
EVALUACIÓN DE LA PRODUCCIÓN DE ÁCIDO GIBERELÍCO DE BACTERIAS NATIVAS DEL GENERO Azospirillum sp AISALADA DE SUELOS DEL
DEPARTAMENTO CORDOBA
CARMEN LUCÍA LUNA RAMOSMIGUEL EDUARDO MONTOYA RAMOS
DIRECTORA:Ph.D CECILIA LARA MANTILLA
UNIVERSIDAD DE CÓRDOBAFACULTAD DE CIENCIAS BÁSICAS
DEPARTAMENTO DE QUÍMICAPROGRAMA DE QUÍMICA
MONTERÍA-2014

OBJETIVOS
OBJETIVO GENERAL
Evaluar la producción de ácido giberelíco de bacterias nativas del genero
Azospirillum sp aisladas de suelos del departamento de Córdoba.
OBJETIVOS ESPECÍFICOS
Multiplicar las bacterias nativas de genero Azospirillum sp existentes en el
banco de muestras del laboratorio de biotecnología teniendo en cuenta las
condiciones nutricionales y factores de crecimiento.
Evaluación de la producción del ácido por el método de reactivo ácido
fosfomolíbdico (folling-wu).
Seleccionar las mejores bacterias teniendo en cuenta su producción de
ácido.

PLANTEAMIENTO DEL PROBLEMA
Actualmente los altos costos de los fertilizantes químicos, así como de los
problemas de contaminación que ocasionan, han generado otras formas de
producción, en donde se incentiven a la práctica de nuevas tecnologías. Estudios
de costos de producción señalan la reducción en la utilización de agroquímicos por
los agricultores en los últimos ocho años, debido a los altos costos que generan
estos insumos, para lograr el máximo rendimiento de los cultivos (Laria et al.,
2003).
Es por esto, que una de las alternativas que puede contribuir con la disminución
de fertilizantes de síntesis química, es la generación de técnicas de producción
agrícola, enfocada al uso eficiente de los recursos que tiendan a una agricultura
sostenible y de bajo impacto agrícola. En este sentido, el interés de adoptar
tecnologías limpias, mediante el manejo de microorganismos con potencial
biofertilizantes, se ha convertido en la mejor alternativa para reducir estos
impactos, y así mismo contribuir con el mejoramiento de las condiciones
medioambientales, económicas y sociales (Laria et al., 2003).

INTRODUCCIÓN
El crecimiento normal de la planta es ocasionado en forma armónica por
sustancias que funcionan como hormonas. Los principales grupos de ellas son las
auxinas, Giberilinas, citoquininas, etileno e inhibidores de crecimiento. Estos
compuestos actúan en bajas proporciones. Durante el desarrollo fenológico de las
plantas, estas varían de acuerdo con el estado de desarrollo, mediante un
delicado balance hormona-inhibidor. Una forma de ataque del patógeno consiste
en alterar ese equilibrio, ya sea mediante la obtención de reguladores de
crecimiento, para aumentar su concentración y así crear los inhibidores de esas
hormonas (Aguilar-Piedras, J.J. 2008).
Las Giberilinas (GAs), como reguladores de crecimiento están estrechamente
asociadas a la promoción de la germinación de las semillas, crecimiento del tallo,
inducir la brotación de yemas y el desarrollo de los frutos. Existen varios tipos de
Giberilinas, siendo las más comunes: Giberelina A1 (GA1), ácido Giberelíco (GA3),
Giberelina A4 (GA4), Giberelina A7 (GA7) y Giberelina A9 (GA9). Químicamente son
un grupo de diterpenoides que se definen más por su estructura que por su
actividad biológica, contrario a lo que ocurre con las auxinas y las citoquininas
(Yamaguchi y Kamiya 2000).
Únicamente las Giberilinas biológicamente activas pueden cumplir con estas
funciones, las Giberilinas no bioactivas existen en el tejido vegetal como
precursores de las formas bioactivas o como metabolitos desactivados. Se ha
dilucidado que existe una necesidad estructural, como requerimiento para la
afinidad con el recientemente descubierto receptor de Giberilinas en arroz (GID1),
y sus homólogos en otras especies (Nakajima et ál. 2006). Parece ser que la
regulación de la biosíntesis de Giberilinas y de sus receptores, y vías de
señalización dependen de la especie de estudio (Yamaguchi 2008).
Teniendo en cuenta lo anterior es importante el estudio de bacterias productoras
de Giberilinas.

ANTECEDENTES
En algunos estudios de campo realizados con cereales se ha llegado a observar
una promoción del crecimiento vegetativo (Kapulnik et al., 1983). También se ha
reportado un efecto a simple vista sobre el crecimiento de varios vegetales como
tomate (Lycopersicon esculentum Mill.), berenjena (Solanum melongena L.),
pimiento (Capsicum annuum) y algodón (Gossypium barbadense) (Bashan et al.
1989)
Bashan & Levanony (1990), Summer (1990), Fages (1994), Okon y Labandera-
González (1994), citados por Dobbelaere et al. (2001), concluyen que la
inoculación con Azospirillum sp. Incrementó significativamente de un 5-30 % en el
rendimiento de materia seca foliar, granos y proteína foliar, así como en el
desarrollo radical, en alrededor de 60 -70 % de los experimentos en campo.
Richards eta, (2001) dice que las Giberilinas son hormonas que regulan el
crecimiento y desarrollo de las plantas; a través de la promoción de la división y
elongación celular Matusa-göttgens y Hedden, (2009).
Weaver (1996) afirma que las Giberilinas son las únicas sustancias químicas
capaces de promover floración en plantas, que de otro modo permanecerían
vegetativas, al remplazar condiciones ambientales específicas que controlan la
formación de flores. Sin embargo, Salisbury y Ross (1994) mencionan que
también las Citoquininas han estimulado la formación de flores en algunas
variedades de crisantemo (Dedranthema sp.) con el empleo de una combinación
de Citoquininas (Benziladenina) y AG5.
Ngamau (2001) probó aplicaciones de BAP (Benzilaminopurina) y/o AG3 sobre
plantas de la variedad ´Green Goddess` de Z. aethiopica. Estas promovieron la
emergencia temprana e incrementaron el número de brotes visibles y el desarrollo
de estos. De Pascale et al. (2003) demostró que la Giberilinas adelantó floración
en aproximadamente 100 días respecto al testigo.

Luria y Weiss (2005) concluyeron que se incrementó el número de brotes y se
obtuvo hasta cinco veces más flores con tratamientos combinados de BA y AG3.
Reiser (1998) reportó que aplicaciones de AG3 en Z. aethiopica y Z. ´Green
Goddess` incrementaron el número de flores por planta de 1.3 a 3.4 y de 1.3 a 3.8,
respectivamente.

JUSTIFICACIÓN
Debido a la problemática mundial existente hoy en día por el uso indiscriminado de
productos fertilizantes de síntesis química, se han implementado con el paso de
los años soluciones amigables y sostenibles que permitan disminuir el efecto
adverso generado por dichas aplicaciones; de allí que nació el uso de productos
biofertilizantes basados en la potencialización de procesos naturales donde los
actores principales son los microorganismos promotores de crecimiento vegetal
Aguilar-Piedras, (2008).
Las plantas para su desarrollo, no solo realizan procesos fotosintéticos o toman
sustancias minerales del suelo, también llevan a cabo en su interior una serie de
procesos fisiológicos que ayudan a dicho desarrollo. De la misma manera, las
plantas también se ven afectadas y de cierta forma reguladas por algunos de los
procesos que ocurren en el suelo, la mayoría de los cuales se llevan a cabo por
los microorganismos (Pedraza et al. 2008; Molina-Favero et al. 2008).
El desarrollo normal de una planta depende de la interacción de factores externos:
luz, nutrientes, agua y temperatura, e internos: hormonas. Una definición global
del termino hormona, es considerar bajo este nombre a cualquier producto
químico de naturaleza orgánica, que sirve de mensajero y que es producido en
una parte de la planta, tiene como “blanco” otra parte de ella (Conney, Nonhebel,
1991).
Dentro de las bacterias que producen se tiene el género Azospirillum; este se
caracteriza por ser un microorganismo promotor de crecimiento vegetal, que ha
sido ampliamente estudiado, a partir de sus capacidades para generar efectos
benéficos por su inoculación a distintas especies vegetales. Sin embargo aún falta
información a nivel genético y bioquímico, de la regulación y capacidad de cada
uno de sus componentes enzimáticos presentes en procesos como la producción
de Auxinas. (Carreño-Lopez et al. 2000; Costacurta et al.. 1994; Ryu & Patten,
2008; Sergeeva, et al. 2007; Pedraza et al. 2004; Contesto et al. 2010).

Azospirillum ha sido ampliamente estudiado como bacteria promotora de
crecimiento vegetal, ya que tiene la capacidad por un lado, de producir sustancias
como sideróforos, definidos como moléculas de bajo peso molecular que
presentan una alta afinidad por Fe generando un efecto quelante en éste,
igualmente tiene la capacidad de producir bacteriocinas como sustancias
antimicrobianas que generan un efecto de control contra patógenos que puedan
llegar a atacar a las plantas (revisado en Baca, 2009; Somers et al. 2005; Borisov
et al. 2007).
Las Giberilinas son un grupo de diterpenoides que se definen más por su
estructura que por su actividad biológica, contrario a lo que ocurre con las auxinas
y las citoquininas (Yamaguchi y Kamiya 2000).
Las Giberilinas biológicamente activas, actúan como reguladores esenciales del
desarrollo de las plantas, y cubren todos los aspectos de la historia de vida de las
plantas, modulando varias respuestas del crecimiento como la germinación de
semillas, el crecimiento del tallo, la partenocarpia, la expansión foliar, la
elongación de la raíz, la floración y la liberación de enzimas hidrolíticas en algunos
tejidos (Ueguchi-Tanaka et ál. 2007). Únicamente las Giberilinas biológicamente
activas pueden cumplir con estas funciones, las giberelinas no bioactivas existen
en el tejido vegetal como precursores de las formas bioactivas, o como
metabolitos desactivado (Castro-Guerrero, et al, 2011).
MARCO TEÓRICO

BACTERIAS PROMOTORAS DE CRECIMIENTO
Definición
Las bacterias promotoras de crecimiento en vegetales (PGPRs) son un grupo de
diferentes especies de bacterias que pueden incrementar el crecimiento y
productividad vegetal (Bashan y Holguín, 1998). Dentro de los microorganismos
promotores de crecimiento vegetal se clasifican una gran variedad de géneros
dentro de los cuales se incluyen Azospirillum, Azotobacter, Pseudomonas,
Bacillus, Rhizobium (Baca, 2009; Klopper, 1993).
Son capaces de colonizar los diferentes sistemas radicales de las plantas y
promover el crecimiento de éstas, pueden alterar el crecimiento de los tejidos
vegetales directa o indirectamente; indirectamente las PGPR juegan un papel
importante en la disminución o prevención de otros fitopatógenos que puedan
atacar las plantas, mientras que las directamente las PGPR promueven el
crecimiento de la planta al sintetizar compuestos (por ejemplo reguladores del
crecimiento vegetal) que le facilita a la planta la toma de nutrientes del ambiente
(Gray & Smith, 2004).
Historia de Azospirillum sp: Spirillum lipoferum, ahora llamado Azospirillum, fue
descrito por primera vez en 1925 por Martinus Willem Beijerinck, luego de lo cual
la bacteria permaneció en el olvido por varias décadas. Las observaciones de
Juan José Peña-Cabriales y Johanna Döbereiner en 1973, inician la época
moderna de este microorganismo.
Hiltner en 1904, observó por primera vez la acumulación de microorganismos en la
zona radical y propuso el término rizósfera. Los exudados radiculares,
conformados por sustancias diversas crean alrededor de las raíces, un ambiente
nutricional enriquecido que favorece el crecimiento bacteriano.

Generalidades Género Azospirillum sp: El género Azospirillum sp, es una
bacteria Gram negativa, perteneciente a la subclase alfa de las proteobacterias, en
el orden de las Rhodospirillaceae, dentro de sus principales características
Las bacterias del género Azospirillum sp., tienen la capacidad de producir auxinas,
citoquininas y giberelinas en medios de cultivo. No obstante, el mecanismo
analizado con mayor amplitud ha sido la producción de auxinas, que puede
modificar el contenido de fitohormonas de las plantas conduciendo a la
estimulación del crecimiento de las mismas, como el caso del Ácido Indol Acético
(AIA), el cual induce al aumento de pelos radiculares, logrando una mayor
captación de nutrimentos.
Metabolismo de Azospirillum sp: Las especies de Azospirillum sp, difieren en su
capacidad para utilizar diferentes compuestos como fuentes de carbono y
nitrógeno. Estas bacterias usan para su crecimiento unos pocos monosacáridos y
disacáridos así como alcoholes polihidroxilados, y principalmente diversos ácidos
orgánicos tales como málico y succínico y algunos aminoácidos (Hartmann &
Burris, 1988). Para el uso de diferentes fuentes de carbono, tanto A. lipoferum
como A. brasilense tienen todas las enzimas de la vía de Embden-Meyerhof-
Parnas y de la vía de Entner-Doudoroff (Westby et al, 1983), así como todas las
enzimas del ciclo de los ácidos tricarboxílicos (Martínez et al, 1984). Tanto A.
lipoferum como A. brasilense tienen la capacidad de crecer autotróficamente con
H2 y CO2, proceso en el que participa la ribulosa-1,5- difosfato carboxilasa (Tilak
et al, 1986).
Efectos de factores fisicoquímicos sobre Azospirillum sp.
Cuando las bacterias se siembran en un medio de cultivo óptimo y bajo
condiciones adecuadas de incubación, ocurre un incremento en el número de
células en períodos muy cortos. En algunas especies se alcanza la población
máxima en 24 horas, en cambio, en otras se necesita un período más prolongado
para alcanzar el máximo desarrollo; para determinar el desarrollo se necesita
medir cuantitativamente la población de células al tiempo de sembrar y
nuevamente después de la incubación. El ciclo de desarrollo de las poblaciones

bacterianas es el procedimiento más importante en la multiplicación de los
microorganismos (Peoples & Craswell, 1992).
pH: Azospirillum sp., tiene un crecimiento óptimo en un rango de pH de 6,5 -7,0.
Si el microorganismo no cuenta con el pH correspondiente, esto provocaría la
inhibición de su crecimiento y desarrollo (Sánchez, 1964). Para Azospirillum
brasilense: pH 6,0-7,8; Azospirillum lipoferum: pH 5,7-6,8; Azospirillum
amazonense: pH 5,7-6,5; Azospirillum halopraeferens: pH 6,0-8,0; Azospirillum
doebereinerae: pH 6,0-7.0.
Temperatura: Muchas especies de Azospirillum sp., requieren una temperatura
óptima de crecimiento cercana a los 30 ºC, excepto para Azospirillum largimobile
que presenta una temperatura de 28 ºC y Azospirillum halopraeferens 41º C. La
modificación de la temperatura, tiene un efecto notable sobre un proceso. Si el
valor utilizado no es adecuado puede disminuir o aún impedir la formación de un
metabolito determinado.
Además, la temperatura puede modificar los requerimientos nutritivos de algún
microorganismo, lo que significa que al modificarse el valor de un factor puede
cambiar los requerimientos de otro.
LA GIBERELINA: Son reguladores del crecimiento de las plantas superiores que
regulan numerosos aspectos del desarrollo vegetal. Ellas promueven el
crecimiento celular debido a que incrementan la hidrólisis de almidón, fructanos y
sacarosa con lo que se originan moléculas de fructosa y glucosa.
Las respuestas que producen o modulan las Giberilinas afectan tanto la regulación
del crecimiento reproductivo de la planta (Azcon & Bieto, 2000). Ellas son
determinantes en el control de la elongación del tallo, también modifican
sustancialmente los procesos reproductivos de los vegetales, participando en el
control del inducción de la floración, en la producción, crecimiento, y desarrollo de

los frutos. Así mismo sustituyen los requerimientos de luz o frio que precisan
muchas de las semillas para germinar (Azcon & Bieto, 2000; Salisbury, 1994).
Para determinar las Giberilinas se usa el método Reactivo de folling-Wu (ácido
fosfomolíbdico); es un método utilizado en la detección de Giberilinas, como un
método sencillo y practico de detección, basado en la reacción de reducción que
presenta el ácido molíbdico frente a soluciones de ácido giberelíco a diferentes
concentraciones. La reacción de reducción ocasiona un cambio de color del
reactivo a azul mostrando la reducción ocasionada por la presencia de Giberilinas
en la solución (Graham & Henderson, 1960)
METODOLOGÍA
EVALUACIÓN DE LOS MICROORGANISMOS

ACTIVACIÓN DE LOS MICROORGANISMOS
Las cepas a utilizar pertenecen al banco de cepas del laboratorio de
Biotecnología GRUBIODEQ de la Universidad de Córdoba. Los microorganismos
serán activados utilizándose como medio de cultivo: Nfb; de acuerdo a lo anterior,
se comenzará con la inoculación de los microorganismos y seguidamente se hará
una dilución de dicho inóculo (Solución madre), para así determinar la cantidad de
células de las que se partirá.
DETERMINACIÓN DE LA CONCENTRACIÓN MICROBIANA: Para esta determinación se emplearán los siguientes métodos:
MEDIO DE CULTIVO NFB (Dobereiner et al, 1997)
El medio de cultivo que se utilizó para realizar estos estudios será el NFB, que se
usará en la multiplicación de bacterias del género Azospirillum sp. El cuál será
preparado de la siguiente manera; Para 500mL de este medio pesamos 0.25g
K2HPO4, 0.025g FeSO4*7H2O, 0.005g MnSO4*H2O, 0.05g MgSO4*7H2O, 0.01g
NaCl, 0.005g CaCl2, 0.001g NA2M0O4, agregamos 500mL de agua destilada y
ajustamos el pH entre 6.6 a 7.0. Luego lo colocamos en autoclave por un tiempo
de 20 minutos a una temperatura de 121°C aproximadamente, dejamos enfriar a
temperatura ambiente e inoculamos los diferentes microorganismos
DETECCIÓN COLORIMETRICA DE GIBERELINAS: REACTIVO ÁCIDO FOSFOMOLÍBDICO (FOLLING-WU)
La determinación de ácido giberélico con el reactivo Folling-Wu, se realizará a
partir de soluciones stock de Giberilinas comercial Merck; a diferentes
concentraciones, buscando establecer una curva de calibración para la detección
de GA3 en base al reactivo.
Se separarán 500mL del reactivo Folling-Wu, a partir de 35g de ácido molibdico y
5g de Tugstanato de Sodio fueron agregados a un Becker de 1L, luego se agregó
200 cc de NaOH al 10% y después 200 cc de agua destilada. Calentar

vigorosamente el contenido del frasco por 40 minutos para remover cualquier
rastro de amonio presente en el ácido molibdico.
Dejar enfriar y diluir la mezcla hasta 350 cc con agua destilada y agregar 125 cc
de ácido fosfórico al 85%. Finalmente la mezcla fue transferida a un matraz de
500 cc y aforada con agua destilada.
Una vez preparadas las soluciones de reguladores de crecimiento se realizó la
evolución del reactivo, a una proporción de 25mL de la muestra por 15mL del
reactivo, el tiempo de relación será de 60 minutos, a baño en ebullición. El tiempo
cero será estimado luego de 2min de sumergidos los tubos, pasados los 60min se
retirarán del baño y se dejan enfriar en un baño de hielo por unos 5 minutos
aproximadamente.
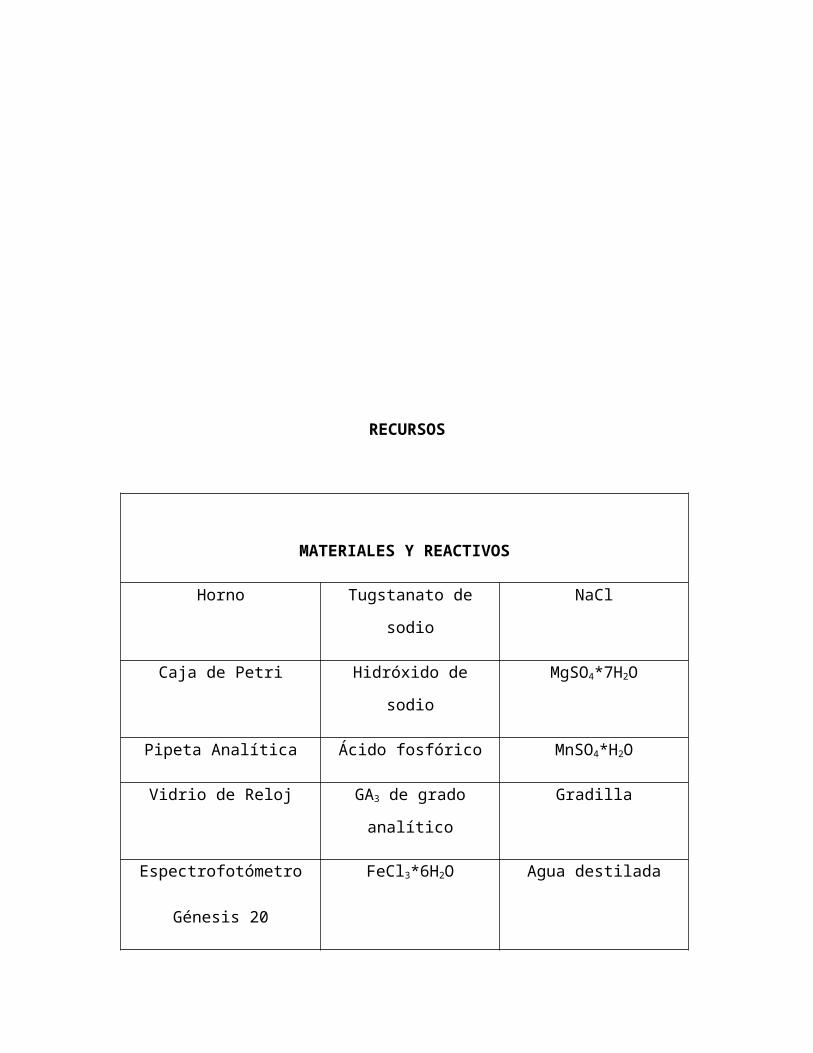
RECURSOS

MATERIALES Y REACTIVOS
Horno Tugstanato de sodio NaCl
Caja de Petri Hidróxido de sodio MgSO4*7H2O
Pipeta Analítica Ácido fosfórico MnSO4*H2O
Vidrio de Reloj GA3 de grado analítico Gradilla
Espectrofotómetro
Génesis 20
FeCl3*6H2O Agua destilada
Pipeta de vidrio HCl Frasco lavador
Autoclave Agar
Becker CaCl2
Tubos de ensayo FeSO4*7H2O
Espátula K2HPO4
Campana de esterilización Agitador de vidrio
Balanza analítica Pera
RECURSO HUMANO
Ph.D Cecilia Lara Mantilla.
INFRAESTRUCTURA

Laboratorio de investigación en biotecnología GRUBIODEQ (Programa de
Química, Universidad de Córdoba).
RECURSOS ECONÓMICOS
GRUBIODEQ (Programa de química, Universidad de Córdoba).
CRONOGRAMA
ACTIVIDAD TIEMPO (MESES)

1 2 3 4 5 6 7 8 9 10 11 12
REVISIÓN BIBLIOGRÁFICA
RECOLECCIÓN DE LA MATERIA PRIMA
CULTIVO DE MICROORGANISMO
S
CUANTIFICACIÓN DE GIBERELÍNAS
CON REACTIVO DE FOLLING-WU
ANÁLISIS E INTERPRETACIÓN DE RESULTADOS
REDACCIÓN DEL INFORME FINAL
PRESUPUESTO

RUBROS JUSTIFICACIÓN VALOR TOTAL
REACTIVOS
Necesario para el cultivo de
las bacterias, extracción y
cuantificación de la GA3.$7’000.000°°
VIDRIERÍA
Necesaria para el desarrollo
total del proyecto.$4’000.000°°
EQUIPOS:
Autoclave,
espectrofotómetro,
horno, balanza
analítica, Campana de
esterilización.
$9’000.000°°
VALOR TOTAL $11’000.000°°
REFERENCIAS

Celis, L. 2008. Estandarización de métodos de detección para promotores
de crecimiento vegetal (ácido indol acético y Giberilinas) en cultivos
microbianos. Trabajo de grado (Microbiólogo agrícola y veterinario).
Pontificia Universidad Javeriana, Bogotá.
Pérez, E. 2012. Inoculación de bacterias promotoras de crecimiento vegetal
en pepino (CucumsativusL.). Tesis (Maestría en ciencias). Institución de
Enseñanza e Investigación en Ciencias Agrícolas, Montecillo-México.
Baca B. E., Elmerich C. 2000. Microbial production of plant hormones.
Associative and endophytic nitrogen-fixing bacteria and cyanobacterial
associations.
Graham H. D., Thomas L. B., 1960. Rapid, Simple Colorimetric Method for
the Determination of Micro Quantities of Gibberellic Acid, Journal of
Pharmaceutical Sciences, Vol 50, N° 1, 44- 48.
Hedden P. 1999. Recent Advances in Gibberellin Biosynthesis. Journal of
Experimental Botany. (50) 553- 563.
Molina-Favero, C; Mónica, C; Simontacchi, M; Puntarulo, S; Lamattina, L.
2008. Aerobic Nitric Oxide Production by Azospirillum brasilense Sp245 and
Its Influence on Root Architecture in Tomato. The American
Phytopathological Society. 21: 1001–1009.
Pedraza, R; Bellone, C; Carrizo de Bellone, S; Fernández, P; Teixeira, K.
2008. Azospirillum inoculation and nitrogen fertilization effect on grain yield
and on the diversity of endophytic bacteria in the phyllosphere of rice rainfed
crop. European J. Soil. doi:10.1016/j.ejsobi.2008.09.007, pp. 1 – 8.
Baca, B.E. 2009. Base moleculares de la interacción planta-bacteria:
Modelo Azospirillum Benemérita Universidad Autónoma de Puebla.
Universidad de Tucumán. Centro en Ciencias Microbiológicas.
Somers, E; Ptacek, D; Gysegom, P; Srinivasan, M; Vanderleyden, J. 2005.
Azospirillum brasilense Produces the Auxin-Like Phenylacetic Acid by Using
the Key Enzyme for Indole-3-Acetic Acid Biosynthesis Applied. Enviromental
Microbiology. 71: 1803-1810.
Borisov, I; Schelud’ko, A; Petrova, L; Katsy, E. 2007. Changes in
Azospirillum brasilense motility and the effect of wheat seedling exudates.
Microbiological Research, doi:10.1016/j.micres.2007.07.003.
Carreño-López, R; Campos-Reales, N; Elmerich, C; Baca, B.E. 2000.
Physiological evidence for differently regulated tryptophan-dependent
pathways for indole-3-acetic synthesis in Azospirillum brasilense. Mol. Gen.
Genet. 264: 521 – 530.
Contesto, C; Milesi, S; Mantelin, S; Zancarini, A; Desbrosses, G;
Varoquaux, F; Bellini, C; Kowalczyk, M; Touraine, B. 2010. The auxin-
signaling pathway is required for the lateral root response of Arabidopsis to
the rhizobacterium Phyllobacterium brassicacearum. Planta. 232: 1455–
1470.
Costacurta, A; Keijers, V; Vanderleyden J. 1994. Molecular cloning and
sequence analysis of an Azospirillum brasilense indole-3-pyruvate
decarboxylase gene. Mol. Gen. Genet. 243: 463–472.
Ryu, R; Patten, C. 2008. Aromatic Amino Acid-Dependent Expression of
Indole-3-Pyruvate Decarboxylase Is Regulated by TyrR in Enterobacter
cloacae UW5. J. Bacteriol. 190: 7200 – 7208.

LARIA, G. et al., Fijador de Nitrógeno. Azospirillum sp. San Salvador, 2003.
Aguilar-Piedras, J.J; Xiqui-Vásquez, M.L; García-García, S; Baca, B.E.
2008. Producción del ácido indol-3-acético en Azospirillum. Rev. Latinoam.
Microbiol. 15: 29 – 37.
Bashan & Levanony (1990), Summer (1990), Fages (1994), Okon y
Labandera-González (1994), citados por Dobbelaere et al. (2001).
Bashan, Y. y G. Holguin. 1998. A proposal for the division of "plant growth-
promoting rhizobacteria" into two classifications: biocontrol-plant growth-
promoting bacteria and plant growth-promoting bacteria. Soil Biol. Biochem.
30: 1225-1228.







