bioseñalización - iib-intech · bioseñalización capitulo 12. lehninger: principios de...
TRANSCRIPT

Bioseñalización
Capitulo 12. Lehninger: Principios de bioquímica / Lehninger, Albert L.; Nelson,
David L.; Cox, Michael M. Lenhinger

Bioseñalización : Generalidades
La capacidad que tienen las células de censar y dar una respuesta ante
un estimulo mas allá de su membrana plasmática es fundamental para la
vida
El crecimiento, la diferenciación y metabolismo de diferentes tejidos y
órganos de nuestro organismo dependen de los estímulos que recibe la
célula

Componentes de la vía de señalización
Bioseñalización : Generalidades
1. Fase intercelular: liberación de una
sustancia portadora de un mensaje (primer
mensajero)a a partir de la célula efectora hasta
la llegada de este al interior de la célula que va
a dar respuesta al mensaje, célula diana
2. Fase intracelular: todos los procesos y las
substancias implicadas en la producción de la
respuesta celular (segundos mensajeros,
enzimas, proteínas estructurales, genes etc)
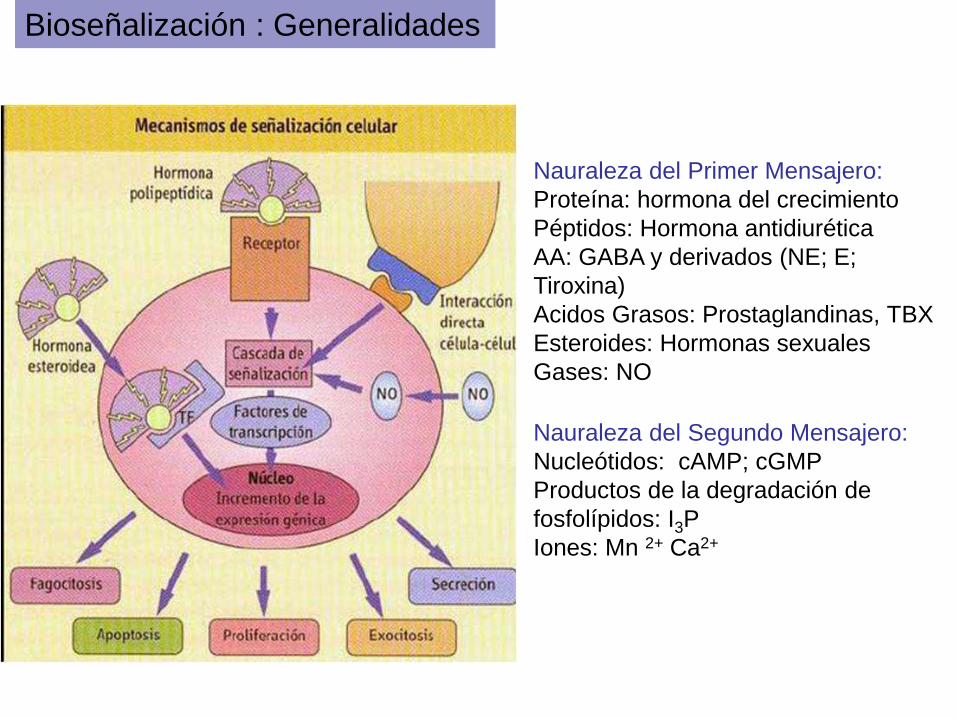
Bioseñalización : Generalidades
Nauraleza del Primer Mensajero:
Proteína: hormona del crecimiento
Péptidos: Hormona antidiurética
AA: GABA y derivados (NE; E;
Tiroxina)
Acidos Grasos: Prostaglandinas, TBX
Esteroides: Hormonas sexuales
Gases: NO
Nauraleza del Segundo Mensajero:
Nucleótidos: cAMP; cGMP
Productos de la degradación de
fosfolípidos: I3P
Iones: Mn 2+ Ca2+

Bioseñalización : Generalidades
1. Especificidad: La molécula señal se acopla al
sitio de unión de su receptor complementario y no
a otros
2. Amplificación: Luego de la
detección de la señal por la
molécula receptora se produce
un segundo mensajero que
incrementa geométricamente la
cascada de señalización.

3. Desensibilización/adaptación : La activación del
receptor pone en marcha un circuito de
retroalimentación que desconecta el receptor o lo
elimina de la superficie de la célula.
4. Integración: Cuando dos señales tienen
efectos opuestos en una característica
metabólica tal el resultado regulador proviene
de la información integrada de ambos
receptores
Bioseñalización : Generalidades

1.Son los traductores de señales mas sencillos. Ej: Canal iónico de receptor de Ach
2.El receptor al ser estimulado por un ligando cataliza la producción de un 2do mensajero. Ej: receptor de Insulina3. Receptores serpentinas que están acoplados a una proteína G que cataliza la producción de un 2do mensajero. Ej:
receptor b- adrenérgico.
4. Receptores nucleares que directamente se unen a una zona especifica del gen y lo modulan
5. Receptores sin actividad enzimática propia, atraen enzimas citosólicas y activan una cascada de señalización hacia el
nucleo . EJ. Receptor Epo
6. Receptores de adhesión que interaccionan con por Ej el colágeno y llevan al citoesqueleto instrucciones para migrar.
Ej: receptor de integrinas
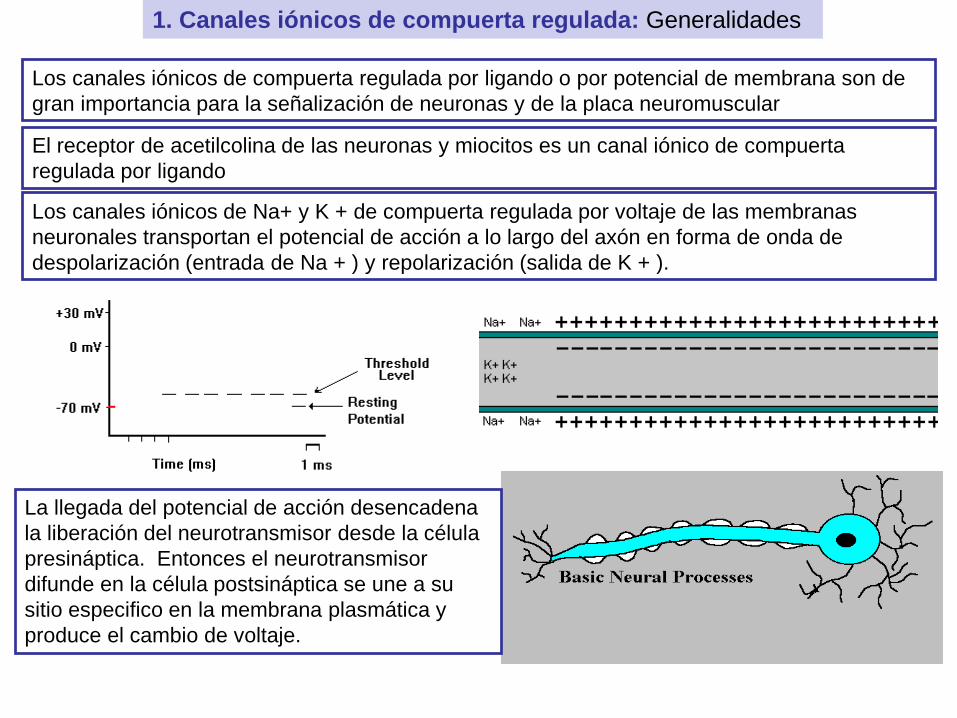
1. Canales iónicos de compuerta regulada: Generalidades
Los canales iónicos de compuerta regulada por ligando o por potencial de membrana son de
gran importancia para la señalización de neuronas y de la placa neuromuscular
El receptor de acetilcolina de las neuronas y miocitos es un canal iónico de compuerta
regulada por ligando
Los canales iónicos de Na+ y K + de compuerta regulada por voltaje de las membranas
neuronales transportan el potencial de acción a lo largo del axón en forma de onda de
despolarización (entrada de Na + ) y repolarización (salida de K + ).
La llegada del potencial de acción desencadena
la liberación del neurotransmisor desde la célula
presináptica. Entonces el neurotransmisor
difunde en la célula postsináptica se une a su
sitio especifico en la membrana plasmática y
produce el cambio de voltaje.

Potencial eléctrico de transmembrana (Vm)
a)La ATP asa Na+/K+ electrogénica
produce un potencial eléctrico de
transmembrana de -60mV (negativo
hacia el interior)
b) Existe un gradiente electroquímico
de iones a desplazarse hacia el
interior de la célula despolarizándola
(Na+ y Ca2+). Y un gradiente de
hiperpolarizacion de salida de Cl- y K+
(en contra de su gradiente de
concentración)
Mantener la osmolaridad
Impulso eléctrico
Traducción de señales

1. Receptor nicotínico de acetilcolina
El receptor de Ach se localiza en la neurona
postsináptica o miocito
La Ach se libera a la hendidura sináptica o la placa
neuromuscluar por la secreción de vesículas inducida por
la entrada de Ca 2+ producida por la despolarización de la
membrana
Esto produce un nuevo potencial de acción y la apertura
de canales iónicos regulados por voltaje (impulso
eléctrico) o la contracción del musculo
Luego actua la acetilcolinesterasa (enzima que degrada la Ach)

3 tipos de canales iónicos regulados por voltaje
Impulso neuronal
1 2
3
1. Canales Na+ regulados por voltaje ubicados a lo largo de
todo el axón. Estan cerrados cuando la membrana esta en
reposo (-60 mv), cuando la membrana se despolariza, se
abren localmente por acciòn de la Ach.
2. La despolarización inducida por la apertura de canales
Na+ produce la apertura de canales K+ voltaje
dependientes con la resultante salida de K+ y
repolarización de la membrana (local).
Un pulso corto de despolarización atraviesa el axón a medida que
la despolarización-repolarización local origina la apertura de
canales vecinos de Na+ voltaje dependientes y es unidireccional
porque el canal tiene un periodo refractario durante el cual no
puede volver a abrirse
3. En el extremo distal del axón hay canales de Ca2+ regulados por voltaje. Cuando la
onda de despolarización los alcanza se abren y el Ca 2+ (actúa como 2do mensajero)
entra y desencadena la liberación de Ach

2. Enzima Receptor: ej Receptor de Insulina
La unión del ligando en la superficie del receptor
produce la autofosforilación de residuos de
tirosina en el sitio activo (actividad tirosina
quinasa).
Estos tipos de receptores tienen un sitio de unión
a ligando hacia el espacio extracelular y un sitio
activo enzimático de cara al citoplasma celular
Esta autofosforilación permite la apertura del sitio
activo permitiendo que la enzima fosforile
residuos tirosina de otras proteínas diana.
Una de esas proteínas diana es el IRS-1
(sustrato-1 del receptor de insulina)
La insulina regula el metabolismo en general (glucosa)
y el crecimiento y desarrollo celular

Cuando el IRS-1 se fosforila en sus residuos
tirosina, se produce la nucleación
La nucleación involucra un complejo
de proteínas que transmiten el
mensaje desde el receptor de
insulina a otras proteínas diana
finales del citosol a través de una
cadena de proteínas intermedias:
Grb2 (por el dominio SH2), que a su
vez recluta a Sos y a su vez a Ras
el cual libera GDP y el sitio de unión
se sustituye por GTP
Asì RAS unido a GTP puede
activar a Raf-1
Raf-1 es la primera de 3 proteínas quinasas (Raf-1,
MEK, ERK) que son miembros de la familia de las
MAPK (mitogen activated phosphorylate kinase) que en
definitiva luego de las cascadas de fosforilación
producen un efecto mitógeno en la célula a nivel génico

La Gbr2 no es la única proteína asociada a IRS-1El IRS-1 fosforilado puede
activar a PI-3K a través
del dominio SH2 y dicha
enzima convierte al PIP2
en PIP3 (localizado en la
membrana plasmática)
PIP3 fosforila a la PKB
que a su vez fosforila a
la GSK3 (glucógeno
sintasa). La forma
activa de la GSK3 es no
fosforilada. Entonces al
fosforilzarse se inhibe.
Entonces la síntesis e
glucógeno se mantiene
activa
Existe un mecanismo de inactivación de esta cascada comandado por PTEN
(fosfatasa específica de PIP3) desfosforila al PIP3 en posición 3 para dar el PIP2
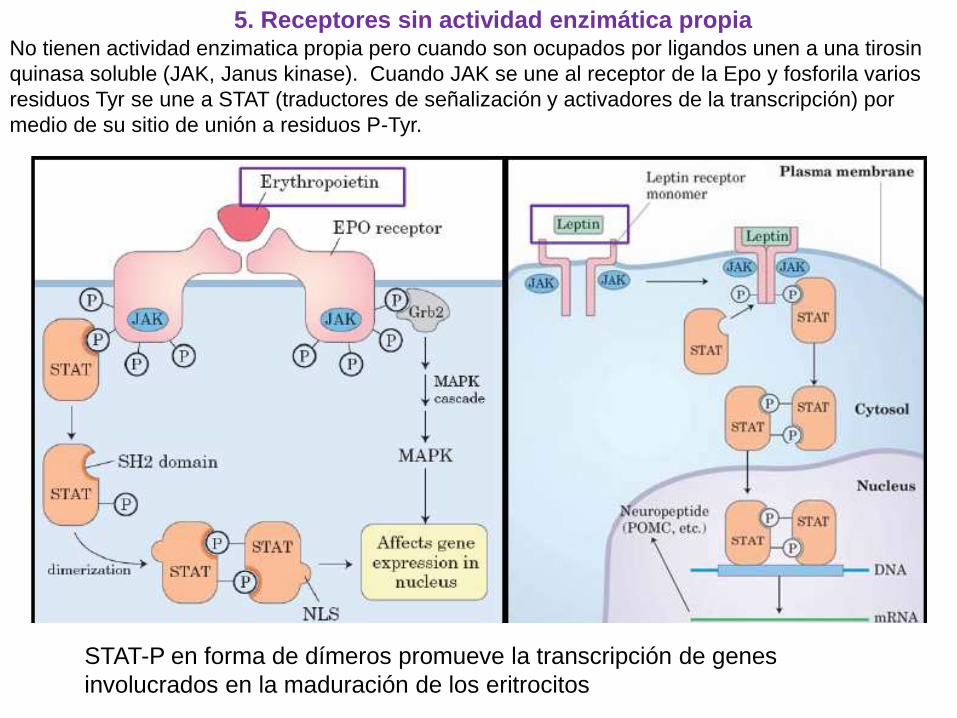
5. Receptores sin actividad enzimática propiaNo tienen actividad enzimatica propia pero cuando son ocupados por ligandos unen a una tirosin
quinasa soluble (JAK, Janus kinase). Cuando JAK se une al receptor de la Epo y fosforila varios
residuos Tyr se une a STAT (traductores de señalización y activadores de la transcripción) por
medio de su sitio de unión a residuos P-Tyr.
STAT-P en forma de dímeros promueve la transcripción de genes
involucrados en la maduración de los eritrocitos

Receptores con actividad GC
Receptores con actividad guanilato ciclasa, es decir,
producen GMPc a partir del GTP
El GMPc como segundo mensajero
activa la proteína quinasa dependiente de
GMPc (PKG) la que dependiendo del
tejido produce el efecto final:
-Excreción renal de Na+
-Relajación del musculo liso vascular
(vasodilatación; aumento del flujo)
-Liberación de Cl- al intestino estimulado
por la guanilina (péptido instestinal).
-Contracción menos enérgica del corazón
Estos receptores pueden estar localizados en la
membrana plasmática (ANF) o solubles en el
citoplasma celular (NO)
Existen PDE (fosfodiesteresas específicas) que convierten al GMPc en 5`GMP
inactivo
Sildenafil (Viagra) es un inhibidor especifico de la PDE de vasos sanguíneos

1.Son los traductores de señales mas sencillos. Ej: Canal iónico de receptor de Ach
2.El receptor al ser estimulado por un ligando cataliza la producción de un 2do mensajero. Ej: receptor de Insulina3. Receptores serpentinas que están acoplados a una proteína G que cataliza la producción de un 2do mensajero. Ej:
receptor b- adrenérgico.
4. Receptores nucleares que directamente se unen a una zona especifica del gen y lo modulan
5. Receptores sin actividad enzimática propia, atraen enzimas citosólicas y activan una cascada de señalización hacia el
nucleo . EJ. Receptor Epo
6. Receptores de adhesión que interaccionan con por Ej el colágeno y llevan al citoesqueleto instrucciones para migrar.
Ej: receptor de integrinas

3. Receptores serpentinas acoplados a proteína G
Componentes:
Un receptor de membrana de 7 pasos de que “serpentean” a lo largo de la
membrana plasmática, con su domino C-t hacia el citoplasma y el N-t al
exterior. (GPCR)
Una enzima localizada en membrana plasmática que produce un segundo
mensajero intracelular (Adenilato Ciclasa, sintetiza AMPc a partir de ATP)
Una tercer proteína que une un nucleótido de guanosina (Proteína G), que
establece el nexo entre el receptor y la producción del 2do mensajero

La unión del ligando al receptor
produce un cambio conformacional
que ocasiona el reemplazo de
GDP por GTP en el dominio Gs (o
subuniddad α (estimulatoria) de la
proteína G
Proteína G ≠ Ras (trimérica ≠ monomérica)
Existe un dominio o subunidad i (inhibitoria) que produce una inhibición de la AC, bajos
niveles de AMPc intracelular (mecanismo de acción de algunas hormonas)
3. Receptores serpentinas acoplados a proteína G
Las subunidades Gβ y Gγ se
disocian entonces Gα-GTP se
puede unir a la AC. Se desplaza
a lo largo del plano de la
membrana (se mantiene unida
por un grupo palmítico)
La AC cataliza la formación de
AMPc . Es autolimitante, ya la
subunidad activa de la Proteina Gs
al donar el Pi se inactiva. Y esta
disponible para actuar nuevamente

Luego de la generación de AMPc
Este puede activar a la PKA (protein
quinasa dependiente de AMPc
Efecto contrario a la insulina


Receptores acoplados a Proteína G
Utilizan como enzima a la PLC (fosfolipasa
C) que origina a partir del 4,5
fosfatidilinositol bifosfato de la membrana
plasmática dos segundos mensajeros:
IP3 (inositol 3 P)
DAG (diacilglicerol)
La subunidad de la proteína G involucrada
se llama Gq
El IP3 es hidrosoluble y disfunde desde la
membrana plasmática hacia el retículo
endoplasmico, se une a receptores
específicos de IP3 y permite la apertura de
canales Ca2+. Esta salida del calcio al
citosol provoca la activación de la PKC
El DAG coopera con el calcio en la
activación de la PKC
Los cambios en los niveles de calcio
intracelular pueden activar a su vez a una
quinasa dependiente de Ca2+/calmodulina
(CAM quinasas

Receptores acoplados a Proteína G
Existe una gran familia de receptores de 7 pasos de transmembrana. Al unir el
ligando el receptor cataliza el intercambio de GTP por GDP a una proteína G
asociada. Esta subunidad viaja por la membrana y estimula/inhibe a una enzima
próxima a la membrana (AC, PLC) alterando la concentración de un segundo
mensajero (AMPc; IP3, DAG, Ca2+)
En el caso del receptor beta adrenérgico al unirse la adrenalina a través de la
subunidad Gs de la proteína G activa a la AC que aumenta la concentración de
AMPc. Este activa la PKA que a su vez esta fosforila otras proteínas.
Algunos receptores están acoplados a PLC que rompe el PIP2 en DAG y IP3, los
cuales liberan calcio y activan a la PKC que finalmente es la que por fosforilación
de proteínas diana especificas. Estas fosforilaciones en algunos casos pueden ser
mediadas por la activación de la calmodulina.
http://www.youtube.com/watch?v=JeFXtpPQXjY&feature=related