ch4 alimentos o orina agua - vet.unicen.edu.ar · principal medio de transporte de todos los...
TRANSCRIPT
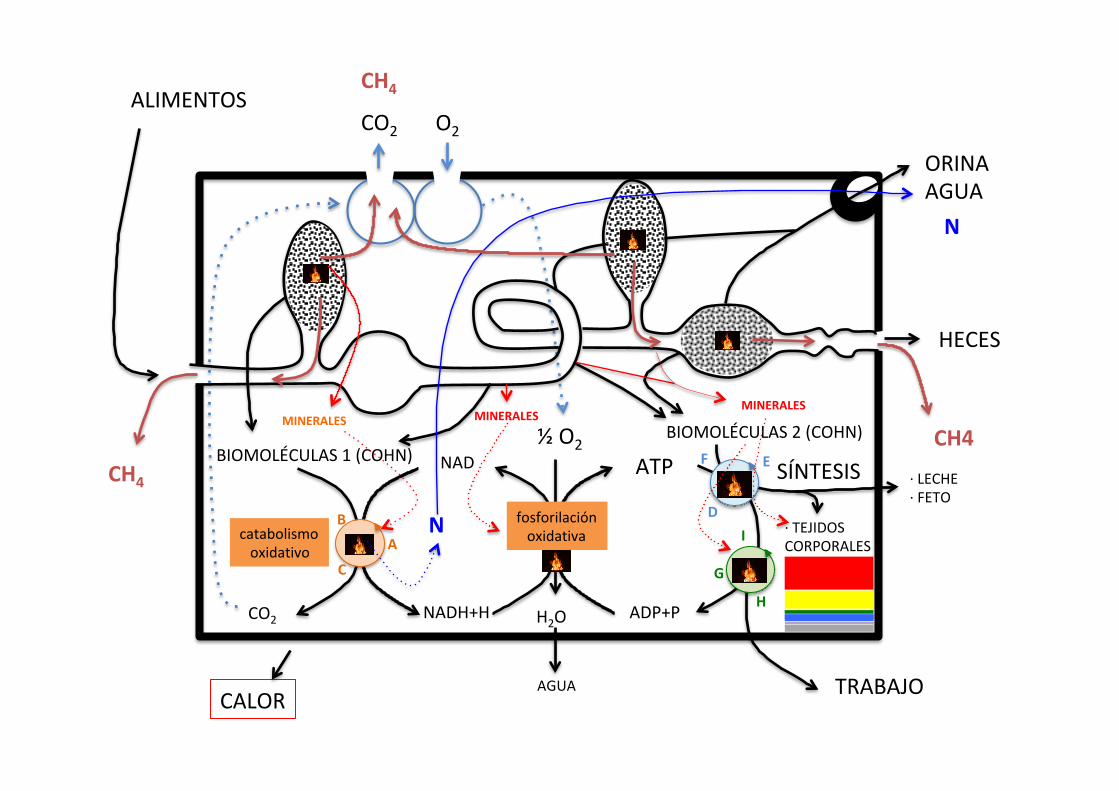
CO2
NAD
NADH+H H2O
½O2
ADP+P
ATP
TRABAJO
SÍNTESIS
ALIMENTOS
·LECHE·FETO
catabolismooxidativo
fosforilaciónoxidativa
AGUA
HECES
AB
C
ORINAAGUA
O2CO2
CALOR
·TEJIDOSCORPORALES
EF
D
G
H
I
CH4
CH4BIOMOLÉCULAS1(COHN)
BIOMOLÉCULAS2(COHN)MINERALES MINERALES
MINERALES
N
CH4
N

Solventedetodosloscompuestosquímicosdelcuerpodentroyfueradelascélulas
Eselprincipalconstituyentedelcuerpo(≅60%delamasacorporal)
Principalmediodetransportedetodoslosnutrientesdelcuerpoentrelostejidosydeexcreciónderesiduosdelmetabolismo
Tieneunelevadocalorespecíficoyaltocalordevaporización->absorbeelcalorqueseoriginaenelmetabolismocelularconmínimocambiodetemperatura->Regulacióndelatemperaturacorporal
Proveelubricaciónparaminimizarlafriccióndelosmovimientosdefluídoscorporalesypartesdelcuerpo(articulaciones)
Regulalaosmolaridadypresiónosmóticadeloscompartimientoscorporales

AGUA 50-70%peso
0
75
70
%peso
50
Inversamenterelacionadocon%degrasa
(agua+minerales+proteína+grasa=1o100)
Edad

Composicióndelamasacorporallibredegrasa(MCLG)dediferentesanimales
ChurchyPond,1994
Novillodelgado
Cerdo(100kg)
Gallina
Caballo
Conejo
Humano
64
49
57
60
69
60
19
12
21
18
18
18
5
3
3
4
5
4
88
65
71
82
93
82
0,73
0,75
0,80
0,73
0,74
0,73
Agua Proteína Minerales MCLG Agua/MCLG
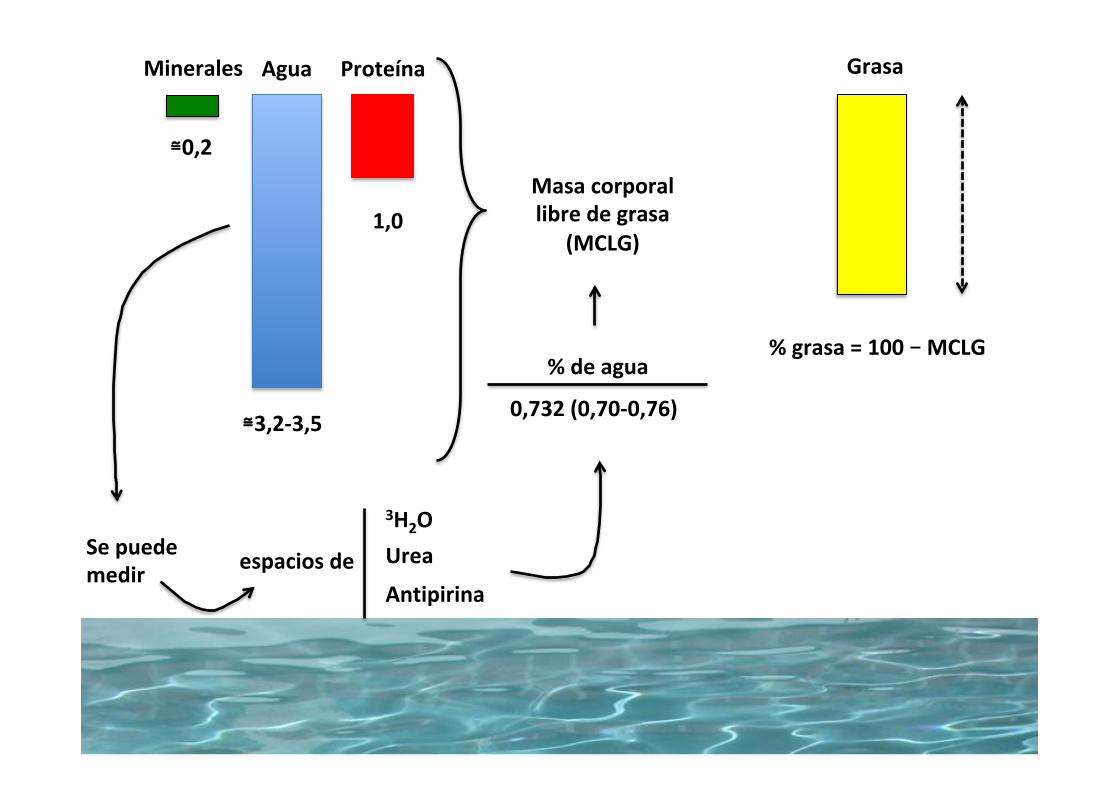
Agua Proteína
≅3,2-3,5
1,0
GrasaMinerales
Sepuedemedir
3H2OUrea
Antipirinaespaciosde
Masacorporallibredegrasa
(MCLG)
%deagua
0,732(0,70-0,76)
≅0,2
%grasa=100–MCLG

2/31/3
Extracelular Intracelular
∆Masacorporal-crecimiento-condicióncorporal
Distribuciónaguacorporal

506 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
OsmosisA critical concept in biology is the concept of osmosis. Water will always try to move from an area or compartment of low solute concentration to an area of high solute concentration (Figure 44.5). Each ion or particle within a solution acts as an osmotic particle, regardless of its charge. The osmolarity of a compartment or solution is determined by the concentration or number of moles of particles in that solution: 1 mol of sodium in a solution will provide just as much osmotic pull for water as 1 mol of albumin. Thinking another way, 10 g of sodium (atomic weight 23) will provide 0.43 osmol of particles to a solution. Adding 10 g albumin (MW 60,000) to the solution provides just 0.00016 osmol of particles to a solution. The key to absorption of water across the intestinal tract is to absorb solute. Water will follow the solute across the cell membranes.
Solvent drag or convection of soluteSmall solutes such as electrolytes can be swept from one compartment to another by the bulk flow of water. The water is moving from one compartment to another due to hydrostatic pressure or osmotic pull. The smaller the particle and the less charge it carries, the greater the chance it can be dragged along with the water into the next compartment.
PinocytosisIn some circumstances, very large or highly charged particles can be moved across cell membranes via a process called endo-cytosis. The substance to be transported comes in contact with the cell membrane and an endocytic membrane forms around the substance. It is taken into the cell as a membrane‐surrounded
vesicle and very often the membrane will cross the membrane on the opposite side of the cell by exocytosis. This process is used to absorb immunoglobulins, very large proteins in colos-trum that provide passive immunity from mother to neonate.
Paracellular versus transcellular transport
Paracellular absorptionAll the mucosal cells lining the intestinal tract have an apical and a basolateral membrane. Adjacent cells are linked to one another on all sides by “tight junctions,” also known as occluding junctions and zonula occludens. The tight junctions are com-posed of several proteins that form a seal between cells that is relatively impermeable to bacteria, viruses, and large molecules that have been ingested. The tight junctions also provide resis-tance to the passage of small ions and water. However, this resis-tance can be overcome if the electrochemical forces driving the ions to the opposite side of a tight junction are great enough. Water channels and ion channels do exist in the tight junction. They provide paths of lower resistance for water and ions that are utilized only when there is a great difference in osmolarity or concentration on one side of the tight junction. Movement across the tight junctions is only significant for the smaller ions such as Na+, Cl–, K+, Ca2+, PO4
– and Mg2+. Absorption of solute across the tight junctions between enterocytes, from the lumen directly into the extracellular fluid, is referred to as paracellular transport (Figure 44.6). It is also possible for solute to move from extracellular fluid to the lumen across the tight junctions. Very high concentrations of some solutes in the lumen of the intestine, such as Ca2+, can put so much electrochemical force on the tight junction that it is damaged and develops holes and becomes leaky.
Transcellular absorptionMost of the nutrients of the body are too large to cross the tight junctions and must be moved across the absorptive enterocytes of the villus by a variety of transport mechanisms (Figure 44.6). Transport proteins can facilitate passive diffusion or allow active transport (pumping) of solute against its electrochemical gra-dient at the expense of ATP. The process involves moving solute from lumen to cytosol of the enterocyte across the apical mem-brane and movement of solute from the cytosol to extracellular fluid across the basolateral membrane. The mechanism used to transport a solute across the apical membrane is often quite different from that used to transport the solute across the basolateral cell membrane.
Compartment 1 Compartment 2 Compartment 1 Compartment 2(A) (B)
Figure 44.5 Osmosis. (A) Before osmosis. Equal volumes of aqueous solutions (solutes represented by black circles and open circles) are placed in compartments that are separated by a membrane permeable to water but not to the solutes (semipermeable membrane). The aqueous solution in compartment 1 has the highest concentration of water (lowest concentration of solute). (B) During osmosis. Osmosis (diffusion of water) occurs from compartment 1 to compartment 2 (highest water concentration to lowest water concentration) and the water level rises in compartment 2. From Reece, W.O. (2009) Functional Anatomy and Physiology of Domestic Animals, 4th edn. Wiley‐Blackwell, Ames, IA. Reproduced with permission from Wiley.
1 How do paracellular and transcellular transport processes differ?
2 Which process works well when there is a large concentration gradient favoring transport? Which system is required to be used if pumping of a substance against its electrochemical gradient is required?
ÓSMOSIS
Cadaiónopartículaenunasoluciónactúacomounapartículaosmótica,enformaindependientedesucargaodesumasa.
Cuandolamembranaqueseparadoscompartimentosesimpermeablealossolutos,peronoalagua,elaguasiempresemuevedesdelazonadebajaconcentacióndesolutosaladealtaconcentracióndesolutos.
Ejemplo:1mmoldeNa+(23mg)ensoluciónejercelamismafuerzaosmóticaqueunmmoldealbúmina(68g)
Paramoveraguaatravésdeunamembranahayquemoversolutos:elagualossigue
(Paracelular)

Chapter 44: Digestion and Absorption of Nutrients 507
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
ABSORPTION OF DIETARY MINERALS AND REABSORPTION OF SECRETED ELECTROLYTES
Dietary electrolytes (Na+, K+ and Cl–) and minerals must be in solution within the fluids of the digestive tract if they are to be absorbed. Fortunately, many minerals that might be in an insol-uble form within the diet components become soluble after exposure to the acid in the stomach. The stomach can absorb some sodium and potassium, but these electrolytes are likely to have been placed in the gastric glands from the blood and not the diet. For monogastric species the bulk of mineral and elec-trolyte absorption occurs in the small and large intestine. The colon can also absorb electrolytes quite well if they are still present in the lumen. The major site of absorption and the mechanisms used to absorb the electrolytes are listed in Table 44.1.
Sodium
Sodium concentration in the lumen of the gastrointestinal tract and in the extracellular fluid is generally higher than inside the enterocyte. The inside of cells is negative in relation to the outside of cells.
Apical membrane transportIntracellular Na+ concentration is about 12–15 mmol/L. Lumen Na+ concentration will vary but except for the final sections of the colon will generally exceed 15 mmol/L. Na+ will be moved
across the apical membrane down its concentration and electrical gradient in most of the tract and down its electrical gradient even when Na+ concentration is very low in the distal colon.1 Cotransported with chloride to maintain electrical balance.
All sections of the small and large intestine have these Na+/Cl– cotransporters (Figure 44.7).
2 Cotransport of Na+ with sugars and amino acids liberated during digestion. The electrochemical force provided by Na+ moving down its electrical and concentration gradient will help push these larger molecules across the apical membrane. These transporter proteins are found in the villous cells of the duodenum and upper jejunum.
Basolateral membrane transportExtracellular Na+ concentration is about 140 mmol/L. Na+ will be moving out of the cell against its electrical and concentration gradient into the extracellular fluid.1 Electrogenic pump: 3Na+ ions will be pumped out of the
cell in exchange for 2K+ ions moving into the cell. This pump will require the energy from an ATP molecule to power it. All cells of the body have electrogenic pumps (Figure 44.7).
2 Na+/Cl– pump: sodium and chloride can be actively pumped across the basolateral membrane at the expense of an ATP. This mechanism is used primarily in the lower intestine (Figure 44.7).
Chloride
Apical membrane transportThe concentration of Cl– in the lumen is variable. Chyme leaving the stomach contains close to 120 mmol/L Cl–. The concentration of Cl– in the distal large intestine will be closer to zero. Intracellular Cl– concentration is variable and can be as low as 4 mmol/L in the colon and as high as 30 mmol/L in the upper small intestine.1 Cotransported with Na+ to maintain electrical balance. All
sections of the small and large intestine have these Na+/Cl– cotransporters (Figure 44.7).
2 One Cl– ion is brought into the cell in exchange for one HCO3
– ion moved into the lumen to maintain electrical neutrality. This process requires the energy of an ATP. This mechanism is especially important in the colon where lumen Cl– concentration is expected to be low (Figure 44.7).
3 Na+/K+/2Cl– cotransporter: the electromotive force of Na+ moving down both its electrical and concentration gradient helps move the more reluctant K+ and Cl– ions across the apical membrane (Figure 44.7).
1 How do sodium atoms cross from the lumen of the gut to the interior of the villous cell?
2 How do sodium atoms cross from the interior of the cell to the extracellular fluid?
Z
Z
X Y
X Y
Figure 44.6 X and Y ions are transported across the intestine by transcellular mechanisms: they have to cross both the apical and basolateral cell membranes. These mechanisms are efficient even if relatively low amounts of solute are present in the lumen compared with extracellular fluid. Ion Z is traversing the tight junction and is absorbed by a paracellular process. Paracellular transport is concentration driven and only functions when solute concentration in the lumen is far higher than ion concentration in the extracellular fluid.
1 How do chloride atoms cross from the lumen of the gut to the interior of the villous cell?
2 How do chloride atoms cross from the interior of the cell to the extracellular fluid?
3 Where is paracellular absorption of chloride most likely to occur and why?
VÍASTRANSCELULARYPARACELULAR
Transcelular TranscelularParacelular
Unionesestrechas(tightjunctions)

104 Section II: Body Fluids and Homeostasis
Sect
ion
II: B
ody
Flui
ds
and
Hom
eost
asis
metals placed side by side. Over time, the two metals will fuse with each. This is still simple diffusion. If a concentration gra-dient (differential) exists, molecules, ions, and colloidal particles tend to move from the area of their higher concentration to the area of their lower concentration. The movement is specific to each substance, i.e., Na+ will diffuse from the area of its higher concentration to the area of its lower concentration regardless of the presence and concentrations of other substances. If the mole-cules and ions are dispersed equally, the random motion continues but does not accomplish net movement or flow; this represents a state of equilibrium. Energy is not required for simple diffusion.
Barriers to diffusion in the animal body are generally the mem-branes of cells. These consist of a lipid bilayer, which is a thin film of lipid only two molecules thick through which fat‐soluble substances (especially carbon dioxide and oxygen) can readily diffuse (Figure 11.1). There might be facilitated diffusion for other substances, in which a carrier is required (Figure 11.2). However, facilitated diffusion for any substance still occurs from the area of its higher concentration to that of its lower concentration and, as in simple diffusion, energy is not required. Because cell membranes are predominantly lipid, they are relatively hydrophobic (water repelling), and the diffusion of water through the lipid bilayer proceeds with difficulty, but water can diffuse through protein channels. Protein channels (Figure 11.1) consist of large protein molecules interspersed in the lipid film; they provide structural pathways (pores) not only for water but also for water‐soluble substances. Some substances may be excluded from diffusion through the pores because of their large size; conversely, diffusion may be facilitated because of other factors, such as a substance’s relatively smaller size, its electrical charge (e.g., negative pore charge assists Na+ diffusion), or the protein channel’s specificity (e.g., specific ion channels). Other protein channels act as carrier proteins for the transport of substances in a direction opposite to their natural diffusion pathway. This is known as active transport. Whereas the trans-port of glucose into most cells of the body is accomplished by facilitated diffusion, exceptions exist in the lumens of the kidney tubules and intestines, where active transport is involved. In these locations, glucose is continually transported into the blood, where
the concentration is high, from the lumen where its concentration may be minute. Loss of glucose from the body is prevented in these locations because of active transport. Active transport requires not only a carrier but also energy.
Protein channelsProtein channels
Lipidbilayer
Figure 11.1 Structure of a cell membrane. The lipid bilayer is represented by a thin film of lipid that is two molecules thick. The protein channels (pores) may be composed of a single protein or a cluster of proteins. The channels may have specificity for certain substances, or they may be restrictive because of size. Virtually all water diffuses through the protein channels. Adapted from Reece, W.O. (2009) Functional Anatomy and Physiology of Domestic Animals, 4th edn. Wiley‐Blackwell, Ames, IA.
Carrierproteinchannel
Carrier receptor protein
Conformation A
Conformation B
Transportedmolecule
Lipid Bilayer
Figure 11.2 A postulated mechanism for facilitated diffusion. (A) The transported molecule enters the protein channel and binds with the receptor at the binding site. (B) Subsequent to binding, the protein channel undergoes a conformational change to open the channel on the opposite side, and the transported molecule is released, causing return of the protein channel to its original conformation. From Reece, W.O. (2009) Functional Anatomy and Physiology of Domestic Animals, 4th edn. Wiley‐Blackwell, Ames, IA. Reproduced with permission from Wiley.
H2OCanalesProteicos(Transcelular)
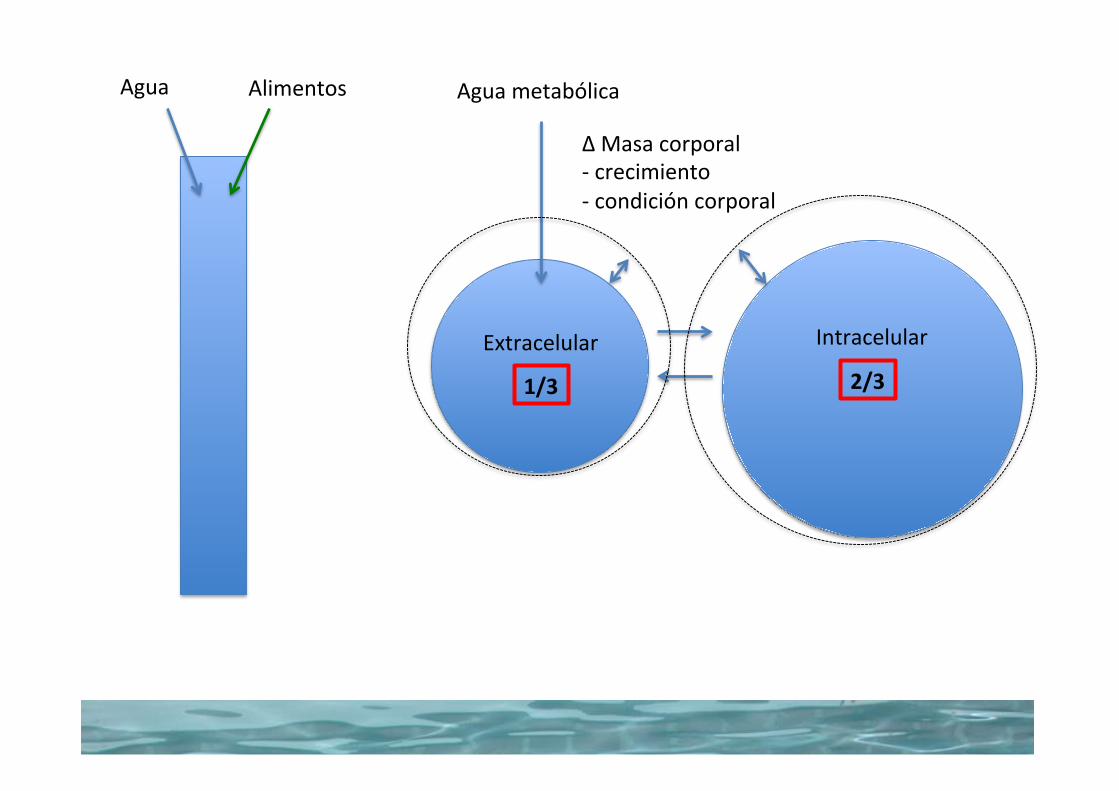
2/31/3
AguametabólicaAgua Alimentos
Extracelular Intracelular
∆Masacorporal-crecimiento-condicióncorporal


McDonald,Edward,GreenhalghyMorgan.NutriciónAnimal,5taEdición
Chapter 1 The animal and its food
4
Table 1.1 Composition of some plant and animal products expressed on a freshbasis and a dry matter basis
Water Carbohydrate Lipid Protein Ash
Fresh basis (g/kg)Turnips 910 70 2 11 7Grass (young) 800 137 8 35 20Barley grain 140 730 15 93 22Groundnuts 60 201 449 268 22Dairy cow 570 2 206 172 50Milk 876 47 36 33 8Muscle 720 6 44 215 15Egg 667 8 100 118 107
Dry matter basis (g/kg)Turnips 0 778 22 122 78Grass (young) 0 685 40 175 100Barley grain 0 849 17 108 26Groundnuts 0 214 478 285 23Dairy cow 0 5 479 400 116Milk 0 379 290 266 65Muscle 0 21 157 768 54Egg 0 24 300 355 321
1.1 WATER
The water content of the animal body varies with age.The newborn animal contains750–800 g/kg water but this falls to about 500 g/kg in the mature fat animal. It isvital to the life of the organism that the water content of the body be maintained: ananimal will die more rapidly if deprived of water than if deprived of food. Waterfunctions in the body as a solvent in which nutrients are transported about the bodyand in which waste products are excreted. Many of the chemical reactions broughtabout by enzymes take place in solution and involve hydrolysis. Because of the highspecific heat of water, large changes in heat production can take place within theanimal with very little alteration in body temperature. Water also has a high latentheat of evaporation, and its evaporation from the lungs and skin gives it a furtherrole in the regulation of body temperature.
The animal obtains its water from three sources: drinking water, water present in itsfood, and metabolic water, this last being formed during metabolism by the oxidation ofhydrogen-containing organic nutrients. The water content of foods is variable and canrange from as little as 60 g/kg in concentrates to over 900 g/kg in some root crops. Be-cause of this great variation in water content, the composition of foods is often ex-pressed on a dry matter basis, which allows a more valid comparison of nutrient content.This is illustrated inTable 1.1, which lists a few examples of plant and animal products.
The water content of growing plants is related to the stage of growth, being greaterin younger plants than in older plants. In temperate climates the acquisition of drink-ing water is not usually a problem and animals are provided with a continuous supply.There is no evidence that under normal conditions an excess of drinking water isharmful, and animals normally drink what they require.
Peso fresco (g/kg) “tal cual”
Nabo
Vaca Lechera
Leche
Músculo
Huevo
Grano de cebada
Maní
Pasto joven (tierno)
Materia seca (g/kg)
Nabo
Vaca Lechera
Leche
Músculo
Huevo
Grano de cebada
Maní
Pasto joven (tierno)
Tabla 1.1: Composición de algunos materiales vegetales y productos animales expresados como peso fresco (“tal cual”) o como materia seca
Agua Carbohidratos Lípidos Proteína Cenizas
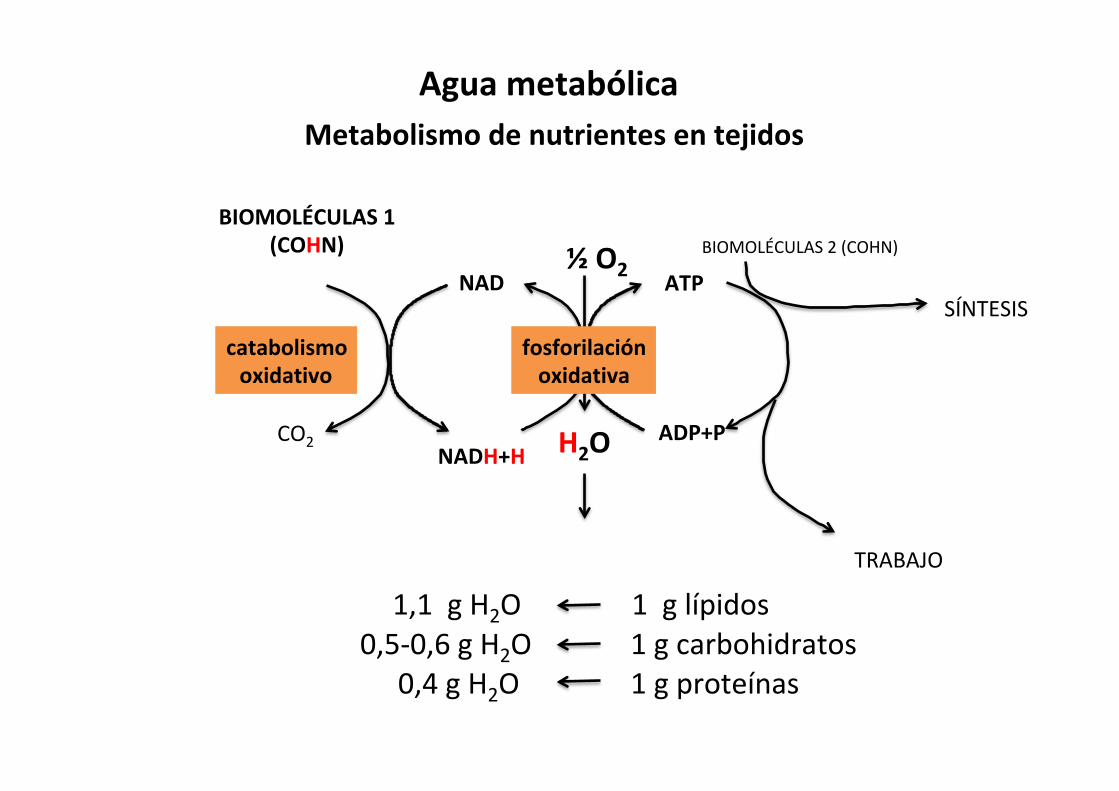
BIOMOLÉCULAS1(COHN)
CO2
NAD
NADH+H H2O
½O2
ADP+P
ATPBIOMOLÉCULAS2(COHN)
catabolismooxidativo
fosforilaciónoxidativa
TRABAJO
SÍNTESIS
MetabolismodenutrientesentejidosAguametabólica
1,1gH2O
1gproteínas
1glípidos1gcarbohidratos0,5-0,6gH2O
0,4gH2O

Chapter 11: Body Water: Properties and Functions 111
Sect
ion
II: B
ody
Flui
ds
and
Hom
eost
asis
reserves associated with their relatively higher needs that calves become distressed more quickly in conditions of uncontrolled water loss (such as diarrhea). It should also be noted that because of the greater surface area relative to the body weight of calves, they will also lose body heat quicker than the cows.
Dehydration, thirst, and water intake
When water losses exceed water gains, a condition known as dehydration develops. The extent is variable and, when mild, physiologic mechanisms may be adequate to reestablish water balance via the thirst mechanism if water is available. Therapeutic measures (fluid replacement, treatment of under-lying cause) may be necessary when water losses are moderate to severe and related to a disease condition.
DehydrationIn dehydration, the immediate source of water lost from the body is the ECF, followed by a shift from the intracellular to the extracellular fluid. A loss of water equal to 10% of body weight is considered to be severe for most animals. The concentrations of electrolytes (ions) in the body fluids do not continue to increase during dehydration, but are excreted by the kidney in proportion to the water loss. With continuing dehydration, water and electrolytes are depleted. Therefore, rehydration requires not only water but also appropriate electrolytes.
Stimulus for thirstWhen water losses exceed water gain, there is an effort on the part of the kidneys to conserve water. Also, animals are provided with a thirst mechanism to recognize the need for water intake greater than that provided by food and metabolic water. Thirst is the conscious desire for water. Central to the thirst mecha-nism is a thirst center located in the hypothalamus of the brain and represented by thirst cells. The thirst cells are stimulated by an increase in their osmoconcentration (loss of water and increased salt concentration). Osmoconcentration of the thirst cells is a consequence of dehydration.
Aminoacids
Fattyacids Glucose
Glycolysis
Acetyl-CoA
OxaloacetateCitrate
Citricacid cycle
Pyruvate
Pyruvatedehydrogenasecomplex
Stage 1Acetyl-CoAproduction
Stage 2Acetyl-CoAproduction
Stage 3Electron transfer
and oxidativephosphorylation
CO2
CO2
CO2
2H+ + 1/2 O2
H2O
e–
e–
e–
e–
e–
e–
e–
e–
e–
NADH,FADH2
(reduced e– carriers)
Respiratory(electron transfer)
chain
ADP ATP+ Pi
Figure 11.9 Catabolism of proteins, fats, and carbohydrates resulting in release of energy. Stage 3, via the electron transfer chain, provides for the oxidative phosphorylation of adenosine diphosphate (ADP) and the production of a high‐energy substance, adenosine triphosphate (ATP). This is the location of oxygen consumption by the body and production of metabolic water. Adapted from Nelson, D.L. and Cox, M.M. (2000) Lehninger Principles of Biochemistry, 3rd edn. Worth Publishers, New York.
1 In dehydration, what is the immediate source (compartment) of water lost from the body?
2 For most animals, what is considered to be a severe loss of body water?
3 With continuing loss of water (dehydration), is there a proportionate loss of electrolytes?
4 Define thirst.
5 Where is the thirst center located?
6 How does dehydration stimulate thirst?
7 How does hypovolemia stimulate thirst?
8 How can thirst be temporarily relieved?
Origendelaguametabólica
40gH2O 100gproteínas
100glípidos
100gcarbohidratos60gH2O
110gH2O
Etapa1ProduccióndeAcetil-CoA
Etapa2OxidacióndeAcetil-CoA
Etapa3Transferenciadee-yfosforilaciónoxidativa

Chapter 11: Body Water: Properties and Functions 111
Sect
ion
II: B
od
y Fl
uid
s an
d H
om
eost
asis
reserves associated with their relatively higher needs that calves become distressed more quickly in conditions of uncontrolled water loss (such as diarrhea). It should also be noted that because of the greater surface area relative to the body weight of calves, they will also lose body heat quicker than the cows.
Dehydration, thirst, and water intake
When water losses exceed water gains, a condition known as dehydration develops. The extent is variable and, when mild, physiologic mechanisms may be adequate to reestablish water balance via the thirst mechanism if water is available. Therapeutic measures (fluid replacement, treatment of under-lying cause) may be necessary when water losses are moderate to severe and related to a disease condition.
DehydrationIn dehydration, the immediate source of water lost from the body is the ECF, followed by a shift from the intracellular to the extracellular fluid. A loss of water equal to 10% of body weight is considered to be severe for most animals. The concentrations of electrolytes (ions) in the body fluids do not continue to increase during dehydration, but are excreted by the kidney in proportion to the water loss. With continuing dehydration, water and electrolytes are depleted. Therefore, rehydration requires not only water but also appropriate electrolytes.
Stimulus for thirstWhen water losses exceed water gain, there is an effort on the part of the kidneys to conserve water. Also, animals are provided with a thirst mechanism to recognize the need for water intake greater than that provided by food and metabolic water. Thirst is the conscious desire for water. Central to the thirst mecha-nism is a thirst center located in the hypothalamus of the brain and represented by thirst cells. The thirst cells are stimulated by an increase in their osmoconcentration (loss of water and increased salt concentration). Osmoconcentration of the thirst cells is a consequence of dehydration.
Aminoacids
Fattyacids Glucose
Glycolysis
Acetyl-CoA
OxaloacetateCitrate
Citricacid cycle
Pyruvate
Pyruvatedehydrogenasecomplex
Stage 1Acetyl-CoAproduction
Stage 2Acetyl-CoAproduction
Stage 3Electron transfer
and oxidativephosphorylation
CO2
CO2
CO2
2H+ + 1/2 O2
H2O
e–
e–
e–
e–
e–
e–
e–
e–
e–
NADH,FADH2
(reduced e– carriers)
Respiratory(electron transfer)
chain
ADP ATP+ Pi
Figure 11.9 Catabolism of proteins, fats, and carbohydrates resulting in release of energy. Stage 3, via the electron transfer chain, provides for the oxidative phosphorylation of adenosine diphosphate (ADP) and the production of a high‐energy substance, adenosine triphosphate (ATP). This is the location of oxygen consumption by the body and production of metabolic water. Adapted from Nelson, D.L. and Cox, M.M. (2000) Lehninger Principles of Biochemistry, 3rd edn. Worth Publishers, New York.
1 In dehydration, what is the immediate source (compartment) of water lost from the body?
2 For most animals, what is considered to be a severe loss of body water?
3 With continuing loss of water (dehydration), is there a proportionate loss of electrolytes?
4 Define thirst.
5 Where is the thirst center located?
6 How does dehydration stimulate thirst?
7 How does hypovolemia stimulate thirst?
8 How can thirst be temporarily relieved?
e-H+
e-H+e-H+
e-H+
Etapa1ProduccióndeAcetil-
CoA

Chapter 11: Body Water: Properties and Functions 111
Sect
ion
II: B
ody
Flui
ds
and
Hom
eost
asis
reserves associated with their relatively higher needs that calves become distressed more quickly in conditions of uncontrolled water loss (such as diarrhea). It should also be noted that because of the greater surface area relative to the body weight of calves, they will also lose body heat quicker than the cows.
Dehydration, thirst, and water intake
When water losses exceed water gains, a condition known as dehydration develops. The extent is variable and, when mild, physiologic mechanisms may be adequate to reestablish water balance via the thirst mechanism if water is available. Therapeutic measures (fluid replacement, treatment of under-lying cause) may be necessary when water losses are moderate to severe and related to a disease condition.
DehydrationIn dehydration, the immediate source of water lost from the body is the ECF, followed by a shift from the intracellular to the extracellular fluid. A loss of water equal to 10% of body weight is considered to be severe for most animals. The concentrations of electrolytes (ions) in the body fluids do not continue to increase during dehydration, but are excreted by the kidney in proportion to the water loss. With continuing dehydration, water and electrolytes are depleted. Therefore, rehydration requires not only water but also appropriate electrolytes.
Stimulus for thirstWhen water losses exceed water gain, there is an effort on the part of the kidneys to conserve water. Also, animals are provided with a thirst mechanism to recognize the need for water intake greater than that provided by food and metabolic water. Thirst is the conscious desire for water. Central to the thirst mecha-nism is a thirst center located in the hypothalamus of the brain and represented by thirst cells. The thirst cells are stimulated by an increase in their osmoconcentration (loss of water and increased salt concentration). Osmoconcentration of the thirst cells is a consequence of dehydration.
Aminoacids
Fattyacids Glucose
Glycolysis
Acetyl-CoA
OxaloacetateCitrate
Citricacid cycle
Pyruvate
Pyruvatedehydrogenasecomplex
Stage 1Acetyl-CoAproduction
Stage 2Acetyl-CoAproduction
Stage 3Electron transfer
and oxidativephosphorylation
CO2
CO2
CO2
2H+ + 1/2 O2
H2O
e–
e–
e–
e–
e–
e–
e–
e–
e–
NADH,FADH2
(reduced e– carriers)
Respiratory(electron transfer)
chain
ADP ATP+ Pi
Figure 11.9 Catabolism of proteins, fats, and carbohydrates resulting in release of energy. Stage 3, via the electron transfer chain, provides for the oxidative phosphorylation of adenosine diphosphate (ADP) and the production of a high‐energy substance, adenosine triphosphate (ATP). This is the location of oxygen consumption by the body and production of metabolic water. Adapted from Nelson, D.L. and Cox, M.M. (2000) Lehninger Principles of Biochemistry, 3rd edn. Worth Publishers, New York.
1 In dehydration, what is the immediate source (compartment) of water lost from the body?
2 For most animals, what is considered to be a severe loss of body water?
3 With continuing loss of water (dehydration), is there a proportionate loss of electrolytes?
4 Define thirst.
5 Where is the thirst center located?
6 How does dehydration stimulate thirst?
7 How does hypovolemia stimulate thirst?
8 How can thirst be temporarily relieved?
e-H+
e-H+
e-H+
e-H+
Etapa2OxidacióndeAcetil-CoA

Chapter 11: Body Water: Properties and Functions 111
Sect
ion
II: B
ody
Flui
ds
and
Hom
eost
asis
reserves associated with their relatively higher needs that calves become distressed more quickly in conditions of uncontrolled water loss (such as diarrhea). It should also be noted that because of the greater surface area relative to the body weight of calves, they will also lose body heat quicker than the cows.
Dehydration, thirst, and water intake
When water losses exceed water gains, a condition known as dehydration develops. The extent is variable and, when mild, physiologic mechanisms may be adequate to reestablish water balance via the thirst mechanism if water is available. Therapeutic measures (fluid replacement, treatment of under-lying cause) may be necessary when water losses are moderate to severe and related to a disease condition.
DehydrationIn dehydration, the immediate source of water lost from the body is the ECF, followed by a shift from the intracellular to the extracellular fluid. A loss of water equal to 10% of body weight is considered to be severe for most animals. The concentrations of electrolytes (ions) in the body fluids do not continue to increase during dehydration, but are excreted by the kidney in proportion to the water loss. With continuing dehydration, water and electrolytes are depleted. Therefore, rehydration requires not only water but also appropriate electrolytes.
Stimulus for thirstWhen water losses exceed water gain, there is an effort on the part of the kidneys to conserve water. Also, animals are provided with a thirst mechanism to recognize the need for water intake greater than that provided by food and metabolic water. Thirst is the conscious desire for water. Central to the thirst mecha-nism is a thirst center located in the hypothalamus of the brain and represented by thirst cells. The thirst cells are stimulated by an increase in their osmoconcentration (loss of water and increased salt concentration). Osmoconcentration of the thirst cells is a consequence of dehydration.
Aminoacids
Fattyacids Glucose
Glycolysis
Acetyl-CoA
OxaloacetateCitrate
Citricacid cycle
Pyruvate
Pyruvatedehydrogenasecomplex
Stage 1Acetyl-CoAproduction
Stage 2Acetyl-CoAproduction
Stage 3Electron transfer
and oxidativephosphorylation
CO2
CO2
CO2
2H+ + 1/2 O2
H2O
e–
e–
e–
e–
e–
e–
e–
e–
e–
NADH,FADH2
(reduced e– carriers)
Respiratory(electron transfer)
chain
ADP ATP+ Pi
Figure 11.9 Catabolism of proteins, fats, and carbohydrates resulting in release of energy. Stage 3, via the electron transfer chain, provides for the oxidative phosphorylation of adenosine diphosphate (ADP) and the production of a high‐energy substance, adenosine triphosphate (ATP). This is the location of oxygen consumption by the body and production of metabolic water. Adapted from Nelson, D.L. and Cox, M.M. (2000) Lehninger Principles of Biochemistry, 3rd edn. Worth Publishers, New York.
1 In dehydration, what is the immediate source (compartment) of water lost from the body?
2 For most animals, what is considered to be a severe loss of body water?
3 With continuing loss of water (dehydration), is there a proportionate loss of electrolytes?
4 Define thirst.
5 Where is the thirst center located?
6 How does dehydration stimulate thirst?
7 How does hypovolemia stimulate thirst?
8 How can thirst be temporarily relieved?
Etapa3Transferenciadee-
yfosforilaciónoxidativa
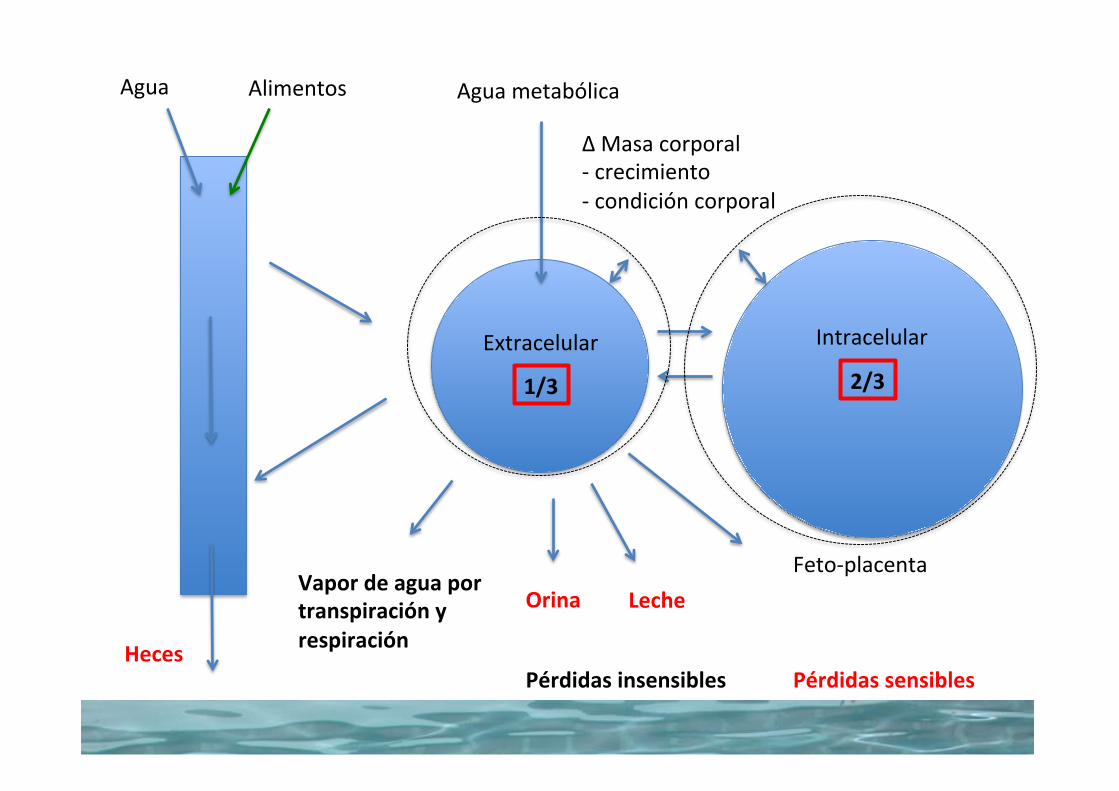
2/31/3
Vapordeaguaportranspiraciónyrespiración
Orina
AguametabólicaAgua Alimentos
Heces
Extracelular Intracelular
LecheFeto-placenta
∆Masacorporal-crecimiento-condicióncorporal
PérdidassensiblesPérdidasinsensibles

110 Section II: Body Fluids and Homeostasis
Sect
ion
II: B
ody
Flui
ds
and
Hom
eost
asis
compartments because no osmoconcentation differences would exist. The infusion of a hypertonic solution would pre-sent a greater effective osmotic pressure in the extracellular compartment than in the intracellular compartment and water would diffuse from the cells to the extracellular compartment. The infusion of a hypertonic solution has been useful in the treatment of head injuries to reduce the swelling (volume) often associated with head injuries.
Water balance
From day to day in any one animal the water content of the body remains relatively constant, with a balance between gains and losses. Water turnover is that amount of water gained by an animal to balance that which is lost. Typical values for lactating and nonlactating cows under moderate environmental condi-tions are shown in Table 11.4. The water turnover for the non-lactating cow is 29 L/day and for the lactating cow is 56 L/day. The water intake in both cases is equal to the output; there is water balance. The “pool size” stays constant, but the water in the pool changes (water turnover). The output of the lactating cow has increased, not only because of the obvious milk produc-tion, but also because of the greater fecal output associated with eating nearly twice as much and because of greater urine and vapor losses associated with increased metabolism.
Water gainWater gains occur by ingestion of water in food and drink and from metabolic water. The food eaten by animals contains a variable amount of water; the usual drink is water or, in the very young, milk. Metabolic water is derived from the chemical reactions of cellular metabolism in the mitochondria. At the end of the electron transfer chain, hydrogen is combined with oxygen to form water; this is metabolic water as shown in Figure 11.9.
The metabolism of proteins, carbohydrates, and fats requires different amounts of cofactors, with the greatest amounts required for fats. Accordingly, the yield of metabolic water is greater for a certain amount of fat than for an equal amount of protein or carbohydrate. For example, the metabolic water yield from each of 100 g of protein, carbohydrate, and fat is 40, 60, and 110 mL, respectively. Energy in the form of adenosine triphosphate (ATP) is formed during the transfer of electrons. The amount of metabolic water formed varies but could be sub-stantial under certain conditions. In domestic animals it is said to average about 5–10% of daily water gain, and it can approach 100% of the water gain for some small rodents.
Water lossWater loss from the body is classified as either an insensible loss or a sensible loss. Insensible losses are associated with vapor losses and occur constantly by evaporation from the skin and by loss of water vapor in exhaled air. Inhaled air becomes saturated with water vapor in the respiratory passages and lungs, but there is no body mechanism to remove moisture from the respiratory gases before exhalation. Sensible losses are the visible losses; they are part of the urine, feces, and body secretions that leave the body and are not subject to evaporation. Sensible losses can become excessive in certain conditions, such as diarrhea, and threaten body stores of water.
Water requirementsNo linear relationship exists between basal water needs and body weight. Accordingly, a 500‐kg cow does not require 10 times more water than a 50‐kg calf. However, the basal daily needs for water (that needed to maintain water balance) are related to caloric expenditure. Under basal metabolism con-ditions (e.g., resting animal, thermally neutral environment, fasting state), caloric expenditure is related linearly to body surface area. The cow might require only three to four times more water than the calf because her body surface area is three to four times greater. If the ECF (20% of body weight) is considered to be that from which emergency water is drawn, the 500‐kg cow has 100 kg of fluid and the 50‐kg calf has only 10 kg. Therefore, the cow has considerably more reserve on which to draw to supply basal needs for water than does the calf. In other words, the cow has ten times more reserve water to supply her needs, and her needs are only three to four times greater than the calf ’s. It is because of the more limited
Table 11.4 Daily water balance of Holstein cows eating legume hay (values in liters).
Balance Nonlactating Lactating
IntakeDrinking water 26 51Food water 1 2Metabolic water 2 3Total 29 56
OutputFeces 12 19Urine 7 11Vaporized 10 14Milk 0 12Total 29 56
Source: Houpt, T.R. (2004) Water and electrolytes. In: Dukes’ Physiology of Domestic Animals, 12th edn (ed. W.O. Reece). Cornell University Press, Ithaca, NY. Reproduced with permission from Cornell University Press.
1 What is meant by water turnover?
2 What is the derivation of metabolic water? Why does 5 g of fat yield more metabolic water than 5 g of protein or carbohydrate?
3 What are examples of insensible water loss?
4 Why are excess water losses (e.g., diarrhea) more critical in young animals than in adults of the same species?
Fuentes Agua de bebida Agua de alimento Agua metabólica Total
Pérdidas Heces Orina Vapor Leche Total
Seca EnLactación
Balancedeaguaenvacasdetamboalimentadasconración
Dukes,2015

2/31/3
Vapordeaguaportranspiraciónyrespiración
Orina
AguametabólicaAgua Alimentos
Heces
Extracelular Intracelular
LecheFeto-placenta
∆Masacorporal-crecimiento-condicióncorporal
PérdidassensiblesPérdidasinsensibles
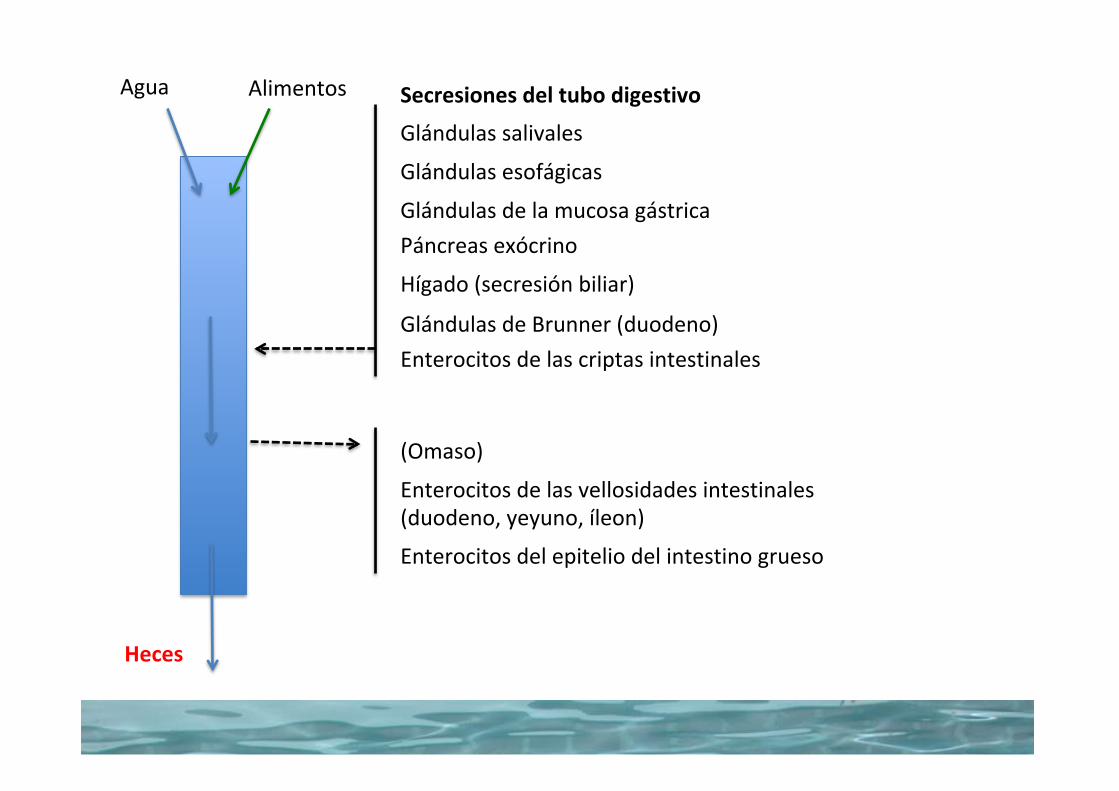
Agua Alimentos
Heces
Glándulassalivales
Glándulasesofágicas
Secresionesdeltubodigestivo
GlándulasdelamucosagástricaPáncreasexócrino
Hígado(secresiónbiliar)
EnterocitosdelascriptasintestinalesGlándulasdeBrunner(duodeno)
(Omaso)
Enterocitosdelasvellosidadesintestinales(duodeno,yeyuno,íleon)
Enterocitosdelepiteliodelintestinogrueso

Chapter 44: Digestion and Absorption of Nutrients 517
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
resistance can be overcome if osmotic forces are great enough on one or another side of the membrane.
A phenomena that aids absorption of water at the villous tip is the countercurrent movement of solute from vein to arteriole in the lamina propria of a villus. Recall that an arteriole carries blood to the tip of the villus and this arteriole is in close proximity to a venule carrying blood away from the tip of the villus. During digestion and absorption of a meal, the postcapil-lary venules bring large amounts of absorbed solute into the venules. As the venule passes down the villus, solute can diffuse from the venule into the arteriole down its concentration gra-dient (see Figure 44.14). The arteriole brings this solute to the villous tip and eventually the osmolarity of the lamina propria at the villous tip can reach 500–600 mosmol, about twice the osmolarity of plasma. This draws water from the lumen of the intestine into the villous tip and on to the extracellular fluid.
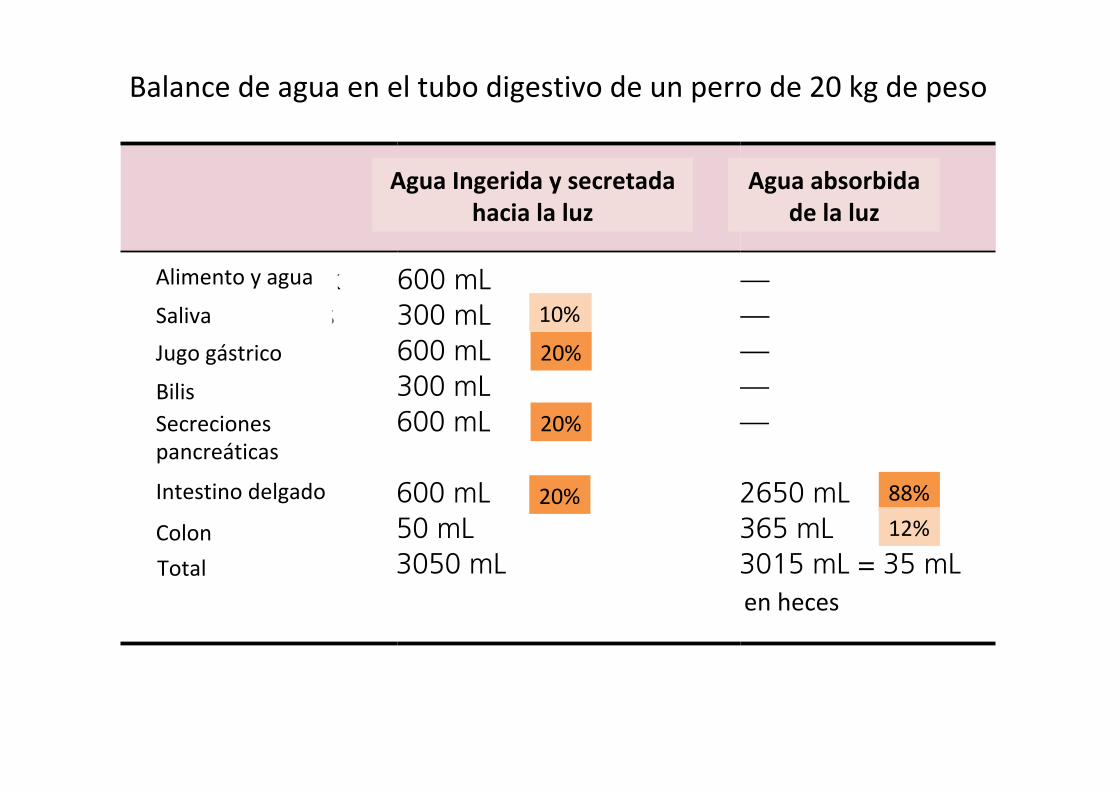
Water balance in the gastrointestinal tractNearly all the water that enters the gastrointestinal tract is removed, so that feces generally contain only small amounts of water. A 20‐kg dog might drink (or ingest with its diet) 600 mL water each day. Salivary glands add 300 mL, gastric juices 600 mL, bile 300 mL, pancreatic secretions 600 mL, small intestine (Brunner’s glands and crypt cell secretions) add another 600 mL, and the colonic mucus adds 50 mL water to the lumen of the gastrointestinal tract for a total of 3050 mL water entering the lumen (Table 44.2). Only about 35 mL, about 1%, of this water exits the tract with the feces. The bulk of the water, about 2650 mL, is absorbed by the small intestine. The duo-denum and jejunum absorb about 1600 mL or 52%. The ileum absorbs about 1060 mL or about 35%, and the colon absorbs 365 mL or 12% of the water entering the gastrointestinal tract. Water is absorbed with close to 99% efficiency. The water balance for the dog is therefore ingested 600 mL, retained 565 mL (which was likely used for respiration humidity and urine production).
Water balance in the horse (and other hindgut fermenters) is somewhat different. A 500‐kg horse would be expected to ingest about 40 L of water each day (Table 44.3). Salivary and gastric juices add 41 L of water, and pancreatic, bile, and small and large intestine secretions add another 63 L for a total of 144 L of water entering the gastrointestinal tract. The small intestine absorbs 79 L of water daily. The cecum absorbs 18 L, the ventral colon absorbs 13 L, the dorsal colon absorbs 18.5 L, and the transverse and descending colon absorb another 8 L of water, for a total of 57.5 L for the large intestine. The horse colon absorbs a much greater amount of water than the monogastric colon. It uses the osmotic force of VFA absorption to absorb much of this water. The net loss of water to the feces is about 7.5 L/day. On balance the horse retains 32.5 L of the 40 L ingested.
Malabsorptive diarrheaBacteria and viruses can damage the tight junctions and/or villous absorptive cells and can interfere with absorption of solute, both the solute that was in the diet and the solute secreted
from crypt cells and in pancreatic, and salivary secretions. The inability to absorb solute will also cause some loss in efficiency of water absorption. If water absorption efficiency in the 20‐kg dog drops from nearly 99% to 90% due to enteritis in a section of the intestine, fecal water content will increase from 35 to 275 mL, which will create a very watery feces. If damage to the gastrointestinal tract is restricted to the colon and no water is absorbed by the colon, fecal water content will only increase by the 365 mL the colon is expected to absorb. However, damage to large segments of the small intestine can cause loss of up to 2650 mL water.
Bacterial and viral infections can not only destroy villous absorptive cells, they can initiate inflammation reactions that generally cause hypersecretion by the crypt and goblet cells in an attempt to flush away the offending pathogens. This can greatly increase the amount of fluid lost with the feces. Bacterial infections tend to be fairly localized and cause local damage to either the small or large intestine. Rotaviruses attack the cells at the tips of villi causing some malabsorption. Coronaviruses, such as the causative agents of transmissible gastroenteritis and porcine epidemic diarrhea, kill villous entrocytes the full length of the villus and are therefore much more severe infections. Parvoviruses causing canine parvo and feline panleukopenia attack rapidly dividing cells – in the intestine these are the crypt
Table 44.2 Water secretion and absorption in a 20‐kg dog.
Into lumen (ingested and secreted)
Out of lumen (absorbed)
Food and drink 600 mL —Salivary glands 300 mL —Gastric juice 600 mL —Bile 300 mL —Pancreatic secretions
600 mL —
Small intestine 600 mL 2650 mLColon 50 mL 365 mLTotals 3050 mL 3015 mL = 35 mL
in feces
Table 44.3 Water secretion and absorption in a 500‐kg horse.
Into lumen (ingested and secreted)
Out of lumen (absorbed)
Food and drink 40 L —Salivary glands 30 L —Gastric juice 11 L —Bile 5 L —Pancreatic secretions
10 L —
Small intestine 47 L 79 LColon 1 L 57.5 LTotals 144 L 136.5 L = 7.5 L
in feces
Alimentoyagua
SalivaJugogástrico
BilisSecrecionespancreáticas
Intestinodelgado
ColonTotal
enheces
Balancedeaguaeneltubodigestivodeuncaballode500kgdepeso
AguaIngeridaysecretadahacialaluz
Aguaabsorbidadelaluz
33%
21%
58%
42%
Chapter 44: Digestion and Absorption of Nutrients 517
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
resistance can be overcome if osmotic forces are great enough on one or another side of the membrane.
A phenomena that aids absorption of water at the villous tip is the countercurrent movement of solute from vein to arteriole in the lamina propria of a villus. Recall that an arteriole carries blood to the tip of the villus and this arteriole is in close proximity to a venule carrying blood away from the tip of the villus. During digestion and absorption of a meal, the postcapil-lary venules bring large amounts of absorbed solute into the venules. As the venule passes down the villus, solute can diffuse from the venule into the arteriole down its concentration gra-dient (see Figure 44.14). The arteriole brings this solute to the villous tip and eventually the osmolarity of the lamina propria at the villous tip can reach 500–600 mosmol, about twice the osmolarity of plasma. This draws water from the lumen of the intestine into the villous tip and on to the extracellular fluid.
Water balance in the gastrointestinal tractNearly all the water that enters the gastrointestinal tract is removed, so that feces generally contain only small amounts of water. A 20‐kg dog might drink (or ingest with its diet) 600 mL water each day. Salivary glands add 300 mL, gastric juices 600 mL, bile 300 mL, pancreatic secretions 600 mL, small intestine (Brunner’s glands and crypt cell secretions) add another 600 mL, and the colonic mucus adds 50 mL water to the lumen of the gastrointestinal tract for a total of 3050 mL water entering the lumen (Table 44.2). Only about 35 mL, about 1%, of this water exits the tract with the feces. The bulk of the water, about 2650 mL, is absorbed by the small intestine. The duo-denum and jejunum absorb about 1600 mL or 52%. The ileum absorbs about 1060 mL or about 35%, and the colon absorbs 365 mL or 12% of the water entering the gastrointestinal tract. Water is absorbed with close to 99% efficiency. The water balance for the dog is therefore ingested 600 mL, retained 565 mL (which was likely used for respiration humidity and urine production).
Water balance in the horse (and other hindgut fermenters) is somewhat different. A 500‐kg horse would be expected to ingest about 40 L of water each day (Table 44.3). Salivary and gastric juices add 41 L of water, and pancreatic, bile, and small and large intestine secretions add another 63 L for a total of 144 L of water entering the gastrointestinal tract. The small intestine absorbs 79 L of water daily. The cecum absorbs 18 L, the ventral colon absorbs 13 L, the dorsal colon absorbs 18.5 L, and the transverse and descending colon absorb another 8 L of water, for a total of 57.5 L for the large intestine. The horse colon absorbs a much greater amount of water than the monogastric colon. It uses the osmotic force of VFA absorption to absorb much of this water. The net loss of water to the feces is about 7.5 L/day. On balance the horse retains 32.5 L of the 40 L ingested.
Malabsorptive diarrheaBacteria and viruses can damage the tight junctions and/or villous absorptive cells and can interfere with absorption of solute, both the solute that was in the diet and the solute secreted
from crypt cells and in pancreatic, and salivary secretions. The inability to absorb solute will also cause some loss in efficiency of water absorption. If water absorption efficiency in the 20‐kg dog drops from nearly 99% to 90% due to enteritis in a section of the intestine, fecal water content will increase from 35 to 275 mL, which will create a very watery feces. If damage to the gastrointestinal tract is restricted to the colon and no water is absorbed by the colon, fecal water content will only increase by the 365 mL the colon is expected to absorb. However, damage to large segments of the small intestine can cause loss of up to 2650 mL water.
Bacterial and viral infections can not only destroy villous absorptive cells, they can initiate inflammation reactions that generally cause hypersecretion by the crypt and goblet cells in an attempt to flush away the offending pathogens. This can greatly increase the amount of fluid lost with the feces. Bacterial infections tend to be fairly localized and cause local damage to either the small or large intestine. Rotaviruses attack the cells at the tips of villi causing some malabsorption. Coronaviruses, such as the causative agents of transmissible gastroenteritis and porcine epidemic diarrhea, kill villous entrocytes the full length of the villus and are therefore much more severe infections. Parvoviruses causing canine parvo and feline panleukopenia attack rapidly dividing cells – in the intestine these are the crypt
Table 44.2 Water secretion and absorption in a 20‐kg dog.
Into lumen (ingested and secreted)
Out of lumen (absorbed)
Food and drink 600 mL —Salivary glands 300 mL —Gastric juice 600 mL —Bile 300 mL —Pancreatic secretions
600 mL —
Small intestine 600 mL 2650 mLColon 50 mL 365 mLTotals 3050 mL 3015 mL = 35 mL
in feces
Table 44.3 Water secretion and absorption in a 500‐kg horse.
Into lumen (ingested and secreted)
Out of lumen (absorbed)
Food and drink 40 L —Salivary glands 30 L —Gastric juice 11 L —Bile 5 L —Pancreatic secretions
10 L —
Small intestine 47 L 79 LColon 1 L 57.5 LTotals 144 L 136.5 L = 7.5 L
in feces
Alimentoyagua
SalivaJugogástrico
BilisSecrecionespancreáticas
Intestinodelgado
ColonTotal
enheces
AguaIngeridaysecretadahacialaluz
Aguaabsorbidadelaluz
Balancedeaguaeneltubodigestivodeunperrode20kgdepeso
20%
20%
20%
10%
88%12%

496 Section VII: Digestion, Absorption, and MetabolismSe
ctio
n V
II: D
iges
tion
, A
bsor
ptio
n, a
nd M
etab
olis
m
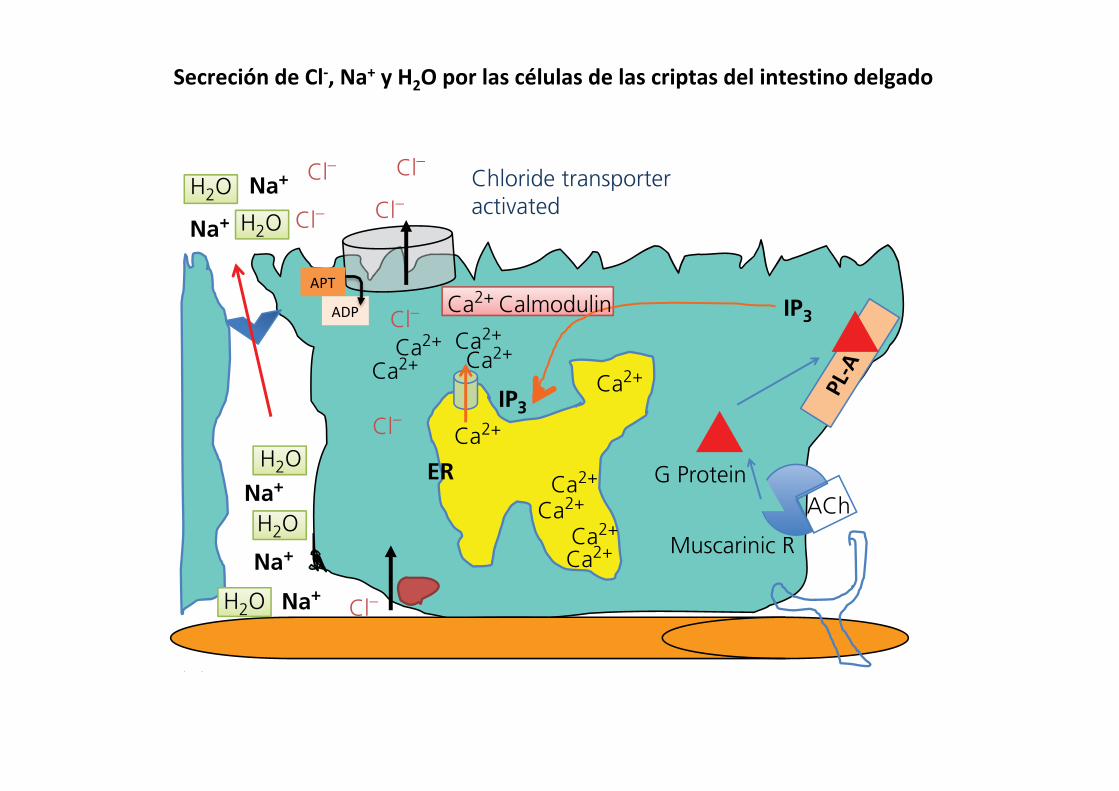
receptors on the basolateral membrane of the crypt cells. These receptors are G protein‐coupled receptors linked to phospho-lipase A, so on activation the intracellular concentration of inositol trisphosphate (IP3) rises. IP3 acts on the membrane of internal cell organelles that store calcium, such as the endoplasmic reticulum, and causes calcium channels to open in the the membrane. This releases Ca2+ to the cytosol of the cell where it becomes bound to calmodulin, an important cell regulatory protein, and causes it to become activated. The Ca2+–calmodulin complex then interacts with a Cl– channel pump protein at the apical membrane and causes it to open. It also causes an ATP to donate the energy of a phosphate bond to supply the energy needed to transport the chloride from the inside of the cell, which has a relatively low Cl– concentration (<30 mmol/L), to the lumen of the crypt where Cl– concentration
is substantially higher. This Cl– channel pump is also known as the cystic fibrosis transmembrane conductance regulator pro-tein. The Cl– pumped into the lumen is rapidly replaced by entry of a chloride into the cell from the extracellular fluid ([Cl–] ~105 mmol/L) across the basolateral membrane, alone or cotransported with Na+ or K+. Once chloride has been secreted into the lumen, the negative charges of the Cl– ions in the lumen of the crypt, together with the high Na+ concentration in the extracellular fluid, cause Na+ ions to move from the extracellular fluid to the lumen across the tight cell junctions separating adja-cent crypt enterocytes. Water then follows the solute into the lumen using water channels in the tight cell junctions. In this way, the secretory activity of the crypt cells is coordinated to occur only at the time the villous cells need sodium ions to accomplish absorptive activities.
ACh
AChACh
ACh
Cl–
Cl–
Cl–
Cl–
Cl– Ca2+
Ca2+
Ca2+ Ca2+
Ca2+
Ca2+ Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Muscarinic R
G Protein
CalmodulinCalmodulin
Inactive chloridetransporter
Na+
Na+
ER
ER ER
Na+
Na+
Na+
H2O
H2O
Cl– Cl–
Cl–Cl–
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Na+
Na+
Na+
Na+
Na+
Cl–
Na+
Na+
Na+
Na+
Na+
(A)
Muscarinic R
G Protein
IP3
IP3
CalmodulinCalmodulin
(B)
Chloride transporteractivated
Chloride transporteractivated
Ca2+ Calmodulin Ca2+ Calmodulin
Muscarinic R
G Protein
IP3 IP3
IP3
(C)
Cl–
Muscarinic R
G Protein
IP3
(D)
H2O
H2O
H2O
H2O
H2O
H2O
H2O
H2O Cl–
Cl–
Cl– Ca2+
Ca2+ Ca2+
Ca2+
Ca2+
Ca2+
Ca2+ Ca2+Ca2+
Ca2+
Ca2+
ER
H2O
H2O
H2O
H2O
H2O
Cl–
Cl–
Cl–
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Na+
Na+
Na+
Na+
Na+
H2O
H2O
H2O
H2O
H2O Cl–
Cl–
Cl– Cl–
Cl–
Cl–
ACh
Figure 43.15 Crypt secretion of chloride, sodium, and water is normally controlled by vagus parasympathetic innervation. (A) The vagus responds to stretch or osmotic changes in the intestine and releases acetylcholine (ACh). The G protein‐coupled muscarinic receptor resides in the basolateral cell membrane. (B) Activation of muscarinic receptor stimulates G‐protein activation of phospholipase A (PL‐A), which catalyzes production of inositol trisphosphate (IP3). (C) The IP3 moves to the endoplasmic reticulum (ER) and binds to an IP3 receptor causing a Ca2+ channel to open in the ER membrane. (D) Ca2+ binds to calmodulin and the Ca2+–calmodulin complex activates the chloride channel to become active. Cl– is actively pumped out of the cell into the lumen at the expense of ATP. Sodium follows through the tight junction between cells to maintain electroneutrality. Water will also cross the tight junction, pulled by the osmotic gradient created by Cl– and Na+ in the lumen.
SecrecióndeCl-,Na+yH2Oporlascélulasdelascriptasdelintestinodelgado
APT
ADP
496 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
receptors on the basolateral membrane of the crypt cells. These receptors are G protein‐coupled receptors linked to phospho-lipase A, so on activation the intracellular concentration of inositol trisphosphate (IP3) rises. IP3 acts on the membrane of internal cell organelles that store calcium, such as the endoplasmic reticulum, and causes calcium channels to open in the the membrane. This releases Ca2+ to the cytosol of the cell where it becomes bound to calmodulin, an important cell regulatory protein, and causes it to become activated. The Ca2+–calmodulin complex then interacts with a Cl– channel pump protein at the apical membrane and causes it to open. It also causes an ATP to donate the energy of a phosphate bond to supply the energy needed to transport the chloride from the inside of the cell, which has a relatively low Cl– concentration (<30 mmol/L), to the lumen of the crypt where Cl– concentration
is substantially higher. This Cl– channel pump is also known as the cystic fibrosis transmembrane conductance regulator pro-tein. The Cl– pumped into the lumen is rapidly replaced by entry of a chloride into the cell from the extracellular fluid ([Cl–] ~105 mmol/L) across the basolateral membrane, alone or cotransported with Na+ or K+. Once chloride has been secreted into the lumen, the negative charges of the Cl– ions in the lumen of the crypt, together with the high Na+ concentration in the extracellular fluid, cause Na+ ions to move from the extracellular fluid to the lumen across the tight cell junctions separating adja-cent crypt enterocytes. Water then follows the solute into the lumen using water channels in the tight cell junctions. In this way, the secretory activity of the crypt cells is coordinated to occur only at the time the villous cells need sodium ions to accomplish absorptive activities.
ACh
AChACh
ACh
Cl–
Cl–
Cl–
Cl–
Cl– Ca2+
Ca2+
Ca2+ Ca2+
Ca2+
Ca2+ Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Muscarinic R
G Protein
CalmodulinCalmodulin
Inactive chloridetransporter
Na+
Na+
ER
ER ER
Na+
Na+
Na+
H2O
H2O
Cl– Cl–
Cl–Cl–
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Na+
Na+
Na+
Na+
Na+
Cl–
Na+
Na+
Na+
Na+
Na+
(A)
Muscarinic R
G Protein
IP3
IP3
CalmodulinCalmodulin
(B)
Chloride transporteractivated
Chloride transporteractivated
Ca2+ Calmodulin Ca2+ Calmodulin
Muscarinic R
G Protein
IP3 IP3
IP3
(C)
Cl–
Muscarinic R
G Protein
IP3
(D)
H2O
H2O
H2O
H2O
H2O
H2O
H2O
H2O Cl–
Cl–
Cl– Ca2+
Ca2+ Ca2+
Ca2+
Ca2+
Ca2+
Ca2+ Ca2+Ca2+
Ca2+
Ca2+
ER
H2O
H2O
H2O
H2O
H2O
Cl–
Cl–
Cl–
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Na+
Na+
Na+
Na+
Na+
H2O
H2O
H2O
H2O
H2O Cl–
Cl–
Cl– Cl–
Cl–
Cl–
ACh
Figure 43.15 Crypt secretion of chloride, sodium, and water is normally controlled by vagus parasympathetic innervation. (A) The vagus responds to stretch or osmotic changes in the intestine and releases acetylcholine (ACh). The G protein‐coupled muscarinic receptor resides in the basolateral cell membrane. (B) Activation of muscarinic receptor stimulates G‐protein activation of phospholipase A (PL‐A), which catalyzes production of inositol trisphosphate (IP3). (C) The IP3 moves to the endoplasmic reticulum (ER) and binds to an IP3 receptor causing a Ca2+ channel to open in the ER membrane. (D) Ca2+ binds to calmodulin and the Ca2+–calmodulin complex activates the chloride channel to become active. Cl– is actively pumped out of the cell into the lumen at the expense of ATP. Sodium follows through the tight junction between cells to maintain electroneutrality. Water will also cross the tight junction, pulled by the osmotic gradient created by Cl– and Na+ in the lumen.
SecrecióndeCl-,Na+yH2Oporlascélulasdelascriptasdelintestinodelgado
APT
ADP
Chapter 43: Secretory Activities of the Gastrointestinal Tract 497
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
Crypt cell secretion as a response to inflammation or pathogens
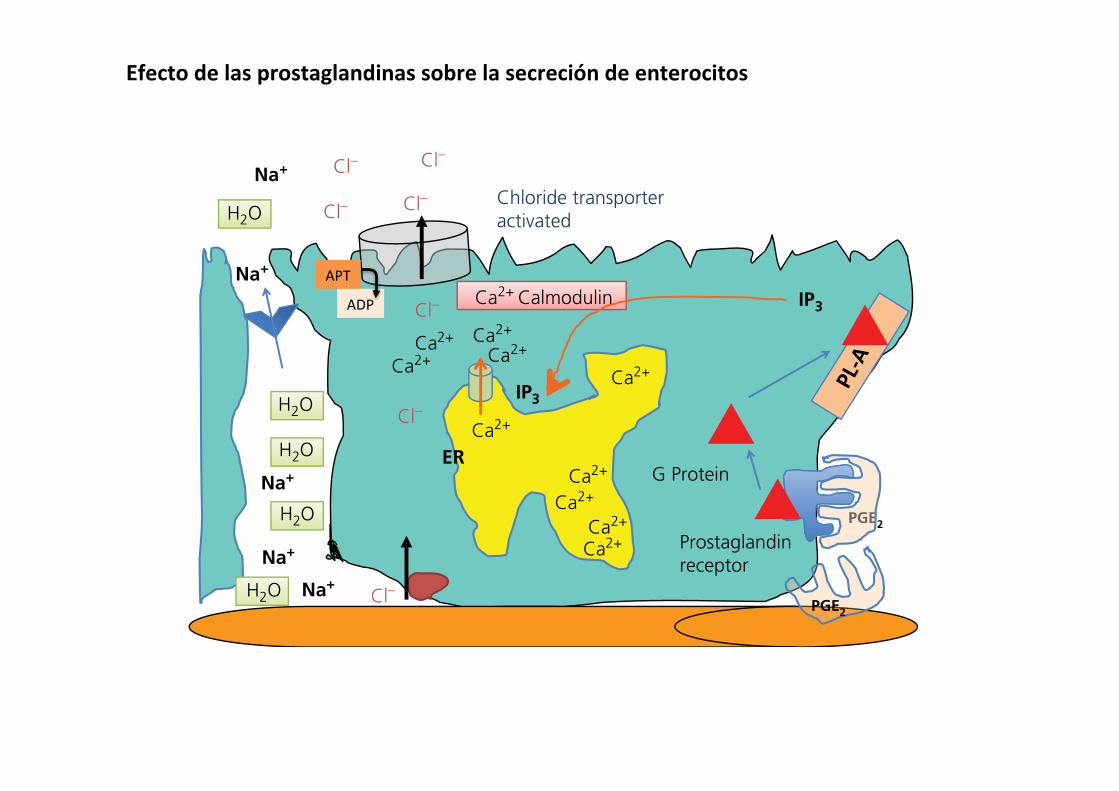
Certain toxins, pathogens, and poisons can cause damage to cells in an area of the intestine. The damaged tissues respond by producing and secreting prostaglandins, primarily PGE2 and PGI2. The prostaglandins diffuse through the lamina propria to reach crypt cells. Crypt enterocytes possess receptors for prosta-glandin on their basolateral membrane. These are G protein‐coupled receptors link to phospholipase A (Figure 43.16). When prostaglandin binds to its receptor, it causes intracellular con-centrations of IP3 to increase in the cytosol of the crypt entero-cyte. This causes Ca2+ channels on the endoplasmic reticulum to open and Ca2+ floods the cytosol. Ca2+–calmodulin complexes form and interact with the apical membrane chloride pump and Cl– is actively pumped out of the cell into the lumen. Extracellular fluid Na+ and water move across the tight junction to follow the Cl– out into the lumen. This action brings large volumes of fluid into the crypt and to surrounding villi to flush the offending toxin away from the area.
Inflammation in a segment of the intestine can also activate crypt secretion activity, presumably to help flush a pathogenic substance away from an area of inflammation. For example, lymphocytes that have become activated by the presence of some “pathogen‐associated molecular pattern” can respond by producing a variety of cytokines. Cytokines such as tumor necrosis factor (TNF)‐α, interleukins, and interferons bind to their respective receptors at the base of the crypt cells and acti-vate adenylyl cyclase or guanylate cyclase. The resulting cyclic AMP or cyclic GMP causes Ca2+ ions to leave intracellular stores and bind to calmodulin. This complex then causes the chloride channel pump to be activated, driving Cl– into the lumen with Na+ and water following through the tight junctions. A slightly different way of stimulating crypt chloride secretion is provided by the action of serotonin (Figure 43.17). Serotonin can be released from enteroendocrine cells in the crypt by the presence of toxins or bacterial cell walls in the lumen of the crypt. The released serotonin diffuses through the lamina propria to
activate nearby crypt cells in a paracrine fashion. Serotonin binds to its receptor, which is linked to a Ca2+ channel in the basolateral cell membrane. The Ca2+ channel opens and extra-cellular Ca2+ floods the cytosol. Again, the Ca2+–calmodulin complex forms and binds to the chloride channel pump, acti-vating secretion of Cl– into the lumen and extracellular Na+ and water cross the tight junction to follow the Cl– into the lumen.
Receptors for all the factors discussed (prostaglandins, cyto-kines, and serotonin associated with cell damage and inflamma-tion) can also be found on goblet cells in the crypts and villi. They respond to these substances by markedly increasing mucus secretion, thought to be a response for flushing away the offend-ing material and coating it with mucus so it will not be as likely to reach the mucosal cells.
Secretory diarrhea caused by bacterial enterotoxinsThe crypt cell secretion activities described so far have been localized to small areas of the intestine that might require Na+ for absorption of sugars and amino acids (described in Chapter 44) or which might use the secretions to flush away pathogens. However, certain bacteria produce toxins that can hijack the normal crypt cell secretion process and cause wide-spread uncontrolled activation of crypt cell secretion. The clas-sic example of this is cholera toxin produced by Vibrio cholerae ingested with contaminated water. The bacteria produce a toxin that is released into the lumen of the small intestine. The cholera toxin binds to proteins (receptors?) at the apical membrane of the crypt enterocyte (Figure 43.18). It is not known why these receptors for cholera toxin exist – it seems logical that there is some natural compound found in the lumen they recognize but none have been identified. Once cholera toxin binds this apical membrane protein, it stimulates activation of guanylate cyclase. Cyclic GMP levels rise inside the cell, causing the Ca2+ channels on the endoplasmic reticulum to open and Ca2+ ions flood the cytosol. This permits Ca2+–calmodulin complexes to form and activate the chloride channel pump. Cl– is secreted into the
PGE2
Chloride transporteractivated
Prostaglandinreceptor
G Protein
IP3
IP3
PGE
PGE2Cl–
Cl–
Cl–
Cl– Cl–
Cl– Cl–
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+ Ca2+
Ca2+
Na+
Na+
Na+ER
Na+
Na+
Ca2+ Calmodulin
H2O
H2O
H2O
H2O
H2O
Figure 43.16 (A) Prostaglandin E (PGE2), produced in response to damage or reactive oxygen species in the area around the crypt cell, binds to a G protein‐coupled PGE2 receptor. This results in activation of phospholipase A (PL‐A) and production of inositol trisphosphate (IP3). The IP3 causes opening of Ca+2 channels in the endoplasmic reticulum (ER) resulting in formation of the Ca2+–calmodulin complex and activation of the apical membrane chloride transporter. This same mechanism can be used by tumor necrosis factors, interleukins, and other cytokines to initiate secretory activity that might help flush away harmful compounds or bacteria. This same mechanism works in goblet cells to stimulate mucus secretion.
Efectodelasprostaglandinassobrelasecrecióndeenterocitos
APT
ADP

498 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
lumen and Na+ and water follow. These toxins spread throughout the intestinal tract and activate huge numbers of crypt cells for a prolonged period. To make matters worse, the toxin can also bind to cholera toxin “receptors” on the villous absorptive enterocytes. Again this causes cyclic GMP production and a rise in intracellular Ca2+ and formation of Ca2+–calmodulin complexes. However, in this instance the Ca2+–calmodulin com-plex binds to the Na+/Cl– cotransporter used to absorb lumen Na+ and Cl– across the apical membrane of the villous cells. Shutting down this mechanism for absorption of Na+ and Cl– also reduces the amount of water that can be absorbed. The end result is that crypts are in a state of tremendous hypersecretion and the villous cells have a reduced capacity to absorb, causing a massive loss of fluids and electrolytes with the feces.
In veterinary medicine, the offending bacteria producing entertoxins are likely to be certain strains of Escherichia coli. At least two enterotoxins have been described. One is heat stable (ST toxin) and when it binds to receptors on the apical mem-brane of crypt cells (and villous cells) it activates cyclic GMP production just like cholera toxin, causing a similar severe watery diarrhea. The other enterotoxin produced by a different strain of E. coli is a heat‐labile toxin (LT). This toxin binds to its receptor on the apical surface of enterocytes (crypt and vil-lus) and activates adenylyl cyclase, causing intracellular cyclic AMP levels to rise, triggering increased secretion of Cl– by crypt enterocytes and decreased absorption of Na+ and Cl– by villous enterocytes (Figure 43.19). A notable difference is that when the LT toxin binds to the receptor protein that recognizes
Cl–
Cl–
Cl–
Cl–
Cl– Cl–
Cl–
Cl–
Cl–
Cl–
Cl–
Cl–
-
EnteroendocrinecellCa2+
Ca2+
Ca2+
Ca2+
Ca2+ Ca2+
Ca2+Ca2+
Ca2+ Ca2+ Ca2+
Ca2+ Ca2+
Ca2+
Calmodulin
(A)
(C) (D)
SR
Serotonin
Release Serotoninto have paracrineaction
Release Serotoninto have paracrineaction
Release Serotoninto have paracrineaction
Toxins
Calmodulin
(B)
Toxins
Calmodulin
Toxins
Chloride transporteractivated
Ca2+ calmodulin
Cl–
Cl–
Cl–
Cl– Cl–
Cl–
Cl– Cl–
Cl–
Cl–
Cl–
SRSR
Cl–
Cl–
Cl–
Cl–
Cl–
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+ Ca2+Ca2+
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+ Ca2+Ca2+
Ca2+
Ca2+Ca2+
Ca2+Ca2+ Ca2+
Ca2+Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+
Ca2+ Ca2+
Ca2+Ca2+ Ca2+
Ca2+
Ca2+
Ca2+
Figure 43.17 (A) An enteroendocrine cell that secretes serotonin when activated neighbors a crypt cell. (B) A bacterial toxin activates receptors at the apical membrane of the enteroendocrine cell causing serotonin vesicles to fuse with the basolateral membrane and secrete serotonin into the lamia propria, where it diffuses toward a serotonin receptor (SR) in the basolateral membrane of the crypt cell. (C) Binding of serotonin causes a conformational change in the receptor allowing it to open Ca2+ channels in the basolateral membrane. Extracellular Ca2+ enters the cell. (D) The rise in cytosolic Ca2+ results in formation of the Ca2+–calmodulin complex and activation of the apical membrane chloride transporter. Sodium and water follow the chloride into the lumen across the tight junctions.
AccióndetoxinasomucopolisacáridosdeparedesbacterianossobrelasecrecióndeCl-,Na+yaguadelosenterocitos
APT
ADP

Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
Figure 43.18 (A) Vibrio cholerae secretes cholera toxin within the lumen of the intestine. A G protein‐coupled receptor that responds to this toxin resides in the apical membrane of the cell. (B) On binding to the receptor, the G protein activates guanylyl cyclase (GC). The GC converts guanosine triphosphate (GTP) to cyclic guanosine monophosphate (cyclic GMP). (C) The cyclic GMP activates kinases and pathways that cause Ca2+ channels in the endoplasmic reticulum to open. This results in formation of the Ca2+–calmodulin complex and activation of the apical membrane chloride transporter. Sodium and water follow the chloride into the lumen across the tight junctions.
G Protein
Calmodulin
Na+
Na+
Na+
Na+
Na+
H2O
H2O
GC
Choleratoxin
(A)
Cl–
Cl–
Cl–
Cl–
Ca2+
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+ Ca2+
Ca2+Ca2+
H2O
H2O
H2O
Na+
Na+
Na+
Na+
Na+
Choleratoxin
G Protein
Calmodulin
GC
GTPcGMP
(B)
Cl–
Cl–
Cl–
Cl–
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+ Ca2+Ca2+
Ca2+
Ca2+
H2O
H2O
H2O
H2O
H2O
Na+
Na+
Na+
Na+
Na+
Cl -
G Protein
(C)
GC
GTPcycGMP
Chloridetransporteractivated
Cl -
Ca2+ Calmodulin
Cl–Cl–
Cl–
Cl–
Cl–
Cl– Cl–
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+Ca2+
Ca2+ Ca2+Ca2+
Ca2+
Ca2+
H2O
H2O
H2O
H2O
H2O
Figure 43.19 (A) Certain strains of Escherichia coli secrete enterotoxin (LT toxin) within the lumen of the intestine. A G protein‐coupled receptor responds to this toxin in the apical membrane of the crypt (and villus) cells. (B) On binding to the receptor, the G protein activates adenylyl cyclase (AC). The AC converts adenosine triphosphate (ATP) to cyclic adenosine monophosphate (cAMP). (C) The cAMP activates kinases and pathways that cause Ca2+ channels in the endoplasmic reticulum to open. This results in formation of the Ca2+–calmodulin complex and activation of the apical membrane chloride transporter. Sodium and water follow the chloride into the lumen across the tight junctions. In villous cells, the toxin’s action is similar but results in blockade of the Na+/Cl–absorption cotransporter in the apical membrane.
G Protein
Calmodulin
(A)
AC
E. coli secretes LT enterotoxin
Na+
Na+
Na+
Na+
Na+
Cl–
Cl–
Cl–
Cl–
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+ Ca2+
Ca2+
Ca2+
H2O
H2O
H2O
H2O
H2O
(B)
G Protein
Calmodulin
AC
ATPcycAMP
LT Enterotoxin
Na+
Na+
Na+
Na+
Na+
Ca2+
Ca2+ Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+
Cl–
Cl–
Cl–
Cl–
H2O
H2O
H2O
H2O
H2O
(C)
Cl-
G Protein
AC
ATPcycAMP
Chloride transporter activated
CClClCl-
LT Enterotoxin
Na+
Na+
Na+
Na+
Na+
Cl– Cl–
Cl–
Cl–
Cl–
Cl–
Cl–
Ca2+
Ca2+Calmodulin
Ca2+
Ca2+Ca2+
Ca2+ Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+Ca2+
Ca2+
Ca2+
H2O
H2O
H2O
H2O
H2OSe
ctio
n V
II: D
iges
tion
, A
bsor
ptio
n, a
nd M
etab
olis
m
Figure 43.18 (A) Vibrio cholerae secretes cholera toxin within the lumen of the intestine. A G protein‐coupled receptor that responds to this toxin resides in the apical membrane of the cell. (B) On binding to the receptor, the G protein activates guanylyl cyclase (GC). The GC converts guanosine triphosphate (GTP) to cyclic guanosine monophosphate (cyclic GMP). (C) The cyclic GMP activates kinases and pathways that cause Ca2+ channels in the endoplasmic reticulum to open. This results in formation of the Ca2+–calmodulin complex and activation of the apical membrane chloride transporter. Sodium and water follow the chloride into the lumen across the tight junctions.
G Protein
Calmodulin
Na+
Na+
Na+
Na+
Na+
H2O
H2O
GC
Choleratoxin
(A)
Cl–
Cl–
Cl–
Cl–
Ca2+
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+ Ca2+
Ca2+Ca2+
H2O
H2O
H2O
Na+
Na+
Na+
Na+
Na+
Choleratoxin
G Protein
Calmodulin
GC
GTPcGMP
(B)
Cl–
Cl–
Cl–
Cl–
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+ Ca2+Ca2+
Ca2+
Ca2+
H2O
H2O
H2O
H2O
H2O
Na+
Na+
Na+
Na+
Na+
Cl -
G Protein
(C)
GC
GTPcycGMP
Chloridetransporteractivated
Cl -
Ca2+ Calmodulin
Cl–Cl–
Cl–
Cl–
Cl–
Cl– Cl–
Ca2+
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+Ca2+
Ca2+ Ca2+Ca2+
Ca2+
Ca2+
H2O
H2O
H2O
H2O
H2O
Figure 43.19 (A) Certain strains of Escherichia coli secrete enterotoxin (LT toxin) within the lumen of the intestine. A G protein‐coupled receptor responds to this toxin in the apical membrane of the crypt (and villus) cells. (B) On binding to the receptor, the G protein activates adenylyl cyclase (AC). The AC converts adenosine triphosphate (ATP) to cyclic adenosine monophosphate (cAMP). (C) The cAMP activates kinases and pathways that cause Ca2+ channels in the endoplasmic reticulum to open. This results in formation of the Ca2+–calmodulin complex and activation of the apical membrane chloride transporter. Sodium and water follow the chloride into the lumen across the tight junctions. In villous cells, the toxin’s action is similar but results in blockade of the Na+/Cl–absorption cotransporter in the apical membrane.
G Protein
Calmodulin
(A)
AC
E. coli secretes LT enterotoxin
Na+
Na+
Na+
Na+
Na+
Cl–
Cl–
Cl–
Cl–
Ca2+Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+ Ca2+
Ca2+
Ca2+
H2O
H2O
H2O
H2O
H2O
(B)
G Protein
Calmodulin
AC
ATPcycAMP
LT Enterotoxin
Na+
Na+
Na+
Na+
Na+
Ca2+
Ca2+ Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+
Cl–
Cl–
Cl–
Cl–
H2O
H2O
H2O
H2O
H2O
(C)
Cl-
G Protein
AC
ATPcycAMP
Chloride transporter activated
CClClCl-
LT Enterotoxin
Na+
Na+
Na+
Na+
Na+
Cl– Cl–
Cl–
Cl–
Cl–
Cl–
Cl–
Ca2+
Ca2+Calmodulin
Ca2+
Ca2+Ca2+
Ca2+ Ca2+
Ca2+
Ca2+
Ca2+
Ca2+Ca2+
Ca2+Ca2+
Ca2+
Ca2+
H2O
H2O
H2O
H2O
H2O
E.ColiToxinatermolábil
E.ColiToxinatermoestable
CanaldeCl-activado CanaldeCl-
activado
AccióndetoxinasodeE.ColisobrelasecrecióndeCl-,Na+yaguadelosenterocitos
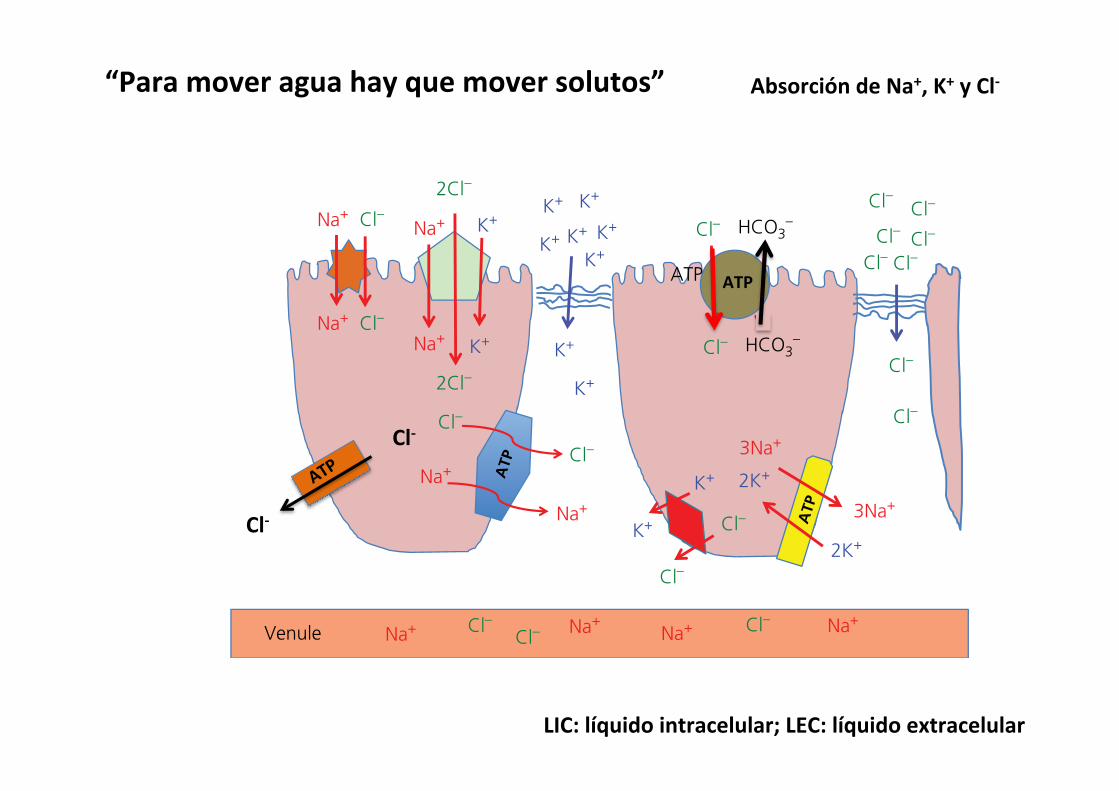
508 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
Basolateral membrane transportChloride concentration in the extracellular fluid is about 102–108 mmol/L. Inside the enterocyte the Cl– concentration might get as high as 30 mmol/L. Cl– will move to the extracel-lular fluid against its concentration gradient but with its electrical gradient.1 Cl–/K+ cotransporter: K+ will be moving into the extracellular
fluid down its concentration gradient but against its electrical gradient. The combined force of K+ moving down its concentration gradient and Cl– moving down its electrical gradient permits both molecules to overcome the forces resisting this movement (Figure 44.7).
2 Cl– pump: chloride can be actively pumped across the baso-lateral membrane at the expense of an ATP. This mechanism is used primarily in the lower intestine (Figure 44.7).
3 Na+/Cl– pump: sodium and chloride can be actively pumped across the basolateral membrane at the expense of an ATP. This mechanism is used primarily in the lower intestine (Figure 44.7).
Paracellular absorption into the extracellular fluidWhen Cl– concentration is very high, as in the uppermost duo-denum (110–120 mmol/L), the concentration gradient between the lumen and the extracellular fluid (102–108 mmol/L) will
allow Cl– to pass through the tight junctions between adjoining enterocytes directly into the extracellular fluid. This mechanism is important in the first few centimeters of the duodenum and quickly removes a large amount of chloride from the chyme; this action also increases the pH of the digesta in the duodenum (Figure 44.7).
Potassium
Paracellular K+ transportThe bulk of K+ absorption occurs across the tight junctions and K+ moves between cells directly into the extracellular fluid, particularly in the lower small intestine. K+ concentration in extracellular fluid is low (4–6 mmol/L) while the concentration of K+ in the lumen can be many fold higher. The concentration of K+ in the lumen actually increases as the digesta moves down
Table 44.1 Major mechanisms utilized for absorption of electrolytes, hexoses, amino acids, and water in various segments of the intestinal tract.
Mechanism Duodenum Upper jejunum Middle jejunum Lower jejunum Ileum Colon
Na+/Cl– cotransporter + + + + ++ +++Na+/hexose cotransporter ++++++ ++++ + + – –Na+/amino acid cotransporter ++++++ ++++ + + – –Cl–/HCO3
– exchange – – – – ++ +++Paracellular Cl– absorption ++++ – – – – –Paracellular K+ absorption – – – – + +++Water +++ +++ + + ++ +
Venule
Na+Na+Cl–
Na+
Na+
Na+
Na+
Na+ Na+Na+ Na+
Cl–
Cl–
Cl–
Cl–
Cl–Cl–Cl– Cl–
Cl–Cl–
Cl–
Cl–
Cl–
Cl–
Cl–Cl–
Cl–
2Cl–
2Cl–
HCO3–
Cl– HCO3–
ATP
3Na+
3Na+
K+
K+
K+ K+ K+
K+
K+
K+
K+
K+ 2K+
2K+
K+ K+
ATP
ATP
Figure 44.7 Major electrolyte absorption mechanisms. The apical membrane has a Na+/Cl– cotransporter, a Na+/K+/2Cl– cotransporter, and a Cl–/HCO3
– ATPase pump to bring electrolytes from the lumen into the cytosol. The basolateral membrane uses a Na+/Cl– ATPase pump, a K+/Cl– cotransporter, and the 3Na+/2K+ exchange ATPase pump (the electrogenic pump) to move electrolytes from the cytosol into the extracellular fluid. In the lower small intestine and colon most of the K+ is absorbed paracellularly. In the uppermost duodenum Cl– can also be absorbed paracellularly.
1 How do potassium atoms cross from the lumen of the gut to the interior of the villous cell?
2 How do potassium atoms cross from the interior of the cell to the extracellular fluid?
3 Where is paracellular absorption of potassium most likely to occur and why?
Cl-
Cl-
ATP
AbsorcióndeNa+,K+yCl-
LIC:líquidointracelular;LEC:líquidoextracelular
“Paramoveraguahayquemoversolutos”

516 Section VII: Digestion, Absorption, and Metabolism
Sect
ion
VII:
Dig
esti
on,
Abs
orpt
ion,
and
Met
abol
ism
mammary gland. The remnant of a VLDL is called a low‐density lipoprotein (LDL). It still contains a large amount of cholesterol and has the unhealthy habit in humans of delivering cholesterol to endothelial cells lining arteries. This is why LDL is often referred to as “bad cholesterol.” The HDL particles released from the liver can actually remove triglyceride and cholesterol from arteries and other peripheral tissues and take these materials to the liver for processing. This property has earned them the moniker of “good cholesterol.”
Absorption of water
Two forces are at work to move water (Figure 44.14): osmotic pressure is generated by the movement of solute from one compartment to another creating an osmotic gradient; hydro-static pressure is a physical force created when water is moved into a confined area. Aquaporin channels exist in the apical and basolateral membranes of enterocytes that allow water to move across the lipid bilayer of the cell membranes to follow the osmotic gradient. These aquaporin channels do not allow any charged ions through them. The key to absorption of water is to absorb solute. Solute is typically absorbed across the apical membrane and crosses the basolateral membrane to enter the lateral space between epithelial cells. This draws water into the lateral space between cells through the aquaporins. As more solute and more water enters this confined space it increases hydrostatic pressure. Distension of the elastic cell membranes of the lateral space also allows the hydrostatic pressure to increase. Under normal circumstances the path of least resistance for this water will be to cross the basement membrane of the fenestrated capillary endothelium to enter the circulation. A critical fact is that the tight junction offers more resistance to flow of water than the fenestrated endothelial basement membrane of the capillary bed. The tight junctions are not impermeable to water, just less permeable than the capillary endothelium. Water chan-nels formed by claudin proteins (not the same as aquaporins) in the tight junction generally resist the flow of water, but this
1 What are aquaporins? Do they allow water to move down or against its osmotic gradient?
2 Why is movement of solute critical for absorption of water?
3 Can water cross the tight junctions? Under what circumstances?
4 How does the flow of blood at the villous tip affect water absorption?
5 Where is the bulk of water absorbed in the digestive tract?
6 Can the colon compensate for malabsorption in the upper intestine? What can it absorb that might help with water absorption?
SoluteNa, Cl, K,sugars,aminoacids
300mosmol
500mosmol
400mosmol
ArterioleVenule
H2OH2O
H2O
H2OH2O
H2O
H2O
H2O
H2O H2O
H2OH2O AA
AA
AA
AA
AAAA
AANa+
Na+
Na+Na+
Na+
Na+ Na+
Na+
Na+
Na+
Na+
Na+Na+ Na+
Na+
AAAA
AA
AA
AA
AA
600mosmol
Capillary bed
1
3
2
H2OH2OH2O
H2OH2OH2O
H2OH2O
H2O
H2O
Figure 44.14 Water absorption. Water is pulled into areas high in solute. (1) In the villous tip the countercurrent flow of arteriole and venule blood causes solute in the descending venule to diffuse into the ascending arteriole to be carried back to the villous tip. The osmolarity at the villous tip can approach 600 mosmol. (2) At the level of individual enterocytes, water utilizes aquaporin to follow its osmotic gradient, created by absorption of solutes such as Na+ and amino acids (AA) into the extracellular fluid through the enterocytes. Aquaporins are specialized water channels that allow water but not charged ions through their channels. (3) As water moves into the lateral spaces, hydrostatic pressure builds up between the cells causing distension and bulging of the cell walls below the tight junction. The hydrostatic pressure then drives the water and solute into the fenestrated capillaries, which have lower resistance to the flow of water than do the tight junctions.
Absorcióndeagua
1)Paraabsorberaguahayqueabsorbersolutos
2)elaguaatraviesalasmembranasapicalyasolateralporcanalesespecíficos(acuaporinas).Efectoosmótico
3)elaguaseacumulaenelespaciolateralentre2enterocitosyaumentalapresiónhidrostática.
4)LauniónestrechaesmásresistentealaumentodelapresiónenesteespacioquelamembranabasalyelaguaatraviesalamembranabasalhaciaelLIC.
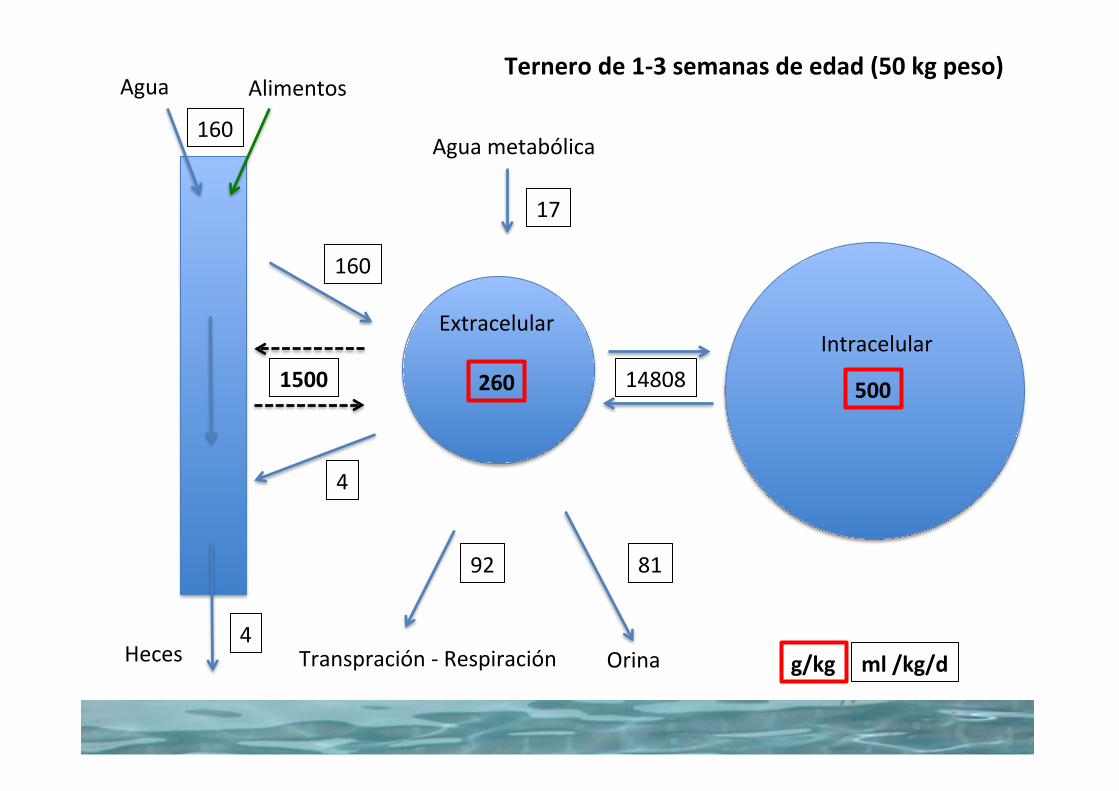
14808
17
8192
160
160
4
4
260 5001500
g/kg ml/kg/dTranspración-Respiración Orina
Aguametabólica
Agua Alimentos
Heces
ExtracelularIntracelular
Ternerode1-3semanasdeedad(50kgpeso)
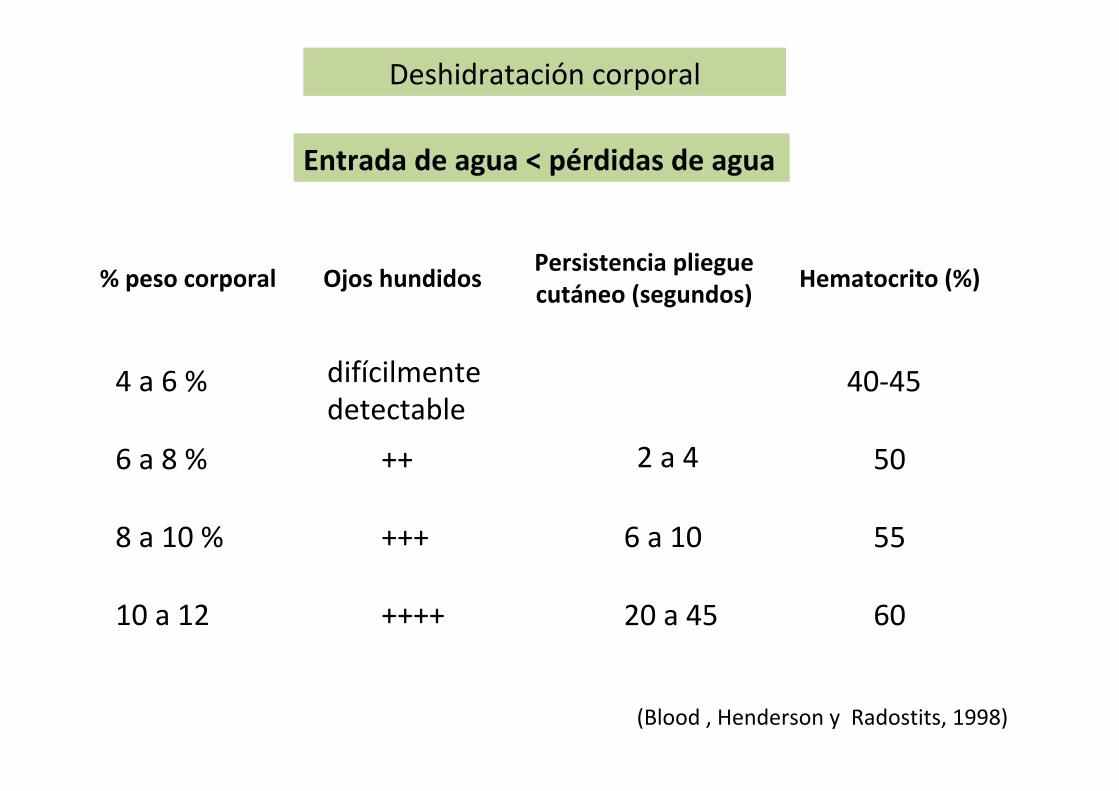
Deshidratacióncorporal
(Blood,HendersonyRadostits,1998)
4a6%
6a8%
8a10%
10a12
difícilmentedetectable
++
+++
++++
2a4
6a10
20a45
40-45
50
55
60
Ojoshundidos Persistenciaplieguecutáneo(segundos) Hematocrito(%)%pesocorporal
Entradadeagua<pérdidasdeagua

Consumodeagua
Presiónosmóticaplasma
Presiónsanguínea
Temperaturacorporal
SecrecióndeADH
AguaenelLEC
Reduccióndeexcrecióndeagualibreenriñón
AguaenelLEC
Hipotálamosed
1000mOsm
100mOsm

RequerimientosMÍNIMOSdeagua
Excreción
-heces=1,6l/kgMSingerida
- orina(@1000mOsm/l)
Termorregulación
Evaporaciónpulmonar
TranspiraciónProductos/tejidosexportados/ables
Leche(900gagua/kg)Feto(2l/día,enelterciofinaldegestación)
Tejidosretenidos/perdidos
(2,5kJ/gagua)
Crecimiento(788gagua/kgtejidomagro)
0,9l/kgMSingerida(bajoNyK+)
2,3l/kgMSingerida(altoNyK+)forrajesenplenocrecimiento
EjemploBovinos
-sensible-insensible
10ml/kgpeso/d(@15C)
15ml/kgpeso/d(@20C)
8ml/kgpeso/d(@10C)
Aguametabólica(5-10%delconsumodeaguaconalimentoydebebida)