bacterias eucariotas - bioinfo.hpcf.upr.edubioinfo.hpcf.upr.edu/biol3705/2-diversidad...
TRANSCRIPT
11
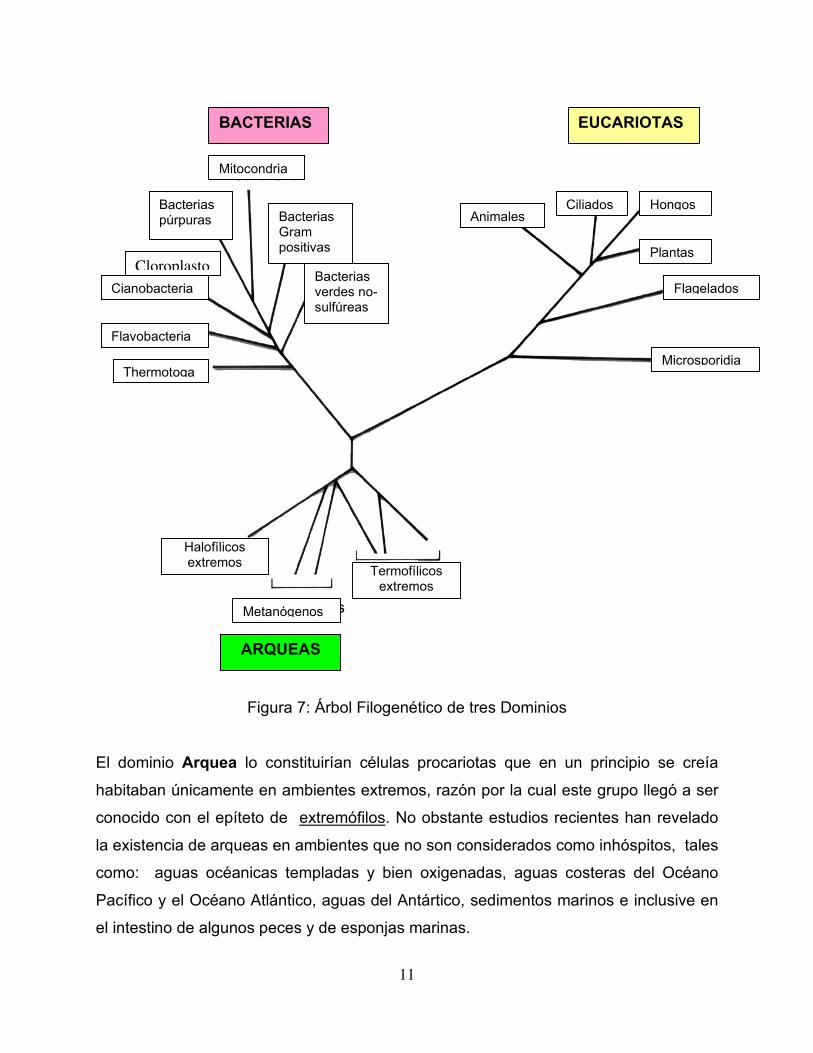
Figura 7: Árbol Filogenético de tres Dominios
El dominio Arquea lo constituirían células procariotas que en un principio se creía
habitaban únicamente en ambientes extremos, razón por la cual este grupo llegó a ser
conocido con el epíteto de extremófilos. No obstante estudios recientes han revelado
la existencia de arqueas en ambientes que no son considerados como inhóspitos, tales
como: aguas océanicas templadas y bien oxigenadas, aguas costeras del Océano
Pacífico y el Océano Atlántico, aguas del Antártico, sedimentos marinos e inclusive en
el intestino de algunos peces y de esponjas marinas.
Hongos
Plantas
Flagelados
Microsporidia
Animales Ciliados
Thermotoga
Flavobacteria
Cianobacteria Cloroplasto
Bacterias púrpuras
Mitocondria
BACTERIAS EUCARIOTAS
Bacterias verdes no-sulfúreas
Bacterias Gram positivas
ARQUEAS
Termofílicos extremos
Halofílicos extremos
Metanógenos

12
Un segundo dominio denominado Bacteria estaría integrado por microorganismos
procariotas que habitan mayormente en ambientes que también son colonizados por
los organismos eucariotas. Los individuos que integran los dominios Arquea y Bacteria
son esencialmente organismos unicelulares que se reproducen asexualmente. Es
conveniente acentuar que hasta el momento no se ha encontrado especie alguna de
procariotas que se reproduzca sexualmente, aunque lleven a cabo procesos de
recombinación genética.
El tercer dominio llamado Eucarya estaría integrado por todos los organismos
eucariotas, tanto unicelulares como multicelulares. En consecuencia, este dominio
reune a los organismos pertenecientes a los reinos: Animalia, Plantae, Hongos y
Protista. El dominio Eucarya será discutido en la siguiente unidad de laboratorio.
El curso de Biología General ha sido estructurado con un enfoque evolutivo, se
presentarán a continuación los grupos que integran a los dominios Arquea y Bacteria
utilizando un esquema filogenético.
Dominio Arquea : En términos anatómicos podemos describir las arqueas como microorganismos que
carecen de núcleo, poseen una pared celular formada por proteínas, conocida como
capa S (“S layer”) y en algunos casos por azúcares aminadas, pero a diferencia de la
pared celular de las Bacterias no contienen mureina (peptidoglucano). Los
microorganismos en este dominio han sido ubicados en tres grupos filogenéticos
distintos, a base del análisis de la secuencia de su rRNA–16S. Estos son los grupos
Crenarchaeota, Euryarcheota y Korarchaeota. Los Crenarchaeota son básicamente
procariotas hipertermofílicos, dependientes de azufre para su crecimiento. El grupo de
los Euryarcheota está integrado por procariotas metanogénicos y los halofílicos
extremos. Finalmente, los Korarchaeota no han podido ser aislados o cultivados. Se
conoce de su existencia a través de la aislación y amplificación de rRNA –16S con una
secuencia de nucleótidos substancialmente diferente a la de los otros dos grupos
filogenéticos ya mencionados. Dichos ácidos nucleicos han sido aislados de ambientes

13
sometidos a altas temperaturas, similares a los ambientes habitados por los
Crenarchaeota.
Los siguientes son rasgos fenotípicos distintivos de los procariotas dentro del dominio
Arquea.
• ausencia de mureina en su pared celular (en muchas de ellas la pared está
formada por capas de proteínas conocidas comos capas S [“S layers”]
• enlace tipo eter en los lípidos de sus membranas (cadenas de isoprenoides
unidas a glicerol a través de un enlace tipo eter)
• ausencia de un residuo de timina en el t-RNA
• amplia resistencia a un gran número de antibióticos que afectan a los
procariotas dentro del dominio Bacteria
• su polimerasa de RNA presenta una estructura compleja integrada por 9 –12
subunidades, a diferencia de la polimerasa de las Bacterias que está formada
por tres subunidades y que se describe como una estructura sencilla)
Basándonos en su fisiología las Arquea pueden ser organizadas en este momento en
cuatro tipos fisiológicos :
• metanógenos (autotrofos, anaerobios estrictos que producen metano)
• halofílicos extremos (heterotrofos, viven en ambientes hipersalinos)
• termofílicos extremos (viven en ambientes con temperaturas elevadas,
algunos llevan a cabo respiración anaerobia utilizando sulfato como
aceptador final de electrones, otros son quimiolitotrofos que obtienen energía
de la oxidación de azufre elemental)
• reductores de sulfato (grupo de amplia distribución, llevan a cabo respiración
anaerobia utilizando sulfato como aceptador final de electrones en cadenas
respiratorias)
Cada tipo fisiológico presenta atributos estructurales y bioquímicos únicos que le han
permitido adaptarse a condiciones extremas de su habitat particular. Los metanógenos

14
son anaerobios obligados que no pueden tolerar una breve exposición al oxígeno. En
consecuencia, estos habitan en ambientes anaerobios, los cuales son fáciles de
encontrar en sedimentos marinos, sedimentos de cuerpos de agua dulce, pantanos,
suelos profundos, el tracto intestinal de los animales, plantas de tratamiento de aguas
usadas (desperdicios sanitarios e industriales), en lagos de oxidación de vaquerías,
porquerizas, etc. Los metanógenos emplean trayectos metabólicos únicos que le
permiten utilizar hidrógeno gaseoso (H2) como fuente de energía y CO2 como su única
fuente de carbono para el crecimiento. Ellos poseen citocromos únicos que le permiten
la reducción de CO2 a metano (CH4) a expensas de H2. En el proceso de generar
constituyentes celulares a partir de la oxidación de H2 estos microorganismos producen
energía en forma de ATP y producen metano. La actividad de los metanógenos es
responsable de la formación de las reservas principales de gas natural en nuestro
planeta. Dichas reservas son utilizadas por el hombre como fuente de energía para
uso doméstico e industrial (eg. generación de energía eléctrica en la planta de
elaboración de rones de la Bacardí). Por otro lado, los metanógenos son pobladores
esenciales de la flora microbiana del rumen de las vacas y de otros rumiantes (ej.
cabras, ovejas, jirafas) y del intestino de insectos que consumen la celulosa en madera
y papel, tales como las termitas y cucarachas. La producción biogénica de metano
excede por mucho su generación por procesos abióticos (ie. actividad volcánica). Una
vaca elimina cerca de 50 litros de metano al día durante el proceso de eruptación. El
metano que eliminan los rumiantes, las termitas y otros organismos vivos junto con el
metano originado de la actividad volcánica escapa a la atmósfera contribuyendo de
forma significativa al llamado efecto invernadero. Las prácticas actuales de
deforestación masiva, ya sea para sostener una mayor actividad agrícola o para
proveer espacio para el pujante urbanismo, la proliferación de grandes hatos de
rumiantes utilizados como fuente de alimento y la generación de gases en plantas de
tratamiento de aguas usadas provocan el que:
• se remueva menos CO2 al reducir la actividad de autotrofos fotosintéticos (el
CO2 es un gas que también abona al llamado efecto invernadero)
• se produza más CO2 y CH4 por la actividad metabólica conjunta de animales
y metanógenos

15
Las arqueas descritas como halofílicas extremas viven en ambientes naturales
hipersalinos, tales como los salitrales de la costa suroeste de Puerto Rico (Cabo Rojo,
Lajas, Guánica), el Mar Muerto y el Gran Lago Salado de Utah. Estos procariotas
Grupos representativos de las Arqueas
halobacterias
metanobacterias
arquea termofílica

16
necesitan una alta concentración de sal para crecer (ie. 4 molar, ± 25% NaCl),
requiriendo particularmente sales de sodio, para estabilizar su pared celular, ribosomas
y enzimas. Muchas de estas células presentan un pigmento conocido como
bacteriorodopsina el cual le imparte a su membrana celular un color púrpura. Cuando
se producen sobrecrecimientos (“blooms”) de estos microorganismos el ambiente
acuático donde viven adquiere colores vivos que oscilan entre el anaranjado, rojo y
rosado. La bacteriodopsina le permite a estos microorganismos protegerse de la alta
intensidad de la radiación solar que predomina en los salitrales solares, al mismo
tiempo que les permite generar energía en forma de ATP mediante la generación de
un gradiente de protones a través de su membrana. Estos microorganismos son
heterotrofos (consumen materia orgánica como fuente de carbono y energía) y
normalmente llevan a cabo respiración aeróbica para generar energía. Aún cuando la
alta concentración de sales que predomina en su habitat reduce la solubilidad de
oxígeno, estos han podido adaptarse a dichas condiciones logrando utilizar luz solar
para generar ATP mediante la bacteriorodopsina. Esto los convierte en fototrofos aún
cuando no emplean clorofilas para la captación de luz solar como fuente primaria de
energía.
Las arqueas termofílicas extremas requieren de altas temperaturas para un crecimiento
óptimo (80 – 105 °C). Pyrodictium, el termofílico más extremo hasta ahora
caracterizado, tiene una temperatura óptima de 105°C. Los termofílicos extremos son
un grupo de procariotas bien diverso. El grupo reúne a procariotas que pertenecen a
diferentes líneas filogenéticas y fisiológicas. Todos sus miembros se caracterizan por
poseer membranas y enzimas con estabilidad no usual a altas temperaturas. Muchas
de estas Arqueas requieren de azufre elemental (S°) para su crecimiento. Algunas son
anaerobias que utilizan azufre como aceptador final de electrones en lugar de oxígeno
(ie. respiración anaerobia). Otras son aerobias que utilizan azufre como fuente de
energía (son quimiolitotrofas). Estas últimas se conocen como termoacidófilas ya que
crecen a pH bajos (ie. pH <2). Acidifican su propio ambiente al oxidar azufre elemental
a sulfato, el cual reacciona con el agua para formar ácido sulfúrico (H2SO4). Las
Arqueas hipertermofílicas habitan en ambientes cálidos, ricos en azufre usualmente
asociados a procesos volcánicos. Los manantiales termales, los géiser del Parque

17
Nacional de Yellowstone, las fumarolas o ventanas termales en el lecho marino de
aguas oceánicas cerca de la Baja California y de las Islas Galápagos son ejemplos de
dicho tipo de escenario.
El grupo de las arqueas reductoras de sulfato esta integrado por procariotas del género
Archaeglobus. Estas son, hasta el presente, el único grupo de arqueas
hipertermofílicas capaces de utilizar sulfato como aceptador final de electrones. En
adición, dicho grupo comparte algunas características metabólicas con los
metanógenos.
Los cuatro patrones metabólicos arriba descritos no constituyen ramas monofiléticas
(ie. taxones filogenéticos separados). Estudios filogenéticos utilizando marcadores
moleculares (ie. secuencias del 16S rRNA) revelan que el grupo de los metanógenos
dio origen a otros grupos fisiólogicos dentro de las arqueas tales como los halofílicos
extremos, los reductores de sulfato y por lo menos dos linajes de termofílicos no
metanogénicos.
Por último es necesario señalar que las Arqueas no son los únicos microorganismos
que pueden colonizar y habitar ambientes extremos, ya que en algunos de esos
escenarios también se pueden encontrar miembros del dominio Bacteria y un número
reducido de microorganismos eucariotas (algas y hongos). En estos últimos el
intervalo de crecimiento óptimo así como el límite máximo de tolerancia para factores
abióticos tales como pH, temperatura o salinidad es menor al registrado en las
Arqueas.
Dominio Bacteria: El dominio Bacteria es integrado por procariotas que presentan en su mayoría una
pared celular cuyo componente estructural principal es la mureina o peptidoglucano. La
mureina ha sido utilizada como marcador fenotípico de las Bacterias, ya que ningún
otro organismo posee dicha macromolécula. Un estudio de la composición química de
la pared bacteriana revela dos tipos de arquitectura y de composición química (Fig. 8).

18
Figura 8 : Diferencias en la Composición Química y Arreglo Estructural de la Pared Celular en Bacterias Gram + y Gram -
Haciendo uso de una tinción diferencial ideada por Christian Gram, hoy día podemos
distinguir fácilmente entre ambos tipos de pared celular. La dicotomía ya establecida
entre Bacterias gram positivas y Bacterias gram negativas alude específicamente a
la existencia de los dos tipos de pared. Bajo el sistema de clasificación de Whittaker (sistema de cinco reinos), las diferencias
marcadas en caracteres anatómicos y fisiológicos fueron utilizadas para clasificar las
Bacterias . Rasgos fenotípicos tales como el tipo de pared (tinción de Gram), morfotipo
(morfología, Fig 9-a), arreglos de las células cuando viven agrupadas, motilidad,
capacidad para producir esporas, necesidad de oxígeno para vivir y capacidades
Tiñen violeta
peptidoglucano
pared celular
membrana plasmática
proteina
lipopolisacárido
Tiñen rojo
membrana plasmática
mem-brana externa
espacio peri- plásmico
proteina
peptidoglucano
Bacteria Gram negativa
Bacteria Gram positiva

19
Figura 9a: Morfotipos bacterianos típicos
cocos

20
metabólicas (ej. fijación de nitrógeno, bioluminiscencia, fermentación de carbohidratos,
etc.) fueron organizados en forma jerárquica para establecer un esquema de
clasificación filogenético (Fig. 9-b). Bajo ese esquema se llegaron a reconocer hasta
33 grupos bacterianos. No obstante, el uso de cronómetros moleculares para la
clasificación de bacterias revela que la catalogación tradicional usando caracteres
fenotípicos heredados de la taxonomía de plantas y animales (ej. forma, tamaño,
composición química de la pared celular, modo nutricional ) no permite descubrir las
relaciones filogenéticas entre los integrantes del dominio Bacteria. Esto se debe a
dos factores:
• La increíble capacidad que exhiben las bacterias para intercambiar
información genética a través de mecanismos de transferencia lateral de
genes (ie. conjugación, transformación y transducción) y la relativamente
alta razón de mutación de algunas regiones de su genoma pueden cambiar
los atributos fenotípicos de éstas. Esto nos puede llevar a separar a dicho
individuo y a su progenie de sus congéneres al perder un atributo (no
esencial) que distinguía al género (ej. pérdida de la capacidad para
fotosintetizar o fijar CO2 o pérdida de motilidad) o al desarrollar una nueva
estructura anatómica o una facultad metabólica que no se asociaba con su
género ( ej. desarrollo de pili del sexo o cápsula, o desarrollo de la capacidad
para oxidar una nueva fuente de carbono).
• Rasgos fenotípicos que nos permitieron elaborar esquemas filogenéticos
válidos en plantas y animales no son del todo apropiados para bacterias.
Rasgos como el morfotipo, tamaño y color pueden variar a nivel de la
especie. Para nosotros resulta fácil distinguir entre un elefante y un lagartijo
o un árbol de pana y un arbusto de acerolas. Las diferencias en formas y
tamaños nos permiten una fácil distinción entre dichas especies. No
obstante, la experiencia acumulada en el cultivo y aislación de especies
bacterianas bajo diferentes condiciones ambientales revela una gran
plasticidad genética, que les permite desarrollar variaciones en morfotipo,
tamaño y color en respuesta a cambios ambientales.

21
Figura 9 : Ejemplo de un esquema de clasificación jerárquico para bacterias
Tincion Gram
Gram positivos Gram negativos
Reacciones por tinciones
Pruebas bioquímicas
bacilos bacilos cocos cocos
esporas presentes
esporas ausentes
catalasa
aerobios (no-fermentadores)
anaerobios facultativos
(fermentadores)
lactosa
positivo negativo
agregados irregulares y tétradas: catalasa postivos
pares o cadenas: catalasa negativos
negativo positivo
Grupos Bacterianos
Grupo I Grupo II Grupo V Grupo Vi Grupo VII Grupo VIII Grupo IX Grupo X

22
Se debe señalar que aún cuando el sistema de clasificación de cinco reinos falla en
establecer una relación evolutiva coherente entre los procariotas e inclusive entre
procariotas y eucariotas, ha permitido el diseño y construcción de sistemas de
identificación automatizados. Estos son de gran utilidad en la medicina y en
actividades industriales donde se emplean cultivos puros de bacterias para generar un
producto o donde los microbios representan un problema de contaminación.
Un análisis filogenético de las Bacterias utilizando el r-RNA-16S como marcador revela
once (11) grupos distintos en este dominio. Dentro de cada grupo filogenético se
ubican organismos con caracteres anatómicos y fisiológicos bien dispares. Los once
filum que se proponen en el nuevo esquema de clasificación de tres dominios son los
siguientes :
Filum 1- Proteobacteria - Este es un grupo diverso de procariotas donde están
representados los cuatro modos nutricionales que conocemos: fotolitotros,
fotoorganotrofos, quimiolitotrofos y quimioorganotrofos. El filum reúne a
bacterias con atributos anatómicos y fisiológicos bien dispares.
Proteobacterias (Bacterias púrpuras)

23
Encontramos en el mismo grupo a:
• bacterias fotosintéticas anoxigénicas (ej. bacterias púrpuras sulfúreas), • bacterias nitrificantes (obtienen su energía de la oxidación de
amoniaco o nitrito, ej. Nitrobacter, Nitrosomonas) • bacterias heterotróficas de vida libre (ej. Pseudomonas)
• bacterias heterotróficas que viven como simbiontes en plantas (ej. Rhizobium- fija nitrógeno atmosférico; Agrobacterium – produce tumores en plantas dicotiledóneas , pero no en monocotiledóneas),
• parásitos intracelulares obligados (Rickettsia)
• bacterias predadoras de otras bacterias (Bdellovibrio).
•
Por último, se ha demostrado que la mitocondria, presente en casi todas las células eucariotas, evolucionó de este filum bacteriano.
Filum 2 – Cianobacterias - Este grupo reúne a los únicos procariotas que
llevan a cabo fotosíntesis oxigénica. Estos presentan diversidad de morfotipos y
arreglos entre los cuales están las formas unicelulares, formas filamentosas
multicelulares y arreglos coloniales. Algunas de las especies en este filum viven
en asociación simbiótica con plantas y animales superiores. Tienen gran
importancia ecológica al desempeñar un rol de importancia en los ciclos
biogeoquímicos del carbono, oxígeno y nitrógeno. De este grupo evolucionó el cloroplasto presente en todos los organismos eucariotas fotosintéticos.
Cianobacterias
heteroquiste- célula diferenciada donde se lleva a cabo la reducción de nitrogeno gaseoso

24
Filum 3 – Cytofagas (Flavobacterium, Cytophaga, Bacteroides) - Este es un
grupo diverso de procariotas heterotróficos que exhibe una gran variedad de
morfotipos. El filum incluye a aerobios y anaerobios estrictos. Algunos géneros
aquí incluídos exhiben motilidad por flagelos, otros son procariotas deslizantes y
otros son sésiles (no-mótiles).
Filum 4 – Planctomicetos – Los procariotas en este grupo se dividen en dos
géneros: Planctomyces y Chlamydia. Las especies dentro del género
Plantomyces incluyen tanto a procariotas unicelulares como a procariotas en
arreglo filamentoso. Los microorganismos en este género se distinguen por su
modo de reproducción; un patrón de gemación poco usual. En adición, estos
carecen de mureina, lo que provoca que tiñan como Gram negativos. Su pared
celular esta formada por capas de proteinas (“S layers”) que le imparten una
estructura crateriforme a su superficie. Los microorganismos en el género
Chlamydia son parásitos intracelulares obligados; en su mayoría patógenos de
humanos y otros animales. Las células en este grupo también carecen de
mureina y no pueden producir su propio ATP dependiendo entonces del ATP
que produce su hospedero.
Planctomyces
Las manchas obscuras son las inclusiones intracelulares de Chlamydia sp.
células eucariotas infectadas
Plantomyces con superficie crateriforme

25
Filum 5 – Espiroquetas - Este grupo se distingue fácilmente por su morfotipo.
Son células cilíndricas con una contorsión helicoidal y que presentan un
filamento axial que se extiende de un extremo al otro del cilindro. Dicho
filamento axial está formado por uno o más flagelos. Dentro de este grupo
encontramos patógenos humanos como Treponema pallidum, agente etiológico
(agente causal) de la sífilis (enfermedad venérea) y Borrelia burgdorferi agente
etiológico de la enfermedad de Lyme.
Filum 6 – Thermotogales (Bacterias Termofílicas Gram Negativas) – Este
filum reune a varios géneros de bacterias gram negativas. Los procariotas en
este grupo son esencialmente heterotróficos, con lípidos inusuales en su pared
celular. Estos presentan un óptimo de crecimiento dentro del intervalo de
temperaturas entre 40° y 100°C. No obstante, su óptimo de crecimiento no
alcanza el intervalo observado en las arqueas termofílicas extremas
corte transversal

26
Filum 7 – Bacterias Gram Positivas - Los procariotas en este filum presentan
una pared rica en mureina con la excepción de las Mycoplasmas. Los miembros
de dicho género carecen de pared celular, por lo que tiñen gram negativo (ie.
tiñen de rojo) cuando son sometidos a una tinción de Gram. El grupo reúne una
gran variedad de morfotipos y de capacidades metabólicas. Encontramos
organismos unicelulares, multicelulares filamentosos (lineales y ramificados) y
algunos con forma de micelios (estructura típica de hongos filamentosos). En su
mayoría son heterotrofos que habitan en suelo y sedimentos. Sin embargo,
también dentro del grupo se encuentran algunos autotrofos. Uno de ellos es un
género de bacterias fotosintéticas anoxigénicas (no produce oxígeno) conocido
como Heliobacterium. Algunos de los miembros del filum producen estructuras
especializadas para la sobrevivencia bajo condiciones ambientales hostiles.
Estas se conocen como endosporas y conidiosporas. Varias especies con la
capacidad para producir esporas producen potentes toxinas que afectan a
humanos y otros animales (ej. Clostridium tetani, C. botulinum, C. perfringes,
Bacillus antracis; estos producen las toxinas que provocan el tétano, botulismo
gangrena gaseosa y antrax, respectivamente). Por otro lado, algunas cepas de
Clostridium presentan la capacidad de fijar nitrógeno atmosférico.
Recientemente se incorporó a este grupo una de las dos especies de bacterias
que pueden ser vistas a simple vista. Esta pertenece al género Epulopiscium .
Epulopiscium fishelsoni
bacilos gram positivos formadores de endosporas enso
endosporas (estructuras ovoides refractiles)

27
Filum 8 – Bacterias Verdes Sulfúreas - Este es un grupo de bacterias
fotosintéticas anoxigénicas. Se distinguen de otros grupos de bacterias
fotosintéticas porque utilizan el ciclo reductivo del ácido tricarboxílico (Ciclo de
Krebs reductivo) para fijar CO2 en lugar del Ciclo de Calvin Benson.
Filum 9 – Bacterias Verdes no-Sulfúreas – Este es otro grupo de bacterias
fotosintéticas anoxigénicas. Los integrantes de este grupo emplean una ruta
metabólica diferente al ciclo reductivo del ácido tricarboxílico y al ciclo de Calvin
Benson para fijar CO2. Las bacterias del género Chloroflexus resultan ser muy
peculiares dado su capacidad para moverse por deslizamiento y por su
versatilidad metabólica. Estas bacterias pueden vivir como autotrofos, llevando
a cabo fotosíntesis y también pueden vivir como heterotrofos.

28
Filum 10 – Deinococcus y Thermus - Este filum está integrado por dos grupos
con atributos fenotípicos bien dispares. De un lado incluye al género
Deinococcus, el cual está integrado por bacterias gram positivas con forma
esférica (cocos). Las bacterias de este género se distinguen de otros géneros de
cocos gram positivos por su gran resistencia a la radiación gamma y a la luz
ultravioleta. El otro grupo corresponde al género Thermus. Estas son bacterias
termofílicas con forma de bastón o cilindro (bacilos). Dentro de dicho género se
encuentra la especie Thermus aquaticus, procariota de donde se aisló la
polimerasa de DNA que es utilizada en la Reacción de Polimerización en
Cadena (“Polymerase Chain Reaction- PCR”). Esta última es la base de toda
una nueva gama de técnicas moleculares dirigidas a amplificar e identificar
secuencias de ácidos nucleicos. Estas técnicas fueron objeto de gran atención
y polémica durante el famoso juicio contra el futbolista y actor O. J. Simpson.
Deinococcus sp.
Thermus aquaticus

29
Filum 11 – Hydrogenobacteria – Dentro de este filum están ubicadas las
bacterias autotróficas que obtienen la energía para fijar CO2 de la oxidación de
hidrógeno gaseoso (H2). El grupo incluye a los géneros Aquifex y el género
Hidrogenobacter. Los miembros del género Aquifex representan el grupo con
mayor capacidad termofílica dentro del dominio Bacteria y al mismo tiempo
representan el grupo más antiguo dentro de dicho dominio.
TAMAÑO: Treinta trillones (30 x10 12) de células procariotas de tamaño promedio pesan alrededor
de 28 gramos. Esto nos da idea de cuán pequeñas son dichas células. La unidad de
medida para éstas células es el micrómetro (µm; 10-6m). Su tamaño oscila entre 0.1 –
0.4 um de ancho a 0.2 – 50 um de largo (tamaño promedio). Recientemente se han
descubierto dos especies de bacterias macroscópicas, las cuales representan la
excepción a la regla. Estas han sido clasificadas como Epulopiscium fishelsoni y
Thiomargarita namibiensis (Fig. 10). La primera es una bacteria heterotrófica aislada de
un pez sargento en el Mar Rojo con unas dimensiones de 600 µm de largo por 80µm
de diámetro. Esta se ha asociado filogenéticamente con los procariotas del género
Clostridium (bacilos gram positivos, anaerobios estrictos, formadores de endosporas).
La segunda es una bacteria quimiolitotrófica que obtiene su energía de la oxidación de
azufre y que fue aislada de sedimentos en las costas de Namibia en Africa. Esta última
resulta ser la bacteria más grande hasta ahora conocida. Si aumentáramos el tamaño
de Thiomargarita namibiensis al de una ballena azul, entonces proporcionalmente una
bacteria común sería un poco más pequeña que un ratón recien nacido y Epulopiscium
fishelsoni sería del tamaño de un león. No obstante, recordemos que la inmensa
Aquifex sp.

30
mayoría de los procariotas que hasta ahora han sido identificados (alrededor de 4000
especies) son todos microscópicos.
Figura 12: Bacterias Macroscópicas
Bacteria Punta de un alfiler
Epulopiscium fishersoni
Thiomargarita namibiensis