enzimas
DESCRIPTION
TRANSCRIPT

ENZIMAS

Las enzimas son proteínas Catalizan reacciones químicas necesarias
para la sobrevivencia celular Sin las enzimas los procesos biológicos
serían tan lentos que las células no podrían existir.
Las enzimas pueden actuar dentro de la célula , fuera de ésta, y en el tubo de ensayo.
E + SE + S ES ESEP EP E + P E + P
E E E E
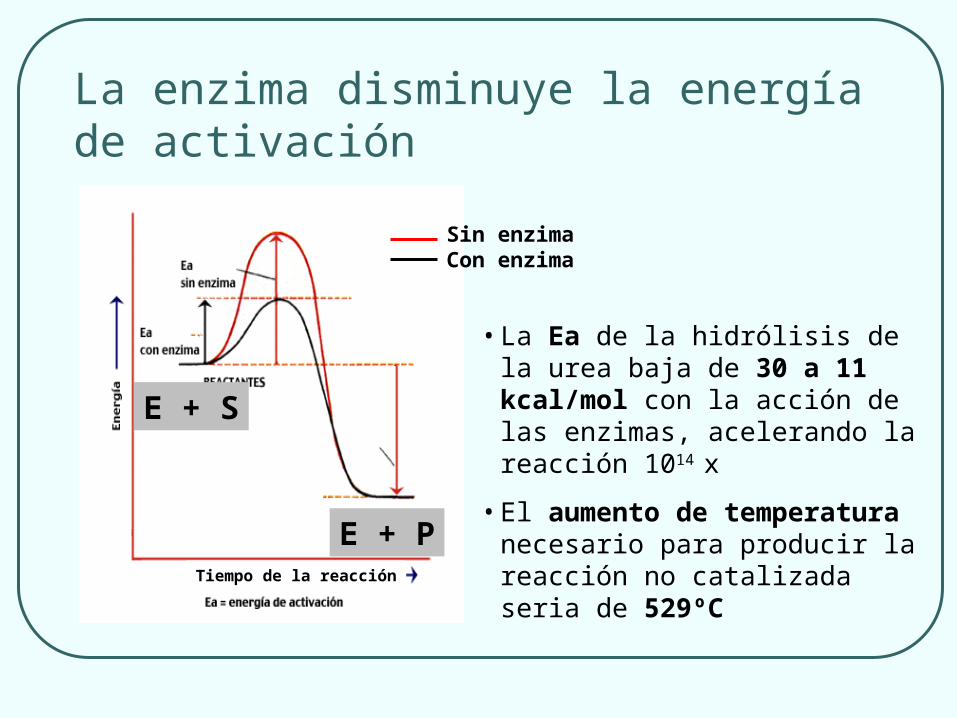
La enzima disminuye la energía de activación
Tiempo de la reacción
E + S
E + P
Sin enzimaCon enzima
• La Ea de la hidrólisis de la urea baja de 30 a 11 kcal/mol con la acción de las enzimas, acelerando la reacción 1014 x
• El aumento de temperatura necesario para producir la reacción no catalizada seria de 529ºC

Enzima - Catalizador Tanto la enzima como el catalizador aceleran
la velocidad de una reacción química. Una enzima puede transformar 1000
moléculas de sustrato/ segundo Las enzimas tienen 3 propiedades que los
catalizadores NO tienen• Especificidad por el sustrato
• Se inactivan por desnaturación
• Pueden ser reguladas


Las enzimas se unen a los reactivos (sustratos) reduciendo la energía de activación
Cada enzima tiene una forma única con un sitio o centro activo en el que se une al sustrato
Después de la reacción, enzimas y productos se separan.
Las moléculas enzimáticas no han cambiado después de participar en la reacción

Las enzimas cumplen su papel catalítico gracias a:• Fijación estereoquímicamente complementaria
del substrato• Transformación catalítica del mismo
En ambas funciones participan:
• Cadenas laterales de los aminoácidos• Grupos o moléculas no proteicas:
Grupos prostéticos Iones metálicos Cofactores

Los siguientes hechos: Especificidad de la reacción enzimática Carácter heterogéneo de la catálisis enzimática
Nos llevan a postular la existencia de un Centro Activo en la molécula de enzima, capaz de:
Fijar específicamente al substrato Transformarlo catalíticamente.
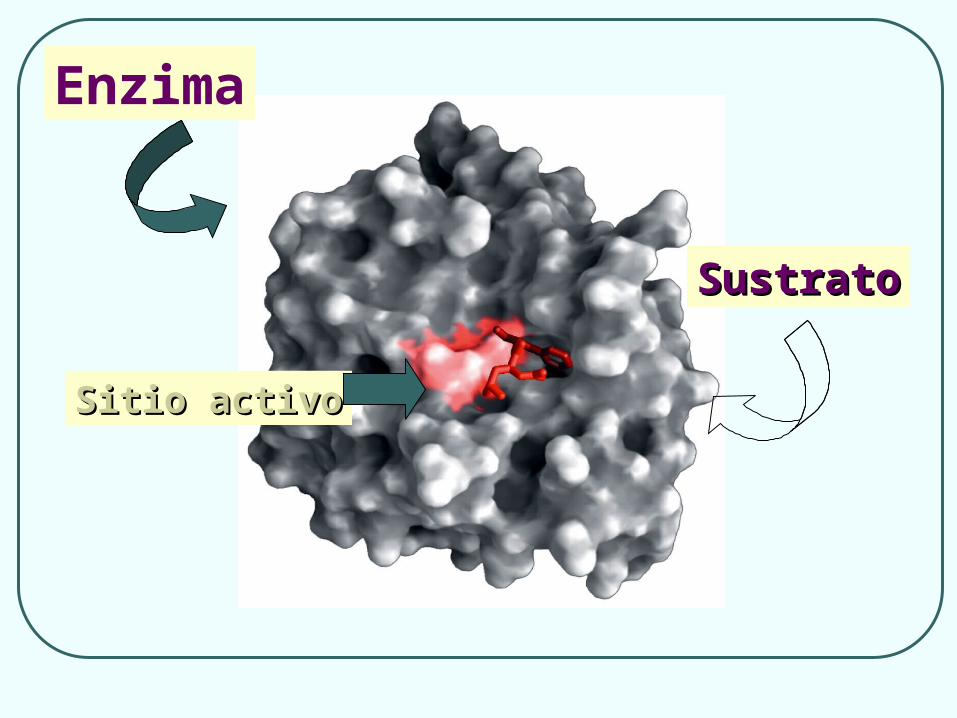
Enzima
Sitio activoSitio activo
SustratoSustrato

Complementariedad geométrica
Complementariedad de cargas, uniones iónicas
Modelos:
Encaje inducido
Llave – cerradura.
Estado de transición
La unión del sustrato es muy específica

Teorías de la acción enzimática, 1
Modelo de Llave y CerraduraModelo de Llave y Cerradura (Emil Fischer) (Emil Fischer)
Substrato y enzima se acoplan de forma estereospecífica,de la misma manera que una llave se ajusta a su cerradura.
Modelo aceptado durante mucho tiempo; hoy se considera insuficiente al no explicar algunos fenómenos de la inhibición enzimática

Teorías de la acción enzimática, 2
Modelo de Ajuste InducidoModelo de Ajuste Inducido (Koshland) (Koshland)
Tanto la enzima como el substrato sufren una alteración en su estructura por el hecho físico de la unión.
Está mucho más de acuerdo con todos los datos experimentales conocidos hasta el momento.

Teorías de la acción enzimática, 3
La teoría del Ajuste Inducido se amplía en la actualidaddefiniendo la acción enzimática como
Estabilización del Estado de TransiciónEstabilización del Estado de Transición
Según lo cual, el Centro Activo enzimático es en realidadcomplementario no al substrato o al producto, sino alestado de transición entre ambos.

Una enzima puede unir dos sustratos en su sitio activo

Clasificación y nomenclatura de enzimas

EC 2.7.1.1
Número Enzyme Commission:
EnzymeComission
Grupo Subgrupo
Nombre común (sustrato+”asa”): Hexokinasa
Clasificación y nomenclatura
ATP: hexosa fosfotransferasa
Nombre sistemático:
Donador Aceptor
Grupo transferido
Tipo de reacción catalizada
Grupos químicos
Enzimas

EC 1.x OxidorreductasasEC 2.x TransferasasEC 3.x HidrolasasEC 4.x LiasasEC 5.x IsomerasasEC 6.x Ligasas
Clasificación de enzimas por Grupos
Clasificación y nomenclatura

Grupo 1: OxidorreductasasCatalizan reacciones de oxidorreducción, en que átomos de oxigeno ó hidrogeno son trasladados entre moléculas:
En las reacciones redox, siempre tienen que estar presentes a la vez el aceptor y el dador electrónico.
AH2 + B A + BH2
Clasificación y nomenclatura
Ared + Box Aox + Bred

Nombre: Dador:Aceptor oxidorreductasa
Nombre común:Glucosa oxidasa
-D-Glucosa : O2 1-oxidorreductasa
Dador Aceptor
EC 1.1.3.4
Clasificación y nomenclatura
Grupo 1: Oxidorreductasas

Nomenclatura del subgrupo en oxidorreductasas:
EC 1.1.x - DeshidrogenasasEC 1.2.x - OxidasasEC 1.3.x - PeroxidasasEC 1.4.x - OxigenasasEC 1.5.x - HidroxilasasEC 1.6.x - Reductasasetc.
Clasificación y nomenclatura
Grupo 1: Oxidorreductasas
Aplicaciones: Ensayos de diagnostico clínico (glucosa oxidasa y colesterol oxidasa
Deslignificación ó Bioblanqueamiento

A-X + B A + B-X
Grupo 2: Transferasas
Catalizan reacciones de transferencia de átomos ó grupo de átomos entre moléculas:
Dador: Aceptor Grupo transferido - transferasa
ATP: D-Hexosa Fosfotransferasa EC 2.7.1.1Nombre común: hexokinasa
Clasificación y nomenclatura

Clasificación de subgrupo de las transferasas:EC 2.1.x - Grupos monocarbonadosEC 2.2.x - Grupos aldehido o cetoEC 2.3.x - AciltransferasasEC 2.4.x - GlicosiltransferasasEC 2.5.x - Alquil- o AriltransferasasEC 2.6.x - Grupos nitrogenadosEC 2.7.x - Grupos fosfatoEC 2.8.x - Grupos sulfatoEC 2.9.x - Grupos selenio
Clasificación y nomenclatura
Grupo 2: Transferasas
Aplicaciones: Síntesis de oligosacáridos

Grupo 3: Hidrolasas
Catalizan reacciones de hidrólisis y también su reverso. Son las más comunes en el dominio de la tecnología enzimática:
A-B + H2O A-OH + H-B
No se suelen utilizar nombres sistemáticos en lashidrolasas. Muchas de ellas conservan el nombrePrimitivo. Ejemplo: Quimosina EC 3.4.23.4.
Clasificación y nomenclatura

Clasificación de las hidrolasas:
3.1.-.- Esterasas (carboxilesterasas, fosfoesterasas, sulfoesterasas)3.2.-.- Glicosidasas3.3.-.- Éter hidrolasas3.4.-.- Péptido hidrolasas3.5.-.- Acil anhídrido hidrolasasetc.
Grupo 3: Hidrolasas
Clasificación y nomenclatura
Aplicaciones: Lipasas → Síntesis de tensioactivos
Proteasas → Fabrico de quesos
Glicosidasas → Clarificación de jugos; liberación de aromas en los vinos; aplicaciones textiles

Caso particular Péptido hidrolasas: clasificación común (no sistemática)
I. Según la situación del enlace atacado:
- Exopeptidasas (extremos de la cadena) (Peptidasas)- Endopeptidasas (interior de la cadena) (Proteinasas)
II. Según el mecanismo catalítico:
- Serin proteinasas- Tiol proteinasas- Aspartil proteinasas- Metaloproteinasas
Grupo 3: Hidrolasas
Clasificación y nomenclatura

Grupo 4: LiasasCatalizan reacciones reversibles de remoción de grupo de átomos del sustrato (este grupo no incluye las hidrolasas):
A-B A + B
Clasificación y nomenclatura
EjemploNombre sistemático: Histidina amonio-liasa (EC 4.3.1.3)
Nombre común: Histidasa

Clasificación de las liasas:
4.1.x - Actúan sobre enlaces C-C4.2.x - Actúan sobre enlaces C-O4.3.x - Actúan sobre enlaces C-N4.4.x - Actúan sobre enlaces C-S4.5.x - Actúan sobre enlaces C-Haluro (S-, Cl-, Br-, I- At-)4.6.x - Actúan sobre enlaces P-O4.99.x - Otras liases
Clasificación y nomenclatura
Aplicaciones: Pectato liasa – Remueve los compuestos indeseables (ceras, pectinas, proteínas) en fibras en la industria textil – “bioscouring”
Grupo 4: Liasas

Grupo 5: Isomerasas
Catalizan reacciones de isomerización moleculares
Clasificación y nomenclatura
A B
Ejemplo:
Glucosa isomerasa (EC 5.3.1.5)

5.1.x - Rasemasas y Epimerasas5.2.x - cis-Trans-Isomerasas5.3.x - Oxidoreductasas Intramolecular5.4.x - Transferasas Intramoleculares (mutases)5.5.x - Liasas Intramoleculares5.99.x - Otras Isomerasas
Grupo 5: Isomerasas
Clasificación y nomenclatura
Clasificación de las isomerasas:

Grupo 6: Ligasas
Catalizan la unión de dos grupos químicos a expensasde la hidrólisis de un nucleótido trifosfato (ATP, GTP, etc.).
A + B + ATP A-B + ADP + Pi
Clasificación y nomenclatura
Ejemplo: Glutationa sintasa (EC 6.2.2.3)
Nombre sistémico:G-L-glutamilo-L-cisteina:glicina ligase

Grupo 6: Ligasas
Clasificación y nomenclatura
Clasificación de las ligasas:
6.1.x - Forman enlaces C-O6.2.x - Forman enlaces C-S6.3.x - Forman enlaces C-N6.4.x - Forman enlaces C-C6.5.x - Forman enlaces ésteres fosfóricos6.6.x - Actúan sobre enlaces N-metal

Cinética Enzimática

Cinética Enzimática La cinética enzimática es el análisis cuantitativo del efecto de
cada uno de los factores que intervienen en la actividad enzimática, que se evalúa a través de la velocidad de la reacción catalizada.
Las variables más importantes son:
• Concentración de enzima, sustratos y productos (incluyendo inhibidores y/o activadores)
• pH
• Temperatura

Ella está basada en la medición de la velocidad de reacción. Así en la reacción:
Se tendrá:
Como se observa en la figura, la velocidad de reacción (pendiente) disminuye continuamente a partir de un valor máximo inicial. Esto se puede deber a:
Desaturación de la enzima con sustrato, por disminución de la concentración del sustrato
Inactivación de la enzima por su inherente inestabilidad
Inhibición por el producto
Desplazamiento del equilibrio, si la reacción es reversible
dt
ds
dt
dpv
ES P

Baja concentración de sustrato
ALTA concentración de sustratoSATURACION

v
[s]
Efecto de la concentración de substrato
..
.. . . . .

dt
t
s
p[ ]
Concepto de velocidad inicial
d[P]v = , t 0

E + S ES E + Pk+1
k-1
k+2
Una cinética hiperbólica implica un proceso saturante:
Hay un número limitado de sitios en la enzimapara fijar substrato; una vez que están ocupadostodos, por mucho que aumente la concentraciónde substrato, la velocidad permanecerá constantetendiendo a un valor asintótico

El sistema de ecuaciones diferenciales que describe la dinámicadel sistema
E + S ES E + Pk+1
k-1
k+2
Sistema No admite solución analítica.
- Simulaciones numéricas- Hipótesis que lo simplifiquen

Hipótesis de Michaelis - MentenHipótesis de Michaelis - Menten
E + S ES E + Pk+1
k-1
k+2
- La primera parte del mecanismo,
E + S ESk+1
k-1Tiene lugar mucho más rápidamente que la segunda:
ES E + Pk+2

K
E SE S
esx
e x s
xm 0
xe s
K sm
0
vk e sK sm
2 0
k e V 2 0 m ax
vV sK sm
m ax
Hipótesis deMichaelis-Menten
Equilibrio rápido
v k x 2

Hipótesis de estado estacionarioHipótesis de estado estacionario
Según veíamos en la simulación numérica, hay un tiemporelativamente prolongado en el curso de la reacción en el que la variación de complejo [ES] es prácticamente iguala cero:
dx/dt = k+1es - (k-1 + k+2) x = 0
A partir de esta expresión podemos llegar a obtener unaexpresión que nos da la velocidad para la concentraciónde substrato

dxd t
k es k k x 0 1 1 2
k es k e x s k k x 1 1 0 1 2
xe s
k kk
s
e sK sm
0
1 2
1
0
vV sK sm
m ax
v k x 2
k e V 2 0 m ax
Hipótesis deestado estacionario

A partir de las suposiciones de Michaelis-Menten y Estado estacionario, llegamos a la ecuación
Km + sv =
Vmax * s
La única diferencia radica en el significado de Km:
Km = k-1/k+1 en mecanismo de M.M.
Km = (k-1 + k+2)/k+1 en mecanismo de E.E.

Significado de la constante Km
1. Constante de equilibrio de disociación del complejo ES (en condiciones de equilibrio rápido)
2. Medida inversa de la afinidad de la enzima por el substrato (en condiciones de equilibrio rápido)
3. Mide la función de fijación (en cond.de equilibrio rápido)
4. Concentración de substrato para la que la velocidad se hace igual a la mitad de la máxima (s0.5)
5. Se define para una pareja enzima-substrato
6. Se mide en unidades de concentración

Significado de la constante Vmax = k+2 e0
1. Velocidad asintótica para s
2. Directamente proporcional a la concentración de enzima (hoy se prefiere caracterizar a la enzima por k+2)
3. Mide función de transformación catalítica
4. Se expresa en unidades de velocidad

Relación entre Km y VmaxMichaelis y Menten
Concentración de Sustrato [S]
Vel
oci
dad
de
la r
eacc
ión
(v)
Km
Vmax
Vmax/2
La Km es la concentración de sustrato donde se obtiene la mitad de la Vmax

Concentración de Sustrato [S]
Vel
oci
dad
de
la r
eacc
ión
(v)
Km
Vmax
Vmax/2
A mayor Km, menor es la afinidad de la enzima por el sustratoA menor Km mayor es la afinidad de la enzima por el sustrato


Representación directa
s
0 20 40 60 80 100
v
0
20
40
60
80
100
Km
Vmax
vV s
K sm x
m
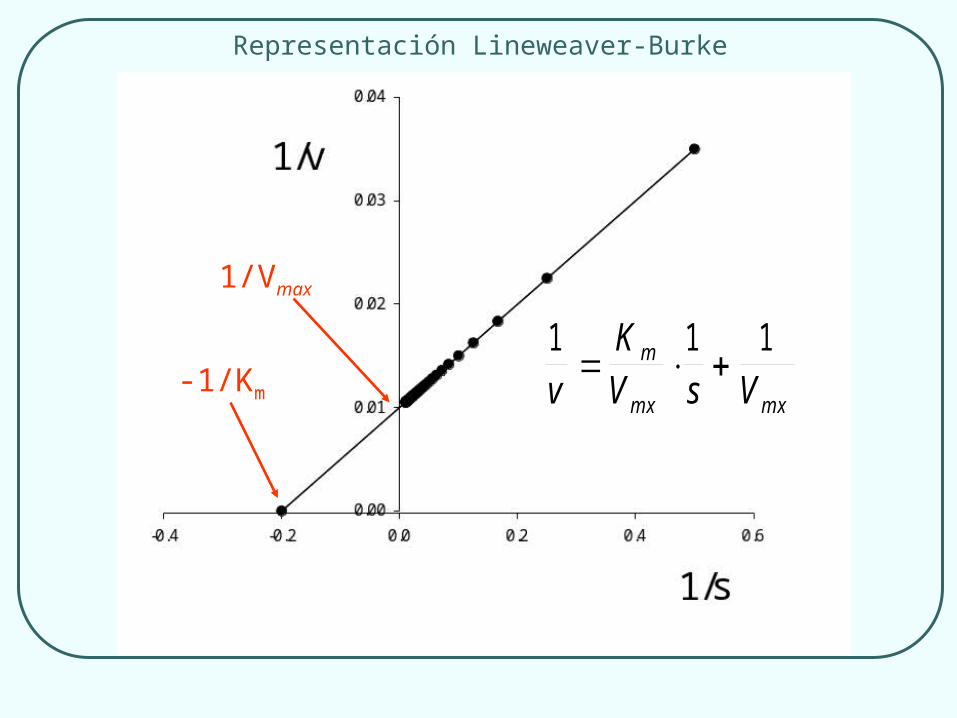
1 1 1v
KV s V
m
m x m x
-1/Km
1/Vmax
Representación Lineweaver-Burke

sv V
sKVm x
m
m x
1
-Km
1/V max
Representación Hanes

v Kvs
Vm m x
Vmax
-Km
Representación de Eadie-Hofstee

Métodos de linealización para determinación parámetros cinéticos
Método Eje Y Eje X Intercepto
Eje Y
Intercepto
Eje x
Pendiente
Lineweaver-Burke
1/v 1/s 1/V -1/K K/V
Hanes s/v s K/V -K 1/V
Eadie-Hofstee
v v/s V V/K -K

Definición Actividad Enzimática
Es el número de moles de sustrato que reaccionan para formar producto, por mol de enzima y por unidad de tiempo. Esto supone que la enzima está plenamente saturada con sustrato y por tanto que la reacción se efectúa con su máxima rapidez
El índice de recambio muestra la eficiencia impresionante de la catálisis enzimática

A fin de normalizar la medición de la actividad, se ocupa el valor de la velocidad inicial de reacción, donde eses factores son despreciables. Así,
Esta velocidad inicial no se espera que sea observada en un proceso enzimático donde, debido a los elevados tiempos de operación, es razonable suponer que esos factores ejercerán su influencia.
000
ttt dt
ds
dt
dpva

Cálculo de Actividad Enzimática
La velocidad de reacción catalizada por 0,1ml de una dilución 1:100 de Fosfatasa Alcalina es 0,3umoles / min. de producto. ¿Cuál es su actividad enzimática?
0,1ml-------- 0,3umoles producto / min. 1,0ml------------3,0umoles producto / min. dil 1:100-------------300umoles producto / min. Respuesta: 300U/ml

Número o Índice de Recambio
Es útil definir una constante de velocidad más general, k cat , para describir la velocidad limitante de cualquier reacción enzimática a saturación
kcat x Et= Vmáx
V0 = k cat x Et x [S] / KM + [S];
Kcat es una constante de velocidad de 1er orden con unidades de tiempo-1, también llamada Número de Recambio

Efecto del pH en la ENZIMAEfecto del pH en la ENZIMA
Cada enzima tiene un pH óptimo para su actividadCada enzima tiene un pH óptimo para su actividadEl pH afecta las interacciones iónicasEl pH afecta las interacciones iónicas

1. Sobre la fijación del substrato al centro activo:- Grupos disociables de la enzima- Grupos disociables del substrato
2. Sobre la transformación catalítica del substrato
3. Sobre la estructura de la proteína enzimática
Efecto del pH

En el efecto de la temperatura hay dos componentes:
1. Aceleración de la reacción según la ecuación de Arrhenius
k = A exp (-Ea/RT)
2. Desnaturalización térmica de la proteína

Efecto de la temperatura en la ENZIMAEfecto de la temperatura en la ENZIMA
Cada enzima tiene una temperatura óptima.Cada enzima tiene una temperatura óptima.
Temperatura
15º 40º 75º
Aumento de la velocidad
Desnaturación por calor

ORGANISMOS TERMÓFILOS
Son organismos que viven a altas tº y realizan sus reacciones enzimáticas a estas altas tº
Ejemplos son los que viven en las aguas termales
Es tema de investigación el aislamiento de las enzimas de estos organismos para desarrollar procesos industriales en condiciones más extremas

CoenzimasCoenzimas
Las coenzimas son pequeñas moléculas Las coenzimas son pequeñas moléculas orgánicas, que se unen a la enzima.orgánicas, que se unen a la enzima.
Las coenzimas colaboran en la reacción Las coenzimas colaboran en la reacción enzimática recibiendo transitoriamente enzimática recibiendo transitoriamente algún grupo químico: Halgún grupo químico: H+ + , OH, CH, OH, CH33 . .
La enzima sin la coenzima recibe el nombre La enzima sin la coenzima recibe el nombre de APOENZIMAde APOENZIMA

El NAD es una coenzima aceptora de H
Sustrato oxidado

Algunas enzimas requieren metales para Algunas enzimas requieren metales para mejorar su actividad mejorar su actividad


Isoenzimas o Isozimas Son formas moleculares diferentes de una
misma enzima Catalizan la misma reacción Ejemplo: Lactato deshidrogenasa Lactato + NAD ==== Piruvato + NADH M4, M3H1, M2H2, M1H3 y H4
Se diferencian por su movilidad electroforética Usadas en clínica: sueros normales y sueros
con alguna patología

Así esta actividad corresponde al máximo potencial catalítico que las enzimas poseen para un determinado conjunto de condiciones ambientales. Luego, estas deben ser consideradas en las expresiones matemáticas representativas del proceso enzimático.
Existen situaciones (sustratos heterogéneos) donde la velocidad inicial de reacción no es representativa del real potencial catalítico de la enzima.
Se asume que la velocidad inicial de reacción es proporcional a la concentración de proteína enzimática.

Inhibición enzimática

Inhibidor:
Efector que hace disminuir la actividad enzimática, a través deinteracciones con el centro activo u otros centros específicos(alostéricos).
Esta definición excluye todos aquellos agentes que inactivan ala enzima a través de desnaturalización de la molécula enzimática
De esta forma, habrá dos tipos de inhibidores:
I. Isostéricos: ejercen su acción sobre el centro activo
II. Alostéricos: ejercen su acción sobre otra parte de la molécula, causando un cambio conformacional con repercusión negativa en la actividad enzimática.

Inhibición enzimática isostérica

Los inhibidores isostéricos pueden ser de dos tipos:
1. Inhibidor reversible: establece un equilibrio con la enzima libre, con el complejo enzima-substrato o con ambos:
E + I EI
2. Inhibidor irreversible: modifica químicamente a la enzima:
E + I E’
ES + I ESI

Inhibición reversible
(a) El inhibidor se fija al centro activo de la enzima libre, impidiendo la fijación del substrato: Inhibición Competitiva
(b) El inhibidor se fija a la enzima independientemente de que lo haga o no el substrato; el inhibidor, por tanto, no impide la fijación del substrato a la enzima, pero sí impide la acción catalítica: Inhibición No Competitiva
(c) El inhibidor se fija únicamente al complejo enzima-substrato una vez formado, impidiendo la acción catalítica; este tipo se conoce como Inhibición Anticompetitiva

Inhibición Competitiva

Inhibidores Competitivos
Compiten con el sustrato por el sitio activo de la enzima
Se une solo a la enzima libre V máx no se altera y K M cambia

Inhibición competitivaInhibición competitiva
Al aumentar la cantidad Al aumentar la cantidad de SUSTRATO el de SUSTRATO el inhibidor competitivo es inhibidor competitivo es desplazado y se forma desplazado y se forma productoproducto


E ES
EI
I
S
E + P
Características:- Las fijaciones de substrato e inhibidor son mutuamente exclusivas- A muy altas concentraciones de substrato desaparece la inhibición- Por lo general, el inhibidor competitivo es un análogo químico del substrato.- El inhibidor es tan específico como el substrato
Se define una constante deequilibrio de disociación delinhibidor:
Ki = [E] [I]
[EI]

En condiciones de equilibrio rápido, llamando y a la concentración del complejo EI, para la inhibición competitiva obtenemos el siste-ma de ecuaciones
vV s
Ki
Ks
m x
mi
1
Ke x y s
x
Ke x y i
y
m
i
( )
( )
0
0
Que resuelto para x nos da
xe s
Ki
Ksm
i
0
1
De donde

Por tanto, en la inhibición competitiva,
1. El efecto cinético del inhibidor es el aumento aparente de la Km, que aparece multiplicada por el factor (1 + i/Ki)
2. La Vmax no aparece modificada; para concentraciones muy altas del substrato, v = Vmax, igual que en ausencia de inhibidor
3. Cuanto más pequeño sea el valor de Ki mayor será la potencia del inhibidor competitivo.

KmSin
inhibidor
Kmcon
inhibidor
Sin inhibidor
Con inhibidor
El inhibidor competitivo aumenta la Km

1/s-0.3 -0.2 -0.1 0.0 0.1 0.2 0.3 0.4 0.5 0.6
1/v
0.00
0.01
0.02
0.03
0.04
0.05
0.06
0.07
-1/Km
-1/(Km(1 + i/Ki))
1/Vmax
Inhibición competitiva - Representación recíproca doble

Ejemplo Inhibidor Competitivo
Ácido succínico + FAD == ácido fumárico + FADH2
El inhibidor competitivo es el ácido malónico
Es un análogo estructuralmente parecido al ácido succínico

Ácido Fólico y Sulfanilamida
La sulfanilamida es un análogo estructural del ácido p - aminobenzoico (PABA)
PABA es el punto de partida para la síntesis de ácido fólico en las bacterias
El ácido fólico es una vitamina esencial para la proliferación (división) bacteriana
Sulfanilamida se usa en el tratamiento algunas infecciones

Metotrexato y Dihidrofolato
El ácido fólico en sus formas de dihidro y tetrahidrofolato es coenzima de la reacción catalizada por la dihidrofolato reductasa
Esta reacción es parte del metabolismo de los nucleótidos para la síntesis de DNA
Metotrexato se usa en la terapia de algunos cánceres, inhibiendo la síntesis de DNA

Inhibidor competitivosu estructura es similar a la del sustrato
No se forma producto



Inhibición No Competitiva

Inhibidor No Competitivo
Se une a un lugar diferente del sitio activo la enzima
Se une a la enzima libre y también al complejo enzima-sustrato
Por acción del inhibidor disminuye la Vm pero el valor de Km no se altera

Inhibición NO competitivaInhibición NO competitivaInhibición NO competitivaInhibición NO competitiva

Inhibidor NO competitivoInhibidor NO competitivoEl inhibidor NO competitivo se une a la enzima en un
sitio diferente del sitio activo

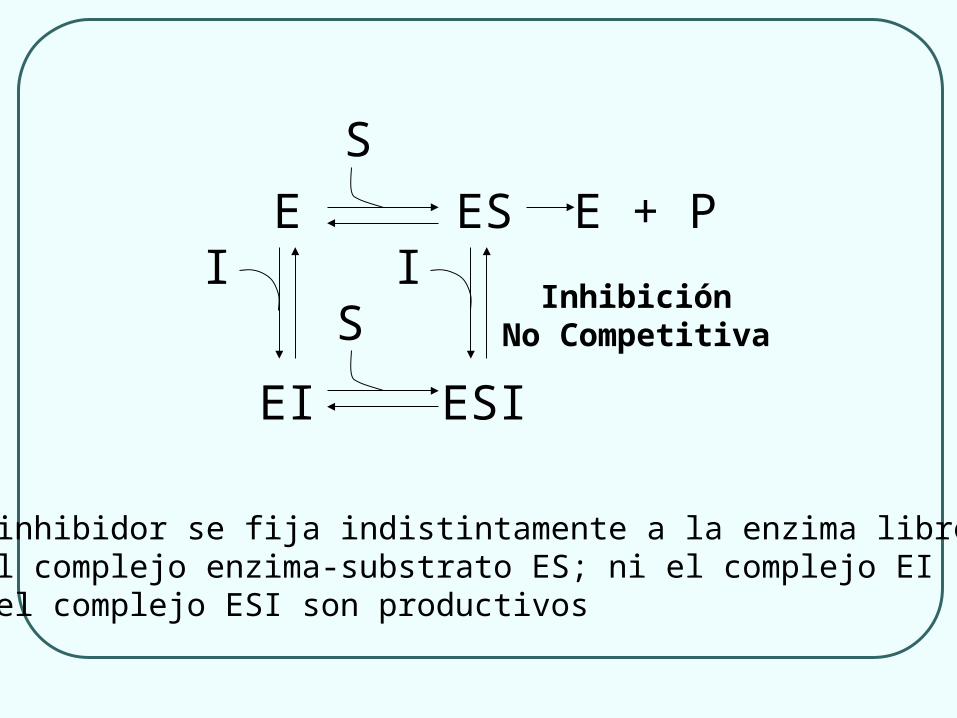
E ES
EI
I
S
E + PI
ESI
SInhibición
No Competitiva
El inhibidor se fija indistintamente a la enzima libre Ey al complejo enzima-substrato ES; ni el complejo EIni el complejo ESI son productivos


Inhibición Anticompetitiva o Incompetitiva

Inhibidor IncompetitivoEn este tipo de inhibición el inhibidor se enlaza al centro activo pero solo después de que el sustrato lo haya hecho y, por tanto, inhibidor y sustrato no compiten. De esta manera, aunque todo el sustrato esté saturando la enzima y toda la enzima esté como complejo ES, el inhibidor puede enlazarse produciendo un complejo inactivo ESI.
Como I solo se une a ES estimula la formación de ES y, por tanto, incrementa la unión del sustrato a la enzima, disminuyendo Km . Sin embargo, el complejo ESI no conduce a productos y Vm disminuye.

Inhibidor Incompetitivo
Se une a un lugar diferente del sitio activo de la enzima
Se une sólo al complejo enzima-sustrato Los efectos que tiene: disminuye el valor
de Km y también el de Vmáx

E ESS
E + PI
ESI
InhibiciónAnticompetitiva
El inhibidor sólo puede fijarse al complejo ES;el complejo ESI no es productivo

Modelo Kap Vap
Inh comp Km(1+i/Ki) Vm
Inh no comp total Km Vm/(1+i/Ki)
Inh anticomp Km/(1+i/Ki) Vm/(1+i/Ki)
Determinación de parámetros cinéticos de inhibición

Inhibición Irreversible
- Los inhibidores irreversibles reaccionan con un grupo químico de la enzima, modificándola covalentemente
- Su acción no se describe por una constante de equilibrio Ki,
sino por una constante de velocidad ki:
E + I E’
- A diferencia de la inhibición reversible, el efecto de los inhibidores irreversibles depende del tiempo de actuación del inhibidor.
- Los inhibidores irreversibles son, por lo general, altamente tóxicos.

Inhibidores Irreversibles
Producen inactivación permanente de la actividad enzimática
Se interfiere con el normal desarrollo de una reacción o vía metabólica
Ejemplos: p-cloromercuribenzoato, yodoacetato, diisopropilfluorfosfato

Algunos tipos de inhibidores irreversibles
1. Reactivos de grupos -SH
2. Organofosfóricos
3. Ligandos de metales
4. Metales pesados

Inhibición enzimática alosterica

Enzimas Alostéricas
Son enzimas cuya estructura proteica está formada de varias subunidades
No se rigen por la cinética de M - M Además del sitio o centro activo tienen sitios
alostéricos o de regulación Sitio activo/sustratos; Sitio alostérico/moduladores o reguladores La relación entre la velocidad de reacción y la
concentración de sustrato sigue cinética sigmoídea

Enzimas alostéricas Las enzimas alostéricas
presentan estructura cuaternaria.
Tienen diferentes sitios activos, unen mas de una molécula de sustrato
La unión del sustrato es cooperativa
la curva de velocidad presenta una forma sigmoidal


Concepto de Cooperatividad La unión de los sustratos al sitio activo de
una subunidad produce un cambio conformacional que por contacto físico se transmite a las subunidades vecinas, facilitando las interacciones
Modelos de unión: secuencial y concertado Ejemplos: unión Hb-O2, enzimas alostéricas
y sus sustratos

Moduladores Alostéricos
También reciben el nombre de efectores y pueden ser positivos o negativos
Se unen al sitio alostérico que puede estar en la misma subunidad que tiene al sitio activo o en las subunidades regulatorias
Su unión produce un cambio conformacional que afecta al sitio activo

Aspartato Transcarbamilasa
Ác L-asp + Carbamil-P === Carbamil-asp + Pi . Primera reacción en la síntesis de pirimidinas
Efector positivo: CTP Efector negativo: ATP
Tiene dos subunidades catalíticas para los sustratos y tres subunidades regulatorias para los efectores


RESUMENRESUMEN
Las enzimas son proteínas que catalizan Las enzimas son proteínas que catalizan las reacciones biológicaslas reacciones biológicas
Presentan especificidad por su sustratoPresentan especificidad por su sustrato Cada enzima presenta dos parámetros Cada enzima presenta dos parámetros
importantes Vmax (saturación de la importantes Vmax (saturación de la enzima) y la Km (medida de la afinidad enzima) y la Km (medida de la afinidad por el sustrato)por el sustrato)

RESUMENRESUMEN La actividad enzimática puede ser inhibida, La actividad enzimática puede ser inhibida,
por inhibidores competitivos (similares al por inhibidores competitivos (similares al sustrato) o por inhibidores no competitivos.sustrato) o por inhibidores no competitivos.
La temperatura y el pH afectan a la enzima La temperatura y el pH afectan a la enzima en su actividad catalítica.en su actividad catalítica.
Algunas enzimas requieren de coenzimas Algunas enzimas requieren de coenzimas y/o cofactores para su actividad.y/o cofactores para su actividad.
