
Centro de lnvestigación Científica de Yucatán, A.C.
Posgrado en Ciencias Biológicas
ESTUDIOS MORFOLÓGICOS Y MOLECULARES
DURANTE LA HISTOGÉNESIS DE LOS
EMBRIONES SOMÁTICOS DE CHILE HABANERO
(Capsi.cum ch/.nense Jacq.) EN PRESENCIA DE2,4-D
Tesis que presenta
SUSANA ALEJANDRA AVILES VIÑAS
En opción al título de
DOCTOR EN CIENCIAS
(Ciencias Biológicas: Opción Bioquímica y Biología Molecular)
Mérida, Yucatán, México
2012


CENTRO DE INVESTIGACIÓN CIENTÍFICA DE YUCATÁN, A. C.
POSGRADO EN CIENCIAS BIOLÓGICAS
OSGRADO EN
CIENCIASBIOLÓGICAS
RECONOcllvllENTO
Por medio de la presente, hago constar que el trabajo de tesis titulado `.ESTUDIOS
MORFOLÓGlcos y MOLEcuLAREs DUFmNTE LA HlsTOGÉNEsls DE Los
EMBRIONES SOMÁTICOS DE CHILE HABANERO (Caps/.cüm chí.nense Jacq.) EN
PRESENCIA DE 2,4-D fue realizado en los laboratorios de la Unidad de Bioquímica y
Biología Molecular de Plantas del Centro de lnvestigacíón Científica de Yucatán, A.C. bajo
la dirección de la Dra. Nancy Santana Buzzy y el Dr. José Juan Zúñiga Aguilar, dentro de
la opcíón de Bioquímica y BÍología Molecular, perteneciente al Programa de Posgrado en
Ciencias Biológicas de este Centro.
Atentamente,
Director Académico
Mérida, Yucatán, México, septiembre 2012.


DECLAFUCION DE PROPIEDAD
Declaro que la información contenida en la sección de Materiales y Métodos
Experimentales, los Resultados y Discusión de este documento proviene de las
actividades de experimentación realizadas durante el período que se me asignó para
desarrollar mi trabajo de tesis, en las Unidades y Laboratorios del Centro de lnvestigación
Científica de Yucatán, A.C., y que a razón de 1o anterior y en contraprestación de los
servicios educativos o de apoyo que me fueron brindados, dicha lnformación, en términos
de la Ley Federal del Derecho de Autor y la Ley de la Propiedad lndustrial, le pertenece
patrimonialmente a dicho Centro de lnvestigación. Por otra parte, en virtud de lo ya
manifestado, reconozco que de Ígual manera los productos intelectuales o desarrollos
tecnológicos que deriven o pudieran derivar de lo correspondiente a dicha información, le
pertenecen patrimonialmente al Centro de lnvestigación Científica de Yucatán, A.C., y en
el mismo tenor, reconozco que si derivaren de este trabajo productos intelectuales o
desarrollos tecnológicos, en lo especial, estos se regirán en todo caso por lo dispuesto por
la Ley Federal del Derecho de Autor y la Ley de la Propiedad lndustrial, en el tenor de lo
expuesto en la presente Declaración.
Firma:
M.C. SUSANA ALEJANDRA AVILES VIÑAS


Este trabajo se llevó a cabo en la Unidad de Bioquímica y Biología Molecular de Plantas
del Centro de lnvestigación Científíca de Yucatán, y forma parte del proyecto titulado"Recalcitrancia del género Caps/.cum: estudios morfológicos, bioquímicos y moleculares
relacionados con los factores que afectan la capacidad de regeneración de plantas de
chile /.n v/.fro (Ciencia básica CONACYT)", bajo la dirección de la Dra. Nancy Santana
Buzzy.


AGRADECIMIENTOS
Le agradezco a la Dra. Nancy Santana Buzzy, por haber creído en mí y por su disposición
para d'irigir en todo momento el presente trabajo de tesis.
AI Dr. José Juan Zúñiga Aguilar por su asesoría y por brindarme su apoyo en la parte
molecular del presente trabajo.
A los integrantes de mi comité tutorial y revisores de tesis: Dra. Lourdes lglesias Andrew,
Dra. Guadalupe López Puc, Dra. Sara Luz Nahuat Dzip, Dra. lleana de la Caridad
Echevarria Machado, Dr. Javier Mijangos Cortés y Dr. Luis Sáenz Carbonell, por sus
valiosas críticas y sugerencias.
A la M. C. Adriana Canto Flick y al M. C. Eduardo Balam Uc, por el apoyo técnico durante
la realización de este proyecto.
A rnis amígos y compañeros de laboratorio: Daniela, Eunice, Carlos L., Jericó, Raúl,
Carlos R., Stephanie, Emily, Alejandrina, Lupita, Karime, Jacobo y Mario, por brindarme
su amistad.
A las familias Avilés Viñas y López Alzina, por su apoyo incondicional durante la
realización de mis estudios de posgrado.


DEDICATORIAS
A MAXIMO, ROMINA Y NICOLAS
LOS AMO


lNDICE
INTRODUcclóN
CAPÍTUL01
ANTECEDENTES
1 GÉNERO CAps/cuM
1.1 lMPORTANCIA DEL GÉNERO CAPS/CUM
1.2 CHILE HABANERO (CAPS/CUM CH/WE~SE JACQ
1.2.1 DESCRIPCIÓN TAXONÓMICA
1.3 CULTIVO DE TEJIDOS VEGETALES
1.3.1 REGENERAclóN DE pLANTAs w v/rRo
1.4 EMBRloGÉNESIS SOMÁTICA
1.4.1 FACTORES INVOLUCRADOS EN LA EMBRIOGÉNESIS SOMÁTICA
.........1
1.5 REGULADORES DE CRECIMIENT0 VEGETAL EN LA INDUCCIÓN DE LA
EMBRIOGÉNESIS SOMÁTICA
1.6 GENES INVOLUCRADOS EN LA EMBRIOGÉNESIS SOMATICA
1.7 REGENERACION DE CHILE (CAps/cuM spp.) w v/rRo
1.8 JUSTIFICACIÓN
1.9 0BJETIVOS
1.9.10BJETIVO GENERAL
1.9.2 0BJETIVOS ESPECÍFICOS
BIBLIOGRAFÍA

CAPíTULO 11 .................
MORPHO-HISTOLOGICAL AND ULTRASTRUCTURAL STUDY ON DIRECT SOMATIC
EMBRYOGENESIS 0F CAPS/CUM CH/~E~SEJACQ. lN LIQUID MEDIUM
2.1 INTRODUCTI0N
2.2 MATERIALS AND METHODS
2.2.1 PLANT MATERIAL SOURCE
2.2.2 SOMATIC EMBRVO INDUCTION
2.2.3 STATISTICAL ANALVSIS
2.2.4 SCANNING ELECTRON MICROSCOPY
2.2.5 HisTOLOGICAL ANALysIS
2.3 RESULTS
2.3.1 EFFECT OF 2,4-D AND CULTURE MEDIUM CONSISTENcy
2.3.2 ANATOMICAL OBSERVATioNS
2.3.3 ULTRASTRUCTURAL OBSERVATIONS
2.4 CONCLUsloNS
2.5 LITERATURE CITED
CAPÍTULO 111
33
33
34
34
35
36
36
36
38
38
43
45
48
50
.........55
ANÁLISIS MORFOLÓGICOS E HISTOLÓGICO DE LOS EMBRIONES SOMÁTICOS DE CHILE
HABANER.O CAPSICUM CHINENSE
3.1 lNTRODUCCIÓN
3.2 METODOLOGÍA
3.2.1 ANÁLISIS MORFOLÓGICOS E HISTOLÓGICO DE LOS EMBRIONES SOMÁTICOS ...... 56

3.2.1.1 PREPARACIÓN DE LAS MUESTRAS PARA INCLUIR EN RESINA
3.2.1.2 PREPARACIÓN DE LAS MUESTRAS PARA INCLUIR EN PARAFINA
3.3 RESULTADOS
3.3.1 CARACTERIZACIÓN MORFOLÓGICA DE ESS DE CHILE HABANERO
3.3.2 CARACTERIZACIÓN HISTOLÓGICA DE ESS MADUROS DE CHILE HABANERO
3.4 DISCUSIÓN
3. 5 CONCLUSIONES
BIBLIOGFMFÍA
CAPÍTULO IV .........,........ .................69
EVALUACIÓN DE LA EXPRESIÓN DE UN GEN TIPO WUSCHEL DUFUNTE LA
EMBRIOGÉNESIS SOMÁTICA DEL CHILE HABANERO
4.1 lNTRODUCCIÓN
4.2 METODOLOGÍA
4.2.1 EVALUACIÓN DE LA EXPRESIÓN DE UN GEN "PO WUS EN ESS DE CHILE HABANERO ............ 70
4.2.2 EXTRACcióN DEL ARN
4.2.3 TFuTAMiENTO DEL ARN CON DNASA 1
4.2.4 CUANTIFICACIÓN Y COMPROBACIÓN DE LA INTEGRIDAD DEL ARN TOTAL
4.2.5 SÍNTESIS DE ADNC: TRANSCRIPCIÓN REVERSA DEL ARN
4.2.6 DISEÑO DE CEBADORES PARA EL GEN TIPO WL/S
4.2.7 AMPLIFICACIÓN DEL GEN TIPO Wl/S POR RT-PCR DE PUNTO FINAL
4.2.8 PURIFICACIÓN DE PCR
4.2.9 ANÁLISIS DE LAS SECUENCIAS

4.3 RESULTADOS
4.3.1 EXTRACCIÓN DE ARN DE ESS yMERISTEMOS DE PLANTULAS DE CAPS/CUM CH/NE~SE ............ 74
4.3.2 AMPLIFICACIÓN DEL TRANSCRITO DEL GEN TIPO WUS DE CAPS/CLW CH/WEWSE CON CEBADORES
HETERÓLOGOS
4.3.3 RESULTADO DE LA SECUENCIACIÓN
4.3.4 ANÁLISIS DE LA EXPRESIÓN GÉNICA DEL GEN TIPO WUS EN ESS DE CHILE HABANERO„...„ „„„ 78
4.4 DISCUSIÓN
4.5 CONCLUSIONES
BIBLIOGRAFÍA
CONCLUSIONES GENERALES
PERSPECTIVAS

LISTADO DE FIGURAS
Figura 1.1 Principios básicos del cultivo de tejidos vegetales (modificado de George ef
a/" 2008)
Figura 1.2 Proceso de embriogénesis somática indirecta (modificado de Pérez ef a/.,
Figura 1.3 Diferentes estadios embrionarios durante el desarrollo de la embriogénesis en
dicotiledóneas
Figura 1.4 Reguladores de crecimiento vegetal (RCVs) utilizados con mayor frecuencia
para la inducción de la embriogénesis somática
Figure 2.1 Efflciency (SEs/ Liquid Medium) of SE in the different treatments evaluated.. .38
Figure 2.2 SE from habanero pepper
Figure 2.3 Histological analysis of SE in Habanero pepper
Figure 2.4 Scanning electron micrographs of SE induction from 0 to 12 d of culture. .„„..46
Figure 2,5 Scanning electron microscopy (SEM) a-d and microphotography e-h of the
explant between 25 and 40 days.
Figura 3.1 Caracterízación morfológica de ESs deformados de chile habanero ............ 57
Figura 3.2 Hjstología del embrión cigótico (EC) de chile habanero en parafina ...„ ....... 58
Figura 3.3 Cortes histológicos longitudinales en parafina de un embrión somático maduro
de chile habanero
Figura 3.4 Cortes hjstológicos longitudinales en parafina de un embrión somático maduro
de chile habanero)
Figura 3.5 Cortes histológicos longitudinales en parafina de un embrión somático maduro
de chile habanero 60

Figura 3.6 Cortes histológicos en resina longitudinales de embriones somáticos
fusionados de chile habanero
Figura 3.7 Cortes histológicos en resina longitudinales de embriones somáticos
fusionados de chile habanero
Figura 4.1 ARN de ESs y meristemos de plántulas de caps/.cum ch/.nense ........... 74
Figura 4.2 Amplificación usando los cebadores diseñados para el gen tipo WUS .... „ ..... 75
Figura 4.3 Alineamiento de la secuencia de aminoácidos obtenidos
Figura 4.4 Análisis de expresión del gen tipo WUS en embriones somáticos de chile
habanero en diferentes estadios de desarrollo
Figura 4.5 Representación gráfica de la expresión del gen tipo WUS durante los
diferentes estadios de desarrollo de los ESs 79

LISTADO DE CUADROS
Table 2.1 Treatments evaluated during the inductíon and development of Habanero
pepper SE
Table 2.2 SEs, per stage of development, in the different treatments evaluated ............ 39
Cuadro 4.1 Mezcla de reacción de DNasa 1
Cuadro 4.2 Cebadores diseñados para el análisis de transcritos de un gen ti.po VVUS en
chile habanero
Cuadro 4.3 Cuantificación de ARN de ESs y meristemos de plantulas de Capsí.cum
chinense
Cuadro 4.4 Secuencia de nucleótidos obtenida de la secuenciación, Fragmento de 74 b.
Cuadro 4.5 Análisis tipo BLAST (Basic Local Alignment Search Tool) de la secuencia
deducida de aminoácidos de 74 b 77


ABREVIATUFUS
Acido 2,4-diclorofenoxiacético
Ácido abscísico
ADN
ADNc
ARN
COT
EC
ES
ESD
Acido desoxirribonucleico
Ácido desox/.rr/.bonuc/e/.co complementario
Ácido giberélico
Ácido indolacético
Acido ribonucleico
Cotiledón
Embríón cigótico
Embriogénesis somática
Embriogénesis somática directa
Embriogénesis somática indirecta
Embrión somátíco
Formaldehído
Hipocotilo
Murashige y Skoog
Reacción en cadena de la polimerasa
Regulador de crecimiento vegetal

SAM Meristemo caulinar
SEM Scanning electron microscopy

RESUMEN
En el presente trabajo se obtuvieron embriones somáticos a de partir hipocotilos de
plantas de Caps/.cL/m chí.nense. Se evaluaron diferentes concentracíones de ácido 2,4-
diclorofenoxiacético (0, 4.50 y 9.05 uM), diferentes tiempos de exposición de los explantes
a esta auxina (15, 30, 45 y 60 días), así como el desarrollo de los embriones en medio de
cultivo semisólido o líquido. Como resultado, se estableció un nuevo sistema de
regeneración a través de embriogénesis somática directa en medio líquido, con una
eficiencia de 1.77xl04 embriones somáticos por litro de medio de cultivo y un 58.7 ± 5.3%
de embriones en estadio cotiledonar. Mediante análisis hlstológicos y de microscopia
electrónica de barrido se identificaron las etapas críticas de la embriogénesis, incluyendo
la adquisición morfogenética de la competencia celular, Ia formación del suspensor y el
desarrollo de los embriones somáticos. Debido a que los embriones somáticos de chile
habanero presentaron deformaciones exclusivamente en el meristemo caulinar y que el
gen homeótico Wusche/ tiene una función importante en la regulación celular durante la
formación del meristemo embrionario, se investigó la expresión de este gen. Los
transcritos de Wusche/ fueron detectados durante las etapas tempranas del desarrollo de
los embriones somáticos, hasta antes de alcanzar el estadio cotiledonar. Estos resultados
ofrecen una perspectiva nueva y prometedora en la regeneración /.n v/.Íro de esta especie
(C. ch/.nense), como una especie con alto potencial embriogénico /.n v/.Íro, contrariamente
a lo que se había repohado para el género Caps/.cum hasta la fecha.


ABSTRACT
ln the present work somatic embryos were produced from the hypocotyls of Caps/.cum
ch/.nense plants. Different concentrations of 2,4-dichlorophenoxyacetic acid (0, 4.50 y 9.05
LiM), several exposure times of the explant to this auxin (15; 30; 45; 60 days) and the
development of somatic embryos cultured in semisolid or liquid medium were evaluated.
As a result, a novel system cif regeneraÑion via direct somatic embryogenesis in liquid
med/.um was established, with an efflciency of 1.77X104 ESs.L-í, and 58.7 ± 5.3% of
embryos reaching the cotyledonary stage. Critical stages of embryogenesis, including
cellular acquisition of morphogenetic competence, formation of suspensor, and
development and maturation of somatic embryos during somatic embryogenesis were
identified by histological analysis and scanning electron microscopy. Because the somatic
embryos of habanero pepper developed abnormalities exclusively in the shoot meristem
and Wusche/ homeobox gene plays an important role in cellular regulation during
formation of the embryonic meristem, we investigated the expression of this gene.
Wuschel mRNA was detected during early stages of development of somatic embryos
before reaching the cotyledonary stage. Our results offered a promising new outlook on
the Í.n v/.fro regeneration of this species (C. ch/.nense). Contrary to what has been reported
to date for the Caps/'cum genus, it is a plant species with high embryogenic potential /.n
vitro.


lNTRODUCCIÓN
lNTRODUCCI0N
El chile habanero (C, ch/.nense Jacq.) es uno de los productos hortícolas más importantes
en el mercado, tanto nacional como internacional. A pesar de ello, no ha sido de los
cultivos más beneficiados por los avances de la ciencia y la tecnología.
El mejoramiento genético convencional es el método más utilizado para generar cultivares
de mayor calidad y mayor potencial productivo en las especies cultivadas. Su limitación
está dada en lo fundamental por el tiempo prolongado requerido para obtener una
variedad mejorada, aunado a la cantidad de mano de obra y de espacio que demanda la
actividad .
La biotecnología abre una amplia gama de altemativas para el mejoramiento genético de
las plantas. La obtención de líneas tolerantes a los factores adversos, o portadores de
atributos superiores relacionados con la calidad y los rendimientos, han sido algunas de
las aplicaciones en las que han sido ocupadas las herramientas biotecnológicas.
La regeneración es el requisito común para cualquiera de los usos que puedan tener las
técnicas del cultivo /n vÍ.fro. El método más eficiente de regeneración de plantas i.n v/.fro es
la embriogénesis somática; sin embargo, existen numerosos reportes que argumentan la
recalcitrancia del género Caps/.cum a la morfogénesis i.n vt.Íno (Ochoa y Ramírez, 2001;
Kothari, 2010) Este fenómeno en el género, se manifiesta a través de: 1) baja eficiencia
de los sistemas de regeneración (Buyukalaca y Mavituna,1996), 2) baja reproducibilidad
de los protocolos de regeneración, 3) alto Índice de embriones somáticos deformados
(Steinitz eí a/., 2003), y 4) baja tasa de germinación y/o conversión de los embriones
somáticos en plantas (Steinitz ef a/., 2003, López e£ a/„ 2006). Tomando en cuenta esta
problemática de la especie y que la auxina sintética 2,4-D ha sido reportada como un
factor determinante en el proceso de ES (Parroft eí a/., 1988), nos propusimos realizar
estudios morfológicos y moleculares durante la histogénesis de los embriones somáticos
de chile habanero en presencia de ácido 2,4-diclorofenoxiacético (2,4-D), con la finalidad
de caracterízar y determinar la posible implicación de diversos factores como lo es la
auxina 2,4-D, diversas condiciones de cultivo y un homólogo del gen M/usche/, en la
recalcitrancia del género a la morfogénesis Í.r) vÍ.Ím.


CAPÍTULO 1
CAPITUL0 I
ANTECEDENTES
1. GÉNERO Caps/cum
1.1 lMPORTANCIA DEL GÉNER0 Capsi.cum
El género Capsí.cum es miembro de la familia de las Solanáceaes, que incluyen el
jitomate, la papa, el tabaco y la petunia El género Capsicum consta de aproximadamente
22 especies silvestres y cinco especies domesticadas: C. annuum, C. baccafum, C.
chí.nense, C. Írtjíescens y C. pubescens (Bosland,1994). México es uno de los principales
centros de origen y domesticación del género Capsícum, en particular de la especie
annuum (Laborde y Pozo,1984). En México se encuentran bajo cultivo C. annut/m en
casi todo el país; C. frufescens en el centro y sureste,. C. ch/'nense en la Península de
Yucatán y C. pubescens en las partes altas de los estados que ocupan la región central
de México.
Los principales mercados nacionales donde se comercializa el chile habanero son las
centrales de abastos del Distrito Federal, Guadalajara, Monterrey, San Luis Potosí,
Morelia, Cancún, Puebla, Mérida y Oxkutzcab. En el mercado internacional se exporta a
los Estados Unidos (California, Florida, Texas, Nueva York, Oregon, Nuevo México y
Massachusetts), Canadá y Cuba. A los mercados europeos, España (Madrid, Barcelona,
Zaragoza y Valencia) e lnglaterra (Londres). En Asia, el principal mercado es Japón, país
que ha iniciado una estrategia sumamente interesante y atractiva, para establecer
negocios relacionados con la industria del chile habanero (Ledón, 2006).
En Yucatán, el cultivo de chile habanero (C. ch/.nense Jacq.) es uno de los más
importantes debído a su demanda tanto local, como nacional e Ínternacional. En la
actualidad, cuenta con 500 hectáreas sembradas de chile habanero, principalmente en 13
municipios, con una producción de cuatro mil toneladas y una derrama de 40 millones de
pesos anuales. Del total de la producción de chile habanero, el 40 por ciento es para
consumo local, el 30 por ciento para el mercado nacional y el 30 por ciento restante se
industrializa para §u exportación a diversos países (SFAYP, 2009).

CAPÍTULO 1
En los últimos años se ha visto un fuerte incremento en la superficie sembrada en campo
y protegida (invernadero). Sin embargo, la producción de chile habanero es afectada en
parte por las situaciones adversas a las que se enfrentan los agricultores (huracanes, falta
de asesoría técnica, disponibilidad de buena semilla, etc.). pero el principal factor que
limita la producción, son las enfermedades de origen viral, inducidas principalmente por
geminivirus transmitidos por la mosca blanca (Bem/.s/.a íabac/. Genn), así como diferentes
virus transmitidos por áfidos que pueden causar pérdidas hasta de un 100% (Latournerie
ef a/., 2006).
1.2 CHILE HABANERO (Caps/.cum chjnense Jacq.)
1.2.1 DESCRIPCIÓN TAXONÓMICA
El cmle habanero pertenece al género Caps/.cum cuyo significado se deriva del griego:
Kapso (picar) y Kapsakes (cápsula) (Nuez ef a/., 2003). Según Tun (2001), se clasifica de
la siguiente manera:
Clase
Subclase
Super orden
Orden
Familia
Género
Especie
Angiosperma
Dicotiledónea
Sympetala
Tubiflorales
Solanaceae
Capsicum
C. chinense Jacq.
1.3 CULTIV0 DE TEJIDOS VEGETALES
El cultivo /.n v/`£ro es una de las herramientas clave de la biotecnología vegetal que
aprovecha la naturaleza totipotente de células vegetales, un concepto propuesto por
Haberlandt (1902) y claramente demostrado por primera vez por Steward eí a/. (1958).

CAPÍTULO 1
Se le llama Cultivo de Tejidos Vegetales (CTV) al conjunto de técnicas que permiten el
establecimiento, mantenimiento y desarrollo de cualquier parte de una planta, desde una
célula hasta un organismo completo, bajo condiciones artificiales, axénicas y controladas
(Pérez ef a/„ 1999). EI CTV comprende, un heterogéneo grupo de técnicas mediante las
cuales un explante se cultiva asépticamente en un medio de composición química definida
y se incuba en condiciones ambientales controladas. Generalmente es común dividir las
técnicas del cultivo de tejidos en dos grandes grupos: a) cultivos en medios semisólidos y
b) cultivos en medios líquidos, los que a su vez pueden ser agitados (mediante el empleo
de agitadores de uso continuo) o estacionarios. También es frecuente dividir al cultivo de
tejidos atendiendo a los niveles de complejidad en cultivo de órganos, cultivos celulares y
cultivo de protoplastos (Roca y Mroginski, 1991 ).
EI CTV comprende una gama de técnicas, métodos y estrategias /.n vÍ`fro que son parte del
grupo de tecnologías llamadas biotecnología de plantas. Además ha sido utilizado para
crear variabilidad genética, mejorar la sanidad y para incrementar el germoplasma
disponible. Actualmente los protocolos de CTV están disponibles para la mayoría de las
especies, aunque estos continúan optimizándose (Brown y Thorpe,1995).
El cultivo /.n vt.íro facilita la multiplicación rápida de clones superiores y es un requisito
previo para la mejora de plantas a través de técnicas de ingeniería genética. El cultivo de
tejidos se ha explotado para crear variabilidad genética a través de la producción de
haploides, variantes somaclonales y gametoclonales. También se utiliza para mejorar el
estado de salud del material vegetal y para aumentar el número de germoplasma
di§ponible para el fitomejorador (Kothari, 2010).
1.3.1 REGENERACIÓN DE PLANTAS /.n v7.fro
El objetivo de la propagación de plantas, a través de cultivo de tejidos, es propagar
plantas como clones. Las plantas obtenidas por cultivo de tejidos se denominan
microplantas y pueden ser obtenidas de tres maneras: 1) de brotes preexistentes o
brotes primordiales (meristemos) que se estimulan para que crezcan y proliferen; 2)
cuando nuevos brotes son inducidos a formarse en los tejidos desorganizados o
directamente sobre el tejido del explante de la planta madre; 3) a través de la formación
de embriones somáticos que se asemejan a los embriones cigóticos y que pueden

CAPÍTULO I
desarrollarse a plántulas de la mísma manera. Este proceso se denomina embriogénesis
somática. Para obtener plantas por los dos primeros métodos, es necesario tratar brotes
de un tamaño adecuado como esquejes e inducir la producción de raíces. La obtención de
nuevas plantas a partir de células las cuales normalmente no habrían formado parte en el
proceso de regeneración, es una muestra de la totipotencialidad celular.
La totípotencia es una característica especial de células en tejidos jóvenes y
meristemáticos. Esta puede ser demostrada tamblén por algunas células diferenciadas,
por ejemplo, las células del cambium y células de empalízada de la hoja, pero no en
aquellas células terminalmente diferenciadas (ejemplo traqueidas) (George ef a/., 2008)
Teóricamente, todas las células vegetales, órganos o plantas, pueden ser clonados, es
decir, producir un gran número de individuos donde todos tengan la misma constitución
genética que la del padre. Actualmente las técnicas de cultivo no permiten esto en cada
caso y a veces se producen irregularidades que resultan en variantes somaclonales
(Larkin y Scowcroft,1981).
En resumen, las vías de regeneración son:
• Multiplicación de brotes de yemas terminales, axilares o laterales. El punto de inicio en
este caso puede estar en los merjstemos, las puntas de los brotes, las yemas, los
nudos o los brotes de las yemas en raíces.
• Organogénesis directa. En este caso, la formación del brote adventicio o de la raíz
ocurre en el explante, o en alguna parte escíndida de la planta.
• Organogénesis indirecta. La formación del brote adventicjo o de la raíz ocurre en el
callo el cual se deriva inicialmente de un órgano, tejido u otra parte de la planta.
• Embriogénesis somática directa. Los embriones se forman directamente en el explante
primario
• Embriogénesis somática indirecta. Los embriones se forman a partir de un callo
embríogénico.

CAPÍTULO 1
En el caso de multiplicación de brotes de yemas terminales, axilares o laterales se parte
de meristemos ya formados para que a partir de ellos se obtengan una o varias plantas
completas, mientras que en la embriogénesis somática y la organogénesis se parte de
tejidos no meristemáticos y se requiere un proceso de desdiferenciación y rediferenciación
de los tejidos, para dar lugar a embriones somátícos (ESs) o brotes adventicios (Pérez e!
a/.,1999).Durante la multiplicación de yemas se ha observado que las nuevas plantas se
originan a partir de estructuras multicelulares (Pérez eí a/.,1999), mientras que durante la
embriogénesis somática se ha demostrado que los embriones somáticos pueden ser tanto
de orjgen unicelular (Quiroz ef a/., 2002) como multicelular (Lee ef a/.,1997).
El esquema básico que resume estos principios se presenta en la Figura 1.1
MCTrstcmo
Nudo9
UO=F06=`,ES SC ?=CTA
rJ•:\Exp'an(evaíloste|i
t` 1 C =` F C C = ` J E S S
`,,=`=.=CTA
•Í4,`\
Planta stock
Cultivo de meristemos
>"
Oiganogénegisdlrec(a
Embrlogér`esigdlre¢taJ®i=rganogénesLsinclir®cta
Embriones somaticogE=ca"\®
#- J_tEmbnogenesisindirecta
ég -''' F,,antu,aEmbnoneg 9omet]cog
Cultlvo en guspenslórl
Figura 1.1 Principios básicos del cultivo de tejidos vegetales (modificado de
George ef a/., 2008).

CAPÍTULO 1
1.4 EMBRloGENESIS SOMATICA
La embriogénesis somática (ES) es un proceso mediante el cual las células somáticas
atraviesan por una etapa de diferenciación para formar embriones somáticos, los cuales
son morfológicamente similares a los embriones cigóticos. Este proceso morfogénico se
produce de manera limitada en condiciones naturales, dentro de óvulos (ejemplo,
Paeonia) y más raramente en hojas (ejemplo, Asp/ení.um y Ka/anchoe). A partir de la
primera observación de formación de embriones somáticos en suspensiones celulares de
Daucus carofa realizada por Steward ef a/. (1958) y Reinert (1958) se ha demostrado el
potencial de la ES. Durante los últimos 40 años, la ES ha sido descrita en un gran número
de especies de plantas. Nuevas especies y protocolos de regeneración por esta vía son
continuamente reportados y su número aumenta continuamente. Los protocolos para la
obtención de este proceso también son constantemente modificados y mejorados. La ES
probablemente pueda lograrse para todas las especies de plantas, siempre que se
empleen el explante, medios de cultivo y condiciones ambientales adecuadas (George eí
a/„ 2008).
La ES es un sistema ideal para investigar los procesos completos de diferenciación de
plantas, así como los mecanismos de expresión de totipotencia en células vegetales.,
Tiene muchas ventajas para su estudio comparada, con la embriogénesis cigótica; por
ejemplo, a) el proceso de embriogénesis es fácilmente monitoreado, b) el ambiente del
embrión puede ser controlado, c) un gran número de embriones puede ser fácilmente
obtenido. Este proceso puede ocurrir por dos vías: directa (ESD) e indirecta (Esl) (Figura
1.1 ). En la ESD, los embriones se originan directamente de las células del explante. En la
Esl,se forma un callo en una primera etapa del cultivo. Si este callo es transferido a un
medio de cultivo líquido, donde es desagregado, se forma una suspensión celular a partir
de la cual son inducidas las células a formar los embriones somáticos. Este es un proceso
mucho más complejo que la ESD. La ESD se da cuando las células del explante original
están predeterminadas a formar embriones somáticos, por lo que sólo se requiere
proporcionar al tejido las condiciones adecuadas para la inducción y diferenciación de los
embriones somáticos. En la Esl, el explante original no tiene células proembriogénicas,
por lo que se requiere primero suministrarle al tejido las condiciones para que ocurra la
inducción y luego cambiarlo a otras que sean propicias para la diferenciación de los

CAPÍTULO 1
embriones. Las células de tejidos muy jóvenes, como por ejemplo los embriones cigóticos
inmaduros, son ya en una buena proporción células proembriogénicas, o si no, son
fácilmente inducibles para que lo sean. Por el contrario las células más diferenciadas o de
tejidos adultos muy difícilmente o casi nunca se llegan a convertir en proembriogénicas.
Esto explica por qué la embriogénesis somática resulta sencilla en algunos tejidos y
prácticamente imposible en otros (Pérez ef a/.,1999).
Aparentemente, los factores químicos más importantes para la embriogénesis somática
son las auxinas exógenas, la fuente y la concentración del nitrógeno y algunas otras
sustancias como la sacarosa. Desde el punto de vista de la propagación, la
embriogénesis somática es el sistema más eficiente, si se considera la eficiencia como el
número de plantas regeneradas por unidad de tiempo. Empleando este sistema se
pueden obtener cantidades virtualmente ilimitadas de plantas, ya que todo hace suponer
que por cada célula suspendida en el medio de cultivo, se está diferenciando una planta
(Roca y Mroginski,1991).
El proceso de embriogénesis somática indirecta consta de varias etapas (Figural .2), en
cada una de las cuales se requieren diferentes estímulos y ocurren fenómenos diferentes
(Merkle ef a/„ 1995). Estas etapas se describen a continuación:

CAPÍTULO 1
E-TEJIDO CALLOSO
MASA DE CELULASPROEMBRloNICAS
EMBRIONES ENESTADIO GLOBULAR
EMBRIONES ENESTADIO COFtAZÓN
EMBRIONES ENESTADIO TOFtpEDO
EMBRIONES EN ESTAD0COTILEDONARIO
•-`._.`.. PLÁNTULAS
INDUCCION
+ AUXINAS
HISTODIFERENCIACIÓN
-AUXINAS
MADURACIÓN
ABA
DESECACIÓN
GERMINACIÓN
Figura 1.2 Proceso de embriogénesis somática indirecta (modificado de Pérez
ef a/., 1999).
Inducción
Las células somáticas de la planta contienen toda la información genética necesaria para
crear una planta completa y funcional. La inducción de la ES, debe constar de la
terminación del modelo actual de expresión genética en el tejido del explante y ser
sustituido por un programa de expresión de genes relacionados con el proceso
embriogénico (George ef a/., 2008).
Este proceso es la conversión de una célula somática a una célula proembriogénica. Se
considera que los factores determinantes para que suceda el proceso de inducción son:
1. Genotipo
2. Grado de diferenciación de las células del explante.
10

CAPÍTULO 1
3. Auxinas. La presencia de auxinas es un factor determinante para que ocurra el
fenómeno de inducción. Se sabe que la presencia de este tipo de reguladores del
crecimiento estimula entre otras cosas, la metilación del ADN, lo cual puede favorecer
el cambio del patrón de expresión genética que se requiere para que ocurra la
inducción.
4. Aislamiento celular. Se sabe que el aislamiento de una célula o de un grupo de células
con respecto al resto del tejido estimula también la inducción o conversión de una
célula somática en pro embriogénica. Este aislamiento sucede cuando hay necrosis en
el tejido adyacente o bien cuando se aplican tratamientos físicos para separar las
células. La presencia de auxinas puede contribuir también a este aislamiento celular, al
promover la friabilidad del tejido (George,1993).
Se necesita la presencia de una auxina para la inducción de embriones o un callo
embriogénico. Usualmente se emplea el 2,4-D, el cual aparentemente brinda el estímulo o
inicia la inducción de la ES en el callo o explante. Sin embargo, la maduración de los
embriones y la germinación no ocurren en presencia de auxinas exógenas; en
consecuencia, hay que remover la auxina, o usarla en concentraciones más bajas. Tanto
la inducción de la embriogénesis somática, como el desarrollo de los estados
subsiguientes de los embriones dependen de la presencia de nitrógeno reducido
(Halperin, 1995). La concentración y la relación entre el nitrógeno oxidado y reducido
pueden influir en la maduración de embriones somáticos (George,1993).
Histodiferenciación
La división celular correcta desempeña un papel crucial durante la embriogénesis, desde
la división de una célula polarizada, el citoplasma y cualquier molécula reguladora
contenida en ellas. Como resultado de las divisiones asimétricas las células hijas pueden
heredar diferentes determinantes citoplasmáticas y de esta manera adquirir diferentes
destinos. (George ef a/., 2008)
Durante la etapa de histodiferenciación, las masas de células proembriogénicas se
diferencian formando embriones somáticos, mediante una división y diferenciación celular
simultáneas. Para que estas células cesen su multiplicación y pasen a una fase de
11

CAPÍTULO 1
diferenciación, se requiere la eliminación de las auxinas exógenas. Uno de los fenómenos
inlciales en esta etapa es el establecimiento de una polaridad en las masas de células
proembriogénicas. Es{a polaridad se mantendrá durante todo el desarrollo del embrión.
Durante la etapa de histodiferenciación, los embriones somáticos pasan por una serie de
estadíos intermedios muy similares a los que ocurren en la embriogénesis cigótica (Pérez
ef a/.,1999). Se ha observado que para el establecimiento de la polaridad y el crecimiento
bipolar en la planta se requieren níveles bajos de auxina. Además, el transporte polar de
la auxi.na es necesario para establecer la simetría bilateral en los embriones
dicotiledonarios (Michalzuk eí a/.,1992).
Un embrión somático experimenta los mismos cambios de estadios que un embrión
cigótico. Las plantas dicotiledóneas atraviesan por las siguientes etapas: globular,
acorazonada, torpedo y cotiledonar (Figura 1.3). Los embriones de las plantas
monocotiledóneas atraviesan por la etapas: globular, coleoptilar y escutelar (Thorpe,
1995).
Preglobular Clobular Corazón Torpedo Plántula
Figura 1.3 Diferentes estadios embrionarios durante el desarrollo de la
embriogénesis en dicotiledóneas.
Maduración
Luego de la división celular y la histodiferenciación, ocurre la maduración, período de
desarrollo del embrión en el cual ocurre la expansión celular y la deposición de las
reservas (Bewley y Black,1985). En los protocolos de inducción de la maduración se han
evaluado diferentes componentes del medio que desempeñan un papel importante en
este proceso, entre ellos fuentes nitrogenadas, carbonadas y reguladores de crecimiento
(Merkle eí a/., 1995).
12

CAPÍTULO 1
Durante la etapa de maduración los embriones somátjcos se someten a cambios
morfológicos y bioquimicos. Cambios en órganos de almacenamiento, cotiledones,
expansión asociada con la deposición de materiales de reserva, represión de la
germinación y adquisición de la tolerancia a la desecación son algunos ejemplos de estos
cambios (Thomas,1993). Se sabe que los embriones somáticos acumulan productos de
almacenamiento que presentan las mismas características que los que se acumulan en
los embriones cigóticos.
Los productos de almacenamiento también están dirigidos a los compartimentos
subcelulares correctos (Merkle ef a/..1995). Sin embargo, la cantidad de un producto de
almacenamiento en particular, así como la coordinación de su acumulación puede diferir
entre embriones somáticos y cigóticos (Merkle eí a/.,1995; Yeung,1995). La síntesis y el
depósito de productos de almacenamiento y proteínas LEA (late embryogenesis
abundant) durante la embriogénesis somática y cigótica generalmente están reguladas a
través de ABA y la expresión de genes inductores de estrés (Dodeman ef a/.,1997).
En esta etapa del proceso el ácido abscísico (ABA), reprime la ES y reduce la frecuencia
de anormalidades de desarrollo como son la formación secundaria de embriones a partir
de embriones somáticos y la germinación precoz (Ammirato,1974). Las concentraciones
de ABA más usadas fluctúan entre 0.1 y 1.0 uM. También, las antiauxinas han sido
efectivas de la misma forma que el ABA (Fujimura y Komamine,1979).
Los embriones somáticos no acumulan la misma cantidad de proteínas /.n v/.vo que las
semillas. Se ha detectado que los embriones somáticos de alfalfa contíenen el 10% del
total de las proteínas que se acumulan en los embriones cigóticos. Los embriones
somáticos son menos vigorosos que las semillas in vivo. En consecuencia, se sugiere que
si se incrementa la acumulación de proteínas, podría ser posible incrementar el vigor de
los embriones somáticos (Stuart eí a/.,1988).
EI ABA tiene al menos dos funciones principales el proceso de maduración de los ESs. La
primera es prevenir la germinación precoz de embriones somáticos cuando se utiliza a
concentraciones de aproximadamente 1 uM (Anandarajah y MCKersie,1990). La segunda,
incrementar la tolerancia a la desecación en el estado de desarrollo cotiledonar a
concentraciones de 10-50 uM (Senaratna ef a/.,1990).
13

CAPÍTULO 1
La función del ABA en la iniciación de la acumulación de reservas no es desconocida.
Cuando se inicia la acumulación de reservas el contenido de ABA endógeno es alto. La
aplicación exógena de ABA a ES en un estado adecuado, puede ayudar a recuperar un
mayor número de embriones somáticos normales (Merkle,1990).
Germinación
Los embriones somáticos son originados vía asexual y durante su desarrollo pasan a
través de las mismas secuencias de maduración que su contraparte cigótica. Sin
embargo, en la germinación de ESs, no ocurre la fase de deshidratación y del estado
quiescente del embrión, presentando un desarrollo continuo como ocurre en semillas
vivíparas y recalcitrantes (Rochon ef a/.,1998; Farnsworth, 2000).
El proceso de germinación comienza con la absorción de agua por la semilla (imbibición)
y finaliza cuando la radícula, se extiende penetrando las estructuras que lo rodea
(Bewley,1997). Para que la germinación ocurra correctamente, los embriones somáticos
deben de tener brotes funcionales y raíces capaces de tener crecimiento meristemático
(Merkle, 1990). En condiciones no apropiadas de cultivo, la germinación puede ocurrir
prematuramente teniendo como consecuencia plántulas débiles o con poca viabilidad
(Merkle,1990).
Para algunas especies, la eficiente conversión a plántula requiere la imposición de una
desecación temporal previa a la germinación. Este procedimiento imita la maduración de
semillas in vivo y puede ser necesario para iniciar los procesos metabólicos necesarios
para la germinación y crecimiento de plántulas /.n v/.íro (Rosenberg y Rinne,1988).
La germinación es el proceso de elongación y reac{ivación metabólica de un embrión
somático maduro para convertirse en una plántula. Para que esto suceda se requieren
estímulos físicos como la luz y químicos como el ácido giberélico o citocininas.
Obviamente, los embriones somáticos carecen de tejidos de reserva por lo que su
germinación sólo ocurre /.n v/.fro, donde el medio de cultivo aporta los nutrientes, o bien,
cuando se les proporcionan depósitos artificiales de nutrientes (semillas artificiales).
Durante la etapa de germinacion, uno de los reguladores de crecimiento vegetal más
utilizado es el ácido giberélico (GA3), el cual, es incorporado al medio con el fin de ayudar
14

CAPÍTULO 1
a la germinación de los embriones somáticos (Kochba ef a/.,1974; Lakshmi ef a/.,1979).
1.4.1 FACTORES INVOLUCRADOS EN LA EMBRloGÉNESIS SOMÁTICA
Los principales factores que afectan a la ES /.n v/.Íro son: la variedad vegetal, el origen del
explante y la composición del medio de cultivo, particularmente con respecto a las
hormonas (Zimmerman eí a/.,1989). De igual manera, se sabe que la fuente nitrogenada
modifica sustancialmente la respiiesta embriogénica de los cultivos (Halperin y Wetherell,
1965).
La adquisición de totipotencia es el paso más crítico en la embriogénesis somática y la
aplicación de hormonas exógena es el factor más impohante que la afecta, debido a que
el uso de auxinas a altas concentraciones es necesario para causar la desdiferenciación y
la estimulación de la totipotencia. Varias auxinas han sido usadas para este propósito: la
auxina natural ácido indolacetico ([AA)y otras sintéticas como el ácido naftalenacético
(ANA), el 2,4-D, picloram, ácido pÍcolínico y kinetina (Finstad eí a/.,1993; Desamero eí a/.,
1994; Litz eí a/.,1998; Xing eí a/., 2000; Cooke ef a/., 2002).
De manera general, el 2,4-D es la auxina más eficiente y la más comúnmente usada para
la promoción de la ES ya que: 1) estimula una rápida división celular; 2) estimula una
dMsión celular sincronizada que da como resultado células proembriogénicas y 3)
estimula la proliferación de células proembriogénicas (Litz ef a/.,1998).
1.5 REGULADORES DE CRECIMIENTO VEGETAL (RCVs) EN LA INDUCCIÓN
DE LA EMBRIOGÉNESIS SOMÁTICA
La composición y la concentración de RCVs en el medio de cultivo determinan tanto la
habilidad del explante para responder, así como también, el tipo de respuesta
morfogénica que se obtiene. Dependiendo de la composición de RCVs en el medio se
obtendrá ES, organogénesis o desarrollo de brotes axílares. Esta característica ha sido
observada y reportada para cultivos de nuez (Fernandez ef a/., 2000), así como para maní
(Victor ef a/.,1999).
Entre diferentes estímulos externos que inducen la embriogénesis somática, los RCVs
como auxinas y citocininas han si.do los más frecuentemente utilizados para la inducción,
15

CAPÍTUL01
ya que regulan el ciclo celular y promueven divisiones celulares (Francis y Sorrell, 2001 ).
Existen algunas especies en las cuales durante su regeneración se ha observado que no
es del 7°/o de los protocolos reportados para la inducción de la ES requirieron de RCVs
para su inducción. Por otra parte, en más de 80 % de los protocolos reporiados para la ES
la inducción se da con la sola presencia de auxinas o en combinación con citocininas. La
auxina más comúnmente usada para la inducción es el 2,4-D (Figura 1.4; Gaj, 2004).
En algimos sistemas se ha reportado que las citocininas en sustitución de las auxinas
pueden ser efectivas para la inducción de la ES (Asano ef a/., 1996; Castillo y Smith,
1997; Carimi ef a/.,1999; Chen y Chang, 2001; Nanda y Rout, 2003). Aunque también
existen reportes en donde la aplicación de citocininas impide el desarrollo de los
embriones somáticos (Choi ef a/.,1998)
2,4-D 2A-D Solo otras Aux Aiot + Cii Cn Libre de RCV
Figura 1.4 Reguladores de crecimiento vegetal (RCVs) utilizados con mayor
frecuencia para la inducción de la embriogénesis somática (modificado de Gaj,
2004). Aux-auxinas; Cit. Citocininas.
Se han reportado moléculas biológicamente activas para la inducción de la ES en
diferentes especies de plantas. Un ejemplo de ellas son las poliaminas, las cuales se han
adicionado al medio de cultivo para mejorar la embriogénesis en algunas plantas (Yadav
y Rajam,1998; Kevers ef a/., 2000; Kelly eí a/., 2002).
16

CAPÍTUL0 1
1.6 GENES INVOLUCRADOS EN LA EMBRIOGENESIS SOMATICA
Existen factores tanto genéticos como fisiológicos que activan la ES /n v/.Ím en diversas
células somáticas vegetales (Karami ef a/., 2009). Las investigaciones sobre los
mecanismos de la ES se han centrado históricamente en la regulación hormonal del
proceso y un gran número de estrategias han sido desarrolladas para regenerar muchas
especies a través de la ES. Sin embargo, las vías de señalización y los genes que se
requieren para inducir y desarrollar embriones somáticos no están bien definidos (Rose y
Nolan, 2006). Actualmente esta situación está cambiando, debido a los avances en la
comprensíón de la embriogénesis cigotica y la diferenciación de los meristemos apicales.
lnicialmente en células embriogénicas de zanahoria fue identificado el gen receptor cinasa
de la embriogénesis somátíca (SERK/ como un marcador de la ES (Schmidt eí a/.,1997).
Lotan e{ a/. (1998) demostraron que era posible inducir ESs después de la expresión
ectópica del gen iEAFY C07-yLEDOM (LEC7), en Arabidopsis. Un aspecto importante
de este hallazgo fue la inducción de la ES en ausencia de reguladores de crecimiento
principalmente auxina. LEC7 es un factor de transcripción necesario para muchos
aspectos de la embriogénesis de Affib/.dops/.s (Lee ef a/., 2003). También se ha es{udiado
el factor de transcripción Wusche/ íWUS) cuya expresión ectópica al igual que LEC7
también induce embriones somáticos en Arabí.dops/.s (Zuo eí a/., 2002).
La expresión de genes durante la ES puede ser evaluada ya sea por aislamiento de
genes expresados en los embriones somáticos y, posteriormente, la identificación de la
función de estos genes o mediante el estudio de la expresión de una variedad de otros
genes aislados a partir de tejidos no embriogénicos, puesto que estos podrían jugar algún
papel en la embriogénesis. Un considerable grupo de genes han sido identificados
durante su expresión en los ESs, y varios de ellos están siendo utilizados para analizar
mecanismos de regulación génica durante la ES (Zimmerman, 1993)
Durante las últimas dos décadas, se han realizado esfuerzos considerables para
identificar los genes con patrones de expresión alterada durante la ES. Varios sistemas
han sido explotados para comprender los mecanismos de regulación génica durante el
SE, y el sistema de ES de zanahoria ha servido como modelo (Chungh y Khurana, 2002;
Feher ef a/., 2003; Zhu y Perry, 2005; lkeda eí a/., 2006; lkeda eí a/., 2006a), Los análisis
del proteoma y transcriptoma han conducido a la identificación y caracterización de ciertos
17

CAPÍTUL0 1
genes implicados en la ES (Raghavan, 1986; Wilson y Wilson, 1993; Giroux y Pauls,
1997; Chuck eí a/.,1998; Zdravkovic-Korac y Neskovic,1999;Thibaud-Nissen e/ a/., 2003;
Ben ef a/„ 2005; lmin e/ a/., 2005; Rensing eí a/., 2005; Che eí a/., 2006; Che ef a/.,
2006a). Sin embargo, la mayoría de estos genes son sobre regulados solamente en
etapas finales del desarrollo, lo que sugiere que no juegan un papel directo en la
transición del estado vegetativo a embriogénico (Karami, 2009).
1.7 REGENERACIÓN /-n v/.Íro DE CHILE (Caps/.cum spp.)
La regeneración en Caps/.cum ha sido difícil de trabajar en comparación con otros
miembros de las solanáceas como: tabaco, jitomate y papa, los cuales son utilizados
frecuentemente como sistemas modelo debido a su alta capacidad de regeneración
(Kothari, 2010). Caps/.cum es considerado como un género recalcitrante a la morfogénesis
i.n vt.íro. Aunado a ello, algunas especies del género Caps/.cum presentan problemas de
incompatibilidad interespecífica e infertilidad en el híbrido F1, durante el mejoramiento
convencional. Todo ello hace que se dificulten los trabajos de mejoramiento genético de
los cultivares que lo requieren.
Actualmente, existen reportes de regeneración de plantas /.n v/.Íro del género Caps/.cum,
particularmente en la especie C. annuum Sin embargo, la gran mayoría son vía
organogénesis directa a partir de hojas cotiledonares e hipoco{ilos (Gunay y Rao,1978;
Fári y Csakó,1981; Agrawal ef a/.,1983; Phillips y Hubstenberger,1985; Ochoa-Alejo e
lreta-Moreno, 1990; Hyde y Phillips, 1996). La ES en C. annuum, ha sido inducida
principalmente a partir de embrión cigótico inmaduro (Harini y Sita, 1993; Binzel ef a/.,
1996; Jo ef a/„ 1996), embrión cigótico maduro (Büyükalaca y Mavituna, 1996), hoja
(Kintzios eí a/., 2001). Khan ef a/. (2006) y Aboshama (2011 ) reportaron recientemente la
obtención de embriones somáticos utilizando yemas apicales y segmentos de tallo.
El chile habanero es una especie perteneciente a un género recalcitrante a la
morfogénesis, en el que la regeneración de plantas a partir de células, tejidos y órganos
no ha logrado grandes avances, lo que limita la aplicación de técnicas de avanzada, así
como la transformacjón genética, para el mejoramiento de la especie (Ochoa y Ramírez,
2001 ; Kothari, 2010). Sin embargo, actualmente se han reportado los primeros protocolos
de regeneración /`n v/.fro (Santana ef a/., 2005 y 2006; López eí a/., 2006; Zapata eí a/.,
18

CAPITULO 1
2007) para esta especíe. De igual manera, Santana-Buzzy ef a/. (2009) reportaron, por
prímera vez en esta especie, un estudio histológico de la ES en diferentes tipos de
explantes (hoja cotiledonar, embrión cigotico germjnado e hipocotilo) de C. ch/.nense.
La problemática común reportada en estos trabajos es el efecto del etileno sobre el
comportamiento de los explantes durante la morfogénesis y el desarrollo de los brotes de
chile habanero, así como el alto Índice de embriones deformados que se obtienen durante
el proceso de ES.
19

CAPÍTULO 1
1.8 JUSTIFICAcloN
El chile habanero es uno de los productos hortícolas más importantes en el mercado,
tanto nacional como internacional. La biotecnología abre una amplia gama de alternativas
para el mejoramiento genético de las plantas. La regenerac.ión es el requisito común para
cualquiera de los usos que puedan tener las técnicas del cultivo /.n v/.fro. Numerosos
reportes argumentan la recalcitrancia del género Caps/.cum a la moffogénesis /.n v/.fro
(Ochoa y Ram'H.ez, 2001; Kothari, 2010): que se manifiesta a través de: 1) la baja
eficienc.ia de los sistemas de regeneración (Buyucalaca y Mavituna, 1996), 2) la baja
reproducibilidad de los protocolos de regeneración, 3) el alto Índice de embriones
somáticos deformados (Steinitz eí a/., 2003), y 4) la baja tasa de germinación y/o
conversión de los embriones somáticos en plantas (Steinitz ef a/., 2003, López ef a/.,
2006). Dada esta problemática de la especie, se vio la necesidad de realizar estudios
morfológicos y moleculares durante la histogénesis de los embriones somáticos de chile
habanero en presencia de 2,4-D, con la finalidad de determinar la .impl.icación de este
RCV así como diversos factores de cultivo en la recalcitrancia del género a la
morfogénesis /.n v/.fro.
20

CApiTULO 1
1.9 0BJETIVOS
1.9.10BJETIVO GENERAL
Caracterizar el desarrollo de los embriones somáticos bajo diversas concentraciones de la
auxina sintética 2,4-D y su implicación en el proceso de la ES del chile habanero.
1.9.2 0BJETIVOS ESPECIFICOS
Determinar la calidad, frecuencia y morfología de los embriones somáticos a diferentes
concentraciones de la auxina 2,4-D y a diferentes tiempos de exposición a esta auxina.
Evaluar el desarrollo de la ES inducida en medio sólido y medio líquido.
Clasificar los diferentes tipos de anormalidades presentes durante la embriogénesis
somática de chile habanero.
Caracterizar mediante técnicas histológicas, las anormalidades presentes en embriones
somáticos de chile habanero.
Caracterizar la expresión del gen Wusche/ (WUS) durante el desarrollo del proceso de
ES.
21

CAPITULO 1
BIBLIOGFtAFiA
AboshamaH.M.S.(2011)DirectSomaticEmbryogenesisofPepper(Caps/.cumamuumL.)
World Journal of Agri.cultural Sciences 7 (6): 755-762.
Agrawal, S. and Chandra N. (1983) Differentiation of multjple shoot buds and plantlets in
cultured embryos of Caps/cum annum L. var. Mathania,Curr. Sci. 52:645-646.
Ammiralo P.V. (1974).The Effects of Abscísic Acíd on the Development of Somatic
Embryos from Cells of Caraway (Carum carvi L.) .Botanical Gazette, Vol. 135,
4:328-337.
Anandarajah, K., and MCKersie (1990). Manipulatlng the desiccation tolerance and vigor of
dry somatic embryos of Medicago sativa L. Wi.th sucrose, heat shock and abscisic
acíd. Plant Cell Reports. 9: 451455.
Asano Y., Katsumoto H., lnokuma C., Kaneko S., Ito Y. and Fujiie A.( 1996). Cytokinin and
thiamine requirements and stimulative effects of riboflavin and a-ketoglutaric acid
on embryogenic callus induction from the seeds of Zoysia japonica Steud. J. Plant
Physjol.14: 413417.
Ben C, Hewezi T, Jardinaud MF, Bena F. Ladouce N, Moretti S, Tamborindeguy C, Liboz
T, Petitprez M, Gentzbittel L (2005) Comparative analysis of early embryonic
sunflower cDNA libraries. Plant Molecular Biology 57, 255-270.
Bewley JD, Black M. Seed: germination, structure and composition.(1985) ln: Bewley JD,
Black M, editors. Physiology of Development and Germination. New York: Plenum
Press; p.1-3.
Binzel, M. L. Sankhla, N., Joshi, S. and Sankhla, D. (1996). Induction of direct somatic
embryogenesis and plant regeneration in pepper (Caps;.cum annuum L.). Plant
Cell Reports.15:536-540.
Bosland, P.W. (1994). Cmles: history, cultivation, and uses. ln: G. Charalambous (ed.),
Spices, herbs, and edible fungi. Elsevier Publ., New York. p. 347-366
Brown, D.C., Thorpe, T.A. (1995) Crop improvement through tissue culture. World Journal
22

CAPÍTULO 1
of Microbiology and Biotechnology.11 : 409-415.
Buyukalaca, S. and Mavituna, F. (1996), Somatic Embryogenesis and plant regeneration
of pepper in liquid media. Plant Cell, Tiss. Org. Cult. 46: 227-235.
Carimi F., Pasquale De F. and Crescimanno F.G.( 1999). Somatic embryogenesis and
plant regeneration from pistn thin cell layers of Citrus. Plant Ceu Rep.18: 935-
940.
Castillo 8. and Smith M.A.I.( 1997). Direct somatic embryogenesis from Begonia gracilis
explants. Plant Cell Rep.16: 385-388.
Che P, Lall S, Nettleton D, Howell SH (2006) Gene Expression Programs during Shoot,
Root, andcallus Development in Arabidopsis Tissue Culture Plant Physiol
141 :620
Che P, Love TM, Frame BR, Wang K, Carriquiry AL, Howell SH (2006)a Gene expression
patterns during somatic embryo development and germination in maize Hi H
callus cultures Plant Mol Biol 62:1-14
Chen J. and Chang W. (2001). Effects of auxins and cytokinins on direct somatic
embryogenesis on leaf explants on Oncid.ium `Gower Ramsey'. Plant Growth
Regul. 34:229-232.
Choi Y.E„ Yang D.C., Park J.C„ Soh W.Y. and Cho.i K.T. (1998). Regenerative ability of
somatic single and multiple embryos from cotyledons of Korean ginseng on
hormone free med.ium. Plant Cell Rep.17: 544-551
Chugh A, Khurana PJ (2002) Gene express.ion during somatic embryogenesis. Recent
advances.Curr Sci 83:715
Cooke, T.J„ Poli, D., Sztein, A. E. Y Cohen, J.D. (2002) Evolutionary patterns in auxin
action. Plant Molecular Biology. 49(34): 319-338.
Desamero, N.V., Rhodes, 8.8., Decoteau, D.R., and Bridges, W.C. (1994). Picolinic acid
induced direct somatic embryogenesis in sweet potato. Plant Cell, Tiss. Org. Cult.
23

CAPÍTULO I
37: 103-111.
Dodeman V.L., Ducreux G. & Krei's M. 1997 Zygotic embryogenesis versus somatic
embryogenesis. J. Exp. Bot. 48,1493-1509.
Fári M. and Czakó M. (1981). Relationship between position and morphogenetic response
of pepper hypocotyls explants cultured /.n v/.Íro. Science Hort.15: 207-213.
Farnsworth E (2000). The ecology and physiology of viviparous and recalcitrant seeds.
Ann. Rev. Ecol. Syst. 31 :107-138.
Feher A, Pastemak TP, Dudits D (2003) Transition of somatic cells to an embryogenic
state. Plant Cell Tissue Organ Cult 74:201
Fernandez H., Perez C. and Sanchez-Tames R.( 2000). Modulati.on of the morphogeníc
potential of the embryonic axis of Juglans regia by cultural conditions. Plant
Growth Regul. 30: 125-131.
Finstad, K., Brown, D. C., and Joy, K. (1993). Characterization of competence during
inductjon of somatic embryogenesis in alfalfa tissue culture. Plant Cell, Tjss. Org.
Cult.34: 125-132.
Francis D. and Sorrell D.A. (2001). The interference between the ceH cycle and plant
growth regulators: a mini review. Plant Growth Regul. 33: 1-12.
Fujimura Tatsuhito and Komamine Atsushi.(1979) Synchronization of Somatic
Embryogenesis in a Carrot Cell suspensíon Culture. Plant Physiol. 64,162-164.
Gaj M. (2004). Factors influencing somatic embryogenesis induction and plant
regeneration with particular reference to A. Íha//.ana (L.) Heynh. Plant Growth
Regul. 43: 27+7.
George Edwin F., HallGeen Michael, Klerk bGeert-Jan,(2008) Plant Propagation by
Tissue Culture 3rd Editionrspringer.
George, E.F. (1993). Plant Propagati'on by Tissue Culture. Segunda ed. vol.1.Edington
Wilts: Exegetics. England.1361 pp.
24

CApiTULO 1
Giroux RW, Pauls KP (1997) Characterization of somatic embryogenesis-related cDNAs
from alfalfa (Med/.cago saf/.va L.) Plant Mol Biol 33:393-404
Gunay A. and Rao P. (1978). /n vi.fro plant regeneration from hypocotyls and cotyledon
explants of red pepper (Caps/.ctJm). Plant Sci. lett. 11 : 365-372.
Haberlandt G. (1902) Culturversuche mit isol.ierten Pflanzenzellen. Sitz-Ber Math-Nat KI
KaisAkad Wiss Wien ;111 :69-92.
Halperin Walter and Wetherell Donald F. (1965) Ammonium Requirement for
Embryogenesis /`n vi.íro Nature 205, 519 -520
Halperin, W. (1995). /n v/.fm embriogénesis: some historical issues and unresolved
problems. ln: /n v/.{ro Embriogénesis in plants. (Ed.) Thorpe TA. Kluwer Academic,
Dordrecht, Netherlands.pp.1 -16
Harini L..and Lakshmi Sita, G. (1993) Direct somatic embryogenesis and plant
regeneration from immature embryos of chili (Caps/'cum annuum L.). Plant Sci.
89: 107-112.
Hyde C. and Phillips G. (1996). Silver nitrate promotes shoot development and plant
regeneration of chili pepper (C. annuum) via organogenesis. /n v/.íro Cen Dev.
Biol. Plant 32: 72-80.
lkeda M, Umehara M, Kamada H (2006) Embryogenesis-related genes; lts expression and
roles dur.ing somatic and zygotic embryogene§is in carrot and Arao/.dops/.s Plant
Biotechnol 23: 153-161
lkeda Y, Banno H, Niu QW, Howen SH, Chua NH (2006) The ENHANCER OF SHO0T
REGENERATION 2 gene in Arabidopsis regulates CUPSHAPED COTYLEDON 1
at the transcriptional level and controls cotyledon development. Plant Cell Physiol
47: 1443-1456
lmin, N.: Nizamidin, M.: Daniher, D.; Nolan, K. E.; Rose, R. J.; Rolfe, 8. G.(2005)
Proteomic analysis of somatic embryogenesis in Med.icago truncatula. Explant
cultures grown under 6-benzylaminopurine and 1-naphthaleneacetic acid
25

CAPÍTULO 1
treatments. Plant Physiol.137:1250-1260.
Jo J., Choi E., Choi D. and Lee, K. (1996). Soma{ic embryogenesi.s and plant regeneration
from immature zygotic embryo culture in pepper (C. annuum L.). J. Plant. Biol.
39:127-135.
Karami 0., Aghavaisj 8., Pour A. M., (2009) Review Molecular aspec{s of somatic-to-
embryogenic transition jn plants J Chem Bjol 2:177-190
Kelly R., Zipf A., Wesenberg D.E. and Sharma G.C.( 2002). Petrescjne-enhanced somatjc
embryos and plant numbers from elite oat (Avena spp. L.) and reciprocal crosses.
/n v/.Íro Cell. Dev. Biol. -Plant 38: 508-512.
Kevers C., LeGal N., Montejro M., Dommes J. and Gaspar T. (2000). Somatic
embryogenesis of Panax ginseng in liquid cultures: a role for polyamjnes and their
metabolic pathways. Plant Growth Regul. 31 : 2019-214.
Khan H., Síddique 1. and Anis M. (2006). Thidiazuron induced somatic embryogenesis and
plant regeneration jn C. annuum. Biologia Plantarum 50, 4: 789-792.
Kintzios S., Drossopolous J. and Lymperopoulos C. (2001). Effect of vitamins and
inorganic micronutrients on callus growth and somatic embryogenesis from leaves
of chillí pepper. Plant Cell Tjss. Org. Cult. 67:55-62.
Kochba J., Button, J. and Bornman C. H. (1974) Fine Structure of Embryoid Development
from Embryogenic Ovular Callus of `Shamouti' Orange (Citrus sinensis Osb.).J.
Exp. Bot. 25: 446457.
Kothari S.L., A. Joshi, S. Kachhwaha and N. Ochoa-Alejo (2010) Chilli peppers -A
review on tissue culture and transgenesis Biotechnology Advances, 28(1): 35-48.
Laborde, J. A. y 0. Pozo. 1984. Presente y pasado del chile en México. Publjcación
especial No 85. lNIA, SARH. 80 p.
Lakshmi Sita G, Raghava Ram NV, Vaidyanathan CS. (1979). Differentiation of embryoids
and plantlets from shoot callus of sandalwood. Plant Scj Lett 15, 265-70.
26

CAPÍTULO 1
Larkin P.J. y Scowcroft W.R. (1981) Somaclonal variation -a novel source of variabmty
from cell cultures for plant improvement. Theor. Appl. Genet. 60,197-214.
Latournerie L. M., Yupit M.E., Tún S. J., Pech M.A.M. y Lozada C.D. (2006).
Compartimiento de materiales criollos de Chile habanero ante el ataque de la
virosis. Uuy u T'aan.(Fundación Produce Yucatán, A.C.) 1 :15-17.
Ledón V. J.C. (2006). lmportancia del chile habanero en Yucatán. Uuy u T'aan.(Fundación
Produce Yucatán, A.C.) 1 :4-5.
Lee K., Zapata K., Brijnner H. and Afza R. (1997). Histology of somatic embryo initiation
and organogenesis from rhizome explants of Musa spp. Plant Cen Tiss. Org. Cult.
51: 1-8.
Lee, H. S., Fischer, R. L., Goldberg, R. 8., Harada, J. J. (2003) Arabidopsis LEAFY
COTYLEDONl represents a functionally specialized subunit of the CCAAT
binding transcription factor. Proc. Natl. Acad. Sci. USA 100:2152-2156.
Litz R. E., Hemdrix, RC., Moon, P.A., y Chavez, V.M. (1998). lnduction of embriogenic
mango cultures as affected by genotype, explanting, 2,4-D and embryogenic
nurse culture. Plant Cell, Tiss. Org. Cult. 53: 13 -18.
Liu W., Parroft W., Hildebrand D., Collins G. and Williams E. (1990). Agrobacterium-
induced gall formation in beH pepper (Caps/.cum annuum L.) and formation of
shoot-like structures expressing introduced genes.Plant Cell Rep. 9:360-364.
López-Puc G., Canto-Flick A., Barredo-Pool F., Zapata-Castillo P., Montalvo-Peniche M.,
Barahona-Pérez F. and Santana-Buzzy N. (2006). Direct Somatic
Embryogenesis: A mghly Efficient Protocol for /n v/.íro Regeneration of Habanero
Pepper (Caps/.cum ch/.nense Jacq.) Hortsc.ience 41 (6) 1-7.
Lotan, T., Ohto, M., Yee, K. M., West, M. A. L„ Lo, R., Kwong, R. W., Yamagishi, K.,
Fischer, R. L., Goldberg, R. 8., Harada, J. J.(1998) Arabidopsis LEAFY
COTYLEDONl is sufflcient to induce embryo development in vegetative cells.
Cell 93:1195-1205,
27

CAPÍTULO I
Meinke D.W.1995 Molecular genetics of plant embryogenesis. Annu. Rev. Plant Physiol.
Plant Mol. Biol. 46, 369-394
Merkle SA, Parroft WA, Flinn 8 S. Morphogenic Aspects of Somatic Embryogenesis
(1995): Thorpe TA, editor. /n v/.fno Embryogenesis in Plants. Kluwer Academic
Publisher. p.155-203.
Merkle, S.A., (1990). Maturation of yellow-poplar somatic embryos. ln: Ahuja, M.R (Ed),
Proceedings of the IUFRO-NATO Advanced Research Workshop on Woody Plant
Biotechnology. Plenum Press, New York.
Merkle, S.A., Parrot, W.A., Flinn, B.S. (1995). Morphogenic aspects Ín somatic
embryogenesis. ln: Thorpe, T.A. (ed.). /n v/.Íro Embryogenesis i.n Plants. Kluwer
Academic Publishers. Dordrecht. pp.155-203.
Michalczuk L., Ribnicky D., Cooke T. and Cohen D. (1992). Regulation of lndole-3-Acetic
Acid Biosynthetic Pathways in Carrot Cell Cultures. Plant Physiol.100:1346-1353.
Nanda R.M. and Rout G.R. (2003). /n ví.fro somatic embryogenesis and plant regeneration
in Acacia arabica. Plant Cell Tjss. Org. Cult. 73: 131-135.
Nuez F., Gil 0. R., Costa J. (2003). El cultivo de pimientos. Chiles y Ajies. Mundi-prensa
España. P 15.
Ochoa-Alejo N. and Ramírez-Malagón R. (2001). /n v/.fro cm pepper Biotechnology. /n
vÍ.Íro Cell. Dev. Biol-Plant. 37(6): 701-729.
Ochoa-Alejo, N. and lreta-Moreno, L. (1990) Cultivar differences in shoot forming capaci.ty
of hypocotyl tissues of chilli pepper (Capsícum annuum L.) cultured /.n v/.fno. Sci.
Hort. 42: 21-28.
Parroft WA, Dryden G, Vogt S, Hi.Idebrand DF, Collins GB, Williams EG (1988).
Optimization of somatic embryogenesis and embryo germination in soybean. /n
vt.Íro Cell Dev Biol 24:817-820.
Pérez Molphe-Bach E.; Ramírez-Malagón, R.; Núñez-Palenius H. y Ochoa-Alejo N.
28

rCAPÍTULO 1
(1999). lntroducción al Cultivo de Tej.idos Vegetales. Universidad Autónoma de
Aguascalientes. Primera edición México,179 pp.
Phillips G. and Hubstenberger J. (1985). Organogenesis in pepper tissue culture. Plant
Cell Tiss. Org. Cult. 4: 261-269.
Quiroz-Figueroa R., Fuentes-Cerda C., Rojas-Herrera R. and Loyola-Vargas V. (2002).
Histological studies on the developmental stages and differentiation of two
different somatic embryogenesis systems of C. arabica. Plant Cell Rep. 20:1141-
1149.
Raghavan, V. (1986). Embryogenesis in Angiosperms: A Developmentaland Experimental
Study. (Cambridge, UK Cambridge University Press).
Reinert J. 1958 Untersuchungen über die Morphogenese an Gewebenkulturen. Ber.
Dtsch. Bot. Ges. 71,15.
Rensing, S. A.; Lang, D.; Schumann, E.; Reski, R.; Hohe, A.( 2005) EST sequencing from
embryogenic Cyclamen persicum ceM cultures identifies a high proportion of
transcripts homologous to plant genes involved in somatic embryogenesis. J.
Plant Growth Regul. 24:102-115.
Roca, W. M. y Mroginski, L. A. , (1991 ), Cultivo de tejidos en la agricultura: fundamentos y
aplicaciones. Centro lnternacional de Agricultura Tropical (CIAT).Cali, Colombia,
953 pp.
Rochon M, Piola F, Deunff E, Mol R, Dumas C (1998). /n vttro development of maize
inmature embryos: A tool for embryogenesis analysis. J. Exp. Bot. 49:839-845.
Rose R. J. And Nolan K.E.,(2006) lnvited review: genetic regulation of somatic
embryogenesis with particular reference to Araó/.dops/.s fha//.ana and Med/.cago
ímncaíu/a /.n v/.fro cell. Dev. Biol.-plant 42:473481
Rosenberg, L.A. and Rinne, R.W. (1988). Protein synthesis suring natural and presosious
soybean seed (Glycine max [L] Merr.) maturation. Plant Physiology. 87: 474478.
29

CAPÍTULO 1
Santana-Buzzy N., Canto-Fli.ck A„ Barahona-Pérez F., Montalvo-Peniche M.C., Zapata-
Castillo P. Y., Solís-Ruiz A., Zaldívar-Collí A., Gutiérrez-Alonso 0. and Miranda-
Ham M. L. (2005). Regeneration of Habanero Pepper (Capsí.cum ch/.nense Jacq.)
Via Organogenesis. Hor{Science 40(6): 829-1831.
Santana-Buzzy N., Canto-Flick A., lglesias-Andreu L. G., Montalvo-Peníche M.C., López-
Puc G. and Barahona-Pérez F. (2006). lmprovement of /n vt.fro Culturing of
Habanero Pepper by lnhjbition of Ethylene Effects. Hortscience 41 (2) 405-409.
Santana-Buzzy N., Guadalupe Lopez-Puc, Adriana Canto-Flick, Felipe Barredo-Pool,
Eduardo Balam-Uc, Susana Aviles-Vi.ñas, Daniela Solís-Marroquin, Carlos
Lecona-Guzman, Jerico Jabin Bello-Bello, Eunice Gomez-Uc and Javjer 0.
Mijangos-Cortes (2009) Ontogenesis of the Somatic Embryogenesis of Habanero
Pepper Ícaps/.cum ch/.nense Jacq.)Hortscience 44(1 ): 113-118.
Schmidt, E. D. L., Guzzo, F., Toonen, M. A. J., de Vri.es, S. C.(1997) A leucineri.ch repeat
containing receptor-like kinase marks somatic plant cells competent to form
embryos. Development 124:2049-2062.
Senaratna, T., MCKersie, 8.0., y Bowely (1990). Artifici.al seeds of alfalfa (Medjcago sativa
L.) lnduction of desiccation tolerance in somati.c embryos. /n vÍ.Íno Cell. Dev. Biol.
16: 85-90.
Steinitz 8., Küsek M., Tabib Y., Paran 1. and Zelcer A. (2003). Pepper (C annuum L.)
regenerants obtajned by direct somatjc embryogenesis fail to develop a shoot. /n
v/.fro Cell Dev. Bi.ol. Plant. 39: 296-303.
Steward F., Mapes M. and Mears K. (1958). Growth and organized development of
cultured cells. 11: Organization in cultures grown from freely suspended cells.
Amer. J. of Bot. 45:705-708.
Stuar, D.A.,Nelse,J.,and Nichol,J.W.(1988) Expressíon of 7S and 11 S alfalfa seed storage
proteins in somatic embryos. J. Plant Physiol.132: 134il39.
Thibaud-Nissen FO, Shealy RT, Khanna A, Vodkin LO (2003) Clusteri.ng of microarray
data reveals transcript patterns associated with somatic embryogenesis jn

CAPÍTULO 1
soybean. Plant Physiol 132: 118-136
Thomas T.L. (1993) Gene expression during plant embryogenesis and germ.ination: An
overview. Plant Cell 5,1401-1410.
Thorpe E. (1995)./n v/.fro embryogenesis in plants. Kluwer Academic Publishers.
Netherlands.
Tun Dzul José (2001) Chile habanero Caracteristicas y Tecnología de producción,
SAGARPA-lNIFAB.:5-16.
Victor J.M.R., Murch S.J., KirshnaRaj S. and Saxena P.K. (1999). Somatic embryogenesis
and organogenesis in peanut: the role of thidiazuron and N6-benzylaminopurine
in the induction of plant morphogenesis. Plant Growth Regul. 28: 9-15
Wilson J.W., Wilson P.M.W. (1993) Mechan'isms of auxin regulation of structural and
physiological polarity in plants, tissues, cells and embryos. Aust J Plant Physiol
20:555-571
Xing, G.M.. Li. S.. Cui, K.R. and Wang, Y.F. (2000). Mechanisms of plant somatic
embryogenesis. Progress ln Natural Science.10(9): 641-649.
Yadav J. and Rajam M. (1998). Temporal regulation of somatic embryogenesis by
adjusting cellular polyamine content in eggplant. Plant Physiol.116: 617J525.
Yeung E.C. (1995) Structural and developmental patterns in somatic embryogenesisin
Thorpe T.A. (ed.) /n vjíro Embryogenesis in Plants. Kluwer Academic Publishers
Dordrecht, Boston, London. . pp. 205-247
Zapata-Castillo Y. P., Canto-Flick A., López-Puc G., Solís-Ruiz A., Barahona-Pérez F. and
Santana-Buzzy N. (2007). Somatic Embryogenesis in Habanero Pepper (C.
chí'nense Jacq.) From Cell Suspensions. Hortscience 42 (2):1-5.
Zdravkovic-Korac S, Neskov.ic M (1999) lnduction and development of somatic embryos
from spinach (Spinacia oleracea) leaf segments Plant Cen Tissue Organ Cult
55: 109-114
31

CAPÍTULO 1
Zhu C, Perry SE (2005) Control of expression and autoregulation of AGL75, a member of
the MADS-box family Plant J 41 :583-594
Zimmerman J. L., (1993) Somati.c Embryogenesis: A Model for Early Development in
Hjgher Plants. The Plant Cell, 5,1411-1423
Zimmerman, J.L.. Apuya, N., Darwish K., afld-O'Carrol. C. (1989). Novel regulation of heat
shock genes during carrot embryo development. The Plant Cell 1 : 1137-1146.
Zuo, J. R., Niu, Q. W., Frugis, G., Chua, N. H.(2002) The WUSCHEl gene promotes
vegetative-to-embryonic transition in Arabí.dops/.s. Plant J. 30:349-359.
32

CAPÍTULO 11
CAPITUL011
MORPHO-HISTOLOGICAL AND ULTRASTRUCTURAL STUDY 0N DIRECTSOMAT]C EIVIBRYOGENESIS OF Capsi.cum cAi.nense Jacq. IN LIQU]DMEDIUM
Susana A. Avilés-V.iñas`, Carlos A. Lecona-Guzmán`, Adriana Canto-Flickí, Stephanie
López-Erosa` , Nancy Santana-Buzzy'*
`Unidad de Bioquímica y Biología Molecular de Plantas, Centro de lnvestigación Científica
de Yucatán. Calle 43 #130. Chuburná de Hidalgo, Mérida, Yucatán. 97200, México.
*Corresponding Author: Email: [email protected]; phone +52 (999) 9428330; fax +52 (999)
9813900.
•:. Este capítulo ha sido aceptado para su publicación en la revista Plant
Biotechnology Reports
33

CAPÍTULO 11
2.1 lNTRODUCTloN
Somatic embryogenesis (SE) can be described as the process through which haploid or
diploid somatic cells develop into different plants through characteristic embryological
stages without fusion of gametes (Wlliams and Maheswaran,1986). Typically, a somatic
embryo (SEs) Ís a plant in its initial stage of development possessjng bipolarity, with shoot
and radical poles opposite jn the same axis, without vascular connec{ion with the mother
tjssue, and from an origin of either unicellular or multicellular nature.
Capsicum SEs has been obtained from immature zygotic embryos (Harinj and Lakshmi
Sita,1993; Binzel ef a/.,1996; Jo ef a/.,1996), mature zygotic embryos (Buyukalaca and
Mavituna,1996), or from fully-expanded first leaves of seedlings (Kintzios ef a/., 2001).
There are only two reports of indirect SE (Buyukalaca and Mavituna, 1996; Zapata-
Castillo ef a/., 2007) working with C. annLíum and C. ch/.nense, respectively. In all other
instances embryos were derived directly from the source tissue. However, {he
recalcitrance of the genus Capsicum to the regeneratjon of whole plants /.n v/.fro has been
amply documented by numerous reports (Harini and Lakshmi Sita, 1993; Binzel ef a/.,
1996; Jo eí a/.,1996; Ochoa and Ramírez, 2001; Steinitz eí a/., 2003; López-Puc eí a/.,
2006; Santana-Buzzy ef a/., 2006; Santana-Buzzy eí a/., 2009; Kothari, 2010; Heidmann ef
a/„ 2011 ).The recalcitrance of the genus Capsicum is manifested by the low efficiency of
regeneration protocols (Buyukalaca y Mavituna, 1996), the hjgh rate of SEs deformed
(Steinitz ef a/., 2003), and the low rate of germination and conversion of SEs into plants
(Steinitz ef a/., 2003, LÓpez eí a/., 2006, Kaparakis and Alderson, 2008). Taking account
this problem, the objective of this study was Study the response from habanero pepper
hypocotyls under different concentratíons and exposure time to 2.4-D, in liquid medium or
semisolid during SE.
2.2 MATERIALS AND METHODS
2.2.1 PLANT MATERIAL SOURCE
Seeds of Habanero pepper var. Mayan Ba'alche were surface-sterilized in 70°/o (v/v)
ethanol (5 min). Subsequently, the seeds were jmmersed for 15 min in 1.8% (v / v) sodium
hypochlorite, and washed four times i.n autoclaved dei.onized water for 5 min. Disinfected
34

CApiTULO 11
seeds were cultured on germination medium containing Murash.ige and Skoog (MS,1962)
mineral salts and vitamins, 3°/o (w / v) sucrose, 0.22% (w / v) gelrite and (1.1 mM) GA3.
ThepHofthemediumwasadjustedto5.8,priortosterilization.Themediumwassterilized
in an autoclave at 121 ° C and 1.5 atm for 15 min. the cultures were incubated in darkness
at 25 ± 2 ° C for a perkxl of 7 days. After this time, the cultures were transferred to 16 h
lightphotoperiod(40-50molm-2s-1),25±2°C,wheretheyremainedfor15-20days.
2.2.2 SOMATIC EMBRYO INDUCTION
Hypocotyl segments (0.3-0.5 cm) from aseptic plants were used as explants for the
induction. The effect of 2,4-D at different concentrations and exposure time, as well as the
effect of the culture medium consistency on the explants were evaluated; the results are
shown in Table 2.1. Fifty explants were used in each treatment, distributed equally in five
magenta vessels. The cultures in liquid medium were maintained in constant agitation (100
rpm). The culture medium was changed every 15 days. SEs formation frequency was
evaluated in each treatment. All treatments were maintained under constant illumination
(40-50 uM.m-2.s-t), at 25 ± 2 oC.
Table 2.1 Treatments evaluated during the induction and development of
Habanero pepper SE.
Concontratlon 2,4-D (uM)
1 5 daysTreaú")ris
30 days 45 days 60 days
(S: semi-solid medium; L: l.iquid medium).
35

CAPITULO 11
For germination, SEs in more advanced stages (torpedo -cotyledonary) were cultured in
ljquid medium (MS) supplemented with AG3 (1.1 uM) as the sole source of growth
regulator. The cultures were incubated in a 16-hour light photoperiod (40 -50 Hmol m-2 s-`),
at 25 ± 2 °C, for two weeks.
2.2.3 STATISTICAL ANALYSIS
Statistícal analysis of the experi.ments was performed using a completely randomized
design. For analysis of variables medíum consistency and 2,4-D concentration in the
culture medi'um, was used a 2 factor experimental design. Factor 1: Medium consistency
with two levels. Level 1: 30 days Soli.d and 30 days líquid (T2, T3, T4) and Level 2: 30
days liquid and 30 days liquid (T5, T6, T7). Fac{or 2: concentration to 2,4-D. Treatments
with three levels. Level 1 : 30 days in 9.05 uM and 30 days in 4.5 uM (T2 and T5), Level 2:
30 days in 9.05 HM and 30 days in 0 uM (T3 and T6) and Level 3: 15 days in 9.05 uM,15
days in 4.5 uM and 30 days in 0 LiM (T4 and T7). The results of the variables analyzed
were subjected to analysis of variance (ANOVA). Differences among the developmental
stages among treatments were revealed through ANOVA. lt was used a Person's chi
square to test differences among treatments regarding the dístribution of SEs.
2.2.4 SCANNING ELECTRON MICROSCOPY (SEM)
During the process of SE, bi.ological samples were taken; these were subsequently
submerged in a glutaraldehyde solution (25%) at 3 % in sodium cacodylate buffer O.1M at
pH 7.2, followed by three washes with the same buffer for 30 min. Dehydration was carried
out in hourly stages with increasing dissolutions of ethanol (50, 70, 90% v/v). The
dehydrated explant tissues were then submerged twi'ce i.n absolute ethanol for 1 hour each
and were dried usíng a Sandri-795 Tousimis® drier, substítu{i.ng ethanol with liquid C02,
and subsequently with gaseous C02. Finally, the dried samples were metalized with a 21 -
nm thick gold cover using a Denton® Vacuum Desk 11 metalizer. The metalized samples
were observed under a JSML6360LV JEOL® Scanning Electron Microscope.
2.2.5 HISTOLOGICAL ANALYSIS
The samples were placed in a fixing solutjon of formali.n aceti.c alcohol containing 5 mL of
96% ethanol,10 mL of 37% formaldehyde, 5 mL acetic acid, and adjusted to a 100 mL
36

CAPÍTULO 11
volume with distilled water, for 24 to 48 h at 10°C, following the Berlin and Miksche (1976)
protocol. After fixing, the samples were dehydrated by successive changes in ethanol
solutions at different concentrations (30%, 50%, 70%, 85%, 99%, and 100% v/v).
Subsequently, after infiltration, the samples were embedded in plastic resin (J84 Glycol
Methacrilato; Polysciences, Warrington, PA). Serial sections of 5 um were cut off in a
rotary microtome HM340E (Microm lntemational GmbH), mounted on cytological glass
slides, and stained with 0.05% toluidine blue (3 to 5 min). Observations and .images wei.e
obtained with an inverted microscope (Axiovert MCSO; Carl Zeiss D-73446).
37

CAPÍTULO 11
2.3 RESULTS
2.3.1 EFFECT OF 2,4-D AND CULTURE MEDIUM CONSISTENCY
The results of variance analysis (ANOVA) showed no signjficant differences (P <0.05) for
the variables of culture medium consistency and concentration of 2,4-D. The analysis of
variance for the developmental stages between each treatment (Table 2.1 ) indicates that
sígni.ficant differences exist in each of the stages of development. According to the
analysis for the distribution of each of the stages of development using chi square the
results indicate that the treatments are different from one another with respect to the
distribu{ion of the SEs, because the value obtajned from chi square is hjgher (370,138)
than the value of the critical chi. square (28.86) and is located outside the acceptance
region. The confidence level was 0.05. Regarding the efflciency, there were significant
differences among the treatments (Figure 2.1 ).
Efflciency
TreatmentsFigure 2.1 Efflciency (SEs/ Liquid Medium) of SE in the different treatments
evaluated. Data recorded after 45 days of culture. Each treatment was
replicated five times, and each replicate consisted of 10 explants. Means having
djfferent letters in superscript are signjficantly different from each other (P <
0.05) according to Duncan's multiple range test.
38

CAPÍTULO 11
The highest response of SEs formation (6.75X104 SEs/L) was observed in treatment T7,
differing significantly from the other treatments. Highest response in the induction of
habanero pepper SEs was obtained when the explants were exposed to liquid medium (60
days) supplemented with 9.05 uM 2,4-D for the first 15 days, with a subsequent reduction
to half this concentration (2,4-D 4.50 LiM) for the following 15 days, and the withdrawal of
this auxin from the medium for the last 30 days of culture. Nevertheless, the SEs formed
did not advance in their development and remained in the globular stage (100%) and died
at 60 days of culture. In contrast, in treatment T2, although efficiency was significantly
lower than in treatment T7 (1.77X104 ESs/L), results showed that it was the only treatment
in which the majority of the SEs reached the cotyledonary stage (58.7%), at 60 days
(Table 2.2).
Table 2.2 SEs, per stage of development, in the different treatments evaluated
Treatment§
100 ± Oa
Stages of development %
33.2 ±5.7C 3.6 ± 0.60a
88.5 ± 8.|b,o ob
100 ± Oa Ob
76`7 ± 1.9b 3.5±0.65a
92.0 ± 2.3b.c ob
100 ±Oa Ob
4.50 ± 0.83aib 58.7 ± 5.3a
4.80 ± 1.3a.b 6.70 ±4.8b
OüOC
7.20 ±0.55a 12.6 ± 1.3b
2.70 ± 1`4b.C 5.20 ± |.|b
(G: globular embryo, H: heart-shaped embryo, T: torpedo embryo, C: cotyledonary
embryo). Means in each column followed by same letters are not significantly different.
ln all probability, the culture regime (see Table 2.1; Materials and Methods) is a
determining factor for SE development. lnduction and development of SEs in a solid
39

CApl'TULO 11
medium for the first 30 days, containing 9.05 uM 2,4-D, and the remai'ning 30 days in a
liquid medium with the concentration of 2,4-D reduced to half (T2) was significantly
favorable to the process. Was interestjng to observe that in treatments T2 and T5, in which
2,4-D was maintained throughout the culture time, were the treatments in which
cotyledonary somatic embryos were obtained and only those observed in embryos heah
stage. These results allow us to infer that exposure time to 2,4-D was an essential factor
for the development of SEs (Figures 2.2a, 2.2b), particularly when they were maintained in
a solid medium for the first 30 days of the process, and in a liquid medium for the following
30 days (Figures 2c, 2d). ln treatments Tl and T4, in addition to obtaining low efficiency
(O.185X104 and 0.253 X104 ESs/L respectively). The SEs that were formed underwent
browning and died three weeks after culture initiation. According to Michalczuk eí a/.
(1992) and Pasternak ef a/. (2002), the embryogenic competency present in some speci.es
is associated with the increase in endogenous IAA levels due to the presence of 2,4-D,
suggesting that this synthetic compound acts upon and alters the metabolism. As shown
in Table 2.2, a significantly higher number (58.7%) of SEs in the cotyledonary stage were
obtained in treatment T2 (Figures 2e, 2f), while a lower frequency of this stage was
observed in treatments T3, T5 and T6 (6.7,12.6 and 5.3%, respectively). The SEs formed
in T2 were characterized by a creamy white color and their remarkable synchrony from the
ea"est stages of their development. Most of the ESs in the cotyledonary stage (75-80°/o)
formed in treatment T2, emitted radicals and cotyledonary leaves (Figures 2.2g, 2.2h). The
results allow to infer that the variables studíed act as a system which both consistency of
the medium as concentration and exposure time to 2,4-D have influence on response of
Habanero pepper during induction and development of SEs. It is weH established that 2,4-
D plays a role in the induction of SE, which is presumably mediated by a signal cascade
triggered by this exogenous auxin (Zuo ef a/., 2002). lt has been reported that the culture
of explants in medium containing 2,4-dichlorophenoxyacetic acid (2,4-D), the classic
induction treatment for many species, increases the endogenous auxin levels in the
responsive explants (Michalczuk ef a/.,1992; Pasternak ef a/., 2002), being this synthesis
one of the crucial signals determiníng fate of cultured cells (Thomas eí a/., 2002). ln spite
of the large amount of research conducted during the last years, knowledge is still vague
in regards to the mechanisms by which plant hormones are involved in regulation of SE.
There is a pattern weH defined for highly responsive species and genotypes, in which the
auxi'n 2,4-D plays a positive role for jnducti.on, while wíthdrawal of this compound triggers
40

CApiTULO 11
expression, allowing development of SEs. However, this scenery is not so clear for the
more recalcitrant genotypes, in which the requirements can vary greatly (Jimenez eí a/.,
2005). ln conclusion, the results obtained show evidence of the dependence on C.
chinense to 2.4-D during induction and histodifferentiation of SEs, contrary to that reported
for most plant species.
41

CAPÍTULO 11
Figure 2.2 SE from habanero pepper: (a) Intense proliferation of SEs developed
entirely in liqui.d medium after 21 d in culture;(b) Cluster of SEs Ín the hypocotyl
after 25 d in culture in liquid medium;(c) SEs in eariy stages developed after 30
days in a solid medium and subsequent transfer to a liqui.d medium; (d) Cluster
of SEs separated from the hypocotyls 14 d after transfer to a liquid medi.um; (e)
SEs in advanced stages developed jn T2 after 35 d in culture; (0 SEs in the
cotyledonary stage developed in T2
42

CAPÍTULO 11
2.3.2 ANATOMICAL OBSERVATIONS
ln a cross section of the hypocotyl, the cortical parenchyma (cp) and provascular zone
(pz) were clearly visible, while deeper inside the hypocotyl, it was possible to distinguish
the tissues conforming the xylem and phloem (Figure 2.3a). Exposure of the explant to the
culture med.ium provoked intense mitotic activity evidenced by successive divisions of the
cells in closest proximity to the vascular bundles (Figure 2.3b). Division of the epidermal
cells never occurred. A very small thickwalled cell, with dense cytoplasm and prominent
nucleus suggests that the ceH will initiate a division cycle to form a two-celled pro-embryo
(Figure 3c), Embryogenic cells (3040 iim length) were observed close to the vascular
tissues (Figure 3d). The first stage of the differentiation of SEs is shown in Figures 3e, f.
Although the protoderm was distinguished .in the early globular stage (Figures 2.3e, f, g, h,
i), it was better defined in the late globular stage (Figures 2.3j, k). By successive divisions
these embryos gave origin to the expected stages of SE in dicotyledons: globular (Figure
2.31), hean (Figures 2.3m, n), and torpedo and cotiledonary shaped embryos where it is
possible to distinguish both meristems, caulinar (Figure 2.3o) and radicular (Figure 2.3p).
These results confirm that the morphogenetic process taking place is undoubtedly direct
SE. The anatomical aspect of SEs has been reported by many authors (Auboiron ef a/.,
1990; Gm and Saxena,1992; Decout ef a/..1994; Suhasini eí a/.,1994). However, only a
few studies have characterized the different ontogenetic stages of these embryos (Barwale
ef a/.,1986; Fransz and Schel,1991; Finer and MCMullen,1991; Kiss ef a/.,1991; Liu eí
a/.. 1992; Sato ef a/., 1993; Toonen ef a/.,1994; Yeung ef a/., 1996). ln the Caps/.cum
genus, histological sections from embryonic structures of de novo formation were reported
by Binzel eí a/. (1996). A more detailed characterization of the ontogenetic route followed
by SEs of Habanero pepper (C. ch/.nense) during their development, was performed
Santana-Buzzy ef a/. (2009).

CAPÍTULO 11
44
Figure 2.3 Histological analysis of SE in Habanero pepper. (a) Horizontal cross
section of hypocotyl at 3 d of culture; (b) Division of somatic cells proximal to
vascular bundles at 6 d of culture; (c, d) Embryogenic cells i.n divi.sion; (e, 0
globular embryos wi.th vascular connection to the explant; (g, h, Í) globular
embryos with suspensor-li.ke structure;( j, k,1) globular embryos wi.thout vascular
connection to the explant and vi'sible protoderm ; (m, n) heart- shaped SEs
containing cotyledonary initi'als; (o) cauljnar apex; (p) radical apex; ci
cotyledonary initials; cp cortical parenchyma; Co cotyledons; pt protoderm; pz
provascular zone; sp suspensor; vz vascular zone.

CAPÍTULO 11
2.3.3 ULTRASTRUCTURAL OBSERVATIONS
Based on SEM observations of the SEs in Habanero pepper hypocotyl cultures, it was
possible to observe that cellular acquisition of morphogenetic competence occurred
between 3 and 6 days of exposing the hypocotyl to the culture medium (Figures 2.4a, b).
During the first four days of culture, an intense proliferation of small isodiametric cells was
observed along the length of the vascular bundles, forming meristematic centers (mc);
these were the forerunners for the formation of proembryogenic sectors (Figures 2.4c, d).
However, the morphological changes in explant tissues became visible after the tenth day
of culture, the moment at which the hypocotyl epidermis spontaneously fractured
longitudinally, exposing a creamy-white proembryogenic mass that emerged from inside
the explant (Figure 2.4e). This proembryogenic mass showed small isodiametric cells with
increasing frequency (Figures 2.4f, g) which subsequently gave origin to embryos in early
stages (FigLires 2.5a, b, e, f). After 40 days of culture SEs were observed in more
advanced stages of development, such as torpedo (Figures 2.5c, g) and cotyledonary
(Figures 2.5d, h).
45

CAPÍTULO 11
Figure 2.4 Scanning electron micrographs of SE induction from 0 to 12 d of
culture. (a) Transverse cross section of hypocotyl at day 0 of culture; (b)
Longitudi.nal cross sectic)n of hypocotyl at day 0 of culture; (c) Longitudinal cross
section of hypocotyl at 4 d of culture; (d) Transverse cross section of hypocotyl
segment at 6 d of culture; (e) Fragmentatíon of hypocotyl with exposure of
proembryogenic mass at 10 d of culture;( f, g) High magnifi.cation showing the
arrangement of isodiametric spherical cells (embryo cells or embryo mother
cells) at 5 and 6 d of culture. mc meristematic centers.
46

CAPÍTULO 11
Figure 2.5 Scanning electron microscopy (SEM) a-d and microphotography e-h
of the explant between 25 and 40 days. (a, e) Explant at 25 days of induction;
(b, 0 SEs in the globular stage; (c, g) SEs in the torpedo stage; (d, h) SEs in
the cotyledonary stage.
47

CAPÍTULO 11
CONCLUSIONS
Among the different auxin, 2,4-D has been the most commonly used to induce SE. Over
65% of recent protocols are based on the use of exogenous 2.4-D as an inductor, alone or
in combination with other PGRs (Gaj, 2004). ln general, competent cells arise from
explants cultured in media supplemented with potent synthetic auxin, like 2,4-D, which can
act directly as an auxin, or modifying the content of lAA .intracellular, and / or metabolism
as agent "stressor" (Feher eí a/., 2003; Quiroz-Figueroa ef a/., 2006). There is a pattern
well defined for highly responsive species and genotypes, in which the auxin 2,4-D plays a
positive role for induction, wmle withdrawal of this compound triggers expression, allowing
development of SEs. Once the stimulus for the development of SEs .is given by the
reduction or elimination of 2,4-D from the culture medium, the endogenous levels of lAA
be reduced to allow the establishment of a polar gradient of auxin. The continued
presence of 2,4-D in the culture medium, prevents the reduction of endogenous auxin
levels resulting in inhibition of the development of SEs (Nissen and Minocha, 1993,
Fischer-lglesias ef a/., 2001). This scenery is not so clear for the more recalcitrant
genotypes, in which the requirements can vary greatly. C. ch/.nense, contrary to what has
been reported for the genus Caps/.cum has proved to be one of those with higher
embryogenic capacity, comparable with species recognized as models of efficiency of SE:
carrot (Halperin and Jansen,1967) and alfalfa (Dos Santos ef a/.,1983). During induction,
the requirements of C. ch/.nense follow the pattern described for most species
embryogenic: the presence of 2,4-D. Contradictorily, after induct.ion this auxin cannot be
suppressed of the culture medium because the development of the SEs is completely
inhibited. Considering that during histodifferentiation the poles (radical and cauline) of SEs
are defined, and form the vascular tissues that will accompany them throughout their lives,
and that 2.4-D is an auxin that can cause genetic alterations through DNA methylation
(Xiao ef a/., 2006), probably the inability of the embryos of C. ch/.nense could be
associated with the presence of this auxin can be as harmful as beneficial, for this
morphogenetic process contrary to what has been reported on the low morphogenetic
potential of the genus Capsicum (Agrawal ef a/., 1989; Ochoa-Alejo and lreta-Moreno,
1990; Valera-Montero and Ochoa-Alejo,1992; Ramírez-Malagón and Ochoa-Alejo,1996;
Husain ef a/., 1999; Venkataiah eí a/., 2003; Steinitz e£ a/., 1999; Ochoa-Alejo and
Ramírez-Malagón, 2001), we have established a promising direct SE system for a highly
48

CAPÍTULO 11
efficient regeneration of Caps/.ci/m ch/.nense, in liquid medium. Probably, the interaction
that occurs between hypocotyl cells, the auxin (2.4-D) and the liqukl medium promote
effect of self-control, resulting in the simultaneous induction of a large number of SEs
directly from explant cells. Similar behavior was observed during induction of somatic
embryogenesis of Daucus carofa hypocotyls cultured in vitro in the presence of 2,4-D
(Guzzoefa/.,1994).ProvascularcellsofHabaneropepperhypocotylssplitandexpanded.
Simultaneously divided were observed along the transverse and longitudinal planes.
Subsequently, these cells proliferated and small isodiametric has emerged as an
embryogen.ic cell line from which somatic embryos were formed. Similar behavior was
observed during induction of somatic embryogenes.is from carrot hypocotyl explants
cultured in vitro, in presence of 2,4-D (Guzzo ef a/.,1994). Provasculars cells of Habanero
pepper divided and expanded simultaneously along the longitudinal and transverse planes
of the explant. Subsequently, these smau and isodiametric cells proliferated form a row of
proembryogenic cells, resulting somatic embryos at four weeks of culture. Unlike most SE
systems have been characterized (Binzel eí a/.,1996; Khalil ef a/., 2002; Maximova ef a/.,
2002), we found no evidence that the high efflciency in the formation of SEs is the result of
secondary SE.Critical stages of embryogenesis, including cellular acquisition of
morphogenetic competence, suspensor formation, and somatic embryo development
during SE from hypocotyl segments of Habanero pepper (Caps/.cum ch/.nense Jacq.) were
identified by SEM. With our SE system, we have managed to overcome one of the
problems that have limited /.n v/'fro regeneration of species of the genus Caps/.cum: Iow
efficiency of the protocols. However, two questions remain unanswered: What factors are
involved in the low rate of conversion of somatic embryos into plants? What is the cause
of the high frequency of deformed SEs? The answers to both these questions wm depend
on further research in order to increase our understanding of the recalcitrance
phenomenon and to find a solut.ion for the inability of SEs of the genus Caps/.cum to
conven into plants. However, our results show a promising new outlook on the /.n vffm
regeneration of this species (C. chí.nense). Contrary to what has been reported to date for
Caps/'cum ch/.nense, it is a species of plants with higher embryogenic potential /.n v/.{ro.
Probably in the not so distant future, it could be one of the most efflcient and useful
regeneration systems for large scale production of plants and genetic improvement of this
Crop.
49

CAPÍTULO 11
LITERATURE CITED
Agrawal S., Chandra N., Kothari S.L.(1989) Plant regeneration in tissue cultures of pepper
(Capsi'cum annuum L. cv. Mathania). Plant CeH Tissue Organ Cult.116,47-55.
Auboiron, E., Carron, M.P., Michauxi Ferriere N. (1990) lnfluence of atmospheric gases,
partícularly ethylene, on somati.c embryogenesis of Hevea brasjliensis. Plant Cell
Tissue Organ Cult. 21,31-37.
Barwale, U.B., Kerns, H.R., Widholm, J.M. (1986) Plant regeneration from callus cultures
of several soybean genotypes via embryogenesis and organogenesis. Planta
167,473481.
Berlin, G.P., Miksche, J.P. (1976) Botanical microtechnique and cytochemistry. 3rd Ed.
lowa State Universjty Press, Ames,.lA.
Binzel, M.L., Sankhla, N., Joshi, S., Sankhla, D. (1996) lnduction of direct somatic
embryogenesis and plant regeneration in pepper (Caps/.cum annuum L.). Plant
Cell Reports 15,536-540.
Buyukalaca, S.,Mavituna, F. (1996) Somatic Embryogenesis and plant regeneration of
pepper in liquid media. Plant Cell, Tiss. Org. Cult. 46, 227-235.
Decout, E., Dubois, T., Guedira, M., Dubois, J., Audran J.C., and Vasseur J. (1994) Role
of temperature as a triggering signal for organogenesis or somatic embryogenesis
in wounded leaves of chicory cultured in vitro. J. Expt. Bot. 45,1859-1865.
Dos Santos AVP, Cutter EG, Davey MR (1983) Origi.n and development of §omatic
embryos in Medjcago sati.va L. (alfalfa). Protoplasma 117,107-115
Feher A., Pasternak T.P., Dudits D. (2003) Transition of somatic cells to an embryogenic
state. Plant Cell Tissue Organ Cult 74(3), 201-228.
Fi.ner, J. J., MCMullen, M.D. (1991) Transformation of soybean via parti.cle bombardment of
Embryogenic suspension culture tissue. ln Vjtro Cell. Dev. Biol. 27,175-182.
Fi'scher-lglesias C., Sundberg 8., Neuhaus G. and Jones A.M. (2001) Auxin distribution
50

CApiTULO 11
andtransponduringembryonicpattemformationinwheat.PlantJ.26,115-129.
Fransz, P.F., Schel, J.H. N. (1991) An ultrastructural study on the early development of
Zea mays somatic embryos. Canadian Journal of Botany. 69 (4), 858-865.
Gaj M. (2004). Factors influencing somatic embryogenesis induction and plant
regeneration with particular reference to A. thaliana (L.) Heynh. Plant Growth
Regul. 43, 2747.
Gill, R., Saxena, P.K. (1992) Direct somatic embryogenesis and regeneration of plants
from seedling explants of peanut (Aracms hypogaea): promotive role of
thidiazuron. Canadian Journal of Botany, 70(6) 1186-1192.
Guzzo, F., Baldan 8., Mariani P., Lo Schiavo F., Terzi M. (1994) Studies on the origin of
totipotent cells in explants of Daucus Carota L. J. Expt. Bot. 45,1427-1432.
Halperin W, Jensen WA (1967) Ultrastructural changes during growth and embryogenesis
in carrot cell cultures. J Ultrastruct Res 18, 428443
Harini, 1., Sita, L. (1993) Direct somatic embryogenesis and plant regeneration from
immature embryos of chilli (C. annL/um L). Plant Sci. 89,107-112.
Heidmann,1., de Lange, 8., Lambalk, J., Angenent, G.C., Boutilier, K. (2011) Efficient
sweet pepper transfomation mediated by the BABY B00M transcription factor.
Plant Cell Rep 30,1107-1115.
Husain S., Jain A., Kothari S.L.(1999) Phenylacetic acid improves bud elongation and in
vitro plant regeneration efflc.iency in Capsicum annuum L..Plant Cell Rep.19, 64-
68.
Jo, J„ Choi, E„ Choi, D„ Lee, K. (1996) Somatic embryogenesis and plant regenerat.ion
from immature zygotic embryo culture in pepper (C. annuum L.). J. Plant. Biol.
39,127-135.
Kaparakis, G. and Alderson, P.G. (2008) Role for Cytokinins in Somatic Embryogenesis of
Pepper (Capsicum annuum L.)?. J Plant Growth Regul. 27.110-114
51

CAPITULO 11
Khalil, S.M., Cheah, K.T., Perez, E.A., Gaskíll, D.A., Hu, J.S. (2002) Regeneration of
banana (Musa spp. AAB cv. Dwari Brazilian) via secondary somatic
embryogenesjs. Plant Cell Rep. 20.1128-1134.
Kintzios, S., Drossopolous, J., Lymperopoulos, C. (2001) Effect of vitamins and inorganic
micronutrients on callus growth and somatic embryogenesis from leaves of chilli
pepper. Plant Cell Tiss. Org. Cult. 67,55-62.
Kiss, E., Heszky, L.E., Gyulai, G., Horváth, H. S., Csillag, A. (1991) Neomorph and leaf
differentiation as alternati've morphogenetic pathways in Soybean ti.s§ue culture.
Acta Biol. Hung. 42,313-321.
Kothari, S.L., Kachhwaha, A.J.S., Ochoa-Alejo, N. (2010) Chilli peppers -A review on
tissue culture and transgenesi.s Biotechnology Advances, 28(1 ), 35-48.
Liu, W., Parrott, W., Hildebrand, D., Colli'ns, G.,Williams E. (1990) Agrobacfe//'wm-induced
gall formation in bell pepper (Caps/.cwm annuum L.) and formation of shoot-Iike
structures expressing introduced genes. Plant Cell Rep. 9,360-364.
López-Puc, G., Canto-Flick, A., Barredo-Pool, F., Zapata-Castillo, P., Montalvo-Penjche.
M., Barahona-Pérez, F., Santana-Buzzy, N. (2006) Direct Somatic
Embryogenesis: A Highly Efflcient Protocol for /n v/.Íro Regeneration of Habanero
Pepper (Capsí.cum ch/`nense Jacq.) Hortscience 41 (6),1-7.
Maximova, S.N., Alemanno, L., Young, A., Ferriere, N.,Traore, A.,Guiltinan, M.J. (2002)
Efficiency, genotypi.c variability, and cellular origin of primary and secondary
Somatic embryogenesjs of theobroma cacao 1. ln Vitro Cem Dev. Bjol. 38,252-
259
Michalczuk, L., Ribnicky, D., Cooke, T., Cohen, D. (1992) Regulation of lndole-3-Acetic
Acid Biosynthetic Pathways in Carrot Cell Cultures. Plant Physiol.100,1346-1353
Murashige, T., Skoog, F.A. (1962) A revised medium for rapid growth and bioassays with
tobacco cultures. Physiol. Plantarum 15 (3),437-497.
Ni'ssen P. and Minocha S.C.1993. lnhibition by 2,4-D of somatic embryogenesis in carrot
52

CAPÍTULO 11
asexploredbyitsreversalbydifluoromethylornithine.Physiol.Plant89,673-680.
Ochoa-Alejo N., lreta-Moreno L.(1990) Cultivar differences in shoot-forming capacity of
hypocotyl tissues of chilli pepper (Capsicum annuum L.) cultured in vitro. Sci
Hortic-Amsterdam. 42, 21 -28.
Ochoa-Alejo, N., Ramirez-Malagón, R. (2001 ) /n v#m chili pepper Biotechnology. /n V/.fm
Cell. Dev. Biol-Plant. 37(6), 701-729.
Pastemak, T., Prinsen, E., Aydin, F„ Mskolcz,i P„ Pofters, G., Asard, H., Van Onckelen,
H., Dudits, D., Fehér, A. (2002) The role of auxins, pH and stress in the activation
of embryogenic ceH division in leaf protoplast-derived cells of alfalfa. Plant
Physiol.129,1807-1819.
Quiroz-Figueroa F.R., Rojas-Herrera R., Galaz-Avalos R.M., Loyola-Vargas V.M. (2006)
Embryo production through somatic embryogenesis can be used to s{udy cell
differentiation in plants. Plant Cell Tissue Org Culture 86 (3),285-301.
Ramírez-Malagón R., Ochoa-Alejo N.(1996) An improved and reliable chim pepper
(Capsicum annuum L.) plant regeneration method. Plant CeH Rep.16,226-231.
Santana-Buzzy, N., Canto-Flick, A., lglesias-Andreu, L. G., Montalvo-Peniche, M.C.,
López-Puc, G., Barahona-Pérez, F. (2006) lmprovement of /n v/% Culturing of
Habanero Pepper by lnhibition of Ethylene Effects. Hortscience 41 (2), 405-409.
Santana-Buzzy, N., Lopez-Puc, G., Canto-Flick, A„ Barredo-Pool, F., Balam-Uc, E.,
Aviles-Viñas, S., Solis-Marroquín, D., Lecona-Guzman, C., Bello-Bello J., Gomez-
Uc E., Mijangos-Cortes, J.O. (2009) Ontogenesis of the Somatic Embryogenesis
of Habanero Pepper (Capsicum chinense Jacq.). Hortscience 44(1),113-118.
Sato, S., Newel,l C„ Kolacz, K., Tredo, L., Finer, J., Hinchee, M. (1993) Stable
transformation via particle bombardment in two different soybean regeneration
systems Plant Cell Repofts 12,408-413
Steinitz, 8., Küsek, M., Tabib, Y., Paran, 1„ Zelcer, A. (2003) Pepper (C a"m L.)
regenerants obtained by direct somatic embryogenesis fail to develop a shoot. ln
53

CAPITULO 11
Vjtro Cell Dev. Biol. Plant 39, 296-303.
Suhasini, K., Sagare, A.P., Krishnamurthy, K.V. (1994) Direct somatjc embryogenesís from
mature embryo axes in chickpea (Cicerarietinum L.). Plant Sci.102,189-194.
Toonen, M.A.J., Hendriks, T., Schmidt, E.D.L., Verhoeve, H.A., Kammen, A., De Vries
S.C. (1994) Descriptjon of somatic-embryoforming single cells in carrot
suspensions cultures employing video cell tracking. Planta 194,565-572.
Valera-Montero L.L., Ochoa-Alejo N. (1992) A novel approach for chili pepper (Capsicum
annuum L) plant regeneration: shoot inducti.on jn rooted hypocotyls. Plant
Scj.84,215-219.
Venkataiah P., Christopher T., Subhash K.(2003) Thjadiazuron induced high frequency
adventitious shoot formati'on and plant regeneratíon in Capsicum annuumL. J
Plant Biotechnol.5,245-50.
Williams, E.G., Maheswaran, G (1986) Somatjc embryogenesís: factors influencing
coordinated behavior of cells as an embryogenic group. Annals of Botany 57,
443462.
Xiao W.. Custard K.D., Roy C. Brown, Lemmon B.E., Harada J.J., Goldberg R.B.,
Fischer R.L.(2006) DNA Methylation ls Critical for Arabidopsis Embryogenesis
and Seed Viability. The Plant Cell.18, 805-814
Yeung, E.C.. Rahman, M.H., Thorpe, T.A. (1996) Comparative development of zygotic and
microspore-derived embryos in Brassi.ca napus L. cv. Topas. 1.
Histodifferentiation. lntl. J. Plant Sci.157,27-39.
Zapata-Castillo, Y. P., Canto-Flick, A., López-Puc, G., Solís-Ruiz, A., Barahona-Pérez, F.,
Santana-Buzzy, N. (2007) Somatic Embryogenesis in Habanero Pepper (C.
ch/.nense Jacq.) From Cell Suspensions. Hortscience 42 (2),1-5.
Zuo, J., Niu, Q.W., Frugis, G., Chua, N.H. (2002) The WUSCHEL gene promotes
vegetatjve-to-embryonic transition in Arabidopsis. The plant Journal 30(3), 349-
359.
54

CApiTULO 111
CApiTULO 111
ANÁLISIS MORFOLÓGICOS E HISTOLÓGICO DE LOS EMBRIONESSOMÁTICOS DE CHILE HABANERO Caps/'cum ch/.nense
3.1 lNTRODUcclóN
Luego del establecimiento de la planta en condiciones asépticas, existe una etapa de
multiplicación, en esta fase los mecanismos por los cuales se producen las nuevas
plantas son muy variados; de am que se requiera de un estudio histológico con el fin de
comprender los procesos morfogénicos que están ocurriendo (Maeda y Thorpe, 1979;
WiHiams y Maheswaran,1986; Lee eí a/.,1997, Lane ef a/.,1998).
En la actualidad existen pocos reportes de estudios histológicos durante la ES en el
género Caps/.cum, en la especie C. annt/um, los dos únicos repor[es realizados por Harini
y Sita (1993) y Binzel ef a/. (1996) no .incluyen un estudio histológico claro y detallado.
Con respecto a la especie C. ch/.nense solamente existe un reporle realizado por Santana
eí a/. (2009) en el que se describe a detalle la ES en diferentes tipos de explantes (hoja
cotiledonar, embrión cigótico germinado e hipocotilo) y a diferencia de los reportes
anteriores se realiza un análisis durante toda la histogénesis y se observan los ESs de
manera dependiente durante todos sus estadios (globular, acorazonado, torpedo y
cotiledonar).
La formación de los meristemos apicales es uno de los principales eventos en la ES. La
calidad en los embriones somáticos y su capacidad para germinar con éxito depende del
desarrollo apropiado de los meristemos. Nickle y Yeung (1993) trabajando con Daucus
carofa realizaron una comparación entre los meristemos de ESs que no logran la
conversión a plantas y los que realizan una conversión exitosa. En los ESs que no logran
convertirse a plantas, Ias células de sus ápices muestran pronunciada vacuolación, estos
embriones no son capaces de formar el meristemo. En cambio, los embriones que se
convierten exitosamente se caracterizan por mantener en los ápices células con núcleo y
citoplasma denso.
55

CAPÍTULO 111
3.2 METODOLOGÍA
3.2.1 ANÁLISIS MORFOLÓGICOS E HISTOLÓGICO DE LOS EMBRIONES
SOMÁTICOS
Con la finalidad de determinar el desarrollo de los embriones somáticos, se tomaron
muestras proveni'entes de los diferentes tratamientos evaluados en el capítulo anterior y
se realizaron los correspondjentes análisis morfológicos e histológicos de estos embriones
embebidos en resina y en parafina. De igual manera, se reali'zó una clasificación
morfológica de las diferentes deformaciones encontradas. El procedimiento que se llevó a
cabo se describe a continuación.
3.2.1.1 Preparación de las muestras para incluir en resina
Las muestras fueron fijadas siguiendo el protocolo de Berlin y Miksche (1976), en solución
FAA por 24 h. (5 mL de etanol al 96°/o, 10ml de formaldehido al 37%, 5ml de ácido
acético, 80 mL de agua destilada). Después de fiiadas, Ias muestras fueron deshidratadas
gradualmente en diferentes concentraciones de etanol (30, 50, 70, 85, 96 y 100%).
Posteriormente se realizó la infiltración y embebido de las muestras en re§inas plásticas
(JB-4 Glycol Methacrilate, Polysciences). Se realizaron secciones seriadas de 5 um
utilizando un micrótomo de rotación HM340E. Los cortes fueron fijados en portaobjetos y
teñidos con azul de toluidina, utilizando el procedimiento de Sakaí (1973), y
posteriormente se montaron en solución de Poly-mount (Polysciences). Las
observaciones y los registros fotográficos fueron realizadas utilizando un microscopio
Axioplan. En la Fig. 3.1 se esquematiza el procedimiento realizado para llevar a cabo el
análjsís histológico.
56

CApiTULO 111
3.2.1.2 Preparación de las muestras para incluir en parafina
Los tejidos fueron fijados en solución FM por 24 h. (5ml de etanol al 96%, 10ml de
formaldehido al 37%, 5 mL de ácido acético, y soml de agua destilada). Después de
fijadas, las muestras fueron deshidratadas gradualmente en diferentes concentraciones
de etanol (30, 50, 70, 85, 96%) y butanol al 100°/o. En este último se realizaron dos
cambios, cada 24 horas. en el segundo cambio se le adicionaron 15 hojuelas de parafina
y se colocaron en agitación toda la noche a temperatura ambiente (40-50 °C).
Posteriormente, las muestras se colocaron en una estufa a 60 °C, ya derretida la
parafina se agregaron más hojuelas cada 2-3 hrs hasta las 24 h. Para la inclusión la
muestras se colocaron en moldes de aluminio y se cubrieron con parafina liquida. Se
realizaron secciones seriadas de 5 um utilizando un micrótomo de rotación HM340E. Los
cortes fueron fijados en portaobjetos y teñidos con ácido periódico-Schiff (APS) + Azul-
Negro de Naftol . Las observaciones y los registros fotográficos fueron realizadas
utilizando un microscopio Axioplan.
3.3 RESULTADOS
3.3.1 CAFUCTERIZACIÓN MORFOLÓGICA DE ESS DE CHILE HABANERO
Se realizó una clasificación de las diferentes defomaciones encontradas en los
embriones somáticos de chile habanero (Figura 3.1 ).
Figura 3.1 Caracterización morfológica de ESs defomados de chile habanero.
Cot.iledones fusionados (a,b); Copa (c,d); Cot.iledones múltiples (e); Haces
vasculares expuestos con cotiledones fus.ionados (f); Haces vasculares
expuestos (g).(la barra corresponde a 5.0 mm)
57

CAPITULO 111
De acuerdo a la clasjficación anterior se puede constatar que morfológicamente las
deformaciones se observan mayormente en el meristemo caulinar.
3.3.2 CARACTERIZACIÓN HISTOLÓGICA DE ESs MADUROS DE CHILE HABANERO
Se realizaron los análisis histológicos en embriones cigóticos y somáticos de chile
habanero para comparar la morfología de é§tos. En el embrión cigótico (Figura 3.2) se
pudo observar la conformación de ambos meristemos caulinar (Figuras 3.2 b y c) y radical
(Figura 3.2e). De igual manera se pudíeron observar los tejidos vasculares (Figura 3.2d).
'JFigura 3.2 Histología del embrión cigótico (EC) de chile habanero en parafina:
a) Embrión cigóti.co completo, b) Corte hi.stológico del EC completo, c y d)
meri.stemo caulinar, e) tejidos vasculares, q meristemo radical. MC (meristemo
caulinar), ZV (zona vascular del embrión), MR (meristemo radicular).
Se realizó la histología de diversos embri.ones somáticos maduros de chile habanero que
presentaron anormalidades en su conversión a plántulas (Figuras 3.3; 3.4; 3.5).
Todos los embriones desarrollaron elementos vasculares. Esto sugiere que son
estructuras independientes del tejido materno (explante). Los haces vasculares formados
58
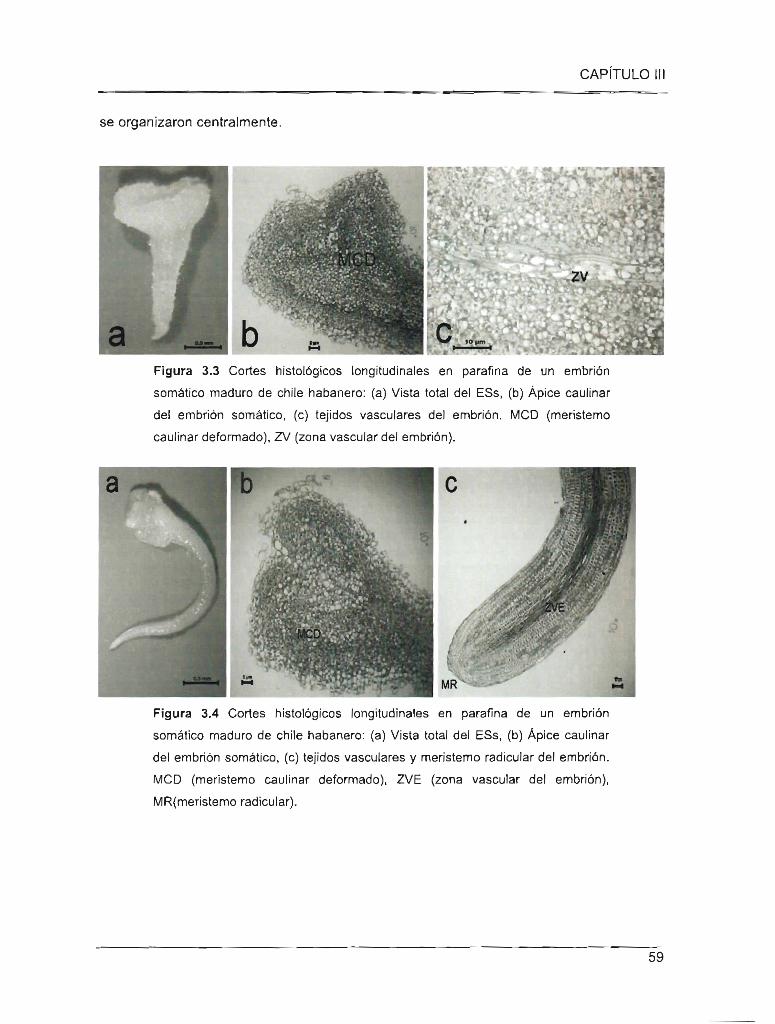
CAPÍTULO 111
se organizaron centralmente.
.t `-: : ,-.-,- vZN ¡ ---
• ,á ,^-, '`,. ':.#;
Figura 3.3 Cortes histológicos longitudinales en parafina de un embrión
somático maduro de chile habanero: (a) V.ista total del ESs, (b) Ápice caulinar
del embrión somático, (c) tejidos vasculares del embrión. MCD (meristemo
caulinar deformado), ZV (zona vascular del embrión).
Figura 3.4 Cortes histológicos longitudinales en parafina de un embrión
somático maduro de chile habanero: (a) Vista total del ESs, (b) Ápice caulinar
del embrión somático, (c) tejidos vasculares y meristemo radicular del embrión.
MCD (meristemo caulinar deformado), ZVE (zona vascular del embrión),
MR(meristemo radicular).

CAPÍTULO IU
Figura 3.5 Cortes histológicos longitudinales en parafina de un embri.ón
somático maduro de chile habanero: (a) Vista total del ESs, (b) Ápice caulinar
del embrión somáti.co. MCD (meristemo caulinar deformado), Z\/E (zona
vascular del embrión).
Se realizaron cortes histológicos de embriones somáticos fusi`onados, observándose
fusi.ones de meristemos caulinares (Figura 3.6) como fusión de meristemo caulinar con
epidermis (Figura 3.7)
Figura 3.6 Cortes hi.stológicos longitudinales de embriones somáticos
fusionados de chile habanero: (a) Vjsta total del ESs, (b y c) Ápice caulinar del
embrión somático. MCF (meristemos caulinares fusionados).
60
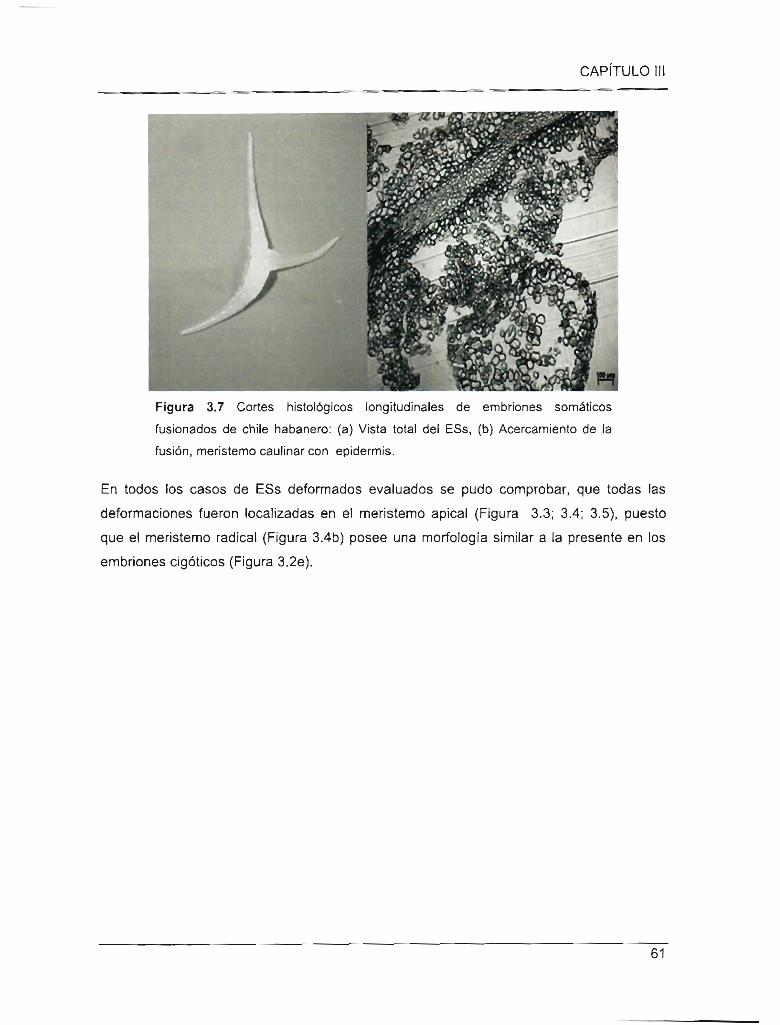
CApiTULO 111
Figura 3.7 Cortes histológicos longitudinales de embriones somáticos
fusionados de chile habanero: (a) Vista total del ESs, (b) Acercamiento de la
fusión, meristemo caulinar con epidermis.
En todos los casos de ESs defomados evaluados se pudo comprobar, que todas las
deformaciones fueron localizadas en el meristemo apical (Figura 3.3; 3.4; 3.5), puesto
que el meristemo radical (Figura 3.4b) posee una morfología similar a la presente en los
embriones cigóticos (Figura 3.2e).
61

CApl'TULO 111
3.4 DISCUSIÓN
A pesar de contar actualmente con protocolos de regeneración de plantas de chile vía ES
(Harini y Lakshmi Sita,1993; Binzel ef a/.,1996; Buyukalaca y Mavituna,1996; Jo ef a/,
1996; Kintzios ef a/., 2001; Steinitz ef a/, 2003; López-Puc ef a/., 2006; Zapata-Castm ef
a/., 2007), al igual que los datos reportados en el presente trabajo (Figuras 3.4 a 3.7),
toda§ Ias especies de este género en el que se ha reportado la ESs tienen un problema
en común el cual se hace evidente cuando los ESs llegan a estadios avanzados y
muestran una amplia gama de malformacjones. Stejnjtz ef a/. (2003) encontraron que las
deformaciones más frecuen{es de los embriones somáticos se pueden clasificar en tres
categorías:
1) Fusión de embriones (Rodriguez y Wetzstejn,1994; Carraway y Merkle,1997; Benelli
eí a/., 2001 ; Stipp ef a/., 2001 ; Tomaz eí a/., 2001 ).
2) Ausencia de cotiledones, un solo cotiledón ó cotiledones malformados. (Carraway y
Merkle, 1997; Jayasankar ef a/., 2002).
3). Anomalías en la hi.stodiferenciación del meristemo caulinar (Nickle y Yeung, 1993;
Suhasini e£ a/., 1996; Stipp ef a/., 2001; Jayasankar ef a/., 2002). Las causas que
provocan estos trastornos en el desarrollo normal de los embriones son todavía
desconocidas.
Se sabe que la recalcitrancia es la incapacidad de las células vegetales, tejidos u Órganos
a responder al cultívo /n vÍ.íro en cualquiera de sus etapas (Benson, 2000). De acuerdo
con los resultados obtenidos en ese trabajo, se muestra una marcada recalcitrancía
durante la histodiferenciación de los ESs, la cual se ve reflejada en las malformaciones
(Figura 3.1) que presentan los ESs en estadios avanzados del desarrollo.
La identidad de los meris{emos y cotiledones se reconoce ya en el dominio apical de un
embrión de 16 células de Arab/.dops/.s (Mayer ef a/.,1998; Schoof el a/., 2000; Schrick y
Laux, 2001). La diferencjación del meristemo caulinar comienza generalmente en la etapa
globular tardía y el meristemo estructuralmente organizado es vi.sible en el estadio
corazón del embrión somático (Thorpe y StasoHa, 2001 ). Mutanles de Araó/.dops/.s con
62

CAPÍTULO 111
cotiledones aberrantes y meristemos apicales anormales han sido aislados (Mayer ef a/.,
1998; Hobbie ef a/., 2000; Budziszewski eí a/., 2001) y durante su manipulación /.n v/tnJ
hanproporcionadoevidenciadequelaauxinaysutransportepolarestánimplicadosenel
desarrollo del meristemo (Schiavone y Cooke,1987; Liu eí a/.,1993; Hadfi ef a/.,1998;
Fischer-lglesias ef a/., 2001). Por lo tanto, una posible causa de las malformaciones
presentes en los meristemos caulinares de los ESs de chile habanero es la dependencia
de la auxina sintética 2,4-D, a la cual están expuestos los ESs durante todo su desarrollo.
Santos ef a/. (1997) trabajaron en el proceso de ES de soya y reportaron la presencia de
ESs con anormalidades en el meristemo caulinar, pero con un procambium bien definido;
dichos embriones se describen con una morfología sim.ilar a las trompetas. En el presente
estudios se observaron ESs con esas mismas características reportadas (Figura 3.4).
Durante el desarrollo de este trabajo se constató que las anomalías encontradas poseen
un factor en común, que es la morfología anormal del meristemo caulinar (Figura 3.2). Es
probable que esto sea el resultado no solamente de las auxinas, pues también pueden
influir factores ambientales, otros RCV de la planta y el estado de desarrollo de ésta. EI
conocimiento de las complejidades de estas interacciones será importante para conseguir
un mejor entendimiento del papel de las aux.mas durante el desarrollo del embrión.
63

CAPÍTULO 111
3.5 CONCLUSIONES
Se pudo constatar mediante el análisis histológico, que en todos los casos los ESs
desarrollaron elementos vasculares y que estos se organizaron centralmente.
En todos los ESs caracteri'zados histológi'camente se observó la formación del meristemo
radicular, el cual se encontró bien definido y sin deformaci.ones.
En todos los casos de ESs deformados evaluados se pudo comprobar que todas las
deformaciones fueron localizadas en el meristemo apical.
64

CAPÍTULO 111
BIBLloGRAFIA
Benelw C., Fabbri A., Grassi S., Lambardi M. and Rugini E. (2001). Histology of somat.ic
embryogenesis in mature tissue of olive (0. et/ropaea L.). J. Hort. Sci. Biotechnol.
76:112-119.
Benson, E. (2000). Special Symposium: /n vffm plant recalcitrance, /n v/.fro plant
recalcitrance: An introduction. /n v/.fro Cell. Dev. Biol.-Plant 36:141-148.
Bernn, G.P. and J.P. Miksche. (1976). Botanical microtechnique and cytochemistry. 3rd
Ed. lowa State University Press, Ames, IA.
Binzel M., Sankhla N., Joshi S. and Sankhla D. (1996). lnduction of direct somatic
embryogenesis and plant regeneration in pepper (C. annt/um L.). Plant CeH Rep.
15:536-540.
Budziszewski, G. J.; Lewis, P. S.; Glover, L. W.; Reineke, J.; Jone§, G.; Ziemnik, L. S.:
Lonowski, J.; Nyfeler, 8.; Aux, G.; Zhou, Q.; MCElver, J.; Patton, D. A.;
Martienssen, R.; Grossniklaus, U.; Ma, H.; Law, M.; Levin, J. Z.(2001) Arabidopsis
genes essential for seedling viability: isolation of insertional mutants and
molecular cloning. Genetics 159:1765-1778.
Buyukalaca, S. and Mavituna, F. (1996), Somatic Embryogenesis and plant regeneration
of pepper in liquid media. Plant Cell, Tiss. Org. Cult. 46: 227-235.
Carraway D. y Merkle S., (1997). Plantlet regeneration from somatic embryos of American
chestnut. Can. J. For. Res. 27:1805-1812.
F.ischer-lglesias, C.; Sundberg, 8.; Neuhaus, G.; Jones, A. M. (2001 )Auxin distribution and
transponduringembryonicpatternformationinwheat.PlantJ.26:115-129.
Hadfi, K.; Speth, V.; Neuhaus, G.(1998) Auxin-induced developmental pattems in Brassica
juncea embryos. Developmer`t 125:879-889.
Harini 1. and Sita L. (1993). Direct somatic embryogenesis and plant regeneration from
immature embryos of chim (C. annuum L). Plant Sci. 89:107-112.
65

CAPíTULO 111
Hobbie, L MCGovern, M.; Hurwi.tz, L. R.; Pi.erro, A.; Liu, N. Y.; Bandyopadhyay, A.;
Estelle, M. (2000) The axr6 mutants of Arabidopsi.s thaliana define a gene
involved in auxin response and early development. Development 127:23-32
Jayasankar S.. Bondada 8., Li Z. and Gray D. (2002). A unique morphotype of grapevine
somatic embryos exhibits accelerated germínation and early plant development.
Plant Cell Rep. 20:907-911.
Jo J., Choi E., Choi D. and Lee, K. (1996). Somatic embryogenesis and plant regeneration
from Ímmature zygotic embryo culture in pepper (C. annuum L.). J. Plant. Biol.
39: 127-135.
Kintzios S., Drossopolous J. and Lymperopoulos C. (2001). Effect of vitamíns and
inorganic micronutrients on callus growth and somatic embryogenesis from leaves
of chilli pepper. Plant Cell Tiss. Org. Cult. 67:55-62.
Lane W.D., lketani H., Hayashi T. 1998. Shoot regeneration from cultured leaves of
japanese pear (Pyrus pyr/.Ío/Í.a). Plant Cell, Tissue and Organ Culture 54: 9-14.
Lee K.S., Zapata-Arias F.J., Brunner H., Afza R. 1997. Histology of somatic embryo
initíation and organogenesis from rhyzome explants of Musa spp. Plan{ Cell,
Tjssue and Organ Culture 51 : 1-8.
Uu, C.-M.; Xu, Z.-H.; Chua, N.-H. (1993) Auxjn polar transpori is essential for the
establishment of bilateral symmetry during early plant development. Plant Cell
5:621-630.
López-Puc G., Canto-Fli.ck A., Barredo-Pool F., Zapata-Castillo P., Montalvo-Peniche M.,
Barahona-Pérez F. and Santana-Buzzy N. (2006). Direct Somatic
Embryogenesi's: A mghly Efficient Protocol for /n v/.(ro Regeneration of Habanero
Pepper (Caps/.cum ch/.nense Jacq.) Hortscience 41 (6) 1 -7.
Maeda E., Thorpe T.A.1979. Shoot histogenesis in tobacco callus cultures. /n v/tno 15(6):
415424.
Mayer K. F., Schoof H., Haecker A., Lenhard M., Jurgens G. and Laux T. (1998). Role of
66

CAPÍTULO 111
Wusche/ in regulating stem cell fate in Arabí.c/ops/.s shoot meristem. Cell, 95, 805-
815.
Nickle C. y Yeung E. (1993). Failure to establish a functional shoot meristem may be a
cause of conversion failure in somatic embryos of D,carofa. Am. J. Bot. 80:128-
1291.
Rodríguez A. y Wetzstein H. (1994). The effect of auxin type and concentration on pecan
(C. /.///.noí.nens/.s) somatic embryo morphology and subsequent conversion into
plants. Plant Cell Reports 13:607-611.
Sakai W. (1973). Simple method for differential staining of paraffln embedded plant
material using toluidine blue 0. Stain Technol. 48: 247:249.
Santana-Buzzy N., Guadalupe Lopez-Puc, Adriana Canto-Flick, Felipe Barredo-Pool,
Eduardo Balam-Uc, Susana Aviles-Viñas, Daniela Solís-Marroquín, Carlos
Lecona-Guzman, Jerico Jabin Bello-Bello, Eunice Gomez-Uc and Javier 0.
Mijangos-Cortes (2009) Ontogenesis of the Somatic Embryogenesis of Habanero
Pepper (Caps/.cum ch/.nense Jacq.)Hortscience 44(1 ):113-118.
Santos K., Mudstock E. and Bodanese-Zanettini M. (1997). Genotype-specific
normalization of soybean somatic embryogenesis through the use of an ethylene
inhibitor. Plant Cell Rep.16: 859-864
Schiavone, F. M.; Cooke, T. J.(1987) Unusual patterns of somatic embryogenesis in the
domesticated carrot: developmental effects of exogenous auxins and auxin
transport inhibitors. Cell Diff. 21 :53-62.
Schoof, H., Lenhard, M., Haecker, A., Mayer, K.F.X., Ju" rgens, G. and Laux, T. (2000)
The stem cell population of Arabidopsis shoot meristems is maintained by a
regulatory loop between the CLAVArA and WUSCHEL genes.Cell, 100, 635-
644.
Schrick, K.; Laux, T.(2001 ) Zygotic embryogenesis: developmental genetics. ln: Bhojwani,
S. S.; Soh, W. Y., eds. Current trends in the embryology of angiosperms.
Dordrecht: Kluwer Academic Publishers; :249-277.
67

CAPÍTULO 111
Steinitz 8., Küsek M., Tabib Y., Paran 1. and Zelcer A. (2003). Pepper (C annuum L.)
regenerants obtained by direct somatic embryogenesis fail to develop a shoot. /n
v/.fro Cell Dev. Biol. Plant. 39: 296-303.
Stipp L., Mendes 8„ Piedad S. and Rodríguez A. (2001). /n v/.Íro morphogenesis of C.
me/on var. inodorus. Plant Cell Tiss. Org. Cult. 65:81-89.
Suhasini K., Sagare A. and Krishnamurthy K., (1996). Study of aberrant morphologies and
lack of conversion of somatic embryos of chickpea (C.ar/.ef/.num L.). /n v/.Íro Cell.
Dev. Biol. Plant 32:6-10.
Thorpe, T. A.; Stasolla, C.(2001) Somatic embryogenesis. ln: Bhojwani, S. S.; Soh, W. Y.,
eds. Current trends in the embryology of angiosperms. Dordrecht: Kluwer
Academic Publishers; 279-336.
Tomaz M., Mendes 8., Filho F„ Demetrio C., Jansakul N. and Rodriguez A. (2001).
Somatic embryogenesis in Citrus spp.: Carbohydrate stimulation and
histodifferentiation. /n v/.fno Cell. Dev. Biol. Plant 37:446J52.
Wílliams E.G., Maheswaran g. 1986. Somatic embryogenesis: factors influencing
coordinated behavior of cells as an embryogenic group. Annals of Botany 57:
443-462
Zapata-Castillo Y. P., Canto-Flick A., López-Puc G., Solís-Ruiz A., Barahona-Pérez F. and
Santana-Buzzy N. (2007). Somatic Embryogenesis in Habanero Pepper (C.
chí.nense Jacq.) From Cell Suspensions. Hortscience 42 (2):1-5.
68

CAPITULO IV
CApiTULO IV
EVALUACIÓN DE LA EXPRESIÓN DE UN GEN TIPO WUSCHE[ DURANTE LAEIVIBRloGÉNESIS SOIVIÁTICA DEL CHILE HABANERO
4.1 lNTRODUCCIÓN
El gen Wusche/ /WUS) es un gen homeótico, el cual codifica un factor transcripcional
(Wt/S). Este gen tiene una función .importante en la regulación celular durante la
formación del meristemo embrionario, manteniendo desdiferenciada§ las células del
meristemo (Laux e£ a/.,1996; Mayer e{ a/.,1998). WUS fue reportado primeramente en
Arabidopsis thaliana y poster.iormente en o\ras plar\\as corno Petunia hibrida, Solanum
/í.copers/.cm Aní/.rrhí.num ma/.us y más recientemente en Zea mays (Nardmann ef a/.,
2006).
WUS fue identificado por medio de mutantes que presentaban defectos en el meristemo
apical del tallo (Shoof Ap/'ca/ Mer/.sfem, SAM) y en las cuales la proteína estaba ausente
(Laux eí a/., 1996). En ese mismo trabajo se observó que también se perdían órganos
centrales en las flores. Con base en esos experimentos se determinó que WUS promovia
la identidad central de SAM y del mer.istemo floral, manteniendo de esta forma íntegra la
función y la estructura de estos tejidos.
Se ha observado que la sobreexpresión de WUS promueve la transición del estado
vegetativo al estado embrionario en explantes de Arab/.dopsí.s, lo cual provee un modelo
para el estudio de muchos aspectos de la biología de las plantas (Zuo ef a/., 2002). En
trabajos previos, Solís ef a/. (2010) sobreexpresaron el gen WUS de Arab/.dops/.s en el
chile habanero y determinaron que en las plantas transgénicas aumentaba la capacidad
morfogénica. De igual manera, se ha comprobado que la correcta expresión de WL/S,
regulada por el nivel crítico de auxinas exógenas, es esencial para la inducción de la ES
(Su ef a/., 2009).
Con base en estos antecedentes que demuestran un papel importante de WUS en el
mantenimiento de la identidad de SAM, y en el hecho de que en nuestro trabajo se
encontraron anormalidades en este tejido, decidimos investigar la expresión de Wl/S
durante la ES del chile habanero.
69

CAPÍTULO IV
4.2 METODOLOGÍA
4.2.1 EVALUAclóN DE LA EXPRESIÓN DE UN GEN TIPO WUS EN ESs DE CHU
HABANERO
Este análisis se llevó a cabo para determjnar el nivel de expresjón de un gen tipo WUS
en embriones somáticos en diferentes estadios de chjle habanero. Para ello, primero se
clonó un fragmen(o del gen homólogo de Capsí.cum chí.nense y posteriormente se analizó
su expresión a lo largo del proceso, medjante la metodología de RT-PCR de punto final.
4.2.2 EXTRACclóN DEL ARN
Se llevó a cabo con la mezcla de fenol: isotiocianato de guanidina, si.guiendo el
procedimíento reportado por Chomczynski y Saccm (1987). Todos los pasos fueron
desarrollados por centrifugaci.ón a 13,000 rpm en un rotor Sorvau #3328 (Sorvall® fresco).
Los ESs (1g) fueron macerados en nitrógeno líquido, a los cuales se les agregó 5 mL de
Trizol (manufacturado en el laboratorio) (38 °/o v/v de Fenol, 12.5 mM de isotiocianato de
guanidina ultrapura, 6.25 mM de tiocianato de amonio,100 mM de acetato de sodio y 5 °/o
de glicerol. Las muestras se incubaron por 10 min a temperatura ambiente. Después de
esta jncubación, las muestras se centrifugaron a 13,000 rpm por 15 min a 4 °C y el
sobrenadante se depositó en un tubo nuevo al cual se le agregó 1/5 de volumen de una
mezcla de cloroformo:alcohol isoamílico (49:1 v/v), se agitaron con vórtex y se incubaron
por 3 min a temperatura ambiente. Se procedi.ó a centrifugar las muestras a 13,000 rpm
por 15 mín a 4 ac para la separación de fases, la fase acuosa (superi.or) se transfirió a un
tubo nuevo, se midió el volumen extraído y se agregó un volumen Ígual de i.sopropanol
fri.o, se incubaron las muestras a 4 °C toda la noche, posteriormente se centrifugaron por
15 min a 4 °C. Las pastillas fueron lavadas con etanol al 70 % frío y cen{rifugadas a
13,000 rpm por 15 min a 4 °C. Este paso se repitió tres veces, con la excepción de que
para el último se empleó etanol al 100% frio. Se eliminó el etanol, y las pastillas se
dejaron secar a temperatura ambiente aproxjmadamente 1 hora y posteriormente se
resuspendíeron en 35 uL de agua Sigma grado bjología molecular tratada con
djetilpirocarbonato (DEPC) y se almacenaron a -80 °C.
70

CApiTULO IV
4.2.3 TRATAMIENTO DEL ARN CON DNasa 1
Durante el análisis de las poblaciones de ARN mensajeros se eliminó el ADN
cromosómico para ev.itar la presencia de resultados falsos positivos, empleando el
protocolo comercial de la casa comercial Ambióm cuya mezcla de reacción se describe el
cuadro 4.1.
Cuadro 4.1 Mezcla de reacción de DNasa 1
Componentes Cantidad
0.5 M Tris-Hcl (pH 7.5) 1 uL
O.5 M Mgc12 1 LiL
H20 estéril 22 HL
DNasa l ( 1 unidad/uL) 1LiL
Volumen final 25 uL
En este protocolo se agrega 0.1 volumen del amortiguador lox de la DNasa 1 y 1 uL de la
enzima DNasa 1 (2 unidades), se mezcla suavemente y se incuba a 37 °C por 20-30 min,
se le agrega 0.1 volumen del reactivo de inactivación de la DNasa 1, se mezcla e 'incuba el
tubo 2 min a TA, por último se centrifuga el tubo por 1-2 min a 13,000 rpm a 4° C.
4.2.4 CUANTIFICACIÓN Y COMPROBAclóN DE LA INTEGRIDAD DEL ARN TOTAL
La cuantificación del ARN total se realizó por medio del espectrofotómetro Nanodrop®.
Para ver.ificar la integridad del ARN, se fraccionó una alícuota mediante electroforesis
nativa en un gel de agarosa al 1.4 %, este ARN se mezcló con amortiguador de carga que
contenía bromuro de etidio en concentración de 1 ug.mL-' .
4.2.5 SÍNTESIS DE ADNc: TRANSCRIPCIÓN REVERSA DEL ARN
Para la síntesis de ADNc fue utilizada la enzima SuperscriptTM 111 Reverse Transcriptase
(lnvitrogen).
71

CAPÍTULO IV
El procedimiento fue el siguíente: se mezclaron 1 LiL de ARN (1 Hg uL-`), 1 uL de Olígo-dT,
1 LJL de dNTP 10 uM y agua hasta un volumen final de 13 uL. La mezcla se incubó a 65°C por 5 min y posteriormente se incubó en hi.elo por 2 min. Seguidamente se le adi.cionó
4 uL del amortiguador 5X first strand, 1 uL de DTT 0.1 M y 1 iJL de la transcriptasa
reversa y se incubó a 50 °C por 60 min, y a 70 ac por 15 min (para inactivar a la enzima).
A continuación, para verificar la integridad fue fraccionada una alícuota por electroforesis
natjva en un gel de agarosa al 1°/o teñjdo con bromuro de etídio (1 ug mL-')
4.2.6 DISEÑO DE CEBADORES PARA EL GEN TIPO Wt/S
Se realizó el di.seño de cebadores con respecto a las secuencias de WUS reportada para
So/anum //.copers/.um, utiljzando el programa OligoperfectTM Desígner de invitrogen
http://tools.invitroaen.com/content. cfm?Daaeid=97±É
4.2.7 AMPLIFICACIÓN DEL GEN TIPO WS POR RT-PCR DE PUNT0 FINAL
Se diseñaron cinco pares de cebadores específicos para analizar por PCR la presencia de
transcritos del gen tjpo WUS (Cuadro 4.2). El gen ribosomal 18S fue utilizado como un
estándar de cargado, en las muestras colectadas de los ESs de chile habanero.
Los cebadores tienen la sjguiente secuencia:
Cuadro 4.2 Cebadores diseñados para el análisis de transcritos de un gen tipo
WUS en chile habanero.
72

CAPÍTULO IV
Las condiciones programadas en la termocicladora para esta reacción fueron; 50°C por 30
min. (1ciclo); 94 °C, 3 min. (1ciclo);[ 94 °C,15 s; 72 °C,30 s], (35 ciclos) ]; 72 °C 10 min, (1
ciclo), 4 °C ci.
Posteriomente se vermcó la calkkid y la integridad de la ampmcación por electroforesis
nativa en un gel de agarosa a 3%. EI ADN se pudo visualizar en el gel por previa tinción
con una solución de bromuro de etidk).
La secuencia del transcrito 18S empleado como referencia no es proporcionada por
Ambion, pero el tamaño esperado del amplicón es de 315 b (el número de catálogo 1718
y d nombre del kft es Quantum RNAtm 18S intema Standards unM3rsal 18S internal
Standards).
42.8 PURIFICACIÓN DE PCR
Una vez obtenido el producto esperado se procedió a purfficar el producto de PCR por
medio del kft QIAquk* PCR Purfflcation (QUIAGEN)
42.9 ANÁLISIS DE LAS SECUENCIAS
El producto purmcado de PCR se mandó a secuenciar a la empresa Davb Sequencing en
Calffomia. Las secuencias recibidas fueron comparadas en bancos de datos de "National
Center for Biotechnology lnformation" (NCBl) con el programa Blast para su k]entificación
y anáiisis en ia síguiente dirección electrónica: (!±±±i2:±Aaa±ea±:D!±i±nlmi±Ég±±£Be£ip9n±±S
sea.htmL).
73

CApl'TULO IV
4.3 RESULTADOS
4.3.1 EXTRACCIÓN DE ARN DE ESs Y MERISTEMOS DE PLÁNTULAS DE Caps/.cum
¢hinense Jacq.
Como se observa en la Figura 4.1, el ARN fraccionado muestra que el método de la
extracción y preparación fueron adecuados y las subunidades 28 S y 18 S se encuentran
íntegras y no presentan degradación.
En el carril 1 se muestran las pozas de ARN de ESs en estadios iniciales, en el carrn 2 de
lol ESs en estadios maduros, en el ca" 3 de los ESs germinados y en el carrn 4 de los
meristemos de plántulas
1234
Figura 4.1 ARN de ESs y meristemos de plántulas de Capsí.cum chí.nense.
Carril 1: ESs tempranos; carril 2: ESs i.ntermedio§; carrn 3: ESs avanzados;
carril 4: meristemos api.cales del tallo de plántulas.
Una vez extraídos los ARN se procedíó a su cuantificación espectrofotométrica. En el
cuadro 4.3 se muestran los resultados para las muestras en las cuales se obtuvo buena
caljdad.
74

CAPÍTULO IV
Cuadro 4.3 Cuantificación de ARN de ESs y meristemos de plántulas de
Capsicum chinense.
Concentración ARN (ng/i.L)
ESs tempranos
ESs intermedios
ESs avanzados
Meristemos de
plántulas
1036.9
1399.1
3288.9
1009.3
4.3.2 AMPLIFICACIÓN DEL TRANSCRITO DEL GEN TIPO WS DE Caps/.cum
ch/.nense CON CEBADORES HETERÓLOGOS
Una vez extraído el ARN de meristemos de plántulas de chile habanero, se procedió a la
amplificación del transcrito del gen tipo Wt/S (Figura 4.2). De los cinco cebadores
diseñados para el análisis de transcritos del gen tipo WUS, el par denomínado WUS3,
correspondiente al carril 3, fue el único en el cual se obtuvo el amplicón esperado de 109
pb.
12345
109 pb.
Figura 4.2 Amplificación usando los cebadores diseñados para el gen tipo
WUS. Carril 1 : WUS 1 ; carril 2: WUS 2; carril 3: VVUS 3; carril 4: WUS 4; carril 5:
WUS 5.
Una vez obtenido el amplicón deseado con el cebador WUS 3 se procedió a purificar el
producto de PCR mediante el kit QIAquick PCR Purification para su posterior
secuenciación.
75

CAPÍTULO IV
4.3.3 RESULTADO DE LA SECUENCIAclóN
Del producto de PCR purificado enviado para su secuenciación a Davis Sequencing, la
secuencia de nucleótidos (Cuadro 4.4) fue comparada en las bases de datos. Los
resultados mostraron que presentaba altos niveles de identidad con secuencias de
aminoácidos reportadas (GenBank, EMBL, etc.)
Cuadro 4.4 Secuencia de nucleótidos obtenida de la secuenciación, Fragmentc)
de 74 b.
CTM GTCTGAG TCTGTAAG CC ATAGWTCTAA G G CAAAAATG
TGTTTTATTG GTTTCAGAAC CATAAAGCTC GTGA
La comparación de su secuencia en bancos de datos usando el sitio de INTERNET del"National Center for Biotechnology lnformation" (httD://blast.ncbj.nlm.nih.aov/Blast.cgí), arrojó
los resultados que se muestran en el Cuadro 4.5. Como se puede observar, la secuencia
obtenida se alinea exclusivamente con la secuencia de la proteína WUS de diversos
mjembros de la familia de las solanáceas, así como otros miembros de otras familias. Los
valores del nivel de confianza umbral (desde E=4xl0-`` hasta E=7xl0-í4) ofrecen la
certidumbre de que la secuencia obtenida corresponde a un transcrito de un gen tipo
WUS de chile habanero.
76

CApiTULO IV
Cuadro 4.5 Análisis t.ipo BLAST (Basic Local Alignment Search Tool) de la
secuencia deducida de aminoácidos de 74 b.
Se muestra únicamente los resultados de 3 secuencias con mayor identidad.
A continuación se alineó la secuencia obtenida con las tres secuencias reportadas para el
gen WUS de jitomate, petunia y uva (Figura 4.3).
Capslcum chlnenseSolanum lya.perslcumPe"nh hybrktaVqtis v]nlftraConsensus
1 10 20 30 40 50 60 70 74l-+-t-.-..-+.-.------lc"TC"mTmmmGm"GGfflmTGT6TT"T"TTT«GmmTmcTC6"CTm"mfl6"6G""TT6mG6ümmTGTGTTT""GTT"6mm"GCTc6"c"6T"m6"GGRm6R"mG6iRflGmT6T6TT"m6TT"6flücfl""ccmnc"6c"G«fl6"Cücfl""6mGGCRRÜRC6TCTT"T"T"fl6mü"mGC""(Tagg€T¡fl6.tim€Ggcfl.R6aT€gmG6CmamiGTSTT"T"TT"GmclmmGctc6"
Figura 4.3 Alineamiento de la secuencia de aminoácidos obtenidos. La
secuencia deducida de aminoácidos se alineó con secuencias de especies
diferentes tanto de la misma familia como de otras. Los residuos idénticos se
muestran en rojo y los residuos conservados en azul.
77

CAPITULO IV
4.3.4 ANALISIS DE LA EXPREslóN GÉNICA DEL GEN TIPO WUS EN ESs DE CHILE
HABANER0
Al analizar la expresión del gen tipo WUS en ESs de chile habanero en diversos estadi.os
(Fígura 4.4) se observó que su expresión se presentó tanto en estadios tempranos como
en estadios intermedios del desarrollo embrionari.o; sin embargo, en estadios avanzados
del desarrollo no se observó expresi.ón alguna de este gen.
1234
WUS
18,Sb---109 pb.
-315pb.
Figura 4.4 Análi.sis de expresíón del gen tipo WL/S en embriones somáticos de
chile habanero en diferentes estadios de desarrollo. Carril 1: estadi.os
tempranos (globular y corazón; carri.1 2: estadio intermedjo (torpedo); carril 3:
estadio avanzado (cotiledonar); carril 4: testigo positivo (meristemos de
plántulas de chile habanero); carril 5: testigo negativo.
Para determinar la intensidad relativa de las bandas obtenidas se utiljzó el software EDAS
Kodak 290 por medio del cual se pudo constatar que la i.ntensidad de las bandas
obtenidas en los estadios tempranos e intermedíos de ESs de chile habanero no varió
significativamente. En la Figura 4.5 se realjzó una representaci.ón gráfica de la expresión
del gen tipo WUS durante los diferentes estadíos de desarrollo de los ESs.
78

CApiTULO IV
Estad:10StemoTanos
Globulár Coíazón
Expresión deWuS ++
Estadio§
EZIZITorpedo
+
Estadios
Co:,`cdc,,J?
Figura 4.5 Representación gráfica de la expresión del gen tipo WUS durante los
diferentes estadios de desarrollo de los ESs de chile habanero. Los signos +
representan intensidad relativa de la expresión.
Los resultados obtenidos indican que la expresión del gen tipo WUS se ve afectada en las
etapas finales de desarrollo de los ESs. Este comportamiento coincide con la presencia
de las anormalidades presentes en el meristemo caulinar de lo ESs.
79


CAPÍTULO IV
4.4 DISCuslóN
El objetM más .irnportante de la investigación actual sobre la ES es identificar los genes
implicados en la inducción de la competenc.ia embriogénica y el desarrollo posterior del
embrión. En los últimos años se ha logrado un gran avance en el análisis molecular de la
embriogénesis con la aplicación de herramientas moleculares modernas que permiten el
análisis rápido y completo de la expresión génica. El evento más crítico en el desarrom
del embrión somático parece ser la formación de los meristemos (Meinke, 1991). Las
células madre dentro del meristemo permiten a las plantas crecer y producir nuevos
órganos (Scheres, 2007), cuya identidad y proliferación se mantienen en parte por
señales que estas células reciben del medio ambiente local. En el meristemo apical de
Arabidopsis, WUS es un regulador crítico que codifica una proteína homeótica requerida
para la formación y mantenimiento de las células madre (Laux ef a/., 1996). En mutantes
de WUS, las plantas son incapaces de mantener una reserva de células madre
indiferenciadas (Laux eí a/.,1996). La expresión de VVUS se I.imita a un pequeño grupo de
células dentro de la zona central del meristemo caulinar, que se encuentra
inmediatamente debajo de la capa apical de las células madre activas (Mayer ef a/.,1998;
Schoof e{ a/., 2000). En cuanto a la expresión de este gen durante la embriogénesis
cigótica, se sabe que su expresión inicia en el embrión conformado por 16 células y
gradualmente se limita a un pequeño grupo de células debajo de las células madre
durante las etapas embriogénicas y post-embriogénicas (Mayer ef a/.,1998; Schoof ef a/.,
2000). Actualmente se desconoce cómo la expresión de WUS es .inicial y
subsecuentemente regulada durante la embriogénesis (De Smet eí a/., 2010); sin
embargo, se sabe que su expresión es regulada tanto por citocininas (Kwon e{ a/., 2005;
Gordon eí a/., 2009) como por auxinas (Su eí a/., 2009).
La ES requiere auxinas y éstas tienen un papel central en el establecimiento del
meristemo apical. Por otro lado, WUS es fundamental para la determinación del dest.ino
de las células madre en el meristemo caulinar de las plantas superiores. Sin embargo, Ia
regulación de la expresión de Wl/S por la auxina durante la embriogénesis somática es
poco conocida. Durante la realización de este estudio se pudo corroborar los resultados
reportados por Su eí a/. (2009), quienes trabajando con el proceso de ES en Arab/.dopsí.s
determinaron que la expresión de WL/S se inducía desde etapas tempranas de la ES (16-
81

CAPITULO IV
24hposterioresalainducción),cuandolosESsnopuedenseridentificadosnimorfológica
ni molecularmente y durante los subsecuentes estadios de desarrollo de los ESs. En chjle
habanero se determi'nó la expresión de este gen desde estadios globulares (Figura 4.4);
no obstante, a diferencia de lo que sucede en Arab/.dops/.s, no se detectó su expresíón en
estadios avanzados del desarrollo embrionario (cotjledonar maduro). Los resultados
anteriores podrían tener djferentes explicaci.ones. Por un lado, es posible que la expresión
de este gen esté afectada por las mismas causas que originan las deformaciones
presentes en lo meristemos caulinares de los ESs del chile habanero, pero también es
posible que estas anomalidades sean resultado de la expresión anómala de WUS. En
cualquiera de estos dos escenarios, el presente trabajo aporta evidencias experimentales
de que los patrones de expresión del gen WUS pueden tener un papel directo o indirecto
en el desarrollo anómalo del meristemo caulinar de los embríones de chile habanero. Esta
hipótesis deberá ser confrontada en estudios posteriores,

CAPÍTULO IV
4.5 CONCLUSIONES
Seaislóunasecuenciaparcialdechilehabaneroquepresentaunaaltahomologíaconlas
secuenciasreportadasparaelgenWUSenotrasespeciesporlotanto,existecerlidumbre
de que los estudios de su expresión pueden constituir evidencias para conocer la
regulación de este gen durante el proceso de ES.
La caracterización de la expresión del gen tipo WUS en diversos estadios de desarrollo de
ESs de chile habanero demostró que los transcritos son detectados en estadios
tempranos, así como en estadios intermedios de desarrollo, pero no se detectaron en
estadios avanzados de desarrollo de los ESs.
83

CAPÍTUL0 lv
BIBLIOGFUFÍA
Chomczynski, P., and Sacchi, N. (1987) Single-step method of RNA isolation by acid
guanidinium thi.ocyana{e-phenol-chloroform extraction Ana/. B/.ochem. 762..156-
159.
De Smet 1., Lau S., Mayer U. and Jürgens G., (2010) Embryogenesis -the humble
beginnings of plant life. The Plant Journal 61, 959-970
Gordon, S.P., Chickarmane, V.S., Ohno, C. and Meyerowitz. E.M. (2009) Multiple
feedback loops through cytokínin signaling control stem cell number wi.thi'n the
Arabidopsis shoot meristem. Proc. Natl Acad. Sci.. USA,106,16529-16534.
Kwon, C.S., Chen, C. and Wagner, D. (2005) WUSCHEl is a primary target for
transcriptional regulation by SPLAYED in dynamic control of stem cell fate in
Arabidopsis. Genes Dev.19, 992-1003.
Laux T., Mayer K. F., Jurgens G. and Berger J. (1996). The Wusche/ gene is required for
shoot and floral meristem integri.ty in Arabí.dops/.s. Development,122, 87-96.
Laux, T., Mayer, K.F.X., Berger, J. and Ju" rgens, G. (1996) The WUSCHEL gene is
required for shoot and floral meristem integrity in Arabídopsis. Development, 122,
87-96.
Mayer K. F., Schoof H., Haecker A., Lenhard M., Jurgens G. and Laux T. (1998). Role of
Wusche/ in regulating stem ceH fate in Araóí.dops/.s shoot meristem. Cell, 95, 805-
815.
Meinke, D.W. (1991) Perspectives on genetic analysis of plant embryogenesis. Plant Cell,
3, 857-866.
Nardmann J. and Werr W. (2006). The shoot stem cell niche in angiosperms: expressi.on
patterns of WUS orthologues in rice and maize imply major modífications in the
course of mono- and djcot evolution. Molecular BÍology and Evolutjon, 23, 2492-
2504.
84

CAPÍTULO IV
Scheres, 8. (2007) Stem-cell niches: nursery rhymes across kingdoms. Nat. Rev. Mol. Cell
Biol. 8, 345-354.
Schoof, H„ Lenhard, M., Haecker, A„ Mayer, K.F.X., Ju" rgens, G. and Laux, T. (2000)
The stem cell population of Arabidopsis shoot meristems is maintained by a
regulatory loop between the CLAVA" and WUSCHEL genes.Cell, 100, 635-
644.
Solís-Ramos L., Nahuath-Dzib S., Andrade-Torres A., Barredo-Pool F.,González-Estrada
T., De la Serna E., (2010) lndirect somatic embryogenesis and morphohistological
analysis in Caps/.cum chinense. Biologia 65:504-511
Su Ying H. , Xiang Y. Zhao, Yu 8. Liu, Chuan L. Zhang, Sharman D. O'Neill2 and Xian S.
Zhang (2009) Auxin-induced WUS expression is essential for embryonic stem cell
renewal during somatic embryogenesis in Arabí.dops/.s. The Plant Joumal , 59,
448460.
Zuo J., Niu Q.W., Frugis G. and Chua N.H. (2002). The WUSCHEL gene promotes
vegetative-to-embryonic transition in Arab/.dops/.s. The plant Journal, 30(3), 349-
359.
85


CONCLUSIONES
CONCLUSI0NES GENERALES
En este estudio, se pudo apreciar que la reducción de la concentración de la auxina (2,4-
D) a la mítad de su concentración y el cambio a medio líquido favoreció el desarrollo y la
sincronía de los embriones somáticos, con un rendimiento de 1.77X104 embriones
somáticos por litro de medio, con un 58.7 ± 5,3°/o de embriones en el estadio cotiledonar.
Los resultados obtenidos durante el desarrollo de este protocolo de embriogénesis
somática en medio líquido para chile habanero muestran la dependencia al 2,4-D durante
la inducción y la histodiferenciación de los embriones somáticos de esta especie, contrario
a lo reportado para la mayoría de las especies vegetales.
La caracterización histológica de los embriones somáticos que se realizó permitió
constatar que en todos ellos se desarrollaron elementos vasculares, los cuales se
organizaron centralmente. De igual manera, se comprobó que las deformaciones
presentes en los embriones somáticos se encuentran localizadas en el meristemo
caulinar,
La caracterización de la expresión del gen tipo WUS en diversos estadios de desarrollo de
ESs de chile habanero demostró que los transcritos son detectados en estadios
tempranos, así como en estadios intermedios de desarrollo, pero no se detectaron en
estadios avanzados de desarrollo de los ESs. Este tipo de comportamiento sugiere una
relación entre la ausencia de la expresión de este gen y las anormalidades presentes en
el meristemo caulinar de los ESs.
87


PERSPECTIVAS
PERSPECTIVAS
La regeneración es un requisito ineludible y común a cualquiera de las aplicaciones de la
biotecnología en plantas. El método más eficiente de regeneración de plantas /'n vÍ.Íro es la
ES. En chile habanero, se observa una recxalcitrancia a la conversión de los embriones
somáticos a plántulas. Este paso, representa la principal limitante en la aplicación y
explotación de la ES como herramienta biotecnológica para el mejoramiento de cultivos.
Por lo cual es de suma importancia continuar investigando los diversos factores que
podrían estar influenciando sobre la recalcitrancia a la conversión de los embriones
somáticos de esta especie.
Un punto a investigar sería determinar el papel de los reguladores del crecimiento vegetal,
principalmente las auxinas y su transporte en el proceso de la ES. Los resultados
obtenidos nos llevarían a elucidar la posible implicación de éstas en la recalcitrancia de
nuestro sistema.
El protocolo de embriogénesis somática desarrollado en el presente trabajo representa un
nuevo punto de vista en cuanto a la regeneración /.n v/.fro de esta especie (C. ch/.nense).
Contrario a lo que ha sido reportado para este género, el chile habanero posee un alto
potencial para la inducción de la ES; sin embargo, es necesario desarrollar estrategias
tanto bioquímicas como moleculares que permitan en un futuro utilizar este sistema de
regeneración para obtener plantas completas resistentes a plagas, bacterias, virus u
hongos.
De igual manera, los resultados derivados de este trabajo de investigación, con respecto a
la expresión del gen Wt/sche/, ofrece evidencias para sustentar el estudio de la expresión
de otros genes homeóticos implicados en el desarrollo de la ES de chile habanero, siendo
interesante la aplicación de técnicas moleculares para el estudio de otros genes
implicados en el desarrollo de la embriogénesis somática como son LEC7, LEC2, FUSCA
3 (FUS3), ABA-INSENSITIVE 3 (A813), CLAVATA 3 (CLV3) y PIN en el desarro\lo de los
embriones somáticos de chile habanero.
89








