evaluacion de factores que afectan la … de grado... · factores relacionados al ciclo estral en...
TRANSCRIPT

UNIVERSIDAD NACIONAL DE CORDOBA
FACULTAD DE CIENCIAS AGROPECUARIAS
ESCUELA PARA GRADUADOS
INSTITUTO DE REPRODUCION ANIMAL CÓRDOBA
(IRAC)
ESPECIALIZACIÓN EN REPRODUCCIÓN BOVINA
EVALUACION DE FACTORES QUE AFECTAN LA
PRODUCCION DE EMBRIONES LUEGO DE UN
TRATAMIENTO SUPEROVULATORIO EN EL
GANADO BOVINO EN ARGENTINA
M.V. Daniel Giovanny Castro López
M.V. Diego Andrés Rodríguez Riveros
Trabajo Final
Para optar al título de Especialista en Reproducción Bovina
Córdoba, 2014

2
INTRODUCCION
SUPEROVULACIÓN EN BOVINOS
El principal objetivo al implementar un protocolo de superovulación en una
producción bovina es el de maximizar y potencializar el valor genético de una
hembra o de un grupo de hembras donantes con importancia genética. La
aplicación del protocolo permite controlar el crecimiento folicular, permitiendo
aumentar el número de ovulaciones por hembra, con la finalidad de incrementar la
cantidad de embriones producidos y transferidos a las hembras receptoras.
El aumento en el número de ovulaciones se logra a través de la aplicación de
hormonas que favorecen el crecimiento de todos los folículos reclutados en una
onda folicular, impidiendo lo que ocurre fisiológicamente en un ciclo estral: que un
solo folículo sea dominante y que los subordinados regresen y se atresien.
CONTROL FARMACOLÓGICO DEL CICLO ESTRAL PARA
LOGRAR LA SUPERESTIMULACIÓN Y SUPEROVULACIÓN EN
LAS HEMBRAS DONANTES
El tratamiento tradicional se inicia con la detección de celo a las donantes, tomado
como Día 0 del tratamiento. Posteriormente, al día 10 se aplica una dosis de
gonadotrofina (ya sea extracto de pituitaria porcina o eCG) se estimula el
desarrollo folicular y la síntesis de estradiol; al día 12 una inyección de
prostaglandina (PGF2α) que induce la regresión del cuerpo lúteo y presentación
de celo al día 14, el cual es respuesta a la acción de la PGF2α (Beal, 1996;
Rathbone et al., 2001) seguida de inseminación a las 12 y 24 horas de observar el
celo. Finalmente, la colecta de embriones se realiza al día 21 del tratamiento.

3
Este protocolo tradicional ha sido modificado con el fin de mejorar la respuesta
superovulatoria. Bó y su grupo de trabajo (1996) estudiaron el efecto de
tratamientos con progestágenos y 17β-estradiol en la sincronización folicular y
encontraron que la respuesta de los individuos sometidos a este tratamiento es
similar al de vacas tratadas en la emergencia de la segunda onda folicular,
encontrando resultados para producción de embriones similar o incluso mayor al
tratamiento tradicional (Bó y Mapletoft, 2012). El uso del estradiol, principal
producto secretado por el ovario, conjuntamente con progestágenos, afecta
directamente el desarrollo folicular ya que la atresia folicular es causada por la
supresión de la FSH circulante y consecuentemente el desarrollo de una nueva
onda folicular (Bó et al., 1991). La atresia es seguida de un pico de FSH y la
emergencia de la nueva onda.
El tratamiento base con estradiol consiste en la inserción de un dispositivo de
liberación lenta de progestágeno, una dosis de 5 mg de estradiol y 100 mg de P4
al día 0, tomado aleatoriamente en cualquier día del ciclo. Se continua con la
superestimulación en dosis decrecientes de Folltropin-V a partir del día 4
aplicadas cada 12 horas, dado por la corta vida media de la FSH (Laster, 1972), el
día 6 se aplica PGF para ocasionar la regresión del folículo dominante de la
primera onda, se retira el dispositivo al día 7, 24 horas después del retiro se aplica
una dosis de GnRH o LH y la inseminación se realiza 36 y 48 horas después de
retirado el dispositivo. La colecta se realiza al día 15 de iniciado el tratamiento (Bó
y Mapletoft, 2014). Otros tratamientos superovulatorios incluyen también sales de
estradiol, en este caso el Valerato de Estradiol (5mg) es aplicado conjuntamente
con 3 mg de progestágeno y un dispositivo subcutáneo también de progestágeno,
al día 5 se inicia la superestimulación con el uso de Folltropin-V. El Valerato de
Estradiol tiene una acción inmediata, comparado con otros derivados del 17β-
Estradiol, por lo cual presenta una divergencia en la presentación de la onda mas
amplía, si se compara con el uso de 17β-Estradiol (3-8 días vs 3-4 días) (Bó et al.,
2006). El uso de Benzoato de Estradiol demostró que asociado a progesterona
(inyectada y en dispositivo de liberación lenta al inicio del tratamiento) tiene mejor
efecto para suprimir el desarrollo folicular y una mejor respuesta a la

4
superestimulación. El tratamiento es similar al utilizado con 17β-Estradiol, solo
que en este caso, la dosis de estrógeno es menor (2,5 mg de Benzoato vs 5 mg
de 17β-Estradiol) (Bó et al., 2006).
FACTORES QUE AFECTAN LA RESPUESTA AL TRATAMIENTO
SUPEROVULATORIO
Los tratamientos superovulatorios y la respuesta que estos generan en la vaca
donante pueden verse afectados por diversos factores tanto intrínsecos como
extrínsecos. Dentro de los intrínsecos encontramos factores relacionados a la
raza y la respuesta de cada animal dependiendo de su estado fisiológico al
momento de iniciar el tratamiento; y a los extrínsecos, como los propios al
tratamiento, el estado nutricional, historial reproductivo, edad, estación climática,
estatus ovárico y la cantidad de tratamientos realizados en cada donante
(Mapletoft et al., 2002). Analizar estos tipos de factores favorece la toma de
decisiones, ya sea cambiando el tratamiento o descartando donantes que posean
poca o nula respuesta. Algunos de estos factores que pueden tener alto impacto
en la respuesta superovulatoria se explican a continuación:
Factores relacionados a la raza
Diversos estudios han demostrado que la dosis a utilizar en el tratamiento de
superovulación dependerá también de la raza con la cual se trabaja. Algunos
estudios sugieren que no se observan diferencias en la respuesta por raza si se
ajustan los protocolos de acuerdo la fisiología de cada especie y raza (Krininger III
et al., 2003). Chupin y colaboradores (1985) sugieren que vacas Holstein
requieren una dosis mayor de FSH si se compara con vacas de raza Charoláis.
En general se describe que las razas Bos indicus necesitan una dosis menor que
las razas Bos taurus (Nasser et al., 2011; González et al., 1990). Las dosis varían
dependiendo entre razas, por ejemplo en vacas Holstein se utilizan hasta 400 mg

5
y en vaquillonas dosis entre 260-300 mg; en razas de Carne Bos taurus la dosis a
utilizar en vacas es entre 300-320 mg y en vaquillonas de 200-260 mg; para
cruzas Bradford y Brangus en vacas se deben utilizar 260 mg y entre 133-160 mg
en vaquillonas; las vacas de raza Brahman deben ser tratadas con 200 mg de
FSH y 140 mg para las vaquillonas; y finalmente, para animales raza Neloré la
dosis recomendada para vacas es de 133-140 mg de FSH y 100 mg para
vaquillonas.
Factores relacionados al ciclo estral
En un animal sometido a un tratamiento de superovulación y que se encuentra
ciclando y con una condición corporal aceptable, se espera que tenga una
respuesta buena al tratamiento, pero estudios han demostrado que existen
diferentes tipos de donantes, las que tienen una buena respuesta a la
superovulación y que generalmente mantienen ese patrón de respuesta, y las
donantes que tienen mala respuesta pero que puede ser causada por poca
estimulación o por un inicio inadecuado del tratamiento, por lo cual no se puede
descartar una donante si no se está totalmente seguro del día del ciclo en el cual
se inicio el tratamiento. Es posible que las donantes tengan una cantidad
adecuada de folículos para responder al tratamiento, pero las gonadotrofinas no
actuaron al momento de la emergencia de la onda folicular donde folículos
dominantes y subordinados se encuentran en fase de crecimiento, sino que más
tarde en la onda folicular. Por ende, el ovario no fue estimulado antes de iniciar la
atresia folicular por parte del folículo dominante. (Chian et al., 2002; Nasser et al.,
1993; Monniaux et al., 1983). Esto indica que se debe tener claro conocimiento
del momento del ciclo estral que se encuentran las donantes. Para tener una
mejor respuesta, el ciclo estral de las donantes debería estar al inicio de una onda
folicular (primera o segunda), lo cual se logra a través del control de la dinámica
folicular.

6
Factores relacionados al control farmacológico de la dinámica folicular
Relacionados con los métodos utilizados para inducir el inicio de la onda
folicular
Para sincronizar el inicio de la onda folicular se han desarrollado métodos
mecánicos y hormonales. Dentro de los métodos mecánicos, la aspiración
folicular es la que mejor respuesta tiene, ya que aumenta significativamente los
niveles de FSH y el inicio del reclutamiento en una onda folicular comienza a las
36 horas (Bergfelt et al., 1994). La desventaja de este método es que necesita de
personal altamente entrenado y con un equipo de ultrasonografía específico para
este procedimiento (Bó et al., 2002).
Dentro de los métodos hormonales, se procura inducir la supresión de folículos
antrales con el uso de estrógenos y prostagenos, o la inducción de la ovulación
mediante el uso de GnRH ó LH, de manera de concluir la onda folicular presente y
originar una nueva onda folicular (Bó y Mapletoft, 2014).
Existen diferentes protocolos hormonales de superovulación para el ganado
bovino pero tradicionalmente se busca iniciar los mismos en la fase luteal, entre
los días 8 a 12, tiempo en el cual se han observado los mejores resultados para
superovulación, comparado con tratamientos iniciados antes o después de estos
días (Bó et al., 2002).
Relacionados con la gonadotrofina utilizada y vía de administración
Dentro de las gonadotrofinas los extractos de pituitaria son los más utilizados,
aunque también se puede encontrar comercialmente gonadotrofina coriónica
equina (eCG) y gonadotrofina menopáusica humana (hMG), la cual por su alto
costo no es utilizada actualmente en tratamientos superovulatorios en la práctica
Veterinaria (Kanitz et al., 2002). Los extractos de pituitaria contienen LH y FSH,
esta última hormona en mayor cantidad que la primera dependiendo de qué tan

7
purificado se encuentre el extracto (Herrler et al., 1991). Al tener una vida media
corta, la FSH se utiliza con intervalos de 12 horas por vía intramuscular (Monniaux
et al., 1983). La eCG posee actividad FSH y LH con una larga vida media, por lo
cual, es recomendado una única dosis por vía intramuscular. Estudios con eCG
encontraron que debido a su larga vida media, puede ocasionar presencia de
folículos anovulatorios, alteraciones en perfiles endocrinos y mala calidad
embrionaria (Mapletoft et al, 2002). La variabilidad de la respuesta superovulatoria
dentro de estos dos tratamientos es causada por la diferente concentración que
existe tanto de FSH como de LH en el producto utilizado, a causa de una posible
acción temprana en la maduración del ovocito preovulatorio por parte de la LH
(Kafi y McGowan, 1997). Se ha comprobado que el uso de FSH pura con baja
concentración de LH resulta en mejor respuesta si se compara con productos que
contienen una concentración alta o similar de FSH y LH (Mapletoft et al, 2002).
Respecto a la vía de administración, se ha demostrado que la inyección
intramuscular genera una mejor respuesta que otras vías, excepto cuando se
utiliza una única dosis aplicada vía subcutánea en la zona posterior de la
escapula, ya que esto genera resultados similares a la aplicación intramuscular en
dosis decrecientes (Bó et al., 1994).
Estos factores que afectan la respuesta a los tratamientos superovulatorios han
sido observados y colectados por diversos investigadores en distintas partes del
mundo. Sin embrago, es interesante notar que no pueden ser generalizados, ya
que estos factores pueden diferir de acuerdo a la ubicación geográfica donde se
estén realizando las superovulaciones. Es por ello que es importante caracterizar
la respuesta superovulatoria en una región, para tener parámetros para comparar
el grado de respuesta obtenidos a posteriori.

8
HIPOTESIS
Hay factores extrínsecos e intrínsecos que afectan la respuesta al tratamiento
superestimulatorio en la hembra bovina, repercutiendo en la producción de
embriones totales y de embriones de mejor calidad (grado 1) de dicho animal
luego de una superovulación.
OBJETIVO
Evaluar y determinar el efecto de factores extrínsecos e intrínsecos al animal
sobre la producción de embriones transferibles y embriones grado 1 en el ganado
bovino posterior a la aplicación de un tratamiento superovulatorio en la provincia
de Córdoba, Argentina.
MATERIALES Y MÉTODOS
Las colectas de embriones fueron realizadas en el IRAC-BIOGEN, empresa de
reproducción animal que está ubicada en la Estación General Paz S/N Paraje
Pozo del Tigre - Zona Rural CP: 5145 – Provincia de Córdoba, Argentina. Se
realizó el siguiente tratamiento: El día 0, Inserción de un dispositivo de liberación
lenta de progestágeno, una dosis de 5 mg de estradiol y 100 mg de P4, el día 4
dosis de FSH aplicadas cada 12 horas, dado por la corta vida media de la FSH, el
día 6 se aplica PGF, el día 7 se realiza el retiro del dispositivo, 24 horas después
del retiro se aplica una dosis de GnRH o LH y la correspondiente inseminación se
realiza 36 y 48 horas después de retirado el dispositivo. Finalmente realizando la
colecta de los embriones al día 15 de iniciado el tratamiento.
Los datos para el análisis fueron obtenidos de superovulaciones y colectas de
embriones desde el mes de febrero del año 2008 hasta el mes de noviembre del

9
año 2013, el acceso a dicha información en físico fue obtenido del archivo privado
del IRAC-BIOGEN. Posterior a esto se procedió a realizar el registro de los datos
mediante el programa Microsoft Excel 2010. Estos datos corresponden a 1578
colectas realizadas en 405 animales.
Al tener toda la información registrada y tabulada se realizó el análisis de los
datos, en el que las variables de estudio consideradas fueron:
Intervalo entre tratamientos (días)
Tratamiento aplicado
Dosis de Folltropin-V o Folltropin-V y eCG
Raza (agrupada en 5 categorías)
Británicas (hereford y angus)
Bonsmara
Cruzas indicas (bradford y brangus)
Continentales (limousin y simmental)
Holando
Estación del año (primavera, verano, otoño e invierno).
En el modelo estadístico, el efecto de las variables independientes sobre la
variable respuesta: cantidad de embriones transferibles (G1 + G2 + G3) o
cantidad de embriones grado 1 (G1), fue evaluado por ANOVA utilizando el
procedimiento GliMMIX de SAS (versión 9.2, SAS institute, Cary, NC).
Las variables independientes en el modelo inicial fueron días de intervalo,
tratamiento, dosis de Folltropin-V y eCG (ambas variables anidadas en
tratamiento) raza, estación y las correspondientes interacciones entre tratamiento
y raza, tratamiento y estación, y por último, estación y raza.
Las variables significativas fueron retenidas en el modelo final, el cual incluyó
tratamiento, raza, estación del año y la interacción entre tratamiento y raza. Una
variable fue considerada significativa si el valor p era menor o igual a 0.05.

10
RESULTADOS
EFECTO DE LAS VARIABLES INDEPENDIENTES SOBRE LA
CANTIDAD DE EMBRIONES TRANSFERIBLES (ET) O LA
CANTIDAD DE EMBRIONES G1 (EG1)
Tanto para la cantidad de ET o EG1, las variables no significativas en el modelo
inicial fueron días de intervalo (ET: p=0.19, EG1: p=0.18), tratamiento (ET:
p=0.63, EG1: p=0.5), y las interacciones raza por tratamiento (ET: p=0.89, EG1:
p=0.77) y raza por estación (ET: p=0.15, EG1: p=0.33). Como la interacción
tratamiento por estación fue significativa en el modelo inicial (ET: p=0.005, EG1:
p=0.0003) la variable tratamiento se retuvo en el modelo final.
En el modelo final todas las variables fueron significativas, tanto para ET como
para EG1. Estas variables fueron: raza (ET: p=0.0004, EG1: p=0.0007),
tratamiento (ET: p=0.02, EG1: p=0.0008), estación (ET: p=0.0016, EG1:
p<0.0001) y la interacción tratamiento por estación (ET: p=0.012, EG1: p=0.0006).
Como esta interacción es significativa, el efecto de tratamiento y estación ya no se
tienen en cuenta, debido a que dicha respuesta está reflejando que el efecto del
tratamiento dependerá de la estación del año.
MEDÍAS POR CUADRADOS MÍNIMOS
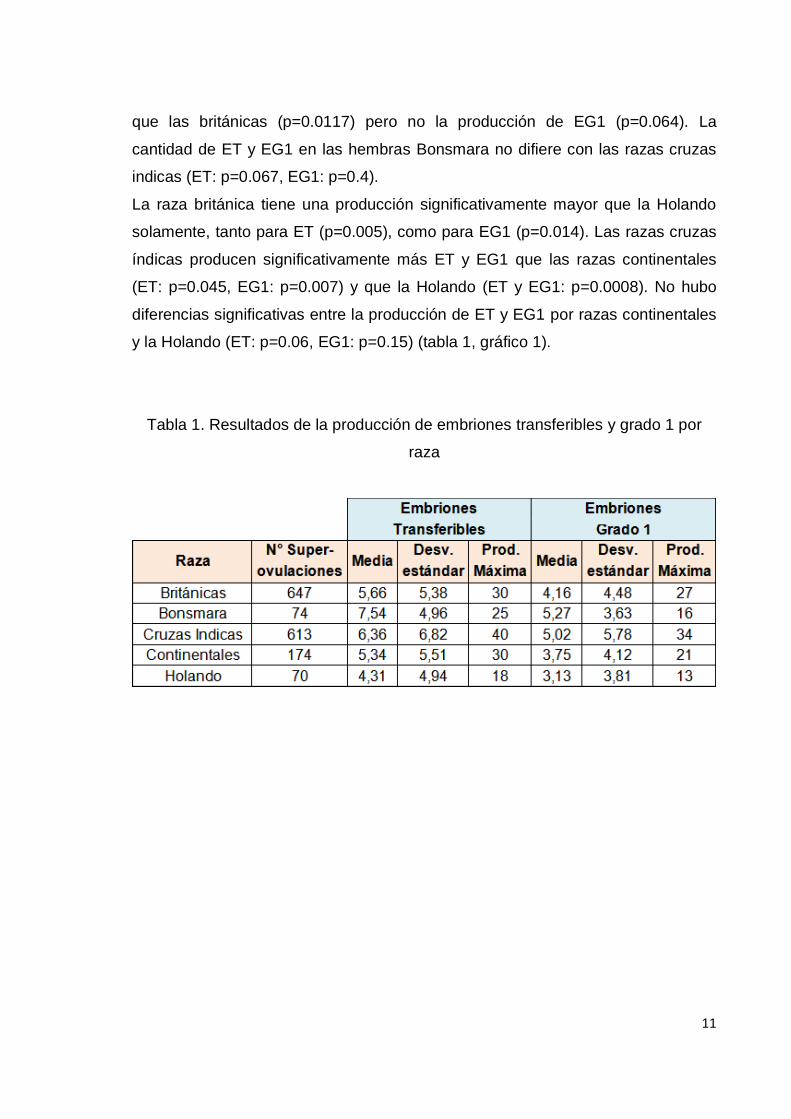
Efecto raza
Las diferencias en las medias por cuadrados mínimos por raza muestra que la
raza Bonsmara tiene una producción de ET y de EG1 significativamente mayor
que las razas continentales (ET: p=0.0035, EG1: p=0.013) y que la raza Holando
(ET: p<0.0001, EG1: 0.001). La producción de ET es significativamente mayor
11
que las británicas (p=0.0117) pero no la producción de EG1 (p=0.064). La
cantidad de ET y EG1 en las hembras Bonsmara no difiere con las razas cruzas
indicas (ET: p=0.067, EG1: p=0.4).
La raza británica tiene una producción significativamente mayor que la Holando
solamente, tanto para ET (p=0.005), como para EG1 (p=0.014). Las razas cruzas
índicas producen significativamente más ET y EG1 que las razas continentales
(ET: p=0.045, EG1: p=0.007) y que la Holando (ET y EG1: p=0.0008). No hubo
diferencias significativas entre la producción de ET y EG1 por razas continentales
y la Holando (ET: p=0.06, EG1: p=0.15) (tabla 1, gráfico 1).
Tabla 1. Resultados de la producción de embriones transferibles y grado 1 por
raza
12
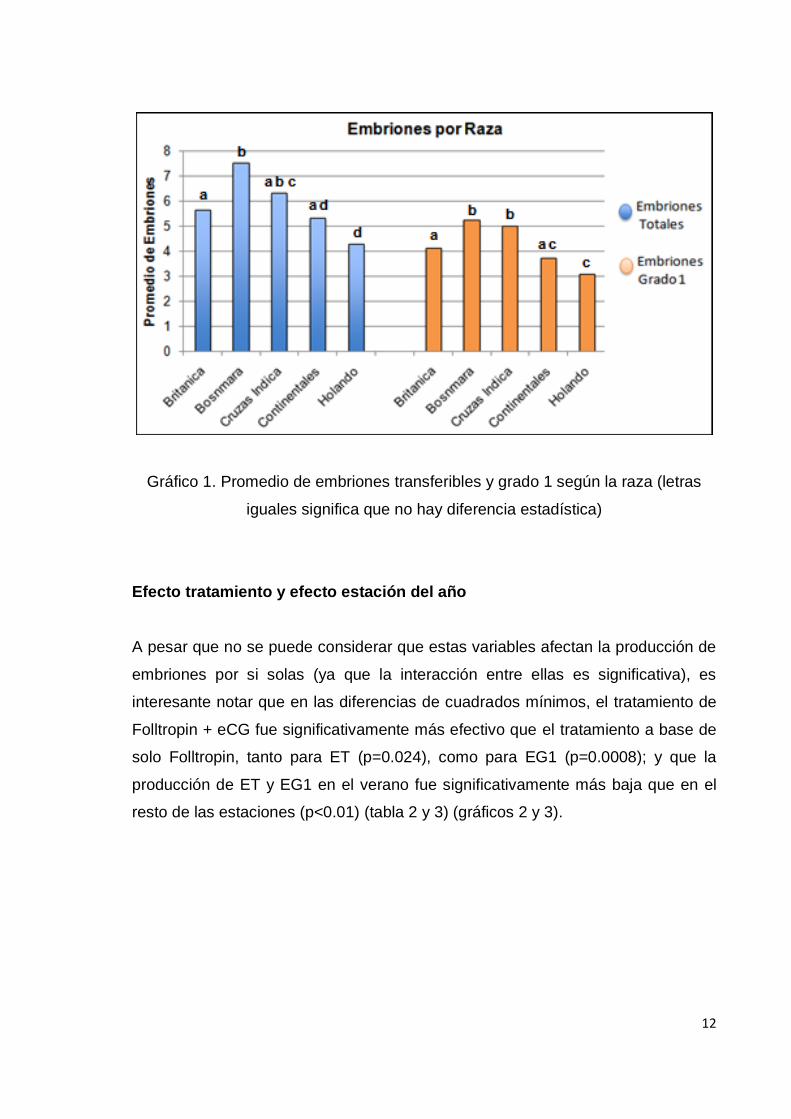
Gráfico 1. Promedio de embriones transferibles y grado 1 según la raza (letras
iguales significa que no hay diferencia estadística)
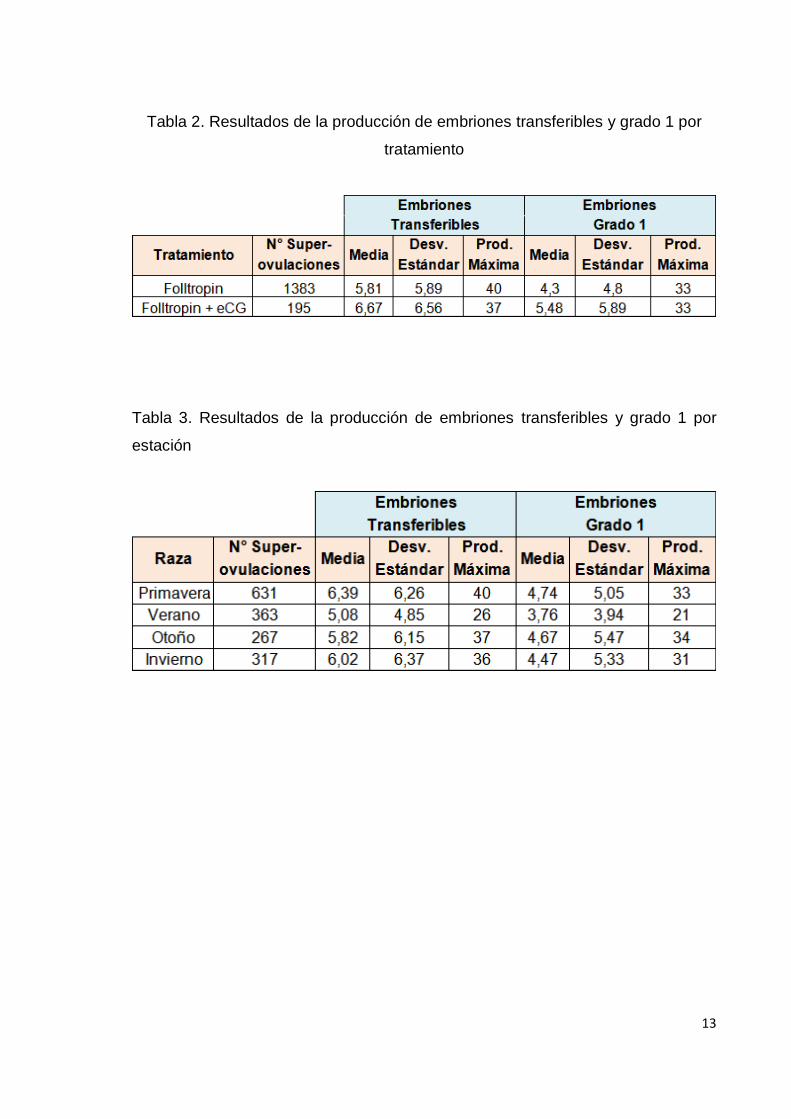
Efecto tratamiento y efecto estación del año
A pesar que no se puede considerar que estas variables afectan la producción de
embriones por si solas (ya que la interacción entre ellas es significativa), es
interesante notar que en las diferencias de cuadrados mínimos, el tratamiento de
Folltropin + eCG fue significativamente más efectivo que el tratamiento a base de
solo Folltropin, tanto para ET (p=0.024), como para EG1 (p=0.0008); y que la
producción de ET y EG1 en el verano fue significativamente más baja que en el
resto de las estaciones (p<0.01) (tabla 2 y 3) (gráficos 2 y 3).
13
Tabla 2. Resultados de la producción de embriones transferibles y grado 1 por
tratamiento
Tabla 3. Resultados de la producción de embriones transferibles y grado 1 por
estación
14
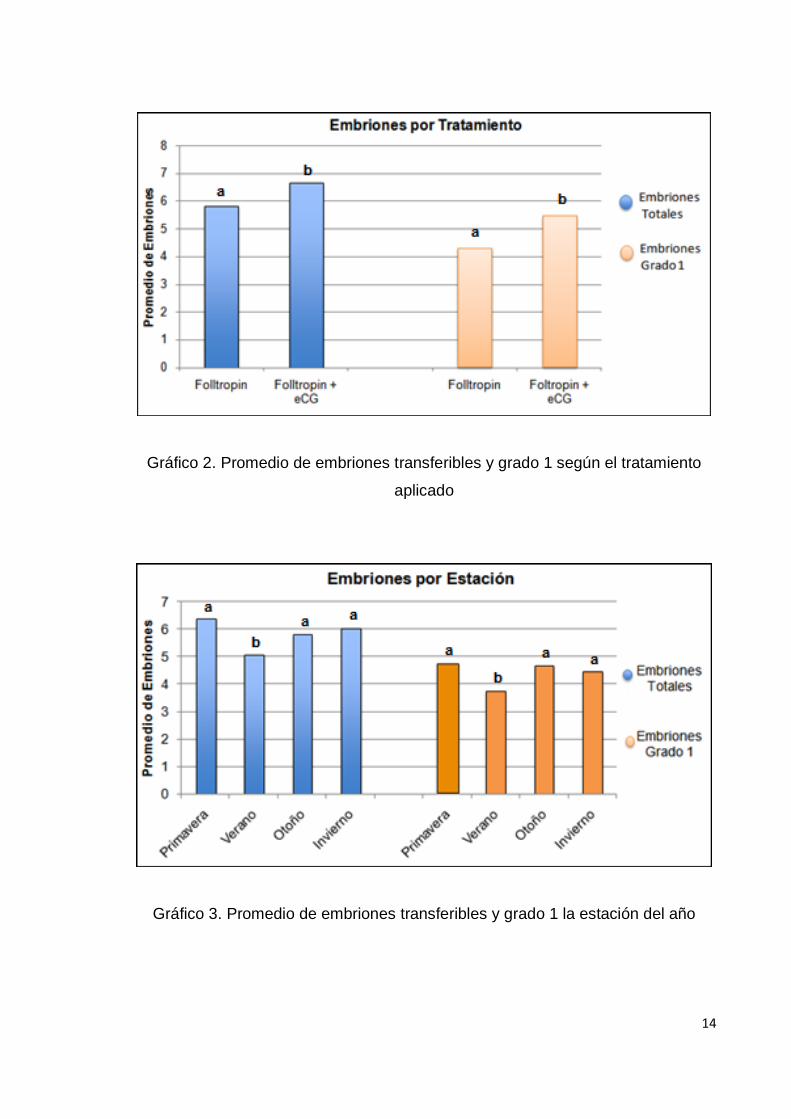
Gráfico 2. Promedio de embriones transferibles y grado 1 según el tratamiento
aplicado
Gráfico 3. Promedio de embriones transferibles y grado 1 la estación del año
15
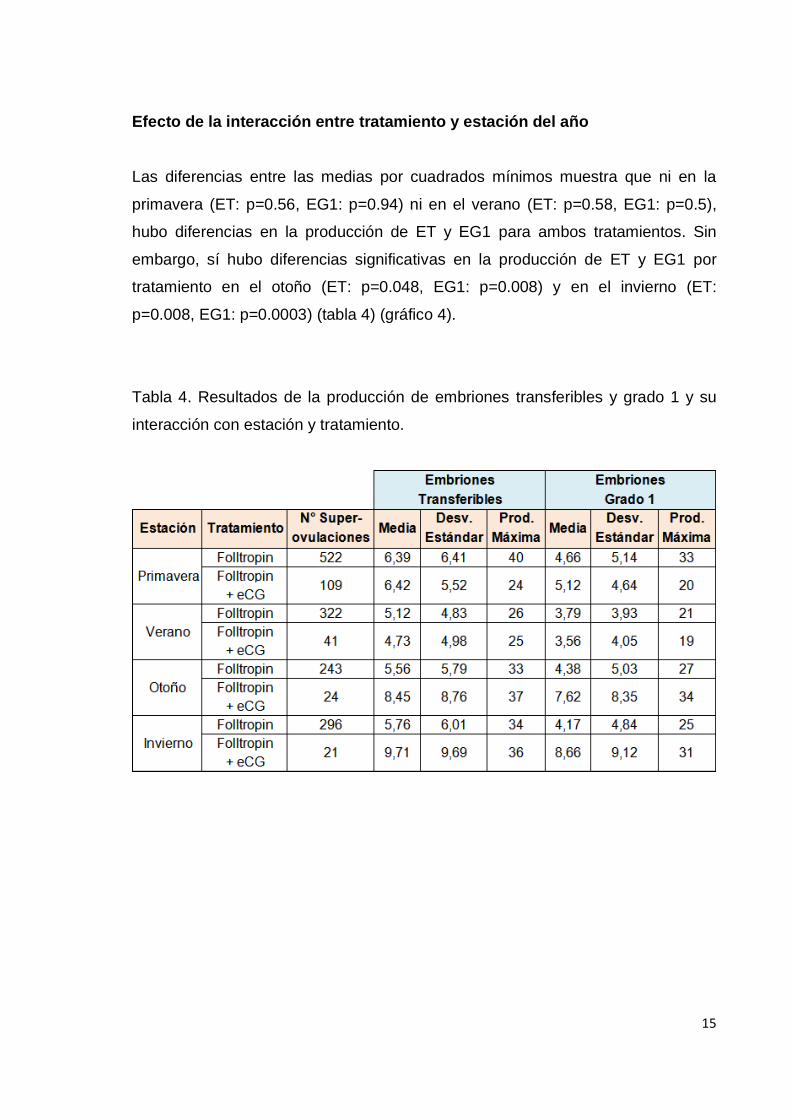
Efecto de la interacción entre tratamiento y estación del año
Las diferencias entre las medias por cuadrados mínimos muestra que ni en la
primavera (ET: p=0.56, EG1: p=0.94) ni en el verano (ET: p=0.58, EG1: p=0.5),
hubo diferencias en la producción de ET y EG1 para ambos tratamientos. Sin
embargo, sí hubo diferencias significativas en la producción de ET y EG1 por
tratamiento en el otoño (ET: p=0.048, EG1: p=0.008) y en el invierno (ET:
p=0.008, EG1: p=0.0003) (tabla 4) (gráfico 4).
Tabla 4. Resultados de la producción de embriones transferibles y grado 1 y su
interacción con estación y tratamiento.
16
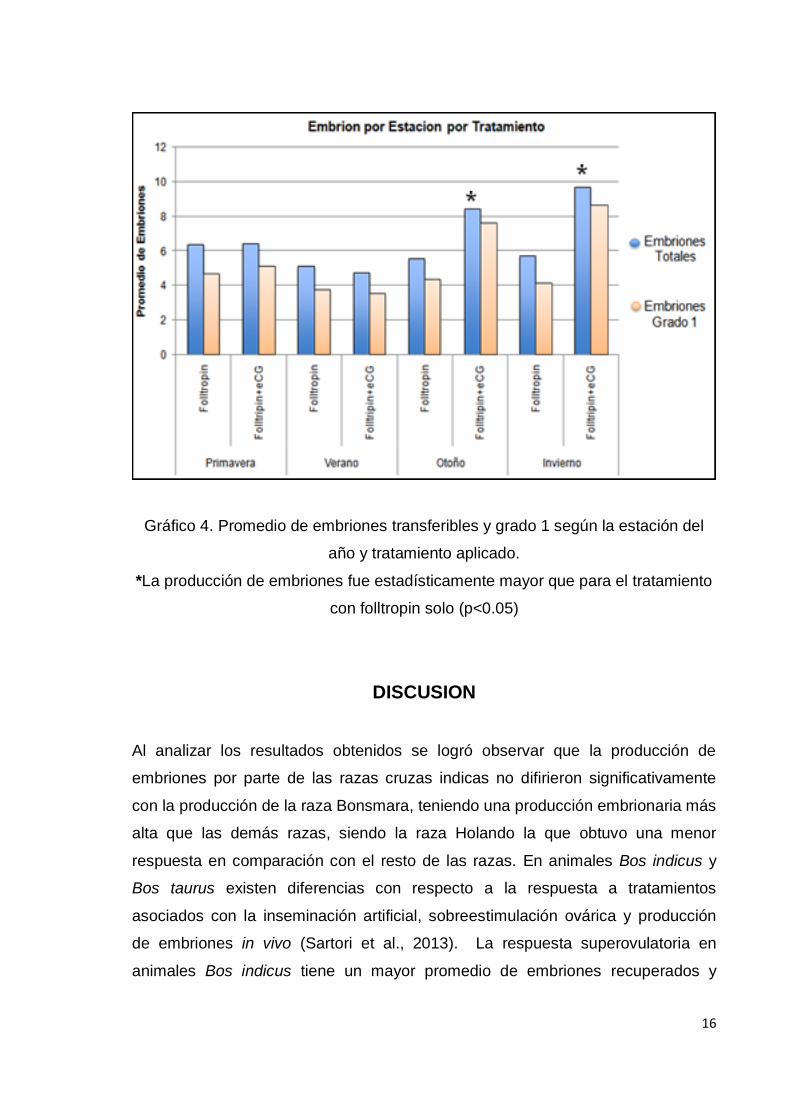
Gráfico 4. Promedio de embriones transferibles y grado 1 según la estación del
año y tratamiento aplicado.
*La producción de embriones fue estadísticamente mayor que para el tratamiento
con folltropin solo (p<0.05)
DISCUSION
Al analizar los resultados obtenidos se logró observar que la producción de
embriones por parte de las razas cruzas indicas no difirieron significativamente
con la producción de la raza Bonsmara, teniendo una producción embrionaria más
alta que las demás razas, siendo la raza Holando la que obtuvo una menor
respuesta en comparación con el resto de las razas. En animales Bos indicus y
Bos taurus existen diferencias con respecto a la respuesta a tratamientos
asociados con la inseminación artificial, sobreestimulación ovárica y producción
de embriones in vivo (Sartori et al., 2013). La respuesta superovulatoria en
animales Bos indicus tiene un mayor promedio de embriones recuperados y

17
embriones transferibles por donadora que los animales Bos taurus, en
condiciones tropicales (Córdova, 2011). También, el ganado Bos indicus responde
con un mayor número de embriones transferibles que las razas británicas (Palma,
2001).
El ganado Holando tiene una respuesta superovulatoria óptima, entre los 2 y 5
años, y hasta los 7 u 8 años sus parámetros son normales, de ahí en adelante la
producción embrionaria se ve afectada por una disminución tanto en el porcentaje
de animales que responden al tratamiento, como en el número y porcentaje de
embriones viables obtenidos (De la Fuente, 2001).
Las condiciones climáticas externas pueden afectar en cierto punto la respuesta
ovulatoria, y que dichas condiciones se presenten en verano o invierno dependerá
de la ubicación geográfica del sitio donde se lleve a cabo el programa de
transferencia de embriones (Bó y Mapletoft, 2014). El hecho de que la producción
embrionaria haya sido menor en la estación de verano señala que el estrés
calórico tiene un efecto directo sobre la respuesta superovulatoria, lo que podría
indicar un efecto negativo sobre la viabilidad del embrión. La pobre respuesta en
época cálida podría estar relacionada en principio con fallas en la ovulación
(evento que ha sido observado en animales expuestos a estrés calórico), aunque
otra de las razones podría ser que los cuerpos lúteos detectados al momento de
la colecta hayan sido en realidad folículos luteinizados (Lozano et al., 2010). A
diferencia de los resultados obtenidos, un estudio realizado en Colombia donde
también se evaluó la producción de embriones durante un periodo de seis años
no encontraron ninguna diferencia significativa sobre el número de estructuras
recuperadas en relación a la estación del año, teniendo en cuenta que es una
zona tropical y no estacional (Jiménez, 2009).
En los programas de mejoramiento genético basados en la implementación de
transferencia de embriones, un factor muy importante es la variabilidad en la
respuesta a los tratamientos superovulatorios. Una alternativa interesante para
mejorar y aumentar la respuesta superovulatoria y la cantidad de embriones
transferibles que son obtenidos por colecta directa es la utilización de un

18
tratamiento superestimulatorio en el que se combina FSH para reclutar los
folículos en la onda y eCG para estimular el crecimiento final de los folículos
reclutados. Se ha demostrado que la administración de este tratamiento tiende a
aumentar la respuesta superovulatoria, dado al reclutamiento de una mayor
cantidad de folículos que ingresan dentro del pool de folículos de 3 a 5 mm de la
onda (Bó et al., 2009).
Al incluir en el tratamiento superovulatorio la eCG se obtiene un mayor número de
Cuerpos lúteos (que son palpados al momento de realizar la colecta),
ovocitos/embriones colectados, embriones congelables (Grado 1 y 2), y
embriones transferibles (Grado 1, 2 y 3) que en tratamientos donde no se usa la
eCG. Siendo este tratamiento una muy buena alternativa incluso en animales que
tienen una historia de baja respuesta superovulatoria (Carballo et al., 2007). La
utilización de la eCG desde el punto de vista práctico ofrece una ventaja, ya que
al tener una vida media de aproximadamente 40 horas, con una sola aplicación es
suficiente para originar una apropiada respuesta ovárica. Hay que tener en cuenta
que una prolongada estimulación con eCG ocasiona un aumento en la cantidad
de folículos anovulatorios al momento de realizar la colecta de embriones (Bó et
al., 2009).
CONCLUSIONES
El análisis efectuado en datos recolectados de más de 1500 tratamientos
superovulatorios a lo largo de 6 años realizados en ganado bovino de la provincia
de Córdoba en Argentina, demuestra que factores intrínsecos y extrínsecos
afectan la producción de embriones transferibles. Dentro de los factores
intrínsecos, la raza de la donante fue la que influyo sobre la producción
embrionaria. Observamos que las razas Bonsmara y cruzas indicas son las que
mejor producción de embriones muestran, mientras que la raza Holando tuvo la
respuesta más pobre. Dentro de los factores externos, el tipo de tratamiento y la
estación del año afectaron la producción embrionaria. En general la respuesta fue

19
menor en el verano que en el resto de las estaciones del año. Con respecto al
tratamiento, la respuesta fue mayor utilizando el tratamiento de Folltropin-V
combinado con eCG, comparado con Folltropin-V solo. Sin embargo, este efecto
no fue observado durante los meses de primavera-verano, probablemente a
consecuencia del estrés calórico.
BIBLIOGRAFIA
Beal W.E. 1996. Application of knowledge about corpus luteum function in control
of estrus and ovulation in cattle. Theriogenology 45: 1399-1411.
Bergfelt D.R., Lightfoot K.C. y Adams G.P. 1994. Ovarian synchronization
following ultrasound-guided transvaginal follicle ablation in heifers. Theriogenology
42:895-907.
Bó G.A., Adams G.P., Pierson R.A., Mapletoft R.J. 1996. Effect of progesteron
plus estradiol-17β treatment on superovulatory response in beef cattle.
Theriogenology 45:897-910.
Bó G.A., Carballo D., Tribulo A., Tribulo H., Tribulo R., Mapletoft R. 2009. Nuevos
tratamientos hormonales para la superovulación de donantes de embriones
bovinos. VIII Simposio Internacional de Reproducción Animal, Córdoba, Argentina.
26 al 28 de Septiembre: CD, Sección Ponencias, 17 páginas.
Bó G.A., Baruselli P.S., Moreno D., Cutaia L., Caccia M., Tribulo R., Tribulo H.,
Mapletoft R.J. 2002. The control of follicular wave development for self-appointed
embryo transfer programs in cattle. Theriogenology 57(1): 53-72.
Bó G.A., Colazo M.G., Martinez M.F., Kastelic J., Mapletoft R.J. 2006.
Sincronización de la emergencia de la onda folicular y la ovulación en animales

20
tratados con progestágenos y diferentes esteres de estradiol. In: SIMPOSIO
INTERNACIONAL DE REPRODUÇAO ANIMAL APLICADA, p. 71-84.
Bó G.A., Hockley D.K., Nasser L.F., Mapletoft R.J. 1994. Superovulatory responde
to a single subcutaneous injection of Folltropin-V in beef cattle. Theriogenology 42:
963-975.
Bó G.A. y Mapletoft R. 2012. The evolution of improved and simplified
superovulation protocols in cattle. Reproduction, Fertility and Develompent,
24:278-283
Bó G.A. y Mapletoft R. 2014. Historical perspectives and recent Research on
superovulation in cattle Theriogenoloy 81:38-48.
Bó G.A., Pierson R.A. y Mapletoft R.J. 1991. The effect of estradiol valerate on
follicular dynamics and superovulatory response in cows with Syncro-Mate-B
implants. Theriogenology 36:169-183.
Carballo D., Tribulo A., Tribulo R., Balla E., Tribulo H., Chesta P., Cutaia L.,
Piccardi M., Reano I., Jerez J. M., Bó G. 2007. Efecto de la aplicación de eCG dos
días previos al inicio de la superestimulación en donantes de embriones con
antecedentes de baja respuesta a los tratamientos tradicionales. VII Simposio
Internacional de Reproducción Animal, Córdoba, Argentina. 29 de Junio al 1 de
Julio: CD, Sección Comunicaciones Cortas, 1 página.
Chian R., Chung J., Downey B.T., Lin S. 2002. Maturational and developmental
competence of immature oocytes retrieved from bovine ovaries at different phases
of folliculogenesis. Reproductive Biomedicie Online, 4(2): 127-132.
Chupin D., Combarnous Y. y Procureur R. 1985. Different effect of LH on FSH
induced superovulation in two breeds of cattle. Theriogenology 23:184.

21
Córdova A. 2011. Protocolos de sincronización y superovulación para
transferencia de embriones bovinos. Monografía pre-grado MVZ. Universidad de
Cuenca, Cuenca, Ecuador, pp. 128.
De la Fuente J. 2001. Transferencia de embriones en ganado bovino. En:
Reproducción Bovina, (ed.). Fundación GIRARZ, Zulia, Venezuela, pp. 375-388.
Gonzalez A., Lussier J.G., Carruthers T.D., Murphy B.D., Mapletoft R.J. 1990.
Superovulation of beef heifers with Folltropin-V: a new FSH preparation containing
reduced LH activity. Theriogenology 33:519-529.
Herrler A., Elsaesser F., Parvizi N., Niemann H. 1991. Superovulation of dairy
cows with purified FSH supplemented with defined amounts of LH. Theriogenology
35(3): 633-643.
Jiménez C. 2009. Superovulación: Estrategias, Factores Asociados y predicción
de la respuesta superovulatoria en bovinos. Rev. Med. Ved. Zoot. 56:195-214.
Kafi M. y McGowan M. 1997. Factors associated with variation in the
superovulatory response of cattle. Animal Reproduction Science, 48:137-157.
Kanitz W., Becker F., Schneider F., Kanitz E., Leiding C., Nohner H.P., Pöhland R.
2002. Superovulation in cattle: practical aspects of gonadotropin treatment and
insemination. Reprod. Nutr. Dev. 42: 586-599.
Krininger III C.E., Block J., Al-Katanani Y.M., Rivera R.M., Chase C.C., Hansen
P.J. 2003. Differences between Brahman and Holstein cows in response to estrus
synchronization, superovulation and resistance of embryos to heat shock. Anim.
Reprod. Sci. 78: 13-24.
Laster D. 1972. Dissapearance of and uptake of I125 FSH in the rat, rabbit, ewe
and cow. J Reprod Fertil. 30:407-15.

22
Lozano R. R., Asprón M. A., Vásquez, C.G., González E., Aréchiga C.F. 2010.
Effect of heat stress on embryo production in superovulated cows and on the
pregnancy rate in recipient cows. Rev. Mex. Cienc. Pecu 1(3):189-203.
Mapletoft R.J., Bennet K. y Adams G.P.. 2002. Recent advances in the
superovulation in cattle. Reprod. Nutr. Dev. 42: 601-611.
Monniaux D., Chupin D. y Saumande J. 1983. Superovulatory responses of cattle.
Theriogenology 19(1): 55-81.
Nasser L.F., Adams G.P., Bó G.A., Mapletoft R.J. 1993. Ovarian superstimulatory
response relative to follicular wave emergence in heifers. Theriogenology 40(4):
713-724.
Nasser L.F., Sá M.F., Reis E.L., Rezende C.R., Mapletoft R.J., Bó G.A., Baruselli
P.S. 2011. Exogenuos progesterone enhaces ova and embryo quality following
superstimulation of the first follicular wave in Nelore (Bos indicus) donors.
Theriogenology 76(2): 320-327.
Palma G.A. 2001. Biotecnología de la reproducción. Ediciones Instituto Nacional
de Tecnología Agropecuaria, Argentina, pp. 705.
Rathbone M.J., Kinder J.E., Fike K., Kojima F., Clopton D., Ogle C.R., Bunt C.R.
2001. Recent advances in bovine reproductive endocrinology and physiology and
their impact on drug delivery system design for the control of the estrous cycle in
cattle. Adv Drug Deliver Rev. 50: 277-320.
Sartori R., Baruselli P.S., Barros C.M., Bastos M.R. 2013. Las diferencias en la
fisiología de la reproducción entre Bos Taurus y Bos Indicus. X Simposio
Internacional de Reproducción Animal, Córdoba, Argentina. 4 al 6 de Julio: CD,
Sección Conferencias Plenarias, 12 páginas.