estudo de interação bioquímica da metaloproteases de
TRANSCRIPT

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE CIÊNCIAS DA SAÚDE
DEPARTAMENTO DE FARMÁCIA
NILTON JOSÉ DA SILVA JÚNIOR
Estudo de interação bioquímica da metaloproteases de
Leishmania spp. e Trypanosoma cruzi com substratos proteicos
NATAL/RN
FEVEREIRO, 2021

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE CIÊNCIAS DA SAÚDE
DEPARTAMENTO DE FARMÁCIA
NILTON JOSÉ DA SILVA JÚNIOR
Estudo de interação bioquímica da metaloproteases de
Leishmania spp. e Trypanosoma cruzi com substratos proteicos
Trabalho de Conclusão de Curso apresentado ao curso de
Graduação em Farmácia da Universidade Federal do Rio
Grande do Norte como requisito parcial à obtenção do título
de Bacharel em Farmácia.
Orientador: Prof. Doutor Marcelo Sousa Silva
Co-orientadora: Dra. Cláudia Gonçalves Moreno
NATAL/RN
FEVEREIRO, 2021

Universidade Federal do Rio Grande do Norte - UFRN
Sistema de Bibliotecas - SISBI
Catalogação de Publicação na Fonte. UFRN - Biblioteca Setorial do Centro Ciências da Saúde - CCS
Silva Júnior, Nilton Jose da.
Estudo de interação bioquímica da metaloproteases de Leishmania spp. e Trypanosoma cruzi com substratos proteicos /
Nilton Jose da Silva Júnior. - 2021. 59f.: il.
Universidade Federal do Rio Grande do Norte, Centro de
Ciências da Saúde, Departamento de Farmácia. Orientador: Marcelo Sousa Silva.
Coorientador: Cláudia Gonçalves Moreno.
1. Tripanossomose - TCC. 2. Metaloproteases - TCC. 3. Gp63 -
TCC. 4. Trypanosoma cruzi - TCC. 5. Leishmania spp. - TCC. 6.
Albumina - TCC. I. Silva, Marcelo Sousa. II. Moreno, Cláudia
Gonçalves. III. Título.
RN/UF/BS-CCS CDU 616.937
Elaborado por ANA CRISTINA DA SILVA LOPES - CRB-15/263

NILTON JOSÉ DA SILVA JÚNIOR
Estudo de interação bioquímica da metaloproteases de
Leishmania spp. e Trypanosoma cruzi com substratos proteicos
Trabalho de Conclusão de Curso apresentado ao curso de
Graduação em Farmácia da Universidade Federal do Rio
Grande do Norte como requisito parcial à obtenção do título
de Bacharel em Farmácia.
BANCA EXAMINADORA:
Prof. Dr. Marcelo Sousa Silva – Orientador
Universidade Federal do Rio Grande do Norte
Prof. – Membro da banca
Universidade Federal do Rio Grande do Norte
Prof. – Membro da banca
Universidade Federal do Rio Grande do Norte
Conceito:
APROVADO EM: / /

AGRADECIMENTOS
Primeiramente, agradeço à Deus pela proteção e força nos momentos difíceis e firmes, e
por me proporcionar a experiência de estudar na Universidade Federal do Rio Grande do Norte,
local onde tive grande aprendizado como pessoa e futuro profissional.
Obrigado aos meus pais e familiares pelo constante carinho, apoio e estimulo ao estudo,
em especial às minhas tias, irmãos e primos.
Obrigado aos meus amigos que me proporcionaram momentos de descontração e conselhos
no decorrer da construção desse trabalho, em especial Mathews pela ajuda durante a criação desse
projeto.
Obrigado para minha namorada Janaína pela preciosa ajuda na organização desse trabalho,
além de forças, afeto e conselhos no decorrer da construção desse trabalho.
Obrigado a todos os colegas do Laboratório de Imunoparasitologia que de alguma forma
contribuíram para conclusão desse trabalho.
Minha eterna gratidão a minha co-orientadora Cláudia pelas preciosas correções, sugestões
de escrita, amizade e companhia agradável. Agradeço por cada palavra que me serviram de
estimulo e inspiração.
Agradeço ao professor Marcelo, pela amizade e ensinamentos que foram essenciais para
minha formação. Obrigado a professora doutora Silvia e doutorando Johny que são exemplos de
profissionais e aceitaram participar da banca examinadora.

RESUMO
Os parasitas do grupo dos Tripanosomatídeos são responsáveis por importantes doenças
negligenciadas de importância humana e animal. Apesar de inúmeros estudos, a Leishmaniose e a
Doença de Chagas representam um importante problema de saúde pública, principalmente no
Brasil. Atualmente, não existe vacinas para o controle das doenças parasitárias humanas e ainda
existe uma grande necessidade de novos fármacos ou novas formulações dos fármacos, seguros,
com maior eficácia e menos tóxicos. As metaloproteases são moléculas de superfície, presente nos
tripanossomatídeos sendo fundamentais para interação parasita-hospedeiro, alterando a
homeostasia da matriz extracelular. Diante disso, o objetivo deste estudo é a caracterização da
atividade proteolítica de metaloproteases de Tripanosomatídeos de importância médica,
causadoras de doenças, tais como a Doença de Chagas e as leishmanioses frente a substratos. A
partir do cultivo axênico foram obtidos os extratos totais dos parasitas, do qual foi feito a
quantificação, selecionando extratos com as melhores concentrações. Foram avaliados os perfis
proteicos, bem como a atividade enzimática e atividade enzimática in vitro de diferentes extratos
brutos dos Tripanosomatídeos. Através de ensaios por SDS-PAGE foi observado clara distinção
entre os perfis dos parasitas. Através de ensaios de zimografia foi confirmado o perfil proteolítico
das proteínas do extrato bruto de Leishmania spp. sobre o substrato gelatina e albumina. Em
relação ao Trypanosoma cruzi, um padrão proteolítico semelhante com o descrito na literatura foi
encontrado para a cruzipaína. Essas proteases foram submetidas a ensaios de degradação frente a
substratos de albumina sérica, em diferentes intervalos de tempo, usando o método de SDS-PAGE,
onde foi possível verificar a afinidade o perfil de atividade proteolítica das espécies selecionadas
para esse estudo.
Palavras-chaves: Tripanossomatídeos; Metaloproteases; gp63; Gelatina; Albumina;
Zimografia.

ABSTRACT
Trypanosomatids parasites are responsible for neglected diseases in humans and with animal
importance. Despite numerous studies, Leishmaniasis and Chagas' disease represent an important
public health problem, especially in Brazil. Currently, there are no vaccines for the control of
human parasitic diseases of these diseases and there is still a great need for new drugs or new drug
formulations, which are safe, more effective, and less toxic. Metalloproteinases are surface
molecules, present in trypanosomatids and are fundamental for parasite-host interaction, changing
the homeostasis of the extracellular matrix. Therefore, the aim of this study is to characterize the
proteolytic activity of metalloproteases of trypanosomatids of medical importance, causing
diseases, such as Chagas' disease and leishmaniasis against substrates. From the axenic cultivation,
the total extracts of the parasites were chosen, from which the quantification was made, extracts
with the best ones were selected. Protein profiles, as well as enzymatic activity and enzymatic
activity in vitro of different crude extracts of the Trypanosomatids, were evaluated. In all assays
performed by SDS-PAGE, a clear distinction was observed between the parasite profiles. Through
zymography tests, the proteolytic profile of the proteins in the crude extract of Leishmania spp. on
the gelatin and albumin substrate. Regarding Trypanosoma cruzi, a proteolytic pattern similar to
that described in the literature was found for cruzipain. These proteases were subjected to
degradation tests against serum albumin substrates, at different time intervals, using the SDS-
PAGE method, where it was possible to verify the affinity of the proteolytic activity profile of the
species selected for this study.
Keywords: Trypanosomatids; Metalloproteinase; gp63; Albumin, Gelatin; Zymography;

LISTA DE SIGLAS E ABREVIATURAS
cDNA - DNA complementar
CR1- Recetor do complemento 1
CR3 - Recetor do complemento 3
C3b - Componente 3b do sistema
complemento
DC – Doença de Chagas
DNA - Ácido desoxirribonucleico
DTN - Doenças Tropicais
Negligenciadas
DTU - Unidade discreta de tipagem
EDTA - Ácido etilenodiamino tetra-
acético
ELISA - Enzyme-Linked
Immunosorbent Assay
SFB – Soro fetal bovino
GPI - Glicosilfosfatidilinositol
gp63 - Glicoproteína de 63kDa
His – Histidina
iC3b - C3b inativado
IFI - Imunofluorescência indireta
IFN-𝛾 - Interferão gama
IgM - Imunoglobulina M
IL-12 - Interleucina-12
IL-4 – Interleucina-4
JAK- Cinase janus
LIT - Liver Infusion Tryptose
LPG – Lipofosfoglicano
LV – Leishmaniose visceral
MAPK - Cinase ativada por
mitogénios
MARCKS – Substrato de proteinase
C rico em alanina miristoilada
MMP – Metaloprotease de matriz de
mamífero
MHC - Complexo Principal de
Histocompatibilidade
MRP – Proteínas relacionadas com
MARCKS
MSP – Pincipal protease de
superfície
NK - Natural Killer
NO – Óxido nítrico
OMS - Organização Mundial de
Saúde
PBS – Tampão fosfato salino
PCR - Proteína c-reativa
PDB – Protein Data Bank
PKC – Substrato da proteína cinase
C
RNA - Ácido ribonucleico
RPMI - Roswell Park Memorial
Institute
SDS - Dodecil sulfato de sódio
Sh – Soro humano
STAT-1 - Transdutor de sinal e
ativador de transcrição-1
TEMED - tetrametiletilenodiamina
Th - T helper
TNF-α - Fator de necrose tumoral
alfa

12
LISTA DE FIGURAS
Figura 1.Prevalência das Doenças Tropicais Negligenciadas por país. Adaptado de MAWSON,
2017. .............................................................................................................................................. 13
Figura 2.Ciclo Biológico Leishmania spp. (Autoria própria). ..................................................... 16
Figura 3.Ciclo Biológico de Trypanosoma cruzi (Autoria própria). ............................................ 17
Figura 4.Estrutura tridimensional da gp63. .................................................................................. 25
Figura 5. Eletroforese em gel de Poliacrilamida (SDS-PAGE) ................................................... 38
Figura 6.Curva padrão, utilizando a proteína Albumina sérica bovina (BSA) ............................ 41
Figura 7.Perfil proteico total dos extratos de Leishmania e as respectivas concentrações por poço,
em géis corados com azul de Coomassie.) .................................................................................... 42
Figura 8.Perfil proteico total dos extratos das cepas de Trypanosoma cruzi e as respectivas
concentrações por poço, em géis corados com azul de Coomassie.) ............................................ 43
Figura 9.Perfil enzimático, em gel de poliacrilamida acoplado com o susbstrato gelatina, na
presença de extratos de Leishmania spp e T. cruzi........................................................................ 44
Figura 10.Perfil enzimático, em gel de albumina, de extratos de Leishmania spp e T. cruzi ...... 46
Figura 11.Perfil de proteico, em gel de poliacrilamida (10%), de substrato Albumina sérica
(BSA) ............................................................................................................................................ 47
Figura 12.Perfil de degradação in vitro da albumina na presença do extrato bruto de Leishmania
spp. e T. cruzi, em gel de poliacrilamida durante 24 horas (A), 48 horas (B) e 72 horas (C). ...... 48

Figura 13.Perfil de degradação in vitro da albumina na presença do extrato bruto de Leishmania
spp. e T. cruzi, em gel de poliacrilamida durante 24 horas (D), 48 horas (E) e 72 horas (F)........ 49

Sumário
1. INTRODUÇÃO ................................................................................................... 12
1.1. Doenças tropicais negligenciadas .................................................................... 12
1.2. Aspectos biológicos da Leishmanioses e Doença de Chagas .......................... 14
1.2.1. Leishmania spp. - morfologia do parasita, ciclo biológico......................... 14
1.2.2. Trypanosoma cruzi - morfologia do parasita, ciclo biológico .................... 16
1.3. Aspectos e aspectos clínicos da Leishmaniose e da Doença de Chagas ....... 19
1.4. Diagnóstico de Leishmaniose e da Doença de Chagas .................................... 20
1.5. Terapêutica atual da Leishmaniose e da Doença de Chagas ............................ 22
1.6. Metaloprotease de superfície de Leishmania spp. e Trypanosoma cruzi.........24
1.5.1. gp63 de Leishmania spp ............................................................................. 27
1.5.2. gp63 de Trypanosoma cruzi ....................................................................... 28
1.6. Principais metodologias utilizadas para a identificação de metaloprotease ..... 29
1.6.1. Zimografia para estudos biológicos ............................................................ 29
2. Justificativa .......................................................................................................... 32
3. OBJETIVOS ........................................................................................................ 33
3.1. Objetivo geral ................................................................................................... 33
3.2. Objetivos específicos ....................................................................................... 33
4. Materiais e Métodos ............................................................................................ 34
4.1. Substratos ......................................................................................................... 34
4.2. Leishmania spp. e Trypanosoma cruzi ............................................................. 34
4.3. Cultura axênico dos parasitas ........................................................................... 36
4.3.1. Cultura de Leishmania spp. ........................................................................ 36
4.3.2. Cultura de Trypanosoma cruzi ................................................................... 36
4.4. Preparação dos extratos .................................................................................... 36
4.5. Determinação da concentração proteica dos extratos brutos ........................... 37

4.6. Identificação das metaloproteinases dos extratos de Leishmania spp. e
Trypanosoma cruzi...................................................................................................................... 37
4.6.1. Caracterização do perfil proteico por SDS-PAGE ..................................... 37
4.6.2. Determinação da atividade enzimática por zimograma .............................. 39
4.7. Otimização e determinação da atividade proteolítica in vitro de extrato bruto de
Leishmania spp e T. cruzi................................................................................................ 39
5. RESULTADOS E DISCUSSÃO ......................................................................... 41
5.1. Perfil proteico de Leishmania e T. cruzi .......................................................... 41
5.2. Atividade proteolítica de extratos Leishmania spp. e T. cruzi ......................... 44
5.3. Atividade proteolítica in vitro de extrato bruto de Leishmania spp. T. cruzi e
sobre o substrato de Albumina .................................................................................................... 47
6. CONCLUSÃO E PERSPECTIVAS .................................................................... 51
7. REFERÊNCIAS BIBLIOGRÁFICAS ................................................................ 53

12
1. INTRODUÇÃO
1.1. Doenças tropicais negligenciadas
A Organização Mundial da Saúde (OMS) fundada, no século XX, mantém a atenção e o
compromisso diante da problemática causada pelas Doenças Tropicais Negligenciadas (DTN).
Solenizou-se, em 30 de janeiro de 2020, o Dia Mundial das Doenças Tropicais Negligenciadas,
onde foi relembrado as conquistas e reafirmado o compromisso de combate à essa problemática
(HOTEZ et al., 2020; MITRA; MAWSON, 2017).
As DTN são o conjunto de doenças causadas por agentes infecto-parasitários que
produzem importante dano físico, cognitivo e socioeconômico em homens, mulheres e crianças.
Estas doenças afetam principalmente, mas não exclusivamente, populações pobres de países
tropicais e subtropicais (Dias et al., 2013), com renda de US$ 1,90 por dia, segundo o Banco
Mundial (HOTEZ et al., 2020; MITRA; MAWSON, 2017).
Atualmente, a OMS identifica 20 condições como DTN distribuídas em 148 países, onde
muito desses países são endêmicos para mais de uma doença negligenciada, como por exemplo, o
Brasil (Fig.1). As mais prevalentes são as infecções por helmintos transmitidos pelo solo
(ascaridíase, infecção por ancilostomíase, tricuríase e estrongiloidíase), seguidas de filariose,
esquistossomose, sarna, leishmaniose, Doença de Chagas e dengue (Aguiar-Santos et al., 2013;
Hotez et al., 2020).
As DTN podem ser classificadas em três categorias, baseado na emergência, controle e
disponibilidade de medicamentos: (1) não está sob controle como a Dengue, Doença do Sono e
Leishmaniose; (2) estratégia de controle disponível como Esquistossomose e Tuberculose; (3) tem
estratégia de combate eficaz como a Hanseníase, Doença de Chagas e Filariose (MALAFAIA,
2009).
A figura 1 mostra a prevalência das DTN nos países, tendo grande impacto em locais
tropicais e subtropicais. O combate as DTN, embora urgente, revela-se uma tarefa difícil pela falta
de investimentos em projetos de desenvolvimento e pesquisa de novos fármacos. Dos 756
fármacos aprovados entre 2000 e 2011, apenas 29 foram destinados exclusivamente para as DTN,
correspondendo em 3,8% (DIAS et al., 2013). Isso mostra o grande desinteresse da indústria
farmacêutica diante dessa problemática.

13
As opções terapêuticas para as DTN, enfrentam outras limitações devido à baixa eficácia,
elevada toxicidade e surgimento de cepas resistentes, além do impacto no perfil de morbidade,
configurando o desafio a saúde pública dos países afetados (Aguiar-Santos et al., 2013; Dias et al.,
2013).
Figura 1.Prevalência das Doenças Tropicais Negligenciadas por país. Adaptado de MAWSON, 2017.
A ordem Kinetoplastida possui protozoários pertencentes à família Trypanosomatidae.
Essa família tem em comum protozoários que possuem junto ao corpo basal do flagelo, o ácido
desoxirribonucleico (DNA, DeoxyriboNucleic Acid) mitocondrial altamente condensado,
designado por cinetoplasto (LISBOA; AVEIRO, 2008; MA et al., 2011). Outro ponto em comum
é o fato de alguns gêneros pertencentes a esta família, Trypanosoma, Leishmania, terem um ciclo
heteroxênico, intercalando seu ciclo de vida entre hospedeiros invertebrados e vertebrados
(LISBOA; AVEIRO, 2008).

14
1.2. Aspectos biológicos da leishmanioses e Doença de Chagas
1.2.1. Leishmania spp. - morfologia do parasita e ciclo biológico
As leishmanioses agrupam um complexo infeccioso de doenças tropicais e subtropicais
causadas por diferentes espécies do parasito intracelular do gênero Leishmania, sendo transmitidas
ara humanos pela picada da fêmea do mosquito do gênero Phlebotomus e Lutzomyia, com
prevalência em países da Europa, Norte da África, Oriente Médio, Ásia e parte da América do Sul
(ARENAS et al., 2017). Leishmania são protozoários da ordem Kinetoplastida, pertencente à
família Tripanosomidae, do gênero Leishmania e subgênero Leishmania ou Viannia, sendo o
agente causador da leishmaniose (PINHEIRO; LUZ; FRANCO, 2008).
É uma doença endêmica em 98 países, sendo que 68 países endêmicos para leishmaniose
visceral e leishmaniose tegumentar, ocorrendo 700.000 a 1 milhão de novos casos anuais
(REBELLO et al., 2019). Ocorre de 50 a 90.000 novos casos anuais de leishmaniose visceral e
600.000 a 1 milhão de novos casos anuais de leishmaniose tegumentar, causando cerca de 70.000
mortes anuais (BURZA; CROFT; BOELAERT, 2018).
A OMS, caracteriza as leishmanioses ( tegumentar e visceral) como uma das sete doença
tropicais mais importantes, com ampla distribuição geográfica e espectro de manifestações clinicas
com desfecho geralmente fatal (ARENAS et al., 2017). As infecções por Leishmania spp têm
distribuição mundial e se limita às áreas de distribuição natural do mosquito flebotomíneo – inseto
vetor (SUNDAR S, 2002). As consequências da globalização como desmatamentos, urbanização
e as constantes migrações por diversos motivos, ajudaram a disseminar a doença e criar novos
reservatórios de Leishmania, incluindo os imunossuprimidos infectados com HIV (DA SILVA-
LÓPEZ, 2010; SUNDAR S, 2002).
Devido a diversidade de espécies de Leishmania, as manifestações clinicas são variadas
e vão desde lesões cutâneas auto curativas até doença visceral, dependendo das características do
parasita, vetor biológico e resposta imune do hospedeiro (CHAGAS et al., 2016). A leishmaniose
pode apresentar duas formas clinicas, a forma tegumentar e visceral. A leishmaniose tegumentar
pode se subdividir em cutânea, mucocutânea e cutânea difusa, sendo causada por pelo menos 13
espécies do protozoário Leishmania, abrangendo manifestações clinicas diferentes (GOTO;
LINDOSO, 2010). A leishmaniose tegumentar cutânea é a mais frequente, e é classicamente
dividida nas formas do Novo e do Velho Mundo.

15
No Brasil, as principais espécies que causam a leishmaniose tegumentar são
Leishmania amazonensis e Leishmania guyanensis. No Velho Mundo a leishmaniose cutânea é
causada pela Leishmania major. A leishmaniose cutânea pode evoluir para cutânea difusa
começando com pápulas localizadas não ulceradas até nódulos cutâneos na face e extremidade,
causada pela L. amazonensis (DA SILVA- LÓPEZ, 2010). A leishmaniose visceral (LV)
antroponótica é uma doença crônica fatal, com letalidade de 10% se não tratada, é prevalente no
Oriente Médio e causada pela Leishmania donavani, podendo afetar todas as faixas etárias de
ambos os gêneros, não tendo reservatório animal conhecido (BURZA; CROFT; BOELAERT,
2018). A LV zoonótica é causada por Leishmania chagasi (infantum) e acomete o homem (na
maior parte das áreas endêmicas 80% dos casos registrados ocorrem em crianças com menos de
10 anos), imunocompetentes e imunossuprimidos. Afeta canídeos sendo o cão o reservatório
doméstico e a raposa como reservatório silvestre (BURZA; CROFT; BOELAERT, 2018;
GONTIJO; MELO, 2004). Frequente na Índia, Sudão, Quênia e Nepal, a leishmaniose dérmica
Pós-Kalazar afeta crianças e adultos jovens, pode ocorrer 6 meses após o tratamento de LV em
até 20% dos indivíduos tratados (BURZA; CROFT; BOELAERT, 2018).
O ciclo biológico de Leishmania spp. é heteroxênico e, morfologicamente, o protozoário
apresenta a forma amastigota, sendo imóvel e intracelular (Figura 2). A amastigota de
Leishmania spp. sobrevive dentro do fagolisossomo do macrófago apesar do meio ácido. A forma
promastigota é móvel, infectante, com cinetoplasto anterior ao núcleo, encontra-se presente
dentro do tubo digestivo do flebotomíneo (CHAGAS et al., 2016).
Como demostra na figura 2, no decorrer do ciclo biológico,as formas promastigotas
metacíclicas, que se localizam nas vulvas faríngeas do vetor, são transmitidas pela picada do
mosquito fêmea flebotomíneo infectado (CHAGAS et al., 2016). Em resposta a inflamação no
local da picada, são recrutados neutrófilos e estes sinalizam para células dendriticas e
macrófagos periféricos iniciarem a fagocitose. Dentro do macrófago as formas promastigotas se
transformam em amastigotas e essas se dividem por fissão binaria até o rompimento do
macrófago, novos macrófagos são recrutados e são infectados (BURZA; CROFT; BOELAERT,
2018). Quando a fêmea do mosquito realizar o repasto sanguíneo no indivíduo infectado, ingere
os macrófagos infectados. Ainda na figura 2, no vetor invertebrado, as formas amastigotas são
liberados no intestino médio ou posterior, onde se transformam em promastigota procíclica e
depois metacíclica por divisão binaria, que migram para as vulvas faringeas do mosquito
(LISBOA; AVEIRO, 2008).

16
Figura 2.Ciclo Biológico Leishmania spp. (Autoria própria). A imagem foi criada no Biorender.com.
1.2.2. Trypanosoma cruzi - morfologia do parasita, ciclo biológico
Em 2009, foi comemorado 100 anos de descoberta da Doença de Chagas. Este fato é um
importante marco na história, quando em 1909, Carlos Justiniano Ribeiro Chagas, através de
estudos minuciosos anunciou a comunidade cientifica a detecção em humanos, do protozoário
Trypanosoma cruzi (T. cruzi), agente causador da doença, assim como o ciclo biológico e a
patologia (MALAFAIA; DE LIMA RODRIGUES, 2010). A doença é considerada uma zoonose
e apresenta uma larga distribuição no continente americano (JANSEN; XAVIER; ROQUE, 2018;
MARTINEZ; ROMANO; ENGMAN, 2020). Atualmente, a doença de Chagas (DC) é classificada
como enfermidade negligenciada e estima-se que aproximadamente 8 milhões de pessoas estejam
infectadas e contabiliza 300.000 casos a cada ano (SIMÕES et al., 2018).
A via clássica da transmissão é a vetorial pelo vetor triatomíneo (Figura 3). Outras
formas de transmissão são, transfusão de sangue de doadores infectados e transmissão vertical de
mães infectadas para seus recém-nascidos (POULOSE, 2016). Contudo, a epidemiologia do T.
cruzi vem mudando, pois atualmente a maioria dos casos ou surtos da doença ocorre, em
cenários
17
regionais, via transmissão oral pela ingestão de alimentos contaminados com as formas
metacíclicas (POULOSE, 2016; YAO, 2010). Presentemente, devido à urbanização e globalização
pelo processo migratório das pessoas infectadas, a DC emerge, tanto nos países endêmicos quanto
não endêmicos, contribuindo assim, para aumentar a complexidade dos cenários epidemiológicos
e patogênese. Adicionalmente, eventos de reativação da doença de Chagas associados a
imunodeficiências têm se tornado desafios emergentes para os sistemas de saúde (DIAS et al.,
2016a; JANSEN; XAVIER; ROQUE, 2018).
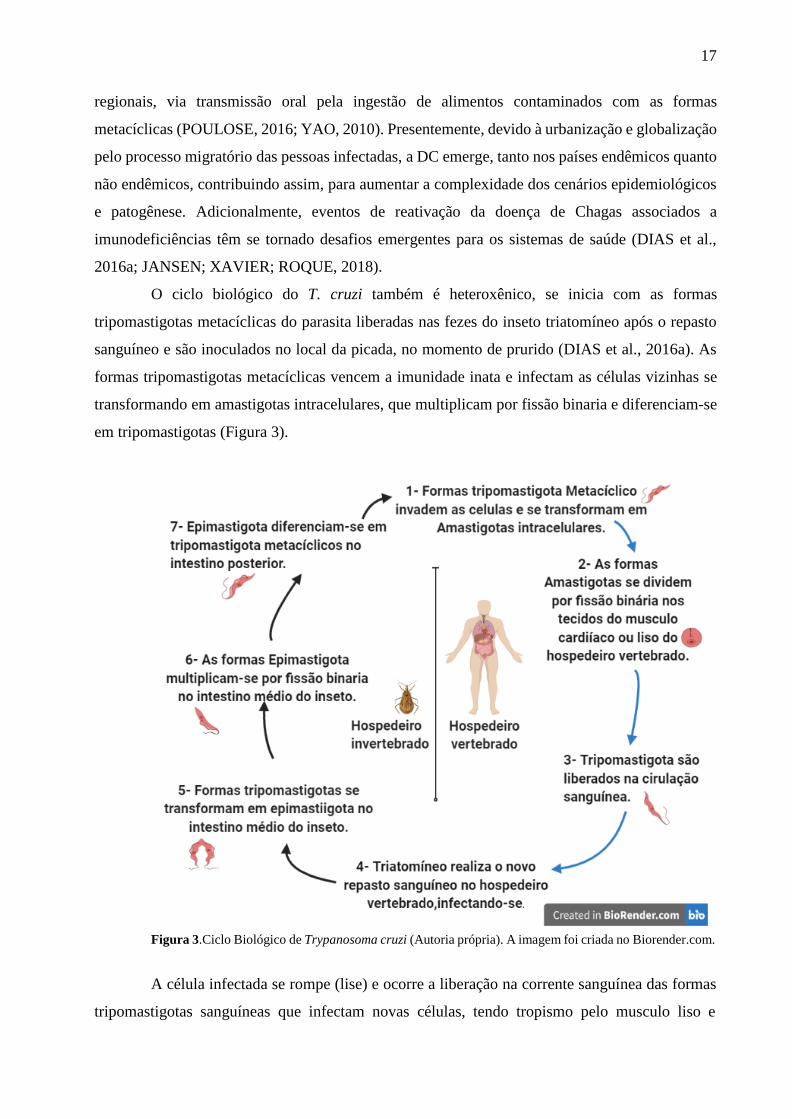
O ciclo biológico do T. cruzi também é heteroxênico, se inicia com as formas
tripomastigotas metacíclicas do parasita liberadas nas fezes do inseto triatomíneo após o repasto
sanguíneo e são inoculados no local da picada, no momento de prurido (DIAS et al., 2016a). As
formas tripomastigotas metacíclicas vencem a imunidade inata e infectam as células vizinhas se
transformando em amastigotas intracelulares, que multiplicam por fissão binaria e diferenciam-se
em tripomastigotas (Figura 3).
Figura 3.Ciclo Biológico de Trypanosoma cruzi (Autoria própria). A imagem foi criada no Biorender.com.
A célula infectada se rompe (lise) e ocorre a liberação na corrente sanguínea das formas
tripomastigotas sanguíneas que infectam novas células, tendo tropismo pelo musculo liso e

18
cardíaco (Figura 3). O inseto vetor ao realizar o repasto sanguíneo no indivíduo infectado,
ingerem as formas tripomastigota metacíclico que vão para o intestino médio do inseto,
transformando-se em epimastigota. Como demostrado na figura 3, epimastigotas migram para o
intestino anterior do inseto onde se transformam em tripomastigotas metacíclico que serão
eliminados nas fezes, reiniciando o ciclo, durante um novo repasto sanguíneo (LISBOA;
AVEIRO, 2008; PÉREZ- MOLINA; MOLINA, 2018).
T. cruzi pertence a ordem Kinetoplastida, da família Tripanosomatidae (Protozoa,
Sarcomastigophora, Kinetoplastida, Tripanosomatidae). É um protozoário unicelular, largamente
distribuído na natureza, durante o ciclo biológico T. cruzi apresenta três formas morfológicas
distintas, a forma amastigotas com cinetoplasto e flagelo interiorizado; epimastigotas com
cinetoplasto justaposto ao núcleo e tripomastigota em que o cinetoplasto está posterior ao núcleo
(LISBOA; AVEIRO, 2008). As variações morfológicas é resultado das diferentes condições em
que o protozoário é exposto durante seu ciclo (MESSENGER; MILES; BERN, 2015).
T. cruzi tem alta diversidade fenotípica sendo subdivido em conjunto das
superpopulações de sete unidades taxonômicas ou Unidades de Tipagem Distintas (DTUs), TcI-
TcVI e Tcbat. É uma classificação de linhagem pura e hibridas, diferenciando pelo reservatório
natural, ciclo biológico, habitat, parasitemia e manifestações clinicas (ZINGALES, 2018;
ZINGALES et al., 2012). Sua circulação ocorre entre 100 espécies de insetos vetores
exclusivamente hematófago (Triatominae, Hemíptera, Reduviidae), albergando mais de 70
gêneros de mamíferos e se proliferando em diferentes biomas do continente americano (DIAS,
2006).

19
1.3. Aspectos e aspectos clínicos da Leishmaniose e da Doença de Chagas
Em relação a leishmaniose, os indivíduos com leishmaniose tegumentar americana
apresentam lesões únicas, profundas e ulceradas que se localizam no local da picada do mosquito,
sendo a leishmaniose cutânea a manifestação clinica mais comum. Geralmente, ocorre aumento
da temperatura no local da picada, formação de pápula eritematosa, pústula e por fim uma lesão
ulcerada, acometendo regiões como orelha, nariz, boca, mãos, antebraço ou pernas (ARENAS et
al., 2017; DE MAGALHÃES et al., 1986). Essas lesões, podem se disseminar através das vias
linfáticas, teciduais e sanguíneas para maior parte da pele do corpo formando nódulos eritematosos
e placas verrucosas, caracterizando a manifestação clinica mais comum, leishmaniose cutânea
difusa (ARENAS et al., 2017). Em áreas endêmicas, pacientes com leishmaniose cutânea podem
desenvolver, após 4-5 anos de cura, a leishmaniose mucocutânea. Essa patologia é caracterizada
por lesões muco-cutânea que afetam, de forma lenta, a região orofaríngea com aspecto necrótico,
rasgadas e descoladas (ARENAS et al., 2017; JENNINGS et al., 2014).
A leishmaniose visceral é a manifestação clinica mais grave e frequentemente letal. Tem
um período de incubação de 3 a 8 meses e afeta principalmente crianças, imunodeprimidos e
desnutridos. Indivíduos acometidos por leishmaniose visceral apresentam febre, anorexia,
fraqueza, sudorese noturna, hepatomegalia, esplenomegalia, palidez, anemia (BURZA; CROFT;
BOELAERT, 2018; GONTIJO; MELO, 2004).
A infecção da Doença de Chagas é complexa devido à alta variabilidade de virulência do
parasita e da susceptibilidade humana à infecção, onde a prevalência das manifestações variam
geograficamente (MARTINEZ; ROMANO; ENGMAN, 2020; ZINGALES, 2018). A
competência para infectar todas as células nucleadas de mamíferos representa um forte mecanismo
de manutenção de T. cruzi (JANSEN; XAVIER; ROQUE, 2018). As variáveis conhecidas que
determinam a duração da parasitemia no indivíduo incluem estado nutricional, a via de infecção e
a infecção concomitante, além do genótipo do parasita (ZINGALES, 2011, 2018; ZINGALES et
al., 2012).
Doença de Chagas apresenta duas fases, a aguda e a crônica. A fase aguda, geralmente é
assintomática devido a intensa atividade de anticorpos e da resposta imune inata mediada por
citocinas TH1, quando sintomático, ocorre sintomas adversos como febre e náuseas e alguns casos
graves com miocardite aguda e meningoencefalite (POULOSE, 2016). A fase crônica é
caracterizada pela forma indeterminada também designada de assintomática, sem anormalidades
na radiografia de tórax e eletrocardiograma(RIBEIRO; ROCHA, 1998; MOLINA; MOLINA
2018). Alguns indivíduos passam da fase aguda para a fase crônica da doença devido à infiltração

20
de amastigotas do T. cruzi nos órgãos). Quando sintomática, a fase crônica apresenta baixa
parasitemia, ocorrendo a destruição progressiva das células musculares do coração, do sistema
nervoso autônomo e afetando o trato digestivo (PÉREZ-MOLINA; MOLINA, 2018). Essa lesões
ocorrem de 10 a 30 anos após a infecção inicial (30 a 40% dos pacientes), sendo caracterizada por
cardiomiopatia (bradicardia, taquicardia) e complicações gastrointestinais, tais como, perda de
peso, disfagia, refluxo esofágico, tosse (SIMÕES et al., 2018).
1.4. Diagnóstico de Leishmaniose e da Doença de Chagas
O diagnóstico da Leishmania spp. é baseado em critérios que consideram dados
epidemiológicos, características clínicas e resultados de exames laboratoriais (GOTO; LINDOSO,
2010).
O diagnóstico para leishmaniose tegumentar se faz inicialmente por critérios
epidemiológicos, onde procura-se relacionar o indivíduo a área endêmica ou não, sendo
informação importante para o diagnóstico de viajantes residentes em áreas não endêmicas, visto
que leishmaniose tegumentar é uma das síndromes dermatológicas diagnosticadas em viajantes
(DE MAGALHÃES et al., 1986; GOTO; LINDOSO, 2010). O diagnóstico parasitológico direto
se faz por meio de amostras biológicas como sangue periférico, coleta de gânglios linfáticos ou
biopsia aspirativa, sendo relativamente rápido, apesar de necessitar de técnicos treinados e ter
baixa sensibilidade (DE MAGALHÃES et al., 1986). O teste cutâneo de Montenegro é utilizado
em estudos epidemiológicos para determinar a prevalência da infecção por Leishmania, no entanto,
o teste não distingue entre infecção presente e passada (GOTO; LINDOSO, 2010). O diagnóstico
sorológico se faz por ensaios de detecção e titulação de anticorpos circulantes como testes rápidos,
aglutinação direta, imunoblotting e ELISA (KARUNAWEERA; FERREIRA, 2018).
O diagnóstico para LV é complexo porque suas características clínicas são
compartilhadas por uma série de outras doenças comuns, como malária, febre tifoide e tuberculose
(SUNDAR S, 2002). Indivíduos infectados com LV apresentam febre prolongada,
esplenomegalia, hepatomegalia, leucopenia, anemia, hipergamaglobulinemia, tosse, dor
abdominal, diarreia, perda de peso e caquexia (GONTIJO; MELO, 2004). Para o diagnóstico
laboratorial, o parasita pode ser visualizado através da punção aspirativa do baço, medula óssea e
fígado para confecção de lâminas. Devido às limitações, diferentes técnicas são usadas para o
diagnóstico sorológico, a detecção de DNA do parasita em amostras de tecido, imunodiagnóstico

21
por detecção de antígeno de parasita em amostras de tecido, além do ensaio de imunidade mediada
por células específicas de Leishmania (GONTIJO; MELO, 2004).
O método padrão ouro usado para o diagnóstico de LV tem sido por aspirado esplênico
ou de medula óssea para demonstração do parasita, com sensibilidade de 95%. É preferível, por
parte do paciente, o aspirado esplênico devido a ser menos doloroso (BOELAERT et al., 1999;
SUNDAR S, 2002). O diagnóstico para pacientes com co-infecção HIV-Leishmania é
essencialmente o mesmo de pacientes não infectados com HIV, onde a análise de PCR mostra-se
útil (ARONSON et al., 2017).
O diagnóstico clínico da Doença de Chagas consiste na análise da história epidemiológica
do paciente, com dados sobre visitas ou transfusão de sangue em áreas endêmicas (DIAS, 2006).
A presença de sinais de porta de entrada como sinal de Romaña, um edema unilateral e indolor
próximo ao olho ocasionado pela resposta inflamatória à saliva do inseto ou o sinal chagoma de
inoculação, uma manifestação cutânea, avermelhada e endurecida que ocorre em qualquer parte
do corpo onde o inseto fez o repasto sanguíneo (PERSOON et al., 2018).
O diagnóstico laboratorial está diretamente relacionado com a fase da doença. Na fase
aguda o exame parasitológico direto é o mais indicado devido a intensa parasitemia, sendo possível
verificar a presença de tripomastigotas no sangue periférico (PÉREZ-MOLINA; MOLINA, 2018).
Outros testes parasitológicos diretos devem ser realizados simultaneamente como o método de
concentração e o de lâmina corada de gota espessa ou delgada (RIBEIRO; ROCHA, 1998). O
diagnóstico sorológico na fase aguda enfrenta dificuldades pela falta de testes comerciais, no
entanto, a verificação de anticorpo anti- T. cruzi da classe IgM é sugestiva, recomendando-se a
metodologia de imunofluorescência indireta (IFI), sendo usada quando os exames parasitológicos
diretos são negativos, porém, as suspeitas clinicas persistem (SIMÕES et al., 2018).
Na fase crônica da doença, o diagnóstico parasitológico direto da doença se faz por meio
de técnicas de isolamento e identificação de T. cruzi, muitas vezes controversas, como o
Xenodiagnóstico de Hemocultura (DIAS et al., 2016a). A técnica de PCR, apesar da falta de
protocolos padronizados, se torna uma sugestão diante de resultados indeterminados ou para
controle de cura (SIMÕES et al., 2018). O diagnóstico sorológico na fase crônica se faz por meio
de dois testes sorológicos com princípios/métodos distintos ou que possuam diferentes preparações
antigênicas, como o caso dos testes de ELISA e IFI, de elevada sensibilidade, e Hemoaglutinação
indireta, de elevada especificidade (PÉREZ-MOLINA; MOLINA, 2018; RIBEIRO; ROCHA,
1998).
Para diagnóstico de gestantes, recomenda-se a triagem sorológica de T. cruzi, para
aquelas vindas de áreas endêmicas, além da triagem sorológica de mulheres receptoras de sangue

22
em áreas endêmicas (MALAFAIA; DE LIMA RODRIGUES, 2010). Recomenda-se a testagem
sorológica anti-HIV nas avaliações do pré-natal, pois a co-infecção T. cruzi e HIV aumenta o risco
de transmissão congênita, morbidade de mortalidade (DIAS et al., 2016b).
1.5. Terapêutica atual da Leishmaniose e da Doença de Chagas
O tratamento medicamentoso para leishmaniose existe desde início do século 20 e embora
a OMS tenha fornecido recomendações para o tratamento, existem diferentes protocolos
terapêuticos devido a diversidade de espécies de Leishmania, diferença de susceptibilidade
medicamentosa e de manifestações clinicas das espécies do Velho e Novo mundo (GONTIJO;
MELO, 2004). Os antimoniais pentavalentes são os medicamentos de primeira linha para o
tratamento da leishmaniose, disponíveis em duas formulações: antimoniato de metaglutamina e
estibogluconato de sódio. Os principais efeitos adversos desse medicamento são cardiotoxicidade,
insuficiência renal e mialgia, não indicado para gestantes (BURZA; CROFT; BOELAERT, 2018).
Em algumas partes do mundo o tratamento de escolha é a Anfotericina B nas diferentes
formulações: anfotericina B desoxicolato, anfotericina lipossomal, anfotericina de dispersão de
colesterol e anfotericina de complexo lipídico. A lesão renal é o principal efeito adverso dessa
terapia (GOTO; LINDOSO, 2010; SILVA et al., 2019).
Tendo em vista a importância das leishmanioses e as dificuldades envolvidas em seus
tratamentos, tais como resistência aos antimoniais, a Miltefosina surgiu como um medicamento
promissor para o tratamento da leishmaniose, sendo administrado por via oral (GONTIJO;
MELO, 2004). Tal medicamento foi originalmente desenvolvido como antineoplásico e tendo
como efeito adverso a teratogenicidade, tem sido testado com bons resultados em formas
amastigotas e promastigotas, interferindo na membrana celular do parasita (DA COSTA FILHO;
LUCAS; SAMPAIO, 2008). Ainda não se tem vacinas para doenças parasitarias humanas
provocadas por protozoários como Trypanosoma cruzi e Leishmania spp., os fármacos presentes
são caros e causam muitos efeitos adversos, sendo ineficazes devido ao surgimento de resistência
(LISBOA; AVEIRO, 2008).
Segundo a OMS, a doença de Chagas e leishmaniose podem ocasionar perdas sociais
importantes nas áreas endêmicas, em termos de mortalidade, incapacidade laboral e custos
médicos os quais são ignorados pelos governos, muito ao fato do desconhecimento da população
exposta ou afetada pelas doenças (MALAFAIA; DE LIMA RODRIGUES, 2010).

23
Desde os anos 1990, o Ministério da Saúde de vários países de Organização Pan-
Americana e a OMS coordenaram iniciativas para controlar a Doença de Chagas, reduzindo a
transmissão por vetores e transfusão de sangue, onde esses esforços levaram a uma redução
substancial da carga da doença na América Latina (DIAS et al., 2016a).
Apenas dois medicamentos são licenciados e disponíveis com ação anti-tripanosomais
para Doença de Chagas e ambos estão longe de serem os ideais, pois requerem tratamento
prolongado, tem ampla gama de efeitos colaterais e são eficazes apenas na fase aguda da doença
(PÉREZ-MOLINA; MOLINA, 2018).
O Benzonidazol é o único medicamento aprovado pela Food and Drug Administration
(FDA) e atualmente usado para a terapêutica da Doença de Chagas no Brasil. É administrado por
via oral em dois ou três doses geralmente por 60 dias, alcançando uma boa efetividade na fase
aguda em crianças e adultos jovens. Contudo, o Benzonidazol apresenta alguns efeitos adversos
como anorexia, distúrbios do sono e digestivo, cefaleia e erupções na pele (PÉREZ-MOLINA;
MOLINA, 2018).O tratamento medicamentoso especifico para T. cruzi em mulheres gestantes ou
em períodos de amamentação é desaconselhado devido a dados de teratogenicidade demostrados
em animais (DIAS et al., 2016a).
Nifurtimox, não licenciado no Brasil devido aos efeitos adversos, foi o primeiro
medicamento usado e é administrado por via oral, em três a quatro doses, durante 60–90 dias. É
um fármaco pouco tolerável, apresenta muitas interações medicamentosas, além de uma gama de
efeitos adversos como anorexia, distúrbios neurológicos, febre, erupções da pele e distúrbios
digestivos incluindo náuseas e vômitos (DIAS et al., 2016a; PÉREZ-MOLINA; MOLINA, 2018).
Às medidas de prevenção, se faz necessário uma otimização e revisão de condutas de
tratamento e acompanhamento nas diferentes formas clinicas dessas patologias, além de uma rede
desenvolvida e hierarquizada no Sistema Único de Saúde (SUS), onde os serviços são distribuídos
de acordo com a endemicidade das doenças (DIAS et al., 2016b).
.

24
1.6. Metaloprotease de superfície de Leishmania spp. e Trypanosoma cruzi.
1.6.1 Metaloprotease de mamífero (MMPs)
As metaloproteases são uma família de endopeptídases proteolíticas, são estrutural e
funcionalmente relacionadas com as endopeptídases dependente de cálcio e contendo zinco, sendo
chamadas de metaloproteases de matriz (MMP). Estão principalmente associadas à degradação da
matriz extracelular (CUI; HU; KHALIL, 2017; NAPOLI et al., 2020). Em humanos, esta família
é composta por 24 proteínas codificadas por diferentes genes, sendo separadas em seis grupos com
base na organização de seus diferentes domínios, sequência de aminoácidos e especificidade de
substrato: colagenases, gelatinases, tipo de membrana, estromelisinas, matrilisinas e outros
(NAPOLI et al., 2020; TAJHYA; PATEL; BEETON, 2017).
Essas proteases são secretadas na forma latente ou inativas chamadas de zimogênios,
requerendo ativação. Em condições fisiológicas, as MMP desempenham importante papel na
proliferação celular, invasão celular, migração (adesão/dispersão), degradação da cartilagem,
remodelação do tecido, cicatrização de feridas, e embriogênese (CUI; HU; KHALIL, 2017;
TAJHYA; PATEL; BEETON, 2017). As MMP podem interagir com algumas moléculas
bioativas e receptores de superfície da proteína G, influenciando o microambiente celular e
possíveis sinalizações, além disso, podem estar envolvidas na apoptose celular, resposta
inflamatória e imunológica (BUNNEY et al., 2017; NAPOLI et al., 2020).
A atividade das MMP é regulada, principalmente, pelos inibidores de metaloprotease
endógenos, nos quais possuem alta heterogeneidade de sequência e consequentemente
propriedades funcionais diferentes. Atualmente, são conhecidos quatro tipos de inibidores de
MMP, com o tamanho aproximado de 23 kDa. Os inibidores se ligam às MMPs, bloqueando o
sitio ativo e impedindo o acesso da protease a seu substrato (DE MEDEIROS; GOMES; FARES,
2012). Em condições normais, o equilíbrio entre MMP e seus inibidores endógenos garante uma
eficaz remodelação tecidual, além de outros aspectos da fisiologia normal. Algumas dessas
proteinases, quando desreguladas, podem ser precursoras de numerosas condições patológicas
onde a síntese desregulada causa uma destruição tecidual ou aumento/diminuição desordenado de
MMP (DE MEDEIROS; GOMES; FARES, 2012; SNOEK-VAN BEURDEN; VON DEN HOFF,
2005).

25
1.6.2. Metaloprotease dos Tripanosomatídeos
Os tripanosomatídeos têm em comum um tipo particular de endopeptidases, as
metaloproteases de superfície presentes no parasita e que desempenham um papel importante em
várias etapas da infecção do hospedeiro, incluindo: adsorção, penetração, sobrevivência
intracelular, replicação, diferenciação, infectividade, evasão imunológica e nutrição (ISNARD;
SHIO; OLIVIER, 2012; SANTOS; BRANQUINHA; D’AVILA-LEVY, 2006).
Uma dessas moléculas é a Major surface protease (MSP) ou glicoproteína gp63, uma
metaloprotease de zinco dependente (Figura 4). Descoberta em 1980, pode estar associada à
membrana do parasita através de lipofosfoglicanos que contém uma âncora de
glicosilfosfatidilinositol (GPI) ou se apresentar na forma secretada, sem sinal de adição de âncora
GPI (CALIXTO; FAGUNDES; OLIVEIRA, 2014). Ambas são consideradas moléculas cruciais
para sobrevivência do parasita nos diferentes ambientes durante seu ciclo de vida (SÁDLOVÁ et
al., 2006). Inicialmente, a gp63 kDa foi descrita como tendo atividade de protease, sendo chamada
de protease de superfície e depois foi especificada como metaloprotease de zinco (ISNARD;
SHIO; OLIVIER, 2012).
Figura 4.Estrutura tridimensional da gp63.À esquerda, a estrutura tridimensional do esqueleto proteico da
gp63. Em azul localiza-se o domínio C-terminal, em verde o domínio central e em vermelho o domínio N-terminal.
O centro catalítico de zinco encontra-se se identificado por um círculo preto e amplificado na imagem à direita. A
molécula foi construída no programa UCSF 1.13, por meio do código de acesso 1LML no PDB.
As metaloproteínas são agrupadas em 3 principais famílias: a Gluzincina, Aspzincina e
as Metzincinas. Esta última agrupando as astacinas, serralisinas, adamalisinas, metaloprotease de
matriz, snapalisina e as leishmanolisinas (CALIXTO; FAGUNDES; OLIVEIRA, 2014).

26
As metaloproteases zinco-dependentes possuem um sítio catalítico composto por uma
sequência de aminoácidos HEXXH coordenada ao átomo de zinco (Figura 4). Membros da
família Metzincina possuem sequencias catalíticas estendidas HEXXHXXGXXX (SÁDLOVÁ et
al., 2006). Os genes da gp63 são altamente polimórficos, porém, a sequência de genes que
codificam os peptídeos funcionais envolvidos na atividade da protease é conservada. O
polimorfismo é mais abundante em segmentos de proteínas que servem como epítopo para
células T e B, e que não participam da interação parasita-hospedeiro (HASSANI et al., 2014).
Como observado na figura 4, a estrutura tridimensional da gp63 é uma estrutura
compacta, predominantemente presente na forma secundaria de folha β, dividida em três
domínios principais, tendo o domínio catalítico formado por duas Alfa-hélices, contra uma folha-
composta por cinco fitas- entrelaçadas, além da afinidade com vários tipos de substratos
proteicos (LISBOA; AVEIRO, 2008). A estrutura da gp63 indica que provavelmente existe um
hemodímero e as formas anfifílicas e hidrofílicas também são encontradas na célula (SANTOS;
BRANQUINHA; D’AVILA-LEVY, 2006).
No domínio N-terminal é o subdomínio que corresponde aos resíduos de aminoácidos de
Valina e Leucina e possui um padrão de enovelamento similar a outros zincos dependentes, além
de uma sequência conservada que contém um resíduo de cisteína ligado ao zinco. Esse domínio
alberga resíduos que compõe o sitio catalítico HGXXHXXGXH, sequência de aminoácidos
característica de metaloprotease zinco-dependentes(CALIXTO; FAGUNDES; OLIVEIRA, 2014;
ISNARD; SHIO; OLIVIER, 2012). O domínio central é constituído por -hélices e folhas-
antiparalelas, ligados ao domínio C-terminal por uma ponte dissulfeto (CALIXTO;
FAGUNDES; OLIVEIRA, 2014). O domínio C-terminal possui seis pontes dissulfeto, sendo um
domínio rígido. A âncora de glicosilfosfatidilinositol (GPI) é adicionada aos últimos resíduos deste
domínio (SÁDLOVÁ et al., 2006).
A síntese da proteína é realizada no réticulo endoplasmático em uma forma inativa ou
zimogênio, onde a sequência de sinal é clivada pós-tradução ou secretada. Um carboidrato é
adicionado ao domínio N e o C-terminal é substituído pela âncora GPI. Ocorre a clivagem de pró-
peptídeo quem tem cisteína para regular a atividade da gp63 (SANTOS; BRANQUINHA;
D’AVILA-LEVY, 2006). A gp63 secretada tem diferentes pesos moleculares devido aos seus
estados glicosilados (ISNARD; SHIO; OLIVIER, 2012). A estrutura tridimensional da gp63 tem
sido alvo de extensos estudos devido a sua importância como fator de virulência, abrindo novas
possibilidades para descoberta de novos alvos terapêuticos, tratamento com agentes
antiparasitários e diagnóstico (ISNARD; SHIO; OLIVIER, 2012).

27
1.6.1. gp63 de Leishmania spp.
A complexidade gênica no género Leishmania é distinta entre os diferentes subgéneros.
As espécies integradas no Subgênero Viannia dispõem de maior variedade de genes MSP, que
poderá ser até quatro vezes superior, relativamente ao subgénero Leishmania (LISBOA; AVEIRO,
2008). Tal gênero é capaz de elaborar uma grande variedade de proteases que são
intra/extracelulares com diferentes especificidades (SANTOS; BRANQUINHA; D’AVILA-
LEVY, 2006).
A protease principal de superfície (PSP) ou gp63 é distribuída, em toda superfície da
forma evolutiva promastigota incluindo o flagelo, tendo seu ápice em promastigotas
metacíclicas. Nas formas amastigotas intracelulares, sua expressão é reduzida (SANTOS;
BRANQUINHA; D’AVILA-LEVY, 2006; YAO, 2010). Estima-se que cada Leishmania na fase
estacionária possui 500.000 copias de gp63, cerca de 1% de todo o conteúdo proteico do
organismo, a maioria presente na superfície, é o restante localizando-se intracelularmente
(SANTOS; BRANQUINHA; D’AVILA-LEVY, 2006).
O elevado número de moléculas de gp63 presente na superfície de promastigotas
metacíclicas, tal como a sua presença nas formas amastigotas, embora de modo menos
representativo, é um requisito obrigatório para a eficaz manipulação do sistema imunitário e
consequente bem-sucedido estabelecimento da infeção, enquanto da sua inoculação no
hospedeiro (LISBOA; AVEIRO, 2008). Patógenos intracelulares geralmente alteram a
organização do citoesqueleto e o maquinário de sinalização celular da célula hospedeira,
contribuindo para sua internalização (ISNARD; SHIO; OLIVIER, 2012).
A gp63 fornece resistência a lise mediada por complemento e facilita o englobamento de
promastigota pelo macrófago. Após a internalização por fagocitose no hospedeiro, as formas
promastigotas do protozoário desativam funções imunológicas e antimicróbicas, através da
inibição da biogênese do fagolisossomo (MATTE et al., 2016). A gp63 permite a sobrevivência
de amastigotas dentro dos macrófagos devido a inibição de fatores pró-inflamatórios (MATTE et
al., 2016). A resolução da doença é necessária a resposta pró-inflamatória, e a progressão da
doença se dá através de atividades anti-inflamatórias (LISBOA; AVEIRO, 2008). Embora os
macrófagos sejam as células hospedeiras primárias de que abrigam as amastigotas após a infecção
inicial, outras células do sistema imune inato são conhecidas por estarem envolvidas na infecção
do parasita, e suas funções comprometidas pela gp63 (CARVALHO et al., 2013).
A gp63 tem ampla especificidade de substrato, hidrolisando diversas proteínas séricas
como: componentes do sistema complemento, fibrinogênio, fibronectina, hemoglobina, colágeno,

28
albumina, gelatina, caseína, além de afetar as vias de sinalização , facilitando a migração do
parasita nos tecidos (SANTOS; BRANQUINHA; D’AVILA-LEVY, 2006). Esta protease afeta
várias proteínas de sinalização do hospedeiro envolvidos no rearranjo do citoesqueleto, além das
funções antimicróbianas e inflamatórias dos macrófagos, mostrando seu papel central como fator
de virulência que contribui para a sobrevivência no início da infecção (ISNARD; SHIO;
OLIVIER, 2012).
O papel da gp63 no inseto vetor carece de mais estudos, mais acredita-se que essa
glicoproteína possibilita o parasita a obtenção de nutrientes através da hidrólise de componentes
proteicos (SANTOS; BRANQUINHA; D’AVILA-LEVY, 2006).
1.6.2. gp63 de Trypanosoma cruzi.
T. cruzi tem uma diversidade que pode estar associada a adaptação e sobrevivência
dentro de vários hospedeiros. Os diferentes genótipos a variabilidade em diferentes
características do parasitismo, como virulência, sintomas e susceptibilidade as drogas
antiparasitárias (GALVAN et al., 2016). T. cruzi é o parasita com maior repertório de genes, que
codificam as metaloproteases de superfície, destes. O protozoário contém mais de 700 genes e
pseudogenes para gp63 (BERNÁ et al., 2017).
Os genes que codificam as metaloprotease de superfície de T. cruzi são agrupados em três
grupos: Tcgp63-I, Tcgp63-II e Tcgp63-III. O grupo I está envolvido no processo infeccioso, onde
a enzima produzida tem local de ancoragem da GPI, ligando-a a membrana do parasita onde é
ativada. Tcgp63-II tem nível de transcritos pouco expressivo e não contém local de adição da
âncora GPI, não sendo expressa na membrana do parasita e Tcgp63-III é constituído de
pseudogenes (LISBOA; AVEIRO, 2008).
T. cruzi contém um grupo de genes de superfície que promove a ativação de linfócitos
policlonais e resposta humoral retardada na fase aguda. A ativação policlonal difunde a resposta
imunológica, impedindo uma resposta específica que neutralize e elimine o parasita
(WATANABE COSTA; DA SILVEIRA; BAHIA, 2016). A função da gp63 em T. cruzi
continua incerto.
De uma forma resumida, as metaloproteases de tripanosomatídeos participam de
importantes vias de sinalização, manutenção do parasita e viabilidade intracelular e extracelular.
Assim, manipulam a resposta imune do hospedeiro e dificulta o desenho de uma vacina eficaz.

29
1.7. Principais metodologias utilizadas para a identificação de metaloprotease
1.7.1. Zimografia para estudos biológicos
Vários métodos têm sido desenvolvidos para avaliação das formas ativas e latentes de
enzimas proteolíticas, entretanto, a zimografia é o método mais usado por ser o mais completo na
biologia molecular (DE MEDEIROS; GOMES; FARES, 2012). Descrita pela primeira vez em
1980 por Heussen e Dowdle, é uma técnica semi-quantitativa e muito sensível, amplamente usada
na identificação e caracterização de protease em células, tecidos ou fluidos biológicos
(WILKESMAN, 2017a).
Por eletroforese em gel sob desnaturação e condições não redutoras, esta técnica fornece
uma análise sensível, quantificável e funcional da atividade de protease por meio da degradação
em diferentes substratos e por seu peso molecular (WILKESMAN, 2017b).
A técnica consiste em uma eletroforese em gel desnaturante de poliacrilamida,
copolimerizado com um detergente aniônico, dodecil sulfato de sódio (SDS) e com o substrato
preferencial da enzima em análise. A aplicação de uma corrente elétrica possibilita a migração das
proteases no gel e sua separação pela diferença do tamanho molecular (DE MEDEIROS; GOMES;
FARES, 2012). As proteases separadas dentro do gel são renaturadas por lavagem com um
detergente não iônico, como Triton X100, devolvendo sua estrutura terciária que irá realizar a
digestão do substrato. O gel é então incubado em um tampão adequado, permitindo que a protease
renaturada realize a digestão do substrato na região circundante de sua eletroforese
(WILKESMAN, 2017a). As metaloproteinases são então identificadas comparando as bandas
claras no gel com padrões de peso molecular e / ou por testes de inibição com inibidores específicos
de certas classes de metaloproteinases.
A zimografia tem a vantagem sobre outras técnicas da biologia molecular por não
requerer anticorpos, tornando-a barata, a migração em condições não redutoras possibilita a
visualização da protease, possibilita a distinção de enzimas latentes e ativas como base no seu peso
molecular, bem como a atividade pode ser quantificada por densitometria (WILKESMAN, 2017a).
A zimografia é usada como o único ensaio para se observar a atividade das MMP ou MSP, bem
como seus inibidores, porém, outras técnicas podem ser usadas para complementar os resultados
obtidos como o ensaio imunoenzimático (ELISA), o Western Blot, a imunohistoquímica e a
imunofluorescência (DE MEDEIROS; GOMES; FARES, 2012).
A zimografia é frequentemente usada pois é um ensaio sensível e quantificável para
analisar a atividade de metaloprotease, porém, alguns problemas precisam ser considerados ao

30
aplicar essa metodologia como a natureza, origem e preparação das amostras, o substrato no gel,
podem comprometer a validade da técnica e complicar a interpretação de resultados (BUNNEY et
al., 2017).
A técnica indica a presença de metaloprotease e seus inibidores e não sua atividade real
dentro do corpo. Amostras com altas concentrações geram bandas distorcidas na matriz do gel.
Existem alguns marcadores de pesos moleculares disponíveis comercialmente em condições com
agente redutor. Quando usados em condições não redutoras, podem indicar pesos moleculares
diferentes. A densitometria é usada para quantificar a atividade das proteases, porém, pode ocorrer
a formação de complexos de metaloprotease (protease mais proteína) ou várias metaloprotease
podem formar complexos que na análise por Western blot configura-se como um anticorpo.
Agentes redutores como B-mercaptoetanol não devem ser adicionados, pois esses agentes
quebram as pontes dissulfeto e evitam algumas formas de dobramentos de proteínas terciarias. A
zimografia in situ não difere metaloproteases, pois os substratos são degradados por mais de uma
protease, sendo necessário o uso de técnicas complementares (HU; BEETON, 2010; SNOEK-
VAN BEURDEN et al., 2005).
Originalmente, a zimografia foi desenvolvida usando gelatina como substrato, sendo
chamada de zimografia de substrato. Todas as outras zimografias de substrato se originaram da
zimografia da gelatina, diferindo apenas do substrato usado, que vai depender do tipo de
metaloprotease e inibidores a serem analisados (SNOEK-VAN BEURDEN et al., 2005).
Algumas MMP não têm afinidade com substrato de gelatina, sendo feitas modificações
na técnica para melhorar a detecção. Incorpora-se substrato de caseína, colágeno, albumina com
aprimoramento com heparina. A zimografia com caseína é ideal para analisar estromelisina,
porém, é menos sensível que de gelatina, além do substrato de caseína correr no gel durante a
eletroforese devido ao seu baixo peso molecular (23kDa), resultando em duas zonas no gel com
caseína e sem caseína (SNOEK-VAN BEURDEN et al., 2005; WILKESMAN, 2017a).
A zimografia reversa é uma modificação da zimografia de substrato, sendo usada para
identificar e analisar inibidores endógenos ou sintéticos (TAJHYA; PATEL; BEETON, 2017). No
gel de poliacrilamida é incorporado um substrato com sua MMP especifica, e os inibidores
aplicados no gel. Após a eletroforese, a MMP é ativada e digere todo o substrato do gel, exceto na
área onde o inibidor se faz presente, e após a coloração, o gel se mostra incolor exceto nos locais
dos inibidores, indicando a não degradação do substrato. Atualmente, para aumentar a
sensibilidade e usar menores quantidades de substrato, usa-se substratos com marcadores
isotiocianato de fluoresceína (SNOEK-VAN BEURDEN et al., 2005).

31
Zimografia in situ permite localizar as metaloproteases em tecidos, sendo uma otimização
ou modificação da zimografia de substrato. Essa metodologia consiste no depósito de um substrato
sobre ou sob um tecido congelado não fixado. Durante a incubação, o substrato é digerido pelas
proteases, e a degradação é detectada por microscopia de luz ou fluorescência (WILKESMAN,
2017b).
Os tripanossomatídeos elaboram uma grande variedade de proteases de superfície, que
são intracelulares e/ou extracelulares, com diferentes especificidades (SANTOS;
BRANQUINHA; D’AVILA-LEVY, 2006). A zimografia é usada em múltiplos estudos que
relacionam as metaloproteases de superfície com enfermidades causadas em vertebrados (DE
MEDEIROS; GOMES; FARES, 2012).

32
2. Justificativa
O estudo, das proteases de microorganismos, particularmente os intracelulares, tem
demonstrado que estas enzimas desempenham importantes funções durante a patogênese. Estas
proteínas participam no processo de invasão ao hospedeiro, podem assimilar os aminoácidos como
fonte nutricional ou para síntese de substâncias orgânicas, na diferenciação e escape ao sistema
imune do hospedeiro (CALIXTO; FAGUNDES; OLIVEIRA, 2014; MATTE et al., 2016).
A metaloproteinase (gp63), possui a capacidade de degradar diferentes peptídeos e
substratos proteicos tais como caseína, azo caseína, gelatina, colágeno, albumina, fibrinogênio e
hemoglobina. Deste modo, a matriz extracelular do macrófago é susceptível à hidrólise pela
enzima em questão, o que favorece a migração do parasita no seu interior. Acredita-se que o
mesmo se verifica em formas recém-inoculadas no tecido subcutâneo, em que no início da
infeção, podem aceder aos fibroblastos, inclusive os dos nódulos linfáticos (ISNARD; SHIO;
OLIVIER, 2012; LISBOA; AVEIRO, 2008; YAO, 2010). Neste contexto, a caracterização
bioquímica das metaloproteinases de T. cruzi (TcMSPs), e Leishmania spp. (LeishMSPs) mostra-
se como uma ferramenta de investigação, através da otimização de técnica da zimografia e suas
diferentes variações. Deste modo, podem ser usadas para avaliar com mais detalhes as
metaloproteases e ajudar o desenvolvimento de possíveis inibidores sintéticos potentes e seletivos
para o controlo da doença de Chagas e Leishmanioses.

33
3. OBJETIVOS
3.1. Objetivo geral
Realizar a otimização da técnica de zimografia e ensaio de degradação in vitro como
metodologia para caracterização bioquímica de metaloproteinases de Trypanosoma cruzi e
Leishmania spp..
3.2. Objetivos específicos
Este trabalho tem como objetivos específicos:
Otimizar substratos para o desenvolvimento da metodologia de zimografia;
Identificar atividade proteolítica do extrato bruto de Leishmania spp. e
Trypanosoma cruzi utilizando protocolo caseiro de zimografia;
Avaliar atividade proteolítica de Leishmania spp. e Trypanosoma cruzi in vitro
utilizando gelatina e albumina como substrato.

34
4. MATERIAIS E MÉTODOS
4.1. Substratos
A Gelatin From Bovine Skin (Sigma aldrich) é uma proteína de origem animal obtida a
partir da hidrólise parcial do colágeno bovino, suíno, frango e de peixes. Apresenta uma cadeia
proteica simples, resultante da degradação química e física das fibras proteicas insolúveis do
colágeno. É formada por 18 diferentes aminoácidos sendo amplamente utilizada em indústrias
farmacêuticas por possuir diversas características funcionais (FERREIRA, 2013).
A Bovine Serum Albumin - BSA (Sigma-aldrich) é uma proteína abundante no sangue
bovino e possui estrutura similar a estrutura HSA (Human Serum Albumin), apresentando
porcentagem de sequência de aminoácidos de 76%. O BSA é uma proteina constituida por uma
cadeia polipeptídica simples de 583 resíduos de aminoácidos com alto número de cisteína, lisina,
arginina, ácido glutâmico e aspártico, além de 3 domínios, podendo formar dímeros,
principalmente em altas concentrações (FERREIRA, 2009).
4.2. Leishmania spp. e Trypanosoma cruzi
Leishmania amazonensis
A espécie, Leishmania amazonensis (L. amazonensis) foi isolada em 1973 de um
paciente com leishmaniose cutânea difusa, no Estado do Pará, no Norte do Brasil, através de
ensaios utilizando anticorpos monoclonais e eletroforese isoenzima no Instituto Evandro Chagas
(Jennings et al., 2014).
Leishmania infantum
Leishmania infantum (L. infantum) foi obtida por citologia aspirativa do gânglio
linfático de um canídeo do canil municipal da região metropolitana de Lisboa, Portugal e mantida
no Instituto de Higiene e Medicina Tropical (IHMT) por passagens sucessivas em camundongos
BALB/c.

35
Leishmania guyanensis
Leishmania guyanensis (L. guyanensis) foi isolado em 2001 de um paciente com
leishmaniose cutânea na microrregião de Santarém, na mesorregião do Baixo Amazonas no
Estado de Pará, Brasil.
Leishmania major -
A espécie é o agente etiológico da leishmaniose cutânea do Velho Mundo. A espécie
foi descrita em 1914 por Vasili Larionovich Yakimoff e Schokhor como Leishmania tropica
major.
Leishmania donovani
A espécie é o agente etiológico da leishmaniose visceral do Velho Mundo. A espécie
foi descrita em 1903 por Alphonse Laveran e Félix Mesnil como Piroplasma donovani.
Trypanosoma cruzi
Cepa Y
A cepa Y foi isolada de um caso agudo humano de doença de Chagas por meio de
xenodiagnóstico por Silva e Nussenzweig, em 1953. Formas tripomastigotas foram mantidas em
camundongos Swiss e formas epimastigotas em meio de cultura LIT.
Cepa Dm28c
Dm28 foi originalmente isolado de opsum, Didelphis marsupialis. O clone Dm28c foi
obtido a partir da infeção de camundongos recém-nascidos com as formas tripomastigotas do
parasito.
Cepa Bolívia
A cepa Bolívia foi isolada de fezes de Triatoma infestans, proveniente de Vitichi-
Bolívia por Funayama e Prado Júnior, 1974.
Cepa CL Brener
Cepa CL Brener que foi isolada de fezes de triatomíneos Triatoma infestans, vetor
estritamente domiciliar (Brener & Chiari, 1963).

36
4.3. Cultura axênica dos parasitas
O estabelecimento de um protocolo de manutenção dos parasitos em meios de cultivo tem
por objetivo promover crescimento in vitro, para obtenção dos extratos proteicos totais, para
posterior análise em ensaios enzimáticos.
4.3.1. Cultura de Leishmania spp.
As formas promastigotas de Leishmania spp. em estudo foram mantidas em meio
desenvolvido por Moore et al. no Roswell Park Memorial Institute - RPMI-1640 (Sigma Aldrish)
contendo L-glutamina (Sigma-Aldrich, Alemanha) suplementado com 0,4 g de bicarbonato de
sódio (NaHCO3, Sigma-Aldrich), 10% de (FBS-Fetal bovine serum inativado, Sigma-Aldrich)
(v/v) ,1,2% (m/v) de cloreto de cálcio (CaCl2, Sigma-Aldrich), 5mM de HEPES (Sigma-Aldrich)
e 0,5% de penicilina/etreptomicina (Sigma-Aldrich) (combinação de 10.000 U.ml-1 de penicilina
com 10.000 g.ml-1 de etreptomicina) a pH 7,2 e a 28 ℃.
4.3.2. Cultura de Trypanosoma cruzi.
A partir da cultura pré-estabelecida (3x105 parasitas/mL), as formas epimastigotas das
cepas de T. cruzi foram transferidas para um tubo falcon ao qual acrescentou-se previamente 5 ml
de meio Liver Infusion Tryptose (LIT). Este meio é constituído por 68 mM de cloreto de sódio
(NaCl, Sigma-Aldrich), 5 mM de cloreto de potássio (KCl2, Sigma-Aldrich), 49mM de fosfato de
sódio (Na2HPO4, Sigma-Aldrich), 11 mM de dextrose (C6H12O6, Sigma-Aldrich), 0,5% de
triptose (Sigma-Aldrich), 0,3% de caldo LB (LB broth, Sigma-Aldrich) e 0,013% de Haemin
(Sigma-Aldrich), cujo pH é ajustado para 7,2. O meio LIT também é suplementado com 10% de
FBS (v/v) e antibióticos (0,5% de estreptomicina/penicilina).
4.4. Preparação dos extratos
Após atingirem a fase estacionária de crescimento, as culturas de Trypanosoma e
Leishmania spp. contendo as formas epimastigotas e promastigotas respectivamente, foram
organizadas para a obtenção dos extratos proteicos totais. Para tal, inicialmente as culturas foram
centrifugadas 3000 x g a 4°C por 10 minutos, desprezado o sobrenadante e seguidamente foram
lavadas 3 vezes com PBS 1x (Phosphate Buffer Saline).

37
Após as lavagens, o pellet foi ressuspendido em tampão de lise sem inibidor de protease
(120mM NaCl, [Sigma-Aldrich], 50mM de tris-HCL [Trizma, Sigma-Aldrich] e 0,5% de NP-
40 [Biochemika, EU], pH 7,8), para a obtenção dos extratos brutos. Por fim, procedeu-se ao
processo de ruptura celular mecânica, através de seis cíclos de congelação e descongelação a -
20°C. Seguidamente, o extrato foi armazenado a -20°C até a sua utilização nos ensaios.
4.5. Determinação da concentração proteica dos extratos brutos
Para a caracterização do perfil eletroforético por SDS-PAGE e determinação da
atividade proteica pelo método de zimografia, os extratos proteicos totais dos parasitas T. cruzi
e Leishmania spp. foram previamente quantificados utilizando a metodologia proposta por
Bradford (Bradford, 1976). A proteína albumina sérica bovina (bovine serum albumin - BSA,
Sigma-Aldrich) foi utilizada como proteína padrão, possibilitando assim a obtenção da equação
da reta para determinação proteica de cada extrato.
Foi utilizado o reagente de Bradford (composto por ácido fosfórico, Comassie blue e
etanol), onde ocorre a ligação da proteína ao complexo ácido fosfórico/Comassie blue e a
mudança de cor ocorre, sendo esse reagente o ideal para determinação da concentração de
proteínas. Para o ensaio de quantificação, foi utilizado uma microplaca de 96 poços
(BrandPlate, Alemanha), sendo adicionado em duplicata, 5µl de extrato proteico dos
tripanosoamtídeos de T. cruzi e espécies de Leishmania. Posteriormente, foi adicionado 250
µl do reagente de Bradford e no controle (branco) foi adicionado 250µl do reagente de
Bradford e 5µl de PBS a 1%. Seguidamente, a placa foi incubada ao abrigo da luz durante 20
minutos a temperatura ambiente e foi realizada a leitura das absorbâncias no comprimento de
onda 570nm, utilizando o leitor de microplaca (BioTek, USA).
4.6. Identificação das metaloproteinases dos extratos de Leishmania spp. e
Trypanosoma cruzi
4.6.1. Caracterização do perfil proteico por SDS-PAGE
Para a visualização do perfil proteico dos extratos, foi realizada uma eletroforese em gel
de poliacrilamida a 10% (SDS-PAGE) de acordo com o método de LAEMMLI (1970). A
preparação do gel consiste na polimerização da acrilamida e da N-metilenobisacrilamida, sendo o

38
gel, quando polimerizado, quimicamente inerte, eletricamente estável e transparente para a
detecção ótica.
Na elaboração do gel foi usado um sistema de eletroforese vertical (Bio-Rad, EUA) sendo
preparado 8 mL solução de gel de corrida (migração de acordo com a massa) em um tubo de
Falcon: 4,1 ml de água destilada; 3,3 ml de acrilamida (30% acrilamiada (m/v) (Sigma-Aldrich) e
0,8% Bis-Acrilamida (v/v) (Sigma-Aldrich); 2,5ml de 1,5M Tris-HCl (Trizma - Sigma-Aldrich);
100μl de dodecil sulfato de sódio (SDS, Sodium dodecyl sulfate) (Sigma-Aldrich) a 10% (m/v);
50 μl Persulfato de amónia a 30% (p/v) (Sigma-Aldrich) e 5 μl de tetrametiletilenodiamina
(TEMED, Sigma-Aldrich), em que é colocado ao conjunto de moldes de vidro, onde depois da
sua polimerização foi preparado o gel de empacotamento 4% constituído por: 2,5 ml de água
destilada, 450μL de acrilamida (30% acrilamiada (m/v); Sigma-Aldrich) e 0,8% bis-Acrilamida
(v/v) (Sigma-Aldrich); 333μL de Tris-HCl 0,6M (Trizma - Sigma-Aldrich); 100 μL de SDS
(Sigma- Aldrich) a 10% (m/v); 50μl de solução saturada de persulfato de amónia e 5μl de TEMED
(Sigma- Aldrich).
Antes da polimerização do gel de empacotamento, é colocado um pente na parte superior
do gel para formar poços, onde foram colocados de 5 a 20 microlitros de amostra, além de 5μl do
padrão de massa molecular (Thermo Fisher scientific) como observado na figura 5.
Figura 5. Eletroforese em gel de Poliacrilamida (SDS-PAGE).
Os extratos quantificados foram solubilizados, na proporção de 1:1, em tampão de
amostra para SDS-PAGE: 3,55mL de água destilada, 1,25 ml de 0,5M Tris-HCl a pH 6.8, 2,5ml
de glicerol (Sigma-Aldrich), 2 ml de SDS 10% (m/v) (Sigma-Aldrich), 200μl de azul de
bromofenol 0,5% (m/v) (Sigma-Aldrich) e 50μl de beta-mercaptoetanol (Sigma-Aldrich). Sendo
este último adicionado no momento de preparação da diluição final da amostra, onde é responsável
pela redução das ligações dissulfeto. Por fim, após diluição, as amostras foram incubadas à
temperatura de 100ºC durante cinco minutos para desnaturação. Na cuba do sistema
eletroforético, os géis polimerizados foram submersos em tampão de corrida constituído por:
25M de Tris, 0,19M de

39
glicina (Sigma-Aldrich) e 0,1% de SDS. Um campo elétrico de 80 a 120 volts foi estabelecido,
promovendo a separação das proteínas com base no tamanho, carga e forma. Após a eletroforese,
os géis foram imersos na solução corante azul de Coomassie (Coomassie Brilliant Blue R-250,
Bio-Rad, EUA) constituído por: metanol 30%; ácido acético 10%; Comassie blue G-250 0,5% e
água destilada até a total coloração. Após a coloração, o corante foi retirado e os géis imersos na
solução Descorante constituída por: metanol 30%; ácido acético 10% e água destilada até a
visualização total das bandas proteicas.
4.6.2. Determinação da atividade enzimática por zimograma
Procedeu-se a avaliação qualitativa da atividade de gelatinase e albuminase dos parasitas
Leishmania e T. cruzi, através de ensaios feitos por zimografia frente a substratos de albumina e
gelatina. Em géis de poliacrilamida a 10% preparados conforme descrito anteriormente (3.6.1), foi
copolimerizado com 0,41% de gelatina e albumina (gelatin from bovine skin; bovine Serum
Albumin – BSA - Sigma-Aldrich).
As amostras foram preparadas de acordo com a concentração proteica presente, os
extratos solubilizados em tampão de amostra não redutor para SDS-PAGE (0,2 M Tris-HCl; 20%
glicerol; 6% SDS; 0,05% de azul de bromo fenol; pH 6,7) foram adicionados nos poços e os géis
submetidos à eletroforese com corrente elétrica constante de 80V a 4°C, evitando a desnaturação
das proteínas. Após concluída a separação proteica, os géis foram transferidos para 50ml de
solução de Renaturação (2,5% Triton X-100) durante uma hora a 37°C. Depois da incubação, os
géis foram lavados três vezes com agua destilada e transferidos para 50ml de solução de
Desenvolvimento (0,5 M Tris-HCl, pH 7,5, 200 mM NaCl, 5 mM CaCl2, 0.02 % (p/v) Brij-35)
overnight a 37°C. Depois do perído da incubação, realizou-se a coloração com Comassie blue (G-
250) e seguidamente a descoloração, como descrito anteriormente (3.6.1).
4.7. Otimização e determinação da atividade proteolítica in vitro de extrato
bruto de Leishmania spp. e Trypanosoma cruzi.
Inicialmente, foi obtido o perfil proteico do substrato de Bovine Serum Albumin – BSA
(Sigma-aldrich). Para tal foi pesado 1 miligrama de substrato e dissolvido em um eppendorf com
0,5ml de PBS (1x). As amostras de substratos nas concentrações decrescentes 20,15,10 e 5ug,

40
respectivamente, foram solubilizadas, na proporção de 1:1, em tampão de amostra. Seguidamente
submetidas a separação em gel de poliacrilamida a 10% em 10 poços foi preparado conforme
descrito anteriormente (4.6.1).
Para determinação da atividade proteolítica in vitro de extrato bruto de Leishmania spp. e
T. cruzi foi necessário realizar a quantificação por meio do método de Bradford (Bradford, 1976),
conforme descrito anteriormente (4.5). Quatro extratos de T. cruzi e cinco de Leishmania que
apresentaram as melhores concentrações foram escolhidos para iniciar-se o ensaio de
degradação. Num tubo de eppendorf foram preparadas amostras contendo 40ul de extrato bruto e
10ul de substrato (BSA) e incubados a 37°C durante 24, 48 e 72 horas. Como controle, foi
utilizado um tubo contendo apenas o substrato sendo incubado nos mesmos intervalos de tempo.
Após as etapas de incubação, as amostras foram solubilizadas em tampão de amostras
na proporção de 1:1, sendo submetidas posteriormente a desnaturação por 5 minutos a 100°C.
Para verificar uma possível degradação do substrato devido a presença do extrato bruto de
tripanosomatídeos, foi preparado um gel de poliacrilamida a 10%, em que no gel de empilhamento
onde contém os poços, foram adicionadas as amostras e o marcador de peso molecular, assim foi
iniciada a eletroforese SDS-PAGE, promovendo a separação proteica. No fim da eletroforese, os
géis foram retirados da cubeta, lavados por 20 segundos com água destilada e imersos na solução
corante azul de Coomassie (Coomassie Brilliant Blue R-250, Bio-Rad, EUA) até a coloração total
do gel. Após a coloração, o corante foi retirado e os géis imersos na solução Descorante até a
visualização das bandas.

41
5. RESULTADOS E DISCUSSÃO
5.1. Perfil proteico de Leishmania spp. e Trypanosoma cruzi
A família tripanosomidae é um grupo cosmopolita de flagelados contendo parasitas que
afetam todos os principais grupos de organismos eucariontes. Eles são distinguíveis de outros
protozoários por características organizacionais distintas, como presença de DNA mitocondrial
cinetoplasto, isolamento de enzimas glicoliticas na organela glicossomo e uso de bolsa flagelar
para tráfego de moléculas (SANTOS; BRANQUINHA; D’AVILA-LEVY, 2006). Os
tripanosomatideos são capazes de adaptar seu metabolismo energético à disponibilidade de
substrato e oxigênio, possibilitando instituir novos ciclos de vida (SÁDLOVÁ et al., 2006)
Os protozoários Leishmania spp. e T. cruzi integram a mesma família taxonômica,
tripanosomatidae, sendo por isso esperado que apresentem proteínas em comum. Após a obtenção
do extrato, realizou-se a quantificação proteica do extrato bruto total pelo método de Bradford.
Para tal foi utilizado como curva padrão, a proteína Albumina sérica bovina – BSA, como
demostrado na figura 6.
Figura 6.Curva padrão, utilizando a proteína Albumina sérica bovina (BSA). A partir da curva foi possível
determinar a concentração proteica do extrato bruto de Leishmania spp e T. cruzi.

42
A eletroforese é uma técnica de separação baseada na migração de moléculas
carregadas, numa solução, em função da aplicação de um campo elétrico. Na eletroforese em gel,
o deslocamento das partículas é retardado pelas interações com o gel da matriz, que constitui um
meio de migração e comporta-se como uma rede molecular. Sendo uma técnica amplamente usada
em vários segmentos da pesquisa como bioquímica clínica, biologia molecular, biotecnologia,
além da análise de DNA (HU; BEETON, 2010).
Os perfis dos extratos de Leishmania spp. e T. cruzi foram analisados pelo método de
eletroforese SDS-PAGE, usando géis de poliacrilamida a 10% corados com Comassie, (Figura
7).
Figura 7.Perfil proteico total dos extratos de Leishmania e as respectivas concentrações por poço, em
géis corados com azul de Coomassie. M – Marcador de pesos moleculares; 1 – L. amazonensis (3,60 µg); 2 – L.
donavani (1,35 µg); 3 – L. guyanensis (1,08 µg); 4 – L. infantum (1,5 µg) e 5 – L. major (3.6 µg).
A coloração por Comassie mostrou-se eficaz devido à grande especificidade por
proteínas, apesar da pouca sensibilidade quando comparada com a coloração por nitrato de prata
(LISBOA; AVEIRO, 2008). Os perfis proteicos dos parasitas estudados são claramente distintos
entre si, contudo, não é possível distinguir entre as diferentes espécies de Leishmania e as
estirpes de T. cruzi (figura 8). Os resultados obtidos demostram a presença de um maior número
de bandas nos extratos brutos de Leishmania, cujo pesos moleculares das bandas variam entre 40
a 80kDa, este resultado foi semelhante com estudos feitos por MONTEIRO (2015) e MORENO
(2019) em que usaram o gel de poliacrilamida na mesma concentração,a mesma coloração e
parasitos.

43
As espécies de L. amazonensis e L. guyanensis apresentaram, além do perfil citado,
bandas proteicas em torno de 100 a 120 kDa, permitindo uma visão mais detalhada. Este fato pode
se dar porque parasita contém uma maior quantidade de glicoproteína 63 em toda sua superfície
que pode expressar diferentes pesos moleculares próximos a 63kDa (figura 7).
As estirpes de T. cruzi apresentaram um perfil menos detalhado com bandas proteicas
mais discretas (Figura 8). Embora este parasita tenha um maior repertório gênico de
metaloprotease, a expressão e a função proteolítica são menores comparativamente às espécies
de Leishmania. O perfil das estirpes de T. cruzi apresentam bandas que giram em torno de 20 -32
kDa e 35 - 60kDa, além de proteínas entre 60 - 120kDa. A principal cisteína-protease de
superfície, a cruzipaína (glicoproteína de 57 kDa) está presente em T. cruzi.
Figura 8.Perfil proteico total dos extratos das cepas de Trypanosoma cruzi e as respectivas concentrações
por poço, em géis corados com azul de Coomassie. M – Marcador de pesos moleculares; 1 – D m28c (0.58 µg); 2 –
QMM9 (3,9 µg); 3 – Cl Brener (2,5 µg); 4 – Bolívia (05,9 µg) e 5 – Y (3,2 µg).
As transformações morfológicas do parasita estão associadas a alterações específicas nos perfis
de glicopeptídeos e polipeptídeos. Os resultados indicam a presença de mucinas, moléculas
abundantes, presentes na superfície dos diferentes estágios do T. cruzi, com peso molecular

44
aparente de 40kDa, além de uma proteína com peso molecular aparente de 54 kDa (MEJÍA;
PALÁU; ZÚÑIGA, 2004).
A cruzipaína, principal cisteína-protease de T. cruzi, é uma proteína altamente homóloga
com outros membros da família de protease da papaína (SIQUEIRA-NETO et al., 2018). Formas
homólogas são encontradas em diferentes cepas do parasita, apesar das diferentes isoformas que
são encontradas no genoma do parasita (SCHNAPP et al., 2002). A virulência e a morfogênese do
parasita dependem da atividade endógena da cruzipaína lisossomal, sendo essencial para
replicação de amastigotas e tem papel crucial nas interações parasita-hospedeiro (CERNY et al.,
2020).
5.2. Atividade proteolítica de extratos Leishmania spp. e Trypanosoma cruzi.
A partir dos resultados dos perfis proteicos, foram feitos ensaios proteolíticos dos extratos
brutos através de protocolo caseiro de zimografia, adaptado aos substratos de gelatina e albumina,
proporcionando uma comparação qualitativa entre as espécies.
Os substratos encontram-se copolimerizados nos géis de poliacrilamida e a atividade dos
extratos está relacionada com a sua especificidade ao substrato.
Os resultados obtidos usando o protocolo caseiro de zimografia, demostra atividade
gelatinolitica e albuminolitica dos extratos brutos do parasita Leishmania. As cinco espécies de
Leishmania escolhidas para esse trabalho apresentaram atividade sobre o substrato de gelatina,
atividade essa mediada por enzimas de pesos moleculares que vão de 50 a 80 kDa, além de bandas
de atividade proteolítica de aproximadamente 100 a 120 kDa (figura 9).
Figura 9.Perfil enzimático, em gel de poliacrilamida acoplado com o susbstrato gelatina, na presença de
extratos de Leishmania spp e T. cruzi. 1 - L. amazonensis; 2- L. donavani; 3 - L. guyanensis; 4 - L. infantum; 5 - L.
major; 6- Cl Brener; 7 – Dm28c; 8 – Y e 9– Bolívia.

45
A observação da atividade frente ao substrato de gelatina por extratos de Leishmania
sugere papel importante das enzimas do parasita frente ao substrato de colágeno e fibronectina do
hospedeiro vertebrado, além de crucial durante os diferentes ciclos de vida do parasita,
concretizando a presença abundante de enzimas nas formas promastigotas metacíclica (Figura 9).
Em extratos de Leishmania, a gp63 é a principal proteína de superfície, tornando provável
que a atividade gelatinolitica observada entre 50 a 80 KDa seja correspondente a proteína. As
espécies de L. donavani L. guyanensis, L. infantum e L. major apresentaram uma proteína com
atividade de peso molecular de 40 kDa, além da glicoproteína gp63. Este resultado foi de acordo
com observado por MONTEIRO (2015) e MORENO (2019). Foi observada a presença de bandas
na parte superior do gel em todas as espécies de Leishmania, podendo ser a forma inativada da
gp63, apresentando maior peso molecular devido ao pró-peptídeo (LISBOA; AVEIRO, 2008).
Em relação aos extratos brutos de T. cruzi, verificou-se atividade proteolítica para as
estirpes de Cl Brener e Dm28c em gel de gelatina que vão de 50 a 60kDa. Em relação as estirpes
de Bolívia e cepas Y foi possível observar atividade semelhante, mesmo que de forma menos
evidente quando comparada com outros extratos da mesma espécie (Figura 9). A proteína
observada pode ser, além da gp63, a principal cisteína-protease de T. cruzi, a cruzipaína que
apresenta peso molecular de 57kDa. A atividade entre 50-60 kDa detectada aqui pode ser
idêntica a enzima cisteína-peptidase que foi amplamente caracterizada por Cazzulo e seus
colegas. Assim como a glicoproteína 82, que parece ter relação entre a interação do protozoário
com hospedeiro vertebrado (SIQUEIRA-NETO et al., 2018).
Na atividade enzimática de T. cruzi, conclui-se que não há diferença na expressão
enzimática entre os estágios de vida do parasita (CERNY et al., 2020). A cruzipaína está presente
em todas as etapas do ciclo de vida e é identificada como antígeno principal do parasita, além de
mucinas e transialidases. Estes glicopeptídeos e polipeptídeo desempenham funções importantes
na diferenciação, metabolismo, evasão da resposta imune e invasão das células hospedeiras
(SIQUEIRA-NETO et al., 2018). T. cruzi apresenta pelo menos duas famílias de genes
amplamente descritas, a transialidase e semelhante à mucina. Essas famílias são compostas por
cerca de 150 e 700 genes, respectivamente, e provavelmente são as proteínas mais abundantes na
superfície do parasita (CUEVAS; CAZZULO; SÁNCHEZ, 2003).

46
A menor a atividade gelatinolítica por parte do extrato de T. cruzi, quando comparado a
Leishmania, pode se dever a menor expressão de gp63 funcional, menor afinidade com o
substrato, além do fato de o protocolo usado seja de origem caseira, onde ainda encontra-se em
processo de otimização para a revelação de atividade enzimática (Figura 10).
Figura 10.Perfil enzimático, em gel de albumina, de extratos de Leishmania spp e T. cruzi; 1 -
L.amazonensis; 2 – L. donavani; 3 – L. guyanensis; 4 – L. infantum; 5 – L. major; 6 –Cl Brener; 7 – Dm28c; 8 – Y e
9 – Bolívia.
No gel de poliacrilamida copolimerizado com albumina foi possível observar uma grande
atividade proteolítica das espécies de Leishmania sobre o substrato (Figura 10). Não foi possível
verificar no gel as bandas de degradação de forma distinta, porém, é possível confirmar afinidade
dos extratos sobre o substrato de albumina. O BSA é uma cadeia polipeptídica simples de 583
resíduos aminoácidos. Possui estrutura muito similar a HSA (Human Serum Albumin),
apresentando sequencia idêntica de aminoácidos de 76% (FERREIRA, 2009). Essa similaridade
confirma a importância dessa endopeptidase nos diferentes estágios de vida do parasita e na
parasitemia no hospedeiro vertebrado.
Assim como na gelatina, no zimograma de albumina foi observado a presença de
degradação na parte superior do gel nas espécies de Leishmania donavani, L. guyanensis, L.
infantum e L. major, podendo ser a forma inativada da gp63, apresentando maior peso molecular
devido ao pró-peptídeo (LISBOA; AVEIRO, 2008). O extrato da espécie de Leishmania
amazonensis não apresentou atividade proteolítica frente à albumina, provavelmente ao fato dessa
espécie ter mais afinidade com substratos de colágeno e fibronectina. Isso pode ser correlacionado
pelo fato dessa espécie ser a principal causadora da leishmaniose cutânea, e reafirmado a atividade
proteolítica frente a substrato de gelatina e colágeno.

47
No que concerne a atividade enzimática de extratos de T. cruzi, conclui-se que não há
diferenças na expressão enzimática em função da forma morfológica do parasita, não ocorrendo
atividade proteolítica sobre a albumina. De fato, há muito poucos estudos sobre a detecção de
peptidases de T. cruzi usando gel de substrato albumina na literatura. Porém, alguns estudos
mostraram padrões de peptidase muito semelhantes entre diferentes cepas, sugerindo que muitas
dessas enzimas são conservadas entre as cepas (MEJÍA; PALÁU; ZÚÑIGA, 2004).
5.3. Atividade proteolítica in vitro de extrato bruto de Leishmania spp.
Trypanosoma cruzi sobre o substrato de Albumina.
Para iniciar o ensaio de atividade proteolitica in vitro, foi necessario otimizar o perfil
proteico do susbtrato de albumina. As amostras de até 20ug foram preparadas em tampão de
amostras, na proporção de 1:1, e realizado a eletroforese por SDS-PAGE, obtendo o perfil proteico
conforme mostra a figura 11.
Figura 11.Perfil de proteico, em gel de poliacrilamida (10%), de substrato Albumina sérica (BSA).
Marcador de pesos moleculares (M). Nos poços foram aplicadas diferentes concentrações de BSA: 1 – 5µg; 2 – 10µg;
3 – 15µg e 4 – 20µg.
A concentração de 10ug de BSA se mostrou nítida e suficiente para visualizar uma
possivel degradação, seja por parte da temperatura durante a incubação ou por ação direta com as
enzimas dos extratos brutos (Figura 11). Assim, foi a concentração escolhida para o estudo de
degradação in vitro.
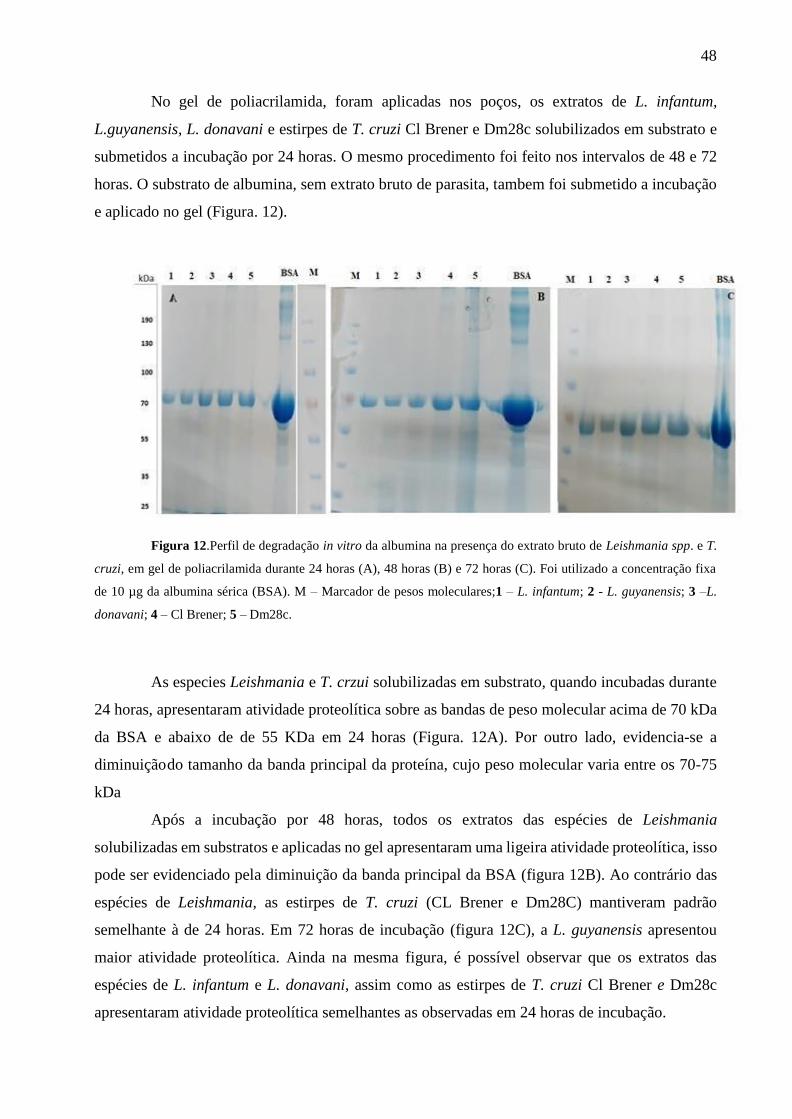
48
No gel de poliacrilamida, foram aplicadas nos poços, os extratos de L. infantum,
L.guyanensis, L. donavani e estirpes de T. cruzi Cl Brener e Dm28c solubilizados em substrato e
submetidos a incubação por 24 horas. O mesmo procedimento foi feito nos intervalos de 48 e 72
horas. O substrato de albumina, sem extrato bruto de parasita, tambem foi submetido a incubação
e aplicado no gel (Figura. 12).
Figura 12.Perfil de degradação in vitro da albumina na presença do extrato bruto de Leishmania spp. e T.
cruzi, em gel de poliacrilamida durante 24 horas (A), 48 horas (B) e 72 horas (C). Foi utilizado a concentração fixa
de 10 µg da albumina sérica (BSA). M – Marcador de pesos moleculares;1 – L. infantum; 2 - L. guyanensis; 3 –L.
donavani; 4 – Cl Brener; 5 – Dm28c.
As especies Leishmania e T. crzui solubilizadas em substrato, quando incubadas durante
24 horas, apresentaram atividade proteolítica sobre as bandas de peso molecular acima de 70 kDa
da BSA e abaixo de de 55 KDa em 24 horas (Figura. 12A). Por outro lado, evidencia-se a
diminuição do tamanho da banda principal da proteína, cujo peso molecular varia entre os 70-75
kDa
Após a incubação por 48 horas, todos os extratos das espécies de Leishmania
solubilizadas em substratos e aplicadas no gel apresentaram uma ligeira atividade proteolítica, isso
pode ser evidenciado pela diminuição da banda principal da BSA (figura 12B). Ao contrário das
espécies de Leishmania, as estirpes de T. cruzi (CL Brener e Dm28C) mantiveram padrão
semelhante à de 24 horas. Em 72 horas de incubação (figura 12C), a L. guyanensis apresentou
maior atividade proteolítica. Ainda na mesma figura, é possível observar que os extratos das
espécies de L. infantum e L. donavani, assim como as estirpes de T. cruzi Cl Brener e Dm28c
apresentaram atividade proteolítica semelhantes as observadas em 24 horas de incubação.

49
Na segunda etapa do ensaio, foram analisadas as espécies de L. major, L. amazonensis,
além das estirpes Bolívia e cepas Y de T. cruzi. Após 24 horas de incubação, a espécie de L. major
apresentou atividade proteolítica, ao contrário da espécie de L. amazonensis e estirpes Bolívia e
cepas Y frente ao substrato BSA (Figura. 13D).
Figura 13.Perfil de degradação in vitro da albumina na presença do extrato bruto de Leishmania spp. e T.
cruzi, em gel de poliacrilamida durante 24 horas (D), 48 horas (E) e 72 horas (F). Foi utilizado a concentração fixa de
10 µg da albumina sérica (BSA). M – Marcador de pesos moleculares. BSA; 1 Cepas Y; 2- Bolívia; 3- L.
amazonensis e 4- L. major.
Em 48 horas de incubação, a L. major apresentou um perfil de degradação semelhando
ao ensaio de incubação de 24 horas. Sendo nítida um aumento da degradação do substrato
dependente do tempo da incubação (Figura 13E). O extrato de L. amazonensis e L. major
apresentou um aumento da atividade proteolítica quando comparada ao perfil de degradação por
24 horas (Fig. 13D-E). As cepas Bolívia e Y apresentaram perfil de degradação.
A preparação e aplicação no gel do substrato de albumina solubilizando em tampão de
amostra sem os extratos de parasitas, foi uma alternativa encontrada para verificar se a degradação
do substrato seria causada pela ação da temperatura durante a incubação ou por ação direta das
enzimas parasitarias. Ainda conforme a figura 12 e 13, o substrato mostrou-se com uma banda
nítida, entre 60 a 66kDa, como observado por FERREIRA e colaboradores (2009). Além disso, a
evidência dessa banda elimina a hipótese de degradação do substrato por parte da temperatura.

50
A leishmaniose tegumentar pode se manifestar em três formas de expressão clínica:
cutânea, mucocutânea e cutânea difusa. A leishmaniose cutânea é a mais frequente e é
classicamente dividida nas formas do Novo e do Velho Mundo. No Brasil, as principais espécies
que causam a leishmaniose tegumentar são L. guyanensis e L. amazonensis, e no Velho Mundo é
causada por L. major (ARENAS et al., 2017).
A leishmaniose cutânea pode evoluir para cutânea difusa começando com pápulas
localizadas e não ulceradas, as amastigotas se disseminam pela pele e produzem nódulos cutâneos
na face e extremidades, sendo associada e L. amazonensis.
Essa grande variedade de espécies que podem causar a forma clinica cutânea da doença
mostra a intensa afinidade do parasita Leishmania com substratos de colágeno, gelatina e
fibronectina, em especial a L. amazonensis. Isso é confirmado pelos ensaios de zimografia com
substratos de gelatina e albumina presentes neste estudo, mostrando intensa atividade proteolítica
de todas as espécies de Leishmania frente a gelatina enquanto que frente ao substrato de albumina,
esses parasitas apresentaram atividade, mesmo que de forma menor ou nenhuma, como a L.
amazonensis.
Além da preferência por substrato de gelatina, a L. amazonensis infecta células no
tegumento, onde a temperatura é de aproximadamente 28°C, e durante o ensaio in vitro, a
temperatura foi de aproximadamente 37°C. Portanto, para além da afinidade pelo substrato, a
temperatura também se mostrou um fator determinante para que enzima apresentasse atividade
máxima (DA SILVA-LOPEZ; GIOVANNI-DE-SIMONE, 2004). Os extratos brutos das estirpes
de T. cruzi apresentaram pouca atividade proteolítica frente a substratos de gelatina e nenhuma
atividade proteolítica visível frente a substrato de albumina, no gel de Zimografia. Este resultado
não está em concordância com os resultados dos ensaios de degradação in vitro, que mostra que
estirpes de Y, Bolivia, Cl Brener e Dm28c apresentaram atividade sobre albumina. A zimografia
e SDS-PAGE são técnicas diferentes, onde a zimografia oferece condições para a renaturação da
proteína com diferentes alterações, no pH. Sendo assim, ensaios futuros com alterações no
protocolo, podem esclarecer a capacidade proteolítica frente a albumina, utilizando a zimografia.
Por outro lado, característica marcante do T. cruzi é a alta diversidade genética e fenotípica
inespecífica. Tal diversidade nos permite hipotetizar a causa da não atividade proteolitica frente
a substrato de albumina (ZINGALES, 2018).A diferença nos perfis proteicos e na atividade
enzimática frente aos substratos entre T. cruzi e Leishmania encontrados neste trabalho, sugere a
existência de múltiplos fatores como proteínas específicas da espécie, estágio morfofisiológico
das populações e sua origem geográfica, indicando uma complexidade no gene de expressão
desses parasitas (MEJÍA; PALÁU; ZÚÑIGA, 2004).

51
6. CONCLUSÃO E PESPERCTIVAS
O presente trabalho permitiu, por meio de ensaios de SDS-PAGE e zimografia, a obtenção
do perfil proteico e atividade proteolítica mediada por proteases presentes nos extratos brutos dos
tripanosomatídeos sobre o substrato gelatina e albumina, respectivamente.
Os perfis proteicos dos parasitas estudados são claramente distintos entre si, não sendo
possível identificar diferença entre as diferentes espécies de T. cruzi e Leishmania. O perfil de
Leishmania se mostrou mais detalhado, tendo uma maior existência de bandas entre 40-80 kDa,
isso é devido a alta capacidade do parasita de expressar a gp63. T. cruzi, porém, tem uma menor
capacidade de expressar glicoproteína com capacidade proteolítica.
A zimografia permitiu a observação da atividade proteolítica mediada por proteases
presentes nos extratos brutos dos tripanosomatídeos. Os resultados apresentam perfis diferentes, o
que aponta para a especificidade e afinidade da atividade enzimática sobre cada substrato. Em
substrato de gelatina, os extratos de Leishmania spp. demostraram a presença de enzimas
localizadas entre 50 - 80 kDa, sendo indicativo da glicoproteína mais abundante de superfície do
parasita, gp63, além de atividade entre 100 – 120kDa. Em relação T. cruzi observou-se atividade
proteolítica das cepas Cl Brener e Dm28c entre 50-60 kDa, enquanto as cepas Y e Bolivia
apresentaram atividade menos evidente. Essa atividade em gel de gelatina das cepas, muito
provavelmente refere-se a cisteína-protease cruzipaína, abundante na superfície das estirpes de
T.cruzi, além de outras glicoproteinas de superficie como trans- sialidase e mucinas.
Verificou-se atividade proteolítica para todas as espécies de Leishmania spp. em gel
de albumina, não sendo possível verificar as bandas de degradação de forma distinta. A
exceção ocorreu na espécie de L. amazonensis que não apresentou atividade. Em relação as
estirpes de T. cruzi, essas não apresentaram atividade frente albumina, em gel de zimografia,
provavelmente devido a diversidade fenotípica e afinidade com substrato, assim como as
condições do protocolo. O ensaio de degradação in vitro permitiu observar, por meio da
incubação a 37°C em diferentes intervalos de tempo, a atividade proteolítica dos extratos
brutos de Leishmania e T. cruzi. Das espécies estudadas nesse ensaio, todas as espécies de
Leishmania e T. cruzi apresentaram atividade, sendo a degradação do substrato mais perceptivel
nos extratos de Leishmania.
Devido a importância das funções bioquímicas e biológicas dessas proteases de superfície
frente ao hospedeiro, a caracterização dos mecanismos biológicos de infecção pode constituir
uma oportunidade para a identificação de novos alvos terapêuticos e novos fármacos.

52
Poranto, é necessário continuar os estudos utilizando uma ampla gama de isolados de
parasitas, além de outros substratos proteicos que nos permita identificar a afinidade de cada
parasita. Além de uma análise de bioinformática, por meio do docking molecular, que nos
permita identificar a atração dos diferentes aminoácidos envolvidos na interação parasita-
hospedeiro ou parasita-substrato.

53
7. REFERÊNCIAS BIBLIOGRÁFICAS
1. ARENAS, R. et al. Leishmaniasis: A review. F1000Research, v. 6, n. May, p. 1–15, 2017.
2. BOELAERT, M. et al. Cost-effectiveness of competing diagnostic-therapeutic strategies for
visceral leishmaniasis. Bulletin of the World Health Organization, v. 77, n. 8, p. 667–674, 1999.
3. BUNNEY, P. E. et al. Orexin activation counteracts decreases in nonexercise activity
thermogenesis (NEAT) caused by high-fat diet. Physiology & Behavior, v. 176, n. 3, p. 139–148,
jul. 2017.
4. BURZA, S.; CROFT, S. L.; BOELAERT, M. Leishmaniasis. The Lancet, v. 392, n. 10151, p.
951–970, 2018.
5. CALIXTO, P. H. M.; FAGUNDES, D. S.; OLIVEIRA, J. C. S. Estrutura Tridimensional da Major
Surface Protease de Leishmania guyanensis Resolvida por Modelagem Comparativa. Biota
Amazônia, v. 4, n. 1, p. 74–80, 2014.
6. CARVALHO, A. M. et al. Immunologic response and memory T cells in subjects cured of
tegumentary leishmaniasis. BMC Infectious Diseases, v. 13, n. 1, 2013.
7. CECÍLIO, P. et al. Deception and manipulation: The arms of Leishmania, a successful parasite.
Frontiers in Immunology, v. 5, n. OCT, p. 1–16, 2014.
8. CERNY, N. et al. Cruzipain and Its Physiological Inhibitor, Chagasin, as a DNA-Based
Therapeutic Vaccine against Trypanosoma cruzi. Frontiers in Immunology, v. 11, n. October,
p. 1–15, 2020.
9. CHAGAS, A. P. et al. Aspectos ecológicos da fauna de flebotomíneos em focos de leishmaniose
na Amazônia Oriental, Estado do Pará, Brasil. Revista Pan-Amazônica de Saúde, v. 7, n. esp, p.
123–132, 2016.

54
10. CUEVAS, I. C.; CAZZULO, J. J.; SÁNCHEZ, D. O. gp63 homologues in Trypanosoma cruzi:
Surface antigens with metalloprotease activity and a possible role in host cell infection. Infection
and Immunity, v. 71, n. 10, p. 5739–5749, 2003.
11. CUI, N.; HU, M.; KHALIL, R. A. Biochemical and Biological Attributes of Matrix
Metalloproteinases. Progress in Molecular Biology and Translational Science, v. 147, n. 617,
p. 1–73, 2017.
12. DA COSTA FILHO, A. V.; LUCAS, Í. C.; SAMPAIO, R. N. R. Estudo comparativo entre
miltefosina oral e antimoniato de N-metil glucamina parenteral no tratamento da leishmaniose
experimental causada por Leishmania (Leishmania) amazonensis. Revista da Sociedade
Brasileira de Medicina Tropical, v. 41, n. 4, p. 424–427, 2008.
13. DA SILVA-LÓPEZ, R. E. Proteases de leishmania: Novos alvos para o desenvolvimento racional
de fármacos. Quimica Nova, v. 33, n. 7, p. 1541–1548, 2010.
14. DE MAGALHÃES, A. V et al. Histopathology of cutaneous leishmaniasis caused by Leishmania
braziliensis. Padroes histológicos e estudo evolutivo das lesoes. Revista do Instituto de Medicina
Tropical de São Paulo, v. 28, n. 6, p. 421–30, 1986.
15. DE MEDEIROS, N. INGRID; GOMES, J. DE A. S.; FARES, R. C. G. Detecção E Quantificação
Da Atividade Enzimática Das Metaloproteinases E De Seus Inibidores Através Da Técnica De
Zimografia. Pós em Revista do Centro Universitário Newton Paiva, v. 2, n. 6, p. 1–4, 2012.
16. DIAS, J. C. P. et al. Aspectos Gerais da Epidemiologia da Doença de Chagas com Especial
Atenção ao Brasil. Epidemiologia e servicos de saude : revista do Sistema Unico de Saude do
Brasil, v. 25, n. spe, p. 7–86, 2016a.
17. DIAS, J. C. P. et al. Mudanças no paradigma da conduta clínica e terapêutica da doença de Chagas:
avanços e perspectivas na busca da integralidade da saúde. Epidemiologia e servicos de saude :
revista do Sistema Unico de Saude do Brasil, v. 25, n. spe, p. 87–90, 2016b.
18. DIAS, J. C. P. Notas sobre o Trypanosoma cruzi e suas características bio-ecológicas, como agente
de enfermidades transmitidas por alimentos. Revista da Sociedade Brasileira de Medicina

55
Tropical, v. 39, n. 4, p. 370–375, 2006.
19. DIAS, L. C. et al. Doenças tropicais negligenciadas: uma nova era de desafios e oportunidades.
Quimica Nova, v. 36, n. 10, p. 1552–1556, 2013.
20. FERREIRA, E. S. Interação da albumina do soro bovino (BSA) com substratos sintéticos. p. 137,
2009.
21. FERREIRA, M. F. Extração E Caracterização De Gelatina Proveniente De Subprodutos Do
Frango : Pés. p. 40, 2013.
22. GALVAN et al. We are IntechOpen , the world ’ s leading publisher of Open Access books Built
by scientists , for scientists TOP 1 %. In: Intech. [s.l: s.n.]. v. ip. 13.
23. GONTIJO, C. M. F.; MELO, M. N. Leishmaniose visceral no Brasil: quadro atual, desafios e
perspectivas. Revista Brasileira de Epidemiologia, v. 7, n. 3, p. 338–349, 2004.
24. GOTO, H.; LINDOSO, J. A. L. Current diagnosis and treatment of cutaneous and mucocutaneous
leishmaniasis. Expert Review of Anti-Infective Therapy, v. 8, n. 4, p. 419–433, 2010.
25. HASSANI, K. et al. Absence of metalloprotease GP63 alters the protein content of leishmania
exosomes. PLoS ONE, v. 9, n. 4, 2014.
26. HOTEZ, P. J. et al. World neglected tropical diseases day. PLoS neglected tropical diseases, v.
14, n. 1, p. e0007999, 2020.
27. HU, X.; BEETON, C. Detection of functional matrix metalloproteinases by zymography. Journal
of Visualized Experiments, n. 45, p. 1–5, 2010.
28. ISNARD, A.; SHIO, M. T.; OLIVIER, M. Impact of Leishmania metalloprotease GP63 on
macrophage signaling. Frontiers in cellular and infection microbiology, v. 2, n. May, p. 72,
2012.
29. JANSEN, A. M.; XAVIER, S. C. D. C.; ROQUE, A. L. R. Trypanosoma cruzi transmission in the

56
Wild and its most important reservoir hosts in Brazil. Parasites and Vectors, v. 11, n. 1, p. 1–
25, 2018.
30. JENNINGS, Y. L. et al. Phenotypic characterization of Leishmania spp. causing cutaneous
leishmaniasis in the lower Amazon region, western Pará state, Brazil, reveals a putative hybrid
parasite, Leishmania (Viannia) guyanensis × Leishmania (Viannia) shawi shawi. Parasite, v. 21,
2014.
31. KARUNAWEERA, N. D.; FERREIRA, M. U. Leishmaniasis: Current challenges and prospects
for elimination with special focus on the South Asian region. Parasitology, v. 145, n. 4, p. 425–
429, 2018.
32. LAMBERTZ, U. et al. Secreted virulence factors and immune evasion in visceral leishmaniasis.
Journal of Leukocyte Biology, v. 91, n. 6, p. 887–899, 2012.
33. LIEKE, T. et al. Leishmania surface protein gp63 binds directly to human natural killer cells and
inhibits proliferation. Clinical and Experimental Immunology, v. 153, n. 2, p. 221–230, 2008a.
34. LIEKE, T. et al. Leishmania surface protein gp63 binds directly to human natural killer cells and
inhibits proliferation. Clinical and Experimental Immunology, v. 153, n. 2, p. 221–230, 2008b.
35. LISBOA, U. N. D. E.; AVEIRO, U. D. E. UNIVERSIDADE NOVA DE LISBOA Instituto de
Higiene e Medicina Tropical. p. 0–4, 2008.
36. MA, L. et al. An evolutionary analysis of trypanosomatid GP63 proteases. Parasitology
Research, v. 109, n. 4, p. 1075–1084, 2011.
37. MALAFAIA, G. A importância do encorajamento de estudos sobre as doenças tropicais
negligenciadas. Revista da Sociedade Brasileira de Medicina Tropical, v. 42, n. 5, p. 609–610,
2009.
38. MALAFAIA, G.; DE LIMA RODRIGUES, A. S. Centenário do descobrimento da doença de
chagas: Desafios e perspectives. Revista da Sociedade Brasileira de Medicina Tropical, v. 43,
n. 5, p. 483–485, 2010.

57
39. MARTINEZ, S. J.; ROMANO, P. S.; ENGMAN, D. M. Precision Health for Chagas Disease:
Integrating Parasite and Host Factors to Predict Outcome of Infection and Response to Therapy.
Frontiers in Cellular and Infection Microbiology, v. 10, n. May, p. 1–11, 2020.
40. MATTE, C. et al. Leishmania major Promastigotes Evade LC3-Associated Phagocytosis through
the Action of GP63. PLoS Pathogens, v. 12, n. 6, p. 1–17, 2016.
41. MESSENGER, L. A.; MILES, M. A.; BERN, C. Between a bug and a hard place: Trypanosoma
cruzi genetic diversity and the clinical outcomes of Chagas disease. Expert Review of Anti-
Infective Therapy, v. 13, n. 8, p. 995–1029, 2015.
42. MITRA, A. K.; MAWSON, A. R. Neglected tropical diseases: Epidemiology and global burden.
Tropical Medicine and Infectious Disease, v. 2, n. 3, 2017.
43. NAPOLI, S. et al. Functional Roles of Matrix Metalloproteinases and Their Inhibitors in
Melanoma. Cells, v. 9, n. 5, p. 1–23, 2020.
44. PÉREZ-MOLINA, J. A.; MOLINA, I. Chagas disease. The Lancet, v. 391, n. 10115, p. 82–94,
2018.
45. PERSOON, M. C. et al. An unusual case of congestive heart failure in the Netherlands. JMM
Case Reports, v. 5, n. 4, p. 1–6, 2018.
46. PINHEIRO, F. G.; LUZ, S. L. B.; FRANCO, A. M. R. Infecção natural por tripanosomatídeos
(Kinetoplastida: Trypanosomatidae) em Lutzomyia umbratilis (Diptera: Psychodidae) em áreas de
leishmaniose tegumentar americana no Amazonas, Brasil. Acta Amazonica, v. 38, n. 1, p. 165–
172, 2008.
47. POULOSE. 乳鼠心肌提取 HHS Public Access. Physiology & behavior, v. 176, n. 1, p. 139–
148, 2016.
48. REBELLO, K. M. et al. Participation of Trypanosoma cruzi gp63 molecules on the interaction
with Rhodnius prolixus. Parasitology, v. 146, n. 8, p. 1075–1082, 2019.

58
49. RIBEIRO, A. L.; ROCHA, M. O. Indeterminate form of Chagas disease: considerations about
diagnosis and prognosis. Revista da Sociedade Brasileira de Medicina Tropical, v. 31, n. 3, p.
301–314, 1998.
50. SÁDLOVÁ, J. et al. Virulent and attenuated lines of Leishmania major: DNA karyotypes and
differences in metalloproteinase GP63. Folia Parasitologica, v. 53, n. 2, p. 81–90, 2006.
51. SANTOS, A. L. S.; BRANQUINHA, M. H.; D’AVILA-LEVY, C. M. The ubiquitous gp63-like
metalloprotease from lower trypanosomatids: In the search for a function. Anais da Academia
Brasileira de Ciencias, v. 78, n. 4, p. 687–714, 2006.
52. SCHNAPP, A. R. et al. Cruzipain induces both mucosal and systemic protection against
Trypanosoma cruzi in mice. Infection and Immunity, v. 70, n. 9, p. 5065–5074, 2002.
53. SILVA, J. A. O. et al. Sandflies in an endemic area for visceral Leishmaniasis in Northeastern
Brazil. Revista Brasileira de Parasitologia Veterinaria, v. 28, n. 4, p. 569–573, 2019.
54. SIMÕES, M. V. et al. Cardiomiopatia da Doença de Chagas. International Journal of
Cardiovascular Sciences, v. 31, n. 2, p. 173–189, 2018.
55. SIQUEIRA-NETO, J. L. et al. Cysteine proteases in protozoan parasites. PLoS Neglected
Tropical Diseases, v. 12, n. 8, p. 1–20, 2018.
56. SNOEK-VAN BEURDEN, P. A M. et al. Técnicas de zimografía para el análisis de
metaloproteasas de matriz y sus inhibidores. BioTechniques, v. 38, n. 1, p. 73–83, 2005.
57. SUNDAR S, R. M. Laboratory diagnosis of visceral leishmaniasis. Clin Diagn Lab Immunol.
Clinacal and Diagnostic Laboratory Immunology, v. 9, n. 9, p. 951–8, 2002.
58. TAFURI, W. L. Pathogenia Da Doenca De ChagasRevista do Instituto de Medicina Tropical
de Sao Paulo, 1987.
59. TAJHYA, R. B.; PATEL, R. S.; BEETON, C. Detection of matrix metalloproteinases by
zymography. Methods in Molecular Biology, v. 1579, p. 231–244, 2017.

59
60. WATANABE COSTA, R.; DA SILVEIRA, J. F.; BAHIA, D. Interactions between Trypanosoma
cruzi secreted proteins and host cell signaling pathways. Frontiers in Microbiology, v. 7, n. MAR,
p. 1–9, 2016.
61. WILKESMAN, J. Zymography. Methods in Molecular Biology, v. 1626, n. 617, p. 25–31, 2017a.
62. WILKESMAN, J. Zymography. Methods in Molecular Biology, v. 1626, p. 25–31, 2017b.
63. YAO, C. Major surface protease of trypanosomatids: One size fits all. Infection and Immunity,
v. 78, n. 1, p. 22–31, 2010.
64. YOSHIDA, N. Molecular mechanisms of Trypanosoma cruzi infection by oral route. Memorias
do Instituto Oswaldo Cruz, v. 104, n. SUPPL. 1, p. 101–107, 2009.
65. ZINGALES, B. et al. The revised Trypanosoma cruzi subspecific nomenclature: Rationale,
epidemiological relevance and research applications. Infection, Genetics and Evolution, v. 12,
n. 2, p. 240–253, 2012.
66. ZINGALES, B. Trypanosoma cruzi genetic diversity: Something new for something known about
Chagas disease manifestations, serodiagnosis and drug sensitivity. Acta Tropica, v. 184, p. 38–
52, 2018.
67. ZINGALES, B. Trypanosoma cruzi: um parasita, dois parasitas ou vários parasitas da doença de
chagas? Revista da Biologia, v. 6b, p. 44–48, 2011.