fisiología del dolor
DESCRIPTION
Fisiología del dolorTRANSCRIPT

FISIOLOGÍA DE DOLORProf. Sabino

CLASIFICACIÓN DEL DOLOR
1. Según el lugar de origen del dolor, puede ser:
Dolor superficial. Si el dolor somático proviene de la piel. Por ej., si se pincha la piel con un alfiler.
Dolor profundo. Si el dolor somático proviene de los músculos, huesos, articulaciones y tejido conjuntivo. Por ej., el dolor articular.
Dolor visceral. Es el dolor que proviene de las vísceras. Por ej., el dolor de la vesícula biliar.
2. Según el tiempo de evolución, se clasifican en:
Dolor agudo. Es el dolor que dura poco tiempo, generalmente menos de dos semanas. Por ej., el dolor de muelas. El dolor está limitado por lo general al lugar de la lesión, por eso este dolor es claramente localizable y la intensidad del mismo depende de la intensidad del estímulo.
Dolor crónico. Es dolor que dura largo tiempo, generalmente más de un año. Por ej., dolor de cardíaco (angina de pecho). Son dolores recidivantes y duraderos.

DOLOR: TIPOS
TIPOS DE DOLOR: SEGÚN EL TIEMPO DE EVOLUCIÓN
Dolor agudo Dolor crónico
< de 2 semanas > de 1 año
A C
Mielínica Amielínica
Localizado Difuso
Vc = 6-30 m/s Vc = 0,5 a 2 m/s
Defensivo Destructivo
Ej., Dolor de muela Ej., Angina de pecho

PICOR
Es otra cualidad sensorial cutánea que está al menos emparentada con el dolor. Posiblemente sea una forma especial de la sensación dolorosa que aparece en determinadas condiciones de estímulos. A favor de ello está que toda una serie de picores conducen a sensaciones dolorosas cuando aumenta la intensidad y que la interrupción de la vía del cordón anterolateral de la médula espinal se acompaña de la pérdida de la sensación de picor, mientras que un trastorno del sentido de presión y tacto (cordones posteriores) no tiene ninguna influencia sobre la sensación de picor. Tampoco se ha podido demostrar que la piel sólo es sensible al picor en determinados puntos como sucede con el dolor.La sensación de picor sólo es provocable desde capas más externas de la epidermis, mientras que el dolor se puede producir también desde las capas profundas de la piel. Finalmente habría que mencionar que para la aparición de la sensación de picor parece ser necesaria la liberación de una sustancia química, quizá la histamina. Una inyección intradérmica de histamina produce un fuerte picor y en lesiones de la piel acompañadas de picor, se libera histamina.

COMPONENTES DEL DOLOR
El dolor presenta cuatro componentes, que son:
1. Componente sensorial. Es cuando el dolor se acompaña de una sensación sensorial. Por ej.,
al introducir una mano en agua fría o tibia se excitan receptores de la piel y pueden
producirnos, una sensación de frío o calor.
2. Componente emocional. Es cuando el dolor se acompaña de una sensación de placer o
displacer. Por ej., cuando nos bañamos en un día muy caluroso de verano con el agua a
25 ºC, sentimos no sólo una sensación de frío en la piel, sino que también nos produce
simultáneamente una sensación refrescante, agradable (placer). Pero si lo hacemos en un
día frío de invierno nos produce entonces una sensación desagradable (displacer). A este
aspecto del dolor se le denomina también componente efectivo.
3. Componente vegetativo. Es cuando la estimulación dolorosa se acompaña de componentes
vegetativos como reacciones cardiovasculares, pupilares... Los componentes vegetativos
pueden ser muy marcados en los dolores viscerales. Por ej., el cólico biliar van
acompañados de vómitos, sudoración...
4. Componente motor. Estos componentes motores del dolor son conocidos como reflejo de
huida o de protección. Por ej., en la meningitis se produce una contracción refleja de la
musculatura del dorso del paciente adquiriendo una postura característica.

TERMINACIONES LIBRES- SUBSTANCIAS ALGÓGENAS
Terminaciones libres
1.- Nociceptores
2.- Mecanonociceptores
3.- Termonociceptores
Substancias algógenas
Bradiquinina
Serotonina
Prostaglandinas (PGE y PGF)
Histamina
Acetilcolina
Ácido láctico
Iones (K+, H+ y Cl-)
Sustancia P
La bradiquinina, serotonina e histamina, administradas en mínimas cantidades, pueden considerarse como fisiológicas, causando dolor, eritema (hiperemia) y edema, signos típicos de la inflamación.

FISIOLOGÍA DEL DOLOR
Nociceptores. Son receptores que responden a estímulos nociceptivos.
La percepción de la lesión o posible lesión se detecta por los nociceptores y se transmite a través de las fibras nerviosas aferentes a las astas posteriores de la médula espinal. En este centro nervioso, estos impulsos se encuentran con un sistema de control de entrada (puerta del dolor). Tal convergencia puede inhibir o exagerar el mensaje transmitido. Neuronas de segundo orden proyectan fibras que conducen la información sobre estímulos dolorosos a través de los haces espinotalámicos hasta amplias regiones del tronco del encéfalo y el tálamo. Otras fibras procedentes de la cabeza y el cuello, tramitan la información de estas regiones (tronco del encéfalo y tálamo), a través del nervio trigémino.
En el tálamo, las fibras transmisoras del dolor de la cabeza arriban al núcleo ventral posteromedial del tálamo, mientras que las fibras transmisoras de la información procedente del resto del cuerpo van a sinaptar con el núcleo ventral posterolateral del tálamo, y de aquí lo proyectan a la corteza cerebral, concretamente a las áreas somestésicas I (áreas 3,1 y 2) y II (áreas 5 y 7).

SISTEMA ANTEROLATERAL DE LA MÉDULA
Se encarga de tramitar la información de la sensibilidad termoalgésica y del tacto, al servicio de la conducta instintiva para el mantenimiento del estado de salud del organismo.
Haz neoespinotalámico (7)
Llamado también vía espinotalámica directa, lemnisco lateral y sistema lateral del dolor, correspondiendo esta última denominación al clínicamente conocido como haz espinotalámico lateral, tramita la sensibilidad termoalgésica o sensibilidad calor-frío y dolor. Este haz es el responsable del aspecto discriminativo, sensorial del dolor. Informa del comienzo de la agresión; es decir, actúa como el mecanismo operativo de un sistema de aviso o de alarma, defensivo en todo caso. Consta de una organización estructural secuenciada a tres órdenes de neuronas: la neurona de primer orden asienta en los ganglios raquídeos; su prolongación periférica recoge la información en los receptores sensoriales o terminaciones sensoriales libres, que son estructuras desnudas, terminales de fibras mielínicas delgadas A, en su mayor parte, y amielínicas C, cuyas velocidades de conducción son de 6-30 m/s y 0,5-2 m/s, respectivamente. La prolongación central penetra hasta las astas posteriores de la médula espinal, distribuyéndose por las láminas I, o Zona marginal de Lissauer, y II, o sustancia gelatinosa de Rolando, en donde abundan interneuronas o neuronas “G”, neuronas de segundo orden, con la que sinaptan muchos de los axones A y C. Los axones de las neuronas de segundo orden cruzan al lado contrario de la médula y ascienden por el cordón lateral hasta sinaptar con el núcleo ventral posterolateral del tálamo (VPL). Finalmente, las neurona de tercer orden citadas envían sus axones a través del cuerpo posterior de la cápsula interna y corona radiada, para terminar en la circunvalación postcentral o áreas somestésicas I (S-I o áreas 3, 1 y 2) y II (S-II o áreas 5 y 7), en donde la información tramitada se torna en mensaje consciente.

VÍAS DEL DOLOR: NEOESPINOTALÁMICO Y PALEOESPINOTALÁMICO

SISTEMA ANTEROLATERAL DE LA MÉDULA
Haz paleoespinotalámico (6)
Es el sistema responsable de los aspectos afectivo-emocionales, que tramitan impulsos dolorosos de carácter difuso, quemantes, siendo responsable de la propagación del dolor visceral.
Conocido también como haz espino-retículo-talámico, es una vía espinotalámica indirecta.. Este haz , filogenéticamente antiguo, es multineural y multisináptico. Consta de una organización estructural secuenciada a tres órdenes de neuronas: la neurona de primer orden asienta en los ganglios raquídeos; su prolongación periférica recoge la información en los receptores sensoriales o terminaciones sensoriales libres, que son estructuras desnudas, terminales de fibras amielínicas C, cuya velocidad de conducción es de 1 m/s. La prolongación central penetra hasta las astas posteriores de la médula espinal, Los axones de las neuronas de segundo orden son cruzados y directos que ascienden por la parte anterolateral de la médula, terminando en núcleos de la sustancia gris periacueductal (SGPA), formación reticular del bulbo, protuberancia y mesencéfalo, en donde sinaptan con neuronas que emiten fibras que alcanzan los núcleos talámicos inespecíficos intralaminares (centromediano y parafascicular) y de la región posterior del tálamo, así como el sistema límbico-hipotalámico. Algunas fibras de os núcleos talámicos citados llegan al área somestésica II (áreas 5 y 7)
Esta vía responsable del dolor no localizado, crónico, lento, ya que los núcleos talámicos en donde asientan sus neuronas son inespecíficos, esto es, del llamado sistema de proyección difusa, carente de organización somatotópica.

MECANISMO PERIFÉRICO: TEORÍA DEL CONTROL DE LA PUERTA DE ENTRADA
La teoría del control de la puerta de entrada del dolor fue propuesta por Melzack y Wall (1965). Esta teoría trata de explicar la función de las vías neuronales, tanto ascendentes como descendentes, para crear un mecanismo de control (una puerta) que regula la transmisión de los impulsos nociceptivos. El mecanismo de puerta de entrada opera en el asta posterior de la médula espinal, en la denominada sustancia gelatinosa de Rolando. La puerta tiene un componente central y otro periférico, que modulan la transmisión del dolor, en las proximidades.Este mecanismo periférico consiste en impedir el paso a los impulsos nociceptivos (dolorosos). Estos, penetran por las astas posteriores de la médula espinal, desde las terminaciones libres (nociceptores, termorreceptores y mecanonociceptores) y se dirigen hacia las neuronas de segundo orden, presentes en las astas posteriores. Este mecanismo periférico está modulado por la acción de las neuronas G.Cuando el dolor es conducido por las fibras amielínicas C, la puerta del dolor está totalmente abierta, debido a que no se estimulan las neuronas G, y por lo tanto no hay influencia sobre el estímulo doloroso. Entonces las fibras amielínicas C sinaptan con las neuronas de segundo orden de la vía ascendente espinotalámica, conduciéndose el impulso doloroso hasta el tálamo. Cuando el dolor es conducido por las fibras mielínicas A, el control de la puerta del dolor está semiabierta, es decir permite el paso del estímulo doloroso, aunque luego lo bloquea. Esto se produce porque las fibras A emiten una colateral para estimular la neurona G, la cual cierra la puerta del dolor después de pasar el estímulo doloroso.Cuando son las fibras mielínicas A las que conducen el estímulo doloroso, la puerta del dolor está cerrada totalmente, debido a que influyen activando las neuronas G de la sustancia gelatinosa de Rolando, suscitando el fenómeno de cierre de la puerta de entrada del dolor.Otros avances importantes en nuestra comprensión del efecto de los factores centrales en la percepción del dolor, es el descubrimiento de que el sistema nervioso contiene endorfinas, sustancias químicas producidas por el cerebro que tienen propiedades muy parecidas a las de los opiáceos, como la morfina.

MECANISMO DE MODULACIÓN DE DOLOR

MECANISMO DE MODULACIÓN DE DOLOR

MECANISMO CENTRAL DE LA MODULACIÓN DEL DOLOR
Dentro de la formación reticular bulbomesencefálica destacan una serie de núcleos, con poder de modulación
del dolor, que son:
1. Sustancia gris periacueductal (SGPA).
2.Núcleo magno de rafe (NMR).
3. Locus ceruleus.
4.Núcleo paragigantocelular.
Las fibras descendentes que proceden de estas regiones sinaptan con interneuronas o neuronas G de la
sustancia gelatinosa de Rolando (lámina II), suscitando la liberación de neuromediadores (serotonina,
encefalinas y noradrenalina, entre otras.
El fascículo dorsolateral. Es una vía serotoninérgica/endorfinonérgica, que parte del núcleo magno del rafe
(NMR) y termina en las astas posteriores de la médula espinal, sinaptando con la neurona “G”, ejerciendo su
efecto modulador, atenuador y hasta supresor del dolor, mediante la liberacion de serotonina, que induce la
descarga de encefalinaspor la neurona “G”, provocando la consabida inhibición. El núcleo del trigémino
recibe igualmente influencia del NMR, por fibras serotoninérgicas.
Por su parte, el NMR está supeditado a la influencia de la SGPA, que a su vez resulta estimulada por las
fibras nociceptivas ascendentes.
En muchos casos de dolor crónico hay un déficit o una falta de activación de esta vía serotoninérgica o
endorfinérgica.
La SGPA cuenta con gran densidad de receptores opioides, activando al NMR mediante la liberación de
endorfinas por las fibras que interconectan ambos núcleos. Por el contrario, la inyección de naloxona en la
SGPA o en el III ventrículo impide o suprime los efectos analgésicos, por la fijación de este antagonista
opiáceo en los receptores de la SGPA.

SUSTANCIAS ALGÓGENAS
El dolor resulta de la activación de los nociceptores por agentes mecánicos, térmicos y químicos, repartidos intersticialmente en torno a la musculatura lisa de los vasos. Entre las sustancias algógenas o agentes químicos que producen dolor, citamos las siguientes: bradiquinina, serotonina, ácido láctico e iones K+, H+ y Cl-. Mención aparte, merece la sustancia P, tanto este undecapéptido es considerado neurotransmisor de las fibras mielínicas A y amielínicas C, propagadoras de impulsos nociceptivos o dolorosos, como por sus interacciones-efectos con los sistemas de modulación y de control del dolor. Igualmente, otra sustancia implicada en la fenomenología del dolor es el glutamato.La bradiquinina favorece la síntesis de prostaglandinas, cuyo precursor es el ácido araquidónico, un componente de los fosfolípidos de las membranas celulares, liberado este compuesto por la acción enzimática de la fosfolipasa A2. Por su parte, el ácido araquidónico liberado se metaboliza precozmente por la actividad de una ciclooxigenasa, con la subsiguiente producción de endoperóxidos cíclicos inestables que se transforman en protaglandinas y prostaciclinas.A la luz de estos sencillos conocimientos, se ha podido instaurar una base terapéutitica eficaz, consistente en la aplicación de fármacos analgésicos-antiinflamatorios-antipiréticos, como son los derivados del ácido acetilsaliciíico, que inhiben la actividad de la enzima ciclooxigenasa, responsable de la conversión de ácido araquidónico en protaglandinas.De esta manera, tendríamos, dos tipos de fármacos para combatir el dolor periférico: los fármacos tipo aspirina y los esteroides.OPIÁCEOS Y RECEPTORES En la década de los 70, se demostró que la inyección de morfina en los núcleos implicados en el mecanismo central de la modulación del dolor (NMR, núcleo ceruleus y otros) ejerce marcados efectos analgésicos. Dicho efecto analgésico selectivo se atribuyó a que la morfina y afines inyectados se ligan a receptores opiáceos, distribuidos por el sistema nervioso central, relacionados con la sensibilidad al dolor como los haces espinotalámicos.

RECEPTORES OPIÁCEOS: TIPOS Y DISTRIBUCIÓN
TIPOS DE RECEPTORES
Receptores (mu) Receptores (delta)
Receptores k (kappa) Receptores (sigma)

MÉDULA ESPINAL: VÍAS DESCENDENTESProf. Sabino
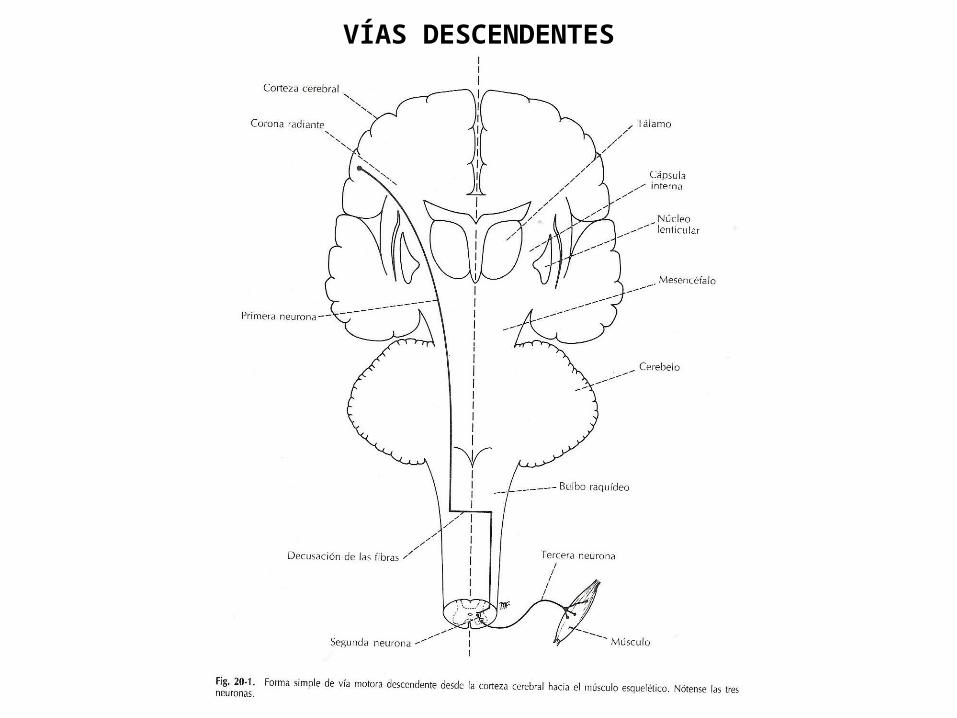
VÍAS DESCENDENTES

VÍAS DESCENDENTES: FASCÍCULO CORTICOESPINAL

VÍAS DESCENDENTES: FASCÍCULO CORTICOESPINALLa corteza motora se reconoce microscópicamente por la presencia de células piramidales gigantes o de Betz. Sin embargo, parten muchas más proyecciones desde las células piramidales pequeñas o de tamaño mediano de esta área, que desde las células de Betz.
La corteza motora se localiza en la parte posterior del lóbulo frontal, por delante de la cisura de Rolando.
Funcionalmente, la corteza motora se divide en:
1. Corteza motora primaria (área 4 de Brodmann).
2. Corteza motora premotora (área 6 de Brodmann).
3. Corteza motora suplementaria (área 8 de Brodmann).
Los tractos corticoespinal (piramidal) o corticobulbar se originan en las células piramidales de la capa 5 de la corteza motora, así como en otras regiones corticales, como la corteza premotora, la corteza motora suplementaria y la circunvolución poscentral.
La corteza motora controla tanto los músculos distales como proximales, sin embargo, las proyecciones corticoespinales y corticobulbares del sistema lateral son fundamentales para el control de los músculos distales de las extremidades contralaterales, de la región inferior de la cara y de la lengua.
La corteza motora primaria es la que manda la orden a las motoneuronas alfa de las astas anteriores de la médula espinal, para la ejecución del movimiento. La corteza motora primaria recibe amplias aferencias desde la corteza somatosensorial (áreas 3, 1 y 2) y desde áreas de integración sensitiva de la corteza parietal (áreas 5 y 7). Las neuronas de la vía piramidal proyectan directamente sobre las motoneuronas del asta anterior de la médula espinal o sobre las interneuronas medulares.

VÍAS DESCENDENTES: FASCÍCULO CORTICOESPINAL
La corteza motora y la corteza motora suplementaria son responsables de la planificación del movimiento, especialmente de la secuencias de movimientos complejos en la musculatura distal, aunque cada una de ellas contribuye en distintos aspectos. Por estudios de tomografía de emisión de positrones, se sabe que la corteza motora suplementaria se activa aproximadamente un segundo antes de la realización del movimiento. Las cortezas motoras suplementarias de ambos hemisferios actúan juntas en la elaboración de un plan de movimientos gracias a sus conexiones, a través del cuerpo calloso. En las lesiones de la corteza motora suplementaria de un hemisferio, el principal déficit observado es la dificultad para realizar movimientos coordinados con ambas manos, por ej., enhebrar una aguja, abrochar los botones, etc.
La corteza premotora, recibe un alto número de aferencias de la corteza visual y se relaciona con la selección de una acción entre las distintas posibles respecto a los estímulos recibidos (por ej., arrancar al ponerse el verde el semáforo).
La corteza premotora y la corteza motora suplementaria, además de elaborar planes de movimientos, también están implicadas en el aprendizaje de patrones de movimientos complejos (por ej., montar en bicicleta, conducir, etc.).
La corteza motora (corteza premotora y corteza motora suplementaria) no primaria desempeña un importante papel en el control, coordinación y regulación de los movimientos complejos.

VÍAS DESCENDENTES: FASCÍCULO CORTICOESPINAL
Las fibras de la vía corticoespinal se originan aproximadamente 1/3 en la corteza motora primaria (área 4), 1/3 en la corteza motora secundaria (área 6) y 1/3, tiene su origen en el lóbulo parietal (áreas 3,1 y 2); por ende, dos tercios de las fibras se originan en la circunvolución precentral y un tercio se origina en la circunvolución postcentral. Es interesante destacar que la mayoría de las fibras corticoespinales son mielínicas y pequeñas, con una conducción relativamente lenta.Las fibras descendentes convergen en la corona radiada y luego atraviesan el brazo posterior de la cápsula interna. Allí, las fibras están organizadas de tal modo que las que se encuentran más cerca de la rodilla se vinculan con porciones cervicales del cuerpo, mientras que las situadas más posteriormente se relacionan con el miembro inferior. Luego el tracto continúa a través de los pedúnculos cerebrales, donde las fibras vinculadas con las porciones cervicales del cuerpo están situadas medialmente, mientras que las relacionadas con la pierna se ubican lateralmente. Al ingresar en la protuberancia el tracto es separado en muchos haces por los núcleos del puente , formándose entonces las fibras pontocerebelosas. En el bulbo raquídeo los haces se reúnen a lo largo del borde anterior para formar un ensanchamiento conocido como pirámide (de ahí el nombre alternativo de la vía piramidal). En la unión del bulbo raquídeo y la médula espinal la mayoría de las fibras (90%) cruzan la línea media en la decusación de la pirámide e ingresan en el cordón lateral de la médula espinal para formar el tracto corticoespinal lateral. Las fibras restantes (10%) no se cruzan en la decusación sino que descienden en el cordón anterior de la médula espinal como el tracto corticoespinal anterior. Estas fibras finalmente cruzan la línea media y terminan en las astas anteriores de la médula espinal en las regiones cervical y torácica superior.El tracto corticoespinal lateral desciende a lo largo de la médula espinal; sus fibras terminan en las astas anteriores de todos los segmentos de la médula espinal.La mayoría de las fibras corticoespinales establecen sinapsis con neuronas internunciales, las que a su vez lo hacen con neuronas motoras alfa y algunas neuronas gamma.Los haces corticoespinales no constituyen la única vía para los movimientos voluntarios . Más bien forman la vía que confiere velocidad y agilidad a esos movimientos y por ende se utiliza para llevar a cabo movimientos hábiles y rápidos. Muchos de los movimientos voluntarios básicos simples están mediados por otros tractos descendentes.
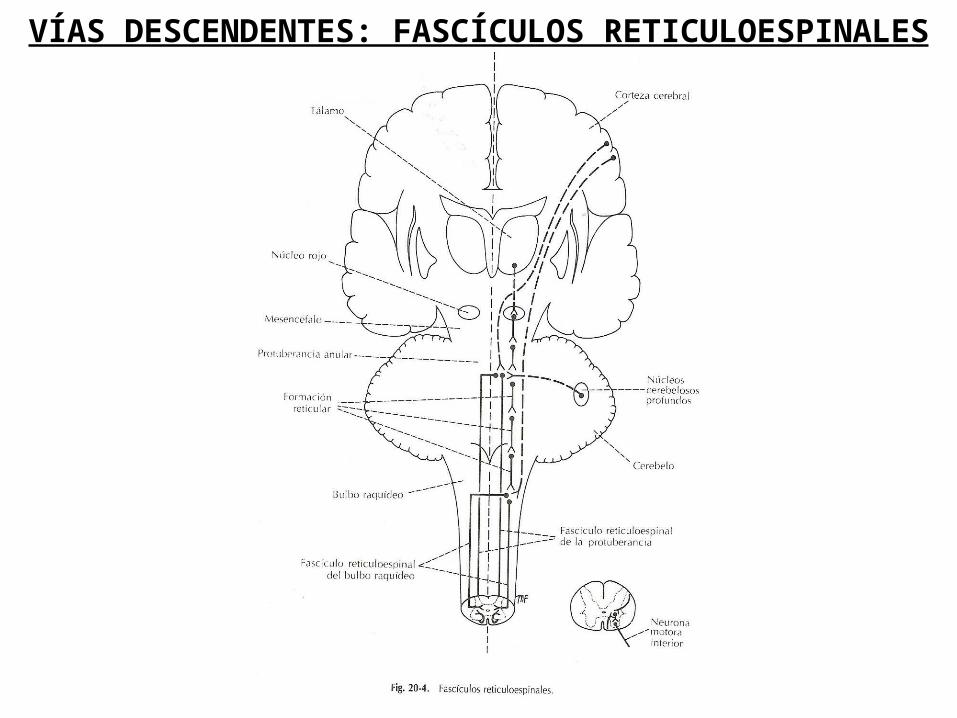
VÍAS DESCENDENTES: FASCÍCULOS RETICULOESPINALES

VÍAS DESCENDENTES: FASCÍCULOS RETICULOESPINALES
En todo el mesencéfalo, la protuberancia y en el bulbo raquídeo existen grupos de neuronas y fibras nerviosas dispersas que, en conjunto reciben el nombre de formación reticular.
Desde la protuberancia, estas neuronas envían axones, la mayoría son directos, hacia la médula espinal y forman el fascículo reticuloespinal de la protuberancia o el fascículo reticuloespinal medial.
Desde el bulbo raquídeo, neuronas similares, envían axones, que son cruzados y directos, hacia la médula espinal y forman el fascículo reticuloespinal bulbar o el facículoreticuloespinal lateral.
Las fibras reticuloespinales, procedentes de la protuberancia, descienden a través del cordón anterior, mientras que las que provienen del bulbo raquídeo descienden por el cordón lateral. Ambos haces de fibras entran en las astas anteriores de la médula espinal y pueden facilitar o inhibir la actividad de las neuronas motoras alfa y gamma. De este modo, los fascículos reticuloespinales influyen en los movimientos voluntarios y en la actividad refleja.
El fascículo reticuloespinal medial estimula las neuronas de los músculos extensores e inhiben a las motoneuronas de los músculos flexores.
El fascículo reticuloespinal medial estimula las neuronas de los músculos flexores e inhiben a las motoneuronas de los músculos extensores.

VÍAS DESCENDENTES: FASCÍCULO TECTOESPINAL

VÍAS DESCENDENTES
FASCÍCULO TECTOESPINAL
Las fibras de este fascículo nacen en las neuronas del tubérculo cuadrigémino superior del mesencéfalo. La mayoría de las fibras cruzan la línea media poco después de su origen y descienden a través del tronco del encéfalo. Este fascículo desciende por el cordón anterior de la médula espinal. La mayoría de las fibras terminan en las astas anteriores en los segmentos cervicales superiores haciendo sinapsis con las neuronas internunciales y éstas con las motoneuronas (alfa y gamma) de la musculatura flexora. Se cree que estas fibras se encargan de los movimientos posturales reflejos en relación con los estímulos visuales.
FASCÍCULO RUBROESPINAL
El núcleo rojo está ubicado en el mesencéfalo, a nivel del tubérculo cuadrigémino superior. Los axones de las neuronas de este núcleo cruzan la línea media a ese nivel y descienden a través de la protuberancia y el bulbo raquídeo para entrar en el cordón lateral de la médula espinal. Las fibras termina haciendo sinapsis con las neuronas internunciales en el asta anterior de la médula espinal y dichas neuronas con las motoneuronas (alfa y gamma).
El fascículo facilita la actividad de los músculos flexores e inhibe la actividad de los músculos extensores o posturales.
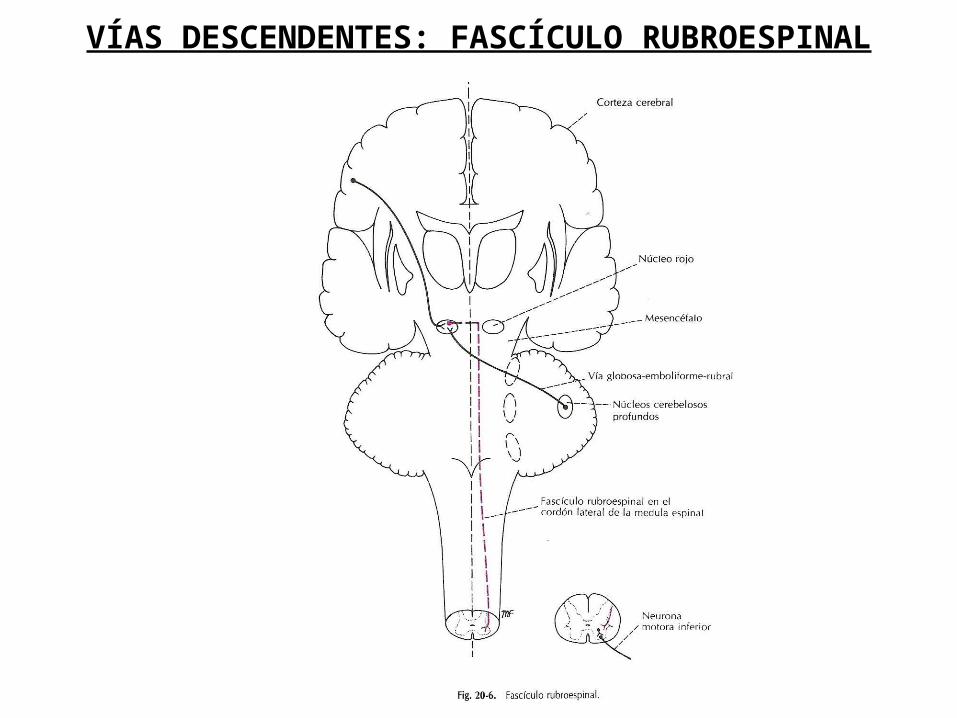
VÍAS DESCENDENTES: FASCÍCULO RUBROESPINAL

VÍAS DESCENDENTES: FASCÍCULO OLIVOESPINAL

VÍAS DESCENDENTES
FASCÍCULO OLIVOESPINAL
El núcleo olivar inferior se halla en el bulbo raquídeo. Las neuronas de este núcleo originan los axones que forman el fascículo olivoespinal. El fascículo cruza la línea media y desciende por el cordón lateral de la médula espinal. Las fibras terminan haciendo sinapsis con neuronas internunciales en la médula espinal y éstas a su vez con las motoneuronas (alfa y gamma).
El fascículo facilita la actividad de los músculos flexores e inhibe la actividad de los músculos extensores o posturales.
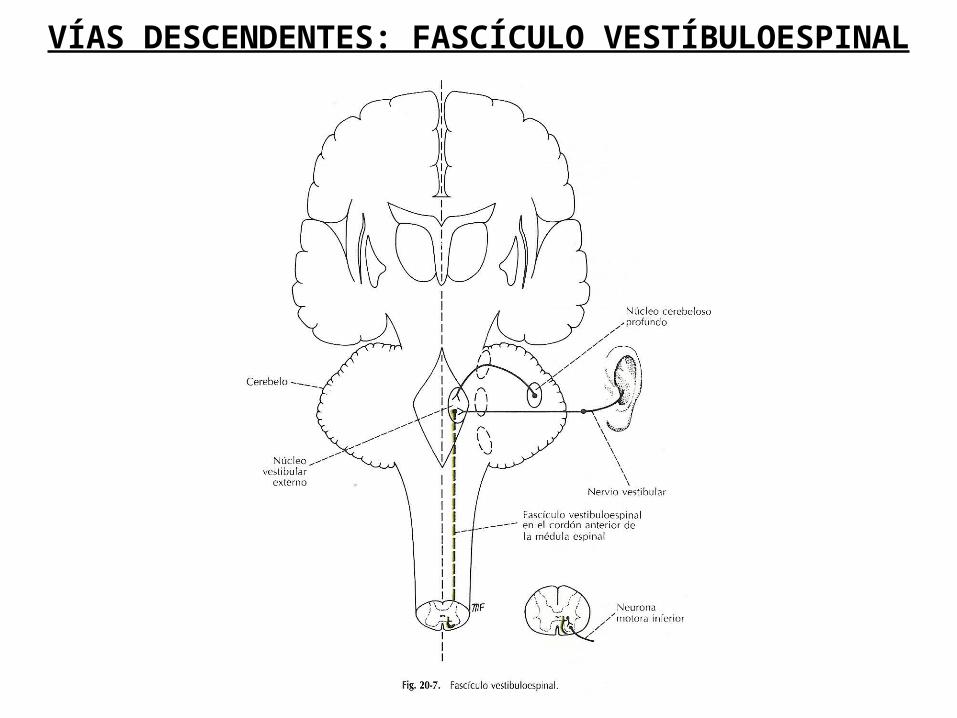
VÍAS DESCENDENTES: FASCÍCULO VESTÍBULOESPINAL

VÍAS DESCENDENTES: HAZ VESTÍBULOESPINAL MEDIAL

VÍAS DESCENDENTES
FASCÍCULO VESTIBULOESPINAL
Los núcleos vestibulares están ubicados en la protuberancia y el bulbo raquídeo por debajo del piso del cuarto ventrículo. Las neuronas del núcleo vestibular lateral o de Deiters dan origen a los axones que forman el fascículo vestibuloespinal lateral. Las fibras terminan haciendo sinapsis con las neuronas internunciales del asta anterior de la médula espinal, las cuales a su vez sinaptan con las motoneuronas (alfa y gamma) de los músculos extensores estimulándolos e inhibiendo a los músculos flexores.
El fascículo vestibuloespinal lateral facilita la excitación de las neuronas motoras (alfa y gamma) de los músculos extensores del tronco y extremidades en respuesta a la aceleración o deceleración lineal o a la inclinación estática de la cabeza.
Las neuronas del núcleo vestibular medial dan origen a los axones que forman el fascículo vesíbuloespinal medial, que proyecta bilateralmente sobre las neuronas internunciales de la médula espinal y éstas a su vez sobre las motoneuronas de los músculos flexores del cuello, estimulando e inhibiendo a la musculatura extensora. Su función consiste en el control de la posición de la cabeza en conjunción con los movimientos oculares (respuesta estática y dinámica).

VÍAS DESCENDENTES: HAZ VESTÍBULOESPINAL LATERAL
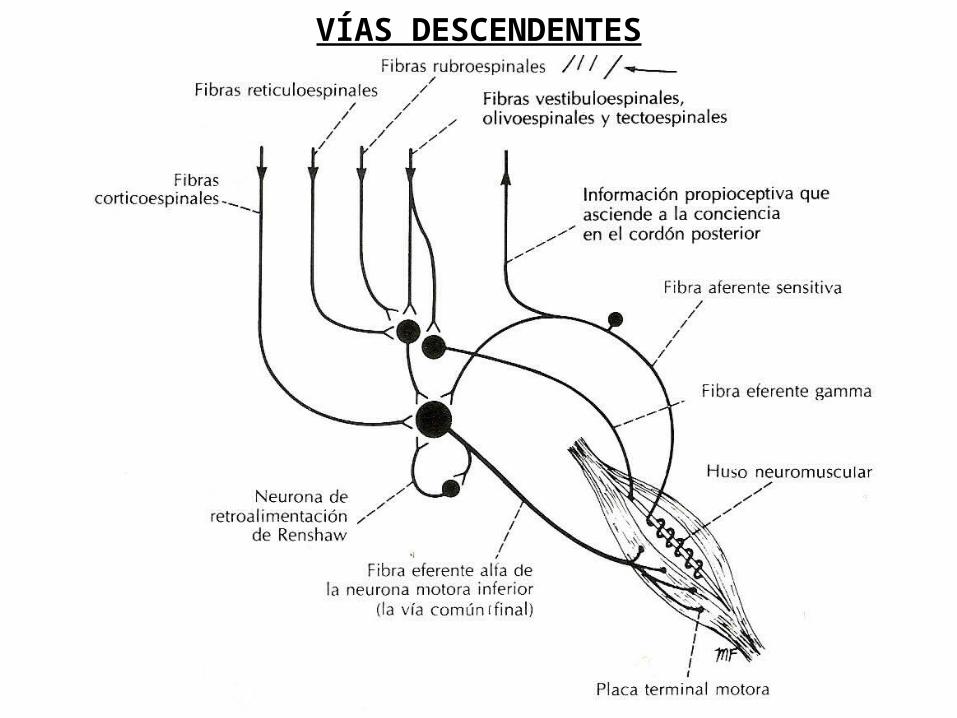
VÍAS DESCENDENTES