e a m a r o n e f e p naeus eallfornienŒisbiblio.uabcs.mx/tesis/te521.pdf · 7 5 coeficiente...
TRANSCRIPT

e A M A R o N
llfornienŒisP naeus ea
e F E
TE5 1
2r

DEDICATORIA
A tí SEÑOR
Porque esta tØsis no habría
GRACIAS
sido posible sin tu ayuda
A tí PapÆ
Por que has sido para mi un gran ejemplo de trabajo animo y
perseverancia en alcanzar las metas que uno se propone Eta
tØsis e el resultado del amor esfuerzo y dedicacion que iempre
me has brindado Gracias te quiero mucho
A tí MamÆ
Por el apoyo
recibido
y conf lanza que aun estando lejos
Siempre he de tí Gracias por el amor tan
grande que siempre me has tenido Yo tambiØn te quiero
A mis Hermanos
Armando te admiro slompre he visto en ti un gran
modelo a seguir Claudia tu caracter me ha ayudado a
fortalecer el m la Raul gracias por tu amor y
preocupaClon por mi primi graclØls por tGdo
A tí Claudia
En verdad eres alguien muy especial para mi y deseo
que lo Sigas siendo
Pepe y Oscar
Esta tambiØn va por ustedes dos
í

AGRADECIMIENTOS
Agradezco a todos los profesores y personas que participaron
en mi formación acadØmica especialmente a la Profra Aida Torres
Dircio Ing Alejandro ChÆvez Prof Samuel Flores y al
Dr Carlos Lechuga D
A la Arq Flori Trejo Barrera por su ayuda en la realización
de las grÆficas de esta tØsis
A los Profesores Giovanni Malagrino y Mario Yoshida por el
tiempo y ayuda en la realización de esta tØsis
A la Universidad Autónoma de Baja California Sur y al Centro
de Investigaciones Biológicas de B C S por las instalaciones
y facilidades otorgadas en la realización de este trabajo
Al Dr Carlos CÆceres Martínez y familia por su amistad A
mis amigos y compaæeros de la carrera
y en forma muy especial al Dr Humberto Villarreal
Colmenares por su amistad tiempo asesoría y orientación eT la
realización de mi Servicio social y trabajo de TØsis
Mil Gracias
ii

1
INDICE
PÆgina
1 Dedica tor iai
B Agradec imientos ii
c 1ndice 1
D Indice de Figuras 4
E 1ndice de Tab1 as 6
l Resœmen 7
2 Justificación 10
3 Introducción 13
Antecedentes3 lo
3 2 Biología de camarón
13
14
3 3 Cultivo de camarón en el mundo 15
3 3 lo
3 3 2
3 3 3
3 3 4
Tipos de cultivo 15
16
16
17
17
sistema
Sistema
Sistema
extensivo
semi intensivo
intensivo
3 4 Países mÆs importantes en el cultivo de camarón
3 4 1 Especies mÆs importantes en el mundo 18
3 5 cultivo de camarón en MØxico 18
203 5 1 Cultivo de camarón en B C S
3 6 Calidad del agua 21

2
3 6 1 Temperatura 23
3 6 2 Sal inidad 25
3 6 3 Oxígeno disuelto 26
3 7 Metabolismo de crustÆceos 29
4 Objetivos generales 32
5 Materiales y mØtodos 33
5 1 Organismos experimentales 33
5 2 Unidades experimentales 33
5 3 Tratamientos experimentales 35
5 3 1 Temperatura 36
5 3 2 Sal inidad 36
5 4 Diseæo experimental 38
5 4 1 Mantenimiento de los organismos 38
5 4 2 Aclimatación de los organismos 39
5 4 3 Desarrollo experimental 39
5 5 AnÆlisis estadístico 40
6 Resultados 42
6 1 Consumo de oxígeno disuelto en postlarvas de
R eal ifarn le nsis 42
6 2 Efecto de la temperatura en el consumo de oxígeno
de postlarvas de E californiensis 49
6 3 Estimación del coeficiente tØrmico o 49
6 4 Efecto de la salinidad en el consumo de oxígeno
de postlarvas de E californiensis 49

3
6 5 Efecto combinado de la temperatura y la salinidad
en el consumo de oxígeno de postlarvas de g
californiensis 50
7 Discusión y conclusiones 57
7 1 Organismos experimentales 57
7 2 Diseæo experimental 57
7 2 1 Mediciones del consumo de oxígeno 57
7 2 2 Tratamientos experimentales 58
7 3 Relación del nivel de oxígeno disuelto con el
consumo de oxígeno de g californiensis 59
7 4 Efecto de la temperatura en la tasa metabólica
rutinaria de postlarvas de californiensis 61
7 5 Coeficiente tØrmico Q10 63
7 6 Efeeto de la salinidad en la tasa metabólica
rutinaria de postlarvas de R californiensis 64
7 7 Efecto de la temperatura y la salinidad en la tasa
metabólica de rutina de postlarvas de R
cal iforniensis 64
8 Literatura citada 68

4
INDICE DE FIGURAS
PÆgina
l Patrón típico de las fluctuaciones del oxígeno disuelto
en estanques de acuacultivo de acuerdo a la hora
del día Modifieado de stickney 1979 y Boyd 1990 28
2 Ubicación geogrÆfica del complejo lagunar Bahía
Magdalena Almejas indicando el lugar de colecta
de los reproductores 240 25 lato Norte 1110 44
long Oeste 34
3 Diagrama del sistema de respirómetros utilizado para
medir el consumo de oxígeno de postlarvas de P
californiensis 37
4 Disminución del OD a travØs del tiempo a 25 ppmil y a
cinco diferentes temperaturas para postlarvas de E
californiensis 44
5 Disminución del OD a travØs del tiempo a 35 ppmil y a
cinco diferentes temperaturas para postlarvas de E
californiensis 45
6 Disminución del OD a travØs del tiempo a 45 ppmil y a
cinco diferentes temperaturas para postlarvas de E
californiensis 46

5
7 Disminución del 00 a travØs del tiempo a 55 ppmil y a
cinco diferentes temperaturas para postlarvas de
californiensis 47
8 Valores del consumo de oXlgeno mg l min en los
respirómetros conteniendo postlarvas de P
californiensis para cada combinación de temperatura y
salinidad experimental 48
9 TØ 3a metabólica rutinaria mg 02 g min para
las 5 temperaturaspostlarvas de E californiensis a
experimentales 51
10 Variaciones en el coeficiente tØrmico QI0 de
postlarvas de E californiensis a diferentes
salinidades 53
11 Tasa metabólica rutinaria mg 02 g min para
postlarvas de E ca 1 iforniens is a las 4
salinidades experimentales 54
12 Efecto combinado
sobre el consumo
de
de
la temperatura y salinidad
oxígeno de postlarvas de E
californiensis 56

6
INDIcE DE TABLAS
PÆgina
l Solubilidad del oxígeno a saturación mg l en el agua
en función de la temperatura y la salinidad tomada
de Hughes 1991 27
2 Valores promedio de los interceptos a y pendientes
b de las regresiones lineales de oxígeno disuelto
mg l contra tiempo min para postlarvas de camarón
cafØ f californiensis 43
3 Consumo de oXigeno mg g min de postlarvas de camarón
cafØ f californiensis a las diferentes salinidades
y temperaturas experimentales 52
4 Estimaeiones del coeficiente tØrmico Q10 a diferentes
salinidades para postlarvas de camarón cafØ P
californiensis 55
5 Tasas metabólicas rutinarias publicadas para algunos
camarones pene idos 67

7
1 RESUMEN
En la aetualidad la acuacultura ha contribuido a satisfacer
las necesidades alimenticias del hombre así como a evitar que
las poblaciones naturales se vean mayormente afeetadas por la
sObre explotación pesquera El enfoque actual en el culti vo de
camarón estÆ dirigido a la producción de semilla de alta calidad
y a la engorda de los organismos en sistemas cuyo desarrollo
tecnológico sea eon base en las condiciones climÆticas donde se
desee llevar a cabo el cultivo Uno de los aspectos mÆs
importantes es la determinación de los intervalos óptimos de los
parÆmetros de cultivo así como la capacidad de adaptación de los
organismos En el presente estudio se define el efecto de
diferentes temperaturas y salinidades sobre el consumo de oxígeno
de postlarvas de Penaeus californiensis
Se establecieron 5 niveles de temperatura 19 23 27 31 Y
350C y 4 niveles de salinidad 25 35 45 Y 55 ppmil de
acuerdo a los valores registrados en granjas comerciales en
Sonora y Sinaloa y en granjas experimentales de cultivo de
camarón en B C S
La evaluación del consumo de oxígeno tasa metabólica
rutinaria de E californiensis para todas las combinaciones de
temperatura y salinidad se realizó en unidades de 250 mI
Los resultados obtenidos indican que existe una relación
directa entre la temperatura y el consumo de oxígeno de

8
postlarvas de E californiensis P 0 01 Por otra parte se
encontró que la salinidad influye en forma directa en el consumo
de oxígeno a 27 31 Y 3 sOc aunque el efecto es
proporcionalmente menor A 19 Y 23oC el eonsumo de oxígeno
permanece constante con el aumento en la salinidad
Se observó por otro lado que la cantidad de
disuelto en el agua no afectó el consumo da oxígeno
postlarvas siendo este constante e independiente
aproximadamente 1 5 mg O2 1
oxígeno
de las
hasta
Los resultados indican que existe un incremento en el
requerimiento de oxígeno de las postlarvas de Penaeus
californiensis para realizar sus funciones metabólicas conforme
aumenta la temperatura Por encima de los niveles reportados en
granj as con cultivos exi tosos a altas sal inidades existe un
incremento adicional que ocasiona que gran parte de la energía
disponible por el organismo sea utilizada en funciones
metabólicas limitando el potencial de crecimiento
Se encontró que existe un intervalo de temperatura 23 a
27oC en el cual el consumo de oxígeno no presentan diferencias
significativas P 0 01 independientemente de la salinidad
siendo este posiblemente el intervalo de temperatura óptimo
para la especie La investigación futura deberÆ ser encaminada
a evaluar el crecimiento y las modificaciones en el balance
energØtico de las postlarvas a diferentes niveles de temperatura
y salinidad para aSl poder determinar con mayor exactitud

9
cuales son los intervalos óptimos de los parÆmetros ambientales
que permitan incrementar la producción en los cultivo de camarón
cafØ californiensis

10
2 JUSTIFICACION
A partir de 1980 la pesquería de camarón en MØxico ha
llegado a su punto de mÆximo rendimiento ubicÆndose en un
promedio de 75 000 toneladas al aæo RPI 1989 Lanza Espino y
Arredondo 1990 Dicha producción no es capaz de satisfacer la
demanda tan elevada que hay de este crustÆceo tanto en el
mercado nacional como en el de exportación Villarreal 1989b
Aproximadamente entre el 80 y 90 del total de camarón que
se obtiene por captura y cultivo se exporta a palses como
Estados Unidos y Japón Barrena 1987 con lo que dicha
actividad representa para el país una de las fuentes de divisas
de mayor importancia Por esta razón se han establecido
recientemente estrategias que permitan obtener un desarrollo
tecnológico acelerado en el cultivo de camarón Dichas
estrategias estÆn enfocadas a la creación de cuadros tØcnicos
nacionales mediante el mejoramiento y acondicionamiento de las
tecnologias ya existentes a nivel mundial para adecuarlas a las
condiciones ambientales del paiS así como a las especies nativas
cICTUS 1982
La camaronicultura se practica con mayor Øxito en paises en
los cuales las condiciones climÆticas son cÆlido humedas en
donde las Øpocas secas y humedas se encuentran bien definidas
I1ughes 1991 En estas condiciones generalmente se obtienen
de 1 a 2 cosechas al aæo y en casos especiales vgr Ecuador se

11
logran hasta 3 cosechas anuales Rosenberry 1989
Las condiciones ambientales que imperan a lo largo del aæo
en el estado de Baja California Sur son típicas de zonas cÆlido
desØrticas por lo que no es posible obtener niveles de
producción cercanos a los que se obtienen en lugares con clima
tropjcal Esto es debido a que la tecnología disponible para el
cultivo se ha desarrollado en base a estas regiones las cuales
son mÆs favorables CIcTUS Q2 cit Hughes QR cit
El desarrollo de nuevas tecnologías nacionales que den como
resul tado una mayor produción y aprovechamiento del recurso
dependerÆ de la selección de la especie que presente una mejor
adaptación a las condiciones físicas químicas y biológicas donde
se desee llevar a cabo el cultivo Por ello es necesaria la
realización de estudios enfocados a obtener información sobre el
comportamiento fisiológico de la especie a cultivar en diferentes
condiciones medio ambientales
Debido a la importancia comercial de los camarones peneidos
en la acuacultura se ha realizado un mayor esfuerzo en
estudiar su nutrición New 1987 aunque mÆs recientemente se
han llevado a cabo un mayor nœmero de investigaciones
relac ionadas con su tasa metaból ica y especificamente con el
efecto de cambios en la salinidad Rao 1958 Kader 1962
Subrahmanyan 1962 Gaudy y Sloane 1981
Durante su ciclo de vida los camarones se enfrentan a

12
amplios intervalos de variación de temperatura y salinidad El
conocimiento de como estos factores afectan a su metabolismo
crecimiento y sobrevivencia es importante en la camaronicultura
ademÆs de ampliar el conocimiento relacionado con su biología y
ecología Darsey 1990
El objetivo principal del presente trabajo de investigación
es el de definir en base a la tasa metaból ica rutinaria los
intervalos óptimos de salinidad y temperatura del agua en los que
las postlarvas de camarón cafØ O cal iforniensis pueden
realizar de manera adecuada sus funciones fisiológicas bÆsicas
Esto contribuirÆ a definir la capacidad de adaptación del
organismo a dichos factores as 1 como proveer información
relevante al potencial de cultivo comercial de dicha especie en
la región cÆlido desØrtica del país

13
3 INTRODUccION
3 1 ANTECEDENTES
La acuacultura se refiere al cultivo de organismos acuÆticos
baj o condiciones controladas Bardach et al 1972 Los
organismos que se utilizan para esta actividad por lo general se
encuentran en densidades bajas en la naturaleza en tanto que en
acuacultura los organlsmos se encuentran en densidades altas
confinados a pequeæos volumenes de agua Villarreal 1984
Existen muy diversos niveles de acuacultivos de acuerdo al
grado de especialización y manejo que presentan abarcando desde
la simple introøucción de organismos acuÆticos en ciertos cuerpos
de agua donde estos no se encuentran presentes hasta sistemas
altamente especializados en los cuales se realiza una
recirculación de agua se provee de alimento artificial a los
organismos se cuenta con mØtodos para la prevención de
enfermadades y parÆsitos y se tiene un control adecuado de la
calidad del agua Villarreal Q2 cit Barrena Q2 cit
El culti vo de peces y de camarones es una industria
importante y de rÆpido crecimiento en muchas naciones tropicales
Arc 1989 Rosenberry Q2 cit Actualmente las tØcnicas para
producir semilla de camarones ya estÆ a disposición de los
acuacultores y los procedimientos para engordarlos en estanques
estÆn altamente desarrollados HughesQJ2 cit

14
El cultivo de camarón se practica principalmente en los
trópicos donde las Øpocas secas y hœmedas estÆn bien definidas
es decir durante la Øpoca 11 uv iosa las descargas de ríos
aumentan y la salinidad baja en los esteras mientras que en la
Øpoca seca la descarga de los ríos baja y la salinidad aumenta
en los esteros Broom 1971
1 2 IOLOGIA DEL cAMARON
El ciclo biológico de la mayoría de los camarones peneidos
se encuentra íntimamente ligado a sistemas estuarinos siendo
estas Æreas altamente productivas Odum 1971 en las cuales el
crecimiento de las etapas intermedias del ciclo de vida del
camarón postlarva juvenil y preadulto se 0e favorec ido
Barrena 9r eit Los pene idos presentan una fase larvilria
planctónica estadios de nauplio protozoea y mysis al
finalizar la cual el organismo pasa al estado de postlarva en
el que el cambio mÆs significativo es la adopción de hÆbitos
bØnticos RPI 912 cit Parece ser que el grado de
tolerancia hacia factores ambientales difiere de acuerdo a la
edad de los organlsmos Krarner 1975 Yagi et 9l 1990 Y
muchas especies estuarinas poseen mecanismos adaptativos capaces
de compensar los cambios de temperatura y salinidad dentro d un
inte valo bastante definido por la especie y su fase de
desarrollo Wolvekamp y Waterman 1960 Prosser y Brown 1968
Nelson et al 1977

15
3 3 CULTIVO DE CAMARON EN EL MUNDO
Debido a la gran aceptación que tiene el camarón a nivel
mundial su cultivo ha ido adquiriendo una mayor importancia en
los países que se dedican a la captura de este crustÆceo Bardach
QQ cit El cultivo de camarón tuvo sus inicios en el sur
este asiÆtico en donde se obtuvieron cosechas incidentales en
estanques de marea destinados a la piscicultura Chiang y Liao
1986 El cultivo de camarón recibió un gran impulso cuando en el
ano de 1934 en Japón Motosaki Fujinaga logró con Øxito la
reproducción y crianza del camarón E j 2gDi s Broom QQ
dt Barrena 9r ctt En los ultimos 20 aæos el cultivo de
camarón se ha desarrollado considerablemente gracias a que las
granjas que se dedican a la engorda de este organismo obtienen
mÆs facilmente postlarvas y juveniles producidos en
l boratorios los cuales son cultivados hasta llegar a su talla
comercial Rosenberry 9Q ctt
En 1990 los productores de camarón en el mundo obtuvieron
una marca rØcord de 663 000 toneladas de camarón entero 17 mÆs
que en el aæo anterior que fue de 565 000 ton lo que equivale
al 25 del producto comercializado a nivel mundial Rosenberry
1991
3 3 1 Tipos de Cultivo
Los camarones son cultivados en muchas naciones del mundo
y por esta razón se han desarrollado diversas tecnologías de
cultivo las cuales varlan dependiendo de las condiciones

16
geogrÆficas y climÆticas especies utilizadas y desarrollo
tecnológico del lugar de cultivo Arce QQ cit
Una de las formas en que se clasifican los sistemas de
e11ti va es con base a la densidad de organismos por unidad de
Ærea o volumen de agua y el cuidado o mantenimiento que se
dedique al sistema Tseng 1987
se definen sistemas de cultivo
intensivo
En base a esa clasificación
extensivo semi intensivo e
3 3 2 Sistema Extensivo
En este tipo de cultivo se utilizan estanques naturales
someros cercanos a bahías y ríos los cuales varía n en tamaæo
yendo de unas cuantas a cientos de hectÆreas y en los cuales se
siembran densidades de 1 000 a 25 000 juveniles ha El recambio
de agua se realiza por medio de las mareas en tasas de 5 a 10
por día y el alimento para los camarones proviene de la
producción natural de los estanques la cual puede estimularse
mediante la utilización de fertilizantes inorgÆnicos u orgÆnicos
La producción que se obtiene en este tipo de cultivos es de 50 a
500 kgjha aæo Roenberry 1990
3 3 3 Sistema Semi intensivo
En este tipo de cultivo se introduce una etapa de crianza o
pre engorda de postlarvas alimentación artificial y sistemas
de bombeo de agua en estanques de 2 a 25 ha El recambio de agua
es generalmente de 10 a 20 por día Los organismos se colocan en
a tas densidades en los estanClues de crianza p 5 000 a 200 000

17
juveniles ha y posteriormente en densidades mås bajas en los
estanques de crecimiento o engorda En el cultivo semi intensivo
se llega a obtener una producción de 500 a 5 000 kg ha aæo RPI
QQ cit
3 3 4 sistema Intensivo
En este sistema se utilizan estanques de 0 1 a 5 ha en los
que se siembran organismos en densidades superiores a 200 000
juveniles ha y en los que se lleva a cabo un control de
parÆmetros de calidad de agua a travØs de recambios de agua de
30 por día o mas Adicionalmente se practica la remoción de
desechos la aereación mecÆnica y la prevención de enfermedades
y parÆsitos En este tipo de cultivo se utilizan dietas
balanceadas nutricionalmente y con un alto valor protØico
CICTUS 92 cit RPI QP ç t obteniØndose producciones de
5 000 a 10 000 k9 h aæo Rosenberry 9Q çit
3 4 PAISES MAS IMPORTANTES EN EL CULTIVO DE CAMARON
Las condiciones climÆticas características de las zonas
tropicales en los países donde se obtienen las producciones mÆs
altas del cultivo de camarón son precipitación pluvial alta
1 000 mm aæo tasas de evaporaclon bajas y temperaturas
c onstantes a lo largo del ano Magallón 1988 Esto hace posiole
que se obtengan 1 ó 2 cosechas al aæo y hasta J para el caso de
Ecuador el cual es el principal productor de camarón cultivado
en LatinoamØrica y que actualmente produce 73 000 toneladas al
aæo Rosenberry 92 çit

18
Los países con mayor consumo de camarón son Estados Unidos
Japón y algunos países de Europa Occidental vgr Francia los
cuales cuentan con la tecnología mÆs avanzada en el cultivo
intensivo de camaron pero su producción es muy pequena Broom
QQ çit Rosenberry QQ cit Actualmente mÆs de 40 palses
cuentan con granJns de cultivo de camarón siendo China
Indonesia Tailandia y Ecuador los principales productores
3 4 1 Especies MÆs Importantes en el Mundo
Existen alrededor de 20 especies de peneidos de importancia
comercial de las cuales Pßn9 us mo o Qn Y E ehinen is cultivados
enAsia y E Y¨B amei y p tyJLlrQ2 ri cultivados en
AmØrica son las espeCles mÆs utilizadas en el cultivo debido
a características particulares de cada una de ellas como son
tamaæo tasa de crecimiento adaptabilidad para reproducción
en cautividad y tolerancia a variaciones en algunos
factores fisicoquímicos como la salinidad y la temperatura
Acuacop 1975 RPI QQ cit 3
5 CULTIVO DE CAMARON EN MEXICO El
cultivo de camarón en MØxico se inició a partir de 1970 Saer
1987 Dugger 1990 basado en dos modelos de nuevo desarrollo
tecnológico uno de tipo intensivoy de ciclo completo de
camarón azul p tLLi ostris en la unidad experimental de Puerto
Peæasco Sonora CICTUS QQ cit y el otro en Nayarit de tipo
semi intensivo y ciclo abierto utilizando un sistema de

19
estanques someros Lanza Espino y Arredondo 22 cit
En nuestro palS existen mÆs de 335 000 hectÆreas con
potencial de uso para cultivo de camarón Barrena QQ c t De
las aproximadamente 70 granjas que actualmente operan en el país
60 utilizan un sistema extensivo de cultivo 35 utilizan un
sistema semi intensivo y solo 5 utilizan el sistema intensivo
Rosenberry
cultivo y un
Dugger Q12
encuentra en
QQ ç tt Actualmente se cuentan con 8 000 ha de
total de 7 laboratorios de producción de semilla
çit El grueso de la industria camaronera se
el Pacífico particularmente en el estado de
Sinaloa Las especies que se cultivan en MØxico son E yann
92 y r stYJjJ Q tri 6 Lanza Espino y Arredondo 9J2
çit Rosenberry 92 cit aunque recientemente E gztec 2 y r
çalÜ9LtÜßn ŒJâ han sido consideradas especies potenciales para el
cultivo en nuestro país Arce QQ cit Villarreal 9p Lt Darsey
92 cit E calìfornien llamado comunmente camarón cafØ
es extraído en las costas del Pacífico Mexicano y su producción
juega un papel importante en el comercio Ki taniy Alvarado
1982 MØxico produce por medio del cultivo de camarón aproximadamente
4000 ton aæo que equivalen al5 5 del total que
se obtenido por medio de su captura RPI 1989 Este nivel de
producción se encuentra muy por debajo de la capacidad instalada
en el pais Barrena Ql ci t Sin embargo lil apertura
del cultivo de camarón al sector privado efectiva a partir
del 10 de abril de 1990 permite la introducción de inversión
extranjera en asociación con capital mexicano Dugger

20
QR cit Rosenberry QR cit lo que mejorarÆ las perspe tivas
de producción en este sector
3 5 1 Cultivo de Camarón en B c S
El estado de Baj a California Sur cuenta con 3 000 ha de
sliperficie potencial disponible para el culti va de camaron
Hasta 1986 solamente 16 ha eran utilizadas por dos granjas de
cultivo de este crustÆceo Barrena QQ it
Algunas de las condiciones ambientales prevalentes en el
estado son una precipitación pluvial anual menor a 250 mm baja
nubosidad altas tasas de evaporación facilitadas por corrientes
de aire que son típicas de las costas orientales de los
continentes Magallón Q12 it Estas condiciones
medioambientales no son consideradas las mÆs adecuadas para el
cultivo de camarón utilizando tØcnicas de cultivo tradicionales
basadas en el uso de estanques someros Por esta razón el Centro
dl Investigaciones Biológicas de B C e ha comenzado la
evaluación del potencial de cultivo de especies nativas de
camarón en la región Tal es el caso de 2 stYJ jrQstri y de E
ç1l iforniensis El cultivo experimental de esta ultima especie
Casillas y Magallón 1988 RPI Q12 2it ha demostrado que
presenta posibilidades de ser explotada comercialmente
HernÆndez y GonzÆlez 1989 encontraron que durante la etapa
de crecimiento del camarón azul 2 tyLÜ Q triŒ en puerto Cha le
B c S el intervalo de temperatura del agua en la región fue
de 19 a 320C y que debido a la falta de aportes de agua dulce

21
la salinidad del agua se mantuvo por arriba de 35 ppmil Por lo
tanto el estudio de la respuesta metabólica del camarón en este
intervalo de temperaturas y salinidades es importante Darsey
QQ c t como un parÆmetro que permita establecer la capacidad
de adaptación de la especie a las condiciones fisicoquímicas
prevalentes en la región
3 6 CALIDAD DEL AGUA
La producción en estanques muchas veces estÆ limitada por la
degradación en la calidad del agua Sin embargo se ha comprobado
que aguas no purificadas pueden ser utilizadas para el cultivo
de varias especies de vida acuÆtica siempre y cuando no esten
contaminadas con altos niveles de químicos agrícolas stickney
1979
En la actualidad hay disponible una gran cantidad de
información sobre el manejo de la calidad del agua en estanques
de agua dulce Este conocimiento ha sido debidamente utilizado en
la camaronicultura ya que muchos de los mØtodos empleados en la
acuacul tura de agua dulce son directamente apl icables a los
cultivos salobres Hughes QQ eit
La mayoría de las grônj as y cr iaderos de camarón se
encuentran en lugares con una buena calidad de agua todo el ano
Menasveta 1980 Esta calidad de agua es un factor importante
que afecta el Øxito de la madurez gonadal en camarones
Primavera 1985 Y por consiguiente los porcentajes de
o 2 6 5 8

22
sobrevivencia y crecimiento de las larvas estÆn igualmente
relacionados con la calidad de la misma Villarrcal QQ cit La
calidad del agua se ve influenciada por una serie de factores
tanto internos como externos Por ejemplo factores externos como
la alimentación excesiva pueden resultar en un aumento de la
materia orgÆnica en oxidación lo que puede originar una
disminución en la cantidad de oxígeno disuelto Tsai 1989 y un
aumento en la concentración de dióxido de carbono amonia y
fósforo Kinne 1970 Bridges y Brand 1980 cuyo efecto
combinado ha demostrado ser precursor de infecciones bacterianas
Villarreal Q ci García y Brune 1989 Boyd 1990
A nivel interno el metabolismo de los organismos estå
influenciado por los paråmetros de calidad de agua Yagi y
Ceccaldi 1984 en un estudio realizado sobre crecimiento larval
del camarón rosa Palaem n serr us observaron que la salinidad
y la temperatura son los parÆmetros mÆs importantes en la
modificación del metabolismo del organismo particularmente en
aquellos de ambientes estuarinos donde su efecto en el ciclo de
vida tiende a opacar otros factores abióticos como la luz y
bióticos como la competencia la cantidad y calidad de alimento
etc Darsey 9P eit Estas respuestas metaból icas se
presentan frecuentemente en otros crustÆceosJ Dall 1986 Dalla
Via 1987a Scelzo y Zuæiga 1987
Debido a las características climjticas de la región seml
Ærida y a la importancia de evaluar el potencial de cultivo de

23
camarón en la zona se considera necesario evaluar el efecto que
tienen en la respuesta metabólica del camarón cafØ Penaeu
californiensis las variaciones en la temperatura y la salinidad
para poder así establecer el nivel de adaptación de la especie
Darsey QL cit Esto repercutirÆ en la optimización de las
tecnicas de cultivo de dicho organismo en una región determinada
3 6 1 Temperatura
virtualmente todos los candidatos para dcuacultivos son
organismos poiquilotermos por lo que sus tasas metabólicas estÆn
determinadas por la temperdtura ambiente del agua Kinne 912
cit
Los efectos de la temperatura son qUiZØlS los de mayor
importancia en la acuacultura Bardach ˙lQ çit Armitage y
Wall 1982 no solo por el efecto en las tasas metabólicas sino
sobre todo por los mecanismos adaptativos que condicionan la
distribución de las especles en sus diferentes habi tats Zuæiga
et al 1982 Algunos de los efectos directamente relacionados
con la temperatura son la cantidad de oXigeno que se puede
disolver en el agua la tasa de difusión de los gases Isai 92
çj t la variación en la tasa metabólica Villarreal 1989a y
el potencial patógeno de algunos organismos Boyd 91 ctt
celzo y Zœæiga Q2 çit encontraron que existe un incremento
en el consumo de oXlgeno con el aumento de la temperatura
ex perim ental para E Ì Ta tLlens is s ituac ió n peviamente
determinada en otros crustÆceos vgr Dall QI cit Liao y
Murai 1986

24
La selección de la especie de cultivo debe estar basada en
el conocimiento de los niveles de temperatura requeridos por la
especie Stickney QQ cit seæala que los organIsmos
utilizados en acuacultivos son usualmente separados en tres
grupos con base en su temperatura óptima 1 De aguas frias con
temperatura óptima de 150C o menores 2 De aguas cÆl idas con
temperatura óptima de 250C o mayores y 3 De temperaturas medias
con un intervalo entre las dos anteriores
Desviaciones repentinas con respecto a la temperatura de
acli atación provocan estrØs tØrmico alterando los
requerimientos respiratorios originando desbalances internos
Æcido base y produciendo disturbios en la regulación
electrolítica de los fluidos prosser y ßrown Q12 cit El
estrØs tØrmico por frío o calor se refiere Øl la respuesta
inducida en animales ectotØrmicos cuando la temperatura a la que
se encontraba el organismo se baja o se eleva en forma repentina
Spotte 1979 Villarreal 912 cLt Entre los efectos debidos a
un aumento repentino en la temperatura se encuentran aumen o de
la actividad motora inactividad enzimÆtica alta producción de
metabolitos tóxicos y una inadecuada osmorregulación Knut
Nielsen 1990 La disminución repentina en la temperatura
provoca en los organismos la pØrdida del equilibrio funciones
enzimÆticas disminuidas y una mayor permeabilidad de las
branquias afectando la osmorregulación Spotte
Vernberg 1970
912 c i t

25
Al elevarse la temperatura las reacciones enz imÆticas se
aceleran Johnson et al 1954 Se han definido varios mØtodos
para comparar cuantitativamente la magnitud del efecto de la
temperatura en las reacciones químicas El mØtodo mÆs empleado
para determinar la sensibilidad tØrmica de la tasa metabólica es
la aproximación de Q10 que se define como el cociente entre la
tasa metaból ica a una temperatura y la tasa metaból ica a una
temperatura 100C menor prosser y Brown Q e t si la tasa se
duplica QIO tiene un valor de 2 SI la tasØl se triplica Q10
vale 3 Y aS1 sucesivamente Los valores de Q10 varian
generalmente de 1 a 4 en el que los valores bajos indican una
alta adaptabil idad tØrmica de los organismos mientras que
valores altos indican una gran sensibi 1 idad a los cambios de
temperatura Knut Nielsen Q cit
Con frecuencia es necesar o calcular QIO para intervalos de
temperatura inferiores a 100C para poder describir adecuadamente
la sensibilidad tØrmica de los organismos Hill 1980 Los
valores de QIO obtenidos de esta manera indican que Sl la
sensibilidad tØrmica permaneciera constante durante un intervalo
de lJoC la tasa metabólica permaneceria tambiØn constante
Johnson QQ cit El QIO de las tasas metabólicas de los
poiquilotermos tiende normalmente a ser próximo a 2 Hill QP
cit
3 6 2 Salinidad
La salinidad se define como la concentración total de
disueltos en el agua y se expresa generalmente en mgjKg
iones
aunque

26
en a uacultura generalmente se expresa en partes por mil ppmil
Los iones que contribuyen mayormente en la sa1inidad son siete
Na K Ca Mg el sulfatos y bicarbonatos Boyd QQ cit
Cada especie marina presenta un intervalo óptimo de
salinidad en el que realiza sus funciones vitales en forma óptima
Bardach 9 cit aSl como su reproducción y producción de
larvas Para el caso de camarones peneidos este intervalo se
encuentra entre 28 y 35 ppmil
especies mÆs resistentes a
concentración salina
I1ughes 1991
las variaciones
Existen algunas
en el nivel de
Debido a las condiciones de la Península de Baja California
en donde se considera que no existe un suministro contínuo de
agua dulce se ve 1 imi tacto el tipo de culti va y las especies
suceptibles de producción acuicola comercial Por esta razón en
el aæo de 1984 el Centro de Investigaciones Biológicas inició un
proyecto piloto de cultivo de camarón en la zona de Puerto Chale
B C S en el que se llevó a cabo la evaluación del potencial
de engorda del camaron azul 2 2J YJ iJo tris en estanquería
somera En ese experimento se observó que la salinidad de los
estanques fue superior a la salinidad media del mar variando en
un intervalo de 35 a 44 ppmil con un promedio de 39 ppmil
Casillas et 9ì 1988
3 6 3 Oxígeno Disuelto
Es la variable de calidad de agua mÆs crítica en la
acuacultura Si los niveles adecuados de oxígeno disuelto 00 no
27
se mantienen los organismos sufren estrØs manifestÆndose en un
rechazo al alimento durante y despuØs del período de disminución
del OD causando por consiguiente una baja en la tasa de
crecimiento y en la eficiencia de conversión del alimento
Villarreal QQ cit
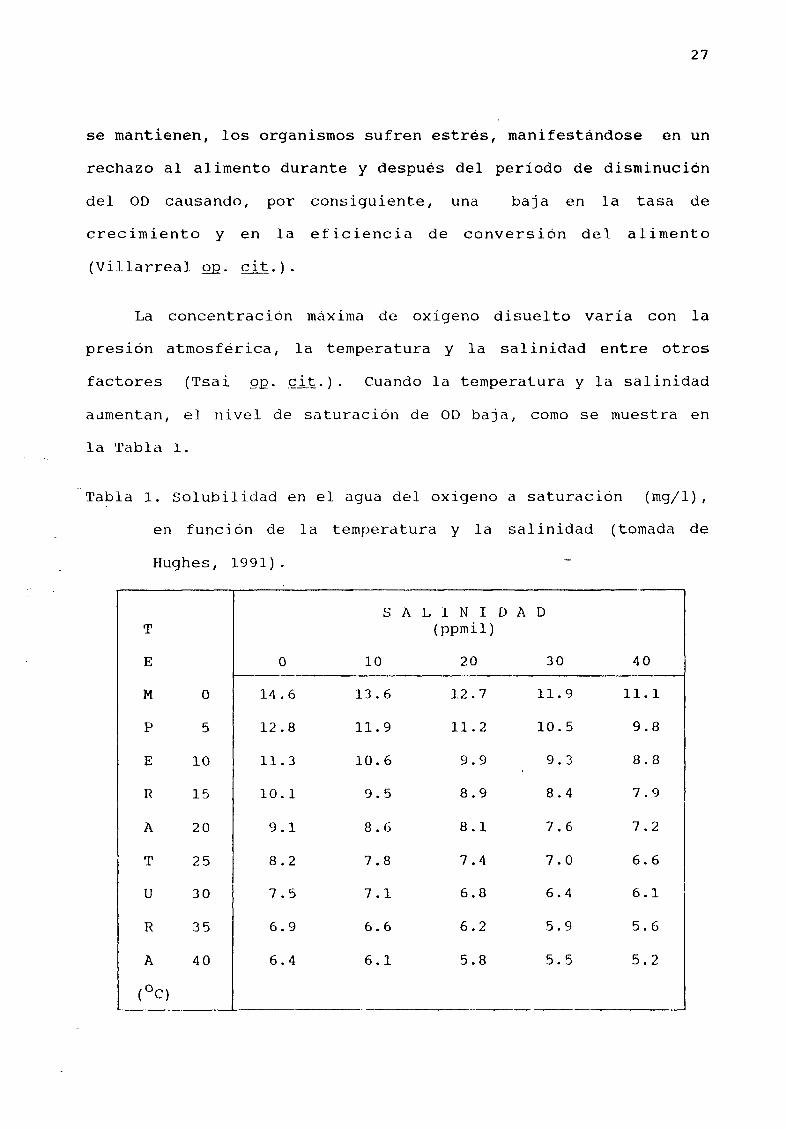
La concentración mÆxima de oXlgeno disuelto varía con la
presión atmosfØrica la temperatura y la salinidad entre otros
factores Tsai QQ cit Cuando la temperatura y la salinidad
aømentan e nivel de saturación de OD baja como se muestra en
la Tabla l
Tabla l Solubilidad en el agua del oX1geno a saturaeión mgjl
en función de la temperatura y la salinidad tomada de
Hughes 1991
S A L 1 N 1 D A D
T ppmil
E O 10 20 30 40
M O 14 6 13 6 12 7 11 9 11 1
P 5 12 8 11 9 11 2 10 5 9 8
E 10 11 3 10 6 9 9 9 3 8 8
R 15 10 1 9 5 8 9 8 4 7 9
A 20 9 1 8 6 8 1 7 6 7 2
T 25 8 2 7 8 7 4 7 0 6 6
U 30 7 5 7 1 6 8 6 4 6 1
R 35 6 9 6 6 6 2 5 9 5 6
A 40 6 4 6 1 5 8 5 5 5 2
oC

28
La influencia del ciclo de variación circadiana del OD sobre
el clecimiento de especies acuícolas es poco conocido pero el
mejor crecimiento se logra cuando la concentración de OD es
superior al 25 30 de saturación durante la noche DallQQ
it Boyd QQ cit La reducción mÆs pronunciada de OD en el
estanque de cultivo se relaciona principalmente con la temporada
de verano y con las primeras horas de la maæana stickney QQ
çJt Figura 1
20
O
X
1G
E 15
NO
D10
1S
U
E
L 5rr
O
mg l
0600
I
1200
I
2400
I
1800
T 1 E M P O Hrs
Figura 1 Patrón tipico de las fluctuaciones del oXlgeno disuelto
en estanques de acuacultivo de acuerdo a la hora
del día Modificado de stickney 1979 y Boyd 1990

29
3 7 METABOLISMO EN CRUSTACEOS
El OD es el mayor factor limitante metabólico debido a que
las reacciones bioquímicas del organismo son controladas por el
nivel de oxígeno el cual es esencial en el metabolismo aeróbico
Neill 1989 La cantidad de oxígeno a disposición de los
organlsmos determina la distribución ecológica y la
sobrevivencia prosser y Brown QQ cit Asimismo les ayuda a
mantener un buen nivel de crecimiento realizar sus funciones
normales de actividØld incrementar su resistencia a parÆsitos y
enfermedades y aumentar su tolerancia a cambios en las
condiciones medioalnbientales Kramer gg ctt Dalla Via 1915
La tasa metaból ica en la cual el proceso catabólico
representa uno de los principales canales de fluj o de energía
Brody 1964 considera el total de las transformaciones
energØticas que tienen lugar y es frecuentemente utilizada como
un indicador del estado interno del organismo Davies 1966 El
metaLolismo se divide en basal rutinario y activo Villarreal
gQ cit La tasa basal es aquel nivel de actividad minlma
compatible con la vida la tasa rutinaria es aquella que se
produce cuando hay una actividad motora mínima y la tasa activa
se da en condiciones de actividad motora sostenida
Es importante definir los intervalos de consumo de oxígeno
para entender algunos de los mecanismos fisiológicos y
metabólicos de los organismos En la acuacultura la estimación
de la tasa metaból ica y factores subordinados tales como el

30
punto críticò Pc permite determinar diversos factores tales
como los intervalos óptimos de aereación y la capacidad mÆxima de
un estanque de cultivo Villarreal QQ c t
Las mediciones del consumo de oXlgeno como una respuesta a
los cambios en la salinidad y temperatura dan como resultado una
estimación de la capacidad adaptativa del organismo Dalla Via
Qfl c t Scelzo y Zuæiga QQ eij or lo general la tasa
metabólica del organismo y su comportamiento se modifican despuØs
4
de presentarse cambios en la salinidad Kutty et al 1971 Yagi
íŒ t 91 9 cit como resultado de las variaciones en el gasto
energØtico debido al trabajo osmótico adicional Kinne 1971
Fletcher 1976 Rennett 1978 Dalla Via 1987b
En general el consumo de oXlgeno estÆ influenciado por
factores ambientales tales como la salinidad la temperatura la
Øpoca del aæo la intensidad luminosa y la concentrarión de
OD en el agua Kinne 912 cit ßridges y Brand 912 eij
Villarreal 912 elt ademÆs de factores intrínsecos de los
organismos tales como el tamaæo la actividad el estado de muda
desarrollo gonadal la inanición y los ritmos biológicos
Wolvekamp y Waterman Qr eit Nelson et fÜ Q1 cit Cockroft
y Wooldridge 1985 Por ejemplo Broom 91 clt observó que
el cama ron cafØ P 1l0 US 9LtIL mantenido en estanques con
concentraciones de 00 menores de 2 rng l mostl aban signos de
estrØs mientras que camarones mantenidos a concentraciones
mayores de 4 mg l no mostraron efectos fisiológicos negativos

31
En la camaronicultura es importante determinar las
condiciones mÆs favorables de cultivo para cada estadio Este
nivel óptimo puede ser determinado directamente por medio del
anÆlisis de la mortalidad y las tasas de crecimiento Yagi et
Ü QQ ç it sin embargo un mØtodo mÆs analítico consiste en
evaluar para cada etapa de desarrollo la tasa metabólica del
organismo dado que a mayor consumo de oXigeno mayor es la
energla que se destina a la respiración la regulación osmótica y
otros procesos vitales y es menor la energía disponible para el
crecimiento Nelson et 91 Qn cit Clifford y Brick FnB
Ydgi y Ceccaldi QQ cit Villarreal QQ cit Yagi t al Qn
c Lt

32
4 OBJETIVOS GENERALES
Los objetivos generales del
inyestigaeión fueron los siguientes
presente proyecto de
l Evaluar el efecto de variaciones en la salinidad y la
temperatura del agua en la tasa respiratoria de postlarvas de
camarón cafØ aliforniensis como un indicador de
modificaciones en la tasa metabólica rutinaria
2 Establecer parÆmetros de referencia que permitan definir los
niveles mÆximos y mínimos de salinidad y temperatura para el
culti va de la especie con base en la respuesta fisiológ ica del
orgarismo

33
5 MATERIALES Y METODOS
5 1 ORGANISMOS EXPERIMENTALES
Se utilizaron postlarvas de camarón cafØ P
cal iforniensis producto del desove de una hembra madura
obtenido en el Laboratorio de Cultivos del Centro de
Investigaciones Biológicas C l B La hembra fue capturada en
el complejo lagunar de Bahía Magdalena Almejas B C S 240 251
latitud Norte 1110 44 longitud Oeste Figura 2
Los estadios larvarios nauplio zoea y mysis se
alimentaron con microalgas ˙hiìs tQceros graçil is y Tetraselmis
ghlJiiJ una dieta comercial Artificial Plancton Nippai con un
contenido protØico de 40 y con nauplios de artemia rtemia
salina siguiendo las tØcnicas de cultivo tipo Galveston de
Ireece y Yates 1990 hasta llegar a postlarva 2 O PL2 O Se
seleccionaron un total de 600 postlarvas de acuerdo a su tamaæo
0 38 l 0 08 g que se determinó con la ayuda de una balanza
digital con precisión de 0 01 g
5 2 UNIDADES EXPERIMENTALES
Existe una gran variedad de diseæos para el moni toreo del
consumo de oxígeno de animales acuÆticos de los cuales los
sistemas cerrados de una o dos cÆmaras son los diseæos mÆs
utilizados Villarreal QP cit Para la medición del consumo de
oXlgeno en decÆpodos los sistemas usados son similares a los que

2e N
211 N
240N
2 UN
34
r r
t
IlaQ W
l1I18W 110
I
B J A
CALIFORNIA
OCi ANO
PACIFICO
Figura 2 Ubicación geogrÆfica del complejo lagunar Bahía
MacJdalena Almejas indicando el 1 uCJ i r d e col ctØl
de 240 25 lato Norte 1110 44los rcproductores
long Oeste

35
se emplean para peees vgr Sutcliffe et al 1975 Bunn y
Horwitz 1980 Hewitt D 1984 Swain g al 1987
En el presente trabajo experimental se utilizaron un total
de 28 matraces Erlenmayer de fondo plano de 250 mI de capacidad
los cuales fueron llenados con 200 mI de agua a la salinidad
experimental filtrada a 5 micras esterilizada con luz
ul trav ioleta y aereada a saturación Pa ra asegurar una
aereìción igual en cada respirómetro se utilizaron mangueras de
plÆstico flexibles de 0 5 cm de diÆmetro cada una con vÆlvulas
para regular el paso del aire En cada unidad experimental
respirómetro se colocaron 4 postlarvas Posteriormente se
cerraron los matraces con tapones de plÆstico y fueron colocados
dentro de un baæo María para mantener constante la temperatura
para cada serie experimental Figura 3
5 3 TRATAMIENTOS EXPERIMENTALES
Como se mencionó anteriormente debido a las condiciones
climÆticas de la región en las que la temperatura provoca
altas tasas de evaporación hay una escasa precipitación pluvial
y por consiguiente un bajo aporte de agua dulce para los cultlVOS
de camarón se seleccionaron 5 temperaturas y 4 salinidades
experimentales las cuales cubren un intervalo de variación
similòr al encontrado en experimentos de engorda realizados por
el personal del C l B en la Granj a piloto de Puerto
Chale B C s Casillas t Ql QQ cit Hernåndez y GonzÆlez

36
QQ t así como las temperaturas y salinidades en granjas de
Sonora sinaloa y Nayarit en donde se concentra el culti vo
comercial Por otro lado se evaluó el efecto de temperaturas y
salillidades altas en el metabol ismo del organismo 350C y 55
ppmil
5 3 1 Temperatura
Las temperaturas experimentales para la determinación de la
tasa metabólica fueron 19 23 27 31 Y 350C El experimento
se llevó a cabo en un cuarto con una temperatura de 23 10C
La temperatura de los respirómetros se controló con agua a la
salinidad experimental a baja temperatura para la pruebò a 19oC
y con calentadores sumergibles a 27 31 Y 350C
El coeficiente tØrmico definido como
QI0 K2 K1IO t2 t1
donde K1 Y K2 representan la tasa metabólica a dos diferentes
temperaturas tI y t2 Hill QQ cit Knut Nielsen QP it
se determinó para grupos de temperatura consecutivos 19 230C
23 270C 27 310C y 3I 35oC Los valores de QI0 representan el
nivel de sensibil idad del orgØlnismo a las variaciones de la
temperatura Yagi et al QQ cit
5 3 2 Salinidad
Las salinidades seleccionadas fueron 25 35 45 Y 55
ppmil las cuales se obtuvieron mediante la dilución del agua de
mar 38 ppmil con agua dulce para 25 y 35 ppmil y la adición

37
SUPlFME Nl0 DE AIHE
I AllUlA E PASO
r J
CAll N1ADO DE AGUA
H S PIROME 1 AO
NiVEL Dl AGUA
OANß MAI IA
DlfUSOA DE AIAE
UilHUcrUHA Úl SOPOHll
0OXIME TRQ
MEDIDon DE OXIGENOOL Ln
AESPIAOMfTAO
NIVEL Of AGUA
à lli Ùilll7
CAMAfIONllANO MAHIA
ILSTHuC1UHA Dl SOPOHll
Figura 3 Diagrama del sistema de respirómetros utilizado para
medir el consumo de oXlgeno de postlarvas de P
californiensis

38
de sal no refinada para 45 y 55 ppmil en las siguientes
proporeiones
25 ppmi 1 diluyendo agua de mar con 60 de agua dulce
35 ppmil diluyendo agua de mar con 12 de agua dulce
45 ppmil aæadiendo 5 g de sal no refinada l de agua de mar
55 ppmil aæadiendo 15 g de sal no refinada l de agua de
mar
5 4 DISEÑO EXPERIMENTAL
5 4 1 Mantenimiento de los organismos
Los organismos se mantuvieron a 23 10C en un tanque de
fibra de vidrio con dimensiones de 2 x 1 5 x 0 6 m en el que se
estableció un recambio de 66 por día con agua de mar a 38
ppmil filtrada 10 micras y desinfectada con rayos vv
Se proporcionó aereación constante manteniendo una
saturación superior a 85
aeuario Cada tercer día
utilizando 6 bombas de aire para
se llevó a cabo un monitoreo de los
siguientes parÆmetros de calidad de agua
a Oxígeno mgjl mediante un oximetro polarogrÆfico YSI
modelo 57
b Temperatura oC mediante un termómetro sumergible mÆximo
mínimo
c Salinidad ppmil mediante un refractómetro ocular

39
5 4 2 Aclimatación de los organismos
Del tanque de fibra de vidrio se tomaron 130 organismos
seleccionados aleatoriamente y se colocaron en unidades de
plÆstico de 0 7 x 0 4 x 0 35 m por un período de aclimatación de
cuando menos 24 horas DespuØs de colocar a los organismos en las
unidades de aclimatación se procedió a ajustar la salinidad del
agua a la de la serie experimental a realizarse 25 35 45 ó 55
ppmil dicho aj uste no fue superior a 8 ppmiljdía Debido a
esto el tiempo requerido para el ajuste de la salinidad fue de
1 día para 35 Y 45 ppmil y 2 días para 25 Y 55 ppmi l Durante
este tiempo se continuó dando al imento a las postlarvas
Posteriormente se ajustó la temperatura por un período no menor
de 24 horas antes de real izar cada prueba experimental El
período de inanición fue simultaneo al ajuste de la temperatura
5 4 3 Desarrollo Experimental
DespuØs del período de adaptación los organismos se
introdujeron en 14 respirómetros donde permanecieron por un
período de aclimatación de 2 horas antes de realizdrse la
evaluación del consumo de oxígeno El nivel de saturación de
oxígeno en el agua de los respirómetros filtrada a 5 micras y
desinfectada con UV se mantuvo cerca del 100 DespuØs del
período de aclimatación se hizo una medición con el oxímetro y
el electrodo polarogrÆfico del nivel de concentración de
oxígeno disuelto en el agua antes de cerrar hermØticamente cada
respirómetro Los camarones de dos de los respirómetros se
removieron y estas unidades experimentales sirvieron como

40
blancos o respirómetros de control para el monitoreo del
consumo de oxígeno de microrganismos y del electrodo La
duración del experimento fue de 60 minutos llevando a cabo
mediciones de la concentración de oX1geno disuelto en los
respirómetros cada 10 minutos Una vez finalizada la evaluación
se pesaron los organismos con una balanza digital con precisión
de 0 01 g Cada tratamiento experimental se evaluó por
duplicado esto es que se realizaron 24 evaluaciones
individuales de consumo de oxigeno para cada combinación de
temperatura y salinidad
5 5 ANALISIS ESTADISTICO
La evaluación estadística del estudio comprendió la
aplicación del anÆlisis de varianza la prueba de rangos
multiples de Tuckey y la prueba de t ponderada Sokal y Rohlf
1984 para la comparación de pendientes de dos 1 íneas de
regresión del consumo de oxigeno mgjgjmin obtenido en base a
la regresión de la saturación de oxígeno disuelto mgjl contra
tiempo min estandarizado al volumen utilizado en los
experimentos y a la biomasa total en el respirórnetro El
anÆllsis se realizó utilizando el paquete estadístico para
computadora STATGRAPHICS statgraphics 1986
La hipótesis
variaciones en la
nula Ho del estudio asume que las
producentemperatura y la salinidad no

41
diferencias en la tasa metabólica rutinaria del camarón La
hipótesis alterna Ha indica que hay diferencias entre los
tratamientos

42
6 RESULTADOS
6 1 CONSUMO DE OXIGENO DISUELTO EN POSTLARVAS DE P
californiensis
Gas figuras 4 5 6 Y 7 muestran la disminución del OD con
respecto al tiempo en los respirómetros conteniendo postlarvas
de camarón cafØ californiensis a las cuatro salinidades y
temperaturas experimentales El ajuste matemÆtico de una línea
recta proporcionó la mejor descripción de la relación de OD y
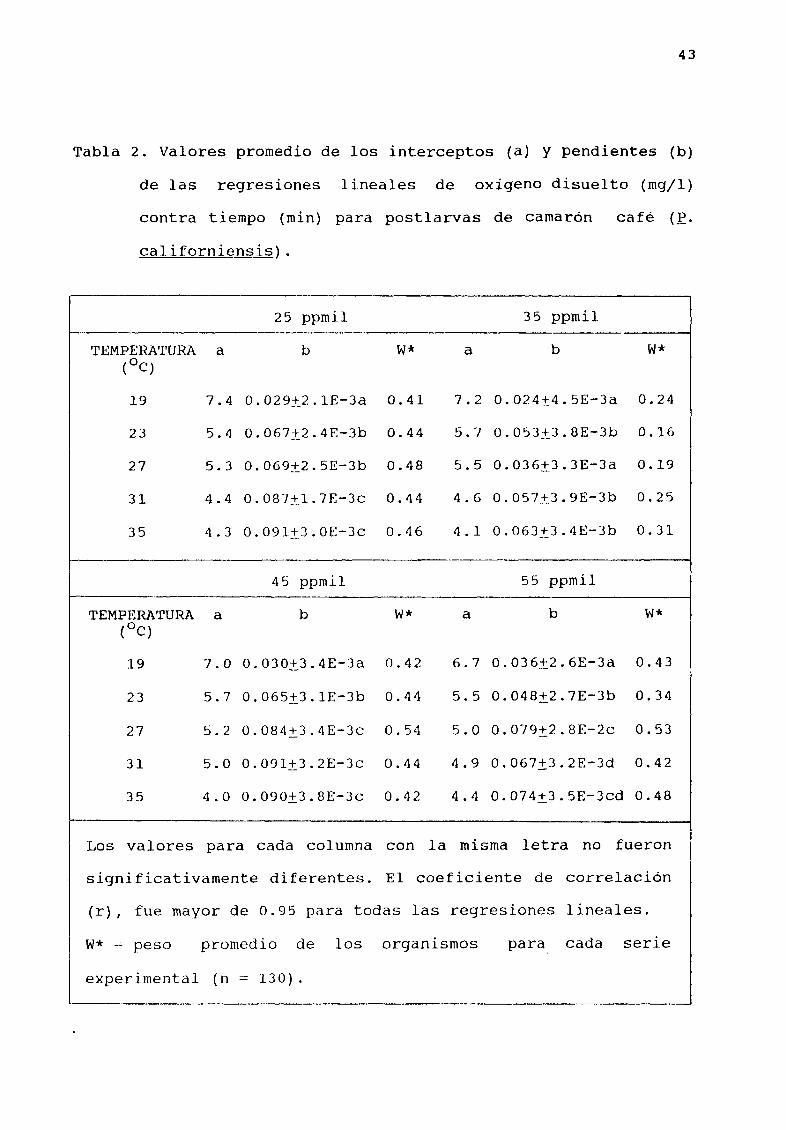
tiempo En la Tabla 2 se muestran los promedios de los
interceptas a y las pendientes b de las regresiones lineales
del OD contra el tiempo para las temperaturas y salinidades
experimentales La figura 8 muestra los valores de las pendientes
b las cuales representan el consumo de oXigeno por mlnuto
mg l min en los respirómetros para las temperaturas
experimentales a diferentes saliniades
El anÆlisis de varianza indicó que existen diferencias
significativas P 0 01 entre las pendientes de los
tratamientos experimentales A 35 ppmil hubo una disminución
generRlizada en la tasa de consumo de oxígeno mg l min a todas
las temperaturas
43
Tabla 2 Valores promedio de los interceptos a y pendientes b
de las regresiones lineales de oxígeno disuelto mgjl
contra tiempo min para postlarvas de camarón cafØ
çaliforniensis
25 ppmil
TEMPERATURA a
oCb w
19 7 4 0 029 2 1E 3a 0 41
23 5 4 0 067 2 4E 3b 0 44
27 5 3 0 069 2 5E 3b 0 48
31 4 4 0 087 1 7E 3c 0 44
35 4 3 0 091 3 0E 3c 0 46
45 ppmil
TEMPERATURA a
oCb w
19 7 0 0 030 3 4E 3a 0 42
23 5 7 0 065 3 1E 3b 0 44
27 5 2 0 084 3 4E 3c 0 54
31 5 0 0 091 3 2E 3c 0 44
35 4 0 0 090 3 8E 3c 0 42
35 ppmil
a b w
7 2 0 024 4 5E 3a 0 24
5 7 0 053 3 8E 3b 0 16
5 5 0 036 3 3E 3a 0 19
4 6 0 057 3 9E 3b 0 25
4 1 0 063 3 4E 3b 0 31
55 ppmil
a b w
6 7 0 036 2 6E 3a 0 43
5 5 0 048 2 7E 3b 0 34
5 0 0 079 2 8E 2c 0 53
4 9 0 067 3 2E 3d 0 42
4 4 0 074 3 5E 3cd 0 48
Los valores para cada columna con la misma letra no fueron
significativamente diferentes El coeficiente de correlación
r fue mayor de 0 95 para todas las regresiones lineales
w peso promedio de los organismos para cada serie
experimental n 130

44
7
6
Q1
E 5
o O44gf1 4wJ
04B
a
O 3zw O 44gCX 2
oo 10
o 190C
20 30
TIEMPO
L 230C 270C
40 50
min
X 31 oc
60
350C
Figura 4 Disminución del OD a travØs del tiempo a 25 ppmil y a
cinco diferentes temperaturas para postlarvas de E
californiensis

45
7
PESO6 PROMEDIO
0 24g
5I
Eo
4W 0 16g
0 19go
3ozw0X 2 0 250C 0 310
oO 10 20 30
TIEMPO
6 230C O 270C
40 50
min
X 31 oc
60
o 190C 350C
Figura 5 Disminución del OD a travØs del tiempo a 35 ppmil y a
cinco diferentes temperaturas para postlarvas de E
californiensis

46
7
6
5o
E
of
4jw
UJ
tJ0 54g
O3
zw
x 2
0 449042 g
oO 10 20 30
TIEMPO
L 230C O 270C
40 50
min
X 31 oe
60
o 190e 350e
Figura 6 Disminución del OD a travØs del tiempo a 45 ppmil y a
cinco diferentes temperaturas para postlarvas de f
californiensis

47
7
6 PESOPROMEDIO
50430
CJl
E
of 4 0 340JW
a
3ozw
x 2 0430o
048 g
o IO 10 20 30 40 50 60
T I E M P O min
O 1 90C 2 Oc O 270C X 31 Oc 350C
Figura 7 Disminución del OD a travØs del tiempo a 55 ppmil y a
cinco diferentes temperaturas para postlarvas de E
californiensis

0 10
0 08
f
J 0 06
o
o
lJJfZ 0 04w
ozwroL
0 02
0 00
48
25 35 45 55
SALlNIDAD ppmil0190C 23oC 0270C X310C 35oC
Figura 8 Valores del consumo de oxígeno mgjljmin en los
respirómetros conteniendo postlarvas de E
aliforniensis para cada combinación de temperatura
y salinidad experimental

49
6 2 EFECTO DE LA TEMPERATURA EN EL CONSUMO DE OXIGENO DE
POSTLARVAS DE californiensis
Se encontró que existe una relación directa entre el consumo
de oxígeno y la temperatura A 45 Y 55 ppmil esta relación se
describe de manera adecuada con el ajuste matemÆtico de una línea
recta por otro lado a 25 y 35 ppmil se observa una zona entre
23 Y 310C en la que el consumo de oxígeno permanece casi
constante independientemente del aumento de la temperatura
Figura 9 En general el consumo de oxígeno aumenta conforme se
incrementa la temperatura encontrÆndose diferencias
significativas entre los tratamientos P 0 001 Tabla 3
6 3 ESTIMACION DEL COEFICIENTE TERMICO Q10
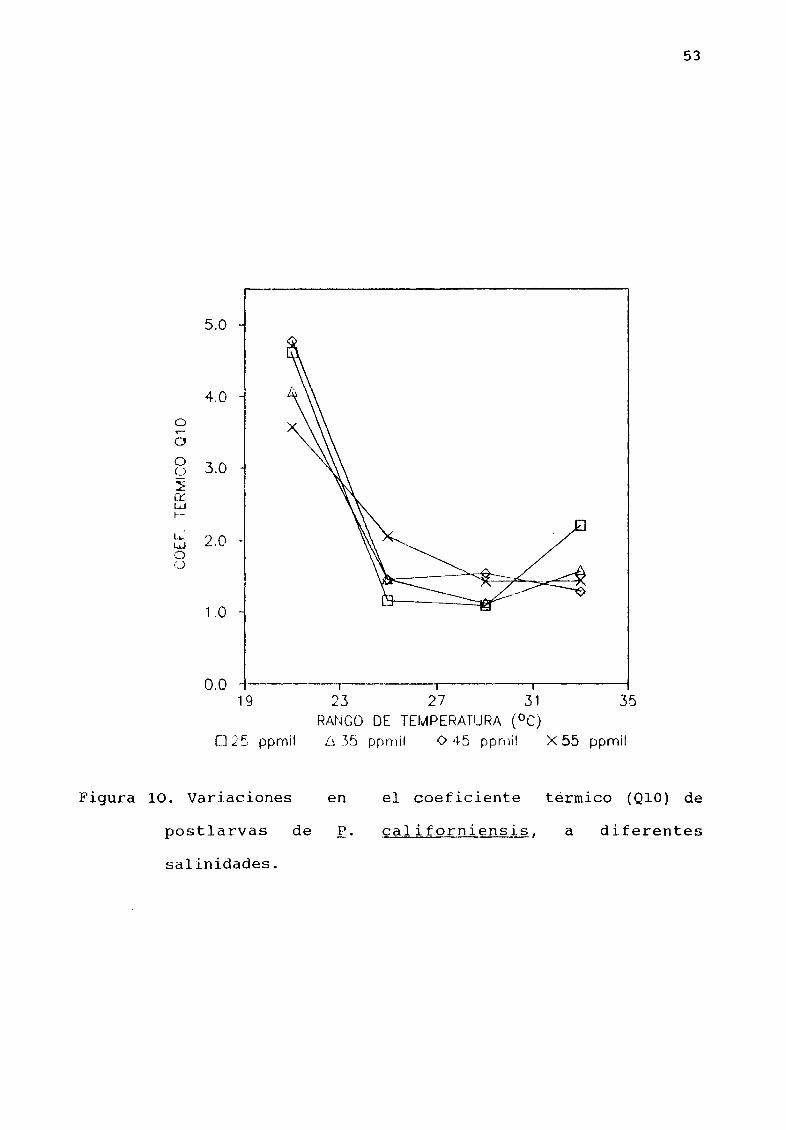
Estimaciones del coeficiente tØrmico Q10 de E
californiensis para las temperaturas experimentales se muestran
en la Tabla 4 y en la Figura 10 Se puede observar que para todas
las salinidades experimentales los valores mÆs altos de Q10
entre 3 5 Y 4 B se presentan en el intervalo de temperatura
experimental mÆs bajo 19 a 23oC reduciØndose en forma
considerable para los intervalos de temperatura de 23 27 27 31 Y
31 350C
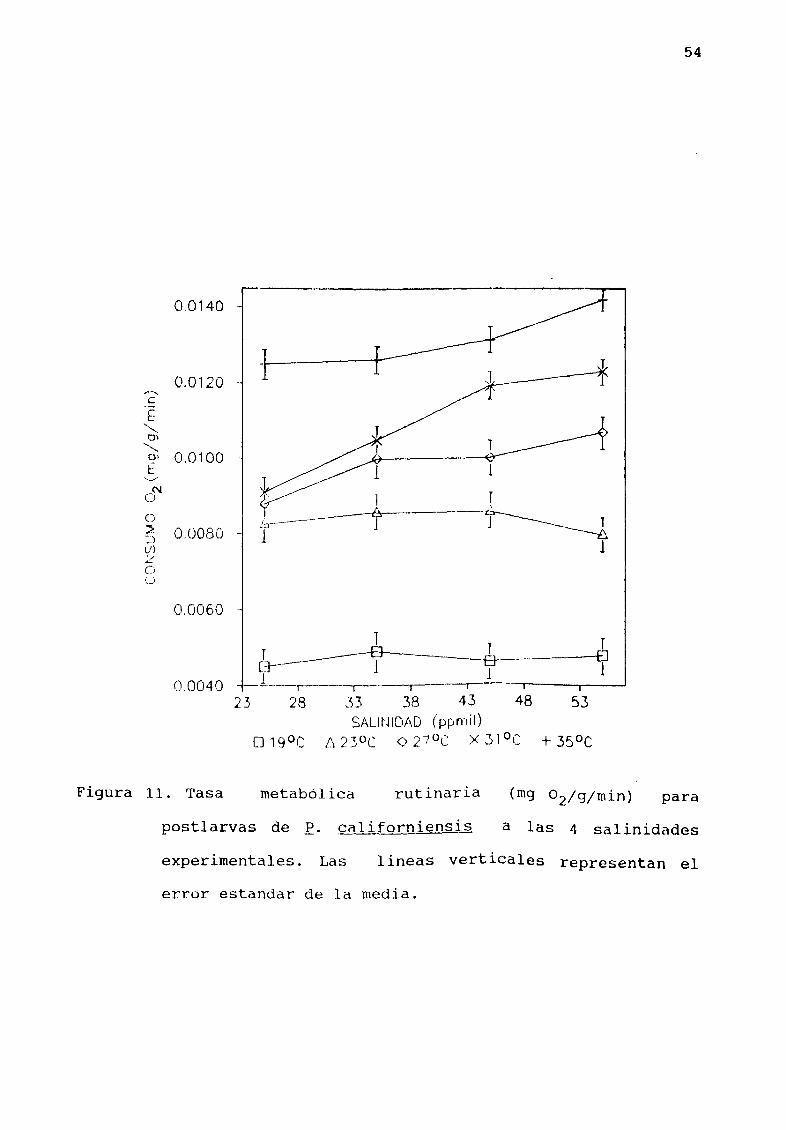
6 4 EFECTO DE LA SALINI DAD EN EL CONSUMO DE OXIGENO DE
POSTLARVAS DE E ealiforniensis
La relación entre la salinidad y el consumo de oXlgeno para

50
las temperaturas experimentales se presenta en la Figura 11 El
anÆlisis estadístico demostró que no hay diferencias
significativas P 0 5 en el consumo de oxígeno a las
diferentes salinidades experimentales a 19 23 Y 350C A 27 Y
310C se encontraron diferencias significativas P 0 001 en el
consumo de oxígeno a 25 ppmil con respecto a las demÆs
salinidades experimentales
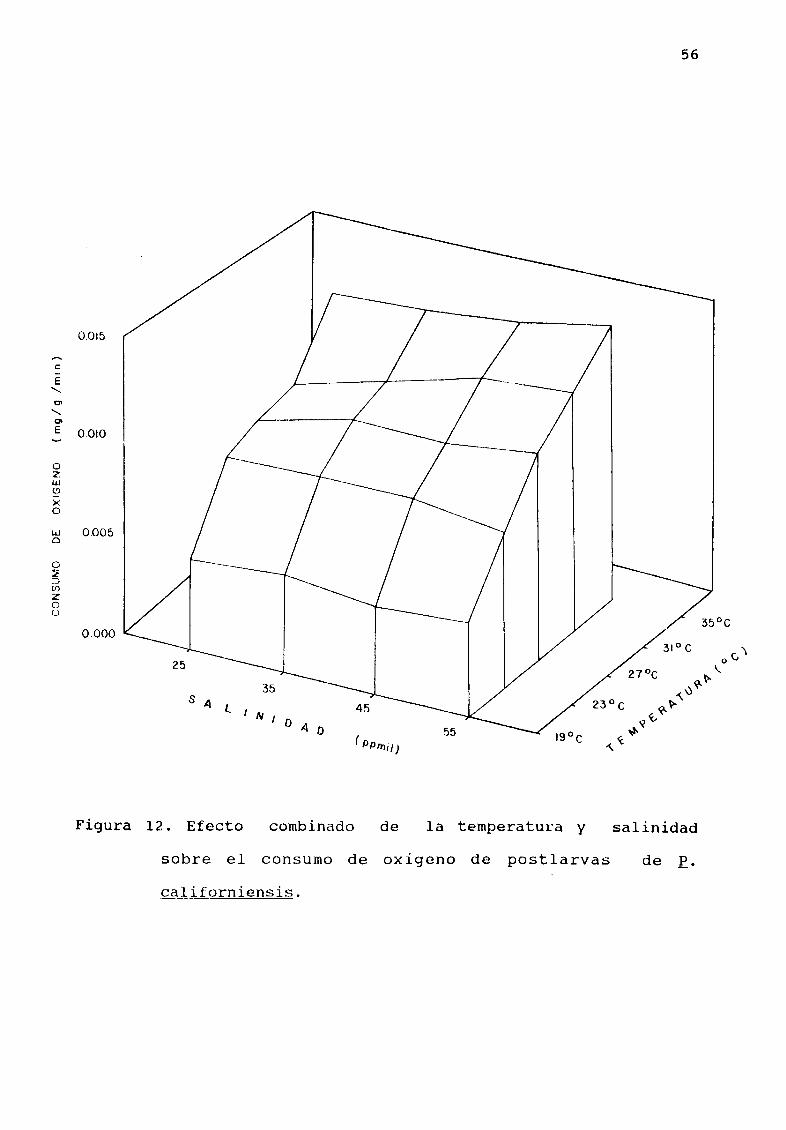
6 5 EFECTO COMBINADO DE TEMPERATURA Y SALINIDAD EN EL CONSUMO DE
OXIGENO DE POSTr RVAS DE E c Liforniensis
El valor mÆs alto en el consumo de oxígeno 0 0142
mgjgjmin correspondió a la combinación de la temperatura y
sal inidad mÆximas 3 50C y 55 ppmi 1 El valor mínimo 0 0045
mgjgjmin correspondió a la combinación de temperatura y
salinidad mAs baja 190C y 25 ppmil A 190C el consumo de
oxígeno es muy bajo media 0 0047 2E 4 Sin embargo al
incrementarse la temperatura a 230C el consumo de oXigeno
casi se duplica media 0 0084 3E 4 Por otro lado a 270C
el consumo de oxígeno es solamente 17 del consumo de oxígeno a
230C Una situción similar se presenta entre las temperaturas de
31 y 350C en las que el consumo de oXlgeno solo aumenta
aproximadamente 20

O 01 40
0 0120e
¸QI
J
E 0 0100
ND
O
J 0 0080lnZ
U
0 0060
0 0040
51
19 23 27 31 35
TEMPERATURA Oe
i 35 pprnil O 45 ppmil X 55 ppmil025 ppmil
Figura 9 Tasa metabóliea rutinaria mg 02 g min para postlarvas
de E califorDiensis a las 5 temperaturas experimentales
Las líneas verticales representan el error estandar de
la media

52
Tabla 3 Consumo de oxígeno mg g min de postlarvas de camarón
cafØ E californiensis a las diferentes salinidades y
temperaturas experimentales
TEMPERATURA oC
19 23 27 31 35
SALINIDAO
25 ppmilConsumo
Oy0 0045 0 0083 0 0088 0 0091 0 0125
mg g min a b b b c
e e 2 8E 4 3 1E 4 3 0E 4 2 9E 4 2 7E 4
35 ppmilConsumo Oy
0 0049 0 0086 0 0100 0 0105 0 0126
mg g min a b be e d
e e 5 7E 4 4 8E 4 4 2E 4 3 8E 4 4 4E 4
45 ppmilconsumo Oy
0 0046 0 0086 0 0100 0 0119 0 0131
mg g min a b c d d
e e 3 5E 4 3 2E 4 3 5E 4 3 3E 4 4 0E 4
55 ppmilconsumo Oy
0 0048 0 0080 0 0107 0 0123 0 0142
mg g mln a b e d e
e e 2 4E 4 2 5E 4 2 6E 4 3 2E 4 3 2E 4
Los valores que presentan la misma letra para
cada renglón no son significativamente diferentes
P 0 05
e e Error estandar
4 0
o
o
O3 0u
Owr
lL 2 0woC
1 0
53
5 0
0 019 23 27 31 35
RN GO DE TEMPERATURA OC
O 5 ppmil l 35 ppmil Ô 45 pprnil X 55 ppmil
Figura 10 Variaciones
postlarvas de E
en el eoeficiente tØrmico Q10 de
californiensis a diferentes
salinidades
0 0140
0 0120e
EIJ
o 0 0100t
NO
S 0 0080LZOU
0 0060
0 0040
Figura 11 Tasa
54
33 38 43 48
SALlt IDAD ppmílL 2 oC 0270 X3tOc
Tr
1I
T
1
23 28
o 190
metabólica rutinaria
postlarvas de E califQrniensis
T
1
T
1
T
J T
1
T
1
T—
1
53
350C
mg 02 g min para
a las 4 salinidades
experimentales Las 1 ineas verticales representan el
error estandar de la media

55
Tabla 4 Estimaciones del coeficiente tØrmico Ql0 a diferentes
salinidades para postlarvas de camarón cafØ
californiensis
S A L 1 N 1 D A D
INTERVALO DE
TEMPERATURA 25 ppmil 35 ppmil 45 ppmil 55 ppmil
19 230C 4 62 4 08 4 78 3 59
23 270C 116 1 46 146 2 07
27 310C 109 113 154 142
31 350C 2 21 158 128 143
El anÆlisis de regresión multiple para definir la proporción
con que cada uno de estos factores actœa dió como resultado la
ecuación
R 0 0059 0 0005 T 0 00005 S
r2 0 77 P 0 0001
donde R es el consumo de oXlgeno T la temperatura en el
intervalo de 19 a 350C y S la salinidad øentro del intervalo de
25 a 55 ppmil
0 015
c
E
01
Cl
E 0010
Oz
UJ
O
UJ 0005o
O
JJZ
Ou
0 000
56
25o
lo
23 oe lo
x
35S A L I N I D 4 o
45
PPfIlil
Figura 12 Efecto combinado la temperatura yde salinidad
sobre el consumo de oxígeno de post larvas de E
ca liforniensis

57
7 DISCUSION y CONCLUSIONES
7 1 ORGANISMOS EXPERIMENTALES
Los camarones utilizados provinieron del desove de una sola
hembra madura asegurando de esta manera que todos se encontraran
en la misma etapa de desarrollo al momento de realizar los
experimentos Por otro lado el alimento proporcionado a los
camarones contenía el nivel de proteína adecuado para el buen
crecimiento de los organismos Treece y Yates QQ cit por lo
que los aspectos nutricionales no tuv ieron un efecto en los
resultados obtenidos
7 2 DISEÑO EXPERIMENTAL
7 2 1 Medición del consumo de oXlgeno
Para las mediciones en el consumo de oxígeno se utilizaron
cÆmaras de tipo abierto por ser el sistema mÆs sencillo Los
resul tados obtenidos de esta manera fueron reproduc ibles y
consistentes en cada una de las series experimentales
Varios autores han seæalado la importancia de usar
blancos como control para los respirómetros Esto permite tener
una estimación de el consumo de oxígeno por microrganismos
Sutcliffe Q12 cit Scelzo y Zuæiga Qp cit y de otros
factores externos como el electrodo y el respirómetro De esta
manera el ajuste del consumo de oxígeno proporciona una mejor
estimación de los requerimientos metabólicos de la especie
Villarreal QQ clt

58
Villarreal 1990 por ejemplo indicó que el consumo del
blanco en un sistema paralelo fue de aproximadamente el 6 del
consumo de oxígeno en ˙h rax tenuimanus Para cada una de las
series experimentales del presente trabajo la disminución del
oxígeno disuelto en los blancos fue menor de 10 del consumo
por parte de los organismos
De acuerdo con Liao y Murai QQ çit es necesario un
período de aclimatación a las unidades experimentales antes de
realizar mediciones de la tasa metabólica de rutina Evitando así
que los valores obtenidos no sean representativos Para reducir
al mÆximo los efectos ocasionados por el manipuleo de los
organismos sobre su consumo de oxígeno el tiempo de aclimatación
en las unidades experimentales fue de 2 horas
7 2 2 Tratamientos Experimentales
Venkataramiahet Ql 1974 demostró que existe una
inter cción de la temperatura y la regulación osmótica con la
respiración pero enfatiza que Østa no debe de ser considerada
como un buen ídice de tolerancia osmótica y salina excepto en el
caso de que la temperatura experimental corresponda a la
condición natural del biotopo
De acuerdo con lo anterior las temperaturas y
salinidades seleccionadas para el presente trabajo son adecuadas
sj tomamos en cuenta que son similares al intervalo de variación
encontrado por
Chale B C S
HernÆndez y GonzÆlez QQ ctt en Puerto
19 a 320C y salinidad superior a 35 ppmil y

59
de datos de salinidad reportados por granjas de cultivo de
camarón en Sonora y Sinaloa 25 a 35 ppmil Por otro lado 350C
y 55 ppmil se seleccionaron para determinar el efecto de
temperaturas y salinidades altas en el consumo de oxígeno de las
postlarvas
7 3 RELACION DEL NIVEL DE OXIGENO DISUELTO CON EL CONSUMO DE
OXIGENO DE p californiensis
El consumo de oxígeno fue constante e independiente de la
saturación de oxígeno durante la prueba experimental Debido a la
duración del experimento 60 min no fue posible determinar el
límite inferior de independencia de Penaeus aliforniensis con
respecto a la saturación de oxígeno para cada una de las
salinidades y temperaturas experimentales Unicamente a las
temperaturas y saliniades mÆs altas se observó que el consumo de
oXlgeno no mantuvo la misma velocidad a concentraciones de OD
menores de 1 5 mg l s in embargo
estadisticamente signi ficat ivas La
estar encaminada a determinar el
experimentos de mayor duración
las diferencias no fueron
investigación futura deberÆ
punto crítico con base en
El grado de independencia metabólica en diferentes animales
es determinado por el punto donde el organismo se vuelve
dependiente de las concentraciones de oxígeno del medio Seidman
y Lawrence 1985 Este punto llamado comunmente punto crítico o
nivel del límite incipiente letal Fry 1947 se encuentra

60
usualmente entre 0 9 Y 2 O mg l OD para los camarones Egusa
1961 Mackay 1974 Liao y Murai 1986
Dall QQ eit encontró que el consumo de oxígeno de E
esculentus se volvió dependiente a concentraciones øe OD de 1 5
mg l 25 de saturación otros estudios vgr Subrahmanyan QQ
cLt Kulkarni y Joshi 1980 sugieren que las condiciones
experimentales son responsables de la aparente dependencia del
consumo de oxígeno de E indicus y E japonicus respectivamente
al nivel de saturación Liao y Huang 1975 y Chen 1985
encontraron que el consumo de oxígeno de postlarvas de E monQdon
es independiente de la concentración de OD por arriba de J 8
mg l de oo Villarreal y Ocampo 1991 por otro lado
encontraron que el consumo de oxígeno de postlarvas y juveniles
de E ç litorniensis era independiente de su concentración por
arriba de 1 8 mg l Con base en esta información y al presente
estudio es posible indicar que el límite incipiente letal pnra
postlarvas de alifo niensis probablemente se encuentra por
debajo de 1 5 mg l OD
La disminución del consumo de oXlgeno mg l min de E
calLforniensis a 35 ppmil puede relacionarse con el hecho de que
esta fue la salinidad de aclimatación y de que probablemente sea
el n ivel óptimo para las postlarvas de esta especie Kutty et ˙ì
9p ç Lt encontraron que el consumo de oxígeno de juveniles de
r ngiç era menor a 35 ppmil que a 7 y 21 ppmil indicando
una condición estresiva para la especie a bajas salinidndes Por

61
su p3rte Hewitt 1992 encontró que la salinidad óptima para
juveniles de E aliforniensis se eneuentra entre 25 y 35 ppmil
Segun Villarreal y Hewitt 1991 desde el punto de vist a
bioenergØtico esto puede relacionarse con la minimizdción de la
energía destinada al metabolismo rutinario y la maximización de
la energía destinada al crecimiento cuando las condiciones
medioambientales son óptimas
7 4 EFECTO DE LA TEMPERATURA EN r TASA METABOLICA RUTINARIA DE
POSTLARVAS DE E calitorniensis
La temperatura es el mayor determinante ambiental de la tasa
metabólica en los invertebrados marinos Scholander et QI 1953
Leffler 1972 Kinne 1977 Armitage y Wall 92 cit sobre
todo en aquellos que viven en ambientes estuarinos Newell 1979
Darsey QJ cit dado que las variaciones de la temperatura son
mayores que en mar abierto en donde los valores de temperatura
son comparativamente mÆs bajos y estables Scelzo y ZuiÜga QQ
cit
Se encontró que existe una relación directa entre el consumo
de oXigeno y la temperatura Esta relación ha sido reportada
previamente para otros crllstº ceos vgr McFarland y Pickens
1965 Haefner 1969 Dimock y Grones 1975 Hart 1980 A 25
ppmil ver Tabla 3 y Figura 9 se presenta un intervalo de
temperatura entre 23 Y 310C en el que no hubo un cambio
significativo en la tasa de consumo de oxígeno Esta zona estable

62
puede relacionarse con el hecho de que a bajas salinidades el
camarón que por naturaleza es hiposmótico realiza un esfuerzo
osmótico menor para mantener la homeostasis Seidman y Lawrence
QD cit permitiendo mantener un nivel metabólico constante para
ese intervalo de temperaturas A 35 ppmil se presenta un patrón
similar entre 27 y 310C
El consumo de oxígeno a 190C para todas las salinidades fue
muy bajo debido quizÆ a que esta temperatura se encuentra fuera
del intervalo óptimo de la especie por lo que el metabol ismo
rutinario se reduce significativamente Aldrich et al 1968
indican que las postlarvas del camarón cafØ aztecus se
muestran inactivas a temperaturas menores de 170C Darsey QJ2
Lt por su parte encontró que a lSoC y salinidades de 10 25
Y O ppmil el aumento en peso longitud producción de tejido y
sobrevivencia para E çalitgrniensis eran considerablemente
menores que los valores obtenidos a temperaturas de 25 y 320C
A 3SoC y a 25 ó 35 ppmil el consumo de oxígeno de p
cqljLorniensis aumentó en forma considerable en relación con los
valores a 310C para las mismas salinidades Esto quizÆ se deba a
que el gasto energØtico que realiza el organismo a 350C es
principalmente para mantener su metabolismo basal y permanecer
vivo bajo estas condiciones Bowler 1963 encontró que las
temperaturas letales afectan la habilidad metabólica y reguladora
de iones de los organismos

63
7 5 COEFICIENTE TERMICO Q10
Los valores de QIO a bajas temperaturas ver Tabla 4
indican una alta sensibilidad a las variaciones de temperatura
Wol vekamp y Waterman QQ cit que se traduce en una
inadecuada capacidad tØrmica reguladora dentro de este intervalo
Zuæiga g al 1982
Por ejemplo Yagi et 9ì Qn cit en su estudio realizado
con EsLlaemon serratus encontraron valores altos de QIO 5 89
dentro del intervalo de temperatura de 13 290C a una salinidad de
13 ppmil Dalla Via CQP cit encontró valores de Q10 entre 3 y
4 en el intervalo de temperatura de 5 150C para postlarvas de
Eenaeuselegans los cuales deerecieron con el aumento de la
temperatura Schlieper 1952 en Yagi et al 91 cit seæalo
que valores de QIO menores de 2 indican una marcada euritermia
Como se mencionó anteriormente para el intervalo de
temperatura de 19 a 230C los valores de Q10 altos muestran una
alta sensibilidad a las variaciones en la temperatura
disminuyendo considerablement e para los intervØllos entre 23 Y
350C A estas temperaturas los valores de Q10 a todas las
salinidades se mantuvieron entre 1 y 2 5 Valores muy similares
de QIO a 35 ppmil pnra el intervalo de temperatura de 19 y 350C
fueron encontrados por Villarreal y Ocampo Qg cit Debido al
ciclo de vida de 1 californiensis PØrez Farfante 1969 es
posible sugerir que las postlarvas han desarrollado una capacidad
adaptativa a variaciones en la temperatura lo cual representa

64
una ventaja para la migración de un medio marino a uno costero
en donde los parÆmetros ecológicos son mÆs inestables Yagi tl
al cit
7 6 EFECTO DE LA SALINIDAD EN LA TASA METABOLICA RUTINARIA DE
POSTLARVAS DE E californiensis
En tØrminos generales la sal inidad no produee di ferencias
significativas en el consumo de oxígeno de E californiensis
aunque existe una tendencia a incrementar dicho consumo a la
mayor salinidad experiment al Figura 11 Scelzo y ZϾiga QQ
cit reportaron resultados similares para E brasiliensis
A 19 Y 230C el consumo de oXigeno se mantiene practicamente
constante con las variaciones en la salinidad Por otro lado a
310C se observan niveles de consumo de oxígeno a 25 y 35 ppmil
similares a los consumos a 270C Esto quizÆ se deba a la
capacidad que tienen los organismos estuarinos de soportar
sal inidades subnormales a temperaturas cercanas a su nivel
incipiente letal Zein Eldin y Aldrich 1965 Sandifer 1973
7 7 EFECTO DE LA TEMPERATURA Y LA SALINIDAD EN TASA METABOLICA
RUTINARIA DE POSTLARVAS DE E c iforniensis
La ecuación obtenida por medio del anÆl isis de regresión
mœltiple indica que la temperatura tiene un efecto mÆs
significativo que la salinidad sobre la tasa metabólica rutinaria
de las postlarvas de J çALtfgrniensis Esta situación ha sido

65
reportada anteriormente en otros crustÆceos
cit Oimoek y Groves QQ cit Liao y Muray
Zuæiga Q cit Yagi et al QQ cit
vgr Leffler Qg
QQ cit Scelzo y
En las Figuras 9 y 12 se observa una zona de temperatura
entre 23 y 270C en la cual el consumo de oxígeno es muy similar
para todas las salinidades lo que parece indicar que e tos
valores de temperatura representan el intervalo óptimo de la
especie Por otro lado la capacidad de adaptación a variaciones
en la salinidad parece disminuir conforme aumenta la sal inidad
por encima de 45 ppmil y 27oC Darsey QQ cit encontró que a
25 ppmil y 250C se producen los mayores incrementos en tamaæo
peso y producción de tej ído así como una mayor sobrevivencia
para postlarvas de E californiensis Sin embargo aun es
necesario llevar a cabo estudios que corroboren estos resultados
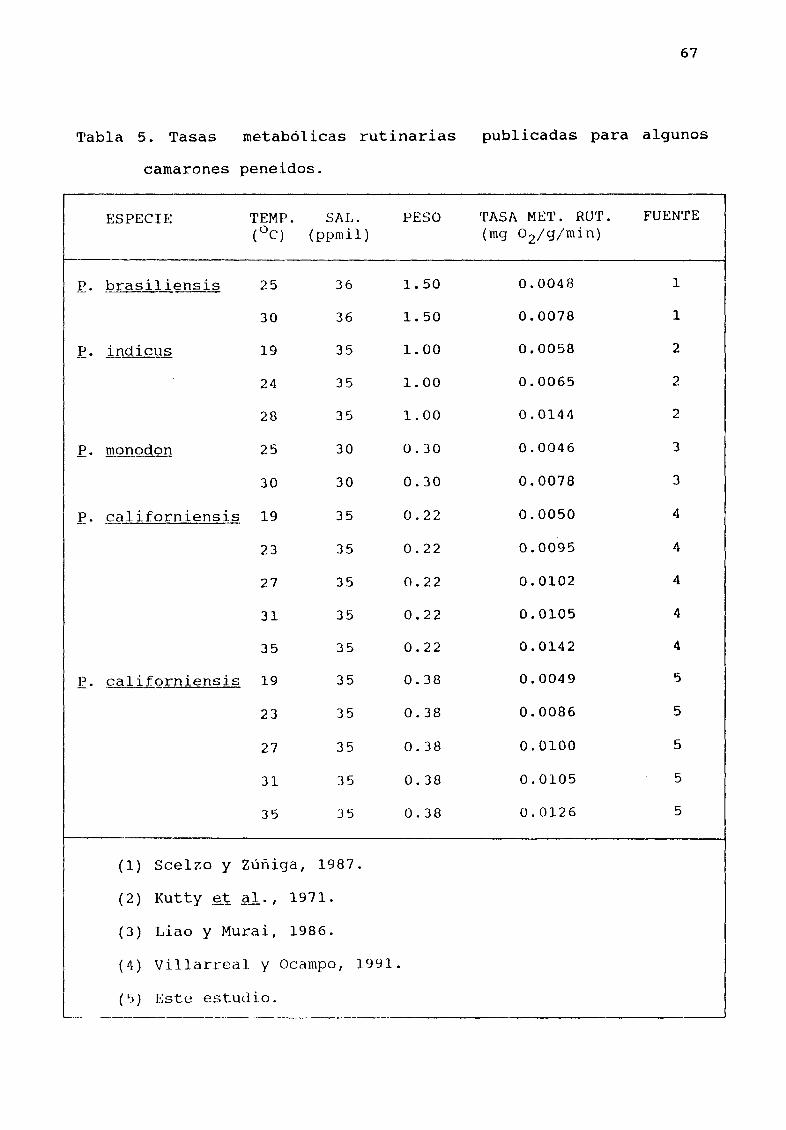
La Tabla 5 presenta una comparación de la tasa metaból ica
rutinaria de E californiensis a 35 ppmil y otros camarones
peneidos En general los valores obtenidos en el presente estudio
se encuentran dentro del intervalo de los valores reportados para
otras especies
Para reducir las posibles variaciones en la tasa
metaból ica de rutina las mediciones del consumo de oxígeno se
real izaron a la misma hora y baj o las mismas condiciones en el
presente experimento sin embargo la determinación del consumo
de oxígeno a corto plazo tiene el inconveniente de que no se
en la tasaincorporan algunos factores que son determinantes

66
metabólica Dall QJ2 cit como son el ritmo cardiaco Sacayanan
y Hirata 1986 y el intervalo metabólico de actividad
Villarreal 912 cit Por esta razón la investigación futura
deberÆ incorporar la realización de experimentos que involucren
el ciclo circadiano
La información obtenida en el presente trabajo indica que el
consumo de oxígeno se ve influenciado significativamente por la
temperatura Sin embargo el organismo muestra una capacidad de
adaptación metabólica a variaciones en la temperatura dentro de
un intervalo definido entre 23 y 270C Investigaciones
posteriores deberÆn enfocarse a la evaluación de otros parÆmetros
intrínsecos y extrínsecos como son la densidad de cultivo
la nutrición y el estado de muda Dall QD cit vi11arreal Q2
cLt ya que la información obtenida permitirÆ optimizar el
cultivo de la especie Wiesepape 1970
67
Tabla 5 Tasas metabólicas rutinarias publicadas para algunos
camarones peneidos
I ESPECIE lEMP SAL PESO TASA MET RUT FUENTE
0C ppmil mg 02 g min
E masiliensis 25 36 1 50 0 0048 1
30 36 150 0 0078 1
E i nQj clJΠ19 35 100 0 0058 2
24 35 1 00 0 0065 2
28 35 100 0 0144 2
E mQJ1Qgon 25 30 0 30 0 0046 3
30 30 0 30 0 0078 3
E liforniensis 19 35 0 22 0 0050 4
23 35 0 22 0 0095 4
27 35 0 22 0 0102 4
31 35 0 22 0 0105 4
35 35 0 22 0 0142 4
califQrniensis 19 35 0 38 0 0049 5
23 35 0 38 0 0086 5
27 35 0 38 0 0100 5
31 35 0 38 0 0105 5
35 35 0 38 0 0126 5
1 Scelzo y ZϾiga 1987
2 Kutty t al 1971
3 Liao y Murai 1986
4 Villarreal y Ocampo 1991
l Este estudio

68
8 LITERATURA CITADA
AQUACOP 1975 Maturation and spawnlng in captivity of penaBid
shrimp Penaeus merguiensis de Man E ponicus Bate E
aztecus Ives MetaJ2enaeus ensis de Haan and Penaeus
emjsulcatus de Haan Eroc Worlg aricul SQ 6 123 132
ALDRICH D V C E WOOD y K N BAXTER 1968 An ecological
interpretation of low temperature responses in P lS
Œz ecus and E 2 tlL L lS postlarvae Bu l Mar Se i 18 1
61 71
ARCE R A 1989 Cultivo larvario de camarones peneidos
Reporte interno del Centro de Investigaciones Biológias de
B C S La Paz MØxico 125 p
ARMITAGE K B Y T J WALL 1982 The effects of body size
starvation and temperature accl imation on the oxygen
consumption of the crayfish Or gnec nais Com iochem
EhYsiol 73A 1 63 68
BARDACH J E J H RYTHER Y W O Mc LARNEY 1972
Aquaculture Th jarming ang husbançlry of LreshY 1Lter
marine grgŒnisms Wiley Interscience Sydney New York
868 p
BARRENA B 1987 La camaronicultura PrÆctica reciente en
MØxico i˙ lay˝sión 8 4 7
BENNETT A F 1978 Activity metabolism in the lower
vertebrates Annu R Y PhysioJ 40 447 469
BOWLER K 1963 A study of the factors involved in acclimation

69
to temperature and death at
pallipes 11 Experiments at
Comp Physiol 62 133 146
BOYD C E 1990 Water Quality management for pond fish
cul ture In DeveloJ2ments in Aquacul turg and Fisher ies
c ence Elsevier New York 482 p
BRIDGES C R y A R BRAND 1980 Oxygen consumption and oxygen
independence in marine crustaceans M EcoJ PrQ Ser
high
the
temperatures in AstaQIJ
tissue level Cell
2 133 14l
BRODY S 1964 Bioenergetics and growth with special reference
to the efficiency complex in domestic animals Hafner
Publishing Co New York 1923 p
BROOM J G 1971 Shrimp culture In proceedings of the first
annual workshop of the QrlA Maricuh SQç 1 63 68
BUNN S y P HORWITZ 1980 The oxygen consumption of Cherfx
n1Jj man t and Che ra x gestr1d tQr a1Q ictl s
Deapoda Parastacidae in response to temperature salinity
and oxygen concentration Honours Thesis University of
Western Australia 71 p
CASILLAS H R Y B J MAGALLON 1988 sustitución de insumos
tradicionales en las dietas para la engorda del camaron
Informe interno C l B 15 p
CASILLAS H R V RUIZ J MORAN F RANGEL B J MAGALLON M
ARVIZU y M Hernandez 1988 Crecimiento y sobrevivencia
del camarón azul E n us stvlirostris bajo seis
tratamientos de cultivo rœstico en Puerto Chale B C S
MØxico Reporte interno C l ß
22p

70
CICTUS 1982 El cultivo del eamarón azœl penaeus stylirostris
Centro de Investigaciones Científicas y Tecnológieas de la
Universidad de Sonora Hermosillo Sonora 126 p
CLIFFORD H C y R W BRICK 1978 Protein utilization in the
freshwater shrimp Macrobrachium rose bergii Proc World
aric lt oc 9 195 208
COCKCROFT A C y T WOOLDRIDGE 1985 The effects of mass
temperature and moulting on the respiration of Kacropetasma
africanus Decapoda penaeida çQ Bjg ç he J Jlysiol
81A 1 143 14 8
CHEN H C 1985 Water quality criteria for farming the grass
shritTIP PenØt llŒ l1gno lQn Proc of the first international
conference on the culture of penaeid prawn shrimps lloilo
Philippines December 4 7 1 165
CHIANG P y l C LIAO 1986 The
P nagl monQiÌQlJ culture in Taiwan
practice of grass prawn
from 1968 to 1984J
Worl 1 Iicul fgh 16 297 315
DALL W 1986 Estimation of routine metabolic rate in a penaeid
prawn penaJ ls SCllIt usJ EI Mar Hol Ecol
96 57 74
DALLA VIA G J 1985 Oxygen consumption and temperature change
in the shr imp Pl Ll9 JIQn JElgans AT icoJ EQ CL ger
26 199 202
DALLA VIA G J 1987a Effects of salinity and temperature on
oxygen consumption in a freshwater population of
pa 1 a llQD t 0Jl J na rj lΠC rustacea Dec apod a COlllJ

71
Biochem Physiol 88A 2 299 305
1987b Salinity responses in brackish water
populations of the freshwater shrimp Palaemonetes
antennarius l Oxygen consumption Co ioçhe Physiol
87A 2 471 478
DARSEY K T 1990 The influence of temperature salinity on
the growth and survival of California brown shrimp Penaeus
califQrnienŒiŒ Holmes M Sc Thesis University of
Arizona 43 p
DAVIES P S 1966 Physiological ecology of P ttell 1 1 rhe
effect of body size and temperature on metabol ic rateL
Mar io 46 647 658
DIMOCK R V y K H GROVES 1975 Interaction of temperature
and salini ty on oxygen consumpt ion of the estuar ine crab
p anope1J1 1erbsJii MarßJol
33 301 308
DUGEER D 1990 Shrimp farming in Mexico yesterday today and
tomorrow Àgu YJLturg Maqazine 16 4 38 53
EGUS1 S 1961 Studies on the respirat ion of the kuruma 11
prawn Pen us iŒPoni us Bate 11 preliminary experiments
on its oxygen consumption B JL J D SQ S fish 27
650 659
FLETCHER C R 1976 A phenomenological description of active
transporto In Davies C Ed Persp tiy s in xperimenta
ZoglQ9Y pergamon Press Oxford 1 55 64
FRY F E 1 1947 Effects of the environment on animal
activity g ßlolr
55 1 62

72
GARCIA A III Y D E BRUNE 1989 Oxygen transport in shrimp
cul ture ponds American Society of Agricul tural Engineers
1989 1 13
GAUDY R Y L SLOANE 1981 Effect of salinity on oxygen
consumption in postlarvae of the penaeid shrimps Eena us
mono on and Pßna us s9Llirostris without and with acclimation
Mar iQ 65 297 301 HAEFNER
P A Jr 1969 Temperature and salinity tolerance ofthe
sand shrimp Cran Qn te spinosa Say hYsiologi alZQolQgy
33 4 288 317 HART
R C 1980 Oxygen consumption in Cari 9Jna ni 1gJJca Decapoda
Atyidae in relation to temperature and size Ereshw
jolo 10 3 215 222 HERNANDEZ
JL Y M GONZALE7 1989 Rendimiento de producción de
camarón azœl E ena lJ Œ tylirostrisJa diferentes densidades
de cultivo semi intensivo en Puerto Chale B
C S MØxico Tesis de Licenciatura U A B C S 98
p HEWITT
DAVID 1984 Growth and bioenergetics of the freshwater prawn
Macrobrachium a lstraltnse Honours Thesis James Cook University
of North Queensland 84 p HEWITT
RICHARD 1992 Efecto de la salinidad en el crecimiento superv
ivencia y el consumo de ox ígeno de j uveni1 es de camarón
cafØ Pen1 lscal iforniensis Holmes 1900 TØsis de Licenciatura
U A B C S 80 p HILL
R W 1980 Fisiología animal comparada un enfo ue

73
ambiental Editorial Reverte S A Espaæa 901 p
HUGHES D G 1991 Manejo de la calidad del agua en estanques
con Ønfasis en camaronicul tura Taller sobre cultiva de
eamarón MazatlÆn Sinaloa MØxico julio 1991 31 p
JOHNSON F A H ERYND Y M J POLISSAR 1954 KiQ tl˙ basis Q
IDoL lar giotQgy John Wiley and Sons 874 p
KADER H A 1962 Ionic influenee on oxygen consumption in
Metal enaellŒ monQçeros FabriciusJ bnim r1ol2hQL Physiol
9 32 40
KINNE O 1970 Temperature Invertebrates Kinne O Ed
Chapter 3 31 of ar Q EcoJ gy Wjley Interscience London
1 1 407 515
KINNE O 1971
Chapter 4 of
Salinity Invertebrates Kinne
ar ne RçglQgy Wiley Interscience
O Ed
London
1 2 821 996
KINNE O 1977 q in EcOlQgy Wiley Interscience N Y 324 p
KITANI H Y J N ALVARADO 1982 The larval developrnent of the
pacific brown shrimp Eena us
cqlifQrn ensis Holmes reared in
the laboratory ßul JÞll Q˙ Sci Fis 48 3 375 389
KNUT NIELSEN S 1990 nirnat r llYsiolQ9Y Cambridge Univürsity
Press New York U S A 602 p
KRAMER G L 1975 Studies on the lethal dissolved oxygen
levels for young brown shrimp P oa s aztecus Proceedings
Worlq
M˙lc Qu h SQ 6 157 167
KULKARNI G K Y P K JOSHI 1980 Some aspects of respiratory
metabol ism of a penaeid prawns E na ld illigniçll Crustacea

74
Decapoda Penaeídae Qbiologia 75 27 32
KUTTY M N G MURUGAPOOPATHY y T S KRISHNAN 1971 Influence
of salinity and temperature on the oxygen consumption in
young juveniles of the Indian prawn Penaeus indicus 1ar
Bio 11 125 131
LANZA ESPINO G Y J L ARREDONDO 1990 La acuacul tura en
MØxico de los conceptos a la producción Instituto de
Biología Universidad Nacional Autónoma de MØxico 315 p
LEFFLER C W 1972 Some effects of temperature on the growth
and metabol ic rate of j uven i1 e b1 ue crab s ˙g11 in ctes
saDi in the laboratory Mgr ßio 14 2 104 110
LIAO 1 C y H J HUANG 1975 Studies on the respiration
of economlC prawns in Taiwan l Oxygen consllmption ílnd
lethal disso1ved oxygen of egg up to young prawn of rßn us IDorr
Qgn FabriciusJ Fis Tniwan 4 1 50 LIAO
l o y T MURAl 1986 Effects of dissolved oxygen
temperature and salinity on the consumption of the grass
shrimpI lΠna cus 1JQEQ90I In J L Maclean LB Dizon
and L V Hosillos Eds Tl llŒfirsJ Astan fisheries Kor
Asian Fisheries Manila Philippines Nov 28 30 1
641 646 McFARI
ND W N y P E PICKENS 1965 The effects of season temperature
and salinity on standard and active oxyqen consumption
ot the grass shrimps J2Al ftlJ 1QJ1 Ji yul˙lgJis Say
Canaqj n J Qplo 43 571 585 McKAY
R D 1974 A note on mínimal levels ofoxygen required

75
to maintain life in Penaeus shrnitti proceedings World
Maric lt Soc 5 451 452
MAGALLON F J 1988 El cultivo del camaron
California SUr Reporte interno
Investigaciones Biológicas de La Paz Baja California Sur
en Baj a
Centro dedel
23 p
MENASVETA P 1980 Effect of ozone treatment of the survival of
prawn larvae Macroþr hium os nQ rgj de Man reared in a
closed recirculating water systern In New M B Ed
J iant prawD fral1LLnI Elsev ier Sc ientific Publ ishing co
Amsterdam 1 95 303
NEILL W H 1989 Effects of temperature and oxygen on biology
of aquatic organisms implications for aquaculture
Abstracts Aquacul ture 89J Worlg Aguacul ture Soc
20 1 59A
NELSON S C D A ARMSTRONG A W KNIGTH y H W LI 1977
The effeets of temperature and sal ini ty on the metabol ic
rate of juvenile Macro 2¨chi1Jm rosenbergii Crustacea
Palaemonidae ç ßjochem hY ol 6A 67 72
NEW M B 1987 Feed and feeding of fish and shrimp
A mIac1Jl t1Jre peY lgpment 9nq ˙Q ˙lrgin9ttQJ1 pQgLill R@grJ
26 FAO Rome Italy
NEWELL R C 1979 Biology of intertidal animals Marirl
E ological S ry s Ltd Faversham Kent 326 p
ODUM E P 1971 fundam ntals QL ecolQgy Saunders
Philadelphia 546 p

76
PEREZ FARFANTE l 1969 Western Atlantic shrimps oi the genus
PenaeYâ U S Fish Wildlife Service Fish BuJ
673 461 591
PRIMAVERA J H 1985 A review of maturation and reproduction
in closed thelycum penaeids In proceedings of the first
international conference on the culture of penaE id
prawns shrimps Iloilo City Philippines December 4 7
1 47 64
PROSSER C L y F A BROWN 1968 Fisiología comparada
Interamericana MØxico 966 p
RAO K P 1958 Oxygen consumption as a function of Size and
salinity in Metapena us rnonQ ceJg Fab From marlne and
brackish water enverionments J esp ßiQJ 35 307 313
ROSENBERRY B 1989 Worlq Shrimp FaImi g 1989 Aquaculture
Digest Annual Report 28 p
ROSENRERRY B 1990 WOJ ld JLrimJ2 Farming 1990 Aquaculture
Digest Annual Report 40 p
HOSENBERRY B 1991 W o Jd ShrÌJ DI FØlrmjJg 1991 Aquaculture
Digest Annual Report 40 p
RPI 1989 Penaeid technology short course CET del Mar La Paz
B C S MØxico
SACAYANAN E B Y H HlRATA 1986 Circadian rythm of feeding
respiration in kuruma prawn Penaeus Ì0J onLQJ2 MinL Rey
D ta FL1 fisJh R 4 63 76
SAENZ M 1987 El cultivo de camaron en MØxico Secretaría
de Pesca MØxico 28 p

77
SANDIFER P A 1973 Effects of temperature and salinity on
larval development of the grass shrimp Pal aemonetes
yulqaris Decapoda Caridea Fish Bu 71 L 115 125
SCELZO A M Y o ZUÑIGA 1987 Consumo de oxígeno en el camarón
rena s brasiliensis Decapoda Penaeidae en relación a la
salinidad y la temperatura Fundación de Ciencias Naturales
La Salle Puerto Madryn Chubut Argentina 47 127 128
SCHLIEPER O 1952 Ubier die Temperatur Stoffwechselrelation
einiger eurythermer Wassertiere YIT hl tsch Zggl çeE
SY2DJ 16 5 267 272
SCHOLANDER P F W FLAGG V
Climatic adaptation in arctic
rhysiol ZQol 26 67 92
SEIDMAN E R Y A L LAWRANCE
WALTERS y L IRVING 1953
poikilothermsand tropical
1985 Growth fced digestibility
and proximate body consumption of juvenile Pen eu y pnal ei
and Pena ls 1gn Qg gn grown at di fferent dissolved oXY0en
levels J ¨ricul S 16 333 346
SOKAL R R Y F J ROHLF 1984 iQ1 ry Freeman Co Ed
San Francisco 776 p
SPOTIE S 1979 ßeayJ fL r Muari Jl1s Th captiyg nvironment
JOhn Wiley and Sons New York 413 p
STATGRAPHICS 1986 9 ati tical GrqQhics y em Statistical
Craphical Corporation Maryland U S A
STICKNEY R R 1979 PLinçJ ples QJ YHlrmW iìt L Muacultlre John
liley and Sons New York 375 p
SUBRAHMANYAM C B 1962 Oxygen consumption in relation to body

78
weight and oxygen tension in the prawn Penaeus indicus
Milne Edwards Proc Ingian Acad sci 55B 152 161
SUTCLIFFE D W rr R CARRICK y W H MOORE 1975 An
automatic respirometer for determining oxygen uptake in
crayfish Austropota obius gallip s over periods of 3 4
days J BiQ 63 673 668
SWAIN R P F MARKER Y A M RICHARSON 1987 Respiratory
responses to hypoxia in stream dwelling ð tacop js
fr nkLlnii and burrowing P9rasta 2otçt 1 asTIaniclJli
parastacid crayfish ˙Q E Biochem Physiol 87A 3
813 817
TREECE D G Y E M YATES 1990 Laboratory manual for the
culture of penaeid shrimp larvae Sea Grant Course Programo
Texas A M University 95 p
ISAI C K 1989 Water qual ity management proceedings of the
Southeast Asia Shrimp Farm Management Workshop July 26
August 11 1989 77 p
TSENG W Y 1987 Shrimp mariculture A practical manual
Departament of Fisheries Chien Chang Publisher University
of Papua New Guinea 305 p
VENKATARAMIAH A G J LAKSMI y G GUNTHER 1974
the effect of sal ini ty and temperature on the
shrimp PenaelJs aztlŒ ç1J1 Ives with spec ial
survival limits growth oxygen consumption
regulation Gulf Co as1 Res J ab Ocean
Studies on
cornmercial
regard to
and ionic
Springs
Miss Rep H 74 134 p

79
VERNBERG F J 1970 oissolved gases animals
Ed Marine Ecology Wiley Interscience
1 1491 1515
VILLARREAL H 1984 Water quality in aquaculture proceedings
o the Conference of aquaculture in Australia
Kooralbyn Qld October 12 13
VILLARREAL H 1989a Feeding growth and energetics of the
freshwater crayfish ˙hera enuim n s with special emphasis
on its potential for commercial culture Ph D Thesis
University of Queensland Australia 249 p
1989b Prawn culture in MØxico Austasia
In Kinne o
New York
Muacultlre Magazin 3 6 17 18
VILLARREAL H 1990 Effect of the temperature on oxygen
consumption and heart rate of the Australian crayfish Ch
gnuimanls smith çomIBlQ
çhem EhYsiQL 91A 1 189 193
VILLARREAL H y R HEWIrr 1991 Effect of salinity on the
oxygen consumption and growth ot juvenile brown shrimp
Penal2 lS ˙ï LliíQKDJ l2nsis Holmes 1900 Poster 22nd Annual
Conference and Exposition WSLtld Ag @culJ J Society San
Juan Puerto Rico June 16 20 1991
VILLARREAL H Y L OCAMPO 1991 Effect of Size and temperature
on the oxygen consumption of the brown shr imp Penae us
ç¨lifQ rni nsis H o1m es 1 9 O O P os ter 2 2 ndAnnua1
Conference and Exposi tion Wgrlg lillL0˙IJJtlJr Qc ieJy San
Juan Puerto Rico June 16 20 1991
WIESEPAPE L M 1970 Effects of temperature and salinity on

80
thermal death in postlarvae brown shrimp Penaeus aztecus
M Sc Texas A M University 85 p
WOLVECAMP N R Y T H WATERMAN 1960 Respiration The
QhysiQJ gy of çru tace Vol 1 Academic Press New York
YAGI H Y H J CECCALDI 1984 Influence combinØ des facteurs
tempØrature et salinitØ sur le mØtabolisme et la croissance
larvaire de la crevette rase PalaemQll serratus
Pennant crustacØ DecapodØ PalaemonidØ Aquaculture
37 73 85
YAGI H H J CECCALDI y R GAUDY 1990 Combined influence of
temperature and salinity on oxygen consumption of the larvae
of the pink shrimp Pa aemQn serr tus Crustacea Decapoda
palaemonidae hql1 CILtur 86 77 92
ZEIN ELDIN Z P y D V ALDRICH 1965 Growth and survival of
postlarval J2 llØl lS azte t under controlled condi tions of
temperature Clnd salinity BiQJ5 111 129 1 199 216
ZUÑIGA o A R A WILSON y E A OYARCE 1982 Consumo de
oxígeno del camarón de roca hyn QQcinete tY2Ys H Milne
Edwards su relación con la temperatura y talla ˙iencia y
Tecnolog˝q de Mar 6 53 60