barbastella 7
DESCRIPTION
ÂTRANSCRIPT



1Barbastella 7 (1) 2014
BarbastellaRevista Científica de Investigación en Quirópteros
Scientific Journal of Bat Research
Barbastella es una revista anual que tiene como objetivo principal ser una plataforma para dar a conocer estudios sobre quirópteros. La revista pretende dar cobertura a estudios llevados a cabo en cualquier localidad del mundo. Se pretende dar visibilidad a estudios realizados con rigor científico pero considerados de ámbito local por las revistas interna-cionales. Asimismo, se espera poder dar cabida a información sobre proyectos destacados y hallazgos de interés en el territorio. Barbastella is an annual journal that wants to provide a platform to publicize studies on bats. The journal aims to cover research carried out in all over the world. It aims to give visibility to studies conducted with scientific rigor, but considered as local by interna-tional journals. Also, we expect to accommodate data about major research projects and findings in the territory.
Juan Tomás AlcaldePresidente de la SECEMU

2 Barbastella 7(1) 2014

3Barbastella 7 (1) 2014
Barbastella 7 (1) Open Access ISSN: 1576-9720 SECEMU www.secemu.org
New records of the Alcathoe bat, Myotis alcathoe (Vespertilionidae) for Italy
Pier Paolo De Pasquale¹*, Andrea Galimberti²
¹ Wildlife consulting, Via G. Saragat 24, 70027 Palo del Colle (Bari), Italy¹ Italian Chiroptera Research Group, Università degli Studi dell’Insubria, via Dunant,3 Varese² ZooPlantLab, Università degli Studi di Milano-Bicocca, Dipartimento di Biotecnologie e Bioscienze, P.za della Scienza 2, 20126 (Milano), Italy
* Corresponding author e-mail: [email protected]
DOI: http://dx.doi.org/10.14709/BarbJ.7.1.2014.01
Spanish title: Nuevas citas de murciélago ratonero bigotudo pequeño, Myotis alcathoe (Vespertilionidae) en Italia
The presence of cryptic species seem to be widespread among vespertilionid bats and especially in the genus Myotis (Jones and Barlow 2004).
Myotis alcathoe is a recently described cryptic species (von Helversen et al. 2001) belonging to the group mystacinus. This small bat is difficult to identify only through morphological analysis.
Although the species can be identified using both morphological characters and genetic markers, only the later provide full confidence about it (Niermann et al. 2007). In Europe population size and trends of M. alcathoe are poorly known.
In Italy the species was recorded for five localities in Abruzzo (central Italy) and Campania (southern Italy). All bats were captured in beech forests (Tereba et al. 2008, Galimberti et al. 2012).
Our records were collected as part of a survey of bats covering the entire territory of the Appennino Lucano Val d’Agri Lagonegrese National Park (south-west of the Basilicata region). We made two capture sessions on 3 and 6 August, 2012 near two small mountain lakes at 1167 and 1304 m a.s.l., respectively. Small lakes are part of a forest reserve dominated by beech (Fagus sylvatica) associated with other tree species, such as Lobel maple (Acer lobelii), sycamore (Acer pseudoplatanus) and Small-leaved Lime (Tilia cordata), fig. 1.
Bats were captured with 2.5 x 12 m mist-nets (50 denier, 38 mm mesh) placed on commuting routes and near small lakes used by bats for drinking and foraging.
The nets were deployed half an hour after sunset and kept in place for 4 hours. Forearm length and body mass of each bat trapped were measured using a digital callipers (± 0.1 mm) and a pesola digital scale (± 0.1 g). Wings were trans-
New records of the Alcathoe bat, Myotis alcathoe (Vespertilionidae) for Italy
Abstract: The Alcathoe bat (Myotis alcathoe) is a recently described cryptic species; in Europe its distribution range is poorly known. In Italy this species has been recorded in a small number of locations in Abruzzo (central Italy) and Campania (southern Italy). Our report refers to three bats captured in a mountainous area dominated by forest habitats in the Appennino Lucano Val d’Agri Lagonegrese National Park (Basilicata region). The iden-tification of bats captured was confirmed by molecular analysis using the technique of DNA barcoding. In this paper we present new recordings that highlight the presence of the species in other regions of southern Italy and that help define its distributional status in Europe.
Key words: Myotis alcathoe, cryptic species, DNA barcoding, coxI.
accepted: October 4, 2013

4 Barbastella 7(1) 2014
illuminated to distinguish juveniles from adults (Antony 1988).
Bats captured were subjected to a skin biopsy using a sterile punch of a 3 mm diameter from the tail membrane (uropatagium) (Worthington Wilmer and Barratt 1996).
Samples were stored in sterile tubes containing 95% ethanol for subsequent molecular analysis.
Total genomic DNA was extracted from tissue samples using 5 PRIME, ArchivePure DNA Purification Kit. A fragment of ca. 650 bp of the mitochondrial subunit 1 of cytochrome c oxidase, suitable for echolocating bats identification (Galimberti et al. 2012), was amplified for the three sampled bats using the primers VF1d 5’- TTCTCAACCAACCACAARGAYATYGG-3’ and VR1d 5’-TAGACTTCTGGGTGGCCRAARAAYCA-3’ from Ivanova et al. (2007).
PCR reactions were performed in 20μl reactions using ca. 1 ng of genomic DNA, 0.2μl of VF1d (0.2 mM), 0.2μl of VR1d (0.2 mM), 2μl of total dNTPs (0.2 mM), 0.1μl of (0.5U) of MasterTaq Eppendorf®, 2μl 1x Buffer including MgCl2 at 1.5 mM and 14.5μl of water. PCR conditions were: 1 min at 94°C, followed by 5 cycles of 30 s at 94°C, 40 s at 50 °C, and 1 min at 72°C, followed by 35 cycles of 30 s at 94°C, 40 s at 55°C, and 1 min at 72°C, and ending with 10 min at 72°C The light strands were sequenced using an ABI3730XL by Macrogen Inc. Chromatographs were checked by eye and sequences were edited, when necessary, using the BioEdit sequence alignment editor (version 7.0.5.3; Hall 1999).
To assess species attribution each sequence was compared using the BLAST algorithm in GenBank, where sequences belonging to almost all the Italian echolocating bats species are available thanks to a previous study (Galimberti et al. 2012). All sequences have been deposited in GenBank (HG325822-23-24).
We caught three lactating females (FAL = 33.6, 32.9 and 33.0 mm respectively; weight = 4.7, 4.9 and 4.0) from the “M. mystacinus group”, two in Marsico Nuovo (altitude 1167 m a.s.l) and one in Calvello (altitude 1304 m a.s.l.). All there caught in beech forest sites.
Molecular analysis of these samples allowed the taxonomic identification of the three bats as belonging to the species M. alcathoe. The high similarity matches with reference deposited sequences excluded the misidentification with other congenerics.
In central and eastern Europe, M. alcathoe seems to prefer deciduous forests with old trees and streams (Niermann et al. 2007; Řehák et al. 2008; Lučan et al. 2009; Bashta et al. 2011). Our data adds further records for the presence of M. alcathoe in beech forests of southern Italy (Tereba et al. 2008).
AcknowledgementsFieldwork was conducted under licence from the Italian
Environment Ministry (prot. PNM-2012-0000644). We are indebted with FEM2 Ambiente srl of the Università of Milano-Bicocca for performing the molecular taxonomic identification. The authors are grateful also to Danilo Russo (University of Naples) for his valuable advice and Antonio Conte for logistic support during fieldwork.
This work was funded by the Appennino Lucano National Park Authority.
ReferencesAgnelli, P., Martinoli, A., Patriarca, E., Russo D.,
Scaravelli, D. & Genovesi, P. (Eds.), 2004. Guidelines for bat monitoring: methods for the study and conservation of bats in Italy. Quad. Cons. Natura, 19, MATTM, Italian Wildlife Institute, Italian Chiroptera Research Group.
Anthony, E. L. P. 1988. Age determination in bats. Pp. 47–58 in Ecological and behavioral methods for the study of bats, ed. T.H. Kunz, Smithsonian Institution Press, Washington, DC.
Bastha, A. T., Piskorski, M., Mislajek, R. W., Tereba, A., Korneliusz, K. & Sachanowicz, K. 2011. Myotis alcathoe in Poland and Ucraine: new data on its status and habitat in central Europe. Folia Zool. – 60 (1): 1-4.
Dietz, C. & Von Helversen, O. 2004. Illustrated identification key to the bats of Europe. Electronic publication, version 1.0, Tübingen.
Pier Paolo de Pasquale & Andrea Galimberti
Fig. 1 – Records of Myotis alcathoe.

5Barbastella 7 (1) 2014
Galimberti, A., Spada, M., Russo, D., Mucedda, M., Agnelli, P., Crottini, A., Ferri, E., Martinoli, A. & Casiraghi, M. 2012. Integrated Operationali Taxonomic Units (IOTUs) in Echolocating Bats: A Bridge Between Molecular and Traditional Taxonomy. PLo S ONE 7(6): e401 22. doi:10.1 371/journal.p one.0040122. DOI: http://dx.doi.org/10.1 371/journal.pone.0040122
Hall, T. A., 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium, 41, 95–98.
Ivanova, N. V., Zemlak, T. S., Hanner, R. H. & Hebert, P. D. N., 2007. Universal primer cocktails for fish DNA barcoding. Mol Ecol Notes 7: 544-548. DOI: http://dx.doi.org/10.1111/j.1471-8286.2007.01748.x
Jones, G. & Barlow, K E., 2004. Cryptic Species of Echolocating Bats, Pp. 345-349 in: Echolocation in Bats and Dolphin, ed. THOMAS J.A., MOSS C.F., VATER M., the University of Chicago Press.
Lučan, R. K., Andreas, M., Benda, P., Bartonička, T., Brezinova, T., Hoffmannová, A., Hulová, Š., Hulva, P., Neckářová, J., Reiter, A., Svačina, T., Šálek, M. & Horáček, I. 2009. Alcathoe bat (Myotis alcathoe) in the Czech Republic: distributional status, roosting and feeding ecology. Acta Chiropterol. 11: 61–69. DOI: http://dx.doi.org/10.3161/150811009X465695
Niermann, I., Biedermann, M., Bogdanowicz, W., Brinkmann, R., Le Bris, Y., Ciechanowski, M., Dietz, C., Dietz, I., Estók, P., Von Helversen, O., Le Houédec, A., Paksuz,
S., Petrov, B., Öskan, B., Piksa, K., Rachwald, A., Roué, S.Y., Sachanowicz, K., Schorcht, W., Tereba, A. & Mayer, F. 2007. Biogeography of the recently described Myotis alcathoe von Helversen & Heller, 2001. Acta Chiropterol. 9 (2): 361–378.
Řehák, Z., Bartonička, T., Bryja, J. & Gaisler, J. 2008. New records of the Alcathoe bat, Myotis alcathoe in Moravia (Czech Republic). Folia Zool. 57: 465–469.
Tereba, A., Russo, D., Cistrone, L. & Bogdanowicz, W. 2009. Cryptic diversity: first record of Myotis alcathoe (Vespertilionidae) for Italy. Acts of the 2nd Italian Bat Congress, pp.157: 85-88.
Von Helversen, O., Heller, K.-G., Mayer, F., Nemeth, A., Volleth, M. & Gombkötö, P. 2001. Cryptic mammalian species: a new species of whiskered bat (Myotis alcathoe n.sp.) in Europe. Naturwissenschaften 88: 217–223.
Worthington-Wilmer, J. & Barratt, E. M. 1996. A nonlethal method of tissue sampling for genetic studies of chiropterans. Bat Research News, 37: 1–3.
New records of the Alcathoe bat, Myotis alcathoe (Vespertilionidae) for Italy

6 Barbastella 7(1) 2014
Ramon Jato, Juan Carlos Albero, Luis Lorente
Barbastella 7 (1) Open Access ISSN: 1576-9720 SECEMU www.secemu.org
Confirmada la presencia y reproducción de Myotis bechsteinii en el Pirineo aragonés
Ramon Jato1*, Juan Carlos Albero1, Luis Lorente2
¹ Departamento de Especies y Hábitats. Sociedad Aragonesa de Gestión Agroambiental. ² Avda Puente del Pilar 23, 6ºB 50014 – Zaragoza
*Corresponding author e-mail: [email protected]
DOI: http://dx.doi.org/10.14709/BarbJ.7.1.2014.02
Spanish title: resence and reproduction of Myotis bechsteinii confirmed in the Aragonese Pyrenees
Abstract: In 2012 we began working to determine the distribution and conservation status of Myotis bechsteinii in Aragon. We surveyed 18 sites with potentially suitable habitat in the Aragonese Pyrenees and 231 individuals of 23 bat species were netted. We confirmed the presence of the Myotis bechsteinii in the Aragonese Pyrenees, providing the first data on breeding in Aragón and the northeast Iberian Peninsula. 16 individuals of Myotis bechsteinii were caught in five locations relatively close to each other. Breeding females and juveniles were captured in a single location, which is also a breeding roost located on an old specimen oak Quercus gr cer-rioides with large diameter in a monospecific open woodland (dehesa), with dominance of ancient hollow trees. Counting during emergence showed that the colony was formed by 64 females and juveniles. We only trapped adult males in the other localities sample all of them within or in the vicinity of well-preserved spots Quercus gr cerrioides. The oak forests are highly degraded in Aragonese Pyrenees, with very few and small patches of healthy forest with adult specimens. This results in a very low habitats suitability for Myotis bechsteinii and a high degree of fragmentation.
Key words: Myotis bechsteinii, breeding roost, oak woodland, Pyrenees, Aragón.
Resumen: En 2012 se iniciaron los trabajos de prospección para determinar la distribución y estatus de conser-vación de Myotis bechsteinii en la Comunidad Autónoma de Aragón. Se muestrearon 18 localidades con hábitat potencial en el Pirineo aragonés donde se trampearon 231 ejemplares de 23 especies. Se confirma la presencia de la especie en el Pirineo aragonés y se presentan los primeros datos de reproducción en el noreste de la Península Ibérica. Se capturaron 16 individuos de Myotis bechsteinii en cinco localidades relativamente próximas entre sí. Capturamos hembras reproductoras y juveniles en una sola localidad, donde también se localizó una colonia de cría en un ejemplar de roble de Quercus gr cerrioides de gran diámetro situado en una mancha monoespecí-fica adehesada, con dominancia de ejemplares centenarios que mostraban abundantes oquedades. La colonia se censó en emergencia, contabilizándose 64 ejemplares entre hembras y juveniles. En el resto de localidades únicamente se trampearon machos adultos, las cuales también estuvieron situadas dentro o en las inmediaciones de manchas bien conservadas de Quercus gr cerrioides. Los robledales aragoneses del Pirineo se encuentran muy degradados, las manchas en buen estado de conservación con ejemplares adultos son muy escasas y de pequeño tamaño lo que redunda en una disponibilidad de hábitat para Myotis bechsteinii muy reducida y con un alto grado de fragmentación.
Palabras clave: Myotis bechsteinii, colonia reproductora, robledal, Pirineos, Aragón.
accepted: January 5 , 2014

7Barbastella 7 (1) 2014
IntroducciónEn Aragón se han citado 28 especies de murciélagos,
según se recoge de la información obtenida de diversos trabajos realizados entre 2001 y 2012 (Woutersen 2001 y 2004; García-González et al. 2003; Alcalde et al. 2003 y 2004; Serra-Cobo 2004; Albero et al, 2010 y 2011). La presión de muestreo en esta última década, ha permitido citar por primera vez nuevas especies y actualmente Aragón es una de las pocas comunidades autónomas con presencia de especies como Plecotus macrobullaris, Myotis cf. nattereri, y Vespertilio murinus (García-Gonzalez et al. 2003; Alcalde et al. 2008; Alberdi et al. 2009). Sin embargo, y pese a ser una comunidad con abundancia y diversidad forestal, únicamente existen tres citas históricas de presencia de Myotis bechsteinii en Aragón, dos en la provincia de Huesca (Woutersen y Bafaluy 2001) consideradas erróneas por otros autores (Alcalde et al. 2008) y otra en los Montes Universales en Teruel (Alcocer & Monsalve, com. pers. en Alcalde et al. 2008).
En Europa M. bechsteinii aparece restringido a bosques maduros de haya y roble con una alta proporción de árboles viejos (Hutson et al. 2012; Dietz y Pir 2009). En la Península ibérica se ha comprobado que la especie se encuentra muy ligada los robledales de Quercus pyrenaica y Q. robur (Napal et al. 2009 y 2010, Goiti et al 2007, López y Hermida 2012), incluso en Cazorla se han encontrado ejemplares en extensos pinares (Arribazalaga et al. 2013 a y b) en los que seleccionaban los rodales de frondosas o árboles caducifolios sueltos dentro de pinares, a pesar de su baja disponibilidad mostrando una importante respuesta a variaciones a nivel de microhábitat dentro de macrohábitats aparentemente homogéneos.
La distancia de más de 200 km. a otras poblaciones conocidas, la escasez de datos en el cuadrante nororiental de
la península con unas pocas citas en Cataluña (Flaquer et al 2010), y el estado de conservación desfavorable de las masas de frondosas, especialmente de las quercíneas, hacían dudar de su presencia en el Pirineo aragonés.
M. bechsteinii es una especie prioritaria de interés europeo (Directiva 92/43/CEE) para la que las comunidades autónomas, en caso de estar presente, deben designar espacios destinados a su conservación. En base a datos erróneos y sin la constancia segura de su presencia en el Pirineo aragonés, la especie se incluyó en los inventarios de varios lugares pirenaicos de la Red Natura 2000 de Aragón. Al tratarse de una especie muy escasa, de requerimientos forestales y detección dificultosa, no suele detectarse en trabajos de inventarios generales, por lo que se ha tendido a dar por presente allí donde el tipo de hábitat así lo hacía sospechar.
En consecuencia, el Gobierno de Aragón decidió iniciar trabajos de prospección para determinar si la especie se encontraba efectivamente presente. En el presente artículo se exponen las observaciones realizadas de esta especie durante los años 2012 y 2013 en el Pirineo oscense, dentro de los trabajos de inventario de quirópteros de interés comunitario (Directiva 92/43/CEE) tanto dentro como fuera de la Red Natura 2000.
Material y métodosSe han prospectado 18 zonas potenciales del Pirineo
de la provincia de Huesca entre el 30 de mayo y el 27 de agosto de 2012, y otra zona en 2013. La ubicación de las zonas de muestreo se realizó seleccionando masas forestales de quercíneas con presencia de pies maduros de gran tamaño (Fig. 1).
Confirmada la presencia y reproducción de Myotis bechsteinii en el Pirineo aragonés
Fig. 1 – Localización de las estaciones de muestreo de Myotis bechsteinii y resultados en el Pirineo aragonés.

8 Barbastella 7(1) 2014
Para la detección de la especie se usaron tres métodos. Por un lado, se colocaron redes de niebla en puntos de agua (remansos de ríos, balsas ganaderas y depósitos). Por otro, también se instalaron redes en el interior del bosque en combinación con dos reproductores de reclamos acústicos (modelo Sussex Autobat, Universidad de Sussex). Dado el pequeño tamaño de las parcelas de bosque a muestrear optamos por un sistema intermedio entre las redes móviles usadas por Goiti et al. (2007) y las estaciones de reclamo con redes fijas, que consistió en la colocación de un número variable de redes fijas dentro del bosque moviendo los reclamos entre ellas. Por último, se realizaron grabaciones de ultrasonidos de manera irregular en bebederos con un detector D240x (Pettersson Elektronik AB, Uppsala, Suecia) y de manera regular con un equipo de detección y grabación EM3+ (WildLife Acoustic, Concord, EEUU) en modo automático tanto en bebederos como en el interior del bosque. Se eligieron los días de trampeo con las condiciones meteorológicas adecuadas, ausencia de viento y precipitaciones, y temperaturas por encima de 10ºC. El muestreo comenzó una hora antes del ocaso y persistió hasta la captura de ejemplares de M. bechsteinii, o bien hasta que la temperatura descendía por debajo de 10ºC.
A los ejemplares capturados (Fig. 2) se les tomaron datos biométricos y se identificaron apoyados por bibliografía especializada (Dietz y Von Helversen 2004). También se tomaron muestras de tejido (biopunch en el plagiopatagio) que fueron depositadas en el Departamento de Zoología de la Universidad del País Vasco/Euskal Herriko Unibertsitatea (UPV/EHU) y en la Estación Biológica de Doñana (CSIC).
El análisis de las grabaciones de ultrasonidos se realizó mediante el software de análisis automático Sonochiro de la empresa Biotope y el programa informático BatSound 3.31 (Pettersson Electronik AB) apoyados en los criterios de identificación publicados por Barataud y Tupinier (2012), teniendo en cuenta las dificultades de discriminar las ecolocalizaciones de algunas especies.
Así mismo, a una hembra reproductora de M. bechsteinii se le adherió un radio emisor (Fig. 3) modelo Picopip de 0,35 g (Biotrack, Dorset, RU), para localizar la colonia. Se tuvo en cuenta que el peso del emisor fuera inferior al 5% del peso del ejemplar (10,6 gramos). El conteo del número de individuos en la colonia se realizó durante la emergencia mediante una cámara de video DCR-TRV14b (Sony, Japón) con foco de luz infrarroja IRLamp6 (Wildlife Engineering, EEUU).
ResultadosEn total, se capturaron 231 ejemplares de 23 especies de
quirópteros en 18 localidades. Respecto a M. bechsteinii se capturaron 16 individuos en cinco localidades, todas relativamente próximas entre sí, situadas en la comarca de la Ribagorza, en el Pirineo Oriental aragonés (Fig. 1). En un enclave (localidad 1), se capturaron hembras reproductoras y juveniles de ambos sexos, en otras tres localidades (2, 3, 4 y 7) fueron trampeados únicamente machos adultos, y en dos localidades (5 y 6) la posible presencia de la especie se basa únicamente en grabaciones (Tabla 1).
En la localidad 1 se capturaron dos hembras lactantes el 11 de julio de 2012. Posteriormente, el 30 de julio de 2012 se capturaron nueve ejemplares más, una hembra adulta y ocho jóvenes. En las otras tres localidades se capturaron machos adultos el 17, el 20 y el 23 de julio de 2012 y el 24 de agosto de 2013. Mediante el análisis de las grabaciones de ultrasonidos se identificaron llamadas de ecolocación compatibles con Myotis bechsteinii en las localidades 1, 5 y 6.
Las capturas se realizaron dentro de manchas de Quercus gr. cerrioides o en bebederos en las inmediaciones de los mismos. Este roble es un híbrido entre Quercus humilis y Quercus faginea y presenta caracteres intermedios entre esas dos especies. Ambas especies y su híbrido son marcescentes.
Ramon Jato, Juan Carlos Albero, Luis Lorente
Fig. 2 – Hembra adulta de Myotis bechsteinii en un tronco de quejigo.
Tabla 1. Resultados por localidades y métodos de muestreo de Myotis bechsteinii en el Pirineo aragonés.Detector Reclamo Bebedero TOTAL
Loc. 1 (1ª visita) Positivo 1 ♀ ad 1 ♀ ad 2Loc. 1 (2ª visita) Positivo 6 ♂ juv, 2 ♀ juv 1 ♀ ad 9Loc. 2 Negativo 1 ♂ ad Negativo 1Loc. 3 1 ♂ ad 1Loc. 4 1 ♂ ad 1Loc. 5 Positivo Negativo -Loc. 6 Positivo -Loc. 7 2 ♂ ad 2

9Barbastella 7 (1) 2014
El marcaje radiotelemétrico de la hembra capturada el 30 de julio de 2012, permitió localizar la colonia de cría en un ejemplar vivo en buen estado de Quercus gr cerrioides. El árbol tenía 100 cm. de diámetro y estaba situado en el medio de una mancha monoespecífica adehesada de 5,8 Ha, con presencia únicamente de ejemplares centenarios (Fig 2). El refugio se encontró en una cavidad realizada por pícidos situada en la cara inferior de una rama horizontal ubicada a 7,25 m. de altura y orientada al sur. Se encuentra a 800 m de altitud y a 275 m. de distancia de un curso permanente de agua. La mancha presenta una densidad de 8,8 árboles/Ha. con una cobertura de copas del 20,2 %, el sotobosque está únicamente compuesto por vegetación herbácea como consecuencia del intenso pastoreo. Alrededor de esta mancha se encuentra un pequeño cauce, vegetación arbustiva, campos de cultivo, desiertos de erosión sobre margas y manchas de roble mucho más densas con un escaso grado de desarrollo. Se contabilizaron 62 y 64 ejemplares en la colonia, los días 31 de julio y 2 de agosto de 2012 respectivamente, comprendiendo adultos y crías que ya eran capaces de volar.
La localidad 2 es una mancha de 3,18 Ha. de Quercus gr cerrioides con un escaso desarrollo del sotobosque que se encuentra representado casi exclusivamente por Buxus sempervirens. La mancha presenta un alto grado de madurez con una buena representación de todas las clases diamétricas con alta presencia de árboles de más de 40 cm de diámetro y algunos ejemplares de más de 80 y 100 cm. de diámetro. La cobertura de copas es alta con presencia de pequeños claros diseminados por la mancha. El ejemplar macho adulto se capturó a 900 m. de altitud y a 150 m. de un curso permanente de agua.
Las localidades 3, 4 y 7 comprenden puntos de agua a una altitud de entre 800 m. y 1.000 m. dentro o muy cerca de manchas de Q. gr. cerrioides de diferentes grados de madurez, que se alternan con prados y bosquetes de coníferas. En estas localidades sólo se capturaron machos adultos de M. bechsteinii.
La localidad 5 son prados de siega en diferente grado de abandono situadas en fajas estrechas. En las márgenes de los prados hay una gran abundancia de robles de gran diámetro que confieren a la localidad un aspecto de bosque adehesado. La altitud es de 950 m. y el curso de agua permanente más cercano se encuentra a 522 m.
La localidad 6 se encuentra en la orilla de un embalse a 1.077 m. de altitud situado junto a una gran mancha forestal mixta con quejigo, avellano y fresno como especies principales y algunas manchas de pino silvestre. Los quejigos en ocasiones forman pequeñas manchas monoespecíficas.
Discusión y ConclusionesSe ha podido confirmar la presencia y reproducción de
M. bechsteinii en el Pirineo aragonés. En la única colonia localizada, se contabilizaron en emergencia nocturna 64 ejemplares entre hembras y crías por lo que la colonia estudiada estaría en concordancia con el tamaño descrito en la literatura (entre 6 y 50 hembras adultas; Napal et al. 2009; Dietz et al. 2009). Podría considerarse como una colonia
mediana-grande, de un mínimo de 32 hembras reproductoras estimado considerando una proporción 1:1 entre hembras y crías (Gessner y Weishaar 2004).
Los tres métodos usados en el presente estudio han arrojado resultados positivos a la hora de detectar M. bechsteinii, aunque con diferente éxito. De los puntos con resultados positivos, la especie se capturó con redes en bebedero en tres de las seis localidades en las que se colocaron; con redes y reclamo en el bosque en tres de las cuatro en las que se usó; por último, se registraron grabaciones compatibles con M. bechsteinii en 3 de las 4 localidades en las que se utilizó este método (Tabla 1). La dificultad para discriminar las ecolocaciones de las especies del género Myotis mediante el análisis de ultrasonidos, hace que las localidades 5 y 6 se deban considerar únicamente como posibles. Además, el uso de este método sólo es recomendable como complementario a las capturas con red, aunque puede ser útil para realizar prospecciones previas y centrar los esfuerzos de captura en las localidades en las que se realicen grabaciones compatibles con los parámetros de los ultrasonidos emitidos por la especie.
El uso de reclamos parece favorecer la captura de M. bechsteinii con redes de niebla dentro del bosque (hembras lactantes y posteriormente crías dependientes en localidad 1, un macho en localidad 2, y dos machos en la localidad 7), aunque es evidente que el uso de redes colocadas en bebederos situados cerca de hábitats favorables también ha resultado efectivo con capturas de hembras reproductoras en la localidad 1, y de machos en las localidades 3 y 4.
La colonia de cría se encuentra en una mancha forestal adehesada rodeada de desierto de erosión, campos de cultivo y manchas de bosque muy joven (Fig. 3). Tanto la colonia de
Confirmada la presencia y reproducción de Myotis bechsteinii en el Pirineo aragonés
Fig. 3 – Hembra adulta de M. bechsteinii con emisor modelo Picopip de 0,35 g (Biotrack, Dorset, RU).

10 Barbastella 7(1) 2014
cría como su entorno más inmediato no se corresponden con los óptimos descritos en otras poblaciones (Dietz y Pir 2009; Napal et al. 2009 y 2010), aunque sí coincide con lo descrito en Kaňuch y Ceľuch (2007) donde encuentran en un bosque adehesado poblaciones de M. bechsteinii asociadas con otras especies forestales.
Las masas de quercíneas del Pirineo aragonés, compuestas por las especies Quercus ilex y Q. gr cerrioides, fueron intensamente aprovechadas para la obtención de leñas y producción de carbón por lo que hoy en día solo se encuentran masas degradadas en proceso de regeneración, con gran abundancia de pies formados a partir de rebrotes vegetativos. Los bosques maduros son casi inexistentes y sólo perviven pequeñas manchas con presencia de ejemplares con un mínimo de madurez. Las zonas con presencia de ejemplares añosos se encuentran normalmente ligadas a pequeñas manchas adehesadas en las que se ha realizado un aprovechamiento tradicional de los pastos, también a agrupaciones de ejemplares trasmochados de gran tamaño, así como a alineaciones de ejemplares que sobrevivieron gracias a su uso como puntos de referencia para delimitar propiedades. Algunas de estas manchas permanecen aisladas o bien diferenciadas de otros hábitats forestales, mientras que en otras ocasiones han quedado inmersas dentro de extensas masas forestales con un escaso grado de madurez general.
Todo parece indicar que solamente en alguna de estas pequeñas zonas es donde pueden encontrarse las últimas colonias del Pirineo aragonés. En concordancia con los resultados de Arribazalaga et al. (2012 y 2013) y Napal et al (2013) parece que Myotis bechsteinii podría estar utilizando en Aragón pequeños parches de hábitat favorable dentro de grandes extensiones con hábitat poco adecuado; incluso que a pesar de ser una especie más abundante en el contexto centroeuropeo que en el área mediterránea podría tener su óptimo ecológico en los bosques templados mediterráneos y que su escasez en la biorregión estaría reflejando el alto grado de fraccionamiento y degradación de los bosques caducifolios mediterráneos (Napal et al, 2013).
De las cinco localidades con resultados positivos, dos de ellas (entre las que se encuentra la única colonia de cría conocida de Aragón) se encuentran fuera de la Red Natura 2000, de las otras 3 dos se encuentran en LIC y la última en una
ZEPA. El alto grado de fragmentación y el pequeño tamaño de las manchas de hábitat potencial hace que la idoneidad del hábitat deba de ser detectada a nivel de microhábitat lo que ha propiciado que las parcelas con hábitat potencial para la especie no hayan sido incluidas en la Red Natura 2000 más que por mero azar o debido a encontrarse dentro de grandes masas forestales que por su extensión han sido expresamente seleccionadas. Para garantizar la protección y gestión adecuada de la especie y sus hábitats es importante la localización de las colonias en futuras prospecciones, la identificación de las manchas de quercíneas con abundancia de pies maduros y su inclusión en la Red Natura 2000.
Ramon Jato, Juan Carlos Albero, Luis Lorente
Fig. 5 – Vista aérea de la mancha en la que se encuentra la colonia de Myotis bechteinii con buffer de 500 m.
Fig. 4 – Vistas general y de detalle de la zona en la que se encuentra la primera colonia hallada de Myotis bechsteinii en el Pirineo aragonés.

11Barbastella 7 (1) 2014
AgradecimientosEste proyecto ha sido financiado por el Servicio de
Biodiversidad de la Dirección General de Conservación del Medio Natural del Gobierno de Aragón, bajo la dirección de Manuel Alcántara.
Inazio Garin y Jose Ramón Aihartza (UPV/EHU), Javier Juste (EBD-CSIC) y Xavier Puig (Associació Galanthus) nos prestaron los equipos de reclamo Autobat, nos asesoraron sobre su uso y compartieron su valiosa experiencia sobre la especie. Los Agentes de Protección de la Naturaleza del Gobierno de Aragón (APN) Mariano Muñoz y Félix Calvo nos ayudaron a localizar manchas forestales bien conservadas. Nos han acompañado en el trabajo de campo Merichel Broto, Fernando Carmena, Ignacio Gómez, Jesús Laín, Quintina Ortiz, Javier Miranda y Juan Ramón Donés de la empresa SARGA, y Félix Calvo APN del Gobierno de Aragón. A Begoña Marquina por la revisión del resumen en inglés. A Inazio Garin por la revisión del manuscrito y sus valiosas aportaciones.
Referencias Alberdi, A., Aihartza, J., Albero, J. C., Aizpurua, O.,
López-Baucells, A., Freixas, L. & Garin, I. (2012). First records of the parti-coloured bat Vespertilio murinus (Chiroptera: Vespertilionidae) in the Pyrenees. Mammalia, 76(1), 109-111. DOI: http://dx.doi.org/10.1515/MAMM.2011.106
Albero J.C., Ballarín I., Carmena F., Rivas J.L., Sanz J. & Hernández D. 2010. Inventario de refugios y especies de quirópteros en el Parque Natural de la Sierra y Cañones de Guara. SODEMASA. Gobierno de Aragón. Informe inédito.
Albero J.C., Broto M., Carmena F., Gómez I., Jato R., Rivas J.L., & Sanz J. 2011. Inventario de refugios y especies de quirópteros en el Parque Natural de Posets-Maladeta. SODEMASA. Gobierno de Aragón. Informe inédito.
Alcalde, J.T., Trujillo, D., Artárcoz, A. & Agirre-Mendi, P.T., 2008. Distribución y estado de conservación de los quirópteros en Aragón. Graellsia, 64(1), 3-16.
Amengual B., Aymerich P., Fontal J., Gosalbez J., Götzens G., López M. & Serra – Cobo J. 2004. Estado de conservación de las poblaciones de quirópteros, mamíferos acuáticos y Rana pyrenaica del Parque Nacional de Ordesa y Monte Perdido. Propuestas para su eficaz gestión. Areambiental 2004. Ministerio de Medio Ambiente. PNOMP. Informe Técnico. DOI: http://dx.doi.org/10.3989/graellsia.2008.v64.i1.51
Arribazalaga, A., Saldamendi, E., Napal, M., Garín, I. Aihartza, J. 2013. Selección del hábitat por Myotis bechsteinii en un pinar al sureste de la Península Ibérica: cuando el macrohábitat enmascara al microhábitat. Barbastella, 6 (1), 95.
Barataud, M. & Tupinier, Y. 2012. Écologie acoustique des chiroptères d’Europe: identification des espèces, étude de leurs habitats et comportements de chasse. Ed Biotope.
Carro, F. 2007. Myotis bechsteinii (Kuhl, 1817). [Ficha de Atlas]. Pp. 171 -173 en L. J. Palomo, J. Gisbert & J. C. Blanco (eds.): Atlas y Libro Rojo de los mamíferos terrestres de España. MMA/DGCN-TRAGSA-SECEM-SECEMU, Madrid.
Dietz, C. & Von Helversen, O. 2004. Claves de identificación de los murciélagos de Europa. Publicación electrónica.
Dietz, M. & Pir, J. B. 2009. Distribution and habitat selection of Myotis bechsteinii in Luxembourg: implications for forest management and conservation. Folia Zoologica, 58(3), 327-340.
Diet, C. Helversen, O. & Nill, D. 2009. Bats of Britain, Europe and Northwestern Africa. A &C Black Publisher Ltd. London.
Fitzsimons, P., Hill, D. & Greenaway, F. 2002. Patterns of habitat use by female Bechstein’s bats (Myotis bechsteinii) from a maternity colony in a British woodland. Report. School of Biological Sciences, University of Sussex.
Flaquer, C., Torre, I. & Arrizabalaga, A. 2007. Selección de refugios, gestión forestal y conservación de quirópteros forestales. Conservación de la biodiversidad, fauna vertebrada y gestión forestal. J.Camprodon y E. Plana (eds). Publicacions i Edicions de la Universitat de Barcelona, 2007.
Flaquer C., Puig X., Fábregas E., Guixé D., Torre I., Ràfols R.G., Páramo F., Camprodon J., Cumplido J.M., Ruíz-Jarillo R., Baucells A.L., Freixas L. & Arrizabalaga, A. 2010. Revisión y aportación de datos sobre quirópteros de Catalunya: propuesta de lista roja. Galemys, 22 (1): 29-61.
García–González, R., Aihartza, J.A., Garín, I., Goiti, U., Trujillo, D., Zabala, J., Gosalbez, J., Luque, J.J., Monreal, E., Aymerich, P., Götzens, G., Couto, S., Herrero, J., Hernández, Y. & Prada, C. 2003. Inventario y criterios de gestión de los mamíferos del PNOMP, OAPN – CSIC, Huesca, Jaca.
Gessner, B. & Weishaar, M. 2004. Fort setzung der Beringung der Bechsteinfledermaus (Myotis bechsteinii) in der Region Trier = Continuing the ringing of Bechstein’s bats (Myotis bechsteinii) in the region of Trier. Dendrocopos, 31, 21-26.
Goiti, U., Aihartza, J., Garin, I. & Salsamendi, E. 2007. Surveying for the rare Bechstein’s bat (Myotis bechsteinii) in northern Iberian peninsula by means of an acoustic lure. Hystrix, the Italian Journal of Mammalogy, 18(2), 215-223. DOI: http://dx.doi.org/10.4404/hystrix-18.2-440
Confirmada la presencia y reproducción de Myotis bechsteinii en el Pirineo aragonés

12 Barbastella 7(1) 2014
Hohti, P., Ceľuch, M., Danko, Š., & Kaňuch, P. 2010. Constraints in the roost-site selection of the tree-dwelling Bechstein’s bat (Myotis bechsteinii). Hystrix, the Italian Journal of Mammalogy, 22(1).
Hutson, A.M., Spitzenberger, F., Tsytsulina, K., Aulagnier, S., Juste, J., Karataş, A., Palmeirim, J. & Paunović, M. 2008. Myotis bechsteinii. In: IUCN 2012. IUCN Red List of Threatened Species. Version 2012.2. <www.iucnredlist.org>. Downloaded on 16 April 2013
Kaňuch, P., & Cel’uch, M. 2007. Bat assemblage of an old pastured oak woodland (Gavurky Protected Area, central Slovakia). Vespertilio, 11: 57-64.
Kerth, G., Wagner, M. & König, B. 2001. Roosting together, foraging apart: information transfer about food is unlikely to explain sociality in female Bechstein’s bats (Myotis bechsteinii). Behavioral Ecology and Sociobiology, 50(3), 283-291. DOI: http://dx.doi.org/10.1007/s002650100352
Kerth, G. & Morf, L. 2004. Behavioural and genetic data suggest that Bechstein’s bats predominantly mate outside the breeding habitat. Ethology, 110(12), 987-999. DOI: http://dx.doi.org/10.1111/j.1439-0310.2004.01040.x
Kerth, G. & Petit, E. 2005. Colonization and dispersal in a social species, the Bechstein’s bat (Myotis bechsteinii). Molecular Ecology, 14(13), 3943-3950.
López, Z. y Hermida, R.X. 2013. Estudio del uso del hábitat por Myotis bechsteinii en los montes de O Morrazo (Pontevedra). Barbastella, 6 (1) 95-96.
Napal, M., Garin, I. , Goiti, U., Salsamendi, E. y Aihartza, J., 2009. Selection of maternity roosts by Myotis bechsteinii (Kuhl, 1817) in the Southwestern Iberian Peninsula. Acta Chiropterologica, 11(2), 425-433.
Napal, M., Garin, I., Goiti, U., Salsamendi, E. & Aihartza, J. 2010. Habitat selection by Myotis bechsteinii in the southwestern Iberian Peninsula. Annales Zoologici Fennici, 47(4), 239-250. DOI: http://dx.doi.org/10.5735/086.047.0402
Napal, M., Garin, I., Goiti, U., Salsamendi, E., & Aihartza, J. (2013). Past deforestation of Mediterranean Europe explains the present distribution of the strict forest dweller Myotis bechsteinii. Forest Ecology and Management, 293, 161-170. DOI: http://dx.doi.org/10.1016/j.foreco.2012.12.038
Reckardt, K. & Kerth, G. 2007. Roost selection and roost switching of female Bechstein’s bats (Myotis bechsteinii) as a strategy of parasite avoidance. Oecologia, 154(3), 581-588. DOI: http://dx.doi.org/10.1007/s00442-007-0843-7
Salsamendi, E., Napal, M., Aihartza, J., Goiti, U., Almenar, D. & Garin, I. 2007. Estudios de seleccion de hábitat en Myotis bechsteinii, Myotis emarginatus y Rhinolophus euryale. Informe final. SECEMU, Junta de Extremadura.
Woutersen, K. & Bafaluy J.J. 2001. Murciélagos del Alto Aragón. Kees Woutersen Publicaciones, Huesca.
Ramon Jato, Juan Carlos Albero, Luis Lorente

13Barbastella 7 (1) 2014
Barbastella 7 (1) Open Access ISSN: 1576-9720 SECEMU www.secemu.org
Murciélagos (Mammalia: Chiroptera) del área urbana del municipio de Boa Vista, Roraima, Brasil
Ubirajara Dutra Capaverde Junior1*, Susi Missel Pacheco2 & Marcos Eugenio Duarte3
1 Instituto de Nacional de Pesquisas Amazônicas, Av. André Araújo, 2936 Aleixo, Caixa Postal: 2223 CEP: 69060-001 Manaus, Amazonas, Brasil.2 Instituto SAUVER, Av. Pernambuco, 2623, sala 404, Bairro Floresta, CEP 90240-005, Porto Alegre, RS, Brasil.3 Agencia de Defesa Agropecuária de Roraima, Rua Coronel Mota, nº 1142, Centro CEP: 69.305-120, Boa Vista , Roraima, Brasil.
*Corresponding author e-mail: [email protected]
DOI: http://dx.doi.org/10.14709/BarbJ.7.1.2014.03
English title: Species of bats (Chiroptera) in urban area of the Boa Vista, Roraima
ResumenLa mayoría de los trabajos con murciélagos en la Amazonía brasileña ha sido realizada en ambientes forestales. Pocos estudios se centran en áreas de sabana amazónica y son rarísimos en las áreas urbanas, lo que resulta en una submuestra de incidencia de esos mamíferos. Esta investigación ha proporcionado la lista de quirópteros que viven en el área urbana de Boa Vista-Roraima y ha contribucido al aumento de datos de distribución y ocur-rencia en Brasil. Se trata de una investigación no experimental del tipo exploratorio y descriptivo, con enfoque cuantitativo. Como resultados se han identificado 23 especies en el área urbana, distribuidas en cinco familias y con dieta predominantemente frugívora. El número de especies es similar al que viene registrándose en los inventarios de otras capitales en Brasil.
Palabras clave: Inventario; Quirópteros; Región Amazónica
Abstract While most work on bat research in the Brazilian Amazon has been held in forests, few studies were placed in Amazonian savannas and even less in urban areas, resulting in a subsampling of these mammals distribution. The study surveyed the bats that inhabit the urban area of Boa Vista, Roraima and contributed to the data growth occurring in Brazil. It is a non-experimental research but exploratory and descriptive, with quantitative focus. We identified 23 species in the urban area, distributed in five families and showing predominantly frugivorous diet. This figure is similar to the number of species that have been recorded in inventories from other Brazilian capitals.
Key words: Amazon Region; Chiroptera; Inventory, Check-list
accepted: Decenmber 21, 2013
Murciélagos (Mammalia: Chiroptera) del área urbana del municipio de Boa Vista, Roraima, Brasil

14 Barbastella 7(1) 2014
Ubirajara Dutra Capaverde junior, Susi Missel Pacheco, Marcos Eugenio Duarte
IntroducciónActualmente el proceso de transformación del paisaje
natural en grandes centros urbanos trae como consecuencia la disminución de la biodiversidad local y, a veces, regional, aunquesegún Yalden (1980), los pequeños mamíferos, entre ellos los quirópteros, podrían sobrevivir en ambientes antropizados. Sin embargo, Pedro (1998) afirmó que solamente el 20% de las especies murciélagos en Brasil serían capaces de mantener poblaciones estables en este ambiente.
Los murciélagos que ocurren en áreas urbanas a menudo presentan mayor habilidad en la búsqueda y en el uso del alimento, agua y de refugios disponibles que les permite mantenerse en este ambiente antropizado y teóricamente hostil (Pacheco et al., 2008).
La mayoría de los murciélagos presentes en ambientes urbanos son insectívoros, y en ambientes urbanos la iluminación pública y de las casas suele atraer insectos. Además hay cierta similitud entre los abrigos naturales de estas especies disponibles en ambiente urbano (Aguita, 2003; Pacheco et al., 2008). Con relación a las especies nectarívoras y frugívoras usualmente detectadas en este ambiente, buscan abrigo y alimentos junto a zonas verdes, en vías públicas, plazas, parques y áreas particulares (Uieda et al., 2008).
En Brasil hay 174 especies de quirópteros registradas pertenecientes a nueve familias (Paglia et al., 2012), de las cuales 63 se ha provado que utilizan el ambiente urbano en un mínimo de 18 ciudades brasileñas (Lima, 2008), siendo Manaus la única ciudad citada para la región Norte con 24 especies. Pacheco et al. (2010) registraron 47 especies para áreas urbanas en seis estados brasileños São Paulo (SP), Santa Catarina (SC), Rio Grande do Sul (RS), Rio de Janeiro (RJ), Paraná (PR), Mato Grosso (MT), además del Distrito Federal (DF). En estas regiones predominaron las especies de la familia Phyllostomidae (17), seguidas de las familias Molossidae (16), Vespertilionidae (11), Emballonuridae (2) y Noctilionidae (1), de las cuales 30 especies son insectívoras, 13 nectarívoras/frugívoras, dos omnívoras, una hematófaga y una piscívora.
Tanto en Brasil como en el resto del mundo, todavía existe poca información respecto a la conservación y ecología de estos mamíferos en áreas urbanas y naturales. El objetivo de ese estudio fue básicamente identificar las especies de quirópteros que ocurren en el ambiente urbano de Boa Vista, Roraima y proporcionar una lista de especies actualizada y completa.
Área de estudioDe acuerdo con SEBRAE (2006), el municipio de Boa
Vista comprende un área de 5.711,9 km², abrigando la capital del Estado de Roraima, y ubicada en la región central del Estado, entre las coordinadas 02°49’11”N y 60°40’24”W (Figura 1), a una altitud de 90 m sobre el nivel del mar. Presenta clima, según Köppen (1948), tropical húmedo del tipo “A”, del subtipo AW: clima tropical lluvioso, , es caluroso y húmedo, con estación lluviosa en el verano con predominio de sabanas.
El estudio se ciñó al área urbana del referido municipio delimitada por las coordinadas geográficas: 2°53’54.53”, 2°44’18.95”, 2°51’23.28”, 2°48’42.31”N; y 60°41’7.64”, 60°43’19.15”, 60°37’36.57”, 60°47’31.85”W. A partir de esos puntos se delineó un polígono imaginario que restringió el área estudiada a 154 km² de área efectivamente urbana, y configurada en zonas: leste, oeste, norte y sur (Figura 2).
Material y Métodos Se realizaron veintitrés noches de muestreo con dos redes
de niebla, ejecutando un esfuerzo de muestra de 4997.7 h/m² según el método de Straube y Bianconi (2002), y siete búsquedas activas de tres horas cada una en construcciones (tejados, paredes de ladrillos, huecos entre edificios), dando un total de 21 h (Tabla 1). Las colectas se realizaron teniendo en cuenta las fases de la luna, intentando realizar el máximo número de noches durante la fase menguante o nueva.
Las redes de niebla fueron dispuestas al nivel del suelo hasta 3 m de altura, próximas a plantas en fructificación, floración, refugios y/o en posibles rutas de vuelo. Los murciélagos se extrayeron de la red inmediatamente después de la captura y fueron mantenidos en sacos de captura. Este método basadoen la baja altura de las redes y al ambiente rico en plantas en fructificación probablemente haya propiciado una mayor captura de especies de la familia Phyllostomidae, en relación a otras especies insectívoras.
Las construcciones humanas se evaluaron esporádicamente en los sitios de muestras descriptos en la Tabla 1, con el objetivo de identificar las especies que usualmente no son capturadas en redes de niebla. Para colectar los murciélagos en estos locales se utilizaron pinzas de 30 cm o redes de mano unidas aun cable extensible de hasta tres metros de largo.
Fig. 1 – Mapa físico del Estado de Roraima, localización en Brasil (recuadro pequeño) y localización del municipio de
Boa Vista (en rojo). Fuente: SEBRAE (2006).

15Barbastella 7 (1) 2014
Murciélagos (Mammalia: Chiroptera) del área urbana del municipio de Boa Vista, Roraima, Brasil
Las identificaciones taxonómicas se basaron en caracteres mofológicos siguiendo a Vizotto y Taddei (1973), Barquez et al. (1993), Gregorin y Taddei (2002) y Aguirre (2009).
Un ejemplar de cada especie fue colectado como material testigo para una identificación específica. Los murciélagos colectados fueroneutanizados y depositados en el Instituto Sauver como material de referencia, en conformidad con la licencia 30045-1/SISBIO. El procedimiento de eutanasia y anestesia fue mediante Isoflurano 1ml/ml (Nombre Comercial: “FORANE”) y posterior acetato de etilo, conforme prescrito por el veterinario Dr. Marcos Eugenio Duarte, responsable técnico de esta investigación. No se cita referencia bibliográfica, debido a la falta de descripción específica para pequeños mamíferos. Se realizaron medidas corporales y cranianas, y fueron verificadas las masas corporales e identificadas las fórmulas dentarias. Tras eso, los especímenes fueron etiquetados y mantenidos congelados en el laboratorio de Zoología de la Facultad Cathedral hasta su envío al Instituto Sauver, donde han sido taxidermizados o conservados en alcohol (70%) e inventariados en la Colección de Referencia de la institución (IS de 0100-00123). Los
especímenes no inventariados fueron liberados, después de su evaluación morfométrica y dentaria, en los mismos puntos de captura durante la misma noche de capturas.
Resultados y DiscusiónSe ha capturado un total de 602 murciélagos (23
colectados), pertenecientes a cinco familias y 23 especies para el área urbana de Boa Vista (Tabla 1), representando el 13% de la riqueza de murciélagos sugerida para Brasil (n= 174) y el 16% de la riqueza de murciélagos sugerida para la Amazonía (n= 146), según Paglia et al. (2012).
Comparando las 23 especies identificadas en Boa Vista con el estudio de Lima (2008), se verifica que las especies Rhynchonycteris naso, Saccopteryx bilineata, Micronycteris minuta, Lophostoma silvicolum, Trinycteris nicefori y Ametrida centurio no estaban registradas para el área urbana de otras ciudades brasileñas. La familia mejor representada fue Phyllostomidae con 353 individuos capturados y 12 especies, seguida de Molossidae 176 individuos y 4 especies,
Fig. 2 – Ciudad de Boa Vista, los puntos negros representan las colectas con auxilio de redes, los puntos blancos, las colectas en refugios y el polígono negro, el límite del área muestreada. Fuente: Google Earth (2013).

16 Barbastella 7(1) 2014
Ubirajara Dutra Capaverde junior, Susi Missel Pacheco, Marcos Eugenio Duarte
Vespertilionidae 5 individuos y 4 especies, Emballonuridae 18 individuos y 2 especies y Noctilionidae 50 individuos y 1 especie (Tabla 1). De acuerdo con Kunz y Kurta (1988), eso puede ser debido a la metodología, concretamente por el uso de redes de niebla que favorece la captura de filostómidos mientras que la búsqueda de refugios en construcciones favorece la captura de molósidos y vespertiliónidos. En el caso de N. albiventris el número de individuos elevado se debe a la captura realizada en la proximidad de la salida del abrigo.
El predominio de la familia Phyllostomidae en el municipio de Boa Vista, confirma los resultados obtenidos en ambientes urbanos por Pacheco et al. (2010) para seis estados de Brasil, que revelan el predominio de la familia Phyllostomidae en las capturas con redes. Sin embargo, el número de especies estrictamente frugívoras (8) es menor que el de las especies insectívoras (12). Se verifica en la
Tabla 1 donde solo la especie A. planirostris, es responsable para la abundancia de la familia Phyllostomidae en relación a las otras, ocurriendo lo mismo para M. molossus y la familia Molossidae.
Las familias Vespertilionidae, Emballonuridae y Molossidae presentaron tasas de captura más bajas que la familia Phyllostomidae. Este se puede deber al diferente esfuerzo de captura en refugios; muchos lugares muestreados o bien no tenían murciélagos ni vestigios de poblaciones, o bien cuando estaban en tejados, eran de difícil captura y observación, es decir, a demasiada altura (superior a 3 m) o bajos (inferior a 50 cm de altura).
Aún presentando diferente esfuerzo de captura en las cuatro zonas de la ciudad de Boa Vista, se prueba que Artibeus lituratus y A. planirostris están presentes en las cuatro zonas urbanas (Tabla 1), así como Molossus molossus,
Tabla 1 – Lista de familias y especies por gremio alimenticio de quirópteros registradas para el área urbana, en las cuatro zonas de muestra en Boa Vista, de agosto de 2011 a julio de 2012.
Especie Nicho tróficoNúmero de individuos
TotalNorte Sur Este Oeste
Emballonuridae 1 16 1 18Rhynchonycteris naso (Wied-Neuwied, 1820) Insectívoro 16 16Saccopteryx bilineata (Temminck, 1838) Insectívoro 1 1 2Noctilionidae 0 50 0 0 50Noctilio albiventris Desmarest, 1818 Insectívoro 50 50Phyllostomidae 17 103 15 218 353Micronycteris minuta (Gervais, 1856) Insectívoro 2 2Phyllostomus discolor Wagner, 1843 Omnívoro 5 7 12Lophostoma silvicolum d’Orbigny, 1836 Insectívoro 1 1Trinycteris nicefori (Sanborn, 1949) Insectívoro 1 1Glossophaga soricina (Pallas, 1766) Polinívoro 2 3 4 9Carollia perspicillata (Linnaeus, 1758) Frugífero 4 9 1 5 19Ametrida centurio Gray, 1847 Frugífero 1 1Artibeus cinereus (Gervais, 1856) Frugífero 1 1Artibeus lituratus (Olfers, 1818) Frugífero 3 22 2 7 34Artibeus planirostris (Leach, 1821) Frugífero 10 65 5 186 266Artibeus obscurus (Schinz, 1821) Frugífero 1 5 6Platyrrhinus lineatus (Geoffroy, 1810) Frugífero 1 1Vespertilionidae 1 4 5Eptesicus brasiliensis (Desmarest, 1819) Insectívoro 1 1Eptesicus furinalis (d’Orbigny, 1847) Insectívoro 2 2Lasiurus egregius Peters, 1870 Insectívoro 1 1Myotis albescens (Geoffroy, 1806) Insectívoro 1 1Molossidae 1 71 22 82 176Cynomops planirostris Peters, 1865 Insectívoro 20 20Molossops temminckii Burmeister, 1854 Insectívoro 1 2 3Molossus molossus (Pallas, 1766) Insectívoro 50 22 80 152Promops nasutus (Spix, 1823) Insectívoro 1 1Total 19 225 53 305 602

17Barbastella 7 (1) 2014
Murciélagos (Mammalia: Chiroptera) del área urbana del municipio de Boa Vista, Roraima, Brasil
con un mayor número de capturas mientras que Saccopteryx bilineata, Lophostoma silvicolum, Trinycteris nicefori, Ametrida centurio, Artibeus cinereus, Platyrrhinus lineatus, Eptesicus brasiliensis, Lasiurus egregius, Myotis albescens y Promops nasutus se capturaron en una única ocasión para el área urbana de Boa Vista.
Los primeros estudios realizados en el estado de Roraima se llevaron a cabo en áreas protegidas (Taddei y Reis, 1980; Mok et al., 1982; Robinson, 1998). Recientemente Souza (2011) realizó un último estudio en área urbana, donde registró para el Estado 18 especies y 11 para Boa Vista. En el presente estudio, han sido capturadas 23 especies, incluyendo 12 nuevas especies para la localidad.
Es preocupante la actual expansión urbana de la ciudad de Boa Vista y se hace necesario conocer y comprender mejor la ecología y el comportamiento de las especies mostradas. A diferencia de otras áreas urbanas del país, la quiropterofauna de Boa Vista aún usa principalmente el ambiente periurbano con refugios próximos a la vegetación, provocando que pocas especies y pocos individuos se encuentren en construcciones. Los estudios se deberían complementar con la realización de una educación ambiental más enfocada a la conservación de los murciélagos, que contribuya a la desmitificación de estos, y que al contrario presente a la comunidad su importancia económica y ecológica.
AgradecimientosAgradecemos a la Facultad Cathedral, al Prof. Alexandre
Castilho, a Ubirajara Dutra Capaverde Neto, a Maria Fernanda Bessa, a Drª Solange Capaverde, al Prof. Dr. Paulo Sergio Ribeiro de Mattos y a Med. Vet. Srª Ana Nóbrega.
ReferenciasAguirre, L.F.; Vargas, A.; Solari, S. 2009. Clave de campo
para la identificación de los murciélagos de Bolivia. Centro de Estudios en Biología Teórica y Aplicada. Cochabamba: Etreus. 38 p.
Aguita, R. 2003. Urbanitas con patas: animales en la gran ciudad. Ambienta, Madrid, v. 20, pp.32-37.
Barquez; M. R.; Giannini, P. N.; Mares, A. M. 1993. Guide to the bats of Argentina. Oklahoma: Museum of Natural History, 282 p.
Gregorin, R.; Taddei, V. A. 2002. Chave artificial para a identificação de molossídeos brasileiros (Mammalia, Chiroptera). Mastozoología Neotropical, Tucumán, v. 9, n. 1, pp. 13-32.
Köppen, W. 1948. Climatologia: con un estudio de los climas de la tierra. Fondo de Cultura Econômica. México. 479p.
Kunz, T. H.; Kurta, A. 1988. Capture methods and holding devices. pp. 1-29. In: (Kunz, T. H. 1998. (Ed). Eco-logical and behavioral methods for the study of bats. Washington: Smithsonian Press.)
Lima, I. P. 2008. Espécies de morcegos (Mammalia, Chiroptera) registradas em parques nas áreas urbanas do Brasil e suas implicações no uso deste ambiente. pp. 71-85. In: (Reis, N. R.; Peracchi, A. L.; Santos, G. A. S. D. (Ed.). Ecologia de morcegos. Londrina: Technical Books.)
Mok, W. Y.; Wilson, D. E.; Lacey, L. A.; Luizão, R. C. C. 1982. Lista atualizada de quirópteros da Amazônia brasileira. Acta Amazônica, Manaus, v. 12, pp. 817-23.
Pacheco, S. M.; Marques, R. V.; Grillo, H.; Marder, E.; Bianconi, G.; Miretzki, M.; Passos I. L.; Rosa, V. A. 2008. Morcegos urbanos da Região Sul do Brasil. pp. 415-426. In: (Pacheco, S. M.; Marques, R. V.; Esbérard, C. E. L. (Org.). Morcegos no Brasil: biologia, sistemática, ecologia e conservação. Porto Alegre: A. Digital.)
Pacheco, S. M.; Sodré, M.; Gama, A. R.; Bredt, A.; Cavallini, E. M.; Marques, R. V.; Guimarães, M. M.; Bianconi, G. 2010. Morcegos urbanos: status do conhecimento e plano de ação para a conservação no Brasil. Chiroptera Neotropical, Brasília, v. 16, n. 1, pp. 629-647.
Paglia, A. P.; Fonseca, G. A. B.; Rylands, A. B.; Herrmann, G.; Aguiar, L. M. S.; Chiarello, A. G.; Leite, Y. L. R.; Costa, L. P.; Siciliano, S.; Kierulff, M. C. M.; Mendes, S. L.; Tavares, V. C.; Mittermeier, R. A.; Patton, J. L. 2012. Lista anotada dos mamíferos do Brasil. 2 ed. Occasional Papers in Conservation Biology, Arlington, n. 6, pp. 1-76.
Pedro, W. A. 1998. Diversidade de morcegos em habitats florestais fragmentados do Brasil. 1998. 128 f. Tese (Doutorado em Ecologia e Recursos Naturais) - Universidade Federal de São Carlos, São Carlos.
Robinson, F. 1998. The bats of the Ilha de Maracá. In: J.A. Ratter, J.A.; Milliken, W. (Eds.). Maracá: the biodiversity and environment of an Amazonian Rainforest. London: John Wiley & Sons. pp. 165-188.
Sebrae – serviço brasileiro de apoio às micro e pequenas empresas. 2006. Série Diagnósticos Municipais: Município de Boa Vista. 3 ed. Boa Vista: Sebrae-RR, 94 p.
Souza, J.R. 2011. Pesquisa do vírus da raiva em quirópteros no estado de Roraima. 63 f. Dissertação (Mestrado em Recursos Naturais) – Universidade Federal de Roraima, Boa Vista.
Straube, F.C. & Bianconi, G. 2002. Sobre a grandeza e a unidade utilizada para estimar esforço de captura com utilização de redes de niebla. Chiroptera Neotropical, Brasília, v. 8, n. 1/2, pp. 150-152.
Taddei, V. A.; Reis, N. R. 1980. Notas sobre alguns morcegos da Ilha de Maracá, Território Federal Roraima (Mammalia, Chiroptera). Acta Amazônica, Manaus, v. 10, pp. 363-68.

18 Barbastella 7(1) 2014
Ubirajara Dutra Capaverde junior, Susi Missel Pacheco, Marcos Eugenio Duarte
Uieda, W.; Bredt, A.; Pinto, P. P. 2008. Dieta, abrigos e comportamento fitófago de Artibeus lituratus (Phyllostomidae) em Brasília, DF, e sua relação com as plantas usadas na arborização urbana. pp. 427-443. In: (Pacheco, S. M.; Marques, R. V.; Esbérard, C. E. L. (Org.). Morcegos no Brasil: biologia, sistemática, ecologia e conservação. Porto Alegre: A Digital.)
Vizotto, L. D.; Taddei, V. A. 1973. Chave para determinação de quirópteros brasileiros. São José do Rio Preto: Francal. 72 p.
Yalden, D. W. 1980. Notes on the diet of urban Kestrels. Bird Study, London, v. 27, pp. 235-238. DOI: https://dx.doi.org/10.1080/00063658009476683

19Barbastella 7 (1) 2014
Barbastella 7 (1) Open Access ISSN: 1576-9720 SECEMU www.secemu.org
Primera cita de nóctulo mediano (Nyctalus noctula) en Aragón (NE de España)
Luís Lorente Villanueva¹*, Carlos LLana Ugalde², José Manuel Sánchez Sanz³
¹ Avda. Puente del Pilar 23, 6ºB, 50014, Zaragoza² Conservación de la Biodiversidad, Servicio Provincial de Zaragoza, Departamento de Agricultura, Ganadería y Medio Ambiente, Pº María Agustín 36, 50071, Zaragoza ³ C/ Avda. de Movera nº 483, casa 5, 50194, Zaragoza
*Corresponding author e-mail: [email protected]
DOI: http://dx.doi.org/10.14709/BarbJ.7.1.2014.04
English title: First record of Noctule Bat (Nyctalus noctula) in Aragón (NE Spain)
ResumenCuatro adultos de nóctulo mediano (Nyctalus noctula) fueron capturados recientemente en un estanque del Mon-asterio de Piedra (Nuévalos, Aragón, NE de España). Estas citas constituyen los primeros datos de su presencia en la Comunidad Autónoma de Aragón y uno de los pocos lugares conocidos con agrupaciones de esta especie en la Península Ibérica.
Palabras clave: Nyctalus noctula, primera cita, Aragón, España.
Abstract Four adults Noctule Bat (Nyctalus noctula) were recently netted over a pond in the Monasterio de Piedra (Nuévalos, Aragón, NE Spain). These are the first unambiguous catches of this species in the Autonomous Com-munity of Aragón. Besides, this site is one of the few localities in the Iberian Peninsula with groups of this bat species.
Key words: Nyctalus noctula, first record, Aragón, Spain.
accepted: Decenmber 21, 2013
Primera cita de nóctulo mediano (Nyctalus noctula) en Aragón (NE de España)
IntroducciónEl nóctulo mediano Nyctalus noctula (Schreber, 1774)
tiene una distribución Paleártica, extendiéndose desde la Península Ibérica hasta Asia Central, y entre el sur de Escandinavia y quizás el norte de África (Simmons 2005). Es una especie migratoria que normalmente forma las colonias de cría en el centro y norte de Europa, y en algunas localidades aisladas en el sur (Ruedi et al. 1998; Dietz et al. 2009). En Europa, numerosas hembras migran hacia el norte para criar, y entre finales del verano y principios de otoño vuelven a las áreas meridionales de apareamiento e hibernación (Dietz et al. 2009; Alcalde 1999). Los territorios de verano pueden estar alejados más de 1000 km de los de invierno (Hutterer et al. 2005).
En España es una especie rara y muy localizada. Se tiene conocimiento de una colonia de cría en Aranjuez (Ruedi
et al. 1998) y de otras tres colonias en Navarra (dos) y Lleida (una) con machos sedentarios durante gran parte del año, y con hembras que acuden para aparearse e hibernar, permaneciendo desde finales de agosto hasta marzo o mayo (Alcalde 1999; Camprodon & Guixé 2013).
Los resultados de este artículo se han obtenido en el marco de un inventario de los murciélagos que se está realizando en el Monasterio de Piedra (Nuévalos, SO Zaragoza), y que está declarado Conjunto de Interés Cultural en la categoría de Jardín Histórico.
Material y métodosPara la captura de los murciélagos se emplearon tres
redes finas de 9 m (Ecotone, Polonia), que fueron colocadas simultáneamente durante cinco noches, dos en otoño de

20 Barbastella 7(1) 2014
Luis Lorente Villanueva, Carlos Llana Ugalde, José Manuel Sanchez Sanz
2012, y tres en verano de 2013, sobre un estanque situado en el interior de los jardines del Monasterio de Piedra a 750 m de altitud (Datum: ETRS89, Huso UTM: 30T X: 602200, Y: 4560740). El entorno del punto de captura está formado por una alineación de bosque de ribera con árboles de especies autóctonas y por varios rodales con árboles ornamentales, entre los que destacan casi un centenar de plátanos de sombra (Platanus hybridus) de gran porte, plantados a finales del siglo XIX. Rodeando este enclave forestal de fondo de valle fluvial se extienden amplias zonas despejadas sobre sustrato calizo cubiertas de matorral mediterráneo y cultivos de secano en cotas superiores.
Resultados y discusiónEn las dos jornadas de trampeo realizadas a mediados del
mes de octubre de 2012 no fue capturado ningún nóctulo. Se repitieron otras dos sesiones en julio y otra en septiembre de 2013. El 18 de julio de 2013 fue capturado un macho adulto de nóctulo mediano, que fue identificado por la longitud del antebrazo, (antebrazo: 53,64 mm, 36,1 g), la morfología del trago y las alas (Dietz et al. 2009). Posteriormente, el 20 de septiembre de 2013 se capturaron dos hembras adultas (antebrazo y peso: 52,53 mm, 31,3 g; 54,86 mm, 36,3 g) y un macho adulto (52,36 mm, 33,3 g) sexualmente activo con glándula bucal característica y testículos escrotales. La Estación Biológica de Doñana (CSIC), confirmó la identificación de todos los individuos mediante análisis molecular de tejido alar obtenido con un “biopunch”.
N. noctula ha sido citado previamente en Aragón por Woutersen & Bafaluy Zoriguel (2001) pero la cita se considera dudosa al carecer de datos biométricos o genéticos que la avale (Alcalde 2008). Por ello, puede considerarse que ésta es la primera cita fiable de esta especie en Aragón.
La abundancia de N. noctula en Europa contrasta con su rareza en la Península Ibérica (Ibáñez et al. 1992). La cita que se aporta se suma a las pocas y dispersas que se han recopilado, y contribuye a mejorar el conocimiento que se tiene de las agrupaciones conocidas de la especie en España (Fig.1). Se observa una distribución claramente septentrional (Alcalde 2007), y en cierta medida existe cierta agregación espacial, considerando que tres de las cuatro provincias donde se han localizado las agrupaciones son adyacentes.
Los datos obtenidos sugieren la presencia de una agrupación de Nyctalus noctula en el Monasterio de Piedra, que podría mostrar el mismo patrón observado en otras colonias (Alcalde 2008; Camprodon & Guixé 2013). Es decir, que ésta podría estar formada por una colonia de machos sedentarios, a la cual se unirían las hembras, y probablemente otros machos, desde finales del verano hasta la primavera, coincidiendo con el apareamiento y la hibernación.
El nóctulo mediano es una especie arborícola que acostumbra a refugiarse en huecos naturales de frondosas. La presencia en el Monasterio de Piedra de viejos plataneros centenarios en los que abundan orificios y superan en su mayoría los 25 m de altura, parece estar relacionada con la presencia de la especie. Una inspección superficial permitió comprobar la presencia de varios refugios potenciales, a veces
Fig. 1 – Distribución de las localidades con agrupaciones conocidas de Nyctalus noctula en España.

21Barbastella 7 (1) 2014
Primera cita de nóctulo mediano (Nyctalus noctula) en Aragón (NE de España)
situados a elevada altura, tal como los que prefiere seleccionar para evitar la depredación (Ruczynski & Bogdanowicz 2005). En España, todas las agrupaciones conocidas se encuentran en parques urbanos (Ruedi et al. 1998; Alcalde 2007; Camprodon & Guixé 2013) muy distantes unos de otros. Esto induce a sospechar sobre la escasez de áreas forestales naturales adecuadas que satisfagan sus preferencias.
La conservación del conjunto de arbolado ornamental del Monasterio de Piedra parece asegurada por el momento debido a la aparente buena condición sanitaria de los árboles y a la ausencia de podas y otro tipo de manejo que pudiera afectarles. Un estudio de la composición y tamaño poblacional, y la selección de refugios de N. noctula en este enclave permitiría evaluar su importancia y prever alternativas de manejo ante eventuales actuaciones sobre el entorno. Por otra parte sería interesante coordinar el seguimiento que se realizan en las colonias conocidas y comprobar si existe alguna relación entre ellas.
AgradecimientosExpresamos nuestro agradecimiento al director del
Centro Turístico del Monasterio de Piedra José Pont y al ex director Javier Rodríguez, así como al resto del personal empleado, por la inestimable ayuda prestada durante nuestra estancia. A la Sección de Conservación de la Biodiversidad del Servicio Provincial de Zaragoza (Departamento de Agricultura, Ganadería y Medio Ambiente del Gobierno de Aragón) por la financiación de algunos de los trabajos. A la Estación Biológica de Doñana (CSIC) a través de Javier Juste por la identificación molecular de los individuos capturados y a Juan Tomás Alcalde por la revisión del manuscrito y sus sugerencias. A Juan Ignacio Llana por su ayuda y asistencia en el campo. A los miembros del Equipo de Edición por sus correcciones y sugerencias.
ReferenciasAlcalde, J. T. 1999. New ecological data on the noctule
bat (Nyctalus noctula Schreber, 1774) (Chiroptera, Vespertilionidae) in two towns of Spain. Mammalia, 63: 273-280. DOI: https://dx.doi.org/10.1515/mamm.1999.63.3.273
Alcalde, J. T. 2007. Nyctalus noctula (Schreber, 1774) In Palomo, L. J., Gisbert, J., Blanco, J. C. (eds.). Atlas y libro rojo de los mamíferos terrestres de España. Dirección General para la Biodiversidad-SECEM-SECEMU, Madrid. Pp. 228-232.
Alcalde, J. T. 2008. El nóctulo mediano (Nyctalus noctula) en Pamplona. Colección Biodiversidad Urbana de Pamplona. Ayuntamiento de Pamplona. 52 p.
Camprodon, J. & Guixé, D. 2013. Estado poblacional, selección de refugios y ecología espacial de las poblaciones de nóctulo grande (Nyctalus lasiopterus) y nóctulo mediano (Nyctalus noctula) en Cataluña. Barbastella 6 (1): 51-59. DOI: http://dx.doi.org/10.14709/BarbJ.6.1.2013.07
Dietz, C. I, Helversen, O. & Nill, D. 2009. Bats of Britain, Europe & Northwest Africa. London: A&C Black Publishers Ltd.
Hutterer. R., Ivanova, T., Meyer-Cords, C. & Rodrigues, L. 2005. Bat migrations in Europe - A review of banding data and literature. Federal Agency for Nature Conservation, Bonn, 94-96.
Ibáñez, C., Guillén, A., Fernández, R., Pérez, J. L. & Guerrero, S. I. 1992. Iberian distribution of some little known bat species. Mammalia, 56 (3): 433-444. DOI: https://dx.doi.orh/10.1515/mamm.1992.56.3.433
Ruczynski, I. & Bogdanowicz, W. 2005. Roost cavity selection by Nyctalus noctula and N. leisleri (Vespertilionidae, Chiroptera) in Bialowieza Primeral Forest, eastern Poland. Journal of Mammalogy 86(5): 921-930. DOI: http://dx.doi.org/10.1644/1545-1542(2005)86[921:RCSBNN]2.0.CO;2
Ruedi, M., Tupinier, Y. & Paz, O. de, 1998. First breeding record for the noctule bat (Nyctalus noctula) in the Iberian Peninsula. Mammalia, 62: 301-304.
Simmons, N.B. 2005. Order Chiroptera. In: Wilson D.E, & Reeder, D.M. (eds) Mammal species of the world, Vol 1, 3rd edn. Johns Hopkins University Press, Baltimore, MD, p 312–529.
Woutersen, K. & Bafaluy Zoriguel, J. J. 2001. Murciélagos del Alto Aragón. Ed. La Val de Onsera, Huesca.
Fig. 2 – Nyctalus noctula.

22 Barbastella 7(1) 2014
Barbastella 7 (1) Open Access ISSN: 1576-9720 SECEMU www.secemu.org
Pasos agrícolas inferiores de carreteras: su importancia para los murciélagos como refugio y lugar para cruzar la vía
Paulo Barros
Laboratorio de Ecologia Aplicada, CITAB - Centro de Investigação e Tecnologias Agroambientais y Biológica. Universidade de Trás-os-Montes e Alto Douro, 5000-801
Corresponding author e-mail: [email protected]
DOI: http://dx.doi.org/10.14709/BarbJ.7.1.2014.05
English title: Agricultural underpasses: their importance for bats as roosts and role in facilitating movement across roads
ResumenCon objeto de ampliar el conocimiento sobre la utilización de los pasos inferiores agrícolas por parte de los murciélagos se han desarrollado trabajos de detección acústica, captura y revisión de refugios en cinco pasos inferiores ubicados a lo largo de 8250m de un tramo de la carretera situada en el Norte de Portugal en una zona rural de uso agrícola y pastoreo. Los pasos tienen las dimensiones medias de 9 m de anchura, 4,25 m de altura y 34 m de profundidad. Los resultados de los registros acústicos y de captura de murciélagos con redes de niebla, permitieron confirmar la utilización de por lo menos 12 especies (Pipistrellus pipistrellus, P. kuhlii, P. pygmaeus, Myotis daubentonii, M. escalerai, M. myotis, Nyctalus leisleri, Plecotus austriacus, Rhinolophus ferrumequinum, R. hipposideros, R. mehelyi/R. euryale y Eptesicus serotinus/E. isabellinus). La inspección y revisión de refugios en los 5 pasos inferiores han permitido constatar la presencia de por lo menos 6 especies (M. daubentonii, M. myotis, N. leisleri, P. austriacus, Eptesicus serotinus/E. isabellinus y Pipistrellus sp.) que usaron fisuras dentro de los túneles de entre 1,2 y 1,9 cm. Para una misma altura (4,2-4,3m) y anchura de túnel (9m), los pasos más utilizados fueron los de mayor longitud. Estos resultados confirman la importancia que los pasos inferiores tienen en la conectividad y minimización de los impactos de las carreteras, así como proporcio-nan nuevos refugios artificiales para los murciélagos. Se incita a promover seguimientos y estudios ecológicos más detallados que profundicen en el conocimiento de la utilización de los quirópteros de estas estructuras así como sus características físicas y su entorno y en otras carreteras.
Palabras clave: Carreteras, Pasos inferiores, Murciélagos, Conservación, Portugal.
Abstract The main goal of this study was to contribute for the knowledge of the use of under-roads passageways by bats in northern of Portugal. We selected five underpasses in a road stretch of 8.250 m located in a rural agricultural and grazing area. The average dimensions of underpasses were, 9 m wide, 4.25 m high and 34 m deep. The data was obtained through acoustic detection, mist net capture and roost inspection. Acoustic and mist netting results confirm the use of the underpasses by at least 12 species (Pipistrellus pipistrellus, P. kuhlii, P. pygmaeus, Myotis daubentonii, M. escalerai, M. myotis, Nyctalus leisleri, Plecotus austriacus, Rhinolophus ferrumequinum, R. hipposideros, R. mehelyi/R. euryale and Eptesicus serotinus/E. isabellinus). Data from roost inspection in all the underpasses showed the presence of at least six species (M. daubentonii, M. myotis, N. leisleri, P. austriacus, Eptesicus serotinus/E. isabellinus y Pipistrellus sp.), they used fissure within the tunnels of between 1.2 and 1.9 cm. For the same height (4.2-4.3 m) and width (9m), the underpasses most used were the longest. These results stress the importance of underpasses in improving the connectivity and permeability between areas, along with its potential to offer new roosts for bats. We encourage further monitoring and detailed ecological studies to bet-ter understand the physical and environmental characteristics that underpin the usage of such structures by bats.
Key words: Roads, Underpasses, Bats, Conservation, Portugal
accepted: January 18, 2014
Paulo Barros

23Barbastella 7 (1) 2014
Pasos agrícolas inferiores de carreteras: su importancia para los murciélagos como refugio y lugar para cruzar la vía
IntroducciónLas carreteras son infraestructuras cada vez más
presentes en los paisajes actuales (Giulio et al. 2009), mejoran la comunicación humana y desempeñan un papel social importante en la sociedad de hoy en día (Trombulak & Frissell, 2000; Hawbaker et al 2004). Sin embargo, tienen una amplia gama de impactos ecológicos negativos en la vida silvestre (Abbott et al, 2012), además de destruir y degradar el hábitat natural (Forman et al., 2003), son un factor importante, causante de la pérdida de biodiversidad en todo el mundo (Giulio et al.2009).
Se han realizado muchos estudios centrados en la mortalidad de mamíferos no voladores (Barthelmess & Brooks, 2010), aves (Gomes et al., 2008), anfibios (Eigenbrod et al., 2008) y reptiles (Woltz et al., 2008). No obstante, los murciélagos son especialmente vulnerables, debido a su gran movilidad y necesidad de recursos alimenticios dispersos en el paisaje, tienden a ponerse frecuentemente en contacto con las carreteras (Abott et al., 2012), sin embargo existen pocos estudios centrados en ellos (Bafaluy, 2000; Gaisler et al., 2009; Lesiński et al., 2011). De hecho, el estudio de mortalidad de las poblaciones de murciélagos en carreteras, es un campo relativamente nuevo en la conservación y la mayoría de la información procede de Europa oriental y central (e. g. Limpens et al , 2005; Lesiński, 2008; Gaisler et al., 2009; Kerth & Melber2009; Lesiński et al., 2011; Berthinussen & Altringham, 2012b). En América del Norte y el sur de Europa, la información sigue siendo muy escasa (González -Prieto et al., 1993; Bafaluy, 2000) y todavía no somos capaces de predecir los lugares exactos donde los murciélagos pueden hallarse particularmente amenazados o las especies que pueden estar en mayor riesgo (Lesiński et al., 2011). Otro impacto negativo descrito de las carreteras en los murciélagos, es la fragmentación del hábitat, en especial los que utilizan las estructuras naturales del paisaje como elementos de orientación (Limpens et al., 2005).
El impacto de las carreteras sobre la vida silvestre, ha llevado a un mayor reconocimiento de la necesidad de construir estructuras que faciliten su cruce en seguridad (Trocmé et al., 2003; Beckmann et al, 2010). Varios estudios han investigado como los vertebrados no voladores, utilizan estructuras viales diseñadas para el drenaje u otros usos humanos (Ascensão & Mira, 2007) y los diseñados específicamente como puntos de cruce para la fauna (Clevenger & Waltho, 2000). Aunque los murciélagos también son conocidos por utilizar los pasos subterráneos para cruzar carreteras anchas (Bach et al., 2004; Kerth & Melber, 2009), raramente se incluyen en las investigaciones. Directrices nacionales para la mitigación de los impactos de tráfico en los murciélagos se han publicado en varios países, debido al estado de conservación comprometido y la protección jurídica de los murciélagos en Europa (Abott et al., 2012). Muchos de ellos recomiendan la implementación de pasos inferiores para facilitar el paso (Limpens et al., 2005, National Roads Authority, 2006; Highways Agency UK, 2008; Sétra, 2008). Sin embargo, el conocimiento del uso de estos pasos por los murciélagos sigue siendo muy limitado (Bach et al., 2004). La capacidad de volar en espacios reducidos es probablemente uno de los principales factores que determinan el uso de los pasos por debajo de la carretera (Abott et al., 2012). Los pasos inferiores, además de proporcionar a los murciélagos seguridad en el cruce de las carreteras, pueden también proporcionar refugios muy adecuados; como en los espacios en cornisas, los huecos en fachadas, las juntas de dilatación, los desagües, la superficie interior del arco de piedra y sus fisuras (Sétra, 2008). De hecho, la pérdida de refugios naturales debido al cambio y uso de la tierra, ha aumentado la importancia de los refugios artificiales y en algunos casos estos son esenciales para la supervivencia de muchas especies de murciélagos (Bat Conservation Trust, 2012). Por ello, el conocimiento sobre el uso de los pasos inferiores de carreteras es de clara relevancia práctica para la planificación de medidas específicas dirigidas a conservar las poblaciones de murciélagos (Abott et al., 2012).
Fig. 1 – Localización de la área de estudio

24 Barbastella 7(1) 2014
A pesar de los posibles efectos positivos que los pasos inferiores de carreteras puedan representar para los murciélagos, el impacto de estas estructuras permanece en gran parte sin cuantificar. Por ello este trabajo tiene como principal objetivo aportar información original y aumentar el conocimiento de la utilización de los pasos inferiores de carreteras por parte de los murciélagos, que podrán ayudar a la hora de poner en práctica medidas de mitigación eficientes para los murciélagos, en la construcción de carreteras.
MetodologíaLa información presentada en este artículo, es el
resultado de los trabajos de campo realizados en los meses de agosto (detección acústica, captura y revisión de refugios) y diciembre (revisión de refugios) del año 2013 en los cinco pasos inferiores ubicados a lo largo de 8250m del tramo construido en 2005, de la carretera N212 (41º 18’ 58.23’’N; 7º 28’ 28.58’’O), situada en el Norte de Portugal (Figura 1), Concelho de Alijó, a una altitud que varía entre los 800 y los 600 m.s.n.m.. Todos los pasos agrícolas inferiores están localizados en zona rural de uso agrícola y pastoreo, con estructuras lineales, como setos, muros de piedra y líneas de árboles que proporcionan conectividad para los murciélagos (Limpens & Kapteyn, 1991; Verboom & Huitema, 1997).
Se realizaron sesiones de captura en los pasos inferiores. La captura se efectuó con redes de niebla de cinco bolsas (14mm), colocadas dentro de los pasos agrícolas inferiores y ajustadas a la dimensión (Vintulis & Šuba, 2010) de cada paso agrícola inferior. Las 5 sesiones de captura (una por paso) realizadas en agosto, tuvieron una duración de 3 horas, las de mayor actividad de murciélagos (Bat Conservation Trust, 2012). La identificación e los ejemplares capturados se realizó en mano, mediante la utilización de caracteres morfológicos y de dentición (Dietz & Helversen, 2004; Dietz et al., 2009); además, se tomaron datos sobre el sexo, así como las mediciones estándar de peso (±0.1 g) y longitud del antebrazo (±0,01 mm). Las redes fueron instaladas al anochecer (en el ocaso) y se revisaron cada 10 minutos, tomando nota de la hora de cada captura. Con el propósito de no duplicar datos, todos los individuos fueron marcados en el codo con esmalte de uñas (Gloza-Rausch et al., 2008; Stawski, 2012) después de ser procesados, fueron puestos en libertad, de inmediato.
Las 5 estaciones de escucha (una por paso) realizadas en agosto, tuvieron una duración de 3 horas a partir del ocaso y fueron realizadas con detectores de ultrasonidos Pettersson Elektronik® AB Mod. D 240X, para grabar
Fig. 2 – Tipología de los pasos (paso IV). Leyenda: A-longitud; B-altura; C-anchura
Fig. 3 – Ocupación del suelo en un buffer de 1km de la carretera
Paulo Barros

25Barbastella 7 (1) 2014
en formato digital y en grabadoras Handy Recorder H2, las llamadas de ecolocación y las llamadas sociales (16 bits/44,1 kHz). El análisis de las grabaciones se realizo con el programa informático BatSound 3.3.1®. Los registros grabados fueron transferidos al programa en formato Wave (.Wav), en una frecuencia de muestra de 44.1kHz, con 16 bits/muestra y un número de muestras de la Transformación Rápida de Fourier de 1024. La identificación de las especies fue realizada teniendo en cuenta las siguientes variables: estructura del pulso; frecuencia inicial (Fi, kHz) y mínima (Ff, kHz); duración del pulso (D, ms); intervalo entre pulso (IPI, ms) y frecuencia máxima de energía (FMaxE, kHz). Los parámetros caracterizadores de cada una de las variables fueron comparados con los descritos en la bibliografía especializada (e. g. Barataud, 1996; Russo & Jones, 2002; Davidson-Watts et al., 2006; Rainho et al., 2011; Walters et al., 2012). Además de las variables indicadas para la identificación de los Pipistrellus, también se han tenido en cuenta las grabaciones con llamadas sociales.
La revisión de refugios en los pasos agrícolas inferiores implicó una búsqueda previa de señales de murciélagos, como excrementos, manchas de orina y de las características potenciales de las fisuras, así como una revisión general de todas las fisuras con la ayuda de unos prismáticos y un frontal. Los murciélagos encontrados en las fisuras fueron contabilizados e identificados mediante el uso de un endoscopio (Andrew et al., 2013) flexible con pantalla LCD (PCE, modelo DE25).
Características de los pasos Todos los pasos tienen una arquitectura tipo bóveda
(Figura 2) con dimensión media de 9 m de anchura (C), 4,25 m de altura (B) y 34 m de profundidad (A) (Tabla 1). Se trata de pasos inferiores destinados al cruce de caminos agrícolas con baja intensidad de tráfico o vías pecuarias. La base del paso es en sustrato natural (tierra) y las bandas laterales no están pavimentadas, lo que puede favorecer su uso como pasos de fauna. Los cinco pasos en estudio son los únicos
Tabla 1 – Características de los pasos inferiores.
Característica Paso I Paso II Paso III Paso IV Paso V
Altura (m) 4,2 4,3 4,3 4,2 4,3
Anchura (m) 9 9 9 9 9
Longitud (m) 28 34 28 38 42
Openness* 1,35 1,13 1,38 0,99 0,92
Hábitat dominante Agrícola Matorral Matorral Viñedos Viñedos
Localización (Km+m) 1+100 2+780 3+600 5+650 7+300
*Openness = (altura X anchura)/ profundidad (Olbrich, 1984)
Tabla 2 – Resultados de utilización de los pasos inferiores como locales de paso por los murciélagos. D – identificación acústica; C – identificación morfológica (captura)
Espécie Paso I Paso II Paso III Paso IV Paso V
Pipistrellus pipistrellus D D C, D C, D C, D
Pipistrellus kuhlii D D C, D C
Pipistrellus pygmaeus D D C, D D C, D
Myotis daubentonii D C, D
Myotis escalerai C, D
Myotis myotis C
Nyctalus leisleri C D
Plecotus austriacus C C
Rhinolophus ferrumequinum D D D D
Rhinolophus hipposideros D
Rhinolophus mehelyi/R. euryale D D
Myotis myotis/M. blythii D
Eptesicus serotinus/E. isabellinus D D
Plecotus auritus/P. austriacus D
Total de especies (mínimo y máximo) 8-10 6 4 7-9 7-9
Número de pases/hora 16 38 28 52 62
Pasos agrícolas inferiores de carreteras: su importancia para los murciélagos como refugio y lugar para cruzar la vía
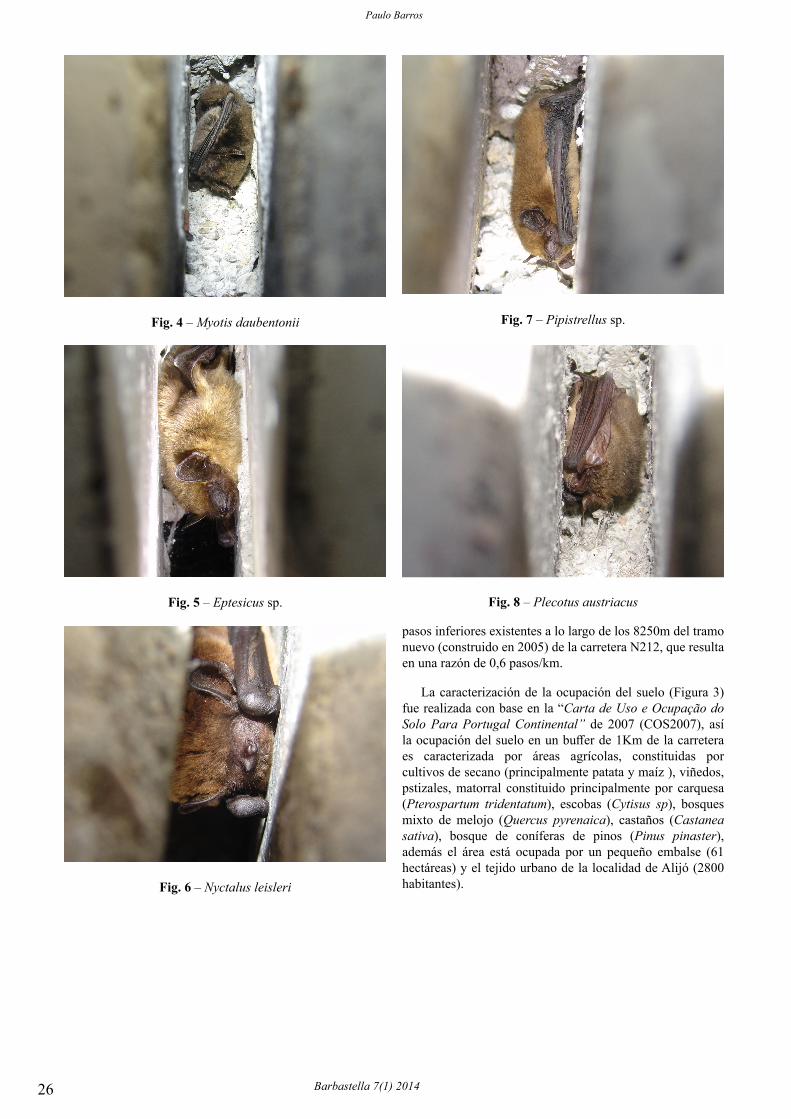
26 Barbastella 7(1) 2014
pasos inferiores existentes a lo largo de los 8250m del tramo nuevo (construido en 2005) de la carretera N212, que resulta en una razón de 0,6 pasos/km.
La caracterización de la ocupación del suelo (Figura 3) fue realizada con base en la “Carta de Uso e Ocupação do Solo Para Portugal Continental” de 2007 (COS2007), así la ocupación del suelo en un buffer de 1Km de la carretera es caracterizada por áreas agrícolas, constituidas por cultivos de secano (principalmente patata y maíz ), viñedos, pstizales, matorral constituido principalmente por carquesa (Pterospartum tridentatum), escobas (Cytisus sp), bosques mixto de melojo (Quercus pyrenaica), castaños (Castanea sativa), bosque de coníferas de pinos (Pinus pinaster), además el área está ocupada por un pequeño embalse (61 hectáreas) y el tejido urbano de la localidad de Alijó (2800 habitantes).
Fig. 4 – Myotis daubentonii
Fig. 5 – Eptesicus sp.
Fig. 6 – Nyctalus leisleri
Fig. 7 – Pipistrellus sp.
Fig. 8 – Plecotus austriacus
Paulo Barros

27Barbastella 7 (1) 2014
ResultadosUso de los pasos inferiores para cruzar la carretera
Los resultados de los 208 registros acústicos y de las 72 captura de murciélagos, permitieron confirmar el cruce de la carretera a través de los pasos inferiores por parte de 12 especies (Pipistrellus pipistrellus, P. kuhlii, P. pygmaeus, Myotis daubentonii, M. escalerai, M. myotis, Nyctalus leisleri, Plecotus austriacus, Rhinolophus ferrumequinum, R. hipposideros, R. mehelyi/R. euryale –probablemente R.euryale- y Eptesicus serotinus/E. isabellinus –probablemente E. serotinus-) y la posibilidad de 4 más (P. auritus, M. blythi, R. mehelyi y E. isabellinus) (Tabla 2). P. pipistrellus y el P. pygmaeus fueron las únicas especies que utilizaron todos los paso inferiores, por lo contrario M. daubentonii, M. escalerai, M. myotis y N. lelsleri fueron las especies capturadas que utilizaron solamente un paso inferior (paso II, IV, II y I, respectivamente), mientras que R. hipposideros, M. myotis/M. blythii y P. auritus/P. austriacus, fueron detectadas acústicamente también en un solo paso inferior (paso IV, I y V, respectivamente) (Tabla 2).
Los resultados obtenidos demuestran que los pasos I, IV y V fueron los que presentaron una mayor riqueza
específica, con un mínimo de 7 y máximo de 10 especies, estos pasos tienen como tipo de uso de suelo dominante áreas agrícolas y viñedos, respectivamente. Por el contario los pasos (II y III) cuyo tipo de uso de suelo dominante es el matorral, demostraron, ser utilizados por un menor número de especies, 6 y 4 respectivamente (Tabla 2 y Tabla 1). No obstante, el número de especies no mantiene una proporción similar con el índice de actividad (número de cruces), ya que por ejemplo en el paso I, con un número de especies elevado (8-10) se obtuvo el índice de actividad más bajo de todos, con 16 cruces/hora. Los datos de índice de actividad apuntan para una proporcionalidad con la Longitud del paso, de hecho los pasos con mayor longitud (II, IV y V) (Tabla 1) presentar una mayor índice de actividad (Tabla 2) e por lo contrario, los pasos más cortos (I y III) (Tabla 1) presentan un número de cruces más reducido (Tabla 2).
Uso de los pasos inferiores como refugioLa inspección y revisión de refugios en los 5 pasos
inferiores en agosto e diciembre han permitido constatar la presencia de por lo menos 4 especies; M. daubentonii (Figura 4), M. myotis, N. leisleri (Figura 6) y P. austriacus) (Figura 8), a las que habría que añadir al menos dos especies más (Tabla 3) de los géneros Eptesicus (Figura 5) y Pipistrellus (Figura 7), cuyos individuos no han podido ser identificados a nivel de especie.
Fig. 9 – Diferencia entre fisuras no ocupadas (A) y ocupadas (B)
Tabla 3 – Resultados de utilización de los pasos inferiores como refugios por los murciélagos. Leyenda: V-verano; I-invierno
EspéciePaso I Paso II Paso III Paso IV Paso V
A D A D A D A D A D
Myotis daubentonii X
Myotis myotis X
Nyctalus leisleri X X
Plecotus austriacus X X
Eptesicus sp. X
Pipistrellus sp. X X X X X X X
Pasos agrícolas inferiores de carreteras: su importancia para los murciélagos como refugio y lugar para cruzar la vía

28 Barbastella 7(1) 2014
La presencia del M. myotis y M. daubentonii, solamente ha podido ser confirmada en la visita de Agosto, mientras que Eptesicus sp, por el contrario, solo se ha observado refugiado en los pasos inferiores en la visita de Diciembre. P. austriacus, N. leisleri y Pipistrellus sp. utilizaron como refugio los mismos pasos inferiores en ambos periodos (agosto e diciembre) (Tabla 3). Todas las especies observadas se refugiaban de forma aislada, aunque en ocasiones, Pipistrellus sp. constituyeron pequeños grupos de 3-6 individuos o bien aislados.Todos los individuos fueron observados en fisuras con una anchura mínima de 1,2 y máxima de 1,9 cm (Tabla 4 y Figura 9), los Pipistrellus sp. ocupaba fisuras más estrechas (1,2-1,4 cm) y las demás especies fisuras más anchas (1,4- 1,9 cm), con una profundidad que variaba entre 17 y 22 cm, indistintamente de la especie.
Discusión El presente trabajo ha permitido comprobar la
utilización de los pasos inferiores por al menos 12 especies de murciélagos (P. pipistrellus, P. kuhlii, P. pygmaeus, M. daubentonii, M. escalerai, M. myotis, N. leisleri, P. austriacus, R. ferrumequinum, R. hipposideros, R. mehelyi/ R. euryale y E. Serotinus/E. isabellinus). La distribución del Eptesicus serotinus y E. isabellinus en la Península Ibérica es de origen alopátrica (Ibáñez et al., 2006). Los individuos del sur pertenecen a la especie E. isabellinus, y las poblaciones del Norte pertenecen a la especie E. serotinus (Ibáñez et al., 2006; Juste et al., 2009), pero existe una zona de contacto entre ambas especies, que hasta el momento, no es claramente conocida, ni sus límites ni su dimensión (Lisón et al., 2010). Así, y a pesar de que E. isabellinus fue recientemente identificado en el Norte de la Beira Interior de Portugal (Barros, 2011), las grabaciones acústicas de este género fueron consideradas como Eptesicus serotinus/E. isabellinus. Teniendo en cuenta la localización del área de estudio, lo más probable es que los individuos detectados pertenezcan a E. serotinus. Lo mismo ocurre con la determinación de la especie de Rhinolophus euryale/R. mehelyi, aunque probablemente la llamada de ecolocación registrada pertenezca a R. euryale, ya que R. mehelyi en el Norte de Portugal es escasa y está desapareciendo en gran parte de su área de distribución (Flaquer et al. 2010, Lisón et al. 2010, 2011).
No obstante, la poca antigüedad de esta carretera (8 años), las 12 especies que utilizan estos pasos representan cerca del
50% de las especies presentes en Portugal Continental, lo que representan una elevada diversidad, cuando comparado con estudios similares (Kerth & Melber, 2009; Abbott et al., 2012). El alto índice de actividad (número de cruces) y la alta diversidad de especies registradas parecen indicar un elevado grado de adaptación de los murciélagos a estas estructuras. La buena densidad de los pasos en el tramo estudiado (0,6 pasos/km) permite una mayor permeabilidad (Ascensão & Mira, 2007), lo que podría explicar la intensidad de uso y riqueza específica observada en estos pasos. No obstante de estos resultados, habría que discernir si los datos sugieren un elevado grado de adaptación o simplemente un uso en la misma intensidad que el resto del entorno, lo que necesitaría la realización de estudios más amplios y detallados de la actividad e riqueza de murciélagos en los entornos de los pasos. De hecho, en Alemania se verifico que las captura en los pasos inferiores eran significativamente más elevadas en algunas especies que en los entornos, lo que sugiere que la eficacia de cruce en los pasos inferiores, puede ser mayor para algunas especies (Kerth & Melber, 2009) y en Irlanda, se observó la tendencia a volar a través de los pasos inferiores en lugar de sobre la carretera, lo que sugiere un grado de adaptación de las especies (Abbott et al., 2012).
La arquitectura del tipo bóveda, es utilizada por un gran número de especies (Sétra, 2008; Abbott et al., 2012), de hecho la relación entre sección de cruce y longitud del paso es de extrema importancia para su utilización por parte de los murciélagos (Limpens et al., 2005). La luminosidad dentro de un paso tiene un efecto contrario, incluso para especies que normalmente se alimentan en torno de iluminación artificial, como por ejemplo los Pipsitrellus (Limpens et al., 2005). La mayoría de los murciélagos tienen una grande capacidad de volar en espacios confinados (Abbott et al., 2012) y evitan espacios amplios y abiertos, visto que se sienten más vulnerables a los predadores (Limpens et al., 2005). De hecho los datos de índice de actividad apuntan para una proporcionalidad con la longitud del paso, los con mayor longitud (II, IV y V) y consecuentemente los más oscuros presentar una mayor índice actividad, lo que reflecte una propensión de cruce en los pasos de “openness” de 1 o próximo.
En los hábitats terrestres el aumento de la urbanización tienen en general un efecto negativo en la riqueza de las especies, aunque este patrón no es universal (McKinney, 2008), de hecho, la proximidad de los pasos I, IV y V a zonas urbanizadas pudo tener influencia en la diversidad
Tabla 4 – Anchuras media (en centímetros) de la fisura registradas para cada especie y número de fisuras.
Espécie Paso I Paso II Paso III Paso IV Paso VMyotis daubentonii 1,5 (2)Myotis myotis 1,9 (1)Nyctalus leisleri 1,6 (3)Plecotus austriacus 1,53 (3)Eptesicus sp. 1,85 (2)Pipistrellus sp. 1,25 (3)Pipistrellus sp. 1,3 (4)Pipistrellus sp. 1,25 (6)Pipistrellus sp. 1,23 (4)
Paulo Barros

29Barbastella 7 (1) 2014
de las especies y en la presencia de especies consideradas antropófilas (e.g. M. escalerai, P. austriacus, Eptesicus sp o Rhinolophus sp.). El índice de actividad de murciélagos (cruces/hora) registrado en los pasos inferiores fue variable entre estos, pero se encuentra recogido en otros estudios (e.g. Bach et al., 2004; Kerth & Melber, 2009; Abbott et al., 2012; Berthinussen & Altringham, 2012a).
Muchas estructuras construidas en carreteras, como pasos inferiores, viaductos y puentes tienen un impacto positivo, proporcionando nuevos abrigos para los murciélagos (Limpen et al., 2005, Seiler & Helldin, 2006; Ceľuch & Ševčík, 2008; Highways Agency UK, 2008; Sétra 2008; Am==orim et al., 2013; Medinas et al., 2013), además, muchas especies demuestran una alta fidelidad a los refugios (Racey & Swift 1985; Entwistle et al., 2000; Senior et al., 2005; Dietz et al., 2009). De hecho, en este trabajo se ha confirmado que los pasos inferiores son utilizados como refugio al menos por 6 especies (M. daubentonii, M. myotis, N. leisleri y P. austriacus, Eptesicus sp. y Pipistrellus sp.), estos resultados, se deben principalmente, a la cantidad de fisuras con anchura de 1,2-1,9 mm existentes entre las piezas prefabricadas del hormigón que representan refugios potenciales.
En conclusión, la importancia que estas estructuras representan para los murciélagos como paso o refugio, resulta fundamental como medida de conservación y a la hora de su implementación, deben ser tenidas en cuenta características como la longitud de los pasos y la cantidad de fisuras con anchura adecuadas para el refugio de murciélagos. Por último, creemos muy necesaria la realización de estudios más detallados que profundicen el conocimiento de la utilización de los quirópteros de estas estructuras así como sus características físicas, su entorno y en otras carreteras.
AgradecimientosDeseo expresar mi agradecimiento a Lúcia Pacheco y Hélia Vale-Gonçalves por su ayuda en la revisión del borrador inicial. Un agradecimiento especial a Manuel Arzúa Piñeiro por la revisión y sus aportaciones para mejorar el manuscrito. Agradezco igualmente los valiosos comentarios de los tres revisores anónimos, los cuales han ayudado a mejorar el manuscrito original.
Todas las capturas se han efectuado con la correspondiente autorización (69/2013/CAPT) del Instituto de Conservação da Natureza e Florestas (ICNF), autoridad responsable de la emisión de los permisos de capturas.
ReferenciasAbbott, I. M., Harrison, S. & Butler, F. 2012. Clutter-
adaptation of bat species predicts their use of under-motorway passageways of contrasting sizes – a natural experimente. Journal of Zoology 287:124–132. DOI: https://dx.doi.org/10.1111/j.1469-7998.2011.00894.x
Amorim, F. Alves, P. & Rebelo, H. 2013. Bridges over the troubled Conservation of Iberian Bats. Barbastella, 6(1):3-12. DOI: http://dx.doi.org/10.14709/BarbJ.6.1.2013.01
Andrews, H. 2013. Bat Tree Habitat Key. AEcol, Bridgwater
Ascensão, F. & Mira, A. 2007. Factors affecting culvert use by vertebrates along two stretches of road in southern Portugal. Ecology Resources, 22:57–66. DOI: https://dx.doi.org/10.1007/s11284-006-0004-1
Bach, L., Burkhardt, P. & Limpens, H. J. G. A. 2004. Tunnels as a possibility to connect bat habitats. Mammalia, 68:411–420. DOI: https://dx.doi.org/10.1515/mamm.2004.041
Bafaluy, J. 2000. Mortandad de murciélagos por atropello en carreteras del sur de la provincia de Huesca. Galemys,12:15–23.
Barataud, M. 1996. The world of bats. Acoustic identification of French bats. Editions Sittelle. France. 47pp.
Barros, P. 2011. Contribución al conocimiento de la distribución de quirópteros en el norte y centro del Por tugal. Barbastella, 5(1): 19-31 DOI: http://dx.doi.org/10.14709/BarbJ.5.1.2012.04
Barthelmess, E. & Brooks, M. S. 2010. The influence of body-size and diet on road-kill trends in mammals. Biodiversity and Conservation, 19:1611–1629. DOI: https://dx.doi.org/10.1007/s10531-010-9791-3
Bat Conservation Trust 2012. Bats and Buildings. Bats and the Built Environment series. Bat Conservation Trust.
Beckmann, J. P., Clevenger, A. P., Huijser, M. P. & Hilty, J. A. 2010. Safe passages: highways, wildlife, and habitat connectivity. Washington: Island Press.
Berthinussen, A., Altringham, J. 2012a. Do Bat Gantries and Underpasses Help Bats Cross Roads Safely? PLoS ONE, 7(6): e38775. doi:10.1371/journal.pone.0038775. DOI: https://dx.doi.org/10.1371/journal.pone.0038775
Berthinussen, A. & Altringham, J. 2012b. The effect of a major road on bat activity and diversity. Journal of Applied Ecolology, 49:82–89. DOI: https://dx.doi.org/10.1111/j.1365-2664.2011.02068.x
Ceľuch, M., Ševčík, M. 2008. Road bridges as a roosts for noctules (Nyctalus noctula) and other bat species in Slovakia (Chiroptera: Vespertilionidae). Lynx, 39(1):47–54
Clevenger, A. P. & Waltho, N. 2000. Factors influencing the effectiveness of wildlife underpasses in Banff National Park, Alberta, Canada. Conservation Biology, 14:47–56. DOI: https://dx.doi.org/10.1046/j.1523-1739.2000.00099-085.x
Davidson-Watts, I., Walls, S. & Jones, G. 2006. Differential habitat selection by Pipistrellus pipistrellus and Pipistrellus pygmaeus identifies distinct conservation needs for cryptic species of echolocating bats. Biologic Conservation, 133:118-127. DOI: https://dx.doi.org/10.1016/j.biocon.2006.05.027
Pasos agrícolas inferiores de carreteras: su importancia para los murciélagos como refugio y lugar para cruzar la vía

30 Barbastella 7(1) 2014
Hawbaker, T, Radeloff, V, Hammer, R & Clayton, M. 2004. Road density and landscape pattern in relation to housing density, land ownership, land cover, and soils. Landscape Ecologic, 20:609-625. DOI: https://dx.doi.org/10.1007/s10980-004-5647-0
Highways Agency UK. 2008. Interim advice note 11608 nature conservation advice in relation to bats. London Highways Agency. 57pp.
Ibáñez, C., García-Mudarra, J. L., Ruedi, M., Stadelmann, B. & Juste, J. 2006. The Iberian contribution to cryptic diversity in European bats. Acta Chiropterologica, 8(2):277-297. DOI: https://dx.doi.org/10.3161/1733-5329(2006)8%5B277:TICTCD%5D2.0.CO;2
Juste, J., Bilgin, R., Muñoz, J. & Ibáñez, C. 2009. Mitochon drial DNA signatures at different spatial scales: from the effects of the Strait of Gibraltar to population structure in the meridional serotine bat (Eptesicus isabellinus). Heredity, 103:178-187. DOI: https://dx.doi.org/10.1038/hdy.2009.47
Kerth, G. & Melber, M. 2009. Species-specific barrier effects of a motorway on the habitat use of two threatened forest-living bat species. Biologic Conservation, 142:270-279. DOI: https://dx.doi.org/10.1016/j.biocon.2008.10.022
Lesiński, G 2008. Linear landscape elements and bat casualties on roads—an example. Annales Zoologici Fennici, 45:277-280. DOI: http://dx.doi.org/10.5735/086.045.0406
Lesiński, G., Sikora, A. & Olszewski, A. 2011. Bat casualties on a road crossing a mosaic landscape. European Journal of Wildlife Research, 57:217-223. DOI: https://dx.doi.org/10.1007/s10344-010-0414-9
Limpens, H. J., Kapteyn, K.1991. Bats, their behaviour and linear landscape elements. Myotis, 29:39-48.
Limpens, H. J., Twisk, .P & Veenbaas, G. 2005. Bats and road construction. Brochure about bats and the ways in which pratical measures can be taken to observe the legal duty of care for bats in planning, constructing, reconstructing and managing roads. Rijkswaterstaat, Dient Weg.en Waterbouwkunde, Delf, the Netherlands and the Vereiging voor Zoogdierkunde en Zoogdierbescherming, Arnhem.
Lisón, F,. Yelo, N. D., Haz, A. & Calvo, J. F. 2010. Contribución al conocimiento de la distribución de la fauna quiropterológica de la Región de Murcia. Galemys, 22(1):11-28.
Lisón, F., Aledo, E. & Calvo, J. F. 2011. Los murciélagos (Mammalia: Chiroptera) de la Región de Murcia (SE España): distribución y estado de conservación. Anales de Biología, 33:79-92.
Dietz, C. & Helversen, O. V. 2004. Illustrated identification key to the bats of Europe (http://www.uni-tuebingen.de/tierphys/Kontakt/mitarbeiter_seiten/dietz.htm).
Dietz, C., Helversen, O. V. & Nill, D. 2009. Bats of Britain, Europe & Northwest Africa. A & C Black Publishers Ltd.
Eigenbrod, F., Hecnar, S. & Fahrig, L. 2008. The relative effects of road traffic and forest cover on anuran populations. Biologic Conservation, 141:35–46. DOI: https://dx.doi.org/10.1016/j.biocon.2007.08.025
Entwistle, A. C., Racey, P. A. & Speakman, J. R. 2000. Social and population structure of a gleaning bat, Plecotus auritus. Journal of Zoology, 252:11–17. DOI: https://dx.doi.org/10.1111/j.1469-7998.2000.tb00815.x
Flaquer, C., Puig, X., Fábregas, E., Guixé, D., Torre, I., Ràfols, R. G., Páramo, F., Camprodon, J., Cumplido, J. M., RuízJarillo, R., Baucells, A. L., Freixas, L. & Arrizabalaga, A. 2010. Revisión y aportación de datos sobre quirópteros de Catalunya: Propuesta de Lista Roja. Galemys, 22 (1) 29-61.
Forman, R. T. T., Sperling, D., Bissonette, J. A., Clevenger, A. P., Cutshall, C. D., Dale, V. H., Fahrig, L., France, R., Goldman, C. R., Heanue, K., Jones, J. A., Swanson, F. J., Turrentine,T. & Winter, T. C. 2003. Road Ecology: Science and Solutions. Island Press, Washington.
Gaisler, J., Rehák, Z. & Bartonička, T. 2009. Bat casualties by road traffic (Brno-Vienna). Acta Theriol, 54:147-155. DOI: https://dx.doi.org/10.1007/BF03193170
Giulio, M, Holderegger, R, Tobias, S. 2009. Effects of habitat and landscape fragmentation on humans and biodiversity in densely populated landscapes. Journal of Environmental Management, 90:2959-2968. DOI: https://dx.doi.org/10.1016/j.jenvman.2009.05.002
Gloza-Rausch, F., Ipsen, A., Seebens, A., Göttsche, M., Panning, M., Drexler, J. F., Petersen, N., Annan, A., Grywna, K., Müller, M., Pfefferle, S. & Drosten, C. 2008. Detection and Prevalence Patterns of Group I Coronaviruses in Bats, Northern Germany. Emerging Infectious Diseases, 14:626-631. DOI: https://dx.doi.org/10.3201/eid1404.071439
Gomes, L., Grilo, C., Silva, C. & Mira, A. 2008. Identification methods and deterministic factors of owl roadkill hotspot locations in Mediterranean landscapes. Ecology Resources, 24:355-370. DOI: https://dx.doi.org/10.1007/s11284-008-0515-z
Gonzalez-Prieto, S., Villarino, A. & Fréan, M. 1993. Mortalidade de vertebrados por atropello en una carretera nacional del NO de España. Ecología, 7:375-389.
Paulo Barros

31Barbastella 7 (1) 2014
McKinney, M. L. 2008. Effects of urbanization on species richness: a review of plants and animals. Urban Ecosystems, 11:161-176. DOI: https://dx.doi.org/10.1007/s11252-007-0045-4
Medinas, D., Marque, J. T. & Mira, A. 2013. Assessing road effects on bats: the role of landscape, road features, and bat activity on road-kills. Ecological Research, 28(2)227-237. DOI: https://dx.doi.org/10.1007/s11284-012-1009-6
National Roads Authority 2006. Best practice guidelines for the conservation of bats in the planning of national road schemes. Dublin: National Roads Authority.
Racey, P. A. & Swift, S. M. 1985. Feeding ecology of Pipistrellus pipistrellus (Chiroptera: Vespertilionidae) during pregnancy and lactation I. Foraging behaviour. Journal of Animal Ecology, 54:205-215.
Rainho, A., Amorim, F., Marques, J. T., Alves, P. & Rebelo, H. 2011. Chave de identificação de vocalizações dos morcegos de Portugal Continental. Versão eletrónica (beta), Abril de 2011.
Russo, D. & Jones, G. 2002. Identification of twenty-two bat species (Mammalia: Chiroptera) from Italy by analysis of time-expanded recordings of echolocation calls. Journal of Zoology, 258:91-103. DOI: https://dx.doi.org/10.1017/S0952836902001231
Seiler, A. & Helldin, J. O. 2006. Mortality in wildlife due to transportation, In The ecology of transportation: managing mobility for the environment. eds J. Davenport, J.L. Davenport, pp. 165 -189. Springer, Dordrecht.
Senior, P., Butlin, R. K. & Altringham, J. D. 2005. Sex and segregation in temperate bats. Proceedings of The Royal Society Biological Sciences, 272:2467-2473. DOI: https://dx.doi.org/10.1098/rspb.2005.3237
Sétra 2008. Rapport Bibliographique, Routes et Chiroptères, État des Connaissances. Bagneux: Sétra.
Stawski, C. 2012. Capture and care of northern long-eared bats (Nyctophilus bifax) and seasonal changes in insect abundance. Australian Mammalogy, 34:245–250. DOI: http://dx.doi.org/10.1071/AM11043
Trocmé, M., Cahill, S., de Vries, J. G., Farrall, H., Folkeson, L. G., Hichks, C. & Peymen, J. (Eds) 2003. COST 341 – habitat fragmentation due to transportation infrastructure: the European review. Luxembourg: Office for Official Publications of the European Communities.
Trombulak, S. & Frissell, C. 2000. Review of ecological effects of roads on terrestrial and aquatic communities. Conservation Biology, 14:18-30. DOI: https://dx.doi.org/10.1046/j.1523-1739.2000.99084.x
Verboom, B. & Huitema, H. 1997. The importance of linear landscape elements for the pipistrelle (Pipistrellus pipistrellus) and the serotine bat (Eptesicus serotinus). Landscape Ecology, 12: 117-125. DOI: https://dx.doi.org/10.1007/BF02698211
Vintulis, V. & Šuba, J. 2010. Autumn swarming of the pond bat Myotis dasycneme at hibernation sites in Latvia. Estonian Journal of Ecology, 59(1):70-80. DOI: https://dx.doi.org/10.3176/eco.2010.1.06
Walters, C. L., Freeman, R., Collen, A., Dietz, C., Brock Fenton, M., Jones, G., Obrist, M. K., Puechmaille, S. J., Sattler, T., Siemers, B. M., Parsons, S. & Jones, K. E. 2012. A continental-scale tool for acoustic identification of European bats. Journal of Applied Ecology, 49(5):1064-1074. DOI: https://dx.doi.org/10.1111/j.1365-2664.2012.02182.x
Woltz, H., Gibbs, J. & Ducey, P. 2008. Road crossing structures for amphibians and reptiles: informing design through behavioral analysis. Biologic Conservation, 141:2745-2750. DOI: https://dx.doi.org/10.1016/j.biocon.2008.08.010
Pasos agrícolas inferiores de carreteras: su importancia para los murciélagos como refugio y lugar para cruzar la vía

32 Barbastella 7(1) 2014
Barbastella 7 (1) Open Access ISSN: 1576-9720 SECEMU www.secemu.org
Recolección de guano de murciélagos de Cabrera (Pipistrellus pygmaeus) por hormigas rojas (Formica rufa)
Juan Tomás Alcalde1*, Inmaculada Antón
1 Plaza Sabicas 5, 2ºB. 31015, Pamplona, Spain.
*Corresponding author e-mail: [email protected]
DOI: http://dx.doi.org/10.14709/BarbJ.7.1.2014.06
English title: Collecting bat guano of soprano pipistrelles (Pipistrellus pygmaeus) by red wood ants (Formica rufa)
Abstract: In June 1999, a colony of soprano pipistrelles (Pipistrellus pygmaeus Leach, 1825) was found in a dead white poplar (Populus alba L., 1753) in Lodosa, Navarre (Northern Spain). More than 717 bats roosted in the same tree. This tree was cut down, and the colony finally disappeared. In 2003, six bat-boxes (Schwegler, 1FF model) were placed in the same area by the regional authorities (Dept. of Environment, Gov. of Navarre) to provide alternative roosts for this species. In July 2008, five bat-boxes were occupied by new breeding colonies of soprano pipistrelles, ranging approximately from 20 to 40 individuals each one. This year, we observed that many red wood ants (Formica rufa L., 1761) collected the bat guano from the ground, below one bat-box, and they took it up the tree trunk, to their nest, located 2 m above the bat-box. Similar behavior has been mentioned by other researchers for some different ant species and other insects, but until now, always in caves. As red wood ants normally feed on other insects, we think that they could collect bat guano as a food resource, in an oppor-tunistic behavior. Thus, soprano pipistrelles have a positive effect on the ant colony.
Keywords: Guano, Formica rufa, bat boxes, Pipistrellus pygmaeus.
accepted: March 21, 2014
Recolección de guano de murciélagos de Cabrera (Pipistrellus pygmaeus) por hormigas rojas (Formica rufa)
En julio de 1999 se localizó una numerosa colonia reproductora de murciélagos de Cabrera, (Pipistrellus pygmaeus Leach, 1825) refugiada en un álamo (Populus alba L., 1753) viejo de Lodosa (Navarra, norte de España), a las orillas de río Ebro.
El árbol refugio fue talado en agosto de ese mismo año y la colonia, de más de 717 ejemplares, desapareció (Alcalde 2000). En 2003, el Departamento de Medio Ambiente del Gobierno de Navarra decidió la colocación de cajas-refugio en árboles de las cercanías, para compensar la pérdida del refugio original y favorecer la permanencia de la población de murciélagos de Cabrera en la zona. Se instalaron 6 cajas planas, del modelo 1FF de la empresa Schwegler. Al carecer de base, estos refugios dejan caer libremente el guano al suelo.
En julio de 2008, cinco cajas estaban ocupadas por pequeñas colonias reproductoras de entre 20 y 40 individuos
adultos aproximadamente. Durante una revisión llevada a cabo el 6 de septiembre del mismo año, se comprobó que todas las cajas seguían estando habitadas por grupos de murciélagos. Bajo todas ellas, a excepción de una, se apreciaba una patente mancha de guano, procedente de las agrupaciones situadas unos metros más arriba. Una observación más detallada en la zona limpia que quedaba debajo de la caja mencionada reveló la presencia de numerosas hormigas rojas (Formica rufa L., 1761) inspeccionando el terreno. Se pudo comprobar también que algunas hormigas recogían el guano fresco, recién caído (Fig. 1), y lo transportaban por el tronco del árbol hasta una grieta situada a 7 m de altura, 2 m por encima de la caja donde se encontraban los murciélagos (Fig. 2). Como consecuencia, el suelo se encontraba casi totalmente vacío de guano a pesar de que la caja contenía en ese momento un mínimo de 13 murciélagos de Cabrera.
Algunos autores mencionan comportamientos similares, aunque siempre referidos a especies cavernícolas como por

33Barbastella 7 (1) 2014
ejemplo: en España, Tinaut (2001) encontró una especie de hormiga (Hypoponera ragusai Emery, 1894), de distribución circunmediterránea, en tres cavidades andaluzas. Dicho autor observó las hormigas asociadas frecuentemente a depósitos de guano, aunque por el momento se desconoce si realmente se alimentan de éste o de otros organismos que viven en el mismo. Moulds (2006) observó hormigas cavernícolas australianas (Iridomyrmex purpureus Smith, 1858), recogiendo guano de murciélagos (Vespadelus findlaysoni Kitchener, Jones & Caputi, 1987) en el interior de cavidades y transportándolo a su nido. No obstante, hasta ahora no se había observado este comportamiento fuera de cavidades.
La hormiga roja es una especie común, que se distribuye por Europa y Asia occidental (Social Insects Specialist Group 1996) y es un insecto oportunista, que puede construir sus nidos tanto en el suelo como en los árboles. Es omnívoro, aunque principalmente carnívoro, devorando áfidos, dípteros y larvas de himenópteros que caza o recoge muertos; en
ciertas ocasiones puede alimentarse de las excreciones azucaradas de los áfidos y la savia de árboles (Skinner 1980).
Aunque hasta ahora no se le había observado aprovechando el guano de murciélagos, éste constituye un importante recurso alimenticio, utilizado por numerosos invertebrados que a su vez mantienen comunidades más diversas (Peck 1982, Ferreira & Martins 1999, Webster & Whitaker 2005, etc.) Se podría suponer por tanto que las hormigas rojas observadas recogían el guano de los murciélagos como alimento y lo transportaban a su nido situado en el álamo, al igual que también recogen los cadáveres de otros insectos muertos entre la vegetación. Teniendo en cuenta la elevada tasa metabólica de los murciélagos de Cabrera, y por tanto la gran cantidad de guano que produce una colonia, estos mamíferos podrían ofrecer un importante suministro de alimento a dichos invertebrados. La ausencia de otras observaciones similares en esta hormiga, cuyas poblaciones son abundantes y muy estudiadas, sugiere que este comportamiento pueda ser ocasional, fruto del oportunismo de estos insectos.
Por otro lado, los murciélagos de Cabrera se alimentan fundamentalmente de pequeños dípteros, mientras que los himenópteros constituyen únicamente entre el 2 y el 3% de sus presas (Barlow 1997, Bartonička et al. 2008). Ello permite deducir que en este caso, las hormigas rojas podrían resultar muy beneficiadas por los murciélagos de Cabrera, al disponer de un recurso alimenticio abundante y de fácil acceso, sin que ello entrañe un riesgo relevante.
ReferenciasAlcalde J. T. 2000. Desaparece una importante colonia
arborícola. Barbastella 1: 10-11.
Barlow K. E. 1997. The diets of two phonic types of the bat Pipistrellus pipistrellus y Britain. J. Zool. Lond. 243: 597-609. DOI: https://dx.doi.org/10.1111/j.1469-7998.1997.tb02804.x
Bartonička T., Řehák Z. & Andreas M. 2008. Diet composition and foraging activity of Pipistrellus pygmaeus in a floodplain forest. Biologia 63 (2): 1-7. DOI: https://dx.doi.org/10.2478/s11756-008-0034-y
Ferreira R. L., Martins R. P. 1999. Trophic structure and natural history of bat guano invertebrate communities, with special reference to Brazilian caves. Tropical Zoology 12: 231-252.
Moulds T. 2006. The first Australian record of subterranean guano-collecting ants. Helictite 39 (1): 3-4.
Peck S. B. 1982. A contribution to the knowledge of the invertebrate cave faunas of Venezuela: invertebrate faunas of tropical American caves, part 4. J. Speleol. 12: 75-81.
Skinner G. J. 1980. The feeding habits of the wood-ant, Formica rufa (Hymenoptera: Formicidae), in limestone woodland in North-West England. Journal of Animal Ecology 49: 417-433.
Juan Tomás Alcalde, Inmaculada Antón
Fig. 1 – Hormiga roja transportando el guano por el árbol hasta su nido.

34 Barbastella 7(1) 2014
Social Insects Specialist Group 1996. Formica rufa. In: IUCN 2013. IUCN Red List of Threatened Species. Version 2013.2. <www.iucnredlist.org>. Downloaded on 1 February 2014.
Tinaut T. 2001. Hypoponera ragusai (Emery, 1985) a cavernicolous ant new for the Iberian Peninsula (Hymenoptera, Formicidae). Graellsia 57 (1): 3-8.
Recolección de guano de murciélagos de Cabrera (Pipistrellus pygmaeus) por hormigas rojas (Formica rufa)
Fig. 2 – Hormiga roja (F. rufa) recogiendo guano de murciélagos de Cabrera (P. pygmaeus) del suelo.

35Barbastella 7 (1) 2014
la especie que nos ocupa no se han encontrado registros documentados, al menos en España, excepto diversos accidentes por choque con aerogeneradores (Camiña 2012, Sánchez et al., 2012).
En este trabajo se describen cinco sucesos similares ocurridos en la ciudad de Valencia en los que un número elevado de murciélagos de esta especie quedan atrapados en diversas estructuras de fachada en edificios altos. En dos de ellos se registra una alta mortalidad. Se discuten las causas que podrían haber causado dichos accidentes.
El primer caso se registró el 5 de octubre de 1995. A raíz de una llamada telefónica avisando del hallazgo de unos murciélagos el personal del centro de recuperación de fauna “La Granja” de El Saler (Valencia) se presentó en una vivienda situada en un 6º planta -coordenadas geografícas 39º28’40,34”N 0º21’26,41”W- y encontraron más de un
Barbastella 7 (1) Open Access ISSN: 1576-9720 SECEMU www.secemu.org
Eventos de mortalidad accidental de Murciélago rabudo Tadarida teniotis en edificios altos de la ciudad de Valencia (España)
Miguel Ángel Monsalve-Dolz1
1 Centros de Recuperación de Fauna-VAERSA-Servicio de Vida Silvestre. Conselleria de Infraestructuras, Territorio y Medio Ambiente. CRF “La Granja” de El Saler. Av Los Pinares 106, 46012, El Saler-Valencia.
Corresponding author e-mail: [email protected]
DOI: http://dx.doi.org/10.14709/BarbJ.7.1.2014.07
English title: Accidental mortality events of the European free-tailed bat Tadarida teniotis in high buildings in Valencia (Spain)
Abstract: Other than events caused by wind turbines, the accidental mortality of European free-tailed bats in Spain has never been reported in the literature. The discovery on five occasions of European free-tailed bats trapped in boxes on a small balcony in the city of Valencia is described here. In two cases, high mortality was recorded. The findings follow a similar pattern: all involve a large number of bats of both sexes, which are found dead in a box with an open top, always in the month of October.
Given the number of cases found over the years in a small area and the number of specimens involved, these mortality events could become a determining factor for some colonies. Keywords: Tadarida teniotis, mortality, boxes, Valencia
accepted: March 21, 2014
Miguel Ángel Monsalve-Dolz
Aunque el estatus de conservación del murciélago rabudo (Tadarida teniotis) es “de Preocupación Menor” en el ámbito mundial (Aulagnier et al 2008) en España la información poblacional es escasa, considerándose su población dispersa aunque no rara (Balmori 2007). En la Comunitat Valenciana no se dispone de información sobre la magnitud ni sobre el estado de las poblaciones (Monsalve 2012).
Al igual que el resto de las especies de murciélagos europeos, el murciélago rabudo es una especie longeva por lo que el conocimiento sobre causas de mortalidad es relevante para evaluar y entender su dinámica poblacional. Si los eventos de mortalidad afectan a un número elevado de individuos y se repiten en el tiempo pueden provocar una regresión de las poblaciones. En general son escasos en la literatura los casos de mortalidad accidental en murciélagos (Manville 1963) y más en los que se vean afectados un grupo numeroso de individuos (p.ej. Jung & Slough 2005). Sobre

36 Barbastella 7(1) 2014
centenar de ejemplares de murciélagos rabudos atrapados en un pequeño macetero vacío (sin contenido) con forma cúbica, orientado al oeste y con dimensiones 40 x 40 x 40 cm. La persona que dio el aviso había estado escuchando unos chirridos provenientes del exterior de la ventana desde hacía unos días. En el momento de la visita casi todos los ejemplares ya habían muerto y se encontraban en avanzado estado de descomposición. Sólo 19 permanecían vivos, y de ellos 2 estaban muy debilitados y murieron a las pocas horas. Los otros 17 fueron inmediatamente liberados. Dado el escaso espacio existente dentro del macetero la causa de la muerte pudo ser el aplastamiento de unos ejemplares sobre otros.
Un año después, el 8 de octubre de 1996, se registró en el mismo edificio el segundo caso, prácticamente idéntico al anterior. Esta vez se rescataron 2 ejemplares vivos que permanecían junto a más de un centenar muertos en un macetero de las mismas características, situación y orientación que el anterior, pero a una altura de un 10º piso (ver figura 1). A los pocos días, el 23 de octubre de 1996, se encontraron 43 ejemplares vivos en el mismo macetero donde se registró la mortalidad el año anterior, en la 6ªplanta. Todos los ejemplares fueron liberados el mismo día.
Estos tres sucesos son muy similares a otro ocurrido en octubre de 2006, a unos 500 m del edificio donde se registraron los accidentes en 1995 y 1996. Esta vez se trató de 56 murciélagos rabudos que habían quedado atrapados en un macetero de cemento (coordenadas geografícas 39º28’50,75”N 0º21’44,71”W). Todos los ejemplares fueron
Fig. 1 – Macetero donde fueron encontrados más de 100 murciélagos rabudos muertos en 1996.
Fig. 2 – Ejemplares encontrados en octubre de 2013 en un balcón de pequeñas dimensiones.
Eventos de mortalidad accidental de Murciélago rabudo Tadarida teniotis en edificios altos de la ciudad de Valencia (España)

37Barbastella 7 (1) 2014
rescatados tras permanecer allí varios días. En este caso no se registró con exactitud la altura de la planta donde se situaba el macetero pero se trataba de un piso alto. Aunque el tamaño de este macetero era mayor que los primeros con una longitud superior a un metro, su anchura y profundidad fueron similares a los anteriores, aproximadamente unos 40 cm. Todos los ejemplares se sexaron (25 machos y 31 hembras), fueron marcados en antebrazo con anilla homologada y liberados en una zona cercana.
El último caso fue observado el pasado 14 de octubre de 2013. Este día fueron hallados un total de 56 murciélagos rabudos atrapados en un pequeño balcón de un 6º piso (ver figura 2) en un edificio situado más al sur -coordenadas geografícas 39º27’52,79”N 0º22’22,18”W-. Las dimensiones del balcón eran de 100 (ancho) x 200 (largo) cm y la altura de la valla de unos 110 cm. En la visita se hallaron un total
de 43 machos y 13 hembras. Ningún ejemplar iba marcado con las anillas colocadas en 2006. Todos los ejemplares se encontraban en el suelo del balcón y la mayoría permanecían escondidos en los únicos refugios disponibles, dentro de unas zapatillas de deporte y detrás de una maceta. Al parecer la barandilla del balcón con paredes de cemento lisas representaba una barrera infranqueable que los murciélagos no podían superar.
En los dos primeros casos descritos probablemente se tardó más en avisar al centro de recuperación de fauna; ese factor pudo provocar que la cantidad de ejemplares afectados fuera mayor y se encontraran ejemplares con buen aspecto general junto a otros con síntomas de inanición y ejemplares muertos ya en descomposición. En ambos se registró alta mortalidad de ejemplares (Tabla 1). Por el contrario, a partir del tercer caso, con menos ejemplares afectados, el tiempo
Fig. 3 – Localización de los lugares donde se encontraron los murciélagos accidentados en la ciudad de Valencia. Se señala el año de los hallazgos.
Tabla. 1 – Fecha, lugar y número de ejemplares afectados en los cinco eventos de mortalidad.
Fecha evento
Estructura donde quedan atrapados
Altura (piso del edificio)
nº ejemplares encontrados
vivos
nº ejemplares encontrados muertos
sex-ratio (relación machos/hembras)
Oct-1995 macetero 6º 19 >100 -
Oct-1996 macetero 10º 2 >100 -
Oct-1996 macetero* 6º 43 0 -
Oct-2006 macetero - 56 0 0,81 (n=56)
Oct-2013 pequeño balcón 6º 56 0 3,31 (n=56)
*mismo macetero que en 1995
Miguel Ángel Monsalve-Dolz

38 Barbastella 7(1) 2014
transcurrido desde el primer ejemplar accidentado hasta el rescate fue menor (estimado entre 4 y 5 días).
Todos los hallazgos siguen un patrón similar, se
ven afectados varias decenas de murciélagos, aparecen numerosos ejemplares de ambos sexos, quedan atrapados en la fachada de un edificio a gran altura, el lugar donde quedan atrapados es una estructura abierta por la parte superior pero sin salida en suelo y paredes, y por último, todos los sucesos se han producido en el mes de octubre.
Al parecer en todos los sucesos los murciélagos fueron quedando atrapados de forma progresiva, seguramente durante varias noches. La persona que se encontró los murciélagos accidentados en 2013 observó como cada día había algunos más que el anterior. Por otra parte, en ningún caso después de la retirada de los ejemplares atrapados hubo nuevos ejemplares accidentados, lo que apunta a que los accidentes estuvieron relacionados con algún comportamiento social que incluía atracción de unos ejemplares hacia otros. Esto viene reforzado por el hecho de que en el momento de la liberación de los murciélagos rescatados alguno de ellos realizó vuelos bajos aproximándose a pocos centímetros de la caja donde permanecía el resto de los individuos, los cuales emitían fuertes sonidos sociales desde el interior de la misma.
La época en la que se producen los accidentes coincide con el celo de la especie, cuando los machos y hembras se agrupan formando harenes, sin embargo las agrupaciones en esta época suelen ser pequeñas. El número máximo de ejemplares controlados en dos refugios de Valladolid a lo largo de tres temporadas fue de 17 (Balmori 2003). El tamaño de las agrupaciones descritas en el presente artículo son mayores y, aunque siempre se observan machos y hembras, la sex-ratio fue diferente en los dos eventos en los que se registró.
En el mismo estudio se observó una agrupación de juveniles muy ruidosa en uno de los refugios durante el mes de octubre (Balmori 2003). En los casos aquí descritos no se vio ningún ejemplar con el característico pelaje claro de los juveniles, no obstante en una exploración de los ejemplares encontrados en 2013 tampoco se observó que los machos presentaran testículos desarrollados, por lo que no hay que descartar la posibilidad de que se trate de individuos jóvenes, nacidos en la primavera, cuya inexperiencia podría explicar, al menos en parte, estos eventos de mortalidad.
Dado el número de casos encontrados a lo largo de los años en un área limitada y la cantidad de ejemplares afectados podría tratarse de un factor de mortalidad determinante para algunas colonias. Sería recomendable recoger mayor información en el futuro y comprobar cómo se producen estos accidentes.
AgradecimientosDavid Almenar y Cati Gerique me acompañaron en
algunas visitas realizadas a los edificios afectados. Además, Cati Gerique fue la veterinaria encargada del CRF La Granja cuando ocurrieron los primeros accidentes. Ella supervisó y determinó la causa de muerte de los cadáveres hallados en 1995 y 1996.
ReferenciasAulagnier, S., Paunovic, M., Karatas, A., Palmeirim, J.,
Hutson, A.M., Spitzenberger, F., Juste, J. & Benda, P. 2008. Tadarida teniotis. En: UICN 2013. Lista Roja de Especies Amenazadas. Versión 2013.2. < www.iucnredlist.org >.
Balmori, A. 2003. Avances en el conocimiento de la biología y organización social del murciélago rabudo (Tadarida teniotis). Galemys 15 (nºespecial), 2003. Pp: 37-53.
Balmori A. 2007 Tadarida teniotis (Rafinesque, 1814) Pp: 267-271. En: L.J. Palomo, J. Gisbert & J.C. Blanco (eds.). Atlas y Libro Rojo de los Mamíferos Terrestres de España. Dirección General para la Biodiversidad. SECEM-SECEMU, Madrid.
Camiña A. 2012. Bat Fatalities at Wind Farms in Northern Spain - Lessons to be Learned. Acta Chiropterologica 14(1):205-212. DOI: http://dx.doi.org/10.3161/150811012X654402
Jung, T.S., & Slough B.G. 2005. Mortality of Little Brown Bats, Myotis lucifugus, in a rodent trap in the boreal forest. Canadian Field-Naturalist 119(4): 589-590.
Manville, R. H. 1963. Accidental mortality in bats. Mammalia. 27: 361-366. DOI: https://dx.doi.org//10.1515/mamm.1963.27.3.361
Monsalve M. A. 2012. Tadarida teniotis Pp: 248-250. En: Jiménez J., Monsalve M.A. & Raga J.A. (Eds) 2012. Mamíferos de la Comunitat Valenciana. Colección Biodiversidad, 19. Conselleria d’Infraestructures, Territori i Medi Ambient. Generalitat Valenciana, Valencia.
Sánchez S, Berrío-Martínez, J. & Ibáñez, C. 2012. Patrones de mortalidad de murciélagos en Parques Eólicos del Sur de la Península Ibérica. Póster, I Congreso Ibérico sobre Energía Eólica y Conservación de la Fauna, Jerez, Cádiz, enero 2012.
Eventos de mortalidad accidental de Murciélago rabudo Tadarida teniotis en edificios altos de la ciudad de Valencia (España)

39Barbastella 7 (1) 2014
Barbastella 7 (1) Open Access ISSN: 1576-9720 SECEMU www.secemu.org
Could overheating turn bat boxes into death traps?
Carles Flaquer 1, Xavier Puig 2, Adrià López-Baucells 1, Ignasi Torre 1, Lídia Freixas 1, Maria Mas 1, Xavier Porres 3, Antoni Arrizabalaga 1
1 Granollers Museum of Natural Sciences, Av. Francesc Macià 51, Granollers 08490. Catalonia. 2 Galanthus Association, Carretera de Juià 46, 17460 Celrà. Catalonia.3 Parc Natural del Delta de l’Ebre, Departament de Medi Ambient i Habitatge, Generalitat de Catalunya, Av. Catalunya 64, 43580 Deltebre. Catalonia.
Corresponding author e-mail: [email protected]
DOI: http://dx.doi.org/10.14709/BarbJ.7.1.2014.08
Spanish title: El calentamiento excesivo puede convertir las cajas-refugio en trampas letales?
Abstract: The placing of bat boxes has become a very popular method of monitoring bats and also an educa-tional tool used by conservation groups and government departments alike to explain the value of bats to human societies. As a result, thousands of bat boxes have been sold or made by bat enthusiasts throughout Europe in past decades. However, very few papers have ever analyzed the importance of temperatures inside bat boxes in light of the expected effects of climate change. This paper is a first attempt to explore the possible effects of extreme heat on bats roosting in boxes.
Based on our experience in monitoring bat boxes since 1999, our research team decided to place 10 data loggers in new bat boxes randomly distributed amongst our established box stations to monitor the maximum recorded temperatures in boxes. We established that there was a risk of overheating when temperatures rose above 40ºC. Throughout the whole summer we recorded box temperatures every hour (17,560 readings). Overheating epi-sodes occurred in six of ten boxes and in 2% of all samples (429 hours). Furthermore, in July 2013 we observed an overheating event in situ when 22 bats fell from a bat box while they were trying to emerge in the evening. Most European bat boxes models are small, have no ventilation and are usually located in places that are exposed to the sun for several hours a day. However, it has been assumed that bats avoid overheating in bat boxes by roost-switching. Nevertheless, in July 2013 we observed bats suffering from overheating in a wetlands where there were many alternative roosting sites (trees and other boxes).
In the Mediterranean area, average summer temperatures are rising annually and we believe it is important to alert bat conservationists to the possibility of overheating events in bat boxes. This preliminary study has indi-cated that very high temperatures can be reached inside boxes and it is not clear to what extent bats will be able to adapt to these events. Future research should address this issue in the coming years. Keywords: bats, boxes, monitoring, roosts, overheating, climate change, Pipistrellus pygmaeus, Mediterranean, Europe.
accepted: March 31, 2014
Carles Flaquer, Xavier Puig, Adrià López-Baucells, Ignasi Torre, Lídia Freixas, Maria Mas, Xavier Porres, Antoni Arrizabalaga

40 Barbastella 7(1) 2014
State of the artBat boxes are a popular management tools that can
mitigate the loss of roost sites. They are an effective way of increasing roost availability and represent an especially interesting occasion for studying bat distribution and ecology (Racey and Kunz 1998). Bat boxes may play an important role during key phases of bat life cycles and provide a good alternative to natural or artificial roost sites for maternity colonies excluded from buildings (Lourenco and Palmeirim 2004; Flaquer, Torre et al. 2006)
Over the last decade, bat boxes have also been used as an awareness-raising tool by conservationists and thousands have been sold by specialized companies or freely distributed by NGOs and governments. As a result of these educational and/or conservation strategies, many bat boxes have been distributed in the Mediterranean region (Agnelli, Maltagliati et al. 2011). In Europe, bat boxes are
mainly small and have few or no compartments (p. ex. Bat Conservation Trust or Schwegler bat box models). In order to improve breeding conditions for maternity colonies, bat boxes need to be placed on southern or eastern exposures (Kerth, Weissmann et al. 2001; Flaquer, Torre et al. 2006; Baranauskas 2013) to increase the sun radiation incidence, whereas western exposures can reach overheating during the mid-day hours in hottest climates (Brittingham and Williams 2000; Flaquer, Torre et al. 2007). Furthermore, some authors suggested using dark-coloured boxes (painted in black) to increase heat absorption (Lourenco and Palmeirim 2004). The correct assessment of both materials and exposures used for bat boxes are also considered to be key factors by according to Bat International Criteria for successful Bat Houses. Differnt colours are recommended regarding climate areas to achive favoralble conditions for maternity colonies in summer, between 27 and 38ºC internal bat box, (www.batcon.org/pdfs/bathouses/bathousecriteria.pdf). Bats’ juvenile growth is dependent on roost temperature during
Fig. 1 – Distribution of bat boxes with data loggers in Catalonia in summer 2013.
Could overheating turn bat boxes into death traps?

41Barbastella 7 (1) 2014
both gestation and lactation periods however, few papers to date have ever analyzed how temperatures in bat boxes influence bat breeding strategies (Lourenço and Palmeirim 2004; Bartonicka and Rehak 2007). Those studies that have been conducted consider around 40ºC to be the maximum temperature tolerated by bats in bat boxes and suggest that bats will switch their roost sites when temperatures are too high (Feyerabend and Simon 2000; Bartonicka and Rehak 2007).
In line with these ideas, in Australia overheating (extreme events over 43ºC) have been found to be the cause of the decimation of flying-fox populations, the most affected individuals being basically lactating females and young bats (Welbergen, Klose et al. 2008). Temperature extremes are inherent expressions of climate change due to their direct link with the increased heat content of the atmosphere (IPCC 2007)
The effect of extreme heat events on microchiroptera in temperate zones is still unknown and their capacity as bioindicators still needs to be properly studied (Flaquer and Puig-Montserrat 2012). During the past two years some exceptional events have alerted us to possible overheating die-offs in the soprano pipistrelle bat Pipistrellus pygmaeus (Leach 1825) breeding in bat boxes. This paper discusses this important subject that could become problematic due to the current trend for rising temperatures.
Collecting evidenceField observation
Our research team has been surveying bat boxes in Catalonia in both natural areas and urban environments since 1999. The climate throughout the region is Mediterranean (mean annual temperatures of ca. 16ºC; average precipitation of 500–700 mm/year). During the 10 years of this monitoring program, we have observed on occasions that some bat breeding colonies remain outside bat boxes and that some
individuals sweat considerably; however, we neither found nor expected any mortality to be linked to this behaviour. No specific effort has ever been carried out to evaluate the effect of high temperatures on bats in artificial roost sites (either on a daily or annual basis).
In summer 2008 almost the whole of a breeding colony of Pipistrellus pygmaeus (42 individuals) were found dead inside bat boxes located in a wetland without any obvious explanation. In the same location but in a different bat box in July 2013 we observed in situ several bats fall out of a bat box while they were trying to emerge. This event happened in the afternoon at 17:00h in a wooden bat box located on a building (UTM: 42.218726, 3.100475), facing west and surrounded by trees (which reduced sun exposure at midday). Of the bats, 22 were collected and sent to a rescue centre where they were rehydrated and released at night (two of them subsequently died).
Small-scale experimentIn 2013 we decided to place 10 data loggers (iButton
DS1922L) inside new small wooden bat boxes (n=9) without compartments (100 x 190 x 200 mm) (Flaquer, Torre et al. 2006) and inside a single Schwegler Bat Box model 1FF. All bat boxes were randomly deployed in eight locations in Catalonia close to bat-box monitoring areas (Fig. 1). Bat boxes were situated from sea level (5 m.a.s.l.) to 990m, and facing to favorable exposures: South, East, and West (Berthinussen, Richardson et al. 2008). Data loggers were programmed to record temperatures hourly every day from July to mid-September 2013, which gave a total of 17,560 temperature readings. Data loggers recorded temperatures on average along 54.1 days ± 19.9 (SD), ranging between 24 and 77 days.
Temperatures recorded in the Schwegler bat box were compared with the wooden bat box under similar circumstances; in addition, data from two wooden bat boxes – one placed on a pole and the other on a wall – at same site
Fig. 2 – Distribution of overheating episodes (sample per hour) recorded at all data loggers (n=10) and the time at which these episodes occurred.
Carles Flaquer, Xavier Puig, Adrià López-Baucells, Ignasi Torre, Lídia Freixas, Maria Mas, Xavier Porres, Antoni Arrizabalaga

42 Barbastella 7(1) 2014
were compared. Since temperature data were non normally distributed, we used the Wilcoxon matched pairs test.
Some clues and preliminary results
60% of all bat boxes recorded temperatures above 40ºC at least once during the experiment (Table 1). Even at high altitudes (990 m a.s.l.) temperatures inside bat boxes rose to 47ºC and overheating episodes occurred from 10.00 to 20.00 h (Fig. 2). Overheating episodes were only observed in bat boxes placed on walls, whereas none of the two bat boxes placed on poles experienced overheating. When the box was placed on a wall, maximum temperatures were 4ºC higher than on a pole (Fig. 3). The Wilcoxon matched pairs test confirmed that Schwegler bat box recorded higher temperatures than Wooden bat box under the same sun exposure situation (Z = 17.27, p < 0.0001, n = 1728). Mean temperatures were 1.5ºC higher in the former box (X = 28.25ºC ± 7.73, range 16.5-54.5ºC, Coefficient of variation CV = 27.3%, and X = 26.83ºC ± 5.05, range 16.5-42.0ºC, CV = 18.8%, respectively). In the Schwegler Bat Box maximum temperatures were 12ºC higher than the wooden boxes under similar situation. Schwegler bat box recorded overheating episodes during 48 days (199 temperature records), representing the 62.3% of the study period, but the wooden bat box only recorded overheating episodes on six days (7.6% of the study period). Air temperature recorded was similar for both bat boxes between 0:00 and 14:00 hours, and differed between 16:00-22:00 hours in the hottest summer months (July and August, Fig. 4). During the first two weeks of September, no significant temperature differences were recorded between both bat boxes.
Could overheating turn bat boxes into death traps?
Tabl
e 1
– D
ata
logg
ers a
ccor
ding
to lo
catio
n an
d re
cord
ings
. The
fina
l col
umn
show
s the
num
ber o
f sam
ples
(hou
rs) d
urin
g th
e su
rvey
in w
hich
the
data
logg
er re
cord
ed te
mpe
ratu
res o
f 40º
C o
r ove
r.
Loca
tion
Alti
tude
(m
snm
)M
odel
Supo
rt O
rient
.Av
erag
eM
àxM
ín>4
0ºC
Day
s O
verh
eatin
gR
ecor
ded
Day
sFr
eq
Are
nys d
e M
unt
121
Schw
egle
r B
at B
ox 1
FFB
uild
ing
SW28
.29
54.5
16.5
199
4877
62.3
4A
reny
s de
Mun
t12
1W
oode
n ba
t bo
xB
uild
ing
SW26
.85
4216
.517
677
7.79
Les
Fran
ques
es23
2W
oode
n ba
t bo
xPo
leW
24.7
937
.514
00
720.
00Sa
nt Ju
lià
de C
.99
0W
oode
n ba
t bo
xB
uild
ing
S23
.347
.510
5122
6036
.67
Del
ta d
e l’E
bre
13W
oode
n ba
t bo
xB
uild
ing
W27
.82
4218
116
3815
.79
Del
ta d
e l’E
bre
13W
oode
n ba
t bo
xPo
leS
26.0
538
17.5
00
380.
00
Bar
celo
na13
Woo
den
bat
box
Bui
ldin
gE
27.7
3720
00
340.
00
Ale
lla92
Woo
den
bat
box
Bui
ldin
gS
26.8
438
180
039
0.00
Aig
uam
olls
de
l’Em
pord
à5
Woo
den
bat
box
Bui
ldin
gS
29.0
845
.513
5018
6030
.00
Espo
lla12
2W
oode
n ba
t bo
xB
uild
ing
S27
.87
45.5
14.5
9727
7635
.53
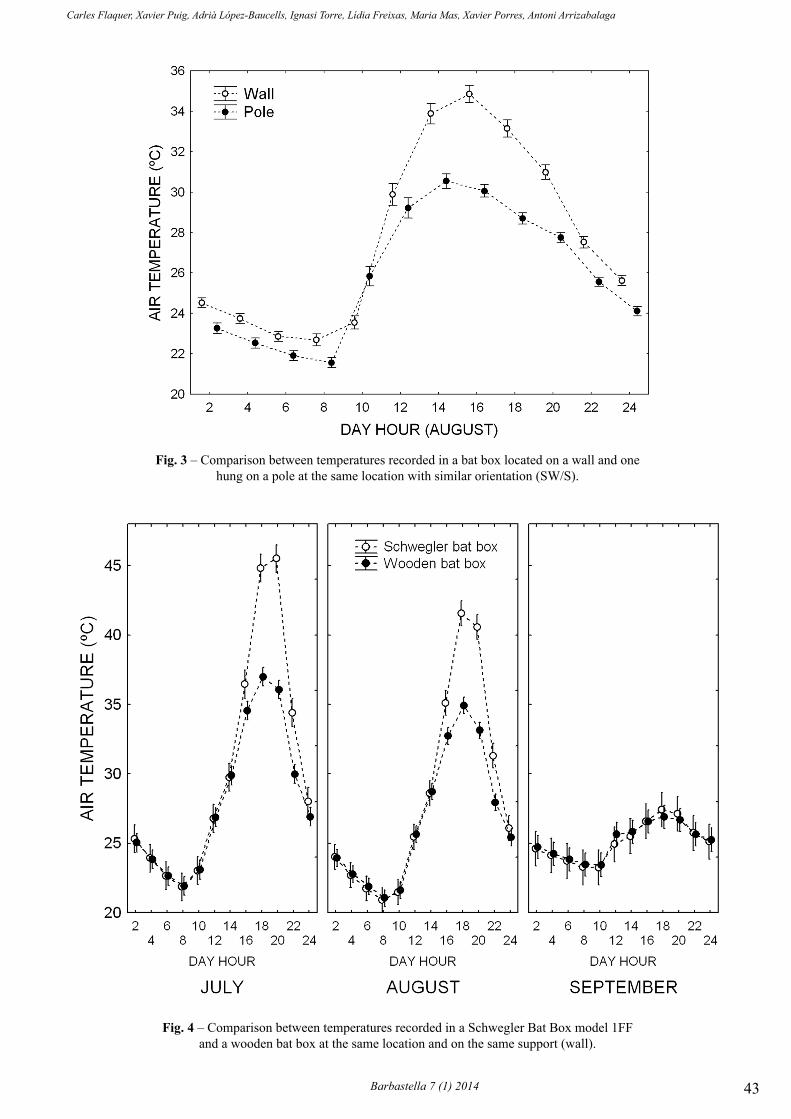
43Barbastella 7 (1) 2014
Fig. 4 – Comparison between temperatures recorded in a Schwegler Bat Box model 1FF and a wooden bat box at the same location and on the same support (wall).
Carles Flaquer, Xavier Puig, Adrià López-Baucells, Ignasi Torre, Lídia Freixas, Maria Mas, Xavier Porres, Antoni Arrizabalaga
Fig. 3 – Comparison between temperatures recorded in a bat box located on a wall and one hung on a pole at the same location with similar orientation (SW/S).

44 Barbastella 7(1) 2014
Discussion and awarenessAs bat conservationists and researchers we have always
considered bat boxes to be an excellent tool for both management and education. According to the bibliography, boxes should be placed from south-east to south-west situations that are well exposed to the sun (http://www.bats.org.uk/pages/bat_boxes.html; (Berthinussen, Richardson et al. 2008), and some authors encourage using dark colours to increase heat absortion (Lourenco and Palmeirim 2004). The models we have been using are inspired and personally designed according to the available information from Europe, and are mostly small and without ventilation. The bat boxes on sale in the Mediterranean are likewise essentially small without any ventilation and, in some cases, are painted black, like the Schwegler bat box.
In the Mediterranean area, average summer temperatures are rising year by year (Fig. 5) and, although there is no doubt that extreme heat events have increased in frequency in the past decade, their potential negative consequences are still unknown. (Welbergen, Klose et al. 2008).
According to (Bartonicka and Rehak 2007), “we cannot assume that box overheating is a possible risk. Box overheating may not be so dangerous if sufficient supplies of alternative roosts, mainly tree cavities, are available in the vicinity”. Yet, we have directly observed an overheating event in a bat box that caused fatalites. It should be taken into account that this episode occurred in a wetland (guaranteed high humidity) and that there were several other bat boxes in the area, as well as trees and buildings. In other Mediterranean wetland (Ebro delta), P.pygmaeus mainly selected bat boxes exposed to the East than to the West (Flaquer et al. 2006).
Fig. 5 – Anomaly temperatures (ºC) recorded in Barcelona from 1780 to 2011 compared with average temperatures. Over the last 30 years almost all these anomalies correspond to hotter events.
Fig. 6 – Bat-box overheating event witnessed in the Aiguamolls de l’Empordà Natural Park in July 2013. All the bats where collected from the ground and then rehydrated..
Could overheating turn bat boxes into death traps?

45Barbastella 7 (1) 2014
Some preliminary records with data loggers in this area showed that bat boxes exposed to the west recorded higher temperatures during mid-day hours, with several episodes of overheating (Flaquer, Torre et al. 2007).
Furthermore, the use of different materials to build up the bat boxes need to be properly examined. Our results point out that bat boxes painted on dark colours and made of woodcrete are extremely hot and the range of temperatures recorded are more variable than those recorded in bat boxes made on wood. The value of the former bat boxes in the conservation of bat populations in Mediterranean habitats can be questioned, despite these are amongst the most selled models in Catalonia.
There are thus a number of unanswered questions that must be addressed and properly studied:
1. How dangerous might box overheating be in light of the trend towards increased temperatures occurring as a result of climate change?
2. Can we be sure that bats use the strategy of roost-switching when temperatures are rising rapidly?
3. Can the models of bat boxes available in Europe properly thermoregulate and so avoid overheating episodes above 40ºC?
4. Should we change recommendations like “... exposed to sunlight for as much of the day as possible to increase bat box internal temperature” (Bat Conservation Trust recommendation to the best place to position the box to general public)?
For the time being not enough data are available to provide answers to these questions, although, when we observed bats falling out of a bat box due to overheating, we realized that our suspicions were well founded (Fig. 6). The results of our tests with data loggers are even more worrying as they show that bat boxes located in Mediterranean mountain areas can reach high temperatures if they are located on south-facing buildings (Table. 1).
We hope that this paper will encourage bat workers and researchers to collect as much information on the subject as possible over the coming years. If necessary, changes in bat-box management protocols may have to be implemented.
AcknowledgementsThis Project received financial support from the Catalan
Office for Climate Change, Diputació de Barcelona, Diputació de Girona, Generalitat de Catalunya and the Granollers Natural History Museum. We are especially grateful to the Ebro Delta and Aiguamolls de l’Empordà natural parks, to l’Albera Natural Area of National Interest, and to the Collserola, Serralada de Marina and Serralada Litoral parks. We are particularly indebted to the support provided by B. Borràs, G. Borràs, L. Cabañeros, T. Curcó, J. Justafré, R. Loire, F. Llimona, B. Minobis, I. Pac, S. Romero, S. Samitier and F. Vidal.
Finally, we would like to make special mention of Dr. Danilo Russo, who has been acting as our mentor during this ongoing, formative period of research.
ReferencesAgnelli, P., Maltagliati, G., Ducci, L., Cannicci, S. 2011.
Artificial roosts for Bats: Education and Research. The “Be a bat’s friend. Project of the Natural History Museum of the University of Florence. Hystrix-Italian Journal of Mammalogy 22(1): 215-223. DOI: https://dx.doi.org/10.4404/hystrix-22.1-4540
Baranauskas, K. 2013. Diversity and Abundance of Bats (Chiroptera) found in Bat Boxes in East Lithuania. Acta Zoologica Lituanica 20(1): 39-44. DOI: https://dx.doi,org/10.2478/v10043-010-0005-8
Bartonicka, T., Rehak, Z. 2007. Influence of the microclimate of bat boxes on their occupation by the soprano pipistrelle Pipistrellus pygmaeus: possible cause of roost switching.” Acta Chiropterologica 9(2): 517-526. DOI: http://dx.doi.org/10.3161/1733-5329(2007)9[517:IOTMOB]2.0.CO;2
Berthinussen, A., Richardson, O. C., Altringham, J. D. 2008. Bat Conservation. Global evidence for the effects of interventions. Synopses of Conservation Evidence. London, Pelagic Publishing. Volume 5.
Brittingham, M. C., Williams, L. M. 2000. Bat boxes as alternative roosts for displaced bat maternity colonies. Wildlife Society Bulletin 28(1): 197-207.
Feyerabend, F., Simonm, M. 2000. Use of roostsand roost switching in a summer colony of 45 kHz phonic type pipistrelle bats (Pipistrellus pipistrellus Schreber,1774). Myotis 38: 51-59.
Flaquer, C., Puig-Montserrat, X. 2012. Proceedings of the International Symposium on the Importance of Bats as Bioindicators. Granollers, Granollers Museum of Natural Sciences Edicions.
Flaquer, C., I. Torre, Arrizabalaga, A. 2007. Selección de refugios, gestión forestal y conservación de los quirópteros forestales. In: Conservación de la biodiversidad, fauna vertebrada y gestión forestal. J. Camprodon and E. Plana Eds. Barcelona, Universitat de Barcelona: 469-488.
Flaquer, C., Torre, I., Ruiz-Jarillo, R. 2006. The value of bat-boxes in the conservation of Pipistrellus pygmaeus in wetland rice paddies. Biological Conservation 128(2): 223-230. DOI: https://dx.doi.org/10.1016/j.biocon.2005.09.030
IPCC (2007). Climate Change 2007: The hisical science basis. Contribution of Working group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge Cambridge University Press.
Carles Flaquer, Xavier Puig, Adrià López-Baucells, Ignasi Torre, Lídia Freixas, Maria Mas, Xavier Porres, Antoni Arrizabalaga

46 Barbastella 7(1) 2014
Kerth, G., Weissmann, K., Konig, B. 2001. Day roost selection in female Bechstein’s bats (Myotis bechsteinii): a field experiment to determine the influence of roost temperature. Oecologia 126(1): 1-9. DOI: https://dx.doi.org/10.1007/s004420000489
Lourenço, S. I., Palmeirim, J. M. 2004. Influence of temperature in roost selection by Pipistrellus pygmaeus (Chiroptera): relevance for the design of bat boxes. Biological Conservation 119(2): 237-243. DOI: https://dx.doi.org/10.1016/j.biocon.2003.11.006
Racey, P., Kunz, T. H. 1998. Bat biology and conservation. Washington, Smithsonian Institution Press.
Welbergen, J., Klose, S., Markus, N., Eby, P. 2008. Climate change and the effects of temperature extremes on Australian flying-foxes. Proceedings of the Royal Society of Biological Sciences 275: 419-425. DOI: https://dx.doi.org/10.1098/rspb.2007.1385
Could overheating turn bat boxes into death traps?
