcción, al inicio del Pleistoceno, hay que atribuirla al homo habilis.
Muy cerca de Olduvai, en Melka Kunturé (Etiopía), también se en-contró una nueva vivienda, esta vez de forma rectangular, descubierta por Jean Chavaillon. En este caso, la protección no estaría hecha por pie-dras, sino por material vegetal que no ha dejado restos. Pero parece razo-nable pensar que allí habría paredes que protegerían la superficie de sue-lo empedrado, aunque no se sabe si sería una cabaña o simplemente un refugio para defenderse de los vientos Esta segunda vivienda posee una antigüedad de aproximadamente un millón de años, y «debería relacio-narse con las actividades domésticas de Homo ergaster o de Homo erec-tus africano»165.
Mucho más moderna es la casa descubierta por H. De Lumley, en Terra Amata (Niza, Francia). Se trata en este caso de una estructura oval, en cuyo interior se encontró un hogar y restos de diversas actividades. La antigüedad de la construcción, de unos 350.000 años, permite atri-buirla al Homo heidelbergensis, y estaría construida por materiales pe-recederos, como vegetales y pieles. De ahí que sólo han pervivido restos de piedra, tanto los bloques que la sujetaban como diversos instrumentos líticos166.
En fechas mucho más recientes, a partir de los 30.000 años (Pleis-toceno Superior), se han encontrado, en Kostienki (Rusia)167, diversas construcciones obra del Homo sapiens, y construidas con defensas de
163 Cfr. ibídem, pp. 85 y ss.; ver en p. 82 el dibujo de esta vivienda primitiva. 164 Ibídem, p. 86. 165 Ibídem, p. 86. 166 Ibídem, p. 86; cfr. Clarke, R., o.c., pp. 177-179.167 Cfr. ibídem, p. 86; véase en pág. 88 dibujos de las viviendas de Kostienki y de Terra
Amata.

324
© Universidad de Deusto - ISBN 978-84-9830-383-4
piedra y con diversos huesos de grandes proboscídeos, como el mamut. También se han hallado habitáculos en forma oval en Ripiceni-Izvor (Rumanía), descubiertos por el profesor Paunescu; esta residencia huma-na estaba «construida por piedras calcáreas, huesos de animales, defen-sas de mamut, piezas de silex y un hogar. Según asegura este colega, las defensas de mamut, clavadas al suelo por su parte más gruesa, servían a modo de palos sobre los que se sujetaban pieles con el objeto de cons-truir un espacio cerrado por los extremos y abierto por la parte superior, por donde comunicaba al exterior gracias a una abertura»168.
En el yacimiento de Molodova, en la orilla del Dniester medio (anti-gua URSS), fue descubierto otro tipo de habitáculo por el profesor Cernis, quien considera que se trata de una tienda cerrada y cubierta. Tenía forma circular, de unos ocho metros de diámetro, y en su interior se han encon-trado hogares, huesos de animales, cráneos, defensas, molares y omopla-tos de mamut169.
En el tiempo de los neandertales, se alternaban épocas templadas con otras más frías. Si en estas últimas viviría en cuevas, en las primeras lo hacía más al aire libre, cerca de los ríos y donde hallaba protección con-tra el viento. En esas épocas, el modo de alimentación procedía de la pesca y, sobre todo, de la caza trashumante; de ahí que tendría un cam-pamento base más o menos permanente junto con la necesidad de hacer expediciones itinerantes, con campamentos más provisionales para viva-quear, como muestran los restos arqueológicos encontrados, y que nos hacen entender que serían campamentos provisionales, donde construía y guardaba a veces las herramientas, y las piezas de caza tomadas, para de regreso transportarlas al campamento central. «El hombre de Neandertal, indica Clarke, tenía la costumbre de vivir entre los cuerpos de los anima-les con los que se alimentaba, que simplemente dejaba arrinconados con-tra las paredes de su refugio. En algunas de esas cuevas, su paso alterna-ba con los de los animales, como hienas y lobos, con los que tal vez tuvo que disputarse los alimentos»170.
Si en épocas muy antiguas los restos encontrados son pocos y ele-mentales, en épocas ya más cercanas se halla mayor abundancia de es-tas construcciones, así como de una mayor amplitud y complejidad, y también una gran variedad de tipos de viviendas, con el paso del tiempo aglomeradas en pequeñas aldeas, anticipo de las futuras or-ganizaciones urbanas. En fechas comprendidas entre hace 25.000 y 20.000 años, hay que situar las cabañas de Paulov (Chequia), donde
168 Ibídem, pp. 86-87. 169 Cfr. ibídem, p. 87. 170 O.c., p. 180.

325
© Universidad de Deusto - ISBN 978-84-9830-383-4
el profesor Klima ha descubierto una gran variedad de viviendas, unas de forma oval y otras circulares, con extensiones entre 4 y 8 metros de longitud. En su interior se descubren en todas ellas restos de hogares, y abundantes restos de materiales diversos, lo que hace pensar que fueron habitadas durante épocas dilatadas de tiempo. Todo parece indicar que estas cabañas, así como las encontradas en Dolnì Vestonice (Chequia), estaban construidas con cubiertas, y poseían una única protección peri-metral. La complejidad de estas construcciones, con un cierto reparto de habitaciones para dedicaciones distintas, nos muestra ya el inicio del reparto de tareas171.
Y en los años que van entre los 15.000 y 14.000 es cuando se pro-ducen los primeros fenómenos de sedentarización, como ya hemos apuntado al hablar de los alimentos. Es un fenómeno seguramente an-terior, como ya dijimos, a la aparición de la agricultura y la ganadería. Hace unos 12.000 años, aparecieron también los primeros poblados y aldeas, momento en el que predomina la denominada cultura natufien-se, localizada en el Oriente Próximo, donde se han encontrado las aldeas más antiguas que se conocen: la de Mallaha (Israel), descubierta y estu-diada por Jacques Perrot, y la de Mureybet, cerca del Éufrates (Siria), descubierta por Jacques Cauvin. La aldea de Mallaha consta de unas veinte cabañas redondas, con los rasgos de ser habitáculos permanentes y organizados en forma de aldea. Las habitarían cazadores y pescadores, siendo muy posible que recolectaran además granos silvestres de cerea-les, que crecían de forma abundante en los entornos. Con el tiempo, la facilidad de la recolección de comida llevó a un aumento de la pobla-ción, y al paso progresivo de recolectar los cereales salvajes a saber pro-ducirlos, dando así origen a la agricultura y al asentamiento definitivo de las residencias humanas172.
Es en esos ámbitos de la cultura natufiense, extendida entre el Nilo y el Éufrates, donde aparece también rastros de albañilería, así como una incipiente carpintería. Se han encontrado muros de piedra sostenidos por mortero de barro, con pilares de madera así como planchas y rodi-llos de madera, modelados gracias a un nuevo utensilio inventado en esta época: la azuela, el más antiguo de los cuales lo descubrió J. Cauvin en Mureybet, a orillas del Éufrates173. También se advierte el cambio de vi-
171 «En esta época, comienzan a aparecer de modo más claro y definido, los restos que evidencian la división del trabajo en los habitáculos. El rincón donde se trabaja el sílex es-taba bien diferenciado del a zona de cocina, cerca del fuego, o de la zona de reposo, donde se han hallado restos de lechos de pieles»: Clarke, R., o.c. p. 181.
172 Cfr. ibídem, pp. 181 y ss.173 Cfr. ibídem, p. 186.

326
© Universidad de Deusto - ISBN 978-84-9830-383-4
viendas circulares a otras rectangulares, que tienen como ventaja el que permiten todo tipo de ampliaciones y añadidos posteriores. Con este tipo de viviendas se permitió la construcción progresiva de aldeas y ciudades, organizando las casas a través de calles y plazas. Supone todo esto el na-cimiento de una sociedad más compleja, que va superando los aislados núcleos familiares y tribales174.
En esta misma época, hace aproximadamente unos 12.000 años, época dominada por la cultura magdaleniense, hay que hacer referencia a los res-tos encontrados en Pincevant (Francia) por el profesor Leroi-Gourhan: unos habitáculos de estructura circular y rodeados por múltiples desechos domés-ticos y varios hogares. Sólo más adelante, a partir de los 8.000 años, los hu-manos irán aprendiendo a emplear todo tipo de materiales y sobre todo las piedras talladas para construir sus casas, en el momento en que empiece la época de las primeras construcciones urbanas. A partir de los 10.000 años empiezan a aparecer las ciudades, una de las más antiguas conocidas es la de Caral-Hüyük, al este de Turquía. Se trata de una ciudad sin calles, tenien-do que caminar por los techos de las casas a las que se accedía desde arriba, a través de una escalera. Se puede advertir que en esa ciudad se cultivaba trigo y cebada, así como guisantes y lentejas. Habían domesticado a ovejas, cabras y perros. También se han hallado restos de enterramientos humanos debajo de las casas. El arqueólogo inglés que la descubrió, James Mellaart, encontró todo tipo de objetos manufacturados de gran valor, y de una anti-güedad de alrededor de 9.000 años. Junto a los enterramientos, también se han hallado indicios de ritos religiosos, con cultos basados en la representa-ción de toros así como cultos a la diosa de la fertilidad175.
«Como vemos, en todas las épocas históricas hay vestigios de las construcciones humanas: las edificaciones son cada vez más complejas y nos informan sobre los adelantos en el uso de los materiales y en la téc-nica constructiva»176. Esto nos confirma, como ya lo habíamos indicado, que los humanos no habitaron en cuevas de forma exclusiva, ni siquiera de modo preferente, sino que su modo de refugiarse de las inclemencias del tiempo y de otros peligros fue muy diverso, en consonancia con la pluralidad de ámbitos ecológicos en que tuvo que asentarse. Pero lo que sí es importante destacar, como conclusión de este apartado, es el hecho de que la construcción de refugios artificiales es lo que nos caracteriza como
174 Cfr. ibídem, pp. 185-190: aquí nos muestra R. Clarke la revolución que supuso para el ser humano la época del Neolítico, tanto en viviendas, costumbres alimenticias, perfec-cionamiento de herramientas, así como el fuerte cambio de mentalidad y de costumbres so-ciales y roles sexuales.
175 Cfr. ibídem, pp. 190-192.176 Carbonell, E./Sala, R., o.c., p. 89.

327
© Universidad de Deusto - ISBN 978-84-9830-383-4
humanos, y un modo de ir consiguiendo nuestra humanización, en la me-dida en que los humanos no nos adaptamos de forma pasiva al entorno, sino que lo transformamos para hacerlo más seguro y confortable, en defi-nitiva, más humano.
2.8. La socialidad: de la familia a los primeros grupos sociales
Hemos ido viendo en repetidas ocasiones la centralidad del grupo para la supervivencia de la especie humana y también para el proceso de hu-manización. Se ha solido también afirmar que la organización social es un fenómeno típicamente humano. Pero tampoco es del todo cierto. En este punto, como en los demás que estamos analizando, hemos de situarnos en la tan repetida síntesis de continuidad y ruptura: la dimensión social de la vida humana es un fenómeno que comparte con las especies animales an-teriores, aunque también vamos a ver que las cualidades de la socialidad humana son específicas y únicas de nuestra especie.
La organización social, como grupo permanente de una especie, for-mada por individuos pertenecientes a varios tipos especializados (obreras, reinas, zánganos, …), que presentan una cierta jerarquización entre ellos y que están ligados por poderosos medios de integración, se da también en diversas especies animales. La sociedad constituye una especie de super-individuo, y es un nuevo nivel de integración, el más reciente y complejo de toda la evolución.
La socialización aparece, de modo más avanzado, en grupos situados en los dos extremos del árbol filogenético: entre los invertabrados, el de los insectos sociales, y, entre los vertebrados, en ciertos mamíferos y el hombre. La diferencia entre ambos grupos está en que, mientras entre las sociedades de insectos los comportamientos sociales se basan en estruc-turas genéticas, en las sociedades de los primates los comportamientos se basan en reglas conductuales más abiertas, y en el ser humano, en lazos culturales orientados por la conciencia y la libertad.
En los mamíferos no primates, se advierten varios tipos de sociedades:
— familia maternal estacional, compuesta por madre e hijos, sólo el tiempo necesario para aprender a alimentarse y a valerse por su cuenta;
— sociedad matriarcal: persisten más los lazos entre madre e hijos, y se pueden mantener unidos durante varias generaciones. Se da aquí una relación de parentesco alrededor de la madre dominante, como puede verse entre los ciervos, elefantes y leones. Los machos se quedan solos en la periferia, y sólo se acercan para el acoplamien-

328
© Universidad de Deusto - ISBN 978-84-9830-383-4
to, teniendo en ello la iniciativa las hembras cuando les llega el pe-ríodo reproductivo, el estro.
— Sociedad bisexuada permanente, donde se da la presencia constan-te de uno o varios machos adultos. Puede adoptar la forma de harén(macho polígamo), como en el caso de las cebras, vicuñas, etc., o la forma de parejas monógamas, como los castores y los lobos, o los grupos con machos comunitarios, como los lycaones de África. En este último caso no se da la fidelidad conyugal, aunque se mantiene la cohesión grupal durante varias generaciones, al estilo de una so-ciedad hippie humana.
Es importante señalar que esta tipografía de sociedades animales son modelos ideales, puesto que nunca se dan en su estado puro, sino que adoptan formas muy variables, pero la mayoría de las veces tienen como elemento común el que la madre siempre ocupa un lugar central y privile-giado, debido a su importante, y casi insustituible, rol de nutrición respec-to a los hijos.
En las sociedades de primates, como veremos más adelante, se man-tienen estos modelos de sociedad, pero innovando en algunos aspectos y flexibilizando las estructuras. Son sociedades más permisivas y menos es-trictas, siendo esto así porque es más favorable selectivamente. Si fuese perjudicial, no se mantendría. Los diversos tipos de estas sociedades res-ponden siempre a motivos ecológicos177.
2.8.1. La sexualidad humana y las primeras células familiares
La dimensión social de la especie humana empieza, como en el resto de los animales, en la célula familiar o similares. En el proceso evoluti-vo, se ha dado un salto en el modo de reproducirse los seres vivos desde un modo de reproducción asexuada, propia de la mayoría de las plantas (gemación), al modo de reproducción sexuada, por fusión de gametos complementarios con intercambio de dotaciones genéticas178. De ahí que en toda especie animal que se reproduce por mezcla del ADN de los dos gametos, masculino y femenino, el nuevo ser aparece y se sitúa dentro de
177 Para esta variedad de sociedades animales, cfr. Melotti, Humberto, El hombre entre la naturaleza y la historia, Barcelona, Península, 1981, pp. 142 y ss.
178 Cfr. Margulis, Lynn/Sagan, Doris, ¿Qué es el sexo?, Barcelona, Tusquets, 1998; Diamond, Jared, ¿Por qué es divertido el sexo? Un estudio de la evolución de la sexua-lidad humana, Madrid, Debate, 1999; Atlan, Anne, «La guerra fría de los cromosomas sexuales», Mundo Científico, 1998, n.º 189, 32-34; Erickson, Robert, «La saga de los es-permatozoides. ¿Cómo se formaron?», Mundo Científico, 1985, n.º 52, 1086-1097.

329
© Universidad de Deusto - ISBN 978-84-9830-383-4
un cierto ámbito familiar. Pero ya hemos visto que los emparejamientos y estrategias reproductivas son muy diversos. Algunos autores (Lovejoy) advierten que se dan dos estrategias diferentes: una de tipo cuantitativo, consistente en producir un número abundante de descendientes (los pe-ces, por ejemplo: la hembra produce una cantidad ingente de huevos, y el macho los rocía con el esperma, dejándolos en la corriente de agua sin preocuparse más por su futuro), sin invertir demasiado esfuerzo en la manutención y cuidado de los descendientes; otra estrategia es más cuali-tativa, en la medida en que se producen menos descendientes, pero se les cuida y protege con mucho más cuidado. Los sociobiólogos hablan de que en cada caso se habría llegado a lo que ellos denominan una estrategiaevolutivamente estable (EEE), en la medida en que cada especie habría alcanzado un equilibrio para saber conjugar la tendencia a maximizar el número de descendientes con la capacidad y esfuerzo que necesita invertir para cuidarlos.
La especie humana sería, dentro del resto de las especies existentes, la que ocuparía el vértice de la tendencia cualitativa en el proceso repro-ductivo: tiene pocos descendientes, pero los cuida con gran acopio de in-versión de tiempo, medios y afecto. Esta estrategia es consecuencia de un elemento específico de nuestra especie: la extrema deficiencia biológicacon la que nacemos179, de modo que se conjuga esta deficiencia biológica extrema con la tendencia diatrófica de la madre para alimentarlo y aco-gerlo180. En este hecho se ha dado un proceso coevolutivo, sobre el que hablaremos en el capítulo siguiente, en el que se han conjugado, dentro de la historia de la aparición del género humano, el aumento del cerebro, el bipedismo y el estrechamiento consecuente de la pelvis femenina, la pre-maturidad de nuestro nacimiento con la consiguiente deficiencia biológi-ca, la neotenia y necesidad de una protección más fuerte y duradera de los hijos, así como el proceso tan largo de nuestra educación e inculturación.
En la especie humana se ha dado también, dentro de la continuidad, una ruptura cualitativa en el comportamiento sexual, sobre todo el feme-nino. Darwin ya habló de la selección sexual como un procedimiento es-tratégico paralelo y complementario al de la selección natural181. Según
179 Cfr. Gehlen, A., El hombre, Salamanca, Sígueme, 1980. 180 Cfr. Rof Carballo, J., El hombre como encuentro, Madrid, Alfaguara, 1973; Id., «El
hombre como encuentro», en VV.AA., Homenaje a Zubiri, vol. II, Madrid, Ed. Moneda y Crédito, 1970, pp. 587-616; Beorlegui, C., «El hombre y sus imágenes. Del narcisismo a la alteridad», Letras de Deusto, 22 (1992), n.º 52, 81-108.
181 Cfr. Darwin, Ch, El origen del hombre, México, Editores Mexicanos Unidos, 1982. El título completo de este libro es El origen del hombre y la selección sexual. Cfr. Buskes, Chris, La herencia de Darwin, o.c., cap. 2.º. La selección sexual; Arsuaga, J. L./Martí-nez, I., La especie elegida, o.c., cap. 11.

330
© Universidad de Deusto - ISBN 978-84-9830-383-4
Ch. Darwin, el dimorfismo sexual que se advierte en la mayoría de las es-pecies, se debería al proceso de selección sexual consistente en que los machos eligen a las hembras en función de ciertas cualidades específicas, al igual que las hembras que se fijan en otras cualidades a la hora de de-jarse fecundar por los machos.
Ahora bien, en la totalidad de las especies animales, las hembras sólo son receptivas desde el punto de vista sexual en períodos determinados y cíclicos de tiempo. Es lo que se denomina el estro. En cambio, en las hembras humanas el período de celo, o estro, desaparece, y de ese modo el momento de la ovulación no es detectable, de tal forma que, «a diferen-cia de las hembras de los antropomorfos, su sexualidad no está regida por el celo e incluye los largos períodos infértiles de la gestación, lactancia y menopausia»182. Según esto, en la hembra humana la sexualidad se halla separada de la mera reproducción, no siendo ésta su única función, sino que posee otros fines diferentes, entre los que el más importante sería for-talecer la unidad y el amor entre la pareja.
A la hora de determinar cómo estuvo conformada las primeras unida-des familiares de la especie humana, no es fácil hallar una respuesta úni-ca y clara, puesto que los comportamientos no dejan restos fósiles, y la pluralidad de formas familiares parece haber sido la norma humana. Por ello, tenemos que hacer referencia a los comportamientos de los primates, como antropomorfos más cercanos a nosotros, así como a las diferentes especies de homínidos, sobre todo los australopitecos.
Los comportamientos de los primates en este punto son bastante di-ferentes, según nos indican las investigaciones de las tres estudiosas más famosas sobre ellos: Jane Goodal183, en el campo de los chimpancés, Dian Fossey (1932-1985)184, en el de los gorilas, y Biruté Galdikas, en el de los orangutanes185. De estos estudios etológicos se desprende que las hembras de los orangutanes tienen su período de celo cada seis años. Igualmente, las de los gorilas y chimpancés lo tienen a su vez entre tres y seis años. Pero el modo de agruparse en cada una de esas especies es diferente. Los
182 Arsuaga, J. L./Martínez, I., La especie elegida, o.c., 199. 183 Cfr. Goodall, J., A través de la niebla, Barcelona, Salvat, 1993; Martínez Pulido,
Carolina, El papel de la mujer en la evolución humana, Madrid, Biblioteca Nueva, 2003, pp. 221-234; Corbella, J., Entrevista a J. Goodall, La Vanguardia, 9 de marzo de 1997.
184 Fossey, Dian, Gorilas en la niebla, Barcelona, Salvat, 1985; Martínez Pulido, C.,o.c., pp. 235-243; Jahme, Carole, Bellas y bestias. El papel de las mujeres en el estudio so-bre primates, Madrid, Ateles Editores, 2002.
185 Cfr. Martínez Pulido, C., o.c., pp. 243-248; Visage, Albert, «La reina de los monos» (reportaje sobre Biruté Galdikas), El País Semanal, verano del 2000; Sanchís, Ima, Entre-vista a Biruté Galdikas, La Vanguardia, 16 de marzo de 2002; Alameda, Sol, «Biruté Gal-dikas. La reina de la selva», El País Semanal, 2002.

331
© Universidad de Deusto - ISBN 978-84-9830-383-4
orangutanes son animales solitarios. El macho se acerca a la hembra en los períodos de ovulación, y tras dejarla fecundada, la hembra queda sola encargada del cuidado de las crías, mientras el macho sigue por su lado buscando su propio sustento, manteniendo un cierto dominio sobre un territorio propio. Igualmente, cada hembra vive sólo en compañía de sus crías, al margen de las demás hembras, situándose dentro del territorio de un solo macho que abarca el territorio de varias hembras. Así, pues, la re-lación entre machos y hembras se reduce a los cortos y muy separados pe-ríodos de estro de las hembras. Pero los machos compiten entre sí por el dominio del territorio y por el acceso a las hembras que viven en él, por lo cual, por los efectos de la selección sexual, se produce en esta especie una gran diferencia de tamaño entre machos y hembras, fenómeno denomina-do dimorfismo sexual, de tal modo que el tamaño de los machos vienen a ser como el doble que el de las hembras.
La organización social de los gorilas es un tanto diferente, puesto que los machos dominantes (los machos de espalda de pelo plateado) con-trolan el acceso a varias hembras que viven en harenes, con sus propias crías. La lucha entre los machos por el acceso a las hembras, de cuya ali-mentación y cuidado se encargan, es más dura que entre los gorilas, por lo que el dimorfismo sexual viene a ser semejante al que se da entre los orangutanes. Los machos jóvenes que no poseen ningún harén, merodean alrededor de alguno de ellos, esperando la decadencia del correspondiente macho dominante o venciéndole con la ayuda de alguno de los otros ma-chos. Cuando uno es vencido, matan o expulsan a los descendientes del macho anterior, hacen abortar a las hembras, y se aseguran de que a partir de ese momento los descendientes futuros llevarán sólo sus genes186.
Entre los chimpancés la situación es diferente, puesto que no hay lu-cha entre los machos por el acceso sexual a las hembras, dándose entre ellos una total promiscuidad sexual. Con lo cual, no se da apenas entre los chimpancés dimorfismo sexual. Cuando se hacen mayores, las hembras se marchan del grupo, pero no así los machos. Cada grupo controla su propio territorio por medio de los machos, teniéndose que enfrentar por esta ra-zón muchas veces de modo violento con otro conjunto de machos de otros grupos vecinos. Pero la promiscuidad sexual que se da dentro de cada grupopermite que cuando una hembra está en período de celo, es copulada por varios machos a la vez.
La cuestión está ahora en saber cómo fueron las primeras sociedades familiares de los homínidos australopitecos, si se puede llegar a saber
186 Cfr. «La conducta de los primates», número monográfico de Investigación y Cien-cia, 2003, n.º 32.

332
© Universidad de Deusto - ISBN 978-84-9830-383-4
algo desde los diferentes registros fósiles. Por un lado, el claro dimor-fismo sexual que se advierte entre ellos permite suponer que su modo de convivir podría ser semejante al de los orangutanes y gorilas. De ahí que, en los yacimientos donde se descubre por primera vez restos de australo-pitecus affarensis, se tendió al principio a confundir la diferencia de ta-maño entre machos y hembras como si fueran individuos pertenecientes a especies diferentes. Pero, por otro lado, hay otros rasgos de su morfolo-gía, como la menor diferencia en el tamaño de los caninos entre machos y hembras (a diferencia de las especies en las que se da fuerte dimorfismo sexual), que parece indicar que no había disputas entre los machos por el apareamiento con las hembras. Por tanto, los especialistas consideran que no tienen pruebas suficientes como para sacar conclusiones contundentes en ninguna dirección187.
Sobre los comportamientos de los primeros seres humanos en este as-pecto no podemos ser demasiado explícitos, ni basarnos tampoco en datos muy objetivos. Por otro lado, no podía ser de otra manera, en la medida en que lo característico de los primeros humanos, en éste como en los de-más caracteres que estamos analizando, es precisamente no tener un tipo de comportamiento único y cerrado, que pudiera considerarse como el rasgo específico de los humanos consecuencia de una herencia genética determinada. Algo así como los rasgos específicos, rígidos y únicos de cada especie, lo que K. Lorenz denominada el etograma. Ya hemos in-dicado, por un lado, la especial característica sexual de las hembras hu-manas, así como las evidencias de la Antropología cultural acerca de la enorme pluralidad de costumbres de cada cultura humana. En líneas ge-nerales, podríamos decir, por tanto, que, dentro de nuestra especie huma-na, la pluralidad de comportamientos culturales hace que se asemeje cada grupo cultural a lo que es una especie biológica en el ámbito de los demás animales.
El cambio tan grande en el comportamiento sexual de las hembras hu-manas suponía poder separar el uso de las relaciones sexuales de la mera reproducción biológica, por lo que muchos antropólogos han visto en ello un elemento de coevolución que ha servido para la conformación de un tipo de relación intersexual monógama. El sociobiólogo Lovejoy, como ya lo vimos en su momento, ha defendido que el caminar erguido de forma permanente se unió con el progresivo uso inteligente de las manos para manejar instrumentos, poder llevar las presas para repartir los alimen-tos, así como para llevar en brazos a las crías por parte de las hembras,
187 Cfdr. Arsuaga, J. L./Martínez, I., La especie elegida, o.c., 208-216; Clarke, R., o.c.,pp. 60-63.

333
© Universidad de Deusto - ISBN 978-84-9830-383-4
de modo que todo ello contribuyó a la formación de una alianza y repar-to de funciones entre machos y hembras, como presión selectiva hacia la conformación de la familia monógama entre los humanos. A ese conjunto de características habría que unir la permanente disposición sexual de las hembras, que serviría para una mayor unidad afectiva entre las diferentes parejas188. Pero, aunque algunos argumentos puedan resultar plausibles, el conjunto de la teoría, como vimos en el capítulo correspondiente al hablar del origen de la bipedestación, resulta más que problemática, en la medida en que, como tenemos que decir reiteradamente, las culturas humanas han sido siempre muy plurales en el modo de organizar los núcleos familiares desde el principio. Más razonable resulta pensar que la especie humana no tiene un comportamiento único, regido por su base genética, sino que su genética le ha dado una gran libertad de posibilidades de establecer las relaciones intersexuales y de conformar los núcleos familiares, en función de las circunstancias de su entorno y de las decisiones culturales que en cada momento se han ido tomando.
2.8.2. El salto de la naturaleza a la cultura: el tabú del incesto
Se ha solido defender, empezando por Cl Lévi-Strauss, que el origen de lo humano en este aspecto que estamos analizando, como auténtico salto de la naturaleza a la cultura, se produce con la implantación del tabúdel incesto y la ley de la exogamia.
La primera tesis que se defendió entre los estudiosos de las primeras configuraciones familiares apuntaba a que los hombres primitivos vivían en promiscuidad total, y posteriormente, por influjo de la moral, se fue poniendo coto a esa forma de vivir hasta llegar a la situación de la vida moderna, apoyada en la familia nuclear y monógama. Así lo defendió John Ferguson McLean, en su libro Primitive Marriage (1865), donde describía el estilo de vida de los hombres primitivos protagonizada por un conjunto de bravos guerreros que se dedicaban a dominar a sus vecinos y a capturar a sus mujeres, con las cuales tendrían descendencia, fundamen-talmente varones, como individuos más valiosos189.
El problema está, a parte de lo poco fundamentadas de estas teorías novelescas, en cómo explicar el supuesto paso de esa vida de promis-cuidad y permanente violencia, en la que dominaba el instinto sexual sin controlar, a un estilo de vida dominado por las ordenaciones culturales.
188 Cfr. ibídem, p. 210. 189 Cfr. Kuper, Adam, El primate elegido, Barcelona, Crítica, 1994, cap. 11, «La prime-
ra familia».

334
© Universidad de Deusto - ISBN 978-84-9830-383-4
La respuesta estaba en el tabú del incesto, con su correspondiente ley de la exogamia. Pero en la explicación de este salto de la naturaleza a la cultu-ra se disputaban tres teorías diferentes: la de Edwar Westermarck, la de Siegmund Freud, y la de Claude Lévi-Strauss.
El estudioso finlandés E. Westermarck desechó la tesis de la promis-cuidad general de los orígenes del ser humano, cuestionando la metodolo-gía de los antropólogos a la hora de propalar como probadas las tesis de esa situación promiscua entre los hombres primitivos. Además, estaba de acuerdo con Darwin sobre el hecho de que los simios no eran especial-mente promiscuos, si descontamos el caso de los chimpancés. Wester-marck consideraba que el matrimonio y la familia eran universales entre los primates, aunque naturalmente la familia humana era bastante diferen-te a la de los primates, aunque sólo sea porque dura más que entre ellos. Entre los primates, cuando un macho crece es expulsado del grupo por el macho dominante, preservándose de ese modo los apareamientos inces-tuosos. La cuestión que se planteaba Westermarck era cuál podía ser el mecanismo que los humanos habían constituido para evitar los incestos, en la medida en que las poblaciones que toleran de modo generalizado el incesto se destruyen a la larga. «Westermarck sugirió que este mecanismo era parte fundamental del proceso mismo de construcción de la vida fa-miliar. Los niños que crecían juntos tendían a experimentar una aversión positiva respecto a la idea de un contacto sexual entre ellos»190. Se sue-len aducir como pruebas de esta tesis del escritor finlandés los estudios hechos en los kibbutz israelitas y en la sociedad china donde se practica el sim pua, o matrimonio de menores. Cuando niños de diferentes sexos viven en una misma casa como si fueran hermanos de sangre, sin serlo, de mayores se suelen oponer a contraer matrimonio y a tener relaciones sexuales entre ellos191.
La postura de S. Freud sobre el tabú del incesto es muy distinta. En su obra Tótem y tabú (1913), comenzó presentando una dificultad evidente a la tesis de Westermarck: ¿por qué convertir algo en tabú si es una tenden-cia biológica, que ya se daría entre los primates, e incluso, como defien-den hoy día los sociobiólogos, en la mayoría o totalidad de los animales? Si se tiene aversión al incesto como rasgo natural, no sería necesario pro-hibirlo, como no se necesita mandar o prohibir algo que lo hacemos de forma natural. Para Freud, la verdad es la contraria: se prohíbe el incesto porque los niños tienen una atracción sexual por los miembros de su fami-lia: son los complejos de Edipo y de Yocasta, como él los denominó. Los
190 Ibídem, p. 160. 191 Ibídem, pp. 160-161.

335
© Universidad de Deusto - ISBN 978-84-9830-383-4
niños sanos superan estos complejos, pero los neuróticos se muestran in-capaces. De ahí las diversas patologías que surgen de ello.
Siguiendo los planteamientos de Haeckel sobre la ontogenia como re-capituladora de la filogenia, Freud defendía que el desarrollo individual de la sexualidad atravesaba las etapas que la especie humana había expe-rimentado. De ahí que el complejo de Edipo, con el deseo de matar al pa-dre, venía a representar para Freud la repetición de la escena primigenia en la que un padre dominante se reservaba el exclusivo acceso a las hem-bras, prohibiéndoselo al resto de los varones. Pero la unión de éstos y la rebelión contra el padre había supuesto la superación de esa fase primera para dar lugar a la sociedad nueva, cimentada sobre la ley de la exogamia y la imposición del tabú del incesto. Así, el nacimiento de la sociedad su-ponía la implantación de la ley básica de la cultura que se imponía sobre las pulsiones naturales de tener relaciones sexuales con el progenitor del sexo opuesto. De este modo, según Freud, la civilización se constituye so-bre la represión de los instintos.
Como podemos ver, para Westermack la cultura se sigue de la natu-raleza, considerando la cultura como un reflejo de una aversión necesa-ria y natural del incesto, mientras que para Freud la cultura se cimenta reprimiendo y controlando a la naturaleza, criticando al finlandés con el argumento de que una aversión natural no requiere ser reforzada con la existencia de una norma. En la actualidad, tanto etólogos como socio-biólogos siguen los planteamientos de Westermack, en la medida en que consideran que la cultura no hace otra cosa que expresar y confirmar la naturaleza. De modo que tratan de mostrar, desde la observación de los comportamientos animales, cómo también entre ellos se daría igualmen-te el rechazo natural al incesto. La verdad es que, de algún modo, ambos planteamientos pueden complementarse. La diferencia está en que ambos autores identifican distintos mecanismos inhibidores del incesto: sea, en el caso de Westermack, que se advierta que los niños que viven juntos in-troyectan un rechazo posterior a casarse, o sea, en el caso de Freud, que la tendencia natural a la relación sexual entre madre-hijo es regulada por la prohibición del padre, los dos consideran a la familia como mecanismo inhibidor y regulador de la tendencia incestuosa. En ambos casos se persi-gue la ventaja genética y psicológica del apareamiento con individuos ex-traños a la familia.
Los planteamientos de Cl. Lévi-Strauss se orientan de modo muy dis-tinto a los de los dos anteriores. Apoyándose en las teorías de E. B. Tylor192,
192 Cfr. Tylor, E. B., «On a Method of Invertigating the Development of Institutions: Applied to Laws of Married and Descent», Journal of the Anthropological Institute, 18 (1888).

336
© Universidad de Deusto - ISBN 978-84-9830-383-4
uno de los grandes antropólogos de la escuela evolucionista, que descri-bía a los hombres primitivos no tanto como hombres en permanente guerra y conflicto, sino como amantes de la paz a base de construir pactos entre ellos, consideraba también que el modo de establecer relaciones estables entre diversos grupos de humanos era a base de establecer relaciones paren-tales, intercambiando en matrimonio a sus respectivas mujeres. Sobre esta base teórica escribió Lévi-Strauss Las estructuras elementales del paren-tesco193, donde defiende que el tabú del incesto se instituye como condición previa para el establecimiento de un sistema de parentesco y de alianzas matrimoniales. Y de ese modo se convierte en el origen de una sociedad es-table y en paz. Por tanto, la ley de la exogamia se erige en la frontera entre la naturaleza y la cultura, viniendo a ser la familia el núcleo de la sociedad, como célula universal, aunque el tipo de familia que se descubre en las di-versas sociedades humanas sea muy plural y diversificada, pudiendo encon-trarse tanto familias patrilineales como matrilineales. Pero, aun así, todo tipo de intercambios se reducía a una de estas tres fórmulas: a) intercambio directo de hermanas entre dos hombres; b) intercambio aplazado: cada uno entregaba su hermana a otro hombre y recibía en la siguiente generación a la hija de él para casarla con su hijo; c) intercambio generalizado, en el que no se realizaba el intercambio directo de hermanas, sino que se entregaban a un segundo grupo, que a su vez las entregaban a un tercero, y éstos a un cuarto, etc., formándose así una cadena abierta, en la que el último grupo entregaba sus mujeres a los hombres del primer grupo.
Esta teoría, que ha tenido un amplio respaldo durante varias décadas, ha recibido también serias críticas, porque se advierten en ella algunas de-ficiencias graves. La primera se refiere al hecho de que no hay evidencias claras de que este tipo de relaciones de parentesco matrimonial sea una institución universal. No lo es entre los cazadores-recolectores, y tampoco en el Islam, donde se permiten en muchos casos la endogamia. En segun-do lugar, como ya indicábamos anteriormente, los etólogos y sociobiólo-gos defienden que también se da la tendencia a evitar el incesto entre los animales, por lo que no sería una ley exclusiva de los humanos194. Es ver-dad que entre los animales domesticados no parece que se evite la endo-gamia, pero entre los que viven en estado natural sí. Y la tercera dificultad provendría del hecho de que en varias culturas humanas se va claramente contra la exogamia, tal es la costumbre entre los faraones, que se casaban
193 Les structures élémentaires de la parenté, París, PUF, 1949 (Las estructuras ele-mentales del parentesco, Buenos Aires, Paidós, 1969); González Echeverria, Aurora, Teo-rías del parentesco. Nuevas aproximaciones, Madrid, Eudema, 1994.
194 Deputte, Bertrand, «La evitación del incesto en los primates», Mundo Científico,1988, n.º 76, pp. 54-64.

337
© Universidad de Deusto - ISBN 978-84-9830-383-4
entre hermanos, y en diversas culturas de Persia y Egipto. De este modo se pondría en cuestión la tesis de que la ley del incesto sea una ley univer-sal de la cultura humana, y el salto definitivo de la naturaleza a la cultura.
La Antropología cultural también ha puesto en entredicho la tesis de que el matrimonio y la familia sean instituciones universales, como pretende Westermack, puesto que se advierte su ausencia, o el diferente modo de entenderlos, en algunas culturas. Tal es la tesis que dedujo Mali-nowski tras sus investigaciones en las Islas Trobriand (Nueva Guinea)195.En esa cultura, se da un parentesco de tipo matrilineal, y supuestamen-te no se advertiría entre ellos el complejo de Edipo. Otro ejemplo que se suele poner es el caso de los nayar, en la India, una cultura de guerreros, descrita ya por Duarte Barbosa en el siglo XVI. Los varones de los nayarno se casan, y sus herederos eran los hijos de sus hermanas, tenidos con un brahmán. Ahora bien, cuando la situación histórica llevó a la desapa-rición de este estilo de vida (un ejército constituido por mercenarios per-manentes que estaban con mucha frecuencia fuera de su entorno familiar), durante la época de la colonia británica, se impuso entre ellos el sistema familiar tradicional196.
Por tanto, aunque se advierten excepciones en la presencia del sistema familiar en algunas culturas, estas excepciones parecen deberse más bien a circunstancias económicas o políticas que obligan a adoptar costumbres diferentes. Pero, cambiadas tales circunstancias excepcionales, se vuel-ve a la norma familiar universal. Por tanto, parecería ser la norma común de la condición humana la existencia de la institución del matrimonio y la familia como una institución que caracteriza de modo universal el modo cultural de convivencia entre los humanos.
Pero, aunque esa afirmación pueda ser cierta, también lo es que los matrimonios y las familias tienen muy diferentes formatos. Muchas veces se piensa que el estilo primero de las familias tendría una estructura am-plia, al estilo de un clan, para después evolucionar hacia familias nuclea-res, formada sólo por padres e hijos, pero esto no siempre ha sido así, y el modo de conformar clanes con diversas familias nucleares dentro ha sido siempre muy plural. Igualmente, la familia ha sido considerada por unos como el cimiento de la sociedad, en la medida en que era el instrumento más adecuado para la transmisión de los valores fundamentales. En cam-bio, para otros ha representado el núcleo del autoritarismo, y el instru-mento de la conservación y transmisión de los típicos valores burgueses,
195 Cfr. Malinowski, Bronislaw, Sex and Represión in Savage Society, Londres, Routledge, 1927, y The Sexual Life of Savages in Noth-Western Melanesia, Londres, Routledge, 1929 (Vida sexual de los salvajes del nordeste de la Melanesia, Madrid, Morata, 1975).
196 Cfr. Kuper, A., o.c., pp. 169-170.

338
© Universidad de Deusto - ISBN 978-84-9830-383-4
con su doble moral: laxa para los varones, y estricta para las mujeres. Así mismo, en las sociedades agrícolas y rurales se daban familias amplias, de tipo clan, mientras que en las sociedades urbanas e industriales se va im-poniendo las familias nucleares.
Incluso, en la época más reciente, estaríamos asistiendo a una fuerte crisis del matrimonio y de la institución familiar, en la medida en que se extienden los divorcios, se prescinde de la institucionalización de la unión matrimonial, proliferando las denominadas «uniones de hecho». E igual-mente, asistimos a la legalización de los matrimonios homosexuales.
A pesar de estos elementos de crisis del matrimonio y la familia tradi-cional, podemos defender que la familia nuclear es una institución univer-sal de la cultura humana, aunque se dé en cada cultura y en cada momento histórico una gran pluralidad de formas y de funciones de la misma.
2.8.3. La formación de sociedades complejas
Sea cual sea lo que se opine sobre la universalidad o no del matrimo-nio y de las formas de la familia, el primer núcleo social del ser humano, al igual que el de los homínidos anteriores fue el núcleo familiar, en un sentido amplio. Esos núcleos familiares, en la medida en que se fueron multiplicando y ampliando, se dividirían y se expandirían por territorios cercanos, puesto que no siempre podrían sobrevivir en un mismo territo-rio limitado un núcleo muy numeroso de individuos. Sólo en situaciones ecológicas muy favorables podrían convivir en un pequeño territorio un núcleo poblacional numeroso.
Además, parece evidente que cada especie animal está capacitada para poder convivir con un número limitado de individuos, en función de las limitaciones de su cerebro para establecer relaciones con otros de sus congéneres197. Así, existe una estrecha relación entre la capacidad cere-bral y la dimensión del grupo social en el que se puede convivir198. A la hora de preguntarse por las ventajas que proporciona el aumento del cere-bro, es evidente que son muchas las respuestas que podemos dar. Y entre ellas está la de dotarnos de la capacidad adecuada para poder interactuar con los demás individuos de la propia especie. Hemos indicado, al hablar del origen de la conciencia y de la mente, que en el ser humano es más determinante, dentro de la lógica de la selección natural y de la lucha por la vida, la relación interpersonal que la relación con el entorno natural
197 Cfr. Waal, Frans de, La política de los chimpancés, Madrid, Alianza, 1993. 198 Cfr. Ayala, F. J./Cela Conde, C. J., Senderos de la evolución humana, o.c., pp. 433-
440.

339
© Universidad de Deusto - ISBN 978-84-9830-383-4
ecológico. De ahí que los humanos nacemos especialmente dotados con la capacidad de interpretar la mente de los demás, de anticiparnos a las in-tenciones y acciones de los otros199. Por eso estamos dotados de las deno-minadas neuronas espejo, recientemente descubiertas en los cerebros hu-manos y de algunos chimpancés200.
En el género humano se habrían dado dos momentos especialmente significativos del aumento del cerebro: el primero se da con la aparición del homo ergaster, sucesor del homo habilis, momento en el que se consi-gue pasar de un cerebro que poseía una dimensión aproximada de un ter-cio de la masa cerebral del hombre actual, hasta alcanzar los dos tercios de nuestro volumen; y el segundo momento se produce con la aparición de los neandertales y el homo sapiens actual, producido hace unos qui-nientos mil años. Ese crecimiento se debió, entre otras cosas, como ya vi-mos, a la incorporación a su dieta alimenticia de cantidades permanentes de proteínas y grasas animales, consecuencia de su capacidad de construir herramientas más refinadas y de cazar en grupos bien organizados.
Por tanto, cabe suponer que ya con el homo ergaster se produciría el fenómeno del nacimiento prematuro del ser humano, con la subsiguien-te necesidad de ser cuidado de forma especial por el entorno familiar, debiéndose dar ya desde este momento un reparto de roles sexuales, de-dicándose la mujer al cuidado de los hijos en los primeros años de vida. Y junto a esta influencia del crecimiento del cerebro en el reparto de roles sexuales, también la tuvo en el aumento de la complejidad social. Los ma-míferos que poseen sociedades más complejas, poseen también cerebros más grandes que los mamíferos similares que viven aislados201. Igual-mente, investigaciones de Aiello y Dunbar han mostrado que «entre las diferentes especies de primates el tamaño relativo del neocórtex respecto del resto del encéfalo está en función directa del tamaño de los grupos so-ciales que forman esas mismas especies. Sin embargo, no se ha encontra-do una relación similar entre el tamaño relativo del neocórtex y el tipo de vida, por lo que la teoría ecológica del origen de nuestra muy desarrolla-da inteligencia pierde fuerza respecto a la teoría social; no obstante, no hay que perder de vista que a partir de los primeros Homo los homínidos entraron en un nuevo nicho ecológico totalmente nuevo para los primates, el de carroñeros y cazadores, que también pudo favorecer el desarrollo intelectual»202.
199 Cfr. Humphrey, N., La mirada interior, Madrid, Alianza, 1993; Id., Una historia de la mente. La evolución y el nacimiento de la conciencia, Barcelona, Gedisa, 1995.
200 Rizzolatti, G./Sinigaglia, C., Las neuronas espejo, Barcelona, Paidós, 2006. 201 Cfr. Arsuaga, J. L./Martínez, I., La especie elegida, o.c., 217.202 Ibídem, 217.

340
© Universidad de Deusto - ISBN 978-84-9830-383-4
Por tanto, parece más que evidente que el desarrollo de nuestra inte-ligencia se produjo como consecuencia de tener que vivir en un ámbito social cada vez más complejo, en el que cada individuo tenía que poner a prueba su inteligencia para colaborar y competir con los otros compañeros de especie. Este juego de relaciones interpersonales obligaba a conformar tácticas y alianzas de unos con otros, tanto basadas en el interés propio y de sus parientes más cercanos, como en el altruismo y la colaboración en-tre grupos más amplios203. En este escenario de comportamientos es don-de se han desarrollado tanto las mentes altruistas como la denominada in-teligencia maquiavélica (A. Whiten y R. Byrne), que son dos alternativas de conducta posibilitadas por el fondo común social con el que está con-formada nuestra mente e inteligencia.
Con esta estructura mental se irían formando y complejificando las primeras sociedades humanas, formadas en una primera fase por grupos humanos emparentados por lazos de sangre y conformados por bandas de cazadores y recolectores, para después pasar, con la llegada del Neolítico, tal y como vimos en su momento, a comenzar una nueva vida basada en la ganadería y la agricultura. Este nuevo estilo de vida permitió el acceso a fuentes más abundantes y seguras de alimentación, posibilitando así el primer importante aumento demográfico del género humano y la apari-ción de las primeras sociedades complejas. Si durante los largos milenios del paleolítico los grupos humanos estaba formados por lazos cercanos de sangre, en las sociedades posteriores más numerosas las relaciones entre los que vivían juntos en las primeras aldeas ya no tenían una relación de sangre tan exclusiva y cercana.
Ya vimos lo referente a la formación de los primeros habitáculos y las primeras aldeas, y no vamos a repetirnos. La organización de las jefaturas y jerarquías dentro de los cazadores-recolectores se haría en función de la po-sesión de cualidades guerreras y de la capacidad para lograr la cohesión del grupo. Posteriormente, se formarían federaciones de bandas, sin que necesa-riamente se diera un jefe supremo que tuviera poder sobre todos. Convivían más o menos pacíficamente, no pudiéndose en muchos momentos detener los brotes de guerra entre ellos. En esos conflictos, unos clanes se unían con otros frente a terceros clanes, y en determinados momentos buscarían qui-zás el arbitraje de otros jefes cercanos. Es el modo como Evans Pritchard describe el estilo de convivencia de los nuer, en Sudán meridional204.
203 Cfr. Ferguson, R. Brian, «La naissance de la guerre», La Recherche Scientifique, 2004, n.º 373, 51-57.
204 Cfr. E. E. Evans-Pritchard, E., The Nuer: A Description of the Modes of Livelihood and Political Institutions of a Neolitic People, Oxford, Clarendon Press, 1940; cfr. Kuper, A., El primate elegido, o.c., cap. 9.º.

341
© Universidad de Deusto - ISBN 978-84-9830-383-4
Los modos de organización social de estos primeros cazadores-re-colectores serían de muy diversa índole, según los factores ecológicos y económicos, así como las tradiciones de cada uno, fruto de sus creencias y de las circunstancias históricas acumuladas205. Sobre estas formas primi-tivas de organización social es sobre las que los teóricos posteriores, entre ellos Hobbes, han proyectado un estado primero de naturaleza (de lucha de todos contra todos) del que se pasaría a otro de cultura, basado en un pacto de no agresión, para conformar los primeros elementos del estado. Pero ya hemos dicho que no hay por qué proyectar sobre esos momentos primeros ni una estructura demasiado uniforme, ni tampoco un esquema de agresión permanente, sino que se alternarían momentos de pactos y co-laboración, con otros de guerra y conflicto.
Marvin Harris, en Jefes, cabecillas, abusones206, indica una secuen-cia de la complejidad en tres fases: tribus en alianza, jefes que van con-formando grupos mayores, y, por último, estados más estables. La pri-mera etapa sería la propia de los cazadores-recolectores, la segunda se iría configurando en los primeros tiempos del neolítico, y las primeras ciudades-estado irían apareciendo entre hace 12.000 y 10.000 años. A par-tir de esos años, se produce una extraordinaria explosión cultural, fru-to de la adopción de la vida sedentaria, con la ganadería y agricultura, y de su correspondiente explosión demográfica, que obligó a reconfigu-rar los núcleos sociales, con la aparición de las primeras ciudades, de la forma en que conocemos por los testimonios escritos207. Ya hicimos referencia, al hablar de los primeros habitáculos primitivos, a los pri-mitivos asentamientos humanos en forma de aldeas, para desembocar en las excavaciones que descubren las primeras ciudades, como Jeri-có o Catal-Hüyük, descubierta en la zona este de Turquía por el inglés James Mellaart, de unos 9.000 años de antigüedad208. Las investigacio-nes arqueológicas de esta ciudad muestran que el desarrollo conseguido por los hombres que la habitaban era considerable, tanto en la técnica de construir y alinear las casas, como en sus herramientas y ajuares, el avance de la ganadería y agricultura, la utilización de vestidos, adornos y cosméticos, su organización social, así como en los aspectos más ideo-lógicos: enterramientos rituales de sus muertos, y restos de ritos religio-
205 Cfr. Johson, Allen W./Earle, Timothy, La evolución de las sociedades humanas,Barcelona, Ariel, 2003.
206 Madrid, Alianza, 1993. 207 Bertranpetit, J./Junyent, Cr., Viaje a los orígenes. Una historia biológica de la es-
pecie humana, Barcelona, Península, 2000, cap. 7, «La génesis de la diversidad humana actual».
208 Cfr. Clarke, R., o.c., pp. 190 y ss.; Hodder, Ian, «La división sexual en Catal-Hüyük», Investigación y Ciencia, 2004, n.º 330, 50-57.

342
© Universidad de Deusto - ISBN 978-84-9830-383-4
sos. Se ha encontrado también en ella el paisaje pintado más antiguo que se conoce: es una pared donde aparece representada la propia ciu-dad. También se han hallado pinturas que representan escenas de caza, con indudables motivaciones mágicas y religiosas. Es muy posible que se diera una importante organización religiosa, porque se han hallado restos de cultos basados en la representación de toros, así como otros re-lativos a la diosa de la fertilidad.
Más adelante, el ser humano sería capaz de alcanzar en su desarro-llo cultural otros logros, como son la alfarería, la metalurgia y la escritu-ra. Todos esos adelantos propiciarían la construcción de sociedades más complejas, dotadas de organizaciones estatales, y, más adelante, federa-ciones de estados e imperios. Pero eso ya nos aleja de las etapas primeras de la humanidad, que es donde nos tenemos que quedar y situar.
En conclusión, la investigación empírica, tanto sobre los restos fósiles como sobre las costumbres de las sociedades primitivas aún existentes, es posible que no nos permita nunca descubrir cual fue la primera y funda-mental forma de organización social y ciudadana. Es propio de la reali-dad humana la no existencia de un único modo de vida y de comportarse, puesto que su condición está abierta a múltiples posibilidades, aunque no a todas. De ahí que veamos en las diferentes épocas y ámbitos geográfi-cos múltiples formas de organización social, sin que se haya descubierto un principio de autoridad legítima aceptado en todas las tradiciones cultu-rales. No hay un sistema de derecho humano que se haya aceptado y re-conocido de modo universal, como para considerarlo un derecho natural propio de la naturaleza humana. Y aunque lo hubiera, podríamos perfecta-mente cambiarlo.
2.9. La aparición de la capacidad estética y de las obras de arte
La capacidad estética siempre ha sido considerada como una incon-testable cualidad natural del ser humano. Pero, al igual que hemos visto con otros aspectos de la cultura, en la actualidad hay quienes consideran que la estética también es una cualidad que se puede atribuir a ciertos animales209, en la medida en que sería una capacidad que permite a los seres vivos optar por determinadas preferencias que resultan más venta-josas en la lucha por la supervivencia. La cuestión estará en determinar si lo que algunos denominan por capacidad estética en el mundo de los
209 Cfr. Buskes, Chr., o.c., cap. 13.º, «Evolución y estética», pp. 361 y ss.; Dutton, De-nis, El instinto del arte. Belleza, placer y evolución humana, Barcelona, Paidós, 2010.

343
© Universidad de Deusto - ISBN 978-84-9830-383-4
animales es algo idéntico, o meramente análogo, a la capacidad estética de los humanos.
2.9.1. ¿Estética animal?
Como en otros aspectos del ámbito cultural, nos inclinamos a pensar que, aunque en el mundo de los animales se pueda atisbar algún indicio de sensibilidad estética, la capacidad de percibir y crear belleza es una cua-lidad exclusivamente humana, que surge en el proceso de humanización como el resto de las demás cualidades culturales.
Pero se discute si el arte es algo que se ha adquirido por aprendiza-je, fruto de la transmisión cultural de unas generaciones a otras, o es una cualidad natural, como parte de nuestra dotación genética. La respuesta ha de ser la misma que respecto al lenguaje y demás cualidades culturales: la capacidad estética es el resultado tanto de una dotación genética como de la educación cultural. La capacidad estética es algo que poseemos como parte de nuestra condición humana, pero no todos los humanos están do-tados de las mismas capacidades para producir obras artísticas, por lo que los más capaces son los que han sido reconocidos como artistas signifi-cativos por la sociedad y han transmitido sus conocimientos y sus obras a las generaciones posteriores, acumulándose a lo largo de la historia un acerbo cultural que constituye la herencia humana.
Ahora bien, quienes atribuyen también capacidad estética a los ani-males, defienden que la percepción de la belleza la recibimos genética-mente y resulta una cualidad que poseemos como resultado del proceso evolutivo, como instrumento de supervivencia. Estas características ge-néticas, nos indica Chr. Buskes, «se manifiestan en circuitos neuronales que, si se estimulan, causan placer o repugnancia al organismo»210. La conclusión que se deduce es que «todos los animales son “estetas” pues la experiencia de la belleza radica en la capacidad biológica antiquísi-ma de poder evaluar cualitativamente los estímulos sensoriales. (…). Por ello, la capacidad de percibir la belleza proviene de objetivos e intereses terrenales»211.
Como puede verse, es clave en esta discusión el concepto que tenga-mos de estética y de belleza. La cuestión a plantear es si hay cosas bellas en sí, o depende su belleza del observador. Lo normal es responder que existe lo bello porque existen seres humanos. Y lo mismo podríamos de-
210 Ibídem, p. 363.211 Ibídem, p. 363. La cursiva y las comillas son del autor.

344
© Universidad de Deusto - ISBN 978-84-9830-383-4
cir de la ética. Para otros, sin embargo, este modo de ver las cosas nos lle-varía a una concepción relativista, puesto que no expresaría nada sobre la realidad del mundo, sino un simple modo de ver la realidad por parte de los seres humanos. Chr. Buskes considera que la postura de Darwin (en Elorigen del hombre) se alinea con los que piensan que es la realidad objeti-va la que posee determinadas cualidades que las hacen más o menos ade-cuadas y apetecibles para que los animales se las apropien de cara a la lu-cha por la supervivencia212.
Para el filósofo de la Universidad de Buckingham, A. O’Hear213, este modo de ver las cosas cae en un inaceptable antropomorfismo, pues las elecciones de los animales por objetos o parejas sexuales supuestamen-te bellos de cara a la supervivencia son resultados de tendencias innatas y adaptativas. Se trata de respuestas preprogramadas, y eso no tiene nada que ver con la estética. O’Hear sigue con estos planteamientos las ideas ya presentadas por Kant, para quien un juicio estético no indica tanto cómo es la cosa en sí, sino cómo reacciona ante el objeto la persona que emite un determinado juicio estético. Y esos juicios estéticos tienen para Kant las características de ser desinteresados, universales y no cognitivos, sino situados en el ámbito de lo emotivo, de los sentimientos214.
De todos modos, O’Hear no coincide del todo con Kant, pues esta for-ma subjetivista de entender la belleza corta sus alas al no poder salir de un evidente relativismo. Su propuesta es una cierta forma de realismo esté-tico, consistente en afirmar que la belleza es en parte producto de la rea-lidad, al mismo tiempo que de la capacidad del sujeto de percibirla. Por tanto, la belleza tiene su consistencia objetiva, pero sólo es percibida por una mente dotada de la correspondiente capacidad para percibirla. Y esa capacidad estética no tiene por qué ser una estructura trascendental al es-tilo platónico, sino un modo de estar estructurada nuestra mente, fruto de la evolución biológica. De ahí que no tienen por qué ser nuestros juicios estéticos totalmente desinteresados, como pensaba Kant, sino que pueden tener claros intereses encaminados a dotarnos de mejor capacidad de su-pervivencia.
Esta es la postura de Chr. Buskes, quien opina que «el instinto y la es-tética son perfectamente compatibles. Así, pues, no hay motivo para negar a los animales un sentido de la belleza, y los deseos naturales pueden ser hermosos y agradables»215. Por tanto, los defensores de la capacidad esté-
212 Cfr. ibídem, pp. 364-366. 213 Cfr. O’Hear, A., Beyond evolution: human nature and the limits of evolutionary
explanation, Oxford, Oxford University Press, 1997. 214 Cfr. Bukes, Chr., o.c., pp. 366-369. 215 Ibídem, p. 372-373.

345
© Universidad de Deusto - ISBN 978-84-9830-383-4
tica de los animales, como Buskes, entienden que el origen del arte no hay que buscarlo en las diferentes especies que forman el género homo, sino en el reino animal, porque tampoco en este punto habría una diferencia esencial entre hombres y animales. Lo que sí aceptan estos autores es que los humanos hemos ampliado considerablemente nuestra capacidad estéti-ca a contextos nuevos con un nivel de complejidad inigualable. Pero eso no va en contra de la tesis por ellos defendida que considera que la expe-riencia de la belleza está asentada en la naturaleza humana, en su base ge-nética y biológica, cuyas expresiones previas se hallan ya en las especies animales anteriores. Por tanto, la belleza hay que situarla dentro del pro-ceso de adaptación al entorno y entenderla como un factor que posibilita y ayuda a la supervivencia216.
No vamos a extendernos más en la discusión de este punto, sino que nos tenemos que limitar a distinguir de nuevo entre la capacidad estéti-ca y las leyes internas de la misma, puesto que son dos aspectos diferen-tes, como tendremos ocasión de verlo en capítulos posteriores, cuando hagamos referencia a la relación entre biología y ética, distinciones que sirven muy pertinentemente para la relación entre biología y estética, y para cualquier dimensión del ámbito de la cultura. Según ello, una cosa es que la condición de posibilidad sea la biología, en la medida en que la capacidad estética de los humanos es resultado del proceso evolutivo, y otra cosa muy distinta es que sea un mero producto de la biología, y, por tanto, sea similar la relación del hombre y los animales con la belle-za. La capacidad estética es una cualidad de la naturaleza humana, pero cuando hablamos de naturaleza no podemos utilizarla de la misma for-ma referida a hombres y a animales, puesto que la naturaleza humana no se refiere sólo a lo genético-biológico, como sí ocurre en los animales. En los seres humanos, la dimensión genético-biológica ha dado de sí, por emergencia, una estructura comportamental nueva, en la que se sitúa la conciencia y la libertad217. Eso es precisamente la cultura: la supera-ción de los constreñimientos de la biología, sin abandonar la dimensión biológica, pero teniendo que completar con actos libres lo que no le ha dado la biología. De ahí que la idea de belleza no pertenece a una con-ducta biológica, sino a una estructura intelectual, cultural, que percibe
216 Cfr. ibídem, pp. 373 y ss. 217 Cfr. Dutton, Denis, El instinto del arte, o.c. En su excelente tratado sobre las bases
biológicas del arte en el ser humano, defiende con rotundidad la diferencia cualitativa entre el arte humano y lo que algunos consideran arte animal: «A pesar de que me considero un amante de los animales, me veo obligado a reconocer que no sirve de nada elevar los de-liciosos garabatos de los chimpancés a la condición de arte en el sentido estrictamente hu-mano (…)»: p. 21.

346
© Universidad de Deusto - ISBN 978-84-9830-383-4
realidades bellas y puede transformar la naturaleza para construir obje-tos naturales en artefactos útiles y bellos, y eso es el arte humano. Y eso es lo que aquí nos interesa, la cuestión del origen del arte, en su sentido estricto, entre los humanos.
2.9.2. Los inicios de los objetos de arte
La cuestión que nos tenemos que plantear es cuándo y por qué surgen los primeros atisbos de arte entre los humanos. Nunca sabremos por qué el hombre primitivo sintió la necesidad de pintar un animal en una gruta, o a esculpir un figurilla de piedra o de barro. ¿Expresaba la necesidad de transmitir un mensaje, una idea, o simplemente era la expresión gratuita de una tendencia lúdica? ¿O poseían tales actividades una simbología re-ligiosa o mágica? Nunca lo sabremos del todo, puesto que no nos dejaron mensajes escritos para explicarlo, y nuestras interpretaciones ante tales expresiones están inevitablemente impregnadas de proyecciones de nues-tro modo de pensar en la actualidad.
Nos quedamos maravillados ante la contemplación de las expresio-nes artísticas de las cuevas de Altamira o de Lascaux, y podemos caer en el error de pensar que ese arte rupestre tan extraordinario surgió de for-ma repentina, cuando la realidad es que, tanto el arte como cualquier otra expresión cultural, ha tenido también sus etapas anteriores, siendo estas ex-presiones más recientes el fruto acabado de una proceso que ha durado muchos miles de años218.
El arte es una expresión más de la capacidad humana de expresar ideas con símbolos. Precisamente consideramos como primeros humanos (homo habilis) a los sujetos capacitados para la fabricación y utilización de las primeras herramientas, hace dos millones y medio de años. Pero hay que distinguir entre una herramienta y otros objetos simbólicos, que serían más bien objetos que no poseen por sí mismos una utilidad prácti-ca. Ahora bien, este modo de entender el objeto simbólico tiene el incon-veniente de que no siempre sabemos la finalidad de un objeto. Aunque damos por hecho que el homo habilis poseía una intencionalidad a la hora de construir y utilizar sus herramientas.
Pero parece evidente que las herramientas del homo habilis eran ins-trumentos que se agotaban en el propósito de la utilidad para la que fueron construidas. En cambio, en las hachas bifaces, construidas más adelante por el homo erectus algunos estudiosos quieren ver una intencionalidad
218 Cfr. Clarke, R., o.c., cap. 8; Carbonell, E./Sala, R., o.c., cap. XII.

347
© Universidad de Deusto - ISBN 978-84-9830-383-4
estética, debido a la simetría con la que fueron fabricadas, sin dejar de poseer su correspondiente eficacia para cortar219. De ahí que Washburn y Lancaster defiendan que los bifaces simétricos achelenses constituyen el primer objeto bello facturado220. Pero no cabe duda de que esta afirma-ción es un tanto problemática, porque queda la duda de si esa simetría es-tética de los bifaces la vemos nosotros como bella o era esa la intención de los que los fabricaron. Puede que, aunque nosotros los encontremos bellos, los motivos que perseguía el homo erectus al fabricarlos fueran muy diferentes.
Los primeros bifaces descubiertos poseen una antigüedad que ronda el millón y medio de años, según M. D. Leaky221. Si comparamos estas supuestas expresiones artísticas con las pinturas rupestres de finales del Paleolítico Superior, la cuestión que se plantean los investigadores es si se ha dado una continuidad progresiva entre esas dos épocas, o más bien las pinturas rupestres son consecuencia de una explosión artística inesperada.
a) Las tesis gradualistas frente a las rupturistas222
Desde los primeros bifaces hasta el arte rupestre más reciente, el lapso de tiempo es muy grande, pudiéndose organizar esa evolución, según la tesis gradualista, en tres momentos o épocas: el primer período de casi un millón de años de los bifaces achelenses, surgidos en África y extendidos después a Europa; un segundo período, de unos 100.000 años, coinciden-te con la glaciación de Würm, la última, en la que se fabrican utensilios en los que algunos autores quieren ver una intencionalidad plasmada de simbolismo, incluso religioso; y la tercer etapa, entre los 40.000 y los 10.000 años antes de nuestra era, en la que se han descubierto de forma masiva múltiples objetos de incontestable sentido estético. Pero, así como nadie discrepa sobre el contenido estético de los objetos de la tercera eta-pa, no ocurre lo mismo con las dos etapas anteriores. En estas apreciacio-nes tan diferentes se muestran las dos posturas en conflicto: la gradualista y la que defiende que el arte ha surgido de forma explosiva en épocas más bien recientes.
219 Cfr. Cela Conde, C./Ayala, F. J., Senderos de la evolución humana, Madrid, Alianza, 2001, p. 461 y ss.
220 Cfr. Washburn, S. L./Lancaster, C. S., «The Evolution of Hunting», en R. B. Lee y I. de Vore (eds.), Man the Hunter, Chicago, III, Aldine Publishing Co., 1968, pp. 293-303.
221 Cfr. Leaky, M. D., «Cultural Patterns in the Olduvai Sequence», en K. W. Butzer y G. L. Isaac (eds.), After the Australopithecines, The Hague, Mouton, 1975, pp. 477-493.
222 Cfr. Cela Conde, C./Ayala, F. J., o.c., pp. 463 y ss.

348
© Universidad de Deusto - ISBN 978-84-9830-383-4
a.1) Las tesis gradualistas
El modelo gradual entiende que se da arte desde los bifaces achelen-ses, desarrollándose estas manifestaciones artísticas de modo gradual has-ta las fases posteriores más excelsas. De modo que, para esta postura, no tendría sentido hablar de un origen del arte, porque la frontera sería muy difusa. Los primeros atisbos de arte se darían con la aparición de elemen-tos ornamentales, que serían la propia elección de los materiales para ha-cer herramientas y la misma fabricación de los bifaces. En relación a lo primero, algunos autores como Oakley223 consideran una expresión ar-tística la presencia de pequeños fósiles dentro de las piedras con las que se ha construido algún bifaz: aunque otros, como Noble y Davidson224,entienden que ese dato no es significativo puesto que encontrar fósiles en los pedernales es más bien un hecho más bien común, no tanto un fenó-meno excepcional.
Otra muestra de este arte primitivo sería las perforaciones y grabacio-nes en hueso y piedra realizadas por neandertales. Pero los críticos con-sideran que tales hechos no aportan una clara intencionalidad artística, pudiéndose interpretarlos como consecuencia de fenómenos naturales225.Pero hay algunos casos en los que las incisiones en huesos tienen todos los visos de ser acciones intencionales, como es el caso de algunas piezas de la cueva de Bilzingsleben (Alemania), en las que se muestran unos di-bujos geométricamente dispuestos, obra del homo erectus y con una anti-güedad de 350.000 años. Otro ejemplo similar es un artefacto encontrado en un yacimiento de Quneitra (Golán, entre Israel y Siria), consistente en una piedra triangular de pedernal de cultura musteriense, y de una anti-güedad de unos 54.000 años. Esta pieza, según los estudios realizados por Marschack226, tiene todos los ingredientes para atribuirle una intenciona-lidad estética. Aunque hay quienes interpretan estas incisiones como mar-cas hechas al azar, los defensores de la intencionalidad de las incisiones entienden que «la relación que existe en este caso entre la distribución espacial de las marcas y el espacio disponible en el hueso (en el caso de Binzingsleben) hacen suponer que no medie una intencionalidad»227. Y lo mismo cabría decir de la lámina de Quneitra.
223 Cfr. Oakley, K., Emergence of higher thougt 3.0-0.2 Ma B.P., Phil. Trans. R. Soc. Lond., 1981, B292, 205-211.
224 Cfr. Noble, W./Davidson, I., Human Evolution. Language and Mind. Cambridge, Cambridge University Press, 1996.
225 Cfr. Cela Conde, C./Ayala, F. J., o.c., pp. 464-465. 226 Cfr. Marshack, A., «A middle paleolitihic Symbolic Composition from the Golan
Heights: The earliest known depictive image», Current Anthropology, 1996, 37, pp. 357-365. 227 Cela Conde, C.,/Ayala, F. J., o.c., 467-468.

349
© Universidad de Deusto - ISBN 978-84-9830-383-4
La conclusión que sacan Cela Conde y Ayala de todo ello es que todos estos objetos supuestamente dotados de intencionalidad estéti-ca cumplen condiciones más que suficientes («son muy antiguos (desde 350.000 a 54.000 años), se deben a una acción antrópica, carecen de utili-dad, muestran una disposición geométrica que indica una cierta intención estética»)228 para considerarlas como las primeras expresiones artísticas de los humanos. Claro que se trata de expresiones minoritarias, y se nece-sitaría encontrar fósiles semejantes en otros lugares del planeta. Una nue-va muestra de un objeto artístico es un arpón tallado en hueso, enconntra-do en Katanda (Zaire), de una antigüedad entre 75.000 y 90.000 años. La duda que siempre se presenta en estos casos es por qué no se han encon-trado otras muestras artísticas semejantes en sitios cercanos.
Una muestra de arte ya cercano a las fechas de las grandes manifes-taciones del arte rupestre clásico es el caballo tallado en marfil y encon-trado en la cueva de Vogelherd (Alemania), muestra del arte auriñacense, con una antigüedad de 32.000 años. En este caso no hay ninguna duda de que la intención de su autor era diseñar un caballo para servir de colgante de un collar229. Este resto arqueológico es un claro precedente de las ma-ravillosas expresiones artísticas del Paleolítico Superior, momento en el que se produce un gran salto de calidad estética. Parece evidente que al-rededor de los 40.000 años antes de nuestra era se produjo una explosión cultural, que no sólo se expresa en el área estética, y que resulta evidente que fue consecuencia de un cambio cognitivo fruto de un crecimiento en el volumen y la complejidad de la estructura cerebral de los humanos, y que traería como consecuencia una proliferación de muestras artísticas de una elevado nivel de calidad y complejidad.
a.2) La tesis rupturista
En este salto cualitativo de las capacidades cognitivas de los huma-nos se apoya el modelo explosivo en la aparición del arte. Es cierto que la aparición de numerosas tallas y pinturas naturalistas a partir de los 38.000 años es un dato contundente. Y no sólo hallamos estas expresio-nes tan impresionantes en las cuevas rupestres, sino que «los objetos de uso cotidiano se encuentran también profusamente decorados con motivos geométricos y con representaciones figurativas cuyo nivel de interpreta-ción naturalista del mundo exterior alcanza unas cotas similares a las que pueden expresar hoy los mejor dotados de nuestros artesanos»230.
228 Cfr. ibídem, 468.229 Cfr. ibídem, 469-470. 230 Ibídem, 471.

350
© Universidad de Deusto - ISBN 978-84-9830-383-4
No nos vamos a detener en estudiar las etapas ni los rasgos específi-cos de las más representativas expresiones del arte de esta época, ni tam-poco sobre los usos y finalidades que se hallaban en la cabeza de los ar-tistas que las diseñaron y realizaron. Lo que nos interesa resaltar aquí es que, aunque «la violenta y extendida aparición del arte figurativo de gran colorido en las cuevas europeas a la vez que se produce una proliferación de objetos decorativos fuera del continente (como los de Enkapune Ya Muto, Kenia) parecen indicar un fenómeno repentino», también hace pen-sar a los defensores de la teoría gradualista que «ese alto nivel técnico y esa muestra de un sentido estético profundo son también interpretados por autores al estilo de Mellars o Van como indicios de que una capacidad tan desarrollada tuvo que evolucionar durante un cierto tiempo»231.
La tesis del origen explosivo del arte da por hecho que la capacidad artística, en su sentido estricto, se limita exclusivamente al homo sapiensreciente, a los cromañones, pero esta afirmación se pone en cuestión con el descubrimiento de muestras artísticas avanzadas entre los neanderta-les232. Aunque es cierto que estas muestras artísticas bien pueden inter-pretarse como un préstamo cultural tomado de los cromañones. Esto se advierte en el yacimiento de la Grotte du Rene, en Arcy-sur-Curte (Fran-cia), atribuido a los neandertales233, estudiado por Hublin y colaboradores, para quienes los artefactos encontrados en este yacimiento son un présta-mo cultural. Pero otros intérpretes (d’Errico y colaboradores234) conside-ran que esos artefactos son obras de los neandertales, a quienes atribuyen una evolución cultural independiente y propia. De la misma opinión es Bahn235, partidario de atribuir a los neandertales una conducta simbólica sofisticada y moderna, y no meros usuarios de algunos elementos ideados por los cromañones.
Pero también es cierto que si el yacimiento de Arcy-sur-Curte no pue-de interpretarse de forma rotunda como un préstamo cultural, algunos ya-cimientos cercanos encontrados en Croacia, como el de Vindiya, habitado por neandertales, y el de Velika Pecina, por humanos modernos, habitados en tiempos coincidentes, parecen indicar que los objetos hallados en am-
231 Ibídem, 472-473. 232 Cfr. Arsuaga, J. L., El collar del neandertal, Madrid, Temas de Hoy, 1999.233 Cfr. Hublin, J. J., et al., «A late neanderthal asociated with upper paleolithic artefacts»,
Nature, 1996, 381, 224-226. 234 Cfr. D’Errico, F., et al., «Neanderthal acculturation in western Europe? A critical
review of the evidence and its interpretation», Current Anthropology, 1998, 39, suplement, 1-44.
235 Cfr. Bahn, P. G., «Neanderthal emancipated», Nature, 1998, 394, 719-720. Cfr. Cela Conde, C./Ayala, F. J., o.c., 473-475; Arsuaga, J. L., El collar del neandertal, Madrid, Te-mas de Hoy, 1999.

351
© Universidad de Deusto - ISBN 978-84-9830-383-4
bos yacimientos se deben o bien a imitación o a intercambio cultural. Esto mostraría que los neandertales eran capaces de construir y usar elementos decorativos, poseyendo capacidad para apreciar a dichos artefactos como cosas bellas236. La verdad es que las prácticas culturales de los neanderta-les eran bastante parecidas a nosotros, en la medida en que podían en-terrar a sus muertos y fabricar objetos con finalidades artísticas, y a veces hacían estas cosas, aunque no de forma continua.
La cuestión es por qué se dio un salto tan grande en estos elementos culturales con la desaparición de los neandertales. Como indican Cela Con-de y Ayala, «incluso si aceptásemos que se trata de un asunto relacionado con la emergencia de una especie nueva, es decir, que la configuración ge-nética de los Homo sapiens les concede, sin más, unas capacidades cogni-tivas superiores, aún quedarían problemas pendientes que resolver»237. El más importante hace referencia al hecho de que las fechas de la aparición de nuestra especie (entre los 150.000 y 100.000 años) son muy anteriores a la explosión del arte del Paleolítico Superior. Por tanto, no se explica de este modo sin más la reciente explosión de estas expresiones artísticas.
b) Una visión gradual más completa
La forma más razonable de explicarlo nos lleva a una interpretación más amplia de la tesis gradualista. Son varios los autores que han pro-puesto la idea de que el origen del arte está íntimamente relacionado con el origen del lenguaje. La base de esta hipótesis se halla en que «el dibujo necesita cierta comunicación previa y luego ese dibujo transforma la co-municación en lenguaje»238. Para ciertas autoras como Sally McBrearty y Alison Brooks239, esta supuesta revolución cognitiva tiene que interpretar-se de otra manera, entendiéndola como la expresión de un largo proceso de avances que empezó mucho antes, teniéndose que interpretar la ruptura entre la supuesta explosión del Paleolítico Superior no como un nacimien-to casi de la nada, sino como consecuencia de la ausencia de hallazgos de restos culturales o fósiles de épocas anteriores. Y para ello hay que tener en cuenta los descubrimientos realizados en zonas fuera de África. Estas autoras, por tanto, proponen la existencia de un proceso más gradual que se remonta a épocas muy antiguas.
236 Cfr. Cela Conde, C.,/Ayala, F. J., o.c., 474.237 Ibídem, 475. 238 Ibídem, 476. 239 Cfr. McBrearty, S./Brooks, A. S., «The revolution that wasn’t: a new interpretation
of the origin of modern human behavior», J. Hum. Evol., 2000, 39, 453-463.

352
© Universidad de Deusto - ISBN 978-84-9830-383-4
En su propuesta se distinguen varios momentos dentro de la historia de la especie homo sapiens, entre los sapiens arcaicos y los más moder-nos, considerando incluso la posibilidad de admitir varias especies en el lapso existente entre los neandertales y los sapiens más modernos. Pero no admiten la tesis de la ruptura en la capacidad artística que supondría la tesis de la explosión aparecida en la fase reciente del hombre moderno. Para ellas, «si ésta se debe a una “revolución cognitiva” propia del Homosapiens, las primeras evidencias acerca de un cambio brusco en las mani-festaciones culturales deberían proceder de África —que es donde se ori-ginan los humanos de aspecto moderno— y tener la edad aproximada que se atribuye a nuestra especie, es decir, al menos 100.000 años»240. Pero es evidente que las cosas no son así.
Se necesita realizar una reinterpretación de los registros fósiles afri-canos, que hasta ahora se han interpretado desde los esquemas de los yacimientos europeos. Esa adecuada interpretación nos llevará a con-cluir que el Homo sapiens mostró diferencias cognitivas desde el princi-pio. Según ello, se advierte que los yacimientos de África muestran ele-mentos de decoración personal mucho más antiguos que los europeos. Lo mismo podemos decir sobre la utilización de pigmentos coloreados para la decoración de objetos, cuyas primeras muestras poseen una anti-güedad de entre 250.000 y 300.000 años241. Pero en algunos yacimien-tos de una antigüedad de millón y medio de años (Gabeb, Etiopía), se han encontrado fragmentos de basalto que, al ser raspados, producen pigmento rojo. El que no hayan aparecido más objetos pintados entre esta fecha y la de los objetos más antiguos hallados, se puede explicar con la tesis de que el primer objeto de decoración fue el cuerpo huma-no, lo que explica que no se hayan hallado evidencias fósiles de obje-tos pintados. Sólo posteriormente se procedió a pintar otros objetos con motivos decorativos.
Si tenemos en cuenta cómo van pintados los humanos pertenecientes en la actualidad a grupos de cazadores-recolectores, advertimos que estas pinturas sobre los cuerpos tienen diversas funciones simbólicas relaciona-das con el orden jerárquico, más allá de intencionalidades decorativas242.Todos estos argumentos llevan a sacar la conclusión de que resulta más evidente y razonable la tesis gradualista, remontándose el origen del arte a las épocas primeras de la aparición de nuestra especie en África, con-secuencia de una coevolución de otros factores que hicieron progresar la cultura humana. Y teniendo en cuenta estas primeras manifestaciones ar-
240 Cela Conde, C./Ayala, F. J., o.c., 477.241 Cfr. ibídem, p. 479. 242 Cfr. ibídem, pp. 479-480.

353
© Universidad de Deusto - ISBN 978-84-9830-383-4
tísticas, la explosión del arte europeo de la época del Paleolítico Superior se explica como la fase final de este largo proceso gradual. Así, «los mu-rales del sur de Francia y el norte de España no son sino la manifestación final de las nuevas capacidades cognitivas. Las pinturas magníficas que allí aparecen pueden obedecer tanto a nuevos rituales como a la posibi-lidad de ocupar las cavernas por la desaparición de otros animales capa-ces de disputárselas a los humanos, pero no tienen por qué indicar ningún cambio mental de último momento»243. Esta explicación parece más razo-nable y más de acuerdo con el conjunto de las evidencias arqueológicas de que hoy disponemos.
2.10. Los primeros vestigios de la religión y de la moral
El origen de la religión es un problema tan complejo como cualquiera de los anteriores aspectos de la cultura humana estudiados con anteriori-dad. Al igual que en el terreno de la estética, del lenguaje y demás capa-cidades humanas, tenemos que distinguir entre la capacidad y los conteni-dos internos de una dimensión cultural.
En relación a la capacidad de plantearse el problema de la religión, tenemos que afirmar que es resultado, como todas las demás capacida-des culturales, del proceso evolutivo: fruto de mutaciones genéticas que, a su vez, han generado la capacidad cerebral de abrirse a la pregunta por el fundamento de la realidad y el sentido de lo todo lo que hay. En la actuali-dad, van apareciendo libros que tratan de localizar la base genética y cere-bral de las creencias y de la religiosidad244.
Pero lo que aquí nos interesa es atisbar, en el proceso histórico de la aparición de los primeros seres humanos, los vestigios del inicio de las creencias y de las costumbres religiosas. De nuevo nos encontramos con la dificultad radical de ponernos en el lugar de los hombres primitivos para adivinar sus creencias y si tenían la capacidad de plantearse la pre-gunta por el sentido de todo, el fundamento de la realidad y la apertura a la dimensión de la trascendencia, características de la religiosidad245. Nos tenemos que limitar a referirnos a los vestigios culturales que mostrarían tal forma de pensar.
243 Ibídem, p. 480. 244 Cfr. Hamer, Dean, El Gen de Dios, Madrid, La Esfera de los Libros, 2006; Alper,
Matthew, Dios está en el cerebro, Barcelona, Granica, 2008; Rubia, Fr. J., La conexión di-vina. La experiencia mística y la neurobiología, Barcelona, Crítica, 2003.
245 Cfr. Armstrong, Karen, En defensa de Dios. El sentido de la religión, Barcelona, Paidós, 2009, cap. 1.º, «El homo religiosus», pp. 27 y ss.

354
© Universidad de Deusto - ISBN 978-84-9830-383-4
a) Los primeros enterramientos
El camino considerado normalmente como el adecuado para aden-trarse en esta investigación es el de los enterramientos. Es la primera y más segura muestra de una inquietud del ser humano sobre lo trascenden-te o el más allá, o simplemente la cuestión sobre el sentido de la vida y la dignidad de la vida humana que nos acontece ante el trauma que supone la muerte de un ser querido, o de un semejante. Lo mismo que en otros ámbitos culturales similares, nos preguntamos si esta sensibilidad ante la muerte es un elemento específico de los humanos, o también se daría en algunas especies animales más inteligentes. La reacción de la mayo-ría de los animales ante los cadáveres de sus congéneres es muy diver-sa: desde no tenerlos en cuenta y abandonarlos, sin más, hasta comerlos. Algunos estudiosos de las costumbres de los grandes simios han querido observar en ellos una cierta conducta especial ante la muerte de un com-pañero de especie, quedándose ante él pensativos, cubriéndolos con ho-jas, etc.246. Pero nos situamos siempre en la misma discusión: peligro de proyección antropomórfica, y dificultad en marcar una línea claramente divisoria entre los atisbos de las especies más dotadas y nosotros. En esto también cabe conjugar la presencia entre los primates de comportamientos que apuntan a la cultura humana con una ruptura cualitativa con la apari-ción del género humano.
Seguimos, por tanto, defendiendo que se da una diferencia cualitativa entre los animales y los humanos, y nos vamos a centrar, por ello, en los datos paleoantropológicos sobre los primeros atisbos de enterramientos humanos con sus también primeras muestras de preocupación religiosa.
Parece evidente que, aunque no sepamos ni tengamos restos ni prue-bas evidentes de ello, «todas las comunidades a lo largo de la historia han demostrado un respeto a los muertos, bien sea mediante la inhuma-ción y, por lo tanto, su conservación en el subsuelo, o bien mediante in-cineración, ceremonia que suele acabar con el depósito subterráneo de las cenizas»247. Y en testimonios escritos de las diferentes tradiciones culturales, como es el caso de la Ilíada de Homero y textos de otras cul-turas, se nos dice que matan y entierran con el difunto diversos restos de
246 Cfr. Clark, R., El nacimiento del hombre, o.c., pp. 110-111, hace referencia al testi-monio de Jane Goodall sobre la lucha entre dos perras, en la que una va matando a los ca-chorros de la otra hasta que, cuando sólo le quedaba uno con vida, ésta se sometió y se dio por vencida. ¿Demuestra esto que la perra poseía una idea del valor de la vida y una cierta idea de la muerte, o era sólo un mero instinto materno de defensa de los hijos, no más allá de la idea que del altruismo paterno-materno, como inversión de los genes en los hijos, nos hablan los sociobiólogos? Nos inclinamos por esta segunda interpretación.
247 Carbonell, E./Sala, R., Planeta humano, o.c., p. 209.

355
© Universidad de Deusto - ISBN 978-84-9830-383-4
animales248 Estas costumbres se descubren ya en muchos enterramientos de los hombres primitivos.
Durante mucho tiempo se consideró que los primeros humanos que tenían por costumbre enterrar a sus muertos dando a esa costumbre un cierto significado religioso pertenecían a la especie del homo erectus.En concreto, se aportaba como prueba de ello el yacimiento de Zuku-dián, cercano a Pekín, perteneciente al denominado Sinanthropus peki-nensis (hoy adscrito al homo erectus), del que hablamos al referirnos al uso y dominio del fuego, y en cuyas investigaciones participó el paleon-tólogo jesuita Teilhard de Chardin. Allí se habrían encontrado, al pare-cer, evidencias de enterramientos rituales y del uso del fuego. Incluso se quiso ver también evidenciada la costumbre de ciertas comidas rituales, en las que se comía parte de la masa encefálica de los fallecidos tras ha-ber quebrado intencionadamente sus cráneos249. Pero ya dijimos en su momento, que, dada la pérdida de los restos de este yacimiento antes de la segunda guerra mundial y el superado nivel técnico de los estudios realizados allí, no resultan en la actualidad válidas sus problemáticas aportaciones.
Las tesis más prudentes y conservadoras solían retrasar hasta la época de los neandertales las fechas más antiguas de restos seguros y fiables de enterramientos intencionales entre los humanos. Pero los descubrimien-tos en la Sierra de Atapuerca, en concreto en la denominada Sima de los Huesos, han cambiado bastante las cosas250. En este yacimiento se nos mostraría que los humanos, hace ya más de 300.000 años, «preservaban a sus muertos y los protegían de la acción de los carnívoros»251. Se trata de cavidades de diversas dimensiones, algunas de ellas pequeñas y ocul-tas, pero otras más grandes y dedicadas a diversas actividades, en las que se amontonaban restos de humanos. El descubrimiento de estas cavidades y el estudio de sus restos, en opinión de dos de sus investigadores, «ha cambiado radicalmente el panorama y la percepción que teníamos hasta entonces del tratamiento de los muertos»252. Entrando por la Cueva Ma-yor se llega a la Sima de los Huesos, de unos doce metros de profundi-dad, donde se han descubierto restos de al menos treinta y tres humanos de ambos sexos, de edades diversas, junto con restos de diversos anima-les, resultando por ello «la necrópolis más antigua de la historia, con más
248 Cfr. ibídem, pp. 209-212.249 Cfr. Clarke, R., o.c., p. 111. 250 Cfr. Arsuaga, J. L., «Los hombres fósiles de la sierra de Atapuerca», Mundo Cientí-
fico, 1994, n.º 143, 167-168; Carbonell, E./Sala, R., o.c., 213 y ss. 251 Carbonell, E./Sala, R., o.c., 213.252 Ibídem, p. 213.

356
© Universidad de Deusto - ISBN 978-84-9830-383-4
de trescientos mil años de edad»253. Los diversos trabajos realizados sobre estos restos permiten concluir de forma razonable que fueron llevados allí de forma intencionada. «Sin embargo, no se trata de un enterramiento pro-piamente dicho: no fue construida ninguna fosa ni los cadáveres presentan disposición especial alguna. En resumen, no existió ningún sepelio, sólo la acumulación de los despojos en el fondo de una sima que los agentes naturales se encargaron de sepultar»254.
Vemos, por tanto, que entre estos humanos se daba una clara separa-ción entre el terreno donde se desarrollaban las diversas actividades de los vivos y la zona reservada a la acumulación de restos mortales. Se tra-ta, pues, de una especialización de espacios y mundos específicos entre los vivos y los muertos. Este es uno de los elementos que van a diferen-ciar los enterramientos de esta especie, homo heidelbergensis, y los de los neandertales. Estos enterraban a sus muertos en los mismos sitios donde habitaban, cosa que no ocurre en el caso de la especie anterior255.
Analizados los restos humanos allí encontrados, llama la atención que la mayoría de ellos sean esqueletos de niños, y sólo dos de más de treinta años. Las cuestiones que se plantean los investigadores son dón-de se enterraron a los demás y quién los enterró. Aunque no se sabe res-ponder a estas cuestiones, parece claro que se produjo allí una selección de los individuos arrojados a esas cavidades. Es evidente que «la Sima de los Huesos es el único conjunto del Paleolítico Inferior que presenta cla-ramente un tratamiento de los muertos»256. En épocas posteriores, las sepulturas abundan. La más antigua, después de la de Atapuerca, es la de Mougharet-es-Skhul, en el Monte Carmelo, y su datación es ya de 120.000 años. Y en fechas posteriores las tumbas encontradas pertenecen en la mayoría de los casos a los neandertales, conformando la época que se denomina el Paleolítico Medio.
Las sepulturas de esta época se hallan situadas en dos grandes áreas. La primera abarca una extensión entre el oeste de Europa y el extremo oc-cidental de Asia. Y la segunda, en Oriente Medio y Próximo. Fuera de es-tas áreas, sólo se han hallado dos casos: el de Border Cave, en Sudáfrica (enterramiento infantil de hace unos 80.000 años), y en Tamasa I (Egipto), la tumba de un anciano, hace unos 50.000 años. Parece una característica común de estos enterramientos el hecho de que se hallan situados en cue-vas y abrigos, y nunca en terreno abierto257.
253 Ibídem, p. 215. 254 Ibídem, p. 215. 255 Cfr. ibídem, p. 215 y ss. 256 Ibídem, p. 217. 257 Cfr. ibídem, p. 217.

357
© Universidad de Deusto - ISBN 978-84-9830-383-4
Ya hemos dicho que la mayoría de los enterramientos del Paleolítico Medio pertenecen a los neandertales. Hay dos casos que escapan a esta regla: Djebel Qafzeh y Moughret-es-Skhul, ambas en el corredor de Pa-lestina, y pertenecientes a individuos de nuestra especie. «Hay que desta-car que ninguna de ellas presenta características especiales, por lo que no existen diferencias culturales perceptibles entre los grupos más antiguos de Homo sapiens y los Neandertales»258. Eso hace concluir que no había grandes diferencias en su comportamiento, ni tampoco en el modo de con-cebir la realidad y quizás de su concepción religiosa.
Pero en esta época, sí se da un cambio esencial del comportamiento, en relación al Paleolítico Inferior, respecto al modo de enterrar y a la con-cepción que se tenía sobre la muerte y el sentido de la vida. Los restos se hallan depositados en un lugar expresamente preparado para ello, en una cavidad dentro de la cueva. Así mismo, se advierte que los cadáveres han sido colocados en una postura estudiada, parecida a la postura fetal («con las piernas plegadas y un brazo, el derecho, plegado igualmente en ángulo recto respecto al cuerpo»), y acompañado de alimentos y objetos fami-liares del fallecido. Esto hace pensar en la existencia de un ritual de en-terramientos, «un ceremonial fúnebre, en un acontecimiento que se consi-deraba importante en la vida de los hombres, que rompía claramente con la indiferencia que, hasta aquel momento, parecía haber sido el comporta-miento regular frente a la muerte»259.
Respecto a los restos encontrados junto al cadáver, como huesos de animales y objetos diversos (herramientas, adornos, restos de polen, con-chas marinas, etc.), pueden hacerse muchas interpretaciones, pero es fácil caer en la equivocación de proyectar nuestro modo de entender las cosas sobre las costumbres de los hombres de esa época260. Las interpretaciones sobre la intencionalidad de estos enterramientos se hallan divididos entre los que les atribuyen una intencionalidad religiosa trascendente, y la de quienes ven en ellos «una respuesta funcional a la necesidad de desem-barazarse de los cuerpos muertos»261. Ahora bien, la presencia de objetos como astas de gamo y mandíbula de jabalí, presentes en los dos enterra-mientos que hemos indicado pertenecen al homo sapiens, parece indicar la presencia de una simbología que apunta a una intencionalidad trascen-dente. Pero hemos de ser conscientes de que no tenemos la seguridad de interpretar adecuadamente el significado simbólico de dichos objetos, ya
258 Ibídem, p. 218. 259 Clarke, R., o.c., p. 105. 260 Cfr. Clarke, R., o.c., pp. 105 y ss.; Carbonell, E./Sala, R., o.c., pp. 217 y ss.; Cela
Conde, C./Ayala, Fr. J., o.c., 455 y ss. 261 Cela Conde, C./Ayala, F. J., o.c., pp. 457-458.

358
© Universidad de Deusto - ISBN 978-84-9830-383-4
que son meras proyecciones nuestras las referencias a símbolos, creencias o ceremonias de enterramiento a partir de tan escasos restos fósiles262.
Hay también algunos datos de gran interés sobre los enterramientos de esa época, como el hallazgo de restos de un recién nacido, de menos de un año de vida; e incluso dos tumbas de fetos, de ocho y siete meses respectivamente, lo que hace pensar que «en las comunidades neander-tales los beneficios del entierro y los rituales inherentes a él no eran res-tringidos, sino que se hacían extensivos a toda la población, incluyendo los fetos»263. Además, en algún caso se han hallado tumbas dobles, cosa más bien rara en esta época, a diferencia del Paleolítico Superior cuando las tumbas múltiples son comunes y numerosas264. Igualmente es intere-sante destacar que determinadas cuevas dedicadas a los enterramientos se utilizan de modo reiterativo, con lo que se supera la idea del nomadismo de estos grupos, restringiéndose sus movimientos más bien a zonas res-tringidas, dedicándose determinados recintos a depositar los cadáveres265.En resumen, parece que «la inhumación debe ser considerada como un fe-nómeno habitual y extendido en el Pleistoceno Superior»266. En esa épo-ca, las inhumaciones son fosas donde se depositaban varios cadáveres, en momentos sucesivos, cubriéndose muchas de esas fosas con montones de piedras encima. «Esta práctica, no sólo constituía una forma de proteger los cadáveres y separarlos, sino que debió de permitir durante un tiempo la identificación del lugar del sepelio»267.
En épocas posteriores, durante el Neolítico268, la riqueza y abundan-cia de tumbas halladas por los arqueólogos nos muestran que el culto a los muertos se fue consolidando como un elemento fundamental de la cul-tura humana, que hace decir a Unamuno que el ser humano es una «ani-mal guarda muertos». Pero, como conclusión, «no parece plausible, hoy por hoy, llevar más allá de Homo heidelbergensis y de los 300.000 años de antigüedad el complejo comportamiento que hemos ideo describien-do. Por lo que sabemos, Homo heidelbergensis inventó el mundo de los muertos»269.
262 Cfr. ibídem, p. 458. 263 Carbonell, E./Sala, R., o.c., p. 223. 264 Cfr. ibídem, p. 223. 265 Cfr. ibídem, pp. 225-226. 266 Ibídem, p. 226. 267 Ibídem, p. 227. 268 Cfr. Gallay, Alain, «El hombre neolítico y la muerte», en VV.AA., Orígenes del
hombre moderno, Barcelona, Prensa Científica (Libros de Investigación y Ciencia), 1993, pp. 124-133; Leroi-Gourhan, André, «Religión y hombres prehistóricos», Mundo Científi-co, 2000, n.º 214, pp. 57-59.
269 Carbonell, E./Sala, R., o.c., p. 239.

359
© Universidad de Deusto - ISBN 978-84-9830-383-4
b) Las primeras creencias (¿La primera «metafísica»?)
Todos estos rasgos intencionales descubiertos en los enterramientos, hacen concluir a los investigadores que «nos encontramos ante los pri-meros indicios de una mitología, de un imaginario que trata de sobrepa-sar, por medio de unos actos repletos de simbolismo, aquellos puramente utilitarios que consistían, desde hace un millón de años, en fabricar armas y utensilios de piedra y de hueso. Y así tenemos los primeros indicios de este esfuerzo del hombre por crear lazos entre los seres, incluso más allá de la muerte»270.
Estas conclusiones, que se ven claramente presentes a partir de los en-terramientos de los neandertales, no significa que no existían en épocas muy anteriores. Pero no tenemos evidencias fósiles sobre ello, aunque es ló-gico pensar que desde muy temprano el ser humano se plantearía, ante el hecho doloroso e inevitable de la muerte, las grandes preguntas sobre el sentido de la vida y de todo que tales experiencias suscitan de modo inevitable. La experiencia de la muerte pone en crisis el sentido de la vida, y plantea cuestiones sobre el sentido del tiempo, la previsión del fu-turo y la existencia de una vida más allá de ésta. Como indica R. Clarke, «tal vez esta idea nueva de la muerte, su realidad más dolorosa percibida de repente, fue la que suscitó en el hombre de hace 50.000 años la oscura necesidad de combatirla, de tratar de compensar aquella pérdida que, de golpe, aparecía como la más grave y más dolorosa de todas las heridas y sufrimientos. ¿Fue esta la primera reflexión “metafísica” del hombre, su primer acto religioso?»271.
Como hemos indicado, el no haber encontrado muestras objetivas en enterramientos en los que aparecen indicios de la presencia en los huma-nos de estas cuestiones tan trascendentales, no prueba que no existieran con mucha mayor antelación. Si atribuimos la conciencia de sí y la ca-pacidad de imaginar y de inteligir al homo habilis, no se ve por qué no atribuirle también una cierta capacidad para empezar ya a plantearse todo este tipo de cuestiones. Pero sobre ello no podemos mas que proyectar nuestras hipótesis imaginarias.
Los indicios más importantes hallados en los enterramientos que ha-cen concluir el significado simbólico y religioso que daban nuestros an-tepasados a la muerte, se refieren, como hemos indicado más arriba, a la postura en que se colocaban los cadáveres, así como a los objetos que se situaban intencionadamente al lado, algunos de los cuales puede interpre-
270 Clarke, R., o.c., pp. 109-110. 271 Ibídem, p. 110.

360
© Universidad de Deusto - ISBN 978-84-9830-383-4
tarse como ayudas para el largo viaje hacia la otra vida. Un elemento es-pecialmente significativo es la presencia de ocre rojo con el que aparecen pintadas determinadas partes del difunto, así como la presencia de estatui-llas de diversos materiales de múltiples tipos de animales, así como figu-ras femeninas. Parece deducirse un tipo de convivencia cultural en el que los humanos desarrollan su vida con una clara coexistencia entre la vida y la muerte, lo cotidiano y la excepcionalidad de la muerte y los enterra-mientos, aunque claramente diferenciados de forma intelectual272.
La cuestión que algunos se plantean, al igual que en relación con el arte, el lenguaje y otros ámbitos de la cultura, es si el culto a los muertos y las creencias sobre el más allá son resultado de la invención creativa de la sociedad, o lo recibimos como fenómeno heredable y parte de nuestra naturaleza genética273. Tenemos que responder de similar manera a como lo hicimos al hablar de la estética y el lenguaje: es consecuencia de ambos elementos, en la medida en que nuestros genes y cerebro nos capacitan para plantearnos estas cuestiones, y el entorno cultural ha sido el caldo de cultivo en el que se han ido dando las diferentes respuestas históricas que han llegado hasta nuestros días.
Pero es importante señalar que, aunque las investigaciones arqueoló-gicas unen los enterramientos con los primeros atisbos de creencias reli-giosas y de fe en una vida después de la muerte, los estudios sobre la his-toria de las religiones separan ambos conceptos. Prueba de ello es que en la cultura judía la fe en su Dios Yahvé es independiente de la fe en la exis-tencia de una segunda vida después de la muerte. Esta creencia sólo apa-rece muy posteriormente, entre los siglos III y II, en la época de los Maca-beos, y va madurando desde la época posterior al Destierro, momento en el que entra en crisis la creencia de que esta vida es el único lugar de la plena realización de la justicia divina ante las conductas buenas y malas de los hombres, crisis que puede verse plasmada en el libro de Job.
Por lo demás, tanto los historiadores como los fenomenólogos de la religión no están de acuerdo en situar las diferentes etapas de los cre-dos religiosos, diferenciándose entre los que estructuran esta historia en tres grandes etapas (animismos, politeísmos y monoteísmos) y quienes entienden que cada cultura ha seguido su propia evolución en el terreno de la historia religiosa274.
272 Cfr. Carbonell, E./Sala, R., o.c., p. 237. 273 Cfr. ibídem, p. 239. 274 Entre la abudantísima literatura que hay sobre todos estos temas, cfr. Gómez Caffa-
rena, J. y Martín Velasco, J., Filosofía de la religión, Madrid, Revista de Occidente, 1973; Martín Velasco, J. D., Fenomenología de la religión, Madrid, Ediciones Cristiandad, 1982 (3.ª ed.).

361
© Universidad de Deusto - ISBN 978-84-9830-383-4
3. CONCLUYENDO
Tras el recorrido realizado por los principales aspectos que conforman la faceta cultural y comportamental del género humano, tenemos que reafir-marnos en las convicciones que hemos ido repitiendo continuamente: el ser humano se sitúa en continuidad a la vez que en ruptura con el conjunto de las especies que conforman la biosfera. Se halla en continuidad evolu-tiva con las especies vivas anteriores, en la medida en que ha surgido por saltos mutacionales que le han ido separando de los australopitecos. Con-tinuidad y ruptura, animal pero de otra manera, animal singular. La faceta de continuidad se advierte, como hemos podido comprobar, en la dificul-tad de establecer una barrera y una distancia nítida entre lo humano y lo pre-humano, teniendo que constatar que el ser humano innova casi nada, pudiéndose encontrar en lo pre-humano atisbos de todas las cualidades humanas, dando con ello suficientes motivos para poder afirmar que en-tre lo humano y lo pre-humano no hay más que una diferencia puramente cuantitativa.
Pero la singularidad de lo humano lo advertimos en que se han supe-rado en él los constreñimientos de lo biológico, naciendo de forma prema-tura, y mostrando su deficiencia biológica, y necesitando por ello poner a prueba su capacidad creativa, gracias a su potente cerebro. De ahí que nos caracterizamos como especie por estar dotados de una extraordinaria capacidad intelectual, la necesidad de optar entre diferentes alternativas de actuación, acumulando las respuestas positivas en estructuras e insti-tuciones sociales, que va transmitiendo de generación en generación a sus herederos. Por tanto, el ser humano es una animal histórico, heredero de sus antepasados y beneficiario de sus aciertos y errores, y a su vez dueño de cambiar y enriquecer la herencia que ha recibido. Y esa herencia que recibe y transmite, es un conjunto de habilidades técnicas para la supervi-vencia (comida, vestido, hábitat, caza, utensilios, etc.), como también un conjunto de modos de organización social (roles sociales, formas de edu-cación y socialización, normas de comportamiento, etc.), y modos de en-tender el sentido de la existencia (filosofía) y las preguntas por la trascen-dencia y el deseo de salvación (religión).
En todo esto se muestra que la naturaleza de nuestra especie no está dominada por las leyes biológicas, sino que posee una naturaleza abier-ta, necesitada de complementar y de realizar, a través de las sucesivas decisiones libres que cada individuo va realizando en el trascurrir de su biografía. Eso nos muestra que podemos definir al ser humano como el animal bio-cultural, el animal que surge en el mundo de la biología, pero que se acomoda en ella de modo artificial, a través de construir un nue-

362
© Universidad de Deusto - ISBN 978-84-9830-383-4
vo nicho ecológico, el nicho cultural, el mundo construido a su medida para poder sobrevivir y perseguir los fines culturales que él mismo se ha conferido. Por eso, lo que define a la especie humana no es la biología de donde viene, sino la cultura en la que se sitúa y hacia la que está cons-tantemente apuntando, puesto que lo cultural no es un estrecho ámbito ya configurado, sino un entorno sin límites que el ser humano va poco a poco colonizando, explorando, y aprendiendo a amueblar como su propia casa, pero sabiendo que es un continente en permanente transformación y enriquecimiento.

363
© Universidad de Deusto - ISBN 978-84-9830-383-4
Capítulo 6
La antropogénesis. El punto de vista ontogenético
1. LA PROBLEMÁTICA RELACIÓN ENTRE FILOGÉNESIS Y ONTOGÉNESIS: UNA VISIÓN EVOLUTIVA DE LA EMBRIOLOGÍA1
La filogénesis se centra en el estudio de la relación entre las dife-rentes especies vivas, dentro de la sucesión evolutiva. Tras las aporta-ciones de Ch. Darwin, los esfuerzos de la paleontología se han encaminado a la búsqueda de los fósiles que pudieran mostrarnos las características de los diversos eslabones perdidos entre las especies animales conoci-das. Y lo mismo hacía la paleoantropología respecto a las diversas es-pecies o subespecies humanas. De este modo, la teoría de la selección natural se aplicaba al campo de la lucha entre las especies y entre los individuos maduros al interior de cada especie. Igualmente, la teoría sintética de la evolución, recogiendo las aportaciones de Darwin y de Mendel, y los genetistas que recogieron su testigo, defenderá que las variaciones genéticas de cada individuo son transmitidas a la genera-ción siguiente, siendo seleccionadas las más aptas por el ambiente, y produciéndose por acumulación de mutaciones las sucesivas nuevas especies.
1 Cfr. Gould, S. J., Ontogeny and Phylogeny, Cambridge, Mass., Harvard University Press, 1977 (Trad. cast.: Ontogenia y filogenia. La ley fundamental biogenética, Barcelona, Crítica, 2010).

364
© Universidad de Deusto - ISBN 978-84-9830-383-4
Pero este modo de ver las cosas ha olvidado con frecuencia el proceso ontogenético, esto es, la ejecución concreta del programa genético en cada uno de los individuos. De este modo, se pasa demasiado directamente del gen al organismo completo, capaz a su vez de reproducirse y de experi-mentar después los efectos selectivos del ambiente.
Los espectaculares avances en el campo de la genética, sobre todo desde el descubrimiento de la estructura del ADN de doble hélice por Watson y Crick, han transmitido el falso convencimiento de que todo estaba ya escrito y conformado en los genes. Por tanto, el genotipo se expresaba directamente y de modo casi automático y determinista en los rasgos fenotípicos, sean de tipo fisiológico como también compor-tamental y cultural. Si a eso se añade el alcance mediático del Proyec-to Genoma Humano, se comprenderá que se haya llegado a pensar que en el genoma está todo lo que se requiere para desentrañar el misterio de lo humano, la naturaleza humana. No es extraño que continuamente aparezcan en los periódicos noticias de haberse encontrado por los cien-tíficos el gen responsable de la enfermedad correspondiente o de algún rasgo comportamental, tanto en especies animales como en el ser huma-no. Pero hoy día, como vamos a ver, se está llamando la atención sobre el proteoma como un concepto tan importante o más que el genoma2, en la medida en que constituye un elemento clave del denominado proceso epigenético3.
Los avances de la embriología y de la biología del desarrollo, son en la actualidad tan grandes que la teoría de la evolución no puede prescindir de sus aportaciones, ni olvidar el proceso ontogenético. Vamos a ir viendo cómo una visión correcta de la evolución nos tiene que llevar a entender la filogénesis y la ontogénesis como dos procesos que están íntimamente relacionados, siendo dos ámbitos que aportan luces y conocimientos mu-tuos el uno al otro, llevándonos de esa manera a un mejor conocimiento de la biosfera y de la realidad humana.
La verdad es que las discusiones sobre la relación entre filogénesis y ontogénesis no siempre han estado muy claras, y los conflictos entre los estudiosos de ambas parcelas se remontan ya a épocas muy tempra-nas. Pero también se han dado precursores que desde tiempos también tempranos hacían ver su íntima interrelación. Así, ya en el siglo XVIII, el naturalista Kielmeyer, y en el siglo XIX Meckel y Serres, advertían se-
2 Cfr. Núñez de Castro, Ignacio, «Análisis antropológico del Proyecto Genoma Hu-mano», 1996, p. 12 (texto del manuscrito de una conferencia, sin publicar todavía); Palma Martínez, J. M., «Proteómica, un nuevo palabro», El País, 2008, 30 de abril, p. 45.
3 Cfr. Esteller, M., «Qué es la epigenética y para qué sirve», El País, 2008, 12 de abril.

365
© Universidad de Deusto - ISBN 978-84-9830-383-4
mejanzas entre rasgos embrionarios de organismos vertebrados superio-res (mamíferos) y aspectos adultos de organismos inferiores (anfibios y peces)4. El resultado de esas observaciones les llevó a generalizar un primer principio de recapitulación, en la medida en que consideraban que los organismos superiores recapitulaban o repetían los rasgos de los inferiores y anteriores en el proceso evolutivo. Pero el conocido em-briólogo estonio Karl Ernst von Baer se apartó de estos planteamientos, ya en 1828, mostrando que el embrión de un animal superior nunca es comparable al adulto de un animal inferior, sino que sólo es comparable al embrión de éste. Para von Baer, «en un determinado estadio del de-sarrollo denominado pharingula todos los vertebrados presentan una si-militud extraordinaria y no es hasta estadios más avanzados cuando las características distintivas de cada animal (ya sea un pez, un pollo o un humano) son morfológicamente obvias»5. Von Baer no aceptaba la reca-pitulación ni tampoco la evolución. Estamos todavía en una época ante-rior a Darwin.
A pesar de esos parecidos evidentes entre diferentes organismos, na-die se preguntaba por la razón de ello, avanzándose claramente en esta reflexión con la aparición de la teoría de la selección natural, iniciándo-se con ello una época nueva respecto a la explicación de los parecidos adulto-embrión. El autor más significativo en este campo fue el zoólogo alemán Ernst Haeckel, quien en 1874 expresó su famosa ley de recapi-tulación con la que trataba de explicar dichos parecidos: «la ontogénesis es la recapitulación de la filogénesis» y «la filogénesis es el motor de la ontogénesis»6. Si la primera parte de esta supuesta ley fuera cierta, bas-taría estudiar el proceso ontogenético para deducir de ahí todo el pro-ceso filogenético, como muchos científicos de entonces lo intentaron. Como el embrión de un mamífero se parece al principio a un pez, luego a un reptil, para llegar al final a poseer los rasgos de un mamífero, pare-cía que la ley de Haeckel estaba justificada y demostrada. Pero, en rea-lidad, la relación y el parecido que se mostraba no era entre adultos y embriones, sino entre unos embriones y otros. Esta fue la aportación de Garstand en 1922, al indicar que «la ontogénesis no recapitula la filogé-nesis; la crea»7.
4 Cfr. Devillers, Charles/Chaline, Jean/Laurin, Bernard, «En defensa de una embriolo-gía evolutiva», Mundo Científico, 1990, n.º 105, 918-924; 919.
5 Izpisua Belmonte, Juan Carlos, «Claves para descifrar el desarrollo humano», El País, 1997, 31 de diciembre, pp. 29-30; 29.
6 Haeckel, E., Histoire de la création des êtres organisés d’aprés les lois naturelles, París, Alfred Costes, 1922, p. 602; Cfr. Devillers, Ch, y otros, o.c., p. 920; Gould, S. J., Ontogenyand Phylogeny, o.c.,
7 Cfr. Devillers, Ch. y otros, o.c., p. 920.

366
© Universidad de Deusto - ISBN 978-84-9830-383-4
Los avances posteriores en el campo de la biología molecular sobre el desarrollo embriológico han subrayado que para comprender a fondo el proceso evolutivo hay que «incluir el estudio de la ontogénesis, por ser ésta la vía por la cual los genes engendran los organismos sobre los cuales opera la selección natural»8. Pero la teoría evolutiva no acaba de tomar en serio a la ontogénesis, devolviéndole ésta con la misma moneda. Sólo con la aparición de la genética del desarrollo, hace un cuarto de siglo, se la ha ido tomando en serio, aprovechándose la teoría de la evolución de sus aportaciones.
El conjunto del proceso ontogenético no se reduce a una sucesión ordenada de estadios o fases de desarrollo, sino que fundamentalmen-te consiste en el desarrollo de un auténtico programa teleonómico. Es-tas diversas fases se interrelacionan de tal forma que un determinado estadio está condicionado por el anterior, y condiciona a su vez a los siguientes, en los que van apareciendo nuevas propiedades. Las prime-ras fases de ese proceso parece que están relacionadas de un modo es-trecho y cerrado, pero «cuanto más avanzado está el desarrollo menos estricto se vuelve el condicionamiento»9. Esta relajación del condicio-namiento entre estadios es lo que permite los fenómenos denominados heterocronías: desplazamiento temporal en la realización de los distin-tos órganos y estructuras10.
¿Cómo se entiende esto? La ontogénesis persigue la ejecución de un programa de construcción de las diversas partes de un organismo vivo. Pero, a medida que se va realizando ese programa, van apareciendo pro-piedades nuevas, que se van uniendo a las instrucciones genéticas, dando lugar a lo que se llama la mecánica epigenética, a la que ya hemos hecho referencia con anterioridad. Por tanto, a las instrucciones genéticas hay que añadir los condicionamientos ambientales que van apareciendo en este proceso de desarrollo y que introducen cambios y variaciones signi-ficativas, en la medida en que aparecen encrucijadas con varias posibili-dades a seguir.
Esto explica que unos mismos genes pueden ser los responsables, en diferentes especies, tanto de las alas como de los brazos, balancines, etc. Y lo mismo puede suceder que los mismos genes puedan ser los respon-sables de cambios radicales de estructuras orgánicas de unas especies a otras.
8 Ibídem, p. 920. 9 Devillers, Ch. y otros, o.c., p. 920. 10 Cfr. Chaline, Jean/Marchand, Didier, «Cuando la evolución cambia el tiempo de los
seres. Paleontólogos y genetistas se interrogan sobre las heterocronías», Mundo Científico,1999, n.º 199, pp. 40-43.

367
© Universidad de Deusto - ISBN 978-84-9830-383-4
Por tanto, lo que todo esto nos muestra es que dentro del proceso on-togenético también se da el fenómeno de la selección natural imperante, en todo el proceso evolutivo. Es el fenómeno que ya en 1881 denominaba W. Roux «la lucha de las partes», y que más tarde denominó Spiegelman la «competencia fisiológica»11. Las células comienzan a competir entre sí desde el inicio del proceso embriológico, siendo esta competencia la cau-sante de la variedad y transformación de los diversos órganos de los seres vivos. Como fenómeno ejemplar de esto, los embriólogos se refieren a la formación de las extremidades anteriores y posteriores, con la diferente composición de formas y de número de dedos12. Si se actúa sobre el de-sarrollo de las células correspondientes, se promueve o se inhibe la forma-ción del número de dedos y su tamaño.
En este ámbito de cosas es donde se producen las denominadas hete-rocronías, es decir, las diferencias en los momentos de expresarse deter-minados rasgos en especies cercanas. «En las secuencias ontogenéticas de especies vecinas, un mismo suceso —por ejemplo la madurez sexual— puede no producirse en el mismo instante. Este desplazamiento del lugar de las secuencias constituye lo que se denomina heterocronía»13. Estos fenómenos podrían explicar las modalidades evolutivas de pasos rápidos y radicales entre especies o líneas de descendencia sin intervención de etapas intermedias. Ya hicimos referencia a las críticas realizadas desde la teoría de los equilibrios puntuados, de S. J. Gould y S. Elredge, con-tra la ortodoxia darwiniana, que defiende la teoría gradualista en el proce-so evolutivo. Estos procesos de heterocronías son ejemplos claros de saltos evolutivos, sin presentar estadios intermedios, como exigiría la tesis evo-lutiva gradualista.
Ahora bien, hay otro aspecto que es importante señalar dentro del proceso del desarrollo ontogenético, y que se deduce de la toma en cuenta de la mecánica epigenética. La tendencia a reducir el proceso embriológico a las orientaciones que surgen del genoma y a las azarosas interrelaciones entre los genes, tiende a pensar que todo se decide en esa combinación azarosa de genes. De ahí se derivaría para cualquier ge-noma una capacidad de variación casi infinita, de tal modo que la vida de la especie estaría llevada por un juego de lotería permanente, con lo cual, si disponemos de tiempo suficiente, parece que todo sería posible, y cualquier forma de vida podría aparecer. Pero los embriólogos con-sideran que eso es entender mal las cosas y defender un planteamiento reduccionista que no se sostiene. No todas las realizaciones genéticas
11 Cfr. ibídem, p. 922. 12 Cfr. ibídem, p. 922. 13 Devillers, Ch. y otros, o.c., 923.

368
© Universidad de Deusto - ISBN 978-84-9830-383-4
son posibles, puesto que hay un número limitado de realizaciones feno-típicas por cada plan de organización. Los genes no actúan como indivi-duos aislados, sino más bien como pertenecientes a grupos, a sociedades de genes, sometidos a planes conjuntos que limitan las veleidades de los planes aislados. Hay, pues, planes posibles y otros imposibles dentro del proceso evolutivo. Así, los caminos seguidos por el desarrollo em-brionario no están totalmente abiertos, sino que siguen patrones determi-nados. Ahora bien, eso no implica que las orientaciones son totalmente fijas y cerradas, porque ello supondría negar la evolución. Las vías que quedan abiertas están interconectadas entre sí, y se influyen mutuamen-te. Hay libertad de elegir diversos caminos, así como de seguir diferen-tes velocidades o ritmos.
Por tanto, como indican las heterocronías, para evolucionar no hay que estar jugando permanentemente a la lotería. Como indica S. J. Gould, al filtro de la selección natural se le presentan no un número infinito de posibilidades, sino un abanico limitado de proyectos de cambio. «La se-lección juega un papel pero no tiene ya el control exclusivo del destino de las especies. La proposición “con el tiempo, todo es posible” ha de inver-tirse en “no todo es posible, por más tiempo que pase”. El continuo no es la única y necesaria modalidad evolutiva»14.
La teoría de la recapitulación de Haeckel se abandonó hace tiempo, pero, como indica Yves Christen, en la actualidad sigue teniendo interés para las investigaciones actuales, en la medida en que sirve de marco y de contraste para la teoría que le ha sustituido: la de la neotenia. Si la teo-ría de la recapitulación veía al ser humano como la especie que recorría de algún modo, como última especie aparecida en el proceso evolutivo, las diferentes fases por las que habían pasado las especies anteriores, la neotenía defiende más bien que la especie humana ha seguido un proceso embriológico lento, deteniéndose su desarrollo en fases iniciales de la on-togénesis. De ahí que «en el estadio adulto seríamos bastante semejantes a fetos de mono. En todo caso, más próximos al feto del mono que al mono mismo»15. El fenómeno consistente en la persistencia de rasgos juveniles es lo que se denomina neotenia.
En definitiva, si la teoría de la recapitulación se fija y hace hincapié en la continuidad de la especie humana con el resto de las especies vi-vas, repitiendo y recordando de algún modo los diferentes estadios por los que la evolución ha ido pasando, la teoría de la neotenía se fija más bien en la ruptura de esa continuidad. La especie humana rompe con la
14 Devillers, Ch. y otros, o.c., p. 924. 15 Christen, Y., El hombre biocultural. De la molécula a la civilización, Madrid, Cáte-
dra, 1989, p. 58.

369
© Universidad de Deusto - ISBN 978-84-9830-383-4
ley general del desarrollo ontogenético, en la medida en que no sigue su lógica común de completar el proceso embriológico hasta el nivel o grado necesario para interactuar con el ambiente de forma madura y ya predeterminada, sino que detiene el proceso embriológico e introduce en el mundo individuos incompletos, inmaduros y deficientes, advir-tiéndose de este modo el peculiar y especial puesto de la especie hu-mana en el proceso ontogenético, como tendremos ocasión de ver más adelante.
2. LAS DIFERENTES FASES DEL PROCESO ONTOGENÉTICO16
El proceso ontogenético o ciclo vital de un ser humano, como con-junto de momentos que van desde la formación de la célula germinal hasta el nacimiento del nuevo ser, se denomina también proceso em-briológico. En este proceso, la especie humana sigue las mismas leyes que el resto de los demás seres vivos, en la medida en que está com-puesto por la misma pasta biológica, organizada desde lo genético a lo molecular de forma muy similar. La especie humana, al igual que las demás que componen el conjunto de la biosfera, sigue ineludiblemen-te la denominada primera ley de la genética: ADN-ARN-PROTEÍNAS, yendo siempre el proceso de izquierda a derecha, no orientándose nun-ca esa relación en la dirección contraria; esto es, sólo se transmiten a la generación siguiente los rasgos que ya se hallan presentes en el ADN, y no los caracteres adquiridos (es lo que defendía la denominada lógica lamarckiana).
En este apartado vamos a describir, en sus líneas generales, las dife-rentes fases o etapas del proceso embriológico, para situar y comparar después este proceso embriológico de la especie humana con el del resto de las especies, y para detenernos finalmente en las cuestiones filosófi-cas de capital importancia que se plantean en este proceso de conforma-ción de un nuevo ser humano. De todos modos, es importante advertir ya de entrada que, aunque se distingan varias fases dentro del conjunto del proceso embrionario, es fundamental entenderlo como un proceso
16 Cfr. Demoulin, Francoise, «Crecimiento fetal humano», en Rebato, E. y otros (eds.), Para comprender la Antropología biológica, Estella (Navarra), Verbo Divino, 2005, 497-511; Lacadena, J. R., Genética y Bioética, Madrid/Bilbao, UPCO/Desclée de Brouwer, 2002, cap. 2.º. Reproducción humana, pp. 51 y ss.; Id., «Biología y humanidad: hominización y humanización», en Masiá, Juan (ed.), Ser humano, persona y dignidad, Madrid/Bilbao, UPCO/Desclée de Brouwer, 2005, pp. 43-89 (79-89).

370
© Universidad de Deusto - ISBN 978-84-9830-383-4
unitario, teniéndose que conjugar adecuadamente la distinción de fases con la unidad del proceso completo y conjunto.
Veamos las diferentes fases que componen este proceso embrionario17.
2.1. Fase previa: de los gametos a la célula germinal (pre-ontogénesis)
Este momento presupone la formación de las células sexuales, pro-ceso en el que se reduce a la mitad el contenido genético de la especie (el número de cromosomas del cariotipo queda reducido a la mitad, veinti-trés), denominado meiosis (reducción meiótica). Es la fase previa a la fe-cundación, momento en el que un espermatozoide fecunda a un óvulo, dándose el proceso de fusión de los dos núcleos de ambas células sexuales (gametos), y recuperándose con ello el conjunto del cariotipo (23 pares de cromosomas). Es un momento que supone un salto drástico, puesto que se pasa de dos realidades diferentes (las células sexuales separadas, gametos) a una nueva y única realidad (la célula germinal o cigoto).
Este momento no es instantáneo, sino que dura alrededor de veinte horas, desde que el espermatozoide penetra en el citoplasma del óvulo, atravesando las células que rodean al ovocito y la zona pelúcida. Como se sabe, el espermatozoide sólo aporta a la fusión su núcleo, provisto de 23 cromosomas, produciéndose la aproximación lenta de los pronúcleos, masculino y femenino, aproximación que culminará en la fusión (sintag-ma) de los dos núcleos, iniciándose posteriormente el proceso de divi-sión celular (mitosis) de esta célula germinal, que contiene ya el cariotipo completo (23 pares de cromosomas) de la especie humana18. Por tanto, el proceso de fecundación termina cuando se ha producido la fusión de los dos pronúcleos y se inicia la fase de replicación, primera división celular (mitosis), de la célula germinal19. Esta fecundación se produce en la parte
17 Cfr. Laín Entralgo, Pedro, El cuerpo humano. Teoría actual, Madrid, Espasa-Calpe, 1989, cap. 3.º, pp. 70 y ss.; Lacadena, José R., Genética y bioética, Madrid/Bilbao, UPCO/Desclée de Brouwer, 2002; Trevijano Echeverría, M., ¿Qué es la bioética?, Salamanca, Sí-gueme, 1999 (cap. 1.º); Gafo, Javier, Problemas éticos de la manipulación genética, Ma-drid, Edic. Paulinas, 1992 (cap. 1.º); Bertranpetit, J./Junyent, Cr., Viaje a los orígenes. Una historia biológica de la especie humana, Barcelona, Península, 2000.
18 Cfr. Lacadena, J. R., Genética y bioética, o.c., cuadro de la pág. 53. 19 Estas precisiones científicas son de enorme importancia de cara a las consideracio-
nes éticas sobre métodos interceptivos (la aceptación o no de la denominada «píldora del días después») como de cara a las nuevas técnicas de reproducción asistida que implican la manipulación de embriones. Así, en algunos países como Alemania, está permitida la con-gelación de cigotos, en la fase anterior a la fusión de los dos pronúcleos, en la medida en que no se habría conformado todavía una célula germinal humana, dado que no se habría producido todavía la fecundación y no estaríamos todavía ante un embrión.

371
© Universidad de Deusto - ISBN 978-84-9830-383-4
superior de las trompas de Falopio. A partir de ese momento, el óvulo fe-cundado inicia un progresivo desplazamiento de unos cuatro días hacia el útero, mientras van produciéndose ya las primeras fases de mitosis.
2.2. Fase zigótica: de la célula germinal o zigoto a la anidación o implantación20
Esta fase viene a durar aproximadamente dos semanas, y comprende a su vez tres fases o etapas:
—Célula germinal: conformada por la fusión de los dos gametos, el óvulo y el espermatozoide, para dar lugar al nuevo ser vivo, dotado de un cariotipo único y bien diferenciado, con los 23 pares de cromosomas propios de la especie humana. A partir de esta fusión de núcleos o fecun-dación, se inicia un dinamismo nuevo, por lo que se advierte con ello la autonomía de este nuevo ser respecto a su entorno, el cuerpo de la madre. El ADN se transforma en ARN (ácido mensajero), fenómeno que se de-nomina transcripción del mensaje generador, momento en el que el ARN pasa del núcleo de la célula, sede del ADN, al citoplasma, sede del riboso-ma, y por eso se llama mensajero (ARNm). En el ADN se halla el código genético, por cuanto en él se encierra la regla que relaciona la secuenciapolinucleótica (la del ADN, cada tríada de las bases nitrogenadas: ade-nina, guanina, citosina y timina) con la secuencia polipeptídica (la de la proteína: cada tríada de bases nitrogenadas corresponde a un aminoácido y proteína)21.
—Mórula: amontonamiento de células, como consecuencia de la re-plicación (mitosis) de la célula germinal. Este proceso de partición es en principio lento, pero irá acelerándose en la medida en que cada célula se irá replicando en otra de modo recurrente. A las 36 horas de la fecun-dación, el zigoto se compone de tan sólo dos células, que empieza ya el camino hacia el útero. A las 60 horas, el zigoto es ya de 4 células, y a los tres días, de entre 6 y 8 células. A los cuatro días, la mórula es ya de 16 células, que forman un todo compacto, y que tienen la virtud de ser totipotentes (células madres o embrionarias), puesto que cada una de ellas puede llegar a ser cualquier parte del cuerpo del futuro ser huma-
20 Cfr. Demoulin, Fr., o.c., p. 497 y ss.; Lacadena, J.R., Genética y bioética, o.c.,pp. 54-56.
21 Cfr. Fox Keller, Evelyn, El siglo del gen. Cien años de pensamiento genético, Barce-lona, Península, 2002; Bertranpetit, Jaume/Junyent, Cristina, Viaje a los orígenes. Una his-toria biológica de la especie humana, Barclona, Península, 2000, cap. 2.º.

372
© Universidad de Deusto - ISBN 978-84-9830-383-4
no. Es el momento en el que la mórula ha recorrido ya las trompas de Falopio y procede a la tarea de fijarse en las paredes del útero, produ-ciéndose entonces la implantación o anidación22, proceso en el que se tarda alrededor de otra semana. En ese momento, la mórula se convierte en blástula.
—Blástula o blastocisto: comienza al final de la primera semana tras la fecundación. En esta fase, las células continúan dividiéndose, llegando a alcanzar la cantidad de unas cien células. Pero ya no forman un apeloto-namiento celular o mórula sencilla, sino que se empieza a formar una ca-vidad central (blastocele), rodeada de una capa externa (trofoblasto: ori-gen de la placenta y otras membranas) de unas 20 o 30 células, pegadas a la pared interior, denominada masa celular interna (MCI) o embrioblasto.Estas células ya no son totipotentes, sino pluripotentes, en la medida en que se ha comenzado un proceso de especialización de las diferentes cé-lulas para dar lugar cada grupo a un tipo diferente de tejido celular. Alre-dedor de los catorce días, el blastocisto ha terminado la anidación, dando lugar el MCI al disco embrionario de 0,5 mm de diámetro, compuesto por unas dos mil células.
La fase de anidación o implantación se ha producido ya al finalizar la segunda semana tras la fecundación (catorce días), constituyendo un momento fundamental del proceso embriológico, razón por la que algu-nos defienden que el embarazo comienza propiamente al final de esta im-plantación, y no tanto en el momento de la fecundación, y hasta que el embrión no está anidado, no es posible hacer un diagnóstico clínico de embarazo. Para cimentar esta postura, se suele aducir que hasta esta fase de anidación se dan un porcentaje de abortos espontáneos, disminuyendo drásticamente este porcentaje a partir de este momento23.
22 Hasta hace no mucho, los embriólogos ignoraban cómo se adhería un embrión a la mucosa uterina (endometrio), pero un grupo de científicos norteamericanos han descubier-to que este proceso se asemeja al proceso de fijación y a la forma en que los glóbulos blan-cos se distribuyen por la pared de los vasos sanguíneos. En ambos casos, se observa una fijación inestable, seguida de una serie de fijaciones más estables, hasta detenerse. La pri-mera adhesión se efectúa gracias a la misma molécula, la L-selectina, que interacciona con los carbohidratos situados en la pared. Los investigadores avanzan la hipótesis de que un defecto en este proceso podría ser la causa de ciertos procesos de esterilidad y aborto natu-ral: Cfr. O. Genbacev et al., Science, 299, 405, 2003.
23 De todos modos, como indica J. R. Lacadena, «la precariedad en que se desenvuel-ve el embrión en las etapas anteriores a la anidación, no tienen, sin embargo, a mi juicio, ni con mucho, el peso genético que ante la problemática del estatuto del embrión humano pueden tener los fenómenos de gemelismo y quimerismo que afectan plenamente a la indi-vidualización del nuevo ser, ya que tal precariedad no es necesariamente expresión de ines-tabilidad o indeterminación genétia», en J. R. Lacadena, «El comienzo de la vida humana», 2008, manuscrito sin publicar, p. 6.

373
© Universidad de Deusto - ISBN 978-84-9830-383-4
En relación al proceso de partición celular y su organización, hay que decir que se trata de un fenómeno azaroso, además de estructural o cam-pal. Decimos que es un fenómeno azaroso, porque, de momento, los gene-tistas y embriólogos no saben quién dirige esa operación, o qué parte del genoma se encarga de dirigir este proceso de especialización celular, ni de qué forma se realiza, aunque se empiezan a conocer algunos aspectos de este fenómeno tan extraordinario24. Lo que parece evidente es que se ini-cia ya en estas primeras células un proceso de reconocerse, base de lo que será el futuro sistema inmunológico.
Pero además se advierte lo que Diego Gracia, basándose en Zubiri, ha denominado la dimensión campal del embrión, dentro de la lógica te-leonómica que se da en el proceso de la ontogénesis. Con esto se quiere resaltar que el desarrollo del embrión no sólo responde a una dinámica interna, sino que depende también de su entorno o campo, que no es otro que el útero materno. X. Zubiri denomina campo, concepto tomado de la física y aplicado a la condición estructural de la realidad25, a la unidad o condición respectiva de todas las cosas, de tal modo que toda cosa o nota está situada en un contexto o fondo que condiciona y orienta su forma de ser y de actuar. Pero es importante indicar que el campo o condición cam-pal de una cosa no es algo externo a ella, sino una dimensión física de la misma26. De ahí que el campo morfogenético no está limitado al territorio del embrión (esto es, su dimensión interna, su estructura genética), sobre el que actúa un inductor normal o artificial, sino que se halla constitui-do por todo lo que, por intermedio de la mucosa uterina, y luego a tra-vés de las formaciones preplacentarias y la placenta, actúa sobre él, como son electrolitos, hormonas, enzimas y nutrientes diversos. Todos estos elementos actúan en el embrión siguiendo un determinado orden tempo-ral. De todo esto parece que hay que concluir que el zigoto no posee en sí mismo una determinación absoluta que garantice que llegará a ser una persona humana. Tal determinación depende de varios aspectos: ante todo de la progresiva actualización de su propia información genética; pero también de la actualización de las informaciones surgidas de novo durante
24 Cr. Shi, Yun-Bo/Stolow, Melisss/Puzianowska-Kuznicka, Monila/Wong, Jiemin, «Los genes de la metamorfosis. Una hormona, director de una orquesta de genes, dirige el remodelaje anatómico», Mundo Científico, 1996, n.º 169, pp. 538-544; Nüsslein-Volhard, Christine, «Gradientes organizadores del desarrollo del embrión», Investigación y Ciencia,1996, n.º 241, 20-27; Gardner, Richard L., «El destino de las células en el embrión», Mun-do Científico, 1991, n.º 110, 146-155.
25 Cfr. Zubiri, X., Estructura dinámica de la realidad, Madrid, Alianza/Fundación Xa-vier Zubiri, 1989.
26 Cfr. Laín Entralgo, P., Nuestro cuerpo. Teoría actual, Madrid, Espasa, 1989, pp. 89 y ss.

374
© Universidad de Deusto - ISBN 978-84-9830-383-4
el proceso embrionario (fenómenos epigenéticos); y también de informa-ciones exógenas, en función de su condición campal. Esto supone aceptar que el cigoto no posee, ni siquiera potencialmente, toda la información necesaria para orientar el desarrollo del embrión. Y la actualización de su potencialidad morfogenética no es enteramente autónoma, y, además de diacrónica, es cualitativamente discontinua. Pero tendremos ocasión más delante de incidir en estos aspectos tan decisivos.
2.3. Fase embrionaria o de gastrulación27
Esta fase dura desde finales de la tercera semana hasta el fin de la oc-tava semana. La gastrulación es el proceso en el que se producen los fe-nómenos de citogénesis y de organogénesis, transformándose el disco em-brionario bilaminar en trilaminar (ectodermo, mesodermo y endodermo).Es lo que se denomina la gástrula, y el embrión crecerá ya hasta tener unas dimensiones de 2,3 mm de longitud, y empezarán a aparecer los atis-bos (primordios) de los principales órganos que conformarán al nuevo ser vivo. Las células del blastocisto se diferencian ahora tanto espacial como potencialmente. En lo espacial se separan y asocian entre sí para formar las tres hojas blastodérmicas ya indicadas. Y potencialmente, porque se va orientando y restringiendo la capacidad morfogenética de cada célula, de tal modo que, como se ha dicho, pasan de ser totipotenciales a pluripoten-ciales. En resumen, se producen tres procesos consecutivos: agrupación,diferenciación y organización de células. En primer lugar, agrupación de células entre sí, mediante un simple proceso de información posicional y de reconocimiento, proceso que, como hemos indicado, no se sabe dónde se sitúa el punto orientador ni la lógica del mismo. En segundo lugar, se produce una progresiva diferenciación, para ir formando cada uno de los tipos hísticos (tejidos celulares) que integran el conjunto de los tejidos del animal adulto. En este proceso se da una progresiva restricción de sus po-tencialidades. Y, en tercer lugar, una incipiente organización, formándose los diversos órganos y regiones del cuerpo del nuevo ser vivo28.
Dentro de ese proceso, como elemento de especial importancia, el ec-todermo dará lugar al tejido de la placa neural, que más tarde se repliega para formar la cuerda espinal y el cerebro (cresta neural)29, la primera
27 Cfr. Demoulin, Fr., o.c., pp. 498-502; Lacadena, J. R., Genética y bioética, o.c., pp. 56-60.
28 Cfr. Laín Entralgo, P., o.c., pp. 99 y ss.29 Cfr. Pujol Gebellí, Xavier, «La cresta neural juega un papel clave en la evolución»
(entrevista a la embrióloga Nicole Ledouarin), El País, 2002, 24 de abril, p. 33.

375
© Universidad de Deusto - ISBN 978-84-9830-383-4
base fisiológica de la futura identidad personal. La cresta neural es el ori-gen de los ganglios, de las células sensoriales, del sistema nervioso peri-férico y del conjunto del sistema nervioso en general, incluido el cerebro. Como indica N. Ledouarin, «la aparición de la cresta neural, como inno-vación evolutiva, habría sido clave para el desarrollo de la cabeza, de ór-ganos sensoriales y de un sistema nervioso y endocrino capaz de mantener el equilibrio con el entorno. Todo ello se ha visto que ha sido clave para la evolución»30. De ahí su importancia como base fisiológica de lo que cons-tituye la apoyatura de la futura identidad y de los caracteres específicos de cada individuo humano. Y esa es la razón por la que algunos pensadores, como veremos más adelante, quieran situar en este momento del proceso embrionario la atribución de personalidad al zigoto, con las consecuencias antropológicas y éticas que ello lleva consigo.
El mesodermo (hoja intermedia o cordo-mesoblasto) formará la cuer-da dorsal, origen del eje esquelético primitivo del embrión, así como los músculos, la dermis e importantes órganos como el corazón, los riñones, los vasos sanguíneos y linfáticos, etc.
Y el endodermo, u hoja interna, dará lugar al epitelio del tubo digesti-vo, al aparato respiratorio (pulmones), al hígado, páncreas y otros órganos importantes. Con esta descripción de la formación de los más importantes órganos del cuerpo humano, parece evidente que al final de esta fase está ya prácticamente formado el diseño del nuevo individuo, pudiéndose re-conocer al embrión como humano.
2.4. Fase fetal
Su duración va aproximadamente desde la 8.ª-9.ª semana hasta el fi-nal del noveno mes, en el que normalmente se produce el nacimiento. Es el momento de la denominada organogénesis, puesto que los diferentes órganos, apuntados en la fase embrionaria, se irán completando y perfec-cionando en esta fase definitiva, hasta el momento de nacer. El cambio de nombre, de embrión a feto, es comprensible, puesto que, aunque nos hallamos en un proceso continuo, también es cierto que el embrión se ha convertido en un ser humano reconocible. Las células embrionarias van a ir cambiando poco a poco sus propiedades, comenzando a sintetizar pro-teínas específicas de cada uno de los órganos de los que van a formar par-te. Un elemento de interés, durante esta fase fetal, es «la relativa ralenti-zación del crecimiento de la cabeza, comparado con el crecimiento muy
30 Ibídem, p. 33

376
© Universidad de Deusto - ISBN 978-84-9830-383-4
rápido del resto del cuerpo. Entre la 11.ª y la 18.ª SA (semana de ame-norrea), la longitud corporal se acelera rápidamente, y se duplica ha-cia la 14.ª SA. La cabeza, que en la 11.ª SA constituía casi la mitad de la longitud fetal, se hace proporcionalmente más pequeña en la 14.ª SA, y continúa su reducción tras este período»31.
Hacia la 14.ª semana van apareciendo los primeros centros de osifi-cación primarios en el esqueleto, entre ellos el cráneo y los huesos lar-gos. A finales de esa 14.ª semana, los miembros inferiores son todavía re-lativamente cortos, aunque las extremidades superiores ya han alcanzado casi sus proporciones definitivas para antes del parto.
Entre las 19-22 semanas, el crecimiento se ralentiza, pero la longitud cráneo-caudal del tronco aumenta alrededor de 50 mm. Las extremidades inferiores aumentan de tamaño, y la madre puede notar ya los primeros movimientos fetales.
El segundo trimestre del embarazo es un período fundamental para realizar diagnósticos precoces sobre posibles malformaciones del feto. «El examen del cariotipo es importante, ya que el 16% de las malforma-ciones se originan por una anomalía cromosómica. Además, los signos morfológicos visibles por ecografía, como el grosor del cuello o la dis-tancia entre las dos órbitas, aportan información suplementaria, y pueden servir como indicaciones sistemáticas para el diagnóstico precoz de ano-malías fetales»32.
Una de las causas de mayor morbilidad es el nacimiento prematuro, puesto que pone en peligro la viabilidad del feto, y está en relación con el peso corpóreo que posee al nacer y la madurez de los diferentes órganos vitales. La viabilidad de un feto consiste en la capacidad para sobrevivir fuera del útero materno, como consecuencia de un nacimiento prema-turo. La viabilidad de los prematuros se está hoy día ampliando con los avances tecnológicos respecto a los cuidados intensivos, pero un prema-turo de 24-27 semanas, aunque pueda sobrevivir, su esperanza de vida en la primera infancia es baja. En cambio, entre las 28-31 semanas, la super-vivencia es más posible en la medida en que el recién nacido tiene ya los pulmones y el resto del sistema respiratorio, así como el sistema nervioso central y el aparato circulatorio, suficientemente maduros para realizar sus funciones de forma adecuada.
Se advierte, por tanto, que un momento fundamental del proceso em-brionario es el del parto y el nacimiento, en la medida en que el recién nacido pasa de una situación de dependencia del útero materno para la
31 Demoulin, Fr., o.c., p. 502. 32 Ibídem, p. 503.

377
© Universidad de Deusto - ISBN 978-84-9830-383-4
realización de las funciones clave para mantenerse vivo y en proceso de crecimiento, a otra en la que tiene que vivir por su cuenta, continuando con los procesos de circulación sanguínea, respiración, ingestión de ali-mentos y excreción de residuos desechables, etc. Lo que está claro es que supone un paso decisivo para cada ser vivo, puesto que constituye el salto de un ámbito protegido, el útero materno, a otro más abierto y peligroso, por lo que algún autor ha hablado del trauma del nacimiento33. Pero no nos vamos a detener aquí en su descripción y en señalar su importancia, puesto que lo volveremos a retomar cuando hagamos referencia a la espe-cial situación de la especie humana en el proceso ontogenético, y veamos la especial peligrosidad que reviste el fenómeno del alumbramiento para las hembras de la especie humana, debido a los efectos producidos por el caminar erguido en el estrechamiento de la pelvis, unido al aumento de la masa encefálica de los individuos de la especie humana.
Pero con el nacimiento no se termina el proceso de formación fisio-lógica del ser vivo, y menos aún en el caso de los seres humanos, proce-so que dura prácticamente toda la vida. De ahí que se hable de una cuar-ta fase.
2.5. Fase de oriogénesis y de gerontogénesis
Cuando se habla de ontogénesis, se está refiriéndose casi en exclusiva a las fases de la conformación del cuerpo humano desde la célula germi-nal hasta el nacimiento. Pero no termina ahí el proceso de conformación tanto fisiológico como psicológica del recién nacido, sino que ese proce-so continúa todavía varios años más, prolongándose tras el nacimiento, como todos sabemos, a lo largo de varias etapas. Estas etapas ulteriores al nacimiento suelen ser denominadas crecimiento infantil, pubertad, ju-ventud, madurez y decrepitud o ancianidad34. Aunque parece admitirse implícitamente que la génesis del individuo humano se acaba con la pri-mera infancia, como si únicamente consistiese en el proceso morfogenéti-co que conduce a un estado terminal en el desarrollo del cuerpo, la verdad es que dicho proceso dura mucho más, pudiéndose defender que dura toda la vida. Es decir, en rigor, el desarrollo del cuerpo viviente no tiene estado terminal. Más bien, tiene sólo un término, la muerte. Por tanto, cabría de-cir que la vida humana es una continua ontogénesis, génesis de una serie continuada de estados, los que le conducen desde el embrión hasta el que
33 Cfr. Rank, Otto, The trauma of birth, Nueva York, 1929. 34 Cfr. Shaffer, David R., Psicología del desarrollo. Infancia y adolescencia, Madrid,
International Thomson Editores, 2000, 5.ª edición.

378
© Universidad de Deusto - ISBN 978-84-9830-383-4
solemos llamar madurez, y los que le llevan a la declinación senil (si no muere antes)35. Tendremos ocasión de hablar del lento proceso de confor-mación del cerebro en los primeros años de la vida extrauterina del recién nacido, así como de los procesos de maduración de los reflejos fisiológi-cos que le permite caminar erguido, manejar las manos y desenvolverse adecuadamente en su entorno ecológico y social, para convertirse en una persona capaz de sobrevivir y de desenvolverse con éxito en su entorno social y cultural.
La ontogénesis del cuerpo humano es, en rigor, la sucesión de una oriogénesis (de orios: lo que está maduro), y una gerontogénesis (geron:viejo), que se solapan entre sí. Uno de los enigmas más difíciles de re-solver por la ciencia es el proceso de envejecimiento celular. Conforme a esta visión de la realidad del proceso vital, deberían describirse los cam-bios psicoorgánicos que denominamos crecimiento juvenil, maduración y envejecimiento. Todo ser vivo huye y teme a la muerte, pero el ser huma-no es la única especie que tiene clara conciencia del fenómeno de la muer-te y de su inevitabilidad. Pero eso no evita que los científicos se afanen por estudiar los procesos químicos y genéticos del envejecimiento, con vistas a prolongar la vida humana36, ya que es imposible evitar la muerte. Aunque siempre sigue habiendo quien crea que el mito de la inmortalidad puede estar al alcance de la investigación científica37.
3. LA ONTOGÉNESIS HUMANA VISTA DESDE LA EMBRIOLOGÍA COMPARADA
A medida en que la embriología ha ido progresando y desvelando los secretos del proceso ontogenético en el conjunto de los seres vivos, se han ido mostrando las grandes similitudes que se dan entre las diversas etapas del proceso embriológico de la diversas especies. Y de ese modo se ha ido mostrando tanto esa similitud de etapas de las diversas especies, como también el paralelismo tan grande existente entre el proceso filogenético y el ontogenético, como ya indicamos en el apartado primero de este capí-tulo. Allí hicimos referencia al principio de E. Haeckel de recapitulación por parte de la ontogénesis del proceso filogenético, así como la supera-
35 Cfr. Laín Entralgo, P., El cuerpo humano. Teoría actual, o.c., cap. III. 36 Cfr. Medina, John J., El reloj de la edad, Barcelona, Crítica, 1997.37 Cfr. Wade, Nicholas, «Los científicos ven la inmortalidad en células que se dividen
indefinidamente. Dos hallazgos de una empresa de EEUU dan un giro al estudio del enve-jecimiento», El País, 1999, 3 de enero, p. 34.

379
© Universidad de Deusto - ISBN 978-84-9830-383-4
ción de este principio, dadas sus evidentes insuficiencias38. Pero eso no quita que sea interesante acercarnos a mostrar la evidencia que tal princi-pio insuficiente haga referencia al hecho evidente de que durante el proce-so embriológico, en cada embrión de la especie humana se van mostrando ciertos atavismos y restos de órganos que nos remiten a las fases anterio-res del proceso evolutivo de las diferentes especies vertebradas, desde los peces a los primates. Ya quedó dicho que, en la actualidad, la teoría reca-pituladora de Haeckel ha sido abandonada, de forma que la aparición de diversos elementos extraños en el proceso evolutivo del embrión humano no se consideran como regresiones, sino, en palabras de Y. Christen, «más bien la demostración de que nuestro programa de desarrollo contiene cier-tas antiguas potencialidades que, normalmente, no se manifiestan. Se tra-ta, en alguna medida, de un simple error en el desarrollo del programa»39.Pero no por ello deja de resultar interesante hacer referencia a esta di-versidad de fenómenos que se suceden en diversas fases del proceso em-briológico, en la medida en que en los diferentes momentos del proceso embriológico van apareciendo en el embrión humano diferentes órganos fisiológicos pertenecientes a otras especies, para desaparecer momentos después, y que vienen como a recordar nuestra línea genealógica en la historia evolutiva, y a hacernos ver que somos descendientes de otras es-pecies, la última especie aparecida en la evolución.
Los ejemplos que se suelen citar y aportar son diversos y muy signifi-cativos40:
a) La pasajera aparición de restos de branquias, en clara referencia a nuestros antepasados los peces, habitantes de las superficies acuá-ticas, para posteriormente salir de ese entorno y transformarse en anfibios, y luego en mamíferos.
b) El paulatino cambio de la posición de los ojos en la cara. Si ad-vertimos la posición que en los peces tienen los ojos, se verá que se hallan situados en ambos lados de la cabeza, muy lejos el uno del otro. Así tienen los ojos no sólo los peces, sino también los la-gartos, los ratones, caballos y otras muchas especies de animales. De ese modo, pueden abarcar un mayor espacio de visión, espacio que se va restringiendo al mismo tiempo que los ojos de los anima-les posteriores se van acercando y situándose en la parte anterior de la cara, produciéndose de ese modo la visión estereoscópica. La
38 Haeckel, E., Histoire de la création des êtres organisés d’aprés les lois naturelles,o.c., p. 602.
39 Christen, Y., El hombre biocultural. De la molécula a la civilización, o.c., p. 58. 40 Cfr. Laín Entralgo, P., Nuestro cuerpo. Teoría actual, o.c., pp. 62-63.

380
© Universidad de Deusto - ISBN 978-84-9830-383-4
pérdida de espacio dominado por la vista se suple con la presencia del oído, al mismo tiempo que se gana, con esa visión estereoscó-pica, la perspectiva y la visión en profundidad.
c) El esbozo de la cola. Se trata del alargamiento del extremo de la co-lumna vertebral, un alargamiento de unos ocho o nueve segmentos vertebrales, que después se transformarán en la especie humana en el coxis, o hueso coxígeo. Aparece en el embrión humano hacia las seis semanas, llega a alcanzar entre 14 y 16 milímetros, y desapare-ce al cabo de unas semanas. Esta circunstancia, que en situaciones normales desaparece antes de nacer, en algunos casos se suele man-tener tras el nacimiento. A un caso de estos, ocurrido en Boston en 1982 y reseñado con gran asombro por la prensa, hace referencia Yves Christen en su libro El hombre biocultural41. Pero, a pesar del asombro producido entre la opinión pública y también en los am-bientes médicos, nos indica Christen que estos casos no son tan ra-ros, puesto que entre 1850 y 1900 se citaron varios centenares, aun-que el último caso anterior databa ya de 1936. El propio Ch. Darwin llegó a interesarse por estos temas en una de sus obras42. Se trata de un atavismo que nos remite a una época muy lejana del proceso evolutivo, en la medida en que, como nos hace recordar Y. Christen, los grandes monos ya no la poseen. De modo que «para encontrar un antepasado común con animales de cola, es preciso indudable-mente retroceder … ¡veinticinco millones de años!»43.
d) El lanugo fetal: aparece entre el séptimo y el noveno mes, y desa-parece poco tiempo después. Así, pues, el lanugo humano es una pasajera recapitulación del sistema piloso de los mamíferos, autén-tico don de la naturaleza para protegerse del frío y otras utilidades, haciéndonos ver en qué medida la ausencia de tal sistema piloso en la especie humana nos hace ser, como indica el título tan sugeren-te como algo demagógico del libro de Desmond Morris, como un mono desnudo44.
e) La oclusión parpebral del embrión. Durante el tercer mes de la gestación, el embrión humano tiene soldados los párpados, duran-do tal situación hasta el séptimo mes. Hay diversas especies, den-
41 Christen, Y., El hombre biocultural. De al molécula a la civilización, o.c. 42 Cfr. Darwin, Ch., The descent of Man, and selection in relation tu sex, Londres, John
Murray, 1971, 2 vols. (trad. cast.: El origen del hombre, México, Editores Mexicanos Uni-dos, 1982).
43 Christen, Y., o.c., p. 58. 44 Cfr. Morris, D., El modo desnudo. Un estudio del animal humano, Barcelona, Plaza
y Janés, 1968.

381
© Universidad de Deusto - ISBN 978-84-9830-383-4
tro de los mamíferos, que nacen con los párpados cerrados, como es el caso del ratón, el conejo, el gato, el perro y los monos infe-riores. A este tipo de especies animales se les denomina nidícolas,por la necesidad de permanecer más tiempo en el nido y bajo la protección de los progenitores, dada su deficiencia ocular. Otras especies, por el contrario, nacen con los párpados abiertos, como es el caso de los caballos, los terneros, las focas y los elefantes. Comprenden el grupo de los nidífugas, caracterizados por la capa-cidad de independizarse más rápidamente de sus progenitores. En relación a esta característica, la especie humana es una mezcla de ambos rasgos, en la medida en que nace con los párpados abiertos, coincidiendo con los nidífugas, pero su desvalimiento biológico45
tras su nacimiento le obliga a depender durante mucho tiempo de la ayuda materna, como ocurre con los animales nidícolas, aunque en mucha mayor medida que ellos. De este modo, los nidífugas re-capitulan a los nidícolas, más antiguos en el proceso evolutivo.
f) Órganos rudimentarios y otras anomalías morfológicas. Como in-dica Laín Entralgo, «desde que se hizo regla general la concepción filogenética de la morfogénesis, la existencia de órganos rudimen-tarios en el organismo humano —el ligamento redondo de la ar-ticulación de la cadera, el músculo piramidal del abdomen— viene siendo interpretada como una residual perduración de estructuras anatómicas que tuvieron forma y función mucho más acusadas en otras especies zoológicas»46. Es el caso también de la aparición de algunas anomalías morfológicas, como son las mamilas supernu-merarias, o también ciertas desviaciones de la norma en el caso de algunos músculos. También es cierto que estas anomalías pueden tener otra explicación: simples accidentes dentro del normal de-sarrollo del proceso embrionario.
4. EL ESPECIAL PUESTO DE LA ESPECIE HUMANA EN EL PROCESO ONTOGENÉTICO
Veíamos también, en el primer apartado de este capítulo, que la tesis de Haeckel de considerar la ontogénesis como una recapitulación de la fi-logénesis, fue sustituida por la teoría más acertada de la neotenia, funda-
45 Cfr. Gehlen, A., El hombre, Salamanca, Sígueme, 1980; Beorlegui, C., Antropología filosófica. Nosotros: urdimbre solidaria y responsable, o.c., pp. 393 y ss.
46 El cuerpo humano. Teoría actual, o.c., p. 63.

382
© Universidad de Deusto - ISBN 978-84-9830-383-4
mentalmente presente en la especie humana. La idea de recapitulación su-pone de algún modo que todas las especies vivas siguen una misma lógica dentro del proceso embriológico: madurar al nuevo ser hasta un momento en el que tendría la capacidad suficiente de sobrevivir de modo satisfacto-rio, encajarse en el ambiente, esto es, en el nicho ecológico propio de su especie, pudiendo defenderse autónomamente con una mínima ayuda de sus progenitores. En la mayoría de las especies vivas así ocurre, resultan-do la especie humana la que nace con una exagerada deficiencia biológi-ca. Si los individuos de la especie humana hubieran seguido esta lógica, el proceso de gestación en el útero materno vendría a durar, como indica Portmann47, alrededor de veintiún meses, es decir, un año más.
Según esto, resulta evidente que el ser humano ha invertido la lógica del proceso ontogenético, puesto que, en vez de alargarlo, lo acorta, na-ciendo antes de tiempo, y siendo un prematuro biológico. Como ya apun-tamos, en el ser humano se da el proceso de neotenia, de inmadurez. De tal forma que si comparamos los diversos momentos de su ontogénesis, en el momento de nuestro nacimiento nos parecemos al feto de un mono,como indica Y. Christen48. No hemos pasado rápidamente por las etapas de desarrollo de las especies anteriores, sino que ha ocurrido lo contrario: somos una especie de desarrollo lento que se ha detenido en las fases ini-ciales del proceso embriológico.
En esta condición biológica deficiente es donde se ha basado A. Gehlenpara construir su peculiar comprensión de la condición humana, en la que se complementa su deficiencia biológica con la necesidad de ser animal cultural49. El ser humano tiene que suplir con su capacidad intelectual las deficiencias biológicas, aprendiendo por una reiterada estrategia de ensa-yo y error a rechazar lo que le perjudica y adoptar lo que le favorece. Y el resultado de todo ese esfuerzo, en el que muchos han dejado la vida, es la cultura, que se va acumulando y transmitiendo a las generaciones siguien-tes. Esta actividad constituye el ejercicio de lo que Gehlen ha denomina-do la ley de la descarga, el esfuerzo por suplir la carencia de instintos por medio del aprendizaje de nuevas conductas.
De ahí la importancia del proceso de educación e inculturación infan-til, y sobre todo durante primer año de vida, el denominado por Portmann año extrauterino del embrión, en la medida en que el entorno familiar se convierte en un auténtico útero cultural. En este fenómeno típicamente
47 Cfr. Portmann, A., Die Biologie und das neue Menschenbild, Berna, 1942; Zoologie und das neue Bild des Menschen, Hambug-Reinbek, 1956; Krogmann, W. M., Child growth,University of Michigan, Ann Arbor, 1972.
48 Cfr. Christen, Yves, El hombre biocultural, o.c., p. 58. 49 Cfr. Gehlen, A., El hombre, o.c.

383
© Universidad de Deusto - ISBN 978-84-9830-383-4
humano se conjugan varios elementos que han experimentado el proceso de co-evolución: la inmadurez biológica, el nacimiento prematuro, la ca-pacidad de las mujeres de acortar y adelantar el alumbramiento, y la nece-sidad de alargar el proceso de ayuda y formación de los recién nacidos.
Aunque el proceso educativo que exige la neotenia se alarga considera-blemente, es decisivo el primero año posparto50, en la medida en que, al fi-nal del mismo, el recién nacido alcanza una cierta madurez en tres aspectos:
— maduración de la mano para ser capaz de manejar los diferentes ob-jetos,
— el dominio de las bases elementales del lenguaje, — y la bipedia, la capacidad de andar erguido.
En cada uno de los tres aspectos, la maduración es incipiente, pues-to que no se detiene ahí sino que dura y se completa a lo largo de varios años. En este proceso de progresiva maduración en estas y otras habilida-des, se conjugan tanto las disposiciones hereditarias como también el fac-tor ambiental, debido a la capacidad de imitar a sus congéneres. Pero para que esto sea posible, es necesario que el nuevo ser humano posea un cere-bro especialmente dotado para que pueda ejercer tan extraordinarias habi-lidades. El cerebro, como ya lo dijimos en su momento (capítulo tercero), aunque nace dotado de prácticamente todas las neuronas que constituyen su dotación personal, no está conformado por las innumerables conexio-nes sinápticas con las que se va a ir estructurando para ser capaz de con-vertirse en el instrumento que rija la conducta del recién nacido.
El cerebro del recién nacido posee alrededor de la mitad de su volumen en el momento de nacer, pero, como indicamos, se desarrollará de forma ex-traordinaria en los primeros años después del nacimiento. Debido a esto, ha sido posible el nacimiento de los fetos humanos, puesto que si naciéramos con las dimensiones cerebrales y craneanas que tenemos en la madurez, no podríamos salir del útero materno, en la medida en que no podría pasar la cabeza por el canal pélvico. La adopción del bipedismo permanente por la especie humana ha hecho que la pelvis de las mujeres se acorte y estreche, con los consiguientes cambios y estrechamiento del canal pélvico51. De he-
50 Cfr. sobre la importancia del primer año tras el nacimiento Michelena, Mariela, Unaño para toda la vida, Madrid, Temas de Hoy, 2002.
51 Cfr. Tortajada Martínez, Miguel, El parto en la evolución humana, Valencia, Uni-versitat de Valencia, 2001; Campillo Álvarez, J. E., La cadera de Eva. El protagonismo de la mujer en la evolución de la especie humana, Barcelona, Crítica (Ares y Mares), 2005; Arsuaga, J. L./Martínez, I., La especie elegida, Madrid, Temas de Hoy, 1998, cap. 10.º; Fisher, Helen, El contrato sexual, Barcelona, Salvat, 1987; Riu, Ester, «El embarazo cam-bió la columna de las mujeres», El País, 11 de marzo de 2008, p. 56 (hace referencia al ar-tículo de Catherine Whitcome y Liza Shapiro y otras, en Nature, sobre este tema).

384
© Universidad de Deusto - ISBN 978-84-9830-383-4
cho, los antropólogos advierten la diferente dificultad del parto de las hem-bras humanas, comparada con las de los chimpancés y las de los australopi-tecos (aunque de estas especies no se tienen fósiles completos de pelvis de hembras en edad de madurez). Las hembras de los chimpancés no necesitan ayuda de sus semejantes para dar a luz, proceso que no resulta especial-mente difícil ni peligroso. En el caso de los australopithecos, como indi-can J. L. Arsuaga e I. Martínez y otros especialistas52, los dos únicos restos fósiles de pelvis de australopithecos (la de Lucy, australopithecus affaren-sis, y la de Sts 14, australopithecus africanus) no nos permiten concluir de forma definitiva si en esas especies el parto se producía con la misma faci-lidad que en las hembras de los chimpancés, o con la dificultad de las hu-manas. La entrada del canal del parto en la pelvis de Lucy es muy ancha en sentido transversal, con lo que ello supondría de facilidad para el momento del parto, pero no podemos saber si se trata de una característica individual o de toda la especie, puesto que en el caso de la pelvis de Sts 14 el canal del parto es más redondeado.
Lo que está claro es que en el caso de la especie humana actual, el par-to es muy complejo, y el bebé tiene que hacer un giro de medio lado para poder sacar adelante la cabeza y la anchura de los hombros. Todo ello ha constituido una presión selectiva sobre la mujer capaz de adelantar el par-to, acortando el proceso de gestación, en la medida en que retrasarlo supo-nía la muerte tanto de la madre como del hijo. De tal modo que las mujeres que aprendían a adelantar el alumbramiento, conseguían un mayor éxito reproductivo en la medida en que tenían hijos más adaptados e inteligen-tes. Esta situación de prematuridad y de adelantamiento del parto conlleva al mismo tiempo un mayor riesgo para el feto, en la medida en que, por el giro que tiene que dar al atravesar el canal pélvico, el bebé nace de espal-das a la madre, con lo que las mujeres necesitan ayuda de otras personas para dar a luz felizmente a sus hijos, superando el riesgo de dañar su médu-la espinal. Este aspecto negativo se compensa, por otro lado, con el carác-ter casi obligadamente social e interpersonal del parto. Así, mientras en los primates el nacimiento es un hecho solitario, en la especie humana es un acontecimiento social. Wenda Trevathan, antropóloga de la Universidad de Nuevo México en Las Cruces, y autora de The Evolution of Human Child-birth (La evolución del parto humano)53, considera que gran parte del do-lor y ansiedad que acompañan al parto en las mujeres es de tipo cognitivo, de tal forma que la previsión de estas dificultades futuras lleva a las muje-res a buscar ayuda para el parto. Así, mientras la mayoría de los animales
52 Cfr. Arsuaga, J. L./Martínez, I., o.c., pp. 192 y ss. 53 Cfr. Angier, Natalie, «La ciencia explica por qué los bebés nacen de espaldas y des-
validos tras nueve meses», El País, 16 de septiembre de 1996, p. 30.

385
© Universidad de Deusto - ISBN 978-84-9830-383-4
buscan la soledad y un lugar tranquilo para dar a luz, las humanas nece-sitan ayuda para ese momento. También es cierto que en algunas culturas las mujeres dan a luz solas, sin ninguna ayuda, sobre todo cuando ya han tenido otros hijos anteriormente, como indica también Karen Rosenberg, de la Universidad de Delaware. Pero, en opinión de Trevathan, las muje-res buscan compañía no sólo por apoyo afectivo, sino por necesidad me-cánica, para que sus hijos nazcan sanos y sin lesiones. En los primates, los recién nacidos nacen con la cabeza por delante y mirando hacia la madre, mientras que en el caso de los humanos el recién nacido da la espalda a la madre, consecuencia, como ya hemos indicado, del proceso de bipedesta-ción que ha introducido en la estructura ósea de los humanos unos cambios significativos que obligan al bebé a realizar un giro en su salida del útero, quedando de espaldas a la madre. «Debido a esta presentación desfavora-ble, la madre no puede ayudar a salir al bebé sin doblarle hacia atrás la co-lumna vertebral, con riesgo de lesión medular o muerte. Tampoco puede desenrollar el cordón umbilical ni limpiar de mucosidad la boca del bebé. Para todas estas operaciones —cada una de las cuales puede suponer la di-ferencia entre la vida y la muerte para su hijo— necesita ayuda. Así surgió la práctica de las comadronas, o su equivalente ancestral»54.
En conclusión, los humanos ocupan un lugar especial en el desarro-llo embriológico porque unen un embarazo largo con un nacimiento in-maduro. Su proceso ontogenético es largo, comparado con el resto de las especies de los chimpancés. Este aspecto estaría en la lógica del proceso evolutivo, que parece necesitar más tiempo de maduración para confor-mar una especie nueva y más compleja. Pero lo curioso es que los hu-manos recién nacidos, como ya lo hemos indicado, son las criaturas más inmaduras, indefensas y desvalidas de todas las que existen, necesitando más que nadie la ayuda materna y del entorno social de su especie. Esto, ya lo hemos dicho también, es la fórmula que ha encontrado la evolución al dilema de un cerebro grande y una pelvis estrecha. El cerebro del feto humano crece hasta donde le es posible dentro del útero materno para po-der atravesar el canal pélvico materno antes de que sea demasiado tarde, y el resto de su maduración lo realiza tras el nacimiento, de modo espe-cial durante el primer año posterior (los recién nacidos siguen siendo fetos durante nueve meses), pero no dejando de crecer durante bastantes años después. Es probable que nuestros antepasados homínidos hubieran dado a luz a descendientes más precoces, pero la fórmula que apareció con los
54 Ibídem, p. 30. Sobre las comadronas y sus vicisitudes históricas en la asistencia a partos y su lucha con los médicos para reivindicar su rol, cfr. González-Crussi, Nacery otras dificultades. Historia cultural del enigma de la vida, Barcelona, Destino, 2008, pp. 139 y ss.

386
© Universidad de Deusto - ISBN 978-84-9830-383-4
humanos se ha centrado en conjugar un período largo de indefensión para los recién nacidos con un fuerte crecimiento cerebral fuera del útero.
Esta característica ontogenética de la especie humana es uno de los elementos que la hacen singular, puesto que su inmadurez cerebral y bio-lógica, la incapacidad de valerse por sí mismo, es consecuencia de la ruptura que en el ser humano se da respecto a las constricciones de tipo comportamental en la que se sitúa el resto de los animales. La inmadurez biológica, que hace que la especie humana no tenga conductas instintivas ni esté determinado en sus comportamientos por su estructura genética y cerebral, es la base de su indeterminación y de su libertad. El ser humano no sabe cómo actuar, teniendo que ser suplido y ayudado en estos prime-ros años por sus progenitores, y, después, en cuanto llega a su madurez psicobiológica, se encuentra necesitado y obligado a elegir entre varias opciones de actuación, teniendo que valorar entre las distintas posibilida-des que se le ofrecen, siguiendo sus propias inclinaciones y los criterios con que su entorno sociocultural le ha educado. Aquí se halla, por tanto, la apoyatura psicobiológica de la libertad y de su condición ética.
5. CUESTIONES FILOSÓFICAS PLANTEADAS EN EL TERRENO DE LA ONTOGÉNESIS
Todo el proceso de conformación ontogenética de un nuevo ser hu-mano plantea una serie de cuestiones filosóficas de enorme importancia y trascendencia, y que contiene decisivas consecuencias en el momento ac-tual dentro del área de la bioética.
Las cuestiones más importantes son tres:
a) La célula germinal, el zigoto, ¿es un ser humano, una persona, o todavía no?
b) ¿Cuándo llega el germen a ser un embrión de una persona? c) ¿Cómo se llega a ser persona?
Vamos a ir desentrañando y tratando de resolver y responder estas im-portantes cuestiones de un modo progresivo y ordenado.
5.1. ¿La célula germinal es un ser humano?
Antes de responder a esta cuestión central, hay que hacer dos preci-siones previas. La primera se refiere al hecho de que nos hallamos, tanto en este apartado como de igual forma en los dos siguientes, ante cuestio-

387
© Universidad de Deusto - ISBN 978-84-9830-383-4
nes que no se resuelven desde el nivel de la ciencia sino de la filosofía. Esto es, nos tenemos que desenvolver no en el terreno de la mera consta-tación de datos científicos, sino en el de las interpretaciones filosóficas y éticas. Eso no significa que la filosofía sea autosuficiente y pueda prescin-dir de los datos científicos. En absoluto. La filosofía depende y tiene que apoyarse en la ciencia, pero su terreno de reflexión se sitúa más allá o por encima de los datos científicos. La ciencia aporta los datos básicos, y la filosofía se esfuerza por interpretar y desentrañar los significados de los avances científicos.
Un segundo aspecto previo se refiere a la distinción que tenemos que hacer entre los conceptos de vida humana y el de persona. Otros si-túan la contraposición entre los conceptos de ser humano y de persona.Aunque defendamos la tesis de que existe la persona desde el momento en que se da vida humana, o ser humano, es lícito y necesario diferen-ciar ambos conceptos, en la medida en que corresponden a dos niveles diferentes de reflexión. Podríamos decir que el concepto de vida huma-na, o de ser humano, pertenece al campo de la ciencia, de la biología, mientras que el de persona se sitúa en el ámbito filosófico y ético. La pregunta acerca de si la célula germinal es ya un ser humano o una per-sona, puede responderse diciendo que sí es vida humana, o ser huma-no, en la medida en que se trata de una realidad que posee el cariotipocompleto de un ser perteneciente a la especie humana, es decir, es una célula que está dotada de 23 pares de cromosomas, como suma de las aportaciones de los dos gametos, el óvulo y el espermatozoide. Ningún genetista niega que la célula germinal sea una realidad nueva, dotada de todos los ingredientes específicos de la vida humana, y autónoma de la madre, aunque necesite del útero materno para sobrevivir. Su programa genético es específico de la especie humana, y se diferencia radicalmen-te de cualquier programa genético de otra especie. No tienen sentido, desde este punto de vista, las afirmaciones de determinadas feministas radicales cuando, para justificar una acción abortiva sin límites éticos, expresan tajantemente: «Yo hago con mi cuerpo lo que quiera». La cé-lula germinal y las demás células que surgen del proceso posterior de mitosis, está en el cuerpo de la madre, pero no es una parte más de su cuerpo, como pueda serlo un tumor o una berruga. Otra cosa diferente es afirmar que sea ya una persona, puesto que, mientras que las posturas más radicales dirán que sí, apoyándose en el hecho de que tiene ya to-dos los elementos potenciales para poder llegar a nacer y a desarrollarse como una persona, otras posturas consideran que la célula germinal no posee todavía los elementos necesarios y suficientes para poder llegar a convertirse en una persona humana. Atribuirle la condición de persona humana implica defender que se trata de un sujeto ético, poseedor de los

388
© Universidad de Deusto - ISBN 978-84-9830-383-4
derechos fundamentales de cualquier persona humana, aunque sea en potencia.
Por tanto, a la cuestión que nos ocupa se puede responder de varias maneras. Una postura dirá que la célula germinal, por ser vida humana,o ser humano, es ya también una persona, aunque todavía lo es sólo en potencia, puesto que, si no se incide de forma negativa y traumática en el proceso del desarrollo embrionario, llegará esa célula germinal a conver-tirse en cigoto y feto, para luego, tras el nacimiento, convertirse en una persona más de la especie humana. Otras posturas defenderán que la célu-la germinal, aunque es una vida humana, un ser humano (posee un carioti-po completo de la especie humana), no es todavía una persona, puesto que no tiene aún la totalidad de los elementos o notas esenciales, suficientes, para convertirse en una persona humana.
De ahí que algunos de los integrantes en el segundo bloque de opinio-nes prefieran hablar de preembrión al referirse a la fase anterior a la im-plantación del cigoto en la pared del útero, denominándolo a partir de en-tonces embrión postimplantatorio. En cambio, los que consideran persona en potencia ya a la célula germinal, hablan de embrión de cuatro células, de dieciséis o de las que sean, insistiendo con ello en la ilicitud de su ma-nipulación o eliminación.
Incluso alguna de las posturas que diferencian entre vida humana y persona vienen a decir que «no todas las personas son humanas ni todos los humanos son personas»55. Eso supone que se considera no persona al cigoto humano («no todos los humanos son personas»), y, al mismo tiem-po, se concede la condición de persona a determinados chimpancés inteli-gentes («no todas las personas son humanas»). Tendremos ocasión de exa-minar en el apartado siguiente las principales afirmaciones de esta postura de los defensores de los derechos de los animales.
La diferencias entre todas estas posturas se verán más explicitadas al presentar los diferentes puntos de vista en relación a la segunda de las cuestiones que tenemos que responder a continuación.
5.2. ¿Cuándo se llega a ser persona?
Ante esta cuestión fundamental se dan cuatro grandes opiniones dife-rentes. Cada una de ellas se apoya tanto en datos científicos como en su correspondiente visión filosófica y ética, deudora de un modelo antropo-lógico y de una cosmovisión determinada.
55 Engelhardt, T., The Foundation of Bioethics, Nueva York/Oxford, 1986, 107 (trad. esp.: Los fundamentos de la bioética, Barcelona, Paidós, 1995).

389
© Universidad de Deusto - ISBN 978-84-9830-383-4
a) La célula germinal es ya una persona
La primera postura defiende que existe la persona desde el principio, desde la célula germinal. Por tanto, para esta postura o no se considera pertinente distinguir entre ser humano y persona, o, si se admite, se con-sidera que coinciden extensivamente ambas realidades. Siempre que hay vida humana hay persona en potencia56. Esta postura la defienden los de-nominados movimientos pro-life y el magisterio oficial de la Iglesia Ca-tólica, aunque no siempre fue así a lo largo de su historia57. En opinión de M. Vidal, a pesar de que defiende la existencia de la persona desde la célula germinal, la postura oficial católica «deja la cuestión antropológi-camente abierta aunque moralmente la cierra siguiendo una orientación tuciorista»58. La apertura en lo antropológico se halla en el reconocimien-to de que «no hay sobre este punto una tradición unánime, y los autores están todavía divididos»59. Y, ante la duda teórica, se adopta la postura más segura en la defensa de la vida humana probable: el momento de la fecundación60.
Las razones científicas en las que se apoya esta postura se hallan en el hecho de que la célula germinal contiene un cariotipo completo y, des-de que el núcleo del espermatozoide se fusiona con el del óvulo, inicia un dinamismo autónomo y específico, aunque dependiente de los nutrientes del útero materno. Además, aunque acepta las diferentes etapas embrio-lógicas que se producen en el desarrollo ontogenético, entiende que todo el proceso de gestación forma un todo único, sin saltos cualitativos, y,
56 Cfr. VV.AA., Identidad y estatuto del embrión humano, Madrid, Ediciones Inter-nacionales universitarias, 2000: López Barahona, Mónica/Lucas Lucas, Ramón (dirs.), Elinicio de la vida. Identidad y estatuto del embrión humano, Madrid, BAC, 1999 (2.ª ed.); Junker-Kenny, Maureen, «El estatuto moral del embrión», Concilium, 1998, n.º 275, 73-87; Vidal, Marciano, «El status humano del embrión. Interrogantes y afirmaciones de la teolo-gía católica actual sobre el comienzo de la vida humana», en Vidal, M, Bioética. Estudios de bioética racional, Madrid, Tecnos, 1998 (3.ª ed.), 37-49; Forment, Eudaldo, «Estatuto ontológico del embrión», Espíritu, LI (2002), n.º 126, 257-270; Fernández Beites, Pilar, «Sustantividad humana: embrión y “actividad pasiva” de la inteligencia», en Murillo, Ilde-fonso (coord.), Filosofía práctica y persona humana, Salamanca, Publicaciones Universi-dad Pontificia de Salamanca, 2004, 244-255.
57 Para una visión completa y sintética de la postura de la Iglesia Católica en este pun-to, cfr. Vidal, M., Bioética. Estudios de bioética racional, o.c., cap. III, 37-49.
58 Ibídem, 45. 59 Declaración de la Sagrada Congregación para la Doctrina de la Fe sobre el aborto,
1974, n.º 13, nota 19. Cfr. Vidal, M., o.c., p.45.60 «Por otra parte, es suficiente que esta presencia del alma sea probable (y jamás se de-
mostrará lo contrario) para que arrebatarle la vida sea aceptar el riesgo de matar a un hom-bre, no solamente en expectativa, sino ya provisto de su alma», Declaración de la Sagrada Congregación ya citada, 1974, n.º 13, nota 19. Cfr. Vidal, M., o.c., p. 45.

390
© Universidad de Deusto - ISBN 978-84-9830-383-4
por tanto, en el cariotipo de la célula germinal, en su ADN, se dan todos los elementos esenciales y suficientes que llegarán a conformar definiti-vamente al nuevo ser vivo.
En el aspecto filosófico, esta postura es deudora, en gran medida, de los planteamientos aristotélicos de potencia y acto, en la medida en que, como hemos indicado, consideran que los componentes bási-cos y suficientes del futuro ser humano se hallarían ya presentes en su ADN. De ahí que la célula germinal sería ya una persona en potencia, poseedora de la densidad ontológica y de la dignidad ética que posee una persona humana adulta. La única diferencia está en que la perso-na adulta lo es en acto, y la célula germinal, en potencia. No conside-ra esta postura que se den saltos cualitativos en las diferentes fases o etapas del desarrollo embrionario conjunto. La única novedad o corte específico que hay que considerar es el inicio realmente nuevo que se da en la conformación de la célula germinal61. Todos los demás proce-sos forman una unidad indisoluble, que no aporta nada esencialmente, porque, «en principio, todos los rasgos emergentes se fundamentan en la unidad autorreproductiva básica del cigoto, de modo que sería arbi-trario afirmar que cualquier etapa en concreto introduce una diferencia cualitativa de tal calibre que sólo a partir de ella habría base suficiente para la personalidad. Es un ser humano único el que posee en sí mismo todas las predisposiciones necesarias para las ulteriores realizaciones; se trata de un potencial marcado por la identidad sobre una base ge-nética y a la vez por la continuidad tanto temporal como sustancial»62.Según esta forma de pensar, es erróneo afirmar que el embrión es po-tencialmente un ser humano; ni tampoco decir que los embriones se desarrollan hasta convertirse en seres humanos, sino que se desarro-llan como seres humanos63. Por tanto, «el quid de la cuestión del ar-gumento del potencial está en que lo decisivo para que se reconozca al embrión el derecho a la vida radica en su capacidad para convertirse en un sujeto moral, no en sus rasgos reales en un determinado momen-to. Este potencial de convertirse en un sujeto moral no se inicia con el nacimiento o con el uso de razón, sino que se despliega sobre la base de todo el proceso natural del desarrollo y forma parte de un contex-to continuo de vida humana a partir de las primeras etapas de la vida embrionaria. Se nos pide que respetemos las futuras oportunidades de vida en cualquier individuo en cualquier punto de su desarrollo a lo lar-
61 Cfr. Schockenhoff, Ethik des Lebens, M. Grünewald, Maguncia, 1993, 307 y ss.; Junker-Kenny, M., o.c., 81-82.
62 Junker-Kenny, M., o.c., pp. 81-82. 63 Cfr. ibídem, p. 82.

391
© Universidad de Deusto - ISBN 978-84-9830-383-4
go del tiempo, aunque carezca aún de un concepto reflexivo de su pro-pia continuidad»64.
La consecuencia ética que se extrae de estos planteamientos filosófi-cos y antropológicos es que cualquier manipulación o daño que se haga a este nuevo ser humano en cualquiera de sus fases, sea la célula germinal o cualquier otra fase del proceso de gestación, es una agresión moral si-milar a la que se comete con un ser humano ya nacido. Incluso en el caso de que se sea sensible a las investigaciones científicas que aportan datos que ponen en duda la tesis de la suficiencia entitativa como persona de la célula germinal, entienden los defensores de esta postura que son apor-taciones provisionales que tienen que probarse de un modo más serio, y que, por tanto, resulta más adecuada para la sensibilidad ética mantener posturas maximalistas (desde la célula germinal) que otras posturas más laxas que se apoyarían en datos científicos todavía no firmes ni suficien-temente contrastados. La razón de fondo, desde el punto de vista moral, para defender esta postura es la intención de defender al máximo toda vida humana, puesto que toda definición teórica tiene siempre de fondo una intencionalidad práctica. Por eso que, como indica M. Junker-Kenny, «si queremos proteger la nueva vida al máximo, habremos de utilizar una definición de mínimos, tal como la de “nueva unidad genética creada por el óvulo y el esperma”. Una definición de máximos de la vida humana, como sería la basada en la capacidad de comunicación, o de actuar inde-pendientemente, asegura una protección mínima en las etapas anteriores a la adquisición de esas capacidades o una vez que se han perdido. Una definición minimalista del inicio de la vida humana personal, capaz de asegurar la máxima protección, podría ser ésta: una vez que existe una unidad autorreproductiva en la que se han fusionado los núcleos del es-perma y el óvulo, ha de admitirse que esta nueva unidad posee la dignidad humana y los derechos humanos a la vida y a la inviolabilidad de su cuer-po. Esta unidad ha de considerarse un “embrión”, no un “pre-embrión”como se dice a veces»65. En definitiva, la vida humana ha de respetar-se, según estos teóricos, como un valor fundamental e integral. Y en este caso, sigue diciendo Junker-Kenny, «“integral” significa que no puede ser dividida en etapas de mayor o menor valor»66.
De todas formas, es importante tener en cuenta, nos indica Junker-Kenny, que «atribuir la personalidad humana al zigoto no implica que
64 Harris, J., «Embryos and Hedgehogs: On the moral status of the embryo», en Dyson, A./Harris, J. (eds.), Experiments on Embryos, Londres, Routledge, 1990, 65-81; 72.
65 Cfr. Junker-Kenny, Maureen, o.c., 80; los entrecomillado son de la autora.66 Ibídem, 81; las comillas son de la autora. Cfr. también Mieth, D., Geburtenregelung.
Ein Konflikt in der katolischen Kirche, Grünwald, Maguncia, 1990, 95-96.

392
© Universidad de Deusto - ISBN 978-84-9830-383-4
haya de ser siempre éste el que “gane” en los dilemas éticos. Lo que ocurre es que esto nos permite plantear esos dilemas y preguntarnos si el dere-cho del embrión a sobrevivir se ha de considerar igual, superior o subor-dinado a los derechos de la madre a la vida y a la autodeterminación»67
b) Persona: entre la anidación y la formación del sistema inmunológico
En la segunda postura se sitúan quienes retrasan varias semanas el inicio de la atribución de personalidad al embrión. Para estos autores, la célula germinal es una vida humana, un ser humano si se quiere, pero todavía no una persona. A la hora de indicar el momento en que se co-menzaría a poder hablar de persona, nos encontramos con posturas dife-rentes tanto en el tiempo como en las razones aducidas para ese retraso. Están quienes entienden que esa frontera se da al final de las dos pri-meras semanas de gestación, cuando se produce la anidación del cigoto en el útero materno, esto es, cuando la blástula o blastocisto se fija en la pared del seno materno. Hay quienes retrasan todavía más este mo-mento fronterizo hasta finales de la sexta u octava semana, dado que en ese momento es cuando se produce la emergencia de la cresta neural y se produce también la conformación definitiva del sistema inmunitario.Vamos a ir viendo las razones en las que se apoyan, tanto en lo científi-co como en lo filosófico y ético, los planteamientos de estas diferentes posturas.
Los defensores de situar el inicio de la persona humana en el mo-mento en que se produce la anidación, recurren a diversas experiencias científicas de no poco valor, y que están poniendo en cuestión la creencia hasta ahora dominante de que la célula germinal sea el único código ge-nético; o más concretamente, que en la célula germinal se dé toda la base genética potencial para el desarrollo posterior del nuevo ser humano. Es-tos datos son muy diversos y no todos del mismo peso, desde el hecho de que en esa fase de cigoto (dos primeras semanas), en el que la mórula va subiendo por las trompas de Falopio hasta fijarse en el útero, se produ-cen una impresionante cantidad de abortos espontáneos (casi la mitad de los casos), hasta los que hacen referencia a la no suficiente determinación específica de las células de esa primera fase, de tal forma que injertadas algunas células de una especie determinada en otras de diferente especie, pueden orientar el resultado de la gestación hacia la formación de seres vivos de la segunda especie, y no de la primera. Estos experimentos (rea-
67 O.c., 82.

393
© Universidad de Deusto - ISBN 978-84-9830-383-4
lizados en especies animales, pero pudiéndose perfectamente extrapolar a la especie humana) nos muestran que se da una mayor afinidad (por su indiferenciación y pluripotencialidad) entre células de diferentes especies de las etapas iniciales del proceso de gestación (cigoto), que entre célu-las de la misma especie pero pertenecientes a cualquier otra etapa poste-rior del proceso embriológico68.
Además, otro aspecto de interés consiste en advertir que el conjun-to de células que forman el blastocisto están ordenadas en dos capas: la externa, denominada trofoblasto o trofoectodermo (compuesto por unas cien células), y la masa celular interna o embrioblasto (formado por entre veinte y treinta células). Mientras la capa externa originará la placenta y otras membranas extraembriónicas, sólo la masa celular interna formará el embrión y posteriormente el feto. Esto plantea la evidencia de que no todas las células que proceden de la mitosis primera a partir de la célula germinal van a formar parte de la realidad humana que se va a ir forman-do hasta desembocar en el nacimiento. Según los datos científicos, tan sólo la quinta parte del blastocisto se convertirán en células que confor-marán el embrión y el futuro feto humano69.
Por tanto, para atribuir a la célula germinal el estatus de persona, se tiene que demostrar que ya desde ese momento se dan los rasgos específi-cos de toda persona, como son la unidad y la unicidad. Ahora bien, ponen en duda tal realidad fenómenos tales como la gemelación o la quimera-ción, y el proceso de conformación del sistema inmunológico70. La geme-lación es un proceso en el que de una sola célula germinal se desdoblan dos individuos (es el caso de los gemelos monocigóticos). Si lo que en un principio parecía se un solo ser humano, después se convierte en dos, hay motivos para concluir que, en esas primeras fases de gestación, se da to-davía una indeterminación sobre las condiciones esenciales que se necesi-tan para ser persona autosuficiente71. Y lo mismo ocurre en el caso de la formación de quimeras, proceso inverso al de la gemelación: donde había dos seres vivos se llega a formar una sola realidad, advirtiéndose también con ello una clara indeterminación en las fases iniciales. En la gemela-ción, no parece darse la unicidad desde el comienzo, y en la quimeración,no se da la unidad.
68 Cfr. Laín Entralgo, P., Nuestro cuerpo. Teoría actual, o.c., cap. 2.º. 69 Cfr. Lacadena, J. R., Genética y Bioética, o.c., cap. 1, pp. 51-71; Abrisqueta, J. A.,
«El embrión humano: estatuto antropológico y ético», en Vidal, M. (dir.), Conceptos fun-damentales de ética teológica, Madrid, Trotta, 1992, 439-455; 441.
70 Cfr. Lacadena, J. R., Genética y Bioética, o.c., 60 y ss.; Id., «Biología y humanidad: hominización y humanización», en Masiá, Juan (ed.), Ser humano, persona y dignidad,Madrid/Bilbao, UPCO/Desclée de Brouwer, 2005, 43-89.
71 Cfr. Lacadena, J. R., «Persona y humanidad», o.c., pp. 84.

394
© Universidad de Deusto - ISBN 978-84-9830-383-4
Un elemento también muy significativo, que apuntala la razonabilidad de esta postura, es el proceso de conformación del sistema inmunológi-co72. Todas las células de un ser vivo poseen el mismo cariotipo, la mis-ma estructura de su ADN, puesto que son replicaciones de una sola célula inicial, la célula germinal, aunque a lo largo del proceso embrionario se han ido especializando en células diversas: neuronas, células sanguíneas, óseas, musculares, etc. En los organismos pluricelulares complejos, el ser vivo ha ido conformando un sistema de defensa del organismo, basado en la capacidad de diferenciar y distinguir entre las células propias del orga-nismo y las diferentes e intrusas, posibles portadoras de peligros e infec-ciones. El sistema inmunológico es el encargado de realizar este proce-so de identificación y de proceder a la defensa y expulsión de cualquier componente peligroso. Los estudiosos del sistema inmunitario calculan entre 6 y 8 semanas el plazo necesario para su conformación dentro del desarrollo embrionario. En la medida en que «el concepto de identidad o mismidad genética de un individuo está íntimamente relacionado con su capacidad genética de distinguir lo propio y extraño»73, función realizada por el sistema inmunitario, el momento de cristalizar la unidad y unicidaddel embrión, parece que se produce en fases muy posteriores a la forma-ción de la célula germinal.
La conclusión que saca J. R. Lacadena es que así como en el caso de los fenómenos de gemelación y quimeración «se hace hincapié en que hacia el día 14 después de la fecundación se podría decir que queda es-tablecida la individualidad del nuevo ser humano al quedar fijadas las propiedades de unicidad y unidad, podemos ahora plantearnos la cues-tión de cuándo en el desarrollo del individuo se actualiza su identidad o mismidad genética»74. Este proceso queda conformado o explicitado con el sistema inmunológico, entre la sexta y octava semana, como hemos in-dicado ya.
Otros autores suelen considerar como dato fundamental para atribuir al embrión la condición de persona la aparición de la cresta neural, esto es, el momento en que aparece la base biológica que dará lugar al cere-bro75. Ya hemos hecho en apartados anteriores referencia a este momento en el proceso embriológico, así como a las características que la apari-ción de dicha cresta neural supone. Esta postura parte de la definición de persona como realidad racional, de ahí que considere fundamental como
72 Cfr. ibídem, pp. 85-87. 73 Ibídem, p. 87.74 Ibídem, p. 87. Las cursivas son del autor. 75 Ledouarin, Nicol, «La cresta neural juega un papel clave en la evolución», El País,
24 de abril de 2002, p. 33.

395
© Universidad de Deusto - ISBN 978-84-9830-383-4
momento de conferir al embrión la dignidad de persona, el que posea al menos la base biológica del órgano por excelencia de la racionalidad, el cerebro. Así que sería en ese momento cuando, para esta teoría, comenza-ría a existir la persona y, por tanto, a partir de ese momento estaría ya do-tado el embrión de toda la dignidad y de los derechos correspondientes.
La teorización antropológica de este conjunto de posturas conside-ra insuficiente e inadecuada la propuesta aristotélica de potencia y acto, basada en una noción de sustancia o esencia humana ya superada, para proponer y apoyarse en otras teorías antropológicas, como la de X. Zubi-ri, basada en la noción de sustantividad y de suficiencia entitativa76. Para esta postura, la dotación genética del nuevo ser humano no está sólo con-formada por el ADN, puesto que el desarrollo del embrión no sólo res-ponde a una dinámica interna, sino que depende también de su entorno o campo (campo morfogenético). Y el campo, o la condición campal del embrión, ya quedó dicho que no es algo externo sino una dimensión física del propio embrión.
Por los datos científicos que se han indicado más arriba, parece con-cluirse que el zigoto no posee en sí mismo una determinación absoluta que garantice que llegará a ser una persona humana. Tal determinación depende, en cada momento, no sólo de la progresiva actualización de su propia información genética, sino también de la actualización de las in-formaciones surgidas de novo durante el proceso embrionario, así como de diversas informaciones exógenas. Por ello, para estos autores no está ahí ya toda la persona en potencia, sino que sólo se puede hablar del ADN como potencia condicionada (Laín Entralgo habla de potentia conditio-nata), en la medida en que el zigoto no posee, ni siquiera potencialmente, toda la información necesaria para orientar el desarrollo del embrión. Y la actualización de su potencialidad morfogenética no es enteramente autó-noma, y, además de diacrónica, es cualitativamente discontinua77. Desde un planteamiento antropológico basado en la filosofía de Zubiri, la sus-tantividad humana en ciernes está conformada por una estructura de notas (unas de tipo somático y otras de tipo psíquico) que sólo tras varias sema-nas de gestación, a tenor de los datos científicos anteriormente señalados, tendrían suficiencia entitativa para existir. En consonancia con esta postu-ra, «yo considero, afirma Carlos Alonso Bedate, que el fin de la cuarta se-
76 Cfr. Gracia, Diego, «Problemas filosóficos en genética y en embriología», en Abel, F./Cañón, C. (eds.), La mediación de la filosofía en la construcción de la bioética, Madrid, UPCO, 1993, 215-254; Beorlegui, C., «Humanismo y bioética (Un nuevo planteamiento del estatuto del embrión desde un nuevo concepto de persona)», Bilbao, Universidad de Deusto (Congreso sobre Humanismo), 2003, publicado en CD (soporte informático).
77 Cfr. Laín Entralgo, P., Nuestro cuerpo. Teoría actual, o.c., cap. 3.º.

396
© Universidad de Deusto - ISBN 978-84-9830-383-4
mana inicia una etapa crucial, en cuanto al valor biológico, y también éti-co del embrión, porque representa la constitución de un sistema en el cual el sujeto que va a nacer empieza a estar definido, aunque no sea más que en forma de esbozo»78.
Los defensores de esta postura consideran que el momento decisivo del proceso embriológico es el paso o conversión del blastocisto en gás-trula, en la medida en que es entonces cuando se establece de manera fir-me la definitiva hominización del embrión humano. Hasta entonces, no era unívoca y definitivamente humana la teleonomía de la génesis; desde entonces, ya lo es79.
La cuestión que ahora se plantea es qué consecuencias de tipo éticose deducen de estas afirmaciones científicas y filosóficas. Los que defien-den que el estatuto antropológico del embrión comienza varias semanas después de la fecundación, no niegan que tengamos obligaciones éticas sobre el cigoto desde sus primeros momentos, sino que el tipo de obliga-ción moral que se tenga será diferente en las primeras semanas que a par-tir de fases posteriores. Siguiendo estos planteamientos, Carlos Alonso Bedate considera que «si la condición de persona es necesaria para poder tener el valor asimilable a la persona, no encuentro argumentos para atri-buir al embrión la condición de dignidad de la persona y el valor que se deriva de la misma. De hecho, conferimos dignidad a elementos no huma-nos. Mi posición es que habrá que determinar si la obligación de respeto es exactamente igual para quien tiene dignidad “inherente” y para quien tiene dignidad “conferida”. Creo que no. Si el embrión es asimilable a la persona no cabe entonces duda que la obligación moral que genera es de respeto»80.
De forma similar se pronuncia Diego Gracia81, razonando desde su específica propuesta de jerarquización de los cuatro grandes principios
78 Cita tomada de Laín Entralgo, P., o.c., p. 91; Alonso Bedate, Carlos, «Reflexiones sobre cuestiones de vida y muerte: Hacia un nuevo paradigma de comprensión del valor ético de la entidad biológica humana en desarrollo», en Abel, Fr./Bone, E./Harvey, J. C. (dirs.), La vida humana: origen y desarrollo, Madrid, UPCO/Barcelona, Instituto Borja de Bioética, 1989, 57-81.
79 Cfr. Abel, Fr./Bone, Edouard/Harvey, John C. (dirs), La vida humana: origen y de-sarrollo, o.c.; Gracia, Diego, «El estatuto del embrión», en Gafo, Javier, Procreación hu-mana asistida: aspectos técnicos, éticos y legales, Madrid, UPCO, 1998, 79-109; Masia, Joan, Bioética y antropología, Madrid, UPCO, 1998;
80 Alonso Bedate, Carlos, «El estatuto ético del embrión humano: una reflexión ante propuestas alternativas», en Mayor Zaragoza, F./Alonso Bedate, C. (coords.), Gen-Etica,Barcelona, 2003, 19-66 (cita tomada de J. R. Lacadena, o.c., p. 88).
81 Cfr. Gracia, Diego, «Problemas filosóficos en Genética y en Embriología», en Abel, F./Cañón, C. (eds.), La mediación de la Filosofía en la construcción de la Bioética, Madrid, 1993, 215-254 (cita tomada de J. R. Lacadena, o.c., p. 89).

397
© Universidad de Deusto - ISBN 978-84-9830-383-4
de la bioética (no maleficencia, beneficencia, autonomía y justicia). Para D. Gracia, el estatuto antropológico y ético del embrión comienza en tor-no a la octava semana, en la medida en que alrededor de ese momento se daría la suficiencia constitucional y entitativa, en terminología zubiria-na. Anteriormente no tendría todavía las notas esenciales suficientes para conformar la estructura completa, esencial, de la realidad humana. Eso no llevaría a la drástica conclusión de que la fase zigótica no poseería ningún valor ético, sino que su densidad ética sería de diferente nivel que a partir de la octava semana. De este modo, si los principios de no maleficencia y de justicia obligan de modo universal, no ocurre lo mismo con los de autonomía y beneficencia, cuyo estatus es más bien de consejo. Por tanto, sólo a partir de la octava semana, cuando la persona está ya conformada con las notas constitucionales, poseería ya la dignidad ética propia de toda persona. Pero en las fases anteriores, el respeto hacia ella sería de tipo de consejo, según la conciencia moral de cada uno82.
c) La postura emotivista y utilitarista
Los planteamientos de esta corriente se apoyan en presupuestos dis-tintos a los de las dos posturas anteriores. Defienden la distinción entre vida humana y persona, como la segunda postura, pero proponen que para poder considerar a alguien como persona, tiene que poseer en acto los rasgos específicos de las personas, como son el sentir y la racionali-dad. No es suficiente considerar que se tiene potencialidad de poseer tales atributos, sino que los tiene que tener en acto. De ahí que los defensores de este planteamiento consideran que «no todas las personas son huma-nos, ni todos los humanos son personas» (T. Engelhardt). Según esto, los embriones humanos no serían personas, y sí lo serían los animales perte-necientes a algunas especies de primates (y otras especies de animales inteligentes, como los delfines o las ballenas), porque muestran eleva-dos niveles de sentimientos y de inteligencia. Es la forma de pensar de P. Singer y de John Harris, y de otros teóricos defensores de los derechos de los animales y partidarios de extender a determinadas especies inteli-gentes la condición de personas.
El relación a la discusión sobre la condición de personas de los em-briones, el filósofo inglés J. Harris, afirma: «Realmente sólo hay dos maneras de abordar esta cuestión: una es en términos de estatuto moral efectivo del embrión en un determinado momento y otras en términos del
82 Cfr. Gracia, D., o.c.; Beorlegui, C., «Humanismo y bioética», o.c.,

398
© Universidad de Deusto - ISBN 978-84-9830-383-4
potencial del embrión para adquirir un estatuto moral o una personalidad que de momento aún no posee». Como puede suponerse, Harris opta por la primera de las opciones, y considera incorrecta la segunda. En sus pro-pias palabras: «Afirmo que el estatuto moral del embrión y, para el caso, para cualquier otro individuo viene determinado por el hecho de que po-sea aquellos rasgos que hacen a los adultos humanos normales moralmen-te más importantes que las ovejas o las cabras o los embriones»83. Esos rasgos que se consideran específicos de las personas son la racionalidad y la autoconciencia.
Ahora bien, los rasgos específicos de la personalidad, para esta corrien-te de pensamiento, no son tanto la racionalidad y autoconciencia, sino sobre todo la capacidad de sentir, de sufrir y de gozar. Precisamente se desmarcan del antropocentrismo y del humanismo de la antropología do-minante en la filosofía occidental, defendiendo que la base para funda-mentar la dignidad y los derechos de los individuos es no tanto la raciona-lidad sino los sentimientos, en la línea de J. Bentham y de J. Stuart Mill. Así, pensadores como Singer y los que firman con él el denominado Pro-yecto Gran Simio84, se sitúan en la línea de la fundamentación emotivistay utilitarista de la moral85. Para el utilitarismo, partiendo de un criterio de igualdad, la bondad o maldad de las acciones se halla en relación con la maximización de los beneficios que consiga, así como con el mayor o menor daño que evite. Por tanto, la norma moral por excelencia consistirá en hacer el máximo bien posible al mayor número de personas. Para ello, prescinde de la bondad intrínseca de los actos a la hora de considerar-los buenos o malos, fijándose más bien en sus consecuencias, esto es, en maximizar su utilidad.
El problema del utilitarismo es que, debido a que no entra en precisiones principialistas sobre qué entendemos por bien y mal, o por esencia inherentea la personalidad, no sabremos distinguir entre el bien y el mal de nues-tras acciones hasta después de que las hayamos realizado (consecuencia-lismo). Por evitar enredarse en cuestiones esencialistas, centrándose en la supuesta rapidez utilitarista, no puede responder a cuestiones como qué tipo de bienes son los que tienen que ser extendidos y los males a evitar, con lo que ponen en cuestión el fin central de su tesis utilitarista.
83 Harris, J., «Embryos and Hedgehogs: On the moral status of the embryo», en Dyson, A./Harris, J. (ed.), Experiments on Embryos, Londres, Routledge, 1990, 65-81; 79 (citas toma-das de Junker-Kenny, M., o.c., 75.
84 Cfr. Singer, P./Cavalieri, P. (eds.), El proyecto «Gran Simio». La igualdad más allá de la humanidad, Madrid, Trotta, 1998.
85 Cfr. Beorlegui, C., «Etica animal e idea de persona», en Guibert Ucin, J. M./Orella Unzué, J. L. (eds.), Preguntas éticas en cuestiones disputadas hoy, San Sebastián, Univer-sidad de Deusto, 2005, 317-335.

399
© Universidad de Deusto - ISBN 978-84-9830-383-4
Peter Singer se ha dado a conocer en el ámbito intelectual por sus te-sis en defensa de los derechos de los animales86, y por el intento de reno-var los planteamientos éticos desde bases diferentes al trasfondo religioso y humanista y antropocéntrico con que se ha solido fundamentar la ética occidental87. Sus planteamientos han sido objeto tanto de entusiasta acep-tación como de críticas de todo tipo88. Pero nos vamos a centrar aquí sólo en sus ideas sobre el estatuto ontológico de los embriones humanos y las consecuencias éticas que extrae de este modo de pensar.
Para Singer, al igual que para Harris, hay una diferencia evidente entre vida humana y persona, y sólo ésta es sujeto de derechos. No se puede negar que hay vida humana desde la célula germinal, pero sólo muy posteriormente se podrá hablar de alguien capaz de sentir, tener una incipiente capacidad cerebral y ser sujeto de intereses que tenemos obli-gación de respetar y defender. Es consciente de que el desarrollo del ser humano es un proceso gradual, en el que resulta muy difícil establecer fronteras y saltos claros89, de modo que no duda en afirmar: «no hay nin-guna línea evidente que divida al óvulo fecundado del adulto, y de ahí el problema»90. Y repite las dificultades esgrimidas normalmente por parte de la segunda postura para oponerse a las tesis de la primera postura ya presentadas aquí, como son la indiferenciación de las células que van a conformar el embrión y las que formarán la placenta, los fenómenos de gemelación y quimeración, la tardía formación del sistema inmunitario, etc. Para Singer, esas razones no son tan fundamentales, considerando que lo central para no reconocer en esa fase a una persona es que el em-brión no tiene capacidad de sufrir, ni la base de lo que luego será su ca-pacidad de conciencia.
86 Cfr. Singer, P., Liberación animal, Madrid, Trotta, 1999. 87 Cfr. Singer, P., Etica práctica, Barcelona, Ariel, 1984; Repensar la vida y la muer-
te: el derrumbe de la ética tradicional, Barcelona, Paidós, 1997; Una vida ética: escritos,Madrid, Santillana, 2000; Un solo mundo. La ética de la globalización, Barcelona, Paidós, 2003.
88 Cfr. Torralba Roselló, Fr., ¿Qué es la dignidad humana? Ensayo sobre Peter Singer, Hugo Tristram Engelhardt y John Harris, Barcelona, Herder; Beorlegui, C., «¿Hacia un humanismo trans-antropocéntrico? Peter Singer y los “derechos” de los animales», Reali-dad (UCA, El Salvador), 2001, n.º 80, 199-236; Id., «Etica animal e idea de persona», en Guibert Ucin, J. M/Orella Unzué, J. L. (eds.), Preguntas éticas en cuestiones disputadas hoy, San Sebastián, Universidad de Deusto, 2005, 317-335; Francione, Gary L., «El error de Bentham (y el de Singer)», Teorema, XVIII/e (1999), 39-60; Lacadena, J. R. (ed.), Losderechos de los animales, Madrid, UPCO/Bilbao, Desclée de Brouwer, 2002.
89 Para ver las ideas centrales sobre este tema, seguiremos su obra Etica práctica, Nue-va York/Melbourne, Cambridge University Press, 1993 (2.ª ed.), cap. 6, «Quitar la vida: el embrión y el feto» (pp. 167-215).
90 Ibídem, p. 170.

400
© Universidad de Deusto - ISBN 978-84-9830-383-4
Es cierto que el embrión es un individuo humano en potencia, Pero, para Signer, no es suficiente la tesis de la potencialidad, porque para con-ceder a algo la categoría de persona y sus derechos consiguientes, no hay que fijarse en su potencialidad sino en la posesión actual de tales rasgos. Y para él, como ya hemos dicho, no sólo tendrán derechos las personas humanas sino también determinados animales inteligentes y sensibles.
Singer advierte en la postura conservadora un silogismo de fondo que vendría a expresarse así:
— Premisa mayor: es malo matar a un ser humano inocente.— Premisa menor: un feto humano es un ser humano inocente. — Conclusión: por tanto, es malo matar a un feto humano.
Las posturas que Singer denomina liberales se esfuerzan en negar la premisa menor, en la medida en que se sitúa tal realidad en fases poste-riores. Pero los conservadores retan a los liberales a explicitar qué etapa del proceso embriológico es capaz de marcar una clara línea divisoria que resulte moralmente significativa. Si no se consigue tal línea divisoria, hay que considerar al zigoto con la misma dignidad que al niño recién naci-do. Singer advierte que todos los intentos de señalar líneas divisorias son argumentaciones sin suficiente éxito probativo. Así, indicar que esa línea puede ser el momento del nacimiento no resulta convincente, pues no se ve por qué tiene más dignidad un recién nacido que un feto al que le fal-tan aún días o meses por nacer. El que a uno lo veamos y al otro no, no es razón para marcar diferencias cualitativas. Dificultades similares tiene la referencia a la viabilidad, en la medida en que es un concepto relati-vo: depende de la tecnología médica, que avanza considerablemente, así como de los medios económicos poseídos. Así, «un feto de seis meses po-dría tener una buena posibilidad de sobrevivir si nace en una ciudad don-de se utiliza la más reciente tecnología médica, y ninguna en absoluto si nace en un pueblo lejano del Chad o de Nueva Guinea»91. Y tampoco tie-ne sentido afirmar que, al ser el feto totalmente dependiente de la madre para su supervivencia, no tiene derecho a la vida independiente de los de-seos de la madre. Pero eso lo mismo podríamos decir de un niño pequeño, y no por eso defendemos la moralidad del infanticidio. Similar debilidad argumentativa tiene referirse como línea divisoria al movimiento del feto: al moverse estaría mostrando que es autónomo respecto a la madre. El problema consiste en que el feto se empieza a mover hacia la sexta sema-na después de la fecundación, pero se empiezan a sentir esos movimien-tos mucho después. Y, por otro lado, «la capacidad de movimiento —o la
91 Ibídem, p. 173.

401
© Universidad de Deusto - ISBN 978-84-9830-383-4
falta de él— no tiene nada que ver con la seriedad de la pretensión de una vida continuada. No vemos la falta de tal capacidad como un argumento para negar a la gente paralítica sus aspiraciones a seguir viviendo»92.
Podría tener mayor peso el argumento de la movilidad si se conside-ra una indicación o anticipo de alguna forma de conciencia, porque para Singer es fundamental como condición de la existencia de la persona la conciencia y la capacidad de sentir dolor o placer. Pero también en este punto es muy difícil señalar fronteras exactas, ya que, aunque se puede advertir cierta actividad cerebral hacia la séptima semana, no se puede es-tar seguro de ello de un modo contundente y objetivo. Desde este punto de vista, reconoce Singer que «la posición conservadora se mueve en un terreno firme al insistir (en) que el paso del embrión al niño es un proceso gradual»93.
El peso de las argumentaciones liberales se orientan no tanto a negar que el feto sea un ser humano inocente, sino más bien a defender que el aborto es permisible, basándose tanto en que las consecuencias de una ley que lo prohíba traerá más abortos y en situaciones peligrosas para la ma-dre (sobre todo en el caso de mujeres sin recursos), como también indi-cando que este tipo de decisiones tan privadas no debiera ser competencia de leyes publicas reguladas por el derecho. El problema de esta segunda argumentación está en que, si bien se puede defender para casos en que esos actos privados no hacen daño a nadie, no tienen víctimas, en el caso del aborto defender que hay o no víctimas depende de si consideramos a los fetos personas humanas o no. Y tampoco valen para Singer las argu-mentaciones feministas apoyadas en que la mujer es dueña de su cuerpo, puesto que no lo es de la nueva vida que está creciendo en él, aunque de-penda su desarrollo y su llegada al nacimiento de la concurrencia necesa-ria del cuerpo de la madre.
Singer presenta su planteamiento considerando que la premisa ma-yor (es malo matar a un ser humano inocente) del silogismo ya indicado más arriba y sobre el que se basa la postura conservadora, no es tan fir-me como se piensa. Es convincente si se acepta la premisa previa de la santidad o valor absoluto de toda vida humana, como así lo defienden las posturas religiosas, humanistas y antropocéntricas. Pero, para Singer, el concepto de humano tiene doble acepción: pertenecer a la especie hu-mana (homo sapiens) y ser persona. En los dos casos, considera Singer que la premisa conservadora es falsa. Si se toma humano en el sentido de persona, es falso, porque «no se puede argumentar que el feto sea racio-
92 Ibídem, p. 175. 93 Ibídem, p. 176.

402
© Universidad de Deusto - ISBN 978-84-9830-383-4
nal o consciente de sí mismo»94. Ya hemos indicado que para Singer no tiene sentido referirse al feto como potencialidad de ser humano. Para definir si algo es persona o no, ese algo tiene que poseer los rasgos (con-ciencia y capacidad de dolor y de placer) de la persona en acto, no como mera potencialidad. Y si se considera humano en el sentido de miembro de la especie homo sapiens, tampoco la premisa será verdadera, porque esa afirmación no tiene pertinencia moral, no pertenece al ámbito de la moral, sino al de la ciencia. Por ello, según Singer, «que un ser sea o no miembro de nuestra especie no es en sí mismo más pertinente al hecho de que matar esté mal que el hecho de que sea o no miembro de nuestra raza»95. Y ello se basa en que afirmar que matar a un individuo está mal por el hecho de pertenecer a nuestra especie, lo considera Singer como es-pecieísmo, válido sólo para los humanistas de orientación religiosa, pero esa afirmación hay que demostrarla con otros argumentos.
Desmontados, pues, esos argumentos, considera Singer que el proble-ma del estatuto del embrión y los problemas morales implicados en ello, hay que situarlos en otro horizonte. Y en primer lugar, como hemos indi-cado ya, hay que considerar al feto por las características reales poseídas en acto, no tanto por sus posibles potencialidades. Según ello, entiende Singer que la postura que se autoproclama pro-vida, tiene una denomi-nación errónea, puesto que sólo defiende las vidas humanas, pero se des-preocupa de otras vidas animales, que, en opinión de los defensores de los derechos de los animales, merecen a veces tanta consideración o más que determinadas vidas humanas. Si consideramos propio de una persona ras-gos como la racionalidad, la conciencia de uno mismo, el conocimiento, la autonomía, el placer, el dolor, y otros similares, veríamos que muchos animales (ya no sólo los primates, sino otros como un cerdo, una ternera o una gallina) «se encuentran muy por delante que el feto en cualquier eta-pa del embarazo, mientras que si hacemos la comparación con un feto de menos de tres meses, un pez mostraría más señales de conciencia»96.
Por tanto, siguiendo la tesis de que no todas las personas son huma-nas, ni todos los humanos son personas, Singer defiende que no hay que dar más valor a la vida del feto humano que a la de un animal no humano, si posee un cierto grado de racionalidad, de autonomía y de capacidad de sufrir y de gozar. Se trata, por tanto, de ampliar el concepto de persona,extendiéndolo a otras especies animales cercanas a la nuestra, así como de restringir la condición de persona entre los humanos a los individuos ca-paces de ejercer las cualidades correspondientes a la personalidad.
94 Ibídem, p. 186. 95 Ibídem, p. 186. 96 Ibídem, p. 187.

403
© Universidad de Deusto - ISBN 978-84-9830-383-4
El problema está en dilucidar en qué momento el feto es capaz de sentir dolor, cualidad que para Singer marca la frontera fundamental de la personalidad. Sólo el feto que tenga esa capacidad tendrá la cualidad de persona, puesto que no es consistente para él, como hemos indicado, para atribuirle al feto un valor intrínseco por el mero hecho de pertenecer bio-lógicamente a la especie homo sapiens. Cuando el feto tenga esa capa-cidad de sufrir, será un sujeto capaz de tener intereses: la defensa de su bienestar. Y sólo a partir de entonces, habrá que discutir sobre los intere-ses del feto y de la mujer que pretende abortar.
En la medida en que Singer se sitúa en posturas éticas emotivistas y consecuencialistas, su enfoque sitúa los problemas comparando los inte-reses de los individuos que se hallan en conflicto, siguiendo el imperativo utilitarista de hacer el mayor bien al mayor número de personas. En fun-ción de ello, defiende la posibilidad del aborto en cualquier momento del embarazo, así como la posibilidad del infanticidio. Eso le llevará a decir, al afrontar las orientaciones éticas para la utilización de animales en el campo de las investigaciones de laboratorio, que «prefiero investigar con un feto humano que con una cobaya»97. No se niega a la utilización de animales como objetos de investigación, con tal de que no se les cause un dolor desproporcionado y se realice la investigación con las condiciones de respeto hacia la vida de los animales, y se saquen unos beneficios ade-cuados de tales prácticas.
Como puede verse, entendemos que el punto central está en que, para Singer, hay que considerar las características reales y no potenciales del feto y de los demás animales98. La cuestión está, por tanto, en ver cuándo podemos atribuir al feto humano algún atisbo de conciencia y de capaci-dad de dolor y de placer. Singer considera que eso hoy día se sabe ya con bastante certeza. «La parte del cerebro asociada con las sensaciones de dolor, y de forma más general con la conciencia, es la corteza cerebral. Hasta las 18 semanas de gestación, la corteza cerebral no está lo suficien-temente desarrollada para que tengan lugar conexiones sinápticas dentro de él, en otras palabras, no se reciben las señales que dan lugar al dolor en un adulto. Entre las 18 y las 25 semanas el cerebro alcanza una etapa en la cual existe algún tipo de transmisiones nerviosas en esas partes asociadas con la conciencia. Incluso entonces, sin embargo, el feto parece estar en estado continuo de sueño y, por tanto, puede no ser capaz de recibir dolor. El feto empieza a “despertarse” a unas 30 semanas de gestación. Natural-mente esto queda bastante más allá de la etapa de viabilidad, y un “feto”
97 Cfr. Sampedro, Javier, Entrevista a Peter Singer, El País (Pabelia), 11 de mayo de 2002, p. 12; cfr. también Singer, Etica práctica, o.c., pp. 209 y ss.
98 Cfr. Singer, P., Etica práctica, o.c., 188 y ss.

404
© Universidad de Deusto - ISBN 978-84-9830-383-4
que estuviera vivo y fuera del útero en esta etapa sería un niño prematuro y no un feto»99.
Volvemos a advertir de nuevo la dificultad de establecer fronteras ní-tidas, y de ahí que Singer, ante la duda, propone que se sitúe el límite para empezar a proteger los intereses del feto en el primer momento en que puede sentir algo, hacia la 18 semana de la gestación. Antes de esa fecha, el feto no tendría intereses que defender, porque antes de esa fecha el feto no puede ser perjudicado, en opinión de Singer. Sólo a partir de entonces, el feto sí necesita ser protegido de posibles daños, «con la misma base que lo necesitan los animales no humanos que sienten, pero que no son cons-cientes de sí mismos»100.
En conclusión, a la cuestión de si vale más la vida de los seres humanos que la vida del resto de los animales, Singer responde que depende de qué humanos y de qué animales. «No tiene sentido decir que la vida de todos y cada uno de los seres humanos sea mejor que la vida de todos y cada uno de los animales no humanos. Porque algunas personas nacen con discapacida-des psíquicas irreparables, trágicamente tan graves, en ocasiones, que ni si-quiera son capaces de responder a sus madres. La vida de estos seres huma-nos no es superior o más valiosa que la de los perros, los cerdos o las vacas. Por tanto, sería mejor preguntar: ¿tiene sentido afirmar que la vida de un ser con las características de una persona normal y madura es más valiosa que la de un cerdo, vaca o perro normal y maduro? En este caso sí cabe respon-der afirmativamente. Es razonable sostener que nuestra vida adquiere valor por nuestra capacidad de planificarla en su conjunto, de educarnos a noso-tros mismos, de realizar proyectos a largo plazo y muchas otras cosas que están fuera del alcance de otros seres de menor capacidad intelectual»101.
d) La persona como entidad relacional: a partir del nacimiento
Hay una cuarta postura, más extrema, que pospone la adscripción de la condición de persona hasta el momento del nacimiento, momento en el que los padres deciden aceptar o no al recién nacido como su hijo y miembro de la sociedad de los humanos. El punto de vista de esta postura se apoya en la concepción relacional de la persona, en la medida en que se entiende como característica central de las personas su capacidad de re-lación y de entrar en comunicación con las demás.
99 Ibídem, p. 204. 100 Ibídem, p. 204101 Sampedro, J., Entrevista a Peter Singer, en El País (Babelia), 11 de mayo de 2002,
p. 12.

405
© Universidad de Deusto - ISBN 978-84-9830-383-4
Una de las que defienden este planteamiento es la teóloga feminista nor-teamericana Marjorie Reiley Maguire102. Parte de defender que la adjudica-ción de la personalidad no depende de los datos biológicos sino de la alian-za que se establece entre los padres y el nuevo ser a partir del nacimiento. «Yo propondría, nos dice, que la única persona capaz de iniciar una relación de amor, de alianza hacia la vida prenatal, haciendo que esta vida entre en la realidad de la comunidad humana y convirtiéndola así en una persona, es la mujer en cuyo seno se ha hecho realidad el embarazo». Y continúa: «Se crea una personalidad cuando la portadora de la vida, es decir, la madre, se-lla la alianza de vivir con la vida que dentro de sí misma se desarrolla has-ta darla a luz… El momento en que empieza a haber una personalidad, por consiguiente, es cuando la madre acepta el embarazo. El momento en que la madre establece un vínculo con el feto, éste se convierte para ella en un tú y deja de ser un ello… Es la madre la que hace del feto una persona. A partir de ese momento, esa vida es sagrada porque lo es para la madre»103.
M. Reiley Maguire coincide en esto con las costumbres de ciertas cul-turas, que en épocas pasadas dejaban en manos de los padres, más bien del paterfamilias, la decisión de aceptar al recién nacido, siendo desecha-do en el caso de que naciera deforme, o simplemente tuviera alguna carac-terística que no gustase a sus progenitores, como, en ocaciones, ser mujer, convirtiéndose también el infanticidio en un modo entre otros de control de natalidad y demográfico104.
Por lo tanto, para Maguire, «desechar algunos óvulos fecundados e in-cluso experimentar con ellos no son comportamientos necesariamente in-morales, puesto que no es propio de las personas flotar en una probeta»105.
Estos planteamientos no dejan de tener serias deficiencias teóricas. Es cierto que la interpersonalidad y la socialidad son rasgos fundamenta-les de la personalidad, y sólo somos personas en un contexto social. Y tal contexto social comienza en la familia, en la pareja paterna, cuya decisión de tener un hijo es básica e imprescindible para que cualquier ser humano pueda llegar a ser concebido, proseguir su proceso embriológico, llegar al nacimiento, y ser protegido y educado posteriormente. Pero para entender adecuadamente esa estructura relacional, hay que tener en cuenta varios aspectos. En primer lugar, si se ha de ser consecuente con la propia teo-
102 Cfr. Reiley Maguire, M., «Personhood, Covenant, and Abortion», en Jung, P./Shannon, T. (eds.), Abortion and Catholicism, Nueva York, Crossroad, 1988, 100-120. Para las ideas centrales de su postura, cfr. Junker-Kenny, M., o.c., 76-78.
103 Ibídem, 109-110. Cita tomada de Junker-Kenny, M., o.c., 76-77.104 Cfr. Harris, M./Ross, E. B., Muerte, sexo y fecundidad. La regulación demográfica
en las sociedades preindustriales y en desarrollo, Madrid, Alianza, 1991. 105 Reiley Maguire, M., o.c., 117.

406
© Universidad de Deusto - ISBN 978-84-9830-383-4
ría relacional, ha de entenderse de forma simétrica y transitiva, cosa que no ocurre en este supuesto, ya que en esa relación entre padres y posible futuro hijo los que intervienen y tienen capacidad de decisión son sólo los padres. Al otro sujeto de la relación no parece tenérsele nunca en cuenta. Es un mero elemento pasivo de las decisiones de los demás.
En el paradigma antropológico de la relación e interpersonalidad, se supone que ambos polos se tratan y se consideran por igual, como yo-tú106,siendo el ámbito del encuentro interpersonal y social la atmósfera en la que surgen y se conforman ambos interlocutores. Pero de ninguna mane-ra se considera que uno de ellos es quien otorga la categoría de persona al otro, ni los derechos a ella vinculados, sino que colabora en la confor-mación de la otra personalidad, al mismo tiempo que va construyendo la suya. En definitiva, resulta correcta la crítica de M. Junker-Kenny a esta postura, al tacharla y considerala más de «relación feudal» que de autén-tico paradigma antropológico relacional, en la medida en que «se trata de una potestad unilateral de elegir ejercida entre humanos»107.
En definitiva, en toda teoría relacional se parte del respeto a la auto-nomía del otro, que posee en sí mismo una dignidad previa y autónoma a la decisión de los otros interlocutores. Autonomía y alteridad son dos po-los del mismo juego de relaciones interhumanas, que no pueden obviarse en lo que tienen de relación, pero también de diferencia y distinción.
De ahí que tampoco tenga peso la conclusión que se quiere sacar de estos presupuestos para apuntalar el derecho al aborto en cualquier mo-mento del embarazo, basándose en el hecho de la total dependencia del nasciturus respecto a la madre. «Creo, afirma M. Reiley Maguire, que nadie tiene derecho a la vida ni que merezca la plena protección de la ley, cuando esa vida depende totalmente del cuerpo de otro ser humano como soporte vital»108.
Esta reflexión podría tener un cierto peso en los casos de violación, en la medida en que una mujer se ve forzada a gestar una vida que no ha de-seado. A veces se suele alargar este argumento para referirse a situaciones en que los progenitores se han visto en tal situación de futura paternidad por ignorancia o descuido109. Pero en una situación normal, el embarazo
106 Cfr. Buber, M., Yo y tú, Buenos Aires, Nueva Visión, 1969; Levinas, E., Totalidad e infinito, Salamanca, Sígueme, 1977; Id., El humanismo del otro hombre, Madrid, Si-glo XXI, 1974; Id., De otro modo que ser, o más allá de la esencia, Salamanca, Sígueme, 1987; Cavada, M., La vigencia del amor, Madrid, Edit. San Pablo, 1994.
107 O.c., 78.108 Reiley Maguire, M., o.c., 192.109 P. Singer analiza con detalle estas diversas situaciones y ejemplos, negando que
sean convincentes para defender el aborto, desde su óptica utilitarista. Cfr. Etica práctica,o.c., pp. 180-185.

407
© Universidad de Deusto - ISBN 978-84-9830-383-4
con todas sus consecuencias es un acto querido y programado por la pa-reja de progenitores. Además, si se lleva el argumento a su extremo, ten-dríamos que decir que podríamos defender la muerte de cualquier ser que dependiera de otro para existir, sea antes o después del nacimiento.
Tras las presentación de estas diversas posturas, parecen claras varias conclusiones. La primera ya la indicamos en la introducción: la respuesta al estatuto ontológico del embrión se sitúa a caballo entre la mirada cien-tífica y la filosófica. La ciencia aporta los datos, pero sólo desde la inter-pretación filosófica podemos dilucidar la condición de persona de la cé-lula germinal o del embrión. Pero la filosofía no puede dar su veredicto sin los datos de la ciencia, y es evidente que la ciencia todavía sabe poco. Prueba de ello es que resulta muy difícil establecer una respuesta contun-dente y convincente a la cuestión de si el proceso entre la célula germinal y el nacimiento es una evolución sin rupturas ni saltos cualitativos, o se dan más bien diferenciaciones cualitativas, hasta llegar un momento en el que el embrión o el feto poseen ya todas las notas esenciales para ha-blar de una persona en potencia. Aunque es muy posible que, tanto en este punto, como en la pregunta por el momento en el que se da el primer ser humano dentro del proceso de hominización (o, lo que es lo mismo, el momento en el que una especie se diferencia de otra contigua), la ciencia nunca nos dará una respuesta clara y contundente, puesto que son cues-tiones sin respuesta nítida, dada la gradualidad y continuidad existente en estos procesos, siendo como un gradiente que no admite distinciones radi-cales entre una etapa o fase y la siguiente.
A partir de ahí, cada teoría hace sus interpretaciones filosóficas y éti-cas que parten y se apoyan en sus respectivos presupuestos cosmovisio-nales, dentro de los cuales, como un ingrediente básico, se halla presente su correspondiente modelo antropológico y sus correspondientes conclu-siones éticas. Con todos estos ingredientes, esto es, los muchos vacíos científicos y el carácter postulatorio de los presupuestos cosmovisionales de cada postura, no es extraño que nos encontremos con un abanico tan amplio de posturas tan dispares y encontradas como las que hemos pre-sentado.
Eso no quiere decir que todas las posturas sean iguales, porque no todo vale ni es igual. Pero el diálogo entre posturas diferentes tiene que hacerse no desde actitudes intransigentes e impositivas, sino desde el respeto a las diferencias y desde el empeño de conseguir unos hábitos de comportamiento social que respeten la dignidad de las personas hu-manas, y acepten las diferencias entre ellas, desde el empeño porque to-dos los afectados se sientan protegidos y respetados. Y todo ello desde el convencimiento de que nadie tiene la verdad absoluta, y, aunque alguien crea poseer la postura mejor fundamentada y razonada, no la puede im-

408
© Universidad de Deusto - ISBN 978-84-9830-383-4
poner a la fuerza a los demás, ya que estamos en un ámbito histórico de permanente provisionalidad, convicción que no supone caer en el relati-vismo moral.
5.3. ¿Cómo se llega a ser persona?
Es otra de las cuestiones fundamentales que se suscitan a la hora de estudiar el proceso evolutivo de la aparición del ser humano. Se trata de un proceso largo y complejo, consecuencia de todo el lento desarrollo evolutivo del que hemos hecho referencia en páginas anteriores. Como en todas las demás cuestiones, no todos responden a este problema de la misma manera. Las diferentes posturas que se alinean ante este problema son las mismas que se dan ante el ya conocido problema de las relaciones alma-cuerpo, o mente cuerpo110. Vamos a presentar, de modo esquemáti-co, las tres posturas principales: la dualista, la monista reduccionista y la emergentista o estructurista.
a) La postura dualista: parte del hecho de que el ser humano está compuesto por dos sustancias ontológicas diferentes, el cuerpo y el alma, y mientras el cuerpo es el resultado del proceso evolutivo, el alma estaría insuflada directamente por Dios en un momento determinado del proceso evolutivo del cuerpo: bien sea desde la célula germinal o bien en un mo-mento posterior del desarrollo embriológico. Estos planteamientos dualis-tas tuvieron en Platón, y después en Descartes, a sus representantes más significativos. Pero también en tiempos más recientes han defendido el dualismo científicos de la talla de Scherrington, Sperry, y sobre todo John Eccles111. El problema de esta postura es cómo demostrar la existencia de una realidad ontológica diferente a lo corpóreo, y cómo entender su in-teracción con el cuerpo. Si quiere esta teoría situarse en el terreno de lo científico y no contradecir los resultados de la misma, el dualismo ontoló-gico resulta difícil de defender. Además, consideramos que las motivacio-
110 Sobre el problema de las relaciones mente-cuerpo, cfr. Priest, S., Teorías y filosofías de la mente, Madrid, Cátedra, 1994; Martínez Freire, P. F., La nueva filosofía de la mente,Barcelona, Gedisa, 1995; Broncano, F. (ed.), La mente humana, Madrid, Trotta, 1995; Dou, A. (ed.), Mente y cuerpo, Bilbao, Mensajero, 1986; Gardner, H., La nueva ciencia de la men-te, Barcelona, Paidós, 1987 (2.ª ed. 2000); Liz, M., Perspectivas actuales en filosofía de la mente, Tenerife, Gobierno de Canarias, 2001; Bunge, M., El problema mente-cerebro. Un enfoque psicobiológico, Madrid, Tecnos, 1985; Churchland, P. M., Materia y conciencia. Introducción contemporánea a la filosofía de la mente, Barcelona, Gedisa, 1999.
111 Cfr. Popper, K./Eccles, J., El yo y su cerebro, Barcelona, Labor, 1980; Eccles, J., Lapsique humana, Madrid, Tecnos, 1986; Eccles, J./Zeier, H., El cerebro y la mente, Barcelo-na, Herder, 1985.

409
© Universidad de Deusto - ISBN 978-84-9830-383-4
nes humanistas que empujan a ciertos teóricos a defender el dualismo112,frente a los reduccionismos materialistas de posturas como la teoría de la identidad, el eliminativismo y ciertos funcionalismos, no implican necesa-riamente acogerse a fórmulas dualistas, puesto que también se puede de-fender la centralidad de lo humano desde posturas no dualistas, como el emergentismo y el estructurismo.
b) El monismo reduccionista parte de la idea de que el ser humano se reduce a su cuerpo, entendido meramente desde el punto de vista biológi-co y genético. Por tanto, el proceso de la conformación de la especie hu-mana y de cada uno de sus componentes individuales no escapa al simple proceso de aparición de una especie nueva, como consecuencia de una acumulación de mutaciones y de los consecuentes procesos de especia-ción, bien sea por factores de aislamiento geográfico o genético. El error básico de los biologismos reduccionistas consiste en no advertir que to-das las realidades, y más aún las biológicas, son estructuras o sistemasen los que el todo no se reduce meramente a la suma de sus partes, por-que el conjunto o sistema posee cualidades o propiedades que no las te-nían sus partes antes de conformar el sistema. Eso es precisamente lo que está implícito en el proceso evolutivo, en la medida en que de la materia surge o emerge un nuevo tipo o sistema de realidad: la vida; y, dentro del continente de la biosfera, van apareciendo o emergiendo posteriormente nuevos y más complejos sistemas vitales, que dan lugar a linajes y espe-cies nuevas. Y como coronación o nivel más complejo de todo el proceso, emerge la vida inteligente y autoconsciente: la especie humana. Para que se dé un sistema o estructura nueva se necesita, como hemos ya indicado, que tenga al menos una cualidad o característica que no esté ya presente en alguna de sus partes. Es lo que ocurre, por ejemplo, con una molécula de agua, respecto a sus dos componentes, el hidrógeno y el oxígeno.
En cambio, para la mentalidad reduccionista, todas las realidades exis-tentes, tanto las realidades vivas como las inanimadas, son una mera suma de elementos que, en último caso, se pueden reducir a las partículas ele-mentales de la materia física. Por tanto, todo se reduce en último término a física. Pero, de ese modo, se pierde la posibilidad de reconocer lo que de novedad tiene el proceso evolutivo, con sus saltos cualitativos emergentes y las novedades reales que se dan dentro del mismo. Prueba de ello es que las leyes del mundo de la vida no pueden reducirse del todo al mundo de la materia inanimada, y la mirada epistemológica que exige la biolo-
112 Cfr. Popper, K./Eccles, J., o.c. En la Introducción, señalan los autores que lo que les ha motivado a escribir el libro es la tendencia generalizada que observan en los textos ac-tuales sobre filosofía de la mente, en los que predomina mayoritariamente la postura mate-rialista reduccionista, que reduce al ser humano a una mera máquina biológica.

410
© Universidad de Deusto - ISBN 978-84-9830-383-4
gía, y más aún la psicología, como estudio de la mente humana, no puede reducirse en último término a las leyes de la física113.
En definitiva, para el biologismo reduccionista, la especie humana no es más que una máquina biológica como las demás formas de vida; má-quina más compleja, pero nada más. Por tanto, la especie humana es una más de las muchas que pueblan la biosfera, y no tiene sentido que busque-mos elementos especiales que la distingan del resto de las especies vivas, en el momento de su aparición como tal. Esta postura, por tanto, reduce lo humano a ser una especie más de la biosfera, sin advertir lo que de es-pecífico y singular posee, fruto también del proceso evolutivo que, en su dar de sí, ha hecho emerger una especie más, vista desde el punto de vista de la composición bioquímica, aunque cualitativamente diferente desde el enfoque sistémico y ontológico.
c) La postura emergentista o estructurista parte de una concepción unitaria del ser humano, conjugando la continuidad de la especie huma-na con el resto de las demás especies vivas emergidas del proceso evo-lutivo, con su condición de especie singular, consecuencia de un salto emergentista específico114. En efecto, la especie humana, como ya hemos ido viendo, se halla inscrita en el proceso evolutivo como el resto de las especies vivas, manteniendo desde este punto de vista una radical y defi-nitiva continuidad con el resto de los demás seres vivos. Pero, al mismo tiempo, toda especie nueva es fruto de la acumulación de mutaciones que llevan a conformar una nueva estructuración biológica, expresada en un nuevo fenotipo morfológico, a la vez que en un nuevo modo de compor-tarse y de acomodarse al entorno. Por tanto, en cada proceso de confor-mación de una nueva especie, se conjugan la suma de sucesivas mejoras respecto de la especie de la que procede, junto con la emergencia de una nueva conformación o solución vital, que se expresa en la emergencia de una nueva estructura esencial, tanto en lo morfológico como en lo com-portamental.
La postura emergentista entiende que en este salto o proceso nove-doso se advierten siempre una serie de notas o características especiales: novedad, impredecibilidad e irreductibilidad. Es decir, la realidad emergi-da, aunque se apoya en el nivel previo (subtensión dinámica, lo denomina
113 Tendremos ocasión de incidir en estos puntos en el capítulo 7.º, dedicado a estudiar las relaciones entre la biología y la cultura.
114 Sobre las posturas emergentistas, cfr. Beorlegui, C., «Los emergentismos sisté-micos: un modelo fructífero para el problema mente-cuerpo», Pensamiento, 62(2006), n.º 234, pp. 391-438; Id., «Filosofía de la mente: visión panorámica y situación actual», Realidad, 2007, n.º 111, 121-160; Id., «Emergentism», Pensamiento, 65 (2009), n.º 246 (Serie especial: Ciencia, Filosofía y Religión, n.º 3), pp. 881-914.

411
© Universidad de Deusto - ISBN 978-84-9830-383-4
Zubiri), es un proceso nuevo, imprevisible desde el nivel anterior, e irre-ductible al mismo en su totalidad, porque, aunque las leyes nuevas que ri-gen el nuevo nivel emergido puedan, en parte, ser reducidas a algunas leyes del nivel anterior, nunca se conseguirá una reducción total115. Una tesis básica de las teorías emergentistas consistirá en defender que toda estructura o sistema emergido tiene que tener al menos una propiedad nueva, distinta a las propiedades de las partes del sistema116.
En el caso del ser humano, se dan estos mismos mecanismos, pero con sus propias peculiaridades. Zubiri, desde posturas cercanas al emer-gentismo (Laín Entralgo define su postura, basada en Zubiri, como es-tructurismo117) habla de que el proceso de conformación de la especie humana es el resultado de una elevación, consecuencia de un proceso de hiperformalización. Dentro del proceso evolutivo, en la aparición de las diversas especies vivas se da siempre un salto cualitativo de emergencia de una nueva estructuración de lo vivo, pero siempre dentro de un mis-mo nivel de complejidad: el ámbito del sentir. Pero, con la aparición de la especie humana, emerge un nuevo tipo de sustantividades cuyo sentir no es ya un mero sentir sino un sentir inteligente. Eso nos indica que el salto cualitativo que se ha producido ahí es de nuevo nivel, y por eso Zubiri lo denomina elevación118. La realidad ha dado de sí en ese momento un ni-vel funcionalmente superior a aquel del que inmediatamente procede. Si lo propio de toda realidad material es estar dando de sí, la elevación su-pone o expresa el paso de un nivel del dar de sí a otro de nivel superior (un sentir inteligente). En el caso de la génesis de la sustantividad huma-na, la elevación cobra un nuevo carácter, que consiste en el acto de bro-tar. Ahora bien, ese brotar de algo, la nueva estructura vital que da origen a una nueva especie, no es un mero surgir algo de algo, porque eso no nos llevaría más que a una psique en el cuerpo (solución dualista), y no a una psique de un cuerpo. Se trata, pues, de algo más radical, en la medida en que es un brotar-desde la estructura de la célula misma. Por tanto, la psi-
115 Cfr. Ayala, Fco. J., «Biology as an Autonomous Science», American Scientist, 1968, n.º 56, pp. 207-221. Edición posterior: «La biología como una ciencia autónoma», en Aya-la, F. J., La evolución de un evolucionista. Escritos seleccionados, Valencia, Universidad de Valencia, 2006, pp. 105-122; Beorlegui, C., «Los emergentismos sistémicos…», o.c., p. 427-428.
116 Cfr. Bunge, M., El problema mente-cuerpo, Madrid, Tecnos, 1985.117 Cfr. Laín Entralgo, P., «El problema alma-cuerpo en el pensamiento actual», en
Mora, Fco. (ed.), El problema cerebro-mente, Madrid, Alianza, 1995, pp. 209-231; Laín Entralgo, P., Qué es el hombre. Evolución y sentido de la vida, Oviedo, Ediciones Nobel, 1999.
118 Cfr. Zubiri, X., Sobre el hombre, Madrid, Alianza/Sociedad de Estudios y Publica-ciones, 1986, pp. 445 y ss.

412
© Universidad de Deusto - ISBN 978-84-9830-383-4
que brota desde la estructura misma de lo corpóreo (sea desde el momen-to de la célula germinal o en fases posteriores, cuando se da la suficienciaentitativa para la existencia de una nueva persona). Y este brotar algo de algo lo entiende Zubiri como el proceso en el que ese primer algo a que da origen el segundo, es algo constituido por el segundo. Por tanto, se nos dice que brotar es ante todo un hacer, que implica un constituir. De tal modo que la nueva estructura de las células de ese nuevo ser hacen o constituyen las notas psíquicas, de modo que la psique está producida por las células somáticas y por sus propias estructuras.
Precisando más, este proceso de constituir que desemboca en la ele-vación a la nueva especie humana, es consecuencia de un fenómeno que Zubiri denomina hiperformalización. Es decir, la estructura desde la que y por la cual brota la psique humana (como subestructura de notas psíquicas dentro del conjunto de la única estructura humana), es el cere-bro (y el cuerpo entero) de la especie anterior a la humana en el proceso evolutivo (australopiteco) que evolutivamente se ha ido complejificando, como consecuencia de acumulación de mutaciones y de un cerebro cada vez más complejo. Como consecuencia de ese enriquecimiento y comple-jificación cerebral, se empuja al proceso evolutivo a la necesidad de hacer brotar otro modelo cerebral capaz de hacer biológicamente viable los lo-gros así adquiridos. De tal manera que no le queda a la evolución más que dos salidas: verse incapaz de dominar desde el mismo tipo de estructura cerebral el aumento de complejidad adquirida, o en su permanente dar de sí hacer emerger una nueva estructura cerebral, la psique humana, que sea capaz desde esta hiperformalización de habérselas con la realidad con un nuevo tipo de habitud: la intelección sentiente119. Esto se aplica y se da tanto en el nivel filogenético como en el ontogenético. Con esta hiperfor-malización, aparece también un modo cualitativamente nuevo de dar de síde la realidad. Si hasta entonces las sustantividades se caracterizaban con conjunto de notas que le correspondían de suyo o en propio, estas nuevas sustantividades (humanos) son realidades que, además de ser de suyo, sona su vez realidades suyas (suidad), esto es, dadas a sí mismas, su ser con-siste en hacerse cargo de la realidad y de su propia realidad, en la medida en que se les ha encomendado la realización de sí mismas a sus decisiones libres y autónomas.
Por tanto, el concepto de psique (la idea tradicional de alma) se confi-gura como la estructura dinámica de la realidad humana, no una sustan-cia diferente a lo material que interacciona con el cuerpo. Esa estructura
119 Cfr. Zubiri, X., o.c., 455 y ss.; Laín Entralgo, P., Qué es el hombre. Evolución y sen-tido de la vida, Oviedo, Ediciones Nobel, 1999.

413
© Universidad de Deusto - ISBN 978-84-9830-383-4
específicamente nueva, la propia del embrión humano, está dotada de mo-dos también nuevos de dar de sí y de actuar ante la realidad: las propieda-des psíquicas propias del ser humano. Cada uno de los seres humanos son una única estructura conformada por dos subestructuras diferentes: la so-mática y la psíquica, poseyendo solamente la estructura total la suficien-cia entitativa para existir.
Ahora bien, esa elevación por hiperformalización, consecuencia del dar de sí de la realidad material a lo largo del proceso evolutivo, no pa-rece que la propia materia lo puede hacer por sí misma, en la medida en que una materia que sólo puede sentir no parece ser capaz de hacer emer-ger un sentir que sea también inteligente. Zubiri se pregunta qué es lo que hace que la materia dé de sí una realidad que no puede hacer brotar por sí misma. Y su respuesta es el cosmos. Pero, para que no se interprete esta respuesta como una salida milagrera, es fundamental entender qué entien-de Zubiri por cosmos. Como ya indicamos más arriba, para las posturas emergentistas y estructuristas, lo característico de un sistema o estructura es que necesita poseer al menos una propiedad específica del sistema o es-tructura, diferente y añadida a las que poseen las partes que la componen. Así, en todo sistema se da un doble nivel de realidad: el todo y las partes, en el que el todo tiene un plus de potencialidad sobre las partes. Si enten-demos que el cosmos, como conjunto de las realidades existentes, es tam-bién un sistema, el macro-sistema del conjunto del universo, tiene que po-seer las correspondientes potencialidades para que, a través de su ya larga trayectoria, y todavía en proceso de realización, capacita a sus diferentes partes de las potencialidades que en cada momento van emergiendo de su interminable dar de sí (es lo que denomina Zubiri el poder de lo real). Así que la emergencia de la vida en este pequeño y suburbial planeta Tierra, y posteriormente la psique humana, es obra de la propia capacidad de dar de sí de la materia, que se hace vida, y luego vida inteligente.
El problema está en mostrar si el propio cosmos, de por sí puede ha-cer emerger una realidad sustantiva como la realidad humana, dotada de la capacidad de inteligir. De ahí que una cuestión complementaria será la que se suscita sobre el fundamento y origen del cosmos. De ese modo en-tramos en el ámbito de la metafísica y de la teología.
En conclusión, para Zubiri la evolución y la elevación son dos poten-cialidades del cosmos mismo. No quiere decirse con esto que con su idea de elevación se sitúe a la especie humana fuera o al margen del proceso evolutivo, sino que la fase evolutiva de la aparición de la especie huma-na (elevación) tiene sus propias y peculiares características. De ahí que la especie humana hay que verla como una fase más del proceso evolutivo (continuidad), pero a la vez es fundamental advertir sus cualitativas dife-rencias (singularidad), como consecuencia de sus específico modo de ser

414
© Universidad de Deusto - ISBN 978-84-9830-383-4
y de habérselas con la realidad (lo que denomina Zubiri habitud: intelec-ción sentiente). Pero, como ya lo hemos dicho, no supone esta concepción singular de la emergencia de lo humano defender la tesis dualista, esto es, no se trata de afirmar que se dan dos génesis, una material y otra psíquica, sino una sola génesis de naturaleza psico-orgánica, desde la concepción misma de un viviente ya psico-somático. Esta génesis tiene su comienzo, según Zubiri, desde la célula germinal (para muchos de sus discípulos, en cambio, el momento de la elevación se daría en momentos posteriores, como defiende la segunda postura indicada en páginas anteriores), conti-nuada a través del proceso embrionario, e incluso tras varios años después del nacimiento. De este modo, la psique, que el embrión ha constituido, constituye a su vez al cuerpo (en la medida en que no está terminado). Así, podemos ver que, en la elevación, lo material se hace más que ma-terial, es psíquico. Pero supone más que la mera toma de conciencia de sí mismo, puesto que la mera reflexibilidad es consecuencia de algo más ra-dical: la nueva estructura de la realidad humana. Sólo por ello hay psique, y, como consecuencia, reflexión, autoconciencia120.
En definitiva, para esta forma de ver las cosas, ser hombre no consis-te en dejar de ser materia, ni en que simplemente ésta sirva a la psique, sino en corporeizar la psique, o también, de modo complementario, en psiquizar el cuerpo, de tal modo que el ser humano se constituye así en unidad psico-somática. Y en esto consiste la hominización y la humaniza-ción, que para Zubiri viene a ser una estricta potenciación de la materia.
120 Este modo de ver las cosas coincide en gran medida, en el enfoque teológico, con la postura defendida por K. Rahner y P. Overhage, en su libro El problema de la homini-zación. Sobre el origen biológico del hombre, Madrid, Cristiandad, 1973. Cfr. también G. Canobbio, Sobre el alma. Más allá de mente y cerebro, Salamanca, Sígueme, 2010, pp. 28-34.

415
© Universidad de Deusto - ISBN 978-84-9830-383-4
Capítulo 7
La singularidad de los humanos: entre el antropocentrismo y el reduccionismo biológico
1. EL ANTROPOCENTRISMO EN CUESTIÓN1
Vimos en el primer capítulo que la propuesta darwiniana de situar al linaje humano en el entorno de la biosfera, como un linaje más, ha ido minando la concepción tradicional antropocéntrica, deudora de una con-cepción del ser humano salido directamente de la acción creadora de Dios y situado en el centro de la tierra y del cosmos. La deriva de los estudios antropológicos posteriores parecería que han ahondado mucho más en esta tendencia trans-antropocéntrica, pretendiendo reducir la especificidad de lo humano no sólo en el ámbito de su origen genético y biológico, sino incluso pretendiendo que su especificidad cultural también fuera pues-ta en entredicho. Es decir, si se impone por evidencia científica el origen biológico de la especie humana por evolución de las especies anteriores, resulta más problemática e inaceptable la pretensión de ciertas teorías an-tropológicas de reducir también su estructura comportamental a un mero reflejo de las directrices genéticas y biológicas presentes en su condición biológica.
En los capítulos anteriores nos hemos detenido en analizar la teoría de la selección de Darwin, los descubrimientos de la genética desde Mendel
1 Este capítulo ha sido publicado, con breves retoques, bajo el título «La singularidad del ser humano como animal bio-cultural», Letras de Deusto, 39 (2009), n.º 125, 59-93.

416
© Universidad de Deusto - ISBN 978-84-9830-383-4
hasta la doble hélice del ADN, para conformar una teoría sintética de la evolución, que está en la actualidad lejos todavía de merecer un consenso absoluto. También hemos visto cómo la paleoantropología y la biología molecular van mostrándonos los secretos de la historia del proceso de ho-minización y los esfuerzos que las diversas especies humanas han realiza-do en su empeño por humanizarse. Y hemos ido viendo en qué medida en cada uno de los rasgos que parecen específicos de la condición humana resultaba difícil marcar una frontera clara y precisa entre los humanos y los demás animales.
De fondo nos quedaba siempre la cuestión de si, entre humanos y ani-males, se da una diferencia claramente cualitativa y de grado, o meramen-te cuantitativa. La respuesta que hemos ido dando se orientaba a conjugar la continuidad y la ruptura entre los humanos y las demás especies vivas. Es el momento de detenernos para encarar directamente esta cuestión. ¿Podemos defender con argumentos precisos la singularidad del ser hu-mano, o tenemos que plegarnos a las afirmaciones de quienes defienden la imagen de lo humano como un animal más, aunque más complejo y elaborado?
Los seres humanos somos la única especie animal autoconsciente de sí misma y, por ello, capaz de preguntarse y cuestionar su propia reali-dad. Y lo ha hecho siempre, de una manera más o menos consciente, más o menos profunda, desde el primer momento de su existencia como espe-cie2. Desde este punto de vista, podríamos definirnos como un animal in-quiriente o animal preguntón3. Este rasgo no es una característica ocio-sa, accidental, de la que el ser humano pueda prescindir, sino algo que corresponde a su propia esencia y condición: necesita saber de sí y tener una idea de sí mismo para orientar su propia realización, en la medida en que, por poseer una naturaleza abierta, su realización depende de sus pro-pias decisiones, entre ellas acertar en el empeño de alcanzar con éxito la meta ideal que sobre sí se ha construido. Se da, por tanto, en la estructura esencial del ser humano una estrecha relación entre autoconocimiento y autorrealización.
Basta echar una mirada a la historia humana para percatarse de la in-terminable secuencia de modelos antropológicos que se han ido propo-niendo y sucediéndose regularmente como consecuencia de los cambios de mentalidad producidos por todo tipo de crisis culturales o cosmovisio-nales, deudores todos ellos de la inevitable evolución de los saberes cien-
2 Cfr. Landmann, M., Antropología filosófica. Autointerpretación del hombre en la his-toria y en el presente, México, UTEHA, 1961.
3 Cfr. Beorlegui, C., Antropología filosófica. Nosotros: urdimbre solidaria y responsa-ble, o.c., pp. 35-37.

417
© Universidad de Deusto - ISBN 978-84-9830-383-4
tíficos acerca de la realidad y del propio ser humano. Si estos avatares se han producido siempre de forma intermitente4, apareciendo en esa histo-ria la radical problematicidad del ser humano, es en la actualidad cuan-do parece haberse percatado de un modo más profundo de su radical e inevitable problematicidad5. Efectivamente, el hombre actual se halla en un momento de seria transformación y de crisis de su propia identidad, de reacomodo de muchos de los rasgos que definían su ser hace algo más de un siglo y medio. Con la aparición del paradigma evolutivo, y más en concreto con las propuestas de Ch. Darwin sobre la selección natural, el modelo antropológico que había predominado en Occidente comenzó a tambalearse, pasando de entenderse como alguien creado directamente por las manos de Dios, a experimentarse como una especie más del ámbi-to de la biosfera, aparecida al igual que las otras como fruto de un similar proceso evolutivo, en el que se conjugan las leyes de la genética con la selección ambiental, según nos indica la teoría sintética de la evolución6.Parecería, pues, que estas aportaciones científicas exigirían defender sin paliativos las tesis del naturalismo filosófico o materialismo fisicalista y reduccionista sobre el ser humano y sobre el conjunto de la realidad.
Posteriores e importantes descubrimientos en diferentes esferas cientí-ficas han seguido minando la cosmovisión religiosa antropocéntrica en la que estábamos aposentados, confirmándonos más fuertemente en este ra-dical cambio de perspectiva y abocándonos a replantearnos la pregunta por nuestra identidad y nuestro puesto en el conjunto del universo. Nos halla-mos en la actualidad en una auténtica edad de oro de disciplinas científicas como la paleoantropología, la genética, la biología molecular, la embrio-logía, la etología y otras muchas, que parecen apuntar hacia una visión del ser humano entroncado en la realidad material y biológica de nuestro mun-do, menos autosuficiente y reducido a entenderse como una especie más del conjunto de la biosfera, producto de leyes evolutivas que parecen regir-se, como dijo J. Monod, por la conjugación del azar y la necesidad7.
Muchas de las aportaciones de las disciplinas científicas señaladas parecen apoyar incluso, según algunos, no sólo la superación de la con-cepción teocéntrica del ser humano, sino incluso una visión humanista antropocéntrica. Según este punto de vista materialista y reduccionista, la
4 Cfr. Buber, M., ¿Qué es el hombre?, México, FCE, 1976 (9.ª ed.). 5 Cfr. García Bacca, J. D., Antropología filosófica contemporánea, Caracas, UCV,
1947, cap. 2.º, «El hombre como tema y como problema» (nueva edición: Barcelona, Anthro-pos, 1982.
6 Cfr. Dobzhanskky, Th., Genetics and the Origin of Species, Columbia University Press, 1937; E. Mayr y W. Provine, The Evolutionary Synthesis, Harvard University Press, 1980; G. Simpson, The major features of evolution, Columbia University Press, 1953.
7 Cfr. Monod, J., El azar y la necesidad, Barcelona, Barral, 1977.

418
© Universidad de Deusto - ISBN 978-84-9830-383-4
ciencia nos abocaría a no tener más remedio que defender una visión re-ductiva y anti-humanista del ser humano, es decir, partidaria de entender a la especie humana como una más del largo y complejo proceso evolutivo, todo lo compleja y maravillosa que se quiera, pero nada más.
Por otro lado, y en paralelo a esta visión reduccionista, parecen cobrar cada vez más fuerza los planteamientos de la liberación animal y de los derechos de los animales, que entienden que, aunque la diferencia entre la especie humana y la mayoría de los animales es cualitativa, de grado, no lo es sin embargo respecto a las especies de los denominados grandes si-mios (chimpancés, bonobos, gorilas y orangutanes), por lo que consideran que hay que ampliar el concepto humanidad y la categoría de persona a los componentes de estas especies, y, en consecuencia, aceptar y cumplir, por parte de los seres humanos, una larga lista de derechos humanos a fa-vor de los grandes simios8.
La cuestión que aquí vamos a plantear es si las propuestas humanistas y religiosas acerca del ser humano están ya superadas y obsoletas, por in-correctas y acientíficas, o si, por el contrario, siguen teniendo vigencia y base científica, lo que nos indicaría, por el contrario, que los planteamien-tos reduccionistas a los que hemos hecho referencia, no son tan evidentes ni justificados como los autores que defienden esos planteamientos nos quieren hacer ver. Necesitamos, por tanto, volver a reflexionar y mostrar aquello que constituye lo específico del ser humano, desde el análisis de las novedosas aportaciones de los diversos saberes científicos, para ver en qué medida nos sirven para asentar una correcta visión del ser humano como realidad específica y cualitativamente diferente al resto de las espe-cies animales, según las tesis de la postura humanista.
No cabe duda de que nos hallamos ante un reto decisivo para la com-prensión humanista, y hasta religiosa, del ser humano, y necesitados de una nueva redefinición de nuestro ser y de nuestro puesto en el cosmos(M. Scheler). La estrategia que tenemos que seguir es presentar una bre-ve síntesis de las diferentes aportaciones científicas que nos permitan rea-lizar un ejercicio comparativo entre el ser humano y el resto de los ani-males, para ver en qué medida tenemos apoyos científicos y filosóficos suficientes para seguir defendiendo el especial puesto del hombre en el
8 Cfr. Singer, P., Liberación animal, Madrid, Trotta, 1999; Id., Una izquierda darwi-niana. Política, evolución y cooperación, Barcelona, Crítica, 2000; Cavalieri, P./Singer, P. (eds.), El proyecto «Gran Simio». La igualdad más allá de la humanidad, Madrid, Trotta, 1998; Beorlegui, C., «¿Hacia un humanismo trans-antropocéntrico? Peter Singer y los de-rechos de los animales», Realidad, 2001, n.º 80, 199-236; Id., «Ética animal e idea de per-sona», en Guibert Uncín, J. M./Orella Unzué, J. L. (eds.), Preguntas éticas en cuestiones disputadas hoy, San Sebastián, Universidad de Deusto, 2005, 317-335.

419
© Universidad de Deusto - ISBN 978-84-9830-383-4
conjunto de la biosfera y de todo el universo, o tenemos que adscribirnos a la postura defendida por el naturalismo filosófico y el materialismo re-duccionista. En este ejercicio comparativo, pretendemos llegar a examinar los ingredientes fundamentales de la compleja y específica conformación biocultural del ser humano, para tratar de concluir que la especie humana es la única que está constituida por una específica conjunción de biología y cultura, conformando ambos componentes una estructura compleja pero unitaria. Esta unidad bio-cultural supone e implica a su vez una específica unidad psico-orgánica, a caballo entre posturas extremas como los dualis-mos interaccionistas y los materialismos reduccionistas, defendidos por el conductismo, la teoría de la identidad y determinados funcionalismos9.En definitiva, pretendemos llegar a la conclusión de que el ser humano se halla tanto en continuidad como en distancia cualitativa con el resto de las especies de la biosfera, pues, aunque es una especie más, sus característi-cas específicas lo sitúan en un nivel de diferencia cualitativa respecto a las especies de las que ha emergido. De este modo, pretendo llegar a concluir que el ser humano, a diferencia del resto de las especies vivas, constituye una irrepetible y compleja unidad bio-cultural y psico-orgánica, dotado de autoconciencia, autonomía, lenguaje, pensamiento complejo, libertad, capacidad ética, apertura a la pregunta por el sentido de su vida y del con-junto del cosmos, y, por eso mismo, en apertura a la pregunta por el fun-damento de la realidad, es decir, al ámbito del Absoluto.
2. LA COMPARACIÓN HOMBRE-ANIMAL: DOS ENFOQUES COMPLEMENTARIOS (CIENTÍFICO Y FILOSÓFICO) Y CUATRO ÁMBITOS DE COMPARACIÓN
2.1. Ciencia y filosofía: dos modos complementarios de acercarse a la realidad y al ser humano
Antes de introducirnos en el ejercicio comparativo de los diversos ele-mentos que diferencian a los seres humanos del resto de los demás anima-les, tenemos que ser conscientes de que hemos de conjugar correctamente dos modos de comprensión y de acercamiento a la realidad: el científico y el filosófico. Cuando nos planteamos la comparación hombre-animal, la discusión se suele reducir con frecuencia a la tarea de presentar los más
9 Cfr. Beorlegui, C., «Filosofía de la mente: visión panorámica y situación actual», Realidad, 2007, n.º 111, 121-160; Id., «Los emergentismos sitémicos: Un modelo fructífe-ro para el problema mente-cuerpo», Pensamiento, 62 (2006), 391-439.

420
© Universidad de Deusto - ISBN 978-84-9830-383-4
diversos datos e investigaciones que continuamente están aportando el amplio abanico de las ciencias, tanto naturales como humanas. Pero el re-sultado de tales disputas no suele traer demasiada claridad al problema, porque no es suficiente apelar a los datos científicos para dilucidar esta compleja cuestión, necesitada ante todo de un adecuado y correcto plan-teamiento. De entrada hay que decir que los datos científicos, en sí mis-mos, no significan nada para elucidar la distinción entre hombres y ani-males. Son mera constatación de hechos, que, por otro lado, nunca van solos, sino acompañados de los inevitables y necesarios aprioris y pre-juicios (presupuestos filosóficos) desde los que sistematizamos los datos científicos para conformar una teoría científica unificadora.
La realidad, y el ser humano dentro de ella, tiene dos dimensiones claramente diferenciadas: la dimensión fáctica y la del sentido o signifi-cado de la misma. Es decir, siguiendo la diferenciación de M. Foucault, no es suficiente para dar cuenta de la realidad humana detenernos en investigar su funcionamiento, en todos sus aspectos (biológico-genético, fisiológico, psicológico, conductual, etc.), sino que se necesita además preguntarnos y tratar de dilucidar su sentido, su dimensión trascendental. Al primer nivel se dedican las ciencias, las antropologías científicas, y al segundo, la filosofía, o mejor, las filosofías, con sus diferentes modelos antropológicos10. La negación o el olvido de esta distinción suele tener consecuencias nefastas, en la medida en que supone en ambos casos apo-yarse, sin tener suficiente consciencia de ello, en una filosofía implícita, el naturalismo filosófico o materialismo reduccionista. Con lo cual, además de presentarse las afirmaciones filosóficas como si fueran científicas, que no lo son, constituyen y representan una mala filosofía. Es decir, no es lo mismo afirmar que el ser humano está configurado, desde los diferentes ámbitos científicos que queramos, por los rasgos y las características que sean (tarea específica de las ciencias), que afirmar que su ser y su sentido se limita a esos simples datos. Esa afirmación, además de no ser científica (no se puede demostrar ni falsar), es equivocada, por insuficiente.
Un ejemplo reciente de este modo tan reduccionista de ver las cosas es el libro de aforismos del profesor español J. Wagensberg, con este título tan significativo: A más cómo, menos por qué11. Lo que parece indicarnos es que cuanto más consigan las ciencias descubrirnos cómo funcionan los diferentes ámbitos de la realidad (ámbito del cómo), menos espacio que-dará para las preguntas de por qué, de sentido. Por el contrario, habría que
10 Cfr. Morey, M., El hombre como argumento, Barcelona, Anthropos, 1987; Lorite Mena, J., La filosofía del hombre, o el ser inacabado, Estella, Verbo Divino, 1992; Beorlegui, C., Antropología filosófica. Nosotros: urdimbre solidaria y responsable, o.c., caps. 2.º y 4.º.
11 Barcelona, Tusquets, 2006.

421
© Universidad de Deusto - ISBN 978-84-9830-383-4
responder más bien, que cuantas más respuestas hallemos a los «cómos», más espacio y ocasiones se abrirán a los «porqués»; o dicho de otra forma: ni mil respuestas a los cómos podrán acallar un porqué, puesto que son dos niveles diferentes, paralelos e inevitables. Como dice significativamente X. Zubiri, la realidad nos da que pensar, y nos empuja a la pregunta por el fundamento, por las cuestiones últimas12. Muchas de las importantes dis-crepancias a cerca del ser humano y de la realidad son consecuencia de la no correcta distinción de planos entre la ciencia y la filosofía, incurriendo en sucesivos reduccionismos: metodológico, epistemológico, ontológico y antropológico, absolutizando una sola dimensión (la científica) y quedán-dose con una parte de la realidad como representativa del todo.
Volveremos más adelante sobre este asunto, puesto que nos va a ir apareciendo en cada uno de los diversos ámbitos de comparación entre hombres y animales que vamos a analizar a continuación.
2.2. Cuatro ámbitos de comparación
Se trata de preguntarnos si la diferencia entre hombre y el resto de los animales es sólo cuantitativa o posee un rango cualitativo. Esta compara-ción podría hacerse en múltiples aspectos, pero se suele considerar que los más importantes y significativos son los cuatro siguientes13:
— El bioquímico y genético; — El anatómico-morfológico; — El embriológico, desde el especial puesto del hombre en el proceso
embriológico u ontogenético; — El referente a la estructura comportamental.
2.2.1. El ámbito bioquímico y genético14
Las investigaciones recientes sobre el Genoma de las diferentes espe-cies vivas van mostrándonos las diferencias y parecidos entre la estructura genética del ser humano y del resto de las especies animales.
12 Cfr. Zubiri, X., Inteligencia y Razón, Madrid, Alianza/Sociedad de Estudios y Publi-caciones, 1983.
13 Cfr. Haeffner, G., Antropología filosófica, Barcelona, Herder, 1986, 20-26. 14 Cfr. Riedley, Matt, Genoma. Autobiografía de una especie en 23 capítulos, Madrid,
Taurus, 2000; Dulbecco, R., Los genes y nuestro futuro, Madrid, Alianza, 1999; Sulston, J. y Ferry, G., El hilo común de la humanidad. Una historia sobre la ciencia, la política, la ética y el genoma humano, Madrid, Siglo XXI, 2003; Núñez de Castro, I., «Análisis antro-pológico del Proyecto Genoma Humano» (manuscrito sin publicar).

422
© Universidad de Deusto - ISBN 978-84-9830-383-4
Si comparamos nuestra composición bioquímica y la del resto de las especies, advertimos que no nos diferenciamos en nada respecto de los de-más seres vivos. Estamos hechos de la misma pasta bioquímica. No po-seemos ninguna sustancia química específica, ni carecemos de ninguna de las que se dan en las otras especies.
Además, la sustancia bioquímica está organizada de similar forma que en el resto de los seres vivos. Nuestra estructura genética se halla codifi-cada en el ADN hereditario, que se organiza y reproduce de la forma co-mún a todas las especies vivas, siguiendo lo que se ha solido denominar (aunque hoy día se está poniendo en cuestión) la ley fundamental de la genética: ADN-ARN-Proteínas. La relación entre estos tres componentes se orienta siempre en esta dirección: de izquierda a derecha, no dándose en la relación de esta estructura genética la denominada lógica lamarckia-na de transmisión de los caracteres adquiridos. Esto es lo que hace que los científicos defiendan la unidad evolutiva de la vida. La vida apareció sólo una vez, y de ese salto cualitativo descendemos todos los seres vivos.
Nuestro ADN está configurado, al igual que en las demás especies, por una estructura de doble hélice (descubierta por Watson y Crick)15,unida por cuatro bases nitrogenadas (Adenina-Guanina, Citosina-Timina,esta última transformada en Uracilo cuando se traduce el ADN en ARN). No es necesario que nos detengamos aquí en explicitar las concreciones de los diferentes procesos que se dan en la reproducción genética de los seres vivos16. Pero sí es importante que veamos algunos aspectos diferen-ciadores en este aspecto entre nuestra especie y las más cercanas a noso-tros. En la conformación de las proteínas, siguiendo esa ley fundamental de la genética, se van construyendo cadenas de aminoácidos, advirtién-dose que en la composición, por ejemplo, de las cadenas alfa y beta que forman la hemoglobina A, la diferencia entre el hombre y el chimpancé en la composición de los 141 aminoácidos de la cadena alfa y en los 146 de la cadena beta es cero, no hay ninguna diferencia. En cambio, entre el hombre y el gorila, hay un aminoácido de diferencia en cada una de las cadenas, y de modo progresivo se van agrandando las diferencias en rela-ción a especies que se hallan más lejanas a nosotros en el proceso evoluti-vo17. Y también podríamos hacer referencia a otras pequeñas diferencias
15 Cfr. Watson, J. D., The double helix (La doble hélice); Id., Genes, chicas y laborato-rios, Barcelona, Tusquets, 2006.
16 Cfr. bibliografía: Ayala, F. J., Origen y evolución del hombre, Madrid, Alianza, 1980, cap. VI; Ruse, M., Filosofía de la biología, Alianza, 237 y ss.; Bertranpetir, J,/Junyent, C., Viaje a nuestros orígenes. Una historia biológica de la especie humana, Barcelona, Península, 2000; Jordan, B., Los impostores de la genética, Barcelona, Península, 2000; Fox Kéller, E., El siglo del gen. Cien años de pensamiento genético, Barcelona, Península, 2002.
17 Cfr. Ayala, F. J., o.c., pp. 158-161.

423
© Universidad de Deusto - ISBN 978-84-9830-383-4
en relación a nuestras enzimas y proteínas, para llegar a la misma conclu-sión: las diferencias son mínimas, aumentándose a medida que las espe-cies se hallan más alejadas en el árbol de la evolución.
Una diferencia clave está en el componente cromosómico de nuestra herencia genética. Mientras la especie humana está dotada de 23 pares de cromosomas, los grandes simios poseen 24 pares. Parece una pequeña diferencia, y todavía se diluye más si tenemos en cuenta que «los cromo-somas de ambos grupos son muy semejantes entre sí, y no sólo uno a uno sino banda a banda, excepto en lo que respecta a algunas mutaciones cro-mosómicas, como la fusión de dos de los cromosomas de los antropoides en uno del hombre»18.
Las investigaciones sobre el genoma humano han servido para pro-fundizar en nuestros parecidos con el resto de las especies vivas. En el año 2001 se hizo público el primer borrador del genoma humano, comple-tándose en 2003 su secuenciación, con una fiabilidad casi total. Dos años más tarde, en 2005, se publicaba también el primer borrador del genoma del chimpancé. Ya en 1995 se había secuenciado el Micoplasma genita-lium (un micoplasma es el organismo más pequeño que puede tener un organismo independiente)19, que posee sólo 470 genes y 580 kpb20. En años posteriores se han ido secuenciando los genomas de otras muchas es-pecies vivas, advirtiéndose una correlación entre el número de genes y de miles de pares de bases y la complejidad de cada especie viva.
Estas investigaciones nos han mostrado que la similitud genómica en-tre el homo sapiens y el chimpancé21es muy grande, llegando al 99% (si nos referimos a la «secuencia», esto es, el orden de los alrededor de 3.200 millones de pares de bases o «letras químicas» que parece componen el conjunto de la estructura del ADN22), siendo esto indicativo del relativa-mente corto espacio de tiempo en que ambas especies se habrían desgaja-do de la especie común anterior: se calcula entre 5 y 7 millones de años.
18 Ayala, F. J., o.c., 160-161. Cfr. Ruffié, J., De la Biologie á la Culture, Paris, Flamma-rion, 1976 (De la biología a la cultura, Barcelona, Muchnik Editores, 1982, 278 y ss.).
19 Cfr. Núñez de Castro, I., o.c., p. 3. 20 Estas letras significan: pb=pares de bases, refiriéndose a los emparejamientos de
las cuatro bases nitrogenadas ya sabidas, mientras que k equivale a la cantidad de pares de bases existentes en el conjunto del genoma, cantidad similar a 10 al cubo. Cuando las cantidades son mayores, se utiliza, en vez de k, el símbolo M, equivalente a 10 a la sexta. Cfr. I. Núñez de Castro, o.c., p. 3.
21 Sobre los datos comparativos entre el hombre y el chimpancé, cfr. The chimpanzee sequencing and análisis Consortium, «Initial séquense of the chimpanzee genome and comparison with the human genome», Nature 437, 2005, pp. 69-87.
22 Cfr. Watson, James D., «The Human Genome Project: past, present and future», Science, 248. 1990, 44-48; Núñez de Castro, I., o.c., pp. 1-12.

424
© Universidad de Deusto - ISBN 978-84-9830-383-4
El Homo sapiens posee 3.200 Mpb y cerca de 25.000 genes, resultando sorpresiva esta cifra tan baja. Se especulaba anteriormente con que pose-yera unos 100.000 genes, y más adelante se rebajó la cifra a 30.000, pero las investigaciones más recientes han dejado la cifra en esos 25.000 (y al-gunos la rebajan incluso a 23.000).
Estos avances tan espectaculares están produciendo no sólo un in-cremento de nuestros conocimientos, sino también nos están obligando a corregir algunos de los denominados dogmas genéticos, entre los que se encuentran, por ejemplo, los conceptos de genoma y de gen. Genomasignifica en la actualidad tanto «el mapa físico de la localización de los genes en los cromosomas», como «la secuencia completa de pares de ba-ses del DNA»; es decir, puede ser entendido como «material genético» o como «información genética y conjunto de genes» característico de una especie determinada23. En relación con el concepto de gen, acuñado por W. Johannsen en 1909 para denominar a los factores hereditarios investi-gados por G. Mendel, en la actualidad tiene una amplia pluralidad de sig-nificados, de modo que «la palabra gen no significa lo mismo para un ge-netista clásico, que para un genetista de poblaciones, que para un genetistadel desarrollo, que para un biólogo molecular. Incluso para un genetista molecular se presentan diferentes definiciones de gen»24. Pero, en defini-tiva, se puede reducir esa pluralidad semántica del gen a dos aspectos bien concretos: «el gen como última unidad de mutación y recombinación» y «el gen como unidad de información»25. Esta segunda acepción del gen es el fundamento en el que se apoya el denominado dogma central de la genética, en la medida en que se daba por hecho que un gen (como unidad del ADN que se convertía en ARN) daba lugar a una proteína, que a su vez realizaba una función (una enzima).
Pero este dogma se ha puesto en cuestión en la actualidad, en la medi-da en que se advierte que se dan fenómenos como la poligenia (varios ge-nes distintos pueden influir en la expresión de un mismo rasgo o compor-tamiento) y la pleiotropía (un solo gen puede influir sobre el desarrollo de varios rasgos físicos o comportamientos)26. Por tanto, aunque por un lado la genética sabe cada vez más acerca de la estructura genómica, ese ma-yor conocimiento le ha mostrado su inesperada complejidad, en la medida
23 Cfr. ibídem, o.c., p. 5.24 Núñez de Castro, I., o.c., p. 6. Cfr. Id., «La indefinición fundamental de las ciencias
biológicas», en Dou, A. (ed.), Fragmentariedad de las ciencias, Bilbao, Mensajero/Biblio-teca Fomento Social, 1985, 143-147; Fontanille, J., «Dieciocho definiciones sobre el gen», La Recherche (Elementos?), 2003, vol. 10, n.º 50, p. 17.
25 Núñez de Castro, I., o.c., pp. 6-7.26 Cfr. Meier, R., Comportamiento animal. Un enfoque evolutivo y ecológico, Madrid,
McGrawHill, 2001. p. 35; Núñez de Castro, I., o.c., p. 9.

425
© Universidad de Deusto - ISBN 978-84-9830-383-4
en que no hay una relación directa entre unidad genómica y unidad feno-típica, y además no todos los genes tienen expresión funcional ni sirven como información para la configuración fenotípica del organismo vivo. Dentro del DNA, se dan segmentos de gen que poseen información y aca-ban representados en una proteína (son los exones), y otros segmentos que aparentemente no tienen utilidad de cara a la conformación de proteínas (intrones), interrumpiendo la secuencia codificadora. Hay, por tanto, una parte del genoma, que se ha denominado por ello genes basura, o DNA intragénico (viene a representar nada menos que el 62,5% del contenido total del genoma), que «no conocemos por ahora cuál sea la función fi-siológica de esta gran cantidad de DNA, cerca de dos tercios a los que no se les ve, por ahora, que contengan ninguna misión en el genoma o en el desarrollo del organismo. Puede ser que esta porción notable del genoma tenga una función tan sutil que al presente se nos escape»27.
Por tanto, frente a la creencia ingenua de la genética clásica, que con-sideraba que en el conjunto del DNA estaba ya encerrada toda la infor-mación que se necesitaba para conformar el fenotipo morfológico y con-ductual del ser vivo, en la actualidad se advierte que esa traducción del genoma al fenotipo es mucho más complicada y abierta de lo que se creía. Y no sólo por los fenómenos ya indicados de la poligenia y la pleiotropía,lo que supone que un solo gen puede originar varias proteínas, sino tam-bién porque cada proteína puede llevar a cabo funciones diferentes. Por eso, en la actualidad va teniendo cada vez más importancia, junto al ge-noma, el concepto de proteoma, «conjunto de proteínas que especifican la naturaleza de todas las reacciones bioquímicas que una célula puede lle-var a cabo en un momento determinado. Así como el genoma es el mismo para todas las células diploides, el proteoma puede variar según el mo-mento de la vida celular»28.
Como puede verse, la complejidad del camino a seguir de un rasgo genómico, a través de la traducción en el abanico proteínico, hasta que llega a expresarse en los diferentes rasgos del fenotipo, es un camino lar-go y complejo que no tiene una única ruta, sino varias posibles, en fun-ción de factores aleatorios que se dan en los diversos recovecos de ese in-trincado camino.
Las conclusiones que de todo lo dicho podemos sacar son varias:
1) La ciencia todavía sabe poco y anda a tientas en su incesante traba-jo de exploración de nuestra estructura genética y de los procesos a través de los cuales se expresa. El trayecto que va del genoma al
27 Núñez de Castro, I., o.c., p. 10. 28 Ibídem, p. 12.

426
© Universidad de Deusto - ISBN 978-84-9830-383-4
fenotipo, campo de estudio de la biología molecular y de la biolo-gía del desarrollo, está lleno de interrogantes y de vacíos sin acla-rar. Por eso, como hemos visto, cada vez está más clara la impor-tancia de ciencias como la biología del desarrollo, o genética del desarrollo, y la embriología, que estudian los fenómenos del pro-ceso de traducción del material informativo del genoma hasta su expresión fenotípica, tanto en sus aspectos físicos y morfológicos como también conductuales, en la medida en que la genómica ya no se reduce al análisis de la secuencia de cada uno de los genes, sino que abarca también la combinación con otros elementos que los encienden, los frenan o los aceleran en su función. Por eso, si el ámbito del genoma es complicado, todavía lo es más el del pro-teoma.
2) La mera descripción de los datos científicos, como ya dejamos di-cho, es insuficiente para la comprensión de lo humano. El punto de vista científico no nos indica más que el funcionamiento de la má-quina corpórea del ser humano, pero no nos indica en sí mismo el sentido de la realidad humana. Afirmar que el ser humano se redu-ce a su genómica o proteómica, no es meramente una afirmación científica sino filosófica, y como tal afirmación filosófica es erró-nea e insuficiente. Si miramos sólo los datos fácticos, y se advierte que sólo nos diferenciamos en un 1% de los chimpancés, claro que se tendría que afirmar nuestro radical parecido con ellos, e incluso defender las tesis de J. Diamond de incluir a los chimpancés den-tro del mismo género en el que los taxonomistas sitúan a la especie humana29. El problema está en que reducir la comparación a ese dato cuantitativo es insuficiente.
3) La complejidad que nos manifiesta el proceso de expresión del ge-noma en el fenotipo, es una mostración de que la naturaleza con pocos cambios es capaz de generar realidades extraordinariamente diferentes. Es como los radios de una circunferencia: en el inicio están cercanos, pero a medida que se prolongan crece su separa-ción y diferencia. Es decir, en el caso del ser humano se advierte que unas pequeñas diferencias cuantitativas son la causa de poste-
29 Jared Diamond, en su libro The tirad chimpanzee: the evolution of the future of the human animal, N. York, Harper Perennial, 1993, propone que al ser humano habría que situarlo dentro de las otras dos especies de chimpancés, el chimpancé común (Pan trogro-dytes) y el chimpancé enano, llamado también bonobo (Pan panicus), o bien, incluir a es-tas dos especies de chimpancés en el género homo, y denominar a los bonobos, Homo pa-nicus, y al chimpancé común, Homo troglodytes. Cfr. J. Diamond, «El tercer chimpancé», en P. Cavalieri y P. Singer, El proyecto «Gran simio». La igualdad más allá de la humani-dad, Madrid, Trotta, 1998, 115-132.

427
© Universidad de Deusto - ISBN 978-84-9830-383-4
riores diferencias de gran significado, como son su cerebro, el len-guaje, el bipedismo permanente, la mano prensil, y en definitiva una conducta cualitativamente diferente.
4) Y aquí está la conclusión más significativa: lo fundamental no está en comparar las diferencias en el punto de origen, sino en sus ex-presiones morfológicas y conductuales. Y este punto de vista lo que nos muestra es que esas pequeñas diferencias son más signifi-cativas de lo que parecen, puesto que son la causa de un modo de ser y de actuar que sí representa una clara diferencia cualitativa, como vamos a tener ocasión de comprobar en otros aspectos que vamos a analizar a continuación. Todo esto nos hace ver, como in-dica J. Ruffié, que la naturaleza innova poco30, y con pocos cam-bios en lo genético es capaz de dar de sí realidades fenotípicas muy diferentes entre sí.
De ahí que podamos concluir que lo que nos diferencia con los anima-les más cercanos a nosotros no es sólo esa pequeña proporción de genes, sino también el itinerario que siguen hasta su expresión fenotípica, tanto morfológica como conductual. De ello tenemos que concluir que la esen-cia o naturaleza del ser humano no se halla encerrada exclusivamente en su ADN, como si fuera la potencia aristotélica que se expresa y se con-vierte en acto en el fenotipo, sino que el desarrollo del genoma desde el ADN hasta su expresión fenotípica es más complejo y decisivo en la con-figuración de un ser humano de lo que hasta ahora creíamos. Aunque to-davía no sepamos del todo identificar el conjunto de pasos que se dan en ese complejo y extraordinario proceso.
2.2.2. El punto de vista anatómico y morfológico31
Los parecidos anatómicos entre nuestra especie y los chimpancés son muy claros y abundantes. «El cuerpo humano, dice F. J. Ayala, está cons-truido con arreglo al mismo plan general que los cuerpos de otros ani-males, siendo más semejante al de los antropoides, los primates, los ma-míferos y los vertebrados, por este orden descendente. La semejanza y correspondencia, hueso a hueso, entre los esqueletos de un antropoide y un ser humano son impresionantes. Algunos de los huesos son tan semejantes entre sí que resulta difícil identificar la especie de procedencia»32.
30 Cfr. Ruffié, J., De la biología a la cultura, Barcelona, Muchnik Editores, 1982.31 Cfr. Ayala, F. J., o.c., 153-158; Laín Entralgo, P., El cuerpo humano. Teoría actual, o.c., 32 O.c., 154.

428
© Universidad de Deusto - ISBN 978-84-9830-383-4
Como es evidente, los parecidos con los antropoides o simios supe-riores es mucho mayor que con otros animales más distantes en la cadena evolutiva. Así, son dignos de señalarse estos parecidos entre los humanos y los chimpancés: la visión estereoscópica y policromática, la inmovilidad de las orejas, la reducción del olfato, la pérdida de pelos táctiles, la exis-tencia de un tiempo menstrual y ausencia de un período estacional de cría, producción de una sola cría por parto como regla común, gran cuidado maternal y dominancia de los machos adultos sobre las hembras e indivi-duos jóvenes33. Pero hemos introducido ya en esta lista elementos conduc-tuales, no sólo aspectos morfológicos y fisiológicos.
Ahora bien, a pesar de estas grandes similitudes, también se dan va-rias diferencias significativas que las vamos a sintetizar en cuatro:
a) el bipedismo, con todos los cambios morfológicos que supuso de cara a la conformación de los huesos de las extremidades inferio-res y la cadera, así como de la columna vertebral, orientadas a po-der acomodarse correctamente a caminar erguido de forma cons-tante34;
b) la mano prensil, con el dedo gordo oponible a los otros cuatro, que es consecuencia de cambiar su función de extremidad delantera para caminar, así como también para asirse a las ramas de los ár-boles, a instrumento para manipular objetos pequeños que exigen gran precisión de movimientos, así como para manejar herramien-tas y construirlas35;
c) la progresiva cerebralización y ampliación de la cavidad cranea-na36. Esta diferencia es un punto clave, y no se limita a la mera cantidad de la masa encefálica, sino a su estructura y composición. El cerebro humano no se reduce a ser más grande que el de los si-mios, sino que posee una estructura propia (mayor proporción de la corteza cerebral y de los lóbulos frontales y temporales, con dis-minución del lóbulo occipital) que lo aleja de la estructura propia de los primates, y más todavía del resto de los animales vivos.
d) y la transformación del conjunto de los rasgos faciales: disminu-ción del hocico y de los músculos que lo sustentan; transforma-
33 Cfr. ibídem, p. 155.34 Cfr. Coppens, Y., La rodilla de Lucy. Los primeros pasos hacia la humanidad, Bar-
celona, Tusquets, 2005.35 Cfr. Wilson, F. R., La mano. De cómo su uso configura el cerebro, el lenguaje y la
cultura, Barcelona, Tusquets, 2002. 36 Cfr. Morgan Llaman, J., El cerebro en evolución, Barcelona, Ariel, 2003; Braillon, M. G.,
El sistema nervioso central, Madrid, Alambra, 1980; Goldberg, E., El cerebro ejecutivo,Barcelona, Crítica, 2002.

429
© Universidad de Deusto - ISBN 978-84-9830-383-4
ción de la dentición37; acortamiento de la cavidad bucal, con re-traimiento de la lengua, que tendrá como efecto la transformación del tracto vocálico y la adecuación de la laringe para emitir soni-dos vocálicos38.
¿Qué conclusiones podemos sacar de estos datos? ¿Son significativos para defender una diferencia cualitativa entre los humanos y demás ani-males, o no da más que para una mera diferencia cuantitativa?
De nuevo tenemos que concluir de forma similar que en el apartado anterior:
— No es suficiente la mera constatación de datos y de hechos, pues-to que podrá decirse, con razón, que la diferencia en lo anatómico-morfológico entre la especie humana y los diferentes primates es menor que la que hay entre éstos y la inmensa mayoría de las espe-cies vivas.
— Ahora bien, para que la comparación sea significativa, tenemos que hacer referencia a la función o soporte que estas diferencias indi-cadas suponen de cara a comportamientos tan importantes y espe-cíficos como pensar (cerebro), hablar (cerebro-laringe), y demás comportamientos inteligentes y libres propios de nuestra especie. Por tanto, si tenemos en cuenta este aspecto, y no podemos dejar de hacerlo, se advierte con claridad que aspectos anatómicos como el cerebro y el conjunto del aparato fonador, por lo que suponen de soporte fisiológico de funciones tan importantes como pensar, ha-blar y actuar libre y responsablemente, constituyen elementos sufi-cientemente significativos como para servir de base a un criterio de diferenciación cualitativa, y no sólo cuantitativa.
2.2.3. La comparación desde el proceso embriológico
Un tercer punto de comparación, que resulta muy significativo, como vamos a ver, es el referente al proceso de ontogénesis, del que se ocupa la embriología. Si en la comparación morfológica veíamos el enorme pare-cido entre el hombre y las especies más cercanas, lo mismo ocurre en las
37 Cfr. Bermúdez de Castro, J. M.ª, El chico de la Gran Dolina. En los orígenes de lo humano, Barcelona, Crítica, 2002, cap. 7, «El reloj biológico de los dientes», pp. 93-109.
38 Cfr. Lieberman, Ph., «On the evolution of language: A unified view», Cognition,1973, n.º 2 («Un enfoque unitario de la evolución humana», en V. Sánchez de Zabala (ed.), Sobre el lenguaje de los antropoides, Madrid, Siglo XXI, 1986, 147-203); Laitman, J. T., «El origen del lenguaje articulado», Mundo Científico, 1986, n.º 64, 1182-1191.

430
© Universidad de Deusto - ISBN 978-84-9830-383-4
diferentes fases del proceso embriológico, según nos indica la embriolo-gía comparada39. Los parecidos son muy significativos, atravesando todo el proceso las mismas tres etapas fundamentales: cigoto (tres primeras se-manas), embrión (hasta el final de la octava semana) y feto (hasta el naci-miento).
Resulta significativo resaltar que en este proceso embriológico, se da una cierta repetición de la historia filogenética, resultando de algún modo certera la teoría de Haeckel de que la ontogénesis repite y resume de cier-ta medida la filogénesis. De este modo, el cuerpo humano en su desarrollo intrauterino viene a ser una recapitulación ontogenética de la filogénesis de su propio género y de toda la evolución.
Prueba de ello es que, en el proceso de desarrollo de la forma total del embrión, como ya indicamos en el capítulo anterior, van apareciendo de forma más o menos transitoria estructuras embrionales y fetales que ha-cen recordar formas de especies que se hallan situadas en el proceso evo-lutivo anterior, desde las amebas hasta los primates. Veamos algunos de estos fenómenos:
— la pasajera aparición de restos de branquias, recuerdo de nuestros antepasados los peces, y luego los anfibios;
— el paulatino cambio de posición de los ojos, desde una postura late-ral (es la posición de los ojos en los peces, lagartos, ratones, caba-llos, etc.) irán evolucionando hasta situarse delante de la cara, mi-rando hacia el frente, para adquirir una mirada estereoscópica;
— el esbozo de la cola (entre 8 y 9 segmentos vertebrales), que luego se transformará en el coxis. Lo posee el embrión de alrededor de seis semanas, y puede alcanzar una longitud entre 14 y 16 mm. En algunos casos especiales, no se supera esta fase y nacen de vez en cuando niños con cola40;
— la aparición del lanugo fetal, que se da entre el séptimo y noveno mes, y luego desaparece, recordándonos el fuerte sistema piloso de los mamíferos, y nos recuerda nuestra condición de «mono desnu-do» (D. Morris);
— la oclusión parpebral del embrión: hacia el tercer mes, los párpa-dos del embrión están cerrados y soldados, y así permanecen hasta el séptimo mes. Teniendo en cuenta este rasgo, se suelen dividir las especies animales en dos clases: nidícolas, predispuestas a alargar su fase de maduración en el nido, y nacen con los párpados cerra-
39 Cfr. Laín Entralgo, P., o.c., pp. 62-63; Lacadena, J. R., Genética y Bioética, Madrid/Bilbao, UPCO/Desclée de Brouwer, 2002, 15-71.
40 Cfr. Christen, Y., El hombre biocultural, o.c., p. 57.

431
© Universidad de Deusto - ISBN 978-84-9830-383-4
dos (es el caso del ratón, conejo, gato, perro y los monos inferio-res), y nidífugas, orientados a una independencia más rápida, na-ciendo con los ojos abiertos (ternero, caballo, foca, elefante). El ser humano participa de un aspecto de ambos grupos: por un lado, nace con los párpados abiertos, como los nidífugas, pero en cambio es la especie que alarga más su permanencia en el nido materno, dado su extrema invalidez, uno de los rasgos específicos de nuestra especie.
Podíamos hacer referencia a otros rasgos menores, que nos recuerdan la persistencia en el proceso embriológico de otros aspectos morfológicos de las especies anteriores, pero con lo dicho es más que suficiente para probar la tesis del resumen en el proceso ontogenético de las fases del proceso filogenético. Con lo dicho, estaríamos apuntalando el aspecto de los parecidos. Pero lo interesante es que en este proceso embriológico es donde se empiezan a ver mayores diferencias con las demás especies, apuntándose así el especial puesto del hombre en el proceso ontogenético general. ¿Dónde se hallarían las diferencias?
El proceso embriológico de todas las especies vivas sigue una lógica básica: los individuos de cada especie nacen en el momento en el que se ha conseguido, por parte del feto, la madurez correspondiente, la propia de la especie, para poder luego desenvolverse con éxito en la tarea funda-mental de adaptarse al entorno ambiental, a su nicho ecológico. Y, en fun-ción de esa lógica, durará más o menos el proceso de gestación y ontogé-nesis. Y, aunque cada animal sigue una lógica propia, podemos decir que todas las especies tienen la capacidad de traer al mundo individuos que pueden desenvolverse con relativa rapidez e independencia en la lucha por la supervivencia.
En cambio, en el ser humano se ha roto esta lógica básica, en la me-dida en que el proceso embriológico se ha detenido, y el recién nacido se parece al «feto de un mono». «Así, según Y. Coppens, en el estado adulto seríamos bastante semejantes a fetos de mono. En todo caso, más próxi-mos al feto del mono que al mono mismo. Esta persistencia de caracteres juveniles o fetales es lo que se llama neotenia»41. La especie humana es, pues, una especie que ha ralentizado su desarrollo, y nace antes de tiem-po, de forma prematura. Formamos una especie de prematuros, de inma-duros biológicos. Necesitaríamos estar doce meses más en el útero ma-terno (21 meses debería durar nuestra gestación) para poder nacer con la madurez biológica y comportamental con la que nace el resto de las espe-cies vivas, según Portmann42.
41 Cfr. Christen, Y., o.c., p. 58.42 Cfr. Die Biologie und das neue Menschenbild, Berna, 1942.

432
© Universidad de Deusto - ISBN 978-84-9830-383-4
Según los antropólogos, en la adquisición de este rasgo se ha dado un proceso de coevolución con otros elementos fundamentales de la especie humana: la postura erguida, que estrechó las caderas y el canal pélvico, hecho fundamental para las mujeres por su relación de cara al parto, lo que favoreció en el proceso evolutivo la tendencia a acortar el tiempo de gestación, para poder dar a luz hijos con una capacidad craneana adecua-da para poder atravesar el canal pélvico y llegar a nacer, dejando la tarea de ampliar la capacidad cerebral a los años posteriores a su nacimiento. De ahí que el nacimiento prematuro, que supone una mayor dependencia de los padres y de su entorno cultural, se tiene que conjugar y completar con un proceso de maduración y de dependencia más largo. Este factor, que representa la cara negativa de la deficiencia y de mayor dependencia, conlleva la ventaja de dotar al recién nacido de una mayor plasticidad, que redunda en su educación y maduración.
Son claves en este cometido las primeras fases posteriores al naci-miento, el denominado «año extrauterino del embrión», en la medida en que en esa fase primera ser producen procesos como la maduración de la vista y de los sentidos en general, la capacidad de caminar, la maduración de la mano prensil, una primera e importante fase de cerebralización (el recién nacido nace con el número completo de neuronas, unos cien mil millones, pero la maduración cerebral se va produciendo y completando después, y sobre todo, a través del logro de las innumerables conexiones sinápticas que serán el soporte cerebral de todas las habilidades conduc-tuales), y el empezar a balbucear un lenguaje elemental.
Por tanto, la deficiencia biológica de los seres humanos (A. Gehlen) le impele a ser necesariamente un animal cultural, a ser moldeado por el útero cultural con objeto de suplir esas deficiencias biológicas. Y en este proceso de maduración y de educación cultural, intervienen tanto las ca-pacidades innatas recibidas en su dotación genética como su capacidad de imitar y aprender de los demás compañeros de especie.
Si evaluamos en su globalidad este rasgo comparativo, vemos que ya las diferencias con los primates y demás especies animales se van agran-dando. En los dos aspectos anteriores (genética y morfología), las diferen-cias eran ciertas y evidentes, pero quizás no tan significativas y explícitas como esta curiosa y específica situación de nuestra especie en el proce-so ontogenético. Pero tenemos que volver a insistir en que, ateniéndonos exclusivamente a los datos científicos, son plausibles y legítimas las dos posturas en conflicto:
— la que interpreta las diferencias como meramente cuantitativas, se puede apoyar en el hecho de que hay más parecidos entre el hom-bre y los simios, que entre éstos y los animales más elementales; y

433
© Universidad de Deusto - ISBN 978-84-9830-383-4
más todavía si tenemos en cuenta que también en los chimpancés podemos advertir una cierta inmadurez y no fijación total, en clave de instintos, de sus pautas comportamentales. Sus presupuestos re-duccionistas no le permite a esta postura conceder más;
— y la que ve ya aquí una diferencia radical y cualitativa, que se com-pletará en el apartado siguiente, cuando hagamos referencia a la es-tructura comportamental específica del ser humano, que se sustenta precisamente en esta deficiencia biológica, siendo su cara positiva. Se advierte, pues, desde este punto de vista, tanto la continuidad como también la ruptura cualitativa entre nuestra especie y el res-to de las especies de la biosfera. Los seres humanos repetimos las mismas fases del proceso embriológico, pero de otra manera, dete-niéndose el proceso de maduración para empujarnos al mundo an-tes de tiempo: hemos dicho ya que parecemos el feto de un mono.Así, a diferencia de la tesis de los estratos de M. Scheler, que par-te de situar al ser humano en un peldaño más arriba del proceso de maduración evolutiva (acentuando la continuidad), la postura de Gehlen hace romper esta lógica, viendo al ser humano no como un animal más perfecto y maduro, sino como un animal diferente,que sigue otra ley y estrategia de maduración: suple con el aporte de aprendizaje cultural la deficiencia biológica. No hay, pues, aquí una superación desde una fase o escalón que subsumiría evolutiva-mente los anteriores, sino que rompe y supera esa lógica a través de un salto cualitativo. No persigue tanto adaptarse al ambiente, aunque fuera con una fórmula más perfecta (postura pasiva propia de las otras especies vivas), sino que adopta una postura activa y transformadora, por cuanto tiene que adaptar obligatoriamente el entorno a sus necesidades. De nuevo vemos que se ha invertido la lógica de la adaptación: el animal tiene ambiente, el hombre tiene mundo, construido artificialmente.
Este aspecto de inmadurez supone ya la base fisiológica de la especí-fica estructura comportamental del ser humano, por lo que no necesitamos detenernos aquí y pasamos a la siguiente faceta comparativa.
2.2.4. Comparación desde la estructura comportamental
Si en el ámbito de lo genético y morfológico, e incluso en el terreno del desarrollo embriológico, podemos apelar a unos datos más o menos objetivos, que necesitan posteriormente su interpretación, en el terreno del análisis del comportamiento o estructura comportamental el esfuerzo in-

434
© Universidad de Deusto - ISBN 978-84-9830-383-4
terpretativo se hace necesario casi desde el primer momento, porque casi no existen meros datos objetivos de comparación. Todos dependen ya de una mirada interpretadora, deudora de un trasfondo antropológico y cos-movisional. Pero, al mismo tiempo, constituye este aspecto el punto más importante y decisivo para el tema que estamos estudiando aquí, en la me-dida en que el punto donde se diferencian más claramente hombre y ani-males es precisamente en su estructura comportamental.
¿En qué se diferencia el comportamiento de los animales del de los humanos? ¿Podemos establecer una división entre los humanos y el resto de los animales, o nuestro comportamiento es sólo un poco más inteligen-te y complejo que el de los primates, habiendo entre éstos y los animales anteriores una diferencia tan grande y significativa como la que hay entre los primates y los humanos?
El empeño por estudiar el comportamiento de los animales se puede encontrar ya en la época de Aristóteles. Pero el estudio científico se co-menzó a realizar a partir de las teorías de Darwin. Es a partir de entonces cuando comienzan a darse dos planteamientos contrarios en la interpre-tación de los comportamientos animales: los innatistas y los ambientalis-tas43. Los primeros intentos serios sobre el comportamiento animal dan lugar a la aparición de la etología, de la mano de tres científicos europeos, Konrad Lorenz, Karl von Frisch y Niko Tinbergen, que recibieron con-juntamente el Premio Nobel en 1973. La etología de estos autores insiste en la dimensión innata de la conducta animal, pero entienden que el com-ponente innato hay que conjugarlo y complementarlo con el aprendiza-je44. En cambio, el conductismo norteamericano, de la mano de Watson y Skinner, defendía que la conducta animal y humana era fruto exclusivo de las influencias ambientales.
La mayoría de los estudiosos del comportamiento, tanto animal como humano, entienden que es el resultado de compaginar lo innato con lo aprendido, aunque el problema radica en cómo conjugar ambos compo-nentes. Pero sigue habiendo partidarios de las dos posturas extremas: el innatismo y el ambientalismo. Referidas al comportamiento humano, am-bas posturas defienden tesis deterministas, aunque sus posturas suelan ser muy ambiguas en este punto, como vamos a ver. Nos detendremos, en pri-mer lugar, en analizar los planteamientos de la sociobiología (determinis-mo genético) y del conductismo (determinismo ambiental), para luego re-ferirnos a las posturas que conjugan lo innato y lo aprendido.
43 Cfr. Maier, R., Comportamiento animal. Un enfoque evolutivo y ecológico, o.c., 4-6.44 Cfr. Thorpe, W. H., Breve historia de la etología, Madrid, Alianza, 1982; Beorle-
gui, C., «El reto de la biología a la antropología: De la etología a la sociobiología», Letrasde Deusto, 16 (1986), n.º 34, 37-69.

435
© Universidad de Deusto - ISBN 978-84-9830-383-4
2.2.4.1. LAS TESIS DETERMINISTAS
a) El determinismo genético de la sociobiología45
La Sociobiología, nacida en los USA a partir de la obra de E. O. Wilsony otros autores, pretende explicar tanto el comportamiento animal como el humano desde las leyes centrales de la genética. Para estos autores, el su-jeto de la evolución no es ni la especie (como defendía K. Lorenz) ni los individuos, sino los genes. El objetivo que persiguen es la supervivencia y estar representados del modo más abundante en las generaciones siguien-tes. En definitiva, la ley que los empuja es la del egoísmo genético (el gen egoísta, R. Dawkins). Y con esta lógica pretenden explicar toda la con-ducta de los seres vivos, sintetizada en cuatro ámbitos fundamentales: la agresividad, la relación entre sexos, la relación paternofilial, y las conduc-tas «altruistas»46.
Es importante advertir que no todos los sociobiólogos defienden la extrapolación de estas pautas de conducta al ámbito humano. Mientras la corriente norteamericana defendería tal extrapolación, la inglesa, con Dawkins a la cabeza, sería partidario de separar ambos terrenos.
En definitiva, para los sociobiólogos la cultura humana no es más que un componente más nacido por evolución y controlado directamente por los genes, y, por tanto, tiene que estar al servicio de los genes y de su ló-gica de egoísmo genético. Y cuando decimos cultura estamos hablando de todos sus componentes, también la ética y la religión. La ética sería, para Wilson, una estrategia más de los genes al servicio de la supervivencia, al igual que a otras especies les ha dotado de alas, branquias o garras para sobrevivir en un nicho ecológico determinado47.
Frente a los evolucionistas clásicos, que pretendían deducir las normas éticas de la lógica evolucionista, cayendo de ese modo en la falacia natu-ralista, Wilson pretende escapar a esa falacia, pero creemos que sin conse-guirlo. Así, cuando afirma que «el comportamiento humano —con las capa-cidades de respuesta emocional que lo impulsan y lo guían— es la técnica indirecta por la cual el material genético humano ha sido, y será, manteni-do intacto. La moralidad no tiene otra función última demostrable»48. Pero, como indica F. J. Ayala, es una afirmación incorrecta tanto si se refiere a la
45 Cfr. Wilson, E. O., Sociobiología. La nueva síntesis, Barcelona, Omega, 1980; Id., Sobre la naturaleza humana, México, FCE, 1980; Ruse, M., Sociobiología, Madrid, Cáte-dra, 1983; Dawkins, R., El gen egoísta, Barcelona, Labor, 1979.
46 Cfr. Ruse, M., Sociobiología, o.c. 47 Cfr. Wilson, E. O., On Human Nature, Cambridge, Massachussets, 1978 (Sobre la
naturaleza humana, México, FCE, 1980). 48 Wilson, E. O., Sobre la naturaleza humana, p. 167.

436
© Universidad de Deusto - ISBN 978-84-9830-383-4
capacidad ética (puesto que los humanos somos éticos por naturaleza, y no porque esto sirva a la eficacia genética) o a los contenidos éticos. Si se re-fiere a estos segundo, sería defender la falacia naturalista.
Las críticas a la Sociobiología suelen venir de tres ámbitos diferentes. Desde el estrictamente científico, los etólogos tienen que comprobar si las tesis sociobiológicas se sostienen, referidas al campo exclusivamente del comportamiento animal. Esto queda en manos de los etólogos, y no todos están de acuerdo con las tesis sociobiológicas, puesto que si en mu-chos campos del comportamiento animal parecen comprobarse sus tesis, no ocurre así en todos los casos. Por ejemplo, en el terreno del altruismo, puesto que no siempre las tesis egoístas se cumplen y son más eficaces, ni tampoco está claro que el gen sea el sujeto de la evolución49. Otro campo de crítica se refiere a la legitimidad o no de extrapolar las tesis biológicas al área del comportamiento humano, puesto que ello supondría negar la li-bertad humana, y la autonomía de la cultura, con todos sus diferentes ám-bitos; así como no poder escapar a la ambigüedad consistente en defender el determinismo y mantener planteamientos éticos (Wilson, en «Sobre la naturaleza humana»)50. Y en tercer lugar, mirando el problema desde la so-ciología del conocimiento, se ha acusado a los sociobiólogos de un claro sesgo ideológico en sus planteamientos51.
b) El determinismo ambiental del conductismo
Se trata de un enemigo que podemos considerar ya superado en su globalidad, desde mitades del siglo XX. Pero nos referimos a él como un modo clásico y ejemplar de entender la condición humana. Frente a la postura sociobiológica, que sitúa el motor de nuestra conducta en los ge-nes, con su lógica egoísta, el conductismo de Watson y de Skinner da por hecho que somos hechura de nuestro entorno. La estructura comporta-mental está configurada por la relación directa estímulo-respuesta (E-R), por más que este sencillo esquema se pueda completar y complejificar por la retroalimentación, según Skinner, de los refuerzos (positivos y negati-vos), que suponen la diversa forma de responder el ambiente (natural y humano) a nuestra respuesta al mismo.
49 Cfr. Sober, E. y Wilson, D. S., El comportamiento altruista. Evolución y psicología,Madrid, Siglo XXI, 2000.
50 Cfr. Ayala, F. J., Origen y evolución del hombre, Madrid, Alianza, 1980, cap. VII; Beorlegui, C., «El reto de la biología a la antropología. De la etología a la sociobiolo-gía», o.c.
51 Cfr. Sahlins, M., Uso y abuso de la biología, Madrid, Siglo XXI, 1982; R. Lewon-tin, S. Rose y L. I. Kamin, No está en los genes. Racismo, genética e ideología, Barcelona, Crítica, 1987.

437
© Universidad de Deusto - ISBN 978-84-9830-383-4
Hoy día son pocos, si es que hay alguno, que defienda el conductis-mo, como postura antropológica determinista, en toda su crudeza, tal y como se dio en la primera mitad del s. XX en los USA. Sus planteamientos perdieron apoyos como consecuencia del progresivo triunfo de las tesis etológicas, que reivindicaban el elemento innato y genético en la conduc-ta, y de las ciencias cognitivistas, que supusieron el retorno del concepto de lo mental.
Así, si es verdad que las circunstancias del ambiente nos influyen, en la medida que sea, sin embargo no nos determinan. Por otro lado, se da en el conductismo, sobre todo en el de Skinner, la misma ambigüedad que en el sociobiólogo Wilson a la hora de querer conjugar determinismo y plan-teamientos éticos (como en su utopía Walden dos), pudiendo advertirse una doble concepción del ser humano: por un lado, lo que podríamos de-nominar el hombre masa, sobre el que teoriza Skinner, hombre que, por hallarse dominado por el ambiente, necesita un salvador que le libere de su dominación y le trasplante a un nuevo paraíso terrenal (Walden dos), y por el otro, el hombre élite, representado por el propio Skinner y cualquier ingeniero de la conducta, que es consciente de esa dominación del am-biente, no está sometido a ella, quiere liberar al hombre masa, y construye un artefacto social para liberarlo. Si no se hace esta distinción antropoló-gica, difícilmente se puede aceptar como coherente la teoría conductista.
2.2.4.2. ENTRE LA GENÉTICA Y EL AMBIENTE
Los estudios sobre el comportamiento animal y humano nos indican que, tanto en el caso de los animales como en el ser humano, el compor-tamiento tiene componentes innatos y aprendidos, llevándonos a superar determinados mitos del pasado, como, en primer lugar, entender el com-portamiento humano como fruto sólo del aprendizaje, como defiende la denominada tesis de la tabla rasa desde J. Locke; o, en segundo lugar, entender el comportamiento animal como fruto exclusivo de un progra-ma innato, mientras que el de los humanos es resultado exclusivo de la experiencia; o también creer que los aspectos genéticos influyen antes del nacimiento y los ambientales sólo a partir del nacimiento; otra idea equi-vocada consiste en pensar que hay aspectos de la conducta humana gené-ticamente determinadas, y otras, sólo influidas por el entorno ambiental. La realidad es que todos los aspectos de la conducta humana son el resul-tado de la interacción entre genes y ambiente. Pero no es posible deter-minar la parte o porcentaje correspondiente a lo genético y qué parte a lo aprendido.
Las investigaciones actuales se orientan en la línea de estudiar casos de hermanos gemelos criados por separado, para ver en qué medida han

438
© Universidad de Deusto - ISBN 978-84-9830-383-4
desarrollado conductas similares o diferentes52. Pero, a parte de la preca-riedad científica de estas investigaciones, en todas ellas se advierte que la conducta humana no está determinada ni por los genes ni por el ambiente, sino por ambos componentes y por la libertad de cada individuo.
En la actualidad, otro modo de acercarse al estudio científico de la con-ducta humana es la denominada Psicología evolucionista, que persigue «afrontar el reto de analizar la conducta humana asumiendo, con todas sus consecuencias, el origen evolutivo de nuestra especie»53. Los pioneros de esta disciplina han sido J. H. Barkow, L. Cosmides y J. Tooby, con su libro fundamental The Adapted Mind: Evolutionary Psychology and the Genera-tion of Culture54. Un autor importante y muy conocido de esta disciplina es S. Pinker, discípulo de Chomsky y uno de los teóricos más interesantes acer-ca del origen del lenguaje humano55. Conscientes de que la conducta huma-na es una síntesis de mecanismos genéticos e influencias del entorno, los psicólogos evolucionistas intentan descubrir los mecanismos psicológicos que se han ido conformando en nuestra especie como resultado de su histo-ria evolutiva, desde el momento en que apareció la especie hace más de dos millones de años. Esos mecanismos psicológicos serían los responsables de los principales rasgos de la conducta humana, como la capacidad intelectual, la agresividad, los comportamientos sexuales, la cooperación en sus dife-rentes niveles, la diversidad cultural y otros más. Se trataría, por tanto, de encontrar ese conjunto de rasgos psicológicos comunes que conformarían la naturaleza humana, conformada fuertemente por ingredientes genéticos. De todos modos, aunque Pinker se desmarca claramente del determinismo ge-nético de la sociobiología, considera que muchos de sus planteamientos son correctos, y se muestra, de modo significativo, más duro con los críticos de la sociobiología, el grupo denominado Science for the People56, que las te-sis deterministas de los sociobiólogos. Eso muestra que en la articulación de lo innato y lo aprendido, Pinker tiende a acentuar en gran medida el primer
52 Cfr. Plomin, R., De Fries, J. C., McCleam, G. E. y McGuffin, P., Genética de la con-ducta, Barcelona, Ariel, 2002; Wright, W., Así nacemos. Genes, conducta, personalidad,Madrid, Taurus, 2000; Ridley, M., Qué nos hace humanos, Madrid, Taurus, 2004.
53 Cfr. Castro, L., López-Fanjul, C. y Toro, M. A., A la sombra de Darwin. Las aproxi-maciones evolucionistas al comportamiento humano, Madrid, Siglo XXI, 2003, 130.
54 New York, Oxford University Press, 1992. 55 Cfr. Pinker, S., The Blank Slate, Nueva York, Penguin, 2002 (trad. cast., La tabla
rasa. La negación moderna de la naturaleza humana, Madrid, Alianza, 2003); Beorle-gui, C., «A vueltas con la naturaleza humana», Arregui, J. V. (ed.), Debate sobre las antro-pologías», Sevilla, Themata, 2005, n.º 35, 139-150.
56 Entre ellos se sitúan Lewontin, Kamin, Rose y S. J. Gould. Cfr. Lewontin/Kamin/Rose, No está en los genes, o.c.; Lewontin, R. C., El futuro del genoma humano y otras ilusiones,Barcelona, Paidós, 2001, cap. 1.º; Gould, S. J., La falsa medida del hombre, Barcelona, Anto-ni Bosch, 1984.

439
© Universidad de Deusto - ISBN 978-84-9830-383-4
componente en detrimento del segundo, sin ser suficientemente consciente de que resulta difícil, por no decir imposible, distinguir la aportación de cada uno de esos dos componentes en aspectos como, por ejemplo, la medición de la inteligencia y en otros ámbitos del comportamiento de los individuos57.
3. LA SINGULAR ESTRUCTURA COMPORTAMENTAL DE LOS SERES HUMANOS
En definitiva, tras todas estas precisiones, parece que podemos con-cluir que la estructura comportamental del ser humano y la del resto de los animales es cualitativamente diferente.
Es cierto, como hemos indicado, que no es correcto generalizar acer-ca de los animales, defendiendo que su conducta se encierra dentro de un esquema innato Estímulo-Respuesta (E-R). Los animales, sobre todo los superiores, además de sus capacidades innatas aprenden del entorno y les influye el ambiente. Pero sus capacidades cognitivas no pasan de un cier-to nivel ni son capaces de una conciencia de segundo nivel como los seres humanos, ni son capaces de romper los constreñimientos biológicos como para poder elegir libremente entre diversas posibilidades de acción.
Si hacemos referencia a las investigaciones con las diferentes especies de grandes simios (chimpancés, bonobos, gorilas y orangutanes), hemos de reconocer la posesión de grandes capacidades cognitivas que con anteriori-dad se minimizaban58. Y, aunque las primeras investigaciones estaban im-pregnadas de errores metodológicos que hacían pensar más en respuestas de amaestramiento que en respuestas con contenido cognitivo autónomo, investigaciones posteriores parecen avalar una riqueza interior que nos deslumbra y rompe muchos de nuestros esquemas anteriores. Todas las es-pecies animales parecen tener su propio sistema de comunicación interna, pero eso no significa que posean una teoría de la mente59, es decir, la ca-pacidad de ponerse en el lugar de otro congénere. Pero es cierto que deter-minados animales superiores utilizan un modo de comunicación a través
57 Cfr. Beorlegui, C., «A vueltas con la naturaleza humana», o.c., pp. 141 y ss. 58 Cfr. De Waal, Frans, La política de los chimpancés, Madrid, Alianza, 1993; Id., Bien natu-
ral. Los orígenes del bien y del mal en los humanos y otros animales, Barcelona, Herder, 1997; Id., Primates y filósofos. La evolución de la moral del simio al hombre, Barcelona, Paidós, 2007.
59 Cfr. Premack, D. y Woodruff, G., «Does the chimpanzee have a theory of mind?», Behavioral and Brain Sciences, 1978, n.º 1, 515-526 (trad. cast.: «¿Tiene el chimpancé una teo-ría de la mente?», en Martí, E., Construir una mente, Barcelona, Paidós, 1997, pp. 137-178);Premack, D., «Does the chimpanzee have a theory of mind?», Byme, R, y Whiten, A. (comps.), Machiavellian Intelligence, Oxford, Science Publication, 1988.

440
© Universidad de Deusto - ISBN 978-84-9830-383-4
de gestos y de gruñidos que parece implicar un cierto contenido semántico. Incluso hay chimpancés que pueden llegar a utilizar el lenguaje de los sor-domudos, o un lenguaje en que se utilizan fichas y teclado de ordenador. Hasta poseen un incipiente grado de autoconciencia, pues parecen recono-cerse ante el espejo. Y hasta parecen capaces de emplear lo que se ha veni-do a llamar conductas maquiavélicas, puesto que tratan de engañar a otros semejantes, percibiéndose en esas conductas un alto grado de capacidad de ponerse en el lugar del otro.
¿Significa esto que podemos deducir que la diferencia entre la capacidad cognitiva y la estructura comportamental entre los hombre y los animales su-periores, los grandes simios, es sólo cuantitativa, pero de ningún modo cua-litativa? Pensamos que no. En este punto, como en los anteriores aspectos mencionados, se da una continuidad junto con una clara ruptura cualitativa. Es evidente que se da una continuidad: los seres humanos no innovan nada, hemos indicado ya con palabras de J. Ruffié, sino que lleva hasta un nivel de perfección extraordinario un conjunto de tendencias y cualidades que apare-cen ya en diversas especies animales anteriores. Pero también se da un salto cualtitativo importante. Lo que nos diferencia respecto a las especies anterio-res, incluso las más cercanas y mejor dotadas, es muy significativo.
a) Una mayor capacidad cognitiva, debido a nuestra extraordina-rio desarrollo cerebral, que nos ha permitido llegar a poseer pen-samiento simbólico, recordar el pasado, anticipar el futuro, crear símbolos arbitrarios, etc.
b) La capacidad lingüística, que nos dota en este punto de una clara diferencia cualitativa no sólo en lo fonético, sino en lo semántico, sintáctico y pragmático60. La diferencia entre el supuesto lengua-je animal y el humano no es gradual, sino cualitativo (Bickerton y sus teorías sobre los lenguajes criollos o pidgin61).
c) La autoconciencia, que no es un simple aumento de la autopercep-ción animal, sino un segundo nivel de conciencia.
d) La libertad y la capacidad moral, por lo que podemos afirmar que sólo los humanos somos animales éticos.
e) La complejidad social: nacemos dotados de la capacidad de po-nernos en el lugar del otro (psicólogos innatos, Humphrey62), de
60 Cfr. Lorenzo, G./Longa, V. M., Homo loquens. Biología y evolución del lenguaje,Lugo, Tris Tram, 2003; Beorlegui, C., «La capacidad lingüística del ser humano: una dife-rencia cualitativa», Thémata. Revista de Filosofía, 2006, n.º 37, 139-168.
61 Cfr. Bickerton, D., Lenguaje y especie, Madrid, Alianza, 1994.62 Cfr. Humphrey, N. L., La reconquista de la conciencia. Desarrollo de la mente hu-
mana, México, FCE, 1987; Id., La mirada interior, Madrid, Alianza, 1993; Id., Una histo-ria de la mente. La evolución y el nacimiento de la conciencia, Barcelona, Gedisa, 1995.

441
© Universidad de Deusto - ISBN 978-84-9830-383-4
modo que nuestro entorno ecológico más cercano y específico son los otros (entorno social), no tanto la naturaleza.
f) La apertura a la pregunta por el sentido (filosofía), la inevitabili-dad de la muerte, la pregunta por el fundamento y el absoluto (reli-gión), etc.
Todos estos ingredientes son suficientemente significativos como para que podamos afirmar que la diferencia entre los hombres y demás anima-les no es sólo cuantitativa sino cualitativa. Por tanto, estamos de acuerdo con Chomsky cuando considera que, a la vista de estas diferencias, no es correcto hablar con idénticas palabras para las acciones animales y las hu-manas, por ejemplo inteligencia o lenguaje, puesto que la diferencia es abismal y cualitativa. Es como decir que los seres humanos vuelan, por-que son capaces de dar grandes saltos y permanecer en el aire durante un breve tiempo. Como tampoco vale creer que los simios podrán con el tiempo, y con el adiestramiento adecuado, llegar a dominar y entender el lenguaje humano y comunicarse adecuadamente con nosotros. Los que así piensan, dice también Chomsky, parecen imaginar que es razonable esperar que haya alguna isla desierta y desconocida en donde habitan aves con capacidad para volar pero que no vuelan; están esperando a que los humanos descubramos esa isla y les enseñemos a volar. Si la selección na-tural ha dotado a tales aves de la capacidad de volar, lo habrá hecho por-que esa cualidad es valiosa para su supervivencia, y resulta absurdo que estén esperando a utilizarla hasta que algún ser humano les enseñe a utili-zarla. Algo así parecen creer algunos sobre la supuesta capacidad intelec-tual y de lenguaje en los chimpancés y demás simios.
El ser humano, como consecuencia del proceso evolutivo, ha sido configurado de un modo tal que posee cualidades y un modo de com-portamiento radicalmente nuevo y cualitativamente diferente al del resto de los animales. Lo esencial está en que nuestro comportamiento no está limitado por un programa genético y biológico, sino que se ha produci-do una ruptura, de modo que no tenemos estructuras comportamentales cerradas sino abiertas. Así, entre los estímulos que percibimos y las res-puestas que tenemos que realizar, se da un hiato o vacío que tenemos que llenar de modo reflexivo. Es decir, tenemos que pensar y elegir entre di-versas formas de responder. Y esa es la condición de posibilidad biológica de nuestra libertad y de nuestra capacidad ética. Es la expresión de nues-tra condición inmadura, como vimos, de nacer antes de tiempo, y de haber roto la lógica del desarrollo embrionario que siguen las demás especies.
Por tanto, no es que los seres humanos seamos más inteligentes o más complejos o capaces de interactuar con el entorno, sino que lo hacemos de otra manera, siguiendo una lógica o una estructura comportamental radi-

442
© Universidad de Deusto - ISBN 978-84-9830-383-4
calmente diferente. Zubiri lo expresa acertadamente diciendo que al ani-mal se le da la realidad bajo la formalidad de estimulidad, mientras que al ser humano, bajo la formalidad de reidad o de realidad. El ser humano está vertido a la realidad, y no a la mera estimulidad. No percibimos me-ros estímulos sino realidades, cosas reales. Y nuestra habitud, o modo de habérnoslas con la realidad es radicalmente diferente. El animal siente, mientras que el ser humano, consecuente con su estructura psicosomáti-ca, intelige sentientemente. Siente pero de modo inteligente. E intelige, pero de modo sentiente. Así, sus actos son complejos pero unitarios. Eso significa que el ser humano ha sido elevado a una nueva estructuraciónesencial, en la medida en que su animalidad ha sido elevada o configurada por una estructura nueva dotada de intelección. Y todos sus actos son al mismo tiempo sensibles e intelectivos, dándose en unos dominancia de lo sensitivo (digerir, respirar), y en otros, dominancia de lo intelectivo (pen-sar, decidir)63.
4. ¿AMPLIACIÓN DE LA «HUMANIDAD» A LOS GRANDES SIMIOS?
Si en los apartados anteriores nos hemos referido a quienes defienden un igualamiento entre la especie humana y los demás animales, merece hacer referencia a quienes en la actualidad también pretenden diluir la di-ferencia hombre-animales pero no tanto por un proceso de abajamiento del ser humano, sino por una elevación de determinadas especies anima-les, las de los denominados grandes simios, al nivel de lo humano. Este movimiento, liderado entre otros por P. Singer y T. Reagan, pretende la liberación animal y la defensa de los derechos de los animales, argumen-tando que la distinción que tradicionalmente se hace entre la especie hu-mana y las de estos grandes simios basada en la racionalidad y la autono-mía no es pertinente, sino que debe hacerse desde la capacidad para sufrir y gozar.
A partir de la época moderna, la ética se ha fundamentado y justifica-do desde la autonomía de la razón y del individuo. Kant nos enseñó que la persona humana, debido a su autonomía y racionalidad, es digna de respe-to y sujeto de derechos, en la medida en que es un fin en sí misma y posee
63 Cfr. Zubiri, X., Sobre la esencia, Sociedad de Estudios y Publicaciones, Madrid, 1972; Id., Inteligencia sentiente, 3 vols., Madrid, Alianza/Sociedad de Estudios y Publica-ciones, 1980-82; Id., Sobre el hombre, Madrid, Alianza/Sociedad de Estudios y Publicacio-nes, 1986; Id., Estructura dinámica de la realidad, Madrid, Sociedad de Estudios y Publi-caciones, 1989.

443
© Universidad de Deusto - ISBN 978-84-9830-383-4
valor y no precio; esto es, no puede ser usada como medio para ningún otro fin o valor. Pero esta fundamentación racional fue puesta en cuestión posteriormente por el emotivismo y el utilitarismo. El emotivismo sostie-ne que la fuente de la moral son los sentimientos, puesto que el ser huma-no actúa moralmente llevado por los sentimientos y emociones, impulsa-do sobre todo por la compasión. Además, el utilitarismo, de la mano de J. Bentham y de J. Stuart Mill, sostiene que la bondad y maldad de los ac-tos no se advierten en sí mismos sino por sus consecuencias.
Por tanto, nos encontramos con dos modelos de ética muy diferentes, apoyados cada uno en un modelo distinto de persona. Mientras la pro-puesta ética kantiana es una propuesta formal y procedimental (no pode-mos llegar a una ética material universal, sino a una forma o procedimien-to universalizable: el imperativo categórico) y deontológico (una ética basada en principios, obtenidos a través del imperativo categórico, de obligado cumplimiento para todos), el utilitarismo es emotivista (defiende una ética basada en sentimientos) y consecuencialista (los principios no valen en sí, sino por sus consecuencias positivas). Igualmente, sus mode-los antropológicos son diferentes. Mientras el modelo kantiano se basa en un humanismo antropocéntrico, basado en la autonomía y en la razón (es la razón y no los sentimientos lo que distingue y caracteriza a las perso-nas), el emotivismo utilitarista se apoya en una idea de persona basada en la capacidad de sufrir y de gozar. Todo ser capaz de sufrir y de gozar ten-drá derecho a que se respeten sus sentimientos y sus intereses, con lo que en la actualidad hay teóricos que pretenden introducir en la clase o grupo de personas a determinados animales dotados de una especial sensibilidad para el goce y el sufrimiento.
De todos modos, los planteamientos de los dos teóricos más influyen-tes en la defensa de los supuestos derechos de los animales, P. Singer y T. Reagan, son un tanto diferentes. P. Singer, apoyado en la tradición emo-tivista y utilitarista, considera que el mal es un mal que debe ser evitado en la medida de lo posible64. En realidad, estos derechos no los extiende a todos los animales, sino a los grandes simios65, por lo que su postura es
64 Cfr. Singer, P., Liberación animal, Madrid, Trotta, 1999; Id., Desacralizar la vida humana. Ensayos sobre ética, Madrid, Cátedra, 2003; Beorlegui, C., «¿Hacia un humanis-mo trans-antropocéntrico? Peter Singer y los “derechos” de los animales», Realidad (UCA, San Salvador), 2001, n.º 80, 199-236; Id., «Ética animal e idea de persona», en Guibert, J. M. y Orella, J. L. (eds.), Preguntas éticas en cuestiones disputadas hoy. In memoriam de Juan Plazaola Artola, S. I., San Sebastián, Universidad de Deusto, 2005, 317-335; Torralba, F., ¿Qué es la dignidad humana? Ensayo sobre Peter Singer, Hugo Tristram Engelhardt y John Harris, Barcelona, Herder/Instituto Borja de Bioética, 2005.
65 Cfr. Cavalieri, P. y Singer, P. (eds.), El proyecto «Gran Simio». La igualdad más allá de la humanidad, Madrid, Trotta, 1998.

444
© Universidad de Deusto - ISBN 978-84-9830-383-4
considerada por los más radicales como reformista. Tacha a las diversas posturas humanistas, sean religiosas o laicas, de especieísmo, puesto que defienden un etnocentrismo humano según él ilegítimo, en la medida en que la diferencia entre los humanos y los animales superiores (los gran-des simios) no sería esencial, sino gradual, y estaría basada en la común capacidad de sufrir y de gozar. Ahora bien, P. Singer es consciente de que en caso de conflicto entre determinados derechos de los humanos y de los simios, los derechos de los humanos pasarían por delante, pero eso no su-pondría tener que afirmar que es preferible cualquier sufrimiento animal respecto al sufrimiento humano, ni que se pueda legitimar cualquier sufri-miento animal para satisfacer cualquier deseo o capricho humano.
T. Reagan, en cambio, defiende los derechos de los animales apoyán-dose en tesis principialistas y deontológicas, en la medida en que entiende que los derechos de los animales poseen una valía intrínseca66. Por tanto, no se trata, como en el caso de P. Singer, de defender los derechos de los animales porque también ellos sufren y gozan, sino porque los animales en general, y más aún los superiores, como los mamíferos, poseen una vida interior muy compleja y evolucionada, que les capacita para ser su-jetos-de-una-vida y les dota de un valor inherente. Esta supuesta inheren-cia ontológica y ética es la que le empuja a Regan a pedir que se respete la dignidad de los animales y a que no se les haga sufrir. Esta petición no tiene recortes ni paliativos, puesto que Reagan aplica un valor inherente a todos los animales, y en general a todos los seres vivos, y por ello todos, hombres y animales, tenemos los mismos derechos morales básicos.
Estaríamos, pues, ante propuestas que defienden, aunque con razo-nes diferentes, una ampliación de la idea de persona, que no abarcaría tan sólo a la especie humana sino que también acogería algunas especies superiores, según la línea de Singer, o la de todos los animales, como propone T. Reagan, e incluso, según los ecologistas más radicales, a to-dos lo seres vivos. Muchos de estos teóricos se apoyan en la propuesta de D. Dennett de entender que el concepto de persona es un concepto borrosoy se puede aplicar a determinados animales67. Las condiciones que, se-gún Dennett, debiera tener quien aspire a ser considerado una personaserían: ser racionales, poseer adscripciones intencionales, ser sujeto de consideraciones morales, capacidad para adoptar una actitud recíproca de respeto, usar lenguaje, y disponer de autoconciencia. Estas cualida-des, que cualquier humanista aceptaría en principio, por parecer que son
66 Cfr. Regan, T., Animal Rights, Berkeley, University of California Press, 1983; Id., «Poniendo a las personas en su sitio», Teorema, XVIII/2 (1999), 17-37.
67 Cfr. Denett, D., «Conditions of Personhood», en Brainstorms. Philosophical Essays on Mind and Psychology, Montgomery, Bradford Books, 1976.

445
© Universidad de Deusto - ISBN 978-84-9830-383-4
suficientemente significativas como para mantener la distancia cualita-tiva respecto al resto de los animales, en el contexto de Dennett sirven para defender una idea borrosa de persona, en la medida en que esas seis condiciones las cumplirían también los animales superiores.
Detenerse a desmontar todos estos presupuestos nos llevaría dema-siado tiempo. Estos planteamientos de Dennett y sus seguidores caen en el mismo error que hemos visto en el apartado anterior referido a la di-ferencia entre hombres y animales en relación a su estructura comporta-mental: insisten en los parecidos y en la incipiente continuidad entre esas características humanas, que en los animales sólo se hallan apuntadas y en embrión. Sólo para quien parta de una concepción filosófica reduccionis-ta y naturalista podrá resultar convincente la visión de una mera diferen-cia cuantitativa entre hombres y animales, como les parecerá también una mera diferencia cuantitativa la existente entre el salto de una persona y el vuelo de un pájaro, según en señalado símil de N. Chomski.
En los animales se hallan presentes, como en embrión, casi todas las cualidades que en los seres humanos llegan a su madurez y concre-ción, desde la conciencia y la comunicación, hasta la capacidad técnica de construir herramientas. Pero de ninguna manera se pueden ignorar las características específicas del ser humano, como son la libertad y la capa-cidad ética, el lenguaje articulado y la apertura a la cuestión del sentido, entre otras. Si no queremos introducir confusiones inaceptables, hemos de reconocer que sólo los individuos que poseen estas características pueden ser considerados como personas.
Eso no quita que no debamos crecer en aprecio y respeto por los ani-males y la naturaleza en general, y proponer normas de respeto y de pro-tección de la biosfera y de las diferentes especies animales, aceptando in-cluso que denominemos a eso derechos de los animales68. Pero otra cosa es que diluyamos el concepto de persona, atribuyendo características ex-clusivas de los humanos a determinados animales. De ahí que pensemos que es totalmente legítimo seguir defendiendo un humanismo antropocén-trico, complementado con la defensa de una normativa que defienda un trato respetuoso con las diferentes especies animales. Igualmente, nos pa-rece también adecuado completar la idea tradicional de persona, no basán-dola exclusivamente en su racionalidad sino también en su sensibilidad, en la medida en que el ser humano es una unidad biocultural, o lo que es lo mismo, una unidad psico-orgánica, en la que no sólo cuenta lo psíqui-co y racional sino también la sensibilidad y los sentimientos. Se trata, por
68 Así lo defiende un etólogo tan cercano a posturas antropomórficas en relación con los primates como Fras De Waal, en Primates y filósofos. La evolcuón de la moral del si-mio al hombre, Barcelona, 2007, pp. 105-111.

446
© Universidad de Deusto - ISBN 978-84-9830-383-4
tanto, de acogernos a un nuevo paradigma en la comprensión de lo huma-no, complementario con un nuevo talante en la relación con los animales, que recoja lo más valioso de los dos paradigmas anteriores, el racional y el emotivo69. Este nuevo paradigma nos permite conjugar el humanismoantropocéntrico con una nueva praxis en relación con los animales, basa-da en el respeto.
5. LA ESPECÍFICA Y SINGULAR ESTRUCTURA ESENCIAL DE LA ESPECIE HUMANA
Como resultado de todo el recorrido que hemos hecho, se desprende de una forma bastante plausible, que el ser humano, a pesar de los pare-cidos inevitables con el resto de los animales, en especial con los que se hallan más cercanos en el proceso evolutivo, representa una especie total-mente singular, con una diferencia no meramente cuantitativa sino cuali-tativa. Lo hemos ido viendo en cada uno de los diferentes apartados de la comparación que hemos ido analizando.
a) En la comparación desde el punto de vista genético, veíamos que los datos de que disponemos parecían favorecer a la postura re-duccionista, si nos limitamos a una mirada meramente naturalista. Las diferencias entre hombres y chimpancés son pocas, sólo un 1% de su genoma. Pero ya veíamos la necesidad de hacer sobre ello varias precisiones: en primer lugar, las investigaciones sobre el genoma nos indican que estamos todavía lejos de haber desen-trañado todos los secretos de su estructura y del modo como pa-recen funcionar los códigos genéticos para expresarse en el ám-bito fenotípico, en la medida en que se halla en cuestión la idea de gen, y se advierte que una gran parte del genoma no se sabe si tiene alguna función y cuál es en concreto; en segundo lugar, cada vez está más clara la insuficiencia del código genético para expli-car la realidad de un ser vivo, cobrando progresiva importancia el trayecto epigenético que une ambos extremos (genotipo y fenoti-po), como nos indican la biología molecular y del desarrollo; ade-más, en tercer lugar, en el ejercicio comparativo de genomas, más significativo que la mera comparación cuantitativa entre genes es la importancia de las consecuencias fenotípicas que producen el
69 Cfr. Gracia. D., «Los animales en la tradición occidental», en Lacadena, J. R. (eds.), Los derechos de los animales, Madrid/Bilbao, UPCO/Desclée de Brouwer, 2002, 134-150.,

447
© Universidad de Deusto - ISBN 978-84-9830-383-4
correspondiente porcentaje diferenciador; en concreto, está claro que ese 1% de singularidad humana es el responsable de las dife-rencias morfológicas, del especial desarrollo embrionario, y, sobre todo, de la específica estructura comportamental del ser humano, y este aspecto es el realmente importante. En definitiva, lo signifi-cativo no es el porcentaje de parecidos y diferencias, sino las con-secuencias tan especiales que esas pequeñas diferencias generan en el fenotipo morfológico, y más aún en el conductual, en la es-pecie humana.
b) Si hacemos referencia al aspecto anatómico-morfológico, volve-mos también a encontrarnos con la dialéctica continuidad-ruptura: los parecidos anatómico-morfológicos son muy amplios, quedando claro que la especie humana está entroncada evolutivamente a la familia de los hominoideos; pero las diferencias son también muy significativas, desde la postura erguida permanente, la utilización inteligente de la mano, y sobre todo la posesión de un cerebro ex-traordinariamente grande y específicamente estructurado que nos dota de especiales capacidades cognitivas y de la posibilidad de utilizar un lenguaje articulado, complementado con la posesión de una laringe adaptada como aparato fonador.
c) En relación al especial modo de desarrollo ontogenético que po-seemos como especie, se advierte que aquí se halla la base de nuestra especial estructura comportamental, en la medida en que nacemos antes de tiempo, como animales prematuros, con el as-pecto del feto de un mono (neotenia) y necesitados de una más prolongada infancia y proceso de aprendizaje, representando esta prematuridad la base biológica de nuestra libertad y la ne-cesidad de poner en marcha nuestras más complejas capacidades cognitivas para dotarnos de cultura y construir un mundo a nues-tra medida. Los defensores de acortar distancias entre hombre y animales siguen aportando supuestas evidencias sobre la existen-cia de culturas animales. Pero volvemos a repetir que tales atisbos de parecido con lo humano es un aspecto más de la tendencia en las especies animales anteriores de poseer aspectos o tendencias embrionarias de lo que después, en la especie humana, serán ca-racterísticas más maduras y complejas, de naturaleza cualitativa-mente diferente.
d) Donde la diferencia es más amplia, explícita e incontestable es en el ámbito de la conducta, de la estructura comportamental. Hemos indicado que la conducta, tanto en el ser humano, y también en la mayoría de los animales superiores, es una síntesis de genética y aprendizaje. Pero la diferencia entre la estructura comportamental

448
© Universidad de Deusto - ISBN 978-84-9830-383-4
humana y animal es abismal: los animales, incluso los más inteli-gentes como los chimpancés y los bonobos, están más sometidos a su herencia genética, y el componente de aprendizaje y de creati-vidad es mucho menor, no advirtiéndose en ellos ningún atisbo de libertad; en cambio, en la especie humana, la herencia genética es más abierta, no está constituida por unas pautas de comportamien-to cerradas, sino que queda en manos de la propia decisión del in-dividuo. De ahí que, como indicaba Gehlen70, entre el estímulo y la respuesta el ser humano tiene que poner, por necesidad, un hia-to, un paréntesis de decisión y de libertad, característica exclusiva de la especie humana.
En definitiva, en el ser humano se da, como ya lo hemos dicho, una síntesis de continuidad y de ruptura evolutiva en relación con el resto de los animales. Y en lo que tiene de ruptura, puede verse el aspecto negati-vo, su deficiencia biológica, pero también el positivo: su extraordinaria capacidad cognitiva y su libertad, que le obligan a decidir en cada mo-mento entre diversas posibilidades de actuación.
El ser humano es, pues, la única especie cuya estructura esencial es una síntesis de biología y cultura. La biología es la condición de posibi-lidad de la cultura, pero no agota ni determina su lógica o racionalidad interna. En el ser humano, la biología se ha abierto y ha sido subsumida por un nivel de realidad nuevo: lo psíquico o espiritual. De tal forma que sin dejar de ser un animal, el ser humano es una estructura esencial nueva: unidad psico-orgánica.
Considero que los planteamientos metafísicos y antropológicos de X. Zubiri siguen siendo uno de los más adecuados para definir la espe-cificidad de lo humano, entendiéndolo como una realidad entroncada y situada dentro del dinamismo total de la realidad71. La realidad es esen-cialmente dinámica, está constitutivamente dando de sí72, configurándose diferentes modos o niveles de dinamismos, desde el dinamismo de la va-riación hasta el de la suidad, con sus tres dimensiones: personal, social e histórica. Todo surge desde la materia, complejificándose sistémicamente
70 Cfr. Gehlen, A., El hombre, Salamanca, Sígueme, 1980; Sequeiros, L., «Arnold Gehlen (1904-1976). La búsqueda de una fundamentación biológica para la antropología filosófica», en Alarcos Martínez, F. J. (ed.)., La moral cristiana como propuesta. Home-naje al profesor Eduardo López Azpitarte, Madrid, San Pablo, 2004, 683-700; Id., «Bio-logía y conducta humana. Reflexiones desde la antropología filosófica de Arnold Gehlen (1904-1976)», texto de discusión con este escrito; Beorlegui, C., Antropología filosófica,o.c., 393-432.
71 Cfr. Zubiri, X., Sobre el hombre, o.c., 72 Cfr. Zubiri, X., Estructura dinámica de la realidad, o.c.,

449
© Universidad de Deusto - ISBN 978-84-9830-383-4
hasta dar de sí la realidad viva, y posteriormente el psiquismo humano. La realidad humana es una conjunto sustantivo de notas, que forma una estructura psico-orgánica única, conformada a su vez por dos subestructu-ras: la psíquica y la orgánica. La subestructura psíquica le hace ser inteli-gente, esto es, su habitud o el modo de habérselas con la realidad consiste en estar abierto a la realidad en cuanto tal. En cuanto realidad psico-orgáni-ca, sus actos son de naturaleza sensible e inteligente al mismo tiempo (in-telección sentiente).
Así, el ser humano siente, como el resto de los demás animales, de tal modo que la realidad se le da en impresión. Pero es un sentir inteligente,en la medida en que su animalidad ha sido elevada a una estructura nue-va: la psíquica, consecuencia de las mutaciones genéticas y la cerebrali-zación. De ahí que el ser humano es el resultado de la emergencia de una nueva estructuración cerebral, que le ha dotado de una mente capaz de es-capar del constreñimiento biológico y genético, para abrirse al nivel de la suidad: la realidad humana es una naturaleza abierta, de una complejidad tal que tiene que hacerse cargo de sí misma, de su propia realidad y cargar libre y responsablemente de ella misma, en diálogo comunitario con las demás realidades humanas.
En definitiva, en función de la específica estructura sustantiva de la realidad humana, posee muchos elementos comunes con los demás ani-males, y advertimos en ellos apuntados y en embrión muchas de las ca-racterísticas que en el ser humano han llegado a su pleno cumplimiento y realización. Por eso que no nos extraña ver en los chimpancés y demás especies de simios, tantas similitudes genéticas, morfológicas e incluso conductuales. Pero tampoco nos debería extrañar que sigamos afirmando al mismo tiempo la distancia cualitativa propia de la condición humana: autoconciencia, libertad y responsabilidad, lenguaje articulado, capacidad de construcción de complejas herramientas, complejo entramado social, capacidad de goce estético, apertura a la pregunta por el fundamento y a la búsqueda del absoluto, etc.
La unidad biocultural del ser humano es una expresión de su unidad psico-orgánica, tal y como la defienden los emergentismos sistémicos,entre cuyos defensores se sitúan M. Bunge, K. Popper, J. Searle, o J. Mon-serrat73, o también el estructurismo dinámico de X. Zubiri y P. Laín En-tralgo74. Este modo de entender las relaciones entre la mente y el cuerpo
73 Cfr. Beorlegui, C., «Los emergentismos sistémicos: un modelo fructífero para el pro-blema mente-cuerpo», Pensamiento, 62 (2006), n.º 234, 391-439.
74 Cfr. Laín Entralgo, P., Cuerpo y alma. Estructura dinámica del cuerpo humano, Ma-drid, Espasa-Calpe, 1991; Id., «El problema alma-cuerpo en el pensamiento actual», en Mora, F. (ed.), El problema cerebro-mente, Madrid, Alianza, 1995, 17-35.

450
© Universidad de Deusto - ISBN 978-84-9830-383-4
permite superar tanto los materialismos o monismos fisicalistas y los dua-lismos, para entender, por el contrario, la realidad humana como una es-tructura o sistema único, resultado de un proceso evolutivo que ha experi-mentado sucesivas complejificaciones, de modo que la mente viene a ser una figura nueva y específica de estructuracion y sistematización del cere-bro y del conjunto del cuerpo humano.
Esta visión dinámica y saltacionista de la realidad está confirmada en todos los niveles de la realidad de nuestro mundo. Pequeños cambios acu-mulados llegan a desencadenar un salto cualitativo, que exige un nuevo ni-vel de realidad, organizado con una nueva estructuración. En estos saltos cualitativos, el nivel inferior es condición de posibilidad del superior, al que subtiende dinámicamente (Zubiri). Pero el nivel superior no se reduce al in-ferior, aunque se apoye en él. De tal forma que la novedad que supone la emergencia del nuevo nivel, es una novedad imprevisible e irreductible. Im-previsible, porque no puede ser prevista desde el nivel inferior, e irreducti-ble, porque las leyes y la lógica con la que está configurado y constituido el nivel emergente no puede ser reducido a las leyes del nivel inferior. Así, la psicología no puede reducirse a la biología, ni ésta a la física.
En consecuencia, esta singularidad y especial densidad ontológica de la realidad humana hace que pueda ser defendida una teoría humanista y antropocéntrica de la realidad humana, que nos permite defender también la centralidad ética del ser humano frente a las teorías que pretenden di-luir la línea divisoria de la humanidad y de la personalidad que separa a hombres y al resto de los animales. Sólo la especie humana está verti-da a la moralidad, porque sólo ella tiene capacidad ética y capacidad de apropiarse de posibilidades para hacerse y realizarse. Eso no quita que las demás realidades, animales, plantas y el resto de la materia, posean su propia densidad ontológica, y, en consecuencia, el ser humano deba comportarse frente a ellos no como un déspota sino como un cuidador y administrador inteligente y responsable. Y bajo este prisma es como tiene sentido la sensibilidad ecológica y proyectos como el Gran Simio y otros semejantes.
En consecuencia, el ser humano puede seguir siendo considerado el centro de la creación, y el único al que se le pueden pedir responsabilida-des, tanto en relación a sí mismo, como también en relación a sus seme-jantes y al resto de la creación. Y es a través de la puesta en marcha de su capacidad de razonar y de llegar a consensos racionales como se podrá establecer cuáles son las obligaciones que nos corresponden en relación al ecosistema y a sus diferentes realidades integrantes, tanto seres vivos como no vivos.
Pensamos, pues, que si los múltiples y complejos avances de las di-ferentes ciencias van representando retos importantes para la compren-

451
© Universidad de Deusto - ISBN 978-84-9830-383-4
sión de nuestro ser como humanos, estos mismos saberes científicos nos aportan suficientes apoyaturas como para seguir defendiendo una visión humanista, y también cristiana, del ser humano. Somos, está claro, una es-pecie más en el conjunto de la biosfera, pero conformada por una singular naturaleza, que nos ha dotado de especial inteligencia, libertad, responsa-bilidad, lenguaje, socialidad, y demás cualidades específicas, con ayuda de las cuales podemos ser conscientes de la acción de Dios que nos ha lla-mado a la existencia y nos ofrece gratuitamente su salvación.

© Universidad de Deusto - ISBN 978-84-9830-383-4

453
© Universidad de Deusto - ISBN 978-84-9830-383-4
Capítulo 8
Entre la biología y la cultura
1. LA EMERGENCIA DE LA CULTURA DESDE LA BIOLOGÍA1
Lo que define la singularidad del ser humano respecto al resto de las especies vivas es, como ya lo hemos subrayado en el capítulo anterior, su cultura, es decir, la superación de los constreñimientos rígidos de la bio-logía, teniendo como consecuencia que decidir en cada momento entre las diferentes alternativas vitales que la realidad le va ofreciendo. Esa na-turaleza abierta supone la posesión de la libertad, la capacidad ética y la consecuente responsabilidad que adquiere ante sus acciones.
En este capítulo vamos a centrarnos en estudiar cómo surge la di-mensión cultural desde el proceso evolutivo de la biología, comparar los dos continentes o dimensiones que componen lo humano, la biología y la cultura, presentar las posturas fundamentales existentes a la hora de entender la unidad bio-cultural que compone lo humano, y concluir con la presentación del modo correcto de conjugar en lo humano lo biológi-co y la ético.
1 Cfr. Ruffié, J., De la biología a la cultura, Barcelona, Muchnick Editores, 1982; Melotti, H., El hombre entre la naturaleza y la historia, Barcelona, Península, 1981; París, Carlos, El animal cultural. Biología y cultura en la realidad humana, Barcelo-na, Crítica, 1994; Christen, Y., El hombre biocultural. De la molécula a la civilización,Madrid, Cátedra, 1989; Ayala, Fr. J., Origen y evolución del hombre, Madrid, Alianza, 1980.

454
© Universidad de Deusto - ISBN 978-84-9830-383-4
1.1. El ser humano, fruto de dos herencias
El atributo específico del ser humano es la cultura, y su condición de posibilidad son la base genética y su capacidad cerebral. Pero esta afir-mación no significa que consideremos lo biológico como mero instru-mento para la aparición de la cultura, para olvidarlo después, sino que entendemos que la aparición de la cultura no suprime la biología, sino que la asume superándola en otra dimensión, desde un nivel superior e integrador nuevo.
Por tanto, el género humano es fruto de dos herencias, la biológica y la cultural. Y de entre las dos, la que ha predominado y orientado la existencia humana es la cultural. Y ahí está lo que le hace original. «La originalidad del hombre, indica Ruffié, no debe buscarse en lo zoológi-co en el sentido tradicional sino en lo psicosocial. Gracias al desarro-llo del cerebro y a la liberación de la mano, la especie humana adqui-rió el alto nivel de conciencia reflexiva que hizo posible la aparición de un medio de comunicación lógica —el lenguaje conceptual— y que permitió la organización de sociedades cada vez más complejas y más logradas»2.
Comparando las dos herencias, advertimos que la biológica, como señala J. Ruffié, es orgánica y endosomática, mientras que la cultural es una herencia superorgánica y exosomática. Es decir, la herencia biológi-ca la recibimos con la dotación cromosómica que conforma nuestra cor-poralidad, mientras que la cultura es una herencia recibida sobre todo, pero no totalmente, desde fuera de nuestra condición corpórea, como fru-to de la adquisición de las generaciones anteriores, a través del lenguaje y del conjunto de las diversas estructuras de educación y de inculturación que poseen las sociedades humanas.
La herencia biológica se comporta en nosotros de forma semejante a como funciona en el resto de las especies animales, esto es, a través de la transmisión de la información genética de la especie, codificada en el ADN, y empaquetada en el conjunto de los cromosomas (23 pares en nues-tra especie) que conforman nuestro cariotipo. Además, la ley que rige la transmisión de esta información genética sigue lo que se denomina la ló-gica darwiniana, expresada en la secuencia ADN-ARN-Proteínas, de tal forma que sólo aparece en el fenotipo lo que existe en el genotipo, aun-que, como sabemos, no toda la información contenida en el genotipo se expresa en el fenotipo. La herencia genética no transmite, pues, rasgos adquiridos durante la vida de un individuo vivo.
2 O.c., p. 278.

455
© Universidad de Deusto - ISBN 978-84-9830-383-4
En cambio, la herencia cultural, exclusiva del ser humano, se basa en la lógica lamarckiana, puesto que transmite caracteres adquiridos en el transcurso de la vida de una persona y de un grupo humano, acumulados de generación en generación, y transmitidos por la enseñanza y el apren-dizaje desarrollados, sobre todo pero no de forma exclusiva, en los prime-ros años de nuestra vida. Esta herencia cultural es autónoma e indepen-diente de la biología en su funcionamiento interno, puesto que sigue sus leyes propias y específicas, como tendremos ocasión de ampliar y precisar más adelante.
Somos conscientes de que cuando hablamos de cultura estamos empleando un concepto polisemántico, que ha tenido muchos proble-mas de definición en el ámbito de la Antropología cultural, de tal modo que se han dado de ella múltiples y muy dispares definiciones sin llegar a un consenso entre los especialistas3. Frente a un concepto de cultu-ra restringido a los frutos superiores e intelectuales del ser humano, la idea de cultura que nos resulta más adecuada para lo que nos ocupa se refiere a todo lo que el ser humano ha tenido que hacer para sobrevivir, como consecuencia de su naturaleza deficiente e inmadura, como vimos en capítulos anteriores. El ser humano posee una naturaleza abierta, de ahí que el salto evolutivo producido en él le obliga a no poder abando-narse en manos de las tendencias o pulsiones biológicas, y tener que inventar las respuestas adecuadas frente al entorno que le puedan ser-vir para sobrevivir y perseguir las metas que se va proponiendo. Esta es la razón por la que se considera que el ser humano no tiene naturaleza, sino que tiene historia (Ortega); o, quizás algo más adecuado a la rea-lidad, que su naturaleza es artificial (artificiosidad natural, Plessner), conjunción y síntesis de biología y cultura, no pudiendo desprenderse de ninguna de las dos.
Cuando hablamos de cultura humana nos estamos refiriendo, en con-secuencia, a todo lo que realiza y produce el ser humano en el ejercicio de existir y de llevar a delante su condición, porque todo lo humano es cultu-ral, fruto de sus decisiones de enfrentarse y transformar la realidad natural:desde la alimentación, vestidos, hábitats, lenguaje, organización social, le-yes, teorías filosóficas, religión, etc.
Pero hoy día se plantea la cuestión de si la cultura es o no un patrimo-nio exclusivo de las especies humanas, o si, por el contrario, no hay que atribuir también, en cierta medida, a algunas especies animales algún gra-do de cultura.
3 Cfr. Kroeber, A., Anthropology, culture patterns and process, Nueva York, Harper Books, 1963; París, C., o.c.; Beorlegui, C., Antropología filosófica, Bilbao, Universidad de Deusto, 2004 (2.ª ed.), pp. 201 y ss.

456
© Universidad de Deusto - ISBN 978-84-9830-383-4
1.2. ¿Cultura animal?
Como hemos ido viendo en los capítulos anteriores, respecto a diver-sos rasgos que caracterizan lo humano, también en el ámbito de la cultura se tendría la pretensión de que no es algo específico de los humanos, sino que también muchas especies animales, sobre todo las que viven en so-ciedades complejas (hormigas, abejas, etc.), y las más cercanas a los hu-manos en el proceso evolutivo, como los grandes simios y en especial los chimpancés, habrían desarrollado también sus respectivas culturas, aun-que posean rasgos muy elementales.
A la hora de dilucidar esta cuestión, tenemos que precisar, como lo hemos hecho en otros momentos, varios aspectos fundamentales. En pri-mer lugar, no es sinónimo la capacidad de generar cultura con la capa-cidad de formar sociedades complejas, puesto que las especies animales organizadas en sociedades complejas conforman su sistema social y de-sarrollan la complejidad de sus roles sociales movidas exclusivamente por sus orientaciones genéticas, sin que se advierta ningún tipo de flexi-bilidad en sus costumbres ni atisbos de conciencia en sus comportamien-tos. Para que se pueda hablar de cultura, en sentido estricto, se necesita poseer una capacidad creativa e innovadora, unida a un nivel de mente cognitiva como para intercambiar mensajes, y la capacidad de transmi-tir exosomáticamente, esto es, de forma distinta a la transmisión genéti-ca, las posibles creaciones e innovaciones que algunos de sus individuos puedan realizar. Esto parece que sólo lo han conseguido algunos chim-pancés en algunas situaciones extraordinarias4. En estos casos, se daría una cierta proto-cultura animal, que nos viene a mostrar de nuevo que en el ámbito de los simios, sobre todo entre los chimpancés, se atisban en embrión muchas de las cualidades y propiedades que caracterizan a la condición humana, y que posee la especie humana en un grado de com-plejidad muy distante del que poseen los chimpancés. De nuevo volve-mos a tener que referirnos a la afirmación de J. Ruffié, cuando nos ad-vierte que el hombre no innova nada, sino que todas las cualidades que adornan su naturaleza o condición están ya presentes o apuntadas en em-brión en las especies anteriores.
De lo que se trata es de advertir tanto la continuidad como la diferen-cia entre las estructuras comportamentales del conjunto de las especies animales y el género humano, tal y como nos muestran las investigaciones
4 Cfr. los datos sobre la diferencia entre hombres y animales en el ámbito de las herra-mientas, del uso del fuego, del lenguaje y demás elementos específicos de la cultura huma-na, a lo largo del capítulo 4.º. Cfr. «Las conductas de los primates», número monográfico de Investigación y Ciencia, 2003, Temas n.º 32.

457
© Universidad de Deusto - ISBN 978-84-9830-383-4
etológicas. Está claro que se da una cierta continuidad entre el conjunto de actividades que las especies animales tienen que realizar en su esfuerzo por sobrevivir en su entorno ambiental o nicho ecológico, y los esfuerzos que los humanos han tenido que hacer en situaciones similares. Cuando se han querido presentar las diferencias, se simplificaba la conducta ani-mal unificándola desde la idea de que todos los animales se comportaban bajo un esquema instintivo, advirtiendo, por otro lado, la ambigüedad del concepto de instinto5.
Pero esas simplificaciones se han ido superando desde hace tiempo. El propio Max Scheler, en su conocido libro El puesto del hombre en el cosmos (1928)6, clasificaba las conductas animales en cuatro estratos progresivamente ordenados: el impulso meramente sentido, el instinto, la memoria asociativa, y la inteligencia. En esa clasificación hacía ver en qué medida los animales no se estancan en una conducta instintiva, sino que la superan hacia un tipo de conducta movida por la costumbre y la memoria asociativa; y, en el caso de los primates, sobre todo los chim-pancés, se descubren actos inteligentes (inteligencia práctica), basados en la capacidad de resolver las dificultades y problemas que aparecen por primera vez a través de un chispazo de inteligencia e improvisación. M. Scheler se apoyaba en las investigaciones etológicas de E. Haeckel con chimpancés en Canarias. Pero desde entonces los estudios de los comportamientos animales han avanzado considerablemente, de tal modo que no se puede negar la enorme complejidad y perfección de los com-portamientos animales.
Al hacer un recorrido desde las plantas a los animales más complejos, pasando por los elementales protozoos, encontramos toda una variada y compleja estrategia de confrontación con el entorno, estrategias que han resultado exitosas para el mantenimiento de las especies desde tiempos ya lejanos7. Si comparamos las diversas estrategias de intercambio entre el animal y su medio con la de los seres humanos, encontramos una evidente similitud y parecido, al mismo tiempo que una clara distancia y diferen-cia. De este modo, podemos afirmar que la técnica humana surge por evo-lución de la animal, innovando y desarrollando enormes potencialidades, y, por tanto, no surgiendo de la nada.
5 Cfr. Heller, Agnes, Instinto, agresividad y carácter, Barcelona, Península, 1980 (2.ª ed.: 1994).
6 Buenos Aires, Losada, 1976, cap. 1.º. 7 Cfr. Griffin, Donald, El pensamiento de los animales, Barcelona, Ariel, 1986; VV.AA.,
«La conducta de los primates», número monográfico de Investigación y Ciencia, 2003, Te-mas n.º 32; París, C., El animal cultural, o.c., parte primera, cap., «Vida y acción sobre el medio. La técnica zoológica», pp. 83 y ss.

458
© Universidad de Deusto - ISBN 978-84-9830-383-4
Tratando de precisar lo que denomina «el paradigma básico de la téc-nica animal», Carlos París considera que ese paradigma posee cuatro ras-gos fundamentales:
1) Se trata, en primer lugar, «de una técnica propia de la especie y, por lo tanto, fundamentalmente uniforme dentro de ella»8. De este modo, como en otras ocasiones lo hemos indicado, se puede estu-diar los rasgos propios de cada especie, bajo la forma de etograma(K. Lorenz), porque por muchos miles y millones de años que so-breviva la especie, siempre se mantendrá el esquema de comporta-miento de forma similar9.
2) La técnica animal es somática, esto es, «utiliza fundamentalmente los recursos corporales (…), su instrumento de acción viene dado por el cuerpo animal»10. Sólo el ser humano utiliza herramientas que complementan la acción del cuerpo, estrategia en la que se le adelantan, de modo muy elemental, algunas de las especies de los simios. C. París hace observar la relación estrecha que hay entre estos dos primeros rasgos: «La unidad entre el primer y el segundo rasgo es evidente, empalmando la doble consideración etológica y morfológica de la especie. Si el cuerpo es el instrumento dirigido por el programa técnico, marcando sus posibilidades y también sus límites, y la corporalidad define a la especie, resulta necesariamen-te que la técnica es unitaria dentro de este esquema»11.
3) La condición corpórea de la técnica animal afecta tanto a los re-cursos activos como también a la teleonomía que se advierte en sus acciones, que están orientadas «a la satisfacción de las ne-cesidades, ligadas a la supervivencia del individuo y de la espe-cie, tal como éstas se plantean en la organización anatómica y fisiológica»12.
4) La transmisión de la técnica se realiza en los animales por vía ge-nética, a través de un programa que se inscribe en los genes. De tal modo que las posibles innovaciones que se puedan dar en algún individuo de la especie, no se transmiten a la generación siguiente. Sólo se producen innovaciones si son consecuencia de mutacio-nes genéticas. Se trata, por tanto, de un programa rígido, que no se
8 Ibídem, p. 98. Las cursivas son del autor.9 Cfr. Lorenz, Konrad, La acción de la naturaleza y el destino del hombre, Madrid,
Alianza, 1989; Lorenz, K./Leyhausen, P., Biología del comportamiento. Raíces instintivas de la agresión, el miedo y la libertad, Madrid, Siglo XXI, 1971 (8.ª ed.: 1991).
10 París, C., o.c., p. 98. 11 Ibídem, p. 98. 12 Ibídem, p. 98. Las cursivas son del autor.

459
© Universidad de Deusto - ISBN 978-84-9830-383-4
puede modificar por las posibles innovaciones de la experiencia. Se impone, por tanto, en el mundo animal lo que se denomina la lógica darwiniana, frente a la lógica lamarckiana que se da en la transmisión de la cultura humana. De todos modos, este cuarto rasgo es el más decisivo, al mismo tiempo que el más problemáti-co, puesto que se advierte, como hemos señalado, que en muchos animales se da un evidente aprendizaje y cambios de conducta, de tal modo que resulta un tanto rígido separar drásticamente lo in-nato y lo aprendido, como bien mostró en su momento K. Lorenz. Ahora bien, aunque en determinados animales se abre lo innato a las posibilidades del aprendizaje, también está claro que aunque su capacidad genética le da posibilidades de aprender, también limita grandemente tales posibilidades, puesto que la apertura que le po-sibilita no es total, sino muy limitada, quedando encerrada dentro del programa genético de su propia especie.
Por tanto, teniendo en cuenta estas apreciaciones, se advierte una pro-gresiva apertura y desbordamiento de la rigidez innata de las pautas con-ductuales a medida que avanzamos en la historia evolutiva, hasta alcanzar el momento máximo de plasticidad en el hombre.
En contraposición al paradigma de la conducta animal, los rasgos del paradigma conductual humano está compuesto por los siguientes rasgos:
1) En el ser humano se da una ruptura de la unidad de la estructu-ra comportamental propio de una especie. El comportamiento de los humanos no es único ni uniforme, sino que se expresa en una gran pluralidad de culturas, de tal modo que algunos antropólogos, como Ericsson13, hablan del fenómeno de subespeciación para re-ferirse a la diversidad de culturas dentro de las poblaciones huma-nas. Es decir, se daría entre los humanos, con esta pluralidad de culturas, un fenómeno similar al de la formación de diversas es-pecies biológicas entre los demás animales. Pero es una similitud más bien lejana y equívoca. Pero, además, como advierte C. París, la diversidad que se da en el ámbito cultural humano «abarca tanto lo sincrónico como lo diacrónico, dibujándose en la especie huma-na el característico progreso técnico»14. Es evidente que la espe-cie humana ha generado una enorme pluralidad de culturas como resultado de la casi infinita capacidad plástica de amoldarse a los diversos ambientes, aunque es más acertado decirlo al revés: por
13 Cfr. Erikson, E.H., «Ontogénie de la ritualisation chez l’homme», en Le comportement rituel chez l’homme et l’ánimal, pp. 139-185 (cita tomada de C. París, o.c., p. 100).
14 París, C., o.c., p. 100.

460
© Universidad de Deusto - ISBN 978-84-9830-383-4
la capacidad de amoldar los ambientes a sus necesidades, siendo también enorme la capacidad de transformación que se da dentro de cada cultura a lo largo de su historia.
2) El segundo rasgo se refiere a la condición de extrasomaticidad de la técnica humana, en la medida en que ha desarrollado una enor-me variedad y complejidad de herramientas que constituyen pro-longaciones de la mano y demás partes de nuestro cuerpo. No en vano vemos que la característica que se utiliza para distinguir la aparición de la primera especie del género humano es la capaci-dad de confeccionar y utilizar herramientas característica del homohabilis. Aunque también vimos en su momento que esta habilidad constituye una frontera un tanto lábil y no muy precisa, puesto que incluso los australopitecos parecen haber demostrado altas posi-bilidades de utilizar herramientas elementales. E incluso, algunos chimpancés. Y también es importante hacer notar, como indica C. París, que la utilización de las herramientas por parte de los hu-manos no suprime su relación con la corporalidad, sino que es en gran medida una prolongación, y no se entiende sin esa referencia a lo corpóreo. Además, «no toda la técnica se vuelca sobre lo exo-somático, sino que de una manera muy característica revierte tam-bién sobre la corporalidad humana y la psique»15, como es el caso de las técnicas corpóreas de meditación del yoga, o la gimnasia y la natación, entre otros muchos ejemplos.
3) En la cultura humana, en tercer lugar, se da una sobredetermina-ción de las necesidades biológicas. Tanto la técnica animal como la humana se orientan hacia la satisfacción de las necesidades de cada especie. Pero, así como las necesidades biológicas de cada especie animal son explicitadas y conformadas por el organismo, el concepto de necesidad humana es mucho más amplio y comple-jo, puesto que abarca desde las necesidades biológicas primarias hasta lo más superfluo, en la medida en que lo considerado por el ser humano como necesidad no está limitado a la mera superviven-cia biológica, sino que amplía artificial y creativamente sus nece-sidades al ámbito ideal y simbólico, de tal modo que se puede de-cir que no hay límite para definirlo. Para ver la complejidad de la idea de necesidad humana, basta tener en cuenta las reflexiones de Ortega sobre su carácter superfluo y de lujo16, como las de Marx sobre la amplitud y diversidad de origen, tanto en el estómago
15 Ibídem, p. 101. 16 Cfr. Ortega y Gasset, J., Meditación de la técnica, Madrid, Espasa-Calpe, 1965.

461
© Universidad de Deusto - ISBN 978-84-9830-383-4
como en la fantasía, así como la capacidad del mercado para ma-nipular psicológicamente la idea de necesidad17. Es evidente que la complejidad del concepto de necesidad depende, en el caso del ser humano, de la idea e imagen que tenga de sí mismo, y, al mis-mo tiempo, de su concepción de la vida, de la idea que tenga de su realización personal y de la felicidad, en función de sus creencias y aspiraciones más hondas.
4) El cuarto rasgo de comparación entre la técnica animal y la huma-na se refiere al modo de transmisión en las que se apoyan ambas. Mientras la técnica animal hemos visto que se transmite genética-mente, endosomáticamente, la técnica humana lo hace exosomáti-camente, a través de la tradición histórica. La técnica humana no se mantiene rígida y repetitiva, como ocurre en el animal, que se limita a explicitar el programa genético con mayor o menor auto-nomía del mismo, la técnica humana posee una amplia plasticidad, una enorme amplitud y capacidad de innovación e inventiva. Y esa capacidad innovadora no depende de saltos mutacionales aleato-rios, sino de la intencionalidad y creatividad de la mente humana. De ahí que el ser humano funcione con ensayos permanentes, con la lógica del ensayo y el error, superando y dejando de lado los errores para quedándose con los aciertos, transmitiéndolos de for-ma acumulativa a las generaciones siguientes, añadiendo paulati-namente a esos aciertos las posteriores innovaciones y creaciones de las generaciones sucesivas. Esto es lo que constituye la condi-ción histórica del ser humano, fruto de la tradición (tradere: trans-mitir), puesto que nace a hombros de las generaciones anteriores y deposita en las siguientes el depósito, con los aumentos correspon-dientes, que le ha transmitido la tradición precedente.
En conclusión, la comparación entre la cultura humana y la culturaanimal nos lleva a la conclusión ya apuntada con antelación: si hay que en-tenderlas como estrechamente relacionadas, puesto que la cultura humana se origina con elementos ya presentes y anticipados en el ámbito animal, también es necesario e inevitable afirmar que la aparición de la cultura hu-mana supone un salto cualitativo y específico en el proceso evolutivo, de modo que rompe totalmente con la lógica del comportamiento y de la téc-nica animal. La condición humana, conformada sobre la deficiencia bioló-gica, y su estructura comportamental, abierta a las decisiones conscientes, libres y responsables de cada uno de los humanos, sitúan al género humano
17 Cfr. Marx, K., El Capital, Madrid, EDAF, 1972, Libro I, cap. I., p. 39 (cita tomada de C. París, o.c., p. 102).

462
© Universidad de Deusto - ISBN 978-84-9830-383-4
en otra dimensión, no pudiéndose limitar simplemente a acomodarse a su entorno, sino que se halla inevitablemente necesitado y obligado a crear li-bre e innovadoramente su entorno mundano, acumularlo y posteriormente transmitirlo a las siguientes generaciones. Y esto es la esencia y estructura de la cultura humana, atisbada es verdad por los balbuceos animales, pero situada en un escalón muy superior de complejidad.
1.3. El salto de la biología a la cultura
La condición cultural del ser humano no nace de modo caprichoso o mágico, sino que es fruto de su especial condición biológica: la defi-ciencia biológica de la que habla A. Gehlen18 y la idea de neotenia sobre la que hemos hablado en el capítulo anterior. El nacimiento prematuro le empuja necesariamente al ser humano a tener que inventar el modo de enfrentarse a la realidad para completar con su inventiva lo que no le ha dado la naturaleza de modo automático e instintivo. De este modo, como indica Portmann, la cultura se convierte en el caso del ser humano en el segundo útero materno, en el auténtico ámbito moldeador de su conducta, tanto frente a la naturaleza como frente al conjunto de sus congéneres, ad-virtiéndose en esa matriz conformadora de lo humano el aspecto biológi-co y la influencia del ambiente, siendo ambos elementos, unidos sistémi-camente, los ingredientes inevitables de la conducta humana.
De este modo, se ve que el hombre es, a la vez, productor de la cul-tura y producto y fruto de la misma. Nacemos en un entorno cultural que nos educa y conforma, ayudándonos incluso a desarrollar nuestra propia autoconciencia y personalidad, y nos capacita también para transformar nuestro entorno cultural interactuando con él creativamente, pudiendo añadir al mismo tiempo nuestras aportaciones al conjunto de la herencia recibida.
Y con el nacimiento de la cultura, esto es, con el nacimiento del géne-ro humano se rompe e interrumpe la dinámica y el ritmo que la biología había mantenido hasta entonces, pudiendo decirse que con la aparición de lo humano se ha trastocado el ritmo y el protagonismo de la selección na-tural. Podemos decir que, de algún modo, la selección natural ya no actúa, o lo hace de modo más lento y de forma diferente a la aparición del ser humano. Al menos es cierto en el ámbito de lo humano. Esto se entiende viendo que en el ser humano ya no se da una estrategia comportamental especializadora, puesto que ante una necesidad no se espera que la selec-
18 Cfr. Gehlen, A., El hombre, o.c.; Beorlegui, C., Antropología filosófica, o.c., pp. 393-432.

463
© Universidad de Deusto - ISBN 978-84-9830-383-4
ción, a través de los saltos mutacionales aleatorios, aporte una novedad, una estrategia instintiva nueva frente al ambiente, sino que es la inteligen-cia la que se le adelanta previendo y creando novedades e inventivas te-leológicamente conformadas. De este modo, la lógica de la cultura escapa a la lógica darwiniana, adoptando la lógica lamarckiana.
Y con ello, también la cultura escapa a la mera función adaptativa que persigue el mundo de la biología. No se limita a solucionar proble-mas inmediatos, sino que imagina y soluciona también problemas fu-turos. Y cuando no puede solucionar determinados problemas, inventa explicaciones y construye soluciones teóricas. Es así como surgen las teorías filosóficas, los mitos y las religiones. De este modo, vemos que la lógica de la cultura escapa a las reglas de la selección natural, inventando sus propias reglas, y construye las nuevas estrategias y orientaciones que dirigirán su comportamiento (la moral), capacidad que es fruto y conse-cuencia de no tener prefijadas por herencia biológica las reglas rígidas de su comportamiento, sino que está obligado necesariamente a elegir entre diversas opciones y caminos. Esta capacidad es parte de su condición na-tural, que tiene que completar con la creación de normas culturales para organizar la convivencia y sobrevivir. Por tanto, la filosofía, la moral y la religión son rasgos específicos de lo humano, como ingredientes supe-riores de nuestra condición cultural.
El paso de lo biológico a lo cultural está marcado por el paso de lo in-nato a lo adquirido, dinámica que va progresivamente aumentando y com-plejificándose hasta llegar al género humano. La biología evolutiva divide a los diferentes grupos de seres vivos en especies, caracterizadas cada una de ellas por su diferente fenotipo así como su distinto modo de compor-tarse. Pero cuando se llega a lo humano, los estudios sobre los hombres primitivos, la paleoantropología y la prehistoria, organizan y dividen a los diferentes grupos humanos en etnias, clasificadas en función de las dife-rentes industrias y artefactos con los que organizan su vida. Las divisiones y avances producidas entre los humanos no son de tipo genético-biológi-co, nacidas de habilidades biológicas, sino de avances técnicos y cultu-rales. Esto nos hace ver que en el género homo ya no hay casi evolución biológica sino psicosocial.
En un principio, las culturas humanas tendieron a adaptar mejor a los humanos a su ambiente, pero después se orientaron más a transformar el entorno para adaptarlo mejor a los humanos. Esa orientación, debido a la progresiva y casi ilimitada capacidad tecnológica de transformar el am-biente, nos ha llevado a la actual crisis ecológica, en la que los seres hu-manos podríamos situarnos en una fase de no retorno respecto a la capaci-dad de degradar las condiciones de vida de nuestra biosfera, de modo que corremos el peligro de destruirla y desaparecer nosotros con ella.

464
© Universidad de Deusto - ISBN 978-84-9830-383-4
Por otro lado, si en un principio se dieron las diversas salidas de Áfri-ca y la dispersión demográfica, con lo que supuso la diversidad cultural progresiva, en la actualidad, en la medida en que hemos poblado toda la superficie del planeta, se está produciendo el fenómeno contrario: la tendencia a la unificación y uniformización cultural, no tanto como con-secuencia de un diálogo convergente entre las culturas actuales, sino a través de una imposición hegemónica de la cultura occidental, o mejor, norteamericana, por medio de su dominancia económica, militar y mas-mediática. Pero, de este modo, caminamos hacia un empobrecimiento progresivo por desaparición de muchas de las culturas actuales (con su bagaje de valores, idiomas, etc.), fenómeno de similares características al empobrecimiento que supone la desaparición progresiva y galopante de miles de especies vivas, tanto vegetales como animales.
Las consecuencias del relevo de la evolución biológica por la cultural son enormes y trascendentales, teniendo en cuenta las características de cada una de ellas.
— Ambas difieren en su naturaleza:
a) La genética está basada en la mutación, fruto del azar y guiada por la necesidad. El precio y la consecuencia de ello es el gasto inmenso que conlleva, puesto que la mayoría de las mutaciones no son viables y, por eso, suelen ser desechadas por la natura-leza, de modo que sus éxitos son muy pequeños, en compara-ción con los múltiples intentos fracasados. Además, sólo aporta soluciones aproximativas; es decir, la adaptación biológica es aceptable, pero siempre imperfecta. De ahí que Fr. Jacob, en su Lógica de lo viviente19, afirme que la evolución funciona de forma chapucera, como el bricolage de un aficionado (bricola-ge genético).
b) En cambio, la lógica de lo cultural es fruto de voluntades cons-cientes y deliberadas, y, por ello, consigue soluciones más per-fectas y ajustadas, aunque no siempre de forma completa. No interviene, por tanto, en el ámbito de lo cultural el azar ni lo aleatorio, al menos en el acto creador de los frutos culturales. De este modo, la especie humana ha conseguido convertirse en la especie más adaptada y adaptable de todas las que existen, superando y evitando los errores y azares de la evolución bio-lógica.
19 Cfr. Jacob, François, La lógica de lo viviente. Una historia de la herencia, Barcelo-na, Laia, 1977.

465
© Universidad de Deusto - ISBN 978-84-9830-383-4
— Además, difieren en el ritmo de sus procesos de avance:
a) La herencia genética, basada en las mutaciones azarosas, consi-gue avances esporádicos, necesitando también para difundirlas el transcurrir de muchas generaciones, a través de múltiples cru-zamientos genéticos.
b) En cambio, la herencia cultural está basada en la inventiva, y sus innovaciones pueden ser comunicadas y transmitidas a una canti-dad ilimitada de individuos, y a una velocidad cada vez más gran-de, debido a los actuales medios de comunicación y a los medios cada vez más rápido de transporte y transmisión de realidades culturales por la superficie de la tierra. De estar en un planeta en el que las distancias se le antojaban al ser humano inabarcables, nos hallamos en la actualidad con una experiencia de los espacios y de los tiempos propios de generaciones que se experimentan como pobladores de una aldea global. De este modo, los frutos de la evolución cultural son muy superiores a los biológicos, en su capacidad adaptativa. Los seres humanos podemos volar, su-mergirnos en los océanos, ir en barco, etc., sin necesidad de que nos hayan salido por mutación alas, branquias, aletas, u otras ca-racterísticas biológicas. Y esto se ha conseguido en el transcurso de muy pocos años, si nos situamos en la escala evolutiva.
— Diferente consistencia y fragilidad. Ahora bien, las ventajas que, desde los puntos de vista anteriores,
advertimos en la cultura respecto a la biología, se invierten en rela-ción a esta característica.
a) Mientras que, en el ámbito de la genética, los avances se fijan en el genoma de modo podríamos decir casi definitivo, y se mantienen ahí hasta que no se produzcan otras mutaciones que las mejoren o las destruyan (aunque la lógica de la selección na-tural evita las mutaciones deletéreas y negativas),
b) los avances culturales son enormemente frágiles y perecederos. Todos los avances, que han sido conseguidos con gran esfuerzo y dedicación, pueden perderse de modo relativamente fácil, de-bido a conflictos, guerras y errores de los humanos, en tan sólo el paso de una generación a otra, pudiéndose detener de este modo el avance de la civilización y retroceder a fases del hom-bre primitivo.
— Diferencias en cuanto a su rigidez y libertad. Pero la contrapartida de lo anterior está en este nuevo punto o rasgo
de ambas.

466
© Universidad de Deusto - ISBN 978-84-9830-383-4
a) Mientras la herencia genética es más estable, también está encerra-do en ello una mayor rigidez, de tal modo que su estabilidad cons-tituye al mismo tiempo una prisión rígida. Es decir, una sociedad de abejas es más perfecta, hasta cierto punto, en la estructuración de los roles y tareas y en la perfección de los panales de miel que construye, que una sociedad de humanos. Pero no se da en ese tipo de sociedad ningún progreso: las mismas especies siempre se organizan socialmente y construyen sus panales o sus habi-táculos de la misma forma, desde que apareció esa especie hasta la actualidad.
b) En cambio, en la sociedad de los humanos pueden darse errores e imperfecciones, pero somos capaces de detectarlos y tenemos capacidad de superarlos, aunque también de destruirlo todo por nuestras diferencias, disputas y envidias. Somos capaces de lo mejor y de lo peor, debido a nuestra libertad y la capacidad y necesidad de elegir en cada momento lo que creemos más ade-cuado a nuestras necesidades.
En conclusión, la seguridad de lo biológico-genético va en detri-mento de su flexibilidad y libertad. Pero la flexibilidad y libertad de las creaciones de lo cultural va también en detrimento de su seguridad y es-tabilidad.
Todo esto nos muestra que el género humano nace sin poseer una es-tructura comportamental cerrada genéticamente, y por ello resulta prodi-giosamente abierta, maleable y educable. Al nacer, como ya lo vimos en los capítulos anteriores, no posee nuestra especie comportamientos inna-tos, sino que es un animal deficiente (Gehlen) y menesteroso. Así que el entorno cultural tiene que suplir las carencias de la biología, y esa suplen-cia es totalmente ventajosa. Los humanos recién nacidos tienen que apren-derlo casi todo, y ese mismo trabajo educativo lo tiene que aprender cada generación y cada niño recién nacido.
La cultura adquirida moldea nuestra personalidad, de modo que so-mos tanto o más fruto y resultado de la educación que de la herencia biológica. Por tanto, no nacemos ya siendo lo que somos por natura-leza biológica, sino que aprendemos a ser y vamos construyendo nues-tro futuro en un entorno cultural que nos moldea progresivamente, hasta que somos capaces de tomar definitivamente las riendas de nuestra pro-pia formación.
Es cierto que ninguna cultura es tan perdurable y consistente como la estructura del ADN, pero la personalidad del ser humano no es nada sin la cultura. Es ella la que define nuestra identidad. El genoma sólo nos aporta un conjunto de predisposiciones, que no son determinismos ni es-

467
© Universidad de Deusto - ISBN 978-84-9830-383-4
tructuras rígidas, y sobre ellas nuestra cultura y nuestras decisiones per-sonales actuarán después. La grandeza de la evolución cultural se halla, por tanto, en el nivel de conciencia y de libertad que implica. El hombre sabe y es consciente de los objetivos que persigue. Eso significa al mismo tiempo que el ser humano es responsable de sus actos; es él quien orienta su futuro y tiene que elegir y asegurar su destino.
En la actualidad, esa responsabilidad es ineludible y de enormes con-secuencias, puesto que tenemos ya en nuestras manos los medios capaces de destruir especies enteras, e incluso a la propia especie humana, en la medida en que poseemos también los medios para degradar gravemente el medio ambiente y destruir el globo terráqueo. Es una situación nueva e inesperada, que no había existido nunca en la historia de la humanidad. La eventualidad del suicidio colectivo, de toda la especie, puede ser des-graciadamente el último avatar de la selección cultural. De hecho, algunas culturas del pasado se han autodestruido, así como muchas especies ani-males han desaparecido como consecuencia de su propio éxito y dominio de su entorno, como es el caso de los dinosaurios.
Esta capacidad que tiene la especie humana de hallarse más allá de los determinismos biológicos, y, por ello, dotada de la capacidad y la necesi-dad de tener que elegir entre diferentes posibilidades de acción, es lo que le convierte en animal cultural y ético, con todas las consecuencias y res-ponsabilidades a las que hemos aludido y de las que cada vez somos más conscientes.
2. LA CONJUGACIÓN DE LA CULTURA CON LA BIOLOGÍA
Esta tesis que estamos defendiendo, consistente en atribuir la cultu-ra en exclusividad a la especie humana, ya hemos dicho que no es acep-tada por todos los estudiosos, apelando a una cierta y supuesta cultura animal. Pero la postura que considera que el ser humano es el único que posee cultura, en sentido estricto, es la que se halla más evidenciada por los datos y las aportaciones más serias de los antropólogos y estudiosos de los comportamientos animales y humanos. De ahí que podamos afir-mar que el ser humano es una animal bio-cultural, síntesis de biología y cultura.
Pero a la hora de estudiar la relación entre ambas dimensiones, nos hallamos ante una pluralidad de planteamientos. Nuestra postura se sitúa entre dos extremos: el reduccionismo bilogista y el culturalismo dualista.El primero aspira a reducir todos los fenómenos culturales a términos y leyes biológicas, reduciendo lo cultural casi a un tipo de comportamiento

468
© Universidad de Deusto - ISBN 978-84-9830-383-4
biológico más complejo. En cambio, para el culturalismo dualista la cul-tura sería una realidad que surge al margen de lo biológico, en unos casos, y en otros, como mucho, desde lo biológico, pero una vez surgida parece que se desentiende de su base biológica y tiende a explicarse como una realidad autónoma y autosuficiente, considerando a la biología como una mera escalera que ha sido necesaria para acceder al nivel superior, pero que, una vez logrado el acceso, se echa abajo y se menosprecia como algo ya inservible y casi inútil.
La postura que consideramos más acertada se sitúa en medio de am-bos extremos, entendiendo que el hombre es un animal bio-cultural, con-formado por una estructura de notas, síntesis de elementos biológicos y culturales, necesitando de ambas dimensiones o vertientes para entender a fondo y en su justas dimensiones la realidad de lo humano. Por tanto, si es insuficiente reducir lo humano a mera biología, también es equivo-cado entenderlo como la suma de dos cosas extrañas, que viven ambas de espaldas a la otra, situando la esencia de lo humano en la dimensión racional, al margen de su base biológica. Por tanto, en su condición de animal bio-cultural, el ser humano no es la suma de dos realidades auto-suficientes, que conviven del modo más decoroso posible, pero dándose la espalda. Somos una unidad psico-orgánica, una única estructura com-puesta por dos subestructuras (somática y psíquica), y dotados de un tipo de actos (nuestra habitud, Zubiri) que son al mismo tiempo sentientes e inteligentes (intelección sentiente, Zubiri), aunque con diferente tipo de dominancia: unos poseen dominancia somática (digerir, respirar, etc.) y otros, dominancia psíquica (pensar, hablar, decidir, etc.).
Vamos a centrarnos en este apartado en presentar las cuatro diferentes posturas que entendemos se dan a la hora de conjugar en la especie huma-na lo biológico y lo cultural:
— postura reduccionista o biologista; — postura subordinacionista o funcionalista; — postura culturalista extrema o dualista; — y postura estructurista o integradora, o culturalismo concordante.
Las dos primeras se inclinan por situar en la biología el centro de gravedad y la importancia básica a la hora de conjugar biología y cultu-ra, aunque discrepen fuertemente en el modo como la biología domina o dirige a la cultura. Las dos posturas últimas consideran que lo cultural es el centro de gravedad de dicha relación, pero discrepan a la hora de diseñar el tipo de conjugación o relación entre ambas dimensiones. Va-mos a ir viendo cada una de las cuatro posturas, ejemplificándolas en algunos autores representativos o en corrientes antropológicas más sig-nificativas.

469
© Universidad de Deusto - ISBN 978-84-9830-383-4
2.1. El reduccionismo biológico
Esta postura sostiene que entre lo biológico y lo cultural no hay rup-tura, porque no hay distinción cualitativa entre ambas, ya que lo cultural vendría a ser un aspecto más de lo biológico. Es decir, para esta postura no hay propiamente conflicto entre lo biológico y lo cultural, en la medi-da en que lo que denominamos cultura, como un rasgo específico de lo humano, sería simplemente un conjunto de rasgos biológicos diferentes, además de entender que no se daría, por tanto, ruptura entre lo cultural en lo humano y otras especies animales situadas en escalones anteriores y cercanos en el proceso evolutivo. En consecuencia, lo mismo que a deter-minadas especies animales les han surgido alas, o pezuñas, u otras carac-terísticas o habilidades específicas que les han capacitado para sobrevivir en el proceso selectivo, al ser humano le ha surgido un cerebro especial-mente voluminoso, que le ha dotado de específicos modos de comportar-se, que han supuesto también unas ventajas significativas en la lucha por la vida. Pero no se vería en ello ninguna diferencia cualitativa respecto al resto de los animales, y digna de ser tenida en cuenta.
De todos modos, hay que distinguir dos tipos diferentes de reduccio-nismos o biologismos: el extremo y el moderado.
a) Biologismo extremo, o naturalismo biológico
En este apartado se sitúan quienes consideran que las leyes de la bio-logía en exclusividad son las que rigen el resto de las realidades huma-nas: lo psicológico, sociológico, ético, etc. Eso supone negar la libertad humana y caer en la denominada falacia naturalista, al confundir el ser,lo que hay, con el deber ser.
Aquí habría que situar, si se quieren tomar en serio sus afirmaciones, los diversos determinismos biológicos, el evolucionismo clásico y, en par-te, la primera sociobiología. Si al ser humano le negamos la libertad, se reduce a ser una especie animal más, dotada de mayores capacidades de interactuar con el entorno ambiental y con el resto de compañeros de es-pecie, pero sus actos son mero reflejo de las estrategias de supervivencia presentes en su genoma.
De ahí que sus planteamientos éticos, consecuentes con esta forma de pensar, no escapan a la falacia naturalista, en la medida en que se tra-taría de deducir las orientaciones morales de las propias leyes biológicas presentes en el proceso evolutivo. De tal modo que los criterios que los autores situados en esta postura proponen para lo bueno y lo malo se tie-nen que extraer o equiparar a los que parece que han orientado a los di-

470
© Universidad de Deusto - ISBN 978-84-9830-383-4
versos seres vivos para conseguir o no el éxito correspondiente en la lu-cha por la supervivencia, según los cánones de la selección natural. Así lo defienden autores como H, Spencer, J. S. Huxley y C. H. Wadington. Para ellos, el progreso de la humanidad tiene que orientarse por los mis-mos criterios que se dan en la lucha por la vida, supuestamente presentes en el proceso evolutivo de la naturaleza biológica20. El problema y la di-ficultad mayor para aceptar estos planteamientos consiste, además de no tener argumentos convincentes para negar la libertad humana, en que es-tos autores parece no ser conscientes de que en el ámbito de la naturaleza no hay más que meros hechos, y no valores éticos, por lo que no acaban de darse cuenta de que caen en un razonamiento circular inaceptable, al pretender ver valores en el ámbito de las leyes naturales, valores que es-tarían supuestamente en el proceso evolutivo porque ellos previamente los han puesto ahí.
b) Biologismo o naturalismo moderado
La sociobiología de E. O. Wilson21 es una teoría compleja y sufi-cientemente ambigua que no hace fácil dilucidar dónde situarla entre las diversas posturas que tratan de conjugar la biología y la cultura en el ámbito de lo humano. Si la ley última que, según los sociobiólogos, rige los comportamientos de todos los seres vivos, incluida la especie huma-na, es la ley del gen egoísta22, se tiene que dar por hecho que no hay po-sibilidades de aceptar la libertad humana, base imprescindible para que podamos hablar de cultura humana como ámbito que supera las leyes biológicas.
A pesar de ello, Wilson no renuncia a hablar de cultura ni de ética, cosa que resulta a todas luces contradictorio, si se quiere ser consecuente con sus tesis centrales y básicas. Tampoco quiere caer en la falacia natu-ralista cuando plantea su teoría ética. Considera, por un lado, que la cul-tura tiene suficiente autonomía respecto a la genética, pero, por otro lado, entiende que está tan atada a las leyes genéticas que no tiene más remedio que seguir las leyes de eficacia genética constituida por lo que denominan el equilibrio evolutivamente estable (EEE). Para ellos, la relación entre la genética y los comportamientos humanos es algo similar a la relación existente entre una boya flotando en el mar y la argolla a la que está aga-
20 Cfr. Ayala, Francisco, Origen y evolución del hombre, o.c., pp. 178-182. 21 Cfr. Wilson, E.O., Sociobiología. La nueva síntesis, Barcelona, Omega, 1980; Id.,
Sobre la naturaleza humana, México, FCE, 1983; Ruse, M., Sociobiología, o.c. 22 Cfr. Dawkins, R., El gen egoísta, o.c.

471
© Universidad de Deusto - ISBN 978-84-9830-383-4
rrada, a través de una cadena, en el fondo del agua. La supuesta libertad del ser humano es, para esta forma de pensar, la ilusión de movilidad que tiene la boya, pero en realidad sus movimientos están orientados y limita-dos por la longitud de la cadena.
En la segunda etapa del desarrollo de la sociobiología, Wilson ha ido evolucionando en sus planteamientos, de la mano de E. Lumsden, tratan-do de conjugar la evolución de los genes y de las unidades culturales, de-nominadas por R. Dawkins memes23, dentro de un proceso que denomi-nan coevolución, siguiendo los procesos de adaptación marcados por lo que denominan las leyes epigenéticas24.
Para Wilson y Lumsden, se da, por tanto, un proceso de coevoluciónentre los genes y los comportamientos culturales, de tal forma que, aun-que se trata de procesos autónomos, se hallan íntimamente relacionados y, en último término, limitados y orientados por las reglas de los genes, de modo que la práctica totalidad de las orientaciones del comportamien-to humano son en gran parte explicables a partir de un determinismo ge-nético. No llegan a defender que los humanos seamos autómatas cuyo comportamiento marcha al compás de nuestros genes, pero entienden que nuestras acciones están mucho más dirigidas por el genoma de lo que nos gustaría admitir.
La dominancia de nuestros genes se da tanto en el nivel biológico como en el cultural. Si nos detenemos en terreno de la visión, se puede advertir en qué medida los genes hacen que los moldes o categorías de nuestra percepción visual sean universales (existen cuatro colores básicos: rojo, amarillo, verde y azul, siendo el resto mezclas posteriores de ellos), consecuencia de los cuatro tipos de receptores cromáticos de nuestra reti-na. De igual modo, consideran que están marcadas por las leyes genéticas costumbres culturales como la prohibición del incesto, o las fobias ante los peligros que amenazaron a los humanos en un estadio más primitivo (fobia a las serpientes, a las ratas, etc.). De la misma manera explican el dominio de la hembra por el macho, la codicia, el altruismo y otros com-portamientos culturales. En la medida en que los etólogos nos muestran que estos comportamientos ya se hallan presentes en las especies animales cercanas a nosotros, aunque expresados de un modo diferente, deducen de
23 Cfr. ibídem. 24 Cfr. Lumsden, E./Wilson, E. O., Genes, Mind and Culture. The Coevolutionary
Process, Cambridge, Mass., Harvard University Press, 1981; El fuego de Prometeo,México, FCE, 1985; París, C., El animal cultural, o.c., cap. 2.º; Id, «La sociobiología y el concepto de cultura», Theoría, Segunda Época, VII (1992), n.º 16-17-18, tomo B, pp. 769-787; Beorlegui, C., o.c., pp. 189 y ss.; Id., «Las Leyes Epigenéticas: ¿el eslabón perdido entre la biología y la cultura?», Zientziartekoak (San Sebastián), II (1987), n.º 2, 102-123.

472
© Universidad de Deusto - ISBN 978-84-9830-383-4
ello que todos esos comportamientos estarían, por tanto, genéticamente esbozados en el repertorio del comportamiento humano, cuando no inclu-so programados en todos los detalles, en la estructura comportamental de la especie humana.
Para explicar y demostrar sus tesis, Wilson y Lumsden proponen lo que denominan el circuito coevolutivo, compuesto de cuatro momentos: molecular, celular, organísmico y poblacional o social25. Así, las leyes del ámbito genético-molecular influyen en el ámbito celular y organísmico, que a su vez orientarán los comportamientos culturales. La relación entre los comportamientos culturales y la base genética a la que están fijados se halla regida por lo que denominan las leyes epigenéticas, que permiten procesos abiertos, pero no entendiéndolos como resultados de decisiones libres, sino más bien como consecuencia de la imprevisión de leyes pro-babilísticas.
El proceso evolutivo ha ido creando especies animales con una ca-pacidad de movimientos y de autonomía cada vez más amplia, siendo la especie humana la máxima expresión de esta movilidad, pero, aun así, en-tienden que eso no debe ser motivo para que podamos hablar de libertad en el sentido absoluto y exagerado de los filósofos humanistas26.
En el ámbito ético, Wilson asegura que no quiere caer en la falacia naturalista, puesto que entiende que el deber ser no se puede deducir de las leyes de la genética. Pero, en realidad, Wilson se mueve entre la ambigüedad y la contradicción a la hora de conjugar sus planteamien-tos cercanos al determinismo y sus pretensiones de seguir defendiendo postulados éticos27. Hablamos de ambigüedad y contradicción porque resulta imposible defender al mismo tiempo planteamientos éticos y el determinismo.
Por otro lado, cuando explicita sus ideas sobre la ética, las entiende como rasgos emanados del proceso evolutivo y al servicio de la mera
25 Cfr. Lewin, Roger, Evolución humana, Barcelona, Salvat, 1986, cap. 30.º, pp. 198-203; Ruse, Michael, Tomándose a Darwin en serio, o.c., cap. 4.º.
26 Para una crítica de los planteamientos sociobiológicos, cfr. Lewontin, R. C./Rose, S./Kamin, L. J., No está en los genes. Racismo, genética e ideología, Barcelona, Crítica, 1987; VV.AA., La biología como arma social, Madrid, Edit. Alhambra, 1982; París, C., El animal cultural, o.c.; cap. 2.º; Id, «La sociobiología y el concepto de cultura», o.c.;Beorlegui, C., «El reto de la biología a la antropología. De la etología a la sociobiología», Letras de Deusto, 16 (1986), n.º 34, pp. 37-69; Id., Antropología filosófica. Nosotros: ur-dimbre solidaria y responsable, o.c., pp. 189 y ss.; Id., «La manipulación de la biología. Función ideológica del determinismo biológico», Zientziartekoak (San Sebastián), I (1986),n.º 2, pp. 71-107; Id., «Las Leyes Epigenéticas: ¿el eslabón perdido entre la biología y la cultura?», o.c.
27 Cfr. Wilson, E.O., Sobre la naturaleza humana, o.c.

473
© Universidad de Deusto - ISBN 978-84-9830-383-4
supervivencia de la especie. Para Wilson, «la moralidad no tiene otra función última demostrable»28. Como bien indica Ayala, esta afirma-ción puede entenderse en el doble sentido que tiene la palabra morali-dad: como capacidad moral o como contenidos morales. Y en ninguno de los dos sentidos del término parece coherente la postura de Wilson. Si se entiende como capacidad ética, hay que decir que el ser humano es ético por naturaleza y no para ser más eficaz de cara a la superviven-cia. Y si se entiende en el sentido de los contenidos morales, Wilson caería en la falacia naturalista, en contra de sus pretensiones explíci-tas29. Por lo tanto, las propuestas sociobiológicas están llenas de erro-res y ambigüedades, por su empeño de eliminar la diferencia cualitativa entre la especie humana y la de los demás animales, diferencia situada en la libertad, su capacidad ética y los diversos elementos que compo-nen su cultura.
Siguiendo los planteamientos de Wilson y Lumsden sobre la coevolu-ción genes-cultura, ha ido apareciendo una interpretación de la cultura en clave evolucionista basada en la teoría de los memes, como unidades de composición y evolución de la cultura30, similar a los genes, y acuñado por R. Dawkins en su célebre y ya citado libro El gen egoísta, y utilizado de forma más amplia y sistemática por Daniel Dennett, Susan Blackmore y Richard Brodie para conformar una interpretación de la cultura basada en los memes y denominada memética31. Se trataría de entender el mundo de la cultura humana como un ámbito dependiente de la genética, nacido y desarrollado en coevolución con el mundo de la genética, y conformado por los memes, que se comportan de modo similar y siguen las mismas leyes que los genes en el ámbito de la genética. Pero, aunque la interpreta-ción de la cultura humana en clave memética puede resultar interesante en algunos aspectos, la supuesta similitud entre genes y memes hace agua por muchos sitios, desde la propia definición de meme, harto ambigua y poco convincente, como unidad cultural, hasta sus comportamientos, pasando por la supuesta comprensión de los memes como unidades autónomas de comportamiento.
La limitación más palmaria de esta teoría es la supresión de la libertad humana, suplida por el dominio de los memes que para Dawkins, Dennett y demás teóricos de la memética serían los autores de nuestras acciones.
28 Wilson, E. O., o.c., p. 167 (edición original). 29 Cfr. Ayala, Fr., o.c., pp. 183-184. 30 Cfr. Aunger, Robert (ed.), Darwinizing Culture: The Status of Memetics as a Science,
Oxford, Oxford Univsersity Press, 2000 (prólogo de D. Denett).31 Cfr. Buskes, Christian, La herencia de Darwin. La evolución en nuestra visión del
mundo, Barcelona, Herder, 2009, cap. 9.º, «La evolución de la cultura», pp. 247-272.

474
© Universidad de Deusto - ISBN 978-84-9830-383-4
Pero, como indica Chr. Buskes, «no somos esclavos obedientes de nues-tros memes, como tampoco lo somos de nuestros genes. Los memes no pueden sobrevivir solos, para eso necesitan de nuestra fuerza mental. Por ello, el determinismo memético es tan improbable como el genético. La evolución nos ha condenado a la libertad, por lo que continuamente he-mos de hacer elecciones y somos considerados responsables de nuestras acciones. Alguien que haga responsable de su conducta a sus memes no puede ser tomado en serio, como tampoco lo será alguien que le eche la culpa a sus genes»32.
Pero, además, la supuesta similitud del comportamiento de genes y memes no se sostiene, porque la lógica de los genes es darwiniana, como ya vimos, y la de los memes, lamarckiana. Y eso es lo que hace que los genes sólo se pueden transmitir a los parientes genéticos, mientras que los memes, si queremos seguir utilizando estas supuestas unidades culturales, se transmiten a todo tipo de componentes de nuestra especie. Además, en la base de las insuficiencias de la memética está la dificultad de definirlos como unidades claras y concretas, razón por la que sus defensores no coin-ciden a la hora de definirlos. Así, mientras unos los consideran como artefac-tos (unidades culturales) que se pueden recordar fácilmente, otros los con-sideran como estructuras neurales de nuestro cerebro, o instrucciones para nuestra conducta33.
Lo que resulta evidente es que el ser humano es un animal dotado de una naturaleza abierta (Zubiri), obligado por ello a hacerse y a realizarse, a construir un mundo artificial a su medida, porque la naturaleza no le ha dado las instrucciones cerradas e instintivas para desenvolverse en medio de la naturaleza. El resultado de esos esfuerzos por sobrevivir y confor-mar un mundo a la medida de las necesidades humanas es lo que deno-minamos cultura, dotación exclusiva, en su sentido estricto, de nuestra especie, consecuencia de nuestra condición de animales inmaduros bioló-gicamente y dotados de libertad y de responsabilidad.
2.2. El biologismo funcionalista o subordinacionista
Aquí se situarían planteamientos como los de Lorenz y Gehlen. El etólogo K. Lorenz, la cultura humana es autónoma y cualitativamente distinta de la biología, dando por hecho que la cultura es resultado de la libertad humana. Pero, a la hora de conjugar lo biológico y lo cul-
32 Buskes, Chr., o.c., p. 263. 33 Para una crítica completa y detenida de la teoría de los memes, cfr. Buskes, Chr., o.c.,
pp. 262-272.

475
© Universidad de Deusto - ISBN 978-84-9830-383-4
tural, entiende que la cultura tiene que aprender de la biología. Frente a quienes consideran que en el mundo animal impera la agresividad y la violencia sin medida, Lorenz defiende la necesidad y bondad de la agresión intraespecífica, porque está controlada por mecanismos de inhi-bición que evitan que una violencia descontrolada lleve a la destrucción de la especie34. En cambio, donde la agresión se halla en peligro de romper todos los moldes es en el ámbito de lo humano, en la medida en que en nuestra especie la cultura ha roto y desbordado los mecanismos inteligentes de la naturaleza, y la aparición de la inteligencia y libertad han dotado a los seres humanos de la capacidad técnica de construir y utilizar mecanismos de agresión que ya no sabemos controlar35. De ahí que, si no queremos perecer víctimas de nuestras desmesuradas capa-cidades, tenemos que ser humildes y aprender de los animales, porque, siguiendo la sabiduría del refrán chino, «aunque no todo el hombre está en el animal, todo el animal está en el hombre». En conclusión, la cul-tura al estar menos estructurada que la biología tiene que aprender y seguir los sabios mecanismos de la biología, porque le va en ello la su-pervivencia36.
Los planteamientos de A. Gehlen son diferentes, pero bastante coinci-dentes a la hora de situar a la cultura en subordinación a la biología37. El enfoque antropológico central de Gehlen define al ser humano como ani-mal deficiente, siendo la cultura el resultado de los esfuerzos que ha teni-do que hacer, generación tras generación, para sobrevivir, en el ejercicio de suplir las consecuencias de tal deficiencia. De ahí que necesite estar en permanente alerta, como si viviese en un bote salvavidas, en permanente peligro de naufragar y perecer38. El esfuerzo por suplir la deficiencia bio-lógica construyendo cultura lo realiza a través del ejercicio de poner en marcha lo que denomina la ley de la descarga, cuyos frutos constituyen las diferentes instituciones con las que se vertebran y constituyen las so-ciedades humanas39.
34 Cfr. Lorenz, K., Sobre la agresión. El así llamado mal, Madrid, Siglo XXI, 1980. 35 Cfr. Id., Los ocho pecados capitales de la humanidad civilizada, Barcelona, Plaza y
Janés, 1975. 36 Cfr Beorlegui, C., «El reto de la biología a la antropología. De la etología a la socio-
biología», o.c., pp. 37-69; Arranz, M., «La antropología de Konrad Lorenz», en Lucas, J. de S. (ed.), Nuevas antropologías del siglo XX, Salamanca, Sígueme, 1994, 43-78.
37 Cfr. Gehlen, A., El hombre, o.c. 38 Sobre el uso de ciertas teorías biologistas de la imagen del bote salvavidas, cfr.
VV.AA., La biología como arma social, Madrid, Edit. Alhambra, 1982. 39 Para una síntesis de las ideas centrales de la antropología y sociología de A. Gehlen,
cfr. Beorlegui, C., Antropología filosófica. Nosotros: urdimbre solidaria y responsable,o.c., pp. 395-432.

476
© Universidad de Deusto - ISBN 978-84-9830-383-4
Desde estos planteamientos, Gehlen considera que la sociedad hu-mana actual se ha ido desarrollando apartándose de los parámetros que le pueden mantener en la existencia, en la medida en que habría olvida-do su dependencia de lo biológico y se habría constituido en una culturade reflexión, olvidándose de que ha surgido de la biología y al servicio de la biología. De ahí que, comparando la cultura elemental pero sólida del hombre primitivo con la actual, la primera sale ganando de la compara-ción, como lo indica en su libro Hombre primitivo y cultura tardía.
Este desvío de la cultura contemporánea lo considera Gehlen como irre-versible, en la medida en que ya no habría salvación posible, no quedándolea los críticos como él más que aceptar lo inevitable, puesto que tiene no vuelta atrás, aunque saben que ya no se puede hacer otra cosa que distan-ciarse y no colaborar con ella, en la medida en que ya no habría otra salida que la decadencia y la progresiva desaparición.
Las críticas a sus planteamientos conservadores40, por otro lado lle-nos de evidentes contradicciones, no han conseguido modificar sus pos-turas sino radicalizarlas todavía más, no pareciendo darse cuenta de que sus planteamientos sobre la deficiencia biológica de la especie humana, posibilitadora de la cultura a través del ejercicio de la ley de la descar-ga, sirven no tanto para mostrar la subordinación de la especie humana a la biología sino para mostrar su autonomía respecto a ella. El ser hu-mano es la especie en la que se han roto los constreñimientos cerrados de lo biológico, dando paso al surgimiento de la libertad y, por ello, a la pluralidad de comportamientos y de respuestas a las condiciones del en-torno ambiental, y a las condiciones de organización de los propios co-lectivos humanos para dar paso a sociedades organizadas de una forma compleja y plural.
2.3. El culturalismo dualista
Las dos posturas siguientes sitúan el centro de la dualidad biología-cultura en el polo cultural, pero se diferencian en el modo como conju-gan esta centralidad. Ambas posturas coinciden en superar el reduccio-nismo biologista, pero difieren en el modo de relacionar la dimensión cultural con la biológica. Mientras el culturalismo extremo, de corte dualista, considera la cultura como un continente autónomo y autosufi-ciente respecto a la biología, no necesitándose apelar a ella para enten-der ni el origen ni el funcionamiento interno de lo cultural, el culturalis-
40 Cfr. la polémica de Gehlen con Habermas y Adorno, en Beorlegui, C., o.c., pp. 472-481.

477
© Universidad de Deusto - ISBN 978-84-9830-383-4
mo estructurista defiende que la cultura representa un salto cualitativo respecto a la biología, pero considera también que la cultura ha surgido de la biología y necesita de la cultura para entender al ser humano en su globalidad.
En resumen, para entender correctamente la relación biología-cultura, se han de distinguir dos leyes o dos afirmaciones que resultan comple-mentarias:
— La cultura depende de la biología en su origen, en la medida en que la cultura es una dimensión humana que ha emergido como fruto de la evolución biológica. Así, los seres humanos no seríamos libres, inteligentes, capaces de hablar, etc, si no se hubiera dado el salto emergente (mutacional y biológico) de lo pre-humano a lo hu-mano, y si no se hubiera producido el progresivo aumento del cere-bro hasta llegar a las proporciones tan extraordinarias que la espe-cie humana actual posee.
— Pero, por otro lado, la cultura no depende de la biología en sus leyes y en su estructura interna, puesto que las leyes que rigen el lenguaje y el pensamiento humano, así como el modo como nos organizamos en sociedad, etc., no dependen de las leyes biológi-cas, aunque no puede olvidarse que no podemos pensar ni actuar si nuestro cerebro no funciona correctamente41.
Estos dos principios son necesarios para entender la relación existen-te entre nuestra dimensión biológica y la cultural. Estos dos principios sostienen la postura que hemos denominado culturalismo estructurista,pero no parecen estar tan claros para la tercera postura, el culturalismodualista, que tiene una idea más independiente de la cultura respecto de lo biológico.
Un representante significativo de esta postura es el antropólogo nor-teamericano Marshall Sahlins42, representante del denominado simbolismocultural y destacado por sus críticas a la primera fase de la sociobiología de E. O. Wilson. Sus planteamientos críticos respecto al reduccionismo biologista de la sociobiología son legítimos y muy acertados, pero nos pa-rece inadecuado el modo de entender el funcionamiento de lo cultural y su autosuficiencia respecto a la biología.
Para Sahlins y los simbolistas culturales, la mejor prueba de la incon-sistencia del determinismo biológico está en la pluralidad de las culturas
41 Sobre la dependencia del pensar respecto a nuestra condición biológica, cfr. Mark Johson, El cuerpo en la mente, Madrid, Debate, 1991.
42 Cfr. Sahlins, M., Uso y abuso de la biología, Madrid, Siglo XXI, 1982; Id., Cultura y razón práctica, Barcelona, Gedisa,

478
© Universidad de Deusto - ISBN 978-84-9830-383-4
humanas43. Si se diera tal determinismo, no se habría dado más que un modo humano de comportarse y de acumular respuestas exitosas para transmitir a sus herederos, mecanismo sobre el que se apoya y constituye la cultura humana. Por el contrario, en cada especie animal no se da más que un modo específico de construirse su relación con el entorno, fruto y consecuencia de un único modo de comportarse, en consonancia con la estructura biológica de cada especie. Esa configuración del comporta-miento de cada especie es lo que denomina K. Lorenz etograma: el con-junto de modos de actuar y de comportarse propios de una especie ani-mal concreta.
Todo esto demuestra que los comportamientos humanos, las culturas, no son consecuencia de la biología, sino que son fruto de creaciones hu-manas arbitrarias, resultado del esfuerzo humano por sobrevivir, al enfren-tarse con las diferentes circunstancias que los grupos humanos han tenido que experimentar a lo largo de su dilatada existencia. Para el simbolismocultural de Sahlins, tanto los diferentes modos como el ser humano ha ido creando respecto a la caza, la recolección de frutos, la agricultura y ga-nadería, como respecto al lenguaje, la organización social, la moralidad, el derecho, las teoría filosóficas y las religiones, son fruto de consensos progresivos y arbitrarios. La condición simbólica que impregna toda la cultura humana está basada en convenciones creativas y arbitrarias de los seres humanos, y no dependen de las condiciones biológicas, que siguen otras leyes y otra lógica.
Sus tesis sobre el origen de la cultura son de gran interés, pero la limita-ción de esta postura está en no ver que se puede, y se debe, defender la con-dición autónoma de la cultura, basada en creaciones inteligentes y libres de los seres humanos, pero complementándola con la dependencia respecto a la biología, tanto en su origen como en su funcionamiento. Si la diversidad de las culturas humanas es un hecho y una prueba contra el determinismo y reduccionismo biológico, no resulta tan acertado defender que las culturas se han originado sin tener en cuenta para nada las condiciones biológicas del ser humano, así como las condiciones materiales de las circunstancias tecnoecológicas y tecnoeconómicas de las que dependen inevitablemente, como defienden los materialistas culturales como Marvin Harris44.
Por tanto, es necesario conjugar la cultura con la biología, y no de modo dualista, como si fueran dos elementos totalmente diferentes, que conviven y colaboran de forma estrecha, sino de modo unitario, formando una estructura compleja pero unitaria.
43 Cfr. Moore, John H., «¿Cómo explicar la diversidad humana?», Mundo Científico,1995, n.º 160, 717-723,
44 Cfr. Harris, Marvin, El materialismo cultural, Madrid, Alianza, 1982.

479
© Universidad de Deusto - ISBN 978-84-9830-383-4
2.4. El culturalismo estructurista
Así es como entienden esta relación biología-cultura los integrantes de esta cuarta postura, en la que se sitúan autores como Francisco Aya-la45, Carlos París46, Xavier Zubiri47, Ignacio Ellacuría48, Diego Gracia49
y otros50.Seguiremos en la exposición de esta postura los planteamientos de
I. Ellacuría, apoyándose en la filosofía de Zubiri. Para esta postura, lo bio-lógico y lo cultural son autónomos, pero desde su mutua relación. Coincide con las dos afirmaciones básicas enunciadas con antelación, consistentes en considerar que la cultura, esto es la dimensión psíquica, depende de la biología en su origen, pero es autónoma en su funcionamiento interno.
Pero su aportación central se encamina a mostrar la correlación in-trínseca de ambas dimensiones, entendiendo la realidad del hombre y, por tanto, la estructura de su comportamiento (su habitud)51 como un sistema o estructura única que comprende dos momentos, el biológico u orgánico, y el psico-social. Por tanto, la biología es un momento constitutivo de la realidad humana. Lo humano es una estructura esencial, compuesta por dos subestructuras: una orgánico-biológica y otra psíquica.
De este modo, las notas biológicas poseen las siguientes características:
— son notas esenciales, no accidentales o adventicias, de la realidad humana;
45 Cfr. Ayala, Francisco J., Origen y evolución del hombre, Madrid, Alianza, 1980, o.c.46 Cfr. París, Carlos, El animal cultural, o.c.47 Cfr. Zubiri, Xavier, Sobre el hombre, Madrid, Alianza/Sociedad de Estudios y Publi-
caciones, 1986. Edición y presentación de Ignacio Ellacuría. 48 Cfr. Ellacuría, Ignacio, «Fundamentación biológica de la ética», ECA, 1979, n.º 368,
pp. 418-425; Id., «Biología e inteligencia», ponencia presentada en el XIV Congreso Na-cional de la Asociación Española de Neuropsiquiatría («Bases biológicas de la psiquia-tría»), Sevilla, octubre, 1977, y editada en Escritos Filosóficos, San Salvador, Edic. UCA, 2001, vol. III, pp. 137-201.
49 Cfr. Gracia, Diego, «Biología e inteligencia», Estudios Centroamericanos (ECA, San Salvador), XXXVIII (1982), n.º 406, pp. 801-816.
50 Cfr. Lewontin, R./Rosen, S./Kamin, L. J., No está en los genes. Racismo, genética e ideología, o.c.; Gutiérrez, A./Ortega, R, «Sobre el hombre biocultural», Letras de Deusto, 21 (1991), n.º 51, 153-165.
51 A la hora de describir el comportamiento humano desde la filosofía de Zubiri, hay que distinguir tres niveles: el de la estructura o sustantividad (el ser humano es una úni-ca estructura con dos subsistemas: lo orgánico y los psíquico), la habitud (el modo de en-frentamiento o modo de habérselas con la realidad, que en el caso de los humanos es for-malidad de realidad), y los actos (los actos humanos están conformados también por una estructura unitaria, psicoorgánica, aunque unos actos poseen dominancia orgánica y otros, psíquica). Cfr. Zubiri, Sobre el hombre, o.c.; Oscar Barroso, Verdad y acción, Granada, Co-mares, 2002, pp. 21 y ss.

480
© Universidad de Deusto - ISBN 978-84-9830-383-4
— pero no agotan, con todo, la realidad humana; — conservan sus características propias, esto es, no dejan de ser bioló-
gicas;— son notas-de, esto es, pertenecen al conjunto de la estructura huma-
na total; — pero pierden su sustantividad (la que poseían cuando eran notas de
un animal no humano), y entran a formar parte de una nueva sus-tantividad, al ser integradas en la estructura única que conforma una nueva realidad humana, como fruto de la elevación de una es-pecie pre-humana a la humana, dentro del proceso evolutivo.
Desde estas afirmaciones es como hay que entender lo cultural como superación de lo biológico. Esa expresión no implica la desaparición o irrelevancia de lo específicamente biológico, sino su desaparición como sus-tantividad autosuficiente, para ser elevada a una parte o componente de la única sustantividad, la realidad de la especie humana (dimensión filogené-tica) y de cada ser humano (dimensión ontogenética).
En definitiva, decir que el hombre es una estructura psico-somática significa que no hay una inteligencia humana sin sensibilidad. Por eso, la inteligencia humana es una intelección o inteligencia sentiente. Pero tam-bién significa, de modo complementario, que la animalidad humana no es viable sin la inteligencia. De ahí que tengamos que hablar de una sensibi-lidad intelectiva.
En consecuencia, ni la persona humana, ni el sujeto humano, pueden ser persona, ni sujeto, más que de un modo constitutivamente biológico. Puede verse, por tanto, que esta postura recoge sintéticamente y permite conjugar unitariamente lo biológico y lo psico-cultural, sin quedarse en posturas unilaterales extremas.
En síntesis, si comparamos las cuatro posturas presentadas, vemos la estrecha relación de esta problemática con la que se refiere a la re-currente polémica sobre las relaciones alma-cuerpo, o mente-cerebro, o mente-cuerpo. Así, mientras los monismos reduccionistas coinciden con las posturas de los biologismos reduccionistas, y los dualismos lo hacen también con ciertos planteamientos del denominado culturalismo simbó-lico, la postura del culturalismo estructurista representa, en la línea de los emergentismos y estructurismo sistémicos, el intento de plantear una vi-sión unitaria de la realidad humana, en la que se conjuguen la visión uni-taria y estructurista con la complejidad psico-somática de esta realidad humana.
Igualmente, la temática biología-cultura se relaciona directamente con la que se ocupa de estudiar la conjugación de lo genético y lo ambiental en la explicación de nuestros comportamientos, en la que se dan también

481
© Universidad de Deusto - ISBN 978-84-9830-383-4
tres posturas: dos extremas (los determinismos genético y ambiental) y la postura integradora, que entiende el comportamiento como la síntesis de ambos factores, sin que se pueden separar de un modo claro y simplista52.Está claro que no es posible que la evolución del homo sapiens haya des-pojado totalmente al hombre de sus bases adaptativas genéticas (contra el ambientalismo extremo). Su gran adaptabilidad y capacidad de cambio frente a su entorno empuja a creer en la supremacía del aprendizaje cultu-ral sobre el determinismo genético, pero sin caer en el determinismo am-biental.
3. LA RELACIÓN DE LA BIOLOGÍA CON LA ÉTICA
La relación entre biología y ética es un aspecto y concreción del tema más amplio de la relación entre biología y cultura, pero lo quere-mos explicitar aquí porque resulta necesario estudiarlo en la actualidad, al mismo tiempo que nos ayuda a comprender mejor y a concreta la pro-blemática general de relación entre la biología y cultura, que acabamos de analizar.
Esta relación biología-ética ha sido tratada y estudiada por múltiples autores y desde muy diversos puntos de vista53. Por eso, resulta imposible y demasiado complicado pretender referirme a toda este amplio abanico de posturas. Me voy a limitar a plantear una síntesis del modo de enfocar
52 Lewontin, R. C./Rose, S./Kamin, L. J., No está en los genes, o.c.; Jordan, B., Los im-postores de la genética, Barcelona, Península, 2001; Sapolsky, El mono enamorado y otros ensayos sobre nuestra vida animal, Barcelona, Paidós, 2007; Salgado, Alfonso, Biologíadel comportamiento. Introducción a la Psicología biológica. Una propuesta para la con-vergencia, Salamanca, Universidad Pontificia, 2007.
53 Cfr. Cela Conde, C. J., Sobre genes, dioses y tiranos, Madrid, Alianza, 1985; Ayala, Fr./Cela Conde, C. J., La piedra que se volvió palabra, Madrid, Alianza, 2006, pp. 21-30; Izquieta, J. L./García Rioboo, M.ª A., «Altruismo y solidaridad. Apuntes antropológi-cos», Estudios Filosóficos, 1987, n.º 36, 459-462; Kitcher, Ph., «El origen de la moral. La tendencia a la simpatía no contradice la teoría de la evolución», Mundo Científico, 1997, n.º 181, 608-611; Gutiérrez Martínez, A., «El fenómeno ético y religioso visto desde la sociobiología. Introducción a la literatura», La Ciudad de Dios, CCXV/1 (2002), pp. 123-154; Franz de Waal, Bien natural. Los orígenes del bien y del mal en los humanos y otros animales, Barcelona, Herder, 1997; Pierre de Puytorac, De la biophilosophie à une éthique de la biologie (La societé face à la biologie), Paris, L’Harmattan, 1998; Tugendhat, E., «La moral en perspectiva evolucionista», en Id., Problemas, Barcelona, Gedisa, 2001, pp. 135-158; Changeux, J-P. (dir.), Fondements Naturels de l’Ethique, París, Odile Jacob, 1993; Ruse, M., Sociobiología, Madrid, Cátedra, 1983, último capítulo; Hauser, Marc D., La mente moral. Cómo la naturaleza ha desarrollado nuestro sentido del bien y del mal, Barcelona, Paidós, 2008.

482
© Universidad de Deusto - ISBN 978-84-9830-383-4
esta problemática de la mano Francisco J. Ayala, uno de los autores que con más acierto la ha tratado54.
Ayala parte del hecho de que la ética se nos muestra como un atributo exclusivo de la especie humana, transmitido genéticamente, como un ele-mento consustancial de nuestra condición de humanos, por lo que cabría entender esta característica como sometida al determinismo de las demás características transmitidas genéticamente. Cabría, pues, sospechar que este rasgo humano fuera algo que depende estrechamente de lo biológi-co. De ahí que tenga sentido, en parte, considerar la ética como algo que depende de la genética, dándose entre ambos elementos una estrecha re-lación. Es evidente que así es, pero nos indica Ayala acertadamente que, para plantearse adecuadamente esta cuestión y resolverla de modo acer-tado, es necesario distinguir previamente dos niveles o dos aspectos muy distintos: la relación, por un lado de la biología con la capacidad ética, y, por otro, la relación de la biología con los códigos morales.
Si nos preguntamos si la capacidad ética depende de la biología, te-nemos que responder que sí, porque somos éticos, es decir, tenemos ca-pacidad de distinguir entre el bien y el mal porque poseemos un cerebro tan complejo y evolucionado que nos capacita para ese tipo de distincio-nes, aunque no todas las culturas ni momentos históricos coincidan de la misma forma lo bueno y lo malo. En cambio, no se advierte que ninguna de las especies animales tengan esta capacidad, aunque algunos chim-pancés muestran ciertas capacidades cercanas a los rasgos éticos de los humanos55.
Según Ayala, lo que nos capacita a los humanos para los discernimien-tos morales son tres rasgos que poseemos en exclusiva: en primer lugar, la capacidad de prever las consecuencias de nuestras acciones; en segundo lu-gar, la capacidad de formular juicios de valor, esto es, el que podamos con-siderar nuestras acciones y las de los demás como buenas o malas: y, por último, la capacidad de elegir entre diferentes modos alternativos de actua-ción56. Por tanto, los humanos tenemos capacidad de evaluar lo correcto o incorrecto de nuestras acciones, y además poseemos libertad para elegir en-tre las diversas alternativas de acción que se presentan ante nosotros.
54 Ayala ha tratado el tema de la relación entre biología y ética en varios de sus traba-jos, pero en todos ellos sigue un mismo esquema fundamntal, por lo que no voy a referir-me más que a uno de sus textos, al capítulo VII, «La ética y la religión», de su libro Origeny evolución humana, o.c., pp. 169-190. Cfr. también Ayala, F. J., «Ensayo sobre las bases biológicas del comportamiento moral», Estudios Filosóficos, LVII (2008), n.º 165, 225-246, donde el autor repite las ideas ya avanzadas en textos anteriores.
55 Cfr. De Waal, Franz, Bien natura. Los orígenes del bien y del mal en los humanos y otros animales, Barcelona, Herder, 1997.
56 Cfr. Ayala, Fr., o.c., pp. 172-173.

483
© Universidad de Deusto - ISBN 978-84-9830-383-4
Estas capacidades dependen de que tenemos una naturaleza abierta, que nos hace nacer prematuramente y con deficiencia biológica de cara a responder de modo ya prefijado a los estímulos del entorno, y, como consecuencia de ello, se nos ha dejado a nuestra libertad elegir entre las diversas alternativas. Esta capacidad cerebral es consecuencia del salto dado, dentro de la historia del desarrollo cerebral, desde el neopalio de los reptiles hasta la formación de la corteza cerebral de los mamíferos, com-pletada y aumentada de forma importante en los humanos.
Se pregunta Ayala si hay alguna especie animal que posea también la capacidad ética, y considera que no, puesto que, aunque se dan en algunos determinados individuos de la especie de los chimpancés algunos indi-cios de conductas similares a las conductas éticas humanas, como lealtad, compasión, engaño, etc., esas conductas están determinadas por su consti-tución genética o por aprendizaje enseñado en algunos laboratorios. Pero no se advierte en esas conductas las tres cualidades indicadas que se ha-llan a la base de la capacidad ética que advertimos en los humanos.
Por tanto, tenemos que aceptar que en la especie humana se ha dado un salto cualitativo, continuando el acercamiento progresivo en anteriores especies animales, hasta haber traspasado un umbral evolutivo, que toda-vía no somos capaces de definir ni explicar, como ocurre con el salto de la no conciencia a la conciencia. Este salto, reconoce Ayala, no es el úni-co que se da en historia de la evolución, donde se pueden advertir otros saltos en los que también se han traspasado su correspondiente umbral crítico, como es el caso del salto desde la materia inanimada a la vida, la complejificación multicelular, la aparición de la reproducción sexual, la emergencia de la autoconciencia, etc. Son saltos cualitativos sobre los que no cabe, desde la mera evidencia empírica de la ciencia, más que consta-tar su aparición, sin saber en muchos casos explicar por qué se da tal salto o cambio, como no sabemos explicar por qué el agua cambia de estado lí-quido a gaseoso a los 100.º C, y no a los 90.º ó a los 120.º57.
Las respuestas que tratan de dar una explicación a estas cuestiones, son propuestas de explicación que pertenecen al ámbito de la filosofía, de la metafísica, cuyo estatus explicativo pertenece a otro nivel epistemoló-gico, en la medida en que sobrepasa el nivel demostrativo de las eviden-cias empíricas propio de las ciencias naturales.
Una vez contestada de forma afirmativa la cuestión de la dependen-cia de la capacidad ética respecto de la biología, pasamos a la cuestión sobre la relación entre la biología y los contenidos de la moralidad. ¿De-penden las normas morales de las reglas y leyes biológicas? Aquí la res-
57 Cfr. ibídem, pp. 174-176.

484
© Universidad de Deusto - ISBN 978-84-9830-383-4
puesta tiene que ser negativa. La moralidad no tiene nada que ver con las leyes de la biología, sino que se construyen siguiendo las pautas racio-nales que se han ido conformando en ámbitos culturales a lo largo de la historia. Eso no significa que el ámbito de la racionalidad, entre el que se encuentra la ética, sea totalmente independiente de la biología, de modo que pueda construir normas morales contrarias a la biología y a la super-vivencia de la especie. Y, por ello, es cierto que la conciencia y la capaci-dad moral ha dotado a los humanos de mayor capacidad para sobrevivir, pero la finalidad de la moral no se reduce a ser un mero instrumento al servicio de la reproducción biológica y al mantenimiento de la especie en la existencia58.
Y también es importante señalar que las normas morales de los hu-manos no han surgido de forma repentina y acabada, sino que han experi-mentado una larga historia tanto en el aspecto filogenético como ontoge-nético. Baste referirse a las investigaciones sobre la evolución de la ética en los niños, realizada por Piaget, Kolhberg y otros59, así como echar mano de las diversas historias de la moralidad60, donde se muestra que la historia de la ética es un progresivo avance en la sensibilidad humana ha-cia lo que es bueno y justo, desde la experiencia de lo malo e injusto, pre-sente en las víctimas de la maldad de los humanos61.
La cuestión es ahora cómo explicar el origen y la lógica de los con-tenidos o leyes de la moralidad, esfuerzo que se suele conocer como la justificación o fundamentación de la ética. Ayala pasa rápido sobre este punto, y no presenta una justificación sistemática ni presenta un abanico completo de los modos de fundamentar la moral existentes en la actuali-dad, porque lo que Ayala pretende en su escrito es sólo demostrar que no es posible fundamentarla desde la biología.
Se ha intentado la fundamentación de la ética tanto desde la religión como desde la filosofía y la ciencia62. Las propuestas desde la religión se encaminan a mostrar la obligación de hacer el bien como expresión de la voluntad de Dios. Pero esto puede justificarse de dos maneras muy dife-rentes: de modo fundamentalista, entendiendo la voluntad de Dios como algo bien conocido y claramente presentado en los libros sagrados de la
58 Cfr. Ruse, M., Tomándose a Darwin en serio, o.c. 59 Cfr. Rubio Carracedo, J., El hombre y la ética. Humanismo crítico, desarrollo mo-
ral, constructivismo ético, Barcelona, Anthropos, 1987 (Parte II: La teoría del desarrollo moral).
60 Cfr. Camps, Victoria y otros (dir.), Historia de la ética, 3 vols., Barcelona, Crítica, 1987-1992; MacIntyre, Alasdair C., Historia de la ética, Barcelona, Paidós, 1982 (1991).
61 Cfr. Marina, J. A./Valgoma, M.ª de la, La lucha por la dignidad: teoría de la felici-dad política, Barcelona, Anagrama, 2001.
62 Cfr. Ellacuría, I., «Fundamentación biológica de la ética», o.c.

485
© Universidad de Deusto - ISBN 978-84-9830-383-4
religión correspondiente, como pretenden las posturas dogmáticas y con-servadoras, sin ser conscientes ni aceptar de buen grado que resulta im-posible saber con claridad total la voluntad de Dios, y que las supuestas expresiones de la misma están siempre mediadas por los que se conside-ran intérpretes de la misma. Por eso, la teología cristiana, aceptando la ra-cionalidad crítica ilustrada, trata de buscar la voluntad de Dios a través de las leyes de la naturaleza y de los acontecimientos históricos, entendiendo que toda afirmación acerca de ella son meras propuestas humanas, que tienen que aceptarse de forma limitada y provisional, y ser mediadas y criticadas por la propia racionalidad humana.
Las propuestas filosóficas son de lo más variadas, desde la que par-te de aceptar la existencia de un mundo autónomo de valores metafísicos, pasando por la que se apoya en la naturaleza humana o en algún rasgo clave de la misma, o bien la propuesta que se suele denominar como irra-cionalismo ético. En el primer caso, en la línea de autores como M. Sche-ler, los valores son realidades en sí autónomas, que existen independien-temente del conocimiento de los humanos, pudiéndose captar a través de una intuición eidética de diferente tipo, bien intelectual o emotivo. Para la segunda opción, la fundamentación de la moral se realiza a partir de la naturaleza humana, considerada como naturaleza abierta, necesitada de permanente realización a partir de la apropiación de posibilidades que nos presenta la realidad (Zubiri), o bien a partir de la reflexión trascendental desde el lenguaje (la facticidad del lenguaje, como homo loquens), dedu-ciendo de tal deducción trascendental los elementos que componen la si-tuación ideal de diálogo (como idea reguladora a perseguir), encontrándo-se ahí las condiciones básicas de una ética dialógica y responsable, de tipo formal y procedimental. La tercera propuesta filosófica la hemos denomi-nado irracionalismo ético, y tiene, a su vez, dos versiones: la positivista y la individualista existencial. Para la postura positivista, la ética pertenece al mundo de lo irracional, o arracional, ya que discutir y dilucidar qué sea bueno o malo no pertenece al ámbito de la racionalidad objetiva (el campo exclusivo de la ciencia, según ellos), sino que es cuestión de preferencias afectivas y sentimentales. Es decir, no se puede decir que algo es bueno o malo, sino simplemente que me gusta o no. En cambio, para el indivi-dualismo existencialista, la ética también es un ámbito irracional, pero de diferente forma, en la medida en que la decisión de qué sea bueno o malo no pertenece al ámbito de lo racional objetivo o intersubjetivo, sino al terreno de la decisión voluntarista de cada individuo, única fuente de sen-tido y de moralidad. Así, para el primer Sartre, en su empeño por defender la exclusiva responsabilidad individual de las acciones afirmaba que «no realizo una acción porque sea buena, sino que es buena porque lo decido yo». El problema de esta postura es que, según eso, todo es bueno (con tal

486
© Universidad de Deusto - ISBN 978-84-9830-383-4
de que lo decida libremente) y nada es malo; pero, por otro lado, no hay modo de hacer juicios morales intersubjetivos cuando lo que uno decide como bueno es malo y perjudicial para otro, de tal forma que no salimos del subjetivismo ético y de la arbitrariedad moral.
Los intentos de fundamentar la ética desde las ciencias, si se entien-de el intento de modo estricto y radical, se tienen que apoyar siempre en la negación de la libertad humana, defendiendo que las acciones humanas están causadas por las leyes de los genes (determinismo genético), o las leyes del funcionamiento cerebral (determinismo neurológico o psicológi-co), o por las leyes del ambiente (determinismo ambiental o sociológico). El problema de estas posturas es que resulta imposible demostrar la ver-dad del determinismo, e igualmente constituye una contradicción querer mantener al mismo tiempo la afirmación del determinismo y el lenguaje ético63. O se afirma el determinismo, y por tanto no tiene sentido hablar de ética, o, al revés, se sigue hablando de ética, y, en definitiva, damos por sentado que somos libres para elegir entre diversas posibilidades morales.
Como hemos dicho, Ayala se detiene sobre todo en mostrar la in-consistencia de las propuestas de fundamentar o justificar la ética desde postulados biologistas de corte evolucionista. Para ello, se refiere en pri-mer lugar a los que denomina evolucionistas clásicos, como H. Spencer, J. S. Huxley y C. H. Waddington, que defienden el naturalismo biológico, para, en segundo lugar, detenerse más ampliamente en la propuesta del sociobiólogo E. O. Wilson64.
La idea que defiende el naturalismo biológico de los denominados evolucionistas clásicos es que la evolución, en su tarea de seleccionar lo apto para sobrevivir frente a lo no viable, va decantando y haciendo emer-ger lo bueno frente a lo que no lo es. De ahí que las reglas que, según ellos, rigen el proceso evolutivo se convierten también en las reglas de la moralidad. En virtud de ello, la máxima regla de la moralidad, tal y como nos enseña la naturaleza, es luchar cada uno por su propio beneficio, pues-to que de ahí se deducirá el bien para el conjunto de la especie, ya que de esa lucha quedarán los mejores, encargados a su vez de ir perfeccionando progresivamente la especie.
Pero, como bien indica Ayala, este planteamiento se apoya en tres erro-res de bulto. En primer lugar, dan por hecho que la tendencia evolucionis-ta introduce un permanente progreso en el desarrollo biológico, cuando en el ámbito de la naturaleza no hay valores sino meros sucesos desprovistos de adscripciones morales. La realidad es que cuando se quieren ver valores
63 Cfr. Berlin, I., Libertad y necesidad en la historia, Madrid, Revista de Occidente, 1958.
64 Cfr. Ayala, Fr., o.c., pp. 177-186.

487
© Universidad de Deusto - ISBN 978-84-9830-383-4
presentes en el proceso evolutivo es porque los humanos los hemos pro-yectado en él, no porque estén ya presentes dentro de dicho proceso. Ade-más, en segundo lugar, considerar que los valores se deducen de las leyes naturales es caer en la falacia naturalista, apelativo que Moore utilizó para la confusión entre el ser con el deber ser, tal y como ya nos había adver-tido D. Hume. Pero además, en tercer lugar, si dejamos que sean las leyes naturales las que dirijan nuestras orientaciones morales, podemos llegar a defender conclusiones tan absurdas e inmorales como no poder imponer remedios médicos ante enfermedades hereditarias o ante cualquier infec-ción de microorganismos dañinos, porque, siendo consecuentes, habría que dejar obrar a la naturaleza y respetar la estricta selección natural. Esto nos hace ver que en la naturaleza hay leyes sin más, sin ninguna connotación moral en sí misma. La moral es una proyección de los seres humanos, que no contradice a la naturaleza, pero que tampoco se deduce de ella.
La crítica que hace Ayala a los sociobiólogos es diferente, porque, como ya dijimos en otro apartado anterior, pretenden evitar en sus juicios morales la falacia naturalista. Pero Ayala trata de mostrar que, al final, Wilson no escapa de ella. En su intento de estudiar sistemáticamente la base biológico-genética de los comportamientos de todos los seres vivos, también de los humanos, los sociobiólogos aceptan la distinción entre biología y cultura, uno de cuyos capítulos es la ética. De este modo, entienden que la evolu-ción no enseña nada acerca de lo que los hombres deben hacer.
A pesar de esa afirmación, Wilson entiende que la biología nos mues-tra que hay que superar la idea de que exista un código ético uniforme y universal, para afirmar que el pluralismo moral es algo que poseemos los seres humanos de forma innata. La biología nos muestra, según Wilson, que ciertos códigos morales, en concreto los que tienen pretensión de uni-versalidad, son incompatibles con la naturaleza humana, y, por tanto, son inaceptables65.
La ética hay que situarla, como el resto de las realidades humanas, dentro del proceso evolutivo. Es consecuencia del desarrollo del cerebro, y éste es, a su vez, consecuencia de las orientaciones programadas en los genes. Así que todos los aspectos de la cultura, la ética entre ellos, tiene que enfocarse y entenderse como algo que surge de la evolución y que tie-ne que ponerse al servicio de la misma. De ahí que afirme que «el com-portamiento humano es la técnica indirecta por la cual el material genéti-co humano ha sido, y será, mantenido intacto. La moralidad no tiene otra función última demostrable»66.
65 Cfr. Wilson, E.O., Sobre la naturaleza humana, o.c. 66 Ibídem, p. 167 (del original inglés).

488
© Universidad de Deusto - ISBN 978-84-9830-383-4
Ayala considera que esta afirmación es ambigua y también errónea en los dos sentidos a los que parece apuntar. Si cuando Wilson habla de mo-ralidad se refiere a la capacidad moral, se trata de una afirmación falsa, porque, como indica Ayala, «los seres humanos son seres éticos por natu-raleza, debido a su eminencia intelectual que les dota de las capacidades de prever las consecuencias de sus acciones, de formular juicios de valor y de libre albedrío; una vez que se dan estas tres condiciones, la evalua-ción moral de las acciones humanas es inevitable. Los hombres son seres éticos necesariamente y por naturaleza, no porque esto sirva para conser-var sus genes o para cualquier otro propósito»67.
Si, por el contrario, cuando menciona Wilson la moralidad quiere re-ferirse a los contenidos morales, significa que se está contradiciendo, en la medida en que nos ha afirmado que no se puede deducir de las leyes biológicas lo que el ser humano tiene que hacer, cayendo, por tanto, en la falacia naturalista. Además, estaría adoptando una postura peligrosa desde el punto de vista social, en la medida en que este modo de pensar serviría para justificar el darwinismo social, el sexismo, el racismo y el genocidio, puesto que estos fenómenos no se podrían criticar desde una moral que defendiera los derechos humanos, en la medida en que se jus-tificarían desde leyes biológico-genéticas contra las cuales no se podría ir ni combatir.
Todas estas críticas fueron presentadas por muchos teóricos contra-rios a la primera etapa de la sociobiología68, ante las cuales se defendieron tanto Wilson como el resto de los sociobiólogos. Pero sus afirmaciones dan pie a estas críticas, y mientras no definan claramente la autonomía de la cultura y de la ética respecto a lo biológico-genético, difícilmente se va a librar de estas críticas, que no se refieren tanto a la intencionalidad de los teóricos que las defienden, sino a los contenidos implícitos de sus afirmaciones, como nos hace ver el punto de vista de la sociología del co-nocimiento, que se pregunta por la función social que realizan las teorías científicas o filosóficas.
Tanto los propios sociobiólogos como sus críticos están de acuerdo en que la piedra de toque para validar o falsar sus afirmaciones sobre la ética
67 Ayala, Fr., o.c., p. 184. 68 Lewontin, R./Rose, S,/Kamin, L. J., No está en los genes. Racismo, genética e ideo-
logía, o.c.; VV.AA., La biología como arma social, o.c.; Sahlins, M., Uso y abuso de la biología, o.c.; Archer, L., La amenaza de la biología, Madrid, Pirámide, 1983; París, C., «La sociobiología y el concepto de cultura», o.c.; Id., El animal cultural, o.c.; Gutiérrez Martínez, A., «Las implicaciones sociales y políticas de la sociobiología: una introducción a la literatura», Revista de Estudios Políticos, 1990, n.º 67, 297-318; Id., «El debate de la sociobiología: una aproximación bibliográfica», Suplementos Anthropos, n.º 10, 170-177; Beorlegui, C., Antropología filosófica, o.c., pp. 186-189.

489
© Universidad de Deusto - ISBN 978-84-9830-383-4
dependen de cómo resolvamos el tema del altruismo. Está claro que si la lógica de los comportamientos de todos los seres vivos es el egoísmo ge-nético, como afirman los sociobiólogos, lo lógico es que el proceso de la selección natural lo tendría que haber eliminado, como contrario a la lógi-ca de lo viviente. Pero vemos que se halla presente de forma importante y constante en todas las especies vivas, no sólo en el ser humano.
La respuesta de los sociobiólogos a este problema es que el supuesto altruismo es un mecanismo que promueve la eficacia biológica inclusiva(inclusive fitness)69, puesto que los comportamientos altruistas no serían tales, sino que siempre promueven objetivos interesados, de modo que «altruismo» lo tenemos que poner entre comillas, o en cursiva. La razón de ello es que se da en el fenómeno del altruismo lo que se denomina la selección de parentesco, esto es, un mecanismo que funciona sólo cuando el objeto de tal altruismo es un pariente genético. Este tipo de altruismo,como el que se da entre la madre y sus hijos, o entre hermanos, no sería más que un mecanismo común de la evolución para preservar los genes propios o cercanos.
El problema estaría en los casos de altruismo en favor de alguien que no es pariente genético cercano. En esos casos, funcionaría lo que denominan los sociobiólogos el altruismo recíproco, un tipo de altruis-mo más exigente y duro, puesto que ya no se realiza en beneficio de pa-rientes genéticos cercanos. Pero en este caso se estaría poniendo en fun-cionamiento la ley de «hoy por mí y mañana por ti», de tal forma que, si se extiende este tipo de comportamiento generoso y desinteresado, saldríamos a la larga todos favorecidos. Pero esto significa que todos los comportamientos de los vivientes, también en el caso de los humanos, estarían siempre orientados por el beneficio egoísta de la conservación y optimización de los propios genes, tal y como los sociobiólogos nos quieren hacer ver.
No cabe duda de que los comportamientos altruistas humanos tienen muchos aspectos que coinciden con el altruismo biológico de los anima-les, en la línea defendida por la sociobiología. Pero los parámetros del al-truismo biológico no son los mismos del altruismo moral, sobre todo por-que el modelo sociobiológico niega en el fondo la hipótesis de la libertad. Si queremos hablar de ética, tenemos que dar por hecho la libertad, como dijimos en su momento. Si queremos reducir los comportamientos huma-nos a meros mecanismos guiados por los genes, como se daría en el ám-bito animal, tendremos que renunciar a emplear el lenguaje moral.
69 Cfr. Hamilton, W. D., «The Genetical Evolution of Social Behavior». Journal of Theoretical Biology, 1964, 7, pp. 1-52.

490
© Universidad de Deusto - ISBN 978-84-9830-383-4
Frente a los sociobiólogos, que defienden el punto de vista de la se-lección individual dentro del proceso evolutivo, hay autores que están de-fendiendo con éxito planteamientos de selección de grupo a la hora de es-tudiar los comportamientos egoístas o altruistas, como es el caso de Eliot Sober y David S. Wilson70. Según Sober y Wilson, el altruismo es más efi-caz que la conducta egoísta cuando se trata de competir entre dos grupos rivales, de tal modo que el grupo que tiene individuos dotados de genes altruistas es más eficaz que el grupo que no los tiene. Un grupo de altruis-tas es a la larga más eficaz que la estrategia puramente egoísta, cuando el grupo de altruistas posee también la capacidad de detectar las estrategias egoístas camufladas, y descubrir a los impostores. De este modo, las tesis de Sober y Wilson resultan debilitadas en el ámbito de individuos que no estén dotados de esta capacidad cognitiva, pero se da precisamente en el ámbito de los humanos. Por tanto, la estrategia de la selección grupal, do-tada por comportamientos altruistas conscientes, resulta más beneficiosa y adaptativa que la contraria71.
Las conclusiones que presenta Francisco J. Ayala de esta discusión so-bre la relación entre biología y ética son tres72. La primera sostiene que, aunque determinados comportamientos humanos puedan coincidir con orientaciones morales, como obedecer a los mayores o amar y ayudar a los demás73, no se da un determinismo de lo biológico sobre lo moral. La condición biológico-genética nos puede predisponer a actuar de determi-nada manera, pero no nos determina de forma obligatoria y cerrada. El ser humano está dotado de libertad, de tal modo que puede aceptar unas ten-dencias y rechazar otras.
La segunda conclusión hace ver que, aunque ciertas normas morales coincidan con las tendencias biológicas propuestas por la selección natu-ral, otras muchas no muestran esa coincidencia. Y en estos casos, la moral nos indica y obliga a no seguirlas y a actuar en la línea de las ordenacio-nes morales. Ayala hace ver, en relación a este punto, que si fuera cierto el planteamiento sociobiológico, el mandato fundamental de la moral sería tener el mayor número posible de hijos, para aumentar la eficacia gené-
70 Cfr. Sober, E./Wilson, D. S., El comportamiento altruista. Evolución y psicología,Madrid, Siglo XXI, 2000.
71 Cfr. Cela Conde, C. J./Capó, M. A./Nadal, M./Gual, J./Fernández, M./Ramos, C., «De homínido a humano», en Feito, Lidia (ed.), Nuevas perspectivas científicas y filosófi-cas sobre el ser humano, Madrid, UPCO, 2007, pp. 67-92, sobre todo: 83-87.
72 Cfr. Ayala, F. J., o.c., pp. 187-188. 73 Esta es la estrategia que sigue M. Ruse para defender la relación entre biología y éti-
ca, en «Evolución y ética: una vieja relación reconsiderada», en Sanmartín, J./Simón, V./García-Merita, M.ª L. (comps.), La sociedad naturalizada. Genética y conducta, Valencia, Tirant Lo Blanch, 1986, pp. 149-169.

491
© Universidad de Deusto - ISBN 978-84-9830-383-4
tica. Y está claro que las orientaciones morales que rigen la convivencia social no es ésa. Lo que nos lleva a concluir que la mera supervivencia y la eficacia biológica no es la ley central de los comportamientos humanos, sino más bien perseguir los valores morales que consideramos más ade-cuados para la realización y el logro de la felicidad y la realización perso-nal y social.
Por tanto, como tercera y definitiva conclusión, «las normas morales están basadas en consideraciones y principios culturales (…), incluyendo los religiosos, no en principios biológicos. Las consideraciones éticas tie-nen que tener en cuenta las realidades biológicas, pero para decidir cuáles deban ser las normas morales de conducta de los seres humanos la biolo-gía por sí misma es palpablemente insuficiente»74.
4. EL SER HUMANO, UNIDAD BIO-CULTURAL
De este modo, podemos seguir afirmando que el ser humano es una estructura compuesta de dos ámbitos estrechamente relacionados: la bio-logía y la cultura. Ambos son esenciales e insustituibles en la composición de lo humano. Pero no somos la suma de dos cosas separadas y autosufi-cientes, sino una única estructura unitaria.
La cultura ha surgido por emergencia del proceso genético y evoluti-vo, y en su expresión y funcionamiento sigue necesitando de lo biológico. Nuestra intelección es sentiente (Zubiri), pero no es un sentir idéntico al meramente animal, sino que es un sentir inteligente y vertido a lo real en cuanto tal. De ahí que somos una especie que venimos de lo biológico, pero hemos saltado la frontera hacia la cultura. Y, por eso, lo que nos define en su más íntima esencia no es aquello de donde venimos (lo biológico), sino aquello a donde vamos y en donde nos encontramos (lo cultural) como un ámbito inacabado e interminable del proceso de humanización.
Así, la biología es condición de posibilidad de nuestra cultura, pero no orienta ni determina nuestros comportamientos culturales y morales. En ese punto, la racionalidad humana es la que orienta el discernimiento cons-tante de lo que tenemos que hacer para satisfacer los cánones morales.
74 Ayala, F. J., o.c., 188.

© Universidad de Deusto - ISBN 978-84-9830-383-4

493
© Universidad de Deusto - ISBN 978-84-9830-383-4
Capítulo 9
Continuidad y ruptura: la especie singular
1. UNA VISIÓN BIO-FILOSÓFICA DE LO HUMANO
La idea central que ha orientado el contenido de este libro ha sido presentar una visión de los rasgos biológicos fundamentales de la espe-cie humana, pero desde un enfoque crítico filosófico. No se ha tratado, por tanto, de un estudio biológico y genético de lo humano, sino de un acercamiento al estudio de lo que define al ser humano desde la óptica fi-losófica. Los espectaculares avances en el ámbito de todos los campos de la biología a lo largo del último siglo y medio han cambiado de modo tan radical la conciencia y la imagen que los seres humanos tenemos sobre nosotros mismos como especie, que hemos pasado de minusvalorar nues-tra condición corpórea hasta situarnos casi en el extremo contrario, en la medida de que para muchos teóricos no somos más que sólo biología, una especie animal más.
Los datos que las diferentes parcelas científicas nos aportan sobre las características genéticas y biológicas de nuestra especie y su comparación con las demás especies vivas, no nos facilitan la tarea de resolver las cues-tiones que más nos interesan: qué es en definitiva el ser humano, cuáles son las características que nos distinguen como especie. Cuestiones que no sólo buscan una respuesta comparativa en el ámbito de los rasgos mor-fológicos, biológicos y genéticos, sino más bien una respuesta que profun-dice en el terreno del sentido de la realidad humana. Somos una compleja realidad bio-cultural, emergidos del ámbito de la biosfera para adentrarnos en un nuevo continente, la cultura, que nos sitúa en una dimensión cuali-tativamente diferente a la del resto de los seres vivos.

494
© Universidad de Deusto - ISBN 978-84-9830-383-4
La tesis que hemos defendido a lo largo de todo el texto es que la es-pecie humana está enraizada en el mundo de la vida como cualquier otra especie, advirtiéndose en ello la radical continuidad con el resto de los animales, aunque al mismo tiempo una distinción cualitativa y de grado respecto a todos ellos. Esta pretensión de conjugar la continuidad y la rup-tura en lo humano, está apoyada en suficientes bases científicas, pero es deudora también de una opción cosmovisional humanista y antropocéntri-ca que no posee el mismo grado de probación empírica que un postulado científico. Prueba de ello es que, basándose en los mismos datos científi-cos, hay muchos autores que defienden posturas opuestas, reduccionistas, anti-humanistas y trans-antropocéntricas. Pero creemos que las posturas humanistas y antropocéntricas poseen suficiente plausibilidad racional y apoyo científico como para considerarlas más fuertemente probadas que las teorías contrarias.
2. EL RECONOCIMIENTO DE NUESTRA CONDICIÓN ANIMAL
Como hemos indicado ya, la compresión que el ser humano ha ido teniendo de sí mismo a lo largo de casi toda su historia ha distado mu-cho de reconocer lo corpóreo como un ingrediente esencial de lo humano. Aunque ya los griegos definían al ser humano como animal racional, el centro y la esencia de esa definición lo constituía el adjetivo, lo racional, mientras que el sustantivo era un mero añadido, o más bien, un estorbo. No cabe duda de que para el dualista Platón el cuerpo representaba una carga añadida, consecuencia de un castigo que los dioses habían impuesto al alma, como consecuencia de una desconocida culpa cometida cuando vivía en estado puro en el paraíso de las ideas. Es verdad que había otras teorías, como el hilemorfismo de Aristóteles y Tomás de Aquino, que se distanciaban del menosprecio dualista hacia lo corpóreo, pero no llegaban a conceder a la dimensión corpórea la importancia que en la actualidad reconocemos. Sólo con el avance de las ciencias médicas y biológicas, y sobre todo con la propuesta darwiniana de la selección natural como ex-plicación del proceso evolutivo, se llegó a reconocer a los humanos como una especie biológica más, sometida como todas al encadenamiento del proceso evolutivo.
El reconocimiento de la esencialidad de la condición genético-bioló-gica de la especie humana se inicia, por tanto, a partir del siglo XIX, con la emergencia de las teorías evolucionistas de Lamarck, y sobre todo con la teoría de la selección natural de Ch. Darwin y Wallace. La teoría de la selección natural suponía hallar la clave esencial del mecanismo de

495
© Universidad de Deusto - ISBN 978-84-9830-383-4
la evolución, al mismo tiempo que se tenía la convicción de que no sólo lasespecies animales no humanas se hallaban entrelazadas en una historia única y continuada, sino que también el género humano, en sus diferentes especies, no escapaba a este mismo proceso.
Se trataba, pues, de una profunda revolución dentro del modelo an-tropológico vigente, puesto que suponía situar a los humanos en medio de las bestias, y poner en cuestión su especial puesto en el cosmos, y su condición de imagen de Dios y representante suyo en el conjunto del pla-neta. Y de ese modo se abría camino la evidencia de que el ser humano se reducía a ser un especie como las demás, planteándose la cuestión de si lo biológico agotaba la totalidad del sentido de lo humano, o podía de alguna manera salvaguardarse todavía algún aspecto de su hasta entonces incon-testada condición privilegiada en medio del universo.
Este cambio de modelo antropológico suponía un duro golpe al nar-cisismo y a la autoestima de nuestra especie, y una puesta en cuestión de las tesis antropocentristas. Este descentramiento de la visión del mundo había comenzado ya en el siglo XVII con el triunfo de la visión cosmológica heliocéntrica propuesta por Copérnico y Galileo, pero el descentramiento biológico representó sin duda un golpe más fuerte si cabe que el anterior, puesto que ya no sólo se hacía evidente que la casa del hombre, el planeta Tierra, no era el centro del universo, sino que se ponía en cuestión y se rompía la pretensión histórica de verse como un ser aparte, creado de un modo especial por Dios, situando en ello su especificidad y su situación de privilegio, en la medida en que se veía hecho a imagen de Dios y encargado de gobernar la tierra y a todos sus pobladores.
Se le obligaba de este modo a la especie humana a bajar de lo etéreo a lo terreno, y a descubrirse cercano y emparentado con el mundo ani-mal, a quien veía siempre como representación de lo irracional y violen-to. Por otro lado, la tesis darwinista suponía hacernos cambiar la idea que teníamos de la naturaleza y la biosfera, puesto que se nos presentaba ahora como un mundo construido por un conjunto de leyes equilibradas y suficientes, ante las cuales no se necesitaba echar mano de la hipótesis divina y de los planteamientos religiosos. Quedaba, por tanto, la cues-tión de si la selección natural como explicativa del proceso evolutivo suponía o no la evidencia empírica de las tesis de la muerte de Dios, al mismo tiempo que el inicio de una concepción de lo humano apoyada en una visión trans-antropocéntrica, en la que se borraba la barrera y distin-ción esencial entre hombres y animales, en beneficio de una concepción de la biosfera en la que la singularidad del género homo se diluía demo-cráticamente, contemplándolo como una rama más del conjunto de los antropoides.

496
© Universidad de Deusto - ISBN 978-84-9830-383-4
La oposición a las tesis darwinianas ya vimos que provenían no sólo del ámbito religioso conservador, sino también del científico y de amplios sectores de la filosofía idealista e historicista. Resultaba muy difícil re-nunciar a todo tipo de orientación teleológica como modelo explicativo de los procesos mundanos, sea que se atribuyera dicha orientación a Dios, al Espíritu hegeliano o a una Naturaleza divinizada.
Ahora bien, entre los que fueron aceptando las tesis de Ch. Darwin, se da una diferencia clara a la hora de sacar las conclusiones sobre la imagen que se desprendía sobre el ser humano. Por un lado se sitúan los que, des-de un olvido, o mala conjugación, de la diferencia de planos epistemológi-cos entre la ciencia y la filosofía y teología, llegan a la conclusión de que la selección natural suponía la total superación de cualquier tesis antropo-céntrica y humanista, en la medida en que dan por hecho la inevitabilidad de echar por tierra tanto las pretensiones religiosas de entender al hombre como imagen de Dios y creado de modo especial por él, como también las tesis humanistas que, sin necesitar apoyarse en planteamientos religiosos, siguen defendiendo la singularidad humana, desde la pretensión de una mayor densidad ontológica y ética de nuestra especie.
En el extremo opuesto a las tesis cientifistas, se situó desde muy tem-prano el creacionismo fundamentalista, no sólo contrario a la distinción y adecuada conjugación de planos epistemológicos, sino también anclado en una interpretación acrítica de la Biblia, y opositor dogmático de toda afirmación sobre la realidad que no se halle acorde con las afirmaciones literales y atemporales de los textos sagrados.
Frente a estos planteamientos extremos, consideramos que la postu-ra más plausible y mejor fundamentada se sitúa en un punto medio, que trata de conjugar las afirmaciones que nos aportan las ciencias con las diversas y plurales interpretaciones filosóficas, entre las cuales se ha-lla, como más plausible y concordante con el conjunto de los datos an-tropológicos, una visión antropocéntrica y humanista del ser humano, acorde incluso con la idea de considerar al ser humano como imagen de Dios y el centro de la creación. En definitiva, entendemos que la visión más acertada de lo humano consiste en reconocer sin reservas la incor-poración de la especie humana en el proceso evolutivo, asumiendo su pertenencia a la biosfera como una especie más, y, al mismo tiempo, en defender la diferencia cualitativa respecto al resto de las demás especies vivas. Por tanto, la especie humana es una especie animal más, pero lo es de un modo cualitativamente diferente. Se da, por un lado, una conti-nuidad evolutiva con el resto de las especies vivas, pero, al mismo tiem-po, hemos de reconocer también que, como consecuencia del propio proceso evolutivo, ha emergido con lo humano una realidad cualitativa-mente nueva.

497
© Universidad de Deusto - ISBN 978-84-9830-383-4
3. EL HOMBRE, FRUTO DE UN LARGO PROCESO FILOGENÉTICO Y ONTOGENÉTICO
El descubrimiento de las evidencias fósiles que han ido confirmando, de un modo incontestable, nuestra pertenencia al proceso evolutivo, ha sido un proceso lento y laborioso, pero acelerado y ampliamente enrique-cido en las últimas décadas. Era fundamental que las tesis evolucionistas, si querían consolidarse como tesis científicas, se fueran confirmando con las correspondientes evidencias fósiles que avalaran la relación evolutiva gradual entre las diferentes especies vivas, y dentro de ese proceso, como una rama más, el género humano y sus diversas especies.
En esta tarea, le ha correspondido a la paleoantropología ser la cien-cia protagonista encargada de aportar los diferentes eslabones intermedios entre los humanos actuales y los chimpancés, como especie viva más cer-cana a nosotros. Los múltiples y variados descubrimientos obtenidos por los paleoantropólogos han ido poco a poco rellenando los agujeros y las sombras de duda que sobre este punto estaban pendientes. La aceleración de los descubrimientos en las últimas décadas ha sido espectacular, aun-que lo que sabemos sobre la historia evolutiva es todavía más bien insu-ficiente y endeble, aunque no por ello deja de ser muy importante y sig-nificativo. Si a ello se unen las aportaciones de la biología molecular y de otras ciencias complementarias, es justo reconocer que nos hallamos ante evidencias que constituyen una apoyatura incontestable a las pretensiones de cientificidad de la teoría de la selección natural. Otra cosa muy distinta es si dicho proceso evolutivo se tiene que entender según las propuestas gradualistas, tal y como defendía Darwin, en clave rupturista y disconti-nua, o desde un punto de vista intermedio, como defiende la teoría de los equilibrios puntuados1.
Ahora bien, a pesar de los grandes avances de la palaoantropología, no sabemos todavía reconocer ni describir con seguridad las diferentes etapas y eslabones que han existido entre los chimpancés y los huma-nos, en la medida en que desde hace siete o más millones de años en que se produjo esa separación, se han descubierto un amplio abanico de restos fósiles de diferentes especies, englobadas todas ellas hasta hace poco bajo la denominación de australopitecos, habiéndose añadido en la actualidad las especies correspondientes a los ardipitecos y los orro-rin, propuestas que todavía tienen que asentarse y demostrar sus preten-
1 Como complemento a lo que ya dijimos sobre este tema en el cap. 2.º, cfr. Shermer, Michael, Las fronteras de la ciencia. Entre la ortodoxia y la herejía, Barcelona, Alba Edi-torial, 2010, 143-184.

498
© Universidad de Deusto - ISBN 978-84-9830-383-4
siones de situarse en nuestra línea evolutiva. Y de igual modo, también tienen que confirmarse y completarse los datos que apuntan a la especie homo habilis como la primera dentro de las diferentes especies del géne-ro homo, puesto que la frontera entre lo prehumano y lo humano (homo habilis) no está totalmente delimitada y aceptada por todos los investi-gadores.
El estudio de las características de cada una de estas especies interme-dias nos confirma, por tanto, nuestra común pertenencia al proceso evolu-tivo, así como nuestras diferencias dentro del mismo. No es fácil estable-cer las fronteras claras de esa separación y singularidad, en la medida en que un proceso evolutivo como el que las ciencias nos presentan, no ad-mite separaciones bruscas ni fronteras claras. Y menos todavía si ese pro-ceso lo entendemos en clave gradualista. De ahí que resulte más cercana a las evidencias fósiles y más convincente para una correcta interpretación del proceso evolutivo las tesis de los equilibrios puntuados, como veía-mos en el capítulo segundo.
Si la tesis de la continuidad y la diferencia se puede defender desde los datos aportados por el proceso filogenético, también se confirma di-cha perspectiva desde el punto de vista del proceso ontogenético. Hemos visto en el capítulo sexto que la visión filogenética y la ontogenética de la antropogénesis han ido durante mucho tiempo separadas y dándose la es-palda, cosa que no ocurre ya hoy. La ontogénesis y la filogénesis se consi-deran en la actualidad como perspectivas complementarias y mutuamente enriquecedoras. También desde el horizonte de la ontogénesis, advertimos la continuidad de la especie humana con el resto de las especies vivas (te-sis de la recapitulación del Haeckel) así como su ruptura y singularidad (neotenia). La especie humana recapitula el proceso evolutivo anterior, pero para asumirlo y reorientarlo de forma específica. Esto explica nuestra prematuridad y el nacer con una deficiencia biológica significativa, rasgos que nos hace tan frágiles y necesitados de una larga y costosa protección de nuestros progenitores, deficiencia complementada y enriquecida, a su vez, con una amplia plasticidad educativa. En consecuencia, si somos fru-to y hechura de la herencia biológico-genética, en mayor medida lo somos de la herencia cultural.
Somos la especie más plástica y más dependiente de nuestro en-torno social, de modo que la cultura tiene que suplir las deficiencias e insuficiencias de nuestra herencia biológica. Y es precisamente este ha-llarnos en permanente auto-conformación, lo que hace insuficiente para definirnos como humanos una mirada meramente descriptiva y asépti-ca, propia del enfoque científico. Nacemos dotados para y necesitados de proyectar sobre las cosas y sobre nosotros mismos todo tipo de valo-raciones ontológicas y éticas. Los entes mundanos no sólo son, sino que

499
© Universidad de Deusto - ISBN 978-84-9830-383-4
valen. Y en la cima de la escala del conjunto de las realidades valiosas están los seres humanos. Los hombres tienen dignidad y no precio. Es lo que les dota de su condición de personas, como realidades singulares e irrepetibles. Pero queda en discusión en qué momento del proceso on-togenético, embriológico, comienza tal pretensión de poseer ese valor irrepetible, de ser una realidad poseedora de las notas esenciales que le dotan de suficiencia entitativa para existir como sustantividad humana autónoma. En la medida en que proyectamos sobre el embrión humano la posesión de un valor que lo hace irrepetible y digno de respeto y dig-nidad ética, reconocemos la diferencia y distancia cualitativa, tanto en su dimensión ontológica como ética, entre los humanos y el resto de las especies animales.
Independientemente de la postura defendida sobre la importante y difícil cuestión acerca del momento en el que situamos, dentro del pro-ceso embriológico, el inicio de su condición de persona, creemos que los datos aportados por la evolución ontogenética de lo humano tienen entidad suficiente como para defender nuestra tesis central ya repetida sobre la continuidad y ruptura de la especie humana dentro del proceso evolutivo.
4. CONTINUIDAD Y EMERGENCIA CUALITATIVA
Pero tenemos que seguir explicitando las razones que nos permiten defender la ruptura dentro de la continuidad. El conflicto se sitúa entre dos posturas contrarias, la reduccionista y la emergentista. Considera-mos que la postura reduccionista no hace justicia a la complejidad de la realidad, pretendiendo explicar los sistemas complejos desde la estrate-gia de desmenuzarlos en sus partes más simples2. Esa misma estrategia la pone en funcionamiento a la hora de explicar la complejidad y singula-ridad de la realidad humana, bien sea reduciéndola, en el nivel de los genes, a un simple mecanismo de supervivencia, o bien, en el nivel cere-bral, explicando el comportamiento humano como un mero resultado de la complejidad del cerebro, que, a su vez, sería visto simplemente como una acumulación de un número extraordinario de neuronas y de conexio-nes sinápticas, reducibles y explicables cada una de ellas desde el juego
2 Sobre las insuficiencias de la estrategia reduccionista frente a un modelo sistémi-co y emergentista en la relación entre la base genética, proteínica y los sistemas orgá-nicos, cfr. Noble, Denis, La música de la vida. Más allá del genoma humano, Madrid, Akal, 2008.

500
© Universidad de Deusto - ISBN 978-84-9830-383-4
de compuestos químicos, y éstos, a su vez, explicables desde el nivel de los átomos. Por tanto, lo psicológico y comportamental quedaría redu-cido a lo biológico, y este nivel, a lo químico y físico. Desde esa óptica explicativa, carecen de importancia las referencias a los diferentes siste-mas y estructuras con las que se habrían organizado los componentes úl-timos de la materia en su historia evolutiva. De ese modo, no se alcanza a explicar convincentemente las razones de las enormes diferencias en-tre el comportamiento de las diferentes especies vivas.
Tampoco resulta convincente la postura dualista contraria, en la medi-da en que explica la complejidad del comportamiento humano atribuyendo la autoría de ello a un alma inmaterial y separada de lo corpóreo, y estando obligada por ello a tener que explicar cómo interactúan entre sí ambas rea-lidades tan diferentes ontológicamente. La distancia de estos planteamien-tos con los parámetros de la ciencia actual hacen tales explicaciones muy poco plausibles. Y, por otro lado, consideramos que no es necesario soste-ner tesis dualistas para defender planteamientos humanistas, con su consi-guiente afirmación de la singularidad y dignidad de lo humano.
Las serias limitaciones de estas dos posturas señaladas, nos hace de-fender la tesis emergentista como fórmula más adecuada para entender y defender la especificidad de lo humano3. El emergentismo explica el proceso evolutivo complementando el punto de vista analítico del re-duccionismo con una concepción sistémica de la realidad. La compleji-ficación de la vida en el proceso evolutivo no sólo se debe a un aumento cuantitativo de proteínas, células y órganos presentes en las diferentes especies vivas, sino que la explicación más convincente se consigue tomando en serio la complejificación sistémica que se da permanente-mente en el proceso evolutivo y sobre todo en la aparición de especies nuevas. Si una especie nueva, como fórmula novedosa que la vida va creando, se explica en parte desde los componentes que la integran, no es menos verdad que para entenderla en su auténtica realidad hay que acudir a las novedades que aporta la nueva sistematización con la que está organizada. Un sistema nuevo supone la aparición de propiedades nuevas (al menos una), propias y específicas del sistema, que no se ex-plican desde las partes del mismo4.
La nueva sistematización de la especie humana da como resultado la psique humana, su personalidad, el centro subjetivo pensante que ex-
3 Cfr. Beorlegui, C., «Los emergentismos sistémicos: un modelo fructífero para el pro-blema mente-cerebro», Pensamiento, 62 (2006), n.º 234, pp. 391-439; Id., «Emeregentism», Pensamiento, 65 (2009), n.º 246, Serie Especial, n.º 3, pp. 881-914.
4 Cfr. Bunge, M., El problema mente-cerebro. Un enfoque psicobiológico, Madrid, Tec-nos, 1985.

501
© Universidad de Deusto - ISBN 978-84-9830-383-4
plica su comportamiento y sus decisiones autónomas. Así, el ser huma-no tiene en común con los demás seres vivos la materia biológica con la que está constituido, con todas sus propiedades y muchas de las subes-tructuras con las que está organizado y conformado, pero su originali-dad es fruto de una nueva sistematización emergida en el mismo pro-ceso evolutivo. Para explicar dicha emergencia se tiene que recurrir, al igual que en cualquier otra novedad de tal proceso, tanto a explicaciones científicas como filosóficas y, si nos parecen adecuadas, religiosas. Des-de la ciencia podremos advertir que la emergencia de esa nueva siste-matización es fruto de un conjunto acumulado de mutaciones azarosas, que en un momento determinado necesitaron ser estructuradas de otra manera, para ascender de ese modo a un nuevo sistema vital que dio lu-gar a nuestra especie. Y desde una perspectiva filosófica y/o teológica podremos acogernos a diversas interpretaciones contrapuestas: desde ver en ello el mero fruto de concatenaciones azarosas, hasta verlo como resultado de una previsión teleológica, como puede ser la acción provi-dente de Dios, que ha dotado a la naturaleza de las propiedades necesa-rias para que pueda desembocar en la emergencia de un tipo de vida tan complejo como el humano.
Desde el planteamiento propio de los emergentismos sistémicos, se advierte también la insuficiencia y esterilidad de las discusiones negado-ras de la especificidad de lo humano apoyadas en investigaciones que se empeñan en descubrir cualidades y comportamientos animales muy simi-lares a las humanas. Dicha estrategia no llega más que a mostrar algo que resulta lógico y aceptable: la familiaridad de lo humano con lo animal, y la cercanía de los comportamientos de los primates con los humanos, puesto que se trata de especies cercanas a la nuestra, y, por tanto, consti-tuyen sistemas vitales que la evolución se dotó en fases previas a lo hu-mano. Pero, junto a ese parecido, la nueva sistematización biológica de la especie humana nos ha dotado de un modo cualitativamente nuevo de encarar la relación con el entorno ecológico y las relaciones sociales con el resto de los componentes de nuestra especie.
Consideramos, pues, que desde la propuesta emergentista se puede entender mejor la conjunción de continuidad y ruptura tanto en el proceso evolutivo, en general (como salto de la materia no viva a la vida, y las su-cesivas complejificaciones de la misma), como en el proceso antropoge-nético de la hominización (en sus dos vertientes, filogenética y ontogené-tica), e igualmente en la específica conjugación de biología y cultura que constituye la realidad de la especie humana.
El proceso evolutivo presenta un encadenamiento entre especies en el que se da tanto la continuidad como la ruptura en cada una de las nuevas especies que van apareciendo. Vimos más arriba que no parece suficiente

502
© Universidad de Deusto - ISBN 978-84-9830-383-4
la estrategia reduccionista a la hora de dar cuenta de las peculiaridades del mundo de la biología, puesto que a esa estrategia reduccionista hay que añadir el aspecto de complejificación de la materia viva, conformada por procesos teleonómicos irreductibles a una simple visión física y química. La capacidad de los seres vivos de orientar sus dinamismos desde sí mis-mos hacia objetivos propuestos, aun sin conciencia refleja de ello, supone aceptar la especial estructuración y sistematización de su forma de vida. La estructura y el sistema nuevo emergido en esa nueva estrategia vital de la naturaleza supone que ese sistema advenido posee alguna propiedad que no se puede explicar con la sola referencia a las diferentes partes que componen el nuevo sistema.
De ahí que resulte una estrategia equivocada, a la hora de negar la di-ferencia cualitativa entre hombres y animales, aportar simplemente datos y más datos, investigaciones tras otras, para intentar borrar la frontera di-visoria entre los hombres y el resto de los animales. Desde la evidencia de la continuidad, y desde las tesis gradualistas de este proceso evoluti-vo, siempre encontraremos eslabones intermedios que apunten a estruc-turas genéticas, componentes morfológicos y estrategias comportamen-tales muy similares a las propias de la especie humana. En esa dirección se mueven los textos que nos presentan abundantes y meticulosas pruebas de los parecidos asombrosos entre los humanos y las especies animales más cercanas a nosotros, como son los primates5, incluso en aspectos tan sensibles como la autoconciencia, los comportamientos sociales o la ca-pacidad ética6. Pero el parecido y cercanía se rompen cuando se hace la comparación desde el conjunto completo de los diferentes aspectos y di-mensiones, llevándonos siempre a la evidencia de que la estructura vital y comportamental humana se sitúa en un nivel cualitativamente superior al del resto de los animales.
Pero también es importante resaltar que tanto nuestra postura que de-fiende la diferencia cualitativa como la contraria, son sólo plausibles y convincentes desde paradigmas o cosmovisiones previas y aprióricas, que sostienen ambos planteamientos y dotan a los datos empíricos de fuerza y consistencia. Se da, pues, un círculo hermenéutico entre los datos y el ho-rizonte interpretativo previo. Y como ocurre en muchos aspectos sobre los que discute la filosofía, el círculo hermenéutico entre datos e interpreta-
5 En su momento ya aportamos bibliografía abundante sobre estos puntos, pero es in-teresante referirse a investigaciones más recientes que confirman esta estrategia investiga-dora: cfr. Guillén-Salazar, Federico (Ed.), Existo, luego pienso. Los primates y la evolución de la inteligencia humana, Madrid, Ateles Editores, 2005.
6 Cfr. Bekoff, Marc/Pierce, Jessica, Justicia salvaje. La vida moral de los animales,Madrid, Turner Publicaciones, 2010.

503
© Universidad de Deusto - ISBN 978-84-9830-383-4
ción no es neutro, sino que supone un compromiso vital y ético de adoptar previamente una postura determinada de aceptación o de rechazo del hori-zonte de interpretación cosmovisional.
Si nos adentramos ahora en el aspecto ontogenético de la antropo-génesis, creemos que ninguna estrategia comparativa podrá negar, en relación a los aspectos que pone en cuestión la postura reduccionista e igualitarista, la especial tendencia en el proceso embrionario de la espe-cie humana hacia la neotenia y el nacimiento prematuro, que nos dota de una condición inmadura, deficiente y necesitada de una ayuda más prolongada de los progenitores y del entorno familiar y social. Este dato es fundamental a la hora de advertir el salto que se da con la especie hu-mana de una estrategia biológica de mayor dependencia entre lo genéti-co y lo conductual a otra fórmula vital en la que la conducta se suelta y se desprende de los automatismos biológicos. La relación entre genética y conducta es en cierto modo similar, aunque muy diferente. Si en el caso de las demás especies animales la relación es de determinación, o de mayor control de lo comportamental por lo genético, en el caso hu-mano la relación entre biología y conducta es distinta: la biología es la condición de posibilidad de la conducta, pero le deja a cada individuo suficiente autonomía como para no depender de los automatismos bio-lógicos, no teniendo otro remedio que pensar y decidir sus actuaciones en libertad. Si en el conjunto de la biosfera los animales están determi-nados a un tipo de conducta cerrada, en el caso humano estamos de-terminados por naturaleza a ser libres y autónomos, a no depender de un modo cerrado de los constreñimientos genético-biológicos. En eso consiste nuestra singularidad, fórmula vital en la que ninguna especie animal nos acompaña, aunque se hallen atisbos y anticipaciones en al-gunos primates. Es importante resaltar que, aunque se abogue por diluir la frontera entre hombre y animales, ninguno de los autores que abogan por los derechos de los animales y la atribución a algunos de ellos de la categoría de persona llega a atribuirles libertad y, por tanto, la necesidad de considerarles responsables de sus acciones.
Lo que constituye nuestra singularidad es consecuencia de la estruc-tura comportamental que poseemos: no nos hallamos determinados por orientaciones rígidas de la genética y la biología, sino por la necesidad de tener que elegir entre diversas opciones de actuación. Nuestra vida está regida por el continuo juego de ensayo-error, en el terreno de la supervi-vencia, y, en el ámbito moral, por la libertad de acción que conlleva la inevitable responsabilidad de nuestros comportamientos.
Vemos, por tanto, que lo que nos hace diferentes y singulares den-tro del mundo de los seres vivos es sobre todo la autoconciencia y la li-bertad. Los humanos no sólo poseemos, como el resto de los animales,

504
© Universidad de Deusto - ISBN 978-84-9830-383-4
una unidad vital interior que unifica las acciones y las guía hacia un objetivo determinado (teleonomía), teniendo un cierto grado de mis-midad (el animal es un autos), sino que somos conscientes de ello, en un grado reduplicado de conciencia. Es una mismidad de segundo gra-do. Y eso es lo que nos permite hacernos preguntas sobre nuestro ser y el de los demás humanos, así como del sentido de la historia y del uni-verso entero. Esa apertura al sentido de la realidad y a la búsqueda de su fundamento es algo que nos constituye y nos conforma como seres especiales, cualidad que no es una mera cuantificación de la capacidad que tienen los animales de evaluar su entorno natural y defenderse de él en su lucha por la supervivencia, sino que constituye una capacidad que constituye en sí misma una diferencia cualitativa. Los animales no han salido del universo biológico, mientras que los humanos somos anima-les culturales.
5. EL ANIMAL BIO-CULTURAL
La condición de animales bio-culturales que somos los humanos puede entenderse de múltiples formas, como hemos visto en el capítu-lo ocho. Desde reducir lo cultural a un simple apartado o rasgo biológico, a subordinarlo a las leyes e intereses de lo biológico, o considerarlo como un simple peldaño, posteriormente desechado, para llegar al nivel cultural. Ninguna de estas tres fórmulas hace justicia a nuestro modo de ser.
La fórmula que mejor describe nuestra condición es la que integra en una estructura única e indivisible lo biológico y lo cultural. Es verdad que la cultura, en la medida en que supone el advenimiento de la autoconcien-cia y la autonomía para orientar nuestras acciones e ir completando nues-tra realización personal, supone una autonomía y dominio sobre lo bioló-gico, pero este aspecto sigue siendo una parte integrante e indispensable del conjunto de la estructura humana.
El sujeto de la autoconciencia y de la libertad es la totalidad del in-dividuo, no una sola parte del mismo, la espiritual, el alma, que estaría añadida al cuerpo, y cuya única relación con él sería necesitarlo como instrumento o mediación para existir y actuar. Las notas o rasgos bioló-gicos forman parte esencial de la persona humana. Somos psique soma-tizada y cuerpo psiquizado, o espíritu encarnado y carne espiritualizada, si lo queremos expresar de esta forma. La relación entre lo psíquico y lo corpóreo no hay que entenderla en clave de dos realidades ontológica-mente autosuficientes, sino como la relación entre dos dimensiones de

505
© Universidad de Deusto - ISBN 978-84-9830-383-4
una única realidad, o como la relación entre el sistema integrador y las diferentes partes del sistema. Ni los genes, ni las proteínas, ni el cerebro actúan o piensan. El que piensa es el yo, esto es, el sistema completo de cada individuo, pero no al margen de su condición biológica y psíquica, sino integrándola como elemento esencial del todo que cada uno de no-sotros somos.
Desde este punto de vista, volvemos de nuevo a visualizar la con-tinuidad junto a la ruptura de la realidad humana respecto al resto de las especies animales. Somos animales que sentimos y percibimos la realidad, esto es, que se nos da lo que hay al modo impresivo, a través de nuestros sentidos. Pero el sentir humano está elevado a un nivel su-perior, puesto que es un sentir inteligente. Esto es, se trata de que las sensaciones impresivas, que en el nivel animal se dan, como dice Zu-biri, bajo la formalidad de estimulidad (se da algo como fuente de estí-mulos), en los humanos se nos dan bajo la formalidad de realidad, esto es, como realidades, como estímulos reales. En definitiva, cada realidad humana, cada individuo, es una única estructura de notas, unas de tipo somático y otras de tipo psíquico, que conforman una sustantividad con suficiencia para existir, y que le dotan al individuo de una peculiar con-sistencia ontológica a la vez que de una específica estructura compor-tamental (habitud) para llegar a ser persona, dotado de autoconciencia (suidad) y libertad. Se advierte, por tanto, la continuidad con la anima-lidad, pero al mismo tiempo el salto cualitativo hacia un nuevo modo de ser y de conformar lo animal. Para la antropología griega se trata de un animal racional; para el planteamiento zubiriano, es un animal de reali-dades, un animal vertido a la realidad, a la formalidad de realidad o de reidad. Esa formalidad de realidad es la que hace que el ser humano se sitúe a un nivel cualitativamente distinto de la pura animalidad, del puro sentir. Las especies cercanas a la humana poseen atisbos y barruntos de ese nuevo nivel que emerge con la especie humana, pero sus carencias les hace situarse todavía en el peldaño anterior, advirtiéndose que la dis-tancia no es de mera cantidad de cualidades, sino de una distancia de grado, cualitativa.
La nueva fórmula que la vida se ha inventado o ha creado en el caso de los humanos, es por tanto una originalidad que se ha conformado con los ingredientes ya aparecidos en los niveles anteriores de la evolución. Pero ello no debe suponer menospreciar el valor, el respeto y el cuida-do que debemos tener respecto al resto de los animales, en la medida en que así lo requiere su complejidad biológica y ontológica. Podemos y de-bemos defender la necesidad de cuidar al conjunto de la biosfera, y del conjunto de la ecosfera, del modo que resulte más adecuado, así como debemos establecer leyes que prohíban el maltrato a los animales y la in-

506
© Universidad de Deusto - ISBN 978-84-9830-383-4
adecuada utilización de los mismos para investigaciones científicas, sin dejar por ello de seguir defendiendo la diferencia ontológica y ética entre hombres y animales, así como seguir manteniendo una visión humanista y antropocéntrica de la realidad.
6. LA LEGITIMIDAD DEL HUMANISMO ANTROPOCÉNTRICO
La defensa de las tesis humanistas y antropocéntricas no supone una incorrecta supravaloración de lo humano, ni tampoco implica menospre-ciar el valor de los animales, en la medida en que la tesis humanista y an-tropocéntrica no implica defender el dominio despótico e irresponsable del hombre sobre la biosfera y la ecosfera, sino que más bien conlleva la obligación de mantenerla y protegerla de forma responsable7. No sólo porque si la daña o destruye se daña a sí mismo y pone en riesgo su pro-pia existencia y la supervivencia de las generaciones futuras, sino tam-bién porque destruye realidades valiosas y dignas de ser respetadas y va-loradas.
Esta singularidad de lo humano posee no sólo relevancia en el terre-no ontológico y ético, sino también en el social y político. La singulari-dad humana implica una densidad ontológica específica, que no rebaja el valor ontológico del resto del universo en su compleja y rica variedad de entes y de componentes vivos, sino que los ordena en función de su com-plejidad y riqueza entitativa. Claro que estas afirmaciones, como ya lo he-mos indicado más arriba, no se hacen desde el mero nivel científico. La ciencia no puede hacer estas valoraciones, teniendo que reconocer que, si nos referimos sólo a los componentes naturales de las diferentes realida-des que se dan en el cosmos, todas valen igual y están compuestas en úl-timo término por los mismos átomos y componentes básicos. La apuesta por una valoración ontológica superior para lo humano se hace desde un horizonte filosófico concreto, desde una cosmovisión antropocéntrica que, aunque se apoya en la complejidad objetiva y científica que las ciencias nos aportan sobre la realidad y sobre lo humano, se sitúa más allá y por encima de ella.
7 Cfr. Jonas, Hans, El principio responsabilidad. Ensayo de una ética para la civili-zación tecnológica, Barcelona, Herder, 1995; Gafo, Javier (dir.), Diez palabras clave en Ecología, Estella (Navarra), Verbo Divino, 1999; Beorlegui, Carlos, «Ética animal e idea de persona», en Guibert, J. M./Orella Unzué, J. L. (eds.), Preguntas éticas en cuestiones dispu tadas hoy, San Sebastián, Universidad de Deusto, 2005, 317-335.

507
© Universidad de Deusto - ISBN 978-84-9830-383-4
Este mismo enfoque cosmovisional es el que atribuye también una mayor densidad ética a los integrantes de la especie humana8. Los datos científicos y objetivos son un apoyo necesario para defender esta densi-dad ética que nos hacen más valiosos que cualquier otra realidad munda-na, pero eso no evita que otras interpretaciones nos nivelen con los de-más animales, e incluso realidades materiales, en el terreno de la ética. Aunque esta interpretación tiene que justificar por qué y qué sentido tie-ne, cuando hablamos de ética, igualar en dignidad a quienes no tienen ca-pacidad y sensibilidad ética y a quienes sí la tienen.
La negación de esta diferencia ética cualitativa nos plantea difíciles problemas a resolver y consecuencias también de no fácil solución. Si no aceptamos la singularidad humana en el terreno de la dignidad ética y de la exclusiva capacidad de responsabilidad de nuestras acciones, tendría-mos que revisar, si no echar abajo, la totalidad del ordenamiento legal y ético de las sociedades humanas. Si defendemos que los animales son tan personas como nosotros, habría que incluirlos como sujetos de derecho, con todo tipo de derechos y también de deberes. Y si la igualdad la enten-demos hacia abajo, es decir, que también al ser humano le negamos su li-bertad y su capacidad ética, tendríamos que negar, por sin sentido, todo el ordenamiento legal, por innecesario. Nada sería susceptible de catalogar como malo y reprobable, y, por tanto, todo lo que constituye la sociedad humana sería consecuencia de factores neutros, amorales, y resultado de leyes biológicas autónomas, o de puro azar.
Las implicaciones sociopolíticas de esta forma de pensar son eviden-tes. Ninguna institución ni componente de la sociedad humana se podría evaluar como negativo o positivo, sino un mero accidente de la natura-leza, como son los mares, las montañas, los fenómenos atmosféricos y otros elementos de la naturaleza. No tendría sentido hablar de socieda-des injustas, de decisiones políticas inmorales, etc. Todo sería fruto del azar, sin posibilidad de poder cambiarlo por otras estructuras diferentes, y por responsables que empujaran el desarrollo y organización de las so-ciedades de otra manera. No tendría sentido hablar en términos morales, en la medida en que determinismo y ética son incompatibles. Si se niega la libertad, tenemos que dejar la ética de lado, o considerar los concep-tos morales como algo ilusorio y metafórico pero sin consistencia real9.
8 Cfr. Cortina, Adela, Etica aplicada y democracia radical, Madrid, Tecnos, 1993; Id, Las fronteras de las persona. El valor de los animales, la dignidad de los humanos, Ma-drid, Taurus, 2009; Gómez Pin, Víctor, El hombre, un animal singular, Madrid, La Esfera de los Libros, 2005.
9 Las reflexiones de I. Berlin, en Libertad y necesidad en la historia, Madrid, Revista de Occidente, 1958, y Sobre la libertad, Madrid, Alianza, 2004, son acertadas y definitivas.

508
© Universidad de Deusto - ISBN 978-84-9830-383-4
Sólo aceptando la libertad se entiende y tiene sentido el mundo de los humanos, las críticas a las situaciones injustas e inhumanas y la apela-ción a la responsabilidad para seguir manteniendo y construyendo un mundo digno de los humanos.
Desde estas convicciones es como se advierte la debilidad de los que niegan la singularidad humana y las tesis humanistas, tachándolas de es-pecieísmo, como es el caso de Peter Singer y los demás defensores de los derechos de los animales e incluso la pertinencia de atribuirles la cate-goría de personas a algunos de ellos10.
7. ¿SOMOS EL RESULTADO DEL PURO AZAR, O PODEMOS APOSTAR POR UN FUNDAMENTO ÚLTIMO Y PROVIDENTE DEL UNIVERSO?
La cuestión de la apuesta por la dignidad y singularidad humana está íntimamente relacionada con la pregunta sobre el sentido último de la vida y del universo. Parece tener poca consistencia, o quedar en entredicho, la defensa de la dignidad humana y considerar, al mismo tiempo, que el mun-do es un sinsentido, que somos un paréntesis entre dos nadas (Sartre). Nos parece que ambas tesis, la dignidad humana y la afirmación de un sentido racional de la realidad, están coherente e íntimamente entrelazados.
Hemos de ser conscientes de que en esta reflexión nos estamos mo-viendo siempre en el ámbito de las propuestas filosóficas, donde se sitúan los empeños por dar valor y sentido a las cosas. Desde el nivel científico, no podemos ir más allá que constatar hechos fácticos y leyes físico-quími-cas que se suceden a lo largo del proceso evolutivo. Estas leyes y avances científicos no las podemos olvidar. Es ahí donde se sitúan las diversas pro-puestas y disputas entre las distintas teorías científicas en su empeño por llegar a descubrir y controlar las leyes y estructuras más convincentes con las que se halla conformada la realidad.
Pero la ciencia encuentra límites inevitables en su empeño por dar cuenta de los interrogantes que la realidad nos plantea. Y ahí es donde emerge la filosofía, que busca no tanto saber cómo funciona la realidad, sino por qué es así y no de otra forma, o para qué existe el mundo en vez de no existir. Son las preguntas en las que tradicionalmente ha asentado la
10 Véanse los diferentes libros citados de P. Singer, en la Bibliografía final. Cfr. Torral-ba Roselló, Fr., ¿Qué es la dignidad humana? Ensayo sobre Peter Singer, Hugo Tristram Engelhardt y John Harris, o.c.; Beorlegui, C., «¿Hacia un humanismo trans-antropocéntri-co? Peter Singer y los “derechos” de los animales», o.c.

509
© Universidad de Deusto - ISBN 978-84-9830-383-4
filosofía la especificidad de su tarea inquisitiva11. Ya vimos que, frente al nivel científico, donde es más factible hallar contrastaciones empíricas en-tre teorías rivales, el ámbito de la filosofía se mueve en un mundo de ma-yor dificultad de comprobación de consistencia y verdad de las diferentes propuestas de sentido. De ahí que sean legítimas y plausibles propuestas contrarias sobre la interpretación finalística de la vida y de la realidad. Si desde el nivel científico la realidad se nos aparece entrelazada con conca-tenaciones causales en las que no cabe más que el determinismo cerrado o el mundo abierto de la probabilidad, en el terreno de la filosofía cabe postular tanto una interpretación azarosa como otra dirigida por un senti-do teleológico. Aunque dentro de esta segunda, tendremos que distinguir siempre entre una teleología en manos de los seres humanos, que orientan la historia a partir del entrelazamiento de sus decisiones históricas, sin que quepa un sentido impuesto a la historia sino el resultado de las decisiones humanas.
Pero también resulta coherente preguntarnos, desde el análisis de la historia del universo, y apoyados en los diversos modos de entender el principio antrópico (capítulo 2.º), si la aparición de una vida inteligente como la humana en un pequeño planeta de nuestro sistema solar, fruto de un ajuste fino que lo ha permitido (principio antrópico débil), es sólo con-secuencia de una combinación azarosa de circunstancias, o resulta legíti-mo y razonable interpretar que el mundo está diseñado intencionalmen-te de tal modo que permitiera la existencia de seres inteligentes y libres como nosotros, como sostiene la cosmovisión religiosa. Volvemos a tener que hacer hincapié que ambas opciones se mueven más allá del terreno científico, en el ámbito filosófico. No tienen más apoyo que una apuesta de sentido, razonables ambas propuestas, pero no demostrables. Ambas se sitúan en el ámbito de la fe, tanto religiosa como epistemológica. Pero no se puede ir más allá. Pero así es como funciona la racionalidad humana.
Desde nuestro punto de vista, cabe y es legítimo advertir, más allá del nivel del entrelazamiento de los factores causales de la materia viva, una fuerza teleológica que da sentido al proceso evolutivo, orientándolo des-de el origen de la vida, pasando por su complejificación desde las formas unicelulares a las pluricelulares, hasta llegar a la especie humana. Se trata de un proceso tan complejo, lleno de tantas maravillas, que resulta plau-sible y legítimo ver en él y apostar por la presencia fundamentadora, pero también respetuosa, de la acción de Dios. Esta propuesta, aparte de situar-se en el terreno de la fe y de la apuesta de sentido, tiene el reto de hacer
11 S. Hawking, en su último libro (escrito en colaboración con L. Mlodinow) El gran diseño, Barcelona, Crítica, 2010, pretende hallar el sentido último de todo desde sólo la perspectiva científica, en la medida en que, según él, «la filosofía ha muerto» (p. 11).

510
© Universidad de Deusto - ISBN 978-84-9830-383-4
plausible el modo de proponer y de entender la acción de Dios dentro del universo. Por ahí discurren las diversas e interesantes propuestas teológi-cas que tratan de reconstruir una visión renovada de Dios y de los diver-sos capítulos de la teología12.
8. CONCLUYENDO
En definitiva, la propuesta antropológica que hemos intentado presentar a lo largo de este libro se conforma con una idea del ser humano que enrai-zado en el proceso evolutivo, y tras un largo proceso de antropogénesis, en el cual se entrelazan estrechamente la dimensión hominizadora y la humani-zadora, ha emergido como una especie singular, conformada por una estruc-tura unitaria en la que converge lo biológico y lo psíquico y cultural.
Esa nueva estructura o sistematización de lo biológico, emergida como consecuencia del proceso evolutivo, hace que los humanos nos vea-mos, en un todo inseparable, tanto como un capítulo más de la evolución, como una especie singular e irrepetible. Esa nueva fórmula vital que no-sotros representamos, es la que nos ha permitido superar los automatis-mos biológicos y encomendar la terminación de nuestra autorrealización a las decisiones libres de nuestra subjetividad, en un entrelazamiento con los demás sujetos de la especie humana.
Nuestra dimensión psíquica e intelectual, como estructura dinámica de lo corpóreo, es la que nos dota de autoconciencia y de libertad, base de nuestra capacidad ética, pero también base de nuestra capacidad de apro-piarnos de las diversas posibilidades que la realidad nos muestra de cara a realizarnos a lo largo de nuestro devenir histórico. De este modo, tanto por el conjunto de elementos que constituyen nuestra constitución genéti-ca como también por la especial e irrepetible concatenación de decisiones libres con las que damos forma y concreción a nuestra personalidad, se hace evidente en qué medida la especie humana es singular e irrepetible, pero también lo somos cada uno de los individuos que la componemos.
12 Entre los numerosos intentos de conjugar los avances de las ciencias con una nue-va teología, cfr. Polkinhorne, J. (ed.), La obra del amor. La creación como kénosis, Estella (Navarra), Verbo Divino, 2008; Edwards, D., El Dios de la evolución. Hacia una teología trinitaria, Santander, Sal Terrae, 2006; Haught, J. F., Cristianismo y ciencia. Hacia una teología de la naturaleza, Santander, Sal Terrae, 2009; Barbour, I., Religión y ciencia, o.c.

511
© Universidad de Deusto - ISBN 978-84-9830-383-4
Bibliografía
ABEL, Fr./BONE, E./HARVEY, J. C. (dirs.), La vida humana: origen y desarrollo,Madrid/Barcelona, UPCO/Instituto Borja de Bioética, 1989.
ABEL, F./CAÑON, C. (eds.), La mediación de la filosofía en la construcción de la bioética, Madrid, UPCO, 1993.
ABRISQUETA, J. A., «El embrión humano: estatuto antropológico y ético», en VI-DAL, M. (dir.), Conceptos fundamentales de ética teológica, Madrid, Trotta, 1992, 439-455.
AGUSTÍ, Jordi, El ajedrez de la vida. Una reflexión sobre la idea de progreso en la evolución, Barcelona, Crítica, 2009.
AGUSTÍ, Jordi/LORDKIPANIDZE, David, Del Turkana al Cáucaso. La evolución de los primeros pobladores de Europa, Barcelona, RBA Libros, 2005.
ALBA, D. M./MOYÁ SOLÁ, A./KÖHLER, Meike, «El origen de la mano humana», Investigación y Ciencia, 2005, n.º 341, 46-53.
ALBERTOS, Jesús E., El mal en la filosofía de la voluntad de Paul Rocoeur, Pam-plona, EUNSA, 2008.
ALCAY, Carlos Lázaro, «Luigi Luca Cavalli-Sforza: Genealogía de la humani-dad», Mundo Científico, 1994, n.º 144, 232-239.
ALEMANY, C./GARCÍA, V. (eds.), El cuerpo vivenciado y analizado, Bilbao, Des-clée de Brouwer, 1996.
ALEMAÑ BERENGER, Rafael, Evolución y creación. Entre la ciencia y la creencia,Barcelona, Ariel, 1996.
ALLEN, J. S./BRUSS, J./DAMASIO, Hanna, «Estructura del cerebro humano», Inves-tigación y Ciencia, 2005, n.º 340, 68-75.
ALLMAN, J. M., El cerebro en evolución, Barcelona, Ariel, 2003.ALONSO BEDATE, Carlos, «Reflexiones sobre cuestiones de vida y muerte: Hacia
un nuevo paradigma de comprensión del valor ético de la entidad biológica humana en desarrollo», en ABEL, Fr./BONE, E./HARVEY, J. C. (dirs.), La vida humana: origen y desarrollo, Madrid, UPCO/Barcelona, Instituto Borja de Bioética, 1989, 57-81.

512
© Universidad de Deusto - ISBN 978-84-9830-383-4
—, «El estatuto ético del embrión humano: una reflexión ante propuestas alter-nativas», en MAYOR ZARAGOZA, F./ALONSO BEDATE, C. (coords.), Gen-Etica,Barcelona, 2003, 19-66.
ALONSO, J. M., Introducción al principio antrópico, Madrid, Ediciones Encuen-tro, 1989.
ALPER, Matthew, Dios está en el cerebro, Barcelona, Granica, 2008.ANGIER, Natalie, «La ciencia explica por qué los bebés nacen de espaldas y des-
validos tras nueve meses», El País, 16 de septiembre de 1996, p. 30.ARANA, Juan (ed.), «La filosofía de los científicos», Sevilla, Thémata. Revista de
Filosofía, 1995, n.º 14.ARCHER, L., La amenaza de la biología, Madrid, Pirámide, 1983.ARDREY, R., La evolución del hombre: la hipótesis del cazador, Madrid, Alianza,
1983 (3.ª ed.).ARENSBURG, B./TILIER, A. M., «El lenguaje del hombre de neanderthal», Mundo
Científico, 1990, n.º 107, nov., 1.144-1.146.ARMSTRONG, Karen, En defensa de Dios. El sentido de la religión, Barcelona,
2009.ARRANZ, M., «¿Un nuevo mecanismo de especiación? En diálogo con Faustino
Cordón», Cuadernos Salmantinos de Filosofía, X (1983), 283-292.—, «La antropología de Konrad Lorenz», en LUCAS, J. de S. (dir.), Nuevas antro-
pologías del siglo XX, Salamanca, Sígueme, 1994, pp. 43-78.ARREGUI, J. V. (ed.), «Debate sobre las antropologías», Thémata. Revista de Filo-
sofía (Sevilla), 2005, n.º 35, 139-150.ARSUAGA, J. L., «Los hombres fósiles de la sierra de Atapuerca», Mundo Cientí-
fico, 1994, n.º 143, pp. 167-168.—, «Canibalismo prehistórico», El País, 11-XII-1996, p. 36.—, «Atapuerca y el origen del hombre», Mundo Científico, 1996, n.º 170,
p. 601.—, «Homo Antecessor», Mundo Científico, 1997, n.º 181, pp. 649 y 651.— y MARTÍNEZ, I., La especie elegida, Madrid, Temas de Hoy, 1998.—, El collar del neandertal, Madrid, Temas de Hoy, 1999.—, «El último neandertal», El País, 22 de diciembre de 1999, p. 47.— y otros, «Los homínidos en la hipótesis de Valverde», Mundo Científico,
2000, n.º 225, pp. 58-63.—, El enigma de la esfinge, Barcelona, Plaza y Janés, 2001.—, Los aborígenes. La alimentación en la evolución humana, Barcelona, RBA
Libros, 2002.—, «Un objeto simbólico», El País, 8 de enero de 2003, p. 26.ASFAW, B./WHITE, T. et al., «Australopithecus garhi: A new Species of Early Ho-
minid from Ethiopia», Science, 1999, 284, 629-635.ATLAN, Anne, «La guerra fría de los cromosomas sexuales», Mundo Científico,
1998, n.º 189, 32-34.AUNGER, Robert (ed.), Darwinizing Culture: The Status of Memetics a a Science,
Oxford, Oxford Univsersity Press, 2000 (prólogo de D. Denett).AYALA, Fco. J., «Biology as an Autonomous Science», American Scientist, 1968,
n.º 56, pp. 207-221 (trad. cast., «La biología como una ciencia autónoma», en

513
© Universidad de Deusto - ISBN 978-84-9830-383-4
AYALA, F. J., La evolución de un evolucionista. Escritos seleccionados (Va-lencia, Universidad de Valencia, 2006), pp. 105-122.
—, Origen y evolución del hombre, Madrid, Alianza, 1980.—, «El prodigio de la evolución», Muy Interesante, 1996, n.º 27, 16-21.—, «El mito de Eva y la evolución humana», en VV.AA, «Innovación y cambio.
Hacia una nueva sociedad», Bilbao, Univ. de Deusto, 1997, vol. II, 121-137.—, La teoría de la evolución. De Darwin a los últimos avances de la genética,
Madrid, Temas de Hoy, 1999.—, «¿Desde África? Una perspectiva de los elementos poblacionales en la apari-
ción de los seres humanos modernos», Ludus Vitalis, VIII (2000), n.º 13, 135-156.
— y CELA CONDE, C., Senderos de la evolución humana, Madrid, Alianza, 2001.—, La evolución de un evolucionista. Escritos seleccionados, Valencia, Universi-
dad de Valencia, 2006, p. 112.— y CELA CONDE, C. J., La piedra que se volvió palabra, Madrid, Alianza, 2006.—, Darwin y el Diseño Inteligente. Creacionismo, cristianismo y evolución,, Ma-
drid, Alianza, 2007.—, «Ensayo sobre las bases biológicas del comportamiento moral», Estudios Fi-
losóficos, LVII (2008), n.º 165, 225-246.BAHN, P. G., «Neanderthal emancipated», Nature, 1998, 394, 719-720.BARAHONA, A./AYALA, Fr. J., El siglo de los genes. Patrones de explicación en
genética, Madrid, Alianza, 2009.BARASH, David P./LIPTON, J. E., El mito de la monogamia. La fidelidad y las infi-
delidades en los animales y en las personas, Madrid, Siglo XXI, 2001.BARBOUR, I. G., Problemas de religión y ciencia, Santander, Sal Terrae, 1972.—, Religión y ciencia, Madrid, Trotta, 2004.—, El encuentro entre ciencia y religión. ¿Rivales, desconocidas o compañeras
de viaje?, Santander, Sal Terrae, 2004.BARROSO, Óscar, Verdad y acción, Granada, Comares, 2002, pp. 21 y ss.BARROW, J. D./TIPLER, F. J., The Anthropic Cosmological Principle, Oxford, Cla-
rendon Press, 1986.GALE, G., «A revised Design: Teleology and Big Questions in Contemporary
Cosmology», en Biology and Philosophy, 2 (1987), 475-492 (trad. esp. en In-vestigación y Ciencia, 1982, n.º 65).
BAUR, M./ZIEGLER, G., La aventura del hombre. Todo empezó en África, Barce-lona, Maeva, 2003, pp. 27-29.
BECK, B. B., Animal tool behaviour: The use and manufacture of tools by ani-mals, Garland Publishing, 1980.
BECHTEL, W., Filosofía de la mente, Madrid, Tecnos, 1991.BEGUN, David R., «The Earliest Hominist. Ist Less More?», Sciencie, vol. 303,
n.º 5663, 5 de marzo de 2004, pp. 1.478-1.480.BEHE, M., Darwin’s Box. The Biochemical Challange to Evolution, N. York,
The Free Press, 1996 (trad. cast.: La caja negra de Darwin. El reto de la bio-química a la evolución, Madrid, Edit. Andrés Bello, 2000).
BEKOFF, Marc/PIERCE, Jessica, Justicia salvaje. La vida moral de los animales,Madrid, Turner Publicaciones, 2010.

514
© Universidad de Deusto - ISBN 978-84-9830-383-4
BEORLEGUI, C., «El reto de la biología a la antropología: De la etología a la socio-biología», Letras de Deusto, 16 (1986), n.º 34, 37-69.
—, «La manipulación de la biología. Función ideológica del determinismo bioló-gico», Zientziartekoak (San Sebastián), I (1986), n.º 2, pp. 71-107.
—, »Las Leyes Epigenéticas: ¿el eslabón perdido entre la biología y la cultura?», Zientziartekoak (San Sebastián), II (1987), n.º 2, 102-123.
—, «De la autonomía a la alteridad. Hacia una nueva racionalidad», Letras de Deusto, 17 (1987), n.º 38, 41-61.
—, «El hombre y sus imágenes. Del narcisismo a la alteridad», Letras de Deusto,22 (1992), n.º 52, 81-108.
—, Antropología filosófica. Nosotros: urdimbre solidaria y responsable, Bilbao, Universidad de Deusto, 1999, 2004 (2.ª); 2009 (3.ª ed.).
—, «¿Hacia un humanismo trans-antropocéntrico? Peter Singer y los “derechos” de los animales», Realidad (UCA, El Salvador), 2001, n.º 80, 199-236.
—, «Humanismo y bioética (Un nuevo planteamiento del estatuto del embrión desde un nuevo concepto de persona)», Bilbao, Universidad de Deusto (Con-greso sobre Humanismo), 2003, publicado en CD.
—, «A vueltas con la naturaleza humana», en ARREGUI, J. V. (ed.), Debate sobre las antropologías, Thémata. Revista de Filosofía (Sevilla), 2005, n.º 35, 139-150.
—, «Etica animal e idea de persona», en GUIBERT UCIN, J M./ORELLA UNZUÉ, J. L. (eds.), Preguntas éticas en cuestiones disputadas hoy, San Sebastián, Uni-versidad de Deusto, 2005, 317-335.
—, «La capacidad lingüística del ser humano: una diferencia cualitativa», Thé-mata (Sevilla), 2006, n.º 37, 139-168.
—, «La capacidad lingüística del ser humano: una diferencia cualitativa», Thé-mata. Revista de Filosofía, 2006, n.º 37, 139-168.
—, «Los emergentismos sistémicos: un modelo fructífero para el problema mente-cuerpo», Pensamiento, 62 (2006), n.º 234, pp. 391-438.
—, «Filosofía de la mente: visión panorámica y situación actual», Realidad, 2007, n.º 111, 121-160.
—, «La singularidad del ser humano como animal bio-cultural», Letras de Deusto, 39 (2009), n.º 125, 59-93.
—, «Emergentism», Pensamiento, 65 (2009), n.º 246, Serie Especial, n.º 3, pp. 881-914.
—, «Ciencia y religión. ¿Incompatibles o convergentes?», Realidad, 2010, n.º 122, pp. 823-852.
BERGSON, Henri, La evolución creadora, Madrid, Espasa-Calpe, 1973.BERLIN, I., «Libertad y necesidad en la historia», Madrid, Revista de Occidente,
1958.—, Sobre la libertad, Madrid, Alianza, 2004.BERMÚDEZ DE CASTRO, J. M.ª, El chico de la Gran Dolina. En los orígenes de lo
humano, Barcelona, Crítica, 2002.—, La evolución del talento. Cómo nuestros orígenes determinan nuestro pre-
sente, Barcelona, Debate, 2010.—, SARMIENTO, S./CUNHA, E., «Dimorfismo sexual en dientes humanos», Mundo
Científico, 2000, n.º 214, 17-21.

515
© Universidad de Deusto - ISBN 978-84-9830-383-4
BERTANPETIT, Jaume (comp.), Orígenes del hombre moderno, Barcelona, Prensa Científica, S.A., 1993.
— y JUNYENT, Cr., Viaje a los orígenes. Una historia biológica de la especie hu-mana, Barcelona, Península, 2000.
BICKERTON, D., Language and species, Chicago, Chicago University Press, 1990 (Lenguaje y especie, Madrid, Alianza, 1994).
—, Language and species, Chicago, University of Chicago Press, 1990.—, Language and human behaviour, London, Taylor and Francis, 1998.BLANC, Marcel, «El origen del bipedismo: ¿por motivos de sexo?», Mundo Cien-
tífico, 1983, n.º 25, 526-529.BLAKESLEE, Sandra, «Sorpresas de la década del cerebro», El País, 12 de enero de
2000.BLUMENSCHINE, R. J./CABALLO, J. A., «Carroñeo y evolución humana», Investiga-
ción y Ciencia, 1992, 195, 70-77.BLOND, Olivier, «Las neuronas», Mundo Científico, 1999, n.º 199, 82-85; 83.BOHANNAN, Paul/GLAZER, Mark (eds.), Antropología. Lecturas, Madrid,
McGraw-Hill, 1992 (2.ª ed.).BOTO, Ángela, «¿Necesitan tomar leche los adultos? Controversia sobre el con-
sumo de lácteos en la edad adulta por su posible relación con ciertos tumo-res», El País, 2004, 21 de diciembre, p. 37.
BOURG, D., L’Homme artífice, París, Gallimard, 1996.BOESCH, Chr./BOESCH-ACHERMANN, H., «Los chimpancés y la herramienta»,
Mundo Científico, 1991, n.º 116, pp. 842-849.BOWLER, Peter J., Charles Darwin. El hombre y su influencia, Madrid, Alianza,
1995.—, The New-Darwinian Revolution (Reinterpreting a historical Myth), Baltimore,
The Johns Hopkins University Press, 1988.BOYD, Robert/SILK, J. B., Cómo evolucionaron los humanos, Madrid, Barcelona,
Ariel, 2001.BRADNER NORRIS, R., «Cosmos and Anthropos: A Philosophical Intepretation of
the Anthropic Cosmological Principle, and Cosmos and Theos: Ethical and Teological Implications of the Anthropic Cosmological Principle, by Errol E. Harris», Zygon, 28 (1993), n.º 4, 531-536.
BRAILLON, M. G., El sistema nervioso central, Madrid, Alhambra, 1980.BRETON, S., «La technique entre nature et culture», Esprit, 1997, n.º 11, 159-180.BROCKMAN, John (eds.), La tercera cultura (Más allá de la revolución científica),
Barcelona, Tusquets, 1996.BRONCANO, F. (ed.), La mente humana, Trotta, Madrid, 1995.BROWN, P. y otros, «A New Small-Bodied Hominin from the Late Pleistocene of
Flores, Indonesia», Nature, vol. 431, 28 de octubre de 2004, pp. 1.055-1.061.BUBER, M., ¿Qué es el hombre?, México, FCE, 1949 (1976: 9.ª ed.).—, Yo y tú, Buenos Aires, Nueva Visión, 1969.BUNGE, M., El problema mente-cerebro. Un enfoque psicobiológico, Madrid, Tec-
nos, 1985.BUSKES, Chris, La herencia de Darwin. La evolución en nuestra visión del
mundo, Barcelona, Herder, 2009.

516
© Universidad de Deusto - ISBN 978-84-9830-383-4
BÚSTER, K. W./ISAAC, G. L. (eds.), After the Australopithecines, The Hague, Mouton, 1975.
CAMPILLO ÁLVAREZ, J. E., La cadera de Eva. El protagonismo de la mujer en la evolución de la especie humana, Barcelona, Crítica (Ares y Mares), 2005.
CAMPS, Guíu, «Los datos bíblicos y en el número de los primeros progenitores», en CAMPS, Victoria y otros (eds.), Historia de la ética, 3 vols., Barcelona, Crí-tica, 1988-1992. .
CANDELA, J. A., «El dualismo interaccionista», en DOU, A. (ed.), Mente y cuerpo,Bilbao, Mensajero, 1986, 33-44.
CANN, R. et al., Nature, 325, 31, enero de 1987.CANO ESPINOSA, Diego, «Teleología en el pensamiento biofilosófico de F. J. Aya-
la», Pensamiento, 65 (2009), n.º 246, Serie Especial (Ciencia, Filosofía y Re-ligión), n.º 3, pp. 915-946.
CANOBBIO, Giacomo, Sobre el alma. Más allá de mente y cerebro, Salamanca, Sí-gueme, 2010.
CAPELLE, Ph., «La relation philosophie-théologie: histoire tourmentée, géographie nouvelle», Esprit, 117 (2007), n.º 171, 1-8.
CARBONELL, E. y otros, «Instrumentos de los homínidos de Atapuerca. Las prime-ras comunidades humanas», Mundo Científico, 1997, n.º 175, 54-59.
— et alii, «Instrumentos de los homínidos en Atapuerca», Mundo Científico,1997, n.º 175, 54-59.
—/ ARSUAGA, J. L. y resto del equipo, «Homo Antecessor y su medio natural. Cómo eran y cómo vivían los europeos más antiguos», Mundo Científico,1998, n.º 192, 42-49.
—/ SALA, Robert, Planeta humano, Barcelona, Península, 2000.—/ SALA, R., Aún no somos humanos. Propuestas de humanización para el tercer
milenio, Barcelona, Península, 2002.CARRASCO I PONS, Silvia, Antropología i alimentació. Una proposta per a l’estudi
de la cultura alimentaria, Barcelona, Universidad Autónoma, 1992.CASSIRER, Ernst, Mito y lenguaje, Buenos Aires, Nueva Visión, 1959.—, Antropología filosófica, México, FCE, 1945.—, Filosofía de las formas simbólicas, 3 vols., 1973.CASTRO NOGUEIRA, L./CASTRO NOGUEIRA, L./CASTRO NOGUEIRA, M. A., ¿Quién
teme a la naturaleza humana?, Madrid, Tecnos, 2008.CASTRO, L./LÓPEZ-FANJUL, C. y TORO, M. A., A la sombra de Darwin. Las
aproximaciones evolucionistas al comportamiento humano, Madrid, Si-glo XXI, 2003.
CASTRODEZA, Carlos, La darwinización del mundo, Barcelona, Herder, 2009.—, Nihilismo y supervivencia. Una expresión naturalista de lo inefable, Madrid,
Trotta, 2007.CAVADA, M., La vigencia del amor, Madrid, Edic. Paulinas, 1994.CAVALIERI, P./SINGER, P. (eds.), El proyecto «Gran Simio». La igualdad más allá
de la humanidad, Madrid, Trotta, 1998.CAVALLI-SFORZA, Luca y Franceso ¿Quiénes somos? Historia de la diversidad
humana, Madrid, Barcelona, Crítica, 1999.CELA CONDE, C. J., Sobre genes, dioses y tiranos, Madrid, Alianza, 1985.

517
© Universidad de Deusto - ISBN 978-84-9830-383-4
—/ AYALA, F. J., Senderos de la evolución humana, Madrid, Alianza, 2001.—/ CAPÓ, M. A./NADAL, M./GUAL, J./FERNÁNDEZ, M./RAMOS, C., «De homínido
a humano», en FEITO, Lidia (ed.), Nuevas perspectivas científicas y filosóficas sobre el ser humano, Madrid, UPCO, 2007, pp. 67-92.
CERVERA, J./ARSUAGA, J. L./BERMÚDEZ, J. M.ª/CARBONELL, E., Atapuerca: Un mi-llón de años de historia, Madrid, Ed. Complutense/Plot Edic., 1999.
CHALINE, Jean/MARCHAND, Didier, «Cuando la evolución cambia el tiempo de los seres. Paleontólogos y genetistas se interrogan sobre las heterocronías», Mundo Científico, 1999, n.º 199, pp. 40-43.
CHANGEUX, J.-P. (dir.), Fondements Naturels de l’Ethique, París, Odile Jacob, 1993.
CHASTEL, Olivier/BONNET, Xavier, «La procreación por años sabáticos», MundoCientífico, 1996, n .º 170, 624-625.
CHAUCHARD, Paul, El cerebro y la mano creadora, Madrid, Narcea, 1972.CHAUVIN, Rémy, Darwinismo. El fin de un mito, Madrid, Espasa-Calpe, 2001.CHEVALIER-SKOLNIKOFF, S./POIRIER, F. E., Primate biosocial development: Bio-
logical, social and ecological determinants, Garland Press, 1977.—, Behavioral and Brain Sciences, 12, 561, 1988.CHRISTEN, Yves, El hombre biocultural. De la molécula a la civilización, Madrid,
Cátedra, 1989.CHOMSKY, N. (1966), Cartesian linguistic. A chapter in the history of rationalistic
thought, New York, Harper and Row (Lingüística cartesiana. Un capítulo de la historia del pensamiento racionalista, Madrid, Gredos, 1969).
—, Language and mind, New York, Harcourt Brace Jovanovich, 1968 (trad. cast.: El lenguaje y el entendimiento, Barcelona, Seix Barral, 1971).
—, Knowledge of language: Its nature, origins and use, New York, Praeger, 1986 (El conocimiento del lenguaje. Su naturaleza, origen y uso, Madrid, Alianza, 1989).
—, Language and thought, Wkafield, Moyer Bell, 1993.—, «Language from an internalist perspective», Conferencia dictada en el King’s
College (Londres), 24 de mayo de 1994 («El lenguaje desde una perspectiva internista», en Una aproximación naturalista a la mente y al lenguaje, Barce-lona, Prensa Ibérica, 1998, 193-230).
CHURCHLAND, P. M., Materia y conciencia. Introducción contemporánea a la filo-sofía de la mente, Barcelona, Gedisa, 1999.
CLARK, G. A., «Origen del hombre: un diálogo de sordos», Mundo Científico,1994, n.º 146, 462-467.
CLARKE, Robert, El nacimiento del hombre, Barcelona, Ediciones Juan Granica, 1983.
COLES, Peter, Hawking y la mente de Dios, Barcelona, Gedisa, 2004.COLLINA-GIRAD, Jacques, Le feu avant les allumettes, París, Editorial de la Mai-
son des Sciences de l’Homme, 1998.COLLINS, Francis S., ¿Cómo habla Dios? La evidencia científica de la fe, Madrid,
Temas de Hoy, 2007.CONGREGACIÓN PARA LA DOCTRINA DE LA FE, Declaración de la Sagrada Congre-
gación para la Doctrina de la Fe sobre el aborto, 1974, n.º 13, nota 19.

518
© Universidad de Deusto - ISBN 978-84-9830-383-4
COPPENS, Yves, «Somos africanos y tenemos 7 millones de años», El País, 19 de diciembre de 1991, pp. 60-61.
—, La rodilla de Lucy. Los primeros pasos hacia la humanidad, Barcelona, Tus-quets, 2005.
—/ SENUT, B. (eds.), Origine(s) de la bipedie chez les hominidés, París, Editions du CNRS, 1991.
CORBALLIS, M. C., From hand to mouth: the origins of language, Princeton, Prin-ceton University Press, 2002.
CORDÓN, Faustino, «La genética, a los cien años de Mendel (1822-1884)», El País, 6 de enero de 1984, 22-23.
—, Cocinar hizo al hombre, Barcelona,. Tusquets, 2009 (7.ª ed.).COSTA-PAU, Marta, «El hallazgo de que las neuronas pueden regenerarse es cru-
cial» (entrevista a Paola Timiras, especialista en envejecimiento), El País, 28 de septiembre de 1999, p. 47.
CORTINA, Adela, Etica aplicada y democracia radical, Madrid, Tecnos, 1993.—, Las fronteras de la persona. El valor de los animales, la dignidad de los hu-
manos, Madrid, Taurus, 2009.CRUSAFONT PAIRO, M./MELÉNDEZ, B./AGUIRRE, Emiliano de (dirs.), La evolución,
Madrid, BAC, 1986,CRUZ CRUZ, J., Alimentación y cultura. Antropología de la conducta alimentaria,
Pamplona, EUNSA, 1991.DAMASIO, Antonio R., El error de Descartes. La emoción, la razón y el cerebro
humano, Barcelona, Crítica, 1996.—, Y el cerebro creó al hombre. ¿Cómo pudo el cerebro generar emociones, sen-
timientos, ideas y el yo?, Barcelona, Destino, 2010.DARLU, Pierre/TASSY, Pascal, «El ADN, África y el hombre», Mundo Científico,
1987, n.º 73, 943-945; 945.DAVIES, M. W., Darwin y el fundamentalismo, Barcelona, Gedisa, 2004.DAVIES, P., Dios y la nueva física, Barcelona, Salvat, 1986.—, Otros mundos, Barcelona, Salvat, 1986.DARWIN, Charles, The descent of Man, and selection in relation tu sex, Londres,
John Murray, 1871, 2 vols. (trad. cast.: El origen del hombre, México, Edito-res Mexicanos Unidos, 1982).
—, The Expression of the Emotions in Man and Animals, London, John Murray, 1872 (La expresión de las emociones en los animales y en el hombre, Alianza, Madrid, 1998; Pamplona, Laetoli, 2009).
—, Autobiografía, Barcelona, Edit. Alta Fulla, 1987.—, Autobiografía, Madrid, Alianza Cien, 1993.—, Autobiografía, Pamplona, Editorial Laetoli, 2008. Trad. de J. L. Gil Arista, e
introducción de Martí Domínguez).—, El origen de las especies por medio de la selección natural, Madrid, Alianza
Editorial, 2009 (prólogo de Fr. J. Ayala, introducción de Diego Núñez y trad. de Antonio de Zulueta).
DAWKINS, R., El gen egoísta, Barcelona, Labor, 1979.—, The blind watchmaker, London, Longman, 1986 (El relojero ciego, Barce-
lona, Labor, 1988).

519
© Universidad de Deusto - ISBN 978-84-9830-383-4
—, River out of Eden. A darwinian view of life, New York, Basic Books, 1995 (Elrío del Edén, Madrid, Debate, 2000).
—, El capellán del diablo, Barcelona, Gedisa, 2005.—, El espejismo de Dios, Madris, Espasa, 2007.—, Evolución, Madrid, Espasa-Calpe, 2009.DE WAAL, Frans, La política de los chimpancés, Madrid, Alianza, 1993.—, Bien natural. Los orígenes del bien y del mal en los humanos y otros anima-
les, Barcelona, Herder, 1997.—, Primates y filósofos. La evolución de la moral del simio al hombre, Barce-
lona, Paidós, 2007.DELOISON, Y., «El pie de los primeros homínidos», Mundo Científico, 1996,
n.º 164, 20-23.DEMARET, J./BARBIER, Ch., «Le principe anthropique en cosmologie», Révue des
questions scientiphiques, 15 (1981), 181-222 y 461-509.DEMOULIN, Françoise, «Crecimiento fetal humano», en REBATO, E. y otros (eds.),
Para comprender la Antropología biológica, Estella (Navarra), Verbo Divino, 2005, 497-511.
DEMBSKI, W., The Design Inference. Eliminating Chance through Small Probabil-ities, Cambridge, University Press, 1995 (trad. esp.: Diseño Inteligente. Res-puesta a las cuestiones más espinosas del diseño inteligente, Madrid, Homo Legens, 2006).
DENNETT, D., «Conditions of Personhood», en Brainstorms. Philosophical Essays on Mind and Psychology, Montgomery, Bradford Books, 1976.
—, Darwin’s dangerous idea, New York, Simon and Schuster, 1995 (La peligrosa idea de Darwin. Evolución y significado de la vida, Barcelona, Galaxa Guten-berg/Círculo de Lectores, 1999).
—, Breaking the Spell. Religión as a natural Phenomenon, Londres, Viking, 2006 (trad. española: Romper el hechizo. La religión como fenómeno natural, Bue-nos Aires, Katz Editores, 2006).
DEPUTTE, Bertrand, «La evitación del incesto en los primates», Mundo Científico,1988, n.º 76, pp. 54-64.
D’ERRICO, F. et al., «Neanderthal acculturation in western Europe? A critical re-view of the evidence and its interpretation», Current Anthropology, 1998, 39, suplement, pp. 1-44.
DESCARTES, R., De método, A. T., VI, 558 (tr. cast., Discurso del método, Madrid, Espasa-Calpe, 1937/1970 (12.ª ed.).
—, Meditaciones IX, 63 (Meditaciones metafísicas, Madrid, Espasa-Calpe, 1937/1970, 12 .ª ed.).
DESMOND, A./MOORE, J., Darwin, N. York/London, W. W. Norton and Company, 1991.
D’ESPAGNAT, B., Une incertaine réalité, París, Gauthier Villar, 1985.DESPORTES, J. P., «El comportamiento de vigilancia en los animales», Mundo
Científico, 1987, n.º 67, 238-245.DEVILLERS, Charles/CHALINE, Jean/LAURIN, Bernard, «En defensa de una embrio-
logía evolutiva», Mundo Científico, 1990, n.º 105, 918-924.

520
© Universidad de Deusto - ISBN 978-84-9830-383-4
DIAMOND, Jared, The tirad chimpanzee: the evolution of the future of the human animal, N. York, Harper Perennial, 1993.
—, «El tercer chimpancé», en P. CAVALIERI y P. SINGER, El proyecto «Gran Si-mio». La igualdad más allá de la humanidad, Madrid, Trotta, 1998, 115-132.
—, ¿Por qué es divertido el sexo? Un estudio de la evolución de la sexualidad humana, Madrid, Debate, 1999.
DOBZHANSKKY, Th., Genetics and the Origin of Species, Columbia University Press, 1937.
DOBZHANSKY, Th./AYALA, Fr. J./STEBBINS, G. L./VALENTINE, J. W., Evolution,San Francisco, W. H. Freeman and Company, 1977 (trad. española: Evolu-ción, Barcelona, Omega, 1983).
DOU, A. (ed.), Fragmentariedad de las ciencias, Bilbao, Mensajero/Biblioteca Fomento Social, 1985.
— (ed.), Mente y cuerpo, Bilbao, Mensajero, 1986.DREES, Willem B., «Creacionismo y evolución», Concilium, 2000, n.º 284, 55-
64. DUPRÉ, John, El legado de Darwin. Qué significa hoy la evolución, Bue-nos Aires, Katz Editores, 2006.
DUFOUR, Jean-Paul, «Las primeras herramientas pudieron ser obras de homíni-dos», El País, 12 de mayo de 1999, p. 43.
DULBECCO, R., Los genes y nuestro futuro, Madrid, Alianza, 1999.DUPRE, J., El legado de Darwin, Buenos Aires, Katz Editores, 2006.DUSART, I./RUBIN, B. C./SCHWAH, M. E., «La regeneración de las fibras nervio-
sas», Mundo Científico, 1993, n.º 41, 1.009-1.014.DUTTON, Denis, El instinto del arte. Belleza, placer y evolución humana, Barce-
lona, Paidós, 2010.DYSON, A./HARRIS, J. (eds.), Experiments on Embryos, Londres, Routledge, 1990,
65-81; 72.EAGLETON, T., Reason, Faith, and Revolution: Reflections on th God Debate,
New Haven, Yale University Press, 2009.ECCLES, J., La psique humana, Madrid, Tecnos, 1986.—, The Human Psyche, Londres, Routledge, 1992.—, Evolution of the Brain. Creation of the Self, Londres, Routledge, 1995.—/ ZEIER, H., El cerebro y la mente, Barcelona, Herder, 1985.EDELMAN, G., Neuronal Darwinism: The Theory of Neuronal Group Selection,
Basic Books, 1987.—, Topobiology: An Introduction to Molecular Embriology, Basic Books, 1988.—, The Remembered Present: A Biological Theory of Consciousnes, Basic Books,
1989.—, Bright Air, Brilliant Fire: On the Matter of the Mind, Basic Books, 1992.—/ TONONI, G., El universo de la conciencia, Barcelona, Crítica, 2002.EDWARDS, Denis, El Dios de la evolución. Una teología trinitaria, Santander,
Sal Terrae, 2006.ELDREDGE, Niles, Darwin. El descubrimiento del árbol de la vida, Buenos Aires,
Katz Editores, 2009.

521
© Universidad de Deusto - ISBN 978-84-9830-383-4
—/ GOULD, S. J. (1972), «Punctuated equilibria: An alternative to phyletic gradu-alism», en T. SCHOPF (ed.), Models in Paleobiology, San Francisco, Freeman, 82-115.
ELLACURÍA, Ignacio, «Fundamentación biológica de la ética», ECA, 1979, n.º 368, pp. 418-425.
—, «Biología e inteligencia», ponencia presentada en el XIV Congreso Nacio-nal de la Asociación Española de Neuropsiquiatría («Bases biológicas de la psiquiatría»), Sevilla, octubre de 1977, y editada en Escritos Filosóficos,San Salvador, Edic. UCA, 2001, vol. III, pp. 137-201.
ENGELHARDT, T., The Foundation of Bioethics, Nueva York/Oxford, 1986, 107 (trad. esp.: Los fundamentos de la bioética, Barcelona, Paidós, 1995).
ERICKSON, Jon, La extinción de las especies. Evolución, causas y efectos, Madrid,McGraw-Hill, 1992.
ERIKSON, E. H., «Ontogénie de la ritualisation chez l’homme», en Le comporte-ment rituel chez l’homme et l’ánimal, pp. 139-185 (cita tomada de C. París, o.c., p. 100).
ERICKSON, Robert, «La saga de los espermatozoides. ¿Cómo se formaron?», Mundo Científico, 1985, n.º 52, 1.086-1.097.
ESPINOSA, Miguel, «René Thom: De la teoría de las catástrofes a la metafísica», en ARANA, J. (ed.), La filosofía de los científicos, Sevilla, Thémata. Revista de Filosofía, 1995, n.º 14, 321-348.
ESTELLER, M., «Qué es la epigenética y para qué sirve», El País, 2008, 12 de abril.
ESTRADA, J. A., La imposible teodicea. La crisis de la fe en Dios, Madrid, Trotta, 1997.
EVANS-PRITCHARD, E., The Nuer: A Description of the Modes of Livelihood and Political Institutions of a Neolitic People, Oxford, Clarendon Press, 1940.
FABIAN, Andrew C. (ed.), Evolución. Sociedad, ciencia y universo, Barcelona, Tusquets, 2001.
FEINER, J., «El origen del hombre», Mysterium Salutis, vol. II, tomo II, Madrid, Cristiandad, 1967, pp. 638-660.
FEITO, Lidia (ed.), Nuevas perspectivas científicas y filosóficas sobre el ser hu-mano, Madrid, UPCO, 2007,
FERGUSON, R. Brian, «La naissance de la guerre», La Recherche Scientifique,2004, n.º 373, 51-57.
FERNÁNDEZ, Domiciano, El problema del pecado original, Madrid, Nueva Uto-pía, 1999.
FERNÁNDEZ BEITES, Pilar, «Sustantividad humana: embrión y “actividad pa-siva” de la inteligencia», en MURILLO, Ildefonso (coord.), Filosofía práctica y persona humana, Salamanca, Publicaciones Universidad Pontificia de Sala-manca, 2004, 244-255.
FERNÁNDEZ MATEOS, L. M., El vínculo afectivo con el niño por nacer, Salamanca, Universidad Pontificia de Salamanca, 2005.
FERNÁNDEZ-RAÑADA, A., Los científicos y Dios, Madrid, Trotta, 2008, pp. 118 y ss.
FISHER, Helen, El contrato sexual, Barcelona, Salvat, 1987.

522
© Universidad de Deusto - ISBN 978-84-9830-383-4
FISCHER, S. R., A history of language, London, Reaktino Books, 1999 (Breve his-toria del lenguaje, Madrid, Alianza, 2003).
FLANDRIN, J.-L./MONTANARI, M. (dirs.), Historia de la alimentación, Gijón, Edic. Trea, S.L., 2004.
FODOR, J. (1975), The language of thought, New York, Thomas Y. Crowell (Ellenguaje del pensamiento, Madrid, Alianza, 1984).
—, The Modularity of Mind, Cambridge, Mass., MIT Press, 1983.FONTANILLE, J., «Dieciocho definiciones sobre el gen», La Recherche Scientifi-
que, 2003, vol. 10, n.º 50, p. 17.FORMENT, Eudaldo, «Estatuto ontológico del embrión», Espíritu, LI (2002),
n.º 126, 257-270.FOSSEY, D., Gorilas en la niebla, Barcelona, Salvat, 1985.FOUCAULT, Michel, Las palabras y las cosas, Madrid, Siglo XXI, 1968.FOX KELLER, Evelyn, El siglo del gen. Cein años de pensamiento genético, Bar-
celona, Península, 2002.FRANCIONE, Gary L., «El error de Bentham (y el de Singer)», Teorema, XVIII/e
(1999), 39-60.GAFO, Javier, Problemas éticos de la manipulación genética, Madrid, Edic. Pau-
linas, 1992.—, Procreación humana asistida: aspectos técnicos, éticos y legales, Madrid,
UPCO, 1998,— (dir.), Diez palabras clave en Ecología, Estella (Navarra), Verbo Divino,
1999.GALE, George, «El principio antrópico», Investigación y Ciencia, 1982, n.º 65,
pp. 93-103.GALLAY, Alain, «El hombre neolítico y la muerte», en VV.AA., Orígenes del
hombre moderno, Barcelona, Prensa Científica (Libros de Investigación y Ciencia), 1993, pp. 124-133.
GARCÍA BACCA, J. D., Antropología filosófica contemporánea, Caracas, UCV, 1947.
—, Curso sistemático de filosofía actual, Universidad Central de Venezuela, Ca-racas, 1969.
GARDNER, Howard, Frames of Mind: the Theorie of Multiple Intelligences,Nueva York, Basic Books, 1983.
—, Inteligencias múltiples. La teoría en la práctica, Barcelona, Paidós, 1995.—, La nueva ciencia de la mente (Paidós, Barcelona,1987; 2.ª ed.: 2000).GARDNER, Martín, ¿Tenían ombligo Adán y Eva? La falsedad de la seudociencia
al descubierto, Madrid, Debate, 2001.GARDNER, Richard L., «El destino de las células en el embrión», Mundo Cientí-
fico, 1991, n.º 110, 146-155.GARTNER, A./GREER, C./RIESSMAN, F. (comps.), Nuevo ataque contra la igualdad
de oportunidades. Cociente intelectual y estratificación social, Madrid, Edi-ciones Morata, 1999.
GAZZANIGA, Michael S., ¿Qué nos hace humanos? La explicación científica de nuestra singularidad como especie, Barcelona, Paidós, 2010.
GEBARA, Ivonne, El rostro oculto del mal, Madrid, Trotta, 2002.

523
© Universidad de Deusto - ISBN 978-84-9830-383-4
GEHLEN, A., Der Mensch, Frankfurt, Athenaion Verlag, 1940 (2.ª ed. 1974) (Elhombre. Su naturaleza y su lugar en el mundo, Salamanca, Sígueme, 1980).
GEVAERT, J., El problema del hombre, Salamanca, Sígueme, 1976.GIRÓN SIERRA, A., Evolucionismo y anarquismo en España (1882-1914), Madrid,
CSIC, 1996.—, En la mesa con Darwin. Evolución y revolución en el movimiento libertario
en España (1869-1914), Madrid, CSIC, 2005.GOLEMAN, D., Inteligencia emocional, Barcelona, Kairós, 1996.GOLSCHMIDT, R. B., The Material Basic of Evolution, Yale University Press,
New Haven, 1940 (El material básico de la evolución, Madrid, Espasa-Calpe, S.A., 1943).
GOMIS, A./JOSA, J, «Creacionismo y evolución. Imágenes de la polémica darwi-nista en España», Mundo Científico, 2002, n.º 233, pp. 20-29.
GOMIS, A./JOSA, J., «Imágenes de la polémica darwinista en España. Creacio-nismo y evolucionismo», Mundo Científico, 2002, n.º 233, pp. 20-29.
GOMIS, Alberto/JOSA, Jaume, Bibliografía crítica ilustrada de las obras de Darwin en España (1857-2008), Madrid, CSIC, 2009.
GOMIS, J., «Sobre el evolucionismo y la evolución de la Iglesia», Iglesia Viva,1996, n.º 186, pp. 555-560; 556.
GROSS, Michael, «El doble juego de una proteína», Mundo Científico, 1999, n.º 204, 28-31.
GEE, Henry, «Explosiones repentinas de cambios. La nueva visión, 21 años des-pués», El País, 8-XII-1993, Futuro, p. 3.
—, «¿Es la fabricación de utensilios un rasgo exclusivo de los humanos?», El País, 12 de mayo de 1999, p. 43.
GOLDBERG, E., El cerebro ejecutivo, Barcelona, Crítica, 2002.GÓMEZ CAFFARENA, J. y MARTÍN VELASCO, J., «Filosofía de la religión», Madrid,
Revista de Occidente, 1973.GÓMEZ PIN, Víctor, El hombre, un animal singular, Madrid, La Esfera de los Li-
bros, 2005.—, Entre lobos y autómatas. La causa del hombre, Madrid, Espasa, 2006.GÓMEZ ROMERO, P., MetaEvolución. La Tierra en el Espejo, Madrid, Celeste Edi-
ciones, 2001.GOMILA, A., «Evolución y lenguaje», en BRONCANO, F. (ed.), La mente humana,
Madrid, Trotta, 1995, 273-288; 276.GONZÁLEZ ÁLVAREZ, Julio, Breve historia del cerebro, Barcelona, Crítica, 2010.GONZÁLEZ ECHEVERRÍA, Aurora, Teorías del parentesco. Nuevas aproximaciones,
Madrid, Eudema, 1994.GONZÁLEZ FAUS, J. I., Proyecto de hermano. Visión creyente del hombre, Santan-
der, Sal Terrae, 1987.—, La autoridad de la verdad, Santander, Sal Térrea, 2006.GONZÁLEZ-CRUSSI, Nacer y otras dificultades. Historia cultural del enigma de la
vida, Barcelona, Destino, 2008, pp. 139 y ss.GOODALL, J., «Nest Building Behaviour in the Free Ranging Chimpanzees», An-
nuary of the New York Academy os Sciences, 1962, 102, pp. 455-467.

524
© Universidad de Deusto - ISBN 978-84-9830-383-4
—, In the shadow of man, Boston, Houghton Mifflin, 1971 (En la senda del hom-bre, Barcelona, Salvat, 1988).
—, The Chimpanzees of Gombe: Patterns of Behavior, Cambridge, Mass., Belk-nap Press, Harvard University Press, 1986.
—, Through a Window, Boston, Houghton Mifflin (trad. cast.: A través de la ven-tana, Barcelona, Salvat, 1994).
GOODMAN, M., «Man’s place in the phylogeny of the primates as reflected in se-rum proteins», en WASHBURN, S. L., Classification and Human Evolution,Metheun an Co., 1964.
GOODWIN, B. (1994), How the leopard changed its sopots. The evolution of com-plexity, New York, Charles Schribner’s Son (Las manchas del leopardo. La evolución de la complejidad, Barcelona, Tusquets, 1998).
GOULD, S. J., Ontogeny and Phylogeny, Cambridge, Mass., Harvard University Press, 1977 (trad. cast.: Ontogenia y filogenia. La ley fundamental biogené-tica, Barcelona, Crítica, 2010).
—/ ELDREDGE, N., «Punctuated equilibria: The tempo and the mode of evolution reconsidered», Paleobiology, 1977, 3, 115-151.
—/ LEWONTIN, R., «The spandrels of San Marcos and the Panglossian paradigm: a critique of the adapatationist programme», Proceedings of the Royal Society of London, 1979. B 205, 581-598.
—/ VRBA, E., «Exaptation: a missing term in the science of form», Palebiology,1981, vol. 8, 4-15.
—, «El equilibrio puntuado y el enfoque jerárquico de la macroevolución», Re-vista de Occidente, 1982, n.º 18-19, 121-148.
—/ LEWONTIN, R. C., «La adaptación biológica», Mundo Científico, 1983, n.º 22, 214-223.
—, La falsa medida del hombre, Barcelona, Antoni Bosch, 1984.—, The panda’s thumb: more reflexions in natural history, London, Penguin, 1990
(El pulgar del panda. Reflexiones sobre historia natural y evolución, Barce-lona, Crítica, 1994).
—, La vida maravillosa, Barcelona, Crítica, 1999.—, «The Structure of Evolutionary Theory», Cambridge (Mas.)/London, Harvard
University Press, 2002 (trad. cast..: La estructura de la evolución, Barcelona, Tusquets, 2004).
—, La falsa medida del hombre, Barcelona, Crítica, 2007 (edición revisada y au-mentada).
—, Ciencia versus religión, Barcelona, Crítica, 2007.GRACIA, Diego, «Biología e inteligencia», ECA (UCA, El Salvador), XXXVIII
(1982), n.º 406, pp. 801-816.—, «Problemas filosóficos en genética y en embriología», en ABEL, F./CAÑON, C.
(eds.), La mediación de la filosofía en la construcción de la bioética, Madrid, UPCO, 1993, 215-254.
—, «El estatuto del embrión», en GAFO, Javier, Procreación humana asistida: as-pectos técnicos, éticos y legales, Madrid, UPCO, 1998, 79-109.
—, «Los animales en la tradición occidental», en LACADENA, J. R. (eds.), Los dere-chos de los animales, Madrid/Bilbao, UPCO/Desclée de Brouwer, 2002, 134-150.

525
© Universidad de Deusto - ISBN 978-84-9830-383-4
GRASA HERNÁNDEZ, R., El evolucionismo: de Darwin a la sociobiología, Madrid, Cincel, 1986.
GREE, Henry, «Explosiones repentinas de cambios. La nueva visión, 21 años des-pués», El País, 8-XII-1993, Futuro, p. 3.
GREGORY, R. L. (ed.), Diccionario Oxford de la mente, Madrid, Alianza, 1995, pp. 149-161.
GRESHAKE, Gisbert, ¿Por qué el Dios del amor permite que suframos?, Sala-manca, Sígueme, 2008.
GRIBBIN, John y GRIBBIN, Mary, Hijos del hielo. Clima y orígenes del hombre,Madrid, Pirámide, 1992.
GRIFFIN, Donald, El pensamiento de los animales, Barcelona, Ariel, 1986.GROSS, Michael Gross, «El doble juego de una proteína», Mundo Científico,
1999, n.º 204, 28-31.GUBERN, Román, El simio informatizado, Barcelona, Fundesco, 1987.GUIBERT UCIN, J M./ORELLA UNZUÉ, J. L. (eds.), Preguntas éticas en cuestiones
disputadas hoy, San Sebastián, Universidad de Deusto, 2005.GUILLAUMIN, Godfrey, Raíces metodológicas de la teoría de la evolución de
Charles Darwin, Barcelona/México, Anthropos/UAM, 2009.GUILLÉN-SALAZAR, Federico (ed.), Existo, luego pienso. Los primates y la evolu-
ción de la inteligencia humana, Madrid, Ateles Editores, 2005.GUITTON, Jean, Dios y la ciencia. Hacia el metarrealismo, Madrid, Debate, 1992
(4.º ed.: 1998).GUTIÉRREZ LOMBARDO, Raúl, «La teoría de la evolución y la Iglesia Católica»,
Ludus Vitalis, X (2002), n.º 17, 111-122.GUTIÉRREZ, A./ORTEGA, R., «Las implicaciones sociales y políticas de la sociobio-
logía: una introducción a la literatura», Revista de Estudios Políticos, 1990, n.º 67, 297-318.
—/ ORTEGA, R., «Sobre el hombre biocultural», Letras de Deusto, 21 (1991), n.º 51, 153-165.
—, «El fenómeno ético y religioso visto desde la sociobiología. Introducción a la literatura», La Ciudad de Dios, CCXV/1 (2002), pp. 123-154.
—, «El debate de la sociobiología: una aproximación bibliográfica», SuplementosAnthropos, n.º 10, 170-177.
—, «El dualismo interaccionista del neurofisiólogo católico John Carew Eccles», La Ciudad de Dios, CCXIV (2001), 189-205 (1.ª parte), 295-346 (2.ª parte).
HAECKEL, E., Histoire de la création des êtres organisés d’aprés les lois naturel-les, París, Alfred Costes, 1922.
HAEFFNER, G., Antropología filosófica, Barcelona, Herder, 1986.HAKEN, H., Fórmulas de éxito en la naturaleza, Madrid, Salvat, 1986.HALDANE, J. B. S., «The Origin of Live», Rationalist Annual, 1929, 148, pp. 3-10.—, »The origin of live», New Biol., 1954, 16, pp. 12-27.HAMER, Dean, El Gen de Dios, Madrid, La Esfera de los Libros, 2006.HAMILTON, W. D., «The Genetical Evolution of Social Behavior». Journal of The-
oretical Biology, 1964, 7, pp. 1-52.HÄRING, H., «La teoría de la evolución como megateoría del pensamiento occi-
dental», Concilium, 2000, n.º 284, 27-40.

526
© Universidad de Deusto - ISBN 978-84-9830-383-4
HARRIS, M., El desarrollo de la teoría antropológica. Una historia de las teorías de la cultura, Madrid, Siglo XXI, 1978.
—, Vacas, cerdos, guerras y brujas. Los enigmas de la cultura, Madrid, Alianza, 1980.
—, El materialismo cultural, Madrid, Alianza, 1987 (2.ª ed.).—, Bueno para comer, Madrid, Alianza, 1989.—, «Embryos and Hedgehogs: On the moral status of the embryo», en DYSON, A./
HARRIS, J. (eds.), Experiments on Embryos, Londres, Routledge, 1990, 65-81.—/ ROSS, E. B., Muerte, sexo y fecundidad. La regulación demográfica en las so-
ciedades preindustriales y en desarrollo, Madrid, Alianza, 1991.HART, S., El lenguaje de los animales, Barcelona, Omega, 1997.HAUGHT, John F., Cristianismo y ciencia. Hacia una teología de la naturaleza,
Santander, Sal Terrae, 2009.HAUSER, Marc D., La mente moral. Cómo la naturaleza ha desarrollado nuestro
sentido del bien y del mal, Barcelona, Paidós, 2008.HAWKING, S., La historia del tiempo. Del big-bang a los agujeros negros, Barce-
lona, Crítica, 1988.—, El universo en una cáscara de nuez, Barcelona, Crítica, 2002.—, La teoría del todo. El origen y el destino del universo, Barcelona, Mondadori,
2007.— y MLODINOV, L., El gran diseño, Barcelona, Crítica, 2010.HEIDEGGER, M., Kant y el problema de la metafísica, México, FCE, 1954; SCHE-
LER, M., El puesto del hombre en el cosmos, Argentina, Losada, 1938.HELLER, Agnes, Instinto, agresividad y carácter, Barcelona, Ed. Península, 1980
(2.ª ed. 1994).HEMLEBEN, Johannes, Darwin, Madrid, Aliaza, 2980 (2.ª ed.).HIERRO-PESCADOR, José, Filosofía de la mente y ciencia cognitiva, Madrid, Akal,
2005.HOLDEN, C. (2005), «Sur les traces du premier langage», Les Dossier de la Re-
cherche, 19, 88-92.HODDER, Ian, «La división sexual en Catal-Hüyük», Investigación y Ciencia,
2004, n.º 330, 50-57.HOWELL, W. W., Journal of Human Evolution, 5, 477, 1976.HUBLIN, J. J. et al., «A late neanderthal asociated with upper paleolithic arte-
facts», Nature, 1996, 381, 224-226.HUMPHREY, Nicholas, Consciousness Regained, Oxford, Oxford University Press,
1983 (La reconquista de la conciencia. Desarrollo de la mente humana,México, FCE, 1987).
—, The Inner Eye, Londres, Faber and Faber, 1986 (trad. cast.: La mirada inte-rior, Madrid, Alianza, 1993).
—, A History of the Mind, Londres, Simon and Schuster, 1992 (trad. cast.: Unahistoria de la mente. La evolución y el nacimiento de la conciencia, Barce-lona, Gedisa, 1995).
ISAAC, G. I., en WENDORF, F./CLOSE, A. (eds.), Advances in World Archaeology,vol. 3, Academic Press, 1984.

527
© Universidad de Deusto - ISBN 978-84-9830-383-4
ISSAR, Aire S., «¿Se llevan bien la Biblia y la ciencia? Las plagas de Egipto y del Éxodo pasadas por el tamiz de la hidrogeología», Mundo Científico, 1996, n.º 166, 240 y ss.
ITARD, J., Victor de L’Aveyron, Madrid, Alianza, 1982 (introducción y comenta-rios de R. Sánchez Ferlosio).
IZPISUA BELMONTE, Juan Carlos, «Claves para descifrar el desarrollo humano», El País, 1997, 31 de diciembre, pp. 29-30; 29.
IZQUIETA, J. L./GARCÍA RIOBOO, M.ª A., «Altruismo y solidaridad. Apuntes antro-pológicos», Estudios Filosóficos, 1987, n.º 36, 459-462.
JACOB, François, Lógica de lo viviente, Barcelona, Laia, 1977.—, La logique du vivant. Une histoire de l’hérédité, París, Gallimard, 1970
(La lógica de lo viviente. Una historia de la herencia, Barcelona, Tusquets, 1999).
—, El desván de la evolución, Valencia, Universidad de Valencia, 2007.JACOBS, Gerald H./NATHAM, Jeremy, «La evolución de la visión de los colores en
los primates», Investigación y Ciencia, 2009, n.º 393, 42-49.JENKINS, Alexander/PÉREZ, Gilad, «Buscando vida en el multiverso», Investiga-
ción y Ciencia, 2010, marzo, n.º 402, pp. 12-19.JOHANSON, Donald y EDEY, Maitland, El primer antepasado del hombre, Barce-
lona, Planeta, 1987 (3.ª ed.).JOHNSON, Allen W., La cuestión esencial. Preguntas y respuestas sobre la evolu-
ción, Barcelona, Planeta, 1990.—/ EARLE, Timothy, La evolución de las sociedades humanas, Barcelona, Ariel,
2003.JOHNSON, Mark, El cuerpo en la mente, Madrid, Debate, 1991.JOHNSON, Ph. E., Juicio a Darwin, Madrid, Homo Legens, 2007.JONAS, Hans, El principio responsabilidad. Ensayo de una ética para la civiliza-
ción tecnológica, Barcelona, Herder, 1995.—, El principio vida. Hacia una biología filosófica, Madrid, Trotta, 2000.JORDAN, B., Los impostores de la genética, Barcelona, Península, 2000.JOULIAN, Frédéric, «Origen del hombre, termitas y chimpancés (¿Eran insectívo-
ros los primeros hombres?)», Mundo Científico, 2001, n.º 230, 14-15.JUAN PABLO II, «Mensaje a los miembros de la Academia Pontificia de Ciencias,
reunidos en Roma para su asamblea plenaria (22-10-96)», Ecclesia, 16 de di-ciembre de 1996, n.º 2.815, pp. 25-26.
JUNG, P./SHANNON, T. (eds.), Abortion and Catholicism, Nueva York, Crossroad, 1988.
JÜNGEL, E., Dios como misterio del mundo, Salamanca, Sígueme, 1984.JUNKER-KENNY, Maureen, «El estatuto moral del embrión», Concilium, 1998,
n.º 275, 73-87.KALIL, Ronald E., «Formación de sinapsis durante el desarrollo del cerebro», In-
vestigación y Ciencia, 1990, n.º 161, 26-34.KAUFFMAN, S., At home in the universe. The search for the laws of self-organiza-
tion and complexity, New York, Oxford University Press, 1995.KEMPF, Heervé, «El nacimiento de los útiles. Peleontólogos, etólogos y psicólo-
gos unen sus esfuerzos», Mundo Científico, 1997, 523.

528
© Universidad de Deusto - ISBN 978-84-9830-383-4
KENNEALLY, Christine, La primera palabra. La búsqueda de los orígenes del len-guaje, Madrid, Alianza, 2009.
KENNEDY, H./DEHAY, C., «Córtex cerebral: el peso de los genes», Mundo Cientí-fico, 1994, n.º 146, 474-475.
KING, Mary-Claire/WILSON, Allan C., «Evolution at twolevels in humans and chimpanzees», Science, 1975, n.º 188, pp. 107-116.
KITCHER, Ph., «El origen de la moral. La tendencia a la simpatía no contradice la teoría de la evolución», Mundo Científico, 1997, n.º 181, 608-611.
KLEIN, Jan/TAKAHATA, N./AYALA, F. J., «Polimorfismo del MHC y origen del hombre», Investigación y Ciencia, 1994, n.º 215, 14-19.
KROEBER, A., Anthropology, culture patterns and process, Nueva York, Harper Books, 1963.
KROGMANN, W. M., Child growth, University of Michigan, Ann Arbor, 1972.KROPOTKIN, Peter, Mutual Aid: A factor on evolution, Londres, 1902 (trad. de
Carlos Díaz, El apoyo mutuo. Un factor de la evolución, Bilbao, Edit. Zero Zyx, 1970; 2.ª ed.: 1978).
—, La moral anarquista, Madrid, 1977.—, La selección natural y el apoyo mutuo, Madrid, CSIC, 2009 (Introducción de
A. Girón Sierra).KÛNG, H., El principio de todas las cosas. Ciencia religión, Madrid, Trotta, 2007.KUPER, Adam, El primate elegido, Barcelona, Crítica, 1994.LACADENA, J. R., Fe y biología, Madrid, PPC, 2001.—, Genética y Bioética, Madrid/Bilbao, UPCO/Desclée de Brouwer, 2002.— (ed.), Los derechos de los animales, Madrid, UPCO/Bilbao, Desclée de
Brouwer, 2002.—, «Biología y humanidad: hominización y humanización», en MASIÁ, Juan
(ed.), Ser humano, persona y dignidad, Madrid/Bilbao, UPCO/Desclée de Brouwer, 2005, pp. 43-89.
LAÍN ENTRALGO, P., El cuerpo humano. Teoría actual, Madrid, Espasa-Calpe, 1989.
—, Cuerpo y alma. Estructura dinámica del cuerpo humano, Madrid, Espasa-Calpe, 1991.
—, «El problema alma-cuerpo en el pensamiento actual», en MORA, F. (ed.), Elproblema cerebro-mente, Madrid, Alianza, 1995, 17-35.
—, «La psicoterapia como somatoterapia: proceso de simbolización», en ALE-MANY, C./GARCÍA, V. (eds.), El cuerpo vivenciado y analizado, Bilbao, Des-clée de Brouwer, 1996, pp. 45-61.
—, Qué es el hombre. Evolución y sentido de la vida, Oviedo, Ediciones Nobel, 1999.
LAITMAN, J. T., «El origen del lenguaje articulado», Mundo Científico, 1986, n.º 64, 1.182-1.191.
LANDMANN, Michael, Antropología filosófica. Autointerpretación del hombre en la historia y en el presente, México, UTEHA, 1961.
LANE, Nock, Los diez grandes inventos de la evolución, Barcelona, Ariel, 2009.LANE CRAIG, W., «Naturalismo y cosmología», en SOLER GIL, F. J. (ed.), Dios y
las cosmologías modernas, Madrid, BAC, 2005, 49-99.

529
© Universidad de Deusto - ISBN 978-84-9830-383-4
LAVOCAT, R./CASELLES, H., «Polygènisme», en Suplément au Dictionnaire de la Bible, París, 1967, col. 90-110.
LEAKY, Meave y WALTER, Alan, «Antiguos fósiles de homínidos en África», In-vestigación y Ciencia, 1997, n 251, 70-75.
LEAKY, R. E./LEWIN, R., Los orígenes del hombre, Madrid, Aguilar, 1980.—, La formación de la humanidad, Barcelona, Edic. Serbal, 1981.LEAKY, M. D., «Cultural Patterns in the Olduvai Sequence», en K. W. BUTZER y
G. L. ISAAC (eds.), After the Australopithecines, The Hague, Mouton, 1975, pp. 477-493.
LEDOUARIN, Nicol, «La cresta neural juega un papel clave en la evolución», El País, 24 de abril de 2002, p. 33.
LEE, R. B./DE VORE, I. (eds.), Man the Hunter, Chicago, III, Aldine Publishing Co., 1968.
LEJEUNE, J., «Adam et Eve ou le monogenisme», en Nouv. Rev. Théol., 1968, n.º 90, p. 191.
—, «Sur les mécanismes de la spéciation», C. R. séances de la Soc. de Biol., 169 (4) 1975, p. 828.
LEITH, B., El legado de Darwin, Barcelona, Salvat, 1986.LEÓN AZCÁRATE, Juan Luis de, La muerte y su imaginario en la historia de las re-
ligiones, Bilbao, Universidad de Deusto, 2007.LEROI-GOURHAN, A., El hombre y la materia (vol. I. Evolución y técnica; vol. II.
Medio y técnica), Madrid, Taurus, 1988/1989.LESLIE, John, «Time and the Anthropic Principle», Mind, 101 (1992), n.º 403,
521-540.LEVINAS, E., El humanismo del otro hombre, Madrid, Siglo XXI, 1974.—, Totalidad e infinito, Salamanca, Sígueme, 1977.—, De otro modo que ser, o más allá de la esencia, Salamanca, Sígueme, 1987.LEVI-STRAUSS, Claude, Les structures élémentaires de la parenté, París, PUF, 1949
(Las estructuras elementales del parentesco, Buenos Aires, Paidós, 1969).LEWIN, Roger, «Un antepasado para el orangután», Mundo Científico, 1982,
n.º 20.—, Evolución humana, Barcelona, Salvat, 1986, cap. 30, pp. 198-203.—, La interpretación de los fósiles, Barcelona, Planeta, 1987.—, «El nacimiento de la Antropología molecular», Mundo Científico, 1991,
n.º 119, pp. 1.222-1.231.LEWONTIN, R. C., «Adaptation», Scientific American, 239: 157169, 1978 (traduc-
ción española en Investigación y Ciencia, 26: 138-149, 1978).—/ ROSE, S./KAMIN, L. J., No está en los genes. Racismo, genética e ideología,
Barcelona, Crítica, 1987.—, El futuro del genoma humano y otras ilusiones, Barcelona, Paidós, 2001.LIEBERMAN, Ph. (1973), «On the evolution of language: A unified view», Cogni-
tion, 2 («Un enfoque unitario de la evolución humana»), en V. SÁNCHEZ DE
ZABALA (ed.), Sobre el lenguaje de los antropoides, Madrid, Siglo XXI, 1976, 147-203).
—, Toward an evolutionary biology of language, Cambridge, Mass., Belknap Press Of Harvard University Press, 2006.

530
© Universidad de Deusto - ISBN 978-84-9830-383-4
LINDBERG, D. L./NUMBERS, R. L. (eds.), God and Nature. Historical Essays on the Encounter between Christianity and Science, Berkeley, CA, University of California Press, 1986.
LINDLEY, David, Incertidumbre. Einstein, Heisenberg, Bohr y la lucha por la esencia de la ciencia, Barcelona, Ariel, 2008.
LIZ, M., Perspectivas actuales en filosofía de la mente, Tenerife, Gobierno de Ca-narias, 2001.
LÓPEZ BARAHONA, Mónica/LUCAS LUCAS, Ramón (dirs.), El inicio de la vida. Identidad y estatuto del embrión humano, Madrid, BAC, 1999 (2.ª ed.).
LORENZ, K. y LEYHAUSEN, P., Biología del comportamiento. Raíces instintivas de la agresión, el miedo y la libertad, Madrid, Siglo XXI, 1971 (8.ª ed.: 1991).
—, Los ocho pecados capitales de la humanidad civilizada, Barcelona, Plaza y Janés, 1975.
—, Sobre la agresión. El así llamado mal, Madrid, Siglo XXI, 1980.—, La acción de la naturaleza y el destino del hombre, Madrid, Alianza, 1989.LORENZO, G./LONGA, V. M., Homo Loquens. Biología y evolución del lenguaje,
Lugo, Edit. Tris Tram, 2003, p. 85.LORITE MENA, José, El animal paradójico. Fundamentos de Antropología filosó-
fica, Madrid, Alianza, 1982, pp. 375-380.—, La filosofía del hombre, o el ser inacabado, Estella, Verbo Divino, 1992.LOVELOCK, J. y otros, Gaia. Implicaciones de la nueva biología, Barcelona, Kai-
rós, 1995 (3.ª ed.).LUISI, Pier Luigi, La vida emergente. De los orígenes químicos a la biología sin-
tética, Barcelona, Tusquets, 2010.LUMSDEN, E./WILSON, E. O., Genes, Mind and Culture. The Coevolutionary Proc-
ess, Cambridge, Mass., Harvard University Press, 1981.—, El fuego de Prometeo, México, FCE, 1985.MALINOWSKI, Bronislaw, Sex and Represión in Savage Society, Londres,
Routledge, 1927,—, The Sexual Life of Savages in Noth-Western Melanesia, Londres, Routledge,
1929 (Vida sexual de los salvages del nordeste de la Melanesia, Madrid, Mo-rata, 1975).
MARGULIS, Lynn/SAGAN, Dorion, ¿Qué es el sexo?, Barcelona, Tusquets, 1998.—, —, Captando genomas. Una teoría sobre el origen de las especies, Barcelona,
Kairós, 2003 (prólogo de Ernst Mayr).MARINA, J. A./VALGOMA, M.ª de la, La lucha por la dignidad: teoría de la felici-
dad política, Barcelona, Anagrama, 2001.MARSHACK, A., «A middle paleolitihic Symbolic Composition from the Golan
Heights: The earliest known depictive image», Current Anthropology, 1996, 37, pp. 357-365.
MARTÍ, E., Construir una mente, Barcelona, Paidós, 1997, pp. 137-178.MARTIN, R. D., «Capacidad cerebral y evolución humana», Investigación y Cien-
cia, 1994, n.º 219, pp. 71-77.MARTÍN VELASCO, J. D., Fenomenología de la religión, Madrid, Ediciones Cris-
tiandad, 1973 (3.ª ed.: 1982).MARTÍNEZ FREIRE, P. F., La nueva filosofía de la mente, Barcelona, Gedisa, 1999.

531
© Universidad de Deusto - ISBN 978-84-9830-383-4
— (ed.), Filosofía actual de la mente (Contrastes, Suplemento 6, Valencia, 2001).
MARTÍNEZ PULICO, Carolina, El papel de la mujer en la evolución humana, Ma-drid, Biblioteca Nueva, 2003.
—, La presencia femenina en el pensamiento biológico, Madrid, Minerva Edicio-nes, 2006.
MARX, K., El Capital, Madrid, EDAF, 1972.MASIÁ, Juan (ed.), Bioética y antropología, Madrid, UPCO, 1998.—, Ser humano, persona y dignidad, Madrid/Bilbao, UPCO/Deslée de Brouwer,
2005.MATABOSCH, A., «El evolucionismo y la Iglesia», La Vanguardia, 25 de noviem-
bre de 1996.MATTEI, Jean-François (coord.), El genoma humano, Madrid, Editorial Complu-
tense, 2002.MAYNARD SMITH, John, La construcción de la vida. Genes, embriones y evolu-
ción, Barcelona, Crítica, 1998.— y SZATHMÁRY, Eörs, Ocho hitos de la evolución. Del origen de la vida a la
aparición del lenguaje, Barcelona, Tusquets, 2001.MAYOR ZARAGOZA, F./ALONSO BEDATE, C. (coords.), Gen-Etica, Barcelona, 2003,
19-66.MAYR, E., Una larga controversia: Darwin y el darwinismo, Barcelona, Crítica,
1992.— y PROVINE, W., The Evolutionary Synthesis, Harvard University Press, 1980.MCBREARTY, S./BROOKS, A. S., «The revolution that wasn’t: a new interpretation
of the origin of modern human behavior», J. Hum. Evol., 2000, 39, 453-463.MCGRATH, A. E./MCGRATH, J. C., The Dawkins Delusion? Atheist Fundamenta-
lism and the Denial of the Divine, Londres, SPCK, 2009.—, «Los ateísmos de superventas. El nuevo cientifismo», Concilium, 2010,
n.º 337, 555-567.MEDINA, John J., El reloj de la edad, Barcelona, Crítica, 1997.MEIER, R., Comportamiento animal. Un enfoque evolutivo y ecológico, Madrid,
McGrawHill, 2001.MELOTTI, Humberto, El hombre entre la naturaleza y la historia, Barcelona, Pe-
nínsula, 1981.MESTRES, F., «Procedimientos utilizados por la evolución molecular para entender
el origen del hombre moderno», Ludus Vitalis, VIII (2000), n.º 13, 163-170.MICHELENA, Mariela, Un año para toda la vida, Madrid, Temas de Hoy, 2002.MIETH, D., Geburtenregelung. Ein Konflikt in der katolischen Kirche, Grünwald,
Maguncia, 1990, 95-96.MITHEN, S. (1996), The prehistory of the mind. A search for the origin of art, re-
ligion and science, London, Thames and Hudson Ltd. (Arqueología de la mente. Orígenes de la religión, el arte y la ciencia, Barcelona, Crítica, 1998).
MOLINA, E., «Los argumentos geológicos y paleontológicos de los creacionis-tas “científicos”: ignorancia y pseudociencia», en MOLINA, E./CARRERAS, A./PUERTAS, J. (eds.), Evolucionismo y racionalismo, Zaragoza, Universidad de Zaragoza, 1998.

532
© Universidad de Deusto - ISBN 978-84-9830-383-4
MONOD, J., El azar y la necesidad. Ensayo sobre la filosofía natural de la filoso-fía moderna, Barcelona, Barral, 1971 (9.ª ed. 1977).
MONSERRAT, J., Epistemología evolutiva y teoría de la ciencia, Madrid, Publica-ciones Universidad de Comillas, 1987.
—, La percepción visual. La arquitectura del psiquismo desde el enfoque de la percepción visual, Madrid, Biblioteca Nueva, 1998.
—, «Engramas neuronales y teoría de la mente», en Pensamiento, 57 (2001), n.º 218, 177-211.
MONTAGU, A., ¿Qué es el hombre?, Buenos Aire, Paidós. 1966.MOORE, John H., «¿Cómo explicar la diversidad humana?», Mundo Científico,
1995, n.º 160, 717-723.MORA, Fco. (ed.), El problema cerebro-mente, Madrid, Alianza, 1995.MOREY, M., «El hombre como argumento», Barcelona, Anthropos, 1987.MORGAN LLAMAN, J., El cerebro en evolución, Barcelona, Ariel, 2003.MORREN, L., «De la loi d’entropie au principe anthropique», Révue Théologique
de Louvain, 1984, pp. 160-183.MORRIS, D., El mono desnudo. Un estudio del animal humano, Barcelona, Plaza
y Janés, 1968.MORWOOD, M. J. y otros, «Archaeology and Age of a New Hominin from Flores in
Eastern Indonesia, Natrure, vol. 431, 28 de octubre de 2004, pp. 1.087-1.091.MOSTERIN, Jesús, «Examen del principio antrópico en cosmología», Diálogos, 79
(2002), n.º 79, 203-236.—, La naturaleza humana, Madrid, Espasa-Calpe, 2006.MOYA, A., «La genética de poblaciones y la trazabilidad genética de la historia de
la vida: el poder de los marcadores moleculares», Ludus Vitalis, VIII (2000), n.º 13, 157-161.
MOYA, C., Filosofía de la mente, Valencia, Universitat de València, 2004.MURILLO, Ildefonso (coord.), Filosofía práctica y persona humana, Salamanca,
Publicaciones Universidad Pontificia de Salamanca, 2004.NAGEL, E., The Structure of Science, Nueva York, Harcourt, Brace and World,
1961.NAPIER, J., Scientific American, 1962, 205, 2.NICOLAU POUS, Fr.. Iglesia y ciencia a lo largo de la historia, Barcelona, Ed. Cla-
ret, 2002 (Barcelona, Ed. SCIRE, 2003).NISHIDA, T., «The social group of chimpanzees in the Mahale Mountains», Pri-
mates, 1968, 9, 167-224.NOBLE, Denis, La música de la vida. Más allá del genoma humano, Madrid, Akal,
2008.NOBLE, W./DAVIDSON, I., Human Evolution. Language and Mind. Cambridge,
Cambridge University Press, 1996.NUMBERS, R. L., Darwinism comes to America, Cambridge, MA, Harvard Uni-
versity Press, 1998.NÚÑEZ, Diego, «La religión y la ciencia (Historia de las controversias entre am-
bas)», Mundo Científico, 1996, n.º 166, 247-256.—, Introducción a Ch. DARWIN, El origen de las especies por medio de la selec-
ción natural, Madrid, Alianza, 2009, pp. 26-33.

533
© Universidad de Deusto - ISBN 978-84-9830-383-4
NÚÑEZ DE CASTRO, Ignacio, Análisis antropológico del Proyecto Genoma Hu-mano, 1996, p. 12 (manuscrito sin publicar).
—, «La indefinición fundamental de las ciencias biológicas», en DOU, A. (ed.), Fragmentariedad de las ciencias, Bilbao, Mensajero/Biblioteca Fomento So-cial, 1985, 143-147.
NUSSBAUM, Martha C., Las mujeres y el desarrollo humano, Barcelona, Herder, 2002.
NÜSSLEIN-VOLHARD, Christiane, «Gradientes organizadores del desarrollo del em-brión», Investigación y Ciencia, 1996, n.º 241, 20-27.
—, Génesis y desarrollo de la vida, Barcelona, Crítica, 2009.OAKLEY, K., «Emergence of higher thougt 3.0-0.2 Ma B.P.», Phil. Trans. R. Soc.
Lond., 1981, B292, 205-211.O’HEAR, A., Beyond evolution: human nature and the limits of evolutionary ex-
planation, Oxford, Oxford University Press, 1997.OPARIN, A.I., Proishkhozhddenie Zhisni, Moskowski Rabocii, 1924.—, The Origin of Live, Dover Publications, 1953 (trad. esp.: El origen de la vida,
Madrid, Akal, 1994).—, The Origin of Live on Erth, Academic Press, 1957, 3.ª ed. (trad. esp.: El ori-
gen de la vida sobre la Tierra, Madrid, Tecnos, 1979).ORTEGA Y GASSET, J., Historia como sistema, Madrid, Espasa-Calpe, 1935
(1971).—, Meditación sobre la técnica, Madrid, Espasa-Calpe, 1965.—, «Ni vitalismo ni racionalismo» (1924), Obras Completas, vol, III (Madrid,
Revista de Occidente, 1966), pp. 270-280.PACHO, Julián, ¿Naturalizar la razón? Alcance y límites del Naturalismo Evolu-
cionista, Madrid, Siglo XXI, 1995.PALEY, W., Natural Theology, Nueva York, American Tract Society, 1802.PALMA MARTÍNEZ, J. M., «Proteómica, un nuevo palabro», El País, 2008, 30 de
abril, p. 45.PANNENBERG, W., Antropología en perspectiva teológica, Salamanca, Sígueme,
1993.PARIS, Carlos, «La sociobiología y el concepto de cultura», Theoría, II época,
vol. VII, 1992, n.º 16-17-18, tomo B, pp. 769-878.—, El animal cultural. Biología y cultura en la realidad humana, Barcelona, Crí-
tica, 1994.PATERSON, Colin, Evolución. La teoría de Darwin hoy, Barcelona, Fontalba,
1991.PENROSE, R., La nueva mente del emperador, Madrid, Mondadori, 1991.—, Las sombras de la mente, Barcelona, Crítica, 1996.PÉREZ DE LABORDA, A., La ciencia contemporánea y sus implicaciones filosófi-
cas, Madrid, Cincel, 1985.—, «El principio antrópico», Diálogo Filosófico, 7 (1991), n.º 1, 46-57.PÉREZ TAPIAS, J. A., Filosofía crítica de la cultura, Madrid, Trotta, 1995 (2000:
2.ª ed.).PINEDA, José Avelino de la, El baile de la ciencia y la metafísica. Respuesta a Ste-
phen Hawking, Madrid, Biblioteca Nueva, 2008.

534
© Universidad de Deusto - ISBN 978-84-9830-383-4
PINKER, S./BLOOM, P. (1990), «Natural language and natural selection», Behavio-ral and Brain Science, 13, 707-727.
PINKER, S. (1994), The language instinct. How the mind creates language,New York, W. W. Norton (El instinto del lenguaje. Cómo crea el lenguaje la mente, Madrid, Alianza, 1995).
—, The Blank Slate, Nueva York, Penguin, 2002 (trad. cast., La tabla rasa. La ne-gación moderna de la naturaleza humana, Madrid, Alianza, 2003).
—, The language instinct, Nueva York, William Morrow, 1994 (El instinto del lenguaje, Madrid, Alianza, 2001); Id., El mundo de las palabras, Barcelona, Paidós, 2007.
PÍO XII, Encíclica Humani Generis, 1950, en DS, 3897 (Documentos bíblicos, n. 703):
PLOMIN, R., DE FRIES, J. C., MECCLEAM, G. E. y MCGUFFIN, P., Genética de la conducta, Barcelona, Ariel, 2002.
POITIER, P. y otros, «La importancia de ser imperfectos», Investigación y Ciencia,1994, n.º 217, 67-79.
POLKINGHORNE, John, Ciencia y Teología. Una introducción, Santander, Sal Te-rrae, 2000.
—, Explorar la realidad. La interrelación de ciencia y religión, Santander, Sal Terrae, 2007.
—, La fe de un físico. Reflexiones teológicas de un pensador ascendente, Estella (Navarra), Verbo Divino, 2007.
— (ed.), La obra del amor. La creación como kénosis, Estella (Navarra), Verbo Divino, 2008.
POPPER, K. R., La miseria del historicismo, Madrid, Alianza, 1973.—, «Darwinism as a metaphysical research programme», sección 37 de la obra de
K. R. Popper Unended Quest, citada en la nota 14, «Evolution and the tree of knowledge», sección 7 de la obra de Popper Objective Knowledge. An Evo-lutionary approach (Oxford University Press, edición revisada, 1974). Trad. cast.: Conocimiento objetivo, Madrid, Tecnos, 1974).
—, El desarrollo del conocimiento científico. Conjeturas y refutaciones, Barce-lona, Paidós Ibérica, 1982.
— y ECCLES, J., El yo y su cerebro, Barcelona, Labor, 1980.—, La sociedad abierta y sus enemigos, Barcelona, Paidós Ibérica, 1982.—, El universo abierto. Un argumento en favor del indeterminismo, Madrid, Tec-
nos, 1984.PORTMANN, A., Die Biologie und das neue Menschenbild, Berna, 1942.—, Zoologie und das neue Bild des Menschen, Hambug-Reinbek, 1956.PREBOSTI, Antonio, «Concepto de especie en el darwinismo actual», Mundo Cien-
tífico, 1993, n.º 141, 1.040-1.049.PREMACK, David, «Minds with and wihtouth language», en WEISKRANTZ, L. (ed.),
Thought without language, Clarendon, 1988, 46-65.— y WOODRUFF, G., «Does the chimpanzee have a theory of mind?», Behavioral
and Brain Sciences, 1978, n.º 1, 515-526 (trad. cast.: «¿Tiene el chimpancé una teoría de la mente?», en MARTÍ, E., Construir una mente, Barcelona, Pai-dós, 1997, pp. 137-178.

535
© Universidad de Deusto - ISBN 978-84-9830-383-4
—, «Is language the key to human intelligencie?», Science, 2003, 303, pp. 318-320.
PRIEST, S., Teorías y filosofías de la mente, Madrid, Cátedra, 1994.PRIETO LÓPEZ, Leopoldo, El hombre y el animal. Nuevas fronteras de la Antropo-
logía, Madrid, BAC, 2008.PRIGOGINE, Ilya-STENGERS, I., La nueva alianza. Metamorfosis de la ciencia, Ma-
drid, Alianza, 1983.—, ¿Tan sólo una ilusión?, Barcelona, Tusquets, 1983.PUJOL GEBELLÍ, Xavier, «La cresta neural juega un papel clave en la evolución»
(entrevista a la embrióloga Nicole Ledouarin), El País, 2002, 24 de abril, p. 33.
PUYTORAC, Pierre de, De la biophilosophie à une éthique de la biologie (La so-cieté face à la biologie), París, L’Harmattan, 1998.
QUERALTÓ, Ramón, «El nuevo orden del tiempo. En torno a la obra de Ilya Prigo-gine», en ARANA, J. (ed.), La filosofía de los científicos, Sevilla, Thémata. Re-vista de Filosofía, 1995, n.º 14, pp. 305-320.
RABOSSI, E. (comp.), Filosofía de la mente y ciencia cognitiva, Barcelona, Pai-dós, 1995.
RADL, E. M., Historia de las teorías biológicas, Madrid, Alianza, 2 vols., 1988.RANK, Otto, The trauma of birth, Nueva York, 1929.REBATO, E. y otros (eds.), Para comprender la Antropología biológica, Estella
(Navarra), Verbo Divino, 2005, 497-511.REGAN, T., Animal Rights, Berkeley, University of California Press, 1983; Id.,
«Poniendo a las personas en su sitio», Teorema, XVIII/2 (1999), 17-37.REILEY MAGUIRE, M., «Personhood, Covenant, and Abortion», en JUNG, P./SHAN-
NON, T. (eds.), Abortion and Catholicism, Nueva York, Crossroad, 1988, 100-120.
RIAZA, José María, La Iglesia en la historia de la ciencia, Madrid, BAC, 1999.RICOEUR, Paul, El mal. Un desafío a la filosofía y a la teología, Buenos Aires,
Amorrortu, 2006.RICHARDS, R. J., Darwin and the emergence of evolutionary theorie of mind and
behavior, Chicago, University of Chicago Press, 1987.RICHTER SHERMAN, Claire, Writing on Hands, Washington, The Folger Shake-
speare Library, 2000.RIEDLEY, Matt, Genoma. Autobiografía de una especie en 23 capítulos, Madrid,
Taurus, 2000.RIU, Ester, «El embarazo cambió la columna de las mujeres», El País, 11 de
marzo de 2008, p. 56.RIVERA, Alicia, «Los dedos son una innovación y no derivan de las aletas de los
peces», El País, 28 de junio de 1995, p. 29.—, «Un hacha hallada en Atapuerca indica que ya había ritos funerarios hace
400.000 años», El País, 8 de enero de 2003, p. 26.RIZZOLATTI, G./SINIGAGLIA, C., Las neuronas espejo, Barcelona, Paidós, 2006.ROCHE, H./TIERCELIN, J. J., Acad. Sc. París, serie D, 1871, 1977.ROF CARBALLO, J., —, «El futuro del hombre», en CRUSAFONT, M. y otros, La
evolución, Madrid, BAC, 1966.

536
© Universidad de Deusto - ISBN 978-84-9830-383-4
—, «El hombre como encuentro», en Homenaje a Zubiri, vol. 2, Madrid, Ed. Mo-neda y Crédito, 1970, pp. 587-616.
—, El hombre como encuentro, Madrid, Alfaguara, 1973.ROGER FISCHER, Steven, Breve historia del lenguaje, Madrid, Alianza, 2003.ROSNY, J. H. (Joseph-Henry-Honoré Boex (1856-1940) y Seraphin-Justin-
François Boex (1859-1948)), En busca del fuego, El león de las cavernas, y Vamireh, Madrid, Valdemar, 2001 (Presentación de Alfredo Lara López).
ROSTAND, J., Introducción a la historia de la biología, Barcelona, Planeta, 1985.RUBIA, Fr. J., La conexión divina. La experiencia mística y la neurobiología, Bar-
celona, Crítica, 2003.RUBIO CARRACEDO, J., «El hombre y la ética», Barcelona, Anthropos, 1987.RUFFIÉ, J., «Les frontières cromosomiques de l’hominisation», en C.-R. Acad.
Sc., 1973, 276, seri D, 1709.—, De la Biologie á la Culture, Paris, Flammarion, 1976 (De la biología a la cul-
tura, Barcelona, Muchnik Editores, 1982).RUIZ, Rosaura/AYALA, Fco. J., El método en las ciencias. Epistemología y darwi-
nismo, México, FCE, 1998.RUIZ DE ELVIRA, Malen, «Recientes experimentos niegan la formación de neuro-
nas nuevas», El País, 13 de enero de 2002, p. 27.RUIZ DE LA PEÑA, J. L., Las nuevas antropologías, Santander, Sal Terrae, 1983.—, Teología de la creación, Santander, Sal Terrae, 1986.—, Qué hay del pecado original, Madrid, Fundación Santa María, 1992.RUSE, Michael, Filosofía de la biología, Madrid, Alianza, 1979.—, Sociobiología, Madrid, Cátedra, 1983.—, Tomándose a Darwin en serio, Barcelona, Salvat, 1987.—, El misterio de los misterios. ¿Es la evolución un constructo social?, Barce-
lona, Tusquets, 2001.—, ¿Puede un darwinista ser cristiano?, Madrid, Siglo XXI, 2007.—, Charles Darwin, Buenos Aires, Katz Editores, 2008.SABATER PI, J., Etología de la vivienda humana. De los nidos de gorilas y chim-
pancés a la vivienda humana, Barcelona, Labor, 1985.SAGAN, Carl, Los dragones del Edén. Especulaciones sobre la evolución de la in-
teligencia humana, Barcelona, Crítica, 1993.SAHLINS, M., Uso y abuso de la biología, Madrid, Siglo XXI, 1982.—, Cultura y razón práctica, Barcelona, Gedisa, 2006.SALGADO, Alfonso, Biología del comportamiento. Introducción a la Psicología
biológica. Una propuesta para la convergencia, Salamanca, Universidad Pon-tificia, 2007.
SAMPEDRO, Javier, Deconstruyendo a Darwin, Barcelona, Crítica, 2002.—, «Entrevista a Peter Singer», El País (Babelia), 11 de mayo de 2002, p. 12.—, El siglo de la ciencia. Nuestro mundo al descubierto, Barcelona, Península,
2009, 58.SAN MARTÍN, Javier, Para una superación del relativismo cultural. Antropología
cultural y Antropología filosófica, Madrid, Tecnos, 2009.SÁNCHEZ CAPDEQUÍ, Celso, Imaginación y sociedad: una hermenéutica creativa
de la cultura, Madrid/Pamplona, Tecnos/UPNA, 1999.

537
© Universidad de Deusto - ISBN 978-84-9830-383-4
SÁNCHEZ DE ZABALA, V. (ed.), Sobre el lenguaje de los antropoides, Madrid, Si-glo XXI, 1976, 147-203.
SANGUINETTI, J. J., «Algunos aspectos de la filosofía de los cosmólogos contem-poráneos», en ARANA, J. (ed.), La filosofía de los científicos, Sevilla, Thé-mata. Revista de Filosofía, 1995, n.º 14, pp. 285- 304.
SANMARTÍN, J./SIMÓN, V./GARCÍA-MERITA, M.ª L. (comps.), La sociedad naturali-zada. Genética y conducta, Valencia, Tirant Lo BlancH, 1986.
SAPOLSKY, El mono enamorado y otros ensayos sobre nuestra vida animal, Bar-celona, Paidós, 2007.
SAVAGE-RUMBAUGH, E. Sue, Kanzi: The ape at the brink the human mind,Nueva York, Wiley, 1994..
— y R. LEWIN, Kanzi: The ape at the brink of the human mind, New York, Wiley, 1994.
— y RUMBAUGH, D. M., «Perspectives on consciousness, language, and other emergent proceses in apes and humans», en HAMEROFF, S. R./KASZNIAC, A. W./SCOTT, A. (eds.), Towards a scicence of consciousness II: The second Tucson discussions and debates, Cambridge, Mass., MIT Press, 1998, pp. 533-549.
— y SHANKER, S./TAYLOR, T. J., Apes, language, and th human mind,Nueva York, Oxford University Press, 1998.
—, «Why are we afraid of apes with language?», en SCHEIBEL, A. B./SCHOPF,J. W. (eds.), The origin of evolution of intelligence, Boston, Jons and Bartlett, 2001, pp. 43-69.
SAVAGE-RUMBAUGH et alii, «Linguistically mediated tool use ad exchange by chimpanzees», Behavioral and Brain Sciences, 1978, 1 (4), 539-545.
— /SHANKER, S./TAYLOR, T. J., «Apes with language», Critical Quarterly, 1996, n.º 38, pp. 45-57.
—, —, —, Apes, language, ande the human mind, Nueva York, Oxford Univer-sity Press, 1998.
SCHIMITZ-MOORMANN, Karl (en colaboración con James F. SALMON, S. J.), Teolo-gía de la creación de un mundo en evolución, Estella, Verbo Divino, 2005.
SCHOCKENHOFF, Ethik des Lebens, M. Grünewald, Maguncia, 1993.SCHOPF, T. (ed.), Models in Paleobiology, San Francisco, Freeman, 82-115.SCHWOERBEL, W., Evolución, Barcelona, Salvat, 1986, pp. 56-60.SCOTT, E. C., Evolution vs. Creationism. An Introduction, Westport, CT, Green-
wood Press, 2004.SEARLE, J., Mente, cerebros y ciencia, Madrid, Cátedra, 1985.—, El misterio de la conciencia, Barcelona, Paidós, 2000.SECHÉ, Andreas, «Así sentamos cabeza», Muy Interesante, 1996, n.º 27, pp. 93 y ss.SENUT, B. et al., «First hominid from the Miocene (Lukeino Formation, Kenia)»,
C. R. Acad. Sci., 2001, 332: 137-144.SEQUEIROS, Leandro, «Arnold Gehlen (1904-1976). La búsqueda de una funda-
mentación biológica para la antropología filosófica», en ALARCOS MARTÍ-NEZ, F. J. (ed.)., La moral cristiana como propuesta. Homenaje al profesor Eduardo López Azpitarte, Madrid, San Pablo, 2004, 683-700.
—, Raíces de la humanidad. ¿Evolución o creación?, Santander/Madrid, Sal Te-rrae/Fe y Secularidad, 1992.

538
© Universidad de Deusto - ISBN 978-84-9830-383-4
—, «Cuando hablamos de evolución biológica, ¿de qué evolución estamos ha-blando? Implicaciones teológicas, Proyección. Teología y mundo actual, LIV (2007), n.º 224, 29-47.
—, «Dioses, monos y humanos: La respuesta de la Ciencia y de la Biblia a los orígenes de la humanidad», en VV.AA., Biblia y Cultura, Bilbao, Universidad de Deusto, 2008, pp. 97 y ss.
—, ¿Puede un cristiano ser evolucionista?, Madrid, PPC, 2009.—, El diseño chapucero. Darwin, la biología y Dios, Madrid, Ediciones Khaf,
2009.SEYES, J. A., Antropología del hombre caído. El pecado original, Madrid, BAC,
1991; Concilium, 2004, n.º 304.SCHMITZ-MOORMANN, K. (con la colaboración de J. F. SALMON, S.J.), Teología de
la creación de un mundo en evolución, Estella, Verbo Divino, 2005.SHAFFER, D. R., Psicología del desarrollo. Infancia y adolescencia, Madrid, In-
ternational Thomson Editores, 2005 (5.ª ed.).SHERMER, Michael, Las fronteras de la ciencia. Entre la ortodoxia y la herejía,
Barcelona, Alba Editorial, 2010.SHI, Yun-Bo/STOLOW, Melisss/PUZIANOWSKA-KUZNICKA, Monila/WONG, Jiemin,
«Los genes de la metamorfosis. Una hormona, director de una orquesta de genes, dirige el remodelaje anatómico», Mundo Científico, 1996, n.º 169, pp. 538-544.
SIMPSON, G. G., «The Principles of Classification and a Classification of Mam-mals», Bulletin of the American Museum of Natural History, 85: 1-350.
—, The major features of evolution, Columbia University Press, 1953.SHMITH, John Maynard, La construcción de la vida. Genes, embriones y evolu-
ción, Barcelona, Crítica, 1998.SINGER, P./CAVALIERI, P. (eds.), El proyecto «Gran Simio». La igualdad más allá
de la humanidad, Madrid, Trotta, 1998.SINGER, P., Etica práctica, Barcelona, Ariel, 1984.—, Repensar la vida y la muerte: el derrumbe de la ética tradicional, Barcelona,
Paidós, 1997.—, Liberación animal, Madrid, Trotta, 1999.—, Una izquierda darwiniana. Política, evolución y cooperación, Barcelona, Crí-
tica, 2000.—, Una vida ética: escritos, Madrid, Santillana, 2000.—, Desacralizar la vida humana. Ensayos sobre ética, Madrid, Cátedra, 2003.—, Un solo mundo. La ética de la globalización, Barcelona, Paidós, 2003.SOBER, E. y WILSON, D. S., El comportamiento altruista. Evolución y psicología,
Madrid, Siglo XXI, 2000.SOLER GIL, F. J. (ed.), Dios y las cosmologías modernas, Madrid, BAC, 2005.SOLER, Manuel, Evolución. La base de la biología, Granada, Proyecto Sur Edi-
ciones, 2002.SOLER, Jean, «Las razones de la Biblia: reglas alimenticias hebreas», en FLAN-
DRIN, J.-L./MONTANARI, M. (dirs.), Historia de la alimentación, Gijón, Edic. Trea, S.L., 2004, 81-94.
SPITZ, R. A., No y sí. Sobre la génesis de la comunicación humana, Buenos Aires, Ediciones Hormé, S.A. E./Paidós, 1972 (3.ª ed.).

539
© Universidad de Deusto - ISBN 978-84-9830-383-4
STRAUSS, Evelyn, «El tiempo variable de los relojes de DNA», Mundo Científico,1999, n.º 204, 20-23.
SULSTON, J. y FERRY, G., El hilo común de la humanidad. Una historia sobre la ciencia, la política, la ética y el genoma humano, Madrid, Siglo XXI, 2003.
SUSANNE, Ch./REBATO, E./DELIGNE, J, «Razas y racismo», en REBATO, E./SU-SANNE, Ch./CHIARELLI, B. (eds.), Para comprender la Antropología biológica. Evolución y biología humana, Estella, Verbo Divino, 2005, pp. 655-664.
SYKES, Bryan, Las siete hijas de Eva, Madrid, Debate, 2001.STRINGER, Ch. B., «¿Está en África nuestro origen?», Investigación y Ciencia,
1991, febrero.SUSMAN, R./MCGRAW, S. A., «¿Quién fabricó las primeras herramientas?»,
Mundo Científico, 1995, n.º 159, 682-683.TAYLOR, Ch., La libertad de los modernos, Buenos Aires/Madrid, Amorrotu Edi-
tores, 2005.TEILLET ROLDÁN, Eduardo, Raza, identidad y ética, Barcelona, Ediciones del Ser-
bal, 2000.THOM, René, Estabilidad estructural y morfogénesis. Ensayo de una teoría gene-
ral de los modelos, Barcelona, Gedisa, 1987.THORPE, W. H., Breve historia de la etología, Madrid, Alianza, 1982.TOBIAS, P., The Brain in Hominid Evolution, Nueva York, Columbia University
Press, 1971.TORO, M. A./LÓPEZ-FANJUL, C., «Indeterminaciones del neodarwinismo», Teo-
rema, XII/3 (1982), 251-274.TORRALBA ROSELLO, Fr., ¿Qué es la dignidad humana? Ensayo sobre Peter Sin-
ger, Hugo Tristram Engelhardt y John Harris, Barcelona, Herder, 2005.TORRES QUEIRUGA, A., Recuperar la creación. Por una religión humanizadora,
Santander, Sal Terrae, 1996.TORT, Patrick (dir.), Dictionnaire du darwinisme et de l’évolution, 3 vols., París,
PUF, 1996.—, Para leer a Darwin, Madrid, Alianza, 2000.TORTAJADA MARTÍNEZ, Miguel, El parto en la evolución humana, Valencia, Uni-
versitat de Valencia, 2001.TOWNSEND, Rosa, «La regeneración cerebral abre nuevas fronteras», El País, 23
de noviembre de 1999, p. 39.TREVARTHEN, C., «Cerebro, desarrollo del», en GREGORY, R. L. (ed.), Diccionario
Oxford de la mente, Madrid, Alianza, 1995, pp. 149-161.TREVIJANO ECHEVERRÍA, M., ¿Qué es la bioética?, Salamanca, Sígueme, 1999.TRINKAUS, Erik, «El cuello del fémur: ¿un indicador social para los antropólo-
gos?», Mundo Científico, 1994, n.º 151, 982-983.TUDGE, Colin/YOUNG, Josh, El eslabón. La historia del fósil de primate más anti-
guo del mundo, Barcelona, Debate, 2010.TUGENDHAT, E., «La moral en perspectiva evolucionista», en Id., Problemas, Bar-
celona, Gedisa, 2001, pp. 135-158.TUTTLE, R. H./WEBB, D. M./TUTTLE, N. I., «Laetoli Footprint Trails and the Evo-
lution of Hominid Bipedalism», en COPPENS, Y./SENUT, B. (eds.), Origine(s) de la bipedie chez les hominidés, París, Editions du CNRS, 1991, 187-198.

540
© Universidad de Deusto - ISBN 978-84-9830-383-4
TYLOR, E. B., «On a Method of Invertigating the Development of Institutions: Applied to Laws of Married and Descent», Journal of the Anthropological In-stitute, 18 (1888).
UBEDA PURKISS, M., «Datos para una antropología del hombre en pie», EstudiosFilosóficos, XL (1991), n.º 113, 7-32.
UDIAS VALLINA, Agustín, El universo, la ciencia y Dios, Madrid, PPC, 2001.—, Ciencia y religión. Dos visiones del mundo, Santander, Sal Terrae, 2010.VALLEJO, Fernando, La tautología darvinista y otros ensayos de biología, Madrid,
Taurus, 2002.VANDERMEERSCH, Bernard, «Homo sapiens sapiens: lo que dicen los fósiles»,
Mundo Científico, 1995, n.º 160, pp. 710-716.VIDAL, Marciano, Pecado estructural y responsabilidad personal, Madrid, Fun-
dación Santa María, 1991.— (ed.),Conceptos fundamentales de ética teológica, Madrid, Trotta, 1992.—, «El status humano del embrión. Interrogantes y afirmaciones de la teología
católica actual sobre el comienzo de la vida humana», en VIDAL, M., Bioética.Estudios de bioética racional, Madrid, Tecnos, 1998 (3.ª ed.), 37-49.
—, Bioética. Estudios de bioética racional, Madrid, Tecnos, 1998 (3.ª ed.).VIDAL, José Manuel, «Darwin, perdónalos. 2.000 años de errores de la Iglesia»,
El Mundo, 1996, 27 de octubre.VINAGRE, Fco./VINAGRE ARIAS, A., Materiales y tecnología en las primeras civili-
zaciones, Calamonte (Badajoz), Edit. Filarias, 2002.VISALBERGHI, E., «Los simios y la herramienta: ¿una cuestión de inteligencia?»,
Mundo Científico, 1989, n.º 97, 1.264-1.267.VRBA, Elisabeth, «Evolution, Species and Fossils: How does Live Evolve?»,
S. Afr. J. Sci., 1980, 76 (2): 61-84.VV.AA., «De la mirada al gesto. Cómo dirige el cerebro la mano», Mundo cientí-
fico, 1998, n.º 192, 25-37.VV.AA., Identidad y estatuto del embrión humano, Madrid, Ediciones Internacio-
nales Universitarias, 2000.VV.AA., Evolución y fe, Concilium, 2000, n.º 284 (número monográfico).VV.AA., «Teología y creación», Iglesia Viva, número monográfico 183, 1996.VV.AA., Evolución hoy, Barcelona, Tusquets, 2001. .VV.AA., «Atapuerca», número monográfico de Journal of Human Evolution, 37
(1999), n.º 3-4, sept.-oct.VV.AA., «La conducta de los primates», número monográfico de Investigación y
Ciencia, 2003, Temas n.º 32.VV.AA., La biología como arma social, Madrid, Edit. Alhambra, 1982.WADE, Nicholas, «Los científicos ven la inmortalidad en células que se dividen
indefinidamente. Dos hallazgos de un empresa de EEUU dan un giro al estu-dio del envejecimiento», El País, 1999, 3 de enero, p. 34.
WAGENSBERG, Jorge (ed.), Proceso al azar, Barcelona, Tusquets, 1986.WALKER, A./SHIPMAN, P., The wisdom of bones: in search of human origins, Lon-
dres, Weidenfeld and Nicolson, 1996.WATSON, J. D., «The Human Genome Project: past, present and future», Science,
248. 1990, 44-48.

541
© Universidad de Deusto - ISBN 978-84-9830-383-4
—, La doble hélice, Madrid, Alianza, 2000.—, Genes, chicas y laboratorios, Barcelona, Tusquets, 2006.WEISKRANTZ, L. (ed.), Thought without language, Clarendon, 1988.WASHBURN, S. L./LANCASTER, C. S., «The Evolution of Hunting», en R. B. LEE e
I. DE VORE (eds.), Man the Hunter, Chicago, III, Aldine Publishing Co., 1968, pp. 293-303.
WHITE, T. D., «Los australopitecinos», Mundo Científico, 1983, n.º 21, pp. 18 y ss.WHITE, Tim D./SUWA, Gen/ASFAW, Berhane, «Australopithecus ramidus, a
new sepcies of early hominid from Aramis, Ethiopia», Nature, 371 (1994), n.º 6.495, 13 de sept., 306-312; cfr. también pp. 313-333 y 280-281.
WILSON, Allan C./CANN, Rebecca L., «Origen africano reciente de los humanos», Investigación y Ciencia, 1992, junio, recopilado por J. BERTRANPETIT (comp.), o.c., 20-25.
WILSON, Frank R., La mano. De cómo su uso configura el cerebro, el lenguaje y la cultura humana, Barcelona, Tusquets, 2002.
WILSON, E. O., Sociobiología. La nueva síntesis, Barcelona, Omega, 1975.—, On Human Nature, Cambridge, Massachussets, 1978 (Sobre la naturaleza hu-
mana, México, FCE, 1980).—, Consilience. La unidad del conocimiento, Barcelona, Galaxia Gutemberg,
1999.WILSON, P. A., «What is the Explanandum in the Anthropic Principle», Am. Phi-
los. Quarterly, 28 (1991), n.º 3, 167-174.WONG, Kate, «El Hombre de Flores», Investigación y Ciencia, abril de 2005,
n.º 343, pp. 22-31.—, «Nueva luz sobre el Hombre de Flores», Investigación y Ciencia, 2010,
marzo, n.º 400, pp. 60-67.WRIGHT, W., Así nacemos. Genes, conducta, personalidad, Madrid, Taurus, 2000.WYN DAVIES. Merryl, Darwin y el fundamentalismo, Barcelona, Gedisa, 2004.YOUNG, David, El descubrimiento de la evolución, Barcelona, Edic. del Serbal,
1998.ZIMMER, Dieter E., Dormir y soñar. La vida nocturna de nuestras vidas, Barce-
lona, Salvat, 1986.ZUBIRI, X., Sobre la esencia, Sociedad de Estudios y Publicaciones, Madrid,
1972.—, Inteligencia sentiente. Inteligencia y realidad, Madrid, Alianza/Sociedad de
Estudios y Publicaciones, 1980.—, Inteligencia y logos, Madrid, Alianza/Sociedad de Estudios y Publicaciones,
1982.—, Inteligencia y razón, Madrid, Alianza/Sociedad de Estudios y Publicaciones,
1983.—, El hombre y Dios, Madrid, Alianza/Sociedad de Estudios y Publicaciones,
1985.—, Sobre el hombre, Madrid, Alianza/Sociedad de Estudios y Publicaciones,
1986.—, Estructura dinámica de la realidad, Madrid, Sociedad de Estudios y Publica-
ciones, 1989.

© Universidad de Deusto - ISBN 978-84-9830-383-4
Carlos BeorleguiLa singularidad de la especie humana
De la hominización a la humanización
El darwinismo supuso un cambio definitivo en la compresión del ser humano, aunque como teoría científica no implica renunciar a una
concepción singular de la especie humana. Somos animales, pero animales bio-culturales, dotados de autoconciencia y libertad, de la capacidad y
necesidad de completar con nuestras decisiones lo que la naturaleza nos ha dado, y de poder plantearnos el sentido de nuestra existencia y abrirnos a la
pregunta por el fundamento de todo lo que hay.
DeustoUniversidad de Deusto