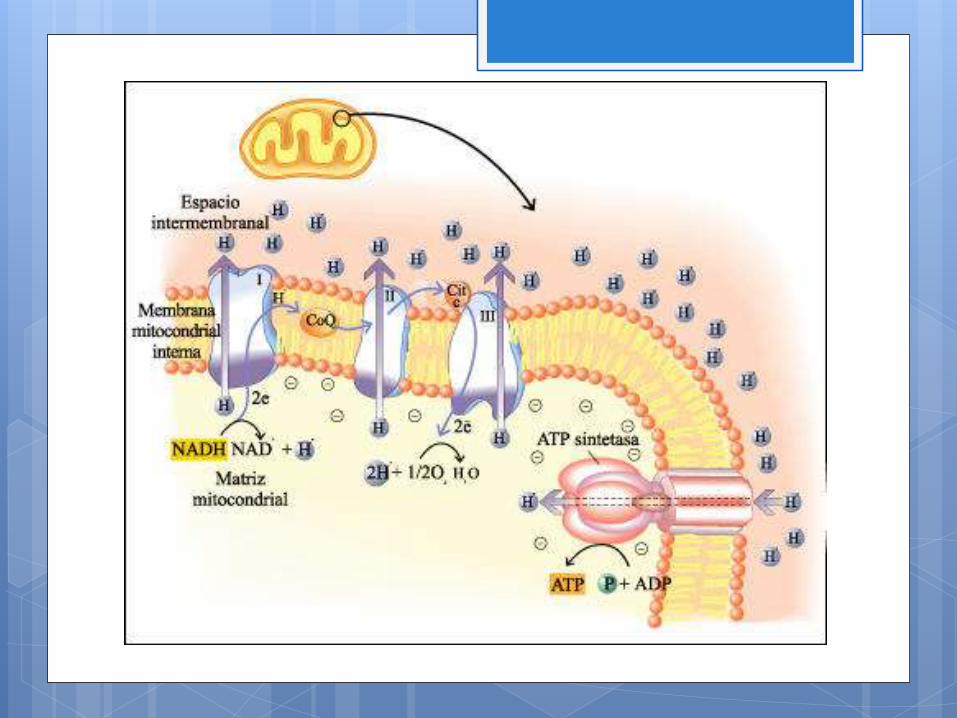
cadena de transporte de electrones
DESCRIPTION
Bioquímica: descripción de la Cadena Transportadora de Electrones y Fosforilación Oxidativa (síntesis de ATP)TRANSCRIPT

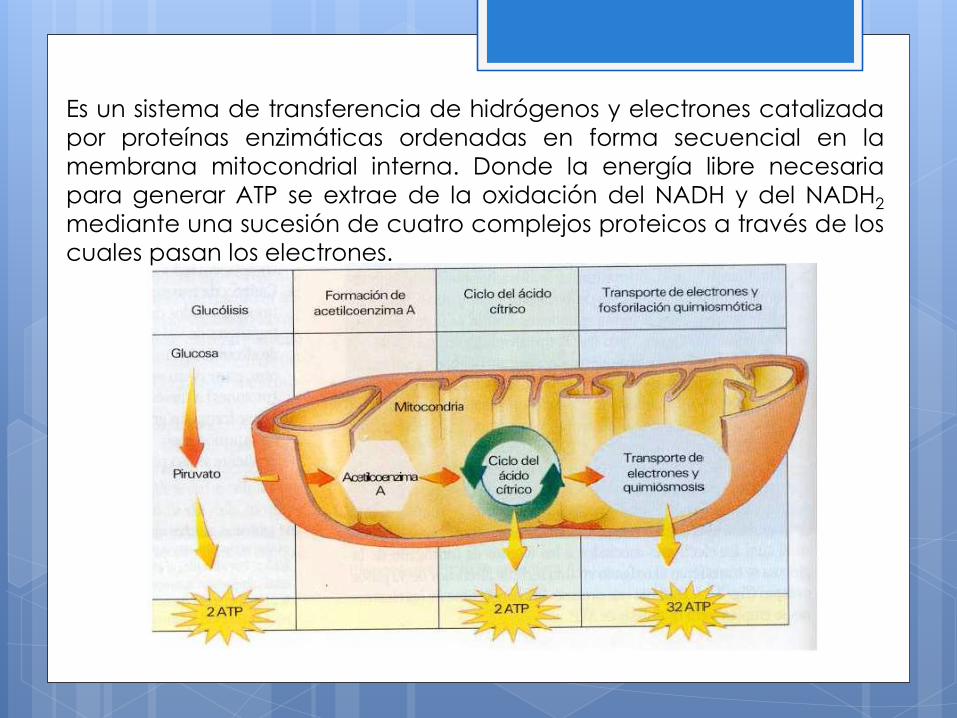
Es un sistema de transferencia de hidrógenos y electrones catalizada
por proteínas enzimáticas ordenadas en forma secuencial en la
membrana mitocondrial interna. Donde la energía libre necesaria
para generar ATP se extrae de la oxidación del NADH y del NADH2
mediante una sucesión de cuatro complejos proteicos a través de los
cuales pasan los electrones.
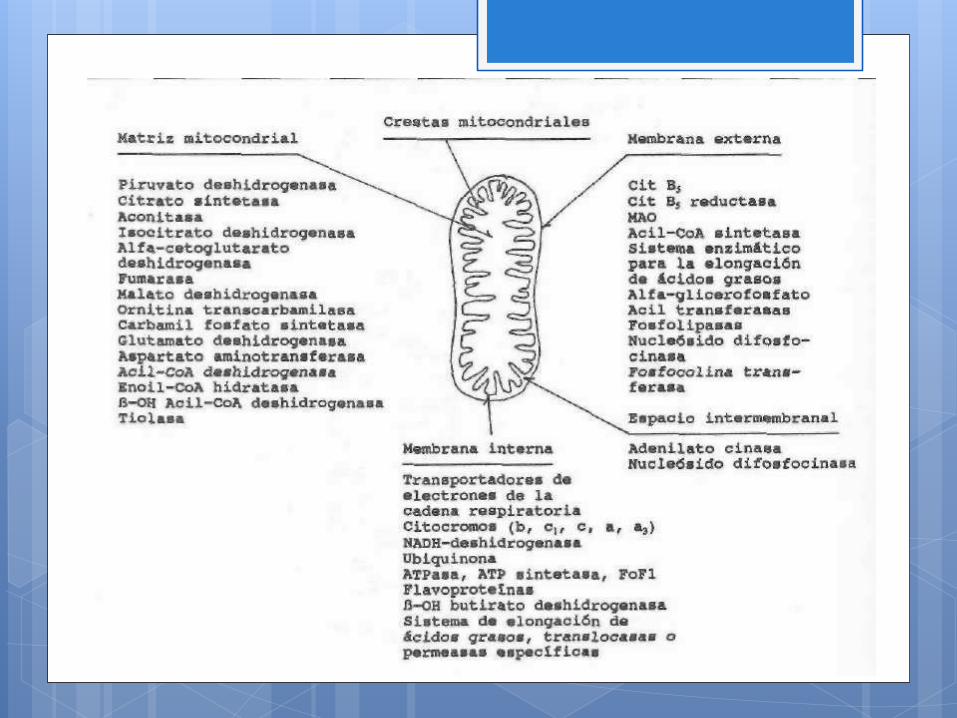

La cadena respiratoria constituye la última etapa de la
oxidación de los principales nutrientes. Es la etapa en la que se
conserva, en forma de ATP, la mayor parte de la energía
contenida en aquéllos y donde el oxígeno actuando como
aceptor final de los electrones aportados por los sustratos de la
cadena respiratoria- se transforma en agua.
La cadena respiratoria está formada por 2 procesos:
El transporte de electrones, que se Lleva a cabo en la
cadena transportadora de electrones -proceso exergónico
que genera un gradiente de protones.
El mecanismo de la fosforilación oxidativa proceso
endergónico acoplado al anterior
Dicho acoplamiento energético ocurre, como ya conocemos, en la
membrana interna de la mitocondria.

Desde los sustratos orgánicos al oxigeno molecular, participan cuatro
tipos de enzimas de oxido-reducción o de proteínas de transferencia
electrónica. Son los siguientes:
• Las deshidrogenasas piridín-dependientes que necesitan NAD o
NADP como coenzima.
• Las deshidrogenasas flavín-dependientes, que contienen
flavínadeníndinucleotido (FAD) o flavínmononucleotido (FMN)
como grupo prostético.
NADH deshidrogenasa y la succínico deshidrogenasa. La primera
tiene como grupo prostético al FMN y la segunda, al FAD. Ambas
flavinas se unen a la enzima de forma covalente. La segunda enzima
no es otra que la enzima que cataliza la sexta reacción del ciclo de
Krebs.

• Las ferrosulfoproteínas. En la mayoría de los complejos, estas
proteínas intervienen como transportadores finales de electrones.
Se encuentran presentes en los complejos I, II y III. El transporte de
electrones se efectúa en uno de los hierros de estos centros,
donde este pasa de la forma Fe3+ a Fe2+.
• Los citocromos, que contienen un grupo prostético ferroporfirina.
Intervienen en el transporte de electrones del complejo IV , pues el
núcleo de cobre cambia su estado de oxidación de Cu2+ a Co+ al
ocurrir el transporte de electrones de ese complejo. Hay uno
asociado al citocromo a y otro al citocromo a3.
Además de estas proteínas, actúa también en el transporte
electrónico la Ubiquinona o coenzima Q, de naturaleza liposoluble.
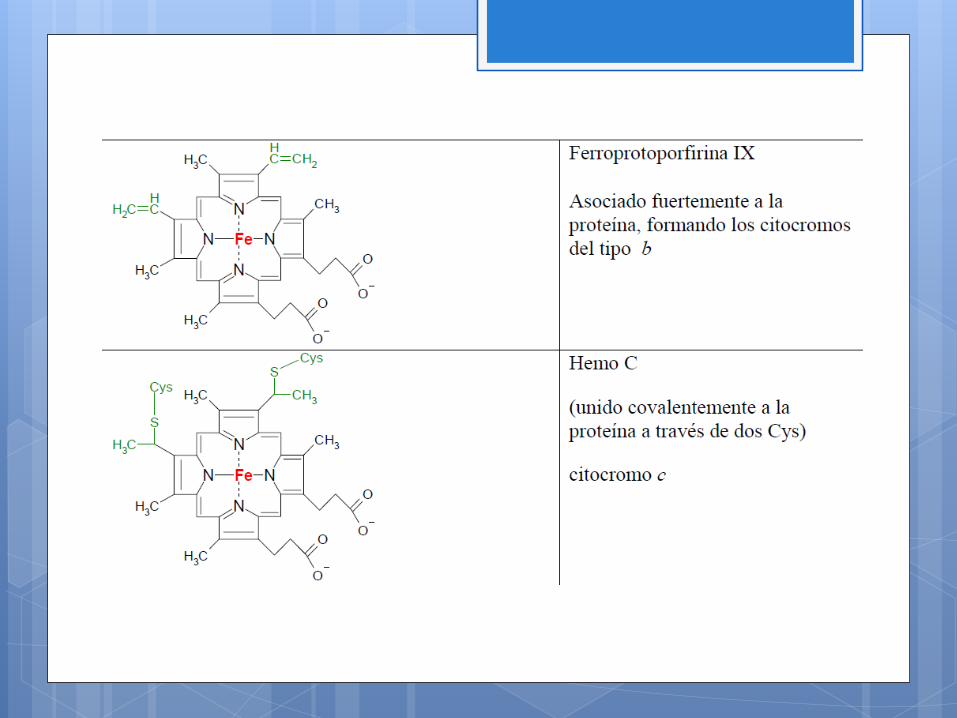
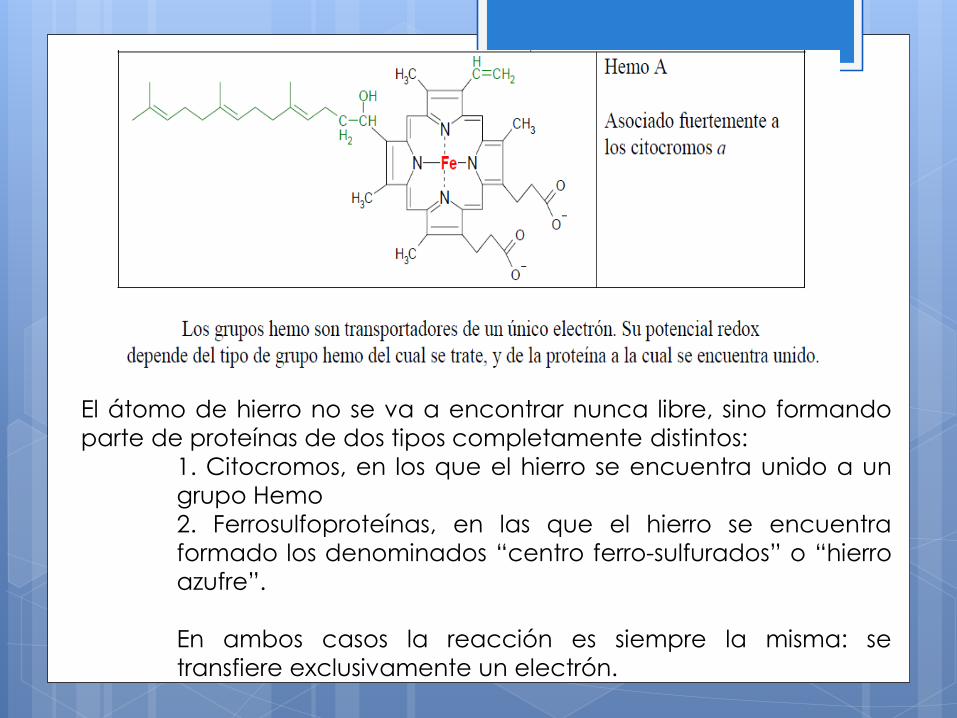
El átomo de hierro no se va a encontrar nunca libre, sino formando
parte de proteínas de dos tipos completamente distintos:
1. Citocromos, en los que el hierro se encuentra unido a un
grupo Hemo
2. Ferrosulfoproteínas, en las que el hierro se encuentra
formado los denominados “centro ferro-sulfurados” o “hierro
azufre”.
En ambos casos la reacción es siempre la misma: se
transfiere exclusivamente un electrón.
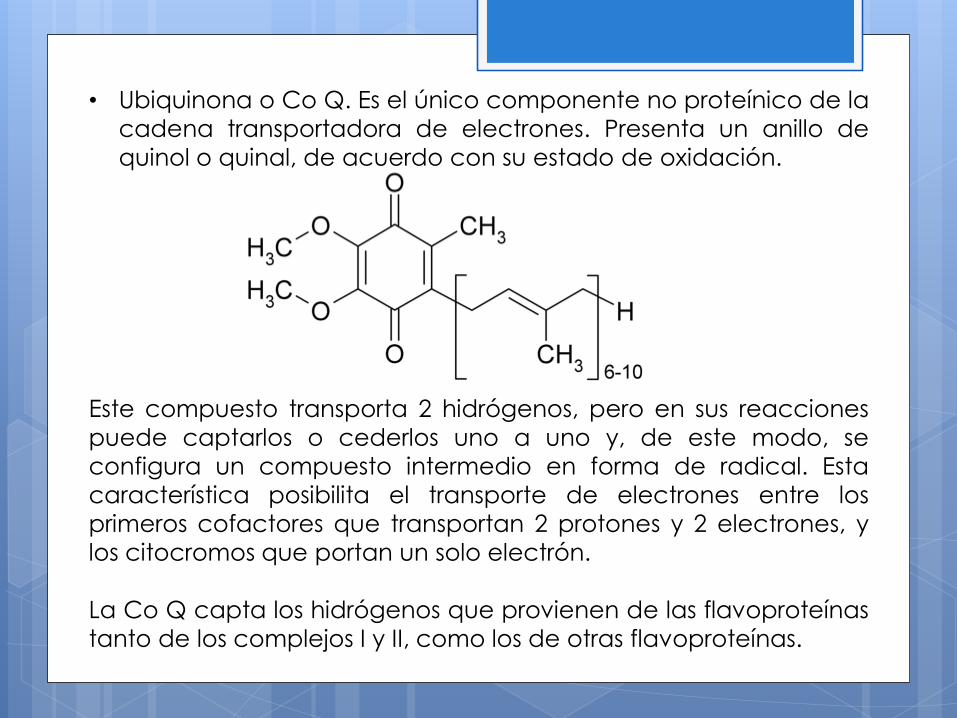
• Ubiquinona o Co Q. Es el único componente no proteínico de la
cadena transportadora de electrones. Presenta un anillo de
quinol o quinal, de acuerdo con su estado de oxidación.
Este compuesto transporta 2 hidrógenos, pero en sus reacciones
puede captarlos o cederlos uno a uno y, de este modo, se
configura un compuesto intermedio en forma de radical. Esta
característica posibilita el transporte de electrones entre los
primeros cofactores que transportan 2 protones y 2 electrones, y
los citocromos que portan un solo electrón.
La Co Q capta los hidrógenos que provienen de las flavoproteínas
tanto de los complejos I y II, como los de otras flavoproteínas.

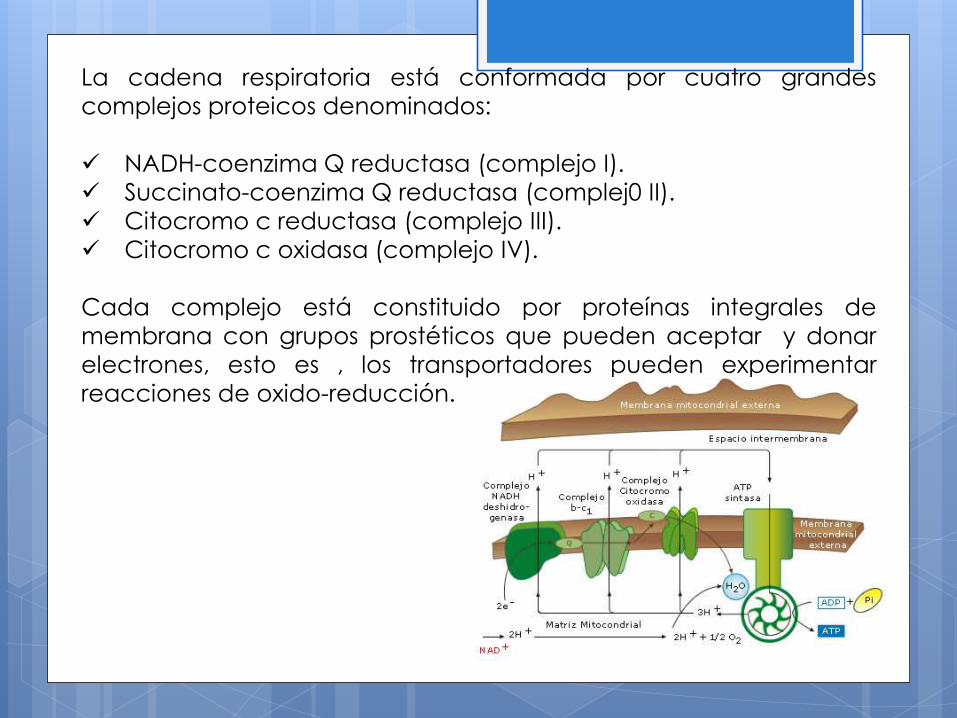
La cadena respiratoria está conformada por cuatro grandes
complejos proteicos denominados:
NADH-coenzima Q reductasa (complejo I).
Succinato-coenzima Q reductasa (complej0 II).
Citocromo c reductasa (complejo III).
Citocromo c oxidasa (complejo IV).
Cada complejo está constituido por proteínas integrales de
membrana con grupos prostéticos que pueden aceptar y donar
electrones, esto es , los transportadores pueden experimentar
reacciones de oxido-reducción.
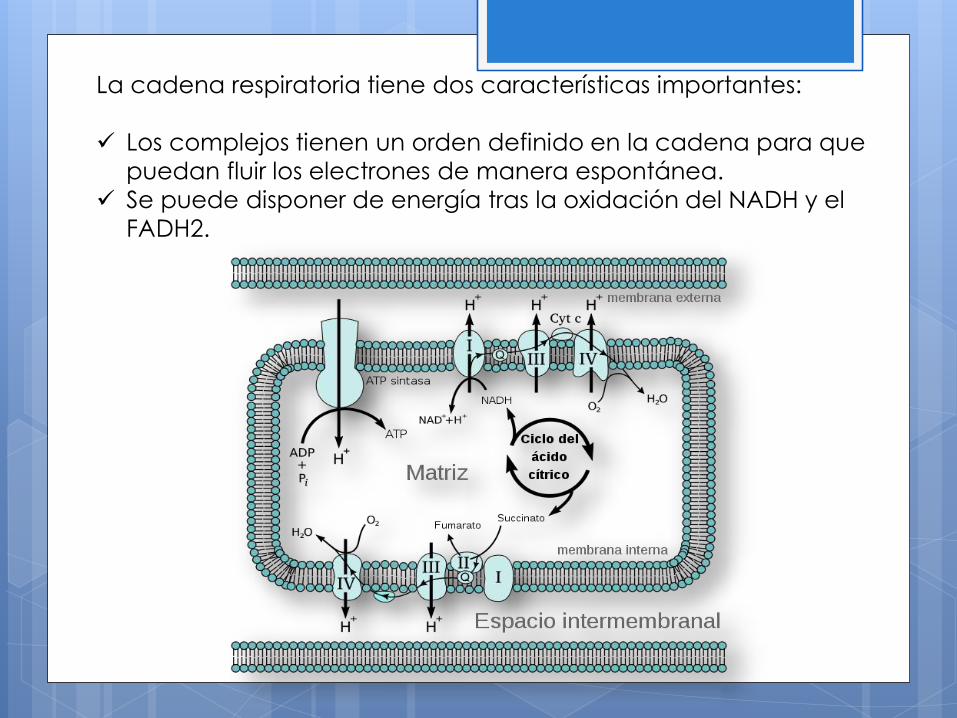
La cadena respiratoria tiene dos características importantes:
Los complejos tienen un orden definido en la cadena para que
puedan fluir los electrones de manera espontánea.
Se puede disponer de energía tras la oxidación del NADH y el
FADH2.
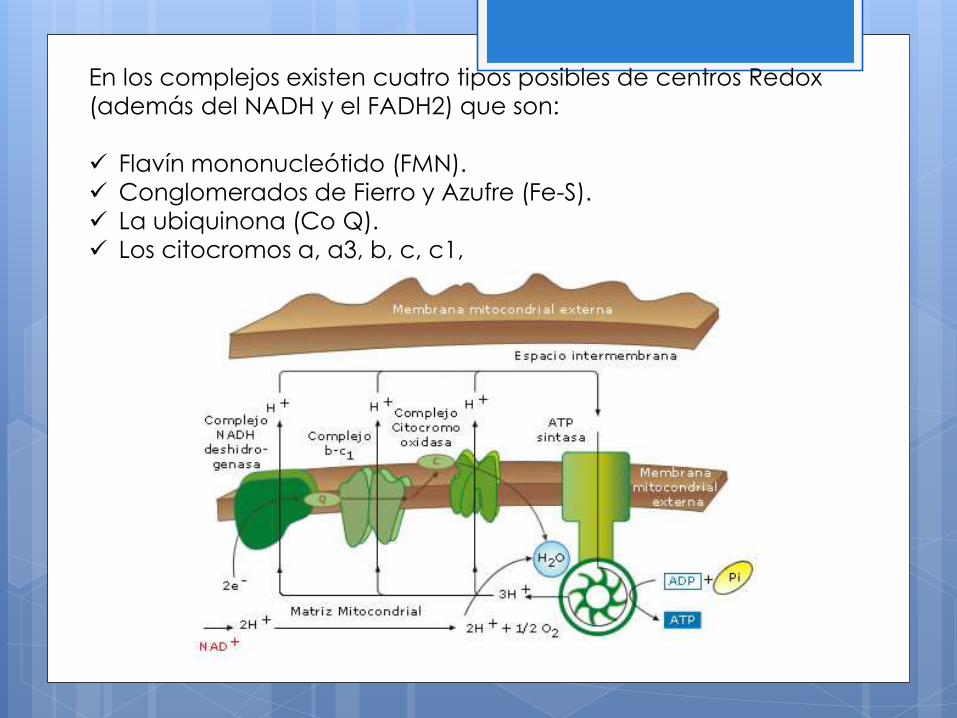
En los complejos existen cuatro tipos posibles de centros Redox
(además del NADH y el FADH2) que son:
Flavín mononucleótido (FMN).
Conglomerados de Fierro y Azufre (Fe-S).
La ubiquinona (Co Q).
Los citocromos a, a3, b, c, c1,

Complejo I (NADH-Coenzima Q Reductasa). Es el más grande y
contiene, a su vez, 3 subfracciones. Una es insoluble en agua, la
llamada HP, a la que quedan asociados los fosfolípidos que forman
parte de este complejo I. Ninguna de sus proteínas es catalítica. Las
otras 2 subfracciones, la FP y la IP, son solubles en agua. La FP es la
que contiene la flavoproteína catalítica. Las 3 subfracciones
contienen las diferentes proteínas Fe-S. La función del complejo les la
de oxidar al NADH y reducir la Co Q.
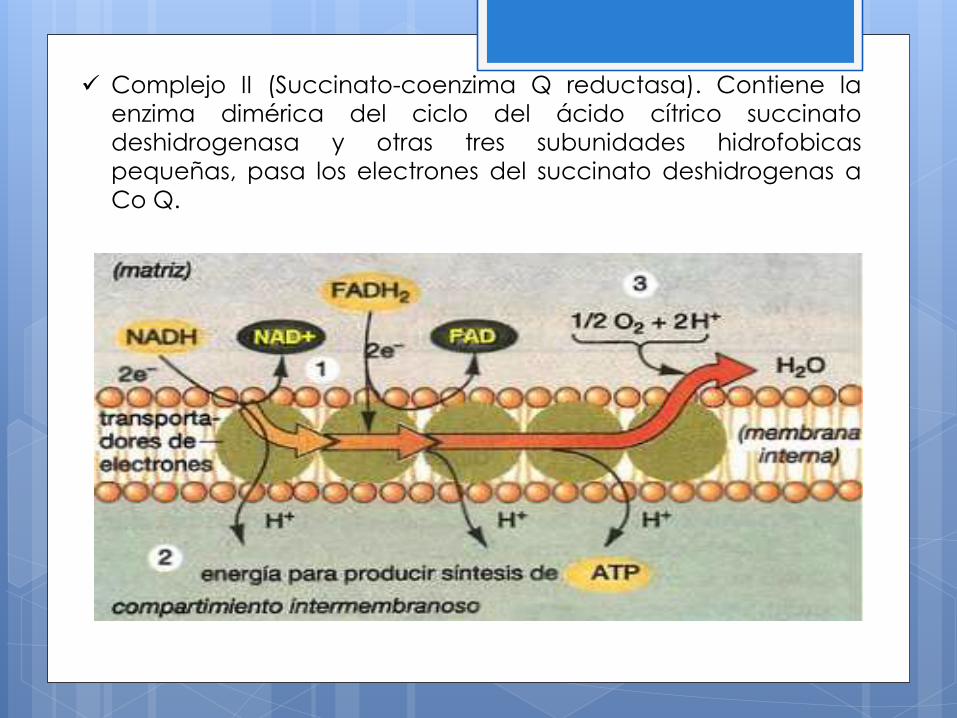
Complejo II (Succinato-coenzima Q reductasa). Contiene la
enzima dimérica del ciclo del ácido cítrico succinato
deshidrogenasa y otras tres subunidades hidrofobicas
pequeñas, pasa los electrones del succinato deshidrogenas a
Co Q.
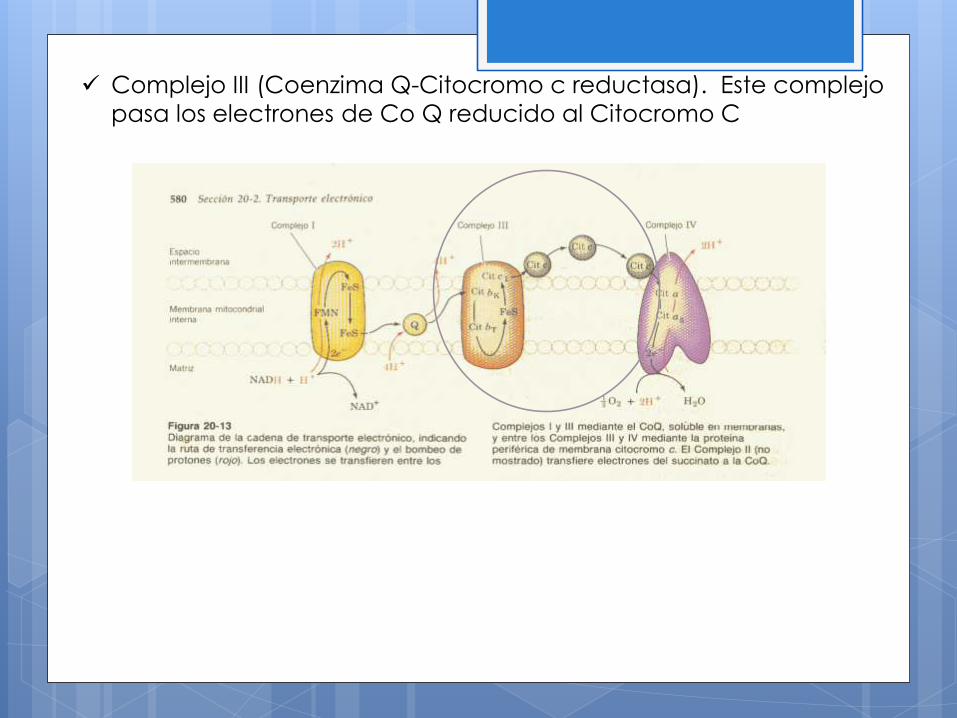
Complejo III (Coenzima Q-Citocromo c reductasa). Este complejo
pasa los electrones de Co Q reducido al Citocromo C
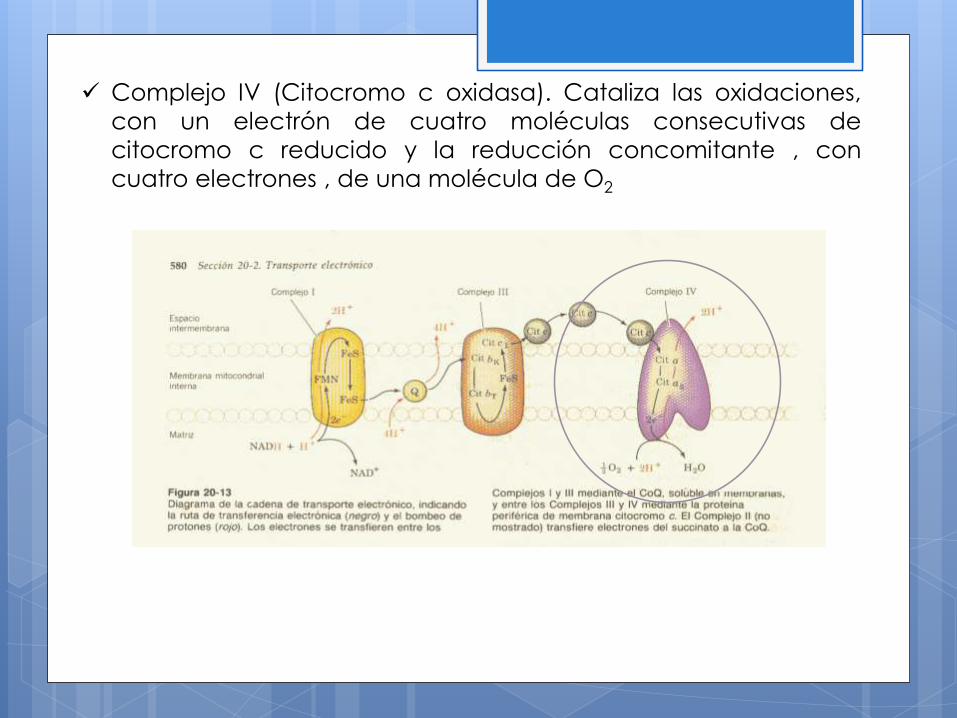
Complejo IV (Citocromo c oxidasa). Cataliza las oxidaciones,
con un electrón de cuatro moléculas consecutivas de
citocromo c reducido y la reducción concomitante , con
cuatro electrones , de una molécula de O2
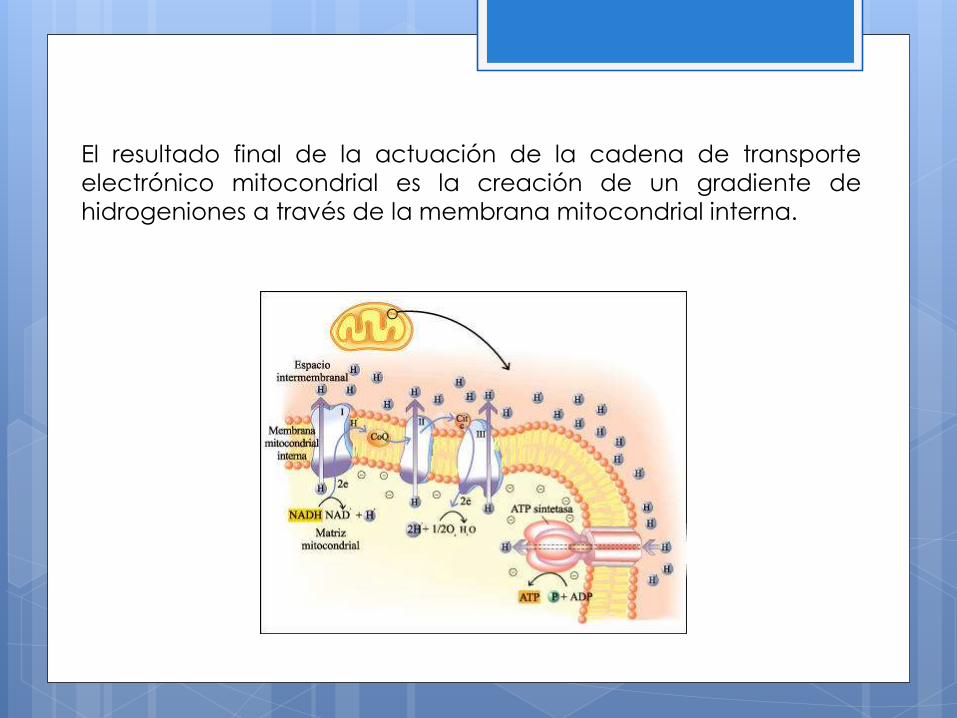
El resultado final de la actuación de la cadena de transporte
electrónico mitocondrial es la creación de un gradiente de
hidrogeniones a través de la membrana mitocondrial interna.

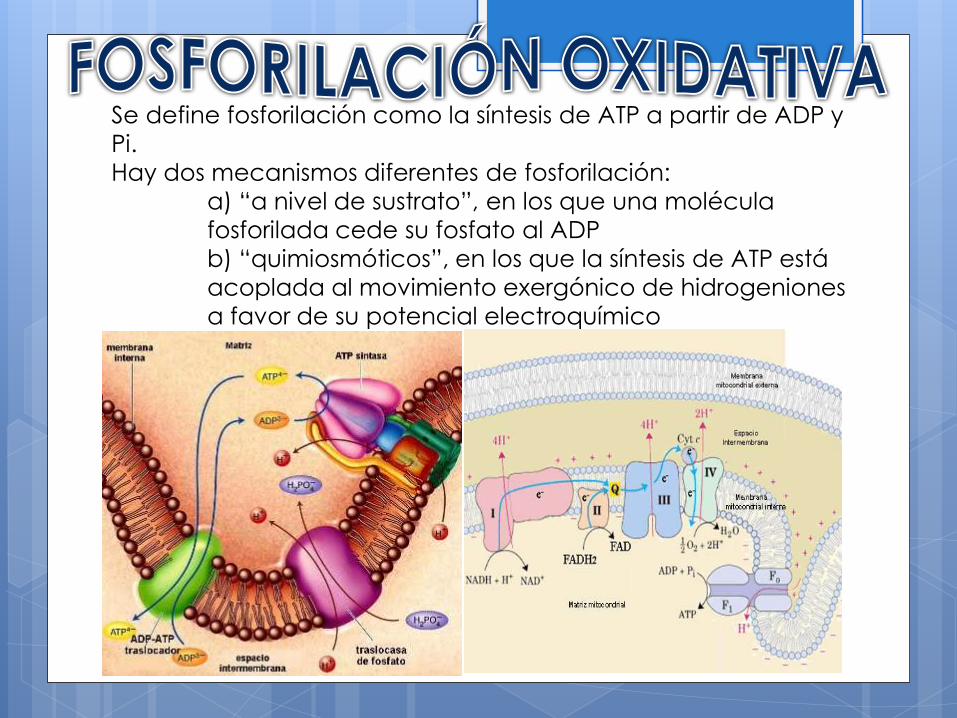
Se define fosforilación como la síntesis de ATP a partir de ADP y
Pi.
Hay dos mecanismos diferentes de fosforilación:
a) “a nivel de sustrato”, en los que una molécula
fosforilada cede su fosfato al ADP
b) “quimiosmóticos”, en los que la síntesis de ATP está
acoplada al movimiento exergónico de hidrogeniones
a favor de su potencial electroquímico

La mayor parte de la energía utilizable obtenida de la degradación
de los hidratos de carbono o de la grasas, deriva de la fosforilación
oxidativa que tiene lugar en el interior del mitocondria.
La degradación de la glucosa mediante la glicolisis y el ciclo del
acido cítrico rinde un total en cuatro moléculas de ATP, diez
moléculas de NADH, y dos de FADH2 son transferidos después al
oxigeno molecular, lo cual está acoplado a la formación de 32 a 34
moléculas de ATP mediante la fosforilación oxidativa.

Se define la fosforilación oxidativa como la síntesis de ATP a partir de
ADP y Pi acoplada a la transferencia de electrones desde un donador
reducido a un aceptor final.
La energía del proceso deriva, precisamente, de este proceso redox.
En eucariotas, el donador último de electrones es siempre un
compuesto orgánico que se oxida por los nucleótidos de nicotinamida
o de flavina.
Tenemos que considerar dos apartados diferentes en la FO:
1) La cadena de transferencia de electrones desde el
donador inicial al aceptor final: es la cadena respiratoria, y
este proceso lo vamos a llamar respiración.
2) La síntesis de ATP propiamente dicha empleando la energía
liberada en la transferencia de electrones.

Propuesta por primera vez en 1961 por Peter Mitchell, que sugirió
que el ATP se genera utilizando la energía almacenada en
forma de un gradiente de protones a través de las membranas
biológicas, en lugar de por una transferencia química directa de
grupos ricos en energía.

l. Al ser transportados los electrones por los complejos dela cadena
respiratoria, se crea un gradiente de protones.
2. La membrana interna dela mitocondria es impermeable a los
protones, puesto que de lo contrario no se forma el gradiente.
3. Los transportadores de electrones o complejos están
organizados en la membrana de forma vectorial de modo que los
protones son extraídos de la matriz hacia el espacio
intermembranoso y así se genera un gradiente de ellos.
4. La ATPasa también está situada de forma vectorial en la
membrana y libera el ATP sintetizado por ella hacia la matriz, y se
pone en contacto en los protones por el espacio
intermembranoso.

Complejo V
ATP sintasa o
ATP sintetasa
Unidad de
fosforilación

FUNCIONES
Situada en la membrana interna de la mitocondria, se han asociado
a este complejo V, 2 funciones, ambas debidas a la actividad
reversible de la enzima:
1. La síntesis de ATP, acoplada a la energía que brinda el gradiente
de protones, y que se forma durante el transporte de electrones.
2. La hidrólisis de ATP, al acoplarse a la traslocación de protones de
la matriz al citosol, con el paso de los cationes como el K', Na*, Ca".
El complejo V lo forman 3 porciones: la cabeza, la base y el cuello.

La cabeza, actualmente conocida como subunidad F1, en ellas se
localiza la actividad de síntesis de ATP. Están unidas por unos tallos,
el cuello, a la membrana, donde se encuentra la tercera parte que
se corresponde con la base. Esta ultima es la subunidad F0 ,
también conocida como el canal de protones, por donde estos
pasan al disiparse el gradiente durante el mecanismo de formación
del ATP.
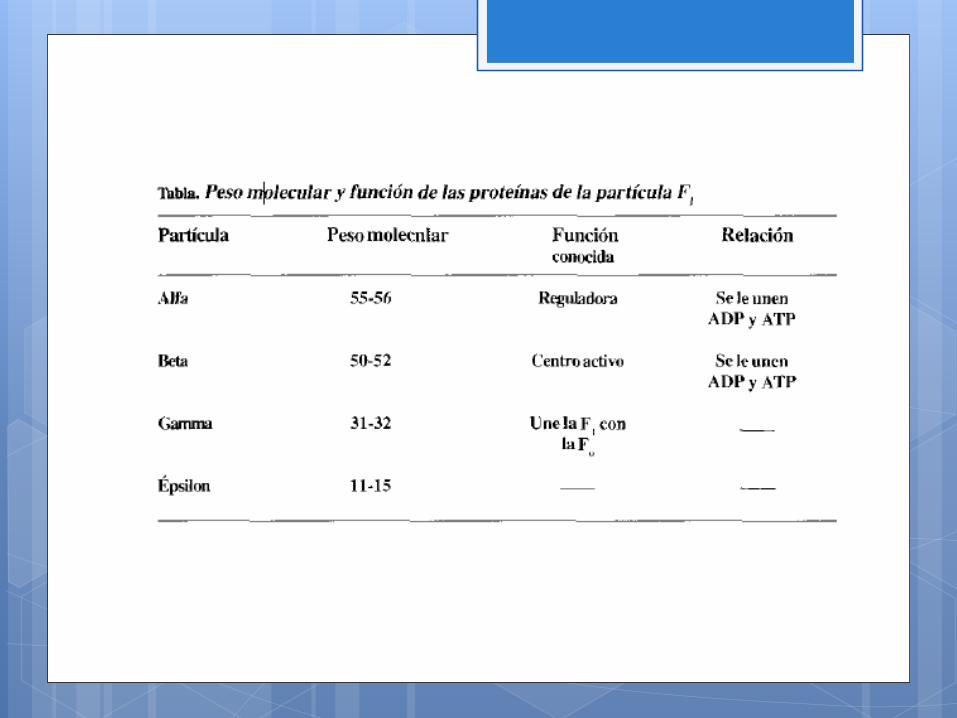
La F1, contiene5 clases de proteínas: dfa, beta, gamma, delta y
épsilon. La relación mediante la que se unen es de alfa, beta,
gamma, delta, épsilon

El tallo o cuello, ausente del complejo V en procariotas y
cloroplastos, pero presente en el complejo de las mitocondrias,
está formado por 2 proteínas: la F6que es indispensable para la
unión de la F1, con la F0.
La base del complejo V, o subunidad F0 unida a la membrana
interna, contiene numerosas proteínas hidrofóbicas, proteolípidos, y
es la traslocadora de protones o canal de protones.