grupo 8 rumiantes menores - alpa.org.ve 15 supl/s_rumiantesmenores.pdf · estómago de los corderos...
TRANSCRIPT

278 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Rumiantes menores
Grupo 8

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•279
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
RESUMEnLa inanición ha sido identificada como la causa más importante de muerte neonatal de corderos en Uruguay (Duran del Campo 1963; Mari, 1979) así como en otros países donde la producción ovina es uno de sus principales rubros (Alexander, 1984). La muerte por inanición puede ser consecuencia de una serie de factores que pueden o no interactuar entre si. Dentro de ellos se encuentran la falta de vigor del cordero recién nacido, falla de la relación madre-hijo, mal comportamiento materno con abandono del cordero por parte de borregas u ovejas con partos laboriosos y prolongados y falta de calostro al momento del parto. Casi todos estos factores se deben a una inadecuada nutrición de la oveja durante el periodo preparto. En éste, la madre tiene altísimos requerimientos para el desarrollo fetal y la glándula mamaria y para la síntesis de calostro. Sin embargo y a pesar del incremento en la demanda de nutrientes, el consumo voluntario de la oveja, sobretodo de forraje, generalmente disminuye durante la última semana de gestación agravando el déficit energético. Esto es aún más grave en condiciones de pastoreo extensivo como las que se registran en los países ovejeros del hemisferio Sur. Una suplementación estratégica de corta duración previo al parto permite revertir el problema ya que las ovejas suplementadas con concentrados energéticos en los últimos días de gestación duplican y hasta triplican la producción de calostro, tienen un mejor comportamiento maternal al parto, el parto puede ser más rápido y en consecuencia sus corderos tienen una mayor sobrevivencia respecto a ovejas alimentadas sólo con forraje.
IntRODUCCIónLa sobrevivencia neonatal de corderos depende de una exitosa interacción entre la madre y su cría. Esta interacción permite a la oveja identificar a su cría y a la cría identificar a su madre. Sin embargo, la creación de este vínculo madre-cría no es suficiente. El cordero necesita además un suministro adecuado de calostro en las primeras horas de vida (Nowak, 1996). La oveja produce calostro durante varias horas
alternativas de maneJo nutriCional para meJorar la supervivenCia de Corderos neonatos
Georgget Banchero*
luego del parto pero el calostro disponible al parto es el mas importante para cubrir los requerimientos de inmunoglobulinas del cordero (Pattinson, 1995). Esto se debe a que la permeabilidad del intestino del cordero, que normalmente es permeable a las macromoléculas de inmunoglobulinas durante las primeras 24 horas de vida, comienza a decrecer a partir de las 6 horas de haber nacido. El calostro también provee al cordero con energía y agua. Los corderos nacen con sus propias reservas de energía (tejido adiposo marrón, glucógeno) pero ésta es limitada y necesita ser reemplazada cuanto antes por otra fuente de energía. Por último, el hecho de que el cordero o se amamante de su madre cuanto antes, también es importante para el establecimiento del vínculo madre-hijo. Goursaud and Nowak (1999) han demostrado recientemente que la presencia de calostro en el estómago de los corderos recién nacidos facilita la habilidad del cordero para reconocer a su madre lo cual es esencial para establecer un vínculo exitoso entre la oveja y su cordero inmediatamente luego del parto disminuyendo las posibilidades de abandono.
Los requerimientos de calostro para el cordero han sido calculados de acuerdo a la energía que éste requiere para evitar movilizar sus propias reservas corporales. por debajo de la cual sus propias reservas de energía deben ser movilizadas. Mellor y Murray (1986) estimaron que un cordero requiere 180 ml de calostro por kg de peso vivo durante sus primeras 18 horas de vida cuando la temperatura ambiente es de 10°C y no hay viento y Robinson et al. (2002) sostienen que aproximadamente un 30% de esta cantidad (50 g/kg de peso vivo) debe estar disponible al parto. Pero si las condiciones ambientales son más adversas con presencia de viento y frío los requerimientos aumentan un 50% (Alexander, citado por McCance y Alexander, 1959).
En estudios realizados en Australia, McNeill et al. (1998) hallaron que un 30% de ovejas Merino bien alimentadas no tenían suficiente calostro para los corderos mellizos y 10% no producían suficiente calostro para los corderos únicos. Veinte por ciento de estas ovejas no presentaban ninguna secreción el día previo al parto y 5% de las ovejas no presentaban DMV, PhD. InIA La Estanzuela. (Uruguay).

280 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
calostro aún una hora luego del parto. Existe una fuerte relación entre la nutrición durante la gestación y el inicio de la lactación. Mellor y Murray (1985a,b) mostraron que una mala alimentación durante las últimas seis semanas de gestación deprime el desarrollo de la ubre y la acumulación prenatal de calostro, así como la producción subsiguiente de leche durante las 18 horas posteriores al parto.
Normalmente, el calostro se acumula rápidamente unos días previos al parto (2 a 3 días) asegurando de este modo la disponibilidad de varios mililitros al momento del nacimiento del cordero. Sin embargo, esta fase llamada lactogénesis II (Hartmann et al., 1973), puede estar reducida a tal grado que en algunas ovejas, particularmente las melliceras, no tengan calostro al momento del parto. Algunas de las posibles razones incluyen una pobre calidad o valor nutritivo de la pastura y/o una reducción en el consumo voluntario de las ovejas durante las últimas semanas de gestación. Weston (1988) sugirió que durante la gestación tardía la compresión del útero y su contenido limitaría el volumen del rumen y esto podría reducir el consumo voluntario, especialmente con dietas a base de forrajes. Esto no sucedería si a la oveja se le suministra un concentrado ya que la densidad energética y/o proteica del mismo siempre puede ser mayor que la de un forraje. De este modo, una suplementación estratégica previo al parto, de corta duración y fácil de aplicar puede contrarrestar este efecto disminuyendo la mortalidad de corderos por inanición.
LACtOGénESIS II O InICIO DE LA SíntESIS DE CALOStROEl comienzo de la lactación se caracteriza por el inicio de la actividad de síntesis de la glándula mamaria durante la mamogénesis. Al principio se producen pequeñas cantidades de componentes de la leche los que permanecen en la luz del alvéolo. Esta fase se llama lactogénesis I y en la oveja este proceso se da en el último mes de gestación (Robinson et al., 1978). Al momento del parto debido a cambios hormonales, la síntesis de leche/calostro aumenta rápidamente lo que se corresponde con una rápida hipertrofia de las células del epitelio mamario. Esta fase llamada lactogénesis II o inicio de la síntesis de calostro en la oveja comienza dos o tres días previos al parto y puede demorarse hasta un día después del parto (Alexander y Davies, 1959) para luego extenderse por varias horas luego de la parición.
Las ovejas gestando corderos mellizos generalmente producen más calostro pero el inicio de la lactogénesis está más demorado que en ovejas gestando corderos únicos (Alexander y Davies, 1959; Geenty, 1986). Esto implica que al momento del parto los corderos mellizos tienen menos calostro disponible que
los corderos únicos. Además, la viscosidad de ese calostro suele ser mayor que en ovejas con corderos únicos (Banchero et al. 2003). Esto dificulta el amamantamiento ya que los corderos tienen que mamar más veces y utilizar más energía para lograr una cantidad adecuada a sus requerimientos (Holst et al. 1996). Esto se debe a que la lactogénesis II esta relacionada negativamente con la concentración plasmática de progesterona (Hartmann et al. 1973). Altos niveles de progesterona durante la gestación bloquean el inicio de la lactación. En la oveja, durante los dos últimos tercios de la gestación, la progesterona es producida mayoritariamente por la placenta (Catchpole, 1991), permaneciendo alta hasta unos días previos al parto cuando comienza a descender lentamente para desaparecer completamente una vez que la misma es expulsada. Dentro de los mecanismos por los cuales la progesterona bloquea la síntesis de calostro se encuentra la inhibición de la síntesis de lactosa. La lactosa es osmóticamente activa y tiene como principal función la de regular el contenido de agua de la leche (Rigour et al., 2002). Cuando el contenido de lactosa no es suficiente, el volumen de calostro es muy bajo o la viscosidad del mismo es muy alta (el calostro parece miel). En la oveja mellicera, la concentración plasmática de progesterona es más alta que en las ovejas que paren corderos únicos (Mc Neill et al. 1998). La presencia de dos placentas en las gestaciones múltiples es parte de la explicación del fenómeno.
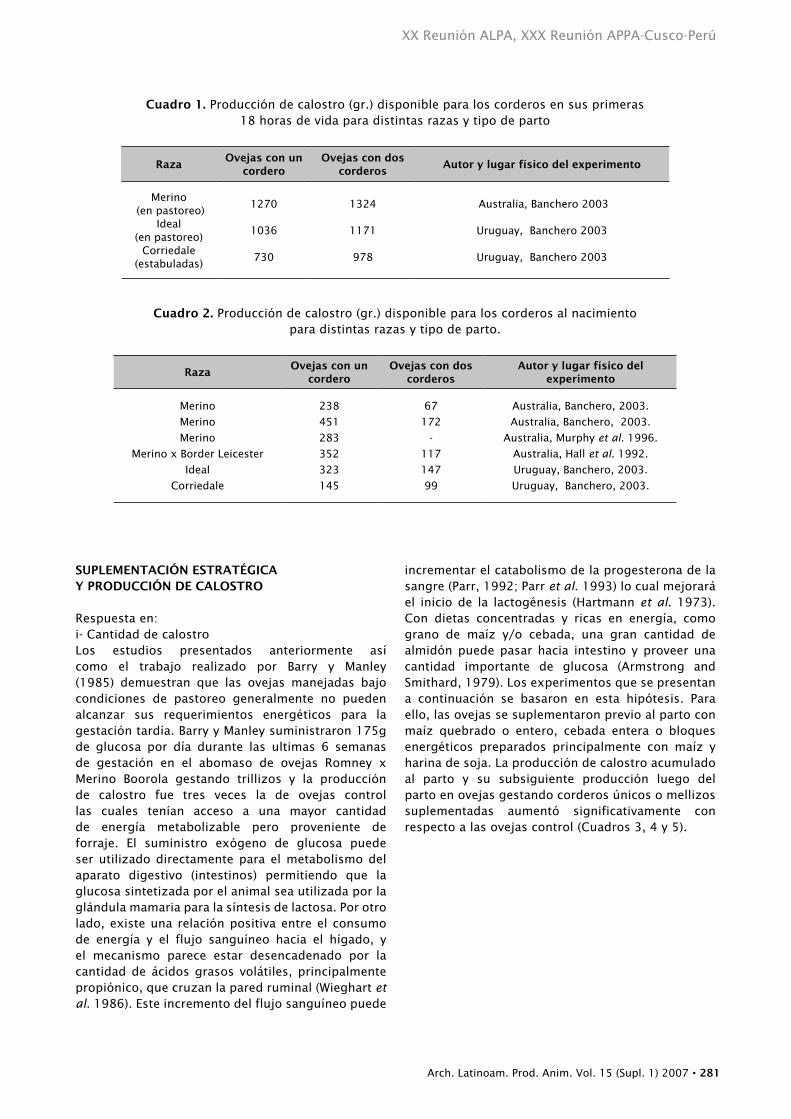
PRODUCCIón DE CALOStRO En OVEJAS ALIMEntADAS COn FORRAJEEn el Cuadro 1 se presenta la cantidad de calostro acumulado al parto y el total producido durante las primeras 18 horas posparto en ovejas pertenecientes a trabajos experimentales realizados en Australia y Uruguay. Las ovejas estaban bajo pastoreo o encerradas pero alimentadas con forraje conservado y la oferta de alimentos fue realizada para cubrir los requerimientos de gestación avanzada de acuerdo a MAFF (1975). Las ovejas con corderos únicos produjeron entre 730 y 1270 gr. lo cual es suficiente para cubrir los requerimientos de la mayoría de los corderos únicos si las condiciones ambientales son favorables (temperatura igual o mayor a 10° C y sin viento). Pero las ovejas melliceras produjeron apenas una mayor cantidad que las únicas, entre 970 y 1320 gr. y si consideramos que cada mellizo obtiene la mitad de esto, podemos afirmar que estos corderos estaban subalimentados ya que sus requerimientos no podían ser cubiertos aún en buenas condiciones climáticas. Más aún, cuando medimos la cantidad de calostro presente en la ubre de la oveja al parto, se observó que muchos corderos tenían muy poco calostro disponible para afrontar sus primeras horas de vida (Cuadro 2).
Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•281
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
SUPLEMEntACIón EStRAtéGICA y PRODUCCIón DE CALOStRO
Respuesta en:i- Cantidad de calostroLos estudios presentados anteriormente así como el trabajo realizado por Barry y Manley (1985) demuestran que las ovejas manejadas bajo condiciones de pastoreo generalmente no pueden alcanzar sus requerimientos energéticos para la gestación tardía. Barry y Manley suministraron 175g de glucosa por día durante las ultimas 6 semanas de gestación en el abomaso de ovejas Romney x Merino Boorola gestando trillizos y la producción de calostro fue tres veces la de ovejas control las cuales tenían acceso a una mayor cantidad de energía metabolizable pero proveniente de forraje. El suministro exógeno de glucosa puede ser utilizado directamente para el metabolismo del aparato digestivo (intestinos) permitiendo que la glucosa sintetizada por el animal sea utilizada por la glándula mamaria para la síntesis de lactosa. Por otro lado, existe una relación positiva entre el consumo de energía y el flujo sanguíneo hacia el hígado, y el mecanismo parece estar desencadenado por la cantidad de ácidos grasos volátiles, principalmente propiónico, que cruzan la pared ruminal (Wieghart et al. 1986). Este incremento del flujo sanguíneo puede
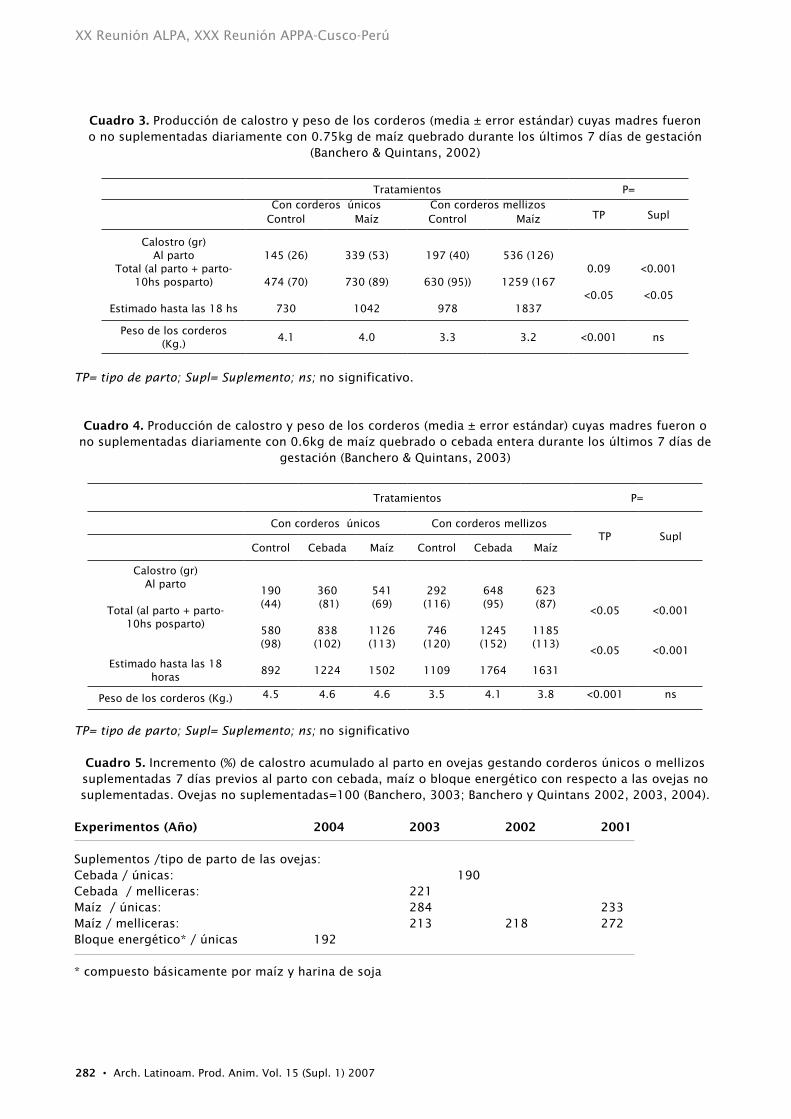
incrementar el catabolismo de la progesterona de la sangre (Parr, 1992; Parr et al. 1993) lo cual mejorará el inicio de la lactogénesis (Hartmann et al. 1973). Con dietas concentradas y ricas en energía, como grano de maíz y/o cebada, una gran cantidad de almidón puede pasar hacia intestino y proveer una cantidad importante de glucosa (Armstrong and Smithard, 1979). Los experimentos que se presentan a continuación se basaron en esta hipótesis. Para ello, las ovejas se suplementaron previo al parto con maíz quebrado o entero, cebada entera o bloques energéticos preparados principalmente con maíz y harina de soja. La producción de calostro acumulado al parto y su subsiguiente producción luego del parto en ovejas gestando corderos únicos o mellizos suplementadas aumentó significativamente con respecto a las ovejas control (Cuadros 3, 4 y 5).
Cuadro 1. Producción de calostro (gr.) disponible para los corderos en sus primeras 18 horas de vida para distintas razas y tipo de parto
RazaOvejas con un
cordero Ovejas con dos
corderos Autor y lugar físico del experimento
Merino (en pastoreo)
Ideal(en pastoreo)
Corriedale(estabuladas)
1270
1036
730
1324
1171
978
Australia, Banchero 2003
Uruguay, Banchero 2003
Uruguay, Banchero 2003
Cuadro 2. Producción de calostro (gr.) disponible para los corderos al nacimiento para distintas razas y tipo de parto.
RazaOvejas con un
cordero Ovejas con dos
corderos Autor y lugar físico del
experimento
Merino
Merino
Merino
Merino x Border Leicester
Ideal
Corriedale
238
451
283
352
323
145
67
172
-
117
147
99
Australia, Banchero, 2003.
Australia, Banchero, 2003.
Australia, Murphy et al. 1996.
Australia, Hall et al. 1992.
Uruguay, Banchero, 2003.
Uruguay, Banchero, 2003.
282 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Cuadro 3. Producción de calostro y peso de los corderos (media ± error estándar) cuyas madres fueron o no suplementadas diariamente con 0.75kg de maíz quebrado durante los últimos 7 días de gestación
(Banchero & Quintans, 2002)
Tratamientos P=Con corderos únicos Con corderos mellizos
TP SuplControl Maíz Control Maíz
Calostro (gr)Al parto
Total (al parto + parto-10hs posparto)
Estimado hasta las 18 hs
145 (26)
474 (70)
730
339 (53)
730 (89)
1042
197 (40)
630 (95))
978
536 (126)
1259 (167
1837
0.09
<0.05
<0.001
<0.05
Peso de los corderos (Kg.)
4.1 4.0 3.3 3.2 <0.001 ns
tP= tipo de parto; supl= suplemento; ns; no significativo.
Cuadro 4. Producción de calostro y peso de los corderos (media ± error estándar) cuyas madres fueron o no suplementadas diariamente con 0.6kg de maíz quebrado o cebada entera durante los últimos 7 días de
gestación (Banchero & Quintans, 2003)
Tratamientos P=
Con corderos únicos Con corderos mellizosTP Supl
Control Cebada Maíz Control Cebada Maíz
Calostro (gr)Al parto
Total (al parto + parto-10hs posparto)
Estimado hasta las 18 horas
190 (44)
580 (98)
892
360 (81)
838 (102)
1224
541 (69)
1126(113)
1502
292 (116)
746(120)
1109
648 (95)
1245(152)
1764
623 (87)
1185(113)
1631
<0.05
<0.05
<0.001
<0.001
Peso de los corderos (Kg.) 4.5 4.6 4.6 3.5 4.1 3.8 <0.001 ns
tP= tipo de parto; supl= suplemento; ns; no significativo
Cuadro 5. Incremento (%) de calostro acumulado al parto en ovejas gestando corderos únicos o mellizos suplementadas 7 días previos al parto con cebada, maíz o bloque energético con respecto a las ovejas no suplementadas. Ovejas no suplementadas=100 (Banchero, 3003; Banchero y Quintans 2002, 2003, 2004).
Experimentos (Año) 2004 2003 2002 2001
Suplementos /tipo de parto de las ovejas:Cebada / únicas: 190 Cebada / melliceras: 221Maíz / únicas: 284 233Maíz / melliceras: 213 218 272Bloque energético* / únicas 192
* compuesto básicamente por maíz y harina de soja

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•283
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
El peso de los corderos al nacimiento no se vio afectado por el corto período de suplementación que recibieron sus madres con la ventaja de que no aumentó la probabilidad de problemas de distocia.
II- VISCOSIDAD DEL CALOStROLa ovejas suplementadas con maíz o con cebada no solo produjeron más calostro sino que éste fue más líquido, lo que hace que el cordero pueda mamarlo más fácilmente que calostros más viscosos o espesos (Holst et al. 1996). La baja viscosidad del calostro en ovejas suplementadas está asociada a altos niveles
de lactosa en el calostro (Cuadros 8 y 9). La lactosa que es osmóticamente activa (Leong et al. 1990) retira agua del torrente sanguíneo y esto disminuye la viscosidad del calostro. Las ovejas melliceras no suplementadas produjeron el calostro de mayor viscosidad y menor concentración de lactosa. Con la falta de suplemento claramente se exacerba el problema de las ovejas melliceras las cuales tienen mucha menor oportunidad que las ovejas gestando corderos únicos de cubrir los requerimientos de glucosa durante el fin de la gestación aún cuando están aparentemente bien alimentadas.
Cuadro 6. Viscosidad y concentración de lactosa (media ± error estándar) del calostro acumulado al parto en ovejas suplementadas o no diariamente con 0.75kg de maíz quebrado durante los últimos 7 días de
gestación (Banchero & Quintans, 2002).
Tratamientos P=
Con corderos únicosCon corderos
mellizos TP Supl TPxSuplControl Maíz Control Maíz
Viscosidad (escore
0-7*)
Lactosa (%)
5.7
(0.23)
1.8 ±
0.18
5.8
(0.30)
2.6 ±
0.26
4.5
(0.40)
1.4 ±
0.27
5.8
(0.11)
2.5 ±
0.20
<0.05
ns
<0.01
<0.001
<0.05
ns
TP= tipo de parto; Supl= Suplemento; ns; no significativo*Mc Cance and Alexander, 1959.
Cuadro 7. Viscosidad y concentración de lactosa (media ± error estándar) del calostro acumulado al parto en ovejas suplementadas o no diariamente con 0.6kg de maíz quebrado o cebada entera durante los
últimos 7 días de gestación (Banchero & Quintans, 2003).
Tratamientos P=
Con corderos únicos Con corderos mellizosTP Supl
Control Cebada Maíz Control Cebada Maíz
Viscosidad (escore
0-7)
Lactosa (%)
4.2
(0.41)
1.6
(0.20)
6.0
(0.43)
2.5
(0.20)
6.3
(0.41)
3.1
(0.20)
4.3
(0.50)
1.4
(0.30)
6.3 (0.50)
2.8 (0.30) 6.1 (0.48)
2.4 (0.20)
ns
ns
<0.001
<0.001
TP= tipo de parto; Supl= Suplemento; ns; no significativo

284 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
III- VALOR EnERGétICO y PROtEICOLos corderos únicos y mellizos hijos de ovejas suplementadas tuvieron cerca del doble de proteína y energía metabolizable disponible en el calostro al parto y durante sus primeras 10 horas de vida comparado con los corderos hijos de ovejas no suplementadas. Tanto la cantidad de energía como la proteína obtenida en el calostro de ovejas suplementadas se debieron a mayor volumen de calostro ya que el porcentaje de sólidos fue menor (25.8 vs 29.4%, p<0.01) que en ovejas no suplementadas. Esto se debe a una menor concentración de grasa y de proteína. Dentro de las posibles causas para esta disminución de grasa y proteína aparece la dilución. Para el caso de las grasas una posible explicación es una disminución en la producción del acido graso C
18 ya sea por
una inhibición de la lipólisis o un incremento en la lipogénesis en ovejas suplementadas con gran cantidad de cereales (Lemosquet et al. 1997).
IV-CAMBIOS hORMOnALES IMPLICADOS En LA REGULACIón DE LA SíntESIS DE CALOStROLa respuesta de las ovejas a la suplementación también se reflejó en cambios hormonales y de metabolitos plasmáticos. La concentración de progesterona durante el período preparto disminuyó en todas las ovejas, pero lo hizo de forma más rápida en los animales suplementados. Hartmann
et al. (1973) sugirieron que el nivel crítico de progesterona plasmática para el inicio de la síntesis de calostro debe ser menor a 1 ng/ml. En las ovejas suplementadas con maíz durante los últimos 7 días de gestación este nivel crítico de progesterona fue alcanzado entre 12 horas preparto y el parto tanto en ovejas melliceras como con cordero único (Figura 1). Los niveles de progesterona de las ovejas no suplementadas gestando corderos únicos cayeron de forma similar que los de ovejas suplementadas, pero las ovejas melliceras demoraron más en obtener la progesterona adecuada para el inicio de la síntesis de calostro, haciéndolo desde 12 horas preparto hasta una hora postparto. Las ovejas de este experimento acumularon la mayoría del calostro disponible al parto durante estos intervalos, pero la gran cantidad de calostro producida por las ovejas suplementadas fue probablemente debido a una mayor disponibilidad de precursores para la síntesis de calostro en estos animales. Esto puede tener su explicación en que la glucosa plasmática de ovejas suplementadas fue superior que la de ovejas no suplementadas, respaldando nuestra hipótesis de que el maíz incrementa la tasa de entrada de glucosa al animal y en consecuencia la toma de la misma por la glándula mamaria con una mayor síntesis de lactosa.
Cuadro 8. Energía metabolizable y proteína (media ± error estándar) secretada en el calostro acumulado al parto y producido en las siguientes 10 hs luego del parto en ovejas suplementadas o no diariamente con
0.75kg de maíz quebrado durante los últimos 7 días de gestación (Banchero 2003).
Con corderos únicos Con corderos mellizos P=
Control Maíz Control Maíz TP Supl
EM en el calostro (MJ)
Al parto
Parto a 10 hs post parto
Total de EM
Proteína secreteada (g)
Al parto
Parto a 10 hs post parto
Total de proteína
0.5 ± 0.09
1.4 ± 0.23
1.9 ± 0.31
24.8 ± 4.5
35.2 ± 4.7
60 ± 8.5
1.1 ± 0.16
1.8 ± 0.29
2.9 ± 0.4
47.3 ± 6.5
38.4 ± 6.9
85.7 ± 11.4
0.8 ± 0.17
2.0 ± 0.32
2.8 ± 0.45
37.8 ± 7.8
53.5 ± 8.4
91.3 ± 14.5
2.1 ± 0.87
3.3 ± 0.5
5.4 ± 1.0
74.9 ± 14.2
73.8 ± 10.5
148.7 ± 17.0
ns
<0.01
<0.01
<0.05
<0.001
<0.001
<0.05
<0.05
<0.01
<0.01
ns
<0.001
TP= tipo de parto; Supl= Suplemento; ns; no significativo

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•285
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Figura 1. Cambios en la concentración plasmática de progesterona y glucosa durante los últimos 7 días de gestación y las primeras 10 hs luego del parto en ovejas suplementadas o no con 0.75kg de maíz por día. (*=P < 0.05 para tipo de parto; * =P < 0.05 para suplementación). (○= unica no suplementada; ●= única suplementada □= mellicera no suplementada ■= mellicera suplementada
Figura 2. Cambios en la concentración plasmática de insulina e IGF-I durante los últimos 7 días de gestación y las primeras 10 hs luego del parto en ovejas suplementadas o no con 0.75kg de maíz por día. (*=P < 0.05 para tipo de parto; * =P < 0.05 para suplementación). (○=mellicera no suplementada; ●= mellicera suplementada □= única no suplementada ■= única suplementada

286 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
La insulina aún cuando no tiene efecto sobre la toma de nutrientes del torrente sanguíneo por la glándula mamaria (Lemosquet et al. 1997), en concentraciones altas pueden incrementar la concentración de IGF-1 en plasma (Bequette et al. 2001). La IGF-I incrementa el crecimiento de la glándula mamaria (Delouis and Richard, 1993) y el flujo sanguíneo hacia la misma (Prosser et al. 1990). Un incremento en el flujo sanguíneo hacia la glándula mamaria puede incrementar la toma de glucosa por la glándula mamaria (Bequette et al. 2001). En nuestro experimento, las ovejas suplementadas tuvieron niveles más altos de insulina e IGF-1, lo que se acompañó de una mayor producción de calostro mientras que el volumen de la glándula mamaria vacía fue 49% superior (P>0.05) en ovejas suplementadas que en ovejas no suplementadas.
COnCLUSIOnESLa muerte de los corderos recién nacidos por inanición sigue siendo la causa más importante de muerte. Uno de los principales factores es la falta de calostro de las ovejas al parto. Una suplementación preparto corta con granos ricos en almidón permite incrementar significativamente la producción de calostro tanto en ovejas con corderos únicos como mellizos. Asimismo, la viscosidad del calostro disminuye sin alterar su calidad, haciéndolo más fácil de consumir por parte del cordero.
Este es el tipo de opciones permite incrementar la sobrevivencia de los corderos únicos y mellizos y asegurar un buen crecimiento posterior. Es importante destacar que con una cantidad limitada o baja de grano por animal (entre 7 y 10 Kg.) se duplica la producción de calostro, se facilita el consumo del mismo y no aumenta los riesgos de distocia ya que no se altera el peso de los corderos al nacer.
REFEREnCIAS
Alexander, G. (1984) Constrains to the lamb survival in: Reproduction in Sheep, D.R. Lindsay and D.T. Pearce (Eds.), Australian Academy of Science and Australian Wool Corporation; Canberra, pp. 199-209.
Alexander, G. and Lloyd Davies, H. (1959). Australian Journal of Agricultural research, 10, 720-724.
Armstrong, D.G. and Smithard, R.R. (1979) Proceedings of the Nutrition Society, 38: 283-294.Banchero, G. (2003). PhD Thesis. The University of Western Australia.
Banchero, G. & Quintans, G. (2002). Actividades de Difusión N° 294, p 32.
Banchero, G. y Quintans, G. (2003). Serie de Actividades de Difusión N° 342: 26-31 Banchero, G. y Quintans, G. (2004). Día de Campo. Octubre 2004. INIA Treinta y Tres.
Banchero, G., Delucci, M. I. y Quintans, G. (2003). Serie de Actividades de Difusión N° 342: 19-25 INIA La Estanzuela.
Barry, T.N. and Manley, T.R. (1985) British Journal of Nutrition, 54: 521-533.
Bequette, B.J., Kyle, C.E., Crompton, C.E., Buchan, V. and Hanigan, M.D., (2001) Journal of Dairy Science, 84: 241-255
Catchpole, H.R. (1991). En: Reproduction in Domestic Animals, 4th Edition. Eds Perry T. Cupp New York, Academic Press., 361-381.
Delouis, C. and Richard, P. (1993) Lactation in: Reproduction in Mammals and Man, C. Thibault, M-C Levasseur and R.H.F Hunter (Eds.), Ellipses; Paris, pp. 503-530.
Durán del Campo, A. (1963). En: Segundo Ciclo de Conferencias y Debates. Centro de Investigaciones Veterinaria Dr. Miguel C. Rubino, Uruguay.
Geenty, K.G. and Sykes, A. R. (1986). Journal of Agriculture Science of Cambridge, 106, 351-367.Goursaud, A-P. and Nowak, R. (1999). Physiology & Behavior, 67: 49-56.
Hall, D.G., Holst, P.J. and Shutt, D.A. (1992) Australian Journal of Agricultural Research, 43: 325-337.
Hartmann, P.E. (1973). Journal of Endocrinology, 59: 231-247.Hartmann, P.E., Trevethan, P. and Shelton, J.N. (1973). Journal of Endocrinology, 59: 249-259.
Holst, P.J., Hall, D.G. and Allan, C.J. (1996). Australian Journal of Experimental Agriculture, 36: 637-640.
Lemosquet, S., Rideau, N., Rulquin, H., Faverdin, P., Simon, J. and Verite, R. (1997) Journal of Dairy Science, 80: 2854-2865.
Leong, W.S., Navaratnam, N., Stankiewicz, M.J., Wallace, A.V., Ward, S. and Kuhn, N. (1990) Protoplasm, 159: 144-156.
MAFF, Ministry of Agriculture, Fisheries and Food (1975). Technical Bulletin 33; London, 79 pp
Mari, J.J. (1979). En: Jornadas Veterinarias de Ovinos, 1: 1-13. Uruguay.

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•287
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
McCance, I. and Alexander, G. (1959). Australian Journal of Agricultural Research, 10: 699-719.
McNeill, D.M., Murphy, P.M. and Lindsay, D.R. 1998. Australian Journal of Agricultural Research, 49, 581-587.
Mellor, D.J. and Murray, L. (1985a). Research in Veterinary Science, 39: 235-240.
Mellor, D.J. and Murray, L. (1985b). Research in Veterinary Science, 39: 230-234.
Mellor, D.J. and Murray, L. (1986). Veterinary record, 118: 351
Murphy, P.M., McNeill, D.M., Fisher, J.S. and Lindsay, D.R. (1996) Proceedings of the Australian Society of Animal Production, 21: 227-230.
Nowak, R. (1996). Applied Animal Behaviour Science 49, 61-72.
Parr, R.A. (1992) Reproduction, Fertility, & Development, 4: 297-300.
Parr, R.A., Davis, I.F., Miles, M.A. and Squires, T.J. (1993a) Research in Veterinary Science, 55: 306-310.
Pattinson, S.E., Davies, D.A.R. & Winter, A.C. (1995). Animal Science, 61: 63.
Prosser, C.G., Fleet, I.R., Corps, A.N., Froesch, E.R. and Heap, R.B. (1990) Journal of Endocrinology, 126: 437-443.
Rigout, S., Lemosquet, J.E., van Eys, J.E. and Blum, J.W. (2002) Journal of Dairy Science, 85: 595-606.Robinson, J.J., McDonald, I., McHattie, I. and Pennie, K. (1978) Journal of Agricultural Science, Cambridge, 91: 291-304.
Robinson, J.J., Rooke, J.A. & McEvoy, T.G. (2002). Sheep nutrition, M. Freer and H. Dove (Eds.), CABI Publishing in association with CSIRO Publishing; Canberra, pp. 189.
Wieghart, M., Slepetis, R., Elliot, J.M. and Smith, D.F. (1986). Journal of Nutrition, 116: 839-850
Weston, R.H. (1988). Australian Journal of Agricultural Research, 39: 659.

288 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
La mortalidad perinatal de corderos es uno de los factores más importantes que limitan la eficiencia biológica y económica de los sistemas de producción ovina en todo el mundo. Las pérdidas derivan no sólo de la muerte de animales, sino también de la mayor utilización de forraje y menor producción de lana de la oveja gestante, así como del menor número de animales disponibles para la selección. En Uruguay, la cifra promedio de mortalidad perinatal se estima en 20% de los corderos nacidos, con una variación del 14 al 32% según los años y los predios.1 Pérdidas similares ocurren en Australia, Nueva Zelanda, Sudáfrica y en otros países con una importante producción ovina. 1, 2
Es significativo que estos niveles de pérdidas no han cambiado substancialmente en las últimas décadas a pesar de los esfuerzos realizados por los investigadores, extensionistas, y productores. En condiciones extensivas de producción la mortalidad es difícil de disminuir más allá del 10%, aún cuando se controlen las enfermedades infecciosas, se mejore la alimentación durante las etapas finales de la gestación, o se implanten montes de abrigo en los potreros de parición, entre otras de las muchas técnicas disponibles.2 Se ha intentado también aumentar la prolificidad de las ovejas con el fin de mejorar el procreo, pero esta práctica es censurable desde el punto ético y del bienestar animal si nuestros sistemas de producción no pueden asegurar la supervivencia de los corderos mellizos. Por estas razones muchos expertos consideran que la mortalidad perinatal en ovinos es un problema frustrante e irresoluble, al punto que las investigaciones sobre el tema han disminuido en todo el mundo o simplemente se han abandonado.
El bajo impacto de las actuales recomendaciones para mejorar la supervivencia de los corderos puede deberse a un error conceptual que impide
nuevos enFoques soBre la mortalidad perinatal de Corderos
A nEW PERSPECtIVE In PERInAtAL LAMB MORtALIty
Fernando Dutra quintela
DMV PhD Dpto. de PatologíaDILAVE Miguel C Rubinotreinta y tres, UruguayE-mail: [email protected]
conocer la naturaleza biológica real de la mortalidad perinatal. Muchos factores genéticos y no genéticos se han incriminado tradicionalmente como causas de mortalidad perinatal, incluyendo distocia, peso al nacer, nutrición pre-parto, comportamiento materno, inanición, predación, malformaciones congénitas, infecciones, y estrés climático, entre otras.2 Genéticamente, la mortalidad perinatal es menor en la raza Texel y mayor en la raza Merino y sus cruzas.3 Es posible sin embargo que existan otras causas de muerte perinatal hasta ahora no reconocidas y sin cuya identificación un avance en la solución del problema es poco probable.
En el hombre4 y en diversas especies animales, incluyendo el equino5 y el cerdo,6 las lesiones cerebrales al momento del parto son una causa significativa de mortalidad y morbilidad perinatal, por lo que sería un absurdo biológico que los corderos no desarrollaran lesiones neuropatológicas perinatales. Los trabajos clásicos de mortalidad perinatal no incluían en su metodología al cerebro por considerarlo poco importante. A principios de los ’70 Haughey en Australia demostró que muchos de los corderos muertos durante el período perinatal presentan hemorragias meníngeas,7 aunque no se realizaron estudios histológicos para evaluar el tipo y severidad de las lesiones en el cerebro. Investigaciones recientes realizadas en Uruguay mostraron que una alta proporción de los corderos muertos en el período perinatal temprano presentaba lesiones cerebrales de encefalopatía hipóxico-isquémica.8 Para estudiar estas lesiones se normalizó un protocolo de investigación que incluye –además del estudio macroscópico- el estudio histológico del sistema nervioso central, ya que la mayoría de las lesiones de encefalopatía hipóxico isquémica son microscópicas.
Las lesiones macro y microscópicas más comunes en el sistema nervioso central de los corderos muertos en el período perinatal son edema cerebral, hemorragias perivasculares e intra parenquimatosas en el bulbo raquídeo y la médula espinal cervical, necrosis isquémica parasagital de la corteza cerebral, infartos agudos de sustancia blanca periventricular, y necrosis isquémica de neuronas en corteza cerebral, núcleos de

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•289
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
la base y tálamo, entre otras regiones neuroanatómicas. Estas lesiones de encefalopatía hipóxico-isquémica se encontraron tanto en corderos mellizos como únicos. Las lesiones más severas son seguramente causa inmediata de muerte de los corderos, mientras que las lesiones más leves probablemente les impide mamar y / o alteran su capacidad de supervivencia y adaptación al medio. Las lesiones constatadas de encefalopatía hipóxico-isquémica son probablemente el resultado de la asfixia y trauma al sistema nervioso central producidas durante el proceso de parto. Lesiones cerebrales similares a las encontradas en corderos de campo se producen en modelos experimentales de corderos sometidos a compresión intermitente del cordón umbilical.9, 10 A este respecto, y a diferencia de la explicación multifactorial tradicional, la curva epidémica de las muertes en los primeros 7 días post-parto es típica de una epidemia a fuente común, con el parto como único factor de exposición. La ausencia de lesiones en los corderos muertos ante-parto y 9-12 días post-parto sugiere también al parto como única causa de las muertes tempranas. La especie ovina parece estar bio-anatómicamente proclive a desarrollar este tipo de lesiones al momento del parto, ya que los corderos tienen al nacer un cuello cilíndrico, largo, y muscularmente muy poco desarrollado, con articulaciones cervicales inestables y sumamente flexibles, que lo predisponen a desarrollar lesiones isquémicas al momento del parto. El desarrollo muscular y la madurez esquelética del cuello varían según el biotipo y la raza de los corderos.11
Los resultados sugieren que para disminuir la mortalidad perinatal se debe hacer mayor hincapié en la facilidad de parto de las ovejas y / o en el biotipo de los corderos, ya sea seleccionando nuevos biotipos o incorporando nuevas líneas genéticas. Nuevas líneas de investigación se están desarrollando en Uruguay en la raza Ideal (lana) y Texel (carne) para estudiar la duración del parto y el biotipo del cordero (examinando por disección su desarrollo muscular y madurez esquelética al nacimiento), y de que manera ello los protege de la asfixia al momento del parto. Otra posible línea de investigación futura sería el uso de alguno de los distintos neuroprotectores usados en medicina humana y experimentalmente en modelos animales, tales como el factor de crecimiento IGF1,12 bloqueadores de calcio, sulfato de magnesio, y antioxidantes como la Vitamina E / Selenio en dosis neuroprotectoras, entre muchas otras posibilidades.En conclusión, la mortalidad perinatal de corderos es un problema aún abierto que requiere de nuevas líneas de investigación.
REFEREnCIAS1. Mari JJ. Pérdidas de corderos . En: Bonino Morlán J, Durán del Campo A, Mari JJ, Editores. Enfermedades de los Lanares. 1st edn., Tomo III, Cap. V, pp. 73-100, Editorial Hemisferio Sur SRL, Montevideo, 1989 vol 3:73-100.
2. Alexander G. Constraints to lamb survival. In: Lindsay DR and Pearce DT, editors. Reproduction in sheep. Australian Academy of Science, Camberra, 1984:199-209.
3. Ganzábal A, Echevarría MN. Análisis comparativo del comportamiento reproductivo y habilidad materna de ovejas cruza. In: Seminario de Reproducción Ovina, INIA Uruguay, Serie de Difusión 401, Abril y Mayo 2005:33-42.
4. Volpe JJ. Hypoxic-ischemic encephalopathy. In: Volpe JJ, editor. Neurology of the Newborn. 4th edn. W.B. Saunders Company, Philadelphia, 2001:217-394.
5. Vaala WE. Peripartum asphyxia syndrome in foals. Proceeding of the American Association of Equine Practitioners 1999, 45:247-253.
6. Herpin P, Hulin JC, Le Dividich J, Fillaut M. Effects of oxygen inhalation at birth on the reduction of early postnatal mortality in pigs. J Anim Sci 2001, 79:5-10.
7. Haughey KG. Vascular abnormalities in the central nervous system associated with perinatal mortality. 1. Pathology. Aust Vet J 1973, 49:1-8.
8. Dutra F, Quintans G, Banchero G. Lesions in the central nervous system associated with perinatal lamb mortality. Austr Vet J 2007, 85:000-000.
9. De Haan HH, Gunn AJ, Williams CE, Gluckman PD. Brief repeated umbilical cord occlusions cause sustained cytotoxic cerebral edema and focal infarcts in near-term fetal lambs. Pediatr Res 1997, 41:96-104.
10. Marumo G, Kozuma S, Ohyu J, et al. Generation of periventricular leukomalacia by repeated umbilical cord occlusion in near-term foetal sheep and its possible pathogenetical mechanisms. Biol Neonate 2001, 79:39-45.
11. Leymaster KA, Jenkins TG. Comparison of Texel- and Suffolk-sired crossbred lambs for survival, growth, and compositional traits. J Anim Sci 1993, 71:859-869.
12. Johnston BM, Mallard EC, Williams CE, Gluckman PD. Insulin-like Growth Factor-1 is a potent neuronal rescue agent after hypoxic-ischemic injury in fetal lambs. J Clin Invest 1996, 97:300-308.

290 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
IntRODUCCIónLa crianza de caprinos en le Perú se inicia en el siglo XVI, con la introducción de diferentes razas de caprinos españoles, aún no completamente definidos, habiéndose encontrados entre ellos las razas Murciana, Granadina, Malagueña, etc. Estos caprinos lentamente se distribuyeron en casi todo el país, con excepción de los trópicos húmedos. En la zona norte y dado la presencia del algarrobo se asentó en forma numerosa. Posteriormente se instalaron en los valles secos de la costa y los valles interandinos de la sierra. Desde el inicio de su crianza, los caprinos estuvieron ligados directamente con los productores agropecuarios de los más bajos recursos económicos y tradicionalmente fueron excluidos de la investigación, la asistencia técnica, el crédito y una adecuada comercialización, lo cual ha frenado notablemente su desarrollo
MAtERIALES y MétODOSEl presente estudio analiza la situación actual de la crianza de caprinos, mediante el uso de los diferentes trabajos de investigación que se han desarrollado en las distintas universidades del país, trabajos de desarrollo de la explotación efectuados por proyectos especiales, acciones de desarrollo de ONGs y la actividad privada de ciertas empresas que se involucraron en esta crianza. En general se han analizado más de 400 diferentes trabajos de investigación y transferencia de tecnologías.
RESULtADOS y DISCUSIón
SItUACIOn ACtUAL PROyECCIOnES
1. Crianza en general
Crianza: La mayor parte de la crianza de caprinos se efectúa en forma extensiva, con desplazamiento de los animales diariamente en busca de alimentos, con un solo corral de crianza. Los empadres son continuos y las crías tienen una reproducción temprana que retarda su crecimiento y presentan una menor producción general. No se planifica ni la producción ni las sacas. Son pocas las crianzas que son de carácter intensivo y con pastoreo cero.
El mejoramiento de la crianza en general de caprinos es convertir la crianza extensiva en una crianza semiiintensiva e intensiva, adonde los animales estén instalados en corrales separados por sexos, se controle los empadres y pariciones y tengan una adecuada alimentación. Se necesita una profunda reingeniería para convertir estas pequeñas crianzas en pequeñas explotaciones de carácter comercial y que rindan utilidades económicas.
Alimentación: Esta se efectúa principalmente con el pasto natural de las lomas de la costa, la pastura nativa de las montañas de la sierra, la pastura eventual de caminos y acequias, rastrojos de cosecha, ramoneo de árboles y arbustos. Especialmente en la costa central existen productores que están suplementando esta alimentación con granos y alimentos concentrados
La alimentación de los animales puede ser efectuada con alimentos verdes sembrados en áreas adecuadas que se pueden manejar conjuntamente con otros productores. Además se puede almacenar subproductos (chala o panca de maíz) de épocas de abundancia para ser utilizadas luego picadas y enmelazadas. También es posible la elaboración y uso de diferentes henos y concentrados.
situaCiÓn aCtual Y proYeCCiones de la CrianZa de Caprinos en el perú
óscar Arroyo. (*)
Ing. zootecnista Msc OnG PROCABRA, Lima, Perú, e-mail: [email protected]

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•291
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Sanidad: Es la parte mas deteriorada de la crianza. No se dispone de calendarios sanitarios, ni vacunaciones ni tratamientos estratégicos. La incidencia de la brucelosis es aún acentuada sobre todo en los lugares mas apartados de las ciudades.. Por las deficientes condiciones de crianza está avanzando el problema de la tuberculosis y la Paratuberculosis, especialmente en la costa central.
Hay una excelente experiencia de la participación del estado, los productores y ONGs para la vacunación de los caprinos contra sus principales enfermedades, como se dio en el Valle del Chillón. Es importante que se continúe con la vacunación contra la brucelosis hasta lograr su control y/o erradicación. Buenos resultados se ha encontrado a la aplicación de calendarios sanitarios.
Mejoramiento genético: Luego de la introducción de algunos ejemplares de las razas Anglo Nubian y Alpinas, y en una menor proporción la Toggenburg, no existen planes de mejoramiento genético de la especie. Los productores no realizan una adecuada selección de los caprinos mas productores, por carencia de registros de producción y solamente intercambian machos reproductores para tratar de evitar la consanguinidad de sus rebaños
Los caprinos que actualmente se crían en el medio requieren mejorar genéticamente para mejorar sus producciones. Introducciones de razas seleccionadas de leche han dado buenos resultados, pero requieren de un seguimiento y adecuada planificación. El uso de la inseminación artificial y la transferencia de embriones serán de mucha utilidad para el rápido mejoramiento de la especie.
2. Producción y transformación de leche
Producción: En promedio nacional la producción de leche de cabra es escasa, llegando a unos 80 Kg. por campaña de 3 a meses. Esta producción se da principalmente por las deficiencias alimenticias de los caprinos. En los criaderos que utilizan suplementación en su alimentación se puede lograr hasta 2.0 y 2.5 Kg. diarios de leche por hembras, en unos 4 a 5 meses de producción.Transformación: Actualmente existe 2 tipos de transformación de los subproductos lácteos: la artesanal y la tecnificada.La artesanal se da en la mayoría de las crianzas, haciendo uso de leche sin pasteurizar, con cuajo natural, sin los cuidados higiénicos adecuados y envases precarios. Estos productos se comercializan en las ferias de pueblo y mercados populares a un costo inferior al del queso artesanal de vaca.En la costa central existen Criadores que producen quesos pasteurizados con alta tecnología y muy buenos estándares de calidad.. Desde hace unos 10 años la Empresa El Bosque produce un queso para untar. El año 2003 el Consorcio PROCABRA/ Asociación de Productores de Caprinos del Chillón ha introducido quesos frescos, semimaduros y de tipo pasta para untar en los Supermercados de la capital. También están produciendo quesos de buena calidad la Empresa Capriole de Chilca y la Familia Igreda del Valle del Chillón. El precio de estos quesos de cabra con alta tecnología son superiores a sus similares de vaca y su mercado va en incremento, lento pero sostenido.
En cuanto a la producción es factible lograr mejores niveles de producción de leche si se logra mejorar las condiciones ambientales de los animales (manejo, sanidad, alimentación, instalaciones), y se tiene una adecuada utilización de razas mejoradas. Esta mejora de la producción tiene que ir acompañada necesariamente de la mejora de la calidad de leche, en cuanto a sus características organolépticas y nutritivas.Referente a la transformación, felizmente es factible acceder a toda la tecnología necesaria para la producción de quesos, yogur y otros subproductos. Se ha probado exitosamente que es factible la producción y comercialización de subproductos lácteos de cabra de alta calidad, con la conjunción del estado, los productores, las ONGs y la actividad privada. Con el actual proceso de regionalización es factible interesar a los gobiernos regionales y locales para el cofinanciamiento de pequeñas Plantas para lácteos caprinos en las zonas del país con mayor producción cabrera y poder desarrollar programas de producción de quesos, leche y otros subproductos, bajo estándares de calidad y también pudieran estar asociadas a los programas sociales de alimentación de las poblaciones más débiles como los niños, las gestantes y los ancianos. De esta forma se podría trazar un importante programa local de lucha contra la desnutrición y el hambre.

292 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
3. Producción de carne
En la costa, otra producción importante de la explotación caprina es la producción de cabritos para carne. Esta es de buena calidad y se consume desde las 4 semanas de edad en diferentes preparaciones culinarias. Lamentablemente en la mayoría de los lugares la producción de cabritos se concentra en 2 épocas del año, adonde los precios caen sensiblemente por la competencia entre los productores. En la sierra también se produce cabritos tiernos, pero dado la lejanía de los mercados hay también una importante producción de cabritos de una mayor edad, de 2 a 6 meses. La carne de los animales adultos, aunque también es consumida por un determinado público, no alcanza la preferencia de la carne de cabrito.Tampoco existe transformación de la carne de caprinos otros subproductos, como embutidos y salchichas.Actualmente en el país no existen razas especializadas de carne ni sistemas de engorde para caprinos adultos.
Sobre la producción de cabritos es necesario planificar su producción para evitar la baja de los precios en determinadas épocas. Es factible la introducción de caprinos de raza Boer para incrementar la producción cárnica de nuestras crianzas, como lo han hecho otros países cercanos como Argentina y Chile.La información procesada permite inferir que la carne de caprino puede ser usada fácilmente para la producción de diferentes tipos de embutidos, hamburguesas, salchichas, etc. que podrían mejorar la utilización de la carne de animales adultos, de una complicada utilización como tal en nuestro medio.Recientemente ha habido una revaloración de la culinaria peruana en general, la misma que se está llegando a exportar a otros países. La carne de los caprinos también puede ser empleada en diferentes preparados, desde platos regionales hasta comida gourmet, con lo que fácilmente puede incluirse en la propuesta global de la culinaria nacional.
4. Producción de pelo, cueros y estiércol
Pelo: Actualmente no existen crianzas dedicadas a la explotación del pelo de cabra. Hace unas décadas hubo una interesante experiencia en la introducción de la raza Angora para la producción de pelo. La crianza se produjo en la zona del Cusco y la poca información que se tubo de ella es que técnicamente fue posible la producción de pelo de caprinos. Lamentablemente esta experiencia fracasó porque no se tuvo asegurado la comercialización.
En los lugares que tienen similitud climática con los lugares de la crianza de las cabras de Cachemira o Angora, es factible la introducción de estos animales para la producción de pelos, la misma que puede darse por empresarios de una amplia visión, en estrecha colaboración de los productores de caprinos.. El círculo de producción estaría completo si estos pelos son tratados y transformados en tejidos y prendas de vestir con un mayor valor agregado.
Cuero: Si consideramos que en país se comercializan anualmente una cantidad cercana a los 500 mil cabritos, lamentablemente la mayoría de las pieles de estos animales, no se utilizan para la curtiembre y su posterior utilización. La razón principal para esta poca utilización es el deficiente manejo inicial de las pieles de los caprinos: sacrificio artesanal, secado al sol y una deficiente conservación.
Es otro rubro que se encuentra prácticamente abandonado. La piel de los cabritos y animales adultos debidamente tratados tienen una amplia gama de utilización en prendas muy finas y otras aplicaciones. Una buena crianza de caprinos podrá lograr buenos cabritos, y animales de saca para la producción de todos los derivados de piel y cuero, con lo cual se mejorarían los ingresos de los productores
Estiércol: El estiércol de los caprinos es del tipo caliente y es un excelente abono para las hortalizas y frutales. Toda la crianza de caprinos puede producir anualmente unas 790 mil TM de estiércol fresco o 9,900 TM de nitrógeno. Lamentablemente este estiércol se deja secar en los criaderos con lo cual se evaporan la mayoría de sus nutrientes. Es ínfima la cantidad de estiércol que se usa en procesos tecnológicos más tecnificados.
En los países en vías de desarrollo el guano de los pequeños rumiantes es una buena fuente de fertilización. La lombricultura, se puede desarrollar fácilmente con el estiércol de los caprinos. Esta naciente industria ya esta exportando ciertos volúmenes a otros países. El uso del estiércol en biodigestores representa otra buena alternativa para lograr mejores ingresos.

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•293
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
5. Mejoramiento de la crianza familiar
En el Perú más de 210,000 familias de pequeños productores agropecuarios se dedican y viven de la crianza de caprinos. Estos pequeños productores en su mayoría ocupan precarias instalaciones para sus crianzas, casi no tienen áreas agrícolas donde sembrar pastos para sus animales. Las crianzas en su mayoría están en manos de personas adultas, porque los jóvenes prefieren migrar a la ciudad para buscar mejores oportunidades de trabajo, que tampoco logran. Estos productores se hallan mas lejos que otros de los centros educativos, el acceso a las Postas de Salud y las principales vías de comunicación. Su vivienda es muy precaria y no disponer de las áreas mínimas para un desarrollo sostenido de su familia. No existen créditos para ninguna de sus actividades económicas no sociales., ni menos sistemas telefónicos ni de internet. Sus organizaciones de productores, con honrosas y poquísimas excepciones no funcionan.
El objetivo central de todos los trabajos de investigación y transferencia de tecnologías en la especie caprina, no debe ser solamente el mejoramiento de esta explotación, sino fundamentalmente el mejoramiento de la familia de criadores. Ningún plan de mejoramiento de la especie tendrá éxito si no van acompañados de planes que mejoren la situación de la familia. Los mejores ingresos que puedan obtener las familias por una mayor producción y venta de sus subproductos deben ser orientado al mejoramiento de la nutrición de la familia, la mejora de su precaria vivienda para hacerla mas higiénica, segura y confortable, la mejora de la sanidad de la familia con prácticas sanitarias adecuadas al medio y finalmente orientadas a una mejor educación de los hijos, la única y real garantía de desarrollo permanente de la familia.
COnCLUSIOnES1. La actual crianza de los caprinos es de carácter artesanal, con poca producción e ingresos, pero con tecnologías medias y poca inversión, puede convertirse en una crianza estabulada o semiestabulada que les permita una mayor producción de subproductos y mejores ingresos económicos.
2. La producción de leche es escasa, pero se ha logrado la producción de quesos de alta tecnología y de venta en las grandes Cadenas de tiendas. Con la participación del estado, los productores, la actividad privada, ONGs y gobiernos locales es factible la organización de Centros de producción de subproductos lácteos de caprinos con mejores tecnologías y a un mejor precio.
3. Con excepción de la carne de cabritos, es escasa la producción de carne de caprinos adultos. Es factible la transformación de esta carne en otros subproductos, que le otorguen un valor agregado. Se debe priorizar la introducción de razas de carne para el mejoramiento de la producción y calidad de esta carne.
4. La producción de cueros y pelos es sumamente escaso, a pesar de las enormes posibilidades de ambos tipos de subproductos. Es recomendable la introducción de caprinos de razas especializadas de pelo, y también es posible la mejora en la obtención, procesamiento y comercialización del cuero de estos animales.
5. Todas las mejoras económicas que se pudieran obtener de la crianza de caprinos deben ser orientadas a mejorar el entorno social de las familias, con
viviendas mas adecuadas, una mejor alimentación y sanidad y fundamentalmente en mejorar la educación de sus hijos.
REFEREnCIASArroyo, O. 1998. Producción de Caprinos. Lima, Codespa – Futuro en marcha/ Procabra/ Ayuntamiento de Madrid. 399 p.
Arroyo, O, Atto, J, Duran, R. y Matossian, C. 2001. Crianza intensiva de caprinos, III: Parámetros reproductivos 1999-2000. In Sevilla, España, Sociedad Española de Ovinos y Caprinos, XXVI Jornadas Científicas. Septiembre 2001.
Arroyo, O. y Matossian, C. 2004. Industrialización de la Leche y subproductos lácteos de caprinos. In Lima, Asociación de Ganaderos Lecheros del Perú, III Congreso Nacional de Producción lechera, PERULACTEA 2004. Octubre 2004. 16 p.
Arroyo, O. y Briones, I. 2005. Manejo ganadero y Mejoramiento genético de los caprinos de los Valles costeños. Lima, Convenio Procabra/ Manos Unidas. Septiembre 2005. 36 p.
Ramet, J.P., Arroyo, O. y Fung. L. 2005. La producción de Leche, la fabricación de quesos y la viabilidad de luna pequeña quesería de quesos de cabra. Lima, Convenio PROCABRA / FAO / MINAG. Diciembre 2005. 39 p.
Nolte, E. 2006. Reingeniería de la producción caprina en el Valle del Chillón. Lima, PROCABRA, Ciclo 2006 de Capacitación a Productores de Caprinos. Enero 2006. 4 p.

294 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
RESUMEnA pesar de que existe diversa información científica, que reporta los beneficios de utilizar caprinos (Capra hircus hircus) para controlar vegetación indeseable, en diversas condiciones ambientales, lamentablemente este enfoque no es utilizado frecuentemente. La preocupación por el ambiente, y los incrementos en los costos de los herbicidas y de los métodos de control de malezas mecánicos, proporcionan nuevas oportunidades para el empleo de caprinos como agentes para el control biológico de vegetación indeseable. Los caprinos tienden a realizar un pastoreo conservativo “autocontrolado”, mejor que otras especies de ganado doméstico, cuando se utiliza un enfoque sustentable; bajo otras circunstancias, los caprinos podrían llegar a ser destructivos. Sin embargo, cualquier riesgo ambiental asociado con los caprinos, esta vinculado, en esencia, a un manejo inadecuado de esta especie por el hombre.
Palabras claves: Caprinos, Ramoneo, Vegetación Indeseable, Pastoreo, Control Biológico.
ABStRACtThe benefits of using goats (Capra hircus hircus) to control undesirable vegetation in many different environmental situations are well documented by research data. Nonetheless, this approach is severely underutilized. Environmental concerns and the increased costs of herbicides and mechanical control methods provide new opportunities to use goats as biological agents for the control of unwanted vegetation. The goat lends itself to conservation grazing better than any other domestic livestock species
utiliZaCiÓn de Caprinos para el Controlde vegetaCiÓn indeseaBle
Luginbuhl J-M1; Pietrosemoli Castagni, S2
Use of goats to control undesirable vegetation
1Departments of Crop Science and Animal Science, College of Agriculture and Live Sciences, north Carolina State University, Raleigh nC, USA. Email: [email protected] de zootecnia, Facultad de Agronomía, La Universidad del zulia, Maracaibo, Venezuela. Email: [email protected]
when utilized in a sustainable manner. Goats can be quite destructive, however, but any environmental hazard associated with goats is essentially a failure in adequate management by man.
Key words: Goats, Browse, Undesirable Vegetation, Grazing, Biological Control
CAPRInOS PARA EL MAnEJO DE VEGEtACIón – LInEAMIEntOS GEnERALES SOBRE EL COMPORtAMIEntO ALIMEntICIO DE LOS CAPRInOS Los caprinos han sido clasificados como una especie con hábitos alimenticios intermedios, adaptados tanto al pastoreo como al ramoneo, exhibiendo cambios en la selectividad de la dieta en función de la disponibilidad de forraje, su valor nutritivo y de la estación (Hoffman, 1989; Van Soest, 1994; Pietrosemoli et al., 2005). De acuerdo a Malecheck and Leinweber (1972), los caprinos son muy flexibles en sus hábitos alimenticios, capaces de ajustarse de una dieta compuesta por un 80% de ramoneo, a una dieta compuesta de 80% de gramíneas, siendo el origen de estos cambios marcadas variaciones en la disponibilidad de forraje. Coblentz (1977) clasifico a los caprinos como “oportunistas genéricos” debido a que ellos consumen la vegetación más nutritiva y palatable disponible, seleccionando una amplia variedad de plantas de una calidad superior que los vacunos y los ovinos (Fraps and Cory, 1940). Comparados con otros rumiantes domésticos, los caprinos seleccionan las partes y porciones más nutritivas de las plantas, y en una selección dada entre gramíneas, hierbas y arbustos, ellos usualmente prefieren dietas con una mayor proporción de arbustos. (Wilson et al., 1975; Bryant et al., 1979). Malecheck and Provenza (1981) señalaron que los caprinos seleccionaron 60% de arbustos, 30% de gramíneas y 10 % de malezas de hoja ancha, comparados con la selección hecha por los ovinos, la cual estuvo compuesta por 20 % de arbustos, 50% de gramíneas y 30% de malezas de hoja ancha. Los caprinos consumen de buena gana gramíneas inmaduras, pero cambian al ramoneo

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•295
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
cuando la calidad del forraje disminuye con el avance de la madurez. Sin embargo, demuestran una preferencia especial hacia las inflorescencias de las gramíneas (Wilson, 1957; Luginbuhl et al., unpublished). Frecuentemente, seleccionan también otras plantas herbáceas que florecen (Malechek and Leinweber, 1972 ).
Los caprinos presentan una mayor masa hepática en relación a los vacunos y a los ovinos, y toleran mayores niveles de compuestos fenólicos tales como taninos (Silanikove, 1997). Esto podría llegar a explicar el porque, los caprinos son generalmente más efectivos que los ovinos, en el control de euphorbia usula L., una dicotiledónea herbácea perenne, con un sistema radicular profundo, que contiene varios compuestos aleloquímicos (Walker et al., 1994). Adicionalmente, los caprinos tienen menores problemas de toxicidad debido a que prefieren consumir una gran variedad de especies de plantas a lo largo del día. La selectividad en la alimentación, y una fuerte preferencia por el ramoneo, permiten a los caprinos reducir los efectos de la variación en la energía y la proteína de la dieta, causada por las condiciones ambientales o de manejo (Fedele et al., 1991). Mas aun, debido a esta versatilidad del comportamiento pastoreo/ramoneo, los caprinos son capaces de controlar efectivamente vegetación invasiva, mientras que al mismo tiempo seleccionan una dieta capaz de satisfacer sus requerimientos nutricionales (Child et al., 1985). Este comportamiento “oportunistico” ha servido muy bien a los caprinos en circunstancias donde otras especies de rumiantes domésticos se hubiesen encontrado en clara desventaja o aun en situaciones de deficiencias nutricionales (Coblentz, 1977; Mackenzie, 1993).
Con el objeto de describir el comportamiento de caprinos en pastoreo en praderas naturales de Uruguay, Perez (1998), señala que los caprinos estan más adaptados que otras especies domésticas a condiciones de vegetación heterogénea, donde pueden desarrollar su capacidad para obtener una dieta de mayor valor nutritivo. Indica que esta especie demuestra preferencia por especies vegetales que realizan altas contribuciones y pueden localizarse en alta frecuencia. Refiere que las gramíneas de porte bajo (stipa spp. y Piptpchaetium spp.), arbustivas, tanto compuestas como umbelíferas, y arbóreas (Acacia cavenm scutia buxifolia y Celtis spinosa) son seleccionadas aun cuando se ubiquen con bajos niveles de disponibilidad, deduciéndose que existe una importante selección en las especies que tienen el mayor aporte de fracción hoja en el estrato medio y alto del sotobosque. La actividad principal del rebaño es la ingestión de alimento, dedicándole entre 58,1 y 92,2 % del tiempo diurno, relacionándose con los tipos de vegetación, y con el grado de homogeneidad de la pradera en términos de calidad y del tipo de especies
que la componen. Durante la mañana, el período de ingestión de alimento se orienta hacia el ramoneo de árboles antes que al pastoreo en los estratos bajo y medio. Esto disminuye a medida que baja la disponibilidad de especies arbóreas, dirigiéndose a la búsqueda de alimento en zonas de estrato bajo. Simultáneamente, la intensidad de pastoreo se vería disminuida por el aumento en la búsqueda de alimento, y el mayor traslado y dispersión de las cabras.
El pastoreo de ganado doméstico puede haber sido la herramienta más temprana de manejo de vegetación, empleada por los humanos. Debido a su inusual preferencia por las hojas y ramitas terminales de las plantas leñosas, los caprinos han sido utilizados como una alternativa a los herbicidas y al corte mecánico en contra de malezas herbáceas invasivas y de especies arbustivas, gracias a que las plantas a controlar les son palatables. La propensión de los caprinos a pararse sobre sus patas traseras, les permite ramonear hasta una altura de 2 metros, en áreas donde se encuentran árboles y bejucos colgantes (Lu, 1988). Owens (1991) indicó que a medida que se incrementaba la densidad de caprinos por unidad de área la utilización, en la comunidad de arbustos de Acacia de alturas entre 0.75 y 1.5 m, se incrementó a una tasa más rápida que en otras zonas de la estructura. Los caprinos inclusive escalan ciertos árboles (García y Gall, 1981). Los caprinos han sido también exitosamente pastoreados en combinación con ganado vacuno (Bos taurus), ovino (ovis aries), o equino (equus caballus).
El control exitoso de malezas y matorrales depende entre otros factores de que los caprinos simplemente consuman la planta (Lambert et al., 1989). En ocasiones, los caprinos ramonearan para variar su dieta ejerciendo poco efecto sobre la población vegetal blanco del programa de manejo. Por el contrario, algunas plantas pueden ser pastoreadas únicamente, cuando los caprinos son forzados a hacerlo ya que se les ofrece como único componente de la dieta. En algunos casos específicos, el papel potencial de la ganadería como diseminador de malezas no puede ser descartado. No existe evidencia que soporte que los caprinos son ambientalmente más riesgosos que otros rumiantes (Harrington, 1982). Sin embargo, la carencia de un programa de manejo adecuado puede provocar la destrucción de toda la cobertura vegetal, debido a que los caprinos no seleccionan exclusivamente la vegetación indeseable (Wilson et al., 1975), pudiendo resultar en una comunidad vegetal dominada por malezas y matorrales. A diferencia de vacunos y ovinos, los caprinos pueden destruir árboles pequeños y vástagos al consumir la corteza, y no se detienen ante la vegetación espinosa (Wood; 1987). De acuerdo a Mellado et al. (2003), décadas de sobrepastoreo en las praderas del desierto de Chihuahua, México, han forzado a los caprinos a

296 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
alterar su patrón de selección de la dieta, consumiendo especies más resinosas, ásperas e inclusive toxicas. Esta adaptación para sobrevivir estuvo asociada con un estatus nutricional más bajo, una ganancia de peso diaria negativa, menor puntaje de condición corporal y menor fertilidad.
Alvarez (1995), refiere que los caprinos representan un papel fundamental en la recuperación de praderas y pastizales invadidos por matorrales. Acota que en pastoreo libre, esta especie nunca consume menos de un 50% de vegetación arbustiva, aun cuando disponga de otras opciones. Señala que los hábitos alimenticios, consistentes en seleccionar las fracciones de mejor contenido nutricional de la vegetación, les imponen el recorrido de grandes áreas en épocas de escasez para satisfacer sus requerimientos nutricionales, lo que logran gracias a la combinación de gran cantidad de especies vegetales: herbáceas, arbustivas e inclusive arbóreas. Los caprinos son capaces de utilizar forrajes de baja calidad, como forrajes con alto contenido de fibras y arbustos leñosos, debido a la gran capacidad de consumo, que puede llegar a un 6 % de su peso vivo/día, gracias a su aparato digestivo que representa aproximadamente el 33% de su peso vivo. La tasa de consumo varia en función de la época, factor que afecta también la composición de la dieta, como consecuencia de variaciones en la fenología de las especies vegetales. Usualmente durante las épocas de mayor humedad se observa una mayor ingestión de especies herbáceas, mientras que en las épocas secas se observa una mayor ingestión de matorrales y arbustos. Otras ventajas de los caprinos en relación a otras especies de animales domésticos son su capacidad de digerir alimentos con mayor contenido de fibra, mayor numero de bacterias celulíticas a nivel ruminal, mayor secreción de saliva, consumo metabólico de agua muy reducido, cualidad que les permite adaptarse a un bajo consumo de agua, y en consecuencia recorrer mayores distancias sin beber agua.
Una herramienta con potencial en la batalla contra la vegetación indeseable, también referida como “pastoreo dirigido” (prescription grazing), es la aplicación de pastoreo en una estación especifica, para una duración e intensidad determinadas, y para alcanzar objetivos de manejo de vegetación muy concretos (Frost and Launchbaugh, 2003). De acuerdo a estos autores, un “pastoreo dirigido” efectivo, debería ocasionar un daño significativo a las plantas blanco del programa de control, limitar el daño irreparable a la vegetación circundante, ser consistente con los objetivos productivos del ganado y, de ser necesario, debería estar incluido con otros métodos de control, como parte de una estrategia integrada de control de vegetación indeseable. Mas aun, una comprensión sólida de ecología vegetal, comportamiento animal y de las interacciones planta animal se requieren
para formular un “pastoreo dirigido” efectivo. En este contexto, los caprinos ofrecen una opción viable para lograr el manejo y control de plantas indeseables, mientras que proveen ingresos adicionales a los productores, en la forma de productos para la venta.
CAPRInOS PARA EL MAnEJO DE VEGEtACIón – EStUDIO DE CASOS.
Control de vegetación de matorral en los pastizales montañosos. Gran parte de las áreas de pastizales de las montañas Appalachian en los Estados Unidos, esta dominada por malezas herbáceas, matorrales y por arbustos de rosa multiflora (Rosa multiflora Thunb.), especie introducida desde Japón en 1886 como patrón de injerto para rosas ornamentales (Mays and Kok, 1988). La invasión de tierras productivas por R. multiflora, se ha incrementado durante los últimos 30 años, y en caso de infestación severa, el acceso a los pastizales y a las áreas recreacionales se ha visto marcadamente restringido. Las semillas de R. multiflora pueden permanecer viable en el suelo por más de veinte años (Harvey, 1996). Nuevos arbustos pueden desarrollarse con semillas transportadas por pájaros, roedores y por el agua. En consecuencia, es necesario llevar a cabo durante años, un programa integrado de manejo efectivo, después de controlar o eliminar las plantas originales (Kay et al., 1995). En el pasado, controlar R. multiflora usualmente implicaba el corte mecánico y el uso de herbicidas. Sin embargo, el incremento en las restricciones en el uso de herbicidas, y los elevados costos del corte mecánico, complican el uso de estos procedimientos de rescate. Adicionalmente, el uso de herbicidas podría afectar a la vegetación deseable. En consecuencia, se requieren alternativas de bajo costo, bajos insumos y ambientalmente aceptables, para proveer a los propietarios de mecanismos que les permitan mantener esos pastizales en producción, y para integrarlos en sistemas de producción sustentables.
Dos proyectos de investigación se llevaron a cabo en las montañas occidentales de Carolina del norte (Luginbuhl et al., 1999 y 2000). Las plantas y bejucos dominantes en el área eran arbustos de R. multiflora, algunos árboles jóvenes de especies maderables, tres especies forrajeras de importancia (Poa pratensis L., Festuca arundinacea L. Schreb., y trifolium repens L.) y seis especies indeseables (Rubus spp., Lonicera japonica Thunberg], mentha L. spp., muhlenbergia schreberi J.F. Gmelin, toxicodendron radicans [L.] Kunt, y Fragaria virginiana Duchesne). En el primer experimento (Luginbuhl et al., 1999) se evaluó la efectividad de utilizar únicamente caprinos criollos (30 cabras/ha) o vacunos con caprinos (17 cabras /ha + 2 a 3 novillos/ha - 225 kg promedio de peso vivo) para recuperar un pastizal de una plantación de

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•297
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
manzanas de 8.4 has abandonada durante 15 años, invadida de malezas. Los pastoreos se realizaron durante 45 a 60 días desde Mayo a Julio, y por otros 24 a 35 días en Septiembre y Octubre. Por más de cuatro estaciones de pastoreo, el manejo de la defoliación origino un incremento sustancial en la cobertura vegetal herbácea, entre 65 y 86% en las parcelas manejadas con caprinos únicamente, y entre 65 a 80% en las parcelas con caprinos y vacunos, mientras que la cobertura vegetal disminuyo de 70 a 22% en las parcelas del tratamiento control (Figura 1). Similarmente, la cobertura de especies de gramíneas se incrementó en las pasturas pastoreadas (caprinos: 16 a 63%; caprinos + vacunos: 13 a 54%), mientras que en las pasturas no pastoreadas alcanzo un 10% en promedio (Figura 2). El cambio en la composición botánica en las pasturas pastoreadas, fue atribuido a la preferencia de los caprinos por las especies de hoja ancha, situación que permite a las gramíneas ser más competitivas. Adicionalmente, los arbustos de R. multiflora fueron prácticamente eliminados, como se demuestra por reducciones en la altura promedio de 2.1 m a 0.6 m (Figura 3), del área promedio de la copa de 12 a 2.5 m2, y por el numero de tallos muertos en ambos casos, en las pasturas con caprinos (100%) o con caprinos y vacunos (92%). Luego de dos años de reposo, vacunos solos (1.7 novillos/ha) o caprinos + vacunos (1.7 novillos/ha y 3.4 caprinos/ha) fueron pastoreados rotacionalmente en el mismo lugar (Luginbuhl et. al., 2000). Durante las cuatro estaciones de pastoreo, la cobertura de las especies herbáceas permaneció alta (81% en promedio) en las pasturas pastoreadas, pero disminuyo a 39% en las pasturas control. Por el contrario, la cobertura de especies herbáceas gramíneas disminuyo linealmente en las pasturas control, de 78% en Mayo 1996 a 39% en Octubre 1999. El cambio en ambos parámetros, cobertura vegetal y composición botánica en las parcelas sin pastoreo fue atribuido al desarrollo de los matorrales, árboles y arbustos de R. multiflora que hicieron sombra sobre la vegetación herbácea del suelo. Los bejucos de Rubus spp. and L. japonica prácticamente fueron eliminados de las parcelas bajo pastoreo. Los arbustos de R. multiflora crecieron hasta un altura de 1.8 m en las pasturas donde pastoreaban vacunos exclusivamente, pero se mantuvieron bajos (0.6 m en promedio) en las pasturas de caprinos + vacunos (Figura 4). Similarmente, la copa de R. multiflora fue bien controlada en las pasturas donde se encontraban caprinos y vacunos (0.5 m2 en promedio), pero se incremento a 7 m2 donde pastoreaban los vacunos. Los árboles de Robinia pseudoacacia L. fueron ramoneados tanto por los caprinos como por los vacunos y fueron prácticamente eliminados de las parcelas en pastoreo (Figura 5).
Pastoreando en una línea de energía eléctrica durante cinco años en Virginia del Oeste, los caprinos
redujeron la cobertura de matorrales de 45 % a 15% en un año, mientras que a los ovinos les tomo tres años alcanzar los mismos resultados (Magadlela et al., 1995). Luego de cinco años de pastoreo, los caprinos habían reducido la cobertura de matorrales al 2%. La efectividad animal, principalmente la de los ovinos, se incremento con el corte mecánico y la aplicación de herbicidas, pero los costos también se incrementaron. Los caprinos solos, fueron el método de eliminación de malezas más económico, con un costo de $ 33/ha, mientras que la aplicación de herbicida seguida de caprinos o de ovinos, $ 593/ha, fue el método más costoso (Tabla 1). El manejo del pastoreo que incluye la defoliación de matorrales temprano en la primavera y repetidamente durante la estación de crecimiento, fue muy efectiva. Por otro lado, la defoliación completa empezando en Agosto/Septiembre no tiene impacto significativo en la vegetación leñosa y el rebrote fue del 100% en la primavera siguiente.
En Virginia, gran parte de la tierra utilizada por operaciones de minería de carbón al aire libre, es restaurada a una condición apropiada para el pastoreo con ganado. Sin embargo, estas tierras son a menudo abandonadas debido a la dificultad en controlar la vegetación leñosa. Una mayor utilización del pastizal fue observada donde los caprinos y los vacuno co-pastoreaban bajo pastoreo rotacional (Webb et al., 2007). En las parcelas co-pastoreadas, la vegetación total fue utilizada más uniformemente que en las pasturas de vacunos solamente, ya que los caprinos agresivamente ramonearon arbustos de olivo otoñal (eleaganus umbellata Thunb.) (Figura 6). Adicionalmente el componente de malezas del pastizal, fue más bajo en las pasturas co-pastoreadas o en las pasturas con los vacunos solamente (14.5% y 16.0%, respectivamente) comparadas con las de control, no pastoreadas (44.4%) al final de la primera estación de pastoreo.
ulex europaeus, un arbusto nativo de Europa central y occidental, y de las islas británicas, fue introducido a Australia y Nueva Zelandia a mediados del siglo 19 como forraje para ovinos y como setos. Para 1900, u. europaeus fue declarado maleza perjudicial en esos países. Hoy en día, se encuentra en la mayoría de las áreas templadas del mundo, y es considerado una maleza en Chile, Irán, Italia, Polonia, Noroeste de España, Estados Unidos y Tasmania. En Nueva Zelandia, los caprinos han sido empleados en el control de esta especie vegetal (Lambert et al., 1989). Una carga continua durante cuatro años, con 33 caprinos por hectárea prácticamente erradico u. europaeus (Tabla 2), mientras que para el año cuatro, u. europaeus, había eliminado el pastoreo de ovinos. Cargas más livianas de hasta 6 caprinos por hectárea, a pesar de tomar un poco más de tiempo para lograr los objetivos, serian más sustentables debido a

298 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
incrementos en la productividad y rentabilidad de los caprinos.
En Nueva Zelandia, los caprinos fueron también utilizados para controlar exitosamente onopordum acanthium L., un miembro de la familia Aster, introducido desde Europa o Asia Oriental. Esta especie puede alcanzar alturas de 2.7 metros y convertirse en un obstáculo infranqueable para el ganado en las praderas y pasturas. Lambert et al. (1989) reportan una rápida reducción en el tamaño de la planta, el numero de inflorescencias y de la sobrevivencia de la planta, cuando caprinos pastorearon o. acanthium (Tabla 3), mientras que aparentemente los ovinos no consumieron las inflorescencias, con tasas de sobrevivencia de las plantas del 48% luego de dos años de pastoreo.
En España, se estudio durante tres años el efecto del pastoreo de caprinos de raza local a baja carga, versus cabras cachemir a cargas altas o bajas. Las cabras locales disminuyeron más la abundancia de plantas leñosas que las de raza Cachemir, en particular en el caso de especies de porte alto como erica australis y e. arbórea, las cuales fueron ramoneadas más intensamente. La presencia de especies herbáceas en la estructura se incremento en los tratamientos con cabras locales. Entre los tratamientos con cabras de raza Cachemir, las mayores cargas provocaron mayores disminuciones en la presencia de especies leñosas que las cargas bajas. Los mayores índices de diversidad fueron observados en los tratamientos con cabras Cachemir a altas cargas, y los más bajos en los tratamientos con la raza Cachemir a cargas bajas, en las cuales la dominancia de los brezos (erica spp.) y del tojo (ulex galii) casi no presento modificación después de las tres estaciones de pastoreo, limitando la presencia de otras especies. Este índice fue intermedio para la raza local a baja carga (Celaya et al., 2005).
Control de Matorrales y Pastoreo en las Praderas Desérticas. En el norte de México, la invasión de vastas áreas de praderas y de pastizales productivos por eysendhartia spinosa y mimosa biuncifera es un problema serio (Fierro et al., 1982). Las cargas animales se establecieron entre 150 y 300 caprinos-día por hectárea, con o sin corte mecánico. Los efectos del ramoneo de los caprinos fueron notables luego del primer año. Después de cuatro años de ramoneo, la mortalidad de e. spinosa fue de 36%, mientras que para m. biuncifera, especie más resistente, la tasa de mortalidad fue solamente del 8%. Sin embargo, más del 60% de las plantas fueron afectadas, reduciendo la cobertura aérea de los matorrales casi por 20%. El mayor impacto de la vegetación de matorral, ocurrió en los pastizales a los que no se les realizo poda mecánica y con la mayor carga. La productividad de
los caprinos fue mayor en las pasturas no podadas debido a una mayor disponibilidad de forraje de calidad.
En las áreas áridas y semiáridas de México, los caprinos son manejados bajo sistemas extensivos tradicionales de la aldea, y son pastoreados en praderas naturales de la comunidad a través del año sin suplementación alimenticia. Existe poca información científica del efecto ecológico de un pastoreo intensivo de caprinos en estos ecosistemas áridos. Mellado et al., (2003) investigaron los efectos de la presión de pastoreo (1.5 ha/caprino vs. 15 ha/caprino) en las praderas del desierto de Chihuahua sobre la cobertura vegetal, la composición botánica de la dieta, los metabolitos sanguíneos y niveles minerales, y la fertilidad de caprinos tipo de leche. Las praderas pastoreadas tuvieron ligeramente una mayor (P < 0.05) cobertura vegetal total (38.6 vs. 30.4%) que aquellas con una sobrecarga. La proporción total de arbustos en la dieta de los caprinos fue mayor (86.4 vs. 72.4 % a finales de la estación seca, 78.6 vs. 42.1% a finales de la estación lluviosa; P < 0.05) en las praderas con una alta carga que en las de carga baja. La proporción de malezas de hoja ancha en la dieta fue menor (P < 0.10) a finales de la estación seca (11.4 vs. 21.5%), inicios (55.4 vs. 64.0%) y finales de la estación húmeda (15.0 vs. 45.8%) en las praderas con alta carga en comparación con los de baja carga. Al inicio de la estación de apareamiento se establecieron substancialmente menores (P < 0.01) valores de glucosa sérica, urea-nitrógeno, zinc y magnesio (Tabla 4) en las cabras en las praderas altamente pastoreadas versus las ligeramente pastoreadas, lo que provoco mayores (P < 0.01) tasas de aborto (22 vs. 12%) y consecuentemente una menor (P < 0.05) tasa de nacimientos (42 vs. 55%).
En las zonas áridas del estado Lara, Venezuela, funcionó un centro experimental para producción de caprinos. Investigaciones realizadas sobre el efecto del pastoreo de esta especie, llevados a cabo por Smith y Rivero (1983) y Garcia, Salazar y Smith (1990), demuestran que el sobrepastoreo provoco disminución en la densidad de los arbustos más altos (más del 80%) y de la diversidad del matorral ralo. Garcia y Salazar (1986), reportan una reducción de 44 a 25 especies de plantas entre una zona protegida y una zona en condiciones de sobrepastoreo. Adicionalmente señalan cambios marcados en la composición botánica, siendo eliminadas completamente por el ramoneo especies de arbustos altos como medelia calycina, Calea bertoroana y Baccharis brachylaenoides. Otros arbustos bajos como Casearia arguta y Cordia steyermarkii resultaron favorecidos, lo que produjo un cambio de la fisonomía de la vegetación hacia un arbustal más bajo. Sin embargo, en ambos casos la cobertura vegetal no se vio sustancialmente

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•299
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
modificada. Smith y Rivero (1983), indican que los caprinos tienen un impacto moderado sobre la cobertura vegetal y su capacidad de reducción de los efectos de la erosión. Sin embargo, al producirse modificaciones en la composición botánica, se produce una unidad forrajera de bajo valor nutricional, afectando la sustentabilidad del sistema.
Con el objeto de establecer la capacidad de carga y la composición botánica de la ingesta de los caprinos de las unidades forrajeras más importantes en un ecosistema árido en Argentina, Allegretti et al. (2005), realizaron un estudio empleando el método de Point Quadrat modificado para el monte, así como análisis microhistológicos de heces. La capacidad de carga anual estimada fue para las unidades forrajeras más importantes: algarrobo (Prosopis flexuosa) 1.2; zampal (Atriplex lampa) 1.7; usillar (tricomaria usillo) 3.5; jarillal (Larrea cuneifolia) 5.8 y medanal 4.3 ha/unidad ganadera caprina, respectivamente. La composición de la dieta varió durante las estaciones, observándose una predominancia de las especies arbustivas, mientras las gramíneas perennes aparecieron en una frecuencia baja en todas las estaciones. El algarrobal y el zampal son las unidades forrajeras de mayor importancia, con una mayor receptividad de forraje y una productividad más estable durante todo el año, en relación a las otras unidades forrajeras analizadas. Mediante la técnica de observaciones visuales, estos autores establecieron que especies como el algarrobo y el usillo contribuyen con material forrajero casi todo el año, durante el verano – otoño época que coincide con su rebrote con hojas, flores y/o frutos, y durante la época seca y fría con material vegetal caído al pie de la planta de donde es aprovechado directamente por los caprinos. Durante todas las estaciones, los arbustos fueron los más consumidos, durante la primavera y el verano las gramíneas prevalecieron, mientras que durante el otoño - invierno las especies arbóreas constituyeron gran parte de la dieta de los caprinos.
Control de vegetación de cobertura en el sotobosque en bosques mixtos leñosos. Los caprinos presentan un gran potencial como agentes biológicos para controlar la vegetación del sotobosque en bosques leñosos, debido a inquietudes ambientales relacionadas con el uso de herbicidas y a la prohibición en contra del herbicida 2,4,5 T como herramienta de manejo.
En Arkansas, se analizaron dietas seleccionadas por cabritos machos, de los tipos de leche y de carne, que fueron rotados entre cuatro parcelas de bosques leñosos de tierras altas de Julio a Diciembre (Hansen et al., 1980; Child et al., 1985). Durante los meses de Julio y Agosto (verano), más del 65% de las dietas seleccionadas por los caprinos (Figure 7) estuvieron compuestas de bejucos, principalmente L. Japonica,
smilax spp. y Berchemia scandens. Los bejucos, se mantuvieron como una porción significativa de las dietas a través del período de estudio. A medida que los bejucos se hicieron menos disponibles, la proporción de otros arbustos y coníferas se incremento en la dieta. Las malezas de hoja ancha desaparecieron de la dieta después de tres meses, mientras que las gramíneas incrementaron su importancia según avanzaba la estación. A medida que se incrementaba la severidad de los períodos de otoño (21 de Setiembre a 20 de Diciembre) e invierno, se incrementó la cantidad de bellotas, hojas muertas y agujas de pino. No se provoco un daño excesivo a los árboles de pino, sino hasta que los otros forrajes fueran consumidos, y durante los meses de otoño e invierno, cuando el forraje verde escaseaba. Las dietas de los cabritos de leche y de carne fueron similares, indicando que la raza no fue un factor de importancia para manipular la composición botánica. La vegetación del sotobosque puede preservarse o destruirse dependiendo de la intensidad de la carga y de la frecuencia de pastoreo.
Más de 988 “días de alimentación por caprino por hectárea” se obtuvieron de un bosque de madera típico, sin destruir la vegetación existente (Langston, 1992). Los caprinos pastorearon los matorrales por dos años consecutivos, una vez en primavera, y una vez a finales del verano, a una intensidad de carga de 86 animales/ha por períodos entre 3 y 5 días. Los caprinos ramonearon de buena gana L. japonica, Rubus spp., Ligustrum vulgare, Pueraria montana y hojas de plantas leñosas y plántulas. Por el contrario, no pastorearon muhlenbergia schreberi, F. virginiana o Phaethusa occidentalis L. independientemente de la intensidad de pastoreo. La densidad de estas tres especies de matorrales, de hecho se incremento durante la duración del experimento.
Control de Pueraria Montana. Pueraria montana es una de las plantas no nativas más agresiva de las tierras boscosas de sudeste de los Estados Unidos. Más de 500.000 hectáreas han sido invadidas por P. montana, un bejuco importado de Japón y China a inicios de 1900 para el control de la erosión (Miller, 2004). Para su control se han empleado herbicidas, pero estos químicos son costosos, y usualmente se requieren aplicaciones repetidas. Adicionalmente, las inquietudes ambientales relacionadas con el empleo repetido de químicos no deben ser minimizadas.
En Alabama, se mantuvieron machos castrados Angora pastoreando P. montana en una plantación de pinos (Pinus taeda y P. Palustris) durante seis semanas (Septiembre 14 hasta Octubre 26); cuando se pastorearon 10 o 20 cabras/ha, ganaron 160 y 180 g/animal/día, respectivamente (Bonsi et al., 1991). El porcentaje de árboles de pino dañados se incremento, a medida que se incrementaba el tiempo en el que

300 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
los animales permanecían en el área de estudio. En otro estudio, caprinos en crecimiento (raza y sexo no reportados) pastoreados rotacionalmente, a una carga de pastoreo de 27 animales/ha, en praderas infestadas con P. montana, ganaron 160 g/d (Woldeghebriel et al., 1992).
En la universidad del Estado de Carolina del Norte, machos cabríos en crecimiento y machos castrados criollos, fueron continuamente pastoreados durante 49 y 26 días a tasas de 59 0 99 cabezas/ha, respectivamente, resultando en 52.9 y 78.3 kg ganancia/ha por la duración del período de pastoreo (Luginbuhl et al., 1996). En un estudio de campo, en el Centennial Campus de la mencionada universidad (Luginbuhl, Spooner y Glennon; unpublished), cabras adultas mestizas Boer fueron pastoreadas a una intensidad de carga de 82 cabras/ha en parcelas de P. montana. Los períodos de pastoreo ocurrieron del 2 al 6 de Junio, del 23 al 26 de Junio, del 16 al 17 de Julio, del 4 al 6 de Agosto, del 2 al 4 de Septiembre y el 6 al 7 de Octubre. La altura de pastoreo inicial varió entre 76 y 140 cm. Al cuantificarse la biomasa vegetal, se establecieron 2.362 kg de hojas y 544 kg de tallos consumibles en promedio por hectárea. Las hojas y tallos presentaron promedios de proteína cruda de 23,3 y 11,9%, fibra neutro detergente de 38,6 y 57,2% y lignina de 3,9 y 7,4 %, respectivamente. Al inicio de la siguiente estación de pastoreo, más del 98% de P. montana había desaparecido, un claro indicador de que esta especie puede ser controlada por caprinos, debido a su sensibilidad a defoliaciones repetidas.
PAStOREO MULtIESPECIES O COMPLEMEntARIOSegún Osoro et al. (2005), la conducta de pastoreo de las especies de herbívoros domésticos varia en forma notable. Así por ejemplo, los vacunos muestran preferencia por especies herbáceas, demostrando los equinos un comportamiento similar; en cambio los caprinos prefieren la vegetación leñosa o arbustiva inclusive las partes más lignificadas de las herbáceas. Los ovinos demuestran un comportamiento herbívoro típico, con la capacidad de seleccionar las especies de mayor calidad nutritiva, como t. repens o los brotes tiernos de las especies leñosas que conforman los matorrales. Se ha observado diferencia en la capacidad de las especies antes mencionadas de modificar su conducta alimenticia en función de la variación de la disponibilidad de la vegetación de su preferencia. Por ejemplo, se ha observado como los caprinos permanecen más tiempo de pastoreo en el brezal (erica spp.) - tojal (ulex galii), que en áreas de pastos mejorados cuando hay disponibilidad abundante de los primeros, incrementando paulatinamente el tiempo de pastoreo en áreas mejoradas cuando se reduce la vegetación natural (Celaya et al., 2005).
Existe la necesidad de evaluar el efecto del pastoreo de caprinos con otras especies herbívoras, en la composición botánica de las pasturas y en la producción del ganado como un todo. Los caprinos pueden pastorear en combinación con vacunos (Bos taurus), ovinos (ovis aries) o equinos (equus caballus). En Carolina del Norte, se ha estimado que la mayoría de productores de vacunos de carne, tendrían forraje subutilizado o en exceso, como para alimentar 1 a 2 caprinos por vaca sin necesidad de realizar ningún aporte alimenticio adicional (Luginbuhl et al., 1995). Los efectos complementarios del pastoreo de vacunos y caprinos en la misma finca, ofrecen una oportunidad para resaltar y expandir la industria de la carne existente. Los otros beneficios es que los caprinos utilizan y en consecuencia suprimen plantas no pastoreadas por los vacunos (Taylor, 1985), permitiendo un mayor crecimiento de las gramíneas para los vacunos. Taylor (1985), también reportó que los vacunos ganaron más peso por cabeza cuando se pastorearon con caprinos y ovinos que cuando se pastorearon solos. du Toit (1972) en el Suroeste Africano, reportó que en un matorral, el ganado presentó un mayor nivel de producción cuando pastoreaba en combinación con caprinos que cuando pastoreaba en conjunto con ovinos. De acuerdo a Rector (1983), los vacunos en una pradera natural de Texas, seleccionaron predominantemente gramíneas (70%), con menores cantidades de malezas de hoja ancha (5%), ramoneo (24%) y Carex spp. (1%). Los ovinos consumieron predominantemente gramíneas (52%), pero más malezas de hoja ancha (15%), ramoneo (31%) y Carex spp. (2%) que los vacunos. Por el contrario, los caprinos prefirieron el ramoneo (70%), las gramíneas (20%), y algunas malezas de hoja ancha (10%). Merrill and Taylor (1981) indicaron que en Texas, la utilización de las praderas y la capacidad de carga fue mejorada 10 a 25% por un pastoreo combinado de caprinos, vacunos y ovinos, un claro indicador de que estas especies de ganado no solamente son compatibles, sino también complementarias. Taylor and Ralphs (1992) reportaron una reducción en las pérdidas de ganado por plantas toxicas debido al co-pastoreo. Finalmente, los beneficios adicionales del pastoreo de multiespecies, puede incluir una disminución de los problemas de parasitosis gastrointestinales cuando vacunos o equinos son pastoreados con caprinos (Hart, 2001).
REDUCCIón DEL RIESGO DE InCEnDIOS FOREStALES MEDIAntE PAStOREO COn GAnADODe acuerdo con investigaciones meteorológicas, los últimos años han sido los más calurosos en los registros climatológicos. Como consecuencia, cada año los incendios forestales queman amplios territorios Europeos, causando no solamente la perdida de vidas humanas, sino devastación incuantificable del

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•301
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
medio ambiente. Países como España, Portugal, Italia, Francia y Grecia, son los miembros de la Comunidad Económica Europea más afectados debido a su posición geográfica y condiciones climatológicas. Un total de 589.669 hectáreas, entre estos cinco países, se quemaron hasta convertirse en cenizas en 2005. Ese año, el numero de incendios sobrepaso los promedios de años anteriores, con cifras abrumadoras de 73.325 casos durante los meses de verano. Portugal y España fueron los más castigados por la devastación contribuyendo con el 88% de los incendios forestales con 85% de las 589.559 hectáreas de superficie afectadas (Babel International, 2007). En los Estados Unidos, las consecuencias de los fuegos forestales han sido desastrosas debido a la debilidad o a la no existencia en algunos casos de leyes de regulaciones de zonificación orientadas a controlar la intrusión de las zonas urbanas en las áreas rurales y silvestres. En consecuencia, como una forma de uso multipropósito de la tierra, los sistemas silvopastoriles pueden reducir el riesgo de fuegos silvestres utilizando el pastoreo para reducir y controlar la vegetación de cobertura del sotobosque en áreas boscosas (Rigueiro-Rodríguez et al., 2005).
En Argentina, Rachene y Bava (2003) reportaron que la conservación de la selva araucaria-nire (nothofagus antarctica) después de los incendios, así como la recuperación de las áreas degradadas es posible a través de la implementación de una prevención activa de fuegos y de estrategias de control. Programas de manejo de combustible para disminuir incendios forestales utilizando ganado en pastoreo fueron revisados por Davison (1996).
El parque regional East Bay del distrito de San Francisco, California, tiene más de 40 años de experiencia utilizando el pastoreo como una herramienta de control de material potencialmente combustible (East Regional Park District. 2007. Regional Parks Online). El programa es conducido bajo una licencia altamente regulada, basada en principios reconocidos de manejo de praderas. Aproximadamente 5.000 vacunos, 1.000 ovinos y 1.000 caprinos están diseminados por más de la mitad de los 65 parques del distrito, para reducir combustibles y matorrales por más de 20.000 hectáreas. La mayoría del pastoreo se realiza durante la primavera e inicios del verano.
En España, Garcia et al., (2005) evaluaron el comportamiento productivo en términos de variaciones de peso y condición corporal de ovejas de raza Gallega y de cabras de raza Autóctona y Cachemir, mantenidas a pastoreo en matorrales dominados por tojo, desarrollados luego de la quema superficial de un brezal (erica spp.) – tojal (ulex galii). Al inicio se establecieron cuatro parcelas que fueron pastoreadas por cada una de las especies independientemente,
a una carga de 10 cabezas/ha. En el tercer año, las parcelas fueron divididas a la mitad, cada una de las cuales fue reasignada a una u a otra de las especies animales. En esta etapa la carga varió entre 6.7 y 10 cabezas/ha. A pesar de que los resultados del primer año de estudio mostraron un mejor comportamiento de los caprinos, la situación se invirtió los años siguientes, por lo menos en las parcelas pastoreadas previamente por ovinos, mientras que no hubieron diferencias significativas entre ovinos y caprinos en las parcelas pastoreadas previamente por caprinos, con interacciones significativas entre el pastoreo previo y la especie animal en el segundo período. Estos autores, señalan que la reducción en la altura de la vegetación herbácea afecta de manera más significativa a los caprinos.
En muchas regiones del mundo existen grandes extensiones de tierra maderable que contienen cantidades significativas de vegetación para ramoneo y de cobertura, que podría sustentar grandes números de caprinos y otras especies de ganado mientras se reduce el riesgo de incendios forestales. Sin embargo, se ha realizado poca investigación para catalogar y examinar la calidad de estos recursos, para determinar su productividad potencial en términos de producto animal, y para cuantificar los efectos del ramoneo por los caprinos en el ciclo de nutrientes, en la productividad de madera, y en la diversidad de plantas y vida silvestre.
COnCLUSIOnESLos caprinos tienen la habilidad distintiva de consumir recursos forrajeros que no pueden ser utilizados efectivamente por otras especies de ganado. Debido a su versatilidad, los caprinos se orientan instintivamente hacia un pastoreo de conservación, mejor que cualquier otra especie de animales domésticos, cuando son utilizados de manera sustentable. Sin embargo, el sobrepastoreo y la degradación ocurren, e inclusive la desertificación, debido a que el hombre permite que la intensidad de carga sobrepase los límites de la capacidad de carga de la vegetación existente. En estas situaciones, los caprinos pueden convertirse en la más destructiva de las especies de ganado. En muchos países aun se culpa a los caprinos por el mal manejo de sus praderas, cuando de hecho es el hombre, el que ha impuesto las condiciones para la destrucción de estos valiosos recursos. Es necesario, llevar a cabo investigaciones con las que se establezca el número de caprinos que permitiría alcanzar el balance entre el ganado en pastoreo y la comunidad vegetal. Las especies leñosas proveerían una fuente continua de material palatable y nutritivo para ramoneo, para los caprinos, los cuales podrían ser controlados para minimizar la perdida de especies forrajeras más favorables, preferidas por otras especies de ganado.

302 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
El manejo del rebaño se convierte entonces en una herramienta de mucha importancia en la sustentabilidad del ecosistema, ya que puede provocar cambios en la estructura del componente vegetal, en su composición botánica y en la diversidad de especies presentes.
REFEREnCIAS
Allegretti, L., C. Passera, J. Paez, A. Ubeda, C. Sartor, y A. B. Robles. 2005. Capacidad sustentadora y composición botánica de la ingesta caprina en un ecosistema árido, La Valle, Argentina. Producciones Agroganaderas: Gestión eficiente y conservación del medio natural. Vol. 1: 221-228. XLV Reunión Científica de la S.E.E.P 2005 Gijón. España.
Alvarez, J. 1995. Dinámica sucesional tras el abandono y recuperación del matorral mediante pastoreo controlado. Experiencia en un sector de la montaña de León. Tesis de Grado. Universidad de Lleida. España. 136 pp. I S B N: 84-89727-64-3.
Babel International. 2007. Over 50,000 forest fires in Europe per year. The European Magazine. URL: http://www.cafebabel.com/en/article.asp?t=t&id=10908Bryant, F. C., M. M. Kothmann, y L. B. Merrill. 1979. Diets of sheep, Angora goats, Spanish goats and White-tail deer under excellent range conditions. J. Range Manage., 32:412-417.
Bonsi, C., E. Rhoden, A. Woldeghebriel, P. Mount, S. Solaiman, R. Noble, y G. Paris. 1991. Kudzu-goat interactions - A pilot study. In: S. G. Solaiman and W. A. Hill (Ed.) Using Goats to Manage Forest Vegetation: A Regional Inquiry. p 84. Tuskegee, AL.
Celaya, R., B. M. Jauregui, U. Garcia, y K. Osoro. 2005. Efectos de la Raza y la Carga ganadera sobre la cubierta Vegetal en Brezales-Tojales pastados por caprino. Producciones agroganaderas: Gestión eficiente y conservación del medio natural. Vol. I: 319-326. XLV Reunión científica de la S.E.E.P 2005 Gijón. España.
Child, R. D., E. K. Byington, y H. Hansan. 1985. Goats in the mixed hardwoods of the southeastern United States. In: F. H. Baker and R. K. Baker (Ed.) Multispecies Grazing. p 149-158. Winrock International, Morrilton, AR.
Coblentz, B. E. 1977. Some range relationships of feral goats on Santa Catalina island, California. J. Range Manage. 30:415-419.
Davison, J. Livestock grazing in wildland fuel management programs. 1996. Rangelands 18:242-245.
du Toit, P. F. 1972. The goat in a bush-grass community. Proc. Grassland Soc. South Africa 7:44-50.
East Regional Park District. 2007. Regional Parks Online. Grazing Animals in the East Bay Regional Park District . URL: http://www.ebparks.org/resources/stewards/grazing.htm#firehaz
Fedele, V., M. Pizzillo, S. Claps, P. Morand-Fehr, y R. Rubino.1993. Grazing behavior and diet selection of goats on native pasture in Southern Italy. Small Rumin. Res. 11:305-322.
Fierro, L. C., F. Gomez, y M. H. Gonzales. 1982. Biological control of undesirable brush species using goats in central Chihuahua, Mexico. In: III International Conf. on Goat Production and Disease. pp 534. Dairy Goat J. Publ. Co. Scottsdale, AZ.
Fraps, G. S., y V. L. Cory, 1940. Composition and utilization of range vegetation of Sutton and Edwards counties. Texas Agric. Exp. Sta. Bull. 586. Texas A&M University. College Station.
Frost, R. A., y K. L. Launchbaugh. 2003. Prescription grazing for rangeland weed management. Rangelands 25:43-47.
Garcia, O., y C. Gall. 1981. Goats in the dry tropics. In: C. Gall (Ed.) Goat Production. pp 515-556. Academic Press. London.
García, R., y M. Salazar. 1986. Loma de León, un Estudio de la Vegetación en una Zona Semiárida de Venezuela y su Relación con el Caprino y la Conservación. Estado Lara en el IV Congreso Venezolano de Conservación. Ponencia. Fundación BIOMA. Caracas. 47 pp.
García, R., M. Salazar, y R. Smith. 1990. Monumento Natural Loma El León. FONAIAP Divulga No. 34.
Garcia, U., R. Celaya, B. M. Jauregui, y K. Osoro. 2005. Comportamiento productivo de ovinos y caprinos pastando sobre brezales-tojales previamente quemados. Producciones Agroganaderas: Gestión eficiente y conservación del medio natural. Vol I: 213-219. XLV Reunion cientifica de la S.E.E.P 2005 Gijon. España.
Hansen, H. H., y R. D. Child. 1980. Goat diets on an upland hardwood ecosystem. J. Anim. Sci. 51 (Suppl. 1): 238.
Harrington, G. N. 1982. Grazing behavior of the goat. In: III International Conf. on Goat Production and Disease. pp 398-402. Dairy Goat J. Publ. Co. Scottsdale, AZ.

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•303
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Hart, S. P. 2001. Recent perspectives in using goats for vegetation management in the USA. J. Dairy Science 84 (Suppl.):E170-E176.
Harvey, T. E. 1996. Mountain pasture renovation and multiflora rose control utilizing goats alone or goats in combination with cattle. Masters of Agriculture Thesis. North Carolina State University, Raleigh, NC.
Hofman, R. R. 1989. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system. Oecologia 78:443-457.
Kay, S.H., W. M. Lewis, y K. A. Langeland. 1995. Integrated management of multiflora rose in North Carolina. 17 p. North Carolina Cooperative Extension Service Publication No. AG-536.
Lambert, M.G., G.A. Jung, H.W. Harpster, y J. Lee. 1989. Forage shrubs in North Island hill country. 4. Chemical composition and conclusions New Zealand Journal Agricultural Research 32: 499-506.
Langston, J. C. 1992. Utilization and elimination of forest understory by goats. Masters of Agriculture Thesis. North Carolina State University, Raleigh, NC. Lu, C. D. 1988. Grazing behavior and diet selection of goats. Small Ruminant Research 1:205-216.
Luginbuhl J-M., J. T. Green, Jr, J. P. Mueller, y M. H. Poore. 1995. Grazing habits and forage needs for meat goats and sheep - Chapter 20. In: D. S. Chamblee (Ed.) Production and Utilization of Pastures and Forages in North Carolina. pp 105-111. North Carolina Agricultural Research Service Technical Bulletin No. 305.
Luginbuhl, J-M., J. T. Green, Jr., J. P. Mueller, y M. H. Poore. 1996. Use of goats to control unwanted vegetation. Presented at International Workshop “Los Arboles en los Sistemas de Producción Ganadera”. November 26-29. Estación Experimental de Pastos y Forrajes Indio Hatuey Matanzas, Cuba.
Luginbuhl, J-M., J. T. Green, Jr., M. H. Poore, y A. P Conrad. 2000. Use of goats to manage vegetation in cattle pastures in the Appalachian region of North Carolina. Sheep & Goat Res. J. 16:124-135.
Luginbuhl, J-M., T. E. Harvey, J. T. Green, Jr., M. H. Poore, y J. P. Mueller. 1999. Use of goats as biological agents for the renovation of pastures in the Appalachian region of the United States. Agroforestry Systems 44:241-252.
Mackenzie, D. 1993. Goat Husbandry. Fifth edition. Faber & Faber. London, UK.
Magadlela, A. M., M. E. Dabaan, W. B. Bryan, E. C. Prigge, J. G. Skousen, G. E. D’Souza, B. L. Arbogast and G. Flores. 1995. Brush clearing on hill land pasture with sheep and goats. J. Agron. & Crop Sci. 174:1.
Malecheck, J. C., y C. L. Leinweber. 1972. Forage selectivity by goats on lightly and heavily grazed ranges. J. Range Management 25:105-111.
Malecheck, J. C., y F. D. Provenza. 1981. Feeding behavior of goats on rangelands. In: P. Morand-Fehr, A. Bourbouze and M. de Simiane (Ed.) Nutrition et Systèmes d’Alimentation de la Chèvre. Vol 1. pp 411-428. ITOVIC - INRA. Tours, France.
Mays, W.J., y L. P. Kok. 1988. Seed wasp on multiflora rose, Rosa multiflora in Virginia. Weed Technol. J. 265-268.
Mellado, M., R. Valdez, L. M. Lara, y R. Lopez. 2003. Stocking rate effects on goats: A research observation. J. Range Management 56:167-173.
Merrill, L. B., y C. A. Taylor. 1981. Diet selection, grazing habits, and the place of goats in range management. In: C. Gall (Ed.) Goat Production. pp 233-252. New York Academic Press, New York.
Miller, J. H. 2004. Nonnative invasive plants of southern forests. A field guide for identification and control. pp 40-41. US Department of Agriculture. Forest Service, Southern Research Station. General Technical Report SRS-62.
Osoro, K., A. Martinez, M. Olivan, U. Garcia, y R. Celaya. 2005. Manejo de los herbívoros domésticos para la biodiversificación y el desarrollo rural sostenible. Producciones Agroganaderas: Gestión eficiente y conservación del medio natural. Vol. I: 45-72. XLV Reunión Científica de la S.E.E.P 2005 Gijón. España.
Owens, M. K. 1991. Utilization patterns by Angora goats within the plant canopies of two Acacia shrubs. .J. Range Management 44:456-461.
Perez, L. 1998. Comportamiento alimentario y actividades de cabras en pastoreo sobre campo natural. URL: htpp://www.capraispana.com/destacados/pastoreou/resumen.htm
Pietrosemoli, S., F. Arenas, D. Bermudez, O. Peley, y A. Casanova. 2005. Goat preference of five tropical legumes. J. Anim. Sci. 83 (Suppl. 1):277.
Rechene, C., y J. Bava. 2003. Los bosques de Araucaria araucana en Chile y Argentina. Parte II. Estudios silvícola y propuestas para su conservación y uso en

304 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Argentina. Program de Ahoy Ecologic (TOEB) (TWF-V/40 s) Ashburn: Deutsche Gesellschaft fur Technische Zusammenarbeit (GTZ) GmbH, p.81-158.
Rector, B. S. 1983. Diet selection and voluntary forage intake by cattle, shee and goats grazing in different combinations. Ph. D. dissertation, Texas A&M Univ., College Station.
Rigueiro-Rodríguez, A. M., R. Mosquera Losada, R. Romero Franco, P. M. González Hernández, y J. J. Villarino Urgiaga. 2005. In: M. R. Mosquera-Losada, J. McAdam, and A. Riguerio-Rodriguez (Ed.) Silvopastoralism and Sustainable Land Management. pp 380-387. CABI Publishing Wallingford, UK.
Silanikove, N. 1997. Why goats raised on harsh environment perform better than other domesticated animals. Options Mediterraneennes 34 (Serie A):185-194.
Smith, R., y A. Rivero. 1983. Los recursos ecológicos de la zona árida de los alrededores de Barquisimeto, Estado Lara – Venezuela. Universidad Centrooccidental Lisandro Alvarado, Instituto Universitario Pedagógico Experimental de Barquisimeto. Barquisimeto. 42 p + cuadros + mapas.
Taylor, C. A., 1985. Multispecies grazing research overview (Texas). In: Proc. Multispecies Grazing. F. H. Baker, R. K. Jones (Eds.) pp 65-83. Winrock International Institute for Agricultural Development, Morrilton, AR.
Taylor, C. A., y M. H. Ralphs. 1992. Reducing livestock losses from poisonous plants through grazing management. J. Range Management. 45:9-12. Van Soest, P. J. 1994. Nutritional Ecology of the Ruminant. 2nd Ed. Cornell University Press, Syracuse, NY.
Walker, J. W., S. L. Kronberg, S. L. Al-Rowaily, y N. E. West. 1994. Comparison of sheep and goat preferences for leafy spurge. J. Range Manage. 47:429-434.
Webb, D. M., A. O. Abaye, C. Teusch, y G. Scaglia. 2007. Assessing the potential of co-grazing small ruminants with beef cattle to improve utilization of margina pasturelands in Virginia. Proc. American Forage and Grassland Council. 23-26 June 2007. State College. Pennsylvania.
Wilson, P. N. 1957. Studies of the browsing and reproductive behavior of the East African Dwarf goat. E. African Agric. J. 23: 138-147.
Wilson, A. D., J. H. Leigh, N. L. Hindley y W. E. Mulham. 1975. Comparison of the diets of goats and sheep on a Casuarina cristata - Heterodendrum oleifolium
woodland community in western New South Wales. Australian J. Experimental Agriculture Animal Husbandry 15:45-53.
Woldeghebriel, A., R. N. Corley, III, y M. R. Murphy. 1992. Rotational grazing model of goats on kudzu-infested forestland. Proc. Internat. Symp. Land Reclamation: Advances in Research and Technology. 50 pp. Nashville.
Wood, G. M. 1987. Animals for biological brush control. Agronomy J. 79:319-321.
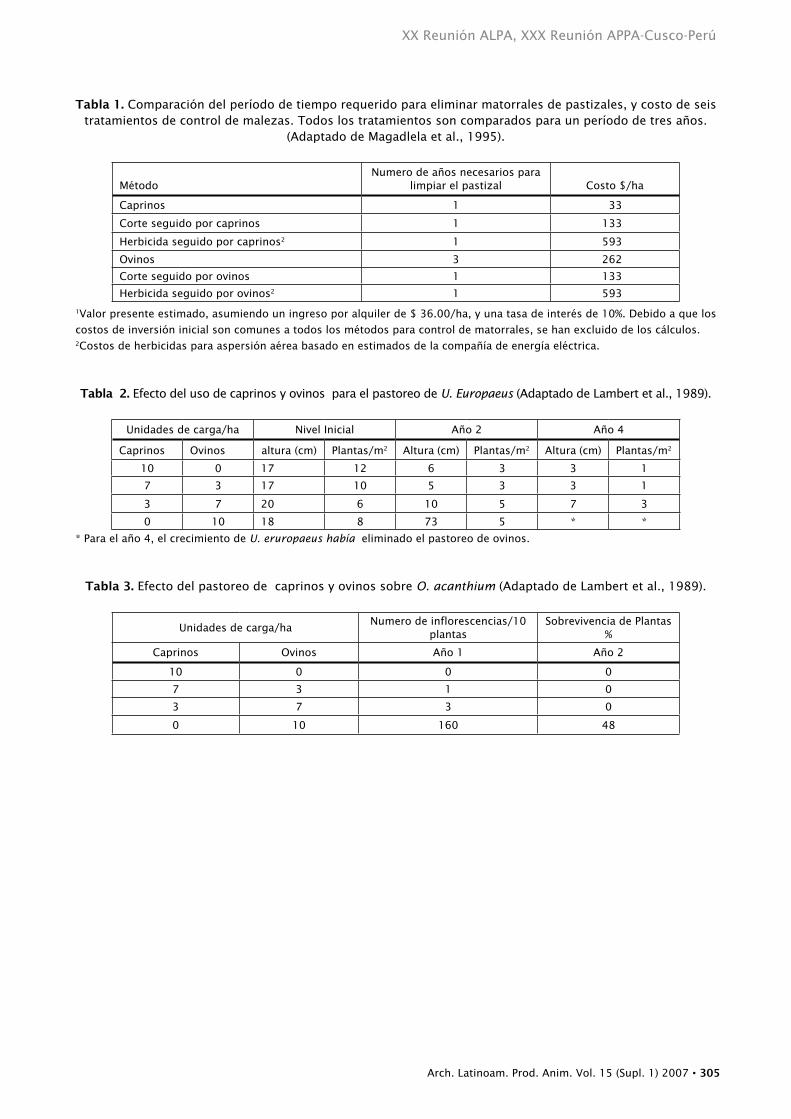
Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•305
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
tabla 1. Comparación del período de tiempo requerido para eliminar matorrales de pastizales, y costo de seis tratamientos de control de malezas. Todos los tratamientos son comparados para un período de tres años.
(Adaptado de Magadlela et al., 1995).
MétodoNumero de años necesarios para
limpiar el pastizal Costo $/ha
Caprinos 1 33
Corte seguido por caprinos 1 133
Herbicida seguido por caprinos2 1 593
Ovinos 3 262
Corte seguido por ovinos 1 133
Herbicida seguido por ovinos2 1 593
1Valor presente estimado, asumiendo un ingreso por alquiler de $ 36.00/ha, y una tasa de interés de 10%. Debido a que los
costos de inversión inicial son comunes a todos los métodos para control de matorrales, se han excluido de los cálculos.2Costos de herbicidas para aspersión aérea basado en estimados de la compañía de energía eléctrica.
tabla 2. Efecto del uso de caprinos y ovinos para el pastoreo de u. europaeus (Adaptado de Lambert et al., 1989).
Unidades de carga/ha Nivel Inicial Año 2 Año 4
Caprinos Ovinos altura (cm) Plantas/m2 Altura (cm) Plantas/m2 Altura (cm) Plantas/m2
10 0 17 12 6 3 3 1
7 3 17 10 5 3 3 1
3 7 20 6 10 5 7 3
0 10 18 8 73 5 * *
* Para el año 4, el crecimiento de u. eruropaeus había eliminado el pastoreo de ovinos.
tabla 3. Efecto del pastoreo de caprinos y ovinos sobre o. acanthium (Adaptado de Lambert et al., 1989).
Unidades de carga/haNumero de inflorescencias/10
plantasSobrevivencia de Plantas
%
Caprinos Ovinos Año 1 Año 2
10 0 0 0
7 3 1 0
3 7 3 0
0 10 160 48

306 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
tabla 4. Comportamiento reproductivo, química sanguínea, puntaje de condición corporal y promedio de ganancia diaria de caprinos pastoreando praderas con alta y baja carga animal (Adaptado de Mellado
et al., 2003).
Carga animal
Variable 1.5 ha/caprino 15 ha/caprino
Características reproductivas Aborto, % 22a 12 Tasa de pariciones, % 42b 55
Preñez, % 60 67
Mortalidad debida a plantas toxicas % 4.2 0Química sanguínea
Glucosa, mg/100 mL 48.9a 64.0
Urea nitrógeno, mg/100 mL 10.8a 12.1
Creatinina, mg/100 mL 0.73a 0.60
Colesterol, mg/100 mL 66c 72
Proteína Total, g/100 mL 6.5 6.8
Calcio, mg/100 mL 11.2 11.0
fosforo, mg/100 mL 4.0c 4.4
Cobre, mg/1,000 g 1.79 1.83
Magnesio, mg/1,000 g 1.93a 2.08
Zinc, mg/1,000 g 1.77a 2.33
Características corporales
Puntaje de condición corporal (Escala 0-5)
2.1a 2.7
Promedio Ganancia diaria -19a 11a = P < 0.01; b = P < 0.05; c = P < 0.10.
Figura 1. Efecto del pastoreo en pastizales montañosos de la región de las Appalachian de los Estados Unidos con caprinos solos, o caprinos con vacunos, sobre el porcentaje de cobertura vegetal durante cuatro estaciones de pastoreo. Contrastes ortogonales: control vs. caprinos y caprinos + vacunos (P < 0.01); caprinos vs. caprinos + vacunos (P < 24); SE = 3.4 (Luginbuhl et al., 1999).

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•307
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Figura 2. Efecto del pastoreo en pastizales montañosos de la región de las Appalachian de los Estados Unidos con caprinos solos, o caprinos con vacunos, sobre el porcentaje de cobertura vegetal como gramíneas durante cuatro estaciones de pastoreo. Contrastes ortogonales: control vs. caprinos y caprinos + vacunos (P < 0.01); caprinos vs. caprinos + vacunos (P < 0.19); SE = 3.4 (Luginbuhl et al., 1999).
Figura 3. Efecto de pastoreo en pastizales montañosos con caprinos solos o con caprinos y vacunos en la altura de Rosa multiflora (m) Contrastes ortogonales: control vs. caprinos y caprinos + vacunos (P < 0.02); caprinos vs. caprinos + vacunos (P < 0.17); SE = 0.2 (Luginbuhl et al., 1999).

308 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Figura 4. Efecto de pastoreo en pastizales montañosos con vacunos solos o con caprinos y vacunos en la altura de Rosa multiflora (m) Contrastes ortogonales: control vs. vacunos y caprinos + vacunos (P < .01); vacunos vs. caprinos + vacunos (P < .01); SE = .28 (Luginbuhl et al., 2000).
Figura 5. Efecto de pastoreo en pastizales montañosos con vacunos solos o con caprinos y vacunos en la altura de Robinia pseudoacacia (m) Contrastes ortogonales: control vs. vacunos y caprinos + vacunos (P < .01); vacunos vs. caprinos + vacunos (P < .87); SE = .41 (Luginbuhl et al., 2000).

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•309
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
Figura 6. Caprinos ramoneando eleaganus umbellata Thunb. en Virginia (Webb et al., 2007).
Figura 7. Composición de la dieta de caprinos de carne, determinada por análisis micro histológicos (Adaptado de Child et al., 1985).

310 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
IntRODUCCIónEl ovino es un animal cuya distribución es amplia por todo el mundo, se le encuentra en todos los climas y ecologías. Gracias a esta especie se ha podido aprovechar extensas áreas de pasturas pobres parar otras especies, especialmente vacunos. Se cree que las ovejas domesticas Ovis aries, (2) son descendientes de estirpes salvajes que aún existen y que son ínter fértiles con las ovejas domesticas.
Las ovejas de pelo han ido evolucionando bajo la influencia selectiva de la naturaleza y del hombre. Su pelaje es similar a los de los bovinos y caprinos. Se adaptan bien a los ambientes tropicales, lo cual es importante para muchos países en vías de desarrollo, donde estas son las principales, sino las únicas, las ovejas. Se estima su número entre 7 al 10% e la población ovina mundial que llega a 1.1 mil millones. Los productos principales de estas ovejas son: animales para la venta, la carne para el consumo familiar y los cueros. Su introducción en el Perú ha concitado interés dadas sus características de adaptación.
SIStEMAS DE PRODUCCIónLas ovejas de pelo contribuyen a varios sistemas de producción en los trópicos. La precipitación pluvial es frecuentemente el factor principal que determina la característica del sistema, aunque los factores socioeconómicos también ejercen influencia. De manera general se dan dos sistemas:
1. Sistemas mixtos de cultivos/ganadería, en áreas agrícolas de potencial mediano alto, y2. Sistemas de pastoreo basados en la ganadería en las zonas de praderas más áridas.
REStRICCIOnES DE LOS SIStEMAS DE PRODUCCIónAunque las características de los sistemas de producción de las ovejas de pelo varían de región en región, la mayoría de los sistemas están sujetos a las
importanCia de los ovinos tropiCales intoduCidos al pais: CaraCterÍstiCas produCtivas Y reproduCtivas
José A. Atto Mendives 1
1 Ing. zootecnista. Docente del Dpto. Prod. Animal de la Fac. de zootecnia - U. nac. de Piura.Miembro del Directorio de la OnG. Procabra. Email: [email protected]
mismas restricciones generales. Las tres categorías de restricciones son en común son la ecológica, la biológica y la socioeconómica.
1. Restricciones EcológicasDentro de este grupo hay dos factores: la tierra con sus características: superficie, topografía y fertilidad del suelo; y, el clima (precipitación, temperatura, estación, etc.) Restringen la producción ovina porque determinan el tipo y rendimiento de los pastos naturales.
Solamente la fertilidad del suelo es susceptible de cambiar, pero solo cuando los rendimientos obtenidos son económicamente factibles.
2. Restricciones BiológicasNutrición.- Restricciones en cuanto a la cantidad como la calidad de nutrientes ingeridos por las ovejas de pelo. El pastoreo diurno y el confinamiento nocturno restringen el consumo de los nutrientes. Además, rara vez, se les proporciona suplementos, su consumo diario de nutrientes posiblemente no sea suficiente para fomentar su potencial productivo real. El calor al medio día y la radiación solar limitan aún más la ingesta de nutrientes de las ovejas en pastoreo. La producción de forraje varía mucho con las estaciones. No se da una adecuada y sostenida conservación de forrajes.
Sanidad.- Se presentan muchas enfermedades principalmente en los trópicos húmedos y limitan la producción: entre las que se mencionan, neumonía, fiebre aftosa, parásitos, pedera. etc. El parasitismo elevado debilita a los animales y permite la transmisión de enfermedades.
La pérdida de animales.- Se presenta robo o predación por parte del hombre o por animales predatores. Las pérdidas directas de animales son menores que las perdidas de productividad que resultan de las practicas de manejo que se precisan emprender para evitar la prelación.
Genotipo.- En ambientes tropicales, el potencial genético para la adaptación tiene prioridad sobre el potencial genético para productividad. Inclusive pueden haber correlaciones genéticas negativas entre las características de adaptación y de producción.

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•311
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
3. Restricciones SocioeconómicasSe consideran factores humanos e institucionales: disponibilidad de mano de obra, y el uso de practicas de manejo; el gusto las preferencias y el poder adquisitivo de los consumidores; el costo y la disponibilidad del crédito; la infraestructura de mercadeo; y las políticas gubernamentales sobre precios, comercio y tenencia de tierras.
CARACtERíStICAS IMPORtAntES PARA LA PRODUCCIón DE OVInOS En LOS tRóPICOS
La productividad y la eficiencia resultan de una combinación de características de producción, sujetas a la influencia de la genética y del medio ambiente. Las tres categorías generales de las características de producción son:
1. Adaptación física: considera las siguientes características:a. Adaptación al estrés ambiental: pelaje, resistencia a enfermedades y parásitos, sobre vivencia de corderos , longevidad, temperamento.b. Adaptación a las fluctuaciones del ambiente.
2. Fertilidad: Incluye características reproductivas:a. Prolificidad: tasa de ovulación, tasa de fertilidad, sobrevivencia de embriones.b. Intervalo entre pariciones: días vacíos, anestro post-parto, periodo de gestación.c. Destete: comportamiento materno, producción de leche, vigor de los corderos.d. Edad de la madurez sexual.e. Características de los machos: libido, calidad del semen.
3. tamaño y Eficiencia: Características del desarrollo corporal:a. Tasas de crecimiento y madurez.b. Pesos corporales: Peso al nacimiento, sobrevivencia de corderos. Peso al sacrificio-rendimiento de carne. Peso adulto, requerimiento de mantenimiento.c. Composición corporal – tejidos comestibles.d. Consumo voluntario de alimentos.e. Composición de la dieta-forrajes, residuos, concentrados.f. Eficiencia de utilización de los nutrientes.
tIPOS y RAzASLas Ovejas de Pelo fueron traídas al Brasil y a las Indias Occidentales durante el siglo XVII, probablemente del África Occidental. En general presentan cola delgada, con la excepción de los Persa de Cabeza Negra. Es probable que las Ovejas de Pelo de cola delgada emigraron del noreste del África, atravesando el centro norte y llegando, finalmente, al África Occidental. En el cuadro 1, se presentan los diferentes tipos de ovejas tropicales.
Ovejas de AméricaLas Ovejas de Pelo americanas son más parecidas al tipo Selva Sabana que al tipo Sahel, aunque, por lo general, son más grandes que las del tipo Selva. Las Ovejas Persa de Cabeza Negra, de grupa ancha, no se parecen a ninguno de estos dos tipos y son claramente de origen diferente. Las poblaciones de Ovejas de Pelo se encuentran en muchas de las Islas del Caribe, en los países de Centro y Sud América, que circundan la cuenca del Caribe, y en el nordeste del Brasil. De las Islas Vírgenes y los Barbados se ha exportado a los Estados Unidos y otros países.
Cuadro 1: Tipos de ovinos tropicales
Características tipo Sahel Selva - Sabana
-Peso de hembras adultas Más de 35 Kg. Selva: 20-30 kg. Sabana: 30-40kg.
-Altura a cruz de hembras adultas Más de 60 cm. Selva: 40-55 cm. Sabana: 55-65 cm.
-Orejas Largas, pendulosas Cortas, horizontal
Cuernos: Machos Frecuentemente largos Cortos forma de media y en espiral luna, algunos mochos
Hembras Generalmente mochas Mochas, a veces cuernos rudimentarios
Melena/papada Ausente Generalmente presente
Fuente: FITZHUGH, H.A. y C.E. BRADFORD, 1983.

312 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
EL OVInO CRIOLLOLos ovinos criollos actuales conservan las características de sus ancestros (Ovinos Merino y Churra), pero con una calidad muy deteriorada. Estos animales al haberse entrecruzado incontroladamente, han dado lugar a una población con una mezcla de características de ambas, pero sin que, en la fibra, se observen finuras intermedias.
En algunos ejemplares, las fibras son más gruesas que las del Merino cuando éstas tienden al Churra; en otros, las fibras son de extrema finura comparados con el Merino original, aunque con zonas peludas, especialmente en los muslos. Este afinamiento de la fibra, no es sino el resultado de la carencia alimenticia, es por eso que se le llama “finura de hambre”
DESCRIPCIóN DE LAS RAZAS INTRODUCIDAS AL PAIS
BLACKBELLy.- Originaria de la Isla de Barbados, Antillas. No posee cuernos. Color castaño rojizo, excepto en el vientre y las patas que son negras. De tamaño mediano, los machos pesan en promedio 60 kg. Y las hembras 45 kg. La fertilidad es alta con 2 a 2.1 corderos por parto; y partos múltiples son frecuentes.
PELIBUEy.- Es un ovino originario de Cuba, y de aquí se ha distribuido a una cantidad de países en la región del Caribe, México, República Dominicana, Colombia y Venezuela. Esta agrupación incluye ovejas de tamaño, color y desempeño similares. Los colores más frecuentes “beige”, blanco y “beige”, y rojo
cereza. Los machos alcanzan pesos adultos de 50 kg. y las hembras de 38 kg. Es una raza también prolífica y rustica.
KAtAhDIn.- Es una nueva raza de los EEUU. Presenta color rojo o blanco con algunas tonalidades entre éstos. Producto del cruce de Ovejas Virgins Islands con carneros de la raza Suffolk y Wiltshire Horn. Los pesos adultos son: 70 kg. para machos y 55 kg. para hembras. Presenta las características de eliminar la cubierta pilífera (especie de muda), quedando una piel fina. Introducida al país en diciembre de 1987 en Piura, como donación de Heifer Project Internacional.SANTA INES.- Raza brasileña. Color rojo o blanco. Es un animal de patas largas, nariz romana y orejas grandes y caídas. Carecen de melena y papada peluda. Se cree que estas características procedan de la cruza con la raza italiana Bergamasca, de lana gruesa. Se clasifica como Ovejas tipo Sahel.
MORADA nOVA.- Procede del Brasil, de color rojo y blanco, se estableció en base al “Pelo do boi”, oveja común en el noroeste del Brasil, de apariencia similar al Pelibuey, La Morada Nova es semejante en tipo de orejas y cola, tamaño y conformación a la mayoría de las Ovejas de Pelo del Caribe. Se han introducido algunos ejemplares a Iquitos.
ASSAF.- Raza lechera de Israel, producto del cruce de la oveja Awassi autóctona de Israel con la oveja East Friesian originaria de Alemania, siendo 5/8 Awassi y 3/8 East Friesian. El cruzamiento se realizó con la finalidad de fusionar en una raza nueva, la característica lechera de Awassi, con la alta prolificidad
Cuadro 2: principales razas que se crían en el país
Ovinos productores de lana: Merino Australiano Ramboullt Americano Merino Argentino
Ovinos productores de carne: Hampshire Down Suffolk Polled Dorsets Merino precoz alemán Black Be lly Pelibuey Asblack en evaluación Criollos
Ovinos de doble propósito: Corriedale Romney Marsh Junín
Ovinos productores de leche: Assaf en fomento

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•313
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
y fecundidad del East Friesian; y de absorber también, del Awassi las características de resistencia a severas condiciones ambientales y a enfermedades propias del medio subtropical. Llegó al Perú en 1988 procedente de Israel, su país de origen.
Es un ovino predominantemente de color blanco e hipermétrico. Con ubres simétricas y de fácil ordeño. Aparte de ser una buena oveja lechera, es reconocida como excelente productora de carne. Las canales de los corderos Assaf son alargados y su grasa bien distribuida e infiltrada, lo que hace que su carne sea aparentemente menos grasosa y de gusto agradable en comparación con las de otras razas. Es de exagerada la acumulación de grasa en la cola (por lo que se les llama “cola gorda”). En el cuadro 3 se dan algunos índices productivos y reproductivos logrados en Israel.
Cruce de Assaf con el Black Belly: ASBLACKDesde 1994 se ha trabajado en el país con la finalidad de conseguir un ovino con características productivas de leche y carne y que se adapte a condiciones tropicales. La tarea emprendida por el recordado e insigne ovinocultor nacional Ing. Rigoberto Calle Escobar, ha dado sus frutos pues en la actualidad se cuenta con el Asblack, un ovino en que se han fusionado dos razas de alta productividad:
El Assaf con alto peso corporal y mayor productor de leche, que significa más kg. de carne y de leche.
El Black Belly, de alta taza reproductiva.
CARACtERIStICAS REPRODUCtIVAS E InDICES DE PRODUCCIónSe ha hecho una evaluación comparativa de las cinco razas tropicales y se ha incluido el Assaf por ser una raza que se adapta a dicho ambiente; se utilizó
los Índices de Productividad de Manada (IPM) y de Eficiencia de Manada (IEM) sugeridos por Fitzhugh y Bradford, ya que involucran muchas características que determinan la productividad y la eficiencia y son de mayor relevancia que comparaciones entre características individuales. Sin embargo, recomiendan trabajar con índices simplificados y sustituir valores que faltan con representantes promedios; aunque señalan que tienen sus limitaciones:
IPM = (tamaño camada x sobrevivencia x PN) / interv. entre parto
IEM = IPM / W.75
El hecho de usar estimadores para los promedios de raza que faltan, introduce errores de magnitud desconocida y reduce la variación entre las razas. No obstante, estos índices si reducen los efectos de muchas características sobre la productividad y la eficiencia de un rebaño.
Analizando las razas del cuadro 4, para el IPM y el IEM resulta la siguiente clasificación: Blackbelly, Assaf, Morada Nova, Pelibuey, Santa Inés, y Katahdin. Hay que señalar que los datos más apreciables son los que corresponden al Blackbelly, Assaf, Pelibuey y Katahdin, ya que son datos nacionales; en cambio la información correspondiente a Morada Nova y Santa Inés, se considera menos relevante por dos motivos: no hay información suficiente de los registros nacionales y el número de cabezas es mínimo en el país, además no han sido traídos a la costa; sin embargo ayudan a una discusión más amplia.
El Assaf y Blackbelly tienen una apreciable ventaja para el IPM que se reduce para el IEM, especialmente en comparación con las ovejas Pelibuey. Esta diferencia para el IPM a favor del Assaf probablemente se debe a que presenta bajo intervalo entre parto (cada 8
Cuadro 3: Rendimiento y producción del Assaf en Israel
LecheRitmo de Crecimiento
Profilicidad Precocidad
Vellón
Producción Composición Peso Característica
300-400
Lit/Leche/año
5.7% grasa
5.11% prot.
17.8% sólidos
50 kg PV. a la comercialización. Incremento 350 g/día
adulto 70kg PV.
1.6 crías/parto
1.4 partos/año
2.3 crías/ovejas/año
Primer servicio a los 8 meses con 50kg PV.
Esquila anual:
Machos 4-5kg
Hembra 3-3.5kg
Semi abierto y desuniforme
Fuente: Tomado y adaptado de Israel Sheep breeders Association. 1994
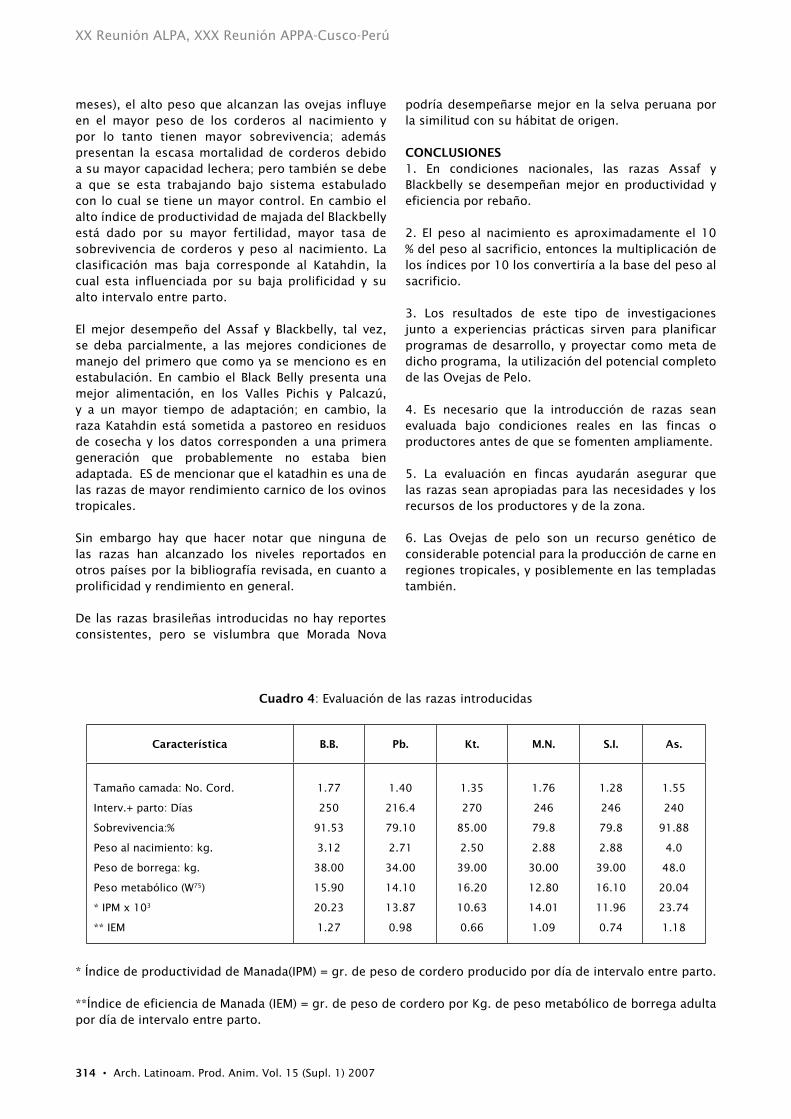
314 • Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
meses), el alto peso que alcanzan las ovejas influye en el mayor peso de los corderos al nacimiento y por lo tanto tienen mayor sobrevivencia; además presentan la escasa mortalidad de corderos debido a su mayor capacidad lechera; pero también se debe a que se esta trabajando bajo sistema estabulado con lo cual se tiene un mayor control. En cambio el alto índice de productividad de majada del Blackbelly está dado por su mayor fertilidad, mayor tasa de sobrevivencia de corderos y peso al nacimiento. La clasificación mas baja corresponde al Katahdin, la cual esta influenciada por su baja prolificidad y su alto intervalo entre parto.
El mejor desempeño del Assaf y Blackbelly, tal vez, se deba parcialmente, a las mejores condiciones de manejo del primero que como ya se menciono es en estabulación. En cambio el Black Belly presenta una mejor alimentación, en los Valles Pichis y Palcazú, y a un mayor tiempo de adaptación; en cambio, la raza Katahdin está sometida a pastoreo en residuos de cosecha y los datos corresponden a una primera generación que probablemente no estaba bien adaptada. ES de mencionar que el katadhin es una de las razas de mayor rendimiento carnico de los ovinos tropicales.
Sin embargo hay que hacer notar que ninguna de las razas han alcanzado los niveles reportados en otros países por la bibliografía revisada, en cuanto a prolificidad y rendimiento en general.
De las razas brasileñas introducidas no hay reportes consistentes, pero se vislumbra que Morada Nova
podría desempeñarse mejor en la selva peruana por la similitud con su hábitat de origen.
COnCLUSIOnES1. En condiciones nacionales, las razas Assaf y Blackbelly se desempeñan mejor en productividad y eficiencia por rebaño.
2. El peso al nacimiento es aproximadamente el 10 % del peso al sacrificio, entonces la multiplicación de los índices por 10 los convertiría a la base del peso al sacrificio.
3. Los resultados de este tipo de investigaciones junto a experiencias prácticas sirven para planificar programas de desarrollo, y proyectar como meta de dicho programa, la utilización del potencial completo de las Ovejas de Pelo.
4. Es necesario que la introducción de razas sean evaluada bajo condiciones reales en las fincas o productores antes de que se fomenten ampliamente.
5. La evaluación en fincas ayudarán asegurar que las razas sean apropiadas para las necesidades y los recursos de los productores y de la zona.
6. Las Ovejas de pelo son un recurso genético de considerable potencial para la producción de carne en regiones tropicales, y posiblemente en las templadas también.
Cuadro 4: Evaluación de las razas introducidas
Característica B.B. Pb. Kt. M.n. S.I. As.
Tamaño camada: No. Cord.
Interv.+ parto: Días
Sobrevivencia:%
Peso al nacimiento: kg.
Peso de borrega: kg.
Peso metabólico (W75)
* IPM x 103
** IEM
1.77
250
91.53
3.12
38.00
15.90
20.23
1.27
1.40
216.4
79.10
2.71
34.00
14.10
13.87
0.98
1.35
270
85.00
2.50
39.00
16.20
10.63
0.66
1.76
246
79.8
2.88
30.00
12.80
14.01
1.09
1.28
246
79.8
2.88
39.00
16.10
11.96
0.74
1.55
240
91.88
4.0
48.0
20.04
23.74
1.18
* Índice de productividad de Manada(IPM) = gr. de peso de cordero producido por día de intervalo entre parto.
**Índice de eficiencia de Manada (IEM) = gr. de peso de cordero por Kg. de peso metabólico de borrega adulta por día de intervalo entre parto.

Arch. Latinoam. Prod. Anim. Vol. 15 (Supl. 1) 2007•315
XX Reunión ALPA, XXX Reunión APPA-Cusco-Perú
REFEREnCIAS
Atto, j.A y m.S. Guarnizo,. Y. 1989, “Comportamiento reproductivo de dos razas de ovinos y un cruce (criollo x suffolk:F2), en condiciones semiextensivas de la provincia de Paita, Departamento de Piura”. Reunión APPA.
Radford, G.E. 1972. “Genetic control of litter size in sheep”. Journal Reproductive and Fertilite. Suppl. 15. pp 23-41.
Calle E., R., 1986, “Ovinos tropicales en los valles Pichis y Palcazú”.Instituto nacional de desarrollo. Proyecto especial Pichis-Palcazú.
Cardi. 1981. Animal production systems in the Eastern Caribean. Consultant Report No. 7. Morrilton, Arkansas. Winrock International.
Epstein, H. 1971. “The original of the domestic animals of Africa” Vol. II. New York, London and Munich. Africana Publishing Co.
Fitzhugh, H.a. Y C.e. Bradford, 1983, “Ovejas de Pelo del África Occidental y de las Amétricas”. Winrock International USA.
Mason, I. L., 1980. “Ovejas prolíficas tropicales”. FAO. Animal Production and Health. Boletin 17. Roma.
Ministerio de Agricultura: 1989. Archivo de permisos zoosanitarios 1985, 1987, 1988.
Universidad Nacional de Piura, 1992. Seminario Taller Internacional. “Producción de Ovinos de Pelo”
Velez N., M., 1984. “La crianza de cabras y ovejas en el trópico”, Escuela Agrícola Panamericana Zamorano, Honduras.