capítulo 1media.axon.es/pdf/66768.pdfratones,la mosca de la fruta drosophila melanogaster y el...
TRANSCRIPT

Capítulo 1
Aspectos generales
La neurociencia comprende una amplia gama de interrogantes acerca de có-mo se organizan los sistemas nerviosos y cómo funcionan para generar la con-ducta. Estos cuestionamientos pueden explorarse por medio de las herramientasanalíticas de la genética, la biología molecular y celular , la anatomía y la f isio-logía de los sistemas, la biología conductual y la psicología. El desafío principalpara un estudiante de neurociencia es integrar el conocimiento diverso derivadode estos distintos niveles de análisis en un conocimiento más o menos coheren-te de la estructura y la función encefálicas (deberíamos hacer algunas salv eda-des a esta afirmación ya que muchas preguntas aún permanecen sin respuesta).Muchas de las cuestiones exploradas con éxito se vinculan con el modo en quelas células principales de todo sistema nervioso –neuronas y glía– realizan susfunciones básicas en términos anatómicos, electrofisiológicos y moleculares.Las variedades de neuronas y de células gliales de sostén que se identificaron es-tán reunidas en conjuntos llamados circuitos neurales, y estos circuitos constitu-yen los componentes primarios de los sistemas nerviosos que procesan tipos es-pecíficos de información. Los sistemas neurales comprenden neuronas y circui-tos en algunas localizaciones anatómicas separadas del encéfalo. Estos sistemascumplen una de tres funciones generales. Los sistemas sensiti vos presentan lainformación acerca del estado del organismo y su entorno, los sistemas motoresorganizan y generan acciones y los sistemas asociati vos vinculan los aspectossensitivos y motores del sistema nervioso y aportan las bases para las funciones“de orden superior” como la percepción, la atención, la cognición, las emocio-nes, el pensamiento racional y otras funciones encefálicas complejas que subya-cen en el núcleo del conocimiento de los seres humanos, su historia y su futuro.
Genética, genómica y encéfalo
La secuenciación recién completada del genoma en los seres humanos, losratones, la mosca de la fruta Drosophila melanogaster y el nematodo Caenor-habditis elegans es tal vez el punto de inicio más lógico para estudiar el encé-falo y el resto del sistema nervioso; después de todo, esta información heredi-taria también es el punto de inicio de cada or ganismo individual. La relativafacilidad para obtener, analizar y correlacionar las secuencias de los genes conlas observaciones neurobiológicas propició muchas ideas nuevas sobre la bio-logía básica del sistema nervioso. En forma paralela con los estudios de lossistemas nerviosos normales, el análisis genético de los pedigríes humanoscon distintas enfermedades encefálicas condujo a la sensación generalizada deque pronto será posible conocer y tratar trastornos que durante mucho tiempose consideraron más allá del alcance de la ciencia y la medicina.
1
Estudio del sistema nerviosode los seres humanos y otrosanimales
A693-01.qxd 8/3/06 11:41 AM Page 1

Un gen presenta secuencias de DNA denominadas exones que se transcri-ben en un RN A mensajero y ulteriormente en una proteína. El conjunto deexones que define el transcripto de cualquier gen está flanqueado por secuen-cias reguladoras corriente arriba (o 5’) y corriente abajo (3’) que controlan laexpresión genética. Además, las secuencias entre los exones –llamadas intro-nes– influyen en la transcripción. De los alrededor de 35.000 genes del geno-ma humano, la mayoría se expresa en el encéfalo en desarrollo y el adulto; lomismo sucede con los ratones, las moscas y los vermes, especies que suelenutilizarse en genética moderna (y cada v ez más en neurociencia) (f ig. 1-1).No obstante, muy pocos genes se e xpresan únicamente en las neuronas, loque indica que las células nerviosas comparten la mayor parte de las propie-dades estructurales y funcionales básicas de otras células. En consecuencia,la mayor parte de la información genética “específ ica del encéfalo” debe re-sidir en el resto de las secuencias de ácido nucleico –secuencias re guladorase intrones– que controlan la oportunidad, la cantidad, la variabilidad y la es-pecificidad celular de la expresión genética.
Una de las consecuencias más promisorias de la secuenciación del geno-ma humano fue el reconocimiento de que uno o algunos genes, cuando se al-teran (mutan), pueden explicar ciertos aspectos de las enfermedades neuroló-gicas y psiquiátricas. Antes de la “era posgenómica” (que comenzó luego decompletada la secuenciación del genoma humano), muchas de las enferme-dades encefálicas más devastadoras seguían siendo misteriosas, porque no seconocía con certeza el modo en que se comprometía la biología normal delsistema nervioso. La identif icación de genes correlacionados con trastornoscomo la enfermedad de Huntington, la enfermedad de Parkinson, la enferme-dad de Alzheimer, la depresión mayor y la esquizofrenia aportó un inicio pro-misorio al conocimiento de estos procesos patológicos de una forma muchomás profunda (y al diseño de terapias racionales).
La información genética y genómica aislada no e xplica por completo elmodo en que el encéfalo funciona normalmente o cómo los procesos patoló-gicos interrumpen su función. Para lograr estos objetivos es esencial por igualconocer la biología celular , la anatomía y la f isiología del encéfalo en la sa-lud y la enfermedad.
2 Capítulo 1
Número de genes0 50.00040.00030.00020.00010.000
Ser humano
Ratón
D. melanogaster
C. elegans
Fig. 1-1. Estimaciones del número degenes en el genoma humano y en los ge-nomas del ratón, la mosca de la frutaDrosophila melanogaster y el nematodoCaenorhabditis elegans.
A693-01.qxd 8/3/06 11:41 AM Page 2

Los componentes celulares del sistema nervioso
A comienzos del siglo XIX la célula se reconocía como la unidad fundamen-tal de todos los organismos vivos. Sin embargo, no fue hasta bien entrado el si-glo XX en que los neurocientíf icos concordaron en que el tejido nervioso, aligual que todos los otros órganos, está formado por estas unidades fundamen-tales. La razón principal fue que la primera generación de neurobiólogos “mo-dernos” del siglo XIX tuvo dificultades para distinguir la naturaleza unitaria delas células nerviosas con los microscopios y las técnicas de tinción celular dis-ponibles entonces. Esta deficiencia se vio agravada por las configuraciones ex-traordinariamente complejas y las ramif icaciones extensas de las células ner-viosas individuales, lo que oscurecía aun más su semejanza con las células deconfiguración más sencilla de otros tejidos (f igs. 1-2 a 1-4). Como resultado,algunos biólogos de esa era arribaron a la conclusión de que cada célula ner-viosa estaba conectada a sus v ecinas por ne xos protoplasmáticos, que forma-ban una red continua de células nerviosas, o retículo. La “teoría reticular” de la
Estudio del sistema nervioso de los seres humanos y otros animales 3
Axón
Cuerpocelular
Dendritas
Dendritas
(C) Célula ganglionar retiniana
(F) Células de Purkinje del cerebelo
Axón
Cuerpocelular
(A) Neuronas en el núcleo mesencefálico del núcleo craneal V
Axones
*
*
Cuerposcelulares
(B) Célula bipolar retiniana
DendritasDendritas
Cuerpo celular
Axón
Cuerpocelular
Axón Cuerpo celular
Dendritas
(D) Célula amacrina retiniana
(E) Célula piramidal cortical
* *
Fig. 1-2. Ejemplos de la rica varie-dad de morfologías de las células ner-viosas halladas en el sistema nerviosohumano. Los trazados provienen de cé-lulas nerviosas reales teñidas por im-pregnación con sales de plata (la deno-minada técnica de Golgi, método utili-zado en los estudios clásicos de Golgi yCajal). Los asteriscos indican que elaxón recorre mucho más de lo que semuestra. Obsérvese que algunas célu-las, como la bipolar de la retina, tienenun axón muy corto, y que otras, comola célula amacrina retiniana, carecen deaxón. Los dibujos no están todos en lamisma escala.
A693-01.qxd 8/3/06 11:41 AM Page 3

comunicación de las células nerviosas, propuesta por el neuropatólogo italianoCamillo Golgi (a partir del cual toma su nombre el aparato de Golgi en las cé-lulas), finalmente perdió favoritismo y se reemplazó por la que llegó a conocer-se como la “doctrina de la neurona”. Los autores principales de esta nueva pers-pectiva fueron el neuroanatomista español Santiago Ramón y Cajal y el f isió-logo británico Charles Sherrington.
Los puntos de vista opuestos representados por Golgi y Cajal generaron acomienzos del siglo XX un debate encendido que sentó las bases de la neuro-ciencia moderna. Sobre la base del examen del tejido nervioso teñido con sa-les de plata con el microscopio óptico según un método propuesto por Golgi,Cajal argumentó persuasivamente que las células nerviosas son entidades se-paradas y que se comunican entre ellas por medio de contactos especializa-dos que Sherrington llamó “sinapsis”. El trabajo que enmarcó este debate fuereconocido con el premio Nobel de Fisiología o Medicina en 1906 tanto aGolgi como a Cajal (el premio conjunto sugiere cierta preocupación que aúncontinúa sobre quién estaba en lo correcto, a pesar de las pruebas abrumado-ras de Cajal). El trabajo ulterior de Sherrington y otros, que demostró latransferencia de señales eléctricas en las uniones sinápticas entre las célulasnerviosas, brindaba apoyo firme a la “doctrina de la neurona”, pero quedabael desafío de explicar la autonomía de las neuronas individuales. No fue has-ta el advenimiento de la microscopia electrónica en la década de 1950 en quese resolvieron todas las dudas acerca de la separación de las neuronas. Loscuadros de alta amplificación y alta resolución que pudieron obtenerse con elmicroscopio electrónico establecieron con claridad que las células nerviosasson unidades funcionalmente independientes; estos cuadros también identifi-caron las uniones celulares especializadas que Sherrington había denomina-do sinapsis (véanse figs. 1-3 y 1-4).
Los estudios histológicos de Cajal, Golgi y un conjunto de sucesores con-dujeron a un mayor consenso en que las células del sistema nervioso puedendividirse en dos cate gorías amplias: células nerviosas (o neuronas) y célu-las de sostén llamadas neuroglia (o simplemente glía; véase fig. 1-5). Las cé-lulas nerviosas están especializadas en el señalamiento eléctrico en lar gasdistancias, y el conocimiento de este proceso representa uno de los éxitos másespectaculares de la biología moderna (y es el tema de la unidad I de este li-bro). Por el contrario, las células de sostén no pueden efectuar el señalamien-to eléctrico; no obstante, tienen varias funciones esenciales en el encéfalo endesarrollo y del adulto.
Neuronas
Las neuronas y la glía comparten el complemento de orgánulos hallado entodas las células en el que se incluye el retículo endoplasmático y el aparatode Golgi, las mitocondrias y distintas estructuras vesiculares. Sin embargo, enlas neuronas estos orgánulos a menudo son más sobresalientes en distintas re-giones de la célula. Además de la distribución de los orgánulos y los compo-nentes subcelulares, las neuronas y la glía en cierta medida son diferentes deotras células en las proteínas f ibrilares o tubulares especializadas que consti-tuyen el citoesqueleto (f igs. 1-3 y 1-4). Aunque muchas de estas proteínas–isoformas de actina, tubulina y miosina, así como varias otras– se encuentranen otras células, su organización distinta en las neuronas es fundamental parala estabilidad y la función de las prolongaciones neuronales y las uniones si-nápticas. Los f ilamentos, los túbulos, los motores vesiculares y las proteínas
4 Capítulo 1
A693-01.qxd 8/3/06 11:41 AM Page 4

Estudio del sistema nervioso de los seres humanos y otros animales 5
MitocondriaRetículoendoplasmático
Axones
Ribosomas
Aparatode Golgi
Núcleo
DendritaSoma
(A) (B) Axón (C) Terminaciones sinápticas (botones terminales)
(D) Axones mielínicos
(G) Axón mielínico y nodo de Ranvier(F) Cuerpo de célula neuronal (soma)(E) Dendritas
F
E
B
DG
C
Fig. 1-3. Características principales de las neuronas al observarlas por microscopioóptico y microscopio electrónico. A. Diagrama de células nerviosas y sus partes compo-nentes. B. Segmento inicial del axón (azul) que entra en la v aina de mielina (dorado).C. Botones terminales (azul) cargados con vesículas sinápticas (puntas de flecha) queforman sinapsis (flechas) con una dendrita (púrpura). D. Corte transversal de axones(azul) envainados por las prolongaciones de los oligodendrocitos (dorado). E. Dendritasatípicas (púrpura) de las células piramidales corticales. F. Cuerpos de células nerviosas (púrpura) ocupados por grandes núcleos redondeados.G. Porción del axón mielínico (azul) que ilustra los interv alos entre segmentos adyacen-tes de mielina (dorado) denominados nodos de Ran vier (flechas). (Microfotografías dePeters y col., 1991.)
A693-01.qxd 8/3/06 11:41 AM Page 5

6 Capítulo 1
(A) (B) (C)
(D)
(E) (G)
(F)
(H) (I)
(J) (K)
Fig. 1-4. Disposición característica de loselementos del citoesqueleto en las neuronas.A. El cuerpo celular, los axones y las dendri-tas se distinguen por la distribución de tubu-lina (verde a través de la célula) a diferenciade otros elementos del citoesqueleto –en estecaso, Tau (rojo), una proteína fijadora de mi-crotúbulos que se encuentra sólo en los axo-nes. B. Se muestra aquí la localización nota-blemente distinta de la actina (rojo) en losextremos en crecimiento de las prolongacio-nes axónicas y dendríticas en una neuronacultivada tomada del hipocampo. C. Por elcontrario, en una célula epitelial cultivada, laactina (rojo) está distribuida en fibrillas queocupan la mayor parte del cuerpo celular . D. En células astrogliales en cultivo, la acti-na (rojo) también se ve en haces fibrilares.E. Se observa tubulina (verde) en todo elcuerpo celular y las dendritas de las neuro-nas. F. Aunque la tubulina es un componenteimportante de las dendritas, que se extiendeen las espinas, la cabeza de la espina es ricaen actina (rojo). G. El componente de tubuli-na del citoesqueleto en células no neuronalesestá dispuesto en redes filamentosas.H-K. Las sinapsis tienen una disposicióndistinta de elementos del citoesqueleto,receptores y proteínas de andamiaje. H. Seobservan dos axones (verde; tubulina) deneuronas motoras que emiten cada uno dosramas a cuatro fibras musculares. El rojomuestra el agrupamiento de receptores post-sinápticos (en este caso para el neurotrans-misor acetilcolina). I. Una imagen de mayorpotencia de una sola neurona motora muestrala relación entre el axón (verde) y los recep-tores postsinápticos (rojo). J. Se muestra enverde el espacio extracelular entre el axón ysu músculo diana. K. Se muestra en verde elagrupamiento de proteínas de andamiaje (eneste caso, distrofina) que localizan receptoresy los conectan a otros elementos del citoes-queleto. (A, Cortesía de YN. Jan; B, cortesíade E. Dent y F. Gertler; C, cortesía de D. Ar-neman y C. Otey; D, cortesía de A. Gonzalesy R. Cheney; E, tomado de Sheng, 2003; F, tomado de Matus, 2000; G, cortesía de T.Salmon y col.; H-K, cortesía de R. Sealock.)
A693-01.qxd 8/3/06 11:41 AM Page 6

de andamiaje de las neuronas dirigen el crecimiento de axones y dendritas; eltráfico y el posicionamiento apropiado de componentes de la membrana, or-gánulos y vesículas, y los procesos activos de exocitosis y endocitosis que sub-yacen a la comunicación sináptica. El conocimiento de las formas en que seutilizan estos componentes moleculares para asegurar el correcto desarrollo yfuncionamiento de las neuronas y la glía aún es un enfoque fundamental parala neurobiología moderna.
La organización celular básica de las neuronas se asemeja a la de otras cé-lulas; sin embargo, se distinguen con claridad por la especialización para lacomunicación intercelular. Este atributo se pone de manif iesto en su morfo-logía general en la or ganización específica de los componentes de la mem-brana para el señalamiento eléctrico y en la complejidad de las estructuras yfunciones de los contactos sinápticos entre las neuronas (véanse figs. 1-3 y 1-4). El signo más e vidente de especialización neuronal para la comunicacióna través del señalamiento eléctrico es la ramificación extensa de las neuronas.El aspecto más sobresaliente de esta ramificación en las células nerviosas tí-picas es la arborización compleja de dendritas que surgen del cuerpo de lacélula neuronal (también llamadas ramas dendríticas o prolongaciones den-dríticas). Las dendritas son la diana primaria de las aferencias sinápticas des-de otras neuronas y también se distinguen por su alto contenido en ribosomasy proteínas específicas del citoesqueleto que reflejan su función en la recep-ción y la integración de la información proveniente de otras neuronas. El es-pectro de configuraciones neuronales varía desde una pequeña minoría de cé-lulas que carecen de dendritas hasta neuronas con arborizaciones dendríticasque rivalizan en complejidad con un árbol maduro (véase fig. 1-2). El núme-ro de aferencias que recibe una neurona particular depende de la complejidadde su arborización dendrítica: las células nerviosas que carecen de dendritasestán inervadas (y por lo tanto reciben señales eléctricas) sólo por una o al-gunas otras células nerviosas, mientras que las que presentan dendritas cadavez más elaboradas están inerv adas por una cantidad comparablemente ma-yor de otras neuronas.
Los contactos sinápticos que se hacen sobre las dendritas (y , menos a me-nudo, sobre los cuerpos de las células neuronales) comprenden una elaboraciónespecial del aparato secretorio que se encuentra en la mayoría de las célulasepiteliales polarizadas. En condiciones típicas, la terminación presináptica esinmediatamente adyacente a una especialización postsináptica de la céluladiana (véase fig. 1-3). En la mayoría de las sinapsis no hay continuidad físicaentre estos elementos presinápticos y postsinápticos. En cambio, los compo-nentes presinápticos y postsinápticos se comunican a tra vés de la secreción demoléculas desde la terminación presináptica que se unen a receptores en la es-pecialización postsináptica. Estas moléculas deben atravesar un intervalo de es-pacio e xtracelular entre los elementos presináptico y postsináptico llamadohendidura sináptica. Sin embargo, esta hendidura no es simplemente un es-pacio a atravesar; más bien, es el sitio de las proteínas extracelulares que influ-yen en la difusión, la unión y la degradación de las moléculas secretadas por laterminación presináptica (véase f ig. 1-4). El número de aferencias sinápticasque recibe cada célula nerviosa en el sistema nervioso humano v aría entre 1 y100.000. Este rango refleja un propósito fundamental de las células nerviosas:integrar la información proveniente de otras neuronas. Por lo tanto, el númerode contactos sinápticos pro venientes de diferentes neuronas presinápticas encualquier célula particular es un determinante en especial importante de la fun-ción neuronal.
Estudio del sistema nervioso de los seres humanos y otros animales 7
A693-01.qxd 8/3/06 11:41 AM Page 7

La información transmitida por las sinapsis a las dendritas neuronales esintegrada y “leída” en el origen del axón, porción de la célula nerviosa espe-cializada en la conducción de señales hacia el sitio siguiente de interacciónsináptica (véanse figs. 1-2 y 1-3). El axón es una extensión singular del cuer-po de las células neuronales que puede viajar algunos cientos de micrómetros(µm; habitualmente llamados micrones) o mucho más lejos, según el tipo deneurona y el tamaño de la especie. Además, el axón posee un citoesqueletocaracterístico cuyos elementos son fundamentales para su inte gridad funcio-nal (véase f ig. 1-4). Muchas células nerviosas del encéf alo humano (y deotras especies) tienen axones cuya longitud no es superior a unos milímetros,y algunas no tienen ningún axón.
Los axones relativamente cortos son una característica de las neuronas decircuito local o interneuronas en todo el encéfalo. Sin embargo, los axonesde las neuronas de proyección se extienden hasta dianas distantes. Por ejem-plo, los axones que se encuentran desde la médula espinal humana hasta elpie tienen una longitud aproximada de un metro. El acontecimiento eléctricoque transporta señales por estas distancias se denomina potencial de acción,que es una onda autorregenerada de actividad eléctrica que se propaga desdesu punto de inicio en el cuerpo celular (llamado botón axónico) hacia la ter-minación del axón donde se hacen los contactos sinápticos. Entre las célulasdiana de las neuronas se incluyen otras células nerviosas en el encéf alo, lamédula espinal y los ganglios autónomos, y las células de músculos y glán-dulas de todo el cuerpo.
El proceso químico y eléctrico por el cual la información codif icada por lospotenciales de acción se transmite en los contactos sinápticos hacia la célula si-guiente en una vía se denomina transmisión sináptica. Las terminaciones pre-sinápticas (también llamadas terminaciones sinápticas, terminaciones axónicaso botones terminales) y sus especializaciones postsinápticas son típicamente si-napsis químicas, el tipo más abundante de sinapsis del sistema nervioso. Otrotipo, la sinapsis eléctrica, es mucho más rara (véase cap. 5). Los or gánulos se-cretorios en la terminación presináptica de las sinapsis químicas son vesículassinápticas (véase fig. 1-3), que en general son estructuras esféricas llenas demoléculas de neurotransmisor. El posicionamiento de las v esículas sinápticasen la membrana presináptica y su fusión para iniciar la liberación del neurotrans-misor es regulado por algunas proteínas que se encuentran dentro de la vesículao se asocian con ella. Los neurotransmisores liberados en las v esículas sinápti-cas modifican las propiedades eléctricas de la célula diana al unirse a recepto-res de los neurotransmisores (fig. 1-4), que se ubican fundamentalmente en laespecialización postsináptica.
Por lo tanto, la actividad compleja y coordinada de neurotransmisores, re-ceptores, elementos relacionados del citoesqueleto y moléculas de transduc-ción de señales conforma la base para que las células nerviosas se comuniquenentre ellas, y con las células efectoras en músculos y glándulas.
Células neurogliales
Las células neurogliales –también denominadas gliales o simplementeglía– son muy diferentes de las células nerviosas. Son más numerosas que lasneuronas en el encéfalo, a las que superan en número en una relación tal v ezde 3 a 1. La distinción principal es que las células neurogliales no participandirectamente en las interacciones sinápticas y en el señalamiento eléctrico,aunque sus funciones de sostén ayudan a def inir contactos sinápticos y a
8 Capítulo 1
A693-01.qxd 8/3/06 11:41 AM Page 8

mantener la capacidad de señalización de las neuronas. Aunque las célulasgliales también tienen prolongaciones complejas que se e xtienden desde suscuerpos celulares, por lo general son menos sobresalientes que las ramif ica-ciones neuronales, y no cumplen los mismos propósitos que los axones y lasdendritas (fig. 1-5).
El término glía (de la palabra griega que significa “pegamento”) refleja lasuposición existente en el siglo XIX de que estas células de alguna forma man-tienen unido el sistema nervioso. La palabra sobrevivió, a pesar de la falta depruebas de que unir las células nerviosas entre ellas es una de las muchas fun-ciones de las células gliales. Las funciones bien establecidas de las célulasgliales son: mantener el medio iónico de las células nerviosas, modular la ve-locidad de propagación de las señales nerviosas, modular la acción sinápticaal controlar la captación de neurotransmisores en la hendidura sináptica ocerca de ella, proporcionar un andamiaje para ciertos aspectos del desarrolloneural y ayudar en la recuperación de la lesión neural (o en algunos casos im-pedirla).
Hay tres tipos de células gliales en el sistema nervioso central maduro: as-trocitos, oligodendrocitos y células microgliales (véase fig. 1-5). Los astroci-tos, que están limitados al encéfalo y la médula espinal, tienen prolongacioneslocales elaboradas que brindan a estas células un aspecto estrellado (de ahí elprefijo “astro”). Una función importante de los astrocitos es mantener, de dis-tintas formas, un entorno químico apropiado para el señalamiento neuronal.Los oligodendrocitos, que también están restringidos al sistema nervioso cen-tral, depositan una envoltura laminada y rica en lípidos llamada mielina alre-dedor de algunos axones, pero no de todos. La mielina también tiene efectosimportantes sobre la v elocidad de la transmisión de señales eléctricas (véase
Estudio del sistema nervioso de los seres humanos y otros animales 9
(B) Oligodendrocito(A) Astrocito
Cuerpocelular Prolongaciones
gliales
(D) (E) (F) (G)
(C) Célula microglial(B) Oligodendrocito(A) Astrocito
Cuerpocelular Prolongaciones
gliales
(C) Célula microglial
Fig. 1-5. Variedades de células neu-rogliales. Trazados de un astrocito (A),un oligodendrocito (B) y una célula mi-croglial (C) visualizados utilizando elmétodo de Golgi. Las imágenes se en-cuentran aproximadamente en la mismaescala. D. Astrocitos en cultivo tisular,marcados (rojo) con un anticuerpo con-tra una proteína específica del astrocito.E. Células oligodendrogliales en culti-vo tisular marcado con un anticuerpocontra una proteína específica de la oli-godendroglia. F. Los axones periféricosestán envainados por mielina (rojo mar-cado) excepto en una región distintallamada nodo de Ranvier. La marcaverde indica canales iónicos concentra-dos en el nodo; la marca azul indicauna región que presenta diferencias mo-leculares llamada paranodo. G. Célulasmicrogliales de la médula espinal, mar-cadas con un anticuerpo específico detipo celular. Recuadro: imagen con ma-yor amplificación de una sola célulamicroglial teñida con un marcador se-lectivo de los macrófagos. (A-C, toma-dos de Jones y Cowan, 1983; D, E, cor-tesía de A.-S. LaMantia; F, cortesía deM. Bhat; G, cortesía de A. Light; recua-dro, cortesía de G. Matsushima.)
A693-01.qxd 8/3/06 11:41 AM Page 9

cap. 3). En el sistema nervioso periférico las células que elaboran mielina sedenominan células de Schwann.
Por último, las células microgliales derivan en mayor medida de célulasprecursoras hematopoyéticas (aunque algunas pueden derivar directamente decélulas precursoras neurales). Estas células comparten muchas propiedadescon los macrófagos que se encuentran en otros tejidos,y son fundamentalmen-te células limpiadoras que eliminan los restos celulares de sitios de lesión o derecambio celular normal. Además, la microglia, como sus análogos, los ma-crófagos, secreta moléculas de señalización –sobre todo una amplia gama decitocinas producidas también por células del sistema inmune– que pueden mo-dular la inflamación local e influir en la supervivencia o la muerte celular. Enefecto, algunos neurobiólogos prefieren categorizar la microglia como un tipode macrófago. Luego del daño encefálico, la cantidad de células microglialesen el sitio de la lesión aumenta de forma espectacular . Algunas de estas célu-las proliferan a partir de la microglia e xistente en el encéf alo, mientras queotras provienen de macrófagos que migran al área lesionada y entran en el en-céfalo a través de interrupciones locales en la vasculatura cerebral.
Diversidad celular en el sistema nervioso
Aunque los componentes celulares del sistema nervioso central son simila-res en varios aspectos a los de otros ór ganos, son inusuales por su número e x-traordinario: se estima que el encéf alo humano contiene 100.000 millones deneuronas y varias veces esa cantidad en células de sostén. Lo que es más impor-tante, el sistema nervioso tiene un rango mayor de tipos celulares distintos –yasea categorizados por morfología, identidad molecular o acti vidad fisiológica–que cualquier otro sistema or gánico (un acontecimiento se presume que puedeexplicar por qué en el sistema nervioso se expresan tantos genes diferentes; véa-
10 Capítulo 1
Fig. 1-6. Diversidad estructural en el sistema nervioso demostrada con marcadores celulares y moleculares. Primera hilera: or-ganización celular de diferentes regiones encefálicas demostrada con tinciones de Nissl, que marcan los cuerpos de las célulasnerviosas y gliales. A. Corteza cerebral en el límite entre las áreas visuales primaria y secundaria. B. Bulbos olfatorios. C. Dife-rencias en la densidad celular en las capas corticales cerebrales. D. Neuronas y células gliales individuales teñidas con Nissl conmayor amplificación. Segunda hilera: enfoques clásico y moderno para ver las neuronas individuales con sus prolongaciones. E. Células piramidales corticales marcadas con técnica de Golgi. F. Células de Purkinje cerebelosas marcadas con técnica de Gol-gi. G. Interneurona cortical marcada mediante la inyección intracelular de un colorante fluorescente. H. Neuronas retinianas mar-cadas mediante la inyección intracelular de un colorante fluorescente. Tercera hilera: enfoques celular y molecular para ver cone-xiones y sistemas neurales. I. En la parte superior, un anticuerpo que detecta proteínas sinápticas en el b ulbo olfatorio; en la base,una marca fluorescente muestra la localización de los cuerpos celulares. J. Zonas sinápticas y localización de los cuerpos de lascélulas de Purkinje en la corteza cerebelosa marcadas con anticuerpos específ icos de la sinapsis (verde) y un marcador del cuerpocelular (azul). K. La proyección desde un ojo hasta el cuerpo geniculado lateral en el tálamo, marcada con aminoácidos radiacti-vos (la marca brillante muestra las terminaciones axónicas del ojo en distintas capas del núcleo). L. Mapa de la superficie corpo-ral de una rata en la corteza somatosensiti va, que se muestra con un marcador que distingue zonas de mayor densidad de sinapsisy actividad metabólica. Cuarta hilera: neuronas periféricas y sus proyecciones. M. Neurona autónoma marcada mediante la in-yección intracelular de un marcador enzimático. N. Axones motores (verde) y sinapsis neuromusculares (naranja) en ratonestransgénitos obtenidos mediante ingeniería genética para e xpresar proteínas fluorescentes. O. Proyección de los ganglios de lasraíces dorsales hacia la médula espinal, demostrada mediante un marcador enzimático. P. Axones de las neuronas receptoras olfa-torias desde la nariz marcadas en el b ulbo olfatorio con un colorante fluorescente vital. (G, cortesía de L. C. Katz; H, cortesía deC. J. Shatz; N, O, cortesía de W. Snider y J. Lichtman; todas las otras, cortesía de A.-S. LaMantia y D. Purves.)
A693-01.qxd 8/3/06 11:41 AM Page 10

se antes). La diversidad celular de cualquier sistema nervioso –incluido el nues-tro– indudablemente subyace a la capacidad del sistema para formar redes cadavez más complicadas que median conductas cada v ez más sofisticadas.
Durante gran parte del siglo XX los neurocientíficos se basaron en el mismoconjunto de técnicas desarrolladas por Cajal y Golgi para describir y categorizarla diversidad de tipos celulares en el sistema nervioso. No obstante, desde finesde la década de 1970 en adelante, tecnologías nuevas que se hicieron posibles
Estudio del sistema nervioso de los seres humanos y otros animales 11
(A) (B) (C) (D)
(E) (F) (G) (H)
(I) (J) (K) (L)
(M) (N) (O) (P)
A693-01.qxd 8/3/06 11:41 AM Page 11

por los adelantos de la biología celular y molecular proporcionaron a los in ves-tigadores muchas herramientas adicionales para discernir las propiedades de lasneuronas (fig. 1-6). Aunque los métodos de tinción de las células generales mos-traron diferencias, principalmente en el tamaño y la distrib ución celulares, lastinciones de anticuerpos y las sondas para RN A mensajero aumentaron muchola apreciación de tipos distintos de neuronas y glía en diferentes regiones del sis-tema nervioso. Al mismo tiempo, los métodos nuevos de rastreo de tractos queutilizan una amplia variedad de sustancias marcadoras permitieron explorar mu-cho más cabalmente las intercone xiones entre grupos específ icos de neuronas.Se pueden introducir marcadores en el tejido vi vo o fijado que se transportan alo largo de las prolongaciones de las células nerviosas para poner en e videnciasu origen y terminación. En época más reciente se combinaron métodos genéti-cos y neuroanatómicos para visualizar la expresión de moléculas de marcadoresfluorescentes o de otro tipo bajo el control de secuencias re guladoras de genesneurales. Este enfoque, que muestra las células individuales en tejido fijado o vi-vo con un detalle notable, permite identificar las células nerviosas tanto por suestado transcripcional como por su estructura. Por último, se pueden combinarlas formas de determinar la identidad y la morfología molecular de las célulasnerviosas con mediciones de su acti vidad fisiológica, lo que demuestra así susrelaciones estructura-función. En la f igura 1-6 se muestran ejemplos de estosdistintos enfoques.
Circuitos neurales
Las neuronas nunca funcionan de forma aislada; están organizadas en con-juntos o circuitos neurales que procesan tipos específ icos de información yaportan las bases para la sensación, la percepción y la conducta. Las conexio-nes sinápticas que def inen estos circuitos se realizan típicamente en una ma-raña densa de dendritas, terminaciones axónicas y prolongaciones de célulasgliales que en conjunto constituyen lo que se denomina neuropilo (el sufijo–pilo proviene de la palabra griega pilos, que significa “sentido”; véase fig. 1-3). Por lo tanto, el neuropilo es la región entre los cuerpos de las células ner-viosas donde se produce la mayor parte de la conecti vidad sináptica.
Aunque la disposición de los circuitos neurales varía mucho según la fun-ción que cumplen, algunos elementos son característicos de estos conjuntos.Es importante la dirección del flujo de información en todo circuito particu-lar, lo que, como es evidente, es esencial para conocer su propósito. Las cé-lulas nerviosas que transportan información hacia el encéfalo o la médula es-pinal (o de modo más central, dentro de la médula espinal o el encéf alo) sedenominan neuronas aferentes; las células nerviosas que transportan infor-mación lejos del encéfalo o la médula espinal (o lejos del circuito en cues-tión) se denominan neuronas eferentes. Las interneuronas o las neuronasde circuito local sólo participan en los aspectos locales de un circuito, sobrela base de distancias cortas sobre las que se e xtienden sus axones. Estas tresclases funcionales –neuronas aferentes, neuronas eferentes e interneuronas–son los componentes básicos de todos los circuitos neurales.
Un ejemplo simple de un circuito neural es un conjunto de células que co-rresponden al reflejo espinal miotático (el reflejo “patelar”; f ig. 1-7). Lasneuronas aferentes del reflejo son neuronas sensitivas cuyos cuerpos celula-res se ubican en los ganglios de las raíces dorsales y cuyos axones periféri-cos terminan en terminaciones sensiti vas en los músculos esqueléticos (losganglios que cumplen esta misma función en gran parte de la cabeza y el cue-
12 Capítulo 1
A693-01.qxd 8/3/06 11:41 AM Page 12

llo se denominan ganglios de los nervios craneales; véase el apéndice A).Los axones centrales de estas neuronas sensitivas aferentes entran en la médu-la espinal donde terminan sobre distintas neuronas centrales vinculadas con laregulación del tono muscular, sobre todo las neuronas motoras que determi-nan la actividad de los músculos relacionados. Éstas son las neuronas eferentesy las interneuronas del circuito. Un grupo de estas neuronas eferentes en el astaventral de la médula espinal se proyecta hacia los músculos flexores de la ex-tremidad y el otro hacia los músculos extensores. Las interneuronas de la mé-dula espinal son el tercer elemento de este circuito. Las interneuronas recibencontactos sinápticos de neuronas aferentes sensiti vas y hacen sinapsis sobrelas neuronas motoras eferentes que se pro yectan hacia los músculos flexores;por lo tanto, son capaces de modular la conexión aferencia-eferencia. Las co-nexiones sinápticas e xcitadoras entre los aferentes sensiti vos y las neuronasmotoras eferentes extensoras producen la contracción de los músculos e xten-sores; a la vez, las interneuronas activadas por los aferentes son inhibidoras ysu activación disminuye la actividad eléctrica en las neuronas motoras eferen-tes flexoras y hace que los músculos fle xores se tornen menos activos (fig. 1-8). El resultado es una acti vación y una inacti vación complementaria de losmúsculos sinergistas y antagonistas que controlan la posición de la pierna.
Un cuadro más detallado de los acontecimientos que subyacen al circuitomiotático o a cualquier otro puede obtenerse mediante el re gistro electrofisio-lógico (fig. 1-9). Hay dos enfoques básicos para medir la actividad eléctrica deuna célula nerviosa: registro extracelular (también denominado re gistro de
Estudio del sistema nervioso de los seres humanos y otros animales 13
Axónsensitivo(aferente)
InterneuronaAxonesmotores(eferentes)
Receptorsensitivodel músculo
Músculoflexor
Músculoextensor
2C
2B
2A1
3A
3B
4
El golpe del martilloestira el tendón, lo que asu vez estira losreceptores sensitivos enel músculo extensorde la pierna
La piernase extiende
(C) La sinapsis de lainterneurona inhibe la neuronamotora de los músculos flexores
(B) La neurona sensitivatambién excita la interneuronaespinal
(A) La neurona sensitiva hacesinapsis con una neurona motorade la médula espinal y la excita
(B) El músculo flexor se relajadebido a que la actividad desus neuronas motorasse inhibió
1. (A) La neurona motoraconduce el potencial de acciónhasta las sinapsis sobre lasfibras del músculo extensor yproduce su contracción
1 2 3 4
Fig. 1-7. Un circuito reflejo simple,la respuesta patelar (En términos másformales, reflejo miotático), muestravarios puntos acerca de la organizaciónfuncional de los circuitos nerviosos. Laestimulación de los sensores periféricos(en este caso un receptor de estiramien-to muscular) inicia los potenciales delreceptor que disparan potenciales deacción que viajan centralmente a lo lar-go de los axones aferentes de las neuro-nas sensitivas. Esta información estimu-la las neuronas motoras espinales pormedio de contactos sinápticos. Los po-tenciales de acción generados por elpotencial sináptico en las neuronas mo-toras viajan de manera periférica en losaxones eferentes, y dan origen a la con-tracción muscular y una respuesta con-ductual. Uno de los propósitos de estereflejo particular es ayudar a manteneruna postura erecta frente a los cambiosinesperados.
A693-01.qxd 8/3/06 11:41 AM Page 13

unidad única), donde se coloca un electrodo cerca de la célula nerviosa de in-terés para detectar su actividad; y registro intracelular, en el que se coloca unelectrodo dentro de la célula. Los re gistros extracelulares detectan fundamen-talmente potenciales de acción, los cambios todo o nada en el potencial a tra-vés de las membranas de las células nerviosas que transmiten información des-de un punto a otro en el sistema nervioso. Este tipo de re gistro es en particularútil para detectar patrones temporales de acti vidad de potenciales de acción yrelacionar estos patrones con la estimulación por otras aferencias, o con episo-dios conductuales específicos. Los registros intracelulares pueden detectar loscambios graduados más pequeños del potencial que disparan potenciales de ac-ción, y permiten así un análisis más detallado de la comunicación entre las neu-ronas dentro de un circuito. Estos potenciales que se disparan de forma gradua-da pueden surgir en receptores sensitivos o sinapsis, y se denominan potencia-les de receptor o potenciales sinápticos, respectivamente.
Para el circuito miotático se puede medir la acti vidad eléctrica fuera de lacélula y dentro de ella y def inir así las relaciones funcionales entre las neuro-nas del circuito. El patrón de actividad de potenciales de acción puede medir-se para cada elemento del circuito (aferentes, eferentes e interneuronas) antes,durante y después de un estímulo (véase fig. 1-8). Si se comparan el inicio, laduración y la frecuencia de la acti vidad de los potenciales de acción en cadacélula, surge un cuadro funcional del circuito. Como resultado del estímulo,laneurona sensitiva se dispara a mayor frecuencia (esto es, más potenciales deacción por unidad de tiempo). Este incremento desencadena una frecuenciamayor de potenciales de acción tanto en las neuronas motoras e xtensoras co-mo en las interneuronas. De manera simultánea, las sinapsis inhibidoras for-madas por las interneuronas sobre las neuronas motoras fle xoras hacen decli-nar la frecuencia de potenciales de acción en estas células. Por medio de unregistro intracelular es posible observar directamente los cambios de potencialsubyacentes a las conexiones sinápticas del circuito del reflejo miotático (véa-se fig. 1-9).
14 Capítulo 1
Axón sensitivo(aferente)
InterneuronaAxonesmotores(eferentes)
Neurona motora(extensora)
Interneurona
Neurona sensitiva
Golpedel martillo
La piernase extiende
Neurona motora(flexora)
Fig. 1-8. Frecuencia relativa de po-tenciales de acción (indicada por líneasverticales individuales) en diferentescomponentes del reflejo miotáticocuando se activa la vía refleja. Obsérvese el efecto modulador de la interneurona.
A693-01.qxd 8/3/06 11:41 AM Page 14

Organización general del sistema nervioso humano
Cuando se consideran en conjunto, los circuitos que procesan tipos similaresde información comprenden sistemas neurales que desempeñan propósitosconductuales más amplios. La distinción funcional más general divide estas co-lecciones en sistemas sensitivos que adquieren y procesan información del en-torno (p. ej., el sistema visual o el sistema auditivo, véase unidad II) y sistemasmotores que responden a esta información generando movimientos y otras con-ductas (véase unidad III). Sin embargo, hay gran cantidad de células y circuitosque se ubican entre estos sistemas aferente y eferente relativamente bien defini-dos. Éstos se denominan en conjunto sistemas de asociación y median las fun-ciones encefálicas más complejas y menos caracterizadas (véase unidad V).
Además de estas distinciones funcionales amplias, los neurocientíficos ylos neurólogos hicieron una división convencional del sistema nervioso de losvertebrados desde el punto de vista anatómico en los componentes central yperiférico (fig. 1-10). El sistema nervioso central, (SNC), comprende el en-céfalo (hemisferios cerebrales, diencéfalo, cerebelo y tronco del encéf alo) yla médula espinal (para mayor información sobre las características anató-micas macroscópicas del SNC véase apéndice A). El sistema nervioso peri-férico (SNP) implica las neuronas sensitivas que conectan los receptores sen-sitivos sobre la superficie del cuerpo o más profundo dentro de él con circui-tos de procesamiento rele vantes en el sistema nervioso central. La porción
Estudio del sistema nervioso de los seres humanos y otros animales 15
(C) Interneurona
Interneurona
Neuronasensitiva
(A) Neurona sensitiva
Neuronamotora (flexora)
(D) Neurona motora (flexora)
Neuronamotora(extensora)
(B) Neurona motora (extensora)
Microelectrodopara medir elpotencial de membranaRegistro
Registro
Registro
RegistroPo
tenc
ial d
em
embr
ana
(mV
)Po
tenc
ial d
em
embr
ana
(mV
)Po
tenc
ial d
em
embr
ana
(mV
)Po
tenc
ial d
em
embr
ana
(mV
)
Tiempo (ms)
Sinapsisexcitadora activa
Sinapsisexcitadora activa
Sinapsisexcitadora activa
Potencialde acción
Potencialde acción
Potencialsináptico
Potencialde acción
Potencialsináptico
Fig. 1-9. Respuestas registradas den-tro de la célula subyacen al reflejo mio-tático. A. Potencial de acción medidoen una neurona sensitiva. B. Potencialdisparador postsináptico registrado enuna neurona motora extensora. C. Po-tencial disparador postsináptico en unainterneurona. D. Potencial inhibidorpostsináptico en una neurona motoraflexora. Estos registros intracelularesson la base para conocer los mecanis-mos celulares de la generación del po-tencial de acción y los potenciales dereceptor sensitivo y sinápticos que desencadenan estas señales conducidas.
A693-01.qxd 8/3/06 11:41 AM Page 15

motora del sistema nervioso periférico a su v ez presenta dos componentes.Los axones motores que conectan el encéf alo y la médula espinal con losmúsculos esqueléticos constituyen la división motora somática del sistemanervioso periférico, mientras que las células y los axones que inerv an losmúsculos lisos, el músculo cardíaco y las glándulas forman la división mo-tora visceral o autónoma.
Los cuerpos de las células nerviosas en el sistema nervioso periférico selocalizan en los ganglios, que son simplemente acumulaciones de cuerpos decélulas nerviosas (y células de sostén). Los axones periféricos se reúnen enhaces llamados nervios, muchos de los cuales están envueltos por las célulasgliales del sistema nervioso periférico denominadas células de Schwann. Enel sistema nervioso central las células nerviosas están organizadas de dos for-mas distintas. Los núcleos son acumulaciones locales de neuronas que tienenconexiones y funciones más o menos similares; estos grupos se encuentrandistribuidos en el cerebro, el tronco del encéf alo y la médula espinal. Por elcontrario, la corteza describe disposiciones laminares de células nerviosas(para obtener información adicional e ilustraciones consulte apéndice A). Lascortezas de los hemisferios cerebrales y el cerebelo proporcionan el ejemplomás claro de este principio de or ganización.
Los axones del sistema nervioso central se reúnen en tractos que son máso menos análogos a los nervios de la periferia. Los tractos que cruzan la línea
16 Capítulo 1
COMPONENTESSENSITIVOS
Hemisferios cerebrales, diencéfalo, cerebelo, tronco del encéfalo y médula
espinal (análisis e integración de lainformación sensitivomotora)
(B)(A)
COMPONENTESMOTORES
MEDIOAMBIENTEINTERNO
Y EXTERNO
Ganglios y nerviossensitivos (divisiones
simpática,parasimpática
y entérica)
SISTEMAMOTOR
VISCERAL
SISTEMAMOTOR
SOMÁTICO
Receptoressensitivos (en la
superficie y el interiordel cuerpo)
Ganglios ynervios
autónomos
Nervios motores
Músculos lisos,músculos cardíacos
y glándulas
Músculosesqueléticos(estriados)
EFECTORES
Sistema
nervioso centralSistem
a nerviosoperiférico
Sistemanervioso central
Sistemanervioso periférico
Nervios craneales
Nervios espinales
Encéfalo
Médula espinal
Fig. 1-10. Componentes principalesdel sistema nervioso y sus relacionesfuncionales. A. SNC (encéfalo y médu-la espinal) y SNP (nervios espinales ycraneales). B. Diagrama de los compo-nentes principales de los sistemas ner-viosos central y periférico, y sus rela-ciones funcionales. Los estímulos pro-venientes del medio ambiente transmi-ten información a los circuitos procesa-dores en el interior del encéfalo y lamédula espinal, que a su vez interpre-tan su significación y envían señales alos efectores periféricos que mueven elcuerpo y adaptan los funcionamientosde sus órganos internos.
A693-01.qxd 8/3/06 11:41 AM Page 16

media del encéfalo se denominan comisuras. Dos términos histológicos am-plios distinguen las re giones ricas en cuerpos de células nerviosas v ersus lasregiones ricas en axones. La sustancia gris se refiere a cualquier acumulaciónde cuerpos celulares y neuropilo del encéf alo y la médula espinal (p. ej., nú-cleos o cortezas), mientras que la sustancia blanca, llamada así por su aspec-to relativamente claro como resultado del contenido de lípidos de la mielina,se refiere a los tractos axónicos y las comisuras.
La organización de la división motora visceral del sistema nervioso peri-férico es un poco más complicada (véase cap. 20). Las neuronas motoras vis-cerales del tronco del encéf alo y la médula espinal, denominadas neuronaspreganglionares, forman sinapsis con neuronas motoras periféricas situadasen los ganglios autónomos. Las neuronas motoras de los ganglios autóno-mos inervan el músculo liso, las glándulas y el músculo cardíaco, y contro-lan así la mayoría de las conductas in voluntarias (visceral). En la divisiónsimpática del sistema motor autónomo, los ganglios se sitúan a lo largo de lacolumna vertebral o por delante de ella y envían sus axones a distintas dianasperiféricas. En la división parasimpática los ganglios se sitúan dentro de losórganos que inerv an. Otro componente del sistema motor visceral, llamadosistema entérico, está formado por ganglios pequeños y neuronas individua-les dispersas en toda la pared del intestino. Éstas influyen en la motilidad yla secreción gástrica.
Terminología neuroanatómica
Describir la organización del sistema nervioso requiere un conocimiento ru-dimentario de la terminología anatómica. Los términos utilizados para especi-ficar la localización en el sistema nervioso central son los mismos que se usanpara la descripción anatómica macroscópica del resto del cuerpo (f ig. 1-11).Así, anterior y posterior indican frente y dorso (cabeza y cola); rostral y cau-dal, hacia la cabeza y la cola; dorsal y ventral, arriba y abajo (dorso y vientre),y medial y lateral, en la línea media o al costado. No obstante, la comparaciónentre estas coordenadas en el cuerpo v ersus el encéfalo puede ser confusa. Pa-ra la totalidad del cuerpo estos términos anatómicos se ref ieren al eje mayor ,que es recto. Sin embargo, el eje mayor del sistema nervioso central tiene unacurva. En los seres humanos y otros bípedos se necesita una inclinación com-pensadora del eje rostrocaudal del encéf alo para comparar de manera correctalos ejes corporales con los encefálicos. Una v ez realizado este ajuste, se pue-den asignar fácilmente los ejes al encéf alo.
La asignación correcta de los ejes anatómicos indica entonces los planosestándar para los cortes histológicos o las imágenes vi vas (véase recuadro A)usadas para estudiar la anatomía interna del encéf alo (véase fig. 1-11B). Loscortes horizontales (también llamados axiales o transversos) se toman para-lelos al eje rostrocaudal del encéfalo; por lo tanto, en un individuo en posiciónerecta estos cortes son paralelos a la tierra. Los cortes tomados en el plano quedivide los dos hemisferios son sagitales, y pueden categorizarse a su vez co-mo mediosagitales y parasagitales, según que el corte se encuentre cerca dela línea media (mediosagital) o más lateral (parasagital). Los cortes tomadosen el plano del rostro se denominan coronales o frontales. Por lo general seutilizan términos diferentes para referirse a los cortes de la médula espinal. Elplano de corte ortogonal a la longitud de la médula espinal se denomina trans-verso, mientras que los cortes paralelos al eje mayor de la médula se denomi-nan longitudinales. En un corte transv erso de la médula espinal humana, los
Estudio del sistema nervioso de los seres humanos y otros animales 17
A693-01.qxd 8/3/06 11:41 AM Page 17
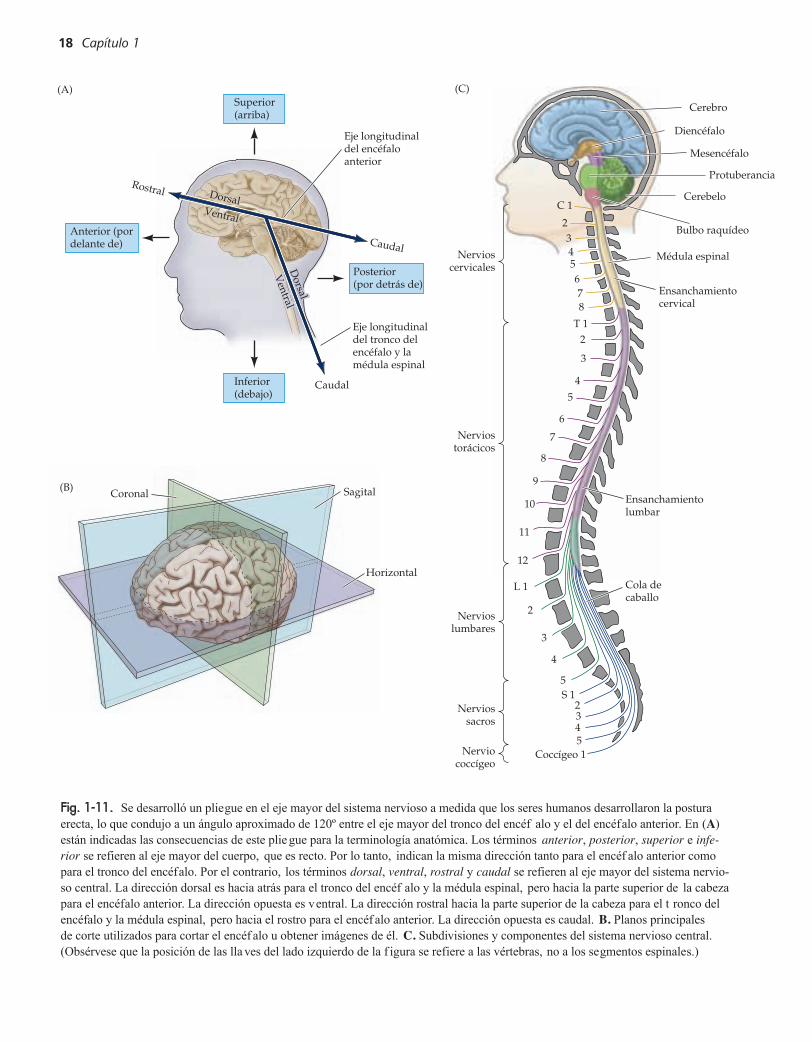
18 Capítulo 1
(B)
Posterior(por detrás de)
Superior(arriba)
Anterior (pordelante de)
Inferior(debajo)
Caudal
Eje longitudinaldel encéfaloanterior
Eje longitudinaldel tronco delencéfalo y lamédula espinal
(A)
Rostral
Caudal
Horizontal
Coronal Sagital
DorsalVentral
Dorsal
Ventral
Médula espinal
Ensanchamientocervical
Ensanchamientolumbar
Cola decaballo
C 1
2345678
T 1
Nervioscervicales
Nerviostorácicos
Nervioslumbares
(C)
Nerviossacros
Nerviococcígeo
T 12
3
4
5
6
7
8
9
10
11
12
L 1
2
3
4
5 S 1
3 4 5
Coccígeo 1
2
Bulbo raquídeo
Protuberancia
Mesencéfalo
Diencéfalo
Cerebro
Cerebelo
Fig. 1-11. Se desarrolló un pliegue en el eje mayor del sistema nervioso a medida que los seres humanos desarrollaron la posturaerecta, lo que condujo a un ángulo aproximado de 120º entre el eje mayor del tronco del encéf alo y el del encéfalo anterior. En (A)están indicadas las consecuencias de este plie gue para la terminología anatómica. Los términos anterior, posterior, superior e infe-rior se refieren al eje mayor del cuerpo, que es recto. Por lo tanto, indican la misma dirección tanto para el encéf alo anterior comopara el tronco del encéfalo. Por el contrario, los términos dorsal, ventral, rostral y caudal se refieren al eje mayor del sistema nervio-so central. La dirección dorsal es hacia atrás para el tronco del encéf alo y la médula espinal, pero hacia la parte superior de la cabezapara el encéfalo anterior. La dirección opuesta es ventral. La dirección rostral hacia la parte superior de la cabeza para el t ronco delencéfalo y la médula espinal, pero hacia el rostro para el encéf alo anterior. La dirección opuesta es caudal. B. Planos principales de corte utilizados para cortar el encéf alo u obtener imágenes de él. C. Subdivisiones y componentes del sistema nervioso central. (Obsérvese que la posición de las llaves del lado izquierdo de la f igura se refiere a las vértebras, no a los segmentos espinales.)
A693-01.qxd 8/3/06 11:41 AM Page 18

ejes dorsal y ventral, y los ejes anterior y posterior indican las mismas direc-ciones (véase fig. 1-11). Aunque esta terminología puede ser tediosa, es esen-cial para conocer las subdivisiones básicas del sistema nervioso (f ig. 1-11C).
Subdivisiones del sistema nervioso central
Por lo general, se considera que el sistema nervioso central (def inido co-mo el encéfalo y la médula espinal) tiene siete partes básicas: la médula es-pinal, el bulbo raquídeo, la protuberancia, el cerebelo, el mesencéfalo, eldiencéfalo y los hemisferios cerebrales (véanse figs. 1-10 y 1-11C). Todasestas subdivisiones son atravesadas por espacios llenos de líquido llamadosventrículos (se puede hallar un informe detallado del sistema v entricular enel apéndice B). Los ventrículos son el remanente de la luz que se forma al ple-garse la placa neural para formar el tubo neural al comienzo del desarrollo(véase cap. 21). Las variaciones en la configuración y el tamaño del espacioventricular maduro son característicos de cada región del encéfalo adulto. Elbulbo raquídeo, la protuberancia y el mesencéfalo se denominan en conjuntotronco del encéfalo y rodean el cuarto ventrículo (bulbo raquídeo y protu-berancia) y el acueducto cerebral (mesencéfalo). El diencéfalo y los hemis-ferios cerebrales se denominan en conjunto encéfalo anterior, y encierran eltercer ventrículo y los ventrículos laterales, respectivamente. Dentro deltronco del encéf alo están los núcleos de los nervios craneales que recibenaferencias de los ganglios sensitivos craneales mencionados antes, a travésde los nervios sensitivos craneales, o dan origen a axones que constituyenlos nervios motores craneales (véase apéndice A).
El tronco del encéfalo también es un conducto para v arios tractos mayoresen el sistema nervioso central que transmiten información sensiti va desde lamédula espinal y el tronco del encéfalo hacia el encéfalo anterior, o transmitenórdenes motoras del encéfalo anterior nuevamente hacia las neuronas motorasen el tronco del encéfalo y la médula espinal. En consecuencia, el conocimien-to detallado de las consecuencias del daño del tronco del encéf alo proporcionaa los neurólogos y otros médicos una herramienta esencial para localizar ydiagnosticar una lesión encefálica. El tronco del encéfalo contiene otros nume-rosos núcleos que participan en muchas funciones importantes,como el controlde la frecuencia cardíaca, la respiración, la presión arterial y el ni vel de con-ciencia. Por último, una de las características más sobresalientes del tronco delencéfalo es el cerebelo, que se extiende sobre gran parte de su cara dorsal. Elcerebelo es esencial para la coordinación y la planif icación del mo vimiento(véase cap. 18) y para el aprendizaje de las tareas motoras y el almacenamien-to de esa información (véase cap. 30).
Hay varias subdivisiones anatómicas del encéf alo anterior. Las estructurasanatómicas más obvias son los hemisferios cerebrales prominentes (fig. 1-12).En los seres humanos los hemisferios cerebrales (cuyas porciones más externasson láminas continuas y muy plegadas de corteza) son proporcionalmente másgrandes que en cualquier otro mamífero, y se caracterizan por circunvolucio-nes (giros) o crestas de tejido cortical plegado y surcos, las hendiduras que di-viden las circunvoluciones entre sí (p. ej., como se observa en la tapa de este li-bro). Aunque los patrones de circunvoluciones y surcos varían de un individuoa otro, hay algunos puntos de referencia sistemáticos que ayudan a di vidir loshemisferios en cuatro lóbulos. Los nombres de los lóbulos derivan de los hue-sos craneales que los cubren: occipital, temporal, parietal y frontal. Una ca-racterística clave de la anatomía de la superficie del cerebro es el surco central
Estudio del sistema nervioso de los seres humanos y otros animales 19
A693-01.qxd 8/3/06 11:41 AM Page 19

20 Capítulo 1
Circunvoluciónprecentral
(A)
(B)
(E)
(D)
(F)
(C)Circunvoluciónpostcentral
Surco central
Surcoparietooccipital
Incisurapreoccipital
Fisuralateral (de Silvio)
Hemisferiocerebral
Cerebelo
Troncodel encéfalo
Médulaespinal
Cerebelo
Circunvolucióncingular
Surcoparietooccipital
Médula espinal
Surcocingular
Diencéfalo
Cuerpocalloso
Comisuraanterior
Troncodelencéfalo
MesencéfaloProtuberancia
Bulbo raquídeo
Surcocalcarino
Surcocentral
Cuerpocalloso
Caudado
Putamen
Cápsulainterna
Sustanciablanca
Quiasmaóptico
Núcleos delencéfalo anterior basal
Comisuraanterior
Lóbulotemporal
Corteza cerebral(sustancia gris)
Amígdala
CuerpocallosoVentrículo
lateral
Fórnix
Tercerventrículo
Hipocampo
Cuerpomamilar
Ventrículolateral(astatemporal)
Tálamo
Caudado Putamen
Globopálido
Cola delnúcleocaudado
Ganglios basales
Cápsulainterna
Lóbulofrontal
Lóbulotemporal
Lóbuloparietal
Lóbulooccipital
Lóbulofrontal
Lóbulotemporal
Lóbuloparietal
Lóbulooccipital
Nivel del corte quese observa en (E)
Nivel del corte quese observa en (F)
A693-01.qxd 8/3/06 11:41 AM Page 20

situado más o menos a mitad de camino entre los polos rostral y caudal de loshemisferios (fig. 1-12A). Este surco prominente divide el lóbulo frontal en el ex-tremo rostral del hemisferio del lób ulo parietal más caudal. A ambos lados delsurco central se destacan las circunvoluciones precentral y postcentral. Estas cir-cunvoluciones también son funcionalmente signif icativas, ya que la primeracontiene la corteza motora primaria importante para controlar el mo vimiento, yla postcentral contiene la corteza somatosensitiva primaria que es importante pa-ra los sentidos del cuerpo (véase más adelante).
Las subdivisiones restantes del encéfalo anterior se sitúan más profundas enlos hemisferios cerebrales (fig. 1-12B). Las que más se destacan es el grupo deestructuras profundas involucradas en los procesos motores y cognitivos que sedenominan en conjunto ganglios basales. Otras estructuras en particular im-portantes son el hipocampo y la amígdala en los lóbulos temporales (se tratade sustratos vitales para la memoria y la conducta emocional, respectivamente)y los bulbos olfatorios (las estaciones centrales para el procesamiento de la in-formación quimiosensitiva que surge en las neuronas receptoras en la ca vidadnasal) sobre la cara anteroinferior de los lób ulos frontales. Por último, el tála-mo se ubica en el diencéfalo y es un área de relevo crítica para la informaciónsensitiva (aunque también tiene muchas otras funciones); el hipotálamo, quecomo su nombre lo indica se sitúa por debajo del tálamo, es la estructura orga-nizadora central para la re gulación de muchas funciones homeostáticas delcuerpo (p. ej., alimentación, ingestión de líquidos, termorregulación).
Esta descripción básica de los puntos de referencia anatómicos más im-portantes proporciona un marco de trabajo para comprender cómo se comu-nican entre ellas las neuronas ubicadas en estructuras encefálicas diferentes yampliamente distribuidas para definir sistemas neurales dedicados a codifi-car, procesar y transmitir tipos específicos de información acerca de aspectosdel entorno del organismo, e iniciar y coordinar luego respuestas conductua-les apropiadas.
Principios de la organización de los sistemas neurales
Estas capacidades perceptivas y motoras complejas del encéf alo reflejan lafunción integrada de distintos sistemas neurales. El procesamiento de la infor-mación somatosensitiva (que se origina en los receptores de piel,tejidos subcu-táneos y sistema musculoesquelético que responden a deformación física en lasuperficie del cuerpo o desplazamiento de músculos y articulaciones) propor-ciona un ejemplo conveniente. Estas estructuras ampliamente distrib uidas queparticipan en la generación de las sensaciones somáticas se denominan sistemasomatosensitivo (fig. 1-13). Los componentes en el sistema nervioso periféri-co son los receptores distribuidos en toda la piel y en músculos y tendones, las
Estudio del sistema nervioso de los seres humanos y otros animales 21
Fig. 1-12. Anatomía macroscópica del encéfalo anterior. A. Anatomía de la superficie de los hemisferios cerebrales que muestra los cuatro lób ulos del encéfalo ylos surcos y las circunvoluciones principales. En esta vista imaginaria también se pue-den ver el sistema ventricular y los ganglios basales. B. Vista mediosagital que muestrala localización de hipocampo, amígdala, tálamo e hipotálamo.
A693-01.qxd 8/3/06 11:41 AM Page 21

22 Capítulo 1
Sistemanervioso central
Sistemanervioso periférico
Receptoressensitivosdel cuerpo
Receptoressensitivosdel rostro
Receptorsensitivo
Tálamo
Corteza cerebralCorteza
somatosensitiva
(A)
(B)
Troncodel encéfalo
Médulaespinal
Tálamo
Corteza cerebralCorteza somatosensitiva
Tronco delencéfalo
Médulaespinal
Ganglios delas raícesdorsales
Ganglios delas raíces dorsales
Sensación mecánica
Gangliostrigeminales
GangliotrigeminalGanglios
trigeminales
Dolor y temperatura
Sensación mecánica
Dolor y temperatura
Cervicales
Torácicos
Lumbares
Sacros
Fig. 1-13. Organización anatómica y funcional del sistema somatosensiti vo. Los com-ponentes del sistema nervioso central del sistema somatosensiti vo se encuentran en lamédula espinal, el tronco del encéfalo, el tálamo y la corteza cerebral. A. La informa-ción somatosensitiva de la superficie del cuerpo se traza según los ganglios de las raícesdorsales, que aquí se esquematizan unidos a la médula espinal. Los distintos tonos depúrpura indican la correspondencia entre las re giones de la superficie corporal y losganglios de las raíces dorsales que transmiten información desde la superf icie corporalhasta el sistema nervioso central. La información pro veniente de cabeza y cuello setransmite hasta el SNC a través de los ganglios trigeminales. B. La información somato-sensitiva viaja desde los receptores sensitivos periféricos a través de vías paralelas paralas sensaciones mecánica y de dolor y temperatura. Estas vías paralelas hacen rele vo através de la médula espinal y el tronco del encéf alo, para enviar finalmente informaciónsensitiva hasta el tálamo, desde donde se transmite a la corteza somatosensiti va en lacircunvolución postcentral (indicada en azul en la imagen del encéf alo entero; RM cor-tesía de L. E. White, J. Vovoydic y S. M. Williams).
A693-01.qxd 8/3/06 11:41 AM Page 22

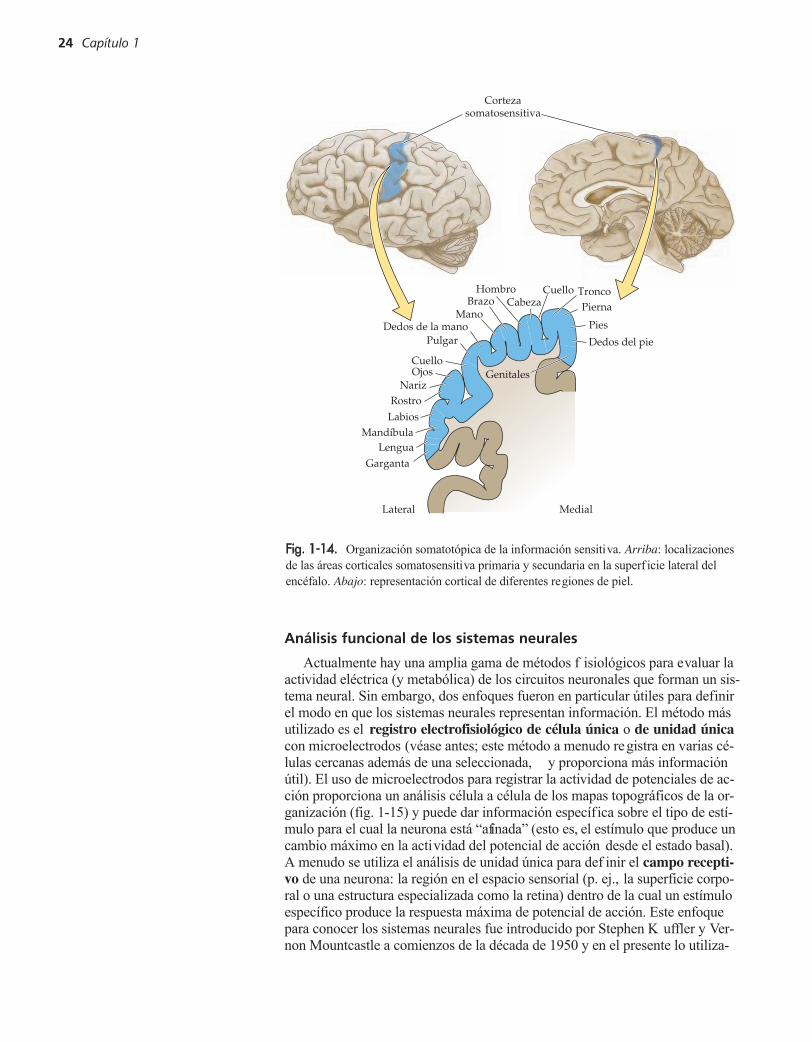
neuronas relacionadas en los ganglios de la raíz dorsal, y las neuronas en algu-nos ganglios craneales. Entre los componentes del sistema nervioso central seincluyen neuronas en la médula espinal; los tractos lar gos de sus axones, quese originan en la médula espinal, cruzan a través del tronco del encéf alo y f i-nalmente terminan en distintos núcleos de relevo en el tálamo en el diencéf a-lo. Las estructuras diana incluso más altas de las neuronas talámicas son lasáreas corticales que rodean a la circun volución postcentral que se denominanen conjunto corteza somatosensitiva. Por lo tanto, el sistema somatosensitivopresenta poblaciones específicas de neuronas prácticamente en todas las subdi-visiones del sistema nervioso.
Otros dos principios de la or ganización del sistema nervioso son e viden-tes en el sistema somatosensiti vo: la organización topográfica y la prepon-derancia de las vías paralelas (véase fig. 1-13). Como su nombre lo indica,topografía se refiere a una función de mapeo –en este caso un mapa de la su-perficie corporal en el que puedan discernirse las distintas estructuras queconstituyen el sistema somatosensitivo. Por lo tanto, se trazan áreas adyacen-tes sobre la superficie corporal a regiones adyacentes en los núcleos, los trac-tos de sustancia blanca y las zonas diana talámicas y corticales del sistema.Comenzando en la periferia, las células de cada ganglio de la raíz dorsal de-finen un dermatoma separado (área de piel inervada por las prolongacionesde células provenientes de una sola raíz dorsal). En la médula espinal,de cau-dal a rostral, los dermatomas están representados en regiones correspondien-tes de la médula espinal de sacros (dorso) a lumbares (piernas) a torácicos(tórax) y cervicales (brazos y hombros) (véanse figs. 1-13 y 1-11C). Esta so-matotopia se mantiene en los tractos somatosensiti vos de la médula espinaly el tronco del encéfalo que transmiten información a las estructuras relevan-tes del encéfalo anterior del sistema somatosensitivo (fig. 1-14).
Las vías paralelas se refieren a la de los axones de las células nerviosasque procesan los distintos atributos del estímulo que comprenden una moda-lidad sensitiva, motora o cognitiva particular. Para la sensación somática, losatributos del estímulo que se transmiten a tra vés de vías paralelas son dolor ,temperatura, tacto, presión y propiocepción (el sentido de posición articularo de las extremidades). Desde los ganglios de las raíces dorsales, a través dela médula espinal y el tronco del encéf alo, y hasta la corteza somatosensiti-va, estas submodalidades se mantienen en gran parte divididas. Por lo tanto,desde el punto de vista anatómico, bioquímico y fisiológico distintas neuro-nas traducen, codifican y transmiten información de dolor, temperatura y me-cánica. Aunque esta información lue go se inte gra para proporcionar la per-cepción unitaria de los estímulos relevantes, las neuronas y los circuitos en elsistema somatosensiti vo están claramente especializados para procesar as-pectos separados de la sensibilidad somática.
Este bosquejo básico de la organización del sistema somático es represen-tativo de los principios pertinentes al conocimiento de cualquier sistema neu-ral. En todos los casos será apropiado considerar la distribución anatómica delos circuitos neurales dedicados a una función particular , el modo en que lafunción está representada o se “mapea” en los elementos neurales dentro delsistema y de qué modo se segregan los distintos atributos del estímulo dentrode los subgrupos de neuronas que comprenden el sistema. Estos detalles pro-porcionan un marco de trabajo para comprender de qué modo la acti vidaddentro del sistema proporciona una representación del estímulo rele vante, larespuesta motora requerida y correlaciones cognitivas de orden superior.
Estudio del sistema nervioso de los seres humanos y otros animales 23
A693-01.qxd 8/3/06 11:41 AM Page 23
Análisis funcional de los sistemas neurales
Actualmente hay una amplia gama de métodos f isiológicos para evaluar laactividad eléctrica (y metabólica) de los circuitos neuronales que forman un sis-tema neural. Sin embargo, dos enfoques fueron en particular útiles para definirel modo en que los sistemas neurales representan información. El método másutilizado es el registro electrofisiológico de célula única o de unidad únicacon microelectrodos (véase antes; este método a menudo registra en varias cé-lulas cercanas además de una seleccionada, y proporciona más informaciónútil). El uso de microelectrodos para registrar la actividad de potenciales de ac-ción proporciona un análisis célula a célula de los mapas topográficos de la or-ganización (fig. 1-15) y puede dar información específica sobre el tipo de estí-mulo para el cual la neurona está “afinada” (esto es, el estímulo que produce uncambio máximo en la actividad del potencial de acción desde el estado basal).A menudo se utiliza el análisis de unidad única para def inir el campo recepti-vo de una neurona: la región en el espacio sensorial (p. ej., la superficie corpo-ral o una estructura especializada como la retina) dentro de la cual un estímuloespecífico produce la respuesta máxima de potencial de acción. Este enfoquepara conocer los sistemas neurales fue introducido por Stephen K uffler y Ver-non Mountcastle a comienzos de la década de 1950 y en el presente lo utiliza-
24 Capítulo 1
Cortezasomatosensitiva
Hombro
Cuello
CabezaCuello
BrazoMano
Dedos de la manoPulgar
OjosNariz
RostroLabios
MandíbulaLengua
Garganta
Dedos del pie
Genitales
Pies
PiernaTronco
Lateral Medial
Fig. 1-14. Organización somatotópica de la información sensitiva. Arriba: localizacionesde las áreas corticales somatosensitiva primaria y secundaria en la superf icie lateral delencéfalo. Abajo: representación cortical de diferentes regiones de piel.
A693-01.qxd 8/3/06 11:41 AM Page 24

ron varias generaciones de neurocientíf icos para evaluar la relación entre estí-mulos y respuestas neuronales en los sistemas sensitivo y motor. En la actuali-dad las técnicas de registro eléctrico a nivel de una única célula se extendierony refinaron para incluir el análisis de una sola célula y a la v ez de numerosascélulas en animales que realizan tareas cogniti vas complejas, los registros in-tracelulares en animales intactos y el uso de electrodos en parches para detec-tar y monitorizar la actividad de moléculas individuales de la membrana que fi-nalmente subyacen al señalamiento neural (véase unidad I).
La segunda área importante en la que se lograron adelantos técnicos nota-bles son las imágenes encefálicas funcionales en seres humanos (y en menorgrado en animales), que revolucionaron el conocimiento funcional de los sis-temas neurales en las dos últimas décadas (recuadro A). Al contrario de losmétodos eléctricos para re gistrar la actividad neural, que son invasores en elsentido de tener que exponer el encéfalo e insertar electrodos en él, las imáge-nes funcionales no son in vasoras y por lo tanto se pueden aplicar a pacientesy sujetos normales. Además, las imágenes funcionales permiten la evaluaciónsimultánea de varias estructuras encefálicas (que es posible pero ob viamentedifícil con métodos de registro eléctrico). Las tareas que pueden evaluarse me-diante imágenes funcionales permiten un enfoque más ambicioso e integradordel estudio de las operaciones de un sistema neural.
En los últimos 20 años estos métodos no invasores permitieron que los neu-rocientíficos evalúen la representación de una cantidad enorme de conductashumanas complejas, y al mismo tiempo aportaron herramientas diagnósticas
Estudio del sistema nervioso de los seres humanos y otros animales 25
(A) (B)
Corteza somatosensitiva
Campo receptivo(periferia)
Actividad de laneurona cortical
Período deestimulación
Surcocentral
Circunvoluciónpostcentral
Registro
Camporeceptivo (centro)
El tacto en la periferiadel campo receptivodisminuye la descargacelular
El tacto fuera delcampo receptivo notiene efecto alguno
El tacto en el centro delcampo receptivo aumen-ta la descarga celular
Fig. 1-15. Registro electrofisiológico de unidad única de una neurona piramidal corti-cal, que muestra el patrón de descarga en respuesta a un estímulo periférico específ ico.A. Disposición experimental típica. B. Terminación de los campos receptivos neuronales.
A693-01.qxd 8/3/06 11:41 AM Page 25

que se utilizan cada vez más de rutina. Muchas de las observaciones resultan-tes confirmaron las inferencias acerca de la localización funcional y la organi-zación de los sistemas neurales que originariamente se basaron en el estudiode los pacientes neurológicos que mostraban una conducta alterada después deun accidente cerebrovascular u otras formas de lesión encefálica. Sin embar-go, otros hallazgos aportaron conocimientos nue vos sobre la forma en quefuncionan los sistemas neurales en el encéf alo humano.
Análisis de la conducta compleja
Muchos de los adelantos más anunciados en la neurociencia moderna invo-lucraron una reducción de la complejidad del encéf alo hasta los componentesque se analizan más fácilmente; esto es, genes, moléculas o células. No obstan-te, el encéfalo funciona como una totalidad y el estudio de las funciones ence-fálicas más complejas (y , para algunos, más interesantes) como percepción,lenguaje, emoción, memoria y conciencia aún es un desafío central para losneurocientíficos contemporáneos. Reconociendo este desafío, en los últimos20 años más o menos surgió un campo llamado neurociencia cognitiva dedi-cado de manera específ ica a comprender estas cuestiones (véase unidad V).Esta evolución también rejuveneció el campo de la neuroetología (dedicado aobservar conductas complejas de animales en sus entornos naturales; p. ej., lacomunicación social en las a ves y los primates no humanos) y estimuló el de-sarrollo de tareas para e valuar mejor la génesis de las conductas complejas enlos seres humanos. Cuando se usan combinadas con imágenes funcionales, lastareas conductuales bien designadas pueden f acilitar la identificación de redesencefálicas dedicadas a funciones complejas específicas, entre las que se inclu-yen habilidades de lenguaje, capacidad matemática y musical, respuestas emo-cionales, juicios estéticos y pensamiento abstracto. También se pueden utilizartareas conductuales cuidadosamente construidas para estudiar la patología delas enfermedades encefálicas complejas que comprometen la cognición, comola enfermedad de Alzheimer, la esquizofrenia y la depresión.
En resumen, los esfuerzos nuevos o revitalizados para estudiar las funcio-nes encefálicas superiores con técnicas cada v ez más poderosas ofrecen for-mas de comenzar a conocer incluso los aspectos más complejos de la conductahumana.
26 Capítulo 1
A693-01.qxd 8/3/06 11:41 AM Page 26

Estudio del sistema nervioso de los seres humanos y otros animales 27
En la década de 1970 la tomogra-fía computarizada o TC abrió unanueva era en las imágenes no invasorasal introducir el uso de la tecnología deprocesamiento computarizado paraayudar a sondear el encéfalo viviente.Antes de la TC, la única técnica porimágenes encefálicas disponible era laradiografía convencional, que tenía es-caso contraste de tejidos blandos ycomprendía una exposición a la radia-ción relativamente alta.
El enfoque de la TC utiliza un hazde rayos X estrecho y una hilera de de-tectores muy sensibles colocados en la-dos opuestos de la cabeza para sondearsólo una pequeña porción de tejido a lavez con exposición limitada a la radia-ción (véase fig. A). Para formar unaimagen, el tubo de rayos X y los detec-tores rotan alrededor de la cabeza pararecoger información de radiodensidadde todas las orientaciones que rodeanun corte estrecho. Las técnicas de pro-cesamiento computarizado calculan
luego la radiodensidad de cada puntodentro del plano de corte, produciendouna imagen topográfica (tomo significa“corte” o “trozo”). Si el paciente esmovilizado lentamente a través del to-mógrafo mientras el tubo de rayos Xrota de esta forma, se puede crear unamatriz de radiodensidad tridimensional,que permite computarizar las imágenespara cualquier plano a través del encé-falo. Las TC permiten distinguir confacilidad la sustancia blanca de la sus-tancia gris, diferenciar muy bien losventrículos y mostrar muchas otras es-tructuras encefálicas con una resolu-ción espacial de varios milímetros.
Las imágenes encefálicas dieronotro gran paso hacia adelante en la dé-cada de 1980 con el desarrollo de laresonancia magnética (RM). La RMse basa en que los núcleos de algunosátomos actúan como imanes que girany que, si se los coloca en un campomagnético fuerte, cubrirán el campo ygirarán a una frecuencia que depende
de la fuerza del campo. Si ellos recibenentonces un pulso breve de radiofre-cuencia ajustado a su frecuencia de gi-ro se alejan de su alineación con elcampo y luego emiten energía en for-ma oscilatoria a medida que se reali-nean gradualmente con el campo. Lafuerza de la señal emitida depende dela cantidad de núcleos que participanen este proceso. Para obtener informa-ción espacial en la RM, el campo mag-nético se distorsiona ligeramente impo-niendo gradientes magnéticos a lo lar-go de tres ejes espaciales diferentes demodo que solo los núcleos en ciertaslocalizaciones están ajustados a la fre-cuencia del detector en cualquier mo-mento dado. Casi todos los resonadoresutilizan detectores ajustados a las ra-diofrecuencias de los núcleos de hidró-geno giratorios en moléculas de agua,y crean así imágenes basadas en la dis-tribución del agua en diferentes tejidos.Una manipulación cuidadosa de losgradientes de campo magnético y lospulsos de radiofrecuencia hace posibleconstruir imágenes extraordinariamentedetalladas del encéfalo en cualquier lo-calización y orientación, con una reso-lución submilimétrica.
El campo magnético fuerte y lospulsos de radiofrecuencia utilizados enla RM son inocuos, lo que convierte aesta técnica en completamente no inva-sora (aunque los objetos metálicos quese encuentran dentro del resonador ocerca de él constituyen una preocupa-ción para la seguridad) (véase fig. B).La RM también es en extremo versátilporque, al modificar los parámetros delresonador, se pueden generar imágenesbasadas en una amplia variedad de dife-rentes mecanismos de contraste. Porejemplo, las imágenes convencionalesde RM aprovechan que el hidrógeno endistintos tipos de tejido (p. ej., sustancia
Fuentede rayos X
Detectorde rayos X
Recuadro A
Técnicas de imágenes encefálicas
(continúa)
A. En la tomografía computarizada, la fuente de rayos X y los detectores se mue ven alrede-dor de la cabeza del paciente. El recuadro muestra un corte horizontal de TC del encéfalode un adulto normal.
A693-01.qxd 8/3/06 11:41 AM Page 27

28 Capítulo 1
gris, sustancia blanca y líquido cefalo-rraquídeo) tiene velocidades de realinea-ción ligeramente diferentes, lo que indi-ca que el contraste de los tejidos blandos puede manipularse simplementeajustando el momento en que se mide laseñal de hidrógeno que se realinea.También se pueden utilizar diferentesregulaciones de los parámetros para ge-nerar imágenes en las cuales las sustan-cias gris y blanca son invisibles, pero enlas que sobresale la vasculatura encefáli-ca con gran detalle. La seguridad y laversatilidad convirtieron la RM en latécnica de elección para obtener imáge-nes de la estructura encefálica en la ma-yoría de las aplicaciones.
También se hizo posible la obten-ción de imágenes de variaciones fun-cionales en el encéfalo viviente con eldesarrollo reciente de técnicas para de-tectar cambios pequeños localizados enel metabolismo o el flujo sanguíneo ce-rebral. Para conservar energía, el encé-falo regula su flujo sanguíneo de modoque las neuronas activas con demandasmetabólicas relativamente altas recibenmás sangre que neuronas relativamenteinactivas. La detección y el trazado deestos cambios locales en el flujo san-guíneo cerebral forma la base para trestécnicas de imágenes encefálicas fun-cionales muy utilizadas: la tomografíapor emisión de positrones (PET), latomografía computarizada por emi-sión de fotón único (SPECT) y la re-sonancia magnética funcional (RMf).
En la PET se incorporan isótoposemisores de positrones inestables en diferentes reactivos (incluidos agua,moléculas precursoras de neurotransmi-sores específicos o glucosa) y se los in-yecta en el torrente sanguíneo. El oxíge-no y la glucosa marcados se acumulancon rapidez en las áreas con mayor ac-tividad metabólica y las sondas de
transmisores marcados se captan selec-tivamente en regiones apropiadas. A medida que el isótopo inestable sedesintegra, conduce a la emisión de dospositrones que se mueven en direccio-nes opuestas. Los detectores de rayosgamma colocados alrededor de la cabe-za registran un “golpe” sólo cuandodos detectores separados por 180º reac-cionan en forma simultánea. Se puedengenerar entonces imágenes de la densi-dad del isótopo en los tejidos (en formamuy similar a la manera en que se cal-culan las imágenes por TC) que mues-tran la localización de las regiones acti-vas con una resolución espacial deunos 4 mm. Según la sonda inyectada,las imágenes de PET pueden utilizarsepara visualizar cambios dependientesde la actividad en flujo sanguíneo, me-tabolismo tisular o actividad bioquími-ca. Las imágenes de SPECT son simi-
lares a las de PET, ya que comprendenla inyección o la inhalación de un com-puesto radiomarcado (p. ej., 133Xe oyodoanfetamina marcada con 123I),que producen protones detectados poruna cámara gamma que se mueve rápi-damente alrededor de la cabeza.
En la actualidad la RM funcional,variante de la RM, ofrece el mejor en-foque para visualizar la función encefá-lica basada en el metabolismo local. LaRMf se basa en que la hemoglobina enla sangre distorsiona ligeramente laspropiedades de resonancia magnéticade los núcleos de hidrógeno en su v e-cindad y el grado de distorsión magné-tica cambia en función de que la hemo-globina tenga oxígeno ligado a ella.Cuando un área encefálica es activadapor una tarea especial comienza a utili-zar más oxígeno y en pocos se gundosla microvasculatura encefálica respon-
Recuadro A (cont.)
Técnicas de imágenes encefálicas
B. En la RM la cabeza se coloca en el centro de un gran imán. Una antena espiral de radio-frecuencia se coloca alrededor de la cabeza para e xcitar y registrar la señal de resonanciamagnética. En la RMf, los estímulos pueden presentarse utilizando gafas de video de reali-dad virtual y audífonos estereofónicos mientras el paciente está en el interior del resonador .
A693-01.qxd 8/3/06 11:41 AM Page 28

Estudio del sistema nervioso de los seres humanos y otros animales 29
de aumentando el flujo de sangre ricaen oxígeno en el área activa. Estoscambios en la concentración de oxíge-no y el flujo sanguíneo conducen a al-teraciones localizadas dependientes delnivel de la oxigenación sanguínea en laseñal de resonancia magnética. Estasfluctuaciones se detectan por medio detécnicas estadísticas de procesamientode imágenes para producir mapas defunción encefálica dependientes de latarea (véase fig. C). Como la RMf uti-liza las señales intrínsecas al encéfalo
sin radiactividad alguna, se pueden rea-lizar observaciones repetidas en el mis-mo individuo: una ventaja importantesobre los métodos por imágenes comoPET. La resolución espacial (2 a 3 mm)y la resolución temporal (algunos se-gundos) de la RMf también son supe-riores a otras técnicas de imágenes fun-cionales. La RM surgió así como latecnología de elección para sondear laestructura y la función del encéfalo hu-mano.
Bibliografía
HUETTEL, S. A., A. W. SONG Y G. MCCARTHY
(2004) Functional Magnetic Resonance Imaging.Sunderland, MA: Sinauer Associates.
OLDENDORF, W Y W. O LDENDORF HIJO (1988) Ba-sics of Magnetic Resonance Imaging. Boston:Kluwer Academic Publishers.
RAICHLE, M. E. (1994) Images of the mind: Stu-dies with modern imaging techniques. Ann. Rev.Psychol; 45: 333-356.
SCHILD, H. (1990) MRI Made Easy (…Well, Al-most). Berlin: H. Heineman.
in progress
Derecha Izquierda Tumor
C. Imágenes por RM de un paciente adulto con un tumor encefálico, con actividad de RMf durante una tarea de mo vimiento de las manossuperpuesta (la actividad de la mano izquierda se muestra en amarillo; la acti vidad de la mano derecha, en verde). A la derecha hay una vistareconstruida tridimensional de superficie de los mismos datos.
Resumen
El encéfalo puede estudiarse mediante métodos que varían desde la genéti-ca y la biología molecular hasta pruebas conductuales en sujetos normales.Además de un conjunto cada vez mayor de conocimientos acerca de la organi-zación anatómica del sistema nervioso, muchos de los éxitos más brillantes dela neurociencia moderna provinieron del conocimiento de las células nerviosascomo la unidad estructural y funcional del sistema nervioso. Algunos estudiosde la arquitectura celular y los componentes moleculares distintos de las neu-ronas y la glía pusieron en evidencia gran parte de sus funciones individuales,y proporcionaron una base para saber de qué modo se or ganizan las células
A693-01.qxd 8/3/06 11:41 AM Page 29

30 Capítulo 1
Lecturas adicionalesBRODAL, P. (1992) The Central Nervous System:Structure and Function. New York: Oxford Uni-versity Press.
CARPENTER, M. B. Y J. SUTIN (1983) Human Neu-roanatomy, 8a ed. Baltimore, MD: Williams andWilkins.
ENGLAND, M. A. Y J. WAKELY (1991) Color Atlasof the Brain and Spinal Cord: An Introduction toNormal Neuroanatomy. St. Louis: Mosby Year-book.
GILSON, G. Y S. MUSE (2001) A Primer of GenomeScience. Sunderland, MA: Sinauer Associates.
HAINES, D. E. (1995) Neuroanatomy: An Atlas ofStructures, Sections, and Systems, 2a ed. Baltimo-re: Urban and Schwarzenberg.
MARTIN, J. H. (1996) Neuroanatomy: Text andAtlas, 2a ed. Stamford, CT: Appleton and Lange.
NATUREVOL. 409, No. 6822 (2001) Issue of Fe-bruary 16. Special issue on the human genome.
NETTER, F. H. (1983) The CIBA Collection of Me-dical Illustrations Vols. I and II. A. Brass and R. V.Dingle (eds.). Summit, NJ: CIBA PharmaceuticalCo.
PETERS, A., S. L. P ALAY Y H. DE F. W EBSTER
(1991) The Fine Structure of the Nervous System:Neurons and Their Supporting Cells, 3a ed. NewYork: Oxford University Press.
POSNER, M. I. Y M. E. R AICHLE (1997) Images ofMind, 2a ed. New York: W. H. Freeman & Co.
RAMON Y CAJAL, S. (1984) The Neuron and theGlial Cell. (T ransl. by J. de la Torre and W. C.Gibson.) Springfield, IL: Charles C. Thomas.
RAMON Y CAJAL, S. (1990) New Ideas on theStructure of the Nervous System in Man and Ver-tebrates. (Transl. by N. Swanson and L W. Swan-son.) Cambridge, MA: MIT Press.
SCIENCE VOL. 291, No. 5507 (2001) Issue of Fe-bruary 16. Special issue on the human genome.
SHEPHERD, G. M. (1991) Foundations of the Neu-ron Doctrine. History of Neuroscience Series No.6. Oxford: Oxford University Press.
WAXMAN, S. G. Y J. DEGROOT (1995) CorrelativeNeuroanatomy, 22a ed. Norw alk, CT: Appletonand Lange.
nerviosas en circuitos y éstos en sistemas que procesan tipos específicos de in-formación pertinente a percepción y acción. Los objeti vos que quedan soncomprender de qué modo los fenómenos genéticos moleculares básicos estánligados a las funciones celulares, de los circuitos y los sistemas; saber cómofallan estos procesos en las enfermedades neurológicas y psiquiátricas, y co-menzar a comprender las funciones especialmente complejas del encéfalo quenos convierten en seres humanos.
A693-01.qxd 8/3/06 11:41 AM Page 30