Índice general resumen i introducciÓn ii objetivos … version de... · capitulo iv resultados y...
TRANSCRIPT

ÍNDICE GENERAL
RESUMEN ........................................................................................................................ I
INTRODUCCIÓN .............................................................................................................. II
OBJETIVOS ..................................................................................................................... III
HIPÓTESIS...................................................................................................................... IV
JUSTIFICACIÓN ............................................................................................................... V
CAPITULO I
MARCO TEORICO
1.1 ANTECEDENTES ...................................................................................................... 1
1.1.1 Internacionales: ........................................................................................................ 1
1.1.2 Nacionales: ............................................................................................................... 3
1.1.3 Locales: ..................................................................................................................... 7
1.2 MARCO CONCEPTUAL .......................................................................................... 11
1.2.1 Bosque montano .................................................................................................... 11
1.2.2 Gradientes ambientales ......................................................................................... 11
1.2.3 Ciclo de carbono en un bosque .............................................................................. 12
1.2.4 Reservorios de carbono en un bosque montano .................................................... 14
1.2.5 RAINFOR ................................................................................................................. 15
1.2.6 Parcela permanente ............................................................................................... 16
1.2.6.1 Dinámica de bosques montanos ................................................................ 16
1. Reclutamiento ....................................................................................................... 16
2. Mortalidad ............................................................................................................. 17
1.2.6.2 Biomasa vegetal ........................................................................................ 19
1. Metodologías para cuantificar biomasa ................................................................ 19
2. Biomasa aérea ....................................................................................................... 23
3. Relación biomasa – carbono ................................................................................. 23
1.2.6.3 Analisis poblacional ................................................................................... 23
1. Composición florística ........................................................................................... 23
2. Importancia ecológica ........................................................................................... 24
3. Diversidad .............................................................................................................. 24
CAPITULO II
ÁREA DE ESTUDIO
2.1. UBICACIÓN .......................................................................................................... 25
2.1.1. Ubicación Política ............................................................................................... 25
2.1.2. Ubicación Geográfica ......................................................................................... 25
2.2. LIMITES ................................................................................................................ 25
2.3. ACCESIBILIDAD ..................................................................................................... 25
2.4. SANTUARIO HISTÓRICO DE MACHUPICCHU .......................................................... 28

2.4.1. Componente físico .............................................................................................. 28
2.4.1.1. Geología .................................................................................................... 28
2.4.1.2. Geomorfología .......................................................................................... 29
2.4.1.3. Fisiografía .................................................................................................. 29
2.4.1.4. Edafología.................................................................................................. 31
2.4.1.5. Hidrografía ................................................................................................ 31
2.4.2. Componente climático ........................................................................................ 33
2.4.3.1. Climadiagrama. ......................................................................................... 33
2.4.4. Componente biológico ........................................................................................ 35
2.4.4.1. Zonas de vida ............................................................................................. 35
2.4.4.2. Sistemas ecológicos ................................................................................... 38
2.4.4.3. Flora .......................................................................................................... 41
2.4.4.4. Fauna ........................................................................................................ 41
CAPITULO III
MATERIALES Y METODOLOGÍA
3.1. MATERIALES ........................................................................................................ 43
3.2. METODOLOGÍA .................................................................................................... 44
3.2.1. Ubicación de las parcelas ................................................................................... 44
3.2.2. Evaluación de las parcelas .................................................................................. 46
3.2.3. Analisis de datos ................................................................................................. 50
3.2.3.1. Dinámica poblacional del bosque .............................................................. 50
1. Reclutamiento ....................................................................................................... 50
2. Mortalidad ............................................................................................................. 51
3. Recambio (% de Turnover) .................................................................................... 51
3.2.3.2. Cuantificación de biomasa aérea almacenada ........................................... 51
1. Estimación de la cantidad de biomasa aérea individual ....................................... 51
3. Estimación de la cantidad de Carbono: ................................................................. 52
4. Prueba no paramétrica: U de Mann Whitney: ...................................................... 52
3.2.3.3. Análisis de composición floristica .............................................................. 53
3.2.3.4. Análisis de importancia ecológica .............................................................. 53
1. Índice de Valor de Importancia por especie (IVI´s) ............................................... 53
2. Índice de Valor de Importancia por familia (IVI´F) ................................................ 55
3. Análisis de la diversidad florística ....................................................................... 55
a) Diversidad alfa (α) ................................................................................................ 55
- Índice de diversidad de Shannon – Weiner (H’) ............................................ 56
- Índice de dominancia de Simpson (D) ........................................................... 57
b) Diversidad beta (β) ............................................................................................... 58
- Índice de similaridad de Morisita – Horn ...................................................... 58

CAPITULO IV
RESULTADOS Y DISCUSIONES
4.1. RESULTADOS ........................................................................................................ 59
4.1.1. Dinámica poblacional (Tasas de mortalidad, reclutamiento y recambio) ......... 59
1. Tipos de mortalidad por parcela ......................................................................... 62
4.1.2. Biomasa aérea y Carbono .................................................................................. 63
1. Biomasa aérea y carbono en cada censo (2013 y 2016) ......................................... 65
2. Distribución de la biomasa aérea y carbono ........................................................... 66
3. Dinámica de biomasa aérea y carbono almacenado ............................................... 68
4. Productividad primaria neta (PPN) .......................................................................... 70
4.1.3. Analisis poblacional ............................................................................................ 71
1. Composición florística ............................................................................................. 71
2. Importancia ecológica (IVI’s e IVIF) ......................................................................... 76
3. Análisis de diversidad .............................................................................................. 80
a) Diversidad alfa (α) ................................................................................................. 80
b) Análisis de riqueza de especies. ............................................................................ 82
c) Diversidad beta (β) ................................................................................................ 83
4.2. DISCUSIONES ....................................................................................................... 84
4.2.1. Dinámica poblacional ......................................................................................... 84
4.2.2. Biomasa aérea y carbono ................................................................................... 85
4.2.3. Analisis poblacional ............................................................................................ 87
4.2.3.1. Composición florística, IVI’s y diversidad ................................................... 87
CONCLUSIONES .............................................................................................................. 89
RECOMENDACIONES Y SUGERENCIAS ............................................................................. 91
BIBLIOGRAFÍA................................................................................................................. 92
ANEXOS ........................................................................................................................ 104

ÍNDICE DE TABLAS
Tabla 1. Datos disponibles sobre contenidos biomasa aérea, subterránea y suelo en
ecosistemas de bosques en la región andina. .................................................................. 14
Tabla 2. Casos especiales donde cambia el POM (Punto Óptimo de Medida) ........................ 21
Tabla 3. Datos geográficos de las parcelas permanentes de Wiñaywayna en el Santuario
Histórico de Machupicchu. ............................................................................................... 25
Tabla 4. Rutas de acceso al área de estudio. ........................................................................... 25
Tabla 5. Datos meteorológicos de 6 años de la estación de Aguas Calientes del Santuario
Histórico de Machupicchu. ............................................................................................... 33
Tabla 6. Zonas de vida consideradas para el Santuario Historico de Machupicchu. ............... 35
Tabla 7. Sistemas ecológicos considerados para el Santuario Historico de Machupicchu. ..... 38
Tabla 8. Familias de flora arbórea con mayor numero de especies registradas para el
Santuario Histórico de Machupicchu. ............................................................................... 41
Tabla 9. Especies de fauna registradas para el Santuario Histórico de Machupicchu. ............ 41
Tabla 10. Categoría de especies de fauna registradas para el Santuario Histórico de
Machupicchu. .................................................................................................................... 42
Tabla 11. Datos geográficos de ubicación de parcelas permanentes de Wiñaywayna ........... 44
Tabla 12. Ecuaciones alométricas utilizadas en los cálculos de biomasa aérea en las parcelas
permanentes de Wiñaywayna. ......................................................................................... 51
Tabla 13. Tasas de mortalidad, reclutamiento y recambio en cada parcela permanente. ..... 59
Tabla 14. Especies reclutadas en la parcela SHM - 01 durante el periodo de evaluación. ...... 61
Tabla 15. Especies reclutadas en la parcela SHM - 02 durante el periodo de evaluación. ...... 61
Tabla 16. Biomasa aérea y carbono almacenadao en cada una de las parcelas permanentes63
Tabla 17. Cambio de biomasa aérea y carbono en el periodo intercensal (2013 – 2016). ..... 68
Tabla 18. Dinámica de biomasa aérea mediante tasas de mortalidad, reclutamiento y
renovación. ....................................................................................................................... 69
Tabla 19. Productividad primaria neta de las parcelas evaluadas ........................................... 70
Tabla 20. Número de Familias, géneros, especies e individuos por parcela evaluada. ........... 71
Tabla 21. Especies registradas en la parcela SHM - 01. ........................................................... 72
Tabla 22. Especies registradas en la parcela SHM - 02. ........................................................... 73
Tabla 23. Número de géneros, especies e individuos por Familias en cada parcela para el
2016. ................................................................................................................................. 74
Tabla 24. Las 15 especies más importantes en la parcela SHM – 01 (Intipunku). ................... 76
Tabla 25. Familias con mayor valor de importancia ecológica en el bosque de Intipunku. .... 77
Tabla 26. Las 15 especies más importantes en la parcela SHM – 02 (Kantupata). .................. 78
Tabla 27. Familias con mayor valor de importancia ecológica en el bosque de Kantupata. ... 79
Tabla 28. Resultado de los Índices de diversidad..................................................................... 80
Tabla 29. Índices de Similaridad (%) de Morissita Horn. .......................................................... 83

ÍNDICE DE FIGURAS
Figura 1. Diagrama de los Stocks y flujos de carbono en un bosque ....................................... 13
Figura 2. Tipos de mortalidad de árboles en parcelas permanentes. ...................................... 19
Figura 3. Representación gráfica de casos especiales donde cambia el POM ......................... 22
Figura 4. Climatodiagrama del Santuario Histórico de Machupicchu. ..................................... 33
Figura 5. Disposición de las parcelas permanentes de 1.0 – ha en Wiñaywayna. ................... 46
Figura 6. Modo de desplazamiento dentro de las parcelas permanentes de Wiñaywayna .... 47
Figura 7. Ubicación de los arboles dentro de la sub parcela ................................................... 47
Figura 8. Ubicación de los individuos dentro de la sub parcela ............................................... 48
Figura 9. Individuos muertos y reclutas por parcela durante el periodo de evaluación. ........ 59
Figura 10. Tasas de mortalidad, reclutamiento y renovación por parcela. ............................. 60
Figura 11. Tipos de Mortalidad en la parcela SHM - 01. .......................................................... 62
Figura 12. Tipos de Mortalidad en la parcela SHM - 02. .......................................................... 62
Figura 13. Promedio de biomasa aérea y carbono acumulado en cada una de las parcelas
permanentes evaluadas. ................................................................................................... 63
Figura 14. Biomasa aérea y carbono almacenado por gradiente altitudinal. .......................... 64
Figura 15. Promedio de biomasa aérea acumulada en cada censo de las parcelas
permanentes. .................................................................................................................... 65
Figura 16. Promedio de carbono acumulado en cada censo de las parcelas permanentes. ... 66
Figura 17. Biomasa aérea y carbono por especies en la parcela SHM - 01 ............................. 66
Figura 18. Biomasa aérea y Carbono por especies de la parcela SHM - 02 ............................. 67
Figura 19. Variación de Biomasa aerea y carbono en el periodo intercensal de evaluación
(2013 – 2016) .................................................................................................................... 68
Figura 20. Tasas de mortalidad, reclutamiento y renovación de Biomasa aérea entre el 2013
al 2016 ............................................................................................................................... 69
Figura 21. Pérdida, ganancia de biomasa aérea y productividad primaria neta. .................... 70
Figura 22. Familias con mayor número de individuos en la parcela SHM - 01, de Intipunku. 75
Figura 23. Familias con mayor número de individuos en la parcela SHM - 02, de Kantupata. 75
Figura 24. Representación gráfica de las especies ecológicamente más importantes en la
parcela SHM – 01 del bosque de Intipunku. ..................................................................... 77
Figura 25. Representación gráfica de las Familias más importantes ecológicamente en el
Bosque de Intipunku. ........................................................................................................ 78
Figura 26. Representación gráfica de las especies más importantes ecológicamente en la
parcela SHM – 02 del Bosque de Kantupata. .................................................................... 79
Figura 27. Representación gráfica de las Familias ecológicamente más importantes en el
Bosque de Kantupata. ....................................................................................................... 80
Figura 28. Representación gráfica de los índices de diversidad en cada una de las parcelas de
Wiñaywayna ...................................................................................................................... 81
Figura 29. Curvas de rarefacción de riqueza de especies en ambas parcelas evaluadas. ....... 82

ÍNDICE DE MAPAS
Mapa 1: Ubicación del área de estudio. ............................................................................... - 26 -
Mapa 2: Santuario Histórico de Machupicchu ..................................................................... - 27 -
Mapa 4: Fisiografía del área de estudio ............................................................................... - 30 -
Mapa 5: Red hidrográfica del Santuario Histórico de Machupicchu ................................... - 32 -
Mapa 6: Zonas de vida del área de estudio ......................................................................... - 37 -
Mapa 7: Sistemas Ecológicos del aérea de estudio ............................................................. - 40 -
Mapa 3: Ubicación de las parcelas permanentes de muestreo ........................................... - 45 -
ÍNDICE DE ANEXOS
1. Documentos de autorización del proyecto de Investigación e ingreso al SHM. ............ 105
2. Ubicación de parcelas permanentes del Santuario Histórico de Machupicchu. ............ 106
3. Protocolo de RAINFOR para la remedición y establecimiento de parcelas. ................... 107
4. Ficha de registro de datos en campo .............................................................................. 109
5. Fichas RAINFOR de códigos de trabajo de campo y de base de datos. .......................... 110
6. Procedimiento para el calculo de Biomasa aérea .......................................................... 111
7. Resumen de tasa de Mortalidad y Reclutamiento ......................................................... 113
8. Especies reclutas en cada una de las parcelas ................................................................ 113
9. Tipos de mortalidad de individuos muertos en cada una de las parcelas ...................... 114
10. Biomasa aérea y carbono almacenado en cada una de las parcelas evaluadas. ........... 115
11. Biomasa y carbono almacenado en cada censo. ............................................................ 116
12. Pruebas de Normalidad con SPSS Statistics 23.0 ............................................................ 116
13. Análisis estadístico de sisgnificancia de biomasa aérea en cada una de las parcelas
con SPSS Statistics 23.0. .................................................................................................. 118
14. Biomasa aérea por especies en cada una de las parcelas. ............................................. 118
15. Biomasa aérea almacenada de muertos y reclutas. ....................................................... 120
16. Dinámica de biomasa aérea. ........................................................................................... 121
17. Productividad primaria. .................................................................................................. 121
18. Composición arbórea de la parcela SHM - 01 (Intipunku) censo 2016. ......................... 121
19. Composición arbórea de la parcela SHM - 02 (Kantupata) censo 2016. ........................ 123
20. Índices de Valor de Importancia en la parcela SHM – 01 ............................................... 124
21. Índices de Valor de Importancia en la parcela SHM – 02 ............................................... 127
22. Índices de diversidad alfa (α) en cada parcela evaluada de Wiñaywayna ..................... 130
23. Datos de análisis de los índices de diversidad beta (β) en parcelas evaluadas de
Wiñaywayna. ................................................................................................................... 130
24. Fotografías de especies arbóreas registradas en las parcelas evaluadas de Wiñaywayna.
......................................................................................................................................... 132

I
RESUMEN
En la presente investigación “DINAMICA, BIOMASA AEREA Y ANALISIS POBLACIONAL DE
DOS PARCELAS PERMANENTES A TRAVES DE UNA GRADIENTE ALTITUDINAL EN
BOSQUES MONTANOS DE WIÑAYWAYNA - SANTUARIO HISTÓRICO DE MACHUPICCHU -
CUSCO” se evaluó la dinámica poblacional, biomasa aérea-carbono almacenado y las
características ecologicas de dos parcelas permanentes en los sectores de Intipunku a
los 2653 m de altitud (SHM – 01) y Kantupata a los 3200 m de altitud (SHM – 02) en
Wiñaywayna dentro del Santuario Histórico de Machupichu, desde setiembre del 2013
a noviembre del 2016, bajo la metodología estandarizada de RAINFOR.
La parcela SHM – 01, presentó 1.18 % de tasa de mortalidad, 0.79 % de tasa de
reclutamiento y 0.99 % de tasa de renovación. La parcela SHM – 02, presentó 1.21 % de
tasa de mortalidad, 2.05 % de tasa de reclutamiento y 1.63 % de tasa de renovación, y
fue la parcela más dinámica con mayor actividad.
La biomasa aérea y carbono almacenado fue mayor en la parcela SHM – 01 de 145 ±
2.80 T ha-1 y 72.99 ± 1.40 T C ha-1 con una productividad de 0.95 T ha-1 año-1 y menor en
la parcela SHM – 02 de 101.06 ± 3.46 Tha-1 y 50.53 ± 1.73 TCha-1 con una productividad
de 1.18 T ha-1 año-1.
Se registró en la parcela SHM – 01 de 1.0 ha, 26 familias, 36 géneros y 45 especies de
árboles en 682 individuos, las especies con mayor IVI fueron Meliosma peytonii con
33.50 %, Hieronyma oblonga con 29.23 % y Gordonia fruticosa con 28.43 %. En la parcela
SHM – 02 de 1.0 ha, se registró a 22 familias, 28 géneros y 31 especies de árboles en 604
individuos, las especies con mayor IVI fueron Weinmannia crassifolia con 70.82 %, Ilex
andicola con 35.65 % y Hedyosmum angustifolium con 35.09 %, las mismas especies
fueron las mas abundantes en cada parcela evaluda. Se registró una alta diversidad de
3.063 y 2.477 según Shanon, 0.93 y 0.87 según Simpson para Intipunku y Kantupata
respectivamente con una muy baja similaridad entre ellas por compartir solo 7 especies
los que se traduce en una composición de especies arbóreas diferente en cada parcela.

II
INTRODUCCIÓN
Los bosques montanos son ecosistemas ubicados por encima de los 1000 m y se
extiende por la ceja de montaña de la vertiente oriental de los andes, pasando a la
vertiente occidental por el norte peruano (Weberbauer 1945). Estas áreas son
ecosistemas frágiles, pues contienen una diversidad biológica caracterizada por su alto
grado de singularidad y rareza (Cuesta et al. 2009), por el endemismo de su diversidad
biológica (Gentry, 1992) y por ser fundamentales en la provisión de servicios
ecosistémicos, principalmente vinculados al agua, a la regulación climática regional y a
la captura y almacenamiento de carbono (Cuesta et al. 2009), sin embargo, su papel en
el ciclo y almacenamiento de carbono es aún poco conocido (Girardin et al., 2013;
Spracklen & Righelato, 2013), pese a que son importantes sumideros. Estos ecosistemas
hoy en día están siendo vulnerados por la degradación antrópica, hechos que los lleva a
ser prioritarios en la conservación.
El Santuario Histórico de Machupicchu es una de las áreas naturales protegidas de
mayor importancia para el estado peruano y reconocida por la UNESCO, como
patrimonio de la humanidad y representa un gran potencial biótico y diversidad de
hábitats, sin embargo, el poco conocimiento sobre la dinámica poblacional y
almacenamiento de biomasa-carbono de sus bosques montanos, generan vacios de
información, por lo que se evaluó en la presente investigación, la dinámica poblacional,
biomasa aérea-carbono almacenado en dos parcelas permanentes pre establecidas en
los bosques montanos de los sectores de Intipunku a los 2653 m (SHM – 01) y Kantupata
a los 3200 m (SHM – 02) en Wiñaywayna dentro del Santuario Histórico de Machupichu,
tomando datos desde el 2013 al 2016, bajo la metodología estandarizada de parcelas
permanentes de RAINFOR.

III
OBJETIVOS
GENERAL
Evaluar la dinámica y biomasa aérea en dos parcelas permanentes de los bosques
montanos de Intipunku y Kantupata en Wiñaywayna dentro del Santuario Histórico de
Machupicchu y su variación a través de una gradiente altitudinal.
ESPECÍFICOS
1. Determinar las tasas de mortalidad, reclutamiento y renovación en cada una de las
parcelas permanentes de los bosques de Intipunku y Kantupata.
2. Cuantificar la biomasa aérea y carbono almacenado, en cada una de las parcelas
permanentes de los bosques de Intipunku y Kantupata.
3. Evaluar la composición florística, Indice de Valor de Importancia y diversidad en
cada una de las parcelas permanentes de los bosques de Intipunku y Kantupata.

IV
HIPÓTESIS
La biomasa y el carbono almacenado por los árboles en los bosques montanos de
Intipunku y Kantupata en Wiñaywayna, esta influenciada por la dinámica poblacional,
altitud y composición florística.

V
JUSTIFICACIÓN
Los bosques montanos de los Andes tienen una importancia global por ser reservorios
de biodiversidad, por sus altos contenidos de carbono, y por sus excepcionales funciones
de regulación hídrica y mantenimiento de una alta calidad del agua (Bubb et al. 2004).
Estos bosques son uno de los ecosistemas menos estudiados y más amenazados de los
Trópicos (Foster 2001, Sala et al. 2005). Por ello realizar estudios de monitoreo a largo
plazo en bosques montanos son una tarea excepcional y preponderante.
En el Santuario Histórico de Machupicchu se alberga un área muy relevante de bosques
montanos, donde se tienen 2 parcelas permanentes instalados por RAINFOR dentro de
los bosques de Wiñaywayna, las cuales fueron monitoreados en el presente estudio
generando información sobre la dinámica poblacional, almacenamiento de carbono en
la biomasa aérea, debido a la productividad, mortalidad y reclutamiento que ocurre
como actividad eco fisiológica del bosque.
Los datos obtenidos en el presente estudio podrán servir de base para la estimación de
biomasa y carbono en todo el Santuario Histórico de Machupicchu, y proveer de esta
manera a las autoridades competentes, las herramientas necesarias para poner en
marcha proyectos de conservación y manejo de bosques así mismo proyectos que
involucre el pago por servicios ambientales o eco sistémicos por la captura de carbono.

1
CAPITULO I
MARCO TEORICO
1.1 ANTECEDENTES
1.1.1 Internacionales:
Ramirez et al. (2012), evaluaron la mortalidad y el reclutamiento de árboles con DAP =
10 cm, en un bosque húmedo montano bajo de la Cordillera de los Andes, Venezuela,
entre 2200 y 2500 m de altitud, mediante 23 mediciones sucesivas (1968 – 2000), de
seis parcelas permanentes de 0,25 ha. Se registraron 55 especies arbóreas incluyendo
palmas, con 622 individuos ha-1 y un área basal promedio de 46 m2 ha-1. Entre las
especies más abundantes se encuentran Myrcia fallax, Graffenrieda latifolia y
Beilschmeidia sulcata, con más de 100 individuos por ha. Las tasas anuales de
reclutamiento y mortalidad de los árboles fueron de 1,64 y 1,59 %, respectivamente,
con diferencias estadísticas entre parcelas solo para el reclutamiento.
Linares (2008), realizó un análisis florístico y estructural de la vegetación en gradiente
altitudinal desde los 2250 m hasta los 3065 m de altitd, en una selva nublada en Mucuy,
del estado de Mérida, Venezuela, en 15 unidades de muestreo de 500m2 (20 x 25m), la
vegetación fue analizada en dos estrados estructurales, los individuos con DAP ≥ 2.5cm
e individuos con DAP < 2.5cm; se reportó un total de 110 especies distribuidas en 45
familias, siendo las más representadas las familias Lauraceae, Melastomataceae,
Rubiaceae, Euphorbiaceae, Myrtaceae, Arecaceae, Araliaceae, Ericaceae y entre los
géneros más importantes Miconia, Ocotea, Nectandra, Myrcia, Myrcianthes, Cinchona,
Guettarda, etc.
Phillips et al. (2011), evaluaron la Biomasa aérea de tres bosques montanos de
Colombia, para el Bosque Húmedo montano reporta un total de Biomasa aérea de 145,5
±17,3 T ha-1; para un Bosque muy Húmedo montano reporta un total de biomasa aérea
de 125,5 ±22 T ha-1 y para un Bosque pluvial montano reporta 106,4 ±7,5 T ha-1 de
biomasa aérea.

2
Bobórquez et al. (2011), evaluaron 0.1 – ha de la selva altoandina de la cordillera central
Colombia a 3100 m de altitud, donde registraron todos los individuos con DAP ≥ 1,5 cm,
y registraron a 42 especies y morfoespecies, distribuidos en 40 géneros y 34 familias en
173 individuos, de los cuales las especies Weinmannia pubescens y Hedyosmum
bonplandianum fueron las especies con mayor importancia ecológica (40,9 % y 26,7 %
respectivamente), y las familias con mayor IVF fueron Rubiaceae 40,4 %, Cunoniaceae
37,2 %, Podocarpaceae 27,5 % y Melastomataceae 23,1 %.
Alvares et al. (2012), Proponen modelos alométricos que usa parámetros como DAP
(diámetro a la altura del pecho), altura y densidad (gravedad especifica de madera) de
los árboles, para realizar estimaciones de biomasa aérea en bosques de alta montaña
(Bosques montanos), de esta manera construyen modelos alométricos que presentan
menos sesgos en la evaluación.
Velásquez et al. (2012), evaluaron los patrones de diversidad, dinámica de biomasa
aérea (AGB) y las tasas de mortalidad y reclutamiento mediante tres censos en un
periodo de 9 años, en dos parcelas permanentes ubicadas en bosques andinos de la
Cordillera Occidental, Colombia, donde se encontraron 1664 individuos con DAP ≥ 10
cm, pertenecientes a 222 especies en 113 géneros y 60 familias, la riqueza fue de 156
especies/ha, las tasas de mortalidad y reclutamiento fue de 0.88% y 1.16%
respectivamente, lo que no permitió establecer ningún efecto externo del
calentamiento global o el cambio climático en la dinámica de los bosques individuales.
La biomasa aérea promedio fue de 243.44 ± 9.82 T/ha con un incremento promedio
anual de 2.9 t/ha.
Álvarez et al. (2013), evaluaron el contenido de carbono en los almacenes aéreos y
subterráneos (Carbono en la biomasa arbórea, estrato herbáceo y arbustivo, material
leñoso muerto, mantillo, raíces finas y suelo mineral) de cinco comunidades de bosque
montano de niebla en la Sierra Norte de Oaxaca, situadas entre 1500 y 2500 m de altitud,
determino en promedio, la reserva de carbono total en biomasa y suelo para los bosques
montanos de niebla fueron de 384.16 ± 40.13 Mg Cha-1, las reservas de carbono en la
biomasa viva (componente arbóreo, arbustivo y herbáceo) fue de 138.3 ± 38.03 MgCha-
1.

3
García (2014), evaluó la Composición y estructura florística del bosque de neblina
montano, del sector “san Antonio de la montaña”, cantón baños, provincia de
Tungurahua, en 5 transectos de 200 m2 cada uno y distribuidos en un gradiente
altitudinal desde los 2500 m hasta los 2700 m de altitud, donde se tomaron datos y se
colectaron individuos con DAP ≥ 10, registrándose un total de 190 individuos
distribuidos en 23 familias y 28 especies, de los cuales Cronton magdalenensis fue la
especie más significativa con 17.69 % y la familia Melastomataceae obtuvo el mayor
peso ecológico con 43.07 %, la especies Oreopanax ecuadorensis fue la más abundante
con 17 individuos y la familia Melastomataceae la más cuantiosa con 44 individuos.
Según los índices de diversidad de Simpson y Shannon – Wiener, el área de estudio tuvo
una alta diversidad por presentar valores de 0.96 y 3.34 según los índices mencionado
respectivamente.
1.1.2 Nacionales:
Palomino (1999), realizó un estudio de diversidad arbórea en el bosque nublado de San
Pedro entre los 1310 m hasta los 2000 m de altitud, ubicados en el Parque Nacional del
Manú. Mediante un muestreo de 10 subparcelas de 25 m x 40 m, en un total de 1 ha,
donde registró 45 familias con 91 géneros y 192 especies distribuidos en 618 individuos
de árboles con DAP ≥10cm. De acuerdo a la composición florística, la familia con mayor
número de especies fue Moraceae con 21 especies, seguidas de Euphorbiaceae con 20
especies, Melastomataceae con 19 especies, Lauraceae con 18 especies, Fabaceae con
15 especies y Rubiaceae con 10 especies, y las familias con mayor valor de importancia
son Moraceae con el 21.94 %, seguida de las familias Euphorbiaceae con el 17.7 % y
Melastomataceae con el 8. 10 %. Las especies con mayor valor de importancia son:
Hiieronima alchomeoides var alchomeoides con el 8%, seguida en importancia por Alza
tea verticillata con el 4.84 % y Tapirira sp con el 4.41 %. Concluye que el bosque nublado
de San Pedro presenta una alta diversidad de especies arbóreas de acuerdo al índice de
diversidad de Simpson.
Quinto e Ibarguen (2009), evaluaron las tasas de mortalidad y reclutamiento de árboles
en una parcela permanente de investigación de un bosque pluvial tropical en dos
mediciones 1998 y 2005. La tasa media anual de mortalidad de árboles fue 1.39 % y el

4
coeficiente de mortalidad exponencial fue 1.41 %; los tipos de mortalidad más comunes
fueron: volcamiento de raíz y muerte en pie. La tasa de reclutamiento anual fue 1.2 % y
el coeficiente de reclutamiento exponencial registró 1.19 %. La similitud en el número
de individuos entre mediciones, así como en los demás parámetros evaluados, sugieren
un posible equilibrio entre mortalidad y reclutamiento del bosque.
Ureta (2009), evaluó las diferencias altitudinales significativas en contenido de carbono
y biomasa arbórea en un bosque montano de la Quebrada de Yanachaga y un bosque
amazónico de la Estación biológica de Paujil dentro del Parque Nacional Yanachaga
Chemillen (PNYCH), en parcelas permanentes de 1ha del Jardín Botánico de Missouri
donde toma datos de árboles con DAP ≥ 10cm, para posteriormente determinar el
contenido de carbono y biomasa arbórea, mediante ecuaciones alométricas. Obtuvo
64.66 TCha-1 de carbono almacenado para el bosque montano de la Quebrada de
Yanachaga a los 3110 m de altitud y 110.12 TCha-1 para el bosque amazónico de la
Estación Biológica de Paujil a los 410 m de altitud.
Bertzky et al. (2010), Indican que los pocos estudios realizados en bosques montanos,
estiman que estos ecosistemas son reservorios importantes de carbono, pues contienen
entre 40 a 80 T ha-1 de biomasa aérea y entre 130 a 240 T ha-1 de biomasa en el suelo, a
pesar de que el conocimiento de la dinámica de carbono en dichos ecosistemas alto
andinos es aún limitado (Gibbon et al. 2010).
Gibbon et al. (2010), realizaron un estudio de biomasa aérea en parcelas permanentes
de los bosques nublados y la puna de la Cordillera Oriental peruana (Parque Nacional
del Manu), reporta que los bosques cerca del límite arbóreo superior, tienen un
contenido promedio de 63.4 ± 5.2 MgCha-1 contenidas en la biomasa sobre el suelo
(árboles, arbustos, hierbas, bambú), el componente arbóreo tiene 53.5 ± 5.7 MgCha-1 y
un total adicional de 13.9 ± 2.8 MgCha-1 en las grandes raíces (raíces y carbono del
suelo).
Girardin et al. (2010), evaluaron la productividad primaria neta y el ciclo del Carbono
(C) a lo largo de un transecto que se extiende desde las tierras bajas de la Amazonía
hasta los altos Andes del Perú (Parque Nacional de Manu). Los valores de biomasa sobre
el suelo se redujeron de 123.50 MgCha a 194 m hasta 47.03 MgCha a 3020 m de altitud,

5
a los 2720m de altitud la biomasa sobre el suelo fue de 65.90 MgCha y a los 3025m de
altitud fue de 64.90 MgCha. La productividad media anual sobre el suelo varía entre 9.50
± 1.08 MgCha a (210 m) y 2.54 ± 0.09 MgCha (2020 m), a 2720m es de 4.03 ± 0.17 MgCha.
Cruzado (2010), cuantificó el carbono almacenado en la biomasa aérea de los bosques
altoandinos de la Concesión para Conservación Alto Huayabamba (CCAH), ubicada en el
distrito de Huicungo, provincia de Mariscal Cáceres, departamento de San Martín,
específicamente en los bosques Altoandinos, entre los 3.000 – 4.600 m de altitud,
mediante cuatro parcelas rectangulares de 0.5 ha, donde se evaluó el componente
arbóreo con DAP≥10cm, sotobosque y herbáceo, se estimó la biomasa mediante
ecuaciones alométricas. Se obtuvo un promedio de biomasa aérea almacenada de
167.11 ± 25.8 Mgha-1 (componente arbóreo, sotobosque, herbáceo, dosel) con 121.18
Mgha-1 de biomasa aérea y 69.77 ± 12.8 MgCha-1 de Carbono para el componente
arbóreo.
García (2011), evaluó la variabilidad de la composición de especies en gradiente
altitudinal desde los 1500m hasta los 3700m de bosques montanos neo tropicales en el
Parque Nacional de Manu a través de 36 parcelas de 50 x 20 para un total de 3.60 – ha,
donde evaluó todos los árboles, helechos y arbustos con DAP > 2.5 a 1.30m de POM y
reportó 9,718 árboles individuales de 540 especies y 86 familias. Entre los 1450 - 3000
m de altitud se evaluaron a 20 parcelas, registrando 5667 Individuos, 435 especies y 81
familias, donde Cyatheaceae y Melastomataceae fueron las familias más abundantes,
además de Rubiaceae, Chloranthaceae y Lauraceae. En transecciones paisajísticas cerca
de la línea de árboles (por encima de 3200 m de altitud) se evaluaron 16 parcelas,
registrando 4051 individuos, 151 especies y 26 familias, siendo Melastomataceae la
familia con más árboles individuales, seguida de Clusiaceae, Cunoniaceae, Myrsinaceae
y Asteraceae la familia más rica en especies. En los tramos de Trocha Unión y San Pedro,
Cyatheaceae fue la familia más común y abundante, en Callanga las familias
Chloranthaceae y Cunoniaceae eran mucho más abundantes. En este estudio la
elevación fue el principal factor que influyó en el desarrollo florístico Composición y
diversidad a través de los gradientes de elevación, incluso a través de pequeñas
elevaciones, y la radiación solar potencial (PSR) tuvo efectos significativos en la riqueza

6
de especies Tanto en los transectos de elevación como en la muestra de paisaje cercana
a la cordillera de los Andes.
Monteagudo et al. (2014), reportan en el primer catálogo de árboles de la Reserva
Comunal El Sira – Perú, desarrollado en cinco parcelas permanentes de 1.0-ha a través
de la gradiente altitudinal desde los 250 a 2230 m, a 527 especies de plantas leñosas
arbóreas, distriobuidas en 211 generos y 76 familias, de las cuales 249 (47.25%) están
identificadas hasta el nivel de especie, 52 (9.87%) definidas como vel sp. aff. (Especie a
fin) o cf. (confrontar con) y 226 (42.88%) solamente identificadas a nivel de género; lo
que nos demuestra que el conocimiento de los árboles a nivel específico es menos del
50%, y concluye que en los bosques Pre Montano y Montano existen una compleja
composición florística y un potencial de especies nuevas y endémicas para el área.
Nina (2014), evaluó la composición, estructura y regeneración arbórea del Bosque
Nublado del Parque Nacional del Manu, en dos parcelas de monitoreo permanente
ubicadas en el sector conocido como Trocha Unión en el distrito de Q’osñipata a 3 450
y 2 750 m de altitud y en parcelas de 1.0 - ha, donde Reporta a los 3450m, 43 especies
arbóreas correspondientes a 22 géneros y 20 familias; y a los 2750m, 85 especies
arbóreas distribuidas en 34 géneros y 26 familias, siendo Weinmannia cochensis con
69.99 %, Symplocos quitensis con 36.09 %, Miconia cf. denticulata con 34.36 % a los
3450m y Cyathea delgadii con 37.13 %, Weinmannia reticulata con 30.42 %, Ocotea sp1
con 18.44 % a los 2750m, las especies más importantes.
Pallqui (2014), en su tesis de investigación realizado en la llanura Amazónica de la
Reserva Nacional de Tambopata – Madre de Dios; evaluó la Dinámica de la Biomasa
aérea y Composición florística de 9 parcelas permanentes, llegó a la obtener una
biomasa aérea almacenada de 296.2 ± 33.9 T ha-1 en promedio, una tasa de mortalidad
y reclutamiento promedio de individuos de 2.12 ± 0.52% y 1.92 ± 0.49%,
respectivamente. La tasa de reemplazo es 2.02% anual. La dinámica de la biomasa
muestra una ganancia neta total de 1.96, 1.69 y –1.23 T ha-1 para cada periodo (2003-
2006, 2006-2008, 2008-2011) respectivamente. Antes de la sequía de 2010 se encontró
un cambio en la biomasa de 1.88 T ha-1 año-1 y post sequía fue de –0.18 T ha-1 año-1 en
promedio. Se registraron 64 familias, 219 géneros y 531 especies, la Composición
florística fue muy similar en todas las parcelas.

7
Bellota (2015), en su tesis de investigación realizado en los bosques andinos de la
Reserva Comunal de “El Sira”, evaluó la variación de la diversidad, biomasa aérea y
dinámica poblacional en 4 parcelas permanentes a través de la gradiente altitudinal,
donde registró 267 especies y 162 géneros distribuidas en 67 familias en total, la
diversidad fue alta en el bosque de transición a los 845m, (4.37 y 0.98) Shannon y Simpson
respectivamente y disminuye conforme aumenta la altitud, la biomasa aérea estimada
en promedio fue mayor en el bosque de transición (845m) y en el bosque montano
(1391m) con 255 T ha-1 y 241.27 T ha-1 respectivamente siendo menor en las partes más
altas a 2230 m sobre un bosque esclerófilo, con sólo 18.73 T ha-1, la dinámica poblacional
de la Reserva Comunal El Sira se expresó con una baja mortalidad y una alta capacidad
de reclutamiento.
1.1.3 Locales:
Vargas (1962), menciona que el Santuario Histórico de Machupicchu comprende una
típica formación vegetal de transición llamada "Ceja de Selva", característica del Perú
donde se encuentra toda una variedad vegetal, como criptógamas y fanerógamas. Las
laderas están cubiertas de bosques, arbustos y matorrales. Además, sus publicaciones
están referidas a la flora ornamental de Machupicchu.
Tupayachi y Galiano (1986), reportaron un estudio preliminar de la flora para el
Santuario Histórico de Machupicchu, mediante la revisión de trabajos realizados por
diferentes autores; donde determinaron 97 familias de dicotiledóneas, 20 familias de
Monocotiledóneas, 02 de Gimnospermas y 08 de Pteridophytos.
Nuñez (1992), realizó un listado de la Flora y Fitogeografía del Santuario Histórico de
Machupicchu, basado en colecciones e intenso trabajo de campo, revisión de
especímenes botánicos, plantas monografiadas, revisión de literatura, visita a herbarios,
consulta a botánicos especialistas, etc. Reporta las siguientes categorías taxonómicas
para el 90 % del total del área del Santuario: Gimnospermas 6, Helechos 264,
Monocotiledóneas 500, Dicotiledóneas 1800, Criptógamas 200, haciendo un total de
2760 especies entre árboles, arbustos y herbáceas.
Dueñas (1992), realizó un estudio en el sector de Wiñaywayna del Santuario Histórico
de Machupicchu, y registro que la diversidad de árboles es alta para 0.1ha, con 30

8
especies distribuidas en 25 familias para 10 parcelas evaluadas, donde se registraron
arboles de DAP ≥ 5cm, siendo las familias Cunoniaceae con 34.37 %, Clethraceae con
11.45 %, Betulaceae con 5.7 %, Ericaceae y Piperaceae con 4.68 % Myrtaceae 4.12 %
Clusiaceae y Proteaceae con 2.08 %, mejor representadas y corresponden a un 82.22 %
de la diversidad arbórea, el 17.78 % lo conforman otras familias, en cuanto se refiere al
número de especies las más importantes fueron: Melastomatáceas con 3 especies,
Myrtáceas y Ericáceas con 2 especies y las otras 22 familias tuvieron una sola especie
cada una.
Galiano y Núñez (1992), en un estudio comparativo, haciendo uso de revisiones de
colecciones botánicas e intenso trabajo de campo durante 10 años, reportan para el
Santuario Histórico de Machupicchu 280 especies en Pteridophytos, 190 especies en
Orchidaceae, 150 especies de árboles, 80 especies en Poaceae, 13 especies en Moraceae
y 20 especies en Ericaceae, haciendo un total de 2670 spp.
Chávez (1993), realizó un estudio de Helechos arborescentes dentro del Santuario
Histórico de Machupicchu, donde se encontró 4 especies: Cyathea delgadii, Cyathea
caracasana, Trichipteris sp. y Nephelea incana, y fuera de los límites del Santuario se
encontraron solo 3 especies, Cyathea caracasana, Trichipteris sp., y Nephelea incana,
siendo Cyathea caracasama la especie más abundante dentro y fuera del Santuario, y
estando mejor conservada fuera de los límites del Santuario.
Palomino (1996), determinó el área mínima para los arboles de Intipunku dentro del
Santuario Histórico de Machupicchu de 2048 m2, por el método de puntos anidados,
donde reporta un total de 24 Familias, 29 Géneros y 38 especies, para un total de 259
individuos de árboles con ≥10 de DAP. Las Familias con mayor importancia son las
Myrsinaceae y Melastomataceae seguido de Euphorbiaceae y Myrtaceae; y la especie
más importante fue Meliosma peytonii con 17.38%.
Monteagudo (1997), evaluó 0.1-ha de tres bosques del Santuario Histórico de
Machupicchu, bosques de Intipunku, Puente Ruinas y margen derecha del rio Aobamba,
divididas en 10 transectos, donde determino la composición florística arbórea,
diversidad, similitud y variables poblacionales. Determinó 96 especies distribuidas en 36
familias y 60 géneros con un total de 483 individuos arbóreos ≥10 de DAP, los tres

9
bosques obtuvieron una alta diversidad (H’) de 5.92 con 96 especies, siendo el Bosque
de Rio Aobamba el más diverso, seguido de Intipunku y Puente Ruinas.
Galiano y Tupayachi (2002), realizaron un listado de especies de flora para cada
localidad del Santuario Histórico de Machupicchu, en áreas que tienen vacío de
información, mediante la instalación de 6 parcelas de 0.10-ha en los sectores de
Cedrobamba, Aguas Calientes, Mandorpampa, Puente Ruinas, Quebrada Wayruro,
Microcuenca de Aobamba, reportándose 24 especies no vasculares, 23 Pteridophytas, 2
Gimnospermas, 15 Liliopsidas, 112 Magnoliopsidas con un total de 1216 especímenes
de los cuales 441 especies fueron nuevos registros para el Santuario Histórico de
Machupicchu y 4 especies nuevas para la ciencia, en el tramo Wiñaywayna-Intipunku
se registraron 8 familias con 12 especies de árboles.
Galiano y Núñez (2011), estimaron la flora arbórea y leñosa del Santuario Histórico de
Machupicchu, en el área núcleo y la zona de amortiguamiento donde reportaron más
de 550 especies de árboles distribuidas en 74 familias y 192 géneros, de las cuales las
familias con más géneros y especies son: Rubiaceae, Solanaceae, Asteraceae,
Melastomataceae, Rosaceae y entre otros. La escasez en investigar a largo plazo afecta
el mal uso y peligro de no conocer bien la flora arbórea que se aplica a la conservación
biológica del país.
Rojas y Peláez (2013), en su tesis de investigacion realizado en la cuenca de Torontoy,
Santuario Histórico de Machupicchu, registraron 778 individuos distribuidos en 33
familias, 49 géneros, 58 especies nativas y 1 especie introducida de árboles mayores a
10 cm de DAP, en 10 parcelas de 0.1 – ha, que hacen 1.0 – ha en total, donde el índice
de diversidad Alpha Fisher es de 14.77, calificándose como de alta diversidad y se
reporta por primera vez 07 especies de árboles para el Santuario: Ageratina gloeoclada
(B.L Rob) R.M King & H. Rob., Critoniopsis sp., Miconia cf. demissifolia Wurdack., Miconia
caerulea (Pav. Ex D. Don) Naudin., Meriania aff. weberbaueri J.F. Macbr., Meriania cf.
hemandii L. Uribe, Iocroma sp.
Valenzuela et al. (2014), tras varias colecciones y revisiones en el Herbario Vargas (CUZ)
de la Universidad Nacional San Antonio Abad del Cusco, de especies arbóreas del
Santuario Histórico de Machupicchu, publicaron 332 especies de árboles, agrupadas en

10
69 familias de angiospermas, 1 en gimnospermas y 1 de pteridofitas, que son registradas
en el libro “Arboles de Machupicchu Cusco – Perú”, de las cuales las familias con mayor
número de especies son: Lauraceae con 18 especies, Melastomataceae con 17,
Solanaceae con 16, Moraceae y Rosaceae con 15 especies cada una, Cunoniaceae y
Asteraceae con 12 especies cada una y las familias Euphorbiaceae, Myrtaceae,
Rubiaceae, Symplocaceae con 11 especies. Ademas se presentan la descripsion de las
100 especies más representativas, siendo algunas de ellas, Nectandra reticulata (Ruiz &
Pav.), Meriania tomentosa (Cogn.) Wurdack, Cedrela angustifolia Sesse & Moc. Ex DC.,
Ficus cuatrecasana Dugand, Morus insignis Bureau, Prumnopitys montana (Humb. &
Bonpl. Ex Willd.), Meliosma peytonii A. H. Gentry, Cinchona bubescens Vahl y entre
muchas otras.

11
1.2 MARCO CONCEPTUAL
1.2.1 Bosque montano
También conocido como la Yunga, Selva Alta o Yunca («valle cálido» en el lenguaje
antiguo de los Andes peruanos), cubierta por selvas y neblinas de montaña, localizado
en la vertiente oriental de los Andes, y es denominado como «Bosques montanos de la
vertiente oriental de los Andes peruanos» (Dinerstein et al. 1995), se extiende a lo largo
de un eje noreste–suroeste, en el rango altitudinal en que la humedad del aire se
condensa y forma nubes, entre los 800 – 1000 y los 3500 – 3600 m sobre el mar, rango
sugerido por gran parte de los que han estudiado las Yungas o los bosques montanos
(Tovar et al. 2010).
Se caracteriza por su vegetación densa y una estructura vertical organizada en múltiples
estratos, que se desarrolla sobre terrenos que presentan fisiografía en extremo
accidentada (por grandes pendientes) y clima variable particularmente húmedo a muy
húmedo, con promedios de temperatura anual que tienden a ser menores en el sur que
en el norte, debido a la influencia de los friajes o surazos (ONERN 1976).
Estos ecosistemas son fundamentales en la provisión de servicios ecosistémicos
principalmente vinculados al agua, a la regulación climática regional y a la captura y
almacenamiento de carbono. En los países andinos han sido catalogados como
ecosistemas frágiles y estratégicos por estar situados en zonas de recarga de cuencas
hidrográficas que proveen agua de la que se benefician más de 40 millones de personas
en Bolivia, Ecuador y Perú. Esto los hace críticos en la adaptación al cambio climático y
en el soporte de los sistemas productivos de los Andes (Cuesta et al. 2009).
1.2.2 Gradientes ambientales
Los gradientes ambientales resultan comúnmente en el cambio gradual de una
comunidad biológica a otra, en las grandes montañas tropicales los cambios altitudinales
en las condiciones ambientales en un rango geográfico corto contribuyen a los cambios
en la estructura del bosque y la composición de las especies, los cambios de
temperatura afectan a la evaporación, a las tasas de crecimiento, a las tasas de
descomposición y una serie de otras funciones ambientales y biológicas (Grubb y
Whitmore, 1966).

12
Los cambios a través de las gradientes altitudinales presentan variaciones en las
distribuciones y/o atributos de muchas variables bióticas como la composición, riqueza
de plantas vasculares (Grubb et al. 1963, Gentry 1988, Kessler 2001, Martin et al. 2007),
en estructura forestal y productividad (Phillips et al. 1994, Weaver 2010), y en la
asignación de carbono (Leetchner et al. 2007, Gibbon et al. 2010). Ademas presentan
patrones excepcionales en el recambio de especies y comunidades debido, en parte, a
la enorme heterogeneidad de hábitats producto de los fuertes gradientes ambientales
(Jorgensen y León-Yánez 1999, Kessler 2002, Kessler et al. 2001). La diversidad de estos
bosques disminuye al incrementarse la elevación por encima de los 1.500 m. Debajo de
este límite, los bosques montanos son tan diversos como los de tierras bajas y presentan
patrones de composición florística similares a éstos (Gentry 1995).
1.2.3 Ciclo de carbono en un bosque
El ciclo del carbono está determinado por procesos de transferencia y almacenamiento
de moléculas de carbono entre los componentes de un ecosistema boscoso. Para poder
entender ello es necesario hacer una clara diferencia entre un Stock de carbono y un
Flujo de carbono.
Se refiere como Stock a todo aquel carbono que se encuentra almacenado en los
diferentes componentes del ecosistema como:
Biomasa viva almacenada en los árboles, arbustos, hierbas y raíces
Necro masa almacenada en la hojarasca y madera muerta
El carbón almacenado en la materia orgánica del suelo
Se refiere flujo de carbono a todos los procesos que afectan el stock como:
Fotosíntesis de hojas y tallos verdes.
Respiración autotrófica en (árboles, arbustos y hierbas) y heterotrófica en
(hojarasca, madera muerta y suelo)
Mortalidad de troncos, ramas, hojas y raíces y reclutamiento de nuevos
individuos.
Descomposición de la madera y la hojarasca por microorganismos.

13
Un incremento neto en la biomasa gracias a los procesos de fotosíntesis excepto la
respiración, se denomina productividad primaria neta (PPN) y se cuantifica al medir el
crecimiento del fuste y la producción de ramas, hojas y raíces.
Al entender la diferencia entre un stock y un flujo de carbono, se puede considerar que
un área determinada de un bosque funciona como un sumidero de carbono, y si la
cantidad almacenada de carbono aumenta con el tiempo, el cambio del stock de
carbono es positivo. (Honorio & Baker, 2010).
Figura 1. Diagrama de los Stocks y flujos de carbono en un bosque
Fuente:(Stock y Flujos de carbono) Modificado de Honorio & Baker, 2010.; (Imagen) Avella,2016.
Las unidades de medida utilizadas en los stoks de carbono, se expresa en términos de
peso de carbono por unidades de área (Mg C ha-1) Megagramos de Carbono por hectárea
y los valores de flujos siempre incluyen la variable tiempo (Mg C ha-1 año-1) Megagramos
de Carbono por hectárea por año, además es importante entender la relación entre las
diferentes unidades usadas en los resultados de los estudios del ciclo del carbono, tales
como:
Suelo
Raíces
Hojarasca
Madera muerta
Fustes y ramas
Hojas
CH4
CO2
CO2
CO2
CO2
= Fotosíntesis - Respiración PPN
Mortalidad Respiración Fotosíntesis Descomposición
STOcK
FLUJOS
CO2
CO2

14
1 Megagramo (Mg) = 1,000 Kilogramos (Kg) = 1 Tonelada (T)
1 Petagramos (Pg) = 1, 000, 000,000 Mg = 1 Gigatonelada (Gt)
1 Hectárea (ha) = 10,000 m2
1.2.4 Reservorios de carbono en un bosque montano
Uno de los ecosistemas menos estudiados en los trópicos en cuanto a sus reservorios y
flujos de carbono son los bosques montanos andinos, a pesar de representar más del 40
% de la vegetación remanente de los andes tropicales (Josse et al. 2009). Los datos
respecto a los contenidos de carbono para estos bosques provienen de regiones muy
puntuales de los Andes en Perú y Ecuador (Tabla 1).
Tabla 1. Datos disponibles sobre contenidos biomasa aérea, subterránea y suelo en ecosistemas de bosques en la región andina.
Fuente Tipo País
Biomasa total
Biomasa aérea
Biomasa subterránea
(Mg C ha-1)
Moser et al. (2011)
Bosques nublados (1.050 m)
Ecuador 154 138,6 15,4
Bosques nublados (2.380 m)
Ecuador 67 48 19
Bosques nublados (3.060 m)
Ecuador 87 53.9 33,1
Gibbon et al. (2010)
Bosques nublados Perú (PN Manu) 77,3 63,4 ±5,2 13,9 ±2,8
Pastizales de puna Perú (PN Manu) <7,5 7,5 ±0,7 -
Zimmermann et al. (2010)
Bosques nublados Perú (PN Manu) 118 Mg ha-1 Biomasa aérea.
Ecotono arbustivo Perú (PN Manu) 147 Mg ha-1
Pastizales de puna Perú (PN Manu) 119 Mg ha-1
Phillips et al. (2011)
Bosque húmedo montano
Colombia 145,5 ±17,3 Mg ha-1
Biomasa aérea.
Bosque muy húmedo montano
Colombia 125,5 ±22 Mg ha-1
Bosque pluvial montano
Colombia 106,4 ±7,5 Mg ha-1
Fuente: Osinaga et al. 2014
Según estos estudios, el único que cuantifica contenidos de carbono por unidad de área
en bosques montanos es el de Moser et al. (2011), que realizo un análisis de variación
de la biomasa aérea y subterránea a lo largo de una gradiente altitudinal de 2.000 m,

15
con intervalos cada 300/400 metros, similar al realizado por Gibbon y colaboradores
(2010). Los valores de carbono más altos se reportaron en los bosques a 1.050 m de
altitud con un promedio de 154 Mg C ha-1, de los cuales el 90 % corresponde a biomasa
aérea y el 10 % al carbono contenido en raíces grandes y finas. Los bosques localizados
a 2380 metros de altitud reportan un total de 67 Mg Cha-1, de los cuales 48 Mg (72 %),
corresponden a la biomasa aérea lo que evidencia también una reducción en la
proporción de BA: BS respecto a los bosques más bajos. Este patrón sugiere que, dentro
de los diferentes compartimentos de la producción neta primaria de un bosque, algunos
de ellos son negativamente controlados por la variación de temperatura y humedad, y
otros son favorecidos conforme la elevación es mayor. Los bosques caracterizados a
3060 m, cerca del ecotono con el páramo, evidencian un repunte en el contenido total
de carbono (87 Mg Cha-1), especialmente por la biomasa subterránea que constituye el
38% del total contenido en estos bosques, esto indica que el carbono contenido en las
raíces de los bosques a esta altura es la fracción con mayor concentración de carbono
por unidad de área respecto a los bosques en gradientes inferiores. (Moser et al. (2011).
1.2.5 RAINFOR
Quiere decir “Red Amazónica de Inventarios Forestales” creada el año 2001, es una red
internacional para la supervisión de la estructura, composición y dinámica de los
bosques amazónicos, utilizando parcelas permanentes a largo plazo para monitorear la
biomasa y la dinámica del bosque y relacionarla con el suelo y el clima, la información
producida, reunida e integrada, puede proveer ideas vitales dentro de los mecanismos
que subtienden a las respuestas actuales de los ecosistemas de la Amazonia frente al
clima y el posible futuro de la Amazonia, bajo el cambio global de los escenarios. (Malhi
et al. 2002).
La importancia de la propuesta de RAINFOR radica en que la compilación y comparación
de cerca de un centenar de estudios de este tipo en una escala regional permite acceder
a un nuevo nivel de información sobre los patrones de composición florística, estructura
y dinámica de los bosques tropicales de la Amazonía y es útil para enriquecer estudios
tendientes a resolver uno de los debates ambientales más trascendentes en la
actualidad en el ámbito mundial, el del bosque tropical y su relación con los cambios
atmosféricos globales (Aguilar et al., 2009).

16
1.2.6 Parcela permanente
Las parcelas permanentes de muestreo son establecidas indefinidamente con el fin de
realizar un monitoreo de procesos dinámicos de manera permanente. La adecuada
demarcación permite la ubicación exacta de sus límites y puntos de referencia a través
del tiempo, así como de cada uno de los individuos que la conforman, donde todos los
arboles existentes han sido identificados, medidos, plaqueados y son evaluados
periódicamente. (Aguilar & Reynel, 2009).
Los estudios demográficos a largo plazo realizados en parcelas permanentes
proporcionan la información para formular sistemas de manejo más eficientes en
bosques naturales. También estas parcelas son muy útiles para la elaboración de
modelos de productividad, de estrategias de manejo sostenible para la conservación de
los bosques tropicales, para la valoración socioeconómica de los recursos procedentes
del bosque, los bienes y servicios que se derivan o pueden derivarse de su utilización,
Londoño & Jiménez, 1999; Philips & Baker, 2002 (citado por Aguilar et al, 2009).
1.2.6.1 Dinámica de bosques montanos
Continuo proceso intrínseco no estático de transferencia de materia y energía, de
renovación y mantenimiento de diversidad de especies en las comunidades de plantas
a través del tiempo que responden a los cambios del medio ambiente e incluyen en los
procesos de sucesión, composición, tolerancia, competición.
1. Reclutamiento
El reclutamiento cuantifica la capacidad que tiene un bosque de incrementar el número
de individuos, árboles en una unidad de área del bosque; es una manifestación de la
fecundidad de las especies y del crecimiento y sobrevivencia de los juveniles, Constituye
uno de los aspectos dinámicos más importantes de una población, Swaine et al. 1987
(citado en Aguilar & Reynel 2009). Para determinar el reclutamiento se considera el
número de árboles que alcanzaron el DAP mínimo de 10 cm entre dos mediciones. Según
Ramírez et al. (2002), el porcentaje de reclutamiento por periodo se calcula por la
diferencia entre el número de árboles reclutados y el número de árboles en el inicio del
periodo.

17
2. Mortalidad
Se considera que un árbol está muerto cuando no tiene hojas vivas ni rebrotes, y se
verifica que el cambium bajo la corteza está seco. Dentro de una parcela permanente,
la mortalidad se refiere al número de árboles que fueron medidos inicialmente y que
murieron durante el período de crecimiento considerado. La mortalidad puede ser
causada por diversos factores, como: edad o senilidad, competición, enfermedades o
plagas, condiciones climáticas adversas, fuego, envenenamiento y corte del árbol
(Sanquetta & Balbinot 2004).
Según Vásquez & Orozco (1992), y Lugo & Scatena 1996, se puede precisar cuatro causas
fundamentales de mortalidad de los árboles:
La primera se debe a procesos endógenos, genéticamente dados, que condicionan
la senescencia.
La segunda se representa por la acción de sustancias toxicas, agentes patógenos,
parásitos o consumidores y puede ser súbita o gradual, y ocurrir local o
masivamente.
La tercera, es ocasionada por cambios en el ambiente que reducen o eliminan una
entrada necesaria de materia o energía como la sequias.
La cuarta causa se representa cuando un bosque es impactado mecánica o
químicamente por una fuerza externa (ej. huracán, incendios, derrame de petróleo,
deslizamientos, etc.). Cada causa tiene diferente periodicidad y opera en escala
espacial distinta.
Una función de la mortalidad es que permite predecir el número de árboles
sobrevivientes después de cierto periodo de tiempo. Los parámetros más usados en
estas funciones han sido la edad y el número de árboles. El modelo más empleado para
determinar la mortalidad es la que determina la mortalidad en términos del número de
árboles sobrevivientes al final de cierto periodo de tiempo, a partir de ecuaciones
diferenciales. (Aguilar & Reynel 2009).

18
a) Tipos de mortalidad de árboles
Muertos quebrados o rotos: Por fuerzas laterales sobre el tronco de un árbol vivo
que no son suficientemente fuertes para dislocar y romper las raíces inferiores,
debido a su débil densidad o a su estructura desequilibrada (Putz 1983), en este caso
el resto del tronco remanente puede permitir que el árbol roto rebrote. La copa
caída de un árbol quebrado, comparado con todo el tronco de un individuo
desenraizado, tiene menor impacto en el bosque.
Muertos en Pie: Un árbol puede “morir en pie” como resultado de senescencia
intrínseca, agentes bióticos extrínsecos, como sombra por competencia en el dosel
y lianas en estratos superiores y ataques de patógenos, o por perturbaciones
fisiológicas abióticas extrínsecas como rayos, sequia e inundaciones (Swaine et
al.1987 y Putz 1983). Y normalmente originan claros pequeños a su alrededor, y una
vez que el tronco está suficientemente descompuesto, se desintegra gradualmente
y cae esparcido en el claro. Este tipo de claros favorece la regeneración de especies
tolerantes a la sombra o se pueden recuperar por expansión de las copas adyacentes
(Whitmore 1978).
Muerto desenraizado: Está sujeto a fuerzas laterales en el tronco (caídas de otros
árboles o perturbaciones catastróficas como vientos o huracanes) que superan la
capacidad de amarre en el suelo por parte de las raíces, pero no rompen el tronco,
esta mortandad puede perturbar las capas del suelo y provocar inversiones en los
horizontes edáficos, lo cual, a su vez, altera el micro-ambiente y tiende hacia la
selección de semillas pioneras del banco del suelo, como las futuras germinadoras
(Putz 1983).
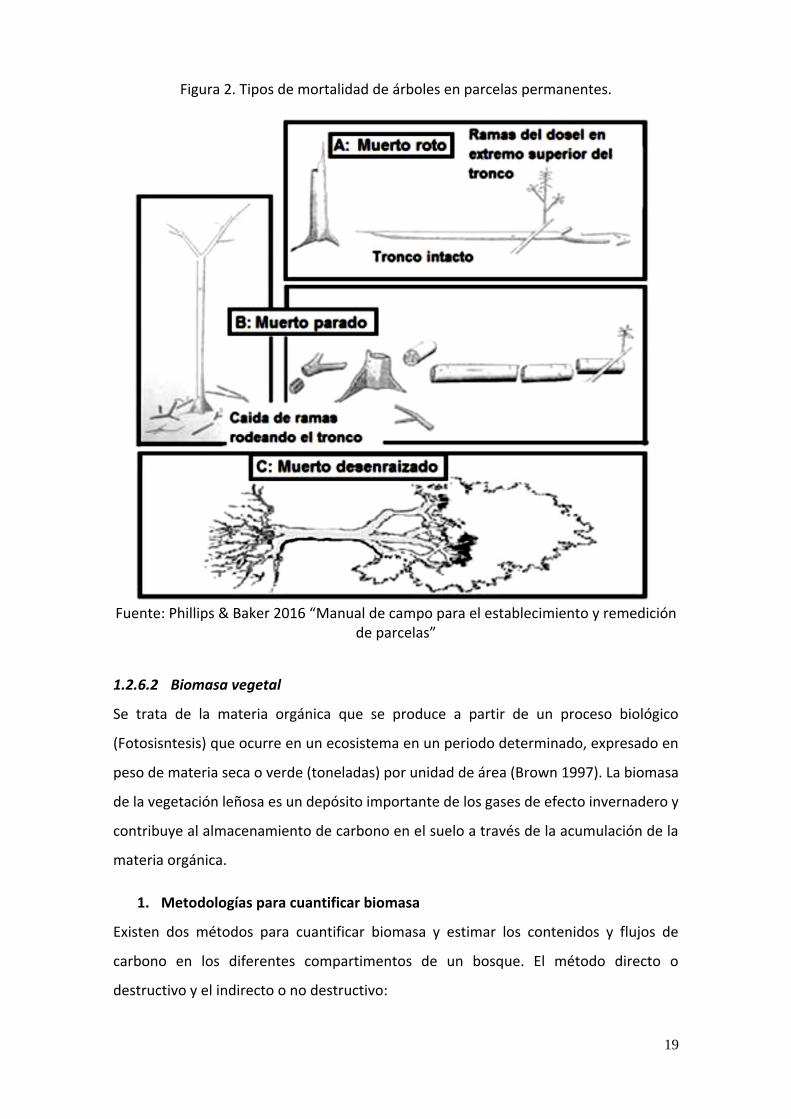
19
Figura 2. Tipos de mortalidad de árboles en parcelas permanentes.
Fuente: Phillips & Baker 2016 “Manual de campo para el establecimiento y remedición
de parcelas”
1.2.6.2 Biomasa vegetal
Se trata de la materia orgánica que se produce a partir de un proceso biológico
(Fotosisntesis) que ocurre en un ecosistema en un periodo determinado, expresado en
peso de materia seca o verde (toneladas) por unidad de área (Brown 1997). La biomasa
de la vegetación leñosa es un depósito importante de los gases de efecto invernadero y
contribuye al almacenamiento de carbono en el suelo a través de la acumulación de la
materia orgánica.
1. Metodologías para cuantificar biomasa
Existen dos métodos para cuantificar biomasa y estimar los contenidos y flujos de
carbono en los diferentes compartimentos de un bosque. El método directo o
destructivo y el indirecto o no destructivo:

20
a) Método directo:
Denominado también método destructivo, implica en la extracción de muestras
(árboles, ramas, hojas, fuste, necromasa aérea, suelo, etc), para luego determinar su
masa seca en laboratorio (biomasa, carbono, necromasa seca). Con estos datos se puede
conocer el contenido de carbono a nivel de individuo, parcela y ecosistema, cuando se
trata de árboles los datos se utiliza para desarrollar ecuaciones alométricas que reflejen
estos contenidos a través de diferentes correlaciones entre los datos de masa seca y
datos obtenidos en campo en un inventario forestal (diámetro del fuste, altura, densidad
de madera), (Honorio y Baker, 2010)
b) Método indirecto:
Denominado no destructivo consiste en utilizar valores en modelos alométricos
generados en otros sitios similares al área de estudio, que permiten relacionar variables
de fácil medición como diámetro del tronco y altura con otras de difícil medición como
densidad específica, área foliar. Estos datos estiman muy bien la biomasa y los
contenidos de carbono en diferentes tipos de bosques.
A partir de este tipo de método, Chambers et al. (2001) obtuvo una marcada similitud
en relación a DAP/biomasa en la mayoría de los bosques amazónicos, relación que
reafirma Phillips et al. (2002), precisando la fiabilidad entre estas dos variables para
obtener un valor robusto de biomasa aérea en estos bosques (Malhi et al., 2002).
Posteriormente se fue incluyendo en los modelos de regresión la variable de densidad
de madera, mejorando la exactitud en el cálculo de la biomasa.
Diámetro a la altura del pecho (DAP)
Esta medición corresponde al diámetro del árbol medido a 1.30 m del nivel del suelo en
condiciones normales es decir cuando el árbol se encuentra en forma perpendicular al
suelo y presenta un fuste recto y cilíndrico. En el campo existen muchos casos
particulares, que ameritan la modificación de dicho procedimiento (Contreras et al,
1999).
Punto óptimo de medida (POM)
Es la medida de 1.3, donde se realiza la medición del DAP (diámetro a la altura del
pecho), cuando el DAP a 1.3 presenta irregularidades que no permiten dar una medida
adecuada, el POM puede variar, según los casos que se presentan a continuación.

21
Tabla 2. Casos especiales donde cambia el POM (Punto Óptimo de Medida)
A) Quebrados
Los árboles con tallos quebrados o que han perdido al
menos la mitad de su copa se miden a 1,30 m (Vallejo y
Joyas et al. 2005).
B) Múltiples
troncos
Todos los troncos mayores de 10 de DAP a 1.3m son
medidos, pintados y registrados.
C) Deformaciones
Si el árbol tiene una deformidad en 1.3m de altura, la
medida se realiza 2cm por encima o por debajo de la
deformidad, es donde se registra la altura del POM (Condit
1998).
D y F)
Pendientes y
árboles caídos
o inclinados
El diámetro a la altura de referencia, siempre se calcula
cuesta abajo, en la dirección de la pendiente del árbol y los
árboles caídos o inclinados son siempre medidos a 1.3m de
altura del lado más cercano al suelo.
E) Contrafuertes
y zancos
Si el árbol presenta contrafuertes, raíces tablares o zancos
a 1.3m, la medida del tallo es a 50cm por encima del final
de los contrafuertes, raíces tablares o zancos donde se
registra en POM (Condit 1998).
G) Tocones
Los tocones deben considerarse si presentan un DAP
adecuado (≥ 10 cm), en cuyo caso se medirá POM, la altura
máxima y la especie, en los casos que sea posible.
H) Arboles
acanalados
Los árboles que son acanalados en todo el tronco se miden
a 1.3m.
I) Rebrotes
En árboles erguidos pero rotos, o individuos caídos el
tronco principal y los rebrotes son medidos a 1.3m de la
base del tronco y solamente son incluidos si están por
encima de 1.3m de la base den tronco.

22
Figura 3. Representación gráfica de casos especiales donde cambia el POM
Fuente: Osinaga et al. 2014
A) B) C)
D) E) F)
G)

23
2. Biomasa aérea
Abarca toda la materia viva sobre el suelo, como árboles vivos, plantas rastreras,
enredaderas y epifitas, así como las plantas del sotobosque y la vegetación herbácea. La
estimación de la biomasa aérea arbórea es esencial para los estudios de almacenes y
flujos de carbono en un bosque (Phillips et al., 1998).
3. Relación biomasa – carbono
El carbono es capturado como dióxido de carbono y almacenado por las plantas vía
fotosíntesis (Aragão et al. 2009), junto con elementos de los suelos y el aire para
convertirlos en biomasa. Aproximadamente el 42 % a 50 % de la biomasa de un árbol es
carbono (Gibbs et al 2007); se asume que el contenido de carbono corresponde al 50%
de la biomasa de los árboles vivos (MacDicken 1977, Fearneside et al. 1999, Clark et al.
2001, Malhi et al. 2004, Chave et al. 2005, Aragão et al. 2009, etc.), por lo que se sugiere
usar el factor de 0,5 para transformar la biomasa aérea a contenido de carbono.
El carbono es un indicador fundamental para analizar la integridad de un ecosistema y
la provisión de bienes y servicios ambientales. Este es sensible a los efectos directos de
la deforestación y la degradación, así como en los impactos esperados por anomalías
climáticas y las posibles alteraciones en la productividad de los ecosistemas (REDD,
2011).
1.2.6.3 Analisis poblacional
1. Composición florística
Se enfoca en la diversidad de especies en un ecosistema la cual se mide por su riqueza
(cantidad de especies), representatividad (balance equitativo de especies) y
heterogeneidad (disimilitud entre riqueza y representatividad) (Cerón, 2003),
resultando de procesos que operan a distintas escalas espaciales y temporales, estos
procesos actúan como filtros que seleccionan a aquellas especies que poseen las
características adecuadas para soportarlos (Díaz et al. 1998).
Generalmente los trabajos de composición florística, se centran en árboles, pues estos,
además de constituir la mayor parte de la biomasa del bosque, determinan en gran parte
su estructura y funcionamiento (Berry 2002). Los estudios sobre la composición florística

24
es una excelente contribución al conocimiento de la sistemática de plantas vasculares
en general, además que permite comparar con otras floras locales (Vásquez et al. 2003).
2. Importancia ecológica
Es el valor que adquiere una especie o familia dentro de una comunidad vegetal,
respecto a las demás especies, que resulta de la suma de las variables relativas como
frecuencia, dominancia y cobertura de cada especie en cada muestra estimada
(Matteucci & Colma 2002) indicando cuán importante es la presencia de una especie y/o
familia en la comunidad vegetal.
3. Diversidad
La diversidad es una característica compleja de los ecosistemas, una composición
interespecifica de las comunidades y se dividen en tres niveles: genético
(intraespecifico), de especies (número de especies) y ecológico (de comunidades) según
Norse et al (1986).
a) Índices de diversidad
La diversidad alfa es el conjunto de especies de una comunidad particular a la que se
considera como un hábitat homogéneo, la diversidad beta analiza el cambio en la
composición de especies en diferentes comunidades o a través de gradientes
ambientales y la diversidad gamma es la composición de especies del conjunto de
comunidades que integran el paisaje (Wittaker, 1960, 1972, Wilson y Molher, 1983).
25
CAPITULO II
ÁREA DE ESTUDIO
2.1. UBICACIÓN
2.1.1. Ubicación Política
El presente estudio se llevó a cabo en las parcelas permanentes instaladas por RAINFOR
en los sectores de Intipunku y Kantupata, en los bosques montanos de Wiñaywayna
dentro del Santuario Historio de Machupicchu, Distrito de Machupicchu, Provincia de
Urubamba y Región del Cusco – Perú.
2.1.2. Ubicación Geográfica
Las dos parcelas permanentes se encuentran ubicadas a través de una gradiente
altitudinal en las siguientes coordenadas:
Tabla 3. Datos geográficos de las parcelas permanentes de Wiñaywayna en el Santuario Histórico de Machupicchu.
PARCELA DATOS
SHM – 01 “Intipunku”
SHM – 02 “Kantupata”
Latitud 13º 10' 37" S 13º 12' 11.3" S
Longitud 72º 32' 13" W 72º 32' 03.5” W
Altitud 2653 m 3200 m
2.2. LIMITES
Por el norte: limita con el rio Vilcanota, la ciudadela de Machupicchu y con el
Distrito de Machupicchu (Aguas calientes)
Por el sur: Limita con los sitios arqueológicos de Phuyupatamarca y Sayaqmarca.
Por el este: Limita con el rio Vilcanota y la cuenca de Pacaymayo.
Por el oeste: Limita con la cuenca del rio Ahobamba.
2.3. ACCESIBILIDAD
Tabla 4. Rutas de acceso al área de estudio.
Tramo - Recorrido Medio de Transporte Vía de Acceso
Cusco - Ollantaytambo Automóvil Particular Asfaltado
Ollantaytambo – Km 107 Tren Ferrocarril
Km 107 – Estación Biológica de WiñayWayna A pie (2 horas) Sendero

- 26 -
Mapa 1: Ubicación del área de estudio.

- 27 -
Mapa 2: Santuario Histórico de Machupicchu

28
2.4. SANTUARIO HISTÓRICO DE MACHUPICCHU
Creada bajo el Decreto Supremo N° 001-81-AA, como Santuario Histórico de
Machupicchu sobre la superficie de 37 302.58 ha; ubicado en el distrito de Machupicchu,
en la Provincia de Urubamba del Departamento de Cusco. Es el sitio de patrimonio
Mundial de mayor importancia en el Perú y fue Inscrito el 09 de diciembre de 1983 en
la lista de sitios del Patrimonio Mundial de UNESCO en la categoría de Patrimonio Mixto,
debido a sus características culturales y naturales.
Los objetivos de conservación del Santuario Histórico de Machupicchu, se sustentan
bajo las prioridades de conservación de valores culturales de los diversos monumentos
arqueológicos, los caminos del inca y sobre todo la sagrada ciudad de Machupicchu
como centro articulador del entorno y la naturaleza como la diversidad de ecosistemas,
especies y la particular geomorfología, reconociéndose de esta manera como
patrimonio mixto de la Humanidad. (Plan Maestro del Santuario Histórico de
Machupicchu 2014)
2.4.1. Componente físico
2.4.1.1. Geología
La mayor parte del Santuario Histórico de Machupicchu se halla sobre un conjunto de
rocas ígneas intrusivas del batolito de Vilcabamba, que está formado principalmente por
granitos y granodioritas. Estas rocas se hallan cortadas por diaclasas y fallas, las que han
jugado un papel determinante modelando la morfología y el aspecto que presentaba la
zona cuando los Incas llegaron al lugar. En el Santuario afloran también rocas
metamórficas del Paleozoico Inferior, rocas sedimentarias de edad Mesozoica y
Cenozoica, y depósitos cuaternarios como coluviales, fluviales y conos de deyección.
Sobre esos diferentes tipos de suelos y, en algunos casos, sobre los mismos
afloramientos rocosos, se ha construido la mayoría de monumentos arqueológicos,
incluyendo la Llaqta de Machupicchu y los caminos del Inca (Carlotto, Cárdenas y Fidel,
2009).

29
2.4.1.2. Geomorfología
La geomorfología de Machupicchu tiene un fuerte control estructural o tectónico y se
caracteriza por un terreno de relieve accidentado y laderas escarpadas. La elevación
mínima es de 1800 m sobre el mar en el valle del Urubamba; mientras que los lugares a
mayores altitudes bordean los 5000 m sobre el mar. En lo más elevado del territorio
perteneciente a la cordillera oriental, se pueden encontrar glaciares con signos de
retroceso y evidencia de glaciaciones anteriores.
Se ha identificado la existencia de dos fallas regionales NO-SE, denominadas Urubamba
y Central Hidroeléctrica de Machupicchu, que forman un corredor estructural dentro del
cual se han desarrollado las fallas NE-SO de Machupicchu, Waynapicchu, Foso Seco y
otras. Esta distribución tectónica ha influenciado la forma del río Urubamba y sus
vertientes y particularmente el graben sobre el cual fue construida la ciudad. Además,
las fallas controlan las laderas empinadas, donde se desarrollan deslizamientos como
del cerro Machupicchu (Carlotto, Cárdenas y Fidel, 2007).
2.4.1.3. Fisiografía
Fisiográficamente se identifican dos grandes paisajes, uno es el paisaje de planicies y el
otro el montañoso, los cuales reciben sus características, de su proceso de formación y
del relieve. El territorio de planicies está conformado por planicies de origen glaciar,
aluvial y coluvial. Mientras el paisaje montañoso, lo conforman montañas de rocas
volcánicas, sedimentarias, metamórficas, e intrusivas.
Los bosques de Wiñaywayna se sitúan en la vertiente suroeste del río Urubamba, y que
se caracteriza por ser muy accidentada, debido principalmente a las laderas con
pendientes empinadas y los grandes desniveles que originó el río sobre el macizo de
rocas intrusivas del batolito de Vilcabamba. (Plan Maestro del Santuario Histórico de
Machupicchu 2014).

- 30 -
Mapa 4: Fisiografía del área de estudio

31
2.4.1.4. Edafología
Teniendo en cuenta los diversos materiales existentes en la zona, se presenta un patrón
distributivo de los suelos según su origen; estos han originado los diferentes suelos
presentes aquí: Suelos derivados de materiales fluvio-glaciales, suelos derivados de
materiales coluvio-aluviales, suelos derivados de materiales aluviales, suelos de origen
antropogénico, suelos derivados de materiales residuales.
En los sectores de Chachabamba, Pacaymayu, Wiñaywayna, parte de la quebrada de
Aobamba hacia el sector de Churubamba, los suelos son poco profundos, superficiales,
predominando los litosoles y cambisoles de coloración pardusca.
En los sectores colindantes con Chachabamba, Pacaymayu y Wiñaywayna los suelos por
lo general son de profundidad media de textura variable entre media y fina de pH sobre
7 siendo calcáreo y puede ser semejante a los cambisoles eútricos, kastonozems y en
algunas áreas phaeozems, los litosoles y las rendzinas aparecen en aquellos lugares de
suelos superficiales y empinados. (Plan Maestro del Santuario Histórico de Machupicchu
2014)
2.4.1.5. Hidrografía
Hidrográficamente, el Santuario Histórico de Machupicchu pertenece a la cuenca
Urubamba, vertiente del Atlántico. Comprende al río Vilcanota desde la quebrada
Misquipuquio (2700 m sobre el mar), y la convergencia aguas abajo con el río Aobamba
(1810 m sobre el mar), que nace aproximadamente a 3500 m sobre el mar, en la
confluencia de los ríos Orcospampa y Rayankancha, que provienen de los deshielos de
los nevados Salkantay, Palqay, Umantay y otros adyacentes, el río Aobamba desemboca
en el río Vilcanota a 1700 m sobre el mar. La hidrografía del Santuario Histórico de
Machupicchu tiene como límites los ríos Aobamba y Cusichaca que desembocan en el
río Vilcanota, el valle Cusichaca tiene una dirección aproximada noreste-suroeste, y las
aguas de este río nacen en los nevados Salkantay y Palqay, que desembocan en el río
Vilcanota, a 2450 m sobre el mar. (Plan Maestro del Santuario Histórico de Machupicchu
2014)

- 32 -
Mapa 5: Red hidrográfica del Santuario Histórico de Machupicchu

33
2.4.2. Componente climático
2.4.3.1. Climadiagrama.
Tabla 5. Datos meteorológicos de 6 años de la estación de Aguas Calientes del Santuario Histórico de Machupicchu.
MESES Temperatura (C°) Precipitación (mm)
Julio 15.66 36.28
Agosto 16.5 56.38
Septiembre 17.42 38.76
Octubre 17.39 155.56
Noviembre 17.14 177
Diciembre 16.47 231.1
Enero 15.99 383.6
Febrero 16.24 247.64
Marzo 16.08 336.82
Abril 16.42 174.5
Mayo 15.9 62.96
Junio 15.51 38.94
Promedio 16.39
Total 1939.54
Fuente: SENAMHI 2012.
Figura. 4. Climatodiagrama del Santuario Histórico de Machupicchu.
Fuente: Elaboración propia
0
40
80
120
160
200
240
280
320
360
400
0
20
40
60
80
100
120
140
160
180
200
Pre
cip
itac
ion
(m
m)
Tem
pe
ratu
ra (
C°)
Estación meteorologica de Aguas calientes (2004 - 2011)
T (C°) P (mm)
T° Pro 16.39 °C Pp 1939.54mm

34
El clima en el Santuario Historico de Machupicchu presenta muchos contrastes debido
a la existencia de diferentes franjas climatológicas relacionadas con los cambios
altitudinales y la variada configuración morfológica del terreno (Plan Maestro – SHM de
2015 a 1019).
En forma general para el Santuario Historico de Machupicchu, el clima está
caracterizado por la alternancia de una estación seca entre los meses mayo a setiembre
y una estación lluviosa entre los meses octubre a abril con tres meses con mucha lluvia
que va de enero a marzo, especialmente en las zonas altas. La precipitación anual es de
1939.54mm y la tempreratura promedio de 16.39°C.
El clima varía de húmedo y cálido en las zonas bajas de 1700 a 2500 m a húmedo -
subhúmedo y templado frío – frío en las zonas mas altas entre 2500 a 4350m (Plan
Maestro – SHM de 2015 a 1019).

35
2.4.4. Componente biológico
2.4.4.1. Zonas de vida
Las zonas de vida están definidas como “un grupo de asociaciones vegetales dentro de
una división natural del clima, que se hacen teniendo en cuenta las condiciones edáficas
y las etapas de sucesión, y que tiene un aspecto físico similar en cualquier parte del
mundo” (Holdridge en 1967). Las características básicas de la zona de vida (vegetación,
tipos de suelo, ciclos de energía y nutrientes, fauna silvestre, otros), esta definidda por
la interrelacion de tres parámetros climáticos fundamentales como la precipitación,
temperatura y humedad atmosférica. (Holdridge 1978).
Para el Santuario Historico de Machupicchu se describieron 10 zonas de vida:
Tabla 6. Zonas de vida consideradas para el Santuario Historico de Machupicchu.
N° PRECIPITACION
(mm)
ALTITUD
(m sobre el mar)
TEMPERATURA
(°C) DESCRIPCION DE ZONA DE VIDA Ha %
1 1700 - 1800 1700 y 1900 m 18° y 24° Bosque húmedo - Premontano
Subtropical (bh – PS) 81 0.22
2 900 - 1000 1900 y 2900 m 12° y 18°, Bosque muy húmedo - Montano
Bajo Subtropical (bh – MBS) 6452 17.26
3 2000 - 2100 2000 y 2300 m 16.5° y 14.5° Bosque húmedo - Montano Bajo
Subtropical (bmh – MBS) 7 0.02
4 1900 - 2000 2900 y 3800 m 6° y 12° Bosque muy húmedo - Montano
Subtropical (bmh – MS) 9730 26.03
5 1000 - 1100 2500 y 2800 m 18° y 12° Bosque seco - Montano Bajo
Subtropical (bs – MBS) 1118 2.99
6 800 - 900 2900 y 3800 m 6° y 12° Bosque húmedo - Montano
Subtropical (bh – MS) 3730 9.98
7 700 - 800 3900 y 4350 m 3° y 6° Páramo - Subalpino Subtropical
(p – SaS) 847 2.27
8 1700 - 1800 3900 y 4350 m 3° y 6° Páramo pluvial - Subalpino
Subtropical (pp – SaS) 7420 19.85
9 1400 - 1500 4350 y 4750 m 3° y 1.5° Tundra pluvial - Alpino
Subtropical (tp – AS) 4 567 12.22
10 1000 - 1100 sobre los 4750m inferior a 1.5° Nival – Subtropical (n – S) 3428 9.17
AREA TOTAL 37320 100
Fuente: Plan maestro – SHM 2005 (INRENA 2003), (Galiano et al. 1995) y (Acurio 1998 y
2000)

36
De las 10 zonas de vida antes mencionadas, las parcelas permanenetes del presente
estudio se encuentran en dos zonas de vida que son:
A. Bosque muy húmedo - Montano Bajo Subtropical (bmh – MBS):
La parcela SHM – 01 de Intipunku, se encuentra dentro de esta zona de vida, que ocupa
el fondo del valle del río Urubamba y las primeras elevaciones de las laderas montañosas
que lo circundan, comprendiendo entre los 1900 a 2900 m, presenta un clima templado
y muy húmedo con una precipitación total anual de 900 a 1000 mm y una temperatura
promedio de 12 a 18°C. La vegetación natural está conformada por un bosque siempre
verde con árboles de hasta 30 m de alto. Entre las especies arbóreas se distinguen al
“Pisonay” Erythrina sp., “Aliso” Alnus acuminata, Ocotea sp., “Toroj” Cecropia sp.,
Podocarpus glomeratus, Inga microcoma, Jacaranda sp., “Cedro” Cedrela odorata,
“Quina” Cinchona officinalis, Eugenia sp., Carica sp., Rubus sp.
B. Bosque muy húmedo - Montano Subtropical (bmh – MS):
La parcela SHM – 02 de Kantupata se encuentra dentro de esta zona de vida, que ocupa
la porción media de las laderas montañosas, entre los 2900 y 3800 m, incluye los grupos
arqueológicos localizados en la margen izquierda del río Urubamba como Intipata,
Wiñaywayna, Phuyupatamarca, Kantupata y Sayacmarca. Presenta un clima frío y pluvial
con una precipitación total anual de 1900 a 2000mm y una temperatura promedio de 6
a 12ºC. La vegetación natural está conformada por árboles de hasta 25 m de alto, entre
los que se puede considerar los siguientes: “Aliso” Alnus acuminata, “Unca” Myrcianthes
indifferens, Polylepis sp., Miconia sp., “Intimpa” Podocarpus glomeratus, “Carapacho”
Weinmania sp., Eugenia sp., “Maqui maqui” Oreopanax sp., Clusia sp., “Laurel” Myrica
pubescens, Chusquea sp., Brachiotum sp. y “Helechos arbóreos” Cyathea sp.
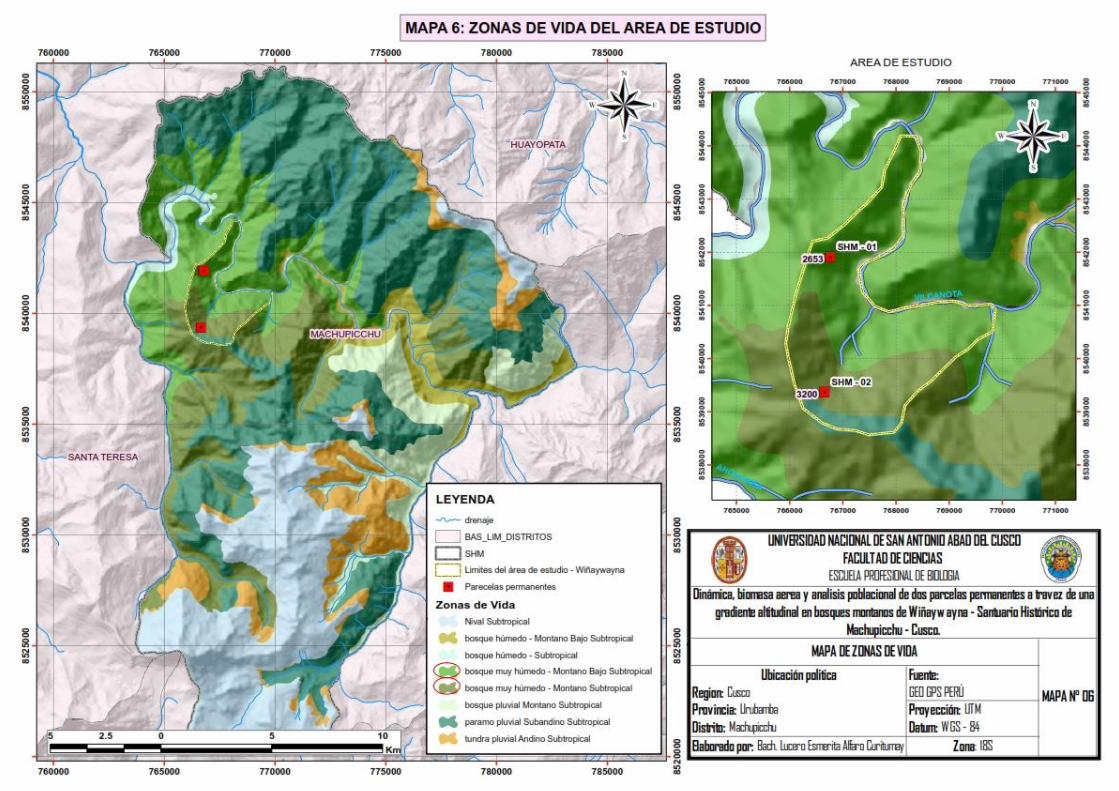
- 37 -
Mapa 6: Zonas de vida del área de estudio

38
2.4.4.2. Sistemas ecológicos
Son unidades naturales de escala intermedia que reflejan la afinidad geográfica de
conjuntos de especies, una fisonomía vegetativa predominante, procesos y condiciones
ecológicas que determinan la ocurrencia conjunta de comunidades naturales en un
ambiente dado (Carmen Josse 2003). Comprende comunidades vegetales que tienden a
co-ocurrir, compartiendo procesos ecológicos, sustratos o gradientes ambientales
(Carmen Josse 2007).
Se registraron un total de 12 sistemas ecológicos para el Santuario Historico de
Machupicchu, que son:
Tabla 7. Sistemas ecológicos considerados para el Santuario Historico de Machupicchu.
N° Altitud
(m sobre el mar) Sistemas Ecológicos (SE) Área (ha)
1 5100 – 5300 Nival. 3720.70
2 2900 a 3700 CES409.059 Pajonal arbustivo altoandino y altimontano pluviestacional de yungas.
4680.54
3 2900 a 4200 CES409.058 Pajonal arbustivo altoandino y altimontano pluvial de yungas.
937.75
4 >3900 - 4100 CES409.084 Pajonal altoandino de la Puna húmeda 10 029.60
5 3100 a 4100 CES409.087 Pajonales y matorrales altimontanos de la Puna húmeda.
1016.08
6 2100 a 2900 CES409.921 Bosque montano pluviestacional subhúmedo de yungas.
1681.05
7 3100 a 3900 CES409.046 Bosque de Polylepis altimontano pluviestacional de yungas.
510.10
8 1900 a 2900 CES409.051 Bosque montano pluviestacional húmedo de yungas.
6009.20
9 1200 a 2100 CES409.054 Bosque basimontano pluviestacional húmedo de yungas.
390.11
10 2900 a 3600 CES409.044 Bosque altimontano pluviestacional de yungas.
5418.17
11 2900 a 3500 CES409.043 Bosque altimontano pluvial de yungas. 1989.38
12 Áreas intervenidas. 919.90
TOTAL 37 302.58
Fuente: Plan Maestro del SHM (2015 – 2019)
De estos sistemas ecológicos, las parcelas permanentes del presente estudio se
encuentran dentro de 2 sistemas ecológicos que son:

39
A. CES409.051 Bosque montano Pluviestacional húmedo de yungas
La parcela SHM – 01 de Intipunku se encuentra dentro de este sistema que presenta
suelos bien drenados en laderas montañosas, con termoclina meso tropical y
ombroclima húmedo. Generalmente presente en la transición de los valles interandinos
secos hacia los valles y laderas pluviales húmedos de las Yungas y con coberturas
extensas sobre sustratos arenosos, se distribuye entre los 1900 m a 2900 m, albergan
bosques con dosel de 15 a 25 m. Se encuentran dominados por especies de
Podocarpaceae o de los generos Weinmannia, Styloceras, Juglans, Cynchona,
Nectandra, Cedrela, Ceroxylon, Prunus, Styrax, Ternstroemia, Podocarpus, Hedyosmun,
Morus, Etc. (Josse y Navarro 2007)
B. CES409.044 Bosque Altimontano Pluviestacional de yungas
La parcela SHM – 02, pertenece a este sistema que presenta bosques altimontanos
propios de las zonas con bioclima Pluviestacional húmedo de las yungas, distribuidos
desde el norte del Perú al centro de Bolivia. Ocupan laderas montañosas altas con suelos
profundos bien drenados, termoclina supratropical inferior y ombroclima húmedo, se
encuentran distribuidos entre los 2900 m a 3600 m, la vegetación es denso siempre
verde estacional con dosel de 10 a 20 m, son bosques de altura media, dominados por
los géneros Podocarpus, Myrcianthes, Weinmania, Oreocallis, Roupala, Clethra,
Escallonia, Oreopanax, Cervantesia, Styloceras, Nectandra, Hesperomeles, Duranta y
Prunus. (Josse 2007)

- 40 -
Mapa 7: Sistemas Ecológicos del aérea de estudio

41
2.4.4.3. Flora
Para el Santuario Histórico de Machupicchu, se registraron hasta el momento unas 3250
especies de plantas entre Angiospermas y Gimnospermas que representan el 15.8% de
la diversidad del Perú, dentro de ella se encuentran más de 370 especies de árboles
pertenecientes a 67 familias y a 45 géneros. Las familias de árboles que presentan mayor
número de especies son:
Tabla 8. Familias de flora arbórea con mayor numero de especies registradas para el Santuario Histórico de Machupicchu.
Familias N° de especies
Melastomataceae 40
Lauraceae 26
Moraceae 17
Euphorbiaceae 14
Rosaceae 14
Solanaceae 14
Rubiaceae 40
Fuente: Plan Maestro del SHM, 2015 – 2019
2.4.4.4. Fauna
El Santuario Histórico de Machupicchu, alberga una cantidad numerosa de especies de
vertebrados, en un porcentaje considerable respecto a la fauna a nivel nacional.
Tabla 9. Especies de fauna registradas para el Santuario Histórico de Machupicchu.
Phylum Clase Total de especies
VERTEBRADOS
Mammalia 77
Aves 443
Osteichthyes 13
Amphibia 16
Reptilia 26
INVERTEBRADOS Insecta 377
Fuente: Plan Maestro del SHM, 2015 - 2019

42
Tabla 10. Categoría de especies de fauna registradas para el Santuario Histórico de Machupicchu.
Clase Especies Nombre común Estado
Mammalia
Tremarctos ornatus
Hippocamelus antisensis
Odocoileus peruvianus
Mazama chunyi
Pudu mephistophiles
Lontra longicaudis
Puma concolor
“oso de anteojos”
“taruca”
“venado cola blanca”
“tanka taruka”
“venado enano”
“mayupuma o nutria”
“puma”
Amenazado
Aves
Vultur gryphus
Andigena hypoglauca
Cinclodes aricomae
Cacicus koepckeae
Asthenes urubambensis.
“cóndor andino”
“tucán andino el churrete real”
“Cacique de Koepcke”
“Canastero frente listada”
Amenazado
Osteichthyes
Acrobycon ipanquianus,
Bryconamericus pachacuti,
Ceratobranchia binghami.
Amphibia
Nannophryne corynetes
Gastrotheca ochoai
Nymphargus pluvialis
“Sapo de Abra Málaga”
“Rana marsupial”
“Rana cristal
Vulnerable
Critico
Vulnerable
Reptilia Bothrocopias andianus. “terciopelo” o “jergón andina” Vulnerable
Insecta
(Lepidóptera)
Lycorea halia pales (“Mariposa monarca”)
Higginsius fasciata,
Styx infernalis,
Thespieus pinda,
Chiodes concinnus.
Morpho lympharis,
Danaus plexippus nigrippus.
Eryphanis zolvizora zolvizora.
Fuente: Plan Maestro del SHM, 2015 - 2019

43
CAPITULO III:
MATERIALES Y METODOLOGÍA
3.1. MATERIALES
Biológico
Muestras botánicas
De Campo
Libretas de campo
Etiquetas de campo
Cintas de plástico
Plumones Sharpie
Cordeles o rafia
Machete
Clavos de aluminio
Placas de aluminio
Lápices de carbón
Linternas frontales
Cinta diamétrica
Equipo de montaña
Wincha de 25m y 50m.
Brújula
Clinómetro
Tijera telescópica
Patas de Loro
Binoculares
Cámara fotográfica
GPS Garmin
De Herborización
Bolsas de polietileno
Sacos vacios de urea
Papel periódico
Alcohol 76%
Tijeras de podar
De Gabinete
- Papel cansón
- Mapas a diferentes escalas
- Microscopio Estereoscópico
- Lupas 10X
- Patrones y claves de identificación
- Computadora (Software Estadístico)
- Bibliografía especializada

44
3.2. METODOLOGÍA
La toma de datos se realizó en dos censos, con un intervalo de 3 años en setiembre del
2013 se llevó a cabo el primer censo y noviembre del 2016 el segundo censo, bajo el
sistema estandarizado de procedimientos para bosques montanos establecido por la
Red Amazónica de Inventarios Forestales (RAINFOR) 2001, donde se incluyeron todos
los individuos mayores e iguales a 10 cm de diámetro.
3.2.1. Ubicación de las parcelas
Las parcelas fueron establecidas en forma aleatoria usando imágenes de Google Earth
como información preliminar, estas imágenes y coordenadas fueron contrastadas en el
campo con las condiciones adecuadas para establecer una parcela permanente.
Se estableció dos parcelas permanentes de 1.0-ha cada una en los sectores de Intipunku
y Kantupata en los bosques montanos de Wiñaywayna dentro del Santuario Histórico de
Machupicchu en setiembre del año 2013, donde se tuvo en cuenta los diferentes
aspectos que garanticen su viabilidad y control adecuado como, una accesibilidad
adecuada, considerable distancia respecto a las vías de ingreso y un área casi
homogénea y no perturbada.
Tabla 11. Datos geográficos de ubicación de parcelas permanentes de Wiñaywayna
Parcela Zona de vida Área (ha)
Latitud Longitud Altitud
(m) Año
SHM-01
(Intipunku)
Bosque muy húmedo
Montano bajo
Subtropical (bmh-MBS)
1.0 766763.43 8541912.86 2653 2013
SHM-02
(Kantupata)
Bosque pluvial Montano
Subtropical (bp-MS) 1.0 766653.80 8539361.09 3200 2013
Fuente: Elaboración propia

- 45 -
Mapa 3: Ubicación de las parcelas permanentes de muestreo

46
3.2.2. Evaluación de las parcelas
A) Orientación y estructura
Las parcelas permanentes se instalaron con las direcciones S/N y W/E para los ejes
principales de manera más conveniente, donde se registró la latitud, longitud y altitud
del centro de la parcela.
Las parcelas consisten en un cuadrado de 100m x 100m (1.0-ha), dividido en 25 sub
parcelas de 20m x 20m, en las cuatro esquinas se colocaron estacas de plástico para una
mejor ubicación (Phillips et al., 2009).
Figura 5. Disposición de las parcelas permanentes de 1.0 – ha en Wiñaywayna.
B) Modo de desplazamiento
Se registraron los datos de forma ordenada, desde el punto (0,0) en las coordenadas (X,
Y) respectivamente, siguiendo la dirección W/E y S/N desde la sub parcela 1, los arboles
fueron plaqueados sistemáticamente moviéndose alrededor de cada sub parcela con el
ultimo árbol plaqueado en cada sub parcela cerca al punto de partida de la próxima sub
parcela (Phillips et al., 2009).

47
Figura 6. Modo de desplazamiento dentro de las parcelas permanentes de Wiñaywayna
C) Ubicación de cada arbol
Se ubicaron los individuos dentro de la sub parcela por medio de coordenadas de
referencia donde “x” indica la distancia generada por la perpendicular entre el árbol y el
eje “y”, mientras que “y” indica la distancia generada por la perpendicular entre el árbol
y el eje “x” (Phillips et al., 2009).
Figura 7. Ubicación de los arboles dentro de la sub parcela
D) Medición del Diámetro a la Altura del Pecho (DAP)
Se midió el diámetro (DAP) en el Punto Óptimo de Medida (POM) que estandarizado es
a 1.30 m sobre el nivel del suelo, con una cinta diamétrica a todos los arboles con DAP
≥ 10 cm en cada parcela. Cada individuo fue marcado con placas de aluminio a 30 cm
5 4 3 2 1
15 14 13 12 11
109876
25 24 23 22 21
2019181716
(100,100)
(0,0)(0,100)
(100,0)
100
100
X
Y
Eje
Eje
Y
(Xi , Yi)

48
encima o debajo del POM según corresponda la estructura del árbol. El perímetro del
tronco del árbol donde se midió el DAP, se marcó con pintura esmalte rojo con el objeto
de garantizar que las posteriores mediciones se realicen en el mismo POM de la primera
medición (Phillips et al., 2009).
Los nuevos reclutas fueron marcados con códigos (.1, .2,… ) respecto al árbol más
cercano, por ejemplo, si el recluta está próximo al árbol 146, su código será 146.1.
Figura 8. Ubicación de los individuos dentro de la sub parcela
E) Casos especiales donde se cambiaron el POM
Deformidades: Si el árbol tuvo una mayor deformidad en 1.3m de altura, la
medida se hizo 2 cm por debajo o por encima de la deformidad donde se registro
el POM (Condit 1998).
Árboles acanalados: Los árboles que son acanalados en todo el tronco deben ser
medidos a 1.3m (Phillips et al., 2009).
Pendientes y árboles caídos o inclinados: El diámetro a la altura de referencia
(DRH) siempre se calcula cuesta abajo, en la dirección de la pendiente del árbol,
y los árboles caídos o inclinados son siempre medidos a 1.3m de altura del lado
del tallo más cercano al suelo. En árboles caídos es difícil definir la base del
tronco cuidadosamente – por lo tanto, medir los árboles 30 cm por debajo de la
placa (Phillips et al., 2009).
Rebrotes: En árboles erguidos, pero rotos, o individuos caídos, el tronco principal
y los rebrotes son medidos a 1.3m de la base del tronco. Un individuo con

49
rebrotes solamente es incluido si los rebrotes están por encima de 1.3m de la
base del tronco (Phillips et al., 2009).
Múltiples troncos: Todos los troncos más grandes de 10cm a 1.3m, son medidos,
pintados y registrados (Phillips et al., 2009).
F) Registro de datos
Se registrarón los siguientes datos para cada Arbol:
1) Número de la placa
2) Número de subparcela
3) Coordenadas (x, y)
4) Familia, nombre científico y número de colección
5) Diámetro a la Altura del Pecho (DAP) (1.30m)
6) Punto Óptimo de Medida
7) Altura del árbol
8) incidencia de luz.
9) Condición del árbol, en base a los códigos RAINFOR de árboles (vivos o
muertos), para el trabajo de campo y para la base de datos.
G) Colecta del material botánico
La muestra vegetal es considerada como el factor de mayor importancia en un trabajo
de tipo florístico, en razón a ello se colectó dos ejemplares de todos los individuos
registrados en los inventarios, las muestras botánicas fueron colectadas con la ayuda del
equipo para trepar árboles, harnees y patas de loro, en cada colecta se trató de obtener
una muestra con flor o fruto para facilitar su determinación tanto en el campo como en
el herbario, y se registró en la libreta de campo las características dendrológicas
consistentes en: forma del fuste, color de la corteza interna y externa, olor, sabor, color
de resina, látex; igualmente la forma, disposición, textura, de las hojas; color de flores,
frutos etc. (Phillips et al., 2009)., estas muestras posteriormente fueron conservadas y
preservadas con alcohol al 75% para su posterior tratamiento en el secadero Fortunato
L. Herrera, de la Escuela Profesional de Biología.

50
H) Determinación de las muestras botánicas
Para la determinación del material botánico, se morfo especió, con ejemplares patrón
identificados y codificados en el Herbario Vargas (CUZ). Se utilizó también para la
identificación, bibliografía especializada, claves dicotómicas para todas las familias,
géneros y especies.
Los nombres científicos fueron revisados y actualizados con listados en la página web de
TRÓPICOS (http://www.tropicos.org/) y Forestplots Database www.foresplots.net. Los
géneros y familias fueron basados en la clasificación filogenética de las angiospermas –
APG IV (2016).
3.2.3. Analisis de datos
3.2.3.1. Dinámica poblacional del bosque
La dinámica poblacional se determino mediante 3 parametros que son la tasa de
reclutamiento (TR), la tasa de mortalidad (TM) y la tasa de renovación (TRe) de
individuos de árboles en cada una de las parcelas, para lo cual se usó las siguientes
ecuaciones.
1. Reclutamiento
Para el cálculo de la tasa anual de reclutamiento se utilizó la siguiente ecuación:
Dónde:
TR: Tasa anual de reclutamiento
Δt: Diferencia entre dos periodos de medición
Nm: Número de Individuos muertos en una determinada área.
No: Número de individuos al comienzo del periodo.
Nr: Número de individuos reclutados en el segundo periodo. (Nebel et al. 2000)
𝐓𝐑 =𝐋𝐧(𝐍𝐨 − 𝐍𝐦 + 𝐍𝐫)– 𝐥𝐧(𝐍𝐨 − 𝐍𝐦)
∆𝐭

51
2. Mortalidad
Para el cálculo de la tasa anual de mortalidad se utilizó la siguiente ecuación:
Dónde:
TM: Tasa anual de mortalidad
Δt: Diferencia entre dos periodos de medición
Nm: Número de Individuos muertos en una determinada área.
No: Número de individuos al comienzo del periodo. (Nebel et al. 2000)
3. Recambio (% de Turnover)
Indica la velocidad de renovación de la biomasa, la cual tiende a disminuir al aumentar
la madurez y complejidad, la cual se da en función a la tasa de mortalidad y
reclutamiento.
(Nebel et al. 2000)
3.2.3.2. Cuantificación de biomasa aérea almacenada
1. Estimación de la cantidad de biomasa aérea individual
Se estimó la biomasa aérea para cada árbol en función al diámetro a la altura del pecho
(d.a.p), altura y a la gravedad específica de madera o densidad (ρ) que sigue el modelo
alométrico seleccionado (Alvarez et al., 2012), según zonas de vida.
Tabla 12. Ecuaciones alométricas utilizadas en los cálculos de biomasa aérea en las parcelas permanentes de Wiñaywayna.
Componente Zona de vida Ecuación alométrica Fuente
Arboles bmh-MS 𝐁𝐀 = Exp (−2,450 + (0,932
∗ (ln((D2) ∗ H ∗ ρ)) Alvarez et al. (2012)
Helechos bmh-MBS 𝐁𝐀 = Exp(−2.286 + 2.471lnD) Sierra et al. (2007)
Palmeras bmh-MBS 𝐁𝐀 = Exp(0.360 + 1.218 lnH) Sierra et al. (2007)
𝐓𝐌 =(𝐥𝐧(𝐍𝐨) – 𝐥𝐧(𝐍𝐨 − 𝐍𝐦)
∆𝐭
𝐓𝐑𝐞 =𝐓𝐌 + 𝐓𝐑
𝟐

52
2. Estimación de la cantidad de biomasa aérea total
Se estimó como la sumatoria de la biomasa arbórea de todos los arboles de la parcela,
Dónde:
AGB est: Biomasa arbórea sobre el suelo.
Área: Tamaño de la parcela en hectárea. (Honorio & Baker 2010)
3. Estimación de la cantidad de Carbono:
Según Pearson et al. 2005, esta aceptado asumir que el 50% del peso seco de biomasa,
es carbono. Para calcular los contenidos de carbono en bosques naturales, se estimó los
valores promedios de biomasa aérea (T ha-1) por tipo de bosque y se multiplicó este
valor por 0,5, y se obtuvo la cantidad de carbono por parcela en TC ha-1.
4. Prueba no paramétrica: U de Mann Whitney:
En estadística, es un método matemático que compara la diferencia significativa entre
las medianas de dos variables independientes, por lo que se basa en rangos, en lugar de
los parámetros de la muestra (media, varianza). Se emplea cuando los datos no siguen
una distribución normal, es decir en lugar del test de Student (Paramétrico). (UA, 2004
-2005)
Para comprobar la normalidad de datos se utilizó la prueba de Kolmogorow Smirnov,
que es una prueba no paramétrica que determina la bondad de ajuste de dos
distribuciones, verificando la normalidad de una distribución.
Para el cálculo de U de Mann Whitney se siguió los siguientes pasos:
Se ordenaron los datos en orden creciente
Se le asignaron un orden a cada dato
Se sumaron los rangos de cada uno de los grupos y se calcularon la suma de los
rangos de los datos de cada uno de los grupos (R1y R2)
Se calcularon los estadísticos U1 y U2 a partir de las siguientes formulas:
𝑩𝑨𝒕 =𝜮𝑩𝑨𝒆𝒔𝒕
𝑨𝒓𝒆𝒂
𝑈1 = 𝑛1 ∗ 𝑛2 +𝑛2(𝑛2 + 1)
2− 𝑅2 𝑈2 = 𝑛1 ∗ 𝑛2 +
𝑛1(𝑛1 + 1)
2− 𝑅1

53
Se obtuvo el Ucal y se escogió el valor más grande entre U1 y U2.
Se comprobó la significancia estadística del estadístico Ucal comparando con el
valor del estadístico Ucrit obtenido a partir de las tablas correspondientes.
Si U cal ≥ U crit (a 0.05 o inferior), se rechaza la hipótesis nula y se acepta
la hipótesis alterna indicando que las medianas son significativamente
diferentes).
Si U cal < U crit (a 0.05), se acepta la hipótesis nula y se rechaza la
hipótesis alterna indicando que las medianas son iguales.
(UA, 2004 -2005)
3.2.3.3. Análisis de composición floristica
Para determinar la composición en cada una de las parcelas, se trabajó con todos
aquellos arboles encontrados vivos, se ordenaron la base de datos por familias y
especies, que fueron revisados y actualizados con listados en la página web de TRÓPICOS
(http://www.tropicos.org/) y Forestplots Database www.foresplots.net.
3.2.3.4. Análisis de importancia ecológica
Para este análisis, se usaron los índices de valor de importancia por especie (IVI’s) y por
familia (IVIF), estos dos índices nos permiten comparar la importancia relativa de cada
especie y familia dentro del tipo de bosque analizado.
1. Índice de Valor de Importancia por especie (IVI´s)
Formulado por Curtis y Macintosh (1951), nos da la importancia ecológica relativa de
cada especie en una comunidad vegetal.
Está constituido por la suma de tres parámetros relativos; la abundancia relativa (Ar),
dominancia relativa (cobertura o área basal) (Dr) y frecuencia relativa (Fr), de la
siguiente manera:
𝐈𝐕𝐈´𝐬 =𝐃𝐫 + 𝐅𝐫 + 𝐀𝐫
𝟑

54
Abundancia:
Es un parámetro que permite conocer la abundancia absoluta (A) de una especie
(Mostacedo 2000) y viene a ser él número de individuos de una especie o familia (N),
presentes en un área determinada. La abundancia relativa (Ar), indica el porcentaje (%)
de participación que tiene cada una de las especies en una determinada área y se
expresa como la relación porcentual entre el número de individuos de una especie
determinada (N) entre el total de individuos en un área determinada (ΣN).
Frecuencia:
Es la probabilidad de hallar un atributo o más en una unidad de área muestreada
(Mateuci & Colma 1982), mide la distribución de las especies sobre el terreno dentro del
habitad.
La frecuencia relativa mide la relación que existe entre la frecuencia absoluta (F) de una
determinada especie, en relación del total de registros para todas las especies (ΣF). Se
expresa por el porcentaje del número de unidades muéstrales en que la especie (mi)
aparece en relación al número total de unidades del área muestreada (M).
Dominancia o área basal:
La dominancia absoluta está determinada por el área basal (AB) de una especie; donde
se considera la medida del diámetro a la altura del pecho (DAP), es decir, a 1.3 m del
suelo en cada individuo. La dominancia relativa constituye la relación del área basal de
una especie con respecto al área basal de todas las especies (Mateuci & Colma 1982).
𝐀 =𝐍𝐢
𝐚 𝐀𝐫 =
𝐍𝐢
𝚺𝐍𝐱𝟏𝟎𝟎
𝐅 =𝐦𝐢
𝐌∗ 𝟏𝟎𝟎; 𝐅𝐫 =
𝐅
𝚺𝐅𝐱𝟏𝟎𝟎
𝐀𝐁 = 𝛑 (𝐃𝐀𝐏
𝟐)
𝟐
; 𝐃𝐫 =𝐀𝐁𝐢
𝚺𝐀𝐁∗ 𝟏𝟎𝟎

55
2. Índice de Valor de Importancia por familia (IVI´F)
Este índice es propuesto por Mateuci & Colma (1982), con la finalidad de posicionar la
importancia de las familias de los arboles dentro de una comunidad vegetal (Mori &
Boom 1987).
Está constituido por la suma de tres parámetros; diversidad relativa (entendida como
riqueza) (DIVr), abundancia relativa (Ar) y la dominancia relativa (Dr) de todos los
individuos de una familia.
Diversidad relativa: Conocida como riqueza de especies, se define como el
número de especies pertenecientes a un determinado grupo (plantas, animales,
etc), se determina como la relación porcentual entre el número de especies de
una determinada familia (N° es) y el número total de especies encontradas (N°
total sp), en una unidad de área determinada.
3. Análisis de la diversidad florística
La diversidad florística para un área determinada, considera al número de especies
(riqueza) y el número de individuos (abundancia). La diversidad permite distinguir dos
comunidades con idéntica riqueza y composición, donde las especies se distinguen por
su abundancia relativa.
Para determinar la diversidad se usaron algunos índices que determinan la
heterogeneidad de una comunidad en bases a su riqueza y abundancia.
a) Diversidad alfa (α)
La diversidad alfa es la riqueza de especies de una comunidad particular a la que
consideramos homogénea. Para obtener parámetros completos de diversidad de
especies, se recomienda cuantificar el número de especies (riqueza) y su
𝐈𝐕𝐈´𝐅 =𝐃𝐈𝐕𝐫 + 𝐃𝐫 + 𝐀𝐫
𝟑
𝐃𝐈𝐕𝐫 =N° sp de una familia
N° total de sp∗ 100

56
representatividad. Para ello existen muchos índices para determinar la diversidad de
una comunidad en particular y cada índice está ligado a la información que se quiere
obtener (Magurran, 1988), para determinar el número de especies de una comunidad,
el índice usado se basa en la riqueza de especies, pero si se quiere medir la equidad se
puede utilizar los índices de abundancia entre todas las especies, o se puede conjuntar
ambos índices, la equidad se refiere a la igualdad en las abundancias de las diferentes
especies y se evalúa tomando en cuenta el número de especies dominantes en relación
con el número de especies poco o nada comunes (Magurran, 2004; Moreno et al., 2006).
La ventaja de los índices es que resumen la información en un solo valor y nos permite
hacer comparaciones rápidas y sujetas a comprobación estadística entre la diversidad y
el número de hábitat, o la diversidad de un mismo hábitat a través del tiempo.
Índice de diversidad de Shannon – Weiner (H’)
Es uno de los índices más usados para determinar la diversidad de plantas en un
determinado lugar (Mostacedo & Friedericksen 2000), este Índice combina la
información de la riqueza de especies y la equidad, en la diversidad o heterogeneidad
(Magurran 2004; Moreno et al., 2006), cuanto mayor sea del número de especies,
incrementa la diversidad, así como la uniformidad (Franco, 1989). Los valores que puede
tomar van de una diversidad baja muy próxima a uno y una diversidad alta, cercana a
cinco.
Este índice se calcula con al siguiente formula:
Dónde:
H’: Indice de Shannon – Wiener
Ln: Logaritmo natural
Pi: Abundancia proporcional de la especie i (ni/Nt)
La uniformidad de la distribución para la comunidad puede medirse comparando la
diversidad observada en ésta, con la diversidad máxima posible para una comunidad
hipotética con el mismo número de especies. Puede demostrarse cuando Pi = 1 /S para
todo pi, y alcanza la uniformidad máxima.
𝐇′ = 𝚺𝐩𝐢𝐥𝐧𝐩𝐢

57
Dónde:
H∫max: Valor máximo posible para la función Shannon - Wiener
S: Número de especies en la comunidad
La Equitatividad de Evenness, basada en la función Shannon –Wiener. Nos indica el
acercamiento a una escala de heterogeneidad relativa en un máximo valor, donde cada
especie de la unidad muestral es representativa por algún número de individuos. Éste
índice se ajusta de 0-1, donde 1 representa una máxima diversidad y 0 una baja
diversidad. (Franco, 1992).
Índice de dominancia de Simpson (D)
Es otro índice comúnmente usado para determinar la diversidad de una comunidad
vegetal, este índice es inverso al concepto de equidad de la comunidad ya que toma en
consideración las especies con mayor importancia ecológica sin considerar al resto de
las especies, y es menos sensible con la riqueza de especie (Krebs 1978, Magurran 1991,
Feinsinger 2003). Además, se basa en la teoría de probabilidades, en una comunidad
biología muy diversa, la probabilidad de que dos organismos tomados al azar sean de
especies similares debe de ser baja. Este índice no solo considera el número de especies
y el número total de individuos sino también la proporción del total que ocurre en cada
especie. La fórmula para el índice de Simpson es:
Dónde:
D= Índice de Simpson
ni= Número de individuos de la especie i en la comunidad 1.
N= Número total de individuos de la comunidad 1. (Moreno 2001)
𝑬 =𝑯′
𝑯∫ 𝒎𝒂𝒙
𝑯∫ 𝒎𝒂𝒙 = 𝐥𝐨𝐠𝟐 𝑺
𝐃 = ∑ [𝐧𝐢(𝐧𝐢 − 𝟏)
𝐍(𝐍 − 𝟏)]

58
La forma más fácil de interpretar el índice de Simpson es utilizando su reciproco el cual
se resta de uno el valor obtenido con el índice de Simpson de esta diferencia mientras
el valor obtenido se aproxime más a uno significará que existe mayor diversidad,
mientras sea más cercano a cero habrá menor diversidad.
b) Diversidad beta (β)
O diversidad entre habitas, es el grado de reemplazamiento de especies o cambio biótico
a través de gradientes ambientales (Whittaker 1972), su medición está basada en
proporciones (Magurran 1988), las que pueden determinarse en base a índices o
coeficientes de similitud, disimilitud o de distancia entre las muestras a partir de datos
cualitativos.
Los índices beta no tienen un valor máximo, entre más alto sea el valor del índice beta
menor será el número de especies compartidas entre las comunidades (Polo 2008).
Índice de similaridad de Morisita – Horn
Índice que calcula la similitud en base a datos cuantitativos y es muy sensible a la
abundancia de las especies (Magurran 1988, Baev & Penev 1995), por lo que Wolda
(1983) propuso una modificación para este índice.
Dónde: aN: Número total de individuos en el sitio A.
bN: Número total de individuos en el sitio B.
ani: Número de individuos de la i-esima especies en el sitio A.
bnj: Número de individuos de la i-esima especie en el sitio B.
𝐂𝐌𝐇 =𝟐𝚺(𝐚𝐧𝐢𝐱 𝐛𝐧𝐣)
(𝐝𝐚 + 𝐝𝐛)𝐚𝐍𝐱𝐛𝐍∗ 𝟏𝟎𝟎% ; 𝒅𝒂 =
𝜮𝒂𝒏𝒊𝟐
𝒂𝑵𝒂𝟐 ; 𝒅𝒃 =
𝜮𝒃𝒏𝒋𝟐
𝒃𝑵𝒃𝟐

59
CAPITULO IV
RESULTADOS Y DISCUSIONES
4.1. RESULTADOS
4.1.1. Dinámica poblacional (Tasas de mortalidad, reclutamiento y recambio)
Tabla 13. Tasas de mortalidad, reclutamiento y recambio en cada parcela permanente.
Parcela Vivos
(2013)
Vivos
(2016) Ti
em
po
Mu
ert
os
Mu
ert
os
/añ
o
Re
clu
tas
Re
clu
tas
/añ
o
TM
(%)
TR
(%)
TRe
(%)
SHM - 01 690 682 3 24 8 16 5 1.18 0.79 0.99
SHM - 02 589 604 3 21 7 36 12 1.21 2.05 1.63
Fuente: Anexo 7., TM(Tasa de Mortalidad), TR(Tasa de Reclutamiento), TRe(Tasa de
Recambio)
Figura 9. Individuos muertos y reclutas por parcela durante el periodo de evaluación.
Fuente: Tabla 13.
Se registraron entre el año 2013 y el 2016, 24 arboles muertos y 15 arboles nuevos o
reclutas en la parcela SHM – 01 y 21 individuos muertos y 36 arboles reclutas en la
parcela SHM – 02.
0
5
10
15
20
25
30
35
40
SHM - 01 SHM - 02
2421
16
36
Nu
me
ro d
e in
div
idu
os
Periodo censal 2013 - 2016
Arboles Muertos en 3 años
Arboles Reclutas en 3 años

60
Figura 10. Tasas de mortalidad, reclutamiento y renovación por parcela.
Fuente: Tabla 13.
Según la figura 10:
Se muestra que la parcela SHM - 01 registró mayor tasa de mortalidad de 1.18 %,
respecto a su tasa de reclutamiento que solo fue de 0.79 % y su tasa de recambio
que fue de 0.99 %. En la parcela SHM - 02 se registró mayor tasa de reclutamiento
de 2.05 % respecto a su tasa de mortalidad que fue de 1.21 %, su tasa de recambio
fue de 1.63 %. Se muestra que la parcela SHM – 02 registro los valores mas altos
que la parcela SHM - 01, quiere decir el bosque de Kantupata tuvo mas actividad
poblacional con mas reclutamiento y menos mortalidad, que el bosque de
intipunku que tuvo mas mortalidad y menos reclutamiento.
Para ser los primeros resultados de dinámica poblacional en los bosques
estudiados, se muestra que ambas parcelas tienen un desequilibrio poblacional,
esto se puede deber a que los bosques son jóvenes o bosques no maduros que
están en proceso de crecimiento y recuperación como es el caso del bosque de
Intipunku que en algún momento ha sufrido intervención antrópica.
1.18 1.21
0.79
2.05
0.99
1.63
0.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
2.25
2.50
SHM - 01 SHM - 02Tasa
s d
e m
ort
alid
ad (
TM),
tas
as d
e r
ecl
uta
mie
nto
(TR
)
y ta
sa d
e r
eca
mb
io(T
Re
)
Periodo censal 2013 - 2016
TM (%) TR (%) TRe(%)

61
Tabla 14. Especies reclutadas en la parcela SHM - 01 durante el periodo de evaluación.
FAMILIAS ESPECIES INDIVIDUOS
Cunoniaceae Weinmannia pubescens Kunth 1
Melastomataceae Miconia media (D. Don) Naudin 1
Melastomataceae Meriania tomentosa (Cogn.) Wurdack 1
Brunelliaceae Brunellia sp1. Ruiz & Pav. 1
Monimiaceae Mollinedia repanda Ruiz & Pav. 2
Moraceae Morus insignis Bureau 1
Phyllanthaceae Hieronyma oblonga (Tul.) Müll.Arg. 3
Primulaceae Myrsine latifolia (Ruiz & Pav.) Spreng. 2
Rubiaceae Cinchona pubescens Vahl 2
Sabiaceae Meliosma peytonii A.H. Gentry 1
Styracaceae Styrax pentlandianus J. Rémy 1
TOTAL 16
Fuente: Anexo 8
Tabla 15. Especies reclutadas en la parcela SHM - 02 durante el periodo de evaluación.
FAMILIAS ESPECIES INDIVIDUOS
Asteraceae Baccharis oblongifolia (Ruiz & Pav.) Pers. vel sp. aff.
1
Chloranthaceae Hedyosmum angustifolium (Ruiz & Pav.) Solms
4
Cunoniaceae Weinmannia crassifolia Ruiz & Pav. 14
Elaeocarpaceae Vallea stipularis L.f. 1
Ericaceae Cavendishia bracteata (Ruiz & Pav. ex J.St.Hil.) Hoerold
2
Escalloniaceae Escallonia myrtilloides L.f. 1
Melastomataceae Meriania vargasii Wurdack 1
Miconia indet 1 1
Primulaceae Myrsine coriacea (Sw.) R.Br. ex Roem. & Schult.
5
Rosaceae Hesperomeles ferruginea (Pers.) Benth. 2
Prunus integrifolia (C. Presl) Walp. 2
Rubiaceae Psychotria indet 1 2
TOTAL 36
Fuente: Anexo 8
Según las tablas 14 y 15, en la parcela SHM – 01 se registró 16 árboles reclutas, que
corresponden a 11 especies, donde la familia con mayor número de individuos
reclutados fue Phyllantaceae con 3 individuos de la especie Hieronyma oblonga (Tul.)
Müll.Arg., seguida de las familias Monimiaceae con Mollinedia repanda Ruiz & Pav.,
Primulaceae con Myrsine latifolia (Ruiz & Pav.) Spreng., y Rubiaceae con Cinchona
pubescens Vahl, con 2 individuos cada una, y el resto de las especies con 1 individuo. En
la parcela SHM - 02 se registró 36 árboles reclutados que corresponden a 12 especies,

62
donde la familia con mayor número de individuos reclutados fue Cunoniaceae con 14
individuos de la especie Weinmannia crassifolia Ruiz & Pav., seguida de la familia
Primulaceae con 5 individuos de la especie Myrsine coriacea (Sw.) R.Br. ex Roem. &
Schult., y Chloranthaceae con 4 individuos de la especie Hedyosmum angustifolium (Ruiz
& Pav.) Solms., y el resto presentó valores por debajo.
1. Tipos de mortalidad por parcela
Figura 11. Tipos de Mortalidad en la parcela SHM - 01.
Fuente: Anexo 9
De la figura 11, del total de árboles muertos en la parcela SHM – 01, el 50 % murieron
quebrados o rotos, el 33.3% murieron en pie y el otro 16.67% murieron desenraizados.
Figura 12. Tipos de Mortalidad en la parcela SHM - 02.
Fuente: Anexo 9
De la figura 12, del total de árboles muertos en la parcela SHM – 02, el 14.29 % murieron
quebrados, el 42.86 % desenraizados, el 42.86 % murieron en pie.
50.00
16.67
33.33
0
10
20
30
40
50
60
Quebrado Desenraizado En pie
po
rce
nta
je d
e m
ort
alid
ad d
e
ind
ivid
uo
s (%
)
14.29
42.86 42.86
0
5
10
15
20
25
30
35
40
45
50
Quebrado Desenraizado En pie
Po
rcen
jate
de
mo
rtal
idad
de
In
div
idu
os
(%)

63
4.1.2. Biomasa aérea y Carbono
Tabla 16. Biomasa aérea y carbono almacenadao en cada una de las parcelas
permanentes
PARCELA Individuos Biomasa
(2013) (T ha-1)
Biomasa (2016) (T ha-1)
Biomasa promedio
(T ha-1)
Desv. Est.
Intervalo de confianza
(95%) 2013 2016
SHM – 01
(2563m) 690 682 144.55 147.41 145.98 1.43 2.80
SHM – 02
(3200m) 589 604 99.29 102.82 101.06 1.77 3.46
Promedio 123.52 - -
Desviación estándar 22.46 - -
Intervalo de confianza, 95% 31.13 - -
PARCELA Individuos Carbono
(2013) Carbono (2016)
Carbono promedio
(T/ha)
Desv. Est.
Intervalo de confianza
(95%) 2013 2016
SHM – 01 (2653m)
690 682 72.28 73.70 72.99 1.01 1.40
SHM – 02 (3200m)
589 604 49.65 51.41 50.53 1.25 1.73
Promedio 61.76 - -
Desviación estándar 11.23 - -
Intervalo de confianza, 95% 15.56 - -
Fuente: Anexo 10
Figura 13. Promedio de biomasa aérea y carbono acumulado en cada una de las parcelas permanentes evaluadas.
Fuente: Tabla 16
0
20
40
60
80
100
120
140
160
SHM 01 SHM 02
Bio
mas
a ae
rea
y C
arb
on
o (
T h
a-1)
Biomasa promedio (T ha-1)
Carbono promedio (T ha-1)
14
5.9
8 ±
2.8
0
72
.99
± 1
.40
10
1.0
6 ±
3.4
6
50
.53
± 1
.73

64
Según la tabla 16 y la figura 13, en la parcela SHM – 01 del bosque de Intipunku, se
registraron un total 682 individuos con DAP ≥ 10, de los que se obtuvo la biomasa aerea
total de 145.98 ± 2.80 T ha-1 y 72.99 ± 1.40 TCha-1 de carbono, en la parcela SHM - 02
del bosque de Kantupata se registraron 604 individuos con DAP ≥ 10, lo que representó
101.06 ± 3.46 T ha-1 de biomasa aérea total y 50.53 ± 1.73 TCha-1 de carbono
almacenado.
Figura 14. Biomasa aérea y carbono almacenado por gradiente altitudinal.
Fuente: Tabla 16
Según la figura 14, se observa una relación negativa de gradiente y biomasa, quiere decir
que a mayor altitud menor cantidad de biomasa aérea, en la parcela SHM – 01 de
Intipunku que se encuentra a 2653 m se registró mayor Biomasa aérea y carbono
acumulado que la parcela SHM – 02 de Kantupata que se encuentra a mayor altitud
(3200 m), lo cual se debe al tipo de bosque, a la dinámica que ocurre en ella, a la altitud
y a la composición florística, pues influyen en el creciemiento diametrico, altura arbórea
y densidad básica de madera.
Este resultado fue corroborado con el análisis de significancia mediante la prueba no
paramétrica U de Mann Whitney previo análisis de normalidad de datos mediante la
bondad de ajuste a la normalidad de Kolmogorow-Smirnov, donde P valor resulto ser <
0.05, que demostró que los datos no presentan una distribución normal (Anexo 12).
Según el estadístico no paramétrico el P valor = 0.001 al 95% resulto ser menor que 0.05
145.98
101.06
72.99
50.53
40
50
60
70
80
90
100
110
120
130
140
150
SHM01 SHM02
2653 m 3200 m
Bio
mas
a ae
rea
y C
arb
on
o a
lmac
en
ado
(T h
a-1
)
Biomasa Carbono

65
(P < 0.05), por consiguiente, se aceptó la hipótesis alterna que indica que existe una
marcada diferencia significativa en las medianas de Biomasa aérea almacenada entre
ambas parcelas (Anexo 13).
1. Biomasa aérea y carbono en cada censo (2013 y 2016)
Figura 15. Promedio de biomasa aérea acumulada en cada censo de las parcelas permanentes.
Fuente: Anexo 11
Biomasa aérea (Figura 15): La parcela SHM - 01 de Intipunku, para el año 2013 con
690 individuos llegó a acumular 144.55 ± 13.83 T ha-1 y para el año 2016 con 682
individuos acumuló 147.41 ± 14.33 T ha-1, en la parcela SHM – 02 de Kantupata,
para el año 2013 con 589 individuos acumuló 99.29 ± 10.70 T ha-1 y para el año
2016 con 604 individuos, acumuló 102.82 ± 11.19 T ha-1.
0
20
40
60
80
100
120
140
160
SHM - 01 SHM - 02
145.98 ± 2.80 T ha-1 101.06 ± 3.46 T ha-1
CU
AN
TID
AD
DE
BIO
MA
SA (
T h
a-1
)
BIOMASA 2013 BIOMASA 2016
14
4.5
5 ±
13
.83
14
7.4
1 ±
14
.33
99
.29
± 1
0.7
0
10
2.8
2 ±
11
.17

66
Figura 16. Promedio de carbono acumulado en cada censo de las parcelas permanentes.
Fuente: Anexo 11
Carbono almacenado (Figura 16): La parcela SHM – 01 para el 2013, almacenó
72.28 ± 6.91 TCha-1, y para el 2016, almacenó 73.70 ± 7.17 T Cha-1, la parcela SHM
– 02 para el 2013 almacenó 49.65 ± 5.35 T Cha-1, y para el 2016 almacenó 51.41 ±
5.59 T Cha-1.
2. Distribución de la biomasa aérea y carbono
Figura 17. Biomasa aérea y carbono por especies en la parcela SHM - 01
Fuente: Anexo 14
0
10
20
30
40
50
60
70
80
90
SHM - 01 SHM - 02
72.99 ± 1.40 50.53 ± 1.73
CA
NTI
DA
D D
E C
AR
BO
NO
(T/
ha)
"C" 2013 (T/ha) "C" 2016 (T/ha)
19.55
19.26
16.23
15.82
13.77
13.13
9.09
6.04
5.37
2.91
9.77
9.63
8.11
7.91
6.89
6.56
4.55
3.02
2.69
1.45
0 2 4 6 8 10 12 14 16 18 20 22 24
Gordonia fruticosa
Aniba coto vel sp. aff.
Hieronyma oblonga
Myrcia fallax vel sp. aff.
Clethra castaneifolia
Meliosma peytonii
Myrsine latifolia
Mollinedia repanda
Meriania tomentosa
Alchornea grandis
Promedio de Carbono (T ha-1) Promedio de biomasa (T ha-1)
72
.28
± 6
.91
73
.70
± 7
.16
49
.65
± 5
.35
51
.41
± 5
.59

67
Según la figura 17, se observa que dentro de la parcela SHM - 01, las especies Gordonia
fruticosa (19.55 T ha-1 y 9.77 TCha-1) y Aniba coto vel sp. Aff. (19.26 T ha-1 y 9.63 T Cha-
1), son los que aportaron mayor biomasa aerea y carbono acumulado, seguido de
Hieronyma oblonga (16.23 T ha-1 y 8.11 T Cha-1) y Myrcia fallax vel sp. Aff. (15. 82 T ha-
1 y 7.91 T Cha-1) y Clethra castaneifolia (13.77 T ha-1 y 6.89 T Cha-1) con Meliosma
peytonii (13.13 T ha-1 y 6.56 T C ha-1), estas especies son las que acumularon en casi el
80% de biomasa aérea y carbono por hectárea en la parcela SHM – 01.
Figura 18. Biomasa aérea y Carbono por especies de la parcela SHM - 02
Fuente: Anexo 15
Según la figura 18 se observa que dentro de la parcela SHM - 02, la especie Weinmannia
crassifolia, tuvo el mayor aporte de biomasa aérea y carbono almacenado de 32.78 T
ha-1 y 16.39 T C ha-1 respectivamente, seguida de Escallonia myrtilloides con 13.51 T ha-
1 de biomasa y 6.75 T C ha-1, Ilex andicola con 12.24 T ha-1 de biomasa y 6.12 T C ha-1,
Hedyosmun angustifolium con 8.24 T ha-1 de biomasa y 4.47 T C ha-1, Weinmannia
banggi con 6.12 T ha-1 de biomasa y 3.06 TC ha-1, Hesperomeles ferruginea con 5.01 T
ha-1 de biomasa y 2.50 TC ha-1, estas especies son las que acumularon mayor biomasa
aérea y carbono por hectárea y representan casi el 89 % del total en la parcela SHM –
02.
32.78
13.51
12.24
8.94
6.12
5.01
4.45
2.70
2.07
1.99
16.39
6.75
6.12
4.47
3.06
2.50
2.22
1.35
1.04
0.99
0 5 10 15 20 25 30 35
Weinmannia crassifolia
Escallonia myrtilloides
Ilex andicola
Hedyosmum angustifolium
Weinmannia bangii
Hesperomeles ferruginea
Clethra ferruginea
Myrsine coriacea
Schefflera herzogii
Clusia trochiformis
Promedio de Carbono (T ha-1) Promedio de biomasa (T ha-1)

68
3. Dinámica de biomasa aérea y carbono almacenado
Tabla 17. Cambio de biomasa aérea y carbono en el periodo intercensal (2013 – 2016).
PARCELA Biomasa (Tha-1) (2013)
Biomasa (Tha-1) (2016)
Tiempo Biomasa cambio (Tha-1)
Carbono cambio (Tha-1)
SHM - 01 144.55 147.41 3 2.85 1.43
SHM - 02 99.29 102.82 3 3.53 1.77
Figura 19. Variación de Biomasa aerea y carbono en el periodo intercensal de
evaluación (2013 – 2016)
Fuente: Tabla 17
En la parcela SHM – 01 de Intipunku, se registró una variación de incremento de 2.85 T
ha-1 de biomasa aerea y 1.43 TCha-1 y en la parcela SHM - 02 de Kantupata se registró
una variación de incremento de 3.53 Tha-1 de biomasa aerea y 1.77 TCha-1 en los 3 años
del periodo intercensal (2013 al 2016), siendo la parcela SHM – 02 la que presentó mayor
variación.
144.55147.41
72.28 73.70
99.29102.82
49.65 51.41
40.0
50.0
60.0
70.0
80.0
90.0
100.0
110.0
120.0
130.0
140.0
150.0
2012 2013 2014 2015 2016 2017
Var
iaci
on
de
Bio
mas
a ae
rea
y c
arb
on
o a
lmac
end
o
(T h
a-1)
SHM - 01 SHM - 02
2.85
1.43
3.53
1.77
B
I
O
M
A
S
A
C
A
R
B
O
N
O
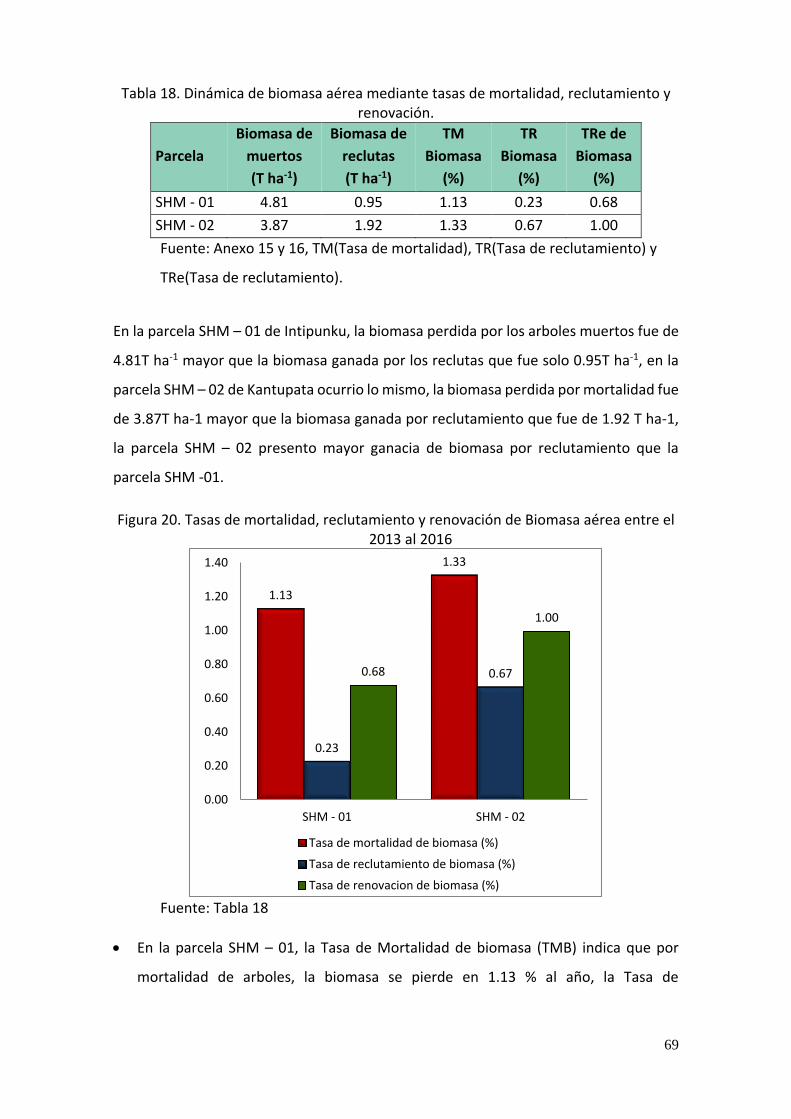
69
Tabla 18. Dinámica de biomasa aérea mediante tasas de mortalidad, reclutamiento y renovación.
Parcela
Biomasa de
muertos
(T ha-1)
Biomasa de
reclutas
(T ha-1)
TM
Biomasa
(%)
TR
Biomasa
(%)
TRe de
Biomasa
(%)
SHM - 01 4.81 0.95 1.13 0.23 0.68
SHM - 02 3.87 1.92 1.33 0.67 1.00
Fuente: Anexo 15 y 16, TM(Tasa de mortalidad), TR(Tasa de reclutamiento) y
TRe(Tasa de reclutamiento).
En la parcela SHM – 01 de Intipunku, la biomasa perdida por los arboles muertos fue de
4.81T ha-1 mayor que la biomasa ganada por los reclutas que fue solo 0.95T ha-1, en la
parcela SHM – 02 de Kantupata ocurrio lo mismo, la biomasa perdida por mortalidad fue
de 3.87T ha-1 mayor que la biomasa ganada por reclutamiento que fue de 1.92 T ha-1,
la parcela SHM – 02 presento mayor ganacia de biomasa por reclutamiento que la
parcela SHM -01.
Figura 20. Tasas de mortalidad, reclutamiento y renovación de Biomasa aérea entre el 2013 al 2016
Fuente: Tabla 18
En la parcela SHM – 01, la Tasa de Mortalidad de biomasa (TMB) indica que por
mortalidad de arboles, la biomasa se pierde en 1.13 % al año, la Tasa de
1.13
1.33
0.23
0.670.68
1.00
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
SHM - 01 SHM - 02
Tasa de mortalidad de biomasa (%)
Tasa de reclutamiento de biomasa (%)
Tasa de renovacion de biomasa (%)

70
Reclutamiento de Biomasa indica que por reclutamiento de arboles se gana un
0.23% al año, la biomasa en esta parcela se renueva en 0.68% anual.
En la parcela SHM -02, se pierde la biomasa por mortalidad de arboles en 1.33% anul
y se gana por reclutamiento de arboles en 0.67% y se renueva en 1 % al año.
Entre ambas parcelas evaludas, la segunda (SHM - 02) presenta mayor dinámica de
biomasa aérea.
4. Productividad primaria neta (PPN)
La productividad neta resulta de las ganancias de biomasa que involucra la biomasa por
reclutamiento y la biomasa por crecimiento menos la biomasa por mortalidad, donde la
PPN se obtiene al realizar los descuentos correspondientes, lo que indica la ganancia
concreta que la parcela permanente almacena.
Tabla 19. Productividad primaria neta de las parcelas evaluadas
Parcelas
Ganancia de biomasa Pérdida de
biomasa PPN
Por reclutamiento
(T ha-1)
Por crecimiento
(T ha-1)
Bruta Por mortalidad
(T ha-1año-1) (%) (T ha-1año-1) (%) (T ha-1año-1) (%)
SHM - 01 0.32 2.24 2.55 1.7 -1.60 1 0.95 0.7
SHM - 02 0.64 1.83 2.47 2.5 -1.29 1.3 1.18 1.2
Fuente: Anexo 17
Figura 21. Pérdida, ganancia de biomasa aérea y productividad primaria neta.
Fuente: Tabla 19
1.7
2.5
-1.1-1.3
0.7
1.2
-2
-1
0
1
2
3
SHM - 01 SHM - 02
Ganancia Bruta de biomasa (%) Biomasa perdida por mortalidad(%)
Productividad Primaria Neta (%)

71
La parcela SHM – 01, tuvo una pérdida de biomasa por mortalidad de 1.1% anual y una
ganancia o productividad bruta (por crecimiento mas reclutamiento) de 1.7% anual, que
resulta en una productividad primaria neta (PPN) de 0.7%.
La parcela SHM – 02, tuvo una pérdida de biomasa por mortalidad de 1.2% al año, una
productividad bruta por crecimiento mas reclutamiento de 2.4% al año y una
productividad primaria neta (PPN) de 1.2% al año, siendo esta parcela la que presentó
mayor PPN.
4.1.3. Analisis poblacional
1. Composición florística
Tabla 20. Número de Familias, géneros, especies e individuos por parcela evaluada.
ULTIMO REGISTRO (2016) INDIVIDUOS
PARCELA Familias Géneros Especies 2013 2016
SHM – 01 (INTIPUNKU) 26 36 45 690 682
SHM – 02 (KANTUPATA) 22 28 31 589 604
COMPARTIDOS ENTRE SI 13 15 7 - -
TOTAL 35 49 69 1279 1286
El registro del numero de individuos disminuyó en la parcela SHM -01 y aumentó en la
parcela SHN – 02 para el censo del 2016, pero el registro del numero de familias, generos
y especies fue la misma en ambos censos (2013 y 2016), por lo que se considero la
composición florística del ultimo censo.
En la parcela SHM - 01 de Intipunku se registró 26 familias con 36 generos y 45 especies
de árboles y en la parcela SHM – 02 de Kantupata se resgistró 22 familias con 28 generos
y 31 especies de árboles, se observa que la composición floristica en ambas parcelas es
muy diferente porque entre ellas solo comparten 7 especies, esto se debe a la diferencia
de gradiente atitudinal de 653m que existe entre ellas y al tipo de bosque.

72
Tabla 21. Especies registradas en la parcela SHM - 01. Familias Especies AB m2/ha
Adoxaceae Viburnum hallii (Oerst.) Killip & A.C. Sm. 0.068
Araliaceae
Oreopanax capitatus (Jacq.) Decne. & Planch. 0.013
Schefflera herzogii Harms 0.130
Arecaceae Ceroxylon pityrophyllum (Mart.) Mart. ex H.Wendl. 0.018
Brunelliaceae Brunellia sp1. Ruiz & Pav. 0.012
Chloranthaceae Hedyosmum angustifolium (Ruiz & Pav.) Solms 0.068
Clethraceae Clethra castaneifolia Meisn. 2.093
Clusiaceae Clusia trochiformis Vesqne 0.363
Cunoniaceae
Weinmannia auriculata D.Don 0.341
Weinmannia pubescens Kunth 0.035
Cyatheaceae
Cyathea conjugata (Spruce ex Hook.) Domin vel sp. aff. 0.340
Cyathea delgadii Sternb. vel sp. aff. 0.050
Cyathea indet1 0.088
Cyathea indet2 0.020
Cyathea indet3 0.048
Cyathea indet4 0.026
Elaeocarpaceae Vallea stipularis L.f. 0.032
Euphorbiaceae
Alchornea grandis Benth. 0.727
Sapium glandulosum (L.) Morong 0.015
Tetrorchidium rubrivenium Poepp. 0.158
Lauraceae
Aniba coto (Rusby) Kosterm. vel sp. aff. 2.323
Nectandra reticulata Mez vel sp. aff. 0.026
Melastomataceae
Meriania tomentosa (Cogn.) Wurdack 1.042
Meriania vargasii Wurdack 0.068
Miconia media (D. Don) Naudin 0.202
Miconia saltuensis Wurdack vel sp. aff. 0.278
Miconia theaezans (Bonpl.) Cogn. vel sp. aff. 0.173
Monimiaceae Mollinedia repanda Ruiz & Pav. 0.987
Moraceae Morus insignis Bureau 0.146
Myrtaceae
Myrcia fallax (Rich.) DC. vel sp. aff. 1.813
Myrcianthes rhopaloides (Kunth) McVaugh 0.142
Pentaphylacaceae Ternstroemia jelskii (Szyszył.) Melch. 0.311
Phyllanthaceae Hieronyma oblonga (Tul.) Müll.Arg. 2.601
Primulaceae
Myrsine coriacea (Sw.) R.Br. ex Roem. & Schult. 0.050
Myrsine latifolia (Ruiz & Pav.) Spreng. 1.278
Rubiaceae
Cinchona pubescens Vahl 0.565
Guettarda crispiflora Vahl 0.022
Palicourea sp1 0.019
Rudgea ciliata (Ruiz & Pav.) Spreng. 0.079
Sabiaceae Meliosma peytonii A.H. Gentry 2.538
Staphyleaceae Staphylea occidentalis Sw. 0.011
Styracaceae Styrax pentlandianus J. Rémy 0.228
Symplocaceae Symplocos melanochroa Sleumer 0.151
Theaceae Gordonia fruticosa (Schrad.) H.Keng 3.158
Urticaceae Cecropia tacuna C.C.Berg & P.Franco 0.028
TOTAL 22.85 m2/ha
Fuente: Anexo 18.

73
En la parcela SHM – 01 se registró 45 especies de árboles con DAP ≥ 10 en 682 individuos
que presenta un área basal total de 22.85m2/ha, de los cuales, las especies con mayor
Área Basal son; Gordonia fruticosa (Schrad.) H.Keng con 3.158 m2/ha, seguida de
Hieronyma oblonga (Tul.) Müll.Arg. con 2.601 m2/ha y Meliosma peytonii A.H. Gentry,
con 2.538 m2/ha. (Tabla 21).
Tabla 22. Especies registradas en la parcela SHM - 02.
Familias Especies AB m2/ha
Aquifoliaceae Ilex andicola Loes. 2.150
Araliaceae
Oreopanax cuspidatus Harms 0.019
Schefflera herzogii Harms 0.412
Asteraceae Baccharis oblongifolia (Ruiz & Pav.) Pers. vel sp. aff. 0.012
Berberidaceae Berberis commutata Eichler 0.062
Betulaceae Alnus acuminata Kunth 0.249
Chloranthaceae Hedyosmum angustifolium (Ruiz & Pav.) Solms 1.955
Clethraceae Clethra ferruginea (Ruiz & Pav.) Link ex Spreng. 0.850
Clusiaceae Clusia trochiformis Vesqne 0.237
Cunoniaceae
Weinmannia bangii Rusby 0.999
Weinmannia crassifolia Ruiz & Pav. 5.805
Cyatheaceae Cyathea indet 0.024
Elaeocarpaceae Vallea stipularis L.f. 0.146
Ericaceae
Cavendishia bracteata (Ruiz & Pav. ex J.St.Hil.) Hoerold 0.051
Gaultheria reticulata Kunth 0.009
Escalloniaceae Escallonia myrtilloides L.f. 1.935
Lauraceae
Nectandra indet 1 0.014
Nectandra indet 2 0.117
Ocotea indet 1 0.022
Melastomataceae
Meriania vargasii Wurdack 0.296
Miconia indet 1 0.124
Miconia indet 2 0.064
Primulaceae Myrsine coriacea (Sw.) R.Br. ex Roem. & Schult. 0.584
Proteaceae Roupala indet 0.036
Rosaceae
Hesperomeles ferruginea (Pers.) Benth. 0.720
Prunus integrifolia (C. Presl) Walp. 0.215
Rubiaceae Psychotria indet 1 0.205
Sabiaceae Meliosma peytonii A.H. Gentry 0.009
Solanaceae
Cestrum conglomeratum Ruiz & Pav. 0.023
Solanum maturecalvans Bitter vel sp. aff. 0.008
Symplocaceae Symplocos quitensis subsp. Boliviensis Brand 0.105
TOTAL 17.46 m2/ha
Fuente: Anexo 19
74
La parcela SHM - 02, registró 31 especies de árboles en 604 indivudos con DAP ≥ 10 y
presentan un área basal total de 17.46m2/ha, de los cuales, las especies con mayor Área
Basal fueron Weinmannia crassifolia Ruiz & Pav., con 5.805 m2/ha, seguida de Ilex
andicola Loes., con 2.150 m2/ha y Hedyosmum angustifolium (Ruiz & Pav.) Solms con
1.955 m2/ha.
Tabla 23. Número de géneros, especies e individuos por Familias en cada parcela para el 2016.
SHM-01 SHM-02
N° Familia Gen. Esp. Indiv. N° Familia Gen. Esp. Indiv.
1 Adoxaceae 1 1 3 1 Aquifoliaceae 1 1 76
2 Araliaceae 2 2 3 2 Araliaceae 2 2 11
3 Arecaceae 1 1 2 3 Asteraceae 1 1 1
4 Brunelliaceae 1 1 1 4 Berberidaceae 1 1 3
5 Chloranthaceae 1 1 6 5 Betulaceae 1 1 1
6 Clethraceae 1 1 22 6 Chloranthaceae 1 1 85
7 Clusiaceae 1 1 6 7 Clethraceae 1 1 23
8 Cunoniaceae 1 1 10 8 Clusiaceae 1 1 4
90 Cyatheaceae 1 6 37 9 Cunoniaceae 1 2 180
10 Elaeocarpaceae 1 1 2 10 Cyatheaceae 1 1 1
11 Euphorbiaceae 3 3 18 11 Elaeocarpaceae 1 1 9
12 Lauraceae 2 2 40 12 Ericaceae 2 2 6
13 Melastomataceae 2 5 73 13 Escalloniaceae 1 1 58
14 Monimiaceae 1 1 47 14 Lauraceae 2 3 7
15 Moraceae 1 1 9 15 Melastomataceae 2 3 24
16 Myrtaceae 2 2 53 16 Primulaceae 1 1 46
17 Pentaphylacaceae 1 1 12 17 Proteaceae 1 1 2
18 Phyllanthaceae 1 1 73 18 Rosaceae 2 2 41
19 Primulaceae 1 1 42 19 Rubiaceae 1 1 17
20 Rubiaceae 4 4 40 20 Sabiaceae 1 1 1
21 Sabiaceae 1 1 104 21 Solanaceae 2 2 3
22 Staphyleaceae 1 1 1 22 Symplocaceae 1 1 5
23 Styracaceae 1 1 13
24 Symplocaceae 1 1 8
25 Theaceae 1 1 55
26 Urticaceae 1 1 2
Las familias que presentaron mayor número de especies en la parcela SHM - 01 fue,
Cyatheaceae con 6 especies, Melastomataceae con 5 especies, Rubiaceae con 4
especies y Euphorbiaceae con 3 especies y en la parcela SHM – 02 fueron Lauraceae con
y Melastomataceae con 3 especies cada una y el resto presentaron entre 1 o 2 especies
por familia.

75
Figura 22. Familias con mayor número de individuos en la parcela SHM - 01, de Intipunku.
Fuente: Tabla 23.
Figura 23. Familias con mayor número de individuos en la parcela SHM - 02, de Kantupata.
Fuente: Tabla 23.
0 20 40 60 80 100 120
Sabiaceae
Melastomatace…
Phyllanthaceae
Theaceae
Myrtaceae
Monimiaceae
Primulaceae
Lauraceae
Rubiaceae
Cyatheaceae
Clethraceae
Euphorbiaceae
Styracaceae
Pentaphylacace…
Cunoniaceae
104
73
73
55
53
47
42
40
40
37
22
18
13
12
10
N° de Individuos
0 50 100 150 200
Cunoniaceae
Chloranthaceae
Aquifoliaceae
Escalloniaceae
Primulaceae
Rosaceae
Melastomataceae
Clethraceae
Rubiaceae
Araliaceae
Elaeocarpaceae
Lauraceae
Ericaceae
Symplocaceae
Clusiaceae
180
85
76
58
46
41
24
23
17
11
9
7
6
5
4
N° de Individuos

76
La figura 22 indica que las familia más abundantes en la parcela SHM – 01 de Intipunku
por presentar un alto número de individuos fueron, Sabiaceae con 104 individuos,
seguida de Melastomataceae y Phyllantaceae con 73 individuos cada una; Theaceae
con 55 individuos, Myrtaceae con 53 individuos, Monimiaceae con 47 individuos y el
resto de las familias presentaron valores por debajo de estas, esto indica que en el
bosque de Intipunku las familias más dominantes son Sabiaceae, Melastomataceae y
Phyllantaceae.
La figura 23 muestra la distribución de individuos por familia en la parcela SHM - 02 de
Kantupata, donde la familia Cunoniaceae fue la Familia más abundante por presentar
180 individuos por ha, seguida de Chlorantaceae con 85 individuos y Aquifoliaceae con
76 individuos, Escalloniaceae con 58 individuos, Primulaceae con 46 individuos,
Rosaceae con 41 individuos y el resto de las familias presentan valores por debajo de
estas familias.
2. Importancia ecológica (IVI’s e IVIF)
Tabla 24. Las 15 especies más importantes en la parcela SHM – 01 (Intipunku).
Indice de Valor de Importancia por especies en SHM - 01 de Intipunku
Especies Abu.
Relativa
Frec.
Relativa
Dom.
Relativa IVI's al 300%
Meliosma peytonii 15.25 7.14 11.11 33.50
Hieronyma oblonga 10.70 7.14 11.38 29.23
Gordonia fruticosa 8.06 6.55 13.82 28.43
Myrcia fallax 7.18 6.55 7.93 21.67
Aniba coto 5.57 5.36 10.16 21.09
Mollinedia repanda 6.89 6.55 4.32 17.76
Myrsine latifolia 5.87 5.65 5.59 17.11
Clethra castaneifolia 3.23 4.17 9.16 16.55
Meriania tomentosa 4.55 4.46 4.56 13.57
Cinchona pubescens 4.69 4.17 2.47 11.33
Alchornea grandis 2.20 3.57 3.18 8.95
Cyathea conjugata 3.23 2.98 1.49 7.69
Miconia saltuensis 2.05 3.27 1.22 6.54
Miconia media 2.35 3.27 0.88 6.50
Ternstroemia jelskii 1.76 2.98 1.36 6.10
Fuente: Anexo 20.

77
Figura 24. Representación gráfica de las especies ecológicamente más importantes en la parcela SHM – 01 del bosque de Intipunku.
Fuente: Tabla 24. En la parcela SHM - 01 del bosque de Intipunku, las especies ecológicamente mas
importantes son Meliosma peytonii con el 33.50 % de IVI’s, Hieronyma oblonga con el
29.23 % de IVI’s, Gordonia fruticosa con el 28.43 % de IVI’s, Myrcia fallax con el 21.67
%, Aniba coto con el 21.09 %., y el resto presenta valores por debajo de estas especies.
En cuanto a la dominancia, frecuencia y abundancia, las especies Meliosma peytonii fue
la mas abundante, la especie Gordonia fruticosa fue la mas dominante y las especies
Meliosma peytonii y Hieronyma oblonga se presentaron con mayor frecuencia.
Tabla 25. Familias con mayor valor de importancia ecológica en el bosque de Intipunku.
Indice de Valor de Importancia por Familia en SHM – 01 de Intipunku
Familias Abu.
Relativa Dom.
Relativa Div.
Relativa IVIF al 300%
Melastomataceae 10.70 7.72 10.87 29.29
Sabiaceae 15.25 11.11 2.17 28.53
Phyllanthaceae 10.70 11.38 2.17 24.26
Theaceae 8.06 13.82 2.17 24.06
Cyatheaceae 5.43 2.51 13.04 20.97
Myrtaceae 7.77 8.56 4.35 20.68
Lauraceae 5.87 10.28 4.35 20.49
Rubiaceae 5.87 3.00 8.70 17.56
Primulaceae 6.16 5.81 4.35 16.32
Clethraceae 3.23 9.16 2.17 14.56
TOTAL 216.71%
Fuente: Anexo 20
15.25
33.50
7.14
29.23
13.82
28.43
0
5
10
15
20
25
30
35
Abu. Relativa Frec. Relativa Dom. Relativa IVI's al 300%
Var
iab
les
po
bla
cio
nal
es
(%)
Meliosma peytoniiHieronyma oblongaGordonia fruticosaMyrcia fallaxAniba cotoMollinedia repandaMyrsine latifoliaClethra castaneifoliaMeriania tomentosaCinchona pubescensAlchornea grandisCyathea conjugataMiconia saltuensisMiconia mediaTernstroemia jelskii

78
Figura 25. Representación gráfica de las Familias más importantes ecológicamente en el Bosque de Intipunku.
Fuente: Tabla 25
En la parcela SHM - 01, las familias que presentaron valores de IVIF más altos son:
Melastomataceae con 29.29 %, Sabiaceae con 28.53 % y Phyllantaceae con 24.36 %,
Theaceae con 24.06 %.
Tabla 26. Las 15 especies más importantes en la parcela SHM – 02 (Kantupata).
Indice de Valor de Importancia por especies en SHM – 02 de Kantupata
Especies Abu.
Relativa Frec.
Relativa Dom.
Relativa IVI's al 300%
Weinmannia crassifolia 26.82 10.75 33.25 70.82
Ilex andicola 12.58 10.75 12.32 35.65
Hedyosmum angustifolium 14.07 9.81 11.20 35.09
Escallonia myrtilloides 9.60 8.41 11.08 29.10
Myrsine coriacea 7.62 9.81 3.35 20.78
Hesperomeles ferruginea 4.64 6.54 4.13 15.30
Clethra ferruginea 3.81 4.67 4.87 13.35
Weinmannia bangii 2.98 3.74 5.73 12.44
Prunus integrifolia 2.15 5.14 1.23 8.53
Psychotria indet1 2.81 4.21 1.17 8.19
Schefflera herzogii 1.66 2.80 2.36 6.82
Meriania vargasii 1.82 2.80 1.69 6.32
Miconia indet1 1.49 3.74 0.71 5.94
Vallea stipularis 1.49 1.87 0.83 4.19
Symplocos quitensis subsp. boliviensis 0.83 2.34 0.60 3.76
TOTAL 276.27 %
Fuente: Anexo 21.
0 5 10 15 20 25 30
Melastomatac…
Sabiaceae
Phyllanthaceae
Theaceae
Cyatheaceae
Myrtaceae
Lauraceae
Rubiaceae
Primulaceae
Clethraceae
29.29
28.53
24.26
24.06
20.97
20.68
20.49
17.56
16.32
14.56IVIF al 300%
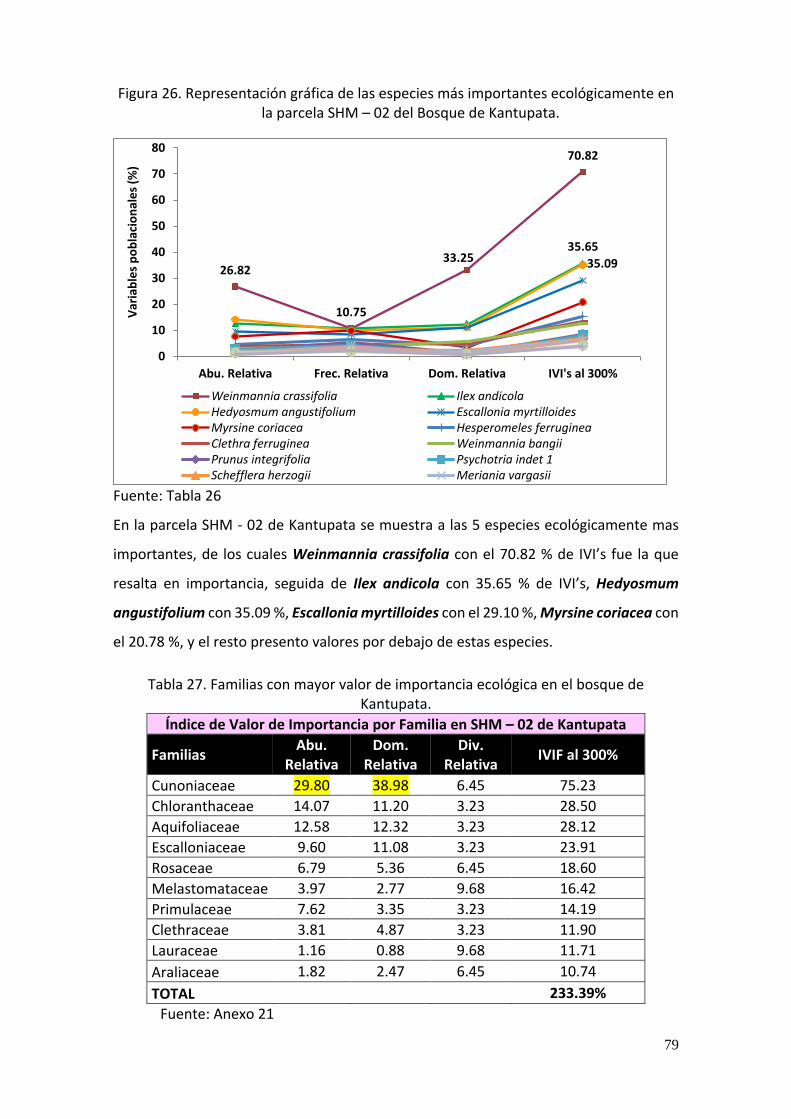
79
Figura 26. Representación gráfica de las especies más importantes ecológicamente en la parcela SHM – 02 del Bosque de Kantupata.
Fuente: Tabla 26
En la parcela SHM - 02 de Kantupata se muestra a las 5 especies ecológicamente mas
importantes, de los cuales Weinmannia crassifolia con el 70.82 % de IVI’s fue la que
resalta en importancia, seguida de Ilex andicola con 35.65 % de IVI’s, Hedyosmum
angustifolium con 35.09 %, Escallonia myrtilloides con el 29.10 %, Myrsine coriacea con
el 20.78 %, y el resto presento valores por debajo de estas especies.
Tabla 27. Familias con mayor valor de importancia ecológica en el bosque de Kantupata.
Índice de Valor de Importancia por Familia en SHM – 02 de Kantupata
Familias Abu.
Relativa Dom.
Relativa Div.
Relativa IVIF al 300%
Cunoniaceae 29.80 38.98 6.45 75.23
Chloranthaceae 14.07 11.20 3.23 28.50
Aquifoliaceae 12.58 12.32 3.23 28.12
Escalloniaceae 9.60 11.08 3.23 23.91
Rosaceae 6.79 5.36 6.45 18.60
Melastomataceae 3.97 2.77 9.68 16.42
Primulaceae 7.62 3.35 3.23 14.19
Clethraceae 3.81 4.87 3.23 11.90
Lauraceae 1.16 0.88 9.68 11.71
Araliaceae 1.82 2.47 6.45 10.74
TOTAL 233.39%
Fuente: Anexo 21
26.82
10.75
33.25
70.82
35.65
35.09
0
10
20
30
40
50
60
70
80
Abu. Relativa Frec. Relativa Dom. Relativa IVI's al 300%
Var
iab
les
po
bla
cio
nal
es
(%)
Weinmannia crassifolia Ilex andicolaHedyosmum angustifolium Escallonia myrtilloidesMyrsine coriacea Hesperomeles ferrugineaClethra ferruginea Weinmannia bangiiPrunus integrifolia Psychotria indet 1Schefflera herzogii Meriania vargasii

80
Figura 27. Representación gráfica de las Familias ecológicamente más importantes en el Bosque de Kantupata.
Fuente: Tabla 27 En la parcela SHM – 02, la familia mas importante fue Cunoniaceae con un valor de IVIF
de 75.23 % por ser la mas abundante y dominante de la parcela (Tabla 30), las familias
que siguen en importancia son Chlorantaceae con 28.50 %, Aquifoliaceae con 28.12 %,
Escalloniaceae con 23.91 %, Rosaceae con 18.60 %, el resto tuvo valores por debajo de
estas.
3. Análisis de diversidad
La diversidad fue determinada a nivel de riqueza y abundancia de especies registradas
en ambas parcelas, es necesario detallar que no todas las especies fueron determinadas
debido a que algunas muestras biológicas colectadas fueron estériles, lo que imposibilitó
su determinación a nivel de especie.
a) Diversidad alfa (α)
Tabla 28. Resultado de los Índices de diversidad
ÍNDICES SHM - 01 (Intipunku)
SHM - 02 (Kantupata)
TOTAL (Bosques de Wiñaywayna)
Riqueza 45 31 69
Individuos 682 604 1286
I. Shannon_H 3.063 2.477 3.432
I. Simpson_1-D 0.9327 0.8702 0.9515
Evenness_e^H/S 0.4752 0.384 0.4482
Equitability_J 0.8045 0.7213 0.8105
Fuente: Anexo 22
0 10 20 30 40 50 60 70 80
Cunoniaceae
Chloranthaceae
Aquifoliaceae
Escalloniaceae
Rosaceae
Melastomataceae
Primulaceae
Clethraceae
Lauraceae
Araliaceae
75.23
28.50
28.12
23.91
18.60
16.42
14.19
11.90
11.71
10.74 IVIF al 300% en SHM - 02

81
Figura 28. Representación gráfica de los índices de diversidad en cada una de las parcelas de Wiñaywayna
Fuente: Tabla 28
El índice de Shannon Wiener en base a la riqueza de especies indica una diversidad
moderadamente alta de 3.063 para la parcela SHM – 01 de Intipunku y de 2.477 para
SHM – 02 de Kantupata y el Índice de Simpson en base a la abundancia indica una
diversidad alta de 0.93 para la parcela SHM - 01 de Intipunku y 0.87 para la parcela SHM
- 02 de Kantupata, según estos resultados la parcela de Intipunku fue mas diversa que
la parcela de Kantupata.
El índice de Equidad de Pielou muestra una distribución proporcional y casi equitativa
del numero de especies por numero de individuos por presentar valores próximos a 1,
0.87
0.93
0.95
2.477
3.063
3.432

82
siendo para la parcela SHM - 01 de Intipunku el valor de 0.80 y para la parcela SHM – 02
de Kantupata el valor de 0.72.
b) Análisis de riqueza de especies.
En la figura 29 se observa que las curvas de los limites de confianza al 95% no se
sobreponen entre si, esto indica que la riqueza de especies en ambas parcelas es
diferente, se observa que la parcelas SHM – 01 (Intipunku) presenta mayor riqueza con
45 especies por 682 individuos (Tabla 28) mientras que la parcela SHM – 02 (Kantupata)
presenta 31 especies por 604 individuos (Tabla 28), entre ambas parcelas el registro de
especies fue de 69 en un total de 1286 individuos (Tabla 28).
Figura 29. Curvas de rarefacción de riqueza de especies en ambas parcelas evaluadas.
Si tenemos mayor área de muestreo (mas parcelas de evaluación) donde se registre
mayor número de individuos es muy probable encontrar un alto número de especies
que indique una alta diversidad dentro del Santuario Histórico de Machupicchu.

83
c) Diversidad beta (β)
Tabla 29. Índices de Similaridad (%) de Morissita Horn.
Morissita Horn SHM 01 SHM 02
SHM 01 1 1.97
SHM 02 1.97 1
Fuente: Anexo 23
Según el Índice de Morissita Horm, las parcelas presentan una similitud muy baja de
solo 1.97 %, quiere decir que son diferentes y entre si solo comparten 7 especies,
esto se debe a la gradiente altitudinal de 653m que existe entre ellas y al tipo de
bosque que condicionan la baja probabilidad de presencia de una especie en ambas
parcelas.

84
4.2. DISCUSIONES
4.2.1. Dinámica poblacional
En la parcela de Intipunku se determino una tasa de mortalidad de 1.18 %, mayor que
las tasas de reclutamiento (0.79 %) y renovación (0.99 %), en la parcela de Kantupata
ocurrió lo contrario, se determinó una tasa de reclutamiento (2.05%) mayor que las
tasas de mortalidad (1.21 %) y renovación (1.63 %).
En ambas parcelas las tasas de mortalidad fueron similares pero las tasas de
reclutamiento y renovación fue mayor en la parcela de Kantupata que se encuentra a
mayor altitud, quiere decir que esta parcela presento mayor actividad poblacional. La
diferencia de las tasas de reclutamiento entre las parcelas se debe principalmente a la
gradiente altitudinal que existe ellas y al tipo de bosque, porque a mayor altitud
(Kantupata a los 3200m) el bosque es de dosel mas bajo y de menor densidad
poblacional (mas ralo), lo que permite mayor incidencia de la luz y genera mayor
competencia en el crecimiento, en Intipunku a los 2653m, ocurre lo contrario por lo se
da un crecimiento lento de los arboles, que esta condicionado por los factores y
condiciones ambientales del bosque, esto generó una baja tasa de reclutamiento. Las
tasas de mortalidad determinadas en ambas parcelas, corresponden a la mortalidad
“Transfondo” que toma valores menores al 5 %, esto comprende eventos de mortalidad
a pequeña escala como muertos en pie, rompimiento del tallo y volcamiento de raíz y
ocasionan cambios graduales (Lugo y Scatena 1996).
Para ser los primeros resultados de dinámica poblacional en los bosques estudiados, se
muestra que ambas parcelas tienen un desequilibrio poblacional, esto se puede deber a
que los bosques son jóvenes o bosques no maduros que están en proceso de
crecimiento y recuperación como es el caso del bosque de Intipunku que en algún
momento ha sufrido intervenciónes antrópicas.
Ramirez, Torres y Serrano 2012, indican un equilibrio entre las tasas de mortalidad y
reclutamiento de 1.59 % y 1.64 % respectivamente, en bosques nublado de Venezuela
(2200 – 2500m), los cuales son menores a los resultados obtenidos de Kantupata y
mayores a los de Intipunku.

85
Velásquez, Maniguaje y Duque 2012, indican una tasa de mortalidad y reclutamiento de
0.88% y 1.16 % respectivamente, en bosques andinos de Colombia, los cuales son
mayores a los resultados de Intipunkuy y menores a los dee Kantupata, y en ambos
estudios lo bosques muestran mas dinamismo con un desquilibrio entre las tasas.
Los resultados de las tasas de mortalidad (1.39%) y reclutamiento (1.2%) reportados por
Quinto et al. 2009, en un bosque pluvial tropical, son mayores a los resultados de
Intipunku y menores a los de Kantupata, esta diferencia se debe principalmente al tipo
de bosque donde se desarrollo cada investigación.
4.2.2. Biomasa aérea y carbono
En este estudio, la biomasa aérea y carbono almacenado del Bosque de Intipunku
(2653m) fue de 145 ± 2.80 T ha-1 y 72.99 ± 1.40TCha-1 respectivamente y en el bosque
de Kantupata (3200) fue de 101.06 ± 3.46 T ha-1 y 50.53 ± 1.73 TC ha-1. Se observa una
clara diferencia de biomasa aérea y carbono almacenado en ambas parcelas, siendo
Intipunku donde hubo mayor acumulación, esta variación se debe principalmente a la
gradiente altitudinal, a la diferente composición florística que se presento en cada una
y al tipo de bosque donde se encuantra establecida cada una.
En otros estudios, Phillips et al. (2011), determinarón 145,5 ±17,3 T/ha de biomasa aérea
para un Bosque Humedo Montano y 125,5 ±22 T/ha para un Bosque muy Humedo
Montano de Colombia, lo cual es similar a lo reportado para el bosque de Intipunku y
mayor a lo reportado para el bosque de Kantupata de este estudio. Mientras que
Rodríguez et al. (2006), reporta un valor de 56.7 Mg C ha-1 de carbono para el bosque
de niebla en la reserva de la biosfera “El Cielo” en Tamaulipas, México, lo cual es
ligeramente mayor a lo registrado en el bosque montano de Kantupata de este estudio.
Velásquez, Maniguaje y Duque (2012), reportaron 243.44 ± 9.82 T/ha de biomasa aérea
en bosques andinos de la Cordillera Occidental, Colombia, valor muy superior a lo
registrado en este estudio.
Al comparar con estudios realizados en los bosques montanos del Perú, el carbono
estimado en la presente investigación es mayor que lo reportado por Girardin et al.
(2010), en los altos Andes del Perú (Parque Nacional de Manu), donde reporta 47.03 T
C ha-1 a los 3020 m, 65.90 T C ha-1 a los 2720m y 64.90 T C ha-1 a los 3025m y parecidos
a lo reportado por Gibbon et al. (2010) en bosques nublados y la puna de la Cordillera

86
Oriental peruana (Parque Nacional del Manu), donde también reportó un promedio de
53.5 ± 5.7 MgCha-1 de carbono. Ureta (2009) estimo la cantidad de carbono almacenado
en la biomasa aérea en el bosque montano de la Quebrada de Yanachaga Chemillen
(PNYCH) a los 3110 m de altitud, de 64.66 TCha-1, lo registrado para Intipunku es mayor
y lo registrdo para Kantupata es menor a este valor. Con lo reportado por Cruzado (2010)
en bosques altoandinos de la Concesión para Conservación Alto Huayabamba (CCAH),
en el departamento de San Martín (3000 – 4600 m de altitud), de 121.18 Tha-1 de
biomasa aérea y 69,77 ± 12,8 TCha-1 de Carbono para el componente arbóreo, lo que se
reporta para Intipunku es ligeramente superior. Mientras que, con lo reportado por
Bellota, T.D. 2015, de 241.27 T ha-1 de biomasa aérea en el bosque montano (1391m de
altitud) de la Reserva Comunal de “El Sira”, este valor es mucho mayor a los valores
registrados en la presente investigación, esta diferencia básicamente se debe al modelo
alométrico aplicado en la determinación de la biomasa aérea en cada investigación y a
la relación de densidad especifica de madera con el DAP de las especies que componen
cada bosque montano descrito anteriormente en las diferentes investigaciones
mencionadas, por lo que regiones con altos valores de biomasa presentan mayor
proporción de especies con alta densidad de madera (Baker et al. 2004).
Es necesario que haya mas áreas de muestreo en los diferentes bosques montanos y
talvez que la estimación de biomasa aérea y carbono en cada uno se aplique con
metodologías similares, para que haya menos segos y/o diferencias de estimación,
además de la prioridad de evaluar los contenidos de carbono en la biomasa total
(biomasa del suelo, de raíces, fuste, ramas y hojas) de los bosques montabos. En el
presente estudio se presento ciertas limitaciones, como menor número de áreas de
muestreo (2 parcelas permanentes) para un área muy grande del Santuario Historico de
Machupicchu y que las parcelas de evaluación fueran recientes, porque los resultados
que se presentan en esta investigación provienen del primer monitoreo de las parcelas
permanente que se encuentran en Wiñaywayna del Santuario Historico de
Machupicchu.

87
4.2.3. Analisis poblacional
4.2.3.1. Composición florística, IVI’s y diversidad
La parcela de Intipunku esta compuesto por 26 familias con 36 generos y 45 especies de
arboles con DAP ≥ 10 en 682 individuos por ha de bosque, que ocupan un área basal de
22.85m2/ha, las especies ecológicamente mas importantes en esta parcela son
Meliosma peytonii (33.50 %) por la mas abundante, Hieronyma oblonga (29.33 %) por
se la segunda especie en abundancia y dominancia, Gordonina fruticosa (28.43 %) por
ser la mas dominante en la parcela. La parcela de Kantupata está compuesto por 22
familias con 28 géneros y 31 especies de arboles con DAP ≥ 10 en 604 individuos por ha,
que ocupan 17.46m2/ha de área basal y las especies ecológicamente mas importantes
son Weinmannia carssifolia (70.82 %) por ser la mas abundante y dominante, Ilex
andicola (35.65 %) y Hedyosmun angustifolium (35.09%) por ser las siguientes mas
abundantes. Se presenta una variación en la composición florística debido a la gradiente
altitudinal de 635m de separación entre las parcelas, al tipo de bosque montano y a las
características en cada zona de vida, siendo la parcela de Intipunku de mayor
composición. La diversidad alfa según los índices de Shannon y Simpson fue alta en
ambas parcelas de estudio, mas la diversidad beta muestra una similitud muy baja entre
ellas por compartir un minimo de 7 especies. Esto demuestra que los bosques de
Intipunku y Kantupata son muy diferentes y que a medida que incrementa la altitud, la
riqueza de especies disminuye.
Por la variabilidad de metodología aplicadas en los diferentes estudios, se compara con
estudios realizados con la misma metodología aplicada en este estudio (arboles con DAP
≥ 10), los valores registrados en el presente estudio son menores a lo reportado por
Velásquez, Maniguaje y Duque (2012), en bosques andinos de la Cordillera Occidental,
Colombia, donde registró 222 especies, 113 géneros y 60 familias en 1664 individuos en
2.0 ha de bosque, con 156 especies/ha, y ligeramente menor a lo de Ramirez, Torres y
Serrano (2012), en bosque húmedo montano bajo (2200 y 2500 m de altitud) de la
Cordillera de los Andes, Venezuela, donde registró 55 especies en 622 individuos en 1.5
ha, donde también registró a Myrcia fallax como la mas representativa.
Respectos a estudios realizados en bosques montanos del territorio peruano, Palomino
(1999), en bosque nublado de San Pedro (Parque Nacional del Manú), reportó 192

88
especies, 91 generos y 45 familias en 618 individuos en 1.0 – ha, donde indica una alta
diversidad y presencia de familias Moraceae (21.94%), Euphorbiaceae (17.7%),
Melastomataceae (8.10%) como las mas importantes, al igual que el presente estudio.
Nina (2014), para los bosques numblados de Trocha Union del Parque Nacional de Manu
reportó 43 especies, 22 géneros y 20 familias a los 3450 m y a 85 especies, 34 géneros y
26 familias a los 2750 m de altitud, estos valores fuerón mayores a lo registrado en el
presente estudio, pero similares en la presencia de algunos generos y especies como
Weinmannia, Cyathea, Myrsine, Miconia, etc.
Estudios realizados próximos al área de estudio como Rojas y Pelaez (2013), reportaron
58 especies, 33 familias y 49 géneros en 788 individuos en 1.0 – ha con una alta
diversidad para la cuenca de Torontoy del Santuario Histórico de Machupicchu, estos
valores son relativamente menores a lo reportado en el presente estudio, pero son
similares en presentar una alta diversidad. Otros estudios próximos, realizados con
metodologías diferentes como los de Dueñas (1992), Monteagudo (1997), Galiano y
Tupayachi (2000), en los bosques de Wiñaywayna, mencionan un alta diversidad y
composición floristica muy similar a lo registrado en el presente estudio.
En cambio, con lo de Galiano y Núñez (2011), quienes estimaron más de 550 especies
de árboles distribuidas en 74 familias y 192 géneros para todo el Santuario Histórico de
Machupicchu y zonas de amortiguamiento con Rubiaceae, Solanaceae, Asteraceae,
Melastomataceae, Rosaceae, como las familias más representativas, lo registrado en el
presente estudio es menor en especies, pues se desarrolló solo para los bosques de
Wiñaywayna, más coinciden las familias y algunas especies reportadas en ambos
estudios al igual que con los de Valenzuela et al. (2014), quienes reportarón mas de 300
especies de arboles para el Santuario Historico de Machupicchu y los describe en el libro
“Arboles de Machupicchu Cusco – Perú”, en cuanto al registro de helechos arbóreos, en
el estudio de Chávez (1993), indica que dentro del Santuario Histórico de Machupicchu
se encontró 4 especies y fuera de los límites del Santuario solo 3 especies, en el presente
estudio se reporta 7 especies de Cyatheas.

89
CONCLUSIONES
1. Las tasas de mortalidad(TM) anual de arboles, fue similar en ambas parcelas evaluadas
(Intipunku y Kntupata) de 1.18 % y 1.21 % respectivamente, las tasas de reclutamiento
y renovación fue mayor en la parcela de Kantupata de 2.05 % y 1.63 %, indicando una
mayor actividad o dinamica en en bosque de Kantupata a los 3200 m de altitud,
respecto al bosque de Intipunku a los 2653 m de altitud donde se dio una menor
actividad con las tasasde 0.79 % y 0.99 % respectivamente, se encontró un
desequilibrio entre las tasa de mortalidad y reclutamiento, lo que mostró una alta
actividad en ambas parcelas por ser bosques no maduros y están en constante
dinámica.
2. El bosque de intipunku a los 2653 m de altitud, registró 145 ± 2.80 T ha-1 de biomasa
aérea con 72.99 ± 1.40 T C ha-1 de carbono almacenado, con una perdida de biomasa
aérea por mortalidad del 1 % anual y una productividad primaria neta (PPN) de 0.7 %
anual, las especies con mayor aporte de biomasa fueron Gordonia fruticosa y Aniba
coto con 19 Tha-1 cada uno. El bosque de Kantupata a los 3200 m de altitud, registró
101.06 ± 3.46 Tha-1 de biomasa aérea con 50.53 ± 1.73 TCha-1 de carbono almacenado,
con una perdida de biomasa por mortalidad del 1.3 % y una productividad primaria
neta (PPN) de 1.2 % anual, la especie con mayor aporte fue Weinmannia crassifolia con
32.78Tha-1 de biomasa aérea.
3. En el bosque de intipunku se registró 45 especies, 36 géneros y 26 familias en 682
individuos, con un área basal de 22.85m2/ha, donde las especies arbóreas
ecológicamente mas importantes fuerón Meliosma peytonii (33.50 %), Hieronyma
oblonga (29.33 %) y Gordonina fruticosa (28.43 %) y las familias mas abuandates
fueron Sabiaceae con 104 individuos, Phyllanthaceae y Melastomataceae con 73
individuos cada una por hectarea. En el bosque de Kantupata, se registró 31 especies,
28 géneros y 22 familias en 604 individuos, con un área basal de 17.46m2/ha, donde
las especies mas importantes fuerón Weinmannia crassifolia (70.82 %), Ilex andicola
(35.65 %) y Hedyosmum angustifolium (35.09 %) y las familias mas abundantes fueron
Cunoniaceae con 180 individuos, Chloranthaceae con 85 individuos y Aquifoliaceae

90
con 76 individuos por hectárea. Hubo mayor registro de especies e individuos en la
parcela SHM – 01, y la diversidad alfa fue alta en ambas parcelas con una muy baja
similaridad entre ellas por compartir solo 7 especies, lo cual se debe a la gradiente
altitudinal de 653m que se presento entre las parcelas y al tipo de bosques montano.

91
RECOMENDACIONES Y SUGERENCIAS
A pesar de los esfuerzos por conocer la ecología de los bosques montanos, aún
queda muchos por hacer, por lo que es necesario continuar con las evaluaciones
y monitoreos a largo plazo, para lo cual se recomienda instalar más parcelas
permanentes en los diferentes bosques montanos que puedan abarcar mayor
área dentro del Santuario Histórico de Machupicchu, donde se evalúen y
monitoreen de manera periódica la dinámica poblacional y el almacenamiento
de biomasa y carbono.
Se recomienda, realizar estudios completos de biomasa y carbono que involucre
no solo a nivel fustal sino, a nivel de ramas, copas, raíces, necromasa y suelo y
así entender mucho mejor la ecología y la dinámica poblacional de los bosques
montanos y la capacidad que tienen de aportar a la reducción de CO2 como
importantes sumideros de carbono.

92
BIBLIOGRAFÍA
AGUILAR, M. y REYNEL C., 2009. Dinámica forestal y regeneración en un bosque
montano nublado de la selva central del Perú, Herbario de la Facultad de Ciencias
Forestales de la Universidad Nacional Agraria-La Molina. 167 pp.
ÁLVAREZ, A. G., García, C. N., Krasilnikov P., García O. F., 2013. Almacenes de Carbono
en bosques montanos de niebla de la sierra norte de Oaxaca, México. Agrociencia 47:
171-180.
ALVAREZ, E., Duque, A., Saldarriaga, J., Cabrera, K., de las Salas, G., del Valle, I., Lema,
A., Moreno, F., Orrego, S. y Rodríguez, L., 2012. Tree above-ground biomass allometries
for carbon stocks estimation in the natural forests of Colombia. Forest Ecology and
Management 267: 297-308.
APG. IV., 2016. An update of the Angiosperm Phylogeny Group classification for the
orders and families of flowering plants: APG IV– The Angiosperm Phylogeny Group – The
Linnean Society of London, Botanical Journal of the Linnean Society, 181:1-20.
ARAGÃO, L., Malhi, Y., Metcalfe, D., Silva-Espejo, J., Jiménez, E., Navarrete, D.,
Almeida, S., Costa, A. et al., 2009. Above- and below-ground net primary productivity
across ten Amazonian forests on contrasting soils. Biogeosciences 6, 2759-2778.
AVELLA M. A., 2016. Los bosques de robles (Fagáceas) en Colombia: composición
florística, estructura, diversidad y conservación.
BAEV P., y L. Penev., 1995. BIODIV: program for calculating biological diversity
parameters, similarity, niche overlap, and cluster analysis. PENSOFT. Sofia-Moscú.
BELLOTA,T.D., 2014. Diversidad florística, biomasa aérea y dinámica poblacional a
través de la gradiente altitudinal en parcelas permanentes en la reserva comunal del Sira
- Huánuco - Perú.
BERRY, P.E., 2002. Diversidad y endemismo en los bosques neotropicales de bajura. In:
Biología y conservación de bosques neotropicales. Eds. 77. MR Guariguata; GH Kattan.
Libro Universitario Regional (LUR). Cartago, CR. 83-96 p.

93
BERTZKY M, Ravilious C, Araujo Navas A, Kapos V, Carrión D, Chíu M, Dickson B. 2010.
Carbon, biodiversity and ecosystem services: Exploring co-benefits. Ecuador:
NEPWCMC, Cambridge, UK.
BOBÓRQUEZ F. A., Sanín O. D., y Silva E. N. 2011. Estructura y composición arbórea de
los bosques del diablo (San Felix, Salamina, Caldas), selva altoandina de la cordillera
central Colombiana. bol.cient.mus.hist.nat. 16 (2): 39 – 52.
BRAKO, L. & S. L. Zaruchii. 1993. Catalogue of the Flowering Plants and Gymnosperms
of Peru in Monographs in Systematic Botani from the Misssouri Botanical Garden
Published by MO USA.
BRAY, R. J. y CURTIS, J. T., 1957. An Ordination of the Upland Forest Communities of
Southern Wisconsin, Ecol. Monogr., 27: 325-349
BROWN, S., 1997. Estimating biomass change of tropical forest: a primer. Forestry Paper
134, FAO, Rome.
BUBB, P., May, I., Miles, L. y Sayer, J., 2004. Cloud forest Agenda. UNEP-WCMC.
Cambridge, UK.
CARLOTTO, V., Cardenas, j. y Fidel,l., 2009. La geología, evolución geomorfológica y
geodinámica externa de la ciudad inka de Machupicchu, Cusco-Perú. Rev. Asoc. Geol.
Argent., vol.65, n.4, pp.
CARLOTTO, V., Cárdenas, R., Fidel, L., 2007. La Geología en la Conservación de
Machupicchu. Boletín INGEMMET, serie I Patrimonio y Geoturismo Nº1, 305p.
CERÓN, C.E., 2003. Manual de Botánica Sistemática, Etnobotánica y Métodos de Estudio
en el Ecuador. Herbario “Alfredo Paredes” QAP, Escuela de Biología de la Universidad
Central del Ecuador. Quito-Ecuador 267-291 p.
CHAMBERS, J. Q., J. dos Santos, R. J. Ribeiro & N. Higuchi, 2001. Tree damage,
allometric relationships, and aboveground net primary production in central Amazon
forest. Forest Ecology and Management 5348:1–12.
CHAVE,J., Andalo, C., Brown, S., Cairns, M. A., Chambers, J. Q., Eamus, D., Folster, H.,
Fromard, F., Higuchi, N., Kira, T. Lescure, J.-P., Nelson, B. W., Ogawa, H., Puig, H., Riera,

94
B. y T. Yamakura, 2005. Tree allometry and improved estimation of carbon stocks and
balance in tropical forests. Ecosystem Ecology. Oecología.145: 87-99.
CHAVEZ H. W. 1994. Estudio Poblacional de Helechos Arborescentes en Wiñaywayna
Santuario Histórico de Machupicchu y Cedropata. Seminario Curricular. Facultad de
Ciencias Biológicas UNSAAC.
CLARK DA, Brown S, Kicklighter DW, Chambers JQ, Thomlinson JR, Ni J., 2001.
Measuring net primary production in forests: concepts and field methods. Ecological
Applications, 11, 356–370.
CONDIT, R., 1998. Tropical forest census plots. Springer Verlag, Berlin.
CONTRERAS, F; Leaño, C; Licona, J.C; Dauber, E; Gunnar, L; Hager, N; Caba, C., 1999.
Guía para la instalación y evaluación de parcelas permanentes de muestreo. Ed. R.
Duchén. Santa Cruz de la Sierra, BO. El país. 80 p.
CRUZADO, B. L., 2010. Determinación de las reservas de carbono en la biomasa aérea
de los bosques altoandinos de la concesión para conservación alto Huayabamba - San
Martín.
CUESTA, F., Peralvo, M. y Valarezo, N., 2009. Los bosques montanos de los Andes
Tropicales. Una evaluación regional de su estado de conservación y de su vulnerabilidad
a efectos del cambio climático. Quito.
SERVICIO NACIONAL DE ÁREAS NATURALES PROTEGIDAS - SERNANP (2014). Plan
Maestro Santuario Histórico de Machupicchu 2015 - 2019
DÍAZ, S., M. Cabido y F. Casanoves, 1998. Plant functional traits and environmental
filters at a regional scale J. Veg. Sci.9:113-122.
DÍAZ F. R., Acosta M. M., Carrillo A. F., Buendía R. E., Flores A. E. & Etchevers B. J.,
2007. Determinación de ecuaciones alométricas para estimar biomasa y carbono en
Pinus patula Schl. et Cham. Madera y Bosques. 13(1) :25-34.
DINERSTEIN, E., D. M. Olson, D. J. Graham, A. L. Webster,S. A. Primm, M. P. Bookbinder
& G. Ledec., 1995. A Conservation Assessment of the Terrestrial Ecoregions of Latin
America and the Caribbean. The World Bank, Washington, D.C.

95
DUEÑAS. L. H., 1992. Estructura diamétrica y diversidad drbórea en Wiñaywayna
(Santuario Histórico de Machupicchu) Tesis para optar el Titulo de Biólogo, Facultad de
Ciencias Biológicas UNSAAC.
FEINSINGER, P., 2003. El diseño de estudios para la conservación de la Biodiversidad.
Eds. FAN. Santa Cruz – Bolivia. 242p.
FONT QUER., P., 1985. Diccionario de Botánica. 9° reimpresión, 1.era ed. Barcelona:
Editorial Labor. 1244 pp.
FOSTER, P., 2001. The potential negative impacts of global climate change on tropical
montane cloud forests. Earth-Science Reviews 55:73-106.
FRANCO, L. J. y et al. 1989. Manual de Ecología, Edit. Trillas Segunda Edición México. PP,
14: 93-99 17: 114-119 18: 122-126 19:128-131
GALIANO, W. 1995. Decálogo Normativo para la Conservación, Desarrollo y manejo del
Santuario Histórico de Machupicchu. Opciones II: 4-7.
GALIANO Y Tupayachi, 2002. Ampliación del Inventario de Biodiversidad (Botanica) del
Santuario Histórico de Machupicchu. PROFONAMPE-PMP.
GALIANO, W. Y Nuñez, P., 1992. Sinopsis de las Especies Arbóreas del Santuario
Histórico de Machupicchu y su zona de Amortiguamiento.
GALIANO, W. y Núñez, P. 2011. Sinopsis de la Especies Arbóreas del Santuario Histórico
de Machupicchu y su Zona de Amortiguamiento Revista El Antoniano N° 117, pág. 178.
GARCÍA C. K., 2011. Spatial variability in species composition in neotropical Montane
tree communities.
GARCÍA G. F, 2014. Composición y estructura florística del bosque de neblina montano,
del sector “San Antonio de la montaña”, cantón baños, provincia de Tungurahua,
Ecuador.
GENTRY, A. H. 1995. Patterns of diversity and floristic composition in neotropical
montane forests. Pp 103-126 en H. Churchill S. P. B., E. Forero y J. L. Luteyn (Eds.).
Neotropical Montane Forest Biodiversity and Conservation Symposium (1993, Bronx,

96
N.Y., USA). Biodiversity and conservation of neotropical montane forests: Proceedings.
New York Botanical Garden, Bronx, New York, USA.
GENTRY, A. H., 1992. A Field Guide to the Families and Genera of Woody Plants of
Northwest South America (Colombia, Ecuador, Perú) with supplementary notes on
herbaceous taxa. 1ra ed. Washington, DC: Conservation International. 920 pp.
GENTRY, A.H., 1988a. Changes in plant community diversity and floristic composition on
environmental and geographical gradients. Annals of the Missouri Botanical Garden 75
(1):1–34.
GIBBON, A., Silman, M., Malhi, Y., Fisher, J., Meir, P., Zimmermann, M., Dargie, G.,
Farfan, W. y Garcia, K., 2010. Ecosystem carbon storage across the grassland-Forest
transition in the high Andes of Manu National Park, Peru. Ecosystems 13: 1097-1111.
GIBBS, H., Brown, S., Niles, J. y Foley, J., 2007. Monitoring and estimating tropical forest
carbon stocks: making REED a reality. Environmental Research Letters 2: 045023.
GIRARDIN C. A. J., Malhi Y.M., Aragao L.E.O.C., Mamani M., Huaraca H. W., Durand L.,
et al., 2010. Net primary productivity allocation and cycling of carbon along a tropical
forest elevational transect in the Peruvian Andes. Global Change Biology.
GRUBB, P.J. & Whitmore, T.C. 1966. A comparison on montane and lowland rainforest
in Ecuador: II. The climate and its effects on the distribution and physiognomy of the
forests. Journal of Ecology, 54, 303-333.
GRUBB, P.J., Lloyd, J.R., Pennington, T.D. & Whitmore, T.C. 1963. A comparison of
montane and lowland rain forest in Ecuador: I. The forest structure, physiognomy, and
floristics. Journal of Ecology, 51, 567-601.
HAMILTON, S. 1995. Una campaña por bosques nublados: Ecosistemas únicos y valiosos
en peligro, UINC.
HOLDRIGE, L.R. 1979. Ecología Basada en Zonas de Vida, Instituto Americano de Ciencias
Agrícolas Costa Rica.

97
HONORIO C. E. N. & Baker. T. R., 2010. Manual para el Monitoreo del ciclo del carbón
en Bosques Amazónicos. Instituto de Investigaciones de la Amazonia
Peruana/Universidad de Leeds. Lima, 54p.
INRENA. 2003. Propuesta de Zonificación y Zona de Amortiguamiento del Santuario
Historico de Machupicchu.
JORGENSEN, P. M. y León-Yánez, S. (Eds.). 1999. Catalogue of the vascular plants of
Ecuador. Missouri Botanical Garden Press, St. Louis.
JOSSE, C., Cuesta, F., Navarro, G., Barrena, V., Cabrera, E., Chacon-Moreno, E., Ferreira,
W., Peralvo, M., SAito, J. y Tovar, A. 2009a. Ecosistemas de los Andes del Norte y
Centro. Bolivia, Colombia, Ecuador, Perú y Venezuela. pp. Secretaría General de la
Comunidad Andina, Programa Regional ECOBONA_Intercooperation, CONDESAN-
Proyecto Páramo Andino, Programa BioAndes, EcoCiencia, NatureServe, IAvH, LTA-
UNALM, ICAE-ULA, CDC-UNALM, RumBOL SRL, Lima.
JOSSE, C., G. Navarro, F. Encarnación, A. Tovar, P. Comer, W. Ferreira, F. Rodríguez, J.
Saito, J. Sanjurjo, J. Dyson, E. Rubin de Celis, R. Zárate, J. Chang, M. Ahuite, C. Vargas,
F. Paredes, W. Castro, J. Maco and F. Reátegui. 2007. Sistemas cológicos de la cuenca
amazónica de Perú y Bolivia. Arlington VA: NatureServe.
JOSSE, C., G. Navarro, P. Comer, R. Evans, D. Faber-Langendoen, M. Fellows, G. Kittel,
S. Menard, M. Pyne, M. Reid, K. Schulz, K. Snow, and J. Teague. 2003. Ecological
systems of Latin America and the Caribbean: s working classification of terrestrial
systems. Arlington VA: NatureServe.
KELLY, D., E. Tanner, E. NicLughadha y V. Kapos. 1994. Floristic and biogeography of a
rain forest in the Venezuelan Andes. Journal of Biogeography 21: 421 – 440.
KESSLER, M. 2002. The elevational gradient of Andean plant endemism: Varying
influences of taxon-specific traits and topography at different taxonomic levels. Journal
of Biogeography 29:1159-1165.
KESSLER, M., Herzog, S. K., Fjeldså, J. y Bach, K. 2001. Species richness and endemism
of plant and bird communities along two gradients of elevation, humidity y land use in
the Bolivian Andes. Diversity & Distributions 7:61-77.

98
KREBS, C. J., 1978. Ecology; the experimental analysis of distribution and abundance.
Harper & Row (eds.). Nueva York. 678p.
LINARES M. A., 2008. Análisis florístico y estructural de la vegetación de una selva
nublada en un gradiente altitudinal, en la Mucuy, estado de Merida, Venezuela.
LUGO, A.E. y F.N. Scatena. 1996. Background and catastrophic tree mortality in tropical
moist, wet and rain forests. Biotropica 28:585-599.
MACDICKEN K., 1997. A guide to monitoring carbon storage in forestry and agroforestry
projects (Arlington, VA: Winrock international).
MAGURRAN A., 1988. Ecological diversity and its measurements. Princeton University
Press. New Jersey-U.S.A. 179 P.
MAGURRAN, A., 1991. Diversidad ecológica y su medición. Vedra (Ed.). Barcelona –
España. 54 – 70 p.
MAGURRAN, A., 2004. Measuring biological diversity. Blackwell Science. Oxford – U.S.A.
256 P.
MALHI, Y., Baker, T., Phillips, O. L., Almeida, S., Alvarez, E., Arroyo, L., Chave, J.,
Czimczik, C., Di Fiore, A., Higuchi, N., Killen, T., Laurance, S. G., Laurance, W. F., Lewis,
S., Mercado Montoya, L. M., Monteagudo, A., Neill, D., Núñez Vargas, P., Patiño, S.,
Pitman, N., Quesada, C. A., Salomao R., Macedo Silva, J. N., Torres Lezama, A., Vasquez
Martinez, R., Terborgh, J., Vicenti, B. y Lloyd, J., 2004. The aboveground coarse wood
productivity of 104 neotropical forest plots. Global Change Biology 10: 563-59.
MALHI, Y., Phillips,O.L., Baker, T., Almeida, S., Fredericksen, T., Grace, J., Higuchi, N.,
Killeen, T., Laurance, W.L., Leaño, C., Lloyd, J., Meir, P., Monteagudo, A., Neill, D.,
Nuñez Vargas, P., Panfil, S., Pitman, N. et al., 2002. An International Network to
Understand the Biomass and Dynamics of Amazonian Forests (RAINFOR). Journal of
Vegetation Science. 13, 439-450.
MATTEUCCI S. D. & Colma A., 1982, “Metodología para el estudio de la vegetación”
Secretaria General de la Organización de los Estados Americanos, Programa Regional de
Desarrollo Científico y Tecnológico, Washington, D:C. USA. P. 21-53.

99
MONTEAGUDO M. A., 1997. Evaluación de la diversidad arbórea en tres bosques
representativos del Santuario Histórico de Machupicchu. Tesis para optar el título de
Biólogo. Facultad de ciencias biológicas. Universidad Nacional de San Antonio Abad del
Cusco – Cusco.
MONTEAGUDO M. A., Valenzuela L.; Vasquez R.; Rojas R., Phillips O., Lopez G., 2014.
Primer catálogo de los árboles y afines de la Reserva Comunal El Sira. Rev. Arnaldoa,
21(1): 127 -164.
MORENO E. C., I. Zuria M., Garcia G., Sanchez I., Castellanos M., Martinez & A. Rojas.
2006. Trends in the measurement of alpha diversity in the last two decades. Interciencia
31: 67 – 71 p.
MORENO E.C., 2001. “Métodos para medir biodiversidad” M&T – manuales y tesis SEA,
vol1, Zaragoza, 84 pp.
MOSSER G., Leuschner C., Hertel D., Graefe S., Soethes N., Iost S., 2011. Elevation
effects on the carbon budget of tropical mountain forests (S Ecuador): the role of the
belowground compartment. Global Change Biology 17: 16.
MOSTACEDO B., Fredericksen T., 2000. “Manual de métodos básicos de muestreo y
análisis en ecología vegetal” BOLFOR, Santa Cruz de la sierra Editora el País Bolivia. 87p.
NEBEL, G; Kvist, L; Vanclay,J & Vidaurre,H. (2000). Dinámica de los bosques de la llanura
alluvial inundable de la Amazonía Peruana: Efectos de las perturbaciones e implicancias
para su manejo y conservación. Folia Amazónica Vol. 11, 65-97.
NINA Q. A., 2014. Composición, Estructura y Regeneración Arbórea en dos Parcelas
Permanentes del Bosque Nublado - Parque Nacional del Manu-Paucartambo – Cusco.
NORSE E.A., et al., 1986. “Conserving biological diversity in our national forests”. The
Wilderness Society, Washington, DC, 116p.
NUÑEZ, M. P., 1992. Flora del Santuario Histórico de Machupicchu, Seminario
Curricular, Facultad de Ciencias Biológicas. UNSAAC-CUSCO.
ONERN. 1976. Mapa ecológico del Perú, Guía explicativa. Oficina Nacional de evaluación
de los recursos naturales. Lima - Perú.

100
OSINAGA, O., Baez, S., Cuesta, F., Malizia, A., Carrilla, J., Aguirre, N. y Malizia, L. 2014.
Monitoreo de diversidad vegetal y carbono en bosques andinos-Protocolo extendido.
Protocolo 2 - Version 1. CONDESAN / IER-UNT / COSUDE. Quito, Ecuador.
PALLQUI, N.C., 2014. Dinámica de un Bosque Tropical: Biomasa Aérea y Análisis en
parcelas permanentes a largo plazo, Reserva Nacional Tambopata – Madre de Dios.
[Tesis de Licenciatura]. Cusco: Universidad Nacional San Antonio Abad de Cusco,
Facultad de Ciencias Biológicas.
PALOMINO, C. W. 1999. Diversidad y Asociación Arbórea del Bosque Nublado de San
Pedro – Reserva de Biosfera del Manu. Tesis para optar el Titulo de Biólogo, Facultad de
Ciencias Biológicas UNSAAC - Cusco - Perú.
PALOMINO, C. W., 1996. Área Mínima de Expresión y Composición Florística Arbórea en
el Bosque de Intipunku Sector de Wiñaywayna Santuario Histórico de Machupicchu.
Seminario Curricular, Facultad de Ciencias Biológicas. UNSAAC-CUSCO.
PHILLIPS, J., Duque, Á., Yepes, A., Cabrera, K., García, M., Navarrete, D., Álvarez, E. y
Cárdenas, D. 2011. Estimación de las reservas actuales (2010) de carbono almacenadas
en la biomasa aérea en bosques naturales de Colombia. Estratificación, alometría y
métodos análiticos. pp. 68. IDEAM, Bogotá.
PHILLIPS, O. & Baker, T. 2002. Field manual for plot establishement and
remeasurement. RAINFOR-Red Amazonica Inventarios Forestales. Informe Inedito,
version Noviembre 2002.
PHILLIPS, O. L. & Gentry, A. H. 1994. Increasing turnover through time in tropical forests.
Science 263, 954–958.
PHILLIPS, O., Baker, T., Feldpausch, T. & Brienen, P. 2016. RAINFOR Manual de Campo
para el Establecimiento y la Remedición de Parcelas, p 28.
PHILLIPS, O., Malhi, Y., Higuchi, N., Laurance, W., Núñez, P., Vásquez, R., Laurance, S.,
Ferreira, L., Stern, M., Brown, S. y Grace, J., 1998. Changes in the Carbon Balance of
Tropical Forests: Evidence from Long-Term Plots. Science (October 1998) 282(16):439-
442.

101
POLO U. C., 2008. Programa de biología aplicada, Facultad de Ciencias, Universidad
Militar “Nueva Granada” Vol. 4 N° 1: 135 – 142 P.
PUTZ, F.E. 1983 Liana biomass and leaf area of a “Tierra Firme” forest in the Río Negro
basin. Venezuela. Biotropica 15(3): 185-189.
QUINTO, H; Ibarguen, R. 2009. Mortalidad y Reclutamiento de Árboles en un Bosque
Pluvial Tropical de Chocó. Rev. Facultad Nacional de Agronomía - Medellin., Vol. 62,
num. 1, pp. 4855 - 4868.
RAMÍREZ, A. H., Torres, L. A., Serrano. J. 2002. Mortalidad y Reclutamiento de Árboles
en un Bosque Nublado de la Cordillera de los Andes, Venezuela. ECOTROPICOS
15(2):177-184.
REDD. 2011. Estimación de los costos de oportunidad de REDD+, Manual de
Capacitación, Versión 1.4.
RODRÍGUEZ L., R; Jiménez P., J.; Aguirre C., O. y E. Treviño. 2006. Estimación de carbono
almacenado en un bosque de niebla en Tamaulipas, México. Ciencia-UANL. Monterrey,
N. L. 9(2): 179-187.
ROJAS V. C. M. y Peláez T. Y., 2013. Diversidad arbórea y cobertura vegetal en la cuenca
de Torontoy, Santuario Histórico de Machupicchu – Cusco.
SHANNON, C.E. & W. WEAVER. 1949. The mathematical theory of Comunication.
Univiversidad of Illinois Press. Illinois.
SIERRA, C. A., Del Valle, J. I., Orrego, S. A., Moreno, F. H., Harmon, M. E., Zapata, M.,
Colorado, G. J., Herrera, M. A., Lara, W., Restrepo, D. E., Berrouet, L. M., Loaiza, L. M.,
& Benjumea, J. F. 2007. Total carbon stocks in a tropical forest landscape of the Porce
region, Colombia. Forest Ecology and Management, 243, 299-309.
SWAINE, M. D., Hall, J. B. y Alexander, I. J. 1987. Tree population dynamics at Kade,
Ghana (1968-1982). Journal of Tropical Ecology 3: 331-345.
TOVAR, A., Tovar, C., Saito, J., Soto, A., Regal, F., Cruz, Z., Véliz, C., Vásquez, P., Rivera,
G. 2010. Yungas Peruanas- Bosques montanos de la Vertiente oriental de los Andes del

102
Perú. Una perspectiva ecorregional de la conservación. Punto Impreso S.A., Miraflores,
Perú.
TUPAYACHI, H. A. Galiano, S. W. 1986. Flora del Santuario Histórico de Machupicchu.
Rev. BIOTA vol. XIV N° 95.
UNIVERSIDAD DE ALCALÁ (UA), Departamento de Ecología, 2004-2005. Métodos de
análisis de datos en ecología. Prácticas en ecología. Licenciatura de Biología y Ciencias
Ambientales. 31 p.
URETA A. M. 2009. Diferencias Altitudinales de Contenido de Carbono y Biomasa
Arbórea en el Parque Nacional Yanachaga Chemillén, Pasco-Perú.
VALENZUELA G. L., Monteagudo M. A., Calatayud H. G., Farfan V. J., Suclli M. E. &
Huamantupa Ch. I., 2014. Arboles de Machupicchu Cusco – Perú. Jardin Botanico de
Missouri a través de RAINFOR (Red Amazonica de Inventarios Forestales).
VALLEJO-Joyas M. I., Londoño-Vega, A. C., López-Camacho, R., Galeano, G., Álvarez-
Dávila, E. y Devia-Álvarez, W. 2005. Establecimiento de parcelas permanentes en
bosques de Colombia. Instituto de Investigacion de Recursos Biologicos Alexander von
Humboldt. Bogota D. C., Colombia. 310 p. (Serie: Metodos para estudios ecologicos a
largo plazo; No. 1).
VARGAS, C. 1962. "Phytomorphic representations of ancient Peruvians". Economic
Botany, 16:106-115.
VASQUEZ, M.R., Rojas G.R. y Rodríguez, E.F., 2003. Adiciones a la Flora Peruana:
especies nuevas, nuevos registros y estados taxonómicos de las Angiospermas para el
Perú. Arnaldoa 9(2): 43-110.
VELÁSQUEZ, R. J., Maniguaje, N. & Duque, A., 2012. Diversidad y dinámica de un bosque
subandino de altitud en la región norte de los Andes colombianos. Rev. Biol. Trop. (Int.
J. Trop. Biol. ISSN-0034-7744) Vol. 60 (2): 943-952.
WEAVER, P.L. 2010a. Forest structure and composition in the lower montane rain forest
of the Luquillo Mountains, Puerto Rico. Interciencia, 35, 640-646.

103
WEBERBAUER, A. 1945 (1911). “El mundo vegetal de los Andes Peruanos”. Ministerio
de Agricultura, Lima.
WHITTAKER, R. H. 1972. Evolution and measurement of species diversity Taxon,
21(2/3):213-251.
WHITTAKER, R. H., 1960. Vegetation of the Siskiyou Mountains, Oregon and California,
Ecol. Monogr., 30: 279-338.
WOLDA, H. 1983. Diversity indices and tropical cockroaches. Oecologia 58: 290 – 298 p.
ZANNE, A.E., López-Gonzalez, G., Coomes, D.A., Ilic, J., Jansen, S., Lewis, S.L., Miller,
R.B., Swenson, N.G., Wiemann, M.C. y Chave, J. (2009). Global Wood density database.
Dryad. Identifier.
ZIMMERMANN, M., Meir, P., Silman, M., Fedders, A., Gibbon, A., Malhi, Y., Urrego, D.,
Bush, M., Feeley, K., Garcia, K., Dargie, G., Farfan, W., Goetz, B., Johnson, W., Kline, K.,
Modi, A., Rurau, N., Staudt, B. y Zamora, F. 2010. No differences in soil carbon stocks
scross the tree line in the Peruvian Andes. Ecosystems 13: 62-74.
PÁGINAS WEB
Boyle, B. et al. 2013. The taxonomic name resolution service: an online tool for
automated standardization of plant names. BMC Bioinformatics 14:16.
http://tnrs.iplantcollaborative.org.
GEO GPS PERÚ, Setiembre 2008, Sistema de Información Geográfica, Instituto
Geográfica Nacional – IGN, http://www.geogpsperu.com
Missouri Botanical Garden – 4344, 2017. TRÓPICOS. Shaw Boulevard - Saint Louis,
Missouri 63110, http://www.tropicos.org/
School of Geography, 2014. FORESTPLOTS.NET University of Leeds,
https://www.forestplots.net/
SERNANP, 2008. Servicio Nacional de Áreas Naturales Protegidas por el Estado,
http://www.sernanp.gob.pe

104
Stevens, P. F., 2001. Angiosperm Phylogeny Website. Version 14, July 2017
http://www.mobot.org/MOBOT/research/APweb/
The Field Museum, (1997 – 2017). Muestras Neotropicales de Herbario, Lake Shore
Drive, Chicago, IL 60605 U.S.A. http://www.fieldmuseum.org
ANEXOS

105
1. Documentos de autorización del proyecto de Investigación e ingreso al SHM.

106
2. Ubicación de parcelas permanentes del Santuario Histórico de Machupicchu.
A, A1: Interior de bosque montano de Intipunku (2653 m) (SHM – 01), las especies dominantes son; Gordonia fruticosa (Schrad.) H.Keng, Hieronyma oblonga (Tul.) Müll.Arg., Meliosma peytonii A.H. Gentry. B, B1: Interior del bosque montano de Kantupata (3200 m)(SHM – 02), las especies dominantes son; Weinmannia crassifolia Ruiz & Pav., Ilex andicola Loes., Hedyosmum angustifolium (Ruiz & Pav.) Solms.
A
B
A.1
B.1

107
3. Protocolo de RAINFOR para la remedición y establecimiento de parcelas.
A B
C D
E
F

108
PROCEDIMIENTO EN CAMPO
A. Instalación de las parcelas permanentes en el SHM.
B. Medición del POM, DAP y toma de datos. C. Codificación y pintado de árboles. D. Codificación de reclutas en una
remedición. E. Colecta de muestras botánicas F. Registro de muestras botánicas
colectadas. G. Prensado de muestras colectadas. H. Secado de muestras I. Determinación y montaje
G
H
I
I

109
4. Ficha de registro de datos en campo
Código de Parcela:
SHM-01
Nombre de la Parcela:
Intipunku Bosque Montano Muy Humedo Fecha: 12 de Julio del 2013 Fecha: 10 al 13 de Noviembre del 2016
Etiqueta
N° T1 X Y Familia Especie
DAP
2013
POM
2013 F1 F2
Altura
2013 F3 F4 LI CI
Comentarios
2013
DAP
2016
POM
2016 F1 F2
Altura
2016 F3 F4 LI CI
Comentarios
2016
Voucher
Code Collected
236 10 20 14 Rubiaceae Guettarda crispiflora 165 1300 cn 1 17 0 0 0 4 166 1400 ce 1 12 0 0 0 4 Cambio de POM MP548 1
237 10 20 15 Clethraceae Clethra castaneifolia 174 1300 cn 1 15 0 0 0 4 193 1300 c 1 11 0 0 0 2c MP551 1
219.1 10 8 0.5 Cunoniaceae Weinmannia pubescens 123 1300 an 1 11 0 0 0 3a no colectado 0
238 11 1.3 1 Styracaceae Styrax pentlandianus 126 1700 an 1 13 0 0 0 3b 126 1700 c 1 12 0 0 0 2b MP536 1
239 11 1.5 1 Myrtaceae Myrcia fallax vel sp. aff. 129 1600 an 1 14 0 0 0 4 133 1600 cq 1 11 0 0 0 2b MP564 1
240 11 8 3 Cyatheaceae Cyathea conjugata vel sp. aff. 112 1300 an 1 8 0 0 0 1 111 1300 a 1 10 0 0 0 2b MP539 1
241 11 8.5 3 Melastomataceae Meriania tomentosa 336 1300 cn 1 15 0 0 0 4 345 1300 c 1 10 0 0 0 3b MP509 1
242 11 2.5 3.5 Moraceae Morus insignis 186 1300 an 1 15 0 0 0 4 186 1300 c 1 12 0 0 0 4 MP523 1
243 11 0.5 4 Clethraceae Clethra castaneifolia 204 1300 bzn 1 12 0 0 0 1 0 0 0 bps b=0.5m
244 11 3.5 7 Primulaceae Myrsine indet 100 1300 cn 1 8 0 0 0 2a 101 1300 c 1 9 0 0 0 3a MP557 1
245 11 6 8.5 Elaeocarpaceae Vallea stipularis 140 1500 cn 1 10 0 0 0 2c 142 1500 c 1 10 0 0 0 3b MP576 1
246 11 7 9.5 Euphorbiaceae Alchornea grandis 108 1600 an 1 10 0 0 0 3b 111 1600 e 1 9 0 0 0 2b MP579 1
247 11 0.5 19 Phyllanthaceae Hieronyma oblonga 295 2000 bcn 1 11 0 0 0 3b b=11m 295 2000 ceq 1 11 0 0 0 4 MP525 1
248 11 8.5 19.5 Cyatheaceae Cyathea indet1 139 1600 an 1 7.5 0 0 0 2a 148 1600 a 1 6 0 0 0 2a MP538 1
249 11 17 20 Cyatheaceae Cyathea indet4 137 1300 an 1 8 0 0 0 2a 143 1300 a 1 8 0 0 0 2a MP580 1
250 11 17 17 Cyatheaceae Cyathea indet4 112 1300 an 1 5 0 0 0 2a 113 1300 a 1 6 0 0 0 2b MP561 1
251 11 15 14 Theaceae Gordonia fruticosa 294 2000 qn 1 15 0 0 0 4 296 2000 eq 1 21 0 0 0 5 MP515 1
252 11 19 8 Urticaceae Cecropia tacuna 107 1300 an 1 13 0 0 0 3b 135 1300 a 1 13 0 0 0 4 MP583 1
253 11 15 4 Cyatheaceae Cyathea indet3 246 1900 an 1 14 0 0 0 4 247 1900 a 1 8 0 0 0 3b MP581 1
254 11 17 2 Cyatheaceae Cyathea indet2 156 1300 an 1 3 0 0 0 2a 160 1300 a 1 3 0 0 0 2c MP584 1
246.1 11 6 8 Melastomataceae Miconia indet 117 1300 cn 1 7 0 0 0 3a no colectado 0
255 12 4.5 2 Theaceae Gordonia fruticosa 130 1300 an 1 14 0 0 0 2b 132 1300 a 1 18 0 0 0 4 MP585 1
256 12 5.5 3 Clethraceae Clethra castaneifolia 337 1600 an 1 21 0 0 0 4 356 1600 a 1 18 0 0 0 5 MP550 1
257 12 9 5 Rubiaceae Cinchona pubescens 171 3000 cn 1 15 0 0 0 2b 176 3000 ce 1 14 0 0 0 3b MP577 1

110
5. Fichas RAINFOR de códigos de trabajo de campo y de base de datos.

111

112
6. Procedimiento para el calculo de Biomasa aérea
1. Se ordenó los datos obtenidos en campo en una base
de datos (Excel)
2. Elección de una ecuación alométrica según área de
estudio y/o zona de vida
3. Obtención de densidad de madera por especie,
género o familia
4. Se ordenó los parámetros de análisis de cada
individuo (DAP, Altura y Densidad de Madera)
5. Se aplicacó de la ecuación alométrica en la determinación de Biomasa aérea

113
7. Resumen de tasa de Mortalidad y Reclutamiento
8. Especies reclutas en cada una de las parcelas
RECLUTAS SHM - 01
FAMILIAS ESPECIES INDIVIDUOS
Cunoniaceae Weinmannia pubescens Kunth 1
Melastomataceae Miconia media (D. Don) Naudin 1
Melastomataceae Meriania tomentosa (Cogn.) Wurdack 1
Brunelliaceae Brunellia sp1. Ruiz & Pav. 1
Monimiaceae Mollinedia repanda Ruiz & Pav. 2
Moraceae Morus insignis Bureau 1
Phyllanthaceae Hieronyma oblonga (Tul.) Müll.Arg. 3
Primulaceae Myrsine latifolia (Ruiz & Pav.) Spreng. 2
Rubiaceae Cinchona pubescens Vahl 2
Sabiaceae Meliosma peytonii A.H. Gentry 1
Styracaceae Styrax pentlandianus J. Rémy 1
TOTAL 16
RECLUTAS SHM - 02
FAMILIAS ESPECIES INDIVIDUOS
Asteraceae Baccharis oblongifolia (Ruiz & Pav.) Pers. vel sp. aff. 1
Chloranthaceae Hedyosmum angustifolium (Ruiz & Pav.) Solms 4
Cunoniaceae Weinmannia crassifolia Ruiz & Pav. 14
Elaeocarpaceae Vallea stipularis L.f. 1
Ericaceae Cavendishia bracteata (Ruiz & Pav. ex J.St.Hil.) Hoerold 2
Escalloniaceae Escallonia myrtilloides L.f. 1
Melastomataceae Meriania vargasii Wurdack 1
Miconia indet 1 1
Parcela Vivos
(2013)
Vivos
(2016)
Tie
mp
o
Mu
ert
os
Muertos
/año
Re
clu
tas
Reclutas
/año
Tm
(%)
Tr
(%)
Tre
(%)
SHM01 690 682 3 24 8 16 5 1.18 0.79 0.99
SHM02 589 604 3 21 7 36 12 1.21 2.05 1.63
TOTAL 1279 1286 6 45 15 52 17 2.39 2.84 2.62
PROMEDIO 639.5 643 3 22.5 8 26 9 1.20 1.42 1.31
MAX 690 682 24 8 36 12 1.21 2.05 1.63
MIN 589 604 21 7 16 5 1.18 1.79 1.49
DES.EST 50.5 39 1.5 0.5 10 3.3 0.02 0.63 0.32

114
Primulaceae Myrsine coriacea (Sw.) R.Br. ex Roem. & Schult. 5
Rosaceae Hesperomeles ferruginea (Pers.) Benth. 2
Rosaceae Prunus integrifolia (C. Presl) Walp. 2
Rubiaceae Psychotria indet 1 2
TOTAL 36
9. Tipos de mortalidad de individuos muertos en cada una de las parcelas
SHM - 01
N° Cod Familia Especie F2 Tipos de mortalidad
1 50 Cyatheaceae Cyathea indet bqv Muerto quebrado en una muerte múltiple y fue matado por otro árbol.
2 58 Indet Indet indet cps Muerto desenraizado, murió solo.
3 92 Indet Indet indet bps Muerto quebrado, murió solo.
4 96 Clusiaceae Clusia trochiformis bps Muerto quebrado, murió solo.
5 123 Rubiaceae Psychotria indet cps Muerto desenraizado, murió solo.
6 127 Rubiaceae Cinchona pubescens aps Muerto en pie, murió solo.
7 150 Lauraceae Aniba coto vel sp. aff. aps Muerto en pie, murió solo.
8 190 Melastomataceae Miconia indet cps Muerto desenraizado, murió solo.
9 222 Rubiaceae Cinchona pubescens aps Muerto en pie, murió solo.
10 228 Melastomataceae Miconia saltuensis vel sp. aff.
bps Muerto quebrado, murió solo.
11 243 Clethraceae Clethra castaneifolia bps Muerto quebrado, murió solo.
12 276 Phyllanthaceae Hieronyma oblonga aps Muerto en pie, murió solo
13 279 Melastomataceae Indet indet cps Muerto desenraizado, murió solo.
14 283 Myrtaceae Myrcia fallax vel sp. aff. bqw Muerto quebrado, en un evento múltiple y fue matado por otro.
15 298 Indet Indet indet bps Muerto quebrado, murió solo.
16 331 Indet Indet indet aps Muerto en pie, murió solo.
17 438 Indet Indet indet bps Muerto quebrado, murió solo.
18 527 Indet Indet indet bqw Muerto quebrado, en un evento múltiple y fue matado por otro.
19 528 Indet Indet indet bqw Muerto quebrado, en un evento múltiple y fue matado por otro.
20 560 Rubiaceae Cinchona pubescens aps Muerto en pie, murió solo.
21 592 Indet Indet indet aps Muerto en pie, murió solo.
22 623 Indet Indet indet bps Muerto quebrado, murió solo.
23 649 Indet Indet indet bps Muerto quebrado, murió solo.
24 652 Melastomataceae Indet indet aps Muerto en pie, murió solo.

115
SHM - 02
N° Cod Familia Especie F2 Tipos de mortalidad
1 1 indet Indet indet cps Muerto desenraizado, murió solo.
2 14 indet Indet indet bps Muerto quebrado, murió solo.
3 51 indet Indet indet aps Muerto en pie, murió solo.
4 70 indet Indet indet aps Muerto en pie, murió solo.
5 85 indet Indet indet aps Muerto en pie, murió solo.
6 105 Chloranthaceae
Hedyosmum angustifolium aps Muerto en pie, murió solo.
7 172 indet Indet indet cps Muerto desenraizado, murió solo.
8 179 indet Indet indet u Muerto y matado por otro.
9 186 indet Indet indet aps Muerto en pie, murió solo.
10 187 indet Indet indet cqw
Muerto desenraizado, en un evento múltiple y matado por otro.
11 216 indet Indet indet cps Muerto desenraizado y murió solo
12 239 indet Indet indet aps Muerto en pie y murió solo.
13 323 indet Indet indet aps Muerto en pie y murió solo.
14 336 indet Indet indet bps Muerto quebrado, murió solo.
15 347 Symplocaceae
Symplocos quitensis subsp. boliviensis cps Muerto desenraizado y murió solo
16 351 indet Indet indet cqw
Muerto desenraizado, en un evento múltiple y matado por otro.
17 379 indet Indet indet aps Muerto en pie y murió solo.
18 454 Cunoniaceae Weinmannia indet aps Muerto en pie y murió solo.
19 506 indet Indet indet cps Muerto desenraizado y murió solo
20 541 indet Indet indet cqu
Muerto desenraizado, en un evento múltiple y matado por otro.
21 573 Rubiaceae Psychotria indet 1 bps Muerto quebrado, murió solo.
10. Biomasa aérea y carbono almacenado en cada una de las parcelas evaluadas.
PA
RC
ELA
Bio
mas
a
(20
13
)
Bio
mas
a (2
01
6)
Bio
mas
a p
rom
ed
io
(T
ha-1
)
De
sv. E
st.
Car
bo
no
alm
ace
nad
o
pro
me
dio
(T h
a-1)
Tie
mp
o
Bio
mas
a ca
mb
io
Bio
mas
a p
or
año
Car
bo
no
ca
mb
io
Car
bo
no
cam
bio
po
r añ
o
SHM - 01 144.55 147.41 145.98 1.43 72.99 3 2.85 0.95 1.43 0.48
SHM - 02 99.29 102.82 101.06 1.77 50.53 3 3.53 1.18 1.77 0.59
PROMEDIO 121.92 125.12 123.52 61.76 3 3.19 1.06 1.60 0.53
DESVIACION ESTANDAR 22.46 11.23 0 0.34 0.11 0.17 0.06
INTERVALO DE CONFIANZA, 95% 31.13 15.56 0 0.47 0.16 0.23 0.08

116
11. Biomasa y carbono almacenado en cada censo.
VARIABLES BIOMASA (T ha-1)
SHM - 01 BIOMASA (T ha-1)
SHM - 02
2013 2016 2013 2016
Individuos 690 682 589 604
Total biomasa 144.55 147.41 99.29 102.82
Promedio por individuo 0.21 0.22 0.17 0.17
Desviacion estandar 0.27 0.28 0.22 0.23
Intervalo de confianza (95%) 0.02 0.02 0.02 0.02
Error (%) 9.56 9.72 10.78 10.88
Error (T/ha) 13.83 14.33 10.70 11.19
VARIABLES CARBONO (T C ha-1)
SHM - 01 CARBONO (T C ha-1)
SHM - 02
2013 2016 2013 2016
Individuos 690 682 589 604
Total carbono 72.28 73.70 49.65 51.41
Promedio por individuo 0.10 0.11 0.08 0.09
Desviacion estandar 0.13 0.14 0.11 0.12
Intervalo de confianza (95%) 0.01 0.01 0.01 0.01
Error (%) 9.56 9.72 10.78 10.88
Error(T/ha) 6.91 7.17 5.35 5.59
12. Pruebas de Normalidad con SPSS Statistics 23.0
PRUEBAS DE NORMALIDAD
PARCELA Kolmogorov-Smirnova Shapiro-Wilk
Estadístico gl Sig. Estadístico gl Sig.
SHM_01 DAP .145 682 0.00 .846 682 .000
biomasa .252 682 0.00 .629 682 .000
SHM_02 DAP .158 604 0.00 .819 604 .000
biomasa .273 604 0.00 .552 604 .000
117

118
13. Análisis estadístico de sisgnificancia de biomasa aérea en cada una de las parcelas
con SPSS Statistics 23.0.
ESTADÍSTICOS DE PRUEBA BIOMASA
U de Mann-Whitney 183286.000
W de Wilcoxon 365996.000
Z -3.418
Sig. asintótica (bilateral) 0.001
Sig. Monte Carlo (bilateral)
Sig. ,001b
Intervalo de confianza al 95%
Límite inferior
0.000
Límite superior
0.002
Sig. Monte Carlo (unilateral)
Sig. ,001b
Intervalo de confianza al 95%
Límite inferior
0.000
Límite superior
0.002
14. Biomasa aérea por especies en cada una de las parcelas.
SHM - 01 de Intipunku
Especie Promedio de biomasa
(T ha-1) Promedio de Carbono
(T ha-1)
Total general 145.98 72.99
Aniba coto vel sp. aff. 19.26 9.63
Gordonia fruticosa 19.55 9.77
Myrcia fallax vel sp. aff. 15.82 7.91
Hieronyma oblonga 16.23 8.11
Clethra castaneifolia 13.77 6.89
Meliosma peytonii 13.13 6.56
Myrsine latifolia 9.09 4.55
Mollinedia repanda 6.04 3.02
Meriania tomentosa 5.37 2.69
Clusia trochiformis 2.82 1.41
Cinchona pubescens 2.90 1.45
Alchornea grandis 2.91 1.45
Weinmannia auriculata 2.06 1.03
Ternstroemia jelskii 1.89 0.95
Cyathea conjugata vel sp. aff. 1.57 0.79
PARCELA N Rango
promedio
Suma de
rangos
BIOMASA
SHM_02 604 605.95 365996.00
SHM_01 682 676.75 461545.00
Total 1286
119
Miconia saltuensis vel sp. aff. 1.57 0.79
Myrcianthes rhopaloides 1.22 0.61
Tetrorchidium rubrivenium 1.03 0.52
Miconia media 1.00 0.50
Miconia theaezans vel sp. aff. 0.93 0.46
Styrax pentlandianus 0.78 0.39
Schefflera herzogii 0.68 0.34
Morus insignis 0.70 0.35
Symplocos melanochroa 0.67 0.34
Rudgea ciliata 0.40 0.20
Cyathea indet1 0.37 0.18
Meriania vargasii 0.30 0.15
Cyathea indet3 0.28 0.14
Myrsine coriácea 0.26 0.13
Hedyosmum angustifolium 0.24 0.12
Vallea stipularis 0.21 0.11
Cyathea delgadii vel sp. aff. 0.21 0.11
Guettarda crispiflora 0.14 0.07
Cyathea indet4 0.11 0.05
Viburnum hallii 0.11 0.05
Weinmannia pubescens 0.13 0.07
Nectandra reticulata vel sp. aff. 0.11 0.06
Palicourea sp1 0.10 0.05
Cyathea indet2 0.09 0.05
Oreopanax capitatus 0.06 0.03
Cecropia tacuna 0.07 0.04
Sapium glandulosum 0.06 0.03
Ceroxylon pityrophyllum 0.05 0.03
Staphylea occidentalis 0.04 0.02
Brunellia sp1 0.03 0.01

120
SHM - 02 de Kantupata
Especie Promedio de biomasa
(T ha-1) Promedio de Carbono
(T ha-1)
Total general 101.06 50.53
Weinmannia crassifolia 32.78 16.39
Escallonia myrtilloides 13.51 6.75
Ilex andicola 12.24 6.12
Hedyosmum angustifolium 8.94 4.47
Weinmannia bangii 6.12 3.06
Hesperomeles ferruginea 5.01 2.50
Clethra ferruginea 4.45 2.22
Myrsine coriácea 2.70 1.35
Schefflera herzogii 2.07 1.04
Clusia trochiformis 1.99 0.99
Meriania vargasii 1.42 0.71
Alnus acuminata 1.40 0.70
Prunus integrifolia 1.32 0.66
Vallea stipularis 0.93 0.46
Psychotria indet 1 0.87 0.44
Nectandra indet 2 0.67 0.34
Symplocos quitensis subsp. boliviensis 0.66 0.33
Miconia indet 1 0.60 0.30
Berberis commutata 0.34 0.17
Miconia indet 2 0.34 0.17
Roupala indet 0.22 0.11
Cavendishia bracteata 0.20 0.10
Cyathea indet 0.12 0.06
Ocotea indet 1 0.11 0.05
Oreopanax cuspidatus 0.09 0.05
Cestrum conglomeratum 0.08 0.04
Nectandra indet 1 0.07 0.03
Gaultheria reticulata 0.04 0.02
Meliosma peytonii 0.03 0.02
Baccharis brachylaenoides vel sp. aff. 0.03 0.01
Solanum maturecalvans vel sp. aff. 0.03 0.01
15. Biomasa aérea almacenada de muertos y reclutas.
Par
cela
Ind
ivid
uo
s
(20
13
)
Bio
mas
a 2
01
3
(t h
a-1)
Mu
ert
os
Bio
mas
a d
e
mu
ert
os
(t h
a-1)
Sob
revi
vi
en
tes
Re
clu
tas
Bio
mas
a d
e r
ecl
uta
s
(t h
a-1)
Ind
ivid
uo
s 2
01
6
Bio
mas
a 2
01
6
(t h
a-1)
HSM - 01 690 144.55 24 4.81 666 16 0.95 682 147.41
SHM - 02 589 99.29 21 3.87 568 36 1.92 604 102.82
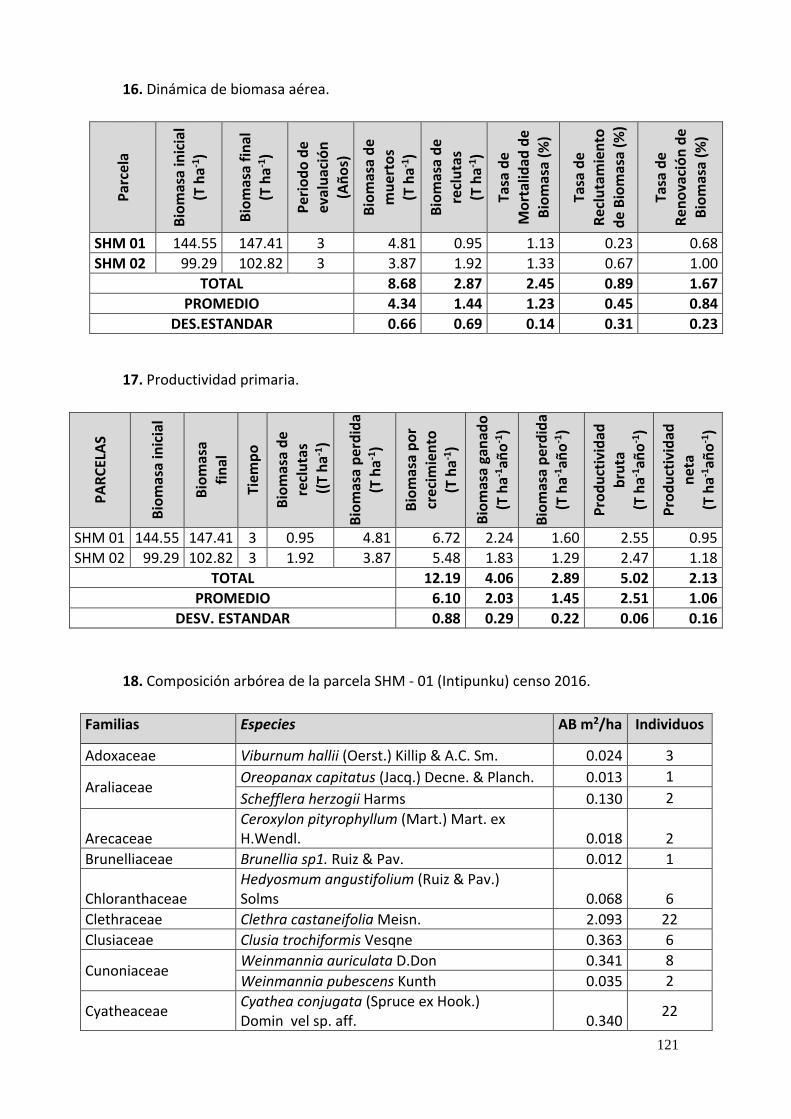
121
16. Dinámica de biomasa aérea.
Par
cela
Bio
mas
a in
icia
l (T
ha-1
)
Bio
mas
a fi
nal
(T
ha-1
)
Pe
rio
do
de
e
valu
ació
n
(Añ
os)
Bio
mas
a d
e
mu
ert
os
(T
ha-1
)
Bio
mas
a d
e
recl
uta
s (T
ha-1
)
Tasa
de
Mo
rtal
idad
de
B
iom
asa
(%)
Tasa
de
Re
clu
tam
ien
to
de
Bio
mas
a (%
)
Tasa
de
R
en
ova
ció
n d
e
Bio
mas
a (%
)
SHM 01 144.55 147.41 3 4.81 0.95 1.13 0.23 0.68
SHM 02 99.29 102.82 3 3.87 1.92 1.33 0.67 1.00
TOTAL 8.68 2.87 2.45 0.89 1.67
PROMEDIO 4.34 1.44 1.23 0.45 0.84
DES.ESTANDAR 0.66 0.69 0.14 0.31 0.23
17. Productividad primaria.
PA
RC
ELA
S
Bio
mas
a in
icia
l
Bio
mas
a
fin
al
Tie
mp
o
Bio
mas
a d
e
recl
uta
s
((T
ha-1
)
Bio
mas
a p
erd
ida
(T h
a-1)
Bio
mas
a p
or
cre
cim
ien
to
(T h
a-1)
Bio
mas
a ga
nad
o
(T h
a-1añ
o-1
)
Bio
mas
a p
erd
ida
(T h
a-1añ
o-1
)
Pro
du
ctiv
idad
bru
ta
(T h
a-1añ
o-1
)
Pro
du
ctiv
idad
n
eta
(T
ha-1
año
-1)
SHM 01 144.55 147.41 3 0.95 4.81 6.72 2.24 1.60 2.55 0.95
SHM 02 99.29 102.82 3 1.92 3.87 5.48 1.83 1.29 2.47 1.18
TOTAL 12.19 4.06 2.89 5.02 2.13
PROMEDIO 6.10 2.03 1.45 2.51 1.06
DESV. ESTANDAR 0.88 0.29 0.22 0.06 0.16
18. Composición arbórea de la parcela SHM - 01 (Intipunku) censo 2016.
Familias Especies AB m2/ha Individuos
Adoxaceae Viburnum hallii (Oerst.) Killip & A.C. Sm. 0.024 3
Araliaceae Oreopanax capitatus (Jacq.) Decne. & Planch. 0.013 1
Schefflera herzogii Harms 0.130 2
Arecaceae Ceroxylon pityrophyllum (Mart.) Mart. ex H.Wendl. 0.018 2
Brunelliaceae Brunellia sp1. Ruiz & Pav. 0.012 1
Chloranthaceae Hedyosmum angustifolium (Ruiz & Pav.) Solms 0.068 6
Clethraceae Clethra castaneifolia Meisn. 2.093 22
Clusiaceae Clusia trochiformis Vesqne 0.363 6
Cunoniaceae Weinmannia auriculata D.Don 0.341 8
Weinmannia pubescens Kunth 0.035 2
Cyatheaceae Cyathea conjugata (Spruce ex Hook.) Domin vel sp. aff. 0.340
22
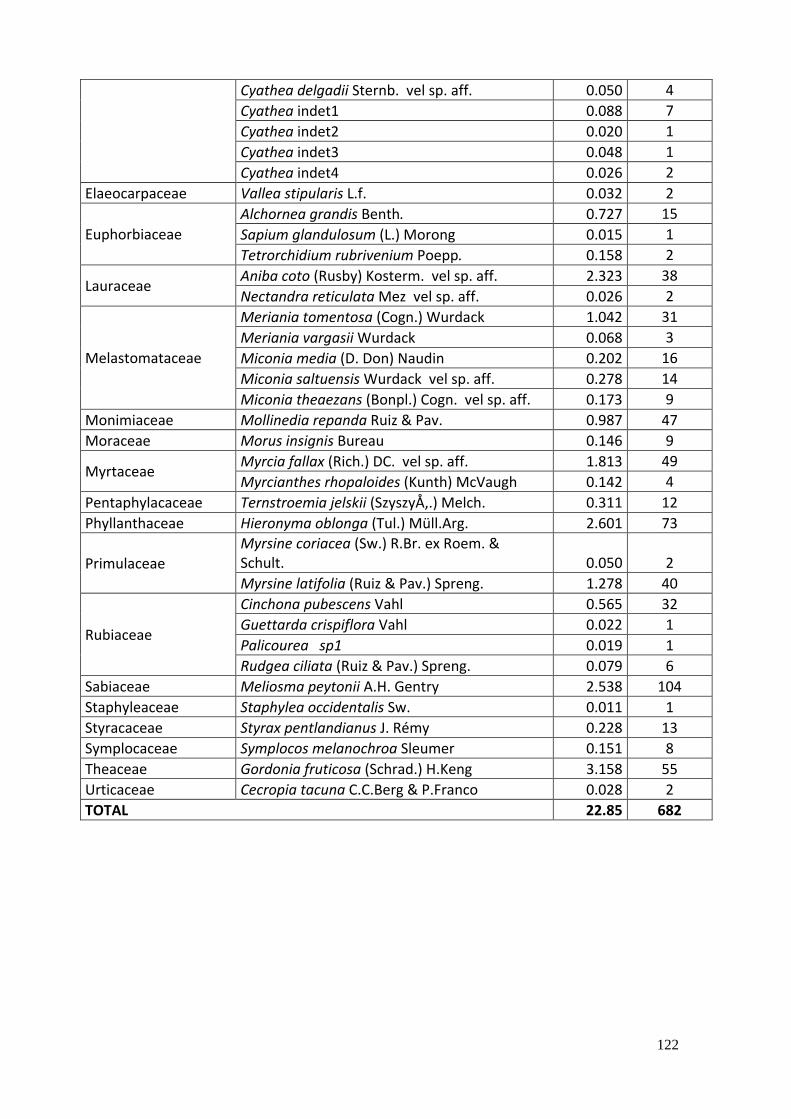
122
Cyathea delgadii Sternb. vel sp. aff. 0.050 4
Cyathea indet1 0.088 7
Cyathea indet2 0.020 1
Cyathea indet3 0.048 1
Cyathea indet4 0.026 2
Elaeocarpaceae Vallea stipularis L.f. 0.032 2
Euphorbiaceae
Alchornea grandis Benth. 0.727 15
Sapium glandulosum (L.) Morong 0.015 1
Tetrorchidium rubrivenium Poepp. 0.158 2
Lauraceae Aniba coto (Rusby) Kosterm. vel sp. aff. 2.323 38
Nectandra reticulata Mez vel sp. aff. 0.026 2
Melastomataceae
Meriania tomentosa (Cogn.) Wurdack 1.042 31
Meriania vargasii Wurdack 0.068 3
Miconia media (D. Don) Naudin 0.202 16
Miconia saltuensis Wurdack vel sp. aff. 0.278 14
Miconia theaezans (Bonpl.) Cogn. vel sp. aff. 0.173 9
Monimiaceae Mollinedia repanda Ruiz & Pav. 0.987 47
Moraceae Morus insignis Bureau 0.146 9
Myrtaceae Myrcia fallax (Rich.) DC. vel sp. aff. 1.813 49
Myrcianthes rhopaloides (Kunth) McVaugh 0.142 4
Pentaphylacaceae Ternstroemia jelskii (Szyszył.) Melch. 0.311 12
Phyllanthaceae Hieronyma oblonga (Tul.) Müll.Arg. 2.601 73
Primulaceae
Myrsine coriacea (Sw.) R.Br. ex Roem. & Schult. 0.050 2
Myrsine latifolia (Ruiz & Pav.) Spreng. 1.278 40
Rubiaceae
Cinchona pubescens Vahl 0.565 32
Guettarda crispiflora Vahl 0.022 1
Palicourea sp1 0.019 1
Rudgea ciliata (Ruiz & Pav.) Spreng. 0.079 6
Sabiaceae Meliosma peytonii A.H. Gentry 2.538 104
Staphyleaceae Staphylea occidentalis Sw. 0.011 1
Styracaceae Styrax pentlandianus J. Rémy 0.228 13
Symplocaceae Symplocos melanochroa Sleumer 0.151 8
Theaceae Gordonia fruticosa (Schrad.) H.Keng 3.158 55
Urticaceae Cecropia tacuna C.C.Berg & P.Franco 0.028 2
TOTAL 22.85 682

123
19. Composición arbórea de la parcela SHM - 02 (Kantupata) censo 2016.
Familias especies AB m2/ha Individuos
Aquifoliaceae Ilex andicola Loes. 2.150 76
Araliaceae
Oreopanax cuspidatus Harms 0.019 1
Schefflera herzogii Harms 0.412 10
Asteraceae Baccharis oblongifolia (Ruiz & Pav.) Pers. vel sp. aff. 0.012 1
Berberidaceae Berberis commutata Eichler 0.062 3
Betulaceae Alnus acuminata Kunth 0.249 1
Chloranthaceae Hedyosmum angustifolium (Ruiz & Pav.) Solms 1.955 85
Clethraceae Clethra ferruginea (Ruiz & Pav.) Link ex Spreng. 0.850 23
Clusiaceae Clusia trochiformis Vesqne 0.237 4
Cunoniaceae
Weinmannia bangii Rusby 0.999 18
Weinmannia crassifolia Ruiz & Pav. 5.805 162
Cyatheaceae Cyathea indet 0.024 1
Elaeocarpaceae Vallea stipularis L.f. 0.146 9
Ericaceae
Cavendishia bracteata (Ruiz & Pav. ex J.St.Hil.) Hoerold 0.051 5
Gaultheria reticulata Kunth 0.009 1
Escalloniaceae Escallonia myrtilloides L.f. 1.935 58
Lauraceae
Nectandra indet 1 0.014 1
Nectandra indet 2 0.117 5
Ocotea indet 1 0.022 1
Melastomataceae
Meriania vargasii Wurdack 0.296 11
Miconia indet 1 0.124 9
Miconia indet 2 0.064 4
Primulaceae Myrsine coriacea (Sw.) R.Br. ex Roem. & Schult. 0.584 46
Proteaceae Roupala indet 0.036 2
Rosaceae
Hesperomeles ferruginea (Pers.) Benth. 0.720 28
Prunus integrifolia (C. Presl) Walp. 0.215 13
Rubiaceae Psychotria indet 1 0.205 17
Sabiaceae Meliosma peytonii A.H. Gentry 0.009 1
Solanaceae
Cestrum conglomeratum Ruiz & Pav. 0.023 2
Solanum maturecalvans Bitter vel sp. aff. 0.008 1
Symplocaceae Symplocos quitensis subsp. Boliviensis Brand 0.105 5
TOTAL 17.46 604

124
20. Índices de Valor de Importancia en la parcela SHM – 01
Por especies en SHM - 01
ESPECIES Abu.
Absoluta Abu. Relativa
Frec.
Absoluta Frec. Relativa
Dom.
Absoluta Dom. Relativa IVI’s al 300% IVI’s al 100%
Meliosma peytonii 104 15.25 24.0 7.14 2.54 11.11 33.50 11.17
Hieronyma oblonga 73 10.70 24.0 7.14 2.60 11.38 29.23 9.74
Gordonia fruticosa 55 8.06 22.0 6.55 3.16 13.82 28.43 9.48
Myrcia fallax 49 7.18 22.0 6.55 1.81 7.93 21.67 7.22
Aniba coto 38 5.57 18.0 5.36 2.32 10.16 21.09 7.03
Mollinedia repanda 47 6.89 22.0 6.55 0.99 4.32 17.76 5.92
Myrsine latifolia 40 5.87 19.0 5.65 1.28 5.59 17.11 5.70
Clethra castaneifolia 22 3.23 14.0 4.17 2.09 9.16 16.55 5.52
Meriania tomentosa 31 4.55 15.0 4.46 1.04 4.56 13.57 4.52
Cinchona pubescens 32 4.69 14.0 4.17 0.57 2.47 11.33 3.78
Alchornea grandis Benth. 15 2.20 12.0 3.57 0.73 3.18 8.95 2.98
Cyathea conjugata 22 3.23 10.0 2.98 0.34 1.49 7.69 2.56
Miconia saltuensis 14 2.05 11.0 3.27 0.28 1.22 6.54 2.18
Miconia media 16 2.35 11.0 3.27 0.20 0.88 6.50 2.17
Ternstroemia jelskii 12 1.76 10.0 2.98 0.31 1.36 6.10 2.03
Styrax pentlandianus 13 1.91 7.0 2.08 0.23 1.00 4.99 1.66
Weinmannia auriculata 8 1.17 6.0 1.79 0.34 1.49 4.45 1.48
Clusia trochiformis 6 0.88 6.0 1.79 0.36 1.59 4.26 1.42
Miconia theaezans 9 1.32 7.0 2.08 0.17 0.76 4.16 1.39
Morus insignis 9 1.32 6.0 1.79 0.15 0.64 3.74 1.25
Symplocos melanochroa 8 1.17 6.0 1.79 0.15 0.66 3.62 1.21
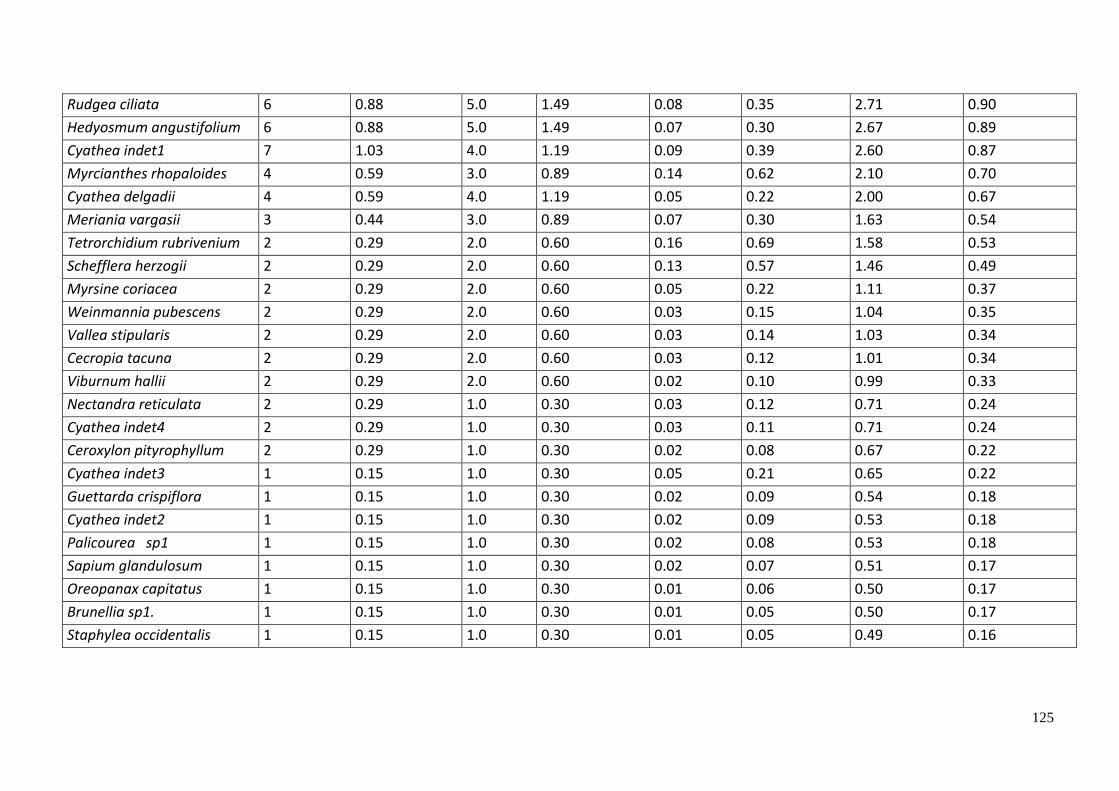
125
Rudgea ciliata 6 0.88 5.0 1.49 0.08 0.35 2.71 0.90
Hedyosmum angustifolium 6 0.88 5.0 1.49 0.07 0.30 2.67 0.89
Cyathea indet1 7 1.03 4.0 1.19 0.09 0.39 2.60 0.87
Myrcianthes rhopaloides 4 0.59 3.0 0.89 0.14 0.62 2.10 0.70
Cyathea delgadii 4 0.59 4.0 1.19 0.05 0.22 2.00 0.67
Meriania vargasii 3 0.44 3.0 0.89 0.07 0.30 1.63 0.54
Tetrorchidium rubrivenium 2 0.29 2.0 0.60 0.16 0.69 1.58 0.53
Schefflera herzogii 2 0.29 2.0 0.60 0.13 0.57 1.46 0.49
Myrsine coriacea 2 0.29 2.0 0.60 0.05 0.22 1.11 0.37
Weinmannia pubescens 2 0.29 2.0 0.60 0.03 0.15 1.04 0.35
Vallea stipularis 2 0.29 2.0 0.60 0.03 0.14 1.03 0.34
Cecropia tacuna 2 0.29 2.0 0.60 0.03 0.12 1.01 0.34
Viburnum hallii 2 0.29 2.0 0.60 0.02 0.10 0.99 0.33
Nectandra reticulata 2 0.29 1.0 0.30 0.03 0.12 0.71 0.24
Cyathea indet4 2 0.29 1.0 0.30 0.03 0.11 0.71 0.24
Ceroxylon pityrophyllum 2 0.29 1.0 0.30 0.02 0.08 0.67 0.22
Cyathea indet3 1 0.15 1.0 0.30 0.05 0.21 0.65 0.22
Guettarda crispiflora 1 0.15 1.0 0.30 0.02 0.09 0.54 0.18
Cyathea indet2 1 0.15 1.0 0.30 0.02 0.09 0.53 0.18
Palicourea sp1 1 0.15 1.0 0.30 0.02 0.08 0.53 0.18
Sapium glandulosum 1 0.15 1.0 0.30 0.02 0.07 0.51 0.17
Oreopanax capitatus 1 0.15 1.0 0.30 0.01 0.06 0.50 0.17
Brunellia sp1. 1 0.15 1.0 0.30 0.01 0.05 0.50 0.17
Staphylea occidentalis 1 0.15 1.0 0.30 0.01 0.05 0.49 0.16

126
Por familias en SHM - 01
FAMILIAS Abu. absoluta Abu. Relativa Dom. Absoluta Dom. Relativa Div. Absoluta Div. Relativa IVIF al 300% IVIF al 100%
Melastomataceae 73.00 10.70 1.76 7.72 5.00 10.87 29.29 9.76
Sabiaceae 104.00 15.25 2.54 11.11 1.00 2.17 28.53 9.51
Phyllanthaceae 73.00 10.70 2.60 11.38 1.00 2.17 24.26 8.09
Theaceae 55.00 8.06 3.16 13.82 1.00 2.17 24.06 8.02
Cyatheaceae 37.00 5.43 0.57 2.51 6.00 13.04 20.97 6.99
Myrtaceae 53.00 7.77 1.96 8.56 2.00 4.35 20.68 6.89
Lauraceae 40.00 5.87 2.35 10.28 2.00 4.35 20.49 6.83
Rubiaceae 40.00 5.87 0.69 3.00 4.00 8.70 17.56 5.85
Primulaceae 42.00 6.16 1.33 5.81 2.00 4.35 16.32 5.44
Clethraceae 22.00 3.23 2.09 9.16 1.00 2.17 14.56 4.85
Monimiaceae 47.00 6.89 0.99 4.32 1.00 2.17 13.38 4.46
Euphorbiaceae 18.00 2.64 0.90 3.94 3.00 6.52 13.10 4.37
Cunoniaceae 10.00 1.47 0.38 1.64 2.00 4.35 7.46 2.49
Araliaceae 3.00 0.44 0.14 0.63 2.00 4.35 5.41 1.80
Pentaphylacaceae 12.00 1.76 0.31 1.36 1.00 2.17 5.30 1.77
Styracaceae 13.00 1.91 0.23 1.00 1.00 2.17 5.08 1.69
Clusiaceae 6.00 0.88 0.36 1.59 1.00 2.17 4.64 1.55
Moraceae 9.00 1.32 0.15 0.64 1.00 2.17 4.13 1.38
Symplocaceae 8.00 1.17 0.15 0.66 1.00 2.17 4.01 1.34
Chloranthaceae 6.00 0.88 0.07 0.30 1.00 2.17 3.35 1.12
Elaeocarpaceae 2.00 0.29 0.03 0.14 1.00 2.17 2.61 0.87
Urticaceae 2.00 0.29 0.03 0.12 1.00 2.17 2.59 0.86
Adoxaceae 2.00 0.29 0.02 0.10 1.00 2.17 2.57 0.86
Arecaceae 2.00 0.29 0.02 0.08 1.00 2.17 2.55 0.85
Brunelliaceae 1.00 0.15 0.01 0.05 1.00 2.17 2.37 0.79
Staphyleaceae 1.00 0.15 0.01 0.05 1.00 2.17 2.37 0.79

127
21. Índices de Valor de Importancia en la parcela SHM – 02
Por especies en SHM - 02
ESPECIES Abu.
Absoluta Abu. Relativa
Frec.
Absoluta Frec. Relativa
Dom.
Absoluta Dom. Relativa IVI's al 300% IVI’s al 100%
Weinmannia crassifolia 162 26.82 23 10.75 5.80 33.25 70.82 23.61
Ilex andicola 76 12.58 23 10.75 2.15 12.32 35.65 11.88
Hedyosmum angustifolium 85 14.07 21 9.81 1.96 11.20 35.09 11.70
Escallonia myrtilloides 58 9.60 18 8.41 1.94 11.08 29.10 9.70
Myrsine coriacea 46 7.62 21 9.81 0.58 3.35 20.78 6.93
Hesperomeles ferruginea 28 4.64 14 6.54 0.72 4.13 15.30 5.10
Clethra ferruginea 23 3.81 10 4.67 0.85 4.87 13.35 4.45
Weinmannia bangii 18 2.98 8 3.74 1.00 5.73 12.44 4.15
Prunus integrifolia 13 2.15 11 5.14 0.22 1.23 8.53 2.84
Psychotria indet 1 17 2.81 9 4.21 0.21 1.17 8.19 2.73
Schefflera herzogii 10 1.66 6 2.80 0.41 2.36 6.82 2.27
Meriania vargasii 11 1.82 6 2.80 0.30 1.69 6.32 2.11
Miconia indet 1 9 1.49 8 3.74 0.12 0.71 5.94 1.98
Vallea stipularis 9 1.49 4 1.87 0.15 0.83 4.19 1.40
Symplocos quitensis subsp. boliviensis 5 0.83 5 2.34 0.10 0.60 3.76 1.25
Clusia trochiformis 4 0.66 3 1.40 0.24 1.36 3.42 1.14
Nectandra indet 2 5 0.83 4 1.87 0.12 0.67 3.37 1.12
Cavendishia bracteata 5 0.83 4 1.87 0.05 0.29 2.99 1.00
Miconia indet 2 4 0.66 4 1.87 0.06 0.37 2.90 0.97
Alnus acuminata 1 0.17 1 0.47 0.25 1.43 2.06 0.69
Berberis commutata 3 0.50 1 0.47 0.06 0.36 1.32 0.44
128
Roupala indet 2 0.33 1 0.47 0.04 0.21 1.01 0.34
Cestrum conglomeratum 2 0.33 1 0.47 0.02 0.13 0.93 0.31
Cyathea indet 1 0.17 1 0.47 0.02 0.14 0.77 0.26
Ocotea indet 1 1 0.17 1 0.47 0.02 0.13 0.76 0.25
Oreopanax cuspidatus 1 0.17 1 0.47 0.02 0.11 0.74 0.25
Nectandra indet 1 1 0.17 1 0.47 0.01 0.08 0.71 0.24
Baccharis brachylaenoides vel sp. aff. 1 0.17 1 0.47 0.01 0.07 0.70 0.23
Gaultheria reticulata 1 0.17 1 0.47 0.01 0.05 0.68 0.23
Meliosma peytonii 1 0.17 1 0.47 0.01 0.05 0.68 0.23
Solanum maturecalvans vel sp. aff. 1 0.17 1 0.47 0.01 0.05 0.68 0.23

129
Por familias en SHM - 02
FAMILIAS Abu. Absoluta Abu. Relativa Dom. Absoluta Dom. Relativa Div. Absoluta Div. Relativa IVIF al 300% IVIF al 100%
Cunoniaceae 180 29.80 6.80 38.98 2 6.45 75.23 25.08
Chloranthaceae 85 14.07 1.96 11.20 1 3.23 28.50 9.50
Aquifoliaceae 76 12.58 2.15 12.32 1 3.23 28.12 9.37
Escalloniaceae 58 9.60 1.94 11.08 1 3.23 23.91 7.97
Rosaceae 41 6.79 0.94 5.36 2 6.45 18.60 6.20
Melastomataceae 24 3.97 0.48 2.77 3 9.68 16.42 5.47
Primulaceae 46 7.62 0.58 3.35 1 3.23 14.19 4.73
Clethraceae 23 3.81 0.85 4.87 1 3.23 11.90 3.97
Lauraceae 7 1.16 0.15 0.88 3 9.68 11.71 3.90
Araliaceae 11 1.82 0.43 2.47 2 6.45 10.74 3.58
Ericaceae 6 0.99 0.06 0.35 2 6.45 7.79 2.60
Rubiaceae 17 2.81 0.21 1.17 1 3.23 7.21 2.40
Solanaceae 3 0.50 0.03 0.18 2 6.45 7.13 2.38
Elaeocarpaceae 9 1.49 0.15 0.83 1 3.23 5.55 1.85
Clusiaceae 4 0.66 0.24 1.36 1 3.23 5.25 1.75
Betulaceae 1 0.17 0.25 1.43 1 3.23 4.82 1.61
Symplocaceae 5 0.83 0.10 0.60 1 3.23 4.65 1.55
Berberidaceae 3 0.50 0.06 0.36 1 3.23 4.08 1.36
Proteaceae 2 0.33 0.04 0.21 1 3.23 3.76 1.25
Cyatheaceae 1 0.17 0.02 0.14 1 3.23 3.53 1.18
Asteraceae 1 0.17 0.01 0.07 1 3.23 3.46 1.15
Sabiaceae 1 0.17 0.01 0.05 1 3.23 3.44 1.15

130
22. Índices de diversidad alfa (α) en cada parcela evaluada de Wiñaywayna
Indices Alfa SHM_01 SHM_02 TOTAL
Taxa S 46 31 70
Individuals 682 604 1286
Dominance D 0.0673 0.1298 0.04852
Shannon H 3.065 2.477 3.433
Simpson 1-D 0.9327 0.8702 0.9515
Evenness e^H/S 0.4662 0.384 0.4424
Menhinick 1.761 1.261 1.952
Margalef 6.897 4.685 9.638
Equitability J 0.8007 0.7213 0.8081
Fisher alpha 11.13 6.918 15.89
Berger-Parker 0.1525 0.2682 0.126
23. Datos de análisis de los índices de diversidad beta (β) en parcelas evaluadas de
Wiñaywayna.
Familias Especies SHM - 01 SHM - 02
Euphorbiaceae Alchornea grandis Benth. 15 0
Betulaceae Alnus acuminata Kunth 0 1
Lauraceae Aniba coto (Rusby) Kosterm. vel sp. aff. 38 0
Asteraceae Baccharis oblongifolia (Ruiz & Pav.) Pers. vel sp. aff. 0 1
Berberidaceae Berberis commutata Eichler 0 3
Brunelliaceae Brunellia sp1. Ruiz & Pav. 1 0
Ericaceae Cavendishia bracteata (Ruiz & Pav. ex J.St.Hil.) Hoerold 0 5
Urticaceae Cecropia tacuna C.C.Berg & P.Franco 2 0
Arecaceae Ceroxylon pityrophyllum (Mart.) Mart. ex H.Wendl. 2 0
Solanaceae Cestrum conglomeratum Ruiz & Pav. 0 2
Rubiaceae Cinchona pubescens Vahl 32 0
Clethraceae Clethra castaneifolia Meisn. 22 0
Clethraceae Clethra ferruginea (Ruiz & Pav.) Link ex Spreng. 0 23
Clusiaceae Clusia trochiformis Vesqne 6 4
Cyatheaceae Cyathea conjugata (Spruce ex Hook.) Domin vel sp. aff. 22 0
Cyatheaceae Cyathea delgadii Sternb. vel sp. aff. 4 0
Cyatheaceae Cyathea indet 0 1
Cyatheaceae Cyathea indet1 7 0
Cyatheaceae Cyathea indet2 1 0
Cyatheaceae Cyathea indet3 1 0
Cyatheaceae Cyathea indet4 2 0
Escalloniaceae Escallonia myrtilloides L.f. 0 58
Ericaceae Gaultheria reticulata Kunth 0 1
Theaceae Gordonia fruticosa (Schrad.) H.Keng 55 0

131
Rubiaceae Guettarda crispiflora Vahl 1 0
Chloranthaceae Hedyosmum angustifolium (Ruiz & Pav.) Solms 6 85
Rosaceae Hesperomeles ferruginea (Pers.) Benth. 0 28
Phyllanthaceae Hieronyma oblonga (Tul.) Müll.Arg. 73 0
Aquifoliaceae Ilex andicola Loes. 0 76
Sabiaceae Meliosma peytonii A.H. Gentry 104 1
Melastomataceae Meriania tomentosa (Cogn.) Wurdack 31 0
Melastomataceae Meriania vargasii Wurdack 3 11
Melastomataceae Miconia indet 1 0 9
Melastomataceae Miconia indet 2 0 4
Melastomataceae Miconia media (D. Don) Naudin 16 0
Melastomataceae Miconia saltuensis Wurdack vel sp. aff. 14 0
Melastomataceae Miconia theaezans (Bonpl.) Cogn. vel sp. aff. 9 0
Monimiaceae Mollinedia repanda Ruiz & Pav. 47 0
Moraceae Morus insignis Bureau 9 0
Myrtaceae Myrcia fallax (Rich.) DC. vel sp. aff. 49 0
Myrtaceae Myrcianthes rhopaloides (Kunth) McVaugh 4 0
Primulaceae Myrsine coriacea (Sw.) R.Br. ex Roem. & Schult. 2 46
Primulaceae Myrsine latifolia (Ruiz & Pav.) Spreng. 40 0
Lauraceae Nectandra indet 1 0 1
Lauraceae Nectandra indet 2 0 5
Lauraceae Nectandra reticulata Mez vel sp. aff. 2 0
Lauraceae Ocotea indet 1 0 1
Araliaceae Oreopanax capitatus (Jacq.) Decne. & Planch. 1 0
Araliaceae Oreopanax cuspidatus Harms 0 1
Rubiaceae Palicourea indet 1 1 0
Rosaceae Prunus integrifolia (C. Presl) Walp. 0 13
Rubiaceae Psychotria indet 1 0 17
Proteaceae Roupala indet 0 2
Rubiaceae Rudgea ciliata (Ruiz & Pav.) Spreng. 6 0
Euphorbiaceae Sapium glandulosum (L.) Morong 1 0
Araliaceae Schefflera herzogii Harms 2 10
Solanaceae Solanum maturecalvans Bitter vel sp. aff. 0 1
Staphyleaceae Staphylea occidentalis Sw. 1 0
Styracaceae Styrax pentlandianus J. Rémy 13 0
Symplocaceae Symplocos melanochroa Sleumer 8 0
Symplocaceae Symplocos quitensis subsp. Boliviensis Brand 0 5
Pentaphylacaceae Ternstroemia jelskii (Szyszył.) Melch. 12 0
Euphorbiaceae Tetrorchidium rubrivenium Poepp. 2 0
Elaeocarpaceae Vallea stipularis L.f. 2 9
Adoxaceae Viburnum hallii (Oerst.) Killip & A.C. Sm. 2 0
Cunoniaceae Weinmannia auriculata D.Don 8 0
Cunoniaceae Weinmannia bangii Rusby 0 18
Cunoniaceae Weinmannia crassifolia Ruiz & Pav. 0 162
Cunoniaceae Weinmannia pubescens Kunth 2 0
Total general 682 604

132
24. Fotografías de especies arbóreas registradas en las parcelas evaluadas de Wiñaywayna.
1 Ilex andicola
AQUIFOLIACEAE
2
Ilex andicola AQUIFOLIACEAE
3
Schefflera herzogii
ARALIACEAE
4
Brunellia sp BRUNELLIACEAE
5
Weinmannia crassifolia CUNONIACEAE
6
Hedyosmum angustifolium CLHORANTACEAE
7
Hedyosmum angustifolium CLHORANTACEAE
8
Clethra ferruginea CLETHRACEAE
9
Vallea stipularis ELAEOCARPACEAE
10
Escallonia myrtillioides ESCALLONIACEAE
10a
Escallonia myrtillioides ESCALLONIACEAE
11
Cavendishia bracteata ERICACEAE
11a
Cavendishia bracteata ERICACEAE
12
Aniba coto LAURACEAE
12a
Aniba coto LAURACEAE
13
Meriania vargasii MELASTOMATACEAE
14
Mollinedia repanda MONIMIACEAE
14a
Mollinedia repanda MONIMIACEAE
15
Ternstroemia jelskii PENTAPHYLACACEAE
16
Myrsine coriacea PRIMULACEAE

133
16a
Myrsine coriacea PRIMULACEAE
17
Myrsine coriacea PRIMULACEAE
18
Prunus integrifolia ROSACEAE
19 Hesperomeles ferruginea ROSACEAE
20 Rudgea ciliata RUBIACEAE
20a
Rudgea ciliata RUBIACEAE
21
Guettarda crispiflora Vahl RUBIACEAE
21a Guettarda crispiflora Vahl RUBIACEAE
22 Psychotria indet 1 RUBIACEAE
23 Meliosma peytonii SABIACEAE
23a
Meliosma peytonii SABIACEAE
24 Symplocos quitensis subsp boliviensis
SYMPLOCACEAE 24a Symplocos quitensis subsp boliviensis
SYMPLOCACEAE 25 Solanum maturecalvans
SOLANACEAE 25a Solanum maturecalvans
SOLANACEAE
26 Cestrum conglomeratum
SOLANACEAE 26a Cestrum conglomeratum
SOLANACEAE 27 Styrax pentlandianus
STYRACACEAE 27a Styrax pentlandianus
STYRACACEAE 28 Gordonia fruticosa
THEACEAE

134