almoraima 35
TRANSCRIPT

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 1/276
TAMBIÉN NOSOTROSESTAMOS POR LA LABOR
P or la hermosa labor de apoyar la culturade nuestra tierra.
Es lo que, con ALMORAIMA, viene realizando laMancomunidad de Municipios de la comarca delCampo de Gibraltar.
Y nosotros también estamos por tan valiosa laborpatrocinando esta revista que es "un medio plural yriguroso que se ocupa de desentrañar los entresijosde la historia y contemporaneidadcampogibraltareñas".
Si toda la provincia es nuestro campo de acción,¿cómo no estar por la labor culturalcampogibraltareña?

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 2/276
J

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 3/276
3
ALMORAIMARevista de Estudios Campogibraltareños
Número 35 - Octubre 2007
EDITA
MANCOMUNIDAD DE MUNICIPIOSDEL CAMPO DE GIBRALTAR
DEPARTAMENTO DE CULTURA
DIRECCIÓN, DISEÑO Y MAQUETA
Rafael de las CUEVAS SCHMITT
SECRETARIA DE DIRECCIÓN
Mª Ángeles ÁLVAREZ LUNA
CONSEJO DE EDICIÓN
Mario Luis OCAÑA TORRESÁngel J. SÁEZ RODRÍGUEZ
Antonio BENÍTEZ GALLARDO
Carlos GÓMEZ DE AVELLANEDA SABIOJuan Emilio RÍOS VERARafael MÉNDEZ PEREA
Rafael FENOY RICOEduardo BRIONES VILLA
FOTOGRAFÍAS
Archivo-Cedidas
REDACCIÓN
MANCOMUNIDAD DE MUNICIPIOS
DEL CAMPO DE GIBRALTAR
DEPARTAMENTO DE CULTURAC/ Del Museo, nº12 · 11207 Algeciras (Cádiz)
Teléfono: 956 580 069Correo electrónico: [email protected]
IMPRESIÓN Y FOTOCOMPOSICIÓN
IMPRESUR, S.L.Avda. de Italia, Blq. 7. Anexo K · 11205 Algeciras
¥ 956 652 051 · Fax 956 587 274Correo electrónico: [email protected]
I.S.S.N. 1133-5319Depósito Legal CA-868-89
Premio LAURISILVA 1996
AGADEN - Campo de Gibraltar
ILUSTRACIÓN PORTADA:
Punta CamarinalFotografía de Orlando Garzón Gómez

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 4/276
4
NORMAS PARA LA PRESENTACIÓN DE COLABORACIONES
Se admitirán únicamente trabajos inéditos relacionados con los temas del Campo de Gibraltar –Historia, Geografía,Arqueología, Artes, Letras, Costumbres, Ciencias, etc– que son fundamento de su contenido.
Los originales se presentarán en CD de ordenador, preferiblemente en formato Word (seleccione "Guardar como…"en su procesador de texto), e impresos por duplicado en hojas tamaño A4, guardando un margen de 2'5 cm por todossus lados.
La extensión de los trabajos no deberá ser inferior a tres folios ni superior a veinte, incluídas notas y bibliografía. Nose admitirán trabajos que no vengan acompañados de las correspondientes referencias documentales y bibliográficas.Se adjuntarán los datos personales del autor: nombre, domicilio, teléfono de contacto.
El tipo de letra a emplear será Times New Roman, tamaño 12 puntos. Se aplicará un cuerpo menos (tamaño 11) encitas de más de 3-4 líneas dispuestas en párrafo aparte.
Las notas irán numeradas de forma consecutiva y reseñadas en página aparte al final del texto. Las fuentesdocumentales y bibliográficas, asimismo en página aparte al final del texto, serán citadas de la siguiente manera:
- Documentos: Título del documento, archivo, sección y legajo.
- Libros: Apellidos (en mayúsculas) y nombre del autor. Título de la obra (en cursiva). Lugar de edición.Editorial. Año. Número de la página citada.
- Revistas: Apellidos y nombre del autor. Título del artículo (entre comillas). Nombre de la revista (en cursiva).Número. Año. Lugar de edición. Editorial. Número de la página citada.
Para lo no especificado en estas normas de presentación de colaboraciones, los autores podrán consultar las Normasde Estilo editadas por el Departamento de Cultura y el Instituto de Estudios Campogibraltareños, solicitándolas enel caso de no disponer de ellas.
Las figuras y fotografías que acompañen al texto deberán tener su referencia claramente anotada en el mismo y, enhoja a parte, se reseñarán los correspondiente pies de cada una. Asimismo, se hará constar el orden de prioridad queel autor estime para su publicación en previsión de que no todas puedan ser incluidas.
Los gráficos y tablas digitalizados deben presentarse en alguno de los siguientes formatos: TIFF, BMP o JPEG (paracualquier tipo de ilustración, incluidos los gráficos estadísticos), y no deben ser incluidos en el archivo de texto.
Las fotografías e ilustraciones que acompañen a los mismos estarán sujetas para su publicación al espacio disponibleen el conjunto de la maqueta de cada número de la Revista, a criterio de los responsables de Diseño y Compaginacióny de la Dirección de la misma.
El Consejo de Edición de la Revista decidirá sobre la publicación en cada caso de los trabajos recibidos que hayansido admitidos previamente por ajustarse a estas Normas. El criterio de oportunidad de publicación valorará laoriginalidad, el rigor científico y la necesaria diversidad de materias que deben ser tratadas en cada número deALMORAIMA.
Las colaboraciones deberán ser enviadas a: ALMORAIMA - Revista de Estudios Campogibraltareños. Mancomu-nidad de Municipios del Campo de Gibraltar. Parque "Las Acacias" s/n. 11207 ALGECIRAS (Cádiz).
ALMORAIMARevista de Estudios Campogibraltareños

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 5/276
5
PALABRAS DE BIENVENIDA DEL DIRECTOR DELINSTITUTO DE ESTUDIOS CAMPOGIBRALTAREÑOS
Sirvan estas primeras palabras de la Dirección del Instituto de Estudios Campogibraltareños para dar la bienvenida a todoslos asistentes a estas nuevas Jornadas de Flora, Fauna y Ecología del Campo de Gibraltar, que van a desarrollarse a lo largode este fin de semana en la ciudad de Jimena de la Frontera en pleno corazón del Parque Natural Los Alcornocales.
Como en otras ocasiones el Comité Técnico se ha tenido que enfrentar con la compleja labor de seleccionar, entre una granvariedad de trabajos, todos ellos de gran calidad científica, novedosos, en unos casos, y continuadores de líneas deinvestigación abiertas en encuentros anteriores en otros, siendo prácticamente insignificante el número de aquellos que se
han rechazado, nunca por razones de contenido, sino por el hecho de no adecuarse a las normas que establece el Institutode Estudios.
Con la perspectiva de varias Jornadas anteriores y con el conocimiento de los trabajos que aquí van a exponerse, cuyostítulos en el programa no constituye más que un avance de los interesantes contenidos que en ellos se manifiestan, nospermitimos, desde la Dirección, adelantar un intenso y largo fin de semana de exposiciones, debates y coloquios que, sinduda, contribuirán, como ya viene siendo habitual, a ampliar el conocimiento que, tanto como los especialistas como losinteresados por los aspectos naturales de nuestra Comarca, obtenemos de estos encuentros bianuales.
Quisiéramos agradecer el apoyo y el respaldo habitual de la Presidencia de la Mancomunidad de Municipios y de la entidadfinanciera Cajasol, que vienen apostando rigurosamente por el proyecto cultural comarcal que supone el IECG y del queestas Jornadas son una muestra fehaciente.
Pero especialmente esta Dirección se felicita por contar con la participación de tantos y tan eminentes investigadores,profesores, naturalistas y estudiosos del medio natural, muchos de ellos integrados en la Sección 10º, presidida por EduardoBriones, que son la columna vertebral de todo el proceso de investigación y exposición de nuevos aspectos de nuestroentorno, en estos tiempos en que los cambios que se están produciendo de forma general en nuestro planeta comoconsecuencia de calentamiento global tienen también un reflejo, y no es este de escasa entidad para los que aquí habitamos,en nuestra Comarca.
No quisiera olvidar la labor del Secretario Coordinador del IECG, Rafael de las Cuevas, que, a partir de este instante seenfrenta a la ardua tarea de convertir estas exposiciones orales y breves, por razón del tiempo del que disponemos, en unvolumen extraordinario que bajo la marca Almoraima recogerá en un plazo que todos esperamos sea breve, el ampliocontenido de todas y cada una de las Comunicaciones aquí expuestas.
Sean todos ustedes bienvenidos a estas Jornadas de Flora, Fauna y Ecología del Campo de Gibraltar.
En Jimena de la Frontera, octubre de 2005.
Mario L. Ocaña Director del Instituto de Estudios Campogibraltareños

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 6/276
J

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 7/276
7
ÍNDICE
MEDIO NATURAL Y CUATERNARIO DE GIBRALTAR EN LOS TEXTOSDE VIAJEROS POR LA PENÍNSULA IBÉRICA DURANTE LOS SIGLOS XVIII Y XIX
José Manuel Recio Espejo ................................................................................................................................................. 11
CONSERVACIÓN FRENTE A DESARROLLO:¿HACIA UN DESARROLLO INSOSTENIBLE EN SAN ROQUE?
Alfonso Gallardo Mayenco / Eric Shaw ............................................................................................................................ 21
CAMBIOS EN LA FAUNA DE VERTEBRADOS DEL CAMPO DE GIBRALTAR
DESDE MEDIADOS DEL SIGLO XIX HASTA LA ACTUALIDAD Arturo Menor / José Prenda .............................................................................................................................................. 31
LA CAZA COMO FACTOR DE CONSERVACIÓN DEL CORZO EN EL PARQUE NATURALLOS ALCORNOCALES. UNA VISIÓN RETROSPECTIVAPablo Fernández-Salguero López / José María Sánchez García ...................................................................................... 43
INFLUENCIA DE LA CONCENTRACIÓN DE HIDROCARBUROS SOBREEL ERIZO COMÚN (PARACENTROTUS LIVIDUS)
José Miguel Ramos Durán / Orlando Garzón Gómez ...................................................................................................... 53
OBSERVACIONES SOBRE EL MEDIO NATURAL DEL CAMPO DE GIBRALTAR YZONAS PRÓXIMAS EN LOS TEXTOS DE VIAJEROS POR ESPAÑA (SIGLOS XVIII Y XIX)
José Manuel Recio Espejo ................................................................................................................................................. 69
EL PROGRAMA MIGRES DE AVES PLANEADORAS. DATOS ACTUALIZADOS A LOS AÑOS 2005 Y 2006 Luis Barrios Jaques / Guillermo Doval de las Heras ....................................................................................................... 77
HUMANS, MONKEYS, AND THE ROCK: THE ANTHROPOGENIC ECOLOGY OFTHE BARBARY MACAQUES IN THE UPPER ROCK NATURE RESERVE, GIBRALTAR
Agustín Fuentes y otros ...................................................................................................................................................... 87
EFECTO DE UN ESCARABAJO PELOTERO (THORECTES LUSITANICUS) SOBRELA REGENERACIÓN DE DOS ESPECIES DE
QUERCUSEN UN BOSQUE MEDITERRÁNEO
Ignacio M. Pérez-Ramos .................................................................................................................................................... 99
EVALUACION Y RECUPERACIÓN DE ZONAS INCENDIADAS EN EL MONTE HOYO DE DON PEDRO(TÉRMINO MUNICIPAL DE LOS BARRIOS, CÁDIZ)
Miguel Cueto Álvarez de Sotomayor y otros ................................................................................................................... 109
NUEVAS APORTACIONES AL CONOCIMIENTO DE LOS MOVIMIENTOS MIGRATORIOSDE LA TORTUGA BOBA (CARETTA CARETTA) A TRAVÉS DEL ESTRECHO DE GIBRALTARA PARTIR DE LA INFORMACIÓN RECOPILADA EN VARAMIENTOS
J. J. Bellido López y otros ................................................................................................................................................ 121

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 8/276
8
ESTUDIO COMPARATIVO ENTRE MUESTREOS DESDE TIERRA Y MARDE LOS CETÁCEOS DE LA BAHÍA DE ALGECIRASFrancisco Ruiz Giráldez y otros ...................................................................................................................................... 129
DISTRIBUCIÓN TEMPORAL DE CETÁCEOS EN EL ESTRECHO DE GIBRALTAR.¿ES REALMENTE UN CANAL DE MIGRACIÓN PARA CETÁCEOS EL ESTRECHO?
Renaud De Stephanis y otros ........................................................................................................................................... 137
DISTRIBUCIÓN ESPACIAL DE CETÁCEOS EN EL ESTRECHO DE GIBRALTARDURANTE LA ÉPOCA ESTIVALSergi Pérez y otros ............................................................................................................................................................ 147
ESTIMACIÓN DE LA ABUNDANCIA ABSOLUTA DE LAS POBLACIONES RESIDENTES YSEMIRESIDENTES DE CETÁCEOS EN EL ESTRECHO DE GIBRALTARPhilippe Verborgh y otros ................................................................................................................................................ 159
INTERACCIONES ENTRE CETÁCEOS Y ACTIVIDADESANTROPOGÉNICAS EN EL ESTRECHO DE GIBRALTAR
Alicia Sánchez y otros ...................................................................................................................................................... 167
LA LAGUNA DE LA JANDA (CADIZ): 10 AÑOS DESPUÉS DEL INICIO DELOS ESTUDIOS BÁSICOS PARA SU RESTAURACIÓN ECOLÓGICA
J. C. Castro Román / J. M. Recio Espejo ......................................................................................................................... 175
DECLINE OF THE LESSER KESTREL (FALCO NAUMANNI ) IN GIBRALTAR:POSSIBLE CAUSES, CONSERVATION AND ACTION PLANKeith J. Bensusan / John E. Cortés .................................................................................................................................. 185
ESTABLECIMIENTO DE UNA DENSIDAD ÓPTIMA PARA EL CORZOEN UNA FINCA DEL PARQUE NATURAL DE LOS ALCORNOCALES
Alejandro Vega Contreras / Ricardo Zamora Diaz / Eduardo Briones Villa .................................................................. 191
DISTRIBUCIÓN Y ESTATUS DE LOS MAMÍFEROS CARNÍVOROSEN EL TÉRMINO MUNICIPAL DE LOS BARRIOS (CÁDIZ)F. J. Gómez Chicano / E. Briones Villa / A. Muñoz Brenes ............................................................................................. 203
EL EMPLEO DE TÉCNICAS DE CONTROL Y ERRADICACIÓN EN LA LUCHA CONTRAESPECIES INVASORAS: CONVENIENCIA DE LA HERBIVORÍA COMO MÉTODO DE BIO-CONTROLDE LA INVASIÓN DE SPARTINA DENSIFLORA BRONGN. EN EL CAMPO DE GIBRALTAREnrique Mateos Naranjo / Susana Redondo Gómez / Jesús M. Castillo / M. Enrique Figueroa .................................. 217
CARACTERIZACIÓN SILVOPASTORAL DE LOS ALCORNOCALES DE LA PROVINCIA DE CÁDIZ Luis Linares García ......................................................................................................................................................... 223

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 9/276
9
CARTOGRAFÍA Y DESCRIPCIÓN DE LA VEGETACIÓN DEL PARAJE NATURAL DELAS MARISMAS DEL RÍO PALMONES Y DEL PARAJE NATURAL DEL ESTUARIO DEL RÍO GUADIAROCarlos J. Luque Palomo y otros ....................................................................................................................................... 233
APLICACIÓN DE UN SISTEMA DE INFORMACIÓN GEOGRÁFICA AL ANÁLISIS DEL MEDIOFÍSICO EN EL PARQUE NATURAL LOS ALCORNOCALES. APROXIMACIÓN A UNACARTOGRAFÍA GEOMORFOLÓGICA A PARTIR DE UN MODELO DIGITAL DE ELEVACIONES
Lorena Martínez-Zavala / Antonio Jordán López / Patricia Illana Sanmiguel .............................................................. 245
CREACIÓN DE UNA NUEVA POBLACIÓN DE LA ESPECIE PROTEGIDA LIMONIUM EMARGINATUM EN EL LITORAL DEL ESTRECHO DE GIBRALTAR
Orlando Garzón / Jesús M. Castillo / Susana Redondo / M. Enrique Figueroa ............................................................ 255MEDIDAS COMPENSATORIAS DEL PROYECTO REMO II DE RED ELÉCTRICA DE ESPAÑA
Manuela de Lucas Castellanos ........................................................................................................................................ 265
SEGUIMIENTO DE LAS COMUNIDADES INTERMAREALES DE LA ISLA DE TARIFA (CÁDIZ):EDUCACIÓN AMBIENTAL Y CONSERVACIÓN
J. M. Guerra García / A. Ruiz Tabares / J. C. García Gómez ......................................................................................... 269

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 10/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 11/276
Comunicaciones
11
Los viajeros que visitaron la península Ibérica durante los siglos XVIII y XIX aportaron interesantes datos sobre la cultura,el ambiente social y el modo de vivir de los habitantes de las ciudades y comarcas que atravesaron; el folclore, losmonumentos, personajes y el orientalismo entre otros aspectos, centraron la atención de los mismos. Las estanciasrealizadas, por lo general de meses e incluso de años como en el caso de Richard Ford, fueron realizadas por diversosmotivos, desde la formación cultural a los motivos personales o políticos, pero siempre atraídos por el mensaje de exotismoy herencia orientalista que ofrecía Andalucía, y que nuestro país exportaba al resto de Europa.
Para casi todos ellos Gibraltar constituyó siempre un objetivo importante de esos itinerarios; estancias efectuadas unasveces en momentos políticos difíciles, y en otros coincidentes con condiciones más favorables que hicieron posible unasdescripciones más, pormenorizadas de este enclave geográfico, tal como aconteció durante casi todo el siglo XIX tras laguerra de la independencia.
De manera indirecta o directa estos viajeros aportaron al mismo tiempo datos importantes sobre el medio natural de losterritorios que visitaron. Aunque a veces esta información naturalista estuviera falta de rigurosidad o casi ausente comoen las descripciones efectuadas por W. von Humbold (1799-1800), Doré y Davillier (1874) o A. Dumas (1846) por ejemplo,otras muchas si fueron escritas con un gran fondo científico. Tales lo fueron las aportadas por Guillermo Bowles, JosephTownsend, Francis Carter, Richard Twiss, este último por ejemplo miembro de la Royal Society de Londres, WilliamsJacob, W.G. Clark, George Borrow o Richard Ford (en su epitafio figura “ Rerum Hispaniae Indagador Acerrimus”).Además de los citados, de los textos literarios de Hans Christian Andersen, Teófilo Gautier y otros autores también ha sidoposible extraer observaciones y deducciones interesantes.
De los trabajos de Guerrero (1990) o Krauel (2004) deducimos que fueron muchos los viajeros que nos visitaron, y que esmuy abundante la información dejada por ellos en forma de cartas y otros escritos. Importantes textos como los de H. Inglis(1831), T. Roscoe (1835) o G. Dennis (1839) aun no traducidos (Alberich, 1976; Jiménez, 2002), están siendo estudiadosactualmente por nosotros. Dejamos toda esta labor para posteriores trabajos, limitándonos en éste a realizar aquelloscomentarios que hemos considerado más interesantes sobre el Cuaternario y el medio natural de Gibraltar, extraídos de lostextos que de una manera u otra han podido ser localizados y estudiados por nosotros hasta la fecha.
MEDIO NATURAL Y CUATERNARIO DEGIBRALTAR EN LOS TEXTOS DEVIAJEROS POR LA PENÍNSULA IBÉRICADURANTE LOS SIGLOS XVIII Y XIX
José Manuel Recio Espejo

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 12/276
Almoraima, 35, 2007
12
I. DENOMINACIONES Y SIGNIFICACIÓN GEOGRÁFICA DE GIBRALTAR
Richard Twist visitó la península Ibérica durante diez meses entre 1771 y 1773, realizando en Gibraltar una estancia de tressemanas; sus comentarios al hablar de Gibraltar son los siguientes: “Omito a propósito la antigua historia, verdadera ofabulosa, de la ciudad y del Hércules egipcio, del que dicen que separó las montañas de Calpe y de Abyla; la primera delas cuales es el peñón de Gibraltar y la otra es la colina de los Monos en Berbería. De esta manera abrió un medio decomunicación entre el océano y el Mediterráneo. Luego levantó dos pilares, según el pensaba, en los límites del mundo”.
Francis Carter, quien residió en Gibraltar en torno a 1775, decía que “El peñón de Gibraltar, su configuración es notableporque se adentra en el mar como una sítula o cubeta, idea que hizo a los griegos llamarle Calpe; tiene casi una legua denorte a sur y anchura irregular, oscilando entre media y tres cuartos de milla; su cima con frecuencia está escondida entrelas nubes”; “La montaña perdió el nombre de Calpe al llegar Tarif; Abdulmalic, historiador de los reyes de Marruecos,deduce su nombre actual de Gibeltah, o montaña de la entrada, por ser la llave que les dejó entrar en España”
Richard Ford, que al igual que Twiss utilizaron su visita a Gibraltar para pasar al norte de África, a la Berbería como sele conocía, y concretamente a las ciudades de Tetuán y Tánger respectivamente, señala en sus extensos comentariosrealizados sobre la Roca lo siguiente: “Los fenicios la llamaron Alube. Abyla es la columna africana. El nombre en españoles Cabo de Bullones.”
El francés Teófilo Gautier, aporta otros topónimos con connotaciones geográficas importantes: “Gibraltar era llamado porlos árabes “Ghiblatah”, es decir “el monte de la entrada”. El monte de Los Monos está al otro lado en África, muy cercade Ceuta”. W. G. Clark en su visita en 1850 aborda su nombre de una forma más familiar afirmando: “¿Qué puedo decirde Gibraltar? ¿No es ya más familiar para nosotros que Plymouth o Chatham?-incluso nos referimos a él con el disminutivo
de “Gib”, mientras nunca decimos “Plym” o “Chat”.Por ultimo, nuevamente R. Ford llegó a deducir que “Calpe ha sido interpretado como Ca-alpe, “la caverna de Dios”; almismo tiempo le llama Gib, “que es como suele abreviarse el nombre de la Roca”.
II. EL MARCO GEOGRÁFICO
Joseph Townsend, quizás uno de los viajeros de mayor formación científica de viaje por la Península, tan solo comenta delos elementos naturales y paisajísticos del entorno más inmediato de Gibraltar lo siguiente: “Antes del anochecer pasamos
junto al estrecho de Gibraltar, donde tuve el placer de observar la altiva roca ante cuya visión cualquier corazón británicodebe sentirse jubiloso recordando a sus bravos defensores...”. Interpretamos que eran momentos de asedios y situaciones
difíciles que dificultaban la visita de Gibraltar, inspirando a veces algún otro tipo de comentarios.Uno de los más interesantes comentarios es el realizado por George Borrow, al que se le atribuye la idea de comparar lamorfología del Peñón con la de un león durmiente:
La travesía por aquellas aguas difícilmente puede carecer de interés ni siquiera para el apático, dado el panorama quese ofrece a ambos lados. Las costas son sumamente altas y escarpadas, en especial las de España que parece querersobrepasar a las moriscas, pero antes Tarifa, doblando hacia el sudoeste, el continente africano adquiere un airesublime y grandioso. Una montaña blanquecina parece atravesar las nubes con su cumbre. Es el monte Abyla, o comose llama en árabe, Gibil Muza, montaña de Muza, debido a que encierra el sepulcro de un profeta llamado así. Estees una de las dos excrecencias de la naturaleza a la que el Viejo Mundo aplicó el título de columnas de Hércules. Susladera y bordes se extienden sobre mucha leguas, en varias direcciones, pero su parte frontal, escarpada y colosal, mira

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 13/276
Comunicaciones
13
de pleno hacia el punto del continente europeo, donde yace Gibraltar, cual monstruo enorme adentrándose en el mar.De las dos colina o pilares la mas interesante, vista de lejos, es la africana Gibil Muza; es una enorme masa informevisible a mayor distancia, pero examinadas desde cerca la columna europea despierta mas admiración.
Quizás la más literaria e intensa, pero al mismo tiempo no exenta de interesantes observaciones de tipo científico se debaa Teófilo Gautier, cuando afirma:
A la altura de Tarifa, Europa y África se acercan y parecen querer darse un beso de alianza. El estrecho está aquí tancerrado, que se descubren a la vez dos continentes. Cuando uno se encuentra en el lugar mismo, es imposible creerque el Mediterráneo no haya sido, en una época no demasiada lejana, un lago interior, un mar aislado como el marCaspio, el mar de Arahal y el mar Muerto. El espectáculo que se presentaba a nuestros ojos era de una magnificenciamaravillosa.
R. Ford, a la altura de Tarifa y desde la actual torre del Cuervo relata:En la torre de La Peña del Ciervo, se abre la magnífica costa africana. África, que nos es tierra de desérticas arenas,se levanta bruscamente del mar con tremendo ímpetu coronada con las nieves eternas del Atlas inferior; ante nosotrosyacen dos continentes; hemos llegado al extremo del mundo antiguo, un agosto golfo divide las tierras delconocimiento, la libertad y la civilización de las regiones vírgenes de la ignorancia bárbara, la esclavitud, el peligroy el misterio.
Y continúa diciendo:
Una corriente constante entra desde el Atlántico a razón de dos millas y media por hora. Algunos han supuesto laexistencia de una corriente submarina, para aliviar al Mediterráneo de esta aportación de agua, además de la quesuponen todos sus ríos, desde el Ebro hasta el Nilo. El doctor Halley sin embargo ha calculado que la cantidad de agua
evaporada y absorbida por el sol es superior a toda la que recibe el Mediterráneo, el cual ciertamente, ha bajado algoen la costa oriental de la península.
Joseph Towsend, previamente en 1791, ya había discutido y reflexionado ampliamente sobre este tema.
III. MORFOLOGÍA DEL PEÑÓN
En 1754 Guillermo Bowles no llegó a visitar Gibraltar ya que sus metas de índole minera no le hicieron tenerlo en suhorizonte; tan solo hizo el siguiente comentario: “Hay montañas enteras en España compuestas de piedra caliza, como lamontaña de Gibraltar, a la cual disolvería seguramente una lluvia de ácido. Lo mismo es la de Morón, donde se hace la mejorcal que yo conozco”.
Con posterioridad R. Twiss proporcionó unos primeros comentarios sobre su morfología, aportando datos sobre lasdimensiones del mismo:
La ciudad esta situada al pie del lado oeste de una montaña o peñón, llamada Calpe. La longitud es de unas dos millasy tres cuartos desde el extremo norte, que elevándose abruptamente desde el pequeño istmo, lo une al continente. Sualtura perpendicular sobre el nivel el mar es de 1.360 pies (aunque el coronel James dice que es 1.403 pies). La parteoriental es casi inaccesible aunque varios oficiales me aseguraron que habían subido hasta la cima por ese lado.
G. Borrow dice al respecto:
En Gibil Muza solo habitan lobos, jabalíes y pequeñas monas, razón por la cual los españoles la llaman montaña delas monas. Gibraltar por el contrario, es la montaña de aspecto más singular del mundo, indescriptible, que los ojos

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 14/276
Almoraima, 35, 2007
14
nunca se cansan de contemplar. Contemplada desde mi posición (desde la proa del barco de llegada desde la bahíade Algeciras) tiene cierta semejanza con un terrible león tendido, cuya magnifica cabeza amenazara a España. Másaun, puesto que el color de sus flancos pedregosos, del espinazo y cerviz, es leonado como la piel del rey del desierto.
William George Clark, que estuvo en Gibraltar en agosto y septiembre de 1850 dice a este respecto: “Subimos a la casade señales. Todos coinciden en que la vista desde este punto es una de las mejores del mundo, y yo estoy de acuerdo. Tanclaro era el aire, que las rocas de África, distantes diez millas, parecen estar a tiro de flecha”. El tres de septiembre dejóla Roca y mientras esperan el barco comentaba:
Engañábamos nuestra impaciencia mirando desde la azotea, a través del llano Terreno Neutral, haciendo dibujos máso menos primitivos, de la gran roca de Gibraltar que se alzaba sobre nosotros como un británico león durmiente. Séque ese parecido se le ocurrió a Mr. Borrow en el mismo lugar; pero él lo describe amenazando a España, mientras
que para mí su cabeza apunta en otra dirección.Y para completar transcribimos los sugerentes comentarios de T. Gautier quien nos dice al entrar en Gibraltar procedentede Cádiz en barco de vapor:
El aspecto de Gibraltar desorienta por completo la imaginación, y ya no sabe uno ni dónde está ni lo que ve. Imaginaduna inmensa roca o más bien una montaña de quinientos pies de altura que surge de repente, bruscamente de mediodel mar en una tierra tan llana y tan baja que apenas se distingue. Nada la prepara, nada la motiva, no está unida aninguna cordillera. Es un monolito monstruoso, lanzado del cielo, un trozo de planeta mellado caído allí con ocasiónde una batalla de astros, un fragmento del mundo roto.¿Quien lo ha depositado aquí?. Sólo Dios y la eternidad lo saben.
Lo que añade aún más el efecto de esta roca inexplicable es su forma: parece como una esfinge de granito enorme,desmesurada, gigantesca, como podrían tallar unos titanes que fueran escultores…
El alargamiento de las patas forma lo que se llama punta de Europa; la cabeza, un poco truncada, está vuelta hacíaÁfrica que parece mirar con una atención soñadora y profunda. Los hombros, los riñones y la grupa se extienden haciaEspaña con unos grandes pliegues descuidados, en bellas líneas sinuosas como las de los leones en reposo.
IV. SOBRE EL ISTMO Y OTROS ASPECTOS DEL LITORAL
Este mismo último autor (T. Gautier) realizó la siguiente afirmación con respecto a esta formación litoral: “Como Cádiz,Gibraltar, situado a la entrada de un golfo en una península, no está unida al continente más que por una estrecha lenguade tierra que llaman “terreno neutro”, en las que están establecidas las líneas de aduanas”.
Algo más científico se mostró Richard Ford al escribir: “Una cinta estrecha y llana de arena, llamada “terreno neutral”,
separa la Roca de la tierra firme; desde aquí, vista a distancia, parece una isla, cosa que, indudablemente, fue en otrotiempo”. Estas observaciones pueden ser aún más matizado con las aseveraciones por ejemplo de Hans Christian Andersencuando afirmaba que:
Salimos al llano istmo que enlaza Gibraltar con España. Todo estaba asolado y seco; aquí y allá veíase alguna adelfa…;el polvo cubría las pesadas hojas, largas como sables… En el camino arenoso por donde íbamos, habían levantadoun campamento…; No cabe duda de que Gibraltar, en otro tiempo, fue un simple peñasco en el mar, hasta que la arenase fue acumulando entre éste y tierra firme, llegando a unirlo a ella, con lo que Gibraltar se convirtió en la punta surde Europa.
El texto de R. Ford aporta comentarios de mayor calado científico:

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 15/276
Comunicaciones
15
La marea se levanta a cosa de cuatro pies. La Roca está compuesta principalmente de piedra caliza gris y mármol; laRoca surgió en una época relativamente reciente, ya que hay una playa marina a cuatrocientos cincuenta pies porencima del nivel del mar”. South Port o Puerto Sur, llegando a la Alameda o Explanada, llamada antes “arenas rojas”,y que fue un desierto ardiente…
F. Carter afirmaba sobre el Peñón:
Se une al continente por un istmo de baja y profunda arena, de su misma anchura, pero que se ensancha bastante alalcanzar la frontera española; este istmo tiene casi una legua de largo, formado con la costa española una excepcional
y segura bahía de ocho millas de ancho; Que el mar cubría antiguamente estas arenas que tenemos delante y que hizoel peñón una isla no admite discusión; es más, podemos saber a qué altura de la roca llegó el agua, pues el mar haminado cavernas y cuevas y la ha descolorido hasta cuarenta pies por encima del nivel actual de las arenas; esto esevidente a simple vista; también es obvio que el mar ha ido bajando gradualmente: la torre del Diablo está construidasobre una roca (sin duda en tiempos recientes), a unos ocho pies por encima del suelo, roca a la que evidentementeantes bañaban las olas del mar.
Sobre las arenas rojas existentes en la línea de costa, Carter observó que “Aunque el peñón de Gibraltar está rodeado demar, se puede encontrar en él agua de pozo, potable y bastante buena, aunque dura y muchas veces salobre; no obstanteel agua de lluvia que baja de la montaña filtrándose por las arenas rojas cerca de Southport es muy buena y saludable y semantiene incorrupta durante mucho tiempo; se almacena en un aljibe y de allí se distribuye a la ciudad”.
Figura 1. Gaucín, mirando hacia Gibraltar y la Costa de Berbería, según David Roberts.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 16/276
Almoraima, 35, 2007
16
Para finalizar este apartado haremos usos de unos comentarios complementarios de R. Twiss: “A la salida de una de ellas(las puertas) vi a algunos oficiales jugando al golf en la arena, de la misma forma que había visto jugarlo en Links”.
V. EL CLIMA
Las especiales condiciones que adquiere el viento del este al soplar y ascender la escasa elevación del peñón, ya fueronpuesto de manifiesto por algunos de estos viajeros. Así, Francis Carter comentó:
El clima de Gibraltar es sano y menos caluroso que el de las demás ciudades de la costa; aun así, ocho meses del añohay vientos de levante que soplan en torbellinos alrededor de la roca, oscurecen el cielo con nubes y neblinas yconvierten en pesado e insoportable el ambiente; causan tanta humedad que todos los muebles se enmohecen, los
objetos de hierro crían orín…; por el contrario cuando soplan los vientos del oeste, el clima cambia de momento, elcielo queda sereno y despejado… El granizo, la nieve y el hielo no se conocen en Gibraltar, aunque se vean próximaslas Serranía de Ronda y montañas de Abila con sus cumbres nevadas de diciembre a marzo”.
Frederick Meyrich llega a comentar: “El día de mi llegada ceñía con un manto de vapor a la poderosa Roca., dejandoúnicamente a la vista aquí y allá hoscos pedazos de granito…”; y Richard Ford llegó a concretar algo más: “El clima esparticularmente malo para los niños durante la primera dentición; pero aparte de esto es sano. Es, sin embargoextremadamente desagradable cuando hay vientos del este, porque entonces cuelga sobre la cima de la Roca un vaporneblinoso y los nervios de las personas y los animales se ponen de punta”.
VI. LA CUEVA DE SAN MIGUEL Y OTRAS CAVIDADES
La cavidad correspondiente a la cueva de San Miguel, fue y sigue siendo uno de los atractivos naturales más sobresaliente deGibraltar. Raro fue el viajero que no la visitó y que realizara comentarios sobre la misma. Estas descripciones nos han servidopara evaluar su estado y la morfología que poseía en esos momentos, pero también han sido útiles para la obtención deinformación relativa a otras cuevas existentes, sobre otros elementos interesantes del karst así como sobre aspectos muycaracterísticos de la vida e historia del Peñón. De esta manera y siguiendo un orden de menor a mayor interés podemoscomenzar con los comentarios efectuados por W. Jacob, quien tan sólo nos legó datos sobre su origen y sus dimensiones: “Ellugar más interesante es la cueva de San Miguel hacia la mitad de la montaña. La entrada a esta cueva es una excavación naturalde treinta pies de ancho y veinticinco de altura. Esta llena de estalactitas de gran tamaño… no pudimos descender…”.
De W. G. Clark entresacamos: “Descendimos hasta la cueva de San Miguel, nos internamos hasta que el barro nos llegóa los tobillos y las gotas de agua tamborileaban sobre los sombreros en medio de la más absoluta oscuridad; decidimosretirarnos y salimos a la luz del sol”. El aspecto más interesante a destacar sería que este viajero la visita en época de verano.R. Ford anteriormente había afirmado: “Volviendo de este punto extremo (bahía Catalana), visítese la cueva de San Miguel,a mitad de camino según se sube por la roca. Entre las muchas cavernas de esta Calpe, o montaña socavada, está la llamada“Del bistec” o “Beefsteak cave”, por encima de las llanuras de Europa”. Aunque no describe pormenorizadamente la deSan Miguel, si da noticias de la existencia de otra cueva.
G. Borrow aporta datos sobre su estado natural y sobre su no utilización:
Me dirigí a la cueva de San Miguel; el camino hasta allí no está orientado en la misma dirección que el que conducea las excavaciones. Esta mira a España mientras que la cueva bosteza ante la faz de África. La entrada es una profundahendidura en la ladera de la montaña, de unos doce pies de alto y otro tanto de ancho. Conduce al interior de la caverna

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 17/276
Comunicaciones
17
una bajada muy en pendiente; Lo mas notable es una columna natural que se alza como el tronco de un enorme roble,como se debiese soportar el techo; el suelo es extremadamente escurridizo pues el continuo gotear del techo hasaturado por entero el terreno, de modo que toda precaución es poca para andar sobre él. Digan lo que digan loshombres con respecto a esta cueva, hay una cosa evidente a quienes se acercan a ella, que la mano del hombre no haterciado allí en absoluto. Algunos has asegurado que en tiempos paganos fue utilizada como templo del Dios Hércules.Baste con observar que en el interior de la cueva no existe nada que pudiera confirmar esta opinión, ni siquiera una
plataforma donde hubiera estado un altar. Por lo que he oído, he llegado a la conclusión de que toda la montaña deGibraltar es como un panal y estoy seguro de que si se profundizara, su interior resultaría estar lleno de abismosinfernales como los de la cueva de San Miguel.
La descripción de Frederick Meyrick es muy interesante, no exenta de elucubraciones; aparte de nombrar una nuevacavidad, evalúa la posible dimensión que alcanza la karstificación de estas calizas gibraltareñas, escribiendo:
Desde las galerías, un sendero empinado conduce a la cueva de San Miguel, un arco abierto en el apanalado interiorde la gran Roca lo bastante amplio como para permitir la entrada y pasear por el comienzo, pero que muy pronto, sehunde en el abismo como un pozo. Es de la misma naturaleza que la famosa gruta de Adelsberg cercana a Trieste. Unay otra vez los aventureros se han arriesgado intentando dentro de ella, encontrar un lugar para descansar; sin embargonunca encontraron más que el abismo que se sumerge quizás hasta el nivel del mar, o si hemos de dar crédito a las
Figura 2. Gibraltar, desde zona neutral, según David Roberts.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 18/276
Almoraima, 35, 2007
18
leyendas, todavía más adentro. Porque de acuerdo con la tradición, no es si no la entrada de un pasaje oculto que correa través de las cuevas de Don Daniel, bajo las raíces del mismo océano y emerge otra vez a la luz del día, entre lasmontañas de Ceuta, en el cielo ya de África.
Sería a través de este pasaje, por donde los monos se abrirían paso hacía África y volverían otra vez, a Europa. Es lacausa de que no se encuentre ninguno en España y por la que a veces, desaparecen de aquí, ya que en cualquiermomento, tiene la facilidad de retirarse con sus congéneres a la costa opuesta.
Termina diciendo que los monos entierran aquí a sus congéneres, y que “los musulmanes cuando llegue el día fijado porAla, encontrarán el pasaje submarino e irrumpirán bruscamente en la cueva de San Miguel, tomando posesión de nuevode la sacrosanta Córdoba, la encantadora Granada y la suntuosa Sevilla”.
Sacando tierra de Land Port, afirmaba F. Carter en 1775:
Han descubierto recientemente una cueva que penetra profundamente en el monte; encima de New Road hay otra muygrande, pero la mas sorprendente de todas es la que se conoce como cueva de San Jorge (o de San Miguel), a mildoscientos pies sobre el nivel del agua de las arenas rojas. Lleva dirección sur hasta el final de la roca, desciendo pocoa poco pero el paso está obstruido por grandes cantidades de piedra viva. La boca es estrecha por fuera pero muy ampliapor dentro y ofrece un retiro agradable; la gente viene a pasar el día aquí. Esta resbaladiza, hay goteras que formanlas cristalizaciones y carámbanos. Estos pilares petrificados aparecen con gran belleza y regularidad formando comoun precioso templo gótico, en las que se distinguen perfectamente naves y capillas asombrosas… Moros y españolesla usan como fortaleza.
Dedujo que antes se podía llegar hasta el final y que la caída de las piedras que lo impiden es muy reciente habiendoacontecido después de Claudio César.
Hans C. Andersen, viene a incidir en la misma cuestión:
Desde lo más alto del peñón, desde allí el camino de bajada pasaba por delante de una enorme gruta en la roca deinexplorada profundidad, cuyas caprichosas estalactitas y estalagmitas son de gran interés. Dice la leyenda, o escreencia popular, que la cueva continua por debajo del estrecho y tiene la salida al otro lado, en la costa africana; losmonos, se cree, pasaron por ese túnel submarino a Gibraltar. En la cueva se encuentran cantidad de esqueletos de esosanimales; los monos entierran ellos mismos a sus muertos.
Por ultimo habría que hacer destacar los comentarios de R. Twiss, quien visita este espacio en 1772, hace sobre otrosaspectos del karst como son las brechas y el relleno de las mismas. Afirmaba este viajero: “Al volar la roca con explosionesen varios lugares del peñón se descubren diariamente muchos trozos de huesos y dientes incrustados en piedras. Se hanenviado algunos de ellos a Inglaterra y han sido depositados en el Museo Británico”.
VII. DATOS FAUNÍSTICOS Y BOTÁNICOS
Fauna y vegetación también sirvieron como objetos de comentarios, la mayoría de ellos ligados a situaciones y formacionescuaternarias. Sin duda alguna la presencia del Macaco sylvana es el exponente máximo de lo que aquí queremos referiry viene a completar en cierta manera lo reseñado en líneas anteriores. Twiss nos trasmite: “En sus cavernas y precipicioshabitan muchos monos, a los que se caza frecuentemente. Se cree que esos animales no se dan en ningún otro lugar deEuropa. Aquí se encuentran los pájaros llamados gorriones solitarios”. “La forma y el aspecto del peñón no sonprometedores ni agradables, tan árido como tosco, sin árboles ni arbustos que asomen por encima de la ciudad; y no es por

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 19/276
Comunicaciones
19
aridez natural, si no por la política de nuestros militares… En mucho sitios abundan las piedras y en otros la roca viva,especialmente hacia el sur…”, comentaba F. Carter en sus escritos.
R. Ford se extiende algo más diciéndonos:
La superficie de la Roca, desnuda y parda en el verano, comienza a reverdecerse con la lluvia de la primavera y elotoño; más de trescientas plantas florecen en estas rocas casi sin tierra. Las codornices y los conejos abundan, y nadielos caza. Los verdaderos leones de Gib son los monos. Merodean por los lugares mas altos y se mueven como gamuzas;apenas se les ven salvo que el viento del oeste o el de levante los fuerce a ir al extremo occidental de la Roca. Robanen los jardines siempre que pueden, pero aparte de esto, viven de las raíces dulces de la palmita. Monsieur Bory, tuvola idea de que la gente vino también, como ellos, de África a España, y de España pasaron a Francia. Ahora bien, porlo que a España se refiere, estos monos están confinados a la Roca
William Jacob, cita una vez más la Colina de Los Monos en el continente africano, y agrega posteriormente: “La partesuperior de la roca de Gibraltar está formada por excelente piedra caliza con una base de granito. Las hendiduras de la rocade gran tamaño son un recurso idóneo para los monos, porque se ocultan cuando el viento sopla del este. A nadie se lespermite dispararle”. Con Gautier terminamos este tipo de comentarios:
Es el único sitio de nuestro continentes donde estos simpáticos cuadrúmanos viven y se multiplican en estado salvaje.Según cambia el viento, pasan de un lado a otro de la roca y sirven así de barómetro. Esta prohibido matarlos bajo penasmuy severas. Abila, si queremos dar crédito a su nombre moderno, debe gozar, en la costa de África, de una poblaciónsemejante.
VIII. CONCLUSIONES
Los distintos topónimos que los diferentes viajeros analizaron y aplicaron a Gibraltar, son portadores siempre deimportantes datos de naturaleza geográfica y natural, y resaltan siempre su alta significación histórica, su importancianatural y su personalidad en el ámbito geográfico del Estrecho y sur de la Península Ibérica. Se contempló este espacio comoun peñón aislado en el mar y posteriormente unido al continente por arenas, altamente karstificado y supuestamenteconectado bajo el mar con el norte de África. Realizaron comentarios sobre el comportamiento de las mareas, del marmediterráneo como mar interior cerrado y de su posterior apertura, de la entrada de una importante corriente de aguasubmarina, y de la existencia de una antigua playa levantada en su cara oriental, deduciendo con ello una edad muy recientede formación para el peñón.
La cueva de San Miguel fue ampliamente visitada y comentada deduciéndose su falta de acondicionamiento en esa época,
y su no anterior utilización por el hombre primitivo. Otras cavidades son citadas como interesantes así como la existenciade brechas calcáreas con abundante contenidos en huesos fosilizados. Desde el punto de vista faunístico hicieron señalarla existencia de una significativa población de conejos, la presencia de especies raras de aves y el carácter relicto de lapoblación de Macaco sylvana de Gibraltar; al mismo tiempo hicieron resaltar la existencia de otra importante poblaciónde esta especie en las elevaciones correspondientes al actual Gebel Muza en Marruecos (antigua colina de Los Monos oSierra Bullones).
De su color pardo y aspecto de león durmiente con que lo vieron, se puede deducir una gran falta de vegetación, presenciade escasos suelos y de abundantes afloramientos de roca en superficie. Las formaciones de arenas rojas cuaternariaslitorales fueron descritas en algunos lugares, así como descrita la morfología del istmo arenoso, donde se llega a notificarincluso que era utilizado como campo de golf.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 20/276
Almoraima, 35, 2007
20
IX. BIBLIOGRAFÍAALBERICH, José. 1976. Del Tamesis al Guadalquivir. Antología de viajeros ingleses en la Sevilla del siglo XIX . Public. Univer. de Sevilla. pp. 256.ANDERSEN, Hans Christian. Viaje por España. Alianza Editorial. 1988. Epílogo y notas de Marísa Rey. pp. 264. La primera edición de su libro fue I Spanien , 1863.BORROW, George. La Biblia en España . Edit. Biblioteca Grandes Viajeros. Prólogo de Emilio Soler Pascual. Traducción de Elena García Ortiz. 2001. pp. 541. Titulooriginal: The Bible in Sapin. 1842BOWLES, Guillermo. Introducción a la Historia Natural y a la Geografía Física de España . Imprenta Real de Madrid. 1789. pp. 554. Tercera edición.CARTER, Francis. Viaje de Gibraltar a Málaga. Edit. Arguval. pp. 336. Título original: A journey from Gibraltar to Málaga . 1777. Traducción de Christina Taylory José Antonio Olmedo López.CLARK, William George. Gazpacho o meses de verano en España. Edit. Horizontes. 1996. Traducción de Fernando Miranda. pp. 247. Titulo original: Gazpacho,o summer months in Spain. Londres. 1850.DORÉ, Gustavo; Davillier, Charles. Viaje por España. Edit. Grech. 1998. I y II: pp. 558 y 470.DUMAS Alexander. De París a Cádiz. Edit. Pre-Textos. 2002. Traducción de Ariel Dilon y Patricia Moinarrieta. pp. 592.FORD, Richard. Manual para viajeros por Andalucía y lectores en casa (1845). Edic.Turner 1988. Reino de Granada (pp. 183); Reino de Sevilla (pp. 356).GAUTIER, Théophile. Viaje a España. Edit. Cátedra. 1998. Edición y traducción de Jesús Cantera Ortiz de Urbina. pp. 384. Titulo original: Tras los montes. 1840.
GUERRERO, Ana Clara. 1990. Viajeros británicos en la España del siglo XVIII . Edit. Aguilar.JACOB, William. Viajes por el Sur. Cartas escritas entre 1809-1810. Introducción y traducción de Rocío Plaza Orellana. Edit. Portada. pp. 391. Título original:Travels in the South of Spain, in letters written a.d. 1809 and 1810. Londres. 1811.KRAUEL, Blanca. 2004. El itinerario andaluz de Washington Irving. En Antonio Garnica (Ed.). Washington Irving en Andalucía. Edit. Fundación José Manuel Lara.pp. 222.JIMÉNEZ, Antonio. 2002. La España pintoresca de David Roberts. El viaje y los grabados del pintor . Public. Univer.de Málaga. pp. 441.MEYRICK, Frederick. Cartas desde Andalucía. Estancia en Málaga y viaje por Andalucía del reverendo James Meyrick (1849-1851). Edit. Miramar. pp. 220.Traducción y estudio introductorio de Antonio Garrido Domínguez. Titulo original: The practical working of the Church of Spain. Oxford 1851.TOWNSEND, Joseph. Viaje por España en la época de Carlos III (1786-1787). Editorial Turner. Prólogo Ian Robertson. Traducción de Javier Portus. 1988. 454 pag.Primera publicación en 1791.TWISS, Richard. Viaje por España en 1773. Edit. Cátedra. 1999. Edición y traducción Miguel Delgado Yoldi. 287 pag. Primera edición en 1775: Travels throughPortugal and Spain in 1772 and 1773.VON HUMBOLT, Wilhelm. Diario de viaje a España, 1799-1800. Edit. Cátedra. 1998. Traducción de Miguel Ángel Vega. 260 pag.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 21/276
Comunicaciones
21
RESUMEN
En estos últimos años el término de San Roque no está siendo ajeno al auge de la construcción que asola gran parte de lacosta española. La creación de grandes complejos hoteleros, urbanizaciones y campos de golf principalmente, unido a laconstrucción de nuevas vías de comunicaciones, está transformando su paraje con los efectos irreversibles que ello puedeocasionar en su biocenosis, patrimonio de todos.
Estos cambios en el paraje inciden negativamente en la red hidrográfica. Así, en lugares en los que en la topografía recienteaparecen representados cursos de agua, muchos de ellos de carácter permanente, hoy día solo se encuentran lechos vacíosa causa de captaciones abusivas, desvíos, soterramientos, etc., cuando no son usados como meros colectores de aguasresiduales. La desaparición de estos cauces no solo supone la pérdida de biodiversidad propia de cada uno de ellos, sino
también, a mayor escala, a la del entorno por disrupciones en la cadena trófica.En este trabajo se enumeran algunos de los cauces alterados encontrados en diferentes puntos del término de San Roque comoconsecuencia de obras de infraestructuras públicas y otras de índole privado, y, considerando la riqueza biocenótica deltérmino, se intenta hacer una valoración de las consecuencias que dichas alteraciones pueden tener sobre su biodiversidad.
Palabras clave: red hidrográfica, biodiversidad, paraje, obras de infraestructuras.
CONSERVACIÓN FRENTE A DESARROLLO:¿HACIA UN DESARROLLOINSOSTENIBLE EN SAN ROQUE?
Alfonso Gallardo Mayenco / Eric Shaw

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 22/276
Almoraima, 35, 2007
22
1. INTRODUCCIÓN
A pesar de que en los años sesenta del pasado siglo comenzó el desarrollo industrial del término municipal de San Roque,básicamente localizado en la bahía de Algeciras, el resto del término ha mantenido un equilibrio aceptable entre eldesarrollo urbano y la conservación de su patrimonio natural, principalmente constituido por monte mediterráneo conformaciones vegetales de gran valor ecológico (alcornoque, acebuche, lentisco y bosque de laurel) y numerososendemismos, y una riqueza faunística no menos importante (guadiaro.html» www.nodo50.org/ecologistas.cadiz/EACadiz/ sitioEACadiz/guadiaro.html; Agenda 21 Local de San Roque). Sin embargo, en los últimos años San Roque no ha sidoajeno al desarrollo urbanístico que asola gran parte de la costa española. La construcción y ampliación de grandes complejoshoteleros, urbanizaciones y campos de golf en el término está alterando de tal forma el paraje, por la destrucción del montey su sustitución por parajes artificiales, que sus consecuencias sobre la biocenosis podrían ser irreparables.
A raíz de un estudio del primer autor de este trabajo sobre la diversidad de macroinvertebrados acuáticos de San Roque,se está haciendo una serie de prospecciones en la red hidrográfica del municipio que están proporcionando algunainformación sobre cómo estos cambios en el paraje están influyendo en dicha red. Así, se han observado cauces secos dondedebería circular agua según la cartografía, o en los que circulan aguas residuales porque están siendo utilizados comocolectores. En cualquier caso, la desaparición o eutrofización de estos cauces no solo supondría la pérdida de labiodiversidad propia de cada uno de ellos, sino también parte de la del entorno por disrupciones en la cadena trófica.
En este trabajo se pretende hacer una valoración preliminar de la situación de parte de la red hidrográfica de San Roque.Para ello, se enumeran los cauces estudiados detallando los que presentan algún tipo de alteración y las posibles causas delas mismas. Por último, se exponen a grosso modo las amenazas que se ciernen sobre el paraje del municipio y sus cauces.
2. EL MUNICIPIO
El municipio de San Roque con unos 140km2 de extensión, comenzaba el año 2003 con una población de algo más de 25.000habitantes y había experimentado en el periodo comprendido entre 1996 y 2003 un incremento de población del 13,5%(www.juntadeandalucia.es/institutodeestadistica/sima/htm/sm11033.htm), estimándose que para el periodo 2002-2012San Roque contribuirá con un 43% al incremento de población en el conjunto del Campo de Gibraltar, siendo solo superada,según estas estimaciones, por el municipio de Los Barrios (VV.AA., 2004).
Las grandes industrias de la zona se localizan entre el área industrial de Puente Mayorga y el polígono industrial deGuadarranque, en torno a la bahía de Algeciras, mientras que el resto del término está ocupado por núcleos urbanos,complejos deportivos, hoteleros y residenciales, zona de cultivos y grandes extensiones de monte mediterráneo, además
de unos diez kilómetros de playa que se reparten casi al 50% entre playas urbanas y no urbanas (VV.AA., 2004).
3. EL DESARROLLO: EL IMPACTO
El caudal de aguas residuales producidas en el municipio de San Roque en el año 2002 fue de 5,89 Hm3, tratadas por cincoEDARs. Análisis efectuados a sus respectivos efluentes en el año 2002 mostraron que su funcionamiento no era correctosegún el decreto 509/1996 que regula los requisitos mínimos de calidad de estos efluentes (VV.AA., 2004). Por otra parte,los habitantes-equivalentes calculados para el municipio de San Roque superan los calculados para el diseño de las EDARsen conjunto. Existen además otras EDARs de propiedad privada de menor capacidad de depuración (para menos de 5.000habitantes-equivalentes) como la del Casino Nuevo de San Roque que vierte al arroyo de la Mujer, la de Asahi Kanko (San

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 23/276
Comunicaciones
23
Figura 1. Situación de los tramos visitados en la red hidrográfica del municipio de San Roque: 1 arroyo de la Mujer, 2 río Cachón, 3 arroyo de Borondo, 4 arroyode Punta Chullera, 5 arroyo de Mantilla, 6 arroyo de los Cortijos, 7 arroyo de Guadalquitón, 8 arroyo de Alaha, 9 arroyo de Cañuelo, 10 río Guadiaro. Hoja 14-47(1075) de la cartografía militar de España (Escala 1:50.000 reducida al 55%).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 24/276
Almoraima, 35, 2007
24
Roque Club) que vierte al arroyo de Borondo, de La Alcaidesa al arroyo de la Bea, y la de (ABB) Alstom Power Españaque vierte al arroyo de Madre Vieja, afluente del río Guadarranque (VV.AA., 2004).
Asociados a algunos complejos hoteleros y residenciales, se encuentran en el término varios campos de golf que ocupan unaextensión variable dependiendo del número de hoyos: Almenara (27 hoyos), San Roque Club (dos campos de 18 hoyos cadauno), Real Club de Golf de Sotogrande (18 hoyos), La Reserva de Sotogrande Golf (18 hoyos), Valderrama (18 hoyos), LaCañada (18 hoyos), y La Alcaidesa (18 hoyos). Este último, con un nuevo campo previsto para 2005, no pertenece a San Roquepero se encuentra en sus límites. Posiblemente sea el consumo de agua uno de los aspectos que más críticas reciben los camposde golf. Sin embargo, algunos de ellos utilizan lagos de captación (Almenara), pozos y agua reutilizada (San Roque Club,Alcaidesa, Sotogrande, La Cañada), y en menor proporción agua procedente de la red general (Alcaidesa y La Cañada). Deotros campos no se tienen datos. En cualquier caso, el consumo de agua de estos campos (sin considerar la procedente de lagos
de captación y reutilizada) se estima en unos 1,09 Hm3/año, lo que supone aproximadamente el 20% del agua consumida parariego en todo el Campo de Gibraltar en el año 2002 (VV.AA., 2004), aunque estos valores deben ser ajustados en la actualidadpor la ampliación y construcción de nuevos campos. Para valorar el papel que juegan en la economía de San Roque estoscampos habría que tener en consideración que el gasto medio diario por turista que practica golf era en el año 2002 de unos180 euros, según estimaciones de la Federación Andaluza de Golf (VV.AA., 2004).
Paralelo a estos campos de golf se amplían los complejos hoteleros y crecen las urbanizaciones de tipo residencial, perotambién se amplían los núcleos urbanos y se construyen nuevas urbanizaciones por necesidades de crecimiento de lapoblación y por el incremento de un turismo atraído por el clima y el entorno natural del término. Este turismo es en muchoscasos temporal (de verano) aunque cada vez es más importante el asentamiento de turistas procedentes, generalmente, depaíses europeos con alto poder adquisitivo que fija aquí su residencia gran parte del año, sino toda. No disponemos de datos
referentes a la extensión construida en los últimos años, pero los procedentes de las principales actividades empresarialespor número de empleados podría ser significativo, de manera que la construcción y las actividades inmobiliarias y dealquiler, están entre las cuatro actividades que ocupan mayor número de empleos en el término (VV.AA., 2004).
4. LA CONSERVACIÓN: LA UTOPÍA
El estuario del río Guadiaro, las sierras Carbonera, del Arca y Almenara, las dunas de Sotogrande, la laguna deTorreguadiaro y las playas son algunos ejemplos de la diversidad paisajística contenida en el término de San Roque. Elestuario del río Guadiaro está protegido con la figura de Paraje Natural, la única del municipio, y ha sido declarada Zonade Especial Protección para las Aves y Lugar de Interés Comunitario. Otra figura de protección es el Plan Especial deProtección del Medio Físico y Catálogo de la Provincia de Cádiz, que atañe al Campo de Gibraltar en cuanto a la figurade Protección Especial Compatible y que se refiere a las zonas en las que se limita la realización de actividades constructivaso transformadoras del medio. Por otra parte, el Plan de Ordenación del Territorio de la Comarca del Campo de Gibraltaraborda la preservación y mejora del medio natural y del paraje de manera que en lo concerniente a San Roque supone algúntipo de protección para la playa de Guadalquitón (como paraje litoral singular), las playas de Guadarranque y punta Mala(como paraje litoral de interés), las sierras del Arca y Carbonera (como parajes de valor ambiental) y Pinar del Rey (comoparaje forestal singular). Por otra parte, el plan urbanístico municipal también incluye algunos lugares de protección(Agenda 21 Local de San Roque).
Todo este conjunto natural concentra una extraordinaria riqueza florística y faunística entre la que destaca el laurel ( Laurusnobilis) y el quejigo (Quercus canadiensis), ambas especies catalogadas como vulnerables por la Junta de Andalucía y la

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 25/276
Comunicaciones
25
UICN, además de la Sideritis perezlarae, catalogada como vulnerable por la Junta de Andalucía y en peligro por la UICN.Además, la conservación del quejigo, por su cubierta protectora, es esencial para garantizar la de otras especies amenazadasasociadas al quejigal (Sánchez García, 2002). Para L. nobilis y S. perezlarae hay que ponderar su presencia para cualquiertipo de actividad que se vaya a realizar allá donde se encuentre, además S. perezlarae es un endemismo exclusivo del litoralgaditano que suele encontrarse en sitios de interés urbanístico, lo que ha contribuido a la desaparición de varias de suspoblaciones (Consejería de Medio Ambiente, 2000). Estas especies junto a otras como el carrizo, castañuela, juncos,tarajes, acebuches, lentisco, zarzales, fresnos, alisos, chopos, alcornoques, ciperáceas, aulagas, herguene, acompañados deuna importante flora endémica (Euphorbia baetica, Genista tridens, Ononis leucotricha, Stauracanthus genistoides)(www.ecologistasenaccion.org/article.php3?id_article=1967), constituyen solo una parte de la cobertura vegetal que sepuede encontrar en estos parajes, y que dan cobijo a una fauna de vertebrados entre la que se puede encontrar elementos
de indudable interés conservativo algunos de ellos en una situación frágil de conservación como el águila pescadora, extintaa nivel de población reproductora y vulnerable como población invernante en Andalucía y en peligro de extinción en elresto de España, espátula y garza imperial, vulnerables en Andalucía y España, avetorillo, vulnerable en Andalucía, vencejocafre, vulnerable en Andalucía y rara en España, (aquí habría que considerar que el estuario del río Guadiaro se sitúa enuna de las rutas migradoras más importantes), el camaleón, en peligro de extinción en España y con un riesgo menor enAndalucía, y grandes herbívoros como el ciervo, muflón y jabalí, y pequeños predadores del bosque y matorralmediterráneo como el tejón, meloncillo, gineta, zorro y nutria, esta última vulnerable en Andalucía y España (Consejeríade Medio Ambiente, 2001). En la Agenda 21 Local de San Roque se encuentra un listado muy completo de la flora y fauna,con sus correspondientes grados de amenaza.
Figura 2. Una imagen que está siendo habitual en San Roque. En la foto obras de desmonte en Borondo en mayo de 2005.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 26/276
Almoraima, 35, 2007
26
El interés por la conservación de tan rico binomio biotopo-biocenosis ha llevado a propuestas como la de creación de uncorredor verde (concepto de reciente actualidad tras el desastre de Aznalcóllar y posterior proyecto de recuperación del ríoGuadiamar) en el estuario del río Guadiaro-Playa de Guadalquitón con el Parque Natural Los Alcornocales para que seincluya en la ley de espacios protegidos de Andalucía (guadiaro.html» www.nodo50.org/ecologistas.cadiz/EACadiz/ sitioEACadiz/guadiaro.html). Esta, sin duda, loable idea que engloba la conservación de numerosas fincas tanto de índoleprivado como público y que afecta a unas 2.500 Ha, más las correspondientes al Pinar del Rey, según los datosproporcionados por sus valedores (Ecologistas en acción), supondría la conservación de un monte mediterráneo con unariqueza florística y faunística inigualable, en palabras de sus promotores y, también sin duda, en las nuestras.
5. EL DESEQUILIBRIO: LA AMENAZA
A pesar de las varias figuras de protección son muchas las amenazas que con distinta magnitud se ciernen sobre los espaciosnaturales de San Roque. Sirvan como ejemplo las dunas de Sotogrande donde el desarrollo turístico amenaza su perdurabi-lidad; los incendios, el desarrollo urbano sin control y la presión ganadera en Sierra Carbonera; el desarrollo urbanístico deSotogrande y la proliferación de los campos de golf en sierra Almenara, y el mismo Paraje Natural del Estuario del río Guadiarodonde el uso público intensivo, la presión urbanística, la invasión de especies vegetales alóctonas y la eutrofización hacen queun 83% de su extensión esté amenazada en grado muy alto-alto (Agenda 21 Local de San Roque).
Para estudiar la biodiversidad de macroinvertebrados y tipificar la calidad de las aguas en el municipio de San Roque, elprimer autor está realizando una prospección en su red hidrográfica. En la revisión bibliográfica previa al estudio (Gallardo-Mayenco, 2002, 2005), no se ha encontrado información alguna respecto a estos organismos en el municipio, por lo que
se desconoce la riqueza y calidad biológica de sus cauces. El primer paso para esta clase de estudios consiste en seleccionarlos cauces que, a priori, podrían resultar más interesantes para muestrear utilizando para ello la cartografía. Para este estudiose ha utilizado la hoja 14-47 (1.075) San Roque de la serie L, escala 1:50.000 de la cartografía militar de España, en la queaparecen representados multitud de arroyos tanto de carácter permanente como temporal o semipermanente. Sin embargo,el contacto directo en el campo ha mostrado otra realidad, puesto que muchos de estos cauces cartografiados han sidoalterados cuando no sólo queda de ellos el lecho seco. Así, de los diez cauces visitados (figura 1) tres estaban secos (arroyosde la Mujer, Cachón en la playa de Guadalquitón y Borondo), dos habían sufrido alteraciones a consecuencia de obras deinfraestructuras (arroyos de Punta Chullera y Mantilla), uno tenía el cauce regulado para regadíos (arroyo de los Cortijos)y dos parecen estar contaminados por aguas residuales (arroyos de Guadalquitón y Alaha). Solo el arroyo de Cañuelo y elrío Guadiaro parecen estar libres de alguna perturbación. La descripción de los sitios en los que se ha encontrado algunaperturbación es la siguiente:
Arroyo de la Mujer a la altura del Km 122 de la E15 (30STF881121). En la cartografía aparece representado como arroyotemporal o semipermanente, sin embargo en todas las visitas estaba seco, excepto en abril de 2005 que se observaronalgunas charcas que posiblemente tenían su origen en los vertidos del Casino de San Roque (quizás su EDAR vierta en esepunto) y en una explotación agraria establecida en la orilla opuesta, ya que aguas arriba el cauce estaba completamente seco.Mucho más arriba del tramo observado se aprecia lo que parecen movimientos de tierra en el monte.
Río Cachón poco después de su unión con el arroyo de Guadalquitón en la playa de Guadalquitón (30STF942153). En lacartografía se representa como arroyo temporal o semipermanente, se encontró seco en una visita en mayo de 2005, sinembargo hay que señalar que el Guadalquitón llevaba en esa misma fecha un caudal apreciable.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 27/276
Comunicaciones
27
Arroyo de Borondo a la altura del Cortijo de Borondo (30STF913148). También representado como temporal osemipermanente, en mayo se encontró seco. Debemos reseñar que en este arroyo vierte la EDAR de San Roque Club aguasmás arriba de donde se ha visitado. No obstante, lo más remarcable es la obra de desmonte que se está llevando a cabo enlos alrededores del tramo visitado (foto 1).
Arroyo de Punta Chullera a la altura del Km 137 de la E15 (30STF972211). En la cartografía se representa un cauce dearroyada que viene a desembocar a la playa de Chullera. Esta arroyada está entubada y soterrada para así salvar la autovía,quedando en el lado opuesto de la autovía, en lo que era el inicio de su desembocadura en la playa, una especie de lagunacubierta por vegetación palustre.
Arroyo de Mantilla a unos 300 metros en la autovía dirección Málaga por detrás de Torreguadiaro (30STF954199). Elcauce está soterrado para salvar la carretera y cuando se visitó estaba seco aunque su curso quedaba perfilado por una densacubierta de Phragmites sp. por lo que se supone que bajo el lecho debía circular agua, por lo que sería interesante ver susituación en la época de lluvias.
Arroyo de los Cortijos a la altura del Km 12 de la carretera entre San Enrique y San Martín del Tesorillo (30STF926222).Este arroyo está regulado por compuertas para el regadío de los cultivos circundantes. De hecho, el tramo estudiado seríaen realidad una acequia alimentada por este arroyo. En la visita realizada en julio a este punto se observó que el cauce estabaseco porque el paso del agua desde el arroyo estaba cerrado, también se observó que este arroyo abastece a otras acequias,estando por lo tanto su caudal regulado.
Arroyo de Guadalquitón a la altura del Km 128 de la carretera auxiliar de la E15 (30STF920158). Aparece como de cursotemporal o semipermanente en la cartografía, sin embargo el seguimiento intensivo que se le está haciendo (un muestreo
mensual) permite afirmar que lleva agua todo el año y con un caudal importante para tratarse de un arroyo relativamentepequeño en el tramo muestreado. Después de algunos muestreos se puede concluir que contiene una comunidad aceptable,detectándose la presencia de Anguilla anguilla, especie con estatus de vulnerable en España aunque con un riesgo menoren Andalucía (Consejería de Medio Ambiente, 2001), que tiene sus principales amenazas en la pesca abusiva, laconstrucción de embalses y la contaminación de los estuarios (Doadrio, 2001). Presenta una abundante vegetación de riberay acuática, destacando en algunas zonas de la orilla por su abundancia Mentha sp. y una desarrollada población deEquisetum telmateia en la zona más sombría del tramo estudiado, y el lecho está formado principalmente por piedras y arenagruesa. No obstante, las poblaciones de macroinvertebrados que dominan por su densidad al resto de la comunidad estáconstituida por tricópteros del género Hydropsyche y simúlidos, grupos que tienen como base alimenticia a la materiaorgánica particulada arrastrada por la corriente. También es significativo que después de nueve muestreos no se ha recogidoni un solo ejemplar de plecópteros, por lo que de todo ello se desprende que muy probablemente este arroyo esté recibiendo
una carga importante de aguas residuales con alto contenido orgánico. Otra agresión que está sufriendo este arroyo en eltramo estudiado es el paso de maquinaria pesada por su cauce, en la zona correspondiente a su paso por el puente de lacarretera y aguas abajo, con la consiguiente destrucción de vegetación de ribera y acuática, de hecho parte de la poblaciónde Equisetum telmateia desapareció, y la alteración del lecho por remoción del sustrato, afectando directamente a lacomunidad de macroinvertebrados.
Arroyo de Alaha en el Pinar del Rey a la altura de la fuente (30STF844125). Presenta en el tramo estudiado un cauceabierto, sin cobertura vegetal, por lo que soporta una importante insolación, con un sustrato formado por arenas gruesasy piedras pequeñas y un aporte de agua continuo procedente de la fuente que se mezcla con la que llega desde aguas arriba.Aquí, los resultados preliminares muestran una comunidad extraordinariamente pobre que no está en consonancia con loesperado según la estructura del hábitat. Posterior a los muestreos, por medio de algunos vecinos de la zona, se supo que

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 28/276
Almoraima, 35, 2007
28
en la zona del Albarracín, aguas arriba de este arroyo, existe un importante núcleo de población (de aproximadamente unasquinientas fincas) que al carecer de red de saneamiento vierten sus aguas residuales a pozos ciegos, por lo que éstas bienvía aguas subterráneas o directamente, llegan al Alaha. En este caso, la contaminación orgánica no solo afecta al arroyo,sino también a los acuíferos.
Los primeros resultados del estudio de biodiversidad de macroinvertebrados acuáticos en el municipio parecen indicar quela comunidad biológica de los arroyos de Guadalquitón y Alaha están en transición hacia la degradación puesta demanifiesto por Allan (2004) como respuesta al incremento de suelo urbanizado.
Aunque no forma parte del estudio de la red hidrográfica, debemos señalar aquí la situación de la laguna de Torreguadiaro.Su visita en febrero de 2005 solo sirvió para constatar el estado de abandono y decrepitud de unas instalaciones que sesupone existen para concienciar al ciudadano en el respeto al medio. Parte de las instalaciones mostraban los restos de unfuego, seguramente provocado al amparo de este abandono.
6. CONCLUSIONES: ¿HACIA UN DESARROLLO INSOSTENIBLE?
Respecto a la red hidrográfica en el municipio de San Roque se ha podido constatar hasta el momento que de diez caucesvisitados (un tramo del río Guadiaro y nueve arroyos, figura 1), tres estaban secos, supuestamente por extracción de agua,dos presentaban el tramo alterado por obras de infraestructuras, dos parecían sufrir descargas de aguas contaminadas pormateria orgánica y uno estaba regulado para su aprovechamiento para regadío, es decir el 80% de los cauces, en sus tramosestudiados, presentaban alguna clase de perturbación que potencialmente incidiría negativamente sobre su biocenosis. Ladesaparición o degradación de los cauces, o tramos de ellos, repercute negativamente en el flujo de la materia y energía.
Los macroinvertebrados acuáticos ocupan todos los eslabones de la cadena trófica, interviniendo en la disgregación de lamateria orgánica descompuesta y en el reciclaje de la materia orgánica disuelta, actuando por tanto como organismosintermedios en la circulación de esta materia y energía hacia elementos situados en niveles superiores de la cadena, por loque la pérdida de estos organismos por desaparición de sus hábitats conlleva a la disrupción del flujo de la materia y energíaen el área donde el cauce ha sido alterado, siendo las especies de anfibios, reptiles, aves y pequeños mamíferos que incluyenen su dieta a estos macroinvertebrados los últimos damnificados por esta ruptura de la cadena.
A una escala mayor, con tal diversidad de parajes en una extensión relativamente tan pequeña, sobre la disyuntiva planteadaen el título de este trabajo cabe preguntarse, ¿qué sentido se ha elegido para el municipio de San Roque? De lo visto hasta ahorasu presente es preocupante: parajes naturales sustituidos por otros artificiales, crecimiento urbanístico desmesurado, montesque desaparecen, cauces que desaparecen o son contaminados, poblaciones animales y vegetales que desaparecen con sus
hábitats. Pero su futuro no parece que vaya a ser mejor: se prevee la construcción de más campos de golf, aunque desde nuestropunto de vista más que los campos en sí lo preocupante son las urbanizaciones y complejos hoteleros que llevan asociados.El ayuntamiento de San Roque pretende urbanizar el área de Guadalquitón, amenazando con ello un bosque mediterráneocostero con consideración de hábitat natural de interés comunitario (los alcornocales de Quercus suber ), (id_article= 1967"www.ecologistasenaccion.org/article.php3?idarticle= 1967), y los fuegos siguen asolando Sierra Carbonera. Más aún, si hoylas depuradoras son insuficientes es fácil predecir que ocurrirá en los próximos años cuando el incremento de poblaciónprevisto sea una realidad junto a la presión turística pareja a los nuevos campos de golf, urbanizaciones e instalacioneshoteleras. Por otra parte, a consecuencia del cambio climático, se predice una disminución de la precipitación que podríaresultar en cambios importantes en el paraje forestal mediterráneo (Jurado Doña, 2002).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 29/276
Comunicaciones
29
Con respecto a la red hidrográfica, encontramos en el municipio la existencia de un eje agricultura-desarrollo urbanísticoen consonancia con el uso del suelo. Así, donde los cauces constituyen un recurso para la agricultura éstos se regulan, ocuando son absorbidos por el crecimiento urbanístico se utilizan como cloacas gratuitas, se les extrae el agua hasta secarlos,o se soterran.
Parece que desde algunos sectores se está alentando un desarrollo, a imitación del practicado por otros municipios, que noes en absoluto alentador, quizás por las expectativas de creación de puestos de trabajo, desarrollo económico y bienestar.Sin embargo, la pérdida del patrimonio y de la propia identidad natural podría en un plazo de tiempo indeterminado volverseen contra de los argumentos que hoy en día la patrocinan, en cuyo caso cabría preguntarnos que ocurrirá cuando, además,patrimonio e identidad natural sean irrecuperables.
BIBLIOGRAFÍAAGENDA 21 LOCAL DE SAN ROQUE. “Avance del documento de diagnóstico ambiental de la Agenda 21 Local de San Roque”, (disponible en www.sanroque.es/ portal/).ALLAN, J. D. “Influence of land use and landscape setting on the ecological status of rivers”, Limnetica, 23 (2004), pp. 187-198.CONSEJERÍA DE MEDIO AMBIENTE (Editor). Libro Rojo de la Flora Silvestre Amenazada de Andalucía. Tomo II: Especies Vulnerables, Junta de Andalucía (2000).CONSEJERÍA DE MEDIO AMBIENTE (Editor). Libro Rojo de los Vertebrados Amenazados de Andalucía, Junta de Andalucía (2001).DOADRIO, I. (Editor). “Atlas y libro rojo de los peces continentales de España”, Dirección General de Conservación de la Naturaleza, Museo Nacional de CienciasNaturales, Madrid, (2001).GALLARDO-MAYENCO, A. “Macroinvertebrados acuáticos de la red hidrográfica del Campo de Gibraltar: una revisión”, Almoraima, 27 (2002), pp. 351-364.GALLARDO-MAYENCO, A. “Nuevas aportaciones al censo de macroinvertebrados acuáticos de la red hidrográfica del Campo de Gibraltar”, Almoraima , 31 (2005),pp. 277-285.JURADO DOÑA, V. Los bosques de las Sierras del Algibe y del Campo de Gibraltar (Cádiz-Málaga). Ecología transformaciones histórcias y gestión forestal,Consejería de Medio Ambiente, Junta de Andalucía (2002).SÁNCHEZ GARCÍA, J. M. “Los bosques de quejigo (Quercus canariensis Willd.) de las Sierras del Aljibe y Campo de Gibraltar: manejo histórico, estado selvícolay propuesta para su gestión”, Almoraima, 27 (2002), pp. 281-292.VV.AA. Estudio de la Calidad Ambiental del Campo de Gibraltar. Medio ambiente acuático y contaminación acústica, Coordinado por el Grupo de Investigaciónde Tecnologías del Medio Ambiente de la Universidad de Cádiz. Segundo informe, revisado y corregido (2004).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 30/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 31/276
Comunicaciones
31
RESUMEN
Desde mediados del siglo XIX hasta la actualidad la fauna de peces, aves y mamíferos del Campo de Gibraltar y su áreade influencia ha perdido al menos 32 especies, muchas de ellas extintas completamente. El grupo más afectado por estas
regresiones es el de las rapaces diurnas ( Accipitridae) al que corresponde el 45% de las pérdidas. Igualmente se ha detectadola incorporación o el aumento significativo de nueve especies de aves y un mamífero. En la mayoría de los casos la actividadantrópica parece ser la responsable de estos cambios faunísticos. Especialmente la desecación de los humedales de La Jandapudo ser el factor primario responsable de la extinción de al menos 17 especies (el 53% de todas las sometidas a regresión).El cambio climático también parece estar ejerciendo una influencia importante sobre la desaparición de especies depreferencias más frías y de la incorporación de otras con hábitos más meridionales.
Palabras clave: Incorporaciones, extinciones, aves, laguna de La Janda, sur de España, cambio climático, Campo deGibraltar, análisis histórico.
CAMBIOS EN LA FAUNA DEVERTEBRADOS DELCAMPO DE GIBRALTAR DESDEMEDIADOS DEL SIGLO XIXHASTA LA ACTUALIDAD
Arturo Menor / José PrendaDepartamento de Biología Ambiental y Salud Pública. Universidad de Huelva

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 32/276
Almoraima, 35, 2007
32
INTRODUCCIÓN
En los últimos tiempos se están produciendo importantes cambios en la biosfera motivados por la actividad humana.Especialmente desde la Revolución Industrial el hombre está transformando drásticamente los hábitats naturales,sobreexplotando las especies, diseminando organismos fuera de sus ámbitos naturales de distribución y provocandocambios ecológicos sin precedentes que promueven extinciones en cadena (Delibes, 2004). Todo ello se traduce enacusadas modificaciones –extinciones y nuevas incorporaciones o sustituciones– en las comunidades naturales. Estefenómeno, de ámbito planetario, puede ser fácilmente seguido a escala local, donde a la vez sería posible, no solo constatar,sino estimar la magnitud real de los cambios acaecidos. Para ello es imprescindible contar con fuentes de informaciónfidedignas que describan la situación de la biodiversidad en condiciones lo menos perturbadas posible, con las quecomparar la situación observada en la actualidad.
La comarca del Campo de Gibraltar, debido a su particular posición geográfica, entre los continentes europeo y africanoy entre el océano Atlántico y el mar Mediterráneo, es un medio idóneo para evaluar estos cambios. Se trata de una zonadonde coexisten áreas profundamente transformadas por el hombre con otras relativamente poco modificadas (Dueñas yRecio, 2000; Clavero et al., 2005). Además es una región de transición climática, donde las predicciones indican que severá fuertemente afectada por el cambio climático y por las especies invasoras (Lavorel et al., 1998, Sala et al., 2000).
En este trabajo se analizan los cambios acaecidos en la fauna de vertebrados del Campo de Gibraltar desde mediados delsiglo XIX hasta la actualidad. Para ello se elabora el catálogo de las especies citadas en fuentes históricas que o bien se hanextinguido localmente, o bien han podido sufrir modificaciones sustanciales en su estatus en la comarca, y se compara conla situación que se observa hoy en día. De este modo es posible realizar una aproximación cualitativa y cuantitativa a lasdimensiones de la transformación ambiental impulsada por el hombre contemporáneo en este enclave natural privilegiado.
METODOLOGÍA
Para la evaluación de los cambios faunísticos se ha recurrido a fuentes bibliográficas pretéritas, fundamentalmente del sigloXIX (tabla 1). La documentación escrita fidedigna existente se refiere fundamentalmente a vertebrados, bien conocidosdesde el inicio de los estudios naturalistas, y con no demasiados problemas taxonómicos que dificulten su interpretación,como podría ocurrir con otros grupos de organismos. La mayor parte de la información se refiere a aves y en menor medidaa mamíferos. Ambos grupos faunísticos son los que tradicionalmente más interés han despertado entre los naturalistas yson también los más fáciles de estudiar/colectar, especialmente las aves. Por contra, no se han encontrado apenasreferencias a anfibios y reptiles y por ello han sido obviados de este estudio.
En el caso particular de la comarca del Campo de Gibraltar, además, existe una buena disponibilidad de datos faunísticosantiguos debido a la presencia histórica de naturalistas británicos afincados en la zona (por ejemplo Irby), así como en elresto de la provincia de Cádiz, y a la labor de una de las figuras decimonónicas más importantes e influyentes de las cienciasnaturales de España, el gaditano Antonio Machado y Núñez (Menor y Prenda, 2004). Las publicaciones de unos y otrosatrajeron a su vez el interés de otros naturalistas españoles, como López Seoane, que también visitaron la zona y dejaronconstancia de sus valores naturales (tabla 1).
Esta información ha sido completada con las citas faunísticas aparecidas en el Diccionario Geográfico-Estadístico- Histórico de España de Pascual Madoz (1845-1850).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 33/276
Comunicaciones
33
Pueden existir dudas en relación a la validez de las identificaciones realizadas por estos naturalistas decimonónicos, yalgunos autores contemporáneos así lo han manifestado en alguna ocasión. No obstante, la gran mayoría de las especiesde vertebrados de Europa fueron descritas en los siglos XVIII y XIX por algunos de los naturalistas aquí considerados opor colegas suyos. La taxonomía vigente es la que ellos crearon. Además, normalmente trabajaban con ejemplarescapturados –en mano–, lo que deja muy pocas dudas respecto a su correcta identificación.
Para los datos actuales se ha compilado la información disponible en los atlas de mamíferos y aves recientementepublicados por el Ministerio de Medio Ambiente (Palomo y Gisbert, 2002; Martí y Del Moral, 2003).
A efecto de su estatus en el Campo de Gibraltar las especies se clasificarán en seis categorías: 1) extintas, 2) extinguidascomo reproductoras o invernantes; 3) virtualmente extinguidas como reproductoras o invernantes; 4) especies cuyo númerode ejemplares reproductores se ha visto significativamente reducido; 5) especies nuevas para la fauna de la zona de estudio,no introducidas por el hombre; 6) especies que han experimentado un aumento en el número de individuos.
ÁREA DE ESTUDIO
El área de estudio lo conforman las siguientes localidades integrantes de la comarca del Campo de Gibraltar: Algeciras,Castellar de la Frontera, Gibraltar, Jimena de la Frontera, La Línea de la Concepción, Los Barrios, San Roque y Tarifa. Porsu proximidad geográfica y similitud ambiental se han tenido en cuenta también los municipios de Barbate, Vejer de laFrontera, Benalup –Casas Viejas– y Alcalá de los Gazules.
En el caso particular de La Línea de la Concepción, los datos más antiguos son prácticamente inexistentes debido a quese hallaba integrada en el municipio de San Roque, no siendo hasta la segunda mitad del siglo XIX, tras su segregación,cuando aparecen las primeras referencias zoológicas de esta localidad.
Autor Años Grupo faunístico Zona geográficaMadoz 1845-1850 vertebrados y otros EspañaMachado 1854-1869 peces, aves y mamíferos AndalucíaLópez Seoane 1861-1870 aves AndalucíaSaunders 1877 aves sur de EspañaMartínez Reguera 1881 mamíferos Montoro (Sierra Morena)Sánchez y García 1885 mamíferos y aves GranadaIrby 1895 aves Campo de GibraltarChapman y Buck 1893 vertebrados y otros EspañaGraells 1897 mamíferos EspañaBernaldo de Quirós 1920 aves La JandaYeates 1946 aves Doñana
Tabla 1. Referencias bibliográficas antiguas utilizadas para confeccionar la lista de vertebrados presentes en el Campo de Gibraltar en condicionesambientales no modificadas sustancialmente por la actividad humana. Se indica el año de publicación de los trabajos, el grupo abordado y la zona geográficaa la que se refiere principalmente.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 34/276
Almoraima, 35, 2007
34
RESULTADOS Y DISCUSIÓN
En los últimos ciento cincuenta años se han extinguido como reproductores o invernantes en el Campo de Gibraltar y suentorno inmediato al menos veinticinco taxones (tabla 2). A ellos habría que añadir otros siete más que se consideranprácticamente extinguidos en esta zona, lo que elevaría a treinta y dos el número de especies cuyo estatus se ha modificadosustancialmente.
Si se asume que la riqueza de especies de vertebrados epicontinentales del área de estudio en el siglo XIX fue similar a laactual, esto es alrededor de 450, al menos el 8% de la riqueza original habría desaparecido. Esto sin contar al menos otrosseis taxones que han visto reducidos significativamente sus efectivos en la zona. Hay que tener en cuenta, además, que elconocimiento que se tiene del pasado es parcial e incompleto (en el siglo XIX no se acometían muestreos sistemáticos ycompletos de la fauna), y que aquí no se consideran ni anfibios, ni reptiles, ni virtualmente los peces continentales. Es porello que la estima del 8% de pérdidas está claramente infravalorando la cifra real de extinciones en el Campo de Gibraltaren los últimos 150 años.
Por familias, la más afectada fue la de las rapaces diurnas ( Accipitridae), que incluyó al 45% de las especies cuyo estatuscambió en el área. Esta familia se muestra especialmente sensible a los cambios ambientales inducidos por la actividad yademás es también la mejor conocida. Le siguieron en importancia, los rálidos (cuatro especies, 13,3%) y las anátidas ysílvidos, ambas con tres especies y un 10,0% del total de especies consideradas (tabla 2).
Es importante considerar que de las 32 especies de aves y mamíferos que han sufrido mermas muy significativas o inclusola extinción en el Campo de Gibraltar, 17 (el 53%) han desaparecido como consecuencia directa de la desecación delconjunto de humedales de La Janda. Esto sin contar las consecuencias directas que ello debió ejercer sobre el resto de la
biodiversidad local, desde invertebrados o plantas, hasta peces, anfibios o reptiles. Estos resultados ponen claramente demanifiesto que el primer paso imprescindible para recuperar al menos una parte de esta biodiversidad perdida, pasanecesariamente por la restauración de los humedales de La Janda.
Las 15 especies restantes analizadas en este trabajo han sufrido las consecuencias de las transformaciones de sus hábitatsnaturales y/o de las persecuciones directas a las que se han visto sometidas. Una única especie, la cabra montés, parecióestar afectada por un factor estocástico natural como es una epizootia.
Respecto a las incorporaciones faunísticas, hay evidencias de la existencia de al menos cuatro especies nuevas llegadas deforma natural (tórtola turca, vencejo cafre, grajilla y golondrina dáurica) (tabla 2), a las que habría que añadir las especiesexóticas introducidas. Asimismo, hay constancia del aumento poblacional de otras cinco especies que antiguamente eranaparentemente muy escasas (garcilla bueyera, cigüeña blanca, ratonero común, gaviota patiamarilla, estornino negro y
meloncillo) (tabla 2). Esto, que podría parecer una buena noticia que compensa, en parte, las pérdidas referidas antes, nolo es, ya que se trata de otra consecuencia más de la alteración del medio, que conlleva la desaparición de especialistas,que son reemplazados, en menor número, por otras especies, normalmente de carácter más generalista.
También es importante señalar el hecho de que cuatro especies, de requerimientos ecológicos más fríos, han desplazadosignificativamente hacia el norte los límites meridionales de su área de distribución, así como que otras dos especies deorigen africano han colonizado nuestras latitudes.
A continuación se exponen los resultados obtenidos agrupando las especies taxonómicamente según el estatus alcanzado.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 35/276
Comunicaciones
35
Tabla
2.Relacióndeespec
iesdepeces,avesymamíferoscuyasituaciónhacambiadosustancialmenteenelCampodeGibraltarenlosúltimos150años.
EXTINTAS
EX
TINGUIDAS
COMO
RE
PRODUCTORAS
O
INV
ERNANTES
VIRTUALMENTE
EXTINGUIDAS
COMO
REPRODUCTORAS
O
INVERNANTES
REDUCCIÓN
SIGNIFI-
CATIVA
DELNÚMERO
DE
REPRODUCTORES
AUMENT
O
EN
EL
NÚMERO
DE
EFECTIV
OS
NUEVAS,NO
INTRODUCIDAS
P E C E S
T h
r e s k i o r n i t h i d a e
Pla
talealeucorodia
A n
a t i d a e
Anseranser
A c
c i p i t r i d a e
Milvusmilvus
Aquilaadalberti
Cir
cusaeruginosus
P a
n d i o n i d a e
Pandionhaliaetus
G r u i d a e
Grusgrus
A c i p e n s e r i d a e
Husohuso
A V E S
P o
d i c e p e
d i d a e
Podicepsnigricollis
A r d e
i d a e
Botaurusstellaris
A n a
t i d a e
Anserfabalis
Anseralbifrons
A c c
i p i t r i d a e
Gypaetusbarbatus
Aegypiusmonachus
Haliaetusalbicilla
T u r n
i c i d a e
Turnixsylvatica
R a
l l i d a e
Fulicacristata
G r u
i d a e
Anthropoidesvirgo
S y
l v i i d a e
Locustellaluscinioides
A r d e i d a e
Ardeapurpurea
R a l l i d a e
Porzanaporzana
Porzanapusilla
O t i d i d a e
Otistarda
S t e r n i d a e
Chlidoniashybridus
Chlidoniasniger
S y l v i i d a e
Acrocephalusscirpac
eus
A c c i p i t r i d a e
Neophron
percnopterus
Aquilachrysaetos
Hieraaetusfasciatus
R a l l i d a e
Rallusaquaticus
S y l v i i d a e
Acrocephalus
arundinaceus
A r d e i d a e
Bubulcus
ibis
C i c o n i d a
e
Ciconiac
iconia
A c c i p i t r i d
a e
Buteobu
teo
L a r i d a e
Larusmichahellis
S t u r n i d a e
Sturnusu
nicolor
C o l u m b i d a e
Streptopeliadecaocto
A p o d i d a e
Apuscaffer
C o r v i d a e
Corvusmonedula
H i r u n d i n i d a e
Hirundodaurica
S o r i c i d a e
Sorexgranarius
C h i r o p t e r a
Barbastellabarbastellus
C a r n i v o r a
Canislupus
Lynxpardinus
R o d e n t i a
Glisglis
Muscardinusavellanarius
M A M Í F E R O S
A r t i o d a c t y l a
Caprapyrenaica
C a r n i v o r a
Herpeste
sichneumon

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 36/276
Almoraima, 35, 2007
36
A. Especies extintas
1. Huso huso
La presencia del esturión beluga originario del mar Negro está documentada por Machado (1857), quien lo cita en “elestrecho de Gibraltar”. La validez de esta referencia parece fuera de toda duda debido a las particulares característicasfenotípicas de esta especie, que hacen que sea difícil confundir con otros esturiones. Machado, además, fue catedráticoen Sevilla (Menor y Prenda, 2004), por lo que sin duda debe conocer al esturión del Guadalquivir ( Acipenser sturio).
Desde entonces hasta la fecha actual no existe constancia de que se haya vuelto a capturar ningún ejemplar de Huso husoen aguas del Estrecho.
2. Podiceps nigricollis
El zampullín cuellinegro era una especie nidificante y muy común en La Janda en el siglo XIX (Saunders en Alonso,1980). La desecación de este humedal probablemente supuso la extinción de la especie en la zona.
3. Botaurus stellaris
El avetoro era otra especie que se encontraba presente en la comunidad de aves nidificantes de La Janda (Irby, 1895)y que se extinguió en la comarca con la desaparición de la misma.
4. Anser fabalis y Anser albifrons
En La Janda, Chapman y Buck (1893) refieren que “en invierno abundan los ánsares” y por la descripción que haceBernardo de Quirós (1920) debieron existir grandes extensiones de castañuela, conformando un hábitat óptimo para la
invernada de estas anátidas. No obstante, Chapman y Buck (1910) apuntan ya un acusado declive de las poblacionesde ánsar campestre ( Anser fabalis) y ánsar careto grande ( Anser albifrons). Esto posiblemente debió estar motivado porlas intensas campañas de desecación de zonas húmedas que se llevaron a cabo en todo el país para erradicarenfermedades como el paludismo y ganar zonas de cultivo. Pero sin duda, lo que agravó este proceso de regresión fuela desecación de La Janda, que seguramente comprometió la supervivencia de las poblaciones de ánsares invernantesen el Campo de Gibraltar, que fueron abocadas a una lenta extinción.
5. Gypaetus barbatus y Aegypius monachus
Irby (1895) apunta a las campañas masivas de envenenamiento para la eliminación del lobo, como la principal causaque motivó la drástica regresión que experimentaron estas dos especies hasta llevarlas a la extinción en la comarca.
6. Haliaetus albicilla
El pigargo es citado en Andalucía por numerosos autores. López Seoane (1870) dice haberla observado cerca deChiclana. Indudablemente, la especie también debió visitar el área del Campo de Gibraltar. Su extinción parece que fuemotivada por la persecución a la que fue sometida la especie.
7. Turnix sylvatica
López Seoane (1870) afirma que esta especie es abundante en todo el litoral andaluz, y Saunders (1871) e Irby (1885)lo citan en varias localidades del Campo de Gibraltar. Se desconocen las causas que han motivado un declive tanacusado, su extinción a nivel local y su casi desaparición en la Península, aunque posiblemente se deba a la reducciónde su hábitat y a la caza.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 37/276
Comunicaciones
37
8. Fulica cristata
Bernardo de Quirós (1920) refiere sobre la focha cornuda, hoy extinta en la zona, que “viven en gran cantidad” en lalaguna del Torero. Como se ha comentado para otras especies, el drenaje de La Janda pudo abocar a esta especie a laextinción en la comarca.
9. Anthropoides virgo
Saunders (1871) señala que la grulla damisela es menos abundante que la grulla común (Grus grus) pero que no es raraen invierno y que no tiene conocimiento de que nidifique en Andalucía. Por su parte, Irby (1895) apunta que haencontrado a esta grulla cerca de Casas Viejas y que no tiene duda de que nidifica en las marismas del Guadalquivir.
10. Locustella luscinioides
La buscarla unicolor debió ser una especie localmente abundante como señala Irby (1895) quien comenta el hallazgode trece nidos de dicha especie en Casas Viejas. Como en casos anteriores, el avenamiento de los humedales queconformaban La Janda pudo haber contribuido a la desaparición de esta especie como nidificante.
11. Sorex granarius
La musaraña ibérica fue citada en la zona con el epíteto específico “araneus” por Irby (1895). Recientemente, a partir dedatos cromosómicos, se ha segregado la especie “granarius” de la “araneus” (Alcantara en Blanco, 1998). La presenciade esta especie en Andalucía también fue documentada por Machado y Núñez (1869) y por Sánchez y García (1885).
Las especies de la subfamilia Soricinae son de distribución Holártica y preferencias norteñas en función de su origenboreal o subboreal. Posiblemente, estos requerimientos ecológicos hayan motivado el desplazamiento latitudinal dellímite de distribución de Sorex granarius hacia áreas más septentrionales, como consecuencia del aumento global delas temperaturas que está aconteciendo en los últimos tiempos.
12. Barbastella barbastellus
Este es un murciélago de carácter centroeuropeo que en España es raro, incluso en las montañas del norte peninsular(Ibáñez en Blanco, 1998). Fue citado por Irby (1895) en la zona de estudio. Asimismo, también lo encontraron enAndalucía Machado y Núñez (1869) y Martínez Reguera (1881). Durante mucho tiempo se ha dudado sobre la veracidadde estas citas (Ibáñez en Blanco, 1998), pero recientemente se ha constatado la presencia de la especie en Jaén (Palomoy Gisbert, 2002) así como en otras zonas de esta comunidad (Migens, comunicación personal) dándoles la razón yhaciendo que cobre mayor fundamento el trabajo de los autores antiguos.
Como en el caso anterior, las variaciones climáticas pueden haber provocado un retroceso de los límites del área dedistribución de esta especie hacia zonas más frías.
13. Canis lupus
El lobo ibérico fue citado por Madoz (1845-1850) en los municipios de Algeciras y Alcalá de los Gazules (Irby, 1895),también nos habla de la presencia de esta especie en la zona del Campo de Gibraltar y escribe que se han exterminadomuchos buitres negros, quebrantahuesos y buitres leonados debido al veneno usado contra los lobos. Esto nos confirmaque la especie desapareció como consecuencia de la presión sistemática a la que fue sometido para lograr su erradicación.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 38/276
Almoraima, 35, 2007
38
14. Lynx pardinus
Citado por Irby (1895) seguramente la especie se vio aquejada en la zona por los mismos problemas que en el resto desu área de distribución, lo que le ha situado al mismo borde de la extinción global.
15. Glis glis y Muscardinus avellanarius
Ambas especies, que resultan inconfundibles, son citadas en el área de estudio por Irby (1895). También refieren supresencia en Andalucía, Machado y Núñez (1869) y Martínez Reguera (1881). Respecto a los datos de estos dos últimosautores, Graells (1897) afirma que no duda de la veracidad de estas citas.
Posiblemente, la extinción de ambas especies se debió a los cambios acontecidos en las condiciones climáticas.
B. Especies extinguidas como reproductoras o invernantes1. Platalea leucorodia
La espátula debió ser una especie relativamente abundante en el Campo de Gibraltar y su entorno, a tenor de los datosaportados por Irby (1895) quien escribe que recolectaron más de setenta huevos en el soto del Torero. Con el drenajede La Janda se extinguió la especie como nidificante en la zona.
2. Anser anser
Ver A. fabalis y A. albifrons
3. Milvus milvus y Aquila adalberti
Las dos especies se han extinguido como nidificantes en la zona seguramente por la conjunción de varios motivos,aunque Irby (1895) señala a las campañas para erradicar al lobo con venenos, como una de las principales causas quemotivaron la drástica regresión que experimentaron.
4. Circus aeruginosus
El aguilucho lagunero es citado como nidificante en La Janda por Irby (1895). La desaparición de este humedal provocóa su vez la extinción, en la Comarca, de la especie.
5. Pandion haliaetus
La reproducción del águila pescadora en la comarca del Campo de Gibraltar está muy bien documentada por variosautores. Su extinción probablemente se debió a la conjunción de diferentes factores como el coleccionismo de huevos
y adultos, otro tipo de molestias en época de nidificación y la caza ilegal, entre otras.6. Grus grus
Chapman y Buck (1893) ya escribieron que la “pequeña colonia” de grulla común de La Janda “va menguando”.Asimismo, Irby (1895) también advierte la situación de declive de esta población, indicando que en breve desaparece-rían de las cercanías de Casas Viejas. Este mismo autor señala como principal causa de esta regresión la presión a la quese vio sometida la población por la acción de los recolectores de huevos. El drenaje de La Janda supuso el episodiodefinitivo para culminar la extinción peninsular de esta especie como nidificante.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 39/276
Comunicaciones
39
C. Especies virtualmente extintas como reproductoras o invernantes o que han sufrido una reducción significativaen el tamaño de su población reproductora
1. Ardea purpurea
La garza imperial era una especie abundante en la Comarca, habiendo dispuesto en el pasado de tres zonas denidificación. Yeates (1946) refiere la existencia de una colonia en la laguna del Torero, e Irby (1895) señala que laespecie crió cerca de Casas Viejas y en una pequeña laguna que existió junto a la carretera de Los Barrios. Una vez más,la desecación de La Janda y la desaparición de la laguna de Los Barrios (Alonso, 1980) a abocado a la especie a su prácticaextinción en el Campo de Gibraltar.
2. Rallus aquaticus , Porzana porzana y Porzana pusilla
La presencia de estas tres especies en La Janda está documentada por Irby (1895) entre otros autores. Actualmente,pueden considerarse prácticamente extinguidas en el Campo de Gibraltar. De continuar reproduciéndose en la zona, supresencia debe ser casi testimonial. La desecación de los humedales de La Janda es lo que debió provocar el declive queexperimentaron estas especies.
3. Otis tarda
La avutarda se encuentra todavía entre la fauna del área de estudio, aunque parece ser que únicamente existe ya un machosolitario (Jiménez, comunicación personal).
El que esta población se halle en el límite del área de distribución de la especie en la península Ibérica, ha podidoocasionar que haya sido especialmente vulnerable a la presión de la caza ilegal y de las transformaciones de su hábitat,
lo que le ha conducido a la situación actual.4. Chlidonias hybridus y Chlidonias niger
Aunque ambas especies de fumareles se pueden observar todavía en la zona de estudio, se les puede considerarvirtualmente extintas desde la desecación de la laguna de La Janda, donde existían ejemplares reproductores (VerAlonso, 1980 e Irby, 1895).
5. Acrocephalus scirpaceus y Acrocephalus arundinaceus
Irby (1895) dejó constancia escrita de la existencia de estas dos especies en dicho humedal. Sin embargo, ambas especieshan reducido enormemente sus efectivos en el área de estudio desde la desecación de La Janda, siendo incluso probableque el carricero común ( A. scirpaceus) haya desaparecido definitivamente como nidificante.
6. Neophron percnopterus
El alimoche era particularmente abundante en el Campo de Gibraltar. Alonso (1980) cita que Verter encontró un nidode esta especie en un árbol. Este caso también es señalado por Sánchez y García (1885), quien afirma que la especienidifica en pinos en Granada. Esto es un claro indicio de lo abundante que debió ser la especie, cuya reducciónpoblacional también pudo estar motivada por el uso de veneno.
7. Aquila chrysaetos y Hieraaetus fasciatus
Ambas especies han visto reducido el número de parejas nidificantes por diversos factores entre los que destacan lasmolestias, el coleccionismo y la persecución a la que se vieron sometidas, en tiempos antiguos, así como a la alteraciónreciente de su hábitat.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 40/276
Almoraima, 35, 2007
40
8. Capra pyrenaica
Según Irby (1895) anteriormente a 1830 la especie estaba presente en las sierras próximas a Algeciras, pero una epizootiamermó enormemente los efectivos de esta especie.
D. Especies cuyos efectivos han aumentado recientemente
1. Bubulcus ibis
La garcilla bueyera contaba con una colonia en la laguna del Torero (Alonso, 1980). Esta especie era especialmenteescasa en el siglo XIX, así Machado y Núñez (1854) indica que el único ejemplar del que tiene noticia procede de PuertoReal y López Seoane (1891) señala que es una especie accidental en Granada y rara en Puerto Real. Por contra,actualmente se trata de una especie muy común que ha experimentado una importante expansión, tanto poblacionalcomo de amplitud de distribución.
2. Ciconia ciconia
Esta especie también ha experimentado un aumento en el número de efectivos debido a cambios en su conductamigratoria y a la explotación de nuevos recursos tróficos, como los proporcionados por los basureros.
3. Buteo buteo
Esta especie no aparece citada en el Campo de Gibraltar en ninguno de los trabajos revisados, e Irby (1895) señalaexpresamente que nunca la ha encontrado criando en las proximidades de Gibraltar. Posiblemente las transformacionesque ha experimentado el medio de la zona hayan favorecido a esta especie.
4. Larus michahellisEsta especie oportunista ha incrementado enormemente sus efectivos en los últimos años aprovechando los nuevosrecursos que le ha proporcionado la actividad humana (vertederos, descartes pesqueros, etc).
5. Sturnus unicolor
Irby (1895) señala la existencia de tres o cuatro parejas en una venta cerca de Casas Viejas. Mientras que Machado yNúñez (1854) y Sanchez García (1885), ni siquiera llegan a recogerlo en sus catálogos de aves. López Seoane (1861)indica que es “poco común” en Granada. Esta especie ha sabido aprovechar los recursos tróficos que le ofrece laactividad humana experimentando una notable expansión.
6. Herpestes ichneumon
Posiblemente la desaparición de dos grandes superdepredadores como el lobo y el lince ha favorecido la expansión queha experimentado el meloncillo en los últimos años. Esta especie fue citada por Irby (1895) en el área de estudio, y sudensidad debió ser muy baja ya que Madoz, 1845-1850 no señala la presencia del mismo en ninguno de los municipiosde la zona.
E. Especies nuevas, no introducidas por el hombre
1. Sterptopelia decaocto
La tórtola turca, procedente de Europa oriental, ha colonizado gran parte del territorio peninsular, con cría confirmadadesde 1974 (Bernis et al. 1985). Es una especie favorecida por la actividad humana, cuya expansión está asociada a laabundancia de recursos tróficos en ambientes urbanos.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 41/276
Comunicaciones
41
2. Apus caffer
Según Alonso, 1980, esta especie procedente del África subsahariana fue vista por primera vez en la península Ibéricapor Bernis en 1962. Desde entonces hasta la fecha ha experimentado un proceso de expansión, favorecida por labenignidad climática y por la colonización de la Península por la golondrina dáurica, especie a la que expulsa de susnidos, para utilizarlos y realizar su propia puesta (Barrios, 1994).
3. Hirundo daurica
La presencia de esta especie en el área de estudio en el siglo XIX no está citada. Al igual que el vencejo cafre, elprogresivo aumento de las temperaturas y el cambio experimentado por el clima en el último siglo y medio, puede haberfavorecido la expansión hacia el norte del límite septentrional de su área de distribución, colonizando nuevos territorios
en la región Paleártica.4. Corvus monedula
Esta especie no está citada como reproductora en el Campo de Gibraltar en el siglo XIX. Alonso (1980) señala que alsur de Cádiz comenzaron a llegar probablemente durante los años cincuenta del siglo XX. Esta especie oportunista seha visto favorecida por los cambios ejecutados sobre el medio como consecuencia de la actividad antrópica.
BIBLIOGRAFÍAALONSO. Avifauna del sur de Cádiz. Campo de Gibraltar y comarca de La Janda. Tesis doctoral. Universidad Complutense de Madrid. 1980. Madrid.BARRIOS, F. “Primeros datos sobre la reproducción del vencejo cafre en España”. Quercus 95: pp. 6-8. 1994.BERNALDO DE QUIRÓS, J. L. “Excursión ornitológica a La Janda”. Boletín de la Real Sociedad Española de Historia Natural, Tomo XX: pp. 236-248. 1920.
BERNIS, F., Asensio, B. y J. Benzal. “Sobre la expansión y ecología de la tórtola turca (Streptopelia decaocto), con nuevos datos del interior de España. Ardeola32: pp. 279-294. 1985.BLANCO, J. C. Mamíferos de España . GeoPlaneta. 1998. Barcelona.CLAVERO M, Blanco-Garrido F. y Prenda J. “Fish-habitat relationships and fish conservation in small coastal streams in southern Spain”. Aquatic Conservation-
Marine and Freshwater Ecosystems 15. pp. 415-426. 2005.CHAPMAN, A. y W. J. Buck. Wild Spain. 1893. Londres.DELIBES, M. La acción humana y la crisis de biodiversidad. En M. Garmendio (Ed.). “ Los retos medioambientales del siglo XXI. La Conservación de la biodiversidad en España”. Fundación BBVA, 2004. Bilbao: pp. 23-38.DUEÑAS, M.A y Recio, J. M. Bases ecológicas para la restauración de los humedades de la Janda (Cádiz, España). Serv. Pub. Univ. 2000. Córdoba.GRAELLS, M. P. Fauna Mastodológica Ibérica. Imprenta Aguado. 1897. MadridIRBY, L. H. L. The ornithology of the Straits of Gibraltar. R. H. Porter. 1895. Londres.LAVOREL, S. et al. “Mediterranean terrestrial ecosystems: research priorities on global change effects”. Global Ecology and Biogeography, 7: pp. 157-166. 1998.LÓPEZ SEOANE, V. “Catálogo de las aves observadas en Andalucía”. Rev. de los Prog. de las C. Exac. Fís. y Nat. de Madrid , (1861): pp. 1-62. 1861.LÓPEZ SEOANE, V. Revisión del catálogo de las aves de Andalucía, 1870. La Coruña.MACHADO Y NÚÑEZ, A. Catálogo de los peces que habitan o frecuentan las costas de Cádiz y Huelva. Librería Española y Extrangera. 1857. Sevilla.
MACHADO Y NÚÑEZ, A. Catálogo de las aves observadas en algunas provincias de Andalucía. Imprenta y taller de encuadernaciones de Juan Moyano. 1854. Sevilla.MACHADO Y NÚÑEZ, A. Catálogo metódico y razonado de los mamíferos de Andalucía. Imp. de Gironés y Orduña. 1869. Sevilla.MADOZ, P. Diccionario Geográfico-Estadístico-Histórico de España. Ámbito/Editoriales Andaluzas Reunidas (Ed. Facsimil, 1988). 1845-1850. Valladolid.MARTÍ, R. y J. C. del Moral. Atlas de las aves reproductoras de España . Dirección General de Conservación de la Naturaleza – SEO. 2003. Madrid.MARTINEZ REGUERA, L. La Fauna de Sierra Morena: catálogo descriptivo de los mamíferos del término de Montoro . Imp. de M. Romero. 1881. Madrid.MENOR, A. y J. Prenda. “Antonio Machado y Núñez, pionero de los inventarios de biodiversidad en España”. Quercus 224: pp. 34-39. 2004.PALOMO, J L. y J. Gisbert. Atlas de los Mamíferos terrestres de España . Dirección General de Conservación de la Naturaleza – SECEM – SECEMU. 2002. Madrid.PERSSON, H. “Invernada de ansares en España”. Quercus 109: pp. 19-22. 1995.PERSSON, H. “Otoño silencioso: el declive del Ansar común en Doñana”. Quercus 129: pp. 39-41. 1996.SALA, O. E. et al. “Global biodiversity scenarios for the year 2100”. Science, 287: pp. 1770-1774. 2000.SANCHEZ Y GARCÍA, J. 1885. Catálogo de los mamíferos y aves observados en la provincia de Granada. Imp. de Indalecio Ventura. Granada.SAUNDERS, H. 1877. “A list of the birds of Southern Spain”. Ibis 1871: pp. 205-225 y pp. 384-402.YEATES, G.R. Bird Life two deltas . Ed. Faber y Faber, Ltd. 1946. Londres.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 42/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 43/276
Comunicaciones
43
RESUMEN
El corzo (Capreolus capreolus L.) es una especie destacada dentro de la fauna salvaje del Campo de Gibraltar. A lapeculiaridad de sus poblaciones, que se encuentran en el límite ecológico y biogeográfico de esta especie de distribuciónEuropea, hay que unir el gran atractivo de este animal como pieza cinegética. En efecto, siendo una especie cazada por elhombre desde tiempos remotos, el corzo se encuentra en este momento en un punto álgido por su atractivo para el cazador.A su vez, esto se está traduciendo en un renovado interés por la conservación y fomento de las poblaciones de este animal.
En la presente Comunicación se pasa revista a la evolución de las poblaciones de corzo en el Parque Natural LosAlcornocales durante las últimas décadas, al tiempo que se expone como ha ido evolucionando la práctica de su caza,haciendo especial hincapié en la relación existente entre ambas cuestiones, y concluyendo que la caza debe ser consideradaun factor importante para la conservación de esta especie de cara al futuro.
Palabras clave: corzo, ciervo, ordenación cinegética, períodos hábiles de caza.
LA CAZA COMO FACTOR DECONSERVACIÓN DEL CORZO ENEL PARQUE NATURAL LOSALCORNOCALES.UNA VISIÓN RETROSPECTIVA
Pablo Fernández-Salguero López / Ingeniero de Montes
José María Sánchez García / Ingeniero de Montes

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 44/276
Almoraima, 35, 2007
44
1. INTRODUCCIÓN
El corzo (Capreolus capreolus L.) es un ungulado perteneciente a la familia de los cérvidos, que encuentra en las sierrasde Cádiz y Málaga, y en concreto en el Campo de Gibraltar, su área de distribución más meridional de Europa Occidental.La particularidad de esta especie radica precisamente en el aislamiento con las que cuentan estas poblaciones del sur frentea las del resto de la península Ibérica. Asimismo, presenta características muy diferentes al resto de cérvidos, es casi una“singularidad” en el ambiente mediterráneo, y por esta razón es tan importante como especie silvestre, y sin duda, comopieza cinegética.
El comportamiento de esta especie es huidizo y silencioso, abandonado en escasas ocasiones la protección del monte, por elque se desplaza fácilmente debido a su pequeño tamaño. Otro aspecto que lo diferencia del ciervo u otros cérvidos es que esterritorial y por tanto, no gregario, apareciendo únicamente juntos el grupo familiar del año, siendo los machos generalmentesolitarios, salvo en el período de celo. En cuanto a su alimentación, es eminentemente ramoneador, precisando de brotesnuevos y tiernos con alto contenido de azúcares y poca fibra, pasando gran tiempo del día alimentándose.
El corzo habita en todo tipo de ambientes forestales, aunque en la provincia de Cádiz prefiere masas de alcornocal-quejigaldensas con abundante cobertura de matorral y diversidad de vegetación para alimentarse y ocultarse. En la época estival,cuando las condiciones son más desfavorables, su preferencia se centra en torno a los “canutos”, típicos del Parque Naturalde Alcornocales, donde encuentra más humedad y variedad de vegetación. Ésta es una de las razones por la que esta especie,de origen boreal y más propia de bosques europeos más húmedos, ha sobrevivido en esta área de distribución tan meridional,en su límite ecológico.
2. LA EVOLUCIÓN DEL CORZOA principios del siglo XX, una vez que el ciervo prácticamente había desparecido en el siglo anterior debido a la acentuadapresión de la caza, el corzo era el único cérvido abundante en la comarca de los alcornocales gaditanos. Sin embargo, comoconsecuencia de la intensa explotación de los espacios forestales, la inestabilidad política y las frecuentes épocas de penuriay hambre para la población, en las que la caza se convierte de nuevo en un recurso para el sustento diario, las poblacionesde especies cinegéticas se hacen cada vez más escasas. El lobo, último gran predador presente en estos montes, fueeliminado sistemáticamente durante las primeras décadas de este siglo con el objeto de evitar los frecuentes daños sobrela cabaña ganadera. La desaparición de este último gran predador tendría más tarde consecuencias en relación con laexpansión de las poblaciones de especies de caza mayor, al quedarse éstas sin enemigos naturales.
A finales de la década de los cincuenta se produce una incipiente recuperación de la caza mayor en esta Comarca, asociada
a la consolidación durante esta época de vedados importantes ( La Almoraima, Montes de Jerez, El Jautor , etc.). En ellosse acometen sucesivas repoblaciones con ciervos, encaminadas a la reintroducción de esta especie. Finalmente, laprogresiva creación de acotados al amparo de la Ley de Caza de 1970 , condujo a la provincia de Cádiz a recuperarprogresivamente el importante papel que en el ámbito de la caza mayor había tenido en siglos pasados.
Este esfuerzo de fomento de la caza, junto con el despoblamiento del medio natural que tuvo lugar a raíz de la crisis delcarbón vegetal (años sesenta), condujo a una expansión de las especies de caza mayor, y en particular del corzo. Comoejemplo, pueden observarse en la tabla siguiente los datos obtenidos en censos realizados en el año 1974, en el coto CA-10.133, que incluye los montes propios de Alcalá de los Gazules, donde el corzo era claramente mayoritario frente al ciervo(se incluyen aquí tanto machos como hembras).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 45/276
Comunicaciones
45
MONTE CORZO VENADOHernan Martín 46 12Montero 46 26Sauzal 30 8Barrancones 20 7Laganes y Laurel 40 12Jota y Zarza 50 15Totales 232 82
Tabla 1. Datos del censo realizado en 1974.
Como consecuencia de ello vinieron los “años dorados” para la caza del corzo, con un progresivo y notable incremento enel número de capturas, y un creciente interés por parte de los titulares de cotos, como se refleja en la evolución en el número
de trofeos homologados.
En los comienzos de los ochenta, se empezó a tomar conciencia de que el éxito inicial en la reintroducción del ciervo estabateniendo consecuencias no deseadas, y en particular que el corzo estaba viéndose afectado por la nueva situación. En efecto,las más de dos décadas de vigilancia y esfuerzo realizado en el fomento de la caza mayor por parte de la Administración y delos titulares de cotos, junto con el progresivo abandono del medio rural por parte de la población al cesar el aprovechamientodel carbón vegetal, habían traído consigo una rápida expansión del ciervo, menos exigente y con mayor capacidad deadaptación que el corzo. Para entonces se había generalizado la caza del ciervo en grandes batidas (monterías y ganchos), conexcelentes resultados; por su parte, también la caza de trofeos a rececho atrajo el interés de los cazadores, lo que aumentó elinterés por esta especie, cuya caza en esta Comarca era aún esporádica hace apenas 20 años, a comienzos de los setenta.
Figura 1. Evolución de los trofeos de corzo homologados en la provincia de Cádiz (1950-75).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 46/276
Almoraima, 35, 2007
46
La conclusión resultaba clara; los cuidados prodigados, junto con la ausencia de enemigos naturales, habían propiciado unexcesivo crecimiento de la población ciervo. Por otra parte, la caza exclusiva de machos y el respeto escrupuloso hacia lashembras y crías, inicialmente aconsejables para el éxito de la reintroducción, se estaba convirtiendo una vez lograda ésta enuna causa del desequilibrio entre sexos y de unas tasas de crecimiento de la población insostenibles por el medio natural.
Resultó evidente, a la vista de la evolución de las poblaciones de ambas especies, que la competencia por el alimento y porel territorio estaba desplazando al corzo, especie de menor talla (con un peso entre cuatro y cinco veces menor que el del ciervo)y más delicada en sus necesidades alimenticias. Como consecuencia de ello, el corzo inició un lento pero progresivo declive,hasta llegar a desaparecer en aquellos enclaves donde el venado había alcanzado mayores densidades poblacionales. Todo ellofue confirmado por los estudios científicos realizados en la Comarca (Braza, 1990). Como resultado, se produjo una mermaimportante en el aprovechamiento cinegético de la especie, con el abandono de la práctica de su caza en batidas (de larga
tradición en la Comarca, pero a las que los escasos resultados obligaron a renunciar) a finales de los años ochenta, e inclusola renuncia a cazarlo a rececho en aquellos cotos donde sus poblaciones se habían visto reducidas a valores residuales.
Una buena referencia de la situación descrita puede encontrarse en la evolución experimentada por los censos de ciervoy corzo en La Almoraima, donde ya en los años sesenta y principios de los setenta, sus titulares procedieron a introducirciervos, gamos, y muflones. Una vez declarado como coto privado de caza en 1974, tras la entrada en vigor de la Ley deCaza de 1970 y su Reglamento, La Almoraima se acogió a la figura de Reglamentación Cinegética Especial, que permitíacon carácter excepcional la caza de hembras y crías como medida para el control de poblaciones. En un principio, solamentese reguló la gestión del ciervo, ya que, el resto de especies introducidas no eran representativas, y en el caso particular delcorzo, no fue hasta 1977 cuando se llevó a cabo la primera reglamentación, pues no se considera necesario atendiendo ala gran densidad de corzos y el elevado número de trofeos que se obtenían anualmente.
Pese a esta larga tradición en ordenación cinegética, el insuficiente cumplimiento de los cupos de descaste previstos paralas hembras de ciervo condujo a lo largo de estos años a una evolución negativa (que reflejamos en la tabla y gráfico quesiguen), con la práctica ausencia del corzo y el crecimiento de la población de ciervo hasta causar daños en la vegetaciónleñosa. Mientras tanto, este coto se manifestó en los sucesivos episodios de mortandad atípica de ciervos (inviernos 89-90 y 95-96) como uno de los “epicentros” de esta mortandad, fruto del exceso de densidad alcanzado.
En la tabla siguiente se representa la evolución comparativa en los censos de corzo y ciervo (1978-2005), estando los datosreferidos a individuos con más de un año en paridera, en el coto de caza CA-10.417 La Almoraima término municipal deCastellar de la Frontera, Cádiz, así como las densidades por cada 100 Ha.
AÑO TOTAL CORZO DENSIDAD TOTAL CIERVO DENSIDAD1976 - - 600 4
1978 573 3,8 1.050 71980 570 3,8 850 5,71983 230 1,6 1.050 71987 132 0,9 1.613 10,91989 - - 2.393 161992 - - 3.606 251997 - - 1.756 12,22005 - - 1.855 12,8
Tabla 2. Evolución en los censos de especies en el coto La Almoraima (1976-2005).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 47/276
Comunicaciones
47
Ante la situación creada, en 1991, la Agencia de Medio Ambiente acometió la elaboración del primer Plan Comarcal deOrdenación Cinegética para la comarca de Los Alcornocales. Entre los objetivos de éste destacaban la restauración de laspoblaciones de corzo y el restablecimiento de un equilibrio adecuado entre herbívoros y vegetación.
En esta línea, las principales medidas contempladas en la ordenación para la consecución de estos objetivos fueron lassiguientes:
Fomento de la especie de mayor interés, el corzo, que se deseaba potenciar como especie más genuina del Parque Natural.Con este fin y a pesar de las propuestas más alarmistas, que proponían suspender la caza del corzo como medida deprotección para sus disminuidas poblaciones, la Administración apostó por mantener el carácter cinegético de la especie,entendiendo que ésta era la mejor forma de incentivar su cuidado por parte de los gestores de caza mayor y de animar aéstos a controlar eficazmente las poblaciones de ciervo. Al mismo tiempo, se modificó el periodo hábil de caza del corzo,
trasladándolo del verano a la primavera, con el objeto de adaptarlo a las condiciones climatológicas de la zona y al ciclobiológico de la especie, haciéndolo al mismo tiempo más atractivo para el cazador.
Corrección del crecimiento poblacional del ciervo y del desequilibrio entre sexos, mediante la caza de descaste sobrehembras y crías, añadiendo a las modalidades de rececho y aguardo la posibilidad de cazarlas en batidas. Todo ello con elobjeto de llegar a una densidad normal de 10-16 animales adultos por cada 100 Ha acotadas, y a una relación de sexos de1:1,5, adecuada para los sistemas de caza en batida tradicionales en la Comarca.
Este primer Plan Comarcal de Ordenación Cinegética tuvo una vigencia de cinco años, siendo revisado y actualizadoposteriormente en dos ocasiones (1997 y 2002). Como resultado de su aplicación, durante los últimos catorce años se ha logradouna sensible disminución en la población de ciervos y una recuperación incipiente, y en algunos cotos destacada, del corzo.
Figura 2. Evolución en los censos de especies en el coto La Almoraima (1978-2005)

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 48/276
Almoraima, 35, 2007
48
En lo que respecta al ciervo y el corzo, el avance en los objetivos perseguidos (control del primero y recuperación delsegundo) se refleja en los datos del seguimiento efectuado durante los últimos años en la Comarca, y que arroja lassiguientes estimaciones:
AÑO Corzos / 100 Ha Ciervo / 100 Ha1989/91 5 342000/04 8 17
Tabla 3. Evolución en la densidad de corzo y ciervo en la provincia.
Por lo que respecta al corzo, su recuperación se ha observado con mayor claridad en aquellos cotos en los que se ha realizadoun intenso control del ciervo, así como mejoras para favorecer al primero de ellos (rozas, comederos, puntos de agua), cuyoefecto resulta muy positivo incluso a corto plazo, dada la alta tasa de crecimiento de las poblaciones de corzo cuando estaespecie se encuentra en condiciones óptimas para su desarrollo.
3. LA CAZA DEL CORZO EN LA COMARCA DE LOS ALCORNOCALES
Desde mediados del siglo XIX, el único cérvido presente en la provincia de Cádiz era el corzo. La cabra montés tambiénhabía sido erradicada debido a la presión humana, de tal modo que las piezas de caza mayor se limitaban al lobo, el jabalí o cerdo asilvestrado y el corzo. Este último se cazaba en rececho y aguardos, pero también en batidas, por el atractivo queha tenido desde siempre esta modalidad de caza. De ello existen testimonios escritos y también gráficos, como lo son lasabundantes fotografías que muestran escenas de las batidas de los años 20 del pasado siglo.
Los convulsos años treinta, la guerra civil y la posguerra, supusieron una etapa de penuria nada favorable para laconservación de la caza; sólo a comienzos de los años sesenta, y coincidiendo con la emigración masiva de los habitantesdel medio natural hacia los pueblos y las grandes ciudades, se producen las condiciones para que prospere la caza mayor.Los grandes propietarios, conscientes de ello, apuestan por este aprovechamiento y realizan, como ya se dijo, las primerasrepoblaciones con ciervos.
Durante los años setenta, el corzo se verá beneficiado por ese esfuerzo de fomento de la caza, y al mismo tiempo favorecidopor la abundancia de recursos tróficos en un medio fuertemente presionado por el hombre hasta fechas recientes. Lavegetación en mosaico, fruto del rebrote del matorral tras el cese del carboneo, y del abandono de la agricultura desubsistencia que se había practicado por los habitantes de estos montes, constituía un hábitat ideal para la especie. Por suparte, el ciervo era aún muy escaso, por lo que no le afectaba su competencia.
Figura 3. Representación de la evolución en la densidad de corzo y ciervo en la provincia.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 49/276
Comunicaciones
49
Durante estos años se produce una creciente presión cinegética sobre el corzo, paralela a la recuperación de sus efectivos,que sin duda fue posible debido al carácter prolífico de la especie. Esta etapa dorada para el corzo se vio interrumpida porla evolución de la población de ciervos, en incesante crecimiento, y probablemente también por la simplificación delhábitat, al envejecer el monte de cabeza y quedar fuera del alcance del diente del corzo buena parte de los recursos tróficos.Por otra parte, la abundancia del venado trajo consigo otro problema, que se sumó a los de competencia por el alimentoy por el territorio: la creciente práctica de la caza del ciervo en batidas, que trajo consigo la importación de rehalas de perrosde agarre, de gran talla, muy diferentes de los pequeños perros podencos de rastro, tradicionalmente empleados para lasbatidas de corzo. El empleo de estos nuevos perros, junto con la fecha tradicional de celebración de las batidas de corzo(marzo-abril), con las corzas a punto de parir, debió ser muy negativo para la evolución de sus efectivos.
De este modo, ya en los comienzos de los ochenta los propios titulares de cotos manifestaron su preocupación a la
Administración, solicitando al ICONA la puesta en marcha de un Plan Especial de Protección aparte el Corzo en la Comarcadel Aljibe. Probablemente los cambios de aquellos años, con el traspaso de competencias a la autonomía andaluza,retrasaron la puesta en práctica de estas iniciativas.
Así se llega a comienzos de los noventa, cuando la escasez de corzos había llevado a abandonar su caza en batidas. Se iniciaentonces un debate sobre el futuro del corzo, esta vez ya contando con los estudios realizados por los científicos de la EBD.Los sectores más conservacionistas propusieron vedar la caza del corzo, debido a su escasez; los titulares de cotos hicieronver que si la especie dejaba de ser cinegética, perdería su atractivo para ellos y se reducirían los esfuerzos dirigidos a surecuperación. Finalmente, la Administración optó por mantener al corzo como especie cazable, pero hizo valer la idea deque sólo controlando eficazmente la población de ciervos sería posible recuperar el corzo. Fruto de esta iniciativa fue laprimera Ordenación Cinegética Comarcal, de la que ya se ha hablado.
Desde entonces hasta esta fecha se han realizado sucesivos ajustes en los periodos y modalidad de caza practicables. Entreellos destaca la recuperación de la caza en batidas en el año 1999, que con limitaciones en cuanto a presión de caza, eltipo y número de perros, etc., parecen estar dando resultados positivos, y convirtiéndose en un aliciente para la conservaciónde la especie.
Igualmente, en lo que respecta a los recechos y aguardos, se ha llegado a un periodo que satisface las necesidades del cazadorsin interferir seriamente en las necesidades biológicas de la especie. Al mismo tiempo, y a escala coto, se ha vinculado laobtención de permisos para la caza del corzo al cumplimiento de los cupos de descaste de ciervo previstos en el Plan Técnicode Caza. Por último, se ha implantado un sistema de marcaje de trofeos mediante crotales, que permite identificar suprocedencia y seguir su rastro desde el coto de origen hasta el taller de taxidermia.
4. EVOLUCIÓN HISTÓRICA DE LOS PERÍODOS HÁBILES DE LA CAZA DEL CORZO EN LA PROVINCIA
La evolución sufrida por las poblaciones de la especie en la provincia ha determinado sin duda, los períodos hábiles de sucaza, así como las modalidades autorizadas. En este apartado se analizará la evolución experimentada por las épocas decaza del corzo desde mediados de los años setenta a partir del inicio de las Reglamentaciones Especiales contempladas enla Ley de Caza de 1970 hasta la presente temporada.
Así, en la tabla en la que se han resumido los períodos, puede observarse claramente como ha existido una gran diversidadde fechas que han ido más allá de los cambios experimentados por los organismos responsables de la elaboración de lasórdenes anuales de veda.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 50/276
Almoraima, 35, 2007
50
AÑO
PERÍODO
HÁBILDELCORZO
MARZO
AB
RIL
MAYO
JUNIO
JULIO
AGOSTO
SEPTIEMBRE
OCTUBRE
1974
Períodode6semanasmáx
.
19
agosto
1octubre
1975
Períodode6semanasmáx
.
19
agosto
1octubre
1976
Períodode6semanasmáx
.
17agosto
5octubre
1977
Períodode6semanasmáx
.
15agosto
3octubre
1978
Períodode6semanasmáx
.
31julio
18septiembre
1979
Períodode6semanasmáx
.
30julio
17septiembre
1980
Períodode6semanasmáx
.
29julio
16septiembre
1981
Períodode6semanasmáx
.
27julio
14septiembre
1982
Períodode6semanasmáx
.
26julio
13septiembre
1983
Períodode6semanasmáx
.
25julio
1
2septiembre
1984
4ºdomingojulio
4ºdomingosept.
1985
4ºdomingojulio
4ºdomingosept.
1986
4ºdomingojulio
4ºdomingosept.
1987
4ºdomingojulio
4ºdomingosept.
1988
4ºdomingojulio
4ºdomingosept.
1989
4ºdomingojulio
4ºdomingosept.
1990
22julio
23septiembre
1991
28julio
22septiembre
1992
15marzo
31mayo
Sólorecechoyaguardo
1993
11marzo
30mayo
Sólorecechoyaguardo
1994
10marzo
29mayo
Sólorecechoyaguardo
1995
11marzo
28mayo
Sólorecechoyaguardo
1996
20mayo
20Julio
Sólorecechoyaguardo
1997
8marzo
25Mayo
Sólorecechoyaguardo
1998
15marzo
24
abril
5julio
26julio
1999
7marzo
23mayo
2000
5marzo
21mayo
5a19marzo:batidas
2001
4a31marzo
4a31demarzo:batidas
2002
3marzo
7abril
3marzoa7abril:batidas
21julio
1sept.
2003
5marzo
15abril
5marzoa5abril:batidas
10julio
15agosto
2004
5marzo
15abril
5marzoa5abril:batidas
10julio
15agosto
2005
5marzo
15abril
5marzoa5abril:batidas
10julio
15agosto

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 51/276
Comunicaciones
51
En los primeros años a los que se hace referencia, la década de 1974 a 1983, la caza del corzo atendía al período establecidopara toda España, que se correspondía con finales de verano y principios de otoño, independientemente de las diferenciasexistentes entre las poblaciones ibéricas. Asimismo, y a petición de los gestores de Cádiz, su caza se pudo ampliar duranteun máximo de seis semanas durante primavera, fecha más favorable para la caza de acuerdo al comportamiento del corzoen estas latitudes, adelantado en su celo y reproducción frente a las poblaciones del centro y norte de la Península.
A partir de 1984 hasta comienzos de los años noventa, se mantiene un único período de dos meses entre finales de julioy septiembre, continuando con lo autorizado en los años anteriores, más acorde a latitudes norteñas, que a las condicionespropias de Los Alcornocales. Este período coincide con la época en la que las poblaciones de corzos se vieron másmermadas en la provincia, llegando sus densidades a valores alarmantes, y como se ha comentado unos años en los quese debieron iniciar medidas de recuperación.
En 1.991 se aprueba la 1ª Ordenación Cinegética Comarcal, y a raíz de las iniciativas establecidas se modificó en granmedida la caza del corzo en la provincia, pasando el período autorizado a la primavera, época más acorde y favorable, quetuvo buena acogida entre titulares y gestores de cotos. Así, en un principio se estableció desde mediados de marzo a finalesde mayo, estando autorizadas únicamente las modalidades de rececho y aguardo, ya que las bajas densidades, no hacíanlas batidas aconsejables ni tampoco atractivas. Este nuevo período autorizado se mantuvo más o menos constante hasta elaño 2000, salvo temporadas puntuales, en las que se modificó sin éxito para volver a la situación anterior. Es de destacarel año 1.998, en el que se vuelve a considerar el período fraccionado dejando vedada la época de reproducción.
A partir del año 2000, y como consecuencia de la cierta recuperación experimentada gracias a las medidas de la OrdenaciónCinegética Comarcal, se inicia una etapa que termina con el actual período hábil autorizado, fraccionado en primavera yverano, y en el que se autoriza la modalidad en batida en aquellos cotos con densidad adecuada.
En la reciente Orden Anual de Vedas, vigente para la actual temporada 2005/06, la caza del corzo sigue con la ciertacontinuidad alcanzada en el último lustro, encontrándose fraccionado en dos períodos hábiles, el primero en verano, desdeel 10 de julio al 15 de agosto, y el segundo, en la primavera siguiente, desde el 5 de marzo al 15 de abril. Aquellos cotosque tienen autorizadas batidas entre las modalidades de caza, las podrán realizar en este período de primavera. Si por algunarazón, estos acotados que cuentan con densidad suficiente de corzos para llevar a cabo batidas optaran únicamente por lamodalidad a rececho, la veda de primavera podría retrasarse hasta el 25 de mayo.
La celebración de batidas de corzos ha supuesto un logro importante en determinadas acotados en los que los gestorescinegéticos han apostado por esta especie, como modalidad tradicional de la provincia, y como alternativa al rececho, sinduda, la modalidad por excelencia de caza de esta especie.
5. CONCLUSIONES
El corzo en la provincia de Cádiz, y en concreto en el Campo de Gibraltar, donde alcanza sus mayores densidades, es unaespecie singular y distintivo propio de Los Alcornocales, así como una cotizada pieza de caza desde la antigüedad.
Los altibajos sufridos por la especie a lo largo del último siglo y la situación actual de lento, pero constante aumento deefectivos, demuestran que la actividad cinegética ordenada y con una adecuada gestión, puede resultar, sin duda, unamedida para recuperar poblaciones que por distintas causas se hayan visto mermadas o en situaciones difíciles. LaOrdenación Cinegética Comarcal y sus sucesivas revisiones, llevadas a cabo en la comarca de los Alcornocales por la

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 52/276
Almoraima, 35, 2007
52
Administración y con la colaboración de titulares y gestores de cotos, ha tenido sus frutos reduciendo las poblaciones deciervo a densidades más adecuadas, y recuperando el corzo como especie cinegética.
Es de esperar que esta situación se mantenga en los próximos años, y que la tendencia actual no se vea alterada, gozandoel corzo de un creciente interés cinegético y natural, siendo la especie “reina” de Los Alcornocales.
6. BIBLIOGRAFÍABRAZA et al. 1990. El Corzo en la Sierra de Cádiz. IARA-AMA. Sevilla.BRAZA et al. 1994. El Corzo Andaluz. Consejería de Agricultura y Pesca, Junta de Andalucía. Sevilla.BRAZA et al. 2004. El Corzo en los Alcornocales. Mapa de Calidad de Hábitats. Consejería de Obras Públicas, Junta de Andalucía. Sevilla.SÁNCHEZ GARCÍA, José María: “La Ordenación Cinegética Comarcal del Parque Natural de Los Alcornocales: Resultados tras doce años de gestión” , Almoraima ,31 (2003), pp. 169-179.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 53/276
Comunicaciones
53
RESUMEN
Los vertidos de hidrocarburos en la Bahía tienen un efecto sobre los organismos que la pueblan, especialmente en los sésiles.Aquí englobamos al erizo común. Se estudia la reacción ante diferentes hidrocarburos y a distintas concentraciones, tantoen su fecundación y desarrollo embrionario, como en la respuesta del sistema inmune.
INTRODUCCIÓNEl trabajo pretende valorar la incidencia de los vertidos de diferentes derivados del petróleo sobre el desarrollo embrionariodel erizo común (Paracentrotus lividus).
El erizo común, nos parecía un buen bioindicador para medir cómo pueden afectar diferentes compuestos y concentracionesal desarrollo embrionario en sus primeros estadios.
De igual modo, se pretende valorar la incidencia de los hidrocarburos sobre el sistema inmunitario del erizo común(Paracentrotus lividus).La respuesta del sistema inmunitario a los contaminantes se midió a través de la hemolinfa de loserizos en las tres pruebas realizadas en nuestro trabajo: medición de los radicales de nitrógeno, quimioluminiscencia yviabilidad; todo ello en condiciones in vitro.
La medición de la concentración de los radicales de nitrógeno se realiza porque es uno de los sistemas de defensa del erizo.Son productos tóxicos y los utiliza para eliminar el patógeno.
La prueba de quimioluminiscencia se realiza debido a que la estimulación de los fagocitos de la hemolinfa produce unaumento del consumo de oxígeno (Adema et al 1991, Anderson 1996A) .El estallido respiratorio provoca la emisión defotones, que son medidos por el Flouroskan.
Por último la viabilidad mide la cantidad de células que sobreviven al tratamiento. El rojo neutro se adhiere a la membranacelular de las células vivas: la intensidad del rojo determina, por tanto, las células supervivientes.
Se pretende determinar como influyen diferentes compuestos en distintas concentraciones in vitro y a semejanza de comose puede dar en el medio marino después de un vertido, en el sistema inmunitario del erizo común.
INFLUENCIA DE LA CONCENTRACIÓN DEHIDROCARBUROS SOBRE EL ERIZOCOMÚN (PARACENTROTUS LIVIDUS)
José Miguel Ramos Durán / Orlando Garzón GómezPablo Camacho Pozo / Ignacio Merlán Llorente

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 54/276
Almoraima, 35, 2007
54
MATERIAL Y MÉTODOS
Desarrollo embrionario
• Características del Erizo común ( Paracentrotus lividus) y otroserizos presentes en el litoral de Algeciras
El erizo común, el más abundante en la zona, se caracteriza por suaspecto globoso, cuyo caparazón está recubierto de espinas que leprotegen y le permiten desplazarse por el fondo marino. Se localiza enla costa a una profundidad entre o y 80m. Como todo equinodermo,tiene capacidad de regeneración. Se halla en todo tipo de colores salvo
en negro. Se encuentra sobre arena y sobre roca. Se recubre de conchas,piedras y algas como defensa y para evitar la luz intensa. Ademásperfora las rocas.
• Método de extracción de los gametos
Se toma un erizo común del acuario y se depositó en un recipiente que posee agua de mar (pasterizada) que lo cubríacompletamente. Aquí fue sometido a la estimulación y liberación de gametos. Se estimuló la secreción de gametosaplicando una corriente eléctrica de 30 V y 50 mA mediante un transformador durante 12 s. En cuanto comienza lasecreción, se tomó con una pipeta de 1 ml y se depositó en un tubo de ensayo. El erizo de mar pasó a un segundo acuario.Todas las muestras de gametos masculinos fueron posteriormente pasadas a un tubo de ensayo mayor. Igualmente lasde óvulos. Conseguimos por tanto un homogenizado de las muestras.
• Preparación de las soluciones base para el desarrollo embrionario.Preparación de las soluciones de cultivo.
Se tomó 0,1 ml de cada compuesto y se vertió en 100 ml de agua de mar previamente pasterizada a 70ºC; de estaconcentración (una vez bien agitada) se tomaron tres muestras de 0,1 ml y se vertieron en 10 ml de agua de mar(concentración 1/100.000), en 50 ml (concentración 1/500.000) y en 100 ml (concentración 1/1.000.000). Posteriormen-te, de cada solución se prepararon 20 preparaciones en los respectivos tubos de ensayo, en los que se depositaronsoluciones que portaban espermatozoides y óvulos. Por tanto los óvulos se fecundaban en las soluciones preparadas.
Al cabo de 72h, se comenzó a tomar muestras de cada tubo de ensayo (dos por tubo) en una cantidad de 1ml y seobservaban a microscopio, contabilizándose la presencia de mórulas y el número de células que aparecían. Se realizaron
un total de 120 observaciones a microscopía, que correspondían a 10 observaciones por cada compuesto orgánico yconcentración. La visualización se realizó de modo ordinario a 100 aumentos.
Los datos observados fueron procesados en los siguientes programas informáticos: Microsoft Excel 2000, Sigmaplot2001, Minitab 13.
Sistema inmunitario
Para su posterior utilización en las pruebas, se realizó una extracción de hemolinfa, en la zona carnosa que se encuentraalrededor de la boca.
Se obtuvieron lentamente 10 ml de hemolinfa que depositábamos en tubos de ensayo colocados en una caja con hielo paraconservar las células. Los erizos a los que se extrajo la hemolinfa, no habían sido expuestos a ningún contaminante, ya que
Figura 1. Erizo común (Paracentrotus lividus)

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 55/276
Comunicaciones
55
los hidrocarburos (gasóleo y aceite base) se les añadirían en los pocillos con las siguientes concentraciones [directa(únicamente contaminante), a 1:2, 1:100, 1:1.000, 1:1.000.000 y la muestra control] y en los dos diluyentes utilizadosDMSO y FSW. El DMSO facilita la dispersión de las sustancias orgánicas, en este caso los hidrocarburos, con la hemolinfa.
Con cada pool se realizaron las siguientes pruebas, haciéndose las mediciones a las 2 y a las 24 horas.
• Medición de la producción de radicales de nitrógeno.
La producción de los radicales de nitrógeno es un método de defensa utilizado por el erizo para eliminar sustanciasnocivas para su desarrollo. El NO es una sustancia toxica tanto para los agentes nocivos que atacan el sistema inmunitariodel erizo, como para el propio erizo, es decir, esta respuesta por parte del erizo si no logra anular el agente nocivo le puedellevar a la autodestrucción.
Se cuantifica la concentración de nitrito utilizando la reacción de Griess, cuyo protocolo seguido por nosotros es el siguiente:
- Se toman 100 ml/ pocillo de hemolinfa.- Se toma un control de 100 ml de FSW (agua de mar) en tres pocillos por cada pool.
- Se espera 30' a 15ºC (para que precipiten los hemocitos).
- Se añaden 100 ml/ pocillo de Zymosan 100 mg/ml (así la concentración de Zymosan final en el pocillo será de50 mg/ml)
- Cada pocillo tendrá de 50 mg/ml.
- Se incubar 2 h a 15ºC.
- Se transfieren 50 ml de cada pocillo a otra placa.
- Se añaden 100 ml de Reactivo A y 100 ml de Reactivo B, se deja 5' a 15ºC y se lee la absorbancia a 540 nm.
- Finalmente se mide en el espectrofotómetro (Labsystem iems Reader MF/ Type 1410/ Serie 14100-137)Se estima la concentración de nitrito presente en los sobrenadantes comparando la absorbancia a la de concentracionesconocidas de NaNO2 (1, 5, 10,25, 50 Y 100 mM). Para comprobar que el patrón utilizado funciona correctamenteañadimos en una placa 50 ml de NaNO2 y a continuación 100 ml de los reactivos A y B.
Reactivo A: 1% sulfanilamide en 2.5% de H3PO4. (50 ml)
Tabla 1. Composición del reactivo A.
Figura 2. Mórulas de 2, 4, 8 y 12 células respectivamente.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 56/276
Almoraima, 35, 2007
56
Reactivo B (Fotosensible): 0.1% N-naphthyl-ethylenediamine en 2.5% de H3PO4 (50 mi)
Tabla 2. Composición del reactivo B.
La medición se realizó con un espectrofotómetro Labsystems iEms Reader MF/ Type 1410 serie 140100_137.
El programa informático utilizado para interpretar los datos fue el Ascent software for EMS Reader MF y para el tratamientoestadístico utilizamos Microsoft Excel. Por último se realizó la misma prueba a las 48h de la exposición al tratamiento.
• Quimioluminiscencia
Los fagocitos del sistema inmunitario del erizo común al ser estimulados por algún agente nocivo o extraño, aumentansu consumo de oxígeno, lo que provoca un estallido respiratorio, generando una emisión de fotones. La medición de laenergía lumínica nos determina la mayor o menor presencia de agentes nocivos.
Este ensayo se hace en una placa de 96 pocillos. El protocolo a seguir es el siguiente:
Añadir 100 ml de hemolinfa/ pocillo.
Esperar 30 minutos a que precipiten los hemocitos (a 15ºC).
Mientras, preparamos la solución de luminol con el estímulo:
- Stock luminol: 0,018 g luminol en 1 ml DMSO. Envolver en papel de plata, y preparar siempre en el momento(no guardar).
- Solución de trabajo de luminol: 15 ml stock luminol en .5 ml FSW. Envolver también en papel de plata.
- Para el estímulo estándar (Zimosán 1 mg/ ml, concentración final en el poc.): 500 ml stock Zimosán en FSW (20mg/ mI) en 4,5 ml de solución de trabajo de luminol. Mantener asimismo en oscuridad.
Añadir 100 ml/ pocillo de la solución de trabajo de luminol con Zymosán, e inmediatamente medir luminiscencia (cuatromedidas consecutivas cada 10 min):
Medida con el Flouroskan Ascent FL Type 374, Serie 374022533-C que realiza medidas cada cinco minutos durante35 minutos.
Este tratamiento se midió a la 24 y 48 horas de la exposición.
• Viabilidad de hemocitos con rojo neutro.La viabilidad mide la cantidad de células que sobreviven al tratamiento. El rojo neutro se adhiere a la pared de lamembrana celular de las células vivas y se mide la intensidad del rojo que determina la cantidad de células vivas. Cuantomayor es la intensidad mayor es la cantidad de células vivas.
- Se añade 100 ml hemolinfa/ pocillo en una placa 96, por triplicado.
- Esperar 30' a 15°C (tiempo de adhesión de los hemocitos al fondo del pocillo).
- Añadir 100 ml de rojo neutro working solution.
- Incubar una hora en oscuridad a 15°C.
- Eliminar el sobrenadante por volcado.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 57/276
Comunicaciones
57
- Lavar con 100 ul de FSW y eliminar de nuevo por volcado.
- Añadir 100 ul de solución de lisis de los hemocitos.
- Incubar 10 minutos en agitación, en el propio Multiscan, y medir a continuación ABS a 540. Para ello, hay quetener el aparato conectado al PC (es el software que permite agitar mejor), y programar la agitación de 10 minutosa 1.020 rpm (vienen esas rpm por defecto) y luego inmediatamente la medida de ABS a 540nm . La concentraciónde hemocitos será muy baja si la absorbancia es menor de 0,2.
Soluciones utilizadas:
- Stock de rojo neutro 0,4%: 20 mg (0,02 g) de rojo neutro + 5 ml FSW. Guardar en oscuridad a RT.
- Rojo neutro Working solution: calcular su concentración para que la [rojo neutro] final en el pocillo sea de 0,004%.En este caso (100 ul de hemolinfa + 100 ul de Working solution), necesitamos que la Working solution sea al0,008% de rojo neutro (hacer una dilución 1:50 del stock en FSW).
- Solución de lisis de los hemocitos: 50% etanol- 1% ácido acético, en dH2O.
Las mediciones se obtuvieron con el espectrofotómetro anteriormente mencionado.
• Recuento de células
Realizamos un recuento de células, distinguiendo las vivas de las muertas para los erizos que se habían expuesto acontaminantes durante 48 horas.
Figura 3. Recuento de células mediante microscopio.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 58/276
Almoraima, 35, 2007
58
Para ello utilizamos estéreomicroscopios ópticos inversos, y colocamos hemolinfa en portas con rejillas. Para realizarlas mediciones dividimos las rejillas en cuatro zonas, turnándonos para realizar la estimación del número de células vivasy muertas, después realizamos la media para verificar los datos de viabilidad obtenidos anteriormente.
Las mediciones se realizaron a las 2 y a las 24 horas.
RESULTADOS
Desarrollo embrionario
• Influencia de la contaminación en el desarrollo embrionario
En el diagrama de barras se representa el promedio de embriones encontrados por ml de cada compuesto. Si describimosla variación del número de embriones según la concentración, se aprecia de mayor a menor variación, el siguiente orden:crudo, aceite base, gasoil y fueloil. Es en este último donde el número de embriones sufre ligeras variaciones de 1,E-06 a 2,E-06. En el crudo si ve afectado el número de embriones al pasar de una concentración a otra. El crecimiento a1,E-05 se da de un modo claro sólo en gasoil, por tanto, el compuesto con un impacto negativo menor a esa concentración.
A continuación se muestran los resultados obtenidos por cada compuesto y según la concentración, el número deembriones encontrados y su estado de desarrollo.
El descenso gradual del número de mórulas según el número de células que lo componen parece normal. Tanto aconcentración 1,E-06 como 2,E-06 se observan ejemplares menos y más desarrollados. Conforme aumenta el númerode células en las mórulas, éstas se presentan en menor número. A concentración 1,E-05 sólo se observan y de modo nosignificativo mórulas de dos células. Dado el valor tan mínimo, consideramos que el dato puede ser debido a lacontaminación.
El descenso gradual del número de mórulas según el número de células que lo componen parece normal. A concentración1,E-06 se observa una disminución gradual del número de mórulas. A 2,E-06 se observa esa gradualidad con la diferenciade apreciarse unos resultados similares de mórulas desarrolladas. A concentración 1,E-05 se observa un crecimientoaleatorio que determina unas condiciones estresantes, pero que al aparecer mórulas desarrolladas, nos da una razón deuna capacidad de desarrollo, aunque mínima. Aquí desechamos darle un valor nulo.
La observación de mórulas con 3 y 6 células puede considerarse anómala. Hemos querido reflejar estos datos a pesarde saber que lo lógico es considerarlas de 4 y 8 células respectivamente. Esta observación puede deberse, bien a noapreciar con nitidez otras células integrantes, bien porque en el proceso mitótico en el que están inmersas, se haya vistobloqueado en alguna fase de su desarrollo.
La gradualidad del número de mórulas comentada en anteriores casos se manifestaría si englobáramos en 4 las de 3 y4, y en 8 las de 6 y 8.
Es destacable el crecimiento tan alto de mórulas a 1,E-06. Sin embargo el crecimiento a 2,E-06 es bajo con respecto ala anterior y es nulo a 1,E-05. A este valor esta claro que la inhibición es total en cuanto al desarrollo.
Aplicando los comentarios sobre el número de células presentes en las mórulas que observamos en la gráfica anterior,se aprecia una gradualidad en el desarrollo, a excepción de las mórulas con 12 células donde existe un importantecrecimiento, tanto a 1,E-06 como especialmente a 2,E-06, donde su presencia es mayor que en 4 y 8 células.
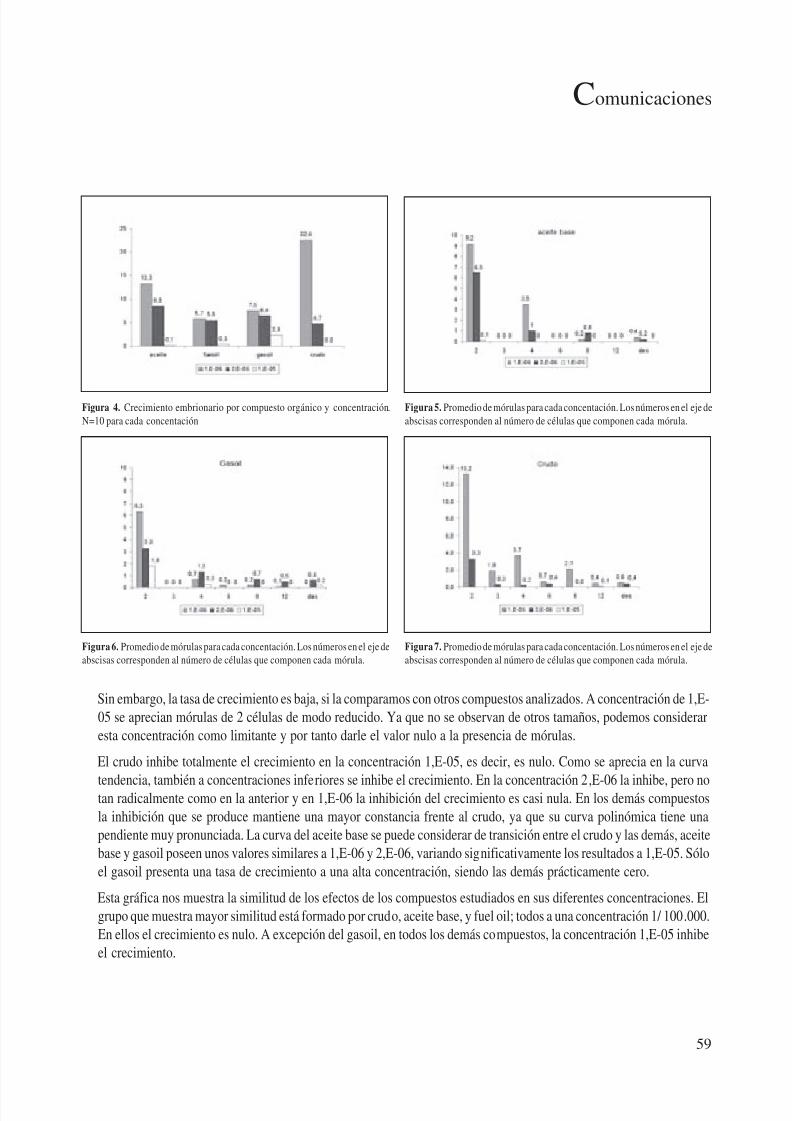
8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 59/276
Comunicaciones
59
Figura 4. Crecimiento embrionario por compuesto orgánico y concentración.N=10 para cada concentación
Figura 5. Promedio de mórulas para cada concentación. Los números en el eje deabscisas corresponden al número de células que componen cada mórula.
Figura 6. Promedio de mórulas para cada concentación. Los números en el eje deabscisas corresponden al número de células que componen cada mórula.
Figura 7. Promedio de mórulas para cada concentación. Los números en el eje deabscisas corresponden al número de células que componen cada mórula.
Sin embargo, la tasa de crecimiento es baja, si la comparamos con otros compuestos analizados. A concentración de 1,E-05 se aprecian mórulas de 2 células de modo reducido. Ya que no se observan de otros tamaños, podemos consideraresta concentración como limitante y por tanto darle el valor nulo a la presencia de mórulas.
El crudo inhibe totalmente el crecimiento en la concentración 1,E-05, es decir, es nulo. Como se aprecia en la curva
tendencia, también a concentraciones inferiores se inhibe el crecimiento. En la concentración 2,E-06 la inhibe, pero notan radicalmente como en la anterior y en 1,E-06 la inhibición del crecimiento es casi nula. En los demás compuestosla inhibición que se produce mantiene una mayor constancia frente al crudo, ya que su curva polinómica tiene unapendiente muy pronunciada. La curva del aceite base se puede considerar de transición entre el crudo y las demás, aceitebase y gasoil poseen unos valores similares a 1,E-06 y 2,E-06, variando significativamente los resultados a 1,E-05. Sóloel gasoil presenta una tasa de crecimiento a una alta concentración, siendo las demás prácticamente cero.
Esta gráfica nos muestra la similitud de los efectos de los compuestos estudiados en sus diferentes concentraciones. Elgrupo que muestra mayor similitud está formado por crudo, aceite base, y fuel oil; todos a una concentración 1/ 100.000.En ellos el crecimiento es nulo. A excepción del gasoil, en todos los demás compuestos, la concentración 1,E-05 inhibeel crecimiento.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 60/276
Almoraima, 35, 2007
60
El siguiente grupo con valores que poseen similitud sería el formado por el gasoil 1/100.000, el crudo 1/500.000, el gasoil1/500.000, el fuel oil 1/1.000.000 y por último, a más distancia, el fuel oil 1/500.000. De esta segunda asociación
podemos destacar la similitud de valores obtenidos en torno a la concentración 2,E-06, tanto en gasoil, crudo comofueloil. Observar el pobre desarrollo embrionario a la baja concentración de fueloil.
El siguiente grupo es el formado por el gasoil 1/1.000.000 y el aceite base 1/500.000.
Los dos compuestos más alejados son aceite base 1/1.000.000 y el crudo 1/1.000.000. Por tanto, el mayor desarrollo abajas concentraciones se da en el siguiente orden: crudo, aceite base, gasoil, fueloil.
F26 y f16 presentan una uniformidad tanto en sus valores globales como en su distribución por el número de célulaspresentes. c16 presenta la mayor el mayor número de mórulas. A16 sería el segundo valor en importancia. g16, g26, c26y a26 presentarían unos valores intermedios con respecto a la mayor tasa de crecimiento y la menor tasa, constituidapor c15, f15, a15 y g15.
Figura 8. Promedio de mórulas para cada concentación. Los números en el eje deabscisas corresponden al número de células que componen cada mórula.
Figura 9. Curvas tendencia de distribución para cada compuesto. Número demórulas / ml. Relacionando la tasa de crecimiento embrionaria con la concentra-ción. Se exponen las funciones polinómicas y el coeficiente de Pearson. Trazocontinuo: cruso; Trazo discontinuo (rayas): aceite base; Trazo discontinuo(puntos): gasoil; Trazo gris: fueloil.
Figura 10. Análisis de clúster, método simple. Diferente comportamiento deldesarrollo embrionario según compuesto y concentración. "c": crudo; "g": gasoil;"a": aceite base; "f": fueloil; "15": 1,E-05; "26": 2,E-06; "16":1,E-06.
Figura 11. Análisis de los componentes principales. Asociándolos por suproximidad en cuanto a la distribución de número de morulas de cada tipo. "c":crudo; "g": gasoil; "a": aceite base; "f": fueloil; "15": 1,E-05; "26": 2,E-06;"16":1,E-06.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 61/276
Comunicaciones
61
Sistema inmunitario
1. A las 2 horas
1.1. Concentración de radicales de nitrógeno
Se observa que en el índice de estimulación del gasóleo a dos horas en DMSO, es mayor a la concentración 1/1000. El índicede estimulación en FSW es mayor a la concentración 1/100, siendo las demás muy inferiores a ésta.
El índice de estimulación del aceite base en DMSO a las dos horas, es mayor en la concentración 1/100. En FSW (aguade mar) es mayor en la concentración directa.
Se aprecia como el Zym genera una respuesta mayor en todos los casos. Con el control, la mayor respuesta se da a laconcentración 1/10 E6. La presencia de DMSO permite una mejor disolución de los hidrocarburos y por tanto una mayorreacción del sistema inmune.
1.2. Quimioluminiscencia/Viabilidad
Mayor concentración en el control de aceite base con agua de mar (FSW). Mayor concentración a 1/2 en aceite base conDMSO.
Figura 12. Eje Y: valores obtenidos en la medición [NO]. Eje X: distintas concentraciones del compuesto para n = 72.
Figura 13. Eje Y: valores obtenidos en la medición [NO]. Eje X: distintas concentraciones del compuesto para n = 72.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 62/276
Almoraima, 35, 2007
62
Figura 14. Eje Y: valores obtenidos en la medición [NO/viab]. Eje X: distintas concentraciones del compuesto para n = 72 en comparación entre Zym y control (FSW).
Figura 15. Eje Y: valores obtenidos en la medición [NO/viab]. Eje X: distintas concentraciones del compuesto para n = 72 en comparación entre Zym y control (FSW)en su caso y Zym/control (DMSO) en otro caso.
Figura 16. Eje Y: valores obtenidos en la medición [Cl/viab]. Eje X: distintas concentraciones del compuesto para n = 72.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 63/276
Comunicaciones
63
Figura 17. Eje Y: valores obtenidos en la medición [Cl/viab]. Eje X: distintas concentraciones del compuesto para n = 72.
Figura 18. Eje Y: valores obtenidos en la medición [NO]. Eje X: distintas concentraciones del compuesto para n = 72. DMSO y FSW.
Figura 19. Eje Y: valores obtenidos en la medición [NO]. Eje X: distintas concentraciones del compuesto para n = 72. DMSO y FSW.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 64/276
Almoraima, 35, 2007
64
Figura 20. Eje Y: valores obtenidos en la medición [NO/viab]. Eje X: distintas concentraciones del compuesto para n = 72. DMSO y FSW.
Figura 21. Eje Y: valores obtenidos en la medición [NO/viab]. Eje X: distintas concentraciones del compuesto para n = 72 en comparación entre Zym/control (FSW)y Zym/control (DMSO).
Figura 22. Eje Y: valores obtenidos en la medición [Cl/viab]. Eje X: distintas concentraciones del compuesto para n = 72.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 65/276
Comunicaciones
65
La mayor concentración: 1/2 con gasóleo en agua de mar (FSW). Mayor concentración: 1/2 con gasóleo en DMSO a lasdos horas.
2. A las 24 horas.
2.1. Concentración de radicales de nitrógeno.
Se observa que en el índice de estimulación del gasóleo a 24 horas en DMSO, es mayor a la concentración 1/100. En elgasóleo a 24 horas en FSW (agua de mar) es mayor a la concentración 1/10 E6.
Se observa que en el índice de estimulación del aceite base a 24 horas en DMSO, es mayor a la concentración 1/2. mientrasque en FSW (agua de mar), se aprecia un mayor valor en la concentración 1/100.
En la concentración 1/1.000 los datos no son fiables debido a un error humano.
2.2. Quimioluminiscencia/Viabilidad
Mayor concentración: directa con gasóleo en DMSO a las 24 horas. Mayor concentración: 1/100 con gasóleo en FSW(aguade mar) a las 24 horas.
Mayor concentración: 1/10 E6 con aceite base en DMSO a las 24 horas. Mayor concentración : control con aceite base enFSW(agua de mar) a las 24 horas.
DISCUSIONES
El compuesto contaminante estudiado con mayor densidad y adherencia es el fueloil, sin embargo, es el compuesto quemenos se dispersa en el medio, mientras que los más dispersantes son el aceite base y el gasoil. Existe una relación entreel aceite base y el gasoil, ya que los valores de densidad, adherencia y dispersión son semejantes.
Desde el punto de vista físico, los compuestos con mayor densidad y adherencia tendrán una dispersión menor y por tantosu capacidad de contaminación se verá reducida en el espacio. Por tanto, el fueloil y el crudo serían los compuestos quecontaminarían más en las zonas donde son vertidos y el gasoil y aceite base, contaminarían en menor intensidad, pero enmayor extensión por su capacidad de dispersión. Al expandirse sobre mayor superficie, su incidencia sobre los organismos
Figura 23. Eje Y: valores obtenidos en la medición [Cl/viab]. Eje X: distintas concentraciones del compuesto para n = 72.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 66/276
Almoraima, 35, 2007
66
será mayor, especialmente sobre los sésiles o los que vivan en el fondo con una capacidad de desplazamiento limitada, comoes el caso de los erizos de mar.
Desde la no tan hipotética situación de un vertido en la bahía de Algeciras, debido al intenso tráfico marítimo, podemosseñalar las repercusiones que para las zonas supralitoral, mesolitoral e infralitoral tendría.
Por su adherencia, especialmente el fueloil y secundariamente el crudo, tendrían un impacto negativo sobre las zonassupralitoral y mesolitoral. Los ecosistemas que ocupan la franja mesolitoral serían destruidos y conforme nos introducimosmar adentro, quedarían seriamente afectados si la concentración es inferior a 1,E-06. La zona infralitoral se vería seriamenteafectada en el lugar del vertido, dadas las características de estos fluidos. En el caso del fueloil, la tolerancia es menor ytendría efectos más negativos que el crudo a igual concentración.
En los casos de gasoil y aceite base, tendrían un efecto más amplio en la zona, puesto que su capacidad de dispersión esmayor a los citados anteriormente. Dada una adherencia menor a los anteriores, su efecto sobre la zona supralitoral ymesolitoral sería menos permanente, aunque su presencia en el medio determinaría a concentraciones altas una toleranciacero. Conviene matizar que el gasoil es el compuesto que permite un crecimiento a 1,E-05 y por tanto unido a lascaracterísticas mencionadas, podría resultar el menos dañino para el medio.
En cuanto al desarrollo embrionario se ha observado que el crudo, posee la distribución más irregular de valores obtenidospara cada una de las concentraciones tratadas. Presenta la mayor tasa de crecimiento embrionario en 1,E-06 con 22,6mórulas por mililitro, la menor tasa de crecimiento a 2,E-06 con 4,7 mórulas por mililitro y en la concentración 1,E-05 unatasa cero. En este compuesto las concentraciones tratadas responden a los valores donde su incidencia es letal para elcrecimiento (1,E-05) y cercana a la inhibición la tasa de 2,E-06. Por tanto, a altas concentraciones la inhibición es total y
a bajas concentraciones la inhibición es nula.El fueloil presenta una tasa de crecimiento similar a las concentraciones 1,E-06 y 2,E-06. Ambos valores representan un 25%aproximado del valor medido en el crudo a 1,E-06, muy por debajo del desarrollo posible. A concentraciones menores se debedar un mayor crecimiento embrionario. A 1,E-05 la tasa embrionaria es nula y por tanto idéntico valor que en el crudo.
El aceite base presenta la segunda mayor tasa de mórulas a 1,E-06 y la mayor tasa a 2,E-06. Considerando los tres tiposde soluciones que han sido estudiadas, el aceite base presenta la mejor proporcionalidad de crecimiento. Se observa unainhibición proporcional en la medida que aumenta la concentración, hasta llegar al crecimiento cero, que es el consideradoa concentración 1,E-05.
El gasoil es el único compuesto de los analizados que presenta un crecimiento de mórulas a 1,E-05. Por tanto es el menoscontaminante de todos, si bien es cierto que, los valores de aparición de mórulas en gasoil en las concentraciones 1,E-06
y 2,E-06 serían los terceros en importancia. La concentración limitante de mórulas se halla por debajo de 1,E-05. Losvalores de crecimiento son proporcionales: a menos concentración de gasoil, mayor número de mórulas.
Por las propiedades físicas, se encuentra una similitud entre el aceite base y el gasoil, sin embargo, las pruebas biológicasmanifiestan claras diferencias en cuanto a la tolerancia al gasoil por parte de las mórulas generadas.
Atendiendo al contenido en azufre de cada uno de los compuestos señalados, se aprecia que es más alto en fueloil, ensegundo lugar en aceite base y en tercer lugar en el crudo. Se puede establecer una concordancia entre estos valores y losresultados obtenidos en los tres compuestos, donde, a concentración 1,E-05, no se aprecia crecimiento. El gasoil manifiestacrecimiento a esa concentración y es, de los cuatro compuestos tratados, el que menos proporción de azufre presenta, portanto, se puede deducir que el azufre es uno de los elementos que inhibe el crecimiento de las mórulas.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 67/276
Comunicaciones
67
Sobre el desarrollo del número de células en las mórulas, los resultados obtenidos muestran una relación que consideramosnormal, puesto que el mayor número de mórulas corresponderá a las que posean menor número de células.
Dado que la fecundación de los óvulos se ha dado en las soluciones preparadas, quedaría por resolver si la inhibición seda en la fecundación, o si ésta se da en las sucesivas divisiones mitóticas del desarrollo embrionario. La presencia demórulas con distinto número de células manifiesta que se da en el crecimiento.
Hemos profundizado en el conocimiento del sistema inmunológico básico del erizo, estudiando la diferente respuesta a losdistintos agentes y concentraciones que actúan sobre los erizos.
Dadas las características de los hidrocarburos, compuestos hidrófobos, podía ser de interés comprobar la incidencia de estoscompuestos sobre los erizos, bien en agua de mar libre (FSW) o bien en DMSO, medio que permite una mayor dispersión
en la hemolinfa. Los compuestos utilizados han sido aceite base y gasoil para el estudio in vitro.- Concentración de radicales de nitrógeno
Se observa que en el índice de estimulación de aceite base en DMSO a dos horas y 24 horas existe una respuesta similar.Muestra una ascenso de directa a 1/2, luego se produce un descenso en concentraciones medias(1/1.000) para, acontinuación, ascender con concentraciones bajas.
En 2 horas, se aprecia que el mayor índice de estimulación corresponde a la concentración 1/100, que se contrapone conel nivel observado en la misma concentración (1/100) en 24 horas.
En el índice de estimulación del aceite base en FSW a dos horas y 24 horas obtenemos que, a las dos horas existe unadisminución de la estimulación conforme se disminuye la concentración (a altas concentraciones no se produce estímulo
alguno; a 1/100 se produce un gran aumento que disminuye progresivamente en concentraciones más bajas), al contrariode la que sucede a las 24 horas(aumento progresivo al disminuir la concentración)
En el índice de estimulación del gasóleo en FSW a dos horas y 24 horas se aprecia que, a las 24 horas se produce uncrecimiento progresivo a menor concentración siendo nulas las concentraciones directa y control . A las dos horasasciende hasta 1/100, baja en la concentración 1/1.000 y una ligera subida en las otras concentraciones.
En el índice de estimulación del gasóleo en DMSO a dos horas, se aprecia una estimulación similar en todas lasconcentraciones, aunque en 1/2 y 1/1.000 la estimulación es ligeramente superior. Respecto a 24 horas se observa unaespectacular estimulación a 1/100 disminuyendo a 1 /1.000 y aumentando en el control.
En cuanto al NO/viabilidad observamos que tanto a las dos y las 24 horas el Zymosán alcanzan mayores valores queel control (DMSO y FSW). Las mayores diferencias se encuentran entre el Zymosán y el control(FSW) en el aceitebase y el gasóleo a 24 horas, siendo el control muy inferior en estas gráficas.
- Quimioluminiscencia/viabilidad
En gasóleo dos horas y 24 horas en DMSO se observa una distribución similar en la mayoría de los casos(decrecimientohasta concentraciones medias y crecimiento a partir de concentraciones inferiores).
En el gasóleo en FSW a las dos horas, los valores obtenidos son muy pequeños, salvo un pico elevado en la concentracióna 1/2, mientras que en 24 horas los valores son mucho más elevados y se mantienen constantes.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 68/276
Almoraima, 35, 2007
68
En el aceite base en DMSO a dos horas y 24 horas, se pueden observar que los resultados que obtenemos son inversos(cuando en 2 horas se produce un aumento, en 24 horas se produce una descenso)
En aceite base en FSW a dos horas y 24 horas, se observa un ritmo casi idéntico en la gráfica (aceite base, gasoil).
Podemos apreciar que el índice de estimulación en presencia de DMSO cuando la concentración es directa, es cero enel aceite base, lo que da entender que es letal mientras que el gasóleo manifiesta respuesta en exposición directa, portanto,es menos nocivo que el aceite base.
Sin embargo en agua libre observamos un comportamiento inverso. El aceite base se manifiesta menos nocivo que elgasóleo como se puede apreciar en las gráficas correspondientes. Por tanto en el mar, el gasóleo a iguales concentra-ciones tiene un carácter mas letal que el aceite base. En combinación con otros compuestos el aceite base se presenta
mas nocivo que el gasoil.
BIBLIOGRAFÍABARBERO et al. Técnicas experimentales de Laboratorio. C.E.P.1994.0 La Línea de la Concepción (Cádiz).CATOIRA, J. L. Prospección, análisis y cartografía de macroalgas y erizo de mar en el litoral de Galicia.1993. Consellería de Pesca e Asuntos Marítimos.CERDEIRA, J. D. El Erizo de mar (Paracentrotus lividus) en la costa gallega. 2001. Delegación territorial de Lugo. Xunta de Galicia.CONSEJERÍA DE PESCA Y AGRICULTURA.Estudio Integral de la Pesquería del Erizo de Mar (Paracentrotus lividus) y la Anémona Marina (Anemonia sulcata)en la Provincia de Cádiz. 2001. Dirección general de pesca y acuicultura. Junta de Andalucía.ENOSA. Prácticas de Microscopía. 1975. Lasgón S.L.DÍAZ DE RADA, V. Técnicas de Análisis Multivariante para Investigación Social y Comercial. 2002. RA-MA Editorial.PEMEX, El Petróleo. 1988. Anuario Estadístico de PEMEX.SALA, E., Ribes, M., Hereu, B., Zabala, M., Alvà, V., Coma R., Garrabou, J. 1998. "Temporal variability in abundance of the sea urchins Paracentrotus lividus and
Arbacia lixula in the northwestern Mediterranean: comparison between a marine reserve and an unproctected area". Marine Ecology 168: pp. 135-145.
SALVADOR FIGUERAS, M. Análisis de conglomerados o cluster , 2001. [en línea] 5campus.org, Estadística <http://www.5campus.org/leccion/cluster>WEISZ, PAUL B. La Ciencia de la Zoología. 1982. Ediciones Omega.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 69/276
Comunicaciones
69
INTRODUCCIÓN
Para casi todos los viajeros que visitaron la península Ibérica durante los siglos XVIII y XIX, Gibraltar constituyó unobjetivo y meta de obligado cumplimiento. Desde o hacía allí, las travesías hacia ciudades como Cádiz o Málaga serealizaron por mar, o mucho más frecuentemente mediante caballerías atravesando la geografía del interior del campo deGibraltar, del litoral, del Estrecho, o de zonas próximas como la Serranía de Ronda o la costa mediterránea. Es por elloquizás por lo que se disponga de esta zona geográfica, una mayor y concisa información ambiental de la misma, frente aotras que fueron visitadas con una menor frecuencia e interés.
En los textos que estos elaboraron se encuentran interesantes comentarios y observaciones sobre el medio natural de laszonas que recorrieron, sobre la fauna y vegetación existentes, las condiciones climáticas reinantes así como interesantesdescripciones de espacios naturales que poco o nada conocidos por entonces, despertaron fuertemente su interés. Es porello por lo que sitios de tan elevado valor geográfico y ecológico como el peñón de Gibraltar, la bahía de Algeciras, la vía
de La Trocha, la antigua laguna de La Janda, el río Guadiaro, los espacios de la cueva de El Gato y el Hundidero, la sierraBermeja o la Hedionda fueron descritos con cierta precisión. Y con un carácter más general también lo fueron las interesanteobservaciones que, sobre el clima y las nieves perpetuas que cubrían por entonces algunas de nuestras montañas máspróximas, efectuaron estos mismos viajeros.
Estas descripciones ayudan a conocer la evolución y transformación sufridas por nuestra naturaleza desde aquella época,así como permitir aproximarnos al pensamiento medioambiental de la sociedad de por entonces, en un país atrasado, exóticoy muy apartado de la modernidad europea, antes de la difusión y consolidación de textos decisivos como El Cosmos deA. Von Humbolt en 1858, el Origen de las especies de C. Darwin (1859) o Los principios de geología de C. Lyell de 1830.
En este trabajo realizamos un recorrido cronológico que nos lleva desde el texto de Guillermo Bowles de 1789, al de A.Chapman y W. S. Buck de 1893, como representantes estos, de los últimos viajeros naturalistas.
OBSERVACIONES SOBRE EL MEDIONATURAL DEL CAMPO DE GIBRALTAR YZONAS PRÓXIMAS EN LOS TEXTOS DEVIAJEROS POR ESPAÑA (SIGLOS XVIII Y XIX)
José Manuel Recio EspejoAula de La Naturaleza Nicolay Masyuk de la Universidad de Córdoba

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 70/276
Almoraima, 35, 2007
70
ANÁLISIS DE TEXTOS
Como hemos comentado la primera obra por nosotros analizada ha sido la de Guillermo Bowles (1789), quien contactó enParis en 1752 con el famoso marino y científico español Antonio de Ulloa, viniendo a España llamado por su compatriotael ministro irlandés de Carlos III Ricardo Wall, para reconocer las minas del reino, en especial las de Almadén. Apoyadoen la obra Viaje de España de Antonio Ponz (1786), con dos discípulos y algunos compañeros recorrió grandes zonas dela Península habiéndonos legado una primera edición de sus experiencias en un texto de 1773. En relación al Campo deGibraltar comenta lo siguiente: “Hay montañas enteras en España compuestas de piedra caliza, como la montaña deGibraltar, a la cual disolvería seguramente una lluvia de ácido. Lo mismo es la de Morón, donde se hace la mejor cal queyo conozco”.
En otro orden de cosas encontramos:
Habiendo caminado dos horas por entre estas montañas blancas calcaricas, entramos en otra cordillera llamada Sierravermeja, que corre al poniente hacia Málaga desde su principio llamada Cresta de gallo. Hay en esta sierra unasingularidad muy rara, y es que extendiéndose paralelas y sus bases se tocan, la una es roxa y la otra blanca. La primeraaunque un poco más alta, no conserva permanentemente la nieve; y la otra esta casi siempre cubierta de ella, de suerteque en el verano surte a todos los países circunvecinos para enfriar las bebidas. La blanca produce solo alcornoquesy encinas, y la roxa no tiene ninguno de estos árboles, y está cubierta de abetes. Aquella contiene únicamente minasde hierro en pelotillas y ésta minas de otros muchos metales, excepto hierro. En fin, las aguas minerales de la blancason marciales y vitriólicas, y las de la roxa sulfúreas, alcalinas y hieden como las de Coterets en los Pirineos de Francia.
Richard Twiss efectuó una estancia en nuestro país de unos diez meses, entre el 17 de noviembre de 1772 y el 7 septiembrede 1773, de los cuales, tres semanas las pasó en Gibraltar. De lo más interesante que encontramos en su texto es lo relativo
a la zona de La Janda, la cual visita el día de17 de julio, comentando :Después de haber recorrido cinco leguas en unas casi ocho horas por un camino montañoso a través de alcornocales,llegamos hasta unas cabañas de barro, a las cuales se dignifica con el nombre del pueblo de Tahivilla. Después denuestra siesta, fuimos a Vejer de la Frontera, que está a tres leguas, atravesando una llanura circular de alrededor unalegua de diámetro, llena de cigarras y saltamontes y dejando una pequeña laguna llamada La Janda, que llegamos aver a la derecha.
Desde Gibraltar hace varios intentos para pasar a Marruecos; cuando el viento lo permite desembarca en la costa de Tetuán,donde nos cuenta: “Una ciudad que está a unas cuatro millas de distancia, situada en la ladera de una colina, como pudeclaramente percibir desde donde nos encontramos”. No le permitieron entrar por orden del gobernador ya que ningúncristiano podía entrar en la ciudad por el asesinato de una mujer mora. Por ello dice finalmente: “Despedí a mi guía y volví
a la playa regresando a Gibraltar.Este autor realizó una de las primeras observaciones que hemos encontrado sobre la cueva de El Gato y el río Guadiaro,afirmando que “A dos leguas de Ronda observé que un torrente vertía en el río desde una caverna que estaba en el costadode una montaña que había enfrente”.
De un carácter más general pero estrechamente relacionado con la Comarca en la que nos centramos, son los interesantecomentarios realizados sobre las condiciones climáticas reinantes por entonces, relativas a las nieves de Sierra Nevada,una constante en casi todos los autores estudiados y que luego serán analizadas de nuevo, sobre las que escribe: “Suscumbres están coronadas con nieve probablemente coetánea a las montañas y son tan altas que desde la cima de algunasque resultan accesibles se puede divisar el estrecho de Gibraltar, la costa de la Berbería y las ciudades de Tánger y Ceuta”.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 71/276
Comunicaciones
71
Francis Carter en su obra A journey from Gibraltar to Málaga recorrió nuestra geografía por el año de 1773. Sobre elEstrecho dice lo siguiente:“El fenómeno más destacado de este paso es la corriente constante que tiene desde el Atlántico”,fenómeno que por entonces era comentado en la comunidad científica; describió los pasos migratorios de aves y sobre laotra orilla, sobre Abila, escribió: “La llaman también los moros actuales Alcudia y los españoles Sierra Jimena o montañade los simios”.
La cueva del Gato vuelve a ser objeto de comentarios por parte de este viajero, esta vez mas extensos y específicos llegandoa afirmar que “merece el primer puesto entre las maravillas de la serranía de Ronda; tiene casi una legua de larga y una alturaincreíble; el río Guadiaro, como ya he dicho, corre en su interior, añadiéndosele una corriente considerable que nace es estamisma cueva…”. Habiendo penetrado un cuarto de legua al parecer tuvieron que detenerse ante la existencia de un profundolago, en cuyas orillas se veían al parecer las ruinas de un gran edificio.
Sobre sierra Blanquilla comenta que eternamente está cubierta de nieves y que al noreste destaca la sierra del Pinal, llamadaJamares por los moros, cuyos pinos proporcionan madera a Ronda y a los pueblos vecinos, constituyendo un refugio parala cabra montés, el jabalí y el lobo.
Joseph Townsend (1791) fue quizás el viajero con una mayor formación científica de todos los que nos visitaron. Estudióen Clare Hall, Cambridge y medicina en Edimburgo. Ordenado en la iglesia y Rector de Pewsey, mostró un interés por lapaleontología y la geología. En 1770 visitó Francia, Holanda y Flandes.
Entra en la península Ibérica desde Francia y en Madrid se hospeda en el mismo hotel de Richard Twiss, 20 años antes.Conoce al director del gabinete de Historia Natural del Buen Retiro de Madrid, Eugenio Izquierdo. De Madrid a Sevillaviaja el 15 de febrero de 1787, efectuando algunos comentarios sobre las recién creadas poblaciones carolingias (nueve
en total). Del viaje de Cádiz a Málaga lo más significativo que hace destacar es lo relativo al régimen de brisas y la posibledesecación del mar Mediterráneo.
Sobre Gibraltar tan solo omenta: “Y antes del anochecer pasamos junto al estrecho de Gibraltar, donde tuve el placer deobservar la altiva roca ante cuya visión cualquier corazón británico debe sentirse jubiloso recordando a sus bravosdefensores…”. Otra discusión científica interesante que aborda lo hace en torno a los suelos y el salitre, pero con unaterminología antigua y poco científica, citando a G. Bowles y con descripciones geológicas muy en la línea de este(esquistos, calizas, descripción de minerales, etc). Varias veces habla de la gran convulsión que ha afectado a la Tierra. Almargen de esto no hemos encontramos en su obra nada significativo sobre la Naturaleza del campo de Gibraltar, ni de losespacios naturales existentes en sus zonas más próximas.
John Carr nos visita en 1809. De su obra ( Descriptive travels in the southern and eastern parts of Spain and the Balearic
Isles in the year 1809), aun no traducida, hemos podido entresacar algunas connotaciones ambientales de sumo interés.Llega a Cádiz desde Londres, en el mes de julio; el día 18 ya puede ver desde el mar la especial forma cónica de la elevaciónde Medina Sidonia, según él denominada Turk´s Cap.
Ya dentro de los límites campogibraltareños se detiene en las peculiaridades del viento de levante comentando: “…and astrong Levanter blowing, one of the effects of which is to cover the elevated summits of the rocks to the north-ward withthick foggy clouds”. Y sobre Gibraltar, lo siguiente:“The travellers will do well to pay a visit to Catalan Bay, situated at the base of the eastern side of the rock, which is there perfectly inaccessible: this spot is truly romantic and beautiful…we dined in refreshing coolness, although it was sultry hot on the other side of the rock.”

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 72/276
Almoraima, 35, 2007
72
Durante su estancia en Cádiz relata algunas de las costumbres sociales de la época, de las cuales una de ellas es muyilustrativa para aproximarnos a entender las condiciones climáticas de por entonces así como la evolución sufrida por estecomo consecuencia de la revolución industrial del siglo XX: “From Alameda, we were invited to an ice-house, called aneveria , the largest and most fashionable in the city, and frequented by ladies of the highest rank… Some were drinkingagras, a delightful beverage made of the juice expressed from the unripe grape and the tendrils, iced”.
Más tarde al hablar de Sevilla vuelve hacer referencia al mismo aspecto afirmando: “ Here as at Cádiz, iced water withsugar-plumb in it is a luxury with the lower orders”. Y en Granada escribió: “The neverias here are much frequented, and the ices are deliciously made. A glass containing about half a pint may be had for the value of sixpence”.
William Jacob (1809-1810) recorrió nuestras comarcas durante la invasión napoleónica. En el prólogo de la edición pornosotros manejada se comenta que Jacob posó su mirada en la naturaleza inhóspita, y en verdad si que lo hizo, siendo unode los viajeros que ha dejado unas de las mejores y mas exactas descripciones de algunos de nuestros espacios naturales,y sobre todo el que más se aproximó a describir y cuantificar las nieves perpetuas que por entonces engalanaban nuestrasmontañas más elevadas.
“Desde Los Barrios empezamos a ascender la sierra, que aunque no es tan alta, es igualmente agreste con aquella cimallamada La Trocha a la espalda de Algeciras. Nos llevó cuatro horas alcanzar la cumbre de la sierra”.
La Trocha constituía una segunda opción para llegar o salir del campo de Gibraltar en dirección a Cádiz, frente al itinerariodel litoral; discurría a través del “bosque de alcornoques” como se comentaba, mucho más corto pero arriesgado por losproblemas de asalto y ataques de bandoleros existentes en esos montes. Partía este camino de La Janda, vía Tahivillaatravesando las areniscosas sierras, y es por ello por lo que provecha para comentar lo siguiente sobre la antigua laguna
de La Janda : “Desde esta maldita venta (Tahivilla), atravesamos una hermosa llanura que las últimas lluvias habíanhumedecido hasta enfangarla, pero que parecía totalmente inculta. Unos cuantos bueyes desperdigados fueron el únicoganado que observé y no pude descubrir ningún vestigio habitable hasta que tras cuatro horas cabalgando alcancé Vejer”.
Sobre las nieves perpetuas llegó a escribir lo siguiente:
Los puntos más altos de toda la sierra son aquellos de la Sierra Nevada junto a esta ciudad. Uno llamado Mulhacéncon una altura de unos 12.762 pies sobre el nivel del mar, y otro Picacho de Veleta que se eleva hasta los 12.456 pies.La línea en la que empiezan la nieves perpetuas es de 9.915 pies sobre el nivel del mar, y desde esta línea hacia arribadesaparece la vegetación completamente.
En otro pasaje de su viaje encontramos:
Alcanzamos El Burgo y ascendimos otra montaña de una altura prodigiosa. En las partes mas altas por las que pasamos,
llegamos a alcanzar justo la línea de las nieves perpetuas, que como ya mencioné con anterioridad, está a unos 10.00pies sobre el nivel del mar y se trata de la elevación más alta que ascendimos. Cuatro de los mas altos picos que enun día claro son visibles desde Cádiz, retiene nieve que se conserva a lo largo del verano en las cavernas, y se envíaempaquetada en barcias para Cádiz, Sevilla y Gibraltar.
Su curiosidad e interés por el medio natural hizo que nos halla dejado unos pioneros y valiosísimos comentarios sobre otrosespacios naturales andaluces tales como El Torcal: (“Al lado derecho vimos todo un espectáculo llamado El Torcal. Estásituado en la cumbre de una elevada montaña y tiene el aspecto de una imponente ciudad en ruinas…”), el karst ypaleoalteritas tropicales de la Serranía de Ronda (“La gran cantidad de mineral de hierro de estas montañas se encuentraen unas pequeñas bolas no mayores que un proyectil…”), sobre Los Infiernos de Loja, años antes que Washington Irving,

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 73/276
Comunicaciones
73
sobre la laguna de Fuente de Piedra y sobre el sistema Hundidero-Gato el cual trascribimos integramente dado su interésy belleza:
Tan pronto como el Guadiaro abandona las rocas de Ronda recibe el caudal tributario del Guadaleví, el Culebras yel Alcobacén, y atraviesa la llanura con este aumento de agua, hasta que a una legua de distancia se precipita desdeunas elevadas rocas, formando una cascada de una belleza sorprendente, y es recibido dentro de una caverna dondese pierde a la vista. La entrada a la caverna que se llama Cueva del Gato, es muy elevada; quienes la habían exploradome informaron de que tras avanzar alrededor de una milla, dentro se abre un amplio lago sobre cuyas orillas hay ruinasde un antiguo edificio. Me dijeron que más allá del lago, que es de una profundidad insondable, el pasaje hecho porel agua es demasiado pequeño para admitir cualquier acceso más allá, y que a veces, la dificultad de descargar todael agua por esta apertura hace que el lago suba hasta el techo. La cueva termina unas cuatro millas desde el comienzo,donde el Guadiaro de nuevo se hace visible, y continúa su curso por Gaucín hasta que entra en el Mediterráneo.
Machenzie A. Slidell viaja en 1826 e Inglis, D. Henry lo hace en 1830 (algunos capítulos de las obras originales han sidotraducidos por Clavijo Provencio). No son autores que aporten abundante información naturalista y geográfica sobrenuestra región, habiendo tan solo encontrado pasajes como estos: “Para viajar de Cádiz a Gibraltar el plan normalmenterecomendado es salir de Cádiz por la tarde, dormir en Chiclana y al día siguiente llegar a Tarifa o a la venta de la Jondal;el tercer día, un corto trayecto lleva al viajero hasta Gibraltar” (Inglis, 1830)
No aporta este autor ningún comentario sobre la laguna de La Janda (¿Jondal?) a pesar de que la recorre durante el mes denoviembre, tan solo esto: “La región por la que pasamos después de partir de la venta donde desayunamos (Vejer) estabaen su mayor parte inculta: por aquí y allá se veía algún campo de maíz y observé dos o tres arados trabajando; algunas vacas,caballos y ovejas daban vida ocasionalmente al paisaje” (¿La Janda?). Si lo hace por el contrario sobre La Trocha: “Haydos caminos para ir a Gibraltar, uno por Tarifa de unas 34 millas, y otro bastante mas corto, por una solitaria venta”.
Lo mismo ocurre con Mackenzie (1831) quien llega a Vejer durante la época de primavera-verano (le llama la atención elcanto de los ruiseñores y las flores aromáticas) y sale de madrugada hacia Los Barrios por La Trocha en dirección aGibraltar. Describe someramente lo abrupto de este recorrido, donde proliferan los barrancos y desfiladeros y las historiasde salteadores. Después de una cabalgata de cincuenta millas llega a dormir en pleno recorrido en una barraca de piedrasy barro cubierta tan solo de paja (¿la Jondal?). Cuando ve Gibraltar comenta: “Hay algo singularmente formidable en laapariencia de la Roca, ya sea vista de cerca o a distancia. Mirándola desde el este o desde el oeste, muchas personas handescubierto en su forma la ruda silueta de un león recostado. No hace falta recordar su fuerza natural o artificial, ni que elleón es el emblema de Gran Bretaña para hacer fácil la asociación”.
El más conocido de los viajeros del XIX es sin duda Richard Ford. Estuvo en España, principalmente en Sevilla y Granada
entre 1830 y 1833, publicando su obra en 1845. Siendo conocedor de la obra de G. Bowles, con sus recorridos a caballose apartó de las rutas tradicionales de los románticos y por ello nos ha dejado una abundante información sobre el paisajey la Naturaleza de la Andalucía de entonces.
En lo que atañe al Campo de Gibraltar y zonas cercanas y excluyendo las observaciones y comentarios efectuados sobrea la Roca analizados por nosotros en otro trabajo, comentó, por ejemplo, sobre el río Guadiaro y el Tajo de Ronda: “El Tajo,o corte, es su característica más marcada. El Guadalvin, el arroyo profundo que mas abajo se llama el Guadairo, rodea aRonda de la misma manera que el Marchán rodea Alhama, el Tajo a Toledo y el Huéscar y el Jucar a Cuenca”. Mas adelantecomenta: “Se pueden hacer excursiones a Ronda La Vieja, a la pintoresca caverna, La cueva del Gato, que se encuentraa dos leguas al noroeste, de donde sale un riachuelo que fluye hasta perderse en el Guadairo”.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 74/276
Almoraima, 35, 2007
74
Sobre el Estrecho retoma el análisis de Towsend comentando:
Una corriente constante entre desde el Atlántico a razón de dos millas y media por hora. Algunos han supuesto laexistencia de una corriente submarina, para aliviar al Mediterráneo de esta aportación de agua, además de la quesuponen todos sus ríos, desde el Ebro hasta el Nilo. El doctor Halley sin embargo ha calculado que la cantidad de aguaevaporada y absorbida por el sol es superior a toda la que recibe el Mediterraneo, el cual ciertamente, ha bajado algoen la costa oriental de la península.
Por otro lado y describiendo el itinerario de Ronda a Gibraltar, podemos encontrarnos: “Súbase la colina (sale de Gaucín)hasta el camino de Ximena y no tardaremos en torcer a la izquierda, por la boca del León, hasta el Bosque de Alcornoques,y desde allí hasta San Roque”.
De Cádiz a Gibraltar, habla in extenso de la bahía de Cádiz, y de La Janda en Vejer (Bekkeh): “Una milla hacia el interiorestá la Laguna de Janda. Cerca de este lago, Tarik el 30 de abril de 711, se enfrentó a don Rodrigo, el último de los godos.Aquí comenzó, el 19 de julio, la batalla que fue decidida el 26 del mismo mes de junio junto al Guadalete, cerca de Jerez”.Y continua diciendo:
En la venta de Taibilla el camino se bifurca; el que sigue a la izquierda conduce a la Trocha. En la torre de La Peñadel Ciervo, se abre la magnífica costa africana. África, que nos es tierra de desérticas arenas, se levanta bruscamentedel mar con tremendo ímpetu coronada con las nieves eternas del Atlas inferior; ante nosotros yacen dos continentes;hemos llegado al extremo del mundo antiguo, un agosto golfo divide las tierras del conocimiento, la libertad y lacivilización de las regiones vírgenes de la ignorancia bárbara, la esclavitud, el peligro y el misterio.
De Gibraltar a Málaga recomienda el camino de herradura que sigue la costa, pasando por La Línea, cruzando el peligrosovalle del Guadairo: “En las colinas a la izquierda está Manilba, la Hedionda, cuya fétida fuente ofende a la nariz y al paladar,
pero beneficia al estómago; el olor y el gusto, según la leyenda local, se atribuye al suspiro de adiós de un demonio del agua,el cual, al ser expulsado por Santiago, se evaporó como un abogado moribundo con un sulfúrico tañido”.
Por último y sin dejar a un lado las alusiones al norte de Marruecos escribió: “No debería perderse el cruce del estrecho,ni dejar de poner el pie en Africa; el contraste es más sorprendente incluso que el cruce de Dover a Calais. La excursióna Berberia es tan fácil como interesante. La caza de codornices y jabalíes es excelente; un vaporcito va de Algeciras a lospuertos vecinos, Tánger y Tetuán”.
Y sobre las nieves de por entonces nos dejó este párrafo:“La Sierra Nevada con su diadema de nieve, se levanta hasta casitrece mil pies de altura sobre el nivel del mar. De esta manera bajo una latitud de treinta y siete grados, la nieve eterna yel clima de África se juntan en un solo lugar”.
William George Clark, pastor protestante del Trinity College de Cambridge, viaja a España en el verano de 1849. El titulode su obra (Gazpacho o meses de verano en España) puede en un principio no ajustarse a nuestros objetivos, pero muy alcontrario en sus páginas hemos podido encontrar una de las más valiosas descripciones del medio natural de la Andalucíaque visitó. Citando la obra de Richard Ford, se siente atraído como todos por las nieves de Sierra Nevada, donde llegaincluso a realizar una escalada y estancia en dicha sierra de varios días, haciendo alusión a la visita que realizó el botánicoalemán Moriz Willkomm:
Cerca de la cima no crece nada, excepto un tipo de hierba acolchonada y una manzanilla enana muy apreciada por losrecolectores. Sin embargo la sierra es rica en tesoros botánicos. Hace dos años, un alemán se estableció en una cuevaen la montaña, con el propósito de recolectar plantas. Empleó a un ayudante de Granada, pero el hombre se marchópronto pensando que su amo estaba embrujado. A pesar de esto, el botánico vivió solo durante algunos meses en lacueva… Varias personas en Granada me mencionaron el asunto, pero ninguna recordaba su nombre.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 75/276
Comunicaciones
75
De sumo valor es la descripción que realiza del complejo kárstico Hundidero-Gato, el cual trascribimos íntegramente porsu valor científico e interés excursionista:
Descendimos por un estrecho desfiladero entre olivos hasta el valle, y luego anduvimos algo más de una legua, juntoa un arroyo, hasta la Cueva del Gato, cuyo solo nombre ya sugiere dificultades. La boca está situada a más de cienpies de altura, y pide a gritos que la dibujen. Un arroyuelo del agua más limpia y pura gotea, deslizándose limpiamente,en modesta contribución a la fertilidad del valle. Es como entrar en una catedral, o mejor aún, como el templo que laNaturaleza ha construido a Dios y a sí misma, su agua es más pura que la bendita por los sacerdotes y sus sonorascascadas más melodiosas que las notas de un órgano. Pero los españoles no son adoradores de la Naturaleza, y la Cuevadel Gato no se usa para el esparcimiento ni para orar.
Ascendimos a Montejaque; en los alrededores hay una gran cantidad de rocas desprendidas, y en un saliente rocoso
cercano, una torre ruinosa, como un centinela que hubiera muerto en su puesto. Girando a la derecha bordeamos unasolitaria masas de roca, llamada el Pico del Zumidero, y alcanzamos la garganta del mismo nombre, que era el objetivode nuestra exploración. Garganta a la que sólo se puede acceder en verano y con tiempo seco, pues aún conservabamarcas visibles del reciente paso de un furioso torrente.
El entorno abrumó mi capacidad de admiración. Nos habriámos paso a través de una grieta en la montaña de no másde veinte o trienta pies de ancho, mientras que los pétreos farallones que se elevaban perpendicualres a cada ladoalcanzan una altura de varios cientos de pies. En mi afán de describirlo no quisiera caer en forzados símiles, pero deuna cosa estoy seguro, ni en los Alpes se puede ver una garganta como la del Zumidero. Nunca antes había oído sunombre, y creo que ésta será la primera vez que aparece impreso. Tras media hora de lucha llegamos a la boca de lacueva, donde el agua se precipita y sale por la cueva del Gato al otro lado de la colina. La boca de la cueva estácompletamente oscura y el ángulo de descenso se aproxima peligrosamente a la vertical. No intentaría la proeza niaunque me asegurasen que encontraría todos los tesoros del sultán Soleymán… Volvimos a ascender y cuandoalcanzamos la cresta de la colina que mira al este, una vista gloriosa estalló en todo su esplendor ante nosotros, cerrandouno de los mejores días de mi vida, delicias que hubieran sido dobles, de compartirlas con un amigo.
Las obras de George Borrow (The Bible in Sapin, 1842) quien nos visitó en 1836, del francés Théophile Gautier (Tras losmontes, 1840), Hans Christian Andersen en su Viaje por España publicado en 1863, o el de Eugéne Poitou (Voyage enEspagne. Alfred Mame et Fils Editeurs 1869) no aportan nada de consideración en la relativo a la naturaleza y los paisajesde la comarca del Campo de Gibraltar. Tan solo destacar las visitas que realizan G. Borrow y H. C. Andersen al norte deÁfrica, la descripción de un enorme incendio que llegan a contemplar en las sierras tarifeñas, o la plaga de langosta queafectaba a esta misma zona. La ascensión a Sierra Nevada que efectúa T. Gautier y los comentarios que por primera vezpor razones cronológicas, efectua E. Poitou sobre la obra de Alexander von Humbold El Cosmos, máximo exponente delos viajeros científico-naturalistas decimonónicos.
Terminamos con la obra de Chapman y Buck (Wild Spain), repleta ya de abundantes descripciones naturalistas de cortemoderno, en cuyo primer párrafo comentan: “Pero en ninguna parte de Europa aparecen ártico y trópico tan cercanos eluno del otro como en España, contrastando, por ejemplo, la austera grandeza de Sierra Nevada, cubierta de nieves eternas,con la exuberancia casi tropical de las playas mediterráneas que se encuentran a sus pies”.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 76/276
Almoraima, 35, 2007
76
SÍNTESIS FINAL
No necesitan los textos antes expuestos el realizar unos especiales comentarios finales, ya que por si solos reflejan conclaridad las impresiones de los autores que la escribieron. Tan solo y a modo de síntesis podríamos hacer resaltar que elpaisaje salvaje de las sierras algecireñas a través de La Trocha impresionó a todos los viajeros que la recorrieron, así comola planitud de la depresión de La Janda y los interesantes humedales por entonces allí existentes.
Los baños de la Hedionda, El Torcal o la laguna de Fuentepiedra fueron espacios naturales visitados expresamente, y sinlugar a dudas todos quedaron maravillados e impresionados por la entidad e importancia del complejo kárstico del ríoGuadiaro, el Hundidero y la cueva del Gato.
Por su especial interés resaltamos las estancias que algunos de estos autores realizaron en las ciudades de Tetuán y Tánger,
antesala de un país difícil de visitar y recorrer por entonces, y con cuyas visitas querían descubrir el máximo exponentey sabor de la cultura árabe y oriental, que habían podido presenciar previamente en Andalucía.
Desde el punto de vista botánico haríamos destacar las menciones a los pinos de las sierra del Pinal, así como a los abetosde sierra Bermeja, es decir el actual pinsapar. Y climatológicamente, las especiales condiciones del viento de levante y lasnumerosísimas observaciones sobre las nieves perpetuas que coronaban nuestras sierras. La existencia de antiguas neveriasy el cálculo realizado sobre la altitud de estas nieves perpetuas, vienen a enriquecer en gran medida nuestros conocimientossobre la Naturaleza de Andalucía así como de la comarca del campo de Gibraltar durante el siglo XIX.
BIBLIOGRAFÍAANDERSEN, C. A. Viaje por España. Alianza Editorial. 1988. Epílogo y notas de Marisa Rey. 264 pag. La primer edición de su libro fue I Spanien, publicada en 1863.
BOWLES, G. Introducción a la Historia Natural y a la Geografía Física de España. Imprenta Real de Madrid. 1789. 554 pag. Tercera edición.GEORGE, G. La Biblia en España . Edit. Biblioteca Grandes Viajeros. Prólogo de Emilio Soler Pascual. Traducción de Elena García Ortiz. 2001. 541 pag.CARR, J. Descriptive travels in the southern and eastern parts of Spain and the Balearic Isles in the year 1809 . Printed for Sherwood, Neely and Jones, and J. M.Richardson. London.CARTER, F. Viaje de Gibraltar a Málaga. Edit. Arguval. 336 pags. Título original: A journey from Gibraltar to Málaga . 1777. Traducción de Christina Taylor yJosé Antonio Olmedo López.CHAPMAN, A.; J.W. Buck. La España agreste. La Caza . Ediciones Giner. 1982. Revisión de la traducción Mauricio González. Prólogo de Luís de Mora-Figueroa. 327 págs.CLARK, G. W. Gazpacho o meses de verano en España. Edit. Horizontes . 1996. Traducción de Fernando Miranda. 247 pag. Titulo original: Gazpacho, or summer months in Spain. Londres 1850.CLAVIJO PROVENCIO, R. 1997. Viajeros Apasionados. Testimonios Extranjeros sobre la Provincia de Cádiz 1830-1930. Public. Excma. Diputación de Cádiz. 167 pags.DELANNOY, J. J. 1987. Reconocimiento biofísico de Espacios Naturales de Andalucía . Public. Casa de Velázquez-Junta de Andalucía. 50 págs.DORÉ, G.; CH. Davillier. Viaje por España. Edic. Grech S.A. (2 vols). 1988.FORD, R. Manual para viajeros por Andalucía y lectores en casa . Edic.Turner 1988. Reino de Granada (183 pag); Reino de Sevilla (356 pag).GAUTIER, T. Viaje a España. Edit. Cátedra. 1998. Edición y traducción de Jesús Cantera Ortiz de Urbina. 384pags. Titulo original: Tras los montes. 1840.GÓMEZ, A.; S. Sánchez, M. Simón, F. Salvador, A. Esteban. 1992. Síntesis de la morfología glaciar y periglaciar de Sierra Nevada. Estudios de Geomorfología
en España. Actas II Reunión Nacional de Geomorfología. Murcia. 379-392.INGLIS, D. Henry. Spain in 1830. In two vols. Whittaker, Treacher and Co. London. 1831. Vol:II: 402 pags.JACOB, W. Viajes por el Sur. Cartas escritas entre 1809-1810. Introducción y traducción de Rocío Plaza Orellana. Edit. Portada. 391 pag. Título original: Travelsin the South of Spain, in letters written a.d. 1809 and 1810. Londres. 1811.MACHENZIE, A.S. A year in Spain, by a young american. John Murray. London. 1831.POITOU, E. Viaje por Andalucía. Edit. Renacimiento. 2004. Traducción de Marie-Christine del Castillo. 260 pag. Título original: Voyage en Espagne. Alfred Mameet Fils Editeurs. 1869.PONZ, A. 1786. Viaje de España. Tomo IX: Trata de Sevilla. 2ª edición. Madrid.TOWNSEND, J. Viaje por España en la época de Carlos III (1786-1787). Editorial Turner. Prólogo Ian Robertson. Traducción de Javier Portus. 1988. 454 pag. Primerapublicación en 1791.TWISS, R. Viaje por España en 1773. Edit. Cátedra. 1999. Edición y traducción Miguel Delgado Yoldi. 287 pag. Primera edición en 1775: Travels through Portugaland Spain in 1772 and 1773.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 77/276
Comunicaciones
77
Luis Barrios Jaques / Coordinador Técnico Fundación Migres
Guillermo Doval de las Heras
EL PROGRAMA MIGRES DEAVES PLANEADORAS.DATOS ACTUALIZADOS A LOS AÑOS 2005 Y 2006.
INTRODUCCIÓN
En Europa los contrastes estacionales, tanto en condiciones meteorológicas como en disponibilidad de alimento, provocanmovimientos masivos de animales que buscan en cada momento las condiciones más adecuadas para su supervivencia. Laestratégica situación geográfica de la comarca del Campo de Gibraltar convierte a la zona en el punto de migración másimportante del Mediterráneo occidental. Así, el estrecho de Gibraltar sirve de puente a millones de aves que cada año locruzan en sus viajes migratorios entre Europa y África, y de puerta entre el Atlántico y el Mediterráneo a un gran númerode especies de peces, reptiles, aves y mamíferos marinos que desarrollan su ciclo vital alternativamente entre ambos mares.
Consciente de la importancia de este fenómeno, en 1995 la Consejería de Medio Ambiente de la Junta de Andalucía encargaa la Estación Biológica de Doñana definir la estrategia de puesta en valor del recurso de la migración en el Estrecho deGibraltar. En 1996 nace la idea del Programa MIGRES, con un doble objetivo: 1) obtener información acerca de laevolución de las poblaciones de distintas especies animales y 2) difundir y poner en valor el fenómeno migratorio en la zona,utilizando como herramienta el seguimiento cuantitativo, cualitativo y a largo plazo de la migración de estas especies a supaso por el estrecho de Gibraltar. Actualmente los trabajos se centran en aves planeadoras, aves marinas y paseriformes,y el objetivo de la Consejería de Medio ambiente es ampliar progresivamente los grupos objeto de estudio, contempladola posibilidad de trabajar también con invertebrados.
Desde 1997 y hasta la fecha se viene desarrollando el Programa MIGRES de Aves Planeadoras, financiado por la Consejeríade Medio Ambiente. El Programa ha sido coordinado por SEO/BirdLife desde 1997 hasta 2005, y desde ese año en adelantela coordinación ha sido asumida por la Fundación MIGRES.
EL PROGRAMA MIGRES DE AVES PLANEADORAS
La primera fase del proyecto (1997-2001) se ha centrado en el ajuste metodológico, incluyendo la determinación del esfuerzode observación. La segunda fase (2002 hasta la actualidad) se centra en el trabajo de seguimiento propiamente dicho.
Para los trabajos de ajuste metodológico se ha partido de la premisa de que el Programa MIGRES debe plantearse con una duraciónindefinida y que los resultados año tras año han de ser comparables entre sí. Para ello los esfuerzos se centraron en sistematizarlos trabajos de conteo, estandarizando al máximo la toma de datos. La base de todo el trabajo es que, por mucho esfuerzo quese realice, los conteos tan sólo representan una muestra del número de aves que utilizan la zona durante sus viajes migratorios.Por ello, el objetivo principal de esta faceta del proyecto ha sido optimizar el esfuerzo obteniendo a su vez datos consistentes.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 78/276
Almoraima, 35, 2007
78
AJUSTE DEL PROTOCOLO DE TRABAJO
Esfuerzo
El control del esfuerzo es fundamental para poder comparar los datos que se obtengan en distintos años de conteos. Lasvariables que definen el esfuerzo de observación son: el número de puestos de observación, el número de observadores porpuesto (Kochenberger & Dunne 1985, Hussell 1985), el horario de observación (Bernis 1980, Sattler & Bart 1985) y elnúmero de días que los observatorios están operativos. Durante la primera temporada de conteo el horario de observaciónse ajustó al ritmo de paso de las aves, y los observatorios utilizados cada día fueron seleccionados en función de lascondiciones meteorológicas con objeto de maximizar los conteos, tomando como base los trabajos de Bernis (1980).
Con los datos de la primera temporada, y para conocer la influencia de las distintas variables que componen el esfuerzo
sobre el número de aves contabilizadas cada día, se seleccionaron aquellos días con más de quinientas aves vistas porpuesto, se ajustó un modelo linear generalizable con un error normal y una función de enlace identidad a la raíz cuadradadel número de aves detectada cada uno de esos días (Crawley 1993). Se utilizaron como variables predictoras el númerode observadores (entre dos y cuatro), el número de horas de observación (entre doce y ocho) y el observatorio.
Con estos datos los resultados muestran que el número de aves contabilizadas no estuvo influido de manera significativapor el número de observadores en cada puesto (F1,127=0,13 ; p=0,71), ni por el número de horas de observación(F1,128=8,09x10-5 ; p=0,99), pero el número de aves contabilizadas era significativamente diferente entre los distintosobservatorios (F14,142=21,88 ; p<0,0001).
Con esta información, durante las temporada de 1998 a 2001 se trabajó con un número fijo de observadores y un horariode observación concreto. Se utilizaron un total de 26 observatorios, aunque se trabajó preferentemente en siete: Bolonia,
Peña, Tráfico, Cazalla, Canteras, Hoya y Algarrobo.Para definir el número mínimo de observatorios necesarios para obtener datos consistentes (de todas las especies excepto lacigüeña blanca, que se trata posteriormente), se han analizado los resultados de estos siete observatorios. Se ha realizado unaregresión múltiple por pasos hacia delante con el número total de aves contabilizadas como variable respuesta y el númerode aves contabilizadas por cada uno de los observatorios utilizados como variable predictora. Para estos análisis se ha ajustadoun modelo lineal generalizable utilizando una función de enlace logarítmica. Se han realizado análisis individuales para unaselección de siete especies (por motivos que se discuten más adelante), y se ha repetido para aves volando hacia el sur y paraaves cruzando. Los resultados indican que, para todas las especies y en ambos comportamientos, con los dos mejoresobservatorios (el principal y el secundario) el ajuste del modelo se considera suficiente para que los datos obtenidos seanconsiderados buenos predictores de los resultados que se obtendrían teniendo operativos todos los observatorios comparados.
Para las aves volando hacia el sur y para las aves cruzadas respectivamente, las tablas 1 y 2 muestran el ajuste del modelo(expresado como porcentaje explicado de la varianza) con el predictor principal más el predictor secundario.
Aves al SurCiconia nigra CAZ+ALG (65%)Pernis apivorus ALG+TRA (71%)Milvus migrans TRA+ALG (66%)Neophron percnopterus CAZ+ALG (67%)Accipiter nisus ALG+CAN (73%)Circaetus gallicus CAZ+ALG (65%)Hieraaetus pennatus ALG+CAN (67%)
Tabla 1. Porcentaje explicado de la varianza por especies, del número de aves volando hacia el sur (S, SE, SW) utilizandoel observatorio principal más el secundario. CAZ: Cazalla, TRA: Trafico, CAN: Canteras, ALG: Algarrobo.
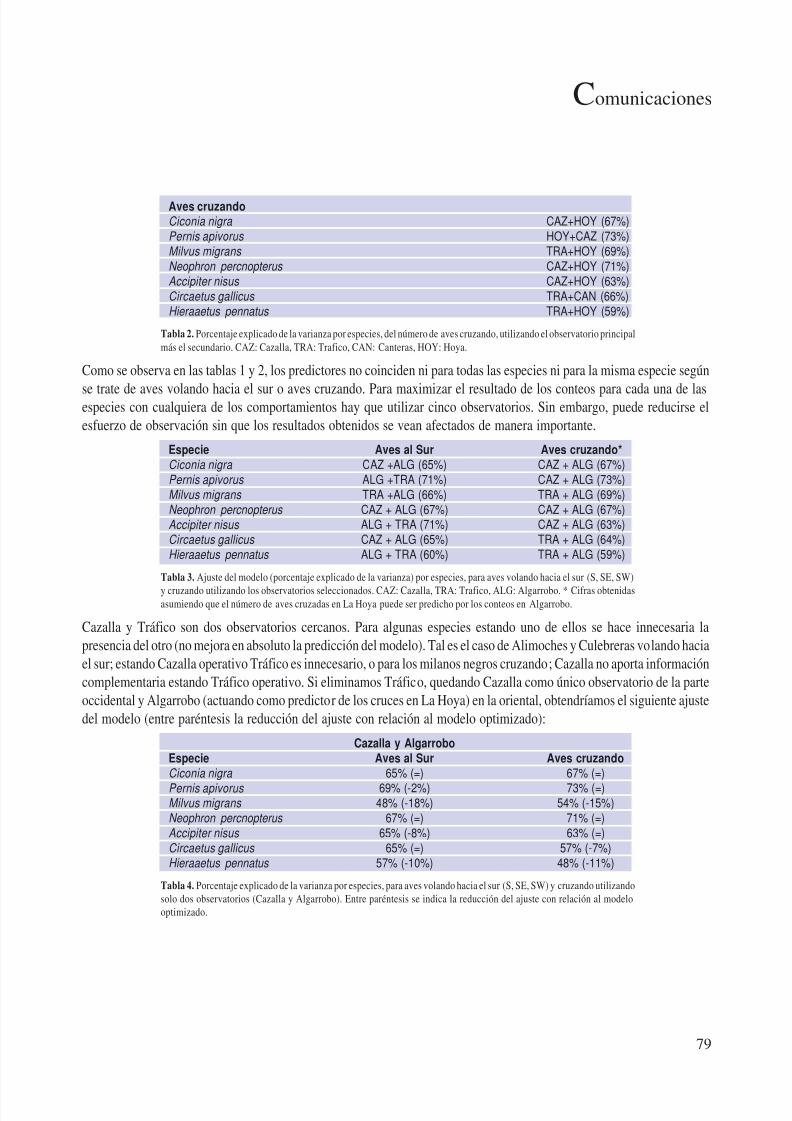
8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 79/276
Comunicaciones
79
Aves cruzandoCiconia nigra CAZ+HOY (67%)Pernis apivorus HOY+CAZ (73%)Milvus migrans TRA+HOY (69%)Neophron percnopterus CAZ+HOY (71%)Accipiter nisus CAZ+HOY (63%)Circaetus gallicus TRA+CAN (66%)Hieraaetus pennatus TRA+HOY (59%)
Tabla 2. Porcentaje explicado de la varianza por especies, del número de aves cruzando, utilizando el observatorio principalmás el secundario. CAZ: Cazalla, TRA: Trafico, CAN: Canteras, HOY: Hoya.
Como se observa en las tablas 1 y 2, los predictores no coinciden ni para todas las especies ni para la misma especie según
se trate de aves volando hacia el sur o aves cruzando. Para maximizar el resultado de los conteos para cada una de lasespecies con cualquiera de los comportamientos hay que utilizar cinco observatorios. Sin embargo, puede reducirse elesfuerzo de observación sin que los resultados obtenidos se vean afectados de manera importante.
Especie Aves al Sur Aves cruzando*Ciconia nigra CAZ +ALG (65%) CAZ + ALG (67%)Pernis apivorus ALG +TRA (71%) CAZ + ALG (73%)Milvus migrans TRA +ALG (66%) TRA + ALG (69%)Neophron percnopterus CAZ + ALG (67%) CAZ + ALG (67%)Accipiter nisus ALG + TRA (71%) CAZ + ALG (63%)Circaetus gallicus CAZ + ALG (65%) TRA + ALG (64%)Hieraaetus pennatus ALG + TRA (60%) TRA + ALG (59%)
Tabla 3. Ajuste del modelo (porcentaje explicado de la varianza) por especies, para aves volando hacia el sur (S, SE, SW)
y cruzando utilizando los observatorios seleccionados. CAZ: Cazalla, TRA: Trafico, ALG: Algarrobo. * Cifras obtenidasasumiendo que el número de aves cruzadas en La Hoya puede ser predicho por los conteos en Algarrobo.
Cazalla y Tráfico son dos observatorios cercanos. Para algunas especies estando uno de ellos se hace innecesaria lapresencia del otro (no mejora en absoluto la predicción del modelo). Tal es el caso de Alimoches y Culebreras volando haciael sur; estando Cazalla operativo Tráfico es innecesario, o para los milanos negros cruzando; Cazalla no aporta informacióncomplementaria estando Tráfico operativo. Si eliminamos Tráfico, quedando Cazalla como único observatorio de la parteoccidental y Algarrobo (actuando como predictor de los cruces en La Hoya) en la oriental, obtendríamos el siguiente ajustedel modelo (entre paréntesis la reducción del ajuste con relación al modelo optimizado):
Cazalla y AlgarroboEspecie Aves al Sur Aves cruzandoCiconia nigra 65% (=) 67% (=)Pernis apivorus 69% (-2%) 73% (=)Milvus migrans 48% (-18%) 54% (-15%)Neophron percnopterus 67% (=) 71% (=)Accipiter nisus 65% (-8%) 63% (=)Circaetus gallicus 65% (=) 57% (-7%)Hieraaetus pennatus 57% (-10%) 48% (-11%)
Tabla 4. Porcentaje explicado de la varianza por especies, para aves volando hacia el sur (S, SE, SW) y cruzando utilizandosolo dos observatorios (Cazalla y Algarrobo). Entre paréntesis se indica la reducción del ajuste con relación al modelooptimizado.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 80/276
Almoraima, 35, 2007
80
Como muestra la tabla 4, las mayores reducciones se producirían para los milanos negros y para las aguilillas calzadas, yen menor medida para las culebreras cruzando y los gavilanes volando hacia el sur. Sin embargo, en todos los casos puedenasumirse dichas reducciones sin que el ajuste del modelo deje de ser satisfactorio.
Por lo tanto, es necesario utilizar un mínimo de dos observatorios (Cazalla y Algarrobo) y el uso de un tercero (Tráfico)mejora sensiblemente los datos (sobre todo para algunas especies).
Recogida y tratamiento de los datos
Para que los resultados sean comparables, además de controlar el esfuerzo de conteo, los datos que se recojan han de serobjetivos, deben ser el reflejo de un suceso claro. Para ello, la información que se ha recogido desde 1998 se refiere a “avessobrevolando el observatorio con componente sur” y a “aves que se lanzan a cruzar el Estrecho”. Ambos datos son
objetivos, son el reflejo de un comportamiento. El número de veces que se observa cada uno de estos comportamientospuede considerarse como un índice objetivo de abundancia para ser comparado a lo largo de los años.
Por último, los observatorios han trabajado desde 1999 sin comunicación entre ellos, tomando muestras independientesdel paso de aves.
Debido al comportamiento migratorio de la cigüeña blanca (Ciconia ciconia), y a la imposibilidad de simultanear su conteocon el de las aves rapaces en los observatorios costeros, se decidió utilizar un único observatorio (Facinas) situado en elinterior y sobre el que tienen lugar la mayor parte de los aflujos de cigüeñas hacia la costa. Por ello, para los resultados deesta especie sólo se refieren a aves volando con componente sur.
Con todo ello, se ha diseñado el siguiente protocolo de trabajo.
Rapaces y cigüeña negra
- Temporada de observación: 25 de julio al 10 de octubre (78 días)
- Horario de observación: 07:00 a 16:00 horas solares
- Número de observadores por puesto: 3
- Puestos de observación: Tráfico y Cazalla (toda la temporada) y Algarrobo (25 de agosto al 10 de octubre)
- Información prioritaria: aves que se lanzan a cruzar el Estrecho en Tráfico y Cazalla y aves volando con componentesur en Algarrobo.
- Sin comunicación entre los distintos observatorios.
Cigüeña blanca
- Temporada de observación: 25 de julio al 24 de agosto (30 días)
- Horario de observación: 07:30 a 15:30 horas solares
- Número de observadores por puesto: 3
- Puestos de observación: Facinas.
- Información prioritaria: aves volando con componente sur.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 81/276
Comunicaciones
81
TRABAJO DE CAMPO
Durante el periodo de estudio 1997-2004, han participado en el proyecto 780 colaboradores voluntarios, y un equipo detres personas para la coordinación. Todos los voluntarios han recibido un cursillo de formación sobre la metodología deltrabajo, especialmente enfocado a la toma de datos, así como un cursillo de identificación de las especies migrantes. Se haninvertido 804 días de trabajo con 91.469 horas de observación, sin incluir el esfuerzo realizado por los miembros delGibraltar Ornithological and Natural History Society (GONHS) desde el observatorio del peñón de Gibraltar.
Para la toma de datos se han utilizado hasta el año 2002 fichas estandarizadas en las que mediante una serie de códigos, setomaron los datos referidos a los bandos o aves individuales con comportamiento migratorio (hora solar del avistamiento,especie, número de individuos, sexo, edad, dirección de procedencia y de vuelo, así como observatorios de procedencia y dedestino). A partir del año 2002 se ha se han utilizado tres agendas electrónicas Palm Zire que contaban con el programa debase de datos HandBase3 y el programa HBForms para el diseño de la interfaz gráfica del capturador de datos mediante ellápiz óptico. Dicha interfaz gráfica fue diseñada y programada por el equipo coordinador del programa a imagen y semejanzade las fichas utilizadas con el mismo objeto durante anteriores campañas. Las observaciones de las aves se han realizado conlos prismáticos y telescopios particulares de los voluntarios y con tres telescopios 20-60x77 y tres prismáticos 8x30. Todoslos días, después de la jornada de campo, los observadores se han reunido con los coordinadores para poner en orden los datosobtenidos y aclarar en la medida de lo posible los problemas que pudieran haber surgido a lo largo del día.
Los datos de velocidad y dirección del viento, temperatura y presión atmosférica, han sido proporcionados por el CentroMeteorológico Territorial de Andalucía Occidental del Instituto Nacional de Meteorología (tomados en la estaciónmeteorológica de Tarifa). Los datos de visibilidad y cobertura de las nubes han sido tomados por los voluntarios a intervalosde una hora, en cada uno de los observatorios.
RESULTADOS
Las tablas 5 y 6 y la figura 1 muestran los resultados de los conteos para la selección de especies entre 1998 y 2006 en elcaso de aves volando al sur. Para las aves cruzando no se presentan los resultados de la temporada de 1998 ya que lametodología empleada entonces no permite comparar los datos.
AVES VOLANDO PUESTO AÑOAL SUR 1998 1999 2000 2001 2002 2003 2004 2005 2006Ciconia nigra Alg 862 820 734 943 1.275 699 607 905 1.033
Caz 882 556 1.031 1.063 1.959 1.661 1.427 980 921Tra 758 554 798 825 1.437 1.176 1.124 742 914
Milvus migrans Alg 7.582 11.935 7.478 5.551 9.477 18.156 13.801 3.674 3.999Caz 45.964 26.901 42.341 51.385 60.345 84.265 74.184 85.696 102.202Tra 37.286 46.119 43.819 51.122 58.004 63.721 60.031 82.750 81.931
Neophron percnopterus Alg 411 672 510 705 514 777 651 584 541Caz 741 699 745 885 967 971 906 972 977Tra 254 494 324 496 596 523 389 673 348
Circus pygargus Alg 80 55 91 80 94 111 141 45 56Caz 84 38 64 86 62 99 180 88 87Tra 116 74 69 158 105 120 138 72 106
Circaetus gallicus Alg 2.718 3.120 3.128 4.615 4.441 8.441 4.385 3.934 4.337Caz 3.295 4.188 6.254 5.512 7.156 5.873 4.468 4.964 4.003Tra 1.871 2.979 4.609 3.556 4.946 4.172 3.211 3.512 3.268

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 82/276
Almoraima, 35, 2007
82
Pernis apivorus Alg 27.195 34.851 53.829 40.777 38.834 42.085 67.249 46.550 38.984Caz 7.544 3.440 13.288 3.954 11.997 14.017 13.391 17.712 6.413Tra 4.320 3.618 10.285 2.386 6.289 8.724 9.456 17.741 5.923
Hieraetus pennatus Alg 5.276 4.551 8.938 10.192 7.098 18.493 13.417 10.197 10.596Caz 1.803 2.543 3.309 4.875 6.333 3.620 3.585 3.315 3.604Tra 1.366 1.961 2.665 4.654 4.789 3.522 2.992 3.417 2.918
Accipiter nisus Alg 450 848 569 596 1.555 1.879 1.241 1.759 1.151Caz 263 249 323 523 550 673 509 577 528Tra 103 176 187 324 356 368 303 674 377
Tabla 5. Resultados de la serie: Relación de aves observadas volando con dirección sur (S, SE, SO) en la migración otoñal en el estrecho de Gibraltar, para las especiesmás numerosas en el periodo 1998-2006. CAZ: Cazalla, TRA: Tráfico, ALG: Algarrobo.
AVES CRUZANDO PUESTO AÑO1999 2000 2001 2002 2003 2004 2005 2006Ciconia nigra Caz 376 347 611 1.673 614 602 801 744
Tra 473 526 636 1.323 734 745 593 795
Milvus migrans Caz 16.336 27.841 33.598 39.429 30.317 60.988 51.911 68.623Tra 37.350 29.570 38.736 44.505 43.858 51.021 70.025 70.368
Neophron percnopterus Caz 259 330 305 607 471 432 627 453Tra 345 223 338 439 399 247 488 236
Circus pygargus Caz 11 24 42 26 42 78 27 33Tra 46 43 95 71 63 91 48 47
Circaetus gallicus Caz 2.289 2.125 1.712 4.861 2.717 1.912 2.833 2.560Tra 2.533 3.135 2.415 4.142 3.472 2.619 2.982 2.644
Pernis apivorus Caz 2.630 11.806 1.707 11.139 12.727 12.464 17.480 5.091Tra 3.517 10.161 2.011 6.148 8.569 9.240 17.632 5.737
Hieraetus pennatus Caz 1.259 1.007 1.242 3.941 1.884 1.352 1.974 2.079Tra 1.540 1.838 2.637 3.635 2.637 2.360 2.952 2.185
Accipiter nisus Caz 132 83 124 307 365 113 398 284Tra 120 125 168 269 285 190 579 246
Tabla 6. Resultados de la serie: Relación de aves observadas cruzando en la migración otoñal en el estrecho de Gibraltar, para las especies más numerosas en el periodo1999-2006. CAZ: Cazalla, TRA: Tráfico.
ESPECIES OBJETO DE ESTUDIO
Es evidente que para un conjunto de especies, el número de efectivos en migración en la zona del Estrecho es muy bajoy escasamente representativo para poder ser utilizado en la monitorización de la evolución de las poblaciones. Debido aesto se han separado en dos grupos las especies objetivo del proyecto, atendiendo al número de efectivos observados enmigración:
1) Especies con número de individuos por año superior a 300 individuos. (10 especies): cigüeña negra, cigüeña blanca,abejero europeo, milano negro, alimoche común, culebrera europea, aguilucho cenizo, gavilán común y aguilillacalzada. Son las especies que han sido utilizadas en los análisis relacionados con la selección de los observatorios.
2) Especies con número de avistamientos por año inferior a 300 individuos. (25 especies, más una subespecie). Son en lamayoría de los casos especies “no estrictamente planeadoras”, que combinan el vuelo “a vela” con el vuelo batido, junto

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 83/276
Comunicaciones
83
Figura 1. Resultados de la serie: Relación de aves observadas volando con dirección sur (S, SE, SO) en la migración otoñal en el Estrecho de Gibraltar, para las especiesmás numerosas en el periodo 1998-2006. Códigos de los puestos: CAZ: Cazalla, TRA: Tráfico, ALG: Algarrobo.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 84/276
Almoraima, 35, 2007
84
con otras especies pocos comunes. En la tabla 7 se indica el número de aves observadas en actitud migratoria para elperiodo 1998-2004.
Familia-Especie NúmeroAccipitridae
Elanio Común (Elanus caeruleus) < 5Milano Real (Milvus milvus) ± 100Quebrantahuesos (Gypaetus barbatus) 1Buitre Negro (Aegypius monachus) < 5Buitre Moteado (Gyps rueppellii) < 30Aguilucho Lagunero Occidental (Circus aeruginosus) ± 300Aguilucho Pálido (Circus cyaneus) ± 50Azor Común (Accipiter gentilis) ± 30
Busardo Ratonero (Buteo buteo) ± 200Busardo Ratonero subesp. Vulpinus (Buteo b. Vulpinus) < 5Busardo Moro (Buteo rufinus) < 25Águila Imperial Ibérica (Aquila adalberti) < 10Águila Real (Aquila chrysaetos) < 5Águila Moteada (Aquila clanga) < 5Águila Pomerana (Aquila pomarina) £ 25Águila Azor Perdicera (Hieraaetus fasciatus) < 25
Pandionidae Águila Pescadora (Pandion haliaetus) ± 100
Falconidae Cernícalo Primilla (Falco naumanni) ± 200Cernícalo Vulgar (Falco tinnunculus) ± 300
Cernícalo Patirrojo (Falco vespertinus) 1Esmerejón (Falco columbarius) < 5Alcotán Europeo (Falco subbuteo) < 100Halcón de Eleonora (Falco eleonorae) < 10Halcón Borní (Falco biarmicus) < 10Halcón Sacre (Falco cherrug) 1Halcón Peregrino (Falco peregrinus) < 100
LA UTILIDAD DE LOS RECUENTOS
Los conteos de migración visible de rapaces en observatorios de migración han dominado el estudio de la migración derapaces en las últimas décadas. (Zalles & Bildstein 1995). En la mayoría de los casos, los estudios de la evolución de los
tamaños poblacionales se realizan mediante censos durante la época de reproducción, sobre aves nidificantes. Con lasespecies que localizan sus nidos en paredes rocosas (como los buitres comunes y los alimoches) está técnica puede serparticularmente efectiva. Sin embargo cuando las zonas de reproducción son muy extensas (en ocasiones varios países) ocuando los hábitats son difíciles de prospectar como los bosques, el seguimiento y monitoreo de poblaciones de rapacesen sus zonas de reproducción puede ser logísticamente difícil y financieramente prohibitivo (Fuller y Mosher 1981,1987).De entre las diez especies con un número de efectivos alto en la migración en la zona de estudio, una se reproducemayoritariamente en bosques (cigüeña negra) y cinco son estrictamente forestales (abejero europeo , milano negro,culebrera europea, gavilán común y aguililla calzada).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 85/276
Comunicaciones
85
El protocolo de trabajo propuesto contempla la utilización de una serie de observatorios elegido por su eficacia en el conteode las seis rapaces forestales y del alimoche. Aunque los censos de alimoche se pueden realizar con cierta precisión durantela nidificación, se ha incluido en los análisis porque la preocupante situación en que se encuentran sus poblaciones aconsejaobtener la máxima información posible sobre la especie.
Para siete especies (cigüeña negra, cigüeña blanca, abejero europeo, alimoche común, culebrera europea, milano negro yaguililla calzada), el número de aves observadas en migración en la zona de estudio es representativo de sus poblacioneseuropeas y debe ser uno de los objetivos principales del programa a largo plazo.
BIBLIOGRAFÍABERNIS, F. 1980. Migración de las aves en el Estrecho de Gibraltar. Volumen I : Aves Planeadoras. Universidad Complutense de Madrid. pp. 481.CRAWLEY, M.J. 1993. Glim for ecologists. Blackwell Scientific Publications. Oxford.FULLER, M.R., MOSHER, J.A. 1981. "Methods of detecting and counting raptos: a review". pp. 235-246 in Estimating numbers of terrestrial birds, ed. by C. J. Ralphand J. M. Scott. Studies in Avian Biology 6.FULLER, M.R., MOSHER, J.A. 1987. Raptor Survey Techniques. Raptor Management Techniques Manual. National Wildlife Federation, Washington, D.C, pp. 420.HUSSELL, D. J. T. 1985. "Analysis of hawk migration counts for monitoring population levels". En: Proceedings of the Fourth Hawk Migration Conference, ed.M. Harwood, pp.243-254. Rochester, NY :Hawk Migration Association of North America.SATTLER, G. & J. BART 1985. "A technique for evaluating observer efficiency in raptor migration counts". pp. 275-280. En : M. Harwood, (ed), Proceedings of
Hawk Migration Conference IV. Hawk Migration Association of North America.ZALLES, J.I & BILDSTEIN, K.L.1995. Hawks Aloft Worldwide. Raptor Migration Watch-site Manual. Hawk Mountain Sanctuary, Kempton, Pennsylvania.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 86/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 87/276
Comunicaciones
87
ABSTRACT
Current perspectives on ecology are increasingly incorporating humans as significant components of dynamic ecosystems.However, these approaches frequently focus on measuring the impact of humans’ habitat alteration, degradation andexploitation on animal or plant populations. In many cases non-human organisms and humans co-exist and interact in amyriad of ways, ranging from competitive to commensal to mutualistic. Understanding these interactive complexes canenable us to increase our assessment and understanding of complex ecological systems. The macaque monkeys ( Macacasylvanus) living in the Upper Rock Nature Reserve, Gibraltar, have co-existed with humans for centuries and currentlyengage in frequent interactions with large numbers of people from across Europe. These interactions involve behavioral,nutritional, physiological, and potentially epidemiological factors. Humans are a core part of these macaques’ ecology andthe ecosystems in the Upper Rock Nature Reserve. In this paper we present an overview of human-monkey interactions
in Gibraltar with a focus on the role that anthropogenic elements play in the behavioral ecology of these monkey andconsider the implications of these patterns for the ecosystems across the Upper Rock Nature Reserve.
Keywords: Anthropogenic, Humans, Macaca sylvanus, Ecosystems, Stakeholders, Gibraltar.
HUMANS, MONKEYS, AND THE ROCK:THE ANTHROPOGENIC ECOLOGY OF THEBARBARY MACAQUES IN THE UPPERROCK NATURE RESERVE, GIBRALTAR
Agustín Fuentes / Nancy O’Neill / Dr. Eric Shaw / Dr. John Cortés

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 88/276
Almoraima, 35, 2007
88
INTRODUCTION
Current perspectives on ecology are increasingly incorporating humans as significant components of dynamic ecosystems.However, these approaches frequently focus on measuring the impact of humans’ habitat alteration, degradation andexploitation on animal or plant populations. In many cases non-human organisms and humans co-exist and interact in amyriad of ways, ranging from competitive to commensal to mutualistic. Understanding these interactive complexes canenable us to increase our assessment and understanding of complex ecological systems.
It is a widely recognized that a substantial portion of the human adaptive success is effectively represented by ecologicalalteration and niche construction (Fuentes 2004, Odling-Smee et al. 2003, Potts 2004). This pattern is described bygeographers and anthropologists as the creation of human "place" (sensu Richardson 1989). If there are other organismsliving in or around this "place", they are impacted by the ecological contexts manipulated, produced, and modified byhuman action. In other words, anthropogenic engagement with the environment changes selection pressures and ecologicalcontexts for all sympatric organisms. Humans may be seen as competitive dominants or foundation species in manyanthropogenic systems. However, humans are not the only stakeholders with agency in ecological systems. Othermammals, plants, and even microorganisms also impact the structure of elements within any given ecosystem. Fullerunderstanding of the relationships between humans and the other animals with which they share their environments canfacilitate a broader and more inclusive view of ecological systems.
As is evident from the rapid and dramatic reduction in numbers and range of many species, most large animals do not dowell when they are a part of human "place". However, in many instances some species, members of the monkey genera
Macaca (the macaques) for example, do quite well around humans (Fuentes et al. 2005). In anthropogenic environments,certain organisms play major roles in the structuring of the local ecology. In the Upper Rock Nature Reserve, Gibraltar,
the relationship between barbary macaques ( Macaca sylvanus) and humans ( Homo sapiens) plays a principle role in thestructure of the environment and therefore the ecological system. In this essay we will examine the role of macaque-humaninteractions in a human "place" in order to gain insight into the anthropogenic ecology of the barbary macaques in the UpperRock Nature Reserve, Gibraltar.
ECOLOGICAL CHALLENGES, NICHE CONSTRUCTION, AND FACILITATION
The basic ecological challenges faced by organisms include acquiring energy, avoiding predators and successfully mating(Denham 1971). These challenges are shaped by variables such as the structure of the local habitat, the type and numberof conspecifics and other organisms in the habitat, the distribution and densities of food sources, and the patterns of climate
change across annual cycles. These variables act as the proximate pressures eliciting the behavioral strategies practiced byorganisms in a given ecological context. In order to examine organisms that successfully exist in highly anthropogenichabitats (human "place") two additional concepts need to be added to these basic premises.
Niche construction- Among the most salient advances in evolutionary biology and ecology is the recognition of the roleof niche construction , resulting in a removing of the traditional organism/environment dichotomy and its replacement witha more integrated and dynamic view of organism/environment interactions and co-evolution. It is becoming increasinglyevident that organisms not only impact their immediate environments but also, in part, shape the selection (ecological/ environmental) pressures that they face (Laland et al. 2001, Stamps 2003). The niche is best characterized as a“multidimensional hypervolume” (Hutchinson 1957), the “way” that organisms make a living and the eco-context in whichorganisms occur. If organisms exhibit agency in the active construction of their niches there arises a need to discard the

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 89/276
Comunicaciones
89
traditional organism/environment dichotomy and replace it with a concept that more accurately reflects the mutuallyengaged set of changes and patterns in organisms and environments that emerge through the course of time. According toLaland et al. (2001) niche construction, per se, can be seen as “when an organism modifies the functional relationshipbetween itself and its environment by actively changing one of the factors in its environment either by physically perturbingthese factors at its current address or by relocating to a different address thereby exposing itself to different factors.” Of interest for this essay, humans exhibit dramatic patterns of niche construction (cultural practices across human populationscertainly attest to this) that can result in significant ecological changes and subsequent behavioral changes in an array of sympatric organisms (Fuentes et al. 2005, Odling-Smee et al. 2003, Potts 2004). Environmental modification/manipulationis an important part of the human niche and its impact extends to more than just the humans living in that environment.
Facilitation- While competition does occur and is a significant factor for organisms on this planet, facilitation, or positive
interactions between species, or groups within an ecosystem, also drives evolutionary change. Research from ecology,especially intertidal and plant ecosystems, demonstrates that the interactions between two or more species co-existing inthe same location and ecological space may alter the selective environments such that each does better when the other isalso sharing the environment/ecology (Bruno et al. 2003). This is seen at the distinction between realized and fundamentalniches (Hutchinson 1957). The fundamental niche is the ecological context in which a species can live indefinitely in theabsence of negative interspecific interactions. The realized niche is that eco-space actually occupied by a species afterexclusion/competition by other species/competitors (Bruno et al. 2003). Research has demonstrated that facilitation canexpand the realized niche of many organisms in a shared system, thereby expanding the ecological success of the organismsvia mutual use of the same environment.
While this research is currently focused on small animals and plants, the theoretical emphasis on examining multi-species,
multi-group, population level interactions for patterns of facilitation can have dramatic impact on our models of whatecological pressures were and are acting on organisms that co-exist in anthropogenic habitats. In this context, someorganisms currently viewed as engaged in commensal (co-existant) or mutualistic (directly co-beneficial) relationshipswith humans could be seen as engaged in a facilitated relationship with humans and/or each other. Facilitation is distinctfrom commensualism or mutualism in that the interactors do not directly assist one another but rather impact the ecologicalcontext such that both groups realized niches expand (and they then receive reduced selective pressures resulting from nicheconstraints). This is different from by-product mutualism in that we are not focusing on single benefits to one organismarising as a by-product of selfish habitat exploitation by another.
Rather than seeing humans and other organisms as exclusively engaging in strict competition with each other overresources, or directly interacting in mutualistic manners, some level of facilitation may be occurring such that by co-existing with humans the overall pressures are reduced compared to living in non-human occupied environments. In a sensethis reinforces the conceptualization of human as a foundation species (sensu Bruno et al. 2003, box 3). Even if there issome level of competition between the humans and the other species, their communal impact on their “living space” maybe enhancing some aspects of survivability. So, rather than seeing other organisms as living alongside humans and simplycompeting with them for resources we can potentially view the interaction as one that changes the selective pressures forone or both species such that evolutionary benefits, and therefore possibly biological changes, accrue via co-existence.Combining the concepts of facilitation and niche construction allows us to create a template in which to assess the impactsand relationships between humans and other sympatric organisms.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 90/276
Almoraima, 35, 2007
90
THE UPPER ROCK NATURE RESERVE. AN ANTHROPOGENIC HABITAT
The local habitat of interest for this essay is the Upper Rock Nature Reserve of Gibraltar. The reserve covers 2.5-3 Km of the middle and upper reaches of a Jurassic limestone uplift, called the Rock of Gibraltar or El Peñón, with the highest pointin the reserve being approximately 424 meters above sea level. The reserve contains nine floral habitat types with unequaldistribution, over 350 plant species, an extensive variety of migratory and some resident bird species, at least 11 mammalspecies, 12 species of reptile, and a diverse array of invertebrates (Cortés 1979, Pérez and Bensusan 2005). Of the variousanimal species in the reserve a few seem to be clearly benefiting from the anthropogenic environment; the barbarymacaques, the yellow-legged gull ( Larus michahellis), feral cats (Felis catus), the black rat ( Rattus rattus alexandrinus)and feral goats (Capra hircus ). Other medium to large mammals including the Spanish Ibex (Carpa pyrenaeica), the redfox (Vulpes vulpes silacea), the wild boar (Sus scrofa) and the small-spotted genet (Genetta genetta) all went extinct in the
reserve area by the middle on the 20 th century. This suggests that specific components of the anthropogenic environmentfavor certain animals on the Rock.
Currently, the reserve contains an extensive road system, two restaurants, a cable car system, a number of private residencesand various tourist attractions ranging from caves, to gun batteries to monkey viewing sites. However, this extensive humanmodification of the environment is not completely recent. The lower reaches of the Rock have experienced continuoushuman habitation since at least the early 8th century AD, with numerous indications that human residents used at least partof the upper reaches of the Rock since then. Beginning in the 15 th century fortifications, buildings and pathways wereconstructed and maintained throughout the upper Rock (Pérez and Bensusan 2005). Today, numerous road systems traversethe reserve and create open areas in between patches of high maquis, cliff faces, and the other habitat types. The numerousfire breaks impact the structure and composition of vegetation types and maintenance of electrical and water services also
create pathways and blockages amongst the habitat types in the middle and upper reaches of the Rock.The main tourist attractions and the roads act to consolidate and direct human movement in the reserve, however, foot trafficalso diverges from the road and main tourist areas. While there is little direct contact between humans and any animals asidefrom the macaques (see below), feral cats congregate and are fed at main tourist sites and feral goats take advantage of theextensive cover provided by the maquis vegetation.
With 718, 919 visitors from more than 19 countries to the upper Rock in 2003, including more than 72,000 a month duringpeaks season (June-September (Pérez and Bensusan, 2005)) a very large number of humans visit the Upper Rock NatureReserve. Humans can access the Rock with on foot, via automobile/coach or by the cable car service. In 2002 alone,1,758,460 motor vehicles (including 12,407 coaches) crossed into Gibraltar from Spain and an estimated 60,000 vehicles(taxis, coaches, and private vehicles) passed through the main tourist sites in the Upper Rock Nature Reserve (Pérez and
Bensusan, 2005). The volume of human presence on the Rock carries with it a diverse array of habitat influencing contexts.The noise and litter from the daily visits to the reserve create a cyclical pattern of rubbish abundance and ambient noisethat varies across the year in intensity (Pérez and Bensusan 2005). The traffic movement throughout the road system alsoacts as a major source of mortality for the larger mammals and the yellow-legged gulls in the reserve (Pérez and Bensusan2005). As nocturnal access is limited to those with permission or resident in the reserve, the density and impact of humanactivity is very low during the hours of darkness. However, as residents of Gibraltar can drive at higher speeds, or lessimpeded speeds, along the roads at night, the potential risk to animals using those roads may be higher. Currently, thereis little evidence of any sustained hunting or capture of animals in the reserve, except for the culling of yellow-legged gulls(Pérez and Bensusan 2005).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 91/276
Comunicaciones
91
AN ABBREVIATED HISTORY OF THE BARBARY MACAQUES
While there is some contention regarding the first appearance of the barbary macaques in Gibraltar, it is generally assumedthat macaques have been on the Rock since the Moorish occupation (711-1462 AD) (Perez and Bensusan 2005). The historyof the macaques on Gibraltar has been well reviewed by Fa (1984), O’Leary and Fa (1993), and Shaw and Cortés (in press),so we will not attempt to provide a thorough historical overview here. However, a brief summary of the provisioning andinteraction history with humans is relevant. From the 19 th century until the mid 1990s there were two main groups of macaques in the population on the Rock; one at Ape’s Den (formerly Queen’s Gate) and another at Princess Caroline’sBattery. By 1946 both groups were provisioned by the British military separately, and by 1970 provisioning for the PrincessCaroline’s Battery group was moved to Middle Hill (a restricted military reservation area) to reduce the macaquesincursions into Gibraltar town (Fa 1984). From 1972 onward the Middle Hill group had minimal interactions with non-
military humans whereas the Ape’s Den group has been regularly visited by tourists at the site since at least 1936 (Fa 1984).Increased human interactions began in 1960 with the promotion of the Gibraltar apes as a tourist attraction. As tourismincreased substantially during the 1980s-90s economic pressures came to bear on the taxi drivers and tour guides ferryingtourists to visit the macaques. This may have led to illegal provisioning by some Gibraltarans at new areas on the Rock of Gibraltar (Pérez and Bensusan, 2005). By the late 1990s this had contributed to a fissioning of extant groups which by 2004had resulted in the permanent presence of approximately 250 macaques in 6 groups on the Rock (Shaw and Cortés in, press).Today the main interaction sites are Ape’s Den, Anglian Way/St. Michael’s cave, Prince Phillips Arch and the cable carstation at the top of the Rock.
It is clear that interactions with humans have been a substantial aspect in the daily lives of the Gibraltar macaques for severalgenerations and that these interactions impact the social behaviour and ecology of the macaques (Fa 1984; Fa and Lind 1996;
O’Leary and Fa 1993; Pérez and Bensusan 2005, Shaw and Cortés,in press). Throughout this history of interactions somerecords on the impact of disease have also been kept. In the period of 1936-1944 gastroenteritis appears to have had asubstantial mortality impact on the Gibraltar macaque population and pneumonia caused high mortality amongst infants inthe late 1980s (Fa 1984; O’Leary and Fa 1993). Both of these cases may reflect pathogen transmission from humans tomacaques. Unpublished observations of intermittent outbreaks of nasal discharge and coughing amongst the macaque groupsalso suggests that the macaques may be acquiring mild respiratory pathogens from humans (A. Fuentes Pers. Observation andPers. Communication Staff members of Gibraltar Ornithological and Natural History Society (GONHS)). To date there areno published records of a viral pathogen transmission event from the Gibraltar macaques to humans, however the transmissionof campylobacter and a macaque louse to humans may be possibilities (Dr. M. Pizarro pers. Comm.).
Currently the feeding, management and care of the macaques is undertaken by the Gibraltar Ornithological and NaturalHistory Society (GONHS) under an agreement with the government of Gibraltar. GONHS provision the macaques at fivelocations in the reserve providing a selection from 15 or 16 kinds of fruits and vegetables plus grains every day of the year(Pérez and Bensusan 2005). Although no formal dietary study has been conducted across this population it appears thatthe macaques derive the majority of their annual diet from provisioning, however, they do feed daily on both vegetationand invertebrates in the reserve.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 92/276
Almoraima, 35, 2007
92
HUMAN-MACAQUE INTERACTIONS
Interactions with humans have been a substantial aspect in the daily lives of the Gibraltar macaques for several generationsand that these interactions impact the social behavior and ecology of the macaques (Fa 1984; Fa and Lind 1996; O’Leary andFa 1993; Pérez and Bensusan 2005, Shaw and Cortés in press). Long-term exposure and subsequent habituation of themacaques to human presence suggests that the macaques are fully acclimatized to being around humans (Fuentes 2006; Shawand Cortés in press) Fa (1984) suggested that human presence and tourist provisioning may have a negative impact on macaquereproduction and lead to other physiologically stressful outcomes. It may be that the current provisioning strategy of feedinggroups a total of over 43,000 kilograms of food annually (Pérez and Bensusan 2005) lessens the impact of humans and thatthe multi-generational exposure to high numbers of humans in the current population of macaques has altered the perceptionsand physiological responses of the macaques relative to earlier times (see Fa 1984 for demographic data 1936-1980).
During 2004 observers collected general data on 5489 human-macaque interactions with 808 of the interactions recordedin full detail during focal follows (Fuentes 2006). Across all of the five interactions sites in the Upper Rock Reserve thereare significantly more contact interactions than non contact interactions (X2=459, p<.001) between humans and macaques.There does not appear to be a simple relationship between the number of humans in proximity to a macaque and theoccurrence of contact interactions and/or aggressive interactions. Across the sites 382 (47%) of the 808 focal interactionsinvolved tourist/taxi driver/tour guide provisioned food. However, each site varies substantially in the relative impact of food presence during interactions, and overall presence of food does not appear to have a direct, linear relationship tointeraction rates or aggression (Anderson et al.unpublished). A comparison of the two main interaction sites in the UpperRock Reserve (Apes Den and Prince Phillips Arch) was made to examine the participation of taxi/coach drivers in theinteractions. Interactions between taxi/coach drivers and the macaques usually involve the taxi driver luring with food, and/
or physically assisting, the macaque onto the head and shoulders of a tourist for a photo opportunity. Additionally, manytaxi/coach drivers feed the macaques, often placing food items directly into the monkey’s mouth. These tourist couriersaccounted for 18.1% of all recorded interactions observed during focal follows at Apes Den and 33.8% percent of allinteractions recorded during focal follows at Prince Phillip’s Arch. As there are usually no more than a few (0-3) of theseindividuals at a given location during a sample period, they make up a maximum of ~8% of humans at an interaction location(Fuentes 2006). Taxi/coach drivers are significantly overrepresented in interactions with the macaques (Apes Den X2 =38.4, p<.001, Prince Phillip’s Arch X2 = 202.8, p<.001).
MODELING AN ANTHROPOGENIC ECOLOGY FOR THE GIBRALTAR MACAQUES
It is obvious that the macaques in the Upper Rock Nature Reserve are doing well. Their population size has grownconsistently across the last 20 years. Can this simply be explained by invoking by-product mutualism; that humanprovisioning has led to increased nutritional intake and increased reproductive rates? Or can we envision these macaquesas participants in an anthropogenic ecosystem (along with the other successful animals) and see this scenario as potentiallyan example of facilitation in a human "place"? What role does assessing the anthropogenic environment play inunderstanding the ecology of this habitat? To answer these questions we will review aspects of the macaques’ behaviour,habitat, ranging and general ecology.
It is fairly clear that, contrary to the assertion of Fa (1984), provisioning and high rates of human interaction are notsuppressing the reproductive rate of the population. However, it may be possible that human patterns and provisioned foodact as selective pressures and are impacting the behaviour and activity patterns of the macaques. Current research suggests

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 93/276
Comunicaciones
93
no linear, or simple, relationship between human density and macaque aggression (Fuentes 2006, Anderson et al.unpublished). Preliminary data also suggest that there is not a clear association between the presence of food and aggressionbetween humans-macaques or macaques-macaques (Fuentes unpublished data). However, these data reflect only peaktourist season, when food sources are extremely plentiful and thus may not accurately reflect the overall impact of provisioned food on the levels of aggression in the macaques.
Ranging patterns appear to have shifted over the last decade in response to human activity. Increased feeding at diverselocations by taxi drivers appears to correlate with the break up of multiple groups. This fractioning of two groups into as manyas six groups signals a significant shift in the demographic structure of the macaque population. Subsequent managementdecisions to provision the new groups may have acted to reinforce these entities as social units as did the earlier change infeeding regimes in 1946. In addition to the provisioning impact on ranging two other elements appear to shape the patterns
with which macaques use the Rock area. As of 2003 at least one group has begun spending substantial amount of time in thewaste dumping site on the Southeast corner of the Limestone uplift. They are tolerated at the site by the local human workersand appear to split their time between the dump area and the habitat around one of the major tourist sites (St. Michael’s Cave).They appear to partition the use of the tourist site with another group (from which they may have split off) (E. Shaw pers.comm). The presence of the trash dump combined with the tolerance of the human workers may have opened up a novel rangingand foraging site for this group. Previous to this, the dump site was not a part of any group’s range. Another impact on rangingis that the reserve is bounded by urban areas (Gibraltar Town and Caleta Bay Village). While individual macaques haveventured into these areas for centuries, consistent use by groups of fringe areas bordering the reserve such as housing estatesand hotels appears to be relatively new. In part this ranging shift by some groups or subgroups may be elicited by humanactivity. Macaques are highly opportunistic and plastic foragers. As new feeding sites appear in proximity to current ranges,macaques are likely to explore them. Active provisioning by humans and access to uncovered and overflowing rubbish binsmay be powerful stimuli eliciting an adaptive foraging response. In both of these cases the shift in macaque ranging must beunderstood both as a foraging response and in the context of human activity and habitat change.
If we can see that a significant portion of the macaques foraging strategies are being impacted by human patterns of provisioning, trash creation and management then we also must consider the nutritional impact. In general human foodsare high in readily utilizable carbohydrates and lipids (fats). If some of the macaques are supplementing their GONHSprovisioned foods with a significant percentage of human trash and handouts, then the energetic balance of some groupsor individuals will be quite different from that of others. This can impact ranging, daily activity cycles and possible overallhealth. The site with the longest history of tourist provisioning, Ape’s Den, does appear to have the highest rate of overweight animals (as assessed by overt skin folds). Understanding the nutritional ecology of these monkeys requires anassessment of the various foraging opportunities and the diversity of food chemistries in these choices.
The physical structure of the sites where macaques spend much of their time (the tourist interaction sites) also might impactmacaque behaviour. Most sites where moneys interact with tourists are along a roadway. However, the highest densityinteraction sites (Ape’s Den and Prince Phillip’s Arch) differ dramatically in structure. Ape’s Den has a large (~10m x 50m)multi-level area where macaques and humans can interact. The macaques can avoid human interaction by moving over aretaining wall onto the western cliff face (low wind, not a sheer cliff) or by moving to the north or south. Humans cannot followin any of these cases. This suggests that when macaques are interacting with humans they are doing so explicitly by choice.At Prince Phillip’s Arch the road way itself is the main interaction location. With a small (~10m x 5m) interaction area anda small (~1m x 2.5m) raised platform (the arch itself). The eastern cliff face is minimally accessible to due its sheer drop andthe frequent high winds that buffet it. To avoid human interactions the macaques must move off of the road to the western slopeor north towards the cable car station and restaurant. It is possible that the human manipulation of physical structure (the roads

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 94/276
Almoraima, 35, 2007
94
and the existing structures at the two sites) combined with the different micro-habitats on the east (Prince Phillip’s Arch)andwest (Ape’s Den) faces of the Rock create a different set of costs and benefits for interacting with humans.
One final and important component of anthropogenic environments deserves attention: the potential risk of pathogen transfer.The frequent overlap of macaques with human use areas suggests the possibility for the acquisition of human pathogens.Macaques are potentially at risk to an array of human pathogens including measles, influenzas and other respiratory diseases(Jones-Engel, et al. 2001). Physical contact and climbing of macaques onto tourists (with encouragement/training from taxi/ coach drivers) places human and macaque faces in close proximity and thus respiratory zones into close contact. Pathogensmost easily transmitted via mucosal contact or aerosol dispersal may pose a significant danger to this macaque population.
THE MACAQUES IN CONTEXTInteractions with humans and the structure and patterns within the anthropogenic environment are a core part of the barbarymacaque ecology and potentially a reason why these macaques are doing so well. However, the macaques themselves area major part of upper Rock ecosystem both because they are large mammals and due to the human use of the reserve. Onecan argue that the Upper Rock Nature Reserve can be viewed as an ecosystem with a few large animals as foundation speciesin the structure of the habitats and thus selection pressures on all of the inhabitants. In this essay we focused only on themacaques and humans as we see these two primates as main elements in structuring the ecology of the reserve. However,the other successful non-human animals in the reserve also contribute to its structure and context.
Although the barbary macaques are large bodied mammals there are no actual comparative data to address their relativecontribution to the overall mammalian biomass in the reserve. However, with approximately 250 individuals in the population
Figure 1

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 95/276
Comunicaciones
95
they contribute over 1000 Kg to the overall mammalian biomass, probably significantly more than the feral goats or cats, andpotentially any other mammals aside from the black rats and possibly rabbits (Oryctolagus cuniculus). However, above andbeyond their biomass contribution, the macaques are integrally tied to the human presence in the reserve. At least 20% of tourists visiting the reserve specifically come to see the macaques and a much larger percentage participate on tours that includea visit the macaque sites (Fuentes unpubl. data). Additionally, local taxi drivers market their tours to visitors to Gibraltar withthe specific reference to seeing and interacting with the macaques. This suggests that the macaques’ presence in the reserveis a major contributor to the human tourist presence and thus contributes to the daily cycle of traffic and rubbish dispersal. Therubbish alone provides forage for many of the other organisms in the reserve including the ubiquitous yellow-legged gulls andthe black rats. The continued presence of tourists also stimulates the Gibraltaran government not only to maintain a relationshipwith GONHS for the continued provisioning of the macaques but also for the support network of road maintenance, water
dispersal and fire control crews. This constant use and managing of the reserve, stimulated in part by the economic benefitsthe macaques provide, acts to maintain relatively static habitat structure and to provide direct human assistance to some species(such as feral cats) and protection to others (feral goats). Even the yellow-legged gulls, who are actively culled in high numbersannually, receive a trade-off in the form of protection for a myriad of predators found in other areas of their breeding rangebut extinct or non-resident in Gibraltar (figure 1).
Is this more than by-product mutualism? The cats, gulls, rats, goats all benefit from structure and context of roads, macaquetourism, and the trash situation. They benefit from the ongoing relationship between the macaques and the humans and itspattern of impact on the Upper Rock Nature Reserve. This may be seen as, in part, some degree of facilitation resulting froma human impacted habitat and active role that the macaques have within it. Other organisms, such as some of the plant species,
Figure 2

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 96/276
Almoraima, 35, 2007
96
migratory birds, the endemic barbary partridge ( Alectoris barbara ), and some resident reptiles may not benefit, and might infact suffer from the extensive human use and management. In this case the anthropogenic environment that is in partconstructed via interaction with the macaques has created sets of ecological challenges (selection pressure) that are beneficialfor some inhabitants and not for others (figure 2). In a sense, we can see a potential expansion of realized niche for a selectgroup of resident species while at the same time a reduction in the realized niche of others. The active and dynamic relationshipsbetween the resident organisms and their environment is structured, shaped and changed in an anthropogenic context.
In the broadest sense we can tackle an assessment of the Upper Rock Nature Reserve from the perspective of the macaquesby envisioning a dynamic ecosystem with two large mammals as major stakeholders; humans and macaques. Therelationships between these two species acts to structure the habitat, provide forage options and opportunities for an arrayof organisms and maintain a high volume human use of the area. Although other organisms also act as significant
components in this ecosystem, the macaques and humans represent two of the major factors (foundation species)influencing the structure of the niches available (figure 3).
WHAT IS THE BENEFIT OF THE PERSPECTIVE PUT FORWARD IN THIS ESSAY?
If niche construction entails an organism modifying “the functional relationship between itself and its environment byactively changing one of the factors in its environment either by physically perturbing these factors at its current addressor by relocating to a different address thereby exposing itself to different factors” (Laland et al. 2001) then one can arguethat we are seeing niche construction by humans as well as the macaques on the Rock. If facilitation entails interactionsbetween two or more species co-existing in the same location and ecological space altering the selective environments such
Figure 3

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 97/276
Comunicaciones
97
that each does better when the other is also sharing the environment/ecology (Bruno et al. 2003), the possibility remainsthat in the Upper Rock Nature Reserve we may be seeing some facilitation via niche expansion amongst various animals.Looking at this ecosystem with both an eye towards management and with an evolutionary perspective, our analyses mayproduce a more comprehensive set of results.
Many researchers would agree that seeing anthropogenic influence as a core component of an environment allows for amore realistic management approach to studying ecosystems. If we take this a step further and include the roles of othersignificant non-human contributors to the shaping of the anthropogenic environment, such as macaques in this case, wecan expand our assessments and move towards a complexity based, multi-stakeholder approach. Thinking about realizedniches and their partitioning within an environment in the context of the active management and construction of localecologies allows for the use of tools from a diverse array of fields. Toolkits from disciplines such as ecology, animal
behavior, anthropology, and resource management can be brought to bear on the same questions in the hopes of producingmore effective and engaged results. Results that then may be of relevance not just to ecologists or resource managers butalso to evolutionary theorists, animal behaviorists, and other researchers trying to understand relationships betweenorganisms in an increasingly anthropogenic world.
BIBLIOGRAPHYANDERSON, M, Fuentes, A., Easterday, N., Eleazer, M., Hogan, K., and Kwiatt, A. (unpublished) Patterns in aggressive interactions between humans and macaquesin Gibraltar.BRUNO, J.F., Stachowicz, J.J., and Bertness, M.D. 2003. "Inclusion of facilitation into ecological theory". Trends in Ecology and Evolution 18(3): pp. 119-125.CORTÉS, J. and Shaw, E. in press. The Barbary Apes in Gibraltar In K. Hodges and J. Cortes Eds. The Barbary Macaque. University of Nottingham Press; NottinghamEngel, G.A., Jones-Engel, L., Suaryana, K.G., Arta Putra, I.G.A., Schilliaci, M.A., Fuentes, A., and Henkel, R. 2002. Human exposures to Herpes B seropositivemacaques in Bali, Indonesia. Emerging Infectious Diseases 8 (8): pp. 789-795.FA, J.E. 1992. Visitor-directed aggression among the Gibraltar macaques. Zoo Biology 11: pp. 43-52.FA, J.E. 1984. "Structure and dynamics of the Barbary macaque population in Gibrlatar". In The Barbary Macaque: a case study in conservation. pp.263-306 J. FaEd. Plenum Press, New York.FA, J.E. and Lind, R. 1996. "Population management and viability of the Gibraltar Barbary Macaques". In Evolution and Ecology of Macaque Societies. pp.235-262J.E. Fa and D.J. Lindburg Eds. Cambridge Press: Cambridge.FUENTES, A. 2006. "Human culture and monkey behavior: Assessing the contexts of potential pathogen transmission between macaques and humans". American
Journal of Primatology 68: pp. 880-896.FUENTES, A. and Gamerl, S 2005. "Disproportionate participation by age/sex class in aggressive interactions between long-tailed macaques (Macaca fascicularis)and human tourists at Padangtegal Monkey Forest, Bali, Indonesia". American Journal of Primatology 66: pp. 197-204.FUENTES, A., Southern, M., and Suaryana, K. 2005. "Monkey forests and human landscapes: is extensive sympatry sustainable for Homo sapiens and Macacafascicularis on Bali". In Commensalism and Conflict: The primate-human interface. Patterson J Ed. American Society of Primatology Publications; Norman, OKHUTCHINSON, G.E. 1957. "Concluding remarks". Cold Spring Harbor Symposium on Quantitative Biology 22: pp. 415-4.JONES-ENGEL, L., Engel, G.A., Schillaci, M.A., Babo, R. and Froelich, J. 2001. "Detection of antibodies to selected human pathogens among wild and pet macaques(Macaca tonkeana) in Sulawesi". American Journal of Primatology 54: pp. 171-178.
LALAND, K.N., Odling-Smee, F.J., and Feldman, M.W. 2001. "Niche construction, ecological inheritance, and cycles of contingency in evolution". In S. Oyama,P.E. Griffiths, and R.D. Gray Eds. Cycles of Contingency: Developmental systems and evolution. MIT Press, Cambridge, Massachusetts. pp. 117-126.ODLING-SMEE, F. John, with Kevin N. Laland, and Marcus W. Feldman 2003. Niche Construction: The Neglected Process in Evolution . Princeton University Press;Princeton.O’LEARY, H. and Fa, J.E. 1993. "Eff ects of Tourists on Barbary Macaques at Gibraltar". Folia Primatologia, 61: pp. 77-91.POTTS, R. 2004. "Sociality and the concept of culture in human origins". In R.W. Sussman and A. Chapman eds. The Origins and Nature of Sociality . Walter deGruyter, New York. pp. 249-269.PÉREZ, C.E. and Bensusan, K.J. (2005) Upper Rock Nature Reserve: A management action plan. A Gibraltar Ornithological & Natural History Society Publication.Gibraltar.RICHARDSON, M. 1989. Place and Culture: two disciplines, two concepts, two images of christ, and a single goal in J.A. Agnew and J.S. Duncan eds. The Power of Place: Bringing Together Geographical and Sociological Imaginations. Unwin Hyman, Winchester. pp. 140-1.STAMPS, J. 2003. Behavioural practices affecting development: Tinbergen’s fourth question comes of age. Animal Behaviour 66: pp. 1-13.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 98/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 99/276
Comunicaciones
99
RESUMEN
La dispersión de semillas hacia micrositios favorables y el fenómeno de la depredación postdispersiva parecen jugar un papelmuy importante como unos de los principales factores que pueden limitar la regeneración de las quercíneas estudiadas(Quercus suber y Quercus canariensis) en los bosques de las sierras orientales de las provincias de Cádiz y Málaga (ParqueNatural los Alcornocales). Estas bellotas constituyen una parte importante de la dieta de muchos animales, fundamentalmenteaves y mamíferos. Lo más novedoso del presente trabajo es que se ha descrito por primera vez la interacción de un coleóptero(Thorectes lusitanicus), que cuando llega el otoño cambia su dieta (fundamentalmente coprófaga) para alimentarse de bellotasque encuentra en el suelo. No se trata de un hecho puramente anecdótico pues se ha encontrado que esta especie de coleópteroes capaz de manipular más de un 40% de las bellotas expuestas experimentalmente en determinados micrositios, actuandocomo un auténtico depredador postdispersivo en nuestro área de estudio. Ahora bien, el impacto que ejerce es bastante
diferente en las dos especies de Quercus estudiadas. Así, la temprana disponibilidad de bellotas en el suelo para el caso delquejigo, que coincide con el pico máximo de actividad del Thorectes, y la aparente menor dureza de la testa que presentansus bellotas hacen que el efecto sobre esta especie sea mucho más negativo que para el caso del alcornoque. En este últimocaso, la acción de estos coleópteros puede llegar incluso a favorecer su proceso de regeneración al dispersar (enterrándolasen el suelo) una importante proporción de bellotas que no llegan a consumir por completo.
Palabras clave: alcornoque, quejigo, Parque Natural Los Alcornocales, depredación, dispersión secundaria.
EFECTO DE UN ESCARABAJO PELOTERO(THORECTES LUSITANICUS)SOBRE LA REGENERACIÓN DEDOS ESPECIES DE QUERCUS ENUN BOSQUE MEDITERRÁNEO
Ignacio M. Pérez-RamosInstituto de Recursos Naturales y Agrobiologia de Sevilla (IRNASE), CSIC

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 100/276
Almoraima, 35, 2007
100
INTRODUCCIÓN
La regeneración natural de cualquier especie leñosa depende en gran parte del éxito en la supervivencia durante las primerasfases de semilla y plántula. Muchas de estas leñosas, como es el caso de las dos especies de Quercus estudiadas en el presentetrabajo (Q. suber y Q. canariensis), producen semillas de gran tamaño y elevado valor nutritivo, constituyendo una parteimportante de la dieta de algunos animales, que pueden llegar a consumir gran parte de la producción anual y comprometerel proceso de regeneración natural. Hasta el momento se ha documentado mucho sobre el papel de aves y fundamentalmentemamíferos como principales responsables de estas enormes pérdidas, pero muy poco se ha especulado sobre el papel delos insectos como posibles depredadores postdispersivos de bellotas.
En este artículo se ha documentado un novedoso y sorprendente caso de interacción entre un escarabajo pelotero (Thorecteslusitanicus) y las semillas de dos especies de quercíneas en los bosques de las sierras del Aljibe y el Campo de Gibraltar.Durante el otoño, Thorectes cambia las heces de herbívoros por las bellotas de Quercus, lo cual supone el paso de una dietafundamentalmente coprófaga hacia otra balanófaga. Este drástico cambio, jamás descrito con anterioridad, debe implicarla posesión por parte del coleóptero de unas fuertes mandíbulas para poder romper la dura testa de la bellota y,probablemente, unas enzimas digestivas muy particulares capaces de procesar este tipo de alimento, tan diferente delhabitual en la comunidad de escarabajos peloteros. Normalmente, cuando Thorectes encuentra una bellota, se entierra conella unos pocos centímetros bajo tierra y permanece ahí durante semanas, protegida en muchas ocasiones dentro del duropericarpo de la semilla. Así, al contrario de lo que ha sido generalmente aceptado en otros estudios, un escarabajo peloteropuede comportarse (al menos estacionalmente) como un consumidor de semillas. Además, durante el lento proceso deroedura del cotiledón, algunas de estas bellotas enterradas pueden ser abandonadas cuando aún están parcialmenteconsumidas o incluso intactas. En estos casos, si el embrión no ha sido dañado, puede ocurrir la emergencia satisfactoria
de plántulas, ejerciendo entonces el escarabajo un importante papel como dispersor secundario de bellotas.En el presente artículo se presentan algunos resultados preliminares de varios experimentos de campo y en cautividad,donde se documentan por primera vez los efectos de un escarabajo pelotero (Thorectes lusitanicus) sobre la regeneraciónde dos especies de quercíneas (Q. suber and Q. canariensis), y se relata cómo un comportamiento fundamentalmentedepredador puede también favorecer la dispersión de semillas.
MATERIAL Y MÉTODOS
Área de estudio
El área de estudio está localizada en los bosques de quercíneas de las sierras del Aljibe y del Campo de Gibraltar, en el sur
de España. La geología dominante está formada por areniscas oligo-miocénicas, con un relieve de suaves lomas con alturasque no superan los 900 m (exceptuando el pico del Aljibe, que cuenta con 1094 m de altitud). El clima es de tipomediterráneo, caracterizado por presentar inviernos suaves y lluviosos alternados con veranos cálidos y secos. Lastemperaturas son suaves en todo el área, con una media anual de 17ºC. Las precipitaciones medias anuales oscilan entre763 y 1180 mm, debido a la influencia del relieve montañoso.
La vegetación está dominada por un extenso bosque de alcornoques perennes (Quercus suber ) que se mezcla con quejigossemidecíduos (Q. canariensis) en los valles y zonas más húmedas. El matorral del sotobosque es muy diverso y rico entaxones endémicos (ver descripción general en Ojeda et al. 2000). La mayor parte del área forestal ha sido protegida comoParque Natural (P. N. Los Alcornocales), que cuenta con una superficie aproximada de 1700 km2.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 101/276
Comunicaciones
101
Los experimentos de campo se llevaron a cabo en dos sitios de bosque, separados unos 40 Km aprox. de distancia: Panera(P) a 36º 31' 54'’N, 5º 34' 29'’W y San Carlos del Tiradero (T) localizado a 36º 9' 46'’ N, 5º 35'’ 39'’W. Estos sitios fueronseleccionados por estar localizados en dos situaciones geográficas muy alejadas (norte y sur del parque, respectivamente)y presentar unas cargas ganadera y cinegética altamente contrastadas. Así, Panera contaba con un mayor número deungulados, tanto silvestres (24.31 ciervos/100 Ha) como domésticos; mientras que Tiradero presentaba un menor númerode ciervos (18.66/100 Ha) y estaba excluido del ganado. En cada uno de estos bosques se delimitó una parcela de una Ha,cercada con malla cinegética para evitar la interferencia de los grandes herbívoros, donde se está estudiando la regeneraciónde ambas especies de Quercus desde 1999-2000 (Pérez-Ramos et al. 2004, Marañón et al. 2004).
Diseño experimental
Experimentos de campo
Las bellotas fueron recolectadas durante el periodo de disponibilidad natural de cada una de las especies, en lasinmediaciones de la zona experimental y procedentes de varias plantas-madre, con el fin de recoger un alto grado devariabilidad genética. Se utilizó una muestra de bellotas completamente sanas y de tamaño similar, desechándose (por elmétodo de flotabilidad) aquellas que aparecían resecas o con síntomas de infestación por larvas de lepidópteros ocoleópteros. Para su conservación, las bellotas se almacenaron durante varias semanas en cámara fría (a una temperaturaconstante de 2-4 ºC), estratificadas en bandejas de plástico usando vermiculita como material inerte, y provistas de humedadsuficiente para evitar su desecación.
Las bellotas, que previamente fueron pesadas y marcadas, se dispusieron sobre el suelo del bosque distribuidas en 48unidades de dispersión, separadas en tres tipos de micrositios: 1) abiertos, 2) bajo árbol, sin matorral y 3) bajo árbol y denso
matorral. Cada unidad de dispersión constaba de 4 bellotas dispuestas en el suelo, en las esquinas de un cuadrado virtualde 0.5 m, haciendo un total de 192 semillas dispersas para cada especie. Todas las bellotas expuestas fueron revisadasperiódicamente, anotándose si aparecían señales de actividad animal. Cuando la bellota no aparecía, se excavó en un radiocercano para verificar la presencia de Thorectes lusitanicus, anotándose el nivel de consumo de la bellota y los dañosocasionados a la misma. Los experimentos de dispersión y posterior monitorización fueron repetidos durante dos ciclos:2003-2004 y 2004-2005. Los censos se llevaron a cabo de manera más asidua durante el primer ciclo: a las 24 horas, a lasemana, a los 15 días y posteriormente de manera mensual (durante tres o cuatro meses); mientras que en el segundo ciclotan sólo se realizaron tres censos: a la semana, al mes y a los tres meses.
Debido al desfase natural existente en la caída de semillas al suelo (más temprana para el caso del quejigo), las bellotas de cadaespecie fueron colocadas en situaciones temporales diferentes, coincidiendo con los picos de máxima producción de cada unade ellas . En primer lugar, las 192 bellotas de Q. canariensis fueron dispersadas a mediados de octubre (2003) o principiosde noviembre (2004), mientras que las bellotas de Q. suber se colocaron a principios de diciembre (2003 y 2004).
Para comprobar si los escarabajos tenían capacidad de discriminar entre bellotas de las dos especies estudiadas, se llevó a caboun segundo estudio experimental a finales de noviembre del 2004 (período en el que solapan, de manera natural, los dos tiposde bellotas en el suelo), en el que se expusieron conjuntamente semillas de ambas especies. Se siguió un diseño similar alanterior, con unidades de dispersión constituidas también por cuatro bellotas, pero esta vez mezclando las dos especies (dosbellotas de cada una). En total, se distribuyeron 24 unidades entre los tres tipos de micrositios comentados con anterioridad.En este experimento, se utilizaron bellotas de peso similar para las dos especies, con el fin de eliminar el efecto del tamaño.
En un tercer experimento, se dispersaron bellotas al otro lado del cercado (sin exclusión de herbívoros) para investigar laposible interferencia de los grandes mamíferos, tales como el ciervo o el ganado. Se utilizó un diseño similar con 24

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 102/276
Almoraima, 35, 2007
102
unidades de dispersión para cada especie (de cuatro bellotascada una), distribuidas entre los tres mismos tipos demicrositios, pero en el exterior de las parcelas experimenta-les. Al igual que en el primer experimento, las bellotas de Q.canariensis fueron dispersas a principios de otoño, mientrasque las de Q. suber se colocaron a finales de otoño-principiosde invierno, tanto en el 2003 como en el 2004. Otro experi-mento mixto, también con 24 unidades, pero mezclandobellotas de ambas especies, fue llevado a cabo durante no-viembre del 2004. La periodicidad de los censos realizados en
el segundo y tercer experimento fue la misma que la descritapara el primer caso.
Estos tres estudios experimentales se replicaron de maneraidéntica en dos sitios de bosque diferentes: Panera y Tiradero;lo cual supone un total de 3072 bellotas de las dos especies,que fueron monitorizadas en dos años diferentes, para deter-minar evidencias de manipulación y enterramiento porThorectes lusitanicus.
Experimentos en cautividad
Para verificar que Thorectes era un verdadero depredador de
bellotas y no un simple carroñero que aprovechaba bellotasparcialmente consumidas por otros animales, como los roe-dores, se llevaron a cabo una serie de experimentos delaboratorio con escarabajos en cautividad.
Los escarabajos fueron capturados manualmente en los mis-mos sitios de bosque donde fueron llevados a cabo los experi-mentos de dispersión. Posteriormente, fueron transportados allaboratorio y colocados, en grupos de cuatro indivíduos, encajas de plástico de 15 x 15 x 25 cm, provistas de una fina capade suelo procedente de la misma zona. De manera regular se les
aportó una cierta cantidad de agua, mediante un difusor, con elfin de mantener unas condiciones de humedad lo más parecidasposibles a las de su hábitat natural. A cada grupo de escarabajosse les suministró un total de cuatro bellotas como únicoelemento de alimentación, que fueron localizadas de maneradispersa en cada una de las esquinas de la caja.
En un primer experimento, para evaluar la capacidad depredadora de los escarabajos, se utilizaron tres tipos de bellotas:1) bellotas intactas, 2) bellotas germinadas (ligeramente abiertas a través de la hendidura provocada por la salida de laradícula); y 3) bellotas totalmente abiertas, con los cotiledones totalmente expuestos. Un total de 15 unidades (de cuatrobellotas cada una) fueron distribuidas entre los tres tipos de tratamientos para cada especie de Quercus, sumando un total
Figura 1. Dinámica temporal de manipulación de bellotas por Thorecteslusitanicus para el rodal de bosque de la parcela de Panera, en los dos añosmuestreados. Porcentajes referidos al total de bellotas expuestas (tanto dentrocomo fuera de cercado), englobando los tres tipos de micrositios.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 103/276
Comunicaciones
103
de 120 escarabajos y 60 bellotas de cada especie. Periódica-mente (cada dos o tres días), durante los 30 días aproximadosque duró el experimento, se cuantificó el número de bellotasque presentaron señales de depredación en cada una de lascajas así como el porcentaje aproximado de biomasa consu-mida de cada una de las bellotas. La tasa de consumo debellotas en los experimentos de cautividad fue medida comoel número de días necesarios para consumir al menos la mitadde la masa de la semilla (T50).
Para ver si los escarabajos mostraban algún tipo de predilec-
ción por una u otra especie de Quercus, se llevó a cabo unsegundo experimento con el mismo diseño que en el casoanterior, pero esta vez mezclando bellotas de las dos especiesen una misma caja. En este caso se utilizaron únicamentebellotas abiertas para testificar posibles preferencias por lacomposición de los cotiledones, no por efecto de la dureza dela testa. En total, se partió de 10 grupos de cuatro escarabajos(diferentes de los utilizados en el experimento anterior, paraevitar posibles efectos de aprendizaje), haciendo un total de40 escarabajos y 20 bellotas de cada especie. A cada uno delos grupos se les suministraron cuatro bellotas: dos de alcor-noque y dos de quejigo, que fueron localizadas de maneraalterna en las esquinas de cada caja.
RESULTADOS
Experimentos de campo
Se han encontrado unas tasas de manipulación de bellotas porThorectes lusitanicus bastante importantes, llegandose a alcan-zar unos valores superiores al 40% en determinados micrositios.Los porcentajes más elevados se recogieron a principios deotoño, viéndose entonces más afectadas las semillas de Q.canariensis, que son dispersadas de manera más temprana(figura 1). El patrón natural y las diferencias entre ambasespecies fueron consistentes para los dos años de estudio. Sinembargo, en el experimento de preferencias alimenticias, no seencontraron diferencias significativas entre las dos especies(p=0.861, mediante test de Kruskal-Wallis).
Por otro lado, usando también el test no paramétrico de Kruskal-Wallis, se ha encontrado un claro efecto del tipo demicrositio, siendo los situados “bajo árbol (sin matorral)” los que presentaron unas tasas mayores de manipulación de
Figura 2. Efectos del tipo de micrositio (A) y la exclusión de grandesherbívoros (B) sobre la tasa de manipulación de bellotas por Thorecteslusitanicus. Las barras negras corresponden a semillas de Q. canariensis y lasblancas de Q. suber .
A

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 104/276
Almoraima, 35, 2007
104
bellotas por Thorectes, en comparación con los sitios abier-tos (p<0.005) así como con los localizados bajo matorral(p<0.005) (figura 2a). Estos resultados se mantienen tantodentro como fuera del cercado (figura 2b). Se ha encontradoademás un efecto del cercado marginalmente significativo(p=0.051), recogiéndose unas tasas de manipulación másaltas en el exterior de las parcelas experimentales, donde losanimales pastorean y defecan libremente (figura 2b).
Es importante tener presente que no todas las bellotasenterradas y manipuladas por Thorectes fueron completa-
mente consumidas, apareciendo un determinado número deellas que aun conservaban intacta su capacidad germinativa(figura 3). Este enterramiento positivo es más importantepara Q.suber (55%) que para Q. canariensis (20%). En estetipo de casos, Thorectes lusitanicus se comporta como unauténtico dispersor secundario de bellotas para las dosespecies de Quercus, aunque al parecer con una capacidaddepredadora potencialmente más eficaz para el quejigo quepara el alcornoque.
Experimentos en cautividad
El resultado principal es que, en condiciones de cautividad,Thorectes lusitanicus era capaz de roer la dura testa de labellota y alimentarse de sus cotiledones, pudiendo llegar aconsumirla por completo en unas pocas semanas. De estaforma, se confirmó el potencial de esta especie de coleópterocomo depredador de bellotas.
Tal y como se esperaba, la tasa de consumo fue superior enlas bellotas abiertas (germinadas o cortadas) en compara-ción con las intactas (p=0.0029, mediante el test de análisisde varianzas, ANOVA) (figura 4). El efecto protector de la
testa fue mayor para las bellotas de Q. suber , medido comoel número de días necesarios para consumir al menos el 50%de la bellota (T50); sin embargo, no se produjo apenas efectopara las bellotas de Q. canariensis, que fueron consumidasmás rápidamente, incluso cuando presentaban el pericarpointacto (figura 4).
Por último, en el experimento de determinación de preferencias alimenticias no se han encontrado diferencias significativasen el porcentaje estimado de biomasa consumida, si comparamos las bellotas de ambas especies (p=0.083, mediante el testde Mann-Whitney U ). Parece ser que, una vez expuestas las bellotas (desprovistas de la testa), los escarabajos no muestranpredilección por una u otra especie, consumiéndolas a una tasa similar.
Figura 3. Proporción de semillas depredadas y dispersadas del total de bellotasmanipuladas por Thorectes lusitanicus.
Figura 4. Resultados de la tasa de consumo de bellotas por Thorectes lusitanicusen condiciones de cautividad. Los valores de T50 representan el tiempo (en nº dedías) necesarios para consumir al menos el 50% de la biomasa de la bellota. Lasbarras negras corresponden a semillas de Q. canariensis y las blancas de Q. suber .

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 105/276
Comunicaciones
105
DISCUSIÓN
Thorectes lusitanicus como depredador de semillas
Nuestro estudio documenta por primera vez cómo un escarabajo pelotero, Thorectes lusitanicus, que habita en un ambienteforestal con abundancia estacional de recursos (bellotas), es capaz de cambiar su dieta, básicamente coprófaga, paraalimentarse de este tipo de semillas tan nutritivas y abundantes durante el otoño. Este descubrimiento contradice la afirmacióncomúnmente aceptada de que “los escarabajos peloteros no son consumidores de semillas” (Vander Wall & Longland 2004,p.157), y confirma la excepción de Thorectes lusitanicus como depredador postdispersivo de semillas de Quercus.
Thorectes lusitanicus Jeckel (Coleoptera, Scarabaeoidea, Geotrupidae) es un escarabajo pelotero áptero, endémico del surde la península Ibérica (España y Portugal) (López-Colón 1981, 1995; Martín-Piera & López-Colón 2000). La poca
información disponible sobre los hábitos tróficos de esta especie sugiere que tienen capacidad para alimentarse de una granvariedad de sustancias (Palestrini y Zunino, 1985). Algunos resultados preliminares sobre el estudio del sistema digestivode Thorectes lusitanicus sugieren la existencia de unas fuertes mandíbulas y una flora microbiana muy diversa, que puedeayudar a comprender el extenso nicho trófico que presenta esta especie, que es incluso capaz de consumir semillas con unduro pericarpo, como es el caso de las bellotas de las dos especies de Quercus estudiadas.
Desde el punto de vista de los Quercus, la proporción de pérdidas de semillas debido a la acción de Thorectes no esdemasiado alta en comparación con la de los roedores (80-90%). En los bosques mediterráneos, los roedores sonprecisamente los principales depredadores de bellotas (e.g., Pérez-Ramos 04; Gómez 03; Pulido 99; Herrera 95; Díaz 92).Sin embargo, el patrón de depredación postdispersiva de semillas por Thorectes es muy variable en el espacio y en el tiempo.Por un lado, los valores más altos se han encontrado en el exterior de las parcelas experimentales, sin exclusión de grandes
herbívoros. Esto puede deberse a que los excrementos de estos ungulados suponen una de sus principales fuentes dealimentación y el medio principal, descrito hasta el momento, que utilizan para nidificar y efectuar sus puestas (Hanski& Cambefort 1991). En general, las poblaciones de escarabajos peloteros estarán reguladas por la densidad de herbívorosde la zona (e.g., Nummelin & Hanski 1989; Halffter et al. 1992; Davis et al. 2001). Así, los bosques que cuenten con mayornúmero de ungulados silvestres o domésticos probablemente contarán con poblaciones más abundantes de escarabajospeloteros (Kadiri et al., 1997) y, por tanto, una mayor proporción de bellotas serán enterradas y consumidas (total oparcialmente) por ellos. Probablemente por esta razón se han encontrado unas tasas de manipulación de bellotas muchomás elevadas en la parcela experimental de Panera (20-40%), que cuenta con una carga de ungulados domésticos y salvajesmucho mayor que la parcela de Tiradero.
La heterogeneidad espacial del bosque es otro factor de variabilidad en la depredación de semillas por escarabajos. AunqueThorectes lusitanicus parece moverse por todo el mosaico de micrositios que ofrece el bosque, cuando las bellotas sondispersadas equitativamente en los tres tipos de micrositios, la tasa de manipulación por escarabajos es mayor en losmicrohábitats localizados “bajo árbol (sin matorral)”. Aunque este patrón debe ser confirmado mediante un estudio delcomportamiento de forrajeo del Thorectes, es probable que este tipo de micrositios puedan llegar a atraer mayor númerode estos coleópteros al presentar una mayor disponibilidad de bellotas en el suelo.
En condiciones naturales, se han encontrado unas tasas de manipulación por escarabajos más altas sobre las bellotas dequejigo que sobre las de alcornoque. Sin embargo, los resultados en el experimento en el que se ofrecieron bellotas de lasdos especies al mismo tiempo, mostraron que los escarabajos no presentaban ningún tipo de preferencia alimenticia entrelas dos especies de Quercus. Por tanto, parece ser que estas diferencias encontradas podrían estar más relacionadas con lafenología diferencial de caída de bellotas al suelo que presentan ambas especies. Así, Q. canariensis comienza a

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 106/276
Almoraima, 35, 2007
106
desprenderse de sus frutos desde principios de otoño, momento en el cual los escarabajos parecen estar más activos. Encambio, las bellotas de Q. suber caen al suelo mucho más tarde, casi entrado ya el invierno, cuando las temperaturas sonmás bajas y los escarabajos ven reducida su actividad, por lo que se registran unas tasas de manipulación mucho menores.
Ahora bien, el daño que Thorectes lusitanicus ocasiona a la bellota parece ser bastante diferente si comparamos ambasespecies de Quercus, lo cual conlleva profundas implicaciones demográficas. Una vez enterrada en el suelo, la probabilidadde que una bellota sea consumida totalmente o casi por completo por estos escarabajos es mucho más alta para Q.canariensis que para Q. suber , encontrándose en este último caso una mayor proporción de bellotas manipuladas pero conel embrión intacto (manteniendo su capacidad germinativa). Estas diferencias entre especies en la tasa de consumo debellotas también ha sido demostrada en los experimentos en cautividad, lo cual parece ser explicado por la mayor durezade la testa que presentan las bellotas de Quercus suber.
Thorectes lusitanicus como dispersor de semillas
El comportamiento innato que presentan muchos escarabajos peloteros al enterrarse junto con porciones de excrementos bajoel suelo parece que está relacionado con la competencia por los recursos, protección frente a condiciones adversas y reduccióndel riesgo de ser devorado por sus depredadores (Hanski & Cambefort 1991). Otros autores han documentado, en bosquestropicales, el papel de algunos escarabajos peloteros como dispersores secundarios de semillas (previamente ingeridas por losherbívoros), que son enterradas junto con las heces que las contienen y, de esta forma, quedan más protegidas frente a losdepredadores (e.g. Andresen 2003; Vander Wall & Longland 2004). En el caso concreto de Thorectes lusitanicus, elcomportamiento enterrador se mantiene e incluso, ocasionalmente, pueden llegar a rodar a poca distancia algunas porcionesde excrementos, ayudándose de sus patas delanteras. Este mismo comportamiento es el que parece seguir con la bellota. Elefecto positivo para la regeneración de las quercíneas estudiadas es que siempre existe una pequeña proporción de esas bellotas
previamente enterradas que no llega a ser consumida por completo, pudiendo llegar a emerger y establecerse como plántula.Un caso similar es el estudiado para algunas aves y roedores, que juegan un doble papel como depredadores y dispersores desemillas, y es aceptado que esa pequeña proporción de semillas viables que entierran en el suelo pero después olvidan es crucialpara la dinámica y mantenimiento de las poblaciones de árboles (e.g. Gómez 03; Díaz 92).
AGRADECIMIENTOS
El presente estudio ha estado financiado por el proyecto HETEROMED (REN2002-04041-C02-02) y una beca FPUconcedida por el Ministerio de Educación y Ciencia. Esta investigación se enmarca en las redes temáticas REDBONE(Andalucía) y GLOBIMED (España).
BIBLIOGRAFÍAANDRESEN, E. (2003). Effect of forest fragmentation on dung beetle communities and functional consequences for plant regeneration. Ecography, 26. pp 87-97DAVIS, A. J et al. (2001). Dung beetles as indicators of change in the forests of northern Borneo. J. Appl. Ecol. 38. pp. 593-616.DÍAZ, M. (1992). Rodent seed predation in cereal crop areas of central Spain: effects of physiognomy, food availability, and predation risk . Ecography 15, pp. 77-85.GÓMEZ, J. M. (2003). Spatial patterns in long-distance dispersal of Quercus ilex acorns by jays in an heterogeneous landscape. Ecography, 26: 573-584.HALFFTER, G., M. E. Favila, and V. Halffter, (1992). A comparative study of the structure of the scarab guild in Mexican tropical rain forest and derived ecosystems .Folia Entomol. Mex. 84, pp. 131-156.HANSKI, I. and Y. Cambefort (1991). Dung beetle ecology. Princeton : Princeton University Press, cop..— XII, 481 p: il. ; 25 cm.HERRERA, J. (1995). Acorn predation and seedling production in a low-density population of cork oak (Quercus suber L.). Forest Ecology and Management 76,pp. 197-201.KADIRI, N., J.M. Lobo, & J.P. Lumaret (1997). Conséquences de l’interaction entre préferences pour l’habitat et quantité de ressources trophiques sur lescommunautés d’insectes coprophages (Coleoptera, Scarabaeidae). Acta Oecologica, 17, 107-119.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 107/276
Comunicaciones
107
LÓPEZ-COLÓN, J.I. 1981. Contribución al conocimiento del género Thorectes Mulsant, 1842 (Coleoptera Scarabaeoidea). Thorectes baraudi, nueva especieespañola. Nouv. Rev. Ent., 11 (3): 277-286.LÓPEZ-COLÓN, J.I. 1995. Ell género Thorectes Mulsant, 1842 (Coleoptera Scarabaeoidea, Geotrupidae) en la fauna europea. G. it. Ent., 7:355-388.MARANÓN T, R. Zamora , R. Villar , M. A. Zavala , J.L. Quero , I. Pérez-Ramos, I. Mendoza , J. Castro (2004). Regeneration of tree species and restoration under contrasted Mediterranean habitats: field and glasshouse experiments. International Journal of Ecology and Environmental Sciences 30: 187-196.MARTÍN-PIERA, F. & J.I. López-Colón, 2000. Coleoptera, Scarabaeoidea I . En: Fauna Ibérica, vol. 14. Ramos, M.A. et al. (Eds.). Museo Nacional de CienciasNaturales. CSIC. Madrid. 526 pp.NUMMELIN, M. & I. Hanski, (1989). Dung beetles of the Kibale forest, Uganda; comparison between virgin and managed forest. J. Trop. Ecol. 5, pp. 349-352.PALESTRINI C. & M. Zunino,1985. Osservazioni sul regime alimentare dell’adulto di alcune specie del genere Thorectes Muls.. Boll. Mus. Reg. Sci. Nat. Torino:3:183-190.PÉREZ-RAMOS, I. M., M. T. Domínguez y T. Marañón (2004). Depredación de semillas de alcornoque (Q. suber) y quejigo (Q. canariensis): una aproximaciónexperimental. Revista de la Sociedad Gaditana de Historia Natural, 4. pp. 175-180.PÉREZ-RAMOS, I. M. y T. Marañón (2004). Limitaciones en la Regeneración Natural de las especies leñosas de un bosque mediterráneo. Almoraima 31 (2004):129-135
PULIDO, F. J. (1999). Herbivorismo y regeneración de la encia (Quercus ilex) en bosques y dehesas. Ph.D. Dissertation. Universidad de Extremadura, Cáceres, Spain.Proyecto de seguimiento y caza mayor en Andalucía (2003). Consejería de Medio Ambiente (Junta de Andalucía)VANDER WALL, S. B. & W. S. Longland. 2004. Diplochory: are two seed dispersers better than one? Trends in Ecology and Evolution 19: 155-161.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 108/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 109/276
Comunicaciones
109
RESUMEN
Durante la última década el monte público Hoyo de don Pedro en el término municipal de Los Barrios ha sufrido variosincendios forestales de importancia afectando a buena parte de su superficie, produciéndose una reincidencia en un períodocorto de tiempo. Esto, junto con la explotación intensiva a la que ha sido sometido a lo largo de su historia, hacen que elHoyo de don Pedro haya sido un monte duramente castigado, perdiendo una parte importante del bosque alcornocal queprimitivamente poseía. No obstante, la situación del monte en el Campo de Gibraltar dentro del Parque Natural LosAlcornocales albergando entre otras formaciones los canutos representativos de la vegetación del terciario, le dan a estemonte un especial valor. El motivo de la comunicación es analizar los siniestros ocurridos en los últimos años con especial
énfasis en el de 1997 y evaluar los daños producidos y las medidas de recuperación de la vegetación propuestas.
Es de resaltar como novedoso el método de estudio de las zonas afectadas y de evaluación de daños basados en tecnologíasde teledetección y su incidencia en las propuestas de recuperación de las formaciones vegetales.
Palabras clave: Monte, Forestal, Incendio, Teledetección.
Frase: Evaluación por teledetección y recuperación de áreas incendiadas.
EVALUACION Y RECUPERACIÓNDE ZONAS INCENDIADAS EN ELMONTE HOYO DE DON PEDRO(TÉRMINO MUNICIPAL LOS BARRIOS, CÁDIZ)
Miguel Cueto Álvarez de Sotomayor / Ingeniero de Montes
Rafael Sánchez Vela / Ingeniero de Montes
Pablo Fernández-Salguero López / Ingeniero de Montes

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 110/276
Almoraima, 35, 2007
110
1. INTRODUCCIÓN Y ANTECEDENTES
Los montes andaluces guardan una parte importante de nuestra propia historia, y el Hoyo de Don Pedro, monte públicoperteneciente al municipio de Los Barrios, no podía ser menos. Este monte ha sido testigo vivo de cientos de años debatallas, guerras, incendios, talas,… Su situación estratégica en el Campo de Gibraltar lo convirtió en el punto de partidade muchos de los asedios que sufrió Algeciras en el siglo XIII y XIV hasta la destrucción total de la ciudad. Asedios quefueron por mar y tierra y que tras más de 700 años han dejado cicatrices en este monte que han perdurado hasta nuestrosdías, como son muchos de los topónimos que hoy conocemos, o el mismo nombre mencionado: Hoyo de Don Pedro.
Si se da una vuelta por el monte y se observa su orografía, nos damos cuenta de que realmente no es un hoyo, ya que estáformado por dos montañas que le dan máxima altura a esta zona, separadas por el arroyo del Prior. Pero, si se escudriñaen el castellano antiguo, esta palabra cobra sentido ya que en época de la Reconquista, hoyo significaba fortaleza con locual el Hoyo de Don Pedro se transforma en la Fortaleza de Don Pedro. Pero… y Don Pedro, ¿quién pudo ser este personajehistórico que le diera nombre al monte con tal fuerza que perdurara hasta nuestros días?
Con el nombre de batalla del Estrecho se conoce al conjunto de operaciones militares y diplomáticas que se llevaron a caboen torno a las plazas de Tarifa, Algeciras y Gibraltar. Los reinos de Aragón, Génova y en menor medida Portugal, estuvieronimplicados en esta prolongada batalla debido a sus intereses económicos en la zona. Un acontecimiento de gran importanciaen esta batalla fue el sitio que Fernando IV puso a Algeciras en 1309. Una empresa que terminó en fracaso y tuvo comocontinuación otras actuaciones militares, también infructuosas. Resultado de este fracaso militar ante Algeciras, la plazade Tarifa se encontró en gran peligro, de ahí que Fernando IV tomara medidas para fortalecer la villa tarifeña ante lasamenazas de los musulmanes que seguían intentando hacerse con ella.
Hay que destacar el interés que por Tarifa, Algeciras y Gibraltar tuvieron los reyes Alfonso X, Sancho IV, Fernando IVy Alfonso XI. El primero trató en varias ocasiones de apoderarse por la vía diplomática de Tarifa, Algeciras y Gibraltarno consiguiéndolo. Llegados a este punto y como represalia al daño que había causado a los cristianos el emir benimerínAbu Yusuf, Alfonso X decidió en 1277 preparar una acción militar contra Algeciras para que “Abén Yuçaf no pudiese pasarpor allí otra vez aquende la mar”. El rey castellano recibió el apoyo de todos sus reinos y mandó aparejar una poderosa flotade “ochenta galeas e veynte et quatro naues, syn las galeotas et lennos e syn los otros nauíos pequennos” , bien pertrechadosde hombres y armas. En el mes de marzo de 1278 ya estaba todo preparado y las tropas reunidas en Sevilla. La flota sedesplazó al Estrecho para bloquear Algeciras. Mientras, el infante Don Pedro y el infante Don Alonso Fernández el Niño,comenzaban el sitio terrestre en el mes de abril, hostigando sin cuartel a la plaza sitiada.
Es más que probable que sea este acontecimiento y sus personajes los que le dieron nombre al monte Hoyo de Don Pedro,(La fortaleza del Infante Don Pedro) desde donde se preparó el sitio terrestre, pero además, si desde el monte se dirige lamirada hacia el noroeste, salvando el arroyo del Tiradero, se divisa la sierra del Niño, topónimo que puede deber su nombreal infante Don Alonso Fernández el Niño.
Todo parecía indicar, en el asedio a Algeciras en 1278, que los Infantes Don Pedro y Don Alonso Fernández el Niñolograrían su propósito, pero un grave incidente ocurrió. Por un desvío de los fondos recaudados por el tesorero real, no sele pudo enviar a la flota los avituallamientos por esto “los onmes de la flota adolecieron de muy grandes dolencias”. Losbarcos fueron abandonados y Abu Yusuf, advertido del estado desastroso de la armada castellana, arremetió contra ella ydestrozó a la flota cristiana, de forma que Algeciras pudo ser abastecida desde Tánger. Viendo inviable seguir el cerco, elinfante Don Pedro se retiró y “dexaron allí los engennos e las armas et otras cosas muchas que non pudieron leuar. Et losmoros sallieron e leuáronlo todo et metiéronlo a la çibdat” .

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 111/276
Comunicaciones
111
De este cerco por mar y tierra es probable que nos haya llegado una parte importante de la toponimia del monte, punto departida para la preparación del sitio a Algeciras llevado a cabo por Alfonso XI. Topónimos como son el arroyo del Priordebido a encontrarse fray Alonso Martín, prior de San Juan, carrera del Moro, el cercano cerro del Fraile o los Castillejoshan perdurado hasta nuestros días; otros como el Regajo de la Cruz del Padre Ricardo, se han ido perdiendo en los últimosaños debido a su falta de uso.
La historia más reciente de este monte no es menos tormentosa, son conocidas por las gentes de la villa, las historias de lascortas indiscriminadas de alcornoques y quejigos que se realizaron a principio del siglo XX. Una de las más significativas yanecdóticas es la narrada por el recientemente desaparecido (marzo de 2005) y jubilado guarda forestal de la Villa de LosBarrios, Cristóbal Pérez Mañé, hombre recto y serio con su trabajo, en el que contaba como dio con sus huesos en la cárcelpor dos días, en el año 1934, a cuenta de las cortas a matarrasa que se estaban llevando a cabo en el Hoyo de Don Pedro.
Pero esta historia anecdótica ha quedado contrastada en la Memoria de la 4ª Revisión y 5º Plan Especial del Proyecto deOrdenación del Grupo de Montes de Los Barrios, en la que aparece reflejado: […] los destrozos causados en tiempos dela república (Tercer Plan Especial 1932-1941), que hicieron bajar los aprovechamientos suberosos de la Sección 4ª de untotal de 2.536 a 128 Qm, dando esto idea de cómo quedó esa Sección y Cuartel.
Figura 1. Plano de situación del monte Hoyo de Don Pedro. Fuente: Elaboración propia.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 112/276
Almoraima, 35, 2007
112
La gran proximidad de éste al pueblo de Los Barrios y Algeciras fue la causa de que en él se cometieran tan ingentes dañoscon preferencia a los demás del Grupo de Los Barrios […]. Posteriormente continúa diciendo: […] la destrucción casicompleta del Hoyo de Don Pedro cuya superficie de descorche bajó de 51.274 m2 que figuraban en el tercer inventario a3.162 m2 que cuenta en el último efectuado con una baja de 48.000 m2 en números redondos.
Es a partir del año 1952 cuando se asume el lamentable estado en el que se encontraba el monte y se planifican, con cargoa la Ley de Auxilio, numerosos trabajos forestales de restauración para el período 1953-1956. Las principales actuacionesacometidas consistieron en repoblaciones de alcornoque tanto de siembra como de plantación que no tuvieron el éxitodeseado, repoblaciones de pino resinero y eucalipto que sí prosperaron, rozas y selección de brotes, rozas y regeneración,y rayas cortafuegos. Las repoblaciones de pino consiguieron una rápida cobertura del terreno que frenó procesos erosivosincipientes, pero no todo fueron ventajas. El monte reúne casi todas las condiciones favorables para la propagación de
incendios: altas pendientes, acceso abrupto y muy dificultoso, exposición a levante sin obstáculos que minimicen la accióndel viento y especies de gran inflamabilidad.
Gracias a los proyectos de ordenación de montes y sus revisiones periódicas cada diez años, hoy se dispone de informaciónmuy valiosa en cuanto al estado en el que se encontraban los montes y las actuaciones que se llevaban a cabo. Así, en el año1976 se aprueba la 6ª Revisión del Proyecto de Ordenación del Grupo de Montes de Los Barrios. En este documento se destacaque se preste atención especial a la regeneración de los tramos de reproducción, y que durante la ejecución del Plan Especial(01-10-1971 a 01-01-1981) se atienda con el máximo cuidado a la defensa de los valores paisajísticos y bioecológicos.
Resulta muy llamativo, si se analizan los datos de producción de corcho del grupo de montes de Los Barrios, que se hicieranestas recomendaciones justamente en el período de mayor producción de corcho de estos montes en el siglo XX. Esto hacepensar que ya en el año 1976 existía una clara conciencia de cual era e iba a ser uno de los mayores problemas del alcornocal
barreño: la falta de regeneración.
2. SITUACIÓN ACTUAL Y DESCRIPCIÓN DEL MONTE
El monte Hoyo de Don Pedro pertenece al grupo de montes de propiedad municipal de Los Barrios, inscrito en el Registrode la Propiedad de San Roque en el año 1932 por el alcalde de Los Barrios en aquel momento, José Castillo Gómez. Endicha inscripción se dice que el ayuntamiento de Los Barrios viene poseyendo estos montes desde tiempo inmemorial,como donación que hicieron los Reyes Católicos sobre los años 1495 a 1502.
La cabida o superficie que abarca es de 761 hectáreas y está declarado monte de Utilidad Pública con el nº 43 del Catálogode Cádiz. Se encuentra deslindado desde el año 1906 y amojonado en el año 1954, estando actualmente en proceso de
revisión su deslinde o definición de límites. Su gestión la realiza la Consejería de Medio Ambiente, habiéndose renovadoen 2002, por cinco años, el convenio de cooperación entre la Consejería y el Ayuntamiento de Los Barrios para la gestiónde sus montes de utilidad pública.
Sus límites son las fincas La Morisca al norte, La Gredera, Benharás, Cancho de Benharás, Herriza del Mesto y El Galeónal este, Botafuegos del Ministerio de Defensa, Las Corzas del ayuntamiento de Algeciras y Dehesa de Ojén al sur y LaZorrilla al oeste.
El monte se encuentra incluido en el Parque Natural Los Alcornocales, y según la zonificación del P.O.R.N. dentro de lasáreas de protección B, “áreas de indudables valores ecológicos, científicos, culturales y paisajísticos, que presentan ciertogrado de transformación antrópica, pues suelen tener algún tipo de aprovechamiento productivo primario que es compatiblecon la preservación de los valores que se pretende proteger”.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 113/276
Comunicaciones
113
Figura 2. Plano de topónimos del monte Hoyo de Don Pedro.Fuente: Elaboración propia.
En cuanto al estado natural, el monte se incluye en el sistemaPenibético, en las estribaciones de la sierra del Aljibe y lassierras del Campo de Gibraltar, y más en concreto en la sierrade La Palma. El monte se encuadra en la cuenca mediterráneaandaluza, y dentro de la cuenca del río Palmones, al que viertesus aguas por medio de la garganta de Benharás, garganta delPrior y garganta del Capitán o Botafuegos.
La vegetación existente se caracteriza por una cubierta arbóreade alcornocal (Quercus suber ), que se extiendemayoritariamente, aunque su continuidad se ve interrumpida
por otros tipos de formaciones, como quejigares de Quercuscanariensis en las zonas más umbrófilas y húmedas, loscanutos; acebuchares (Olea europaea var. sylvestris) y pasti-zales donde afloran las arcillas dando lugar a los bujeos;masas artificiales procedentes de repoblación, como pinares(Pinus pinea y Pinus pinaster ) y eucaliptares, donde el alcor-nocal se había perdido por los reiterados incendios; herrizas,matorral denso donde predomina la quejigueta (Quercuslusitanica) y brezos (Erica spp.) en las cotas altas debido alazote del viento y al escaso desarrollo del suelo; etc.
Destacar brevemente la importancia que, en las últimas décadas,está alcanzando la Seca, proceso de decaimiento y muerte queafecta al alcornocal, y que ha provocado que existan ampliaszonas desarboladas, hoy cubiertas de matorral de degradación, omasas con un estado sanitario preocupante, con alto grado dedefoliación, y afectadas por plagas y enfermedades.
De cara al posterior análisis de la vegetación afectada y las consecuentes mejoras selvícolas a realizar, se han diferenciadolas siguientes estructuras vegetales: alcornocal, alcornocal-quejigal, pinar de piñonero, pinar de negral, matorral deericáceas, matorral de oleolentisco, herriza, acebuchal, pastizal y eucaliptar.
En el Hoyo de Don Pedro, exceptuando la zona sur del monte, el alcornocal presente está constituido por bornizos, que
no tienen un aspecto demasiado envejecido pero que en gran parte, presentan los daños producidos por los reiteradosincendios que se han producido. Asimismo, existen repoblaciones de pino negral de los años 1958 (localizado entre lagarganta del Prior y la de Benharás) y en 1970 el restante. No obstante, por los sucesivos incendios ha desaparecido granparte de los pies originarios de las repoblaciones artificiales y se ha instalado abundante regeneración natural. Una masaimportante de eucaliptar (Eucaliptus globulus y E. camaldulensis) se localiza en la linde oriental del monte, cruzada porel arroyo del Prior, que fue implantado en 1960. Actualmente, al no contar con aprovechamiento alguno, está siendosustituido por una repoblación de alcornoque y quejigo. Entre las especies de flora destacadas en el monte aparecen elavellanillo (Frangula alnus subsp. baetica), el laurel ( Laurus nobilis) y el acebo ( Ilex aquifolium), escaso en los canutos.
El estado en que se encuentra el monte no permite que se realicen otros aprovechamientos forestales que los que contribuyena su regeneración, y sean compatibles con la serie evolutiva de la vegetación. Así, tras el incendio de 1997 se ha suspendido

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 114/276
Almoraima, 35, 2007
114
toda actividad ganadera en el período de diez años que abarca la ordenación del monte aprobada en el año 2002, y tampocoestá prevista la extracción de corcho. Las cortas de madera se reducen a la eliminación del eucaliptar con destino a berlingasy leñas estimadas en 40.000 kg. Los brezos suponen para el decenio unos 6.000 kg de taramas y el aprovechamiento apícolarepartido por el monte asciende a 40 colmenas.
En relación al aprovechamiento cinegético, el monte constituye el coto de caza CA-10.443, siendo el titular el ayuntamientode Los Barrios, y aunque está cedido a la sociedad de caza municipal La Gineta, no se lleva a cabo actividad cinegética enel mismo.
3. SERIE HISTÓRICA DE INCENDIOS FORESTALES EN EL MONTE
Los incendios forestales son acontecimientos frecuentes en el ámbito mediterráneo, especialmente durante el períodoestival, en el cual, las altas temperaturas y el estrés hídrico que sufre la vegetación, favorecen el inicio del fuego y suposterior propagación. Estos fenómenos suceden de forma periódica ya sea por origen natural o por la acción humana, quedesde la antigüedad ha empleado el fuego como herramienta agrícola, ganadera y forestal.
La vegetación mediterránea ha ido evolucionando con la ocurrencia de incendios forestales reiterados, lo que ha supuestouna adaptación al fuego bien mediante mecanismos de defensa (cortezas gruesas, yemas durmientes, rebrote de raíz o cepa,etc.) o bien de forma activa (liberación de semillas encerradas en frutos o desactivación de inhibidores de germinación).
En este sentido, se debe tener en cuenta la frecuencia con la que se producen los incendios en una zona o área determinada(índice de recurrencia), pues es fundamental para evaluar su incidencia. Así, si entre dos fuegos consecutivos, la vegetaciónno ha tenido tiempo para recuperar esas estructuras o cortezas defensivas, generar nuevas yemas durmientes o producir unnuevo banco de semillas, el efecto será aún mayor. Esta situación, en general, disminuye la diversidad florística, reducela presencia de especies arbóreas, y favorece, sin duda, a aquellas formaciones arbustivas pirofíticas, que se handesarrollado con fuegos muy frecuentes.
La necesidad de medidas preventivas, así como la restauración de las áreas afectadas, debe acometerse como situaciónprevia a una dinámica de incendios cíclicos, por la presencia de especies que necesitan quemarse para asegurar susupervivencia y evitar la competencia de otras menos adaptadas al fuego.
En el caso particular del monte Hoyo de Don Pedro, desde los años sesenta, hasta la actualidad, ha sufrido varios incendiosde cierta importancia que han afectado a su superficie, reseñándose a continuación los más importantes. El primero del quedispone información veraz, ocurrido en el año 1967, interesó únicamente a la zona sur del monte y resultó afectada una
superficie de 160,24 hectáreas. El incendio sucedido en el año 1973 afectó a 138,09 Ha de la zona central del monte,superficie que fue nuevamente quemada siete años después, en un fuego en 1980, de mayor envergadura y que recorrió287,34 Ha, siempre referidas al monte objeto de trabajo. En 1989, de nuevo en el extremo sur del monte incendiado en elaño 1967, sufrió un fuego aunque de menor importancia, recorriendo 65,34 Ha.
Por último, el más importante y también el más reciente, ocurrió en el año 1997, afectando a una extensión total de 941,82Ha, de las cuales casi la mitad, 415,95 pertenecen al monte Hoyo Don Pedro. La importancia de este último radica tantoen la superficie ardida como en la recurrencia del fuego en el monte. En la figura 2 puede observarse como resultaronafectadas zonas que ya previamente se habían quemado, como es el área central y sur. En la tabla 1 se resumen los incendiosreseñados anteriormente, con sus correspondientes datos.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 115/276
Comunicaciones
115
Año Incendio Superficie afectada del monte (Ha) Zona afectada Área quemada previamente1967 160,24 Sur -1973 138,09 Central -1980 287,34 Central SI (-7 años)1989 65,34 Sur SI (-22 años)1997 415,95 Central-Sur SI (-17 y -8 años)
Fuente: Elaboración propia.
Tabla 1. Reseña histórica de los incendios ocurridos en el monte “Hoyo Don Pedro”.
Así, únicamente en un período de 30 años, se han producido en el monte cinco incendios de cierta importancia, ydeterminadas superficies han ardido hasta tres veces, siendo numerosas las que se han visto afectadas en dos ocasiones,
como puede observarse en la tabla 2.Recurrencia de incendios Superficie afectada del monte (Ha) Porcentaje de superficie afectadaTerrenos no afectados 152,32 21,07Incendiados 1 vez 235,66 32,60Incendiados 2 veces 199,78 27,64Incendiados 3 veces 135,06 18,69Superficie total 722,82 100,00
Fuente: Elaboración propia.
Tabla 2. Superficie del monte desglosada según la recurrencia de incendios.
4. EVALUACIÓN DE LAS ZONAS INCENDIADAS MEDIANTE TELEDETECCIÓNEl incendio ocurrido entre los días 6 y 10 de agosto de 1997 (con una duración hasta su total extinción de 131 horas) supusoel de mayor importancia de los anteriormente mencionados que han recorrido el monte. Los medios disponibles en este año(medición con GPS con las aeronaves de extinción) permitían tener una información fiable de la cuantía superficial yafectación del incendio. Pero se consideró oportuno disponer de una información más profunda de lo acontecido dada sumagnitud. Así, mediante un acuerdo entre la Consejería de Medio Ambiente y la Universidad de Córdoba (E.T.S.I. deMontes) se desarrolló un trabajo que se aplicó en este monte consistente en evaluar las superficies y niveles de afectaciónocasionados por el incendio; el desarrollo de modelos de evaluación de la regeneración para las cubiertas vegetales despuésde un incendio; y elaborar propuestas de restauración.
4.1. Metodología
En el análisis se ha empleado un método novedoso en este campo, consistente en la utilización de técnicas de teledeteccióncon empleo de las imágenes de satélite Landsat-TM. Con esta información se puede realizar una valoración de los dañoscausados a la vegetación y utilizarse como punto de partida para la regeneración posterior del incendio.
En una primera fase, se realiza la delimitación inicial del incendio mediante GPS instalado en un helicóptero tras laextinción determinándose las hectáreas afectadas, ya que como la frecuencia de imágenes del Landsat es de 16 días y puedehaber interferencia de nubes, no se dispone de forma inmediata de la imagen de satélite posterior al incendio. Se realizana continuación trabajos de campo en la zona incendiada, en parcelas de muestreo representativas de los niveles de daño (sindaños, moderado, elevado y extremo) y vegetación afectada (cartografía de vegetación previa al incendio: mapas de usosy coberturas vegetales de Andalucía). Asimismo, se tipifica la respuesta de la vegetación al fuego lo que permite junto conlos datos previos un primer pronóstico de regeneración potencial.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 116/276
Almoraima, 35, 2007
116
En una segunda fase, se hace la evaluación de los daños una vez recibidas las imágenes anterior y posterior al incendio. Paraello se realiza la delimitación definitiva del perímetro del incendio (y superficie recorrida por el fuego) con la imagen satélitegeorreferenciada que ajusta el perímetro del GPS sobre la composición en color dicha imagen. Esto permite disponer de unainformación de detalle para poder superponer a perímetros de anteriores incendios y evaluar la frecuencia a escala local.
La cartografía de vegetación previa al incendio se realiza mediante técnicas clásicas de fotointerpretación, delimitando yclasificando teselas homogéneas de cubierta vegetal a partir de fotografías de vuelo fotogramétrico infrarrojo color de mayode 1997 a escala 1:15.000 de la Consejería de Medio Ambiente.
Una vez delimitada, se elabora la cartografía de niveles de afectación a partir de los niveles de daño definidos por imágenessatélite anterior y posterior al incendio (clasificación supervisada de las bandas originales que asigna a cada píxel un nivelde daño suministrado por las parcelas de muestreo de campo). La imagen posterior al incendio corresponde al 20 de octubre,la más próxima al muestreo previo. A las imágenes una vez clasificadas, se le aplica un proceso de filtrado para eliminarzonas que por su pequeño tamaño no tienen especial significación.
Por último, se obtiene la cartografía sintética de propuesta de actuaciones forestales (cruce de cartografía de daños y devegetación previa) para diagnosticar regeneración potencial según tipo de vegetación. Este paso constituye el objetivo finaldel sistema de evaluación pues supone cartografiar las recomendaciones de cara a la toma de decisiones para la futurarestauración de la zona incendiada.
4.2. Aplicación al incendio del Hoyo de Don Pedro de 1997
Se resumen a continuación los resultados obtenidos en el proceso de evaluación del grado de afectación y alternativas derestauración en el monte Hoyo de Don Pedro aplicando la metodología antes descrita y realizada en marzo de 1998. Los
trabajos de campo efectuados a finales de septiembre de 1997 se realizaron para estudiar la superficie total, y consistieronen levantamiento de 30 parcelas de control en áreas homogéneas asignándoles vegetación presente, estado de regeneracióny grado de afectación.
Superficies (Ha) Estimación inicial (GPS) Evaluación Final (satélite)Arbolada 694,99 709,9Matorral 49,34 28,22Otros usos 194,47 203,7Total 938,88 941,82
Fuente: Evaluación de los grados de afectación de los grandes incendios de 1997.
Tabla 3. Estimación inicial y evaluación final de superficie recorrida por el fuego de 1997.
De esta superficie total, el fuego recorrió 415,95 Ha del monte Hoyo de Don Pedro, siendo las restantes de los montesprivados La Gredera, Benharás, Matavacas, El Galeón, La Torre, Herriza de Mestos y Acebuche.
Formaciones afectadas Superficie (Ha)Masa de Pinus pinaster 270,37Alcornocal adehesado–matorral 103,99Eucaliptar 41,59Total 415,95
Fuente: Evaluación de los grados de afectación de los grandes incendios de 1997.
Tabla 4. Vegetación afectada por el incendio en el monte Hoyo de Don Pedro.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 117/276
Comunicaciones
117
Figura 3. Superficies incendiadas en los sucesivos fuegos del monte (1967-1997).Fuente: Elaboración propia.
Las propuestas de restauración pretenden fijar actuaciones basándose en un análisis previo sobre los tipos de vegetación,su nivel de afectación, y su respuesta y capacidad de adaptación ante el fuego. Las medidas propuestas y distribuidas enel espacio, permitirán minimizar riesgos inmediatos (erosión del suelo) y orientar en las actuaciones de restauración.
Formación Vegetación Superficie Niveles de daño (% de Has totales)1 2 3 4 5
Arbolada P.pinaster joven 169,48 80 1 8 10 1
P.pinaster adulto 70,46 34 4 36 16 10Mixta Qs-Pp 92,65 61 - 19 20 -Q. suber adulto 87,35 39 - 32 28 1Mosaico Q. suber 260,7 44 4 11 39 2Canutos 23,77 26 - 20 25 29Eucaliptar 5,5 - - 64 36 -
Matorral Matorral 28,82 64 - 10 17 9Otros Pastizal y otros 203,7 - - - - -TOTAL 941,82 53 2 17 25 3
Nota. 1. Extremo 2. Elevado 3. Moderado 4. Escaso 5. Sin afectarFuente: Evaluación de los grados de afectación de los grandes incendios de 1997.
Tabla 5. Niveles de afectación para la superficie total incendiada en 1997.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 118/276
Almoraima, 35, 2007
118
4.3 Restauración
Del análisis global de la zona del incendio se proponen las siguientes actuaciones encaminadas a la restauración del monte,así como la no actuación en 234 Ha, correspondientes a las zonas de matorral y pastizal, de rápida regeneración natural:
- Cortas sanitarias en 33 Ha.
- Repoblación forestal en 174 Ha.
- Clareo con densificación en 13 Ha.
- Ayuda a regeneración, cortas y claras 29 Ha.
- Roza entre dos tierras 158 Ha.
- Cortas, podas sanitarias y desbroces selectivo en 199 Ha.
- Desbroce selectivo 30 Ha.
5. RESTAURACION DE LAS ZONAS INCENDIADAS.
Después de los incendios que han recorrido el monte, la administración gestora, la CMA, ha venido ejecutando distintosproyectos de restauración para paliar los efectos negativos del fuego sobre el ecosistema, resumiéndose a continuación losaspectos más destacables.
En 1991 en un proyecto de restauración del Parque Natural Los Alcornocales se aborda el área incendiada en septiembre
de 1989, que se correspondía con una zona repoblada entre los años 1960-65 por el distrito forestal de Cádiz de unas 100Ha de superficie en este monte y 25 Ha del cercano Las Corzas, y afectando a Pinus pinaster (90 Ha), alcornocal (10 Ha)y resto a matorral. Tras el incendio, la zona presentaba una cubierta de brezo, madroño, jaras y aulagas, y al ser la vegetaciónclímax el alcornocal, pero existiendo etapas de degradación en distintas zonas, se realizó unas rozas de regeneración sobrelos pies existentes y densificación en los claros con Q. suber de una savia hasta alcanzar los 800 pies/Ha en los suelos másricos, menos degradados y orientaciones mas protegidas de los vientos. En las zonas más degradadas repoblación con P.
pinaster a densidades de 1.100 pies/Ha hasta pendientes del 30% disminuyendo las densidades hasta las pendientes del60%. En zonas de transición se efectúa repoblación mixta de Q. suber y P. pinaster al 50% a densidades de 830 pies/Ha.
En 1996, un proyecto de restauración de zonas incendiadas incluye actuaciones consistentes en la eliminación de residuosen 64 Ha, restauración mediante casillas y siembra en 21 Ha con P. pinaster ; y plantación en ahoyado manual en 64 Ha
con Q. suber y P. pinaster.En el año 2000, un proyecto de restauración aborda las zonas incendiadas en 1997. Se realizan mejoras en las vías de accesoen 7.072 metros, eliminación de residuos en 97,5 Ha, repoblación en 175 Ha, y cerramientos para aislar del ganado y lafauna cinegética para favorecer la regeneración natural con un longitud de 29 Km.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 119/276
Comunicaciones
119
Figura 4. Plano de vegetación afectada por el incendio de 1997. Fuente: Evaluaciónde los grados de afectación de los grandes incendios de 1997.
Figura 5. Planos de niveles de afectación de la vegetación en el incendio de 1997.Fuente: Evaluación de los grados de afectación de los grandes incendios de 1997.
6. RESULTADOS Y CONCLUSIONES
El monte Hoyo de Don Pedro ha resultado duramente castigado durante la reciente historia por incendios recurrentes que hanafectado a gran parte de su superficie. En el más importante de todos de 1997, se estudió en profundidad su nivel de afectación,y no solo la superficie recorrida (941,8 Ha), destacando que únicamente el 53% de la misma presentaba un daño extremo, yun 45% daños moderados, escasos o sin afectar. Los canutos, las áreas de mayor valor ecológico, fueron afectadas en 24 Hapero solo la cuarta parte con daños extremos. Los mayores porcentajes de daños se produjeron en masas jóvenes de pinar,presentando niveles de daños parecidos en las masas adultas de pinar y alcornocal al ser recorridas por el fuego.
Las técnicas de teledetección aplicadas han permitido mejorar la precisión de los daños, y confirmar que la mediciónrealizada por GPS al final de la extinción presenta una fiabilidad considerable. Al haberse realizado las mediciones conteledetección sobre 30 parcelas (revisadas también en campo) y no sobre toda la superficie recorrida por el fuego, las tablasy parámetros de resultados son susceptibles de mayor fiabilidad conforme se aumente la superficie muestreada (másparcelas) para estudios posteriores.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 120/276
Almoraima, 35, 2007
120
BIBLIOGRAFÍACrónica de Alfonso X , edición de Manuel González Jiménez, Real Academia de Alfonso X El Sabio, Murcia, pp. 195-204. 1998.VÉLEZ RUBIO, Ricardo et al. La Defensa contra Incendios Forestales: Fundamentos y Experiencias. Ed. McGraw-Hill. Madrid. 2000.Evaluación de los grados de afectación de los grandes incendios de 1997. Alternativas de Restauración. CMA-Universidad de Córdoba. 1998. Sin editar.Proyecto de Ordenación del Grupo de Montes de Los Barrios. CMA. 2002.Proyectos de Restauración de zonas incendiadas. CMA. 1991, 1996, 2000.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 121/276
Comunicaciones
121
INTRODUCCIÓN
El litoral andaluz se caracteriza por formar parte de la zona de encuentro entre el mar Mediterráneo y el océano Atlántico.La frontera geográfica entre ambas masas de agua se establece en el estrecho de Gibraltar, aunque la influencia del Atlánticosobre las características físico-químicas del Mediterráneo se percibe muchos kilómetros al este del Estrecho. La franjamarítima de Andalucía no es una entidad homogénea, sino que está separada en varias fracciones con características propiasy distinguibles entre sí. Así, la costa de Andalucía se puede dividir en tres zonas principales en función de las aguas quela bañan: la vertiente atlántica de Huelva y Cádiz; la zona de transición denominada costa peninsular del mar de Alborán,que comprende parte de Cádiz, Málaga, Granada y parte de Almería (hasta la capital), y el levante de Almería, que ya es
una zona típicamente Mediterránea (Rodríguez, 1982).Asimismo, el estrecho de Gibraltar también se considera una frontera biogeográfica que limita la región mediterránea dela región lusitana, ambas comprendidas en la provincia biogeográfica atlanto-mediterránea (Casanova, 1977). Cabedestacar la especial importancia del mar de Alborán, pues es aquí donde se establece el límite de distribución para lasespecies estrictamente atlánticas y mediterráneas, como es el caso, por ejemplo, de la Posidonia oceánica, que es unendemismo mediterráneo cuyas poblaciones no llegan mas allá del estrecho de Gibraltar (Moreno et al., 2004).
Entre los reptiles marinos, la tortuga boba (Caretta caretta) es la especie de quelonio mas frecuente en el litoral andaluz,representando el 90% de los casos de varamientos de tortugas marinas en la costa de Andalucía (CREMA, 2004). Aún no seconoce con detalle el uso que esta especie hace de la costa de Andalucía, aunque los estudios existentes hasta la fecha apuntana que es lugar de paso hacia zonas de forrajeo en el mediterráneo interior y de salida hacia las zonas de puesta del oeste Atlántico
NUEVAS APORTACIONES ALCONOCIMIENTO DE LOS MOVIMIENTOSMIGRATORIOS DE LA TORTUGA BOBA(CARETTA CARETTA) A TRAVÉS DEL
ESTRECHO DE GIBRALTARA PARTIR DE LA INFORMACIÓNRECOPILADA EN VARAMIENTOS
J. J. Bellido López / J. J. Castillo Martín / J. J. Martín Jaime / J. L. Mons ChecaCentro de Recuperación de Especies Amenazadas, Málaga

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 122/276
Almoraima, 35, 2007
122
(Camiñas y De la Serna, 1995). El estudio del registro de varamientos del Centro de Recuperación de Especies MarinasAmenazadas (CREMA) muestra que los varamientos de tortuga boba se producen en todo el litoral andaluz y en todas lasépocas del año, aunque con importantes diferencias entre las distintas estaciones del año y zonas costeras (CREMA, 2004).
El presente estudio se centra en los datos obtenidos en el entorno del estrecho de Gibraltar, presentando los resultados delos registros obtenidos en las provincias de Málaga y Cádiz, que son las provincias de Andalucía más afectadas por lainfluencia del estrecho de Gibraltar.
METODOLOGÍA
Para la realización del presente estudio se ha divido el litoral de las provincias de Cádiz y Málaga en varias zonas en función
de diversas características como la orientación y el tipo de litoral. Así, en Cádiz se han establecido cuatro zonas: CA1 quecomprende todo el entorno de la bahía de Cádiz; entre Chiclana y Tarifa, que es una zona de similar orientación y tipo deplayas, se han establecido dos zonas, CA2 y CA3, fijando la separación en Barbate, punto donde se produce un leve cambioen la orientación de la costa. Finalmente la parte mediterránea de Cádiz ha sido denominada como CA4. La provincia deMálaga se ha dividido en tres zonas, MA1, MA2 y MA3, en función de los cambios de orientación del litoral.
Cádiz
CA1: Sanlúcar de Barrameda-San Fernando.
CA2: Chiclana-Vejer de la Frontera.
CA3: Barbate-Tarifa.
CA4: Algeciras-San Roque. Málaga
MA1: Manilva-Marbella.
MA2: Mijas-Málaga.
MA3: Rincón de la Victoria-Nerja.
La distribución final de las diferentes zonas se refleja en la figura 1.
La diversidad física del litoral andaluz, que ha permitido establecer esta zonificación del mismo, y la existencia dediferencias geográficas en la distribución de los varamientos, nos ha llevado a plantear la hipótesis de que pueden existirdiferencias en la tasa de varamientos de tortuga boba en cada una de las zonas establecidas.
Para el presente estudio se han seleccionado un total de 533 varamientos de tortuga boba registrados en las costas de Cádizy Málaga entre los años 1995 y 2004. Se han elaborado tablas en la que se reflejan el número de varamientos registrados encada una de las zonas establecidas, diferenciando entre varamientos de ejemplares muertos y varamientos de ejemplares vivos.
Una vez que se han establecido las diferentes divisiones del litoral y se ha determinado el número de varamientos deejemplares vivos y muertos en cada una de ellas, se ha realizado un estudio comparativo mediante el uso del métodoestadístico de la χ2 (chi cuadrado). Este test permite determinar si un fenómeno observado, en este caso los varamientosde tortuga boba a lo largo del litoral andaluz, difiere estadísticamente o no de lo esperado por el azar a partir de la siguienteexpresión matemática:
χ2 = Σ (Observado – Esperado)2
Esperado

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 123/276
Comunicaciones
123
Utilizando este método comparativo se han cotejado las zonas establecidas en las dos provincias entre sí, con objeto dedeterminar la existencia de diferencias significativas en la distribución de los varamientos de ejemplares vivos y muertos.
A continuación se describe cómo se han obtenido los datos esperados para la realización de la prueba, considerando dos
grandes zonas en el área de estudio, la vertiente atlántica (litoral atlántico) y la mediterránea (mar de Alborán). En primerlugar se ha obtenido la proporción de varamientos vivos y muertos para toda la zona estudiada dividiendo de formaindependiente el total de varamientos vivos (TV) y muertos (TM) por el total de varamientos (TT) (ver tabla 3). Los valoresesperados para vivos y muertos en cada vertiente se han generado multiplicando la proporción de vivos (P1) y muertos (P2)por el número total de tortugas varadas en cada vertiente.
Finalmente, se ha elaborado una tabla en la que se presenta el total de varamientos por zona y la época del año en el quese han producido.
RESULTADOS
A priori, se pueden observar importantes diferencias en la distribución longitudinal de los varamientos. La vertienteatlántica acumula mayor número de varamientos totales, aunque con una baja incidencia de ejemplares vivos. Sin embargo,en la zona de Alborán se produce menor número de varamientos totales pero con mayor incidencia de ejemplares vivos.
En la tabla 1 se presentan los varamientos producidos en las siete zonas establecidas en el área de estudio diferenciandoentre los casos vivos y muertos.
CA1 V CA1 M CA2 V CA2 M CA3 V CA3 M CA4 V CA4 M34 103 7 31 19 73 30 20
MA1 V MA1 M MA2 V MA2 M MA3 V MA3 M31 26 60 56 25 18
Tabla 1. Distribución de los varamientos de tortuga boba en las siete zonas establecidas en las provincias de Cádiz y Málaga. V: vivos, M: muertos.
Figura 1. Representación sobre el mapa de Andalucía de las siete zonas costeras establecidas en las provincias de Cádiz y Málaga.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 124/276
Almoraima, 35, 2007
124
En la figura 2 se muestra el número total de tortugas bobas, así como los ejemplares vivos y muertos, que se han encontradovarados en cada una de las siete zonas consideradas para este estudio.
Las siete zonas establecidas en el área de estudio se pueden agrupar en dos zonas geográficas claramente definidas: el litoralatlántico y el mar de Alborán. Cada una de ellas estaría integrada por las siguientes zonas:
Litoral atlántico: CA1, CA2 y CA3.
Mar de Alborán: CA4, MA1, MA2, MA3.
La distribución de los varamientos, diferenciando entre ejemplares vivos y muertos, en cada una de estas dos zonasgeográficas se muestra en la tabla 2.
Atlántico V Atlántico M Alborán V Alborán M
60 207 146 120
Tabla 2. Distribución de los varamientos de tortuga boba en las dos grandes zonas geográficas del área de estudio. V: vivos, M: muertos.
Con la información contenida en la tabla 2 se han obtenido los parámetros necesarios para la realización del test de la χ2
(chi cuadrado) (tabla 3) y así, poder comparar las diferencias en la distribución de ejemplares vivos y muertos entre lavertiente atlántica (litoral atlántico) y la mediterránea (mar de Alborán).
Vivas Vivas Total Muertas Muertas Total Total Total TotalAtl Alb Vivas (TV) Atl Alb Muertas Tortugas Tortugas Tortugas
(TM) Atlántico Mediterráneo (TT)Observado 60 146 206 207 120 327 267 266 533Proporciones TV/TT TM/TT
P1: 0,39 P2: 0,61Generación P1*267 P1*266 P2*267 P2*266valores esperadosEsperado 103.2 102.8 163.8 163.2
Tabla 3. Método de obtención de los parámetros necesarios para la realización de la prueba χ2.TV= Total Vivas. TM= Total muertas. TT: Total tortugas. P1 y P2= proporciones.
Los resultados obtenidos en la prueba χ2 al comparar los varamientos producidos en la vertiente atlántica y la vertientemediterránea (tabla 4) ponen de manifiesto que existen diferencias estadísticamente significativas. Concretamente, existeuna confianza superior al 99% de que las diferencias observadas no se deban al azar. Ello implica que en la vertiente atlánticavaran más ejemplares muertos de lo que cabría esperar por azar. Sin embargo, en la vertiente mediterránea del área deestudio ocurre lo contrario, es decir, varan más ejemplares vivos de los esperados por azar. En la figura 2 se aprecia cómoen todas las zonas atlánticas los varamientos muertos son superiores a los vivos, mientras que en la vertiente mediterráneala tendencia, en la mayor parte de las zonas, es a registrar más varamientos vivos que muertos.
grados de libertad χχχχχ2 SignificaciónLitoral atlántico/ litoral Alborán 3 59.01 P< 0.01
Tabla 4. Resultados de la χ2 al comparar los varamientos de las vertientes atlántica y mediterránea.
Finalmente se ha diseñado una tabla en la que los varamientos se agrupan por la zona y la época del año en que se hanproducido (tabla 5).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 125/276
Comunicaciones
125
QUINCENA CA 1 CA 2 CA 3 CA 4 MA 1 MA 2 MA 3Q 1 (1-1 / 14-1) 3 0 4 4 0 6 0Q 2 (15-1 / 28-1) 1 0 2 4 0 0 1Q 3 (29-1 / 11-2) 3 0 2 1 2 2 0Q 4 (12-2 / 25-2) 4 0 3 1 5 5 0Q 5 (26-2 / 11-3) 8 2 1 1 0 3 2Q 6 (12-3 / 25-3) 4 1 4 2 4 5 2Q 7 (26-3 / 8-4) 2 3 1 1 0 5 0Q 8 (9-4 / 22-4) 3 4 0 1 2 3 0
Q 9 (23-4 / 6-5) 6 0 3 0 1 6 1Q 10 (7-5 / 20-5) 7 1 14 2 4 3 1Q 11 (21-5 / 3-6) 13 7 7 1 1 7 3Q 12 (4-6 / 17-6) 10 0 5 0 3 6 3Q 13 (18-6 / 1-7) 7 3 6 3 2 7 3Q 14 (2-7 / 15-7) 9 5 7 1 3 7 6Q 15 (16-7 / 29-7) 3 0 10 3 5 12 7Q 16 (30-7 / 11-8) 12 1 8 6 4 12 5Q 17 (12-8 / 25-8) 12 3 3 1 5 7 3Q 18 (26-8 / 9-9) 8 1 6 2 3 5 1Q 19 (10-9 / 23-9) 7 0 1 5 8 4 3Q 20 (24-9 / 7-10) 4 1 0 0 1 1 0Q 21 (8-10 / 21-10) 9 1 1 1 0 1 0Q 22 (22-10 / 4-11) 3 0 1 2 0 3 2Q 23 (5-11 / 18-11) 3 0 0 0 0 2 0Q 24 (19-11 / 2-12) 2 0 2 0 1 0 0Q 25 (3-12 / 16-12) 2 1 0 2 1 1 1Q 26 (17-12 / 31-12) 0 2 1 2 2 0 0
Tabla 5. Varamientos totales por zonas y quincena del año en la que se han producido. En negrita aparecen reflejados los valores máximos.
Figura 2. Representación gráfica por zona de los varamientos de ejemplares vivos y muertos, así como de los totales.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 126/276
Almoraima, 35, 2007
126
DISCUSIÓN
La tortuga boba es una especie de marcado carácter migratorio, especialmente en su etapa juvenil, que es el momento enel que realiza los desplazamientos de mayor envergadura. Así, en esta etapa abandona su playa de nacimiento y vaga poraguas libres hasta que alcanza el estado subadulto, pudiendo recorrer miles de kilómetros. De esta forma, tortugas nacidasen las costas de Florida pueden llegar a la cuenca mediterránea. Una vez alcanzada la madurez sexual, las tortugas retornana sus áreas de puesta y los movimientos migratorios posteriores no son tan extensos, quedando reducidos desde la zona depuesta a la de forrajeo y viceversa (Musick y Limpus, 1997).
En el curso de estos movimientos numerosos ejemplares entran y salen del mar Mediterráneo a través del estrecho de Gibraltar,pasando frente a las costas de Andalucía (Camiñas y De la Serna, 1995). En la cuenca occidental mediterránea se localiza unárea de forrajeo utilizada tanto por tortugas de origen atlántico como mediterráneo (Margaritoulis et al., 2003).
Una parte de estos ejemplares que transitan frente a las costas de Andalucía sufren diversos accidentes que provocan queterminen varando en el litoral, vivos o muertos y, en este último caso, en distintos estados de descomposición. Losvaramientos producidos en la costa de Andalucía son registrados y almacenados por el CREMA, lo que ha dado lugar auna completa base de datos sobre los mismos (CREMA, 2004). La tasa de varamientos no es constante para toda la líneadel litoral, sino que existen variaciones longitudinales, que pueden estar afectadas por distintos factores como son lasfluctuaciones en el esfuerzo pesquero, la disposición de las corrientes o los temporales. Por estos motivos, aunque no seauna tarea sencilla, los análisis comparativos de la información recogida durante los varamientos nos permite diferenciarzonas que se caracterizan por un determinado ritmo y tipología de varamientos e inferir así un posible patrón de varamientosde tortuga boba en el litoral andaluz.
Los resultados obtenidos muestran que existen importantes diferencias en el comportamiento de los varamientos de tortugaboba en ambos lados del estrecho de Gibraltar. Así, mientras que en la vertiente atlántica existe un claro predominio devaramientos muertos sobre los vivos, en la vertiente mediterránea el comportamiento es el contrario, se registra mayornúmero de ejemplares vivos que muertos (tabla 2). Este hecho tiene una base estadística sólida según los resultadosobtenidos en el test de la χ2 (tabla 4), que pone de manifiesto la existencia de diferencias significativas en el comportamientode la tasa de varamientos en ambas zonas.
En la figura 2 se observa como en la vertiente atlántica las diferencias entre ejemplares muertos y vivos son muy grandesen todas las zonas consideradas, mientras que en la vertiente mediterránea las diferencias son ligeramente favorables hacialos animales vivos en todas las zonas establecidas.
Esta distribución nos sugiere que el mar de Alborán, situado entre el estrecho de Gibraltar y el frente Almería-Orán (este
último se puede considerar ya como la puerta al Mediterráneo abierto) funciona como un corredor para las tortugas,concentrando el número máximo de ejemplares, pues todas las tortugas están obligadas a pasar por ahí para entrar o salirdel mediterráneo. Esto se reflejaría en la alta incidencia de varamientos de ejemplares vivos, pues a diferencia de otras áreas,la estrechez de la franja marítima facilitaría que las tortugas alcanzaran la costa vivas o en buen estado de conservación.
Las áreas adyacentes al mar de Alborán participan también de este flujo de tortugas, pero en menor medida, pues al ser zonasmas abiertas se reduce la concentración de animales, lo que se refleja en la cantidad total de varamientos vivos.
También hay que señalar el papel que juega el litoral atlántico de Cádiz como costa de depósito, al constituir la zona dondeacaban muchos de los ejemplares muertos durante su travesía por el océano Atlántico. Este hecho explicaría la altaincidencia de ejemplares varados muertos. Un aspecto a tener en cuenta es el hecho de que este efecto depósito sobre CA3

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 127/276
Comunicaciones
127
(y sobre CA1 y CA2) apenas afecta a los varamientos vivos, pues existe una importante diferencia entre ambos tipos devaramientos a favor de los ejemplares muertos (figura 2). Parece que las tortugas vivas, mientras que conservan una mínimacapacidad de movimiento dirigido, no tienden a varar en esta zona. Sin embargo una vez dentro de Alborán la incidenciade ejemplares vivos se eleva de manera patente. Es muy posible que las tortugas vivas busquen la corriente de agua atlánticaque penetra en el mar Mediterráneo, y sigan el curso de esta hacia el interior. Por el contrario los ejemplares que derivanmuertos no podrían mantenerse en esta corriente y terminarían por varar en las costas de Cádiz o bien, perdidos en el interiordel mar Mediterráneo.
En la tabla 5 se constata cómo la presencia de varamientos en las costas del estrecho de Gibraltar y zonas adyacentes esconstante a lo largo del año, lo que muestra una presencia continua de ejemplares de tortugas. Sin embargo se observa quehay importantes variaciones a lo largo del año, con máximos entre los meses de mayo y septiembre, siendo estos, a priori,
los meses de máxima actividad de tortugas marinas frente al litoral andaluz.Una vez más, se hace patente la enorme importancia que tiene el estrecho de Gibraltar como corredor natural para el tránsitode especies entre las cuencas mediterránea y atlántica.
BIBLIOGRAFÍABOLTEN A.B. Y B.E. WitheringtonI. (editores). Loggerhead Sea Turtles . Smithsonian Intistution Press. Washington D.C. 2003.CAMIÑAS J.A. Y J.M. De la Serna. The loggerhead distribution in the western Mediterranean Sea as deducted from captures by the Spanish long line fishery. Scientia
Herpetologica 1995: pp. 316-323CASANOVA, J.P. La faune pélagique profonde (zooplancton et micronecton) de la province atlanto–méditerranéenne. Thése Doct. Sciences Naturelles, Universidadde Provence. 1977.CENTRO DE RECUPERACIÓN DE ESPECIES MARINAS AMENAZADAS, (CREMA). Memoria anual 2004 de la consultoría y asistencia para la realización yevaluación de un estudio sobre la migración y el estado sanitario de las tortugas marinas en el litoral andaluz. Consejería de Medio Ambiente, Junta de Andalucía. 2004.LUTZ P. L. y J.A. Musick. (editores) The Biology of Sea Turtle.Vol I . CRC Press. 1997.LUTZ P. L., J.A. Musick y J. Wyneken. (editores) The Biology of Sea Turtle. Vol II . CRC Press. 1997.MARGARITOULIS D., R. Argano, I. Baram, F. Bentivegna, M.N. Bradai, J.A. Camiñas, P. Casale, G. De Metrio, D.G. Gerosa, B.J. Godley, D.A. Haddoud, J.Houghton, L. Laurent Y B. Lazar. Loggerhead turtles in the Mediterranean sea. En: Loggerhead sea turtles , ed. BOLTEN A.B. y WITHERINGTON B.E. Florida,USA. 2003.MORENO, D., A. Luque, y J. Templado. Las praderas de Posidonia oceánica. Distribución en Andalucía. En: Praderas y Bosques Marinos de Andalucía. LUQUEA. Y TEMPLADO J. Consejería de Medio Ambiente, Junta de Andalucía. 2004.RODRÍGUEZ, J. Oceanografía del Mar Mediterráneo. Ed. Pirámide. 1982.SOKAL, R.R. y F.J. Rohlf. Introducción a la Bioestadística. Ed. Reverté. 1984.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 128/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 129/276
Comunicaciones
129
RESUMEN
Este trabajo se ha desarrollado en el ámbito del proyecto “Estudio y Seguimiento de Cetáceos en el Estrecho de Gibraltar”incluido en el Programa de Gestión Sostenible de Recursos para la Conservación del Medio Marino Andaluz de laConsejería de Medio Ambiente de la Junta de Andalucía, y se centra en la bahía de Algeciras. Las campañas de investigaciónse implementaron entre los meses de julio y septiembre de los años 2003 y 2004, ejecutándose de manera simultáneamuestreos desde tierra y en el mar. Se registraron cuatro especies: delfín común ( Delphinus delphis), delfín listado (Stenellacoeruleoalba), delfín mular (Tursiops truncatus) y calderón común (Globicephala melas). En ambos años y tipos demuestreos, el delfín común fue la especie más avistada y con mayor número de individuos por manada, seguido del delfínlistado. No se han encontrado diferencias significativas en el tamaño de manada de D. delphis y S. coeruleoalba entre losdos tipos de muestreos. Aunque los patrones etológicos registrados en ambos tipos de muestreos fueron similares,
existieron diferencias en algunas pautas de comportamiento; así, por ejemplo, la frecuencia de alimentación fuesobreestimada en los muestreos desde tierra frente a los llevados a cabo en el mar. Los datos obtenidos en el presente estudioindican que ambos tipos de muestreos son igualmente válidos para determinar el tamaño medio de manada y la diversidadde especies de cetáceos que existen en la bahía de Algeciras. En cambio, existen ciertas restricciones en la determinacióndel comportamiento en los muestreos desde tierra debido a la dificultad de observación de algunas pautas.
Palabras clave: cetáceos, comportamiento, tipos de muestreo, bahía de Algeciras.
ESTUDIO COMPARATIVO ENTREMUESTREOS DESDE TIERRA Y MARDE LOS CETÁCEOS DELA BAHÍA DE ALGECIRAS
Francisco Ruiz Giráldez / Rafael Gálvez César / José Manuel Guerra García Aurora Montero Chacón / Sara Gil Coronil / José Carlos García Gómez

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 130/276
Almoraima, 35, 2007
130
INTRODUCCIÓN
El presente trabajo se enmarca dentro del proyecto denominado “Estudio y Seguimiento de Cetáceos en el Estrecho deGibraltar” integrado en el Programa de Gestión Sostenible de Recursos para la Conservación de Medio Marino Andaluz,financiado por la Consejería de Medio Ambiente de la Junta de Andalucía y articulado por EGMASA.
La bahía de Algeciras, por su profundidad, riqueza en ictio y malacofauna (principalmente cefalópodos) y su condiciónde refugio del estrecho de Gibraltar, es una zona propicia para albergar cetáceos (Ruiz-Giráldez et al., en prensa). A pesarde ello, la Bahía constituye un enclave muy antropizado. Así, el estudio de las poblaciones de cetáceos posibilita laaplicación directa de la investigación en las diferentes herramientas de gestión y conservación del medio ambiente, que,en el caso de los cetáceos en Europa, son entre otras, la Directiva Hábitats, el Convenio de Barcelona, el Convenio de Bernay el acuerdo de ACCOBAMS del Convenio de Especies Migratorias.
El Laboratorio de Biología Marina de la Universidad de Sevilla viene realizando desde el año 2002 campañas de muestreo,principalmente estivales, destinadas al estudio de las poblaciones de cetáceos de la bahía de Algeciras (pautas decomportamiento, tamaños de manada, etc.), ejecutándose de manera simultánea muestreos desde tierra y mar. El objetivode este estudio es comparar los resultados obtenidos mediante los dos tipos de muestreos durante los años 2003 y 2004,para explorar las ventajas e inconvenientes de cada técnica de muestreo en futuros estudios de cetáceos.
MATERIAL Y MÉTODOS
En este estudio se han desarrollado dos tipos de muestreos en la bahía de Algeciras: muestreos desde tierra y muestreosen el mar.
Para que estos muestreos sean comparables, hay que tener en cuenta las condiciones meteorológicas, ya que la capacidadpara poder identificar los cetáceos depende de la claridad del agua, estado del mar y el comportamiento del animal (Daviset al., 2002). Por tanto, se consideran como esfuerzo nulo o datos no representativos, aquellos que se toman cuando el estadodel mar es de marejada o superior (a partir de 3 en la escala de Douglas) o la fuerza del viento es superior de 3-4 en la escalade Beaufort, o cuando la visibilidad es mala (SEC, 1999; Cañada et al., 2002).
Los datos se obtuvieron entre los meses de julio y septiembre en campañas estivales de 2003 y 2004. En ambos tipos demuestreo (mar y tierra) se anotaron todos los avistamientos realizados, se estimó el comportamiento a partir del porcentajede número de observaciones de las diferentes pautas registradas (alimentación, natación, socialización, remoloneo, etc.)(SEC, 1999) y se anotó el tamaño de manada en cada avistamiento.
MUESTREOS DESDE TIERRA
El punto fijo elegido para los muestreos desde tierra fue Punta Europa (Gibraltar) (05º 21,03´W/36º 06,75´N). Este puntose localiza en la boca de la bahía y a cierta altura, aproximadamente a 25 metros sobre el nivel del mar lo que permite, ademásde conocer las entradas y salidas de cetáceos en la bahía, sus movimientos dentro de ella.
El número de observadores osciló entre un mínimo de dos y un máximo de seis. Se empleó un telescopio Kowa TSN-820(20x60W) para los rastreos, consistentes en fijar el objetivo a una altura determinada, el cual se desplazaba de derecha aizquierda cubriendo el mayor arco posible, a una velocidad que permitiera observar perfectamente todo lo que aparece en

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 131/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 132/276
Almoraima, 35, 2007
132
RESULTADOS
Avistamientos
Tierra
De los 188 avistamientos registrados en Punta Europa en la campaña de 2003, 96 correspondieron a Delphinus delphis(delfín común), seis a Tursiops truncatus (delfín mular) cuatro a Stenella coeruleoalba (delfín listado), uno a manadasmixtas (delfín común y listado) y 81 a delfínidos no identificados (figura 1). El tiempo total de observación invertido enlos muestreos en 2003 fue de 229 horas y 23 minutos.
En la campaña de 2004 se contabilizaron de 161 avistamientos, 50 de delfín común, 15 de delfín listado, dos de calderóncomún, seis de manadas mixtas (delfín común y listado) y 88 de delfínidos sin identificar (figura 1). Se invirtió un tiempototal de 115 horas y 52 minutos.
Mar
En la campaña de 2003 se anotaron 78 avistamientos, 53 de delfín común, 11 de delfín listado, tres de delfín mular, ochode manadas mixtas (delfín común y listado) y tres delfínidos no identificados (figura 1), con un tiempo total de estudio de93 horas y 10 minutos.
En la campaña de 2004 se realizaron 100 avistamientos, de los cuales 69 correspondieron a delfín común, 16 a delfín listado,dos a calderón común, tres a manadas mixtas (delfín común y listado) y 10 a delfínidos no identificados (figura 1), con untiempo de estudio de 93 horas y 18 minutos.
Figura 1. Porcentaje del número de avistamientos en 2003 y 2004 desde tierra y mar.
TIERRA
2003 2004
MAR

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 133/276
Comunicaciones
133
Tamaño de manada
Tierra
En 2003 se observaron manadas de delfines comunes, listados y mulares, siendo mayores las del delfín común, con unamedia de 54 individuos por manada. Sin embargo, no se encontraron diferencias significativas entre el tamaño de lasmanadas de delfín común, listado y mular (Kruskal-Wallis, K=5,02; p>0,05) (figura 2).
El delfín común en 2004, con 31 individuos/manada, fue la especie con mayor número medio de individuos por manada,seguido del delfín listado (24 individuos/manada). Como en el caso anterior, no se encontraron diferencias significativasentre el tamaño de manada de las tres especies observadas en Punta Europa (K= 4,52; p>0,05) (figura 2).
Mar
Las manadas más numerosas avistadas en 2003 fueron las de delfín común (52 individuos/manada), aunque no seencontraron diferencias estadísticamente significativas entre las manadas de delfín común y listado, mientras que lasmanadas de delfín mular fueron significativamente diferentes a las del resto de las especies (Kruskal-Wallis, K=7,5975,p< 0,05) (figura 2).
En 2004, las manadas más numerosas también fueron las de delfín común (46 individuos/manada). Sin embargo, no seencontraron diferencias significativas entre el tamaño de manada del delfín común, delfín listado ni calderón común(K=4,8567; p>0,05) (figura 2).
TIERRA
2003 2004
MAR
Figura 2. Número medio de individuos por manada registrados en 2003 y 2004 desde tierra y mar. Las barras indican el error estándar de la media.
NÚMEROSDEINDIVIDUOS
NÚMEROSDEINDIVIDUOS
NÚMEROSDEINDIVIDUOS
NÚMEROSDEINDIVIDUOS

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 134/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 135/276
Comunicaciones
135
A pesar de la dificultad en los muestreos desde tierra para distinguir las pautas de comportamiento desde la distancia (comocaso más significativo es la socialización que se produce únicamente bajo agua) parece claro, que tanto desde tierra comoen el mar para los dos años de campaña estival, la alimentación es la conducta más observada. Aunque los patronesetológicos registrados en ambos tipos de muestreos fueron similares, existieron diferencias en algunas pautas decomportamiento. Así, por ejemplo, la frecuencia de alimentación fue sobreestimada en los muestreos desde tierra frentea los ejecutados en el mar, especialmente en el año 2004.
Los datos obtenidos en el presente estudio indican la idoneidad del punto fijo de muestreo desde tierra ubicado en Punta
Europa para determinar las diferentes especies que se adentran en la bahía de Algeciras y su tamaño de manada, mientrasque los muestreos en el mar son necesarios para comprobar su comportamiento. En futuros trabajos, se podrá optar por unau otra metodología dependiendo de los objetivos del estudio.
REFERENCIASALTMANN, J. "Observational study of behavior: sampling methods". Behaviour 49: pp. 227-267. 1974.BUCKLAND, S.T., D.R. Anderson, K.P. Burnham, y J.L. Laake. Distance Sampling: Estimating Numbers of Biological Populations . Chapman y Hall, 1993. Londres.CAÑADAS, A., R. Sagarminaga, y S. García-Tiscar. "Cetacean distribution related with depth and slope in the Mediterranean waters off southern Spain". Deep-searesearch 49: pp. 2053-2073. 2002.DAVIS, R.W., J.G. Ortega-Ortiz, C.A. Ribic, W.E. Evans, D.C. Biggs, P.H. Resseler, R.B. Cady, R.R. Leben, K.D. Mullin y B. Würsig. "Cetacean habitat in thenorthern oceanic Gulf of Mexico". Deep-sea research 49: pp. 121-142. 2002.
TIERRA
2003 2004
MAR
%
DELNÚMERODEOBSERVACIONES
Figura 3. Porcentaje del número de observaciones de las diferentes pautas de comportamiento obtenidas desde tierra y en el mar en los años 2003 y 2004(AL=alimentación; SO=socialización; Nr=natación rápida; Nm=natación media; Nl=natación lenta; Nsd=natación sin determinar; RE=remoloneo; R=reposo).
%
DELNÚMERO
DEOBSERVACIONES
%
DELNÚMERODEOBSERVACIONES
%
DELNÚMERO
DEOBSERVACIONES

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 136/276
Almoraima, 35, 2007
136
DIXON, W. J. BMDP Statistical Sofware. Univ. California Press, Berkeley. 1983.FORCADA, J y P. Hammond. "Geographical variation in abundance of striped and common dolphins of the western Mediterranean". Journal of sea research 39: pp.313-325. 1998.GERRODETTE, T. y J. Forcada. "Non-recovery of two spotted and spinner dolphin populations in the eastern tropical Pacific Ocean". Marine Ecology Progress Series291: pp. 1-21. 2005.HIBY, A. R. y P. S. Hammond. "Survey Techniques for Estimating Abundance of Cetaceans". Rep. Int. Whal. Commn. Special issue II. 1989.RUIZ-GIRÁLDEZ, F., A. Montero, R. Gálvez, y J.C. García-Gómez, En prensa. "Resultados preliminares del estudio de las poblaciones de cetáceos en la bahía deAlgeciras". Almoraima .SEC. 1999. Recopilación, análisis, valoración y elaboración de protocolos sobre las labores de observación, asistencia a varamientos y recuperación de mamíferosmarinos de las Aguas Españolas. Informe técnico. Sociedad Española de Cetáceos.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 137/276
Comunicaciones
137
INTRODUCCIÓN
El estrecho de Gibraltar es la única conexión natural existente entre el mar Mediterráneo y el océano Atlántico. Por ello, seríael único punto de unión para las poblaciones animales presentes en el Atlántico y el Mediterráneo. El estrecho de Gibraltarse conoce como uno de los puntos donde se desarrollan las migraciones de gran número de especies, ya sea de norte a sur, o
de este a oeste. Aves, poblaciones piscícolas e incluso tortugas marinas realizan movimientos migratorios o pasos a través delEstrecho. Entre ellas, cabe destacar especies de aves, como el milano negro ( Milvus migrans) o la cigüeña blanca (Ciconiaciconia,), de entre las planeadoras, y por ejemplo, de entre las no planeadoras, muchas de ellas pequeñas paseriformes, elverderón común (Carduelis chloris) o el vencejo común ( Apus apus), las cuales atraviesan el Estrecho por el eje Norte-Sur.Por otra parte, aves marinas como el alcatraz atlántico ( Morus bassanus) o la pardela balear (Puffinus mauretanicus) pasana través del eje Este-Oeste (Barros D. y Ríos Esteban D., 2002, y Paterson, A. 2002). En el caso de las poblaciones piscícolas,el atún rojo, (Thunnus thynnus), realiza movimientos a través del Estrecho para reproducirse en el Mediterráneo, hecho quees aprovechado por pescadores para atraparlos, ya sea gracias a redes laberínticas llamadas almadrabas, o por medio de lallamada pesca con piedra (FAO-Copemed 1998, FAO-Copemed 1999, FAO-Copemed 2000).
La distribución temporal de cetáceos en el estrecho de Gibraltar está pobremente descrita y se limita a unas pocas fuentes.Una de ellas proviene de datos de las actividades comerciales balleneras que tuvieron lugar entre 1921 y 1954 desde
estaciones balleneras en tierra, en Getares (España) y Benzou (Marruecos) y desde barcos factoría. Se calcula que 826cachalotes (Physeter macrocephalus), 4.516 rorcuales comunes ( Balaenoptera physalus) y 189 rorcuales boreales( Balaenoptera borealis) fueron capturados durante este período (Aloncle 1964, Aguilar y Lens 1981, Bayed y Beaubrun1987, Sanpera y Aguilar 1992). Un estudio llevado a cabo desde los ferries que unían España y Ceuta, desde marzo a mayode 1999, demostró que la mayor parte de las especies encontradas en el sector Este del Estrecho eran delfines listados(Stenella coeruleoalba), delfines comunes ( Delphinus delphis), y, ocasionalmente, delfines mulares (Tursiops truncatus),calderones comunes (Globicephala melas) y cachalotes (Roussel 1999). Asimismo, Silvani et al. (1999), en un estudiosobre capturas accidentales de redes a la deriva en el Mediterráneo occidental, pone de relevancia la abundante presenciade delfines comunes y delfines listados en el Estrecho. Sin embargo ninguno de estos estudios hace referencia a ladistribución temporal de las especies de cetáceos que se encuentran a lo largo del estrecho de Gibraltar, ni al grado deresidencia que las especies observadas tienen en el mismo.
DISTRIBUCIÓN TEMPORAL DE CETÁCEOSEN EL ESTRECHO DE GIBRALTAR.¿ES REALMENTE UN CANAL DE MIGRACIÓN
PARA CETÁCEOS EL ESTRECHO?
Renaud De Stephanis, Renaud / Philippe Verborgh / Sergi Perez Alicia Sánchez / Christophe Guinet / Bruce Mate

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 138/276
Almoraima, 35, 2007
138
Diferentes órganos administrativos, tanto nacionales como autonómicos, grupos conservacionistas, ecologistas, prensaescrita, radiofónica o televisiva, siempre han hablado de una posible migración de grandes cetáceos a través del Estrecho.Sin embargo, no existe ninguna referencia bibliográfica que demuestre este hecho. De las especies de cetáceos de grantamaño que podrían migrar a través del Estrecho, estaría el cachalote y el rorcual común, si nos centramos en las especiesde gran tamaño que están presentes en el Mediterráneo y el Atlántico. El rorcual común es el depredador de más tamañoobservado en el Mediterráneo. Según Berubé (1995 y 1998), las poblaciones de rorcual común del Atlántico norte, y lasdel Mediterráneo, son genéticamente distintas. Cada verano se estima que entre 1.000 y 2.000 individuos se concentranen la parte norte de la cuenca occidental mediterránea (mar de Liguria y golfo de León) (Forcada J., Aguilar A. 1996), endonde esta especie se alimenta de zooplancton ( Meganyctiphanes norvegica) (Orsi Relini et al., 1992 y Viale (1991). Sinembargo, no existe una imagen clara de la distribución de rorcuales comunes en esta zona a lo largo del tiempo, ni se sabe
si existe una migración de rorcuales comunes a través del Estrecho. Algunos autores han sugerido que podría existir unaabundancia máxima de rorcuales comunes durante el periodo estival (Duguy et Vallon, 1976). Marini et al. (1995) indicaa partir de observaciones en el mar que la población de rorcuales comunes podría invernar en las costas norte africanas.Viale (1977) a y b, y emitió la hipótesis (no demostrada a día de hoy), que la cuenca mediterránea podría ser utilizada poruna población de verano para su alimentación, y luego por otra población invernal para la reproducción. Referente a loscachalotes del mediterráneo, no se tiene información referente a su abundancia absoluta, distribución espacial o temporal.
Por ello, este estudio tiene como objetivos contestar a las siguientes preguntas: 1) ¿Cuál es la abundancia relativa de lasespecies de cetáceos presentes en el estrecho de Gibraltar a lo largo del año? 2) ¿Cuál es el grado de fidelidad hacia la zonapor parte de los cetáceos presentes en el estrecho de Gibraltar, a lo largo del año? 3) ¿Existe alguna especie que migre a travésdel Estrecho? En caso afirmativo, ¿Cuál es su origen y destino?
MATERIAL Y MÉTODOS
Área de estudio
La zona de estudio es el área del estrecho de Gibraltar, comprendida entre los 5º y 6º de longitud Oeste, intentando cubrir todael área hasta las aguas del reino de Marruecos. El estrecho de Gibraltar (figura 1) tiene aproximadamente 60 Km de largo. Sufrontera oeste está localizada entre el cabo de Trafalgar, en España, y el cabo Espartel, en Marruecos, separados por 44 Km.El Estrecho se estrecha hacia el este llegando a una distancia mínima entre Tarifa y Punta Cires de 14 Km. Su frontera esteestá localizada entre Punta Europa (Gibraltar) y Punta Almina (África) y están separadas por 23 Km (Parrilla et al., 1988).La batimetría del Estrecho se caracteriza por ser un cañón que va de este a oeste con aguas poco profundas (200-300 m) quese pueden encontrar en la cara atlántica y aguas más profundas (800-1000 m) en la cara mediterránea (figura 1).
Origen de los datos
Los datos de este estudio provienen de dos fuentes. Se realizaron muestreos desde embarcaciones de whale watching deuna hora y cincuenta y siete minutos cada uno de media, con investigadores experimentados a bordo entre julio de 1998y diciembre de 2000. Todos estos trayectos fueron realizados de manera independiente, e intentaban cubrir toda la zonade estudio a lo largo de una semana. Entre febrero de 2001 y julio de 2004, se tomaron datos desde el barco de investigaciónELSA, propiedad del Grupo de Investigación CIRCE; que es una motora de 11 metros de eslora, 2,80 metros de manga,y que tiene una plataforma que permite posicionar observadores a 4 metros sobre el nivel del mar. Está provista de dosmotores Perkins de 82 CV de potencia, que alimentan un convertidor eléctrico que permite distribuir corriente eléctricaa 220 voltios y 300 watios, desarrollando una onda sinusoidal pura, que alimentan un ordenador de abordo que toma la ruta

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 139/276
Comunicaciones
139
y la posición de la embarcación cada minuto. Los trayectos no siguieron un diseño sistemático y fueron llevados a caboaleatoriamente en el Estrecho durante los muestreos. La estrategia de muestreo fue idéntica durante todo el periodo deestudio y para los dos tipos de plataformas de investigación. La zona de estudio fue muestreada a una velocidad media de5,3 nudos. El esfuerzo de búsqueda era parado cuando se contactaba con un grupo de cetáceos y volvía a empezar otra vezcuando el avistamiento se terminaba, volviendo al rumbo establecido previamente. Los observadores se situaban en unaplataforma de observación a 4 metros sobre el nivel del mar. Dos observadores entrenados ocupaban el puesto deobservación en turnos de una hora, durante la luz del día, cuando la visibilidad superaba las 3 millas náuticas (5,6 Km) yprovistos de unos prismáticos 8 x 50, cubriendo 180º por delante del barco. El esfuerzo de búsqueda fue medido como elnúmero de kilómetros navegados con condiciones de avistamiento adecuadas (con un estado de mar menor de 4 en la escalaDougglas y con dos observadores en el puesto de observación). Se siguió en todo momento los protocolos de observacióndesarrollados en 1999 por la SEC, Sociedad Española de Cetáceos (SEC 1999). La posición geográfica de la embarcación
era grabada continuamente en un ordenador portátil desde un GPS y a través de un programa informático (IFAW DataLogging Software Logger 2000 versión 2.20) provisto por la International Fund For Animal Walfare). Un servidor NMEAgrababa automáticamente la hora, la posición, el rumbo, y la velocidad cada minuto en una base de datos. Datos relativosa la hora, especie, número de individuos, comportamiento y otros datos relevantes durante los avistamientos fuerongrabados con otros datos ambientales relevantes. Un avistamiento fue definido como un grupo de animales de la mismaespecie observados a la misma hora que llevan un comportamiento similar y que están a menos de 1.000 m de distanciaentre ellos (SEC 1999). Cuando un grupo de animales era observado se grababa la posición de la embarcación, la distanciay la demora con respecto a los cetáceos. La posición exacta de los animales era grabada cuando se establecía el contactocon los mismos (a menos de 100 m de los animales). Algunos grupos de animales eran observados pero no contactados.
Figura 1. Área de estudio.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 140/276
Almoraima, 35, 2007
140
El esfuerzo de observación no tuvo en cuenta los kilómetros navegados cuando se estaba siguiendo un grupo de animales(desde que eran contactados hasta que se abandonaban).
Distribución temporal de cetáceos
La distancia en kilómetros de búsqueda en cada mes fue entonces calculada utilizando un sistema de informacióngeográfico: Arc-view 3.2 de ESRI y su extensión Animal Mouvement (Hooge and Eichenlaub 2000) proporcionados porel CEBC-CNRS (Centre D´Etudes Biologiques de Chizé del Centre Nationale de Recherche Scientifiques).
Para cada mes se calculó un parámetro, la tasa de encuentro de individuos, definidos como AI (Abundance Index Rate),y cuya formula es:
Abundance Index Rate (AI) (ind/Km.) fue definido como: AI = (Ind / Eff) x 100
Ind: número de individuos de una especie dada, observados con esfuerzo de búsqueda en la zona de estudio.
Eff: Distancia (Km) cubierta con esfuerzo de búsqueda.
Grado de residencia de las especies en el Estrecho
Durante los encuentros, se tomaron fotografías de aletas dorsales de los animales avistados, para el caso de delfines mulares,calderones comunes, orcas, y de aletas caudales para cachalotes. Las fotografías tomadas entre 1998 y 2003, están en formatode diapositivas. A partir de finales de 2003, las fotografías pasan a ser digitales. Se levantaron catálogos de fotoidentificaciónsiguiendo la metodología de Würsig et al., (1990). Cada individuo tiene una aleta dorsal diferente, y por tanto a partir de estasfotografías podremos levantar un catálogo que represente a todos los individuos “capturados” a partir de estas fotografías(Wilson et al. 1999). Comparando las fotos año tras año, y mes tras mes, podremos por tanto comparar los animales presentes
cada temporada, y por tanto, podremos saber el grado de residencia que tiene cada especie catalogada.Migración de rorcual común a través del Estrecho: análisis de isótopos estables en barbas de rorcual común
Para averiguar en qué tipo de aguas se han alimentado rorcuales comunes durante el tiempo, se procedió a analizar relacionesporcentuales de isótopos estables de carbono (13C/12C) y nitrógeno (15N/14N) a lo largo de diez barbas de rorcual comúnvarados en el mediterráneo franco-español y en tres rorcuales varados en la costa atlántica o mar de Alborán. Estos valoresse compararon con medidas de estas relaciones de la dieta de los rorcuales comunes del Mediterráneo y el Atlántico para podercompararlos y poder saber así si hay alternancias en las relaciones a lo largo del tiempo, y por tanto si existen diferentes puntosde alimentación para esta especie, así como ver si hay movimientos de ésta a través del estrecho de Gibraltar.
Migración de rorcual común a través del Estrecho: despliegue de balizas ARGOS, de seguimiento por satélite, en
Rorcual comúnPara poder confirmar si existe un paso de rorcuales a través del Estrecho, se ha procedido también a marcar once rorcualescomunes en la cuenca mediterránea occidental durante el mes de agosto de 2003 con emisores por satélite. Estos emisoresdan una posición del individuo marcado cada tres días, así se tendría una idea clara de si existen movimientos o no de estosanimales a través del Estrecho.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 141/276
Comunicaciones
141
RESULTADOS
Muestreos realizados
Se ha recorrido un total de 22.649 kilómetros durante los cuales el esfuerzo de búsqueda era el más adecuado para esteestudio, cubriendo prácticamente todo el estrecho de Gibraltar (figura 2 y tabla 1). Tan sólo los meses de enero, febreroy diciembre se quedaron con menos de 300 kilómetros muestreados, por lo que los resultados de estos meses se toman concautela. Asimismo, se realizó un total de 2.288 avistamientos de diez especies de mamíferos marinos con un total de 72.842individuos observados (tabla 2).
Distribución temporal de cetáceos
Para realizar los análisis sobre distribución temporal, el estudio se va a centrar en las siete especies de cetáceos másabundantes en el mismo. Estos resultados se pueden observar en las figuras 3 a 9.
Grado de residencia de las especies en el Estrecho
Se tomaron fotografías de cinco de las especies más comúnmente observadas en el Estrecho, delfines mulares, calderonescomunes, orcas, cachalotes y rorcuales comunes. Asimismo, se tomaron fotografías de delfines comunes. Sin embargo paraesta especie, no se levantaron catálogos de fotoidentificación debido a la gran abundancia de individuos. Para delfineslistados, se optó por no tomar fotografías debido también a su gran abundancia, y a que las marcas en las aletas dorsalesno son tan evidentes como para el resto de las especies estudiadas. En la siguiente tabla se resume el número de fotografíastomadas y el número de individuos identificados para cada especie.
Figura 2. Muestreos realizados en el Estrecho entre 1998 y 2004. En rojo se representan los seguimientos a cetáceos.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 142/276
Almoraima, 35, 2007
142
ESPECIES AVISTAMIENTOS INDIVIDUOS OBSERVADOSBalaenoptera acutorostrata 2 2Balaenoptera musculus 1 1Balaenoptera physalus 26 41Cistophora cristata 2 3Delphinus delphis 451 15.993Glovicephala melas 576 14.717Orcinus orca 50 255Physeter macrocephalus 348 446Stenella coeruleoalba 435 34.076Tursiops truncatus 397 7.308Total general 2.288 72.842
Tabla 2. Avistamientos realizados en el Estrecho entre 1998 y 2004.
Migración de rorcual común a través del Estrecho: análisis de isótopos estables en barbas de rorcual común
Los análisis de los patrones de los isótopos 15N/14N indicaron que estas ballenas se alimentaban básicamente en nivelestróficos secundarios, observándose variaciones en los niveles de N a lo largo de la barba.
Migración de rorcual común a través del Estrecho: despliegue de balizas de seguimiento por satélite ARGOS enrorcual común
Las marcas emitieron desde nueve rorcuales con una duración máxima de ocho meses para uno de los individuos. En uno delos casos, el animal cruzó el Estrecho durante el mes de noviembre, y estuvo moviéndose por antiguas zonas de caza ballenera.
DISCUSIÓN
Por los resultados obtenidos en las figuras 3 a 10, se puede apreciar que los meses de diciembre a febrero producen unaserie de distorsiones en los resultados, probablemente debido a la falta de datos (esfuerzo de búsqueda), que le den robusteza los mismos. Por ello, en esta discusión tan solo se podrá hablar de lo que pasa durante los meses primaverales a otoñales,e intuir con la tendencia de las gráficas lo que podría pasar en invierno. Las especies observadas de forma común en elEstrecho son los delfines listados, comunes y mulares, los calderones comunes, las orcas, los cachalotes y los rorcualescomunes, mientras que rorcuales aliblancos, rorcuales azules y focas de casco lo harían de forma casual, y probablementedebido a procesos de desorientación. Dentro de las especies comúnmente observadas, se puede asimismo separar tresgrupos: por un lado las especies que están presentes todo el año, que son las cuatro primeras mencionadas anteriormente,
MES ENERO FEBRERO MARZO ABRIL MAYO JUNIO1998-2000 Km Navegados 22'75 22'75 126'40 499'37 662'72 994'272001-2004 Km Navegados 178'70 240'88 326'97 427'16 515'90 1.089'271998-2004 Km Navegados 201'45 263'63 453'37 926'53 1.178'62 2.083'54
MES JULIO AGOSTO SEPTIEMBRE OCTUBRE NOVIEMBRE DICIEMBRE1998-2000 Km Navegados 3.314'38 4.521'78 2.485'78 1.301'91 0'00 145'852001-2004 Km Navegados 2.152'95 1.880'66 839'46 831'93 355'10 0'001998-2004 Km Navegados 5.467'33 6.402'44 3.325'24 2.133'84 355'10 145'85
Tabla 1. Kilómetros muestreados en el Estrecho entre 1998 y 2004.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 143/276
Comunicaciones
143
Figura 5. Distribución temporal de AI en delfín mular.
Figura 3. Distribución temporal de AI en delfín común.
Figura 4. Distribución temporal de AI en delfín listado.
por otro lado las que solamente se observan durante ciertaparte del año, que son cachalotes, orcas, y rorcuales comu-nes. Los delfines mulares y los calderones comunes, sonresidentes, según los datos obtenidos a través de sus catálo-gos de fotoidentificación, aunque se puede apreciar que hayvariaciones a lo largo del año, siendo más abundante laprimera especie durante la primavera, y durante el final delverano e invierno para la segunda especie (variación menosabrupta en este caso). Para el caso de los delfines comunesno se realizaron catálogos de fotografías, aunque observa-
ciones en el mar confirman que algunos individuos estánpresentes durante todo el año. Su distribución a lo largo delaño presenta un máximo a finales de primavera. Para el casode los delfines listados, no se pudo identificar a ningúnindividuo, debido a su gran abundancia (grupos de más dequinientos ejemplares en algunos casos), por lo que no sepuede hablar de un grado de residencia importante para estaespecie. Sin embargo, es fácil pensar que una parte de losgrupos observados están presentes a lo largo de todo el año,mientras que existe una población flotante menos sedentariaque ocuparía el Estrecho a finales del verano y durante los
meses de otoño, procedente probablemente del mar deAlborán o del Atlántico contiguo. En todos los casos, (enmenos grado para calderones y delfines comunes), se ve quehay variaciones a lo largo del año, que pueden ser debidas ados posibilidades: por un lado se podría pensar que algunasespecies vienen al Estrecho para reproducirse, y por otrolado, podría ser para alimentarse únicamente. El primero delos casos es fácilmente descartable, debido a las condicionesmeteorológicas del Estrecho, sin embargo, la segunda razónes la más plausible, debido a que por un lado el Estrecho esuna zona rica en nutrientes, y, por tanto, también en presas
para estas especies. Por otro lado, es una zona de paso paradiferentes especies piscícolas que son presa de cetáceos. Alproducirse un embudo, sería una zona fácilmente explotablepor éstos a la hora de capturarlos. La colonización temporaldel Estrecho por estas poblaciones flotantes se haría portanto desde el mar de Alborán o del Atlántico contiguo.
En el caso de los cachalotes, se observa que tan solo veintiúnindividuos surcan el Estrecho desde finales del inviernohasta mediados del verano, con máximos en primavera. Esta

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 144/276
Almoraima, 35, 2007
144
población es por tanto muy reducida, viniendo al Estrechosiempre los mismo individuos, probablemente debido aprocesos alimenticios. Es posible la existencia de algúnfenómeno migratorio de sus presas, probablementecefalópodos, que haga que esta población venga al Estrechodurante esta estación del año. Este hecho descarta cualquierproceso migratorio a través del Estrecho por parte de cacha-lotes, siendo la especie por tanto semi-residente en el Estre-cho. De los veintiún individuos, tan sólo dos presentabanmarcas pronunciadas en sus aletas caudales, lo que hace
pensar que esta especie está en una zona carente dedepredadores naturales hacia ellas el resto del año también,como puedan ser tiburones u orcas depredadoras de mamí-feros marinos. Esto podría hacer pensar que esta poblaciónde cachalotes proviene del Mediterráneo. En seis indivi-duos, se pudo obtener muestras de piel, las cuales han sidoanalizadas en la Universidad de Durham, y mostraron quelos seis eran machos. El tamaño medio de los ejemplares erade unos 12 metros, lo que hacía pensar que son grupos demachos sub-adultos del Mediterráneo que vienen a alimen-tarse al Estrecho durante la primavera.
Las orcas únicamente fueron observadas en la zona de estudiodurante los meses de julio y agosto. En el 90% de los casos,fueron observadas asociadas a los pescadores de atún conpiedra de Tarifa. Se las observó alimentándose de atunes entodos los casos. Según se ve en la figura 2, la zona de estudiose limita al centro del Estrecho. En los muestreos realizados
por CIRCE en las costas del golfo de Cádiz, se pudieron observar orcas durante los meses de abril a septiembre, y los datosque se presentan en este estudio, confirman también la presencia de esta especie durante el mes de noviembre. Por tanto, lasorcas estarán presentes en el centro del Estrecho, asociadas a las pesquerías de atún, durante los meses de julio y agosto,aprovechando el esfuerzo de pesca de los pescadores tarifeños, y el resto del año estarían presentes en el golfo de Cádiz.
Durante el invierno, no se ha observado la especie, pero este hecho puede ser debido a la falta de esfuerzo por parte de losestudios de CIRCE durante esta época del año.
ESPECIES FOTOGRAFÍAS TOMADAS INDIVIDUOS IDENTIFICADOSBalaenoptera physalus 450 10Delphinus delphis 1.032 0Globicephala melas 9.865 192Ornicus orca 3.834 26Physeter macrocephalus 548 21Stenella coeruleoalba 0 0Tursiops truncatus 4.567 151
Tabla 3. Fotografías tomadas e individuos identificados por especie.
Figura 6. Distribución temporal de AI en calderón común.
Figura 7. Distribución temporal de AI en cachalote.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 145/276
Comunicaciones
145
Finalmente, el rorcual común se ha confirmado como laúnica especie que se observa transitando el Estrecho, sinquedarse a alimentarse. En todas las observaciones realiza-das entre abril y octubre, se intentó hacer un seguimiento delos mismos, y en todos los casos, los animales transitaban agrandes velocidades (entre 5 y 8 nudos) hacia el Atlántico.En los avistamientos realizados durante el invierno se pudoobservar a los rorcuales alimentándose lentamente. La pocatasa de encuentro durante esos meses, hace pensar que estetipo de alimentación es ocasional y podrían ser individuos
provenientes del Atlántico contiguo según los datos obteni-dos por isótopos estables y de seguimiento por satelite de losmismos. Los análisis de los patrones de los isótopos 15N/ 14N indicaron que estas ballenas se alimentaban básica-mente en niveles tróficos secundarios, tanto en el Mediterrá-neo, como para las barbas obtenidas del mar de Alborán yAtlántico contiguo. La comparación de los patrones de losisótopos 13C/12C de las barbas y muestras de krill delMediterráneo y Atlántico sugieren que estos animales pre-sentan un tipo de comportamiento migratorio. Ocho de losrorcuales demostraron ser residentes (en cuanto a su alimen-
tación al menos) en el Mediterráneo, mientras que tresindividuos mostraron grandes variaciones en sus relacionesde 13C/12C que sugieren migraciones regulares hacia elAtlántico. Los datos obtenidos a partir de las balizas Argosconfirman los resultados mencionados anteriormente, y portanto confirman que una parte de los rorcuales comunes delMediterráneo migra regularmente al Atlántico a través delestrecho de Gibraltar. La causa de que sea tan sólo una parte de esta población la que migra regularmente a las zonas decaza ballenera de los años cincuenta y sesenta, podría estar en una posible pérdida cultural de estas poblaciones debido ala caza ballenera que se realizó durante los años veinte y cincuenta en el Estrecho de Gibraltar, y durante los cincuenta ysesenta en el Atlántico contiguo. Así pues, se produciría una importante disminución en las posibilidades de transmisión
y permanencia de los conocimientos sobre migraciones de una generación a otra de rorcuales comunes. Por otra parte,también podría explicar el hecho que exista una diferenciación genética entre las poblaciones de rorcual común delMediterráneo y Atlántico norte.
CONCLUSIÓN
Por un lado, se ha demostrado que existen siete especies presentes en el Estrecho de forma común. Cuatro de estas, losdelfines listados, comunes, mulares y calderones serían residentes durante todo el año, dos de ellas, cachalotes y orcas,serían semiresidentes y, finalmente, una especie, el rorcual común, utilizaría el Estrecho como canal de paso para susmigraciones. Las cuatro primeras especies mencionadas anteriormente, tendrían variaciones en cuanto a su abundancia
Figura 9. Distribución temporal de AI en rorcual común.
Figura 8. Distribución temporal de AI en orca.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 146/276
Almoraima, 35, 2007
146
relativa a lo largo del año, lo que podría hacer pensar en una población residente en y una pequeña población flotanteproveniente del Atlántico contiguo o mar de Alborán.
En el caso de los cachalotes, se estaría hablando de un grupo de machos sub-adultos mediterráneos que vienen a alimentarseal Estrecho durante los meses primaverales.
Las orcas estarían presentes en las partes centrales del Estrecho, durante los meses de julio y agosto, y en el golfo de Cádizel resto del año, desconociéndose su paradero durante los meses de invierno.
Finalmente, una especie, el rorcual común, migraría a través del Estrecho, hacia zonas de alta productividad del Atlánticocontiguo. Este estudio demuestra también que debe de existir una pérdida cultural en el ámbito de la población de rorcualcomún del Mediterráneo, debido a la sobreexplotación a la que esta especie se vio sometida en los últimos siglos.
BIBLIOGRAFÍAAGUILAR A, Lens S, "Preliminary report on spanish whaling activities". Rep Int Whal Commn 31: pp. 639-643. 1981.ALONCLE H, "Premières observations sur les petits Cétacés des côtes marocaines". Bull Inst Pech Maroc 12: pp. 21-42. 1964.BARROS D. y Ríos Esteban D. Guía de Aves del Estrecho de Gibraltar , Ed. Ornitour, S.L., La Línea de la Concepción. 2002.BAYED A, Beaubrun PC, "Les mammifères marins du Maroc: Inventaire préliminaire". Mammalia 51(3): pp. 437-446. 1987.BÉRUBÉ M., Palsboll P., Larsen F., Sears R., Notarbartolo di Sciara G., Aguilar A., Sigurjonsson J., Urban Ramirez J., Dendanto D. Genetic structure of the North
Atlantic fin whale, Balaenoptera physalus L.: analysis of mitochondrial and nuclear D.N.A. Abstracts, eleventh Biennal Conference on the Biology of MarineMammals, 14-18 Dec., Orlando, Florida. 1995.BÉRUBÉ, M., Aguilar, A., Dendanto, D ;, Larsen, F., Notarbartolo di Sciara G., Sears R., Sigurjonsson J., Urban R J., Palsboll, P.J. "Population genetic structureof North Atlantic, Mediterranean Sea and Sea of Cortez fin whales, Balaenoptera physalus (Linnaeus 1758): analysis of mitochondrial and nuclear loci". Molecular Ecology 7, pp. 585-599. 1998.DUGUY R., Vallon D. "Le Rorqual commun ( Balaenoptera physalus) en Méditerranée Occidentale: Etat actuel des observations". XXVème Congrès-Assemblée
plénière de Split , 22-30 Octobre 1976, Comité des Vertébrés marins et Céphalopodes. 1976.FAO-COPEMED. 1998. Project FAO-COPEMED / Gibraltar98 / Aim7: discussion and conclusions.FAO-COPEMED. 1999. Projet FAO-COPEMED/Tunidos99/Rapport final du Maroc 1999.FAO-COPEMED. 2000. Projet FAO-COPEMED/Grandes Pelagicos’2000.FORCADA J., Aguilar A. "Distribution and abundance of fin whales ( Balaenoptera physalus) in the Western Mediterranean Sea during the Summer". J. Zool., Lond.238: pp. 23-34. 1996.HOOGE PN, Eichenlaub B, Animal movement extension to Arcview, ver 2.0. Alaska Science Center - Biological Science Office, U.S. Geological Survey, Anchorage,AK, USA (http://www.absc.usgs.gov/glba/gistools/index.htm#ANIMAL MOVEMENT). 2000.MARINI L., Villetti G., Consiglio C. 1995. "Wintering areas of fin whales ( Balaenoptera physalus) in the Mediterranean Sea: a preliminary survey". Ninth EuropeanCetacean Society Meeting, Lugano, 9-11 February 1995.ORSI Relini L., Relini G., Cima C., Fiorentino F., Palandri G., Relini M., Torchia G. Una zona di tutela biologica ed un parco pelagico per i cetacei del Mar Ligure.Boll. Mus. Ist. Biol. Univ. Genova. 56-57: pp.247-28, 1990-1991. 1992.PARRILLA G, Kinder TH, Bray NA, "Hidrología del agua mediterránea en el Estrecho de Gibraltar durante el Experimento Gibraltar (octubre 1985-octubre 1986)".Seminario sobre la oceanografía física del Estrecho de Gibraltar (Madrid, 24-28 de octubre 1988), pp. 95-121. 1988.PATERSON, A. Aves Marinas de la Península Ibérica, Baleares y Canarias, Ed. Edilesa, León. 2002.
ROUSSEL E., Les cétacés dans la partie orientale du Détroit de Gibraltar au printemps: indications d’écologie. Master thesis. Ecole Pratique des Hautes Etudes,Montpellier, France. 1999.SANPERA C, Aguilar A, "Modern Whaling off the Iberian Peninsula during the 20th Century". Rep Int Whal Commn 42: pp. 723-730. 1992.SEC. Sociedad Española de cetáceos, Recopilación, análisis, valoración y elaboración de protocolos sobre las labores de observación, asistencia a varamientos yrecuperación de mamíferos y tortugas marinas de las aguas españolas. Ministerio de Medio Ambiente Español. Secretaria General de Medio Ambiente, TechnicalReport Sociedad Española de Cetáceos Avaible from SEC, Nalón 16, E-28240 Hoyo de Manzanares, Madrid, Spain. 1999.SILVANI, L., M. Gazo and A Aguilar. “Spanish driftnet fishing and incidental catches in the western Mediterranean.” Biological Conservation 90: pp. 79-85. 1999.VIALE D. "Contribution à l’étude des cétacés en Méditerranée et sur les côtes atlantiques d’Espagne". Mammalia, 41 (2): pp. 197-206. 1977 a.VIALE D. 1977 b. Ecologie des cétacés en Méditerranée Nord Occidentale, leur place dans l’écosystème ; leur réaction à laVIALE D. "Une méthode synoptique de recherche des zones productives en mer: détection simultanée des cétacés, des fronts thermiques et des biomassessousjacentes". Ann. Inst. Oceanogr. Paris. 67 (1): pp. 49-62. 1991.WILSON, B., Hammond, P.S., and Thompson, P.M. "Estimating size and assessing trends in a coastal bottlenose dolphin population". Ecol. Appl. 9: pp. 288-300. 1999.WÜRSIG, B., and T. A. Jeffersen. "Methods of photo-identification for small cetaceans". Reports of the International Whaling Commission: pp. 43-52. 1990.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 147/276
Comunicaciones
147
INTRODUCCIÓN
El Estrecho de Gibraltar es la única conexión natural existente entre el mar Mediterráneo y el océano Atlántico (figuras1 y 2). La circulación de las masas de agua en el Estrecho está caracterizada por un sistema de corrientes este-oeste. El mar
Mediterráneo presenta un balance hídrico negativo debido a que las elevadas pérdidas por evaporación exceden a lasganancias debidas a precipitaciones y pocos aportes fluviales. El flujo de corrientes será por tanto el responsable de queexista un equilibrio hídrico-salino en las cuencas mediterráneas y está compuesto de:
- Un flujo superficial de aguas atlánticas entrando al Mediterráneo.
- Una contracorriente profunda de aguas mediterráneas saliendo hacia el Atlántico, cada una con salinidades y caudalesdiferentes (Lacombe and Richez 1982).
La interfase entre las aguas superficiales atlánticas y las aguas profundas mediterráneas, generalmente se sitúa aprofundidades comprendidas entre 50 y 200 metros, dependiendo de la localización y los flujos de marea. A medida quenos acercamos a la costa marroquí la frontera entre las aguas atlánticas y mediterráneas es cada vez más profunda (Garrett
1996, Reul et al. 2002). Está situada aproximadamente a 100 metros en 5°20 W y baja a 300 metros a 6
°W en la parte centraldel Estrecho. (Kinder et al. 1988). La mayor parte de la biomasa de plancton es transportada al mar Mediterráneo por las
aguas atlánticas. Las concentraciones más altas eran observadas en la parte norte del Estrecho, donde circulan aguasatlánticas superficiales enriquecidas (Van Geen y Boyle 1988) pero debido a las altas velocidades de las corrientes, la mayorparte de la importación de biomasa tenía lugar en la parte central y sur del Estrecho.
La distribución de cetáceos y su abundancia en el estrecho de Gibraltar están pobremente descritas y se limitan a unas pocasfuentes. Un estudio llevado a cabo desde los ferries que unían España y Ceuta, desde marzo a mayo de 1999, demostró quela mayor parte de las especies encontradas en el sector este del Estrecho eran delfines listados (Stenella coeruleoalba),delfines comunes ( Delphinus delphis), y, ocasionalmente, delfines mulares (Tursiops truncatus), calderones comunes(Globicephala melas) y cachalotes (Roussel 1999).
DISTRIBUCIÓN ESPACIAL DE CETÁCEOSEN EL ESTRECHO DE GIBRALTAR
DURANTE LA ÉPOCA ESTIVALSergi Pérez / Renaud De Stephanis / Philippe Verborgh / Alicia Sánchez / Christophe Guinet

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 148/276
Almoraima, 35, 2007
148
Estudios sobre la ecología de depredadores superiores en el mar, indican que su distribución y abundancia está a menudorelacionada con procesos oceanográficos y productividad marina.
Los objetivos de este estudio son: 1) Estimar la abundancia relativa de las especies de cetáceos presentes en el estrecho deGibraltar. 2) Investigar cómo sus distribuciones pueden ser relacionadas con los parámetros batimétricos del estrecho deGibraltar. 3) Examinar las asociaciones o segregaciones espaciales interespecíficas en el estrecho de Gibraltar en época estival.
METODOLOGÍA
Zona de estudio y muestreos
La zona de estudio es el área del estrecho de Gibraltar, comprendida entre los 5º y 6º de longitud Oeste. El estrecho deGibraltar (figura 1) tiene aproximadamente 60 Km de largo. Su frontera oeste está localizada entre el cabo de Trafalgar enEspaña y el cabo Espartel en Marruecos, separado por 44 Km. Su frontera este está localizada entre Punta Europa (Gibraltar)y Punta Almina (África) y están separadas por 23 Km (Parrilla et al., 1988). La batimetría del Estrecho se caracteriza porser un cañón que va de este a oeste con aguas poco profundas (200-300 m) que se pueden encontrar en la cara atlántica yaguas más profundas (800-1000 m) en la cara mediterránea (figura 1).
Trayectos aleatorios fueron llevados a cabo desde el barco de investigación Elsa durante los meses de julio, agosto yseptiembre 2001 y 2002. La embarcación Elsa es una motora de 11 metros de eslora, 2,80 metros de manga, y que tieneuna plataforma que permite posicionar observadores a 4 metros sobre el nivel del mar. La zona de estudio fue muestreadaa una velocidad media de 5,3 nudos.
Los observadores eran situados en una plataforma de observación a 4 metros sobre el nivel del mar. Dos observadoresentrenados ocuparon el puesto de observación en turnos de una hora, durante la luz del día, cuando la visibilidad superabalas 3 nm (5,6 Km) y provistos de unos prismáticos 8 x 50, cubriendo 180º por delante del barco. El esfuerzo de búsquedafue medido como el número de kilómetros navegados con condiciones de avistamiento adecuadas (con un estado de marmenor de 4 en la escala Douglas y con dos observadores en el puesto de observación).
La posición geográfica de la embarcación era grabada continuamente en un ordenador portátil desde un GPS y a través deun programa informático (IFAW Data Logging Stware Logger 2000 versión 2.20) provisto por la International Fund ForAnimal Walfare). Datos relativos a la hora, especie, número de individuos, comportamiento y otros datos relevantes durantelos avistamientos fueron grabados con otros datos ambientales relevantes. Un avistamiento fue definido como un grupode animales de la misma especie observados a la misma hora que llevan un comportamiento similar y que estaban a menos
de 1.000 metros de distancia entre ellos (SEC 1999).El área de estudio fue dividida en cuadrículas con una resolución de dos minutos de latitud por dos minutos de longitud.La distancia en kilómetros de búsqueda en cada cuadrícula fue entonces calculada utilizando un sistema de informacióngeográfico: Arc-view 3.2 de ESRI. Tan sólo fueron utilizados para los análisis, las cuadrículas de los trayectos cubiertoscon esfuerzo de al menos 3 Km en total.
Presencia de cetáceos y abundancia
Para poder comparar los datos obtenidos en la zona de estudio con otras zonas de estudio dos parámetros fueron definidos:
- El Encounter Rate (ER), es el número de avistamientos de una especie dada por cada 100 kilometros navegados y es definido:
ER = (Sigh / Eff) x 100

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 149/276
Comunicaciones
149
Sigh: número de avistamientos realizados de una especie en esfuerzo de búsqueda en la zona de estudio.Eff: Distancia (Km) cubierta con esfuerzo de búsqueda.
Sigh incluye los avistamientos de cetáceos en donde los animales fueron contactados y los avistamientos de los cetáceosen donde los animales no fueron contactados.
- El Abundance Index Rate (AI) (ind/Km) también fue definido como:
AI = (Ind / Eff) x 100
Ind: número de individuos de una especie dada, observados con esfuerzo de búsqueda en la zona de estudio.
Eff: Distancia (Km) cubierta con esfuerzo de búsqueda.
Ind incluye los avistamientos de cetáceos en donde los animales fueron contactados y los avistamientos de los cetáceosen donde los animales no fueron contactados
Distribución espacial y batimetría
El Encounter Rate para cada especie fue calculado para cada cuadrícula como número de avistamientos por especies y porkilometro buscado. Sólo los avistamientos para los cuales el contacto fue establecido (de la que se tiene posición precisa)fueron usados para este análisis.
Figura 1. Área de estudio.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 150/276
Almoraima, 35, 2007
150
Dos parámetros batimétricos fueron analizados: la profundidad y la pendiente. La profundidad media y la pendiente fueroncalculadas para cada cuadrícula. La profundidad media fue calculada como la media entre la profundidad máxima y laprofundidad mínima de cada cuadrícula. La pendiente fue calculada como:
Pendiente (m/Km.) = (Dmax - Dmin) / DI
Dmax es la profundidad máxima en la cuadrícula
Dmin
es la profundidad mínima en la cuadrícula
DI es la distancia (Km) entre los puntos de profundidad máxima y mínima de la cuadrícula.
Para determinar si las diferentes especies de cetáceos estaban distribuidas no uniformemente respecto a la profundidad, lapendiente, la latitud y la longitud (positiva hacia el este del meridiano 0º de Greenwich y negativa al oeste), se aplicó un
modelo lineal generalizado (GLM), usando el Software R (Ihaka y Gentleman 1996), para analizar las relaciones entre elER de cada especie de cetáceo para cada cuadrícula en relación con la profundidad, la pendiente y la posición (latitud ylongitud en el medio) de la cuadrícula. Dada la naturaleza de la variable respuesta, con valores de “0” para el ER ennumerosas cuadrículas, los datos fueron transformados usando una distribución de quasi Poisson para evitar sobredispersión.
Uso del hábitat y superposición de áreas de campeo
Para analizar el uso del hábitat por parte de las diferentes especies de cetáceos, se observó el uso de toda la zona de estudiorealizado por cada especie, y fue definido como el número de cuadrículas en donde una especie dada fue observada. Paracada especie se calculó la profundidad media de su hábitat, como la media de las profundidades de cada cuadrícula visitadapor esta especie. El grado de superposición entre dos especies fue definido, como el número de cuadrículas utilizadas por
Figura 2. Distribution of the observation effort in km spent by quadrate over the study area and during the study period.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 151/276
Comunicaciones
151
las dos especies divididas por el número total de cuadrículasusadas por cada especie y expresadas en porcentaje. Paraanalizar más precisamente el entrecruzamiento en el uso delhábitat entre dos especies también se tomó en consideración elER (expresado en porcentaje) para cada cuadrícula por cadaespecie. El sobrecruzamiento fue entonces calculado como lasuma del mínimo ER común para una cuadrícula dada visitadapor las dos especies.
RESULTADOS
Esfuerzo de búsqueda en la zona de estudio
Un total de 4.202 Km de transectos fueron realizados en la zonade estudio. De éstos, 3.396 Km fueron realizados con esfuerzode búsqueda adecuado, y 3.356 Km. de trayectos con esfuerzode búsqueda adecuado fueron retenidos en la zona de estudiodurante los meses de julio, agosto y septiembre 2001 y 2002.Representaron 120 cuadrículas (con suficiente esfuerzo deobservación) lo que representa un área de 1.340 km2 (figura 2).
Presencia de cetáceos y abundancia relativa
Se realizaron 455 avistamientos en la zona de estudio. De estos,399 fueron realizados con un esfuerzo de observación apropiado,y fueron por tanto usados para el cálculo de ER y AI, y en 345 deellos los animales fueron contactados (tabla 1).
Se observaron siete especies de cetáceos. En términos de avistamientos, las especies más comúnmente observadas fueron eldelfín común y el cachalote. Las menos frecuentemente observadas fueron el rorcual común con sólo un avistamiento y la orca(Orcinus orca) (tabla 1). Si se toma en cuenta el número de individuos medio en un grupo, la especie más abundante era eldelfín listado y el delfín común, mientras que las especies menos observadas fueron el rorcual común y el cachalote (tabla 1).
Especie Nº Avistamientos Avistamientos Encounter Rate Abundancecon contacto Index
Delphinus delphis 92 87 2,74 101,49Stenella coeruleoalba 85 78 2,53 273,49Tursiops truncatus 35 31 1,04 21,43Globicephala melas 62 59 1,85 66,94Physeter macrocephalus 91 60 2,71 3,53Orcinus orca 21 19 0,63 5,69Balaenoptera physalus 1 1 0,03 0,03No id 12 4 0,36 -TOTALES 399 339 11,89 472,60
Tabla 1. Número de avistamientos, ER y AI, calculado en relación al esfuerzo de observación (3.396 km) en la zona de estudio durante el verano.
Figure 3. Means and standard deviation for the number of individuals inthe groups and the depth for common, striped and bottlenose dolphins,long-finned pilot whales, sperm whales and killer whales over the studyarea and during the study period.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 152/276
Almoraima, 35, 2007
152
Distribución en relación con parámetros batimétricos
Debido a su bajo ER, los rorcuales comunes fueron retirados de estos análisis. La distribución espacial de ER para las seis
especies más comúnmente observadas se representa en la figura 4. La profundidad media del área frecuentada por cadaespecie de cetáceos se representa en la figura 3. El análisis de la distribución espacial de las especies de cetáceos a travésdel Estrecho se resume en la tabla 2.
Los delfines comunes fueron observados en el 37,5% de las cuadrículas muestreadas. La distribución del delfín común fueasociada positivamente a la pendiente y a la latitud. Los delfines listados fueron observados en el 37,5% de las cuadrículasmuestreadas. La distribución del delfín listado fue explicada positivamente por la profundidad y por la pendiente. Delfinesmulares, calderones comunes y cachalotes fueron encontrados respectivamente en el 8,3%, 13,3% y 9,2% de las cuadrículasmuestreadas. La distribución de estas tres especies fue explicada positivamente por la profundidad y negativamente porla latitud. Las orcas se encuentran en el 6,5% de las cuadrículas muestreadas. La distribución de esta especie fue explicadanegativamente por la profundidad y la longitud.
Especies Valores Esfuerzo Profundidad Pendiente Lat. Long. Null Residual r_(en%)deviance deviance
Delphinus Estimate 9.87 * 10-6 - 7.70 * 10-3 4.66 - 208.12 156.84 24.6delphis t value 3.20 1.231 3.28 2.67 0.50
P <0.001 ns < 0.001 0.009 nsStenella Estimate 1.71 * 10-5 1.97 * 10-3 6.01 * 10-3 - - 187.44 118.47 36.8coeruleoalba t value 6.52 3.35 2.65 0.75 -1.31
P <0.001 <0.001 0.009 ns nsTursiops Estimate 3.21* 10-5 4.28 * 10-3 - -12.82 - 173.41 92.421 46.7truncatus t value 5.59 2.58 0.07 -1.97 0.14
P <0.001 0.011 Ns 0.050 nsGlobicephala Estimate 3.89 * 10-5 5.38 * 10-3 - -22.63 - 288.58 71.814 75.1
melas t value 12.89 5.60 -0.20 -6.17 -1.55P <0.001 <0.001 Ns <0.001 nsPhyseter Estimate 3.53 * 10-5 5.73 * 10-3 - -15.39 - 340.53 153.17 55.0macrocephalus t value 7.197 3.81 -1.67 -2.62 0.00
P <0.001 <0.001 Ns 0.009 NsOrcinus orca Estimate 6.90 * 10-5 -9.16 * 109-3 - -68.42 12.43 129.37 27.324 78.9
t value 8.22 -3.84 0.354 -5.89 -5.88P <0.001 <0.001 Ns <0.001 <0.001
Tabla 2. GLM de la distribución de ER de seis especies de odontocetos comúnmente encontrados en el estrecho de Gibraltar, en relación a la profundidad, pendiente,el esfuerzo de observación y la situación geográfica.
Especies Delphinus Stenella Tursiops Globicephala Physeter Orcinus delphis coeruleoalba truncatus melas macrocephalus orca
Delphinus delphis 100.00 34.56 1.67 4.35 2.22 3.23Stenella coeruleoalba 66.67 100.00 6.05 10.19 6.80 3.42Tursiops truncatus 13.33 15.56 100.00 53.50 59.29 5.07Globicephala melas 22.22 24.44 50.00 100.00 47.38 14.28Physeter macrocephalus 15.56 17.78 63.63 56.25 100.00 3.18Orcinus orca 11.11 8.89 20.00 25.00 9.09 100.00
Tabla 3. Entrecruzamiento de las áreas de campeo calculado con el porcentaje de ocurrencia (en negrita) en cada cuadrícula muestreada, y en función del porcentajede ER (en cursiva) de cada cuadrícula visitada por las seis especies de odontocetos más comunes, encontradas en el estrecho de Gibraltar.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 153/276
Comunicaciones
153
Figure 4. Distribution of encounter rates of common, striped and bottlenose dolphins, long-finned pilot whales, sperm whales and killer whales over the studyarea and during the study period.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 154/276
Almoraima, 35, 2007
154
Asociación y segregación espacial entre especies de cetáceos
Los datos se resumen en la tabla 3. Tres grupos de cetáceos pueden ser diferenciados:
- Delfines comunes y delfines listados tendían a estar asociados espacialmente entre ellos. El entrecruzamiento entredelfines comunes y, en menor medida, entre listados y otras especies de cetáceos era limitado, lo que era mucho másobvio cuando se considera el entrecruzamiento espacial de los ER por cuadrícula.
- Delfines mulares, calderones comunes y cachalotes comparten una gran parte de su hábitat en el Estrecho.
- El menor entrecruzamiento con otras especies de cetáceos fue observado con orcas.
DISCUSION
El estrecho de Gibraltar se caracteriza por una alta diversidad de cetáceos con siete especies observadas durante el periodode este estudio. Se podría sugerir que esta alta diversidad de cetáceos observada en el estrecho de Gibraltar podría estarrelacionada con un alto número de cetáceos transitando dentro y fuera del Mediterráneo a través del Estrecho. Sin embargo,trabajos de foto-identificación (Bigg et al. 1990, Brain 1990, Defran et al. 1990, Hansen y Defran 1990) a largo plazoindican que los individuos de cachalotes, calderones comunes, delfines mulares, orcas y delfines comunes son al menosestacionalmente residentes en el Estrecho (de Stephanis, datos no publicados) mientras que el estatus del delfín listadonecesita ser precisado. La hipótesis más probable para explicar esta alta densidad de cetáceos en el Estrecho, debe ser ladisponibilidad de presas a lo largo del mismo.
Este estudio indica que los parámetros batimétricos, así como la localización en el Estrecho juega un papel significante enla distribución de seis especies de cetáceos. Las variables con una mayor influencia en la distribución de cetáceos en elEstrecho fueron la profundidad, y en menor medida la pendiente. Tres grupos distintos de especies fueron identificadosen función de su distribución a través del Estrecho. Delfines comunes y delfines listados tienen una distribución bastanteamplia, y se sitúan sobre todo en la parte norte de la zona de estudio, concentrados sobre todo en las zonas de aguasprofundas y a lo largo del borde norte del canal septemtrional del Estrecho (figura 2). Los delfines mulares, calderonescomunes y cachalotes comparten una gran parte de su zona de alimentación y estas tres especies fueron encontradas sobretodo en aguas profundas en el canal principal del Estrecho. El tercer grupo es el de las orcas y fueron más observadas enla parte oeste de la zona de estudio. Estas diferencias en las distribuciones espaciales de estos tres grupos podrían estarrelacionadas con sus respectivas estrategias de alimentación, y por el hecho de que se alimentan en diferentes masas de agua.De acuerdo con otros estudios, tanto delfines comunes como delfines listados parecen ser especies bastantes oportunistas,
(Klinowska 1991, Young and Cockcrdet 1994, Gannier 1995), cazando sobre todo pequeños peces y cefalópodos neríticos.Los pequeños cefalópodos y myctophidos mesopelágicos, que componen la dieta de estos dos delfines parecen ser másimportantes en listados, (Blanco et al. 1995, Kenney et al. 1995, Santos et al. 1996, Pauly et al. 1998) que en delfinescomunes (Young and Cockcrdet 1994, Kenney et al. 1995, Cordeiro 1996, Santos et al. 1996) y estas diferencias podríancontribuir y explicar la mayor afinidad por aguas profundas de delfines listados comparado con delfines comunes. Estudiossobre el comportamiento de inmersión llevados a cabo en delfines comunes y en dos especies de stenellas demostraron queestas especies no suelen realizar inmersiones de más de 150 metros de profundidad. (Evans 1974, Scott et al. 1993, 1995,Davis et al. 1996). Esto sugiere que tanto delfines comunes como delfines listados restringirían la mayoría, o casi todassus actividades de alimentación a las aguas superficiales atlánticas. Además la interfase de aguas atlántico-mediterráneaspodría actuar como una frontera para sus presas (peces y cefalópodos) y esta interfase se sitúa a menor profundidad en laparte norte del Estrecho, lo que podría aumentar el éxito en sus estrategias de alimentación.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 155/276
Comunicaciones
155
Respecto al segundo grupo de cetáceos, dos especies, el calderón común y el cachalote, son considerados como cazadoresde calamares y realizan inmersiones a grandes profundidades. Por ello, la distribución espacial en verano de estas dosespecies a lo largo del Estrecho, podría ser indicativa de una distribución de grandes especies de calamares. Sin embargo,no queda claro por qué el delfín mular, cuya dieta se basa sobre todo en peces (Gunter 1942, Tomilin 1957, Evans 1980,Barros y Odell 1990, Gannier 1995) tiende a estar espacialmente asociado con cachalotes y calderones comunes. Elcontenido estomacal de tres cachalotes capturados durante las operaciones balleneras en 1925 en el estrecho de Gibraltarcontenía sobre todo restos de calamar y algunos restos de elasmobranquios bentónicos (Cabrera 1925). Varios estudiossobre cachalotes han demostrado que suelen realizar inmersiones regularmente a profundidades mayores de 1.000 metros(Watkins et al. 1993) y por lo tanto son capaces de llegar al fondo marino en toda la zona de estudio. Los calderones comunestambién suelen realizar inmersiones profundas llegando a profundidades generalmente comprendidas entre 200 y 600
metros, con una profundidad máxima de 828 m (Heide-Jøgersen et al. 2002, Baird et al. 2002). Los calderones comunesse alimentan básicamente de calamares neríticos y oceánicos y en una mayor medida de peces que suelen ser comunes aprofundidades comprendidas entre los 100 y los 1.000 metros. La distribución espacial en verano de calderones comunesy cachalotes indica que ambas especies se alimentarían de calamares y, posiblemente, de peces, asociados con el flujoprofundo de aguas mediterráneas. Es interesante el ER de cachalotes es el más elevado justo al oeste de la cresta centralque separa el canal norte del canal principal. El otro área de mayor ER para cachalotes se encuentra al este de la crestaKamara. Esto sugiere que los cachalotes podrían alimentarse en áreas donde se producen contracorrientes profundas(inducidas por la topografía marina), y que podría por tanto actuar en la distribución de las presas. La concentración decalderones comunes durante el día sugiere que sus presas se concentran principalmente en las aguas del canal principal ya la entrada del cañón de Ceuta. Los delfines mulares presentan una distribución similar a la de los cachalotes. Sin embargo,como sus habilidades de inmersión son limitadas, deberían de restringir sus procesos de alimentación a las aguas atlánticas
superficiales. Los delfines mulares se alimentan de forma oportunista y se considera que tienen una dieta basadaprincipalmente en presas demersales (Barros y Odell 1990, Gannier 1995), lo que es improbable en este estudio en relacióncon su distribución en las aguas muy profundas del estrecho de Gibraltar, donde una alimentación pelágica parece ser laúnica estrategia de alimentación posible. Es interesante que prácticamente no existe entrecruzamiento entre la distribucióndel delfín mular y del delfín común, lo que sugiere algún mecanismo de exclusión entre estas dos especies. Los delfinesmulares suelen matar marsopas en Escocia (Ross and Wilson 1996, Patterson et al. 1998) y los delfines comunes podríanevitar las áreas donde los delfines mulares se encuentran.
La distribución de orcas en el Estrecho es la más diferente comparada con la del resto de las especies de cetáceos que ocupanel estrecho de Gibraltar. Las orcas suelen interactuar con las pesquerías de palangre de atún rojo ( Thunnus thynnus) (deStephanis et al. 2001), que migran fuera del Mediterráneo después de completar la puesta. La distribución espacial en
verano de las orcas está estrechamente asociada a la localización geográfica de esta pesquería, que se concentra al este dela cresta Kamara para la flota marroquí, y en el paso entre monte Seco y monte Tartesos para la flota española.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 156/276
Almoraima, 35, 2007
156
BIBLIOGRAFÍAAGUILAR A., Lens S., "Preliminary report on spanish whaling activities". Rep Int Whal Commn 31: pp. 639-643. 1981.AGUIAR DOS SANTOS R., Haimvici M., "Cephalopods in the diet of marine mammals stranded or incidentally caught along southeastern and soutern Brazil (21-34ºS)". Fish Res 52: pp. 99-112. 2001.ALLUÉ C, Lloris D, Rucabado J, Guerra A, Morales E., "Fichas de identificación de especies, Atlántico Oriental. Estrecho de Gibraltar-Cabo Verde. II ClCephalopoda. Datos Inf de Inv Pesq nº3. 1977.ALONCLE H., "Premières observations sur les petits Cétacés des côtes marocaines". Bull Inst Pech Maroc 12: pp. 21-42. 1964.BAIRD R.W., Borsani J.F., Hanson M.B., Tyack P., "Diving and night-time diving behaviour of long-finned pilot whales in the ligurian sea". Mar Ecol Prog Ser 237:pp. 301-305. 2002.BARROS N.B., Odell D.K., "Food habits of bottlenose dolphins in the Southeastern United States". In: Leatherwood S , Reeves RR (eds) The bottlenose dolphin.Academic Press, Inc San Diego, pp. 309-328. 1990.BAUMGARTNER M.F., "The distribution of Risso’s dolphin (Grampus griseus) with respect to the physiography of the northern Gulf of Mexico". Mar Mamm Sci13(4): pp. 614-638. 1997.BAYED A., Beaubrun P.C., "Les mammifères marins du Maroc: Inventaire préliminaire". Mammalia 51(3): pp. 437-446. 1987.
BEARZI, G., Reeves, R.R., Notarbartolo di Sciara, G., Politi, E., Cañadas, A., Frantzis, A. & Mussi, B. "Ecology, status and conservation of short-beaked commondolphins ( Delphinus delphis ) in the Mediterranean Sea". Mammal Review 33(3): pp. 224-252. 2003.BIANCHI G., Carpenter K.E., Roux J.P., Molloy F.J., Boyer D., Boyer H.J. Field guide to the living marine resources of Namibia. FAO species identification guide
for fishery purposes . FAO, Rome: pp. 265. 1999.BIGG M.A., Olesiuk P.F., Ellis G.M., Ford J.F.K., Balcomb K.C. "Social organization and genealogy of resident killer whales (Orcinus orca) in the coastal watersof British Columbia and Washington State". In: Hammond P., Mizroch S., Donovan G. (eds) Individual recognition of cetaceans: Use of Photo-Identification and Other Techniques to Estimate Population Parameters, Vol Special Issue 12. International Whaling Commission, Cambridge, pp. 383-406. 1990.BJØRKE H., Gjøsæter H. Who eats the larger Gonatus fabricii (Lichtenstein) in the Norwegian Sea? International Council for the Exploration of the Sea, CM/M:10. 1998.BLANCO C., Aznar J., Raga J.A. "Cephalopods in the diet of the striped dolphin Stenella coeruleoalba from the western Mediterranean during an epizootic in 1990".
J Zool Lond 237: pp. 151-158. 1995.BOYLE P.R. Cephalopod life cycles. Academic Press, London. pp. 475. 1983.BRAIN D. "Examining the validity of inferences drawn from photo-identification data, with special reference to studies of the killer whale (Orcinus orca) in BritishColumbia". In: Hammond P, Mizroch S, Donovan G (eds) Individual recognition of cetaceans: Use of Photo-Identification and Other Techniques to EstimatePopulation Parameters, Vol Special Issue 12. International Whaling Commission, Cambridge, pp. 93-100. 1990.CABRERA A. Los grandes cetáceos del estrecho de Gibraltar, su pesca y explotación . Trabajos del Museo Nacional de Ciencias Naturales. Serie Zoologica N° 52 :
pp. 64. 1925.CANO N., García Lafuente .J, Hernández Guerra A., Blanco J.M., Escánez J. "Hidrología del Mar de Alborán en Julio de 1993". Publicaciones Especiales InstitutoEspañol de Oceanografía 24: pp. 9-26. 1997.CAÑADAS A., Sagarminaga R., García-Tiscar S. "Cetacean distribution related with depth and slope in the Mediterranean waters off southern Spain". Deep Sea ResPart I 49(11): pp. 2053-2073. 2002.CLARKE M.R. "Cephalopod biomass-Estimation from predation". Memoirs of the National Museum of Victoria: Proceedings of the Workshop on the Biology and
Resource Potential of Cephalopods , Melbourne, Australia, 9-13 March, 1981. 1983.CLARKE M., Goodall N. "Cephalopods in the diets of three odontocete cetacean species stranded at Tierra del Fuego, Globicephala melaena (Traill, 1809),
Hyperoodon planifrons (Flower, 1882) and Cephalorhynchus commersonii (Lacepede, 1804)". Antarct Sci 6(2): pp. 149-154. 1994.CLARKE M.R., Santos M.B., Pierce G.J. "The Importance of Cephalopods in the Diets of Marine Mammals and Other Top Predators". International Council for theExploration of the Sea: The impact of Cephalopods in the Food Chain and Their Interaction with the Environment , CM/M:8. 1998.CORDEIRO M. Contribuição para o conhecimento do regime alimentar e da biologia da reprodução do golfinho-comum,Delphinus delphis Linnaeus , 1758, na costaPortuguesa. Master thesis. Depart de Zoologia e Antropologia. Facultad de Ciências da Universidade de Lisboa. 1996.DAVIS R.W., Worthy G.A.J., Wursig B., Lynn S.K., Townsend F.I. "Diving behavior and at sea movements of an Atlantic spotted dolphin in the Gulf of Mexico".
Mar Mamm Sci 12: pp. 569-581. 1996.
DAVIS R.W., Fargion G.S., May N., Leming T.D., Baumgartner M., Evans W.E., Hansen L.J., Mullin K., "Physical habitat of cetacean along the continental slopein the north-central and western Gulf of Mexico". Mar Mamm Sci 14(3): pp. 490-507. 1998.DE STEPHANIS R., Perez Gimeno N., Salazar Sierra J., Poncelet E., Guinet C. "Interactions between killer whales (Orcinus orca) and red tuna (Thunnus thynnus)fishery in the Strait of Gibraltar", Proceedings of the the Fourth International Orca Symposium and Workshop, Chize, France, pp. 138-142. 2002.DEFRAN R.H., Schultz G.M., Weller D.W. "A technique for the photografic identification and cataloging in dorsal fins of the bottlenose dolphin ( Tursiopstruncatus)". In: Hammond P., Mizroch S., Donovan G. (eds) Individual recognition of cetaceans: Use of Photo-Identification and Other Techniques to EstimatePopulation Parameters, Vol Special Issue 12. International Whaling Commission, Cambridge, pp. 53-56. 1990.DEGNER E. "Cephalopoda". Rep “Dana” Exp 2: pp. 1-94. 1925.DESPORTES G., Mouritsen R. "Preliminary results on the diet of long-finned pilot whales off the Faroe Islands". Rep Int Whal Commn. Special Issue n°14, pp. 305-324. 1993.DIETRICH G., Kalle K., Krauss G., Siedler G. Allgemeine Meerkeskunde-Einführung in die Ozeanographie . Eds Borntraeger, Berlin, Stuttgart, Pp2. 1975.ECHEVARRÍA F., García Lafuente J., Bruno M., Gorsky G., Goutx M., González N., García C.M., Gómez F., Vargas J.M., Picheral M., Striby L., Varela M., AlonsoJ.J., Reul A., Cózar A., Prieto L., Sarhan T., Plaza F., Jiménez-Gómez F., "Physical-biological coupling in the Strait of Gibraltar". Deep-Sea Res Part II 49: pp. 4115-4130. 2002.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 157/276
Comunicaciones
157
EVANS W.E. "Radio-telemetric studies of two species of small odontocete cetaceans". In: Schevill WE (ed) The whale problem. Harvard University Press, Cambridge,Mass, pp. 385-394. 1974.EVANS P.G.H. "Cetaceans in British waters". Mammal Review 10: pp. 1-52. 1980.FAO, (1987). Fiches FAO d’identification des éspèces pour les besoins de la pêche. Mediterranée et Mer Noire. Zone de Peche 37. Révision 1. Vol. II: Vertebres,FAO, Rome.GANNIER A. Les cétacés de Méditerranée nord-occidentale: estimation de leur abondance et mise en relation de la variation saisonnière de leur distribution avecl’écologie du milieu. Master thesis. Ecole Pratique des Hautes Etudes, Montpellier, France. 1995.GANNON D., Read A., Craddock J., Fristrup K., Nicoles J. "Feeding ecology of long-finned pilot whales Globicephala melas in the western North Atlantic". Mar Ecol Prog Ser 148: pp. 1-10. 1997.GARRETT C. "The role of the Strait of Gibralatar in the evolution of the Mediterranean water, properties and circulation". Bull Inst Oceanogr de Monaco , N° spécial17: pp. 1-18. 1996.GIL de Sola L. Las pesquerías demersales del mar de Alborán (sur-mediterráneo ibérico),evolución en los últimos decenios . Inf Tec Inst Esp Oceanogr 142, pp. 179.GONZÁLEZ M., Fernández-Casado M., Rodriguez M.P., Segura A., Martin J.J., "First record of the giant squid Architeuthis sp (Architeutidae) in the MediterraneanSea". J Mar Biol Ass U K 80: pp. 745-746. 1993. 2000.
GONZÁLEZ M., Sánchez P. "Cephalopods assemblages caught with the trawling along the Iberian Peninsula Mediterranean coast". In: Abelló P., Bertrand J.A., Gilde Sola L., Papaconstatinou C., Relini G., Soplet A. (eds) Mediterranean Marine Demersal Resources: The MEDITS International Trawl Survey (1994-1999) Sci Mar 66 (suppl.2): pp. 199-208. 2002.GUERRA A. "Cefalópodos capturados en la campaña 'Golfo de Cádiz-81'". Res Exp Cient 10: pp. 17-49. Abril, 1982.GUERRA A. "Fauna Ibérica. Vol. 1. Mollusca. Cephalopoda". In: Ramos M.A. et al. (eds) Fauna Ibérica, Vol.1. Museo Nacional de Ciencias Naturales. ConsejoSuperior de Investigaciones Científicas, Madrid, pp. 327. 1992.GUNTER G. "Contributions to the natural history of the bottlenose dolphin,Tursiops truncatus (Montagu),on the Texas coast,with particular reference to food habits".
Journal of Mammology 23: pp. 267-276. 1942.HANSEN E.S., Defran R.H. "A comparison of photo-identification studies of California coastal bottlenose dolphins". In: Hammond P., Mizroch S., Donovan G. (eds)
Individual recognition of cetaceans: Use of Photo-Identification and Other Techniques to Estimate Population Parameters, Vol Special Issue 12. InternationalWhaling Commission, Cambridge, pp. 101-104. 1990.HEIDE-JØRGENSEN M.P., Bloch D., Stefanson E., Mikkelsen B., Ofstad L.H., Dietz R. "Diving behaviour of long-finned pilot whales Globicephala melas aroundthe Faroe Islands". Wildlife Biology 8: pp. 307-313. 2002.HOOGE P.N., Eichenlaub B. Animal movement extension to Arcview, ver 2.0. Alaska Science Center - Biological Science Office, U.S. Geological Survey,Anchorage, AK, USA (http://www.absc.usgs.gov/glba/gistools/index.htm#ANIMAL MOVEMENT). 2000.
IHAKA R., Gentleman R. "R: a language for data analysis and graphics". Journal of Computational and Graphical Statistics 5: 299-314 (www.creem.st-and.ac.uk). 1996.KENNEY R.D., Scott G.P., Thompson T.J., Winn H.E. "Estimates of prey consumption and trophic impacts of cetaceans in the USA northeast continental shelf ecosystem". J Northwest Atl Fish Soc 22: pp. 155-171. 1995.KINDER T.H., Parrilla G., Bray N.A., Burns D.A. "The hydrographic structure of the strait of Gibraltar". In: Almazan J.L., Bryden H., Kinder T., Parrilla G. (eds)Seminario sobre la oceanografía física del Estrecho de Gibraltar , Madrid, 24-28 October 1988, pp. 55-67. 1988.KLINOWSKA M.M. Dolphins, porpoises and whales of the world. The IUCN Red Data Book . Cambridge, United Kingdom & Gland: IUCN, Switzerland andCambridge, UK, pp. 429. 1991.LACOMBE H., Richez C. "The regime of the Strait of Gibraltar". In: Nihoul JCJ (ed) Hydrodinamics of semi-enclosed seas. Elsevier, Amsterdam, p 13-73. 1982.OKUTANI T. Cuttlefish and squids of the world in color . Publication for the 30th anniversary of the foundation of the National Cooperative Association of SquidProcessors: pp. 185. 1995.PATTERSON I.A.P., Reid R.J., Wilson B., Grellier K., Ross H.M., Thompson P.M. "Evidence for infanticide in bottlenose dolphins: An explanation for violentinteractions with harbour porpoises?" Proc R Soc Lond, B 265(1402): pp. 1167-1170. 1998.PARRILLA G., Kinder T.H., Bray N.A. "Hidrología del agua mediterránea en el Estrecho de Gibraltar durante el Experimento Gibraltar (octubre 1985-octubre 1986)".Seminario sobre la oceanografía física del Estrecho de Gibraltar (Madrid, 24-28 de octubre 1988), pp. 95-121. 1988.PAULY D., Trites A.W., Capuli E., Christensen V. "Diet composition and trophic levels of marine mammals". ICES J Mar Sci 55: pp. 467-481. 1998.
REUL A., Vargas J.M., Jiménez-Gómez F., Echevarría F., Garcia-lafuente J., Rodriguez J. "Exchange of plantonic biomass through the strait of Gibralatar in latesummer conditions". Deep-Sea Res Part II 49: 4131-4144. 2002.ROPER C.F.E., Sweeney M.J., Nauen C.E. Cephalopods of the world . Food and Agriculture Organization, Rome, Italy. Vol. 3: pp 277. 1984.ROSS H.M., Wilson B. "Violent interactions between bottlenose dolphins and harbour porpoises". Proc R Soc London, B 263 (1368): 283-286. 1996.ROUSSEL E. Les cétacés dans la partie orientale du Détroit de Gibraltar au printemps: indications d’écologie. Master thesis. Ecole Pratique des Hautes Etudes,Montpellier, France. 1999.RUBÍN J.P. El ictioplacton y el medio marino en los sectores norte y sur del mar de Alborán, en junio de 1992. Inf Tec Inst Esp Oceanogr 146, pp. 92. 1994.SANPERA C., Aguilar A. "Modern Whaling off the Iberian Peninsula during the 20th Century". Rep Int Whal Commn 42: pp. 723-730. 1992.SANTOS M.B., Pierce G.J., López A., Barreiro A., Guerra A. Diets of small cetaceans stranded NW Spain. International Council for the Exploration of the Sea, C.M.1994/N: 11. 1996.SANTOS M.B., Pierce G.J., Boyle P.R., Reid R.J., Ross H.M., Patterson I.A.P., Kinze C.C., Tougaard S., Lick R., Piatkowski U., Hernández-García V. "Stomachcontents of sperm whales Physeter macrocephalus stranded in the North Sea 1990-1996". Mar Ecol Prog Ser 183: pp. 281-294. 1999.SANTOS M.B., Clarke M.R., Pierce G.J. "Assessing the importance of cephalopods in the diets of marine mammals and other top predators: problems and solutions".Fish Res 52: pp. 121-139. 2001.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 158/276
Almoraima, 35, 2007
158
SANTOS M.B., Pierce G.J., Hartmann M.G., Smeenk C., Addink M.J., Kuiken T., Reid R.J., Patterson I.A.P., Lordan C., Rogan E., Mente E. "Additional notes instomach contents of sperm whales Physeter macrocephalus stranded in the north-east Atlantic". J Mar Biol Ass U K 82: pp. 501-507. 2002.SANZ J.L., Acosta J., Herranz P., Palomo C., San Gil C. Mapa Batimétrico del Estrecho de Girbaltar. Proyecto Hércules 1980-1983, Instituto Español de Oceanografía. 1988.SCOTT M.D., Chivers S.J., Olson R.J., Lindsay R. "Radiotracking of spotted dolphins associated with tuna in the eastern tropical Pacific". Abstracts of the 10 thBiennial Conference on the biology of Marine Mammals, Galveston , Texas, p. 97. Society for Marine Mammalogy, Lawrence, Kansas. 1993.SCOTT M.D., Olson R.J., Chivers S.J., Holland K. "Tracking of spotted dolphins and yellowfin tuna in the eastern tropical Pacific". Abstracts of the 11 th BiennialConference on the biology of Marine Mammals, Orlando, Florida, p. 102. Society for Marine Mammalogy, Lawrence, Kansas. 1995.SEC. Sociedad Española de cetáceos. Recopilación, análisis, valoración y elaboración de protocolos sobre las labores de observación, asistencia a varamientos yrecuperación de mamíferos y tortugas marinas de las aguas españolas. Ministerio de Medio Ambiente Español. Secretaria General de Medio Ambiente, TechnicalReport Sociedad Española de Cetáceos Avaible from SEC, Nalón 16, E-28240 Hoyo de Manzanares, Madrid, Spain. 1999.SILVANI, L., M. Gazo and A Aguilar. "Spanish driftnet fishing and incidental catches in the western Mediterranean". Biological Conservation 90: pp. 79-85. 1999.SMITH S., Whitehead H.Y. "The Diet of Galapagos Sperm whales Physeter macrocephalus as indicated by fecal sample analysis". Mar Mamm Sci 16(2): pp. 315-325. 2000.TOMILIN A.G. "Mammals of the U.S.S.R. and Adjacent Countries". Vol.IX: Heptner VG Ed, Cetacea, Nauk USSR, Moscow. English translation, 1967, IsraelProgram for Scientic Translations, Jerusalem, pp. 523-549. 1957.
VAN GEEN A., Boyle E. "Atlantic water masses in the Strait of Gibraltar : inversión of trace metal data". In: Almazan J.L., Bryden H., Kinder T., Parrilla G. (eds.)Seminario sobre la oceanografía física del estrecho de Gibraltar . Madrid, 24-28 October 1988, pp. 68-80. 1988.VILLANUEVA R., Collins M.A., Sánchez P., Voss N.A. "Systematics, distribution and biology of the cirrate octopods of the genus Opisthoteuthis (Mollusca,Cephalopoda), in the Atlantic Ocean with description of two new species". Bull Mar Sci 71(2): pp. 933-985. 2002.WATKINS W.A., Daher M.A., Fristrup K.M., Howald T.J., Nortobartolo di Sciara G. "Sperm whales tagged with transponders and tracked underwater by sonar".
Mar Mamm Sci 9: pp. 55-67. 1993.YOUNG D.D., Cockcroft V.G. "Diet of common dolphins ( Delphinus delphis ) off the south-east coast of southern Africa: opportunism or specialization?". J Zool
Lond 234: pp. 41-53. 1994.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 159/276
Comunicaciones
159
INTRODUCCIÓN
El estrecho de Gibraltar es la única conexión entre el mar Mediterráneo y el océano Atlántico. La circulación de las masasde agua se caracteriza por el sistema de corrientes oeste-este. El mar Mediterráneo tiene un balance hídrico negativo debidosu alta pérdida a través de la evaporación, que excede las ganancias por las precipitaciones y el aporte de los ríos. Lascorrientes en el estrecho de Gibraltar son por tanto la única regulación para este desequilibrio. Los flujos están formadospor una corriente superficial de aguas atlánticas entrando en el mar Mediterráneo y una corriente profunda de aguasmediterráneas en dirección al Atlántico, cada uno con su correspondiente salinidad (Lacombe and Richez, 1982). La mezclade aguas también se produce a través de los afloramientos, su fuerza dependerá de las mareas y la batimetría. Todo estocrea un ambiente rico en nutrientes, en el cual los cetáceos son abundantes (de Stephanis et al., 2005a).
Hasta la actualidad, la distribución y abundancia de cetáceos en el estrecho de Gibraltar están pobremente descritas y estálimitado a unas pocas referencias. Un estudio que se llevó a cabo desde los ferris que unían la Península con Ceuta, desdemarzo a mayo de 1999, demostró que muchas de las especies encontradas en la parte este del Estrecho eran delfines listados(Stenella coeruleoalba), delfines comunes ( Delphinus delhis), y ocasionalmente delfines mulares (Tursiops truncatus),calderones comunes (Globicephala melas) y cachalotes (Physeter macrocephalus) (Roussel 1999). Silvani et al. (1999)en un estudio de capturas accidentales con redes de deriva en el Mediterráneo occidental demostró la abundante presenciade delfines comunes y listados en el Estrecho, aunque ninguno de estos estudios demostró la densidad relativa, abundanciarelativa o absoluta de cetáceos. El primer estudio sobre densidad relativa y distribución de cetáceos en el estrecho deGibraltar fue llevado a cabo por de Stephanis et al. (2005a), el cual demostró que siete especies de cetáceos pueden serobservadas en verano y seis de ellas regularmente durante todo el año. Estas son el delfín listado, el delfín común, el delfín
ESTIMACIÓN DE LA ABUNDANCIAABSOLUTA DE LAS POBLACIONESRESIDENTES Y SEMIRESIDENTESDE CETÁCEOS ENEL ESTRECHO DE GIBRALTAR
Philippe Verborgh / Renaud De StephanisSergi Pérez / Alicia Sánchez / Christophe Guinet

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 160/276
Almoraima, 35, 2007
160
mular, el calderón común, cachalotes y orcas. El rorcual común es la única especie migratoria observada a través delEstrecho (Salazar Sierra et al. 2004), mientras que las otras especies usan este como zona de alimentación (Laplanche et al. 2004, de Stephanis et al. 2005a y de Stephanis et al. 2005b).
El objetivo de este estudio es obtener una estimación de la población de cetáceos presentes en el estrecho de Gibraltar. Lasestimaciones de poblaciones de calderón común (Globicephala melas) y delfín mular (Tursiops truncatus) se realizaronmediante metodos de captura-recaptura, mientras que las poblaciones de orcas (Orcinus orca) y cachalotes (Physeter macrocephalus) fueron estimadas mediante el número total de indivuduos foto-identificados.
MATERIAL Y MÉTODOS
El área de estudio es el estrecho de Gibraltar y está comprendida entre los 5º y 6º de longitud Oeste, intentando cubrir todala zona hasta las aguas de Marruecos. El estrecho de Gibraltar tiene aproximadamente 60 Km de largo. La frontera oesteesta localizada entre el cabo de Trafalgar en España y cabo Espartel en Marruecos, separados por 44 Km. En la parte este,el Estrecho alcanza su mínima amplitud con 14 Km entre Tarifa y Punta Cires. La frontera este se localiza entre Gibraltary punta Almina (Marruecos) separados por 23 Km (Parrilla et al., 1988). La batimetría del Estrecho se caracteriza por uncañón que va de oeste a este, con aguas más someras (200-300 m) que se pueden encontrar en la zona atlántica y aguas másprofundas (800-1000 m) en la cuenca mediterránea.
Desde 1998, se han tomado fotos de las aletas dorsales de calderón común, delfín mular y orcas y de aletas caudales ydorsales de cachalotes en el estrecho de Gibraltar. Desde 1998 hasta 2000, las fotos se tomaron desde una embarcación deavistamiento de cetáceos comercial con una cámara Nikon equipada con un objetivo de 100-300 mm. En 2001, las fotos
se realizaron desde la embarcación de investigación Elsa, una motora de 10 metros, de la asociación CIRCE (Conservación,Información e Investigación de Cetáceos) con la misma cámara. En 2002 y 2003 se utilizó una Canon EOS-3 con un objetivoCanon EF 100-400 mm con un estabilizador de imagen. Durante 2004, una cámara digital reflex Canon 10D fue usada conel mismo objetivo que en 2003.
Para las especies de delfín mular y calderón común se obtuvo fotografias del lado izquierdo de la aleta dorsal aproximandosetan cerca como fue posible. Generalmente, este lado es el más accesible debido a que los animales nadan la mayoría deltiempo contra la corriente predominante para permanecer en el mismo lugar, situandose el sol al sur, evitando así problemasde contraluz. Todos los individuos durante los avistamientos fueron fotografiados independientemente de su nivel demarcas para tener la misma probabilidad de capturar a todos los animales. El mismo método esta siendo usado desde 1998para que todas las fotos puedan ser analizadas de la misma manera.
Las orcas no nadan frecuentemente en la misma dirección, y el lugar donde salen a la superficie es difícil de predecir. Porlo tanto, ambos lados de la aleta dorsal y la silla de montar, la cual se puede apreciar en la parte trasera de la aleta dorsal,se fotografiaron para maximizar la probabilidad de captura de todos los individuos.
Para los cachalotes la aproximación se realiza lentamente desde detrás para fotografiar las aletas caudales cuando el animalse esta sumergiendo. Se tomaron fotos de la aleta dorsal antes de que el animal se sumergiera, para identificar los individuosque no pueden ser aproximados desde atrás.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 161/276
Comunicaciones
161
Catálogo de Foto-Identificación
Todas las fotos desde 1998 hasta 2003 se revelaron como diapositivas en color mientras que en 2004 todas las fotografiasfueron digitales. Todas las diapositivas fueron revisadas con una lupa de 8 aumentos y una tabla de luz. Cada foto fueanalizada e introducida en una hoja de Excel. Los datos se basan en información general como número de avistamiento,nombre del grupo, número de carrete, número de la foto, número total de animales en la foto, número del individuoanalizado en la foto (puede haber más de un individuo en la foto); y de información de cada individuo con la exposiciónde la aleta (fuera o no del agua), ángulo, calidad del individuo (Q), código del individuo, veracidad de la identificación,proporción de la parte trasera expuesta del individuo y comportamiento.
Un rango de calidad en una escala de 0 a 2 (mala y excelente) es asignado a cada imagen de aleta, basado en las característicasde la imagen: enfoque, tamaño, orientación, exposición y porcentaje de la aleta expuesta en la fotografía:
Q 0: aletas dorsales individuales de forma inusual, cuando está borrosa, demasiado lejos o si el ángulo esta entre 330º y30º o 150º y 210º.
Q 1: fotografías de calidad media donde aparece parte de la aleta dorsal o entera.
Q 2: fotografías de alta calidad con toda la aleta dorsal.
Delfines mulares y calderones comunes fueron identificados a partir de muescas y deformaciones sólo para maximizar lasoportunidades de recaptura, ya que los rasguños y cicatrizes tienden a desaparecer con el tiempo (Wilson et al. 1999).
Cada individuo en el catálogo obtiene un nivel de marca de 0 a 3:
M 0: individuos sin ninguna marca en la aleta dorsal, pero con una forma específica.
M 1: individuos con pequeñas muescas que sólo pueden ser vistas mediante fotografías de alta calidad.
M 2: individuos con un tamaño medio de muescas o muchas de pequeño tamaño.
M 3: individuos con muescas muy características, por ejemplo, con muescas muy profundas o con amputaciones parcialeso totales de la aleta dorsal. Éstas se pueden reconocer mediante fotografías de calidad muy baja. Se ha creado un historialde las muescas de cada individuo para poder hacer un seguimiento de la evolución de estas. Las muescas de la aleta dorsalse conservan durante años, pero pueden evolucionar, por ejemplo si una muesca más grande se hace encima de una vieja.
Las orcas fueron identificadas a partir de muescas o cicatrices en la aleta dorsal y también a partir de cicatrices en la sillade montar (Bigg 1982 and Bigg et al., 1983). Los individuos son identificados principalmente a partir de las muescas desus aletas dorsales lo cual facilita la unión de ambos lados. Cuando la aleta dorsal no tiene ninguna muesca, entonces seusa la forma y el color de la silla de montar, ya que es única en cada individuo (Baird and Stacey, 1988). Por lo tanto, unabuena calidad de foto de la silla de montar es importante para una buena identificación, especialmente en el Estrecho deGibraltar donde la mayoría de los individuos están muy poco marcados. Los nuevos individuos son incluidos en el catálogosi ellos están marcados mediante marcas M2 y M3 y deberán haber sido vistos por los menos dos veces si tienen una calidadde marcas de 0 y 1.
Los cachalotes fueron identificados a partir de marcas y patrones de color blancos en sus aletas (Whitehead and Arnbom 1987).Se utilizaron identificaciones adicionales de marcas y patrones en la aleta dorsal para identificar individuos a los que no seles pudo aproximar desde detrás. Sin embargo el catálogo sólo se realizó a partir de identificaciones de la aleta caudal.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 162/276
Almoraima, 35, 2007
162
Análisis de captura-recaptura para calderones comunes y delfines mulares
La proporción de todo los individuos marcados (M0, M1, M2 y M3) en la población es estimado para poder corregir laestimación realizada por CAPTURE. Se calcula un factor de corrección (c)̂ par cada año:
Ecuación nº 1:c =̂ número de imágenes de aleta de mejor calidad (Q2) de todos los individuos
número de imágenes de aleta de mejor calidad (Q2) de los individuos marcados
Esta estimación asume que de media, se tomaron el mismo número de fotografias de la mejor calidad (Q2) de animales bienmarcados como de animales poco marcados o sin marcas (Ottensmeyer and Whitehead, 2003).
Los modelos de captura-recaptura sólo estiman el tamaño de animales bien marcados en la población ( N ̂). Para obtener eltamaño total de la población se corrigio mediante el factor de corrección (c)̂ (ver Ecuación 1):
Ecuación nº 2: N ̂' = N ̂ x c^
Lo mismo ha sido aplicado para obtener los limites de los intervalos de confidencia (CI) del 95% por el programaCAPTURE:
Ecuación nº 3: L.CI(N ̂') = L.CI(N ̂) x c^
U.CI(N ̂') = U.CI(N ̂) x c^
donde L.CI es el límite inferior del 95% del CI y el UC.I es el límite superior del 95% del C.I.
Los modelos de captura-recaptura para poblaciones cerradas se usaron para estimar el número de calderones comunes ydelfines mulares presentes en el estrecho de Gibraltar. Estos modelos provienen del modelo M
0que se basa en cuatro
hipotesis (Otis et al., 1978; Pollock et al., 1990):- la población está demográficamente y geográficamente cerrada.
- todos los individuos tienen la misma probabilidad de recaptura en cada ocasión de captura.
- todas las marcas son claramente observadas y guardadas en cada captura.
- las marcas no se pierden y todos los individuos en el avstamiento son fotografiados indistintamente de su nivel de marcaje.
Asumimos que la población era aproximadamente cerrada, por ejemplo sin mortalidad, sin nacimientos, emigración einmigración durante el periodo estudiado (meses de verano). A partir del estudio de de Stephanis et al. (2005a) se puedeobservar que la distribución de calderones comunes y delfines mulares está principalmente limitada al canal central delEstrecho. Por lo tanto, su distribución geográfica también está probablemente cerrada. Todas las imágenes de aletas decalidad Q0 no fueron usadas en los análisis para poder disminuir la alta probabilidad de indivduos M3 capturados.
La hipótesis de la misma probabilidad de captura puede ser modulada por tres tipos de variación del modelo M 0:
- variación temporal (t)
- heterogeneidad entre individuos (h)
- respuesta del comportamiento de los individuos a sus primeras capturas (b)
Estos modelos pueden ser utilizados solos o combinados como: M0, Mt, Mh, Mb, Mth, Mtb, Mbh y Mtbh.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 163/276
Comunicaciones
163
El programa MARK tiene integrado el programa CAPTURE que permite la selección de todos los modelos y da un criteriopara la elección del modelo correcto. Estos modelos se clasifican en una escala de 0 a 1, siendo 1 el mejor modelo a utilizar.Aunque estos criterios no son absolutos, se recomienda considerar modelos coherentes a las condiciones de observacióny a la biología de las especies estudiadas (Pollock et al., 1990).
Los modelos se han elegido en función de su compatibilidad con los datos. La respuesta en el comportamiento influyedespués de la primera captura, por ejemplo, el animal se verá atraido, mostrará indiferencia o evitará la embarcación. Elcomportamiento “trap shy” (da bajas probabilidades de captura a partir de marcajes) obtendrá un tamaño de poblaciónsobrestimado, debido a que los animales evitarán la embarcación; el comportamiento trap happy (da altas probabilidadesde captura a partir de marcajes) obtendrá un tamaño de población subestimado (Wilson et al., 1999). Debido a que se usanmarcas existentes para la foto-identificación, no se requiere de ninguna interacción física entre animal e investigador enrelación con el marcaje y por lo tanto la respuesta del comportamiento de este tipo no puede ocurrir. (Wilson et al., 1999).Todos los modelos que tienen en cuenta el comportamiento (M b, Mbh, Mtb y Mtbh) no son considerados.
RESULTADOS
Calderon común
Se analizaron un total de 9.865 individuos de calderón común (en 5.319 fotografias), de los cuales 6.723 eran aletasdorsales, en 107 avistamientos durante 76 dias en el estrecho de Gibraltar. De estas, 3.142 eran de calidad Q0, 1.864 deQ1y 1.717 de Q2. Entre 1999 y 2004, se identificaron un total de 192 individuos en el catálogo con un individuo con marcasM0, 108 con marcas M1, 58 con marcas M2 y 25 con marcas M3.
La tabla 1 muestra como el esfuerzo fotográfico aumenta a lo largo de los años. Las estimaciones de poblaciónes desde 1999a 2001 no han sido consideradas debido a la falta de datos. El modelo de captura-recpatura de Chao M
thda una estimación
de 272 (95% CI: 209-401) individuos de calderón común en 2002, 290 (95% CI: 228-407) en 2003 y 278 (95% CI: 249-328) en 2004.
Delfín mularLa estimación de población ha sido realizada sólo en 2004. Un total de 153 individuos han sido indentificados en el catálogo.El modelo de Darroch M
tda una estimación de 264 (95% CI: 222-334) individuos de delfín mular. Los mismos individuos
han sido recapturados a lo largo de los años y en diferentes épocas del año.
Orcas
Se han analizado un total de 3.834 individuos de orca, en 3.134 fotografías.
La población de orcas está compuesta como mínimo de 25 individuos identificados en el catálogo. Otros ocho individuosfueron identificados pero no se incluyeron en el catálogo ya que estos solo fueron vistos en un avistamiento y no tenian
1999 2000 2001 2002 2003 2004 1999-2004Aletas totales analizadas 163 564 811 1508 1864 1813 6723Aletas totales en Q1y Q2 102 146 327 595 861 1550 3581Número total de individuos identificados 25 22 46 76 93 159 192
Tabla 1. Esfuerzo fotográfico y número total de individuos analizados por año.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 164/276
Almoraima, 35, 2007
164
marcas características en la aleta dorsal o en la silla de montar que permitieran recapturas. Por lo tanto, puede haberalrededor de 33 individuos en el estrecho de Gibraltar, de los cuales nueve han sido identificados en Barbate y cuatro nofueron incluidos en el catálogo en primavera de 2002, 2003 y 2004. Otros 16 individuos identificados residen en la zonadel centro-oeste del Estrecho, además de cuatro individuos no identificados.
Cachalotes
Un total de 548 aletas caudales y dorsales fueron analizadas. Al menos 21 individuos que fueron identificados en el catálogoutilizan el estrecho de Gibraltar como zona de alimentación. Recapturas de los mismos individuos a lo largo de los añosen el Estrecho demuestran su fidelidad al lugar.
DISCUSIÓN
La estima de población de calderón común de 278 individuos (95% CI: 249-328) en 2004 es la más aproximada con el errorestandar más bajo de los diferentes años. Esto es debido a la mejor calidad de las fotos tomadas y al mayor número deavistamientos de calderón común en 2004 (ver tabla 1). Esto causa el aumento del número de recapturas durante el periodode muestreo y por lo tanto la disminución del error estandar.
La primera estima de delfín mular en el estrecho de Gibraltar de 264 (95% CI: 222-334) individuos es interesante, ya que seaproxima a la estima de calderón común. Los mismos calderones comunes y delfines mulares han sido observados en elestrecho de Gibraltar durante diferentes épocas y a lo largo de los años. Esto sugiere que estas especies pueden ser residentesen el estrecho de Gibraltar, o al menos que es una importante zona de alimentación para ellos durante todo el año.
Las mismas orcas han sido observadas desde 1998 en el centro del estrecho de Gibraltar. La estima de 33 individuosidentificados probablemente se aproxima a la realidad. El Estrecho es una zona de alimentación para estas orcas por lomenos en primavera alrededor de Barbate y en verano en la zona centro-oeste del Estrecho.
La actividad de la industria ballenera tuvo lugar entre 1921 y 1954 desde las estaciones balleneras de Getares, en Españay de Benzou, en Marruecos y también desde barcos factoria. Se calculó que 826 cachalotes ( Physeter macrocephalus)fueron capturados en este periodo (Aloncle 1964, Aguilar y Lens 1981, Bayed y Beaubrun 1987, Sanpera y Aguilar 1992).El número de cachalotes identificados (21) comparado al número de animales capturados por los balleneros (826) indicael declive de los stocks. Aunque los balleneros estuvieran capturando ballenas en el mar de Alborán y el Atlántico contiguo,el número de cachalotes era probablemente mayor antes de este periodo. Además, los problemas de colisión con ferris (deStephanis et al 2003) y probablemente con cargueros no ayudan en la recuperación de la población.
Todas estas estimaciones de abundancia absoluta muestran la importancia del Estrecho de Gibraltar para cuatro especiesde cetáceos. Estas estimaciones y análisis de tendencias de población son básicas para la gestión y conservación de laespecie de la mejor manera posible, aunque se necesitan estudios a largo plazo de al menos ocho años, (Wilson et al. 1999)para conocer las tendencias poblacionales ya que se trata de animales longevos.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 165/276
Comunicaciones
165
AGRADECIMIENTOS
Este estudio ha sido financiado por la Ciudad Autónoma de Ceuta, entre 2000 y 2001. Entre 2002 y 2005, el ProyectoLIFE02NT/E/8610 "Conservación de Cetáceos y Tortugas Marinas de Murcia y Andalucía", coordinado por la SociedadEspañola de Cetáceos, financiado en un cincuenta por ciento por la Comisión Europea y que cuenta entre sus socios ycofinanciadores la Consejería de Medio Ambiente de la Junta de Andalucía, la conserjería de Industria y Medio ambientede la Región de Murcia, Ministerio de Medio ambiente (DGB y DGC), la Secretaría General de Pesca Marítima delministerio de Agricultura Pesca y Alimentación, la Universidad de Cádiz, y el Instituto Español de Oceanografía. Así mismo, este estudio ha contado con la donación de 123 voluntarios de CIRCE.
BIBLIOGRAFÍAAGUILAR A., Lens S. "Preliminary report on Spanish whaling activities". Rep Int Whal Commn 31: pp. 639-643. 1981.ALONCLE H. "Premières observations sur les petits Cétacés des côtes marocaines". Bull Inst Pech Maroc 12: pp. 21-42. 1964.BAIRD, R. W., and P. J. Stacey. "Variation in saddle patch pigmentation of killer whales (Orcinus orca) from British Columbia, Alaska, and Washington State".Canadian Journal of Zoology 66: pp. 2.582-2.585. 1988.BAYED A., Beaubrun P.C. "Les mammifères marins du Maroc: Inventaire préliminaire". Mammalia 51(3): pp. 437-446. 1987.BIGG, M. "An assessment of killer whale (Orcinus orca) stocks off Vancouver Island, British Columbia". Reports of the International Whaling Commission 32: pp.655-666. 1982.BIGG M., I. MacAskie, and G. Ellis. "Photo-identification of individual killer whales". Whalewatcher 17: pp. 3-5. 1983.BUISSON L. "Evaluation des effectifs, de la distribution spatiale et de l’organisation sociale de l’orque (Orcinus orca) dans la région du détroit de Gibraltar". DEAThesis, Université de Strasbourg, France. 2004.DE STEPHANIS, R., Salazar-Sierra ,J.M., Pérez-Gimeno, N., Verborgh, P., Tellez, E., Rueda, L. "Collision between a sperm whale ( Physeter macrocephalus) anda ferry in the Strait of Gibraltar". Poster at the ECS Congress in Las Palmas, Canary Islands 2003. 2003.DE STEPHANIS, R., Perez Gimeno, N., Salazar Sierra, J., Fernandez , M. C., and Guinet, C. Summer spatial distribution of cetaceans in the Strait of Gibraltar inrelation to the oceanographic context . Marine Ecology Progress Series, In revision. 2005a.
DE STEPHANIS, R., Guinet, C., Buisson, L., Verborgh, P., Dominici, P. "Population status, social organisation and feeding strategies of killer whales (Orcinus orca)in the Strait of Gibraltar". Poster at the ECS Congress in La Rochelle, France 2005. 2005b.ECHEVARRÍA F., García Lafuente J., Bruno M., Gorsky G., Goutx M., González N., García C.M., Gómez F., Vargas J.M., Picheral M., Striby L., Varela M., Alonso J.J.,Reul A., Cózar A., Prieto L., Sarhan T., Plaza F., Jiménez-Gómez F. "Physical-biological coupling in the Strait of Gibraltar". Deep-Sea Res Part II 49: pp. 4.115-4.130. 2002.FERNÁNDEZ-CASADO M., de Stephanis R., Pérez Gimeno N. and Gozalbes P. "Evidences about site fidelity of the Strait of Gibraltar for sperm whales (Physeter macrocephalus)". Poster at the ECS Congress in Rome, Italy 2001. 2001.LACOMBE H. and Richez C. "The regime of the Strait of Gibraltar". In: Nihoul JCJ (ed) Hydrodynamics of semienclosed seas. Elsevier , Amsterdam, pp. 13-73. 1982.LAPLANCHE, C., De Stephanis, R., Adam, O., Lopakta, M., Demoulin, X., Savanier, O., Rondepierre, F., Pérez Gimeno N., Verborgh P., Guinet, C. and Motsch,J.F. "The Strait of Gibraltar as a feeding ground for sperm whale (Physeter macrocephalus)". Poster at the ECS Congress in Kolmarden, Sweden 2004. 2004.OTIS, D.L., Burnham, K.P., White, G.C. and Anderson, D.R. "Statistical inference from capture data on closed animal populations". Wildlife Monographs 62: pp.1-135. 1978.OTTENSMEYER, C.A. and Whitehead, H. "Behavioural evidence for social units in long-finned pilot whales". Canadian Journal of Zoology 81: pp. 1.327-1.338. 2003.POLLOCK, K.H., Nichols, J.D., Brownie, C. and Hines, J.E. "Statistical inference for capture-recapture experiments". Wildlife Monographs 107: pp. 1-97. 1990.PARRILLA, G., Kinder, T. H., Bray, N. A. "Hidrología del agua mediterránea en el Estrecho de Gibraltar durante el Experimento Gibraltar (octubre 1985-octubre1986)". Seminario sobre la Oceanografía Física del Estrecho de Gibraltar (Madrid, 24-28 de octubre 1988). pp. 95-121. 1988.REUL, A., Vargas, J.M., Jiménez-Gómez, Echevarría, Garcia-lafuente, J., Rodriguez, J. "Exchange of plantonic biomass through the strait of Gibralatar in late summerconditions". Deep-Sea Research.II , 49: pp. 4.131-4.144. 2002.ROUSSEL E. Les cétacés dans la partie orientale du Détroit de Gibraltar au printemps: indications d’écologie. Master thesis. Ecole Pratique des Hautes Etudes,Montpellier, France. 1999.SALAZAR Sierra J., De Stephanis R., Cañadas A., Verborgh P., Perez-Gimeno N., Sagarminaga, R., Guinet C. "Evidences of exchanges of fin whales through theStrait of Gibraltar". Poster at the ECS Congress in Kolmarden, Sweden. 2004.SANPERA C., Aguilar A. "Modern Whaling off the Iberian Peninsula during the 20th Century". Rep Int Whal Commn 42: pp. 723-730. 1992.SILVANI, L., M. Gazo y A. Aguilar. "Spanish driftnet fishing and incidental catches in the western Mediterranean". Biological Conservation 90: pp. 79-85. 1999.WHITEHEAD, H. y T. Arnbom. "Social organization of sperm whales off the Galapagos Islands, February-April 1985". Canadian Journal of Zoology 65: pp. 913-919. 1986.WILSON, B., Hammond, P.S., and Thompson, P.M. "Estimating size and assessing trends in a coastal bottlenose dolphin population". Ecol. Appl. 9: pp. 288-300. 1999.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 166/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 167/276
Comunicaciones
167
INTRODUCCIÓN
El estrecho de Gibraltar es el único canal natural de comunicación entre el mar Mediterráneo y el Atlántico. Se trata de unimportante enclave militar, además la economía de la región depende en gran medida de los recursos pesqueros de susaguas, de la industria y del sector turístico, con una nueva actividad emergente: el avistamiento de cetáceos comercial
(whale watching).
Estudios presentados en este foro, demuestran la presencia de siete especies de cetáceos en la región del estrecho deGibraltar. Cuatro especies, el delfín común ( Delphinus delphis), el delfín listado (Stenella coeruleoalba), el delfín mular(Tursiops truncatus) y el calderón común (Globicephala melas) están presentes en el Estrecho todo el año. La poblaciónde cachalotes (Physeter macrocephalus) está formada por machos subadultos, y está presente en el Estrecho durante losmeses de primavera, la población de orcas (Orcinus orca) está presente en el Estrecho durante los meses de julio y agostoy, finalmente, se producen fenómenos migratorios por parte del rorcual común ( Balaenoptera physalus), entre las aguasmediterráneas y atlánticas. Estas especies están incluidas en el Catálogo Nacional de Especies Amenazadas con lassiguientes categorías:
- Vulnerable: Delphinus delphis, Tursiops truncatus, Physeter macrocephalus, Balaenoptera physalus
- De Interés Especial: Stenella coeruleoalba, Globicephala melas, Orcinus orca
En vista de los diferentes tratados, acuerdos y normativas en el marco de la conservación de la biodiversidad en el mediomarino, tanto a nivel nacional como internacional (ACCOBAMS, Directiva Hábitats, Convención del Mediterráneo yConvenio de Berna), es conveniente profundizar en los factores antropogénicos que condicionan este área.
En el presente trabajo, se proponen cuatro factores como principales causantes de la posible problemática ambiental paralos cetáceos del Estrecho. Esta región es la segunda zona más transitada del mundo en cuanto a tráfico marítimo. Existeuna importante actividad pesquera, una creciente actividad de avistamiento de cetáceos comercial y problemas geopolíticos,debido a la proximidad entre Gibraltar, Ceuta, España y Marruecos. Se analizará como cada uno de ellos puede afectar alas poblaciones de cetáceos presentes en el estrecho de Gibraltar, tanto a corto, medio y largo plazo.
INTERACCIONES ENTRE CETÁCEOS YACTIVIDADES ANTROPOGÉNICAS ENEL ESTRECHO DE GIBRALTAR
Alicia Sánchez / Philippe Verborgh Renaud De Stephanis / Sergi Pérez / Christophe Guinet

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 168/276
Almoraima, 35, 2007
168
MATERIAL Y MÉTODOS
Área de estudio
La zona de estudio es el área del estrecho de Gibraltar,comprendida entre los 5º y 6º de longitud Oeste, intentandocubrir toda el área hasta las aguas del reino de Marruecos. Elestrecho de Gibraltar (figura 1) tiene aproximadamente 60 Kmde largo. Su frontera oeste está localizada entre el cabo deTrafalgar, en España, y el cabo Espartel, en Marruecos, sepa-rados por 44 Km. El Estrecho se hace más angosto hacia el este,llegando a una distancia mínima entre Tarifa y Punta Cires de14 Km. Su frontera este está localizada entre Punta Europa(Gibraltar) y Punta Almina (África) y están separadas por 23Km (Parrilla et al., 1988). La batimetría del Estrecho secaracteriza por ser un cañón que va de este a oeste con aguaspoco profundas (200-300 m), que se pueden encontrar en lacara atlántica, y aguas más profundas (800-1000 m) en la caramediterránea (figura 1).
Interacciones con embarcaciones de avistamiento decetáceos
Para analizar las interacciones existentes entre los cetáceos ylas embarcaciones presentes en el Estrecho, se analizaron,por un lado, las reacciones que los animales tienen conrespecto a las embarcaciones de whale watching, y, por otro,la evolución que está teniendo la actividad en el Estrecho enlos últimos años. Los datos utilizados para este estudioproceden de los avistamientos realizados en una campaña,desde embarcaciones de avistamientos de cetáceos en Tarifa,
entre el 25 de abril y el 31 de octubre de 1999, desde dos barcos de 7 y 9 metros, pertenecientes a las empresas FIRMMy Mar de Ballenas S.L. Se tomaron datos sobre las especies encontradas, número de individuos avistados y reacción a laembarcación en el momento de ser contactados por la misma (se tomó como “atracción”, cuando al menos un animal del
grupo se acercaba, “huída” cuando los animales mostraban claros síntomas de alejamiento a la embarcación, e“indiferencia” cuando no se observó cambio de comportamiento alguno).
Interacciones con el tráfico marítimo
Se analizó la evolución que el tráfico marítimo ha tenido en el Estrecho en los últimos años a partir de los datos obtenidospor el puerto de Algeciras y por medio de datos cedidos por el CZCS Tarifa, que identificó entre 85% y 90% de lasembarcaciones que transitaron por el Estrecho en 1998 y 1999. Por otro lado, se trató de identificar las reacciones que lasdiferentes especies de cetáceos tienen con respecto a los ferris que unían Ceuta con Algeciras, a partir de un estudiorealizado por Roussell en 1999. En este estudio se realizó un total de 122 embarques, durante los meses de marzo, abril ymayo de 1999 en los ferris que unían las dos orillas del Estrecho, y se tomaron datos de avistamientos de cetáceos, así como
Figura 1. Área de estudio.
Figura 2. Muestreos realizados en el Estrecho entre 1998 y 2004. En rojo serepresentan los seguimientos a cetáceos.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 169/276
Comunicaciones
169
de sus reacciones a las embarcaciones. Finalmente, se coteja-ron las rutas que las embarcaciones tienen en el Estrecho conrespecto a las especies que más podrían sufrir colisiones oimpactos negativos. Se plasmaron también las nuevas rutasque el nuevo puerto de Tánger introducirá en el Estrecho a lolargo de los próximos años.
Interacciones con el sector pesquero
La evaluación de la actividad pesquera ha sido realizada através del censo de la flota que faena en esta zona, publicadoen el BOE 192 del 12 de agosto de 1998. Asimismo, setomaron datos obtenidos a lo largo de las campañas queCIRCE lleva desarrollando en el estrecho de Gibraltar desde1998. Se ha realizado un total de 22.649 kilómetros durantelos cuales el esfuerzo de búsqueda era el más adecuado paraeste estudio, cubriendo prácticamente todo el estrecho deGibraltar (figura 2). Asimismo, se realizaron un total de 2.288avistamientos de 10 especies de mamíferos marinos con untotal de 72.842 individuos observados (tabla 1). Los datosobtenidos a lo largo de estos muestreos se analizaron para estetrabajo. Finalmente, una campaña de 1.200 entrevistas a
pescadores en el puerto de Tarifa proporcionará datos realessobre las interacciones que existen en el estrecho de Gibraltarentre cetáceos y pesquerías.
Problemas geopolíticos
Se analizó la situación geopolítica del Estrecho, y los proble-mas que ello conlleva en la conservación de los cetáceos delestrecho de Gibraltar.
Figura 3. Evolución del número de embarcaciones de avistamiento de cetáceoscomercial a lo largo del tiempo.
Figura 4. Evolución de la capacidad total de las embarcaciones de avistamientode cetáceos comercial a lo largo del tiempo.
Especies Avistamientos Individuos observadosBalaenoptera acutorostrata 2 2Balaenoptera musculus 1 1Balaenoptera physalus 26 41Cistophora cristata 2 3Delphinus delphis 451 15.993Globicephala melas 576 14.717Ornicus orca 50 255Physeter macrocephalus 348 446Stenella coeruleoalba 435 34.076Tursiops truncantus 397 7.308Total general 2.288 72.842
Tabla 1. Avistamientos realizados en el Estrecho entre 1998 y 2004.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 170/276
Almoraima, 35, 2007
170
RESULTADOS
Interacciones con embarcaciones de avistamiento decetáceos
A finales de 2004, había un total de 13 embarcaciones whalewatching entre los puertos de Tarifa, Algeciras, La Línea yGibraltar, con una capacidad total de 864 plazas. En lasfiguras 3 y 4 se puede apreciar la evolución del número deembarcaciones de avistamientos de cetáceos así como sucapacidad en el Estrecho entre 1982 y 2004.
Durante la campaña desde embarcaciones de avistamientocomercial, se realizaron un total de 336 avistamientosdurante los cuales se pudieron tomar los datos de atraccióna las embarcaciones. En el 48% de los avistamientos, losanimales fueron atraídos por la embarcación. Los resulta-dos por especies se pueden observar en la figura 5.
Interacciones con el tráfico marítimo
El estrecho de Gibraltar es la segunda vía marítima mástransitada del mundo (Gárate J., com pers). Dada la impor-tancia del transporte marítimo, 75% del volumen nacional
e internacional, esta zona presenta una evidente relevanciacomercial. Por otro lado, sustancias potencialmente peli-grosas son transportadas a diario a través del Estrecho, así como la posible existencia de una importante contamina-ción acústica.
En 1999 se registraron un total de 83.856 embarcacionescruzando el Estrecho, sin incluir embarcaciones de recreo,pesqueros ni embarcaciones de avistamiento de cetáceos.De estos, el 36% representaba tráfico de pasajeros entre lasdos orillas del Estrecho (tabla 2). En cuanto a la evolución
del tráfico marítimo, se puede apreciar en la figura 6, cómoéste casi se ha doblado en los últimos 5 años.
GIBREP’S* (E-W) 53.336Ferrys (N-S) 13.473EAV** (N-S) 17.047Total de buques identificados 83.856
* Mercantes, petroleros y porta-contenedores. ** Embarcación de alta velocidad.NOTA: No se incluyen embarcaciones menores (pesqueros, embarcaciones de recreo…)
Tabla 2. Tráfico marítimo en 1999 en el estrecho de Gibraltar.
Figura 5.Reacciones de atracción a los barcos de avistamiento de cetáceos por especies.
Figura 6. Evolución de tráfico de pasajeros, comparativa entre el año 2000 y 2005.
Figura 7. Reacciones de atracción a los ferris por especies.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 171/276
Comunicaciones
171
Los datos obtenidos desde los ferris en 1999 se puede apreciar en la figura 7.
En las figuras 8 y 9, se pueden apreciar las rutas de los ferris y fast ferris presentes y previstas con la creación del nuevopuerto de Tánger, y la distribución de cachalotes en la zona de estudio.
Se ha podido confirmar un total de dos colisiones entre embarcaciones y cachalotes en el Estrecho, una de las cuales fueobservada en directo por el equipo de CIRCE en septiembre de 2002. Asimismo, se ha confirmado la colisión entre dosembarcaciones y dos rorcuales comunes y entre por lo menos un calderón común y una embarcación.
Interacciones con el sector pesquero
Se realizó un total de 1220 entrevistas durante el año 2004, en el puerto pesquero de Tarifa, y se concluyó durante estasentrevistas, que la única interacción que podía existir entre cetáceos y las pesquerías del Estrecho se producía entre las orcasy los pescadores de atún con piedra. Los datos de las entrevistas reflejaron que el 17% de las capturas realizadas por lospescadores de Tarifa eran robadas por grupos de orcas en el Estrecho, durante los meses de julio y agosto. Asimismo, a partirde las observaciones realizadas por el equipo de investigación de CIRCE, se ha observado que hay interacciones entre lospescadores de curricán y los calderones comunes del Estrecho. Los pescadores de curricán, pesqueros deportivos, cruzanlos grupos de calderones comunes, con sus líneas, cuyos anzuelos pueden clavarse en los cuerpos de estos cetáceos.
Problemas geopolíticos
El área de estudio esta localizada en un importante punto estratégico donde confluyen intereses de diversas naciones. Lasaguas están bajo soberanía de España y Marruecos, y además la colonia británica de Gibraltar complica aún más la situacióngeopolítica. Además, se conoce la existencia de tres zonas de tiro, y el tránsito de todo tipo de embarcaciones militares(incluidos submarinos, tanto en superficie como sumergidos), así como ejercicios militares de la OTAN (operacionesNEOTAPON), lo que acarrea posibles problemas para los cetáceos (Frantzis, A.& Cebrian,D.,1998; Rendell L.E. &Gordon,J.C.D., 1996; Tyack,P.L. & Clark, C.W. 1998;). Finalmente, existe una descoordinación entre las administracionespúblicas a la hora de ordenar, y sobre todo gestionar lo referente a cetáceos, desconociendo algunos de estos órganos losestudios que se están realizando en la zona, y financiando estudios ya realizados.
DISCUSIÓN
Los resultados de este estudio demuestran que existe una serie de problemas entre cetáceos y actividades antropogénicasen el Estrecho. Las actividades pesqueras, de avistamiento de cetáceos, las pesquerías, y los problemas geopolíticos de lazona, pueden llegar a afectar tanto negativamente como positivamente a las poblaciones de cetáceos.
En cuanto a las interacciones positivas, siempre desde el punto de vista de los cetáceos, en este caso las orcas, los pescadoresde atunes del Estrecho están proporcionando alimento extra, con un gasto energético mínimo por parte de la especie. Sinembargo, esto tiene efectos negativos de cara a los pescadores, ya que éstos ven reducidos sus beneficios con este tipo deinteracciones, y podría generar algún tipo de repulsa por su parte hacia las orcas, resultando en interacciones negativas haciaellas. El hecho que la pesca deportiva de curricán afecte a las poblaciones de calderones comunes debe tenerse muy en cuentatambién, ya que a pesar de producir pequeñas heridas y molestias en los grupos que descansan en la superficie, a la larga podríatener un impacto negativo en sus comportamientos, y por tanto podría afectar negativamente en su abundancia absoluta Elavistamiento de cetáceos de tipo comercial, siempre y cuando se realice de forma correcta, siguiendo unos códigos éticosadecuados para la zona, y con proyectos de educación ambiental a bordo de calidad son beneficiosos para las especies decetáceos, ya que dan a conocerlas, y permiten que se apliquen, a la larga, medidas de gestión para ellas. No hay nada peor que

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 172/276
Almoraima, 35, 2007
172
Figura 8. Rutas de ferris y fast ferris y distribución de cachalotes en el Estrecho.
Figura 9. Rutas de ferris y fast ferris previstas en la zona con la construcción del nuevo Puerto de Tánger y distribución de cachalotes.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 173/276
Comunicaciones
173
una especie en peligro que no se conozca, ya que la presión social no accedería a tomar medidas protectoras hacia ellas. Enel caso del Estrecho los códigos éticos se siguen, a pesar de no haberse legislado ningún decreto para ello por parte delMinisterio de Medio Ambiente, o Consejería de Medio Ambiente de la Junta de Andalucía. Los resultados referentes a loscomportamientos de acercamiento así lo demuestran. Sin embargo, no hay ningún proyecto de educación ambientalcoordinado entre todas las empresas a bordo de las embarcaciones, a pesar de que haya naturalistas que informen al pasajede lo que están viendo, por lo que muchas veces la información que lo pasajeros reciben es dispar y a veces contradictoria.
Uno de los problemas actuales más apremiantes para la conservación de las especies de cetáceos en el Estrecho, y enparticular los de gran tamaño, como son el rorcual común y el cachalote, es la construcción del nuevo puerto de Tánger,situado al oeste del cabo de Punta Cires. Como se ve en los mapas de distribución de cachalotes, y la predicción de nuevasrutas de ferris y fast ferris en el Estrecho, la probabilidad de que haya alguna colisión entre esta especie y las embarcaciones
de transporte de pasajeros, aumentará de forma drástica, al cruzar de pleno las zonas de máxima abundancia de la especie.Para el caso de calderones comunes y delfines mulares, las rutas cruzarán también sus zonas de máxima presencia en elEstrecho. A pesar de que es poco probable que existan colisiones con estas especies, y en particular con delfines listadosy delfines comunes, u orcas (si nos atenemos a los datos de acercamiento obtenidos desde ferris), lo que si que se produciráes un aumento drástico de la contaminación acústica en la zona, que podría producir molestias tanto a corto como a medioplazo, podría influir en la abundancia de estas especies, al poder reducirse sus tasas de natalidad a lo largo del tiempo.
Otro problema que existe, son los problemas geopolíticos. Los problemas a la hora de legislar, y sobre todo a la hora degestionar, vienen dado, por las diferentes soberanías que existen en la zona. Así pues, por ejemplo, a pesar de que ya hayaempezado a construirse el puerto nuevo de Tánger, el estudio sobre el impacto del mismo, se está realizando actualmentey, por tanto, no podrá tener en cuenta ninguna medida correctora, ni podrán evaluarse los impactos en su fase deconstrucción. Además de los problemas internacionales, existen problemas locales, ya que hay una terrible descoordinaciónentre las diferentes administraciones públicas. Asimismo, hay un total desconocimiento por parte de administracionesandaluzas por la labor que entes privados puedan estar desarrollando en la zona, y por tanto, recursos económicos quepodrían sumar conocimientos y propuestas de medidas de gestión, y que al final se quedan en saco roto, repitiendo estudiosya realizados con fondos privados.
CONCLUSIÓN
Como conclusiones a este estudio, se puede apreciar que es urgente desarrollar una serie de medidas que ayuden a controlarestos problemas:
Por un lado, se debería limitar la utilización de curricán por parte de embarcaciones de recreo, sobre todo en zonas depresencia de cetáceos. Por otro lado, un decreto que regule las actividades de avistamiento de cetáceos comercial se presentacomo posible medida urgente en la actualidad.
Se hace necesario un estudio tanto a corto como largo plazo de los impactos que el nuevo puerto de Tánger producirá sobrelas especies de cetáceos residentes, semi-residentes o migratorias en la zona, ya que un aumento drástico del tráficomarítimo en el Estrecho, llevaría probablemente a la desaparición de los grupos de cachalotes en la zona, y podría poneren peligro también las migraciones de rorcuales comunes.
Además, un estudio en profundidad de las actividades de investigación que se están realizando en el Estrecho es necesario,para poder coordinar todos los esfuerzos para una correcta protección de las poblaciones de cetáceos presentes en la zona.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 174/276
Almoraima, 35, 2007
174
AGRADECIMIENTOS
Este anuncio ha sido financiado por la Ciudad Autónoma de Ceuta, entre 2000 y 2001. Entre 2002 y 2005, el ProyectoLIFE02NT/E/8610 "Conservación de Cetáceos y Tortugas Marinas de Murcia y Andalucía, coordinado por la sociedadEspañola de Cetáceos, financiado en un cincuenta por ciento por la Comisión Europea y que cuenta entre sus socios ycofinanciadores la Consejería de Medio Ambiente de la Junta de Andalucía, la Consejería de Industria y Medio Ambientede la Región de Murcia, Ministerio de Medio Ambiente (DGB y DGC), la Secretaría general de Pesca Marítima delMinisterio de Agricultura, Pesca y Alimentación, la Universidad de Cádiz, y el Instituto Español de Oceanografía. Así mismo, este estudio ha contado con la donación de 123 voluntarios de CIRCE.
BIBLIOGRAFÍAFRANTZIS, A. y Cebrian, D. "A rare,atypical mass stranding of Cuvier’s beaked whales. Cause and implications for the species’biology". 332-333. In: European Research on Cetaceans -12 . Proc. 12th Ann. Conf. ECS, Monaco, 20-24 Jan, 1998. (Eds. P.G.H. Evans and E.C.M. Parsons). European Cetacean Society, Valencia,Spain. 1998. pp. 436.PARRILLA G., Kinder T.H., Bray N.A. "Hidrología del agua mediterránea en el Estrecho de Gibraltar durante el Experimento Gibraltar (octubre 1985-octubre 1986)".Seminario sobre la oceanografía física del estrecho de Gibraltar (Madrid, 24-28 de octubre 1988), pp. 95-121. 1988.RENDELL L.E. y Gordon, J.C.D. "Vocal responses of long-finned pilot whales(Globicephala melas) to military sonar pulses". pp. 69-71. In: European Research onCetaceans - 10: Proc. 10th Ann. Conf. ECS, Lisbon, Portugal, 11-13 Mar, 1996. (Ed. P.G.H. Evans). European Cetacean Society, Kiel, Germany. pp. 334. 1996.ROUSSEL E. "Les cétacés dans la partie orientale du Détroit de Gibraltar au printemps: indications d’écologie". Master thesis. Ecole Pratique des Hautes Etudes,Montpellier, France. 1999.SEC. Sociedad Española de cetáceos. Recopilación, análisis, valoración y elaboración de protocolos sobre las labores de observación, asistencia a varamientos yrecuperación de mamíferos y tortugas marinas de las aguas españolas. Ministerio de Medio Ambiente Español. Secretaria General de Medio Ambiente, TechnicalReport Sociedad Española de Cetáceos Avaible from SEC, Nalón 16, E-28240 Hoyo de Manzanares, Madrid, Spain. 1999.TYACK,P.L. y Clark, C.W. Quick Look, Phase II, Playback of low frequency sound to gray whales migrating past the central California coast, January, 1998.Unpublishedreport, department of Biology, Woods Hole Oceanographic Institute, Woods Hole, MA 02543. 1998.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 175/276
Comunicaciones
175
RESUMEN
La laguna de La Janda constituyó la laguna más importante y representativa de la península Ibérica. Las lagunas y áreaspalustres de Janda, Jandilla, El Torero, Tapatana, Tapatanilla, Rehuelga y Espartinas constituyeron las áreas másimportantes de este extenso humedal. Su desecación representó un punto álgido del desarrollismo agrícola español de ladécada de los sesenta del siglo pasado. Hoy día a pesar de las obras hidráulicas realizadas y a las grandes modificacionessufridas por la dinámica natural y condiciones ecológicas de esta laguna, grandes áreas de lo que constituyeron sus antiguosfondos continúan aún inundándose y conservando gran parte de los valores ambientales primigenios. Tras diez añosrealizándose trabajos de investigación y actividades científicas tendentes a recuperar parte de los antiguos humedales queen ella existieron, la Administración aun no ha realizado obras tendentes a la restauración de los mismos.
Palabras clave: laguna, Janda, humedal, restauración ecológica.
SUMMARY
La Janda lake was once the most important and representative lake within the Iberian Peninsula. The lakes and natural vegetationwater of Janda, Jandilla, El Torero, Tapatana, Tapatanilla, Rehuelga and Espartinas were the most important areas of this vastwetland and their subsequent reclamation coincided with the zenith of spanish agricultural development in the 1960s. Despitethe completion of drainage works and transformations in both the natural landscape and the ecological conditions of the lake, largeareas of the old lake bottom continue to flood and have retained many of their primitive environmental attributes. A series of studies and scientific projects currently under way have set out to regenerate part of the former wetlands. Today, ten years later,the autonomic goberment still haven´t developed regeneration events on these wetlands.
LA LAGUNA DE LA JANDA (CADIZ):10 AÑOS DESPUÉS DEL INICIO DE LOSESTUDIOS BÁSICOS PARA SURESTAURACIÓN ECOLÓGICA
J. C. Castro Román / J. M. Recio Espejo

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 176/276
Almoraima, 35, 2007
176
I. INTRODUCCIÓN
Uno de los humedales más importantes y extensos de la península Ibérica fue la antigua laguna de La Janda. Su importanciay relevancia cultural e histórica se ha visto reflejada en el acervo cultural de los habitantes de la zona. La importancia de laspinturas rupestres de las cavidades y abrigos de su entorno, su arqueología y megalitismo (Cabre y Hernández Pacheco,1914;Mass,1994; Mass et al., 1995), o incluso los primeros datos que sobre su importancia ornitológica nos dejaron los primerosnaturalistas ingleses que la visitaron a principios de siglo XX, demostraron a todos la importancia e interés natural del mismo(Irby,1895; Verner,1909; Chapman y Buck,1910; Bernaldo de Quiros, 1920; Yates,1945) (figura 1).
Pero a pesar de todos estos valores y al igual que aconteció en otros humedales de nuestra Península, y según el modo deactuar de la Administración y la sociedad de aquella época, sus valores e importancia ecológica no fueron tenidos en cuenta.Tras la ejecución de las obras de desecación los suelos presentes en sus cubetas, las conocidas Tierras Negras Andaluzas(Hernández Pacheco,1915; Cano et al.,1994; Díaz del Olmo y Recio,1995), fueron puestos en cultivos de regadíosaprovechándose los propios canales para efectuar los riegos en la época de estío (Castro et al., 1994)
Pero como ocurre en muchas ocasiones las condiciones naturales primitivas se resisten a ser modificadas y a desaparecerpor completo y no dejan de manifestarse en cuanto las condiciones naturales le son ligeramente favorables. En el caso dela laguna de La Janda a pesar de las grandes obras ejecutadas la depresión continúa inundándose en los inviernos lluviososque con frecuencia acontecen en la comarca, y una extensa lámina de agua continua formándose a pesar de la acción dedrenaje efectuada por la canalización y a los represamientos realizados en la cabecera de los ríos.
Aunque ésta es efectiva en un plazo de unos quince o veinte días, ciertas zonas internas de la depresión coincidentes conlas zonas menos transformadas agrícolamente continúan con unas condiciones hidromorfas más prolongadas y duraderas
(Castro y Recio, 1990; Recio et al. 1995)
II.1. MEDIO FÍSICO Y CONDICIONAMIENTOS NATURALES
La laguna de La Janda se encuentra ubicada en una depresión tectónica cercana a la costa, en la fachada atlántica peninsulardel sur de la provincia de Cádiz. Localidades como Vejer de la Frontera, Medina Sidonia, Benalup-Casas Viejas o Tarifase encuentran estrechamente ligadas a ella.
De génesis compleja, la depresión tectónica de La Janda no llega a funcionar como humedal hasta hace unos 7000 años,cuando un progresivo ascenso del nivel del mar provocó la formación de un ambiente litoral tipo estuarino-marismeño,originando una bahía interior a donde darían término los cursos principales de agua (ríos Barbate y Almodóvar). Hace 3800
años se produciría el cierre de la conexión entre el mar abierto y la citada bahía (Luque et al., 1999). La progresiva bajadadel mar haría evolucionar a la zona hacia un ambiente palustre más continental afectado por grandes inundaciones fluvialesy controlado en cierta manera por los niveles de base que imponen la pleamar y bajamar del mar Atlántico cercano. Laantropización del territorio sería la causa principal y final de la génesis y colmatación del espacio marismeño actuallocalizado en la desembocadura del río Barbate antes comentado (Zazo, y Obejero, 1976; Borja,1992; Recio et al. 1997a,Luque et al., 1999).
II.2. Aspectos climáticos
Su proximidad al estrecho de Gibraltar hace al viento de levante un factor de suma importancia ya que va a ser el responsablede controlar no solo el régimen de precipitaciones y temperaturas y por tanto de la evaporación, si no que también lo vaa ser de todo el desarrollo de la vegetación del entorno.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 177/276
Comunicaciones
177
Las precipitaciones de la zona pueden considerarse cuantiosas del orden de los 700-1.000 mm anuales. La temperaturasmedias son del orden de 17ºC y los valores de evapotraspiración son superiores a 900 mm (Ibarra, 1993 y 1995). El áreade la depresión de La Janda presenta por tanto un ombroclima subhúmedo y pertenece al piso bioclimático termomediterráneo
inferior. Las lluvias se reparten principalmente en la época invernal donde son frecuentes los temporales de procendenciaatlántica que dejan en la zona sustanciosas e intensas precipitaciones repartidas en pocos días de lluvias. Todo ello facilitaenormente el incremento rápido de los caudales fluviales y la formación de grandes avenidas que acumuladas en ladepresión tectónica constituyen la base de los aportes y formación de la antigua lámina de agua. La impermeabilidad delsustrato y la altitud del fondo de la llanura aluvial próximo a los 3 m.s.n.m., serían coautores de la formación de la mismay favorecedores de la acumulación del agua (Recio y Dueñas, 2000).
II.3. Importancia ornitológica
La importancia ornitológica de La Janda se debió fundamentalmente a su posición geográfica estratégica entre el continenteeuropeo y el africano, al servir de área clave para las migraciones pre y posnupciales de un gran conjunto de aves,
Figura 1. Ubicación y evolución histórica de La Janda. A. Península Ibérica, B. Marruecos, C. Andalucía, D. Provincia de Cádiz, a. Cádiz, b. Vejer, c. Benalup, d.Tarifa, e. Barbate, 1. Laguna de La Janda, 2. Aldea de Casas Viejas, 3. Aldea de Tahivilla, 4. CN-340, 5. Aldea de Facinas, 6. Marismas del Barbate, 7. Red de canalesde desecación, 8. Embalse del Celemín, 9. Embalse del Barbate, 10. Embalse de Cabrahigos,11. Embalse del Milagro, 12. Embalse del Almodóvar, 13. Parque Naturalmaritimo-terrestre del Pinar, Acantilado y Marismas de Barbate.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 178/276
Almoraima, 35, 2007
178
principalmente anátidas y limícolas (Verner, 1912). Al tratarse de un humedal muy heterogéneo era posible encontrarmultitud de especies diversas cada una de ellas adaptadas a las diferentes características que poseían las lagunas yhumedales en ella existentes. Además al estar rodeada por grandes masas de alcornocales y acebuchales entremezcladoscon grandes roquedos (Laja de Aciscar por ejemplo) veía incrementado el número y diversidad de especies no sólo lasorníticas sino las de otros tipos zoológicos (Castro et al., 1996)
En la actualidad la fauna esta formada principalmente por una importante comunidad de aves que aunque no tansobresaliente como la que existió en el pasado, sigue manteniendo su interés; abundantes poblaciones de grulla (Grus grus),cigüeña blanca (Ciconia ciconia), calamón (Porphyrio porphyrio), cigüeña negra (Ciconia nigra), y una gran variedad deaves acuáticas y limícolas siguen siendo frecuentes en toda la zona (Finlayson, 1992; Castro et al., 1996, Mosquera yFinlayson, 1997).
Las anátidas y ardeidas no sólo nidificaban entre la vegetación que existía sino que además descansaban y recuperabanfuerzas para el salto definitivo a sus cuarteles invernales. Patos cuchara ( Anas clypeata), silbones ( Anas penelope), porrones(Aythya spp.), cercetas ( Anas spp.), tarro blanco (Tadorna tadorna) y ánsares ( Ansar ansar ), constituyeron las poblacionesmás numerosas; algunas de ellas siguen observándose actualmente en época de inundación en La Janda o aprovechandocanales de desecación y arrozales. La comunidad de ardeidas también fue y sigue siendo una de las más interesantes de estesingular humedal con la presencia de avetoro ( Botaurus stellaris), garcilla cangrejera ( Ardeola ralloides), bueyera( Ardeola ibis), martinetes ( Nycticorax nycticorax), garcetas (Egretta garzetta), garzas reales ( Ardea cinerea) e inclusomorito (Plegadis falcinelus) (Castro et al., 1996; Dueñas y Recio, 2000).
III.1. GÉNESIS Y FUNCIONAMIENTO DEL ANTIGUO HUMEDALA los condicionamientos tectónicos, litológicos y paleogeográficos antes comentados hemos de sumar el comportamientode los cursos principales de la red fluvial generadora de la antigua laguna y responsable de sus aportes hídricos. Dos seríanlos cursos fluviales principales responsables de su génesis: por un lado el río Barbate junto al Celemín y de otro la corrientefluvial que en sentido este-oeste recorría todo el fondo de la depresión constituyendo el río Almodóvar.
La lámina de agua así generada llegaba a superar una extensión de unas 4.000 hectáreas constituyendo todo una gransuperficie que persistía durante prácticamente toda la época otoño-primaveral, conformando un extensísimo espaciohúmedo que denominamos como laguna de La Janda (Recio et al., 1996; Recio et al., 1997a). La extensión de esta ampliasuperficie fascinó y atrajo la atención del hombre primitivo, y los árabes le denominaron "La Albuhera" (albufera)(Saavedra, 1881)
Con posterioridad a las fases de máximas avenidas fluviales y niveles mas elevados alcanzados por las aguas y al drenajeejercido por la corriente fluvial principal, se producía la fragmentación de la extensa lámina de agua formada. Sería en estasituación cuando diferentes lagunas y áreas palustres de importante valor ecológico se independizaban unas de otras paraevolucionar independientemente en base a los diferentes mecanismos de génesis que los formaban. Estos humedalesvendrían a representar lo denominados por nosotros como humedales anexos a Janda (Dueñas y Recio, 2000)
De esta forma y asociada a un paleotrazado del río Barbate quedaba constituida la laguna de Rehuelga. Aguas abajo, esta vezen la margen derecha y a favor de los diques fluviales formados y a los aportes de aguas recibidos de las surgencias existentesen el borde de las mesas pliocenas, se constituía con carácter casi permanente la laguna de Espartinas; también en la margenderecha y desconectada de la gran lámina de agua se generaba la laguna de Jandilla. La laguna del Torero era otro pequeñohumedal asociado a la margen izquierda del Barbate, con cerca de 3 m. de profundidad y una gran riqueza botánica.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 179/276
Comunicaciones
179
Junto a estas lagunas de gran extensión habría otras de menores dimensiones tales como las lagunas de Canteruelas, elÁguila y la de Los Hierros, las cuales eran mantenidas y diferenciadas gracias a los aportes procedentes de la sierra de Retíno a los procedentes de la mesa de El Cañar respectivamente. La laguna de Los Lobos era otra pequeña laguna generada comoconsecuencia de los derrames del Tragante-Celemín.
Asociado al discurrir del rio Almodóvar y a sus levées delimitadores de su cauce, quedaba formada la zona palustre deTapatana, conocida en parte como laguna de Tarifa y en menor medida la laguna de Tapatanilla. Ya en plena depresión yallí donde esta corriente fluvial carecía de poder de evacuación del agua, ésta quedaba almacenada en diferentes zonasdepresionarias constituyendo la laguna de Janda propiamente dicha. El "charco de los ansares" constituía aquí un área muyimportante con una profundidad en torno a los 3 metros.
A todo este gran complejo palustre habríamos de añadir un conjunto de lagunas de pequeño tamaño que se localizan en elentorno de La Janda en lo alto de las plataformas calcareníticas de la zona (lagunas de Alcalá, Cantarranas o de Las Habas)(Dueñas y Recio, 2000) (figura 2).
Figura 2. Área de ubicación de las antiguas lagunas. 1. Mesa de Las Lomas, 2. Área de Jandilla, 3. Río Barbate, 4. Río Celemín, 5. Embalse del Celemín, 6. Mesadel Mirador, 7. Arroyo Juan de Sevilla, 8. Arroyo del Trimpancho, 9. Arroyo del Torero, 10. Arroyo Culebras, 11. Río Almodóvar, 12. Curva de nivel 10 metros, 13.Carretera Nac.340 (Vejer-Tarifa), 14. Canal colector del este, 15. Marismas del Barbate. * Inicio de la canalización Sentido de los flujos.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 180/276
Almoraima, 35, 2007
180
III.2. Síntesis histórica
Las obras de desecación de la laguna comienzan en 1822 fecha en la que se concede a Segismundo Moret un primer proyectoe intento para drenar y canalizar las aguas de la misma. El ingeniero Larramendi se encarga de las obras y son construidos losprimeros canales de San Fernando y San José con los que evacuar las aguas del río Barbate y río Almodóvar. La construcciónde dichas canalizaciones no fueron del todo eficientes y en 1929 al amparo de la Ley General de Obras Públicas se inicia unprimer estudio sobre la viabilidad de desecación de la laguna de La Janda por parte de la empresa Colonias Agrícolas, S.A.En 1935 se realiza y actualiza un primer proyecto elaborado por el ingeniero Suarez Sinova en 1929. En 1946 el Estado otorgala concesión de desecación correspondiente y en 1947 la empresa adjudicataria solicita la realización del deslinde de losterrenos de dominio público. Éste es realizado en 1949 por la Confederación Hidrográfica del Guadalquivir y el InstitutoGeográfico Nacional es el organismo responsable de efectuar su nivelación y delimitación taquimétrica.
Al mismo tiempo que se desata una gran discusión entre los propietarios en relación al deslinde efectuado, en 1954 sedeclara incursa la concesión otorgada a la empresa Colonias Agrícolas, S.A. al no ser ejecutado y finalizado ellevantamiento del plano parcelario correspondiente. Por ello en 1957 el Consejo de Ministros concede a la empresa Lagunasdel Barbate, S.A. la concesión administrativa de desecación de la laguna, realizándose en el mismo año la aprobación delplano parcelario. Durante todo el año de 1958 el resto de propietarios de la zona presentan recursos en relación a la concesiónadministrativa otorgada a la empresa adjudicataria antes citada.
Durante la década de los sesenta se especifican las diferentes etapas por las que han de pasar las obras, y se dicta de urgenteocupación los terrenos afectados por la desecación de la laguna. Sin embargo al mismo tiempo que en 1964 se produce elrescate por el Estado de la concesión que fue otorgada mediante Decreto del 29/11/1949, acontece por un lado el desarrollode la segunda fase del proyecto inicial, y por otro la creación de una comisión específica que evalúa los gastos efectuados
hasta la fecha por Lagunas del Barbate, S.A. Por último en 1965 se autoriza al concesionario la continuación de las obrasy concesión otorgada hasta que se consuma el rescate por parte del Estado.
Por ello a finales de la década de los sesenta se efectuó la canalización de las aguas que abastecían al humedal, arrancaday aniquilada la vegetación palustre que se asociaba a estos ecosistemas y la rica ornitofauna desapareció mayoritariamentede todo el enclave. La madre vieja del Barbate fue canalizada, conectada al cauce nuevo y borrada casi en su totalidad engran parte de su recorrido, a la vez que fueron edificados diques de contención en la zona de Jandilla.
Se establece una red principal de canales (canal colector del este y otros), capturados y canalizados toda la red de pequeñosarroyos que en ella desembocaban, y es embocado y conducido en dirección este el río Almodóvar a través del puente deTapatana. Mediante la construcción de un gran canal-túnel, las aguas se dirigieron de una manera totalmente artificial ysubterráneamente hacia la zona noreste de la cercana marisma del Barbate, la cual comenzó a recibir aportes hídricos deaguas dulces experimentando por ello un gran cambio en sus características biológicas (figura 3).
No obstante y a pesar de todas estas faraónicas obras hidráulicas bien de desecación de la depresión bien de construcciónde grandes embalses, la Janda volvía a inundarse y a alcanzar su plenitud laminar año tras año (Recio et al., 1997b).
Ello animó al grupo de Ecología Aplicada de la Universidad de Córdoba a iniciar en 1990 una serie de acciones encaminadasa posibilitar la recuperación de parte de aquel extenso humedal. A través de reuniones científicas de carácter internacional ynacional que recabaron el interés y la opinión de expertos, y mediante la realización de nuevas investigaciones y trabajos enrelación con el humedal se intentó dar a conocer el proyecto. Con la presentación del mismo a los fondos LIFE de la ComunidadEuropea se intentó dar un paso a nivel internacional para conseguir sus objetivos (Castro y Recio, 1990; Castro et al., 1993).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 181/276
Comunicaciones
181
En 1995 el gobierno autónomo de Andalucía a través de su Consejería de Medio Ambiente manifestó poseer un importanteinterés sobre La Janda y sobre sus posibilidades reales de regeneración parcial, apostando por una nueva alternativacomplementaria a las actividades que de tipo agrícola y ganadero se llevan a cabo en la zona. Por ello y bajo la ayuda económicade la misma desarrollamos un proyecto de investigación tendente a sentar unas primeras bases ecológicas que permitieran laregeneración de humedales en lo que fue la antigua laguna de La Janda (Consejería de Medio Ambiente, 1995). Éste se vio
complementado con otras actividades culturales alternativas tales como una exposición itinerante por las localidades de la zonao con la celebración de unas primeras jornadas técnicas celebradas en Vejer de la Frontera, en octubre de 1996.
Con lo señalado más arriba se quiso poner de manifiesto el interés que existe por restaurar parte de lo que fue un importantey antiguo humedal, restablecer sus condiciones primigenias y recuperar parte de nuestro patrimonio natural perdido,haciéndolo compatible en todo momento con la explotación agro-ganadera de la comarca. Para ello fue necesario tanto unaevaluación ecogeográfica de la depresión donde se ubicaba La Janda y de los condicionamientos ambientales actuales comouna reconstrucción lo más fidedigna posible de los antiguos ecosistemas palustres existentes (Recio et al., 1997b)
Por ello en 1995 comenzaron los estudios del Área de Ecología de la Universidad de Córdoba encaminados a vislumbrarla factibilidad de la restauración de este singular ecosistema palustre. Desde entonces la Consejería de Medio Ambiente
Figura 3. Canalizaciones de drenaje en la antigua laguna de Janda. Aerofotograma de 1992.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 182/276
Almoraima, 35, 2007
182
realizó un esfuerzo de difusión de los valores ambientales de la Janda llevando a cabo una campaña de concienciaciónpública en todos los municipios de la comarca. Además se han publicado decenas de artículos científicos y una tesisdoctoral, todo ello sobre los valores naturales y la viabilidad de la restauración de la laguna de La Janda. Hay que añadirque entre 1999 y 2000 se realizó también un estudio de viabilidad económica del citado proyecto analizándose incluso elnúmero de puestos de trabajo perdidos o ganados. En 2002 se publicó por parte de la citada consejería un anteproyecto derestauración cuantificándose la inversión para junio del citado año en 1.150 millones de €; en la actualidad tan sólo se haninvertido 231.000 € a través de fondos INTERREG, repoblando con vegetación algunos tramos de los ríos Barbate, Celemíny Almodóvar (Consejería de Medio Ambiente, 2005). Todo ello dista bastante del proyecto de restauración parcial de loshumedales de La Janda. Eso sí en 2004 se acometen obras para un nuevo canal de desagüe que circula en su tramo finalparalelo al anterior y que hará posible drenar a mayor velocidad el vaso lagunar.
III.3. Régimen de la propiedadEn la actualidad la comarca de La Janda se encuentra sometida en su mayor parte a un uso agrícola intensivo dedicado ala explotación de cultivos de verano y ciclo corto; algodón, sorgo, girasol y plantas hortícolas son las plantaciones másusuales y abundantes. Esta orientación agrícola comenzó no obstante en la zona después de la desecación de la gran laguna,ya que el uso tradicional había sido hasta la fecha la explotación de la cabaña ganadera tanto de reses retintas como bravaal hacer uso de los frescos pastos de la zona. Este cambio reciente ha provocado una modificación importante en el régimende la propiedad de la tierra y en la actualidad son las grandes fincas agrícolas las posicionadas en el fondo lagunar. La antiguaactividad ganadera no ha desaparecido sin embargo en su totalidad y se mantiene en grandes superficies que aúnpermanecen poco o nada transformadas. En 1999 llegó el cultivo del arroz de forma extensiva a la Janda y desde entoncesy hasta la fecha extensas superficies han sido reconvertidas al cultivo de esta gramínea la cual y según informes de la Unión
Europea debe o bien adaptarse al campo del cultivo ecológico o la PAC no lo hará económicamente viable, sobre todo porla competencia en precios y mercados con los procedentes de Asia.
Unas 1.200 hectáreas se encontraban sumergidas por la lámina de agua y 2.500 estaban ocupadas por una abundante yespesa vegetación palustre, repartida mayoritariamente entre diferentes propiedades tales como Canteruelas, Derramaderosy La Mediana, las cuales representaban las zonas ecológicamente mejor conservadas. Hoy día muchas de ellas siguensumergidas y utilizadas para el cultivo del arroz (Recio y Dueñas, 2000)
IV. DATOS PARA LA RESTAURACIÓN ECOLÓGICA
Las indagaciones sobre las condiciones primitivas que aquí reinaron así como la evolución posteriormente experimentada
vienen apoyadas firmemente por un estudio detallado de los suelos y formaciones superficiales existentes, actuando estoscomo verdaderos testigos almacenadores de la información del pasado. Las variaciones y modificaciones experimentadaspor las antiguas topografías de los fondos lagunares se muestran también como una herramienta muy eficaz.
Dada la evolución geomorfológica reciente detectada en la zona y los cambios habidos en la dinámica fluvial, constituyenestos aspectos unos pilares básicos donde fundamentar las acciones futuras de restauración de las antiguas zonas palustres.Las más favorables antes citadas, parecen coincidir con las zonas menos transformadas agrícolamente estando por tantodedicadas en la actualidad a la explotación ganadera o bien al cultivo del arroz.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 183/276
Comunicaciones
183
V. PROBLEMAS DE CONSERVACIÓN
Si bien intrínsecamente este humedal presenta en primer lugar una gran problemática para ser restaurado, no deja de estaral margen sin embargo de grandes impactos ambientales ligados a la explotación agrícola actual o a las grandes obras deinfraestructura que pudieran realizarse en la zona.
Si pensamos desde este último punto de vista y en su parcial restauración, sería de sumo interés el efectuar una revisióny actualización de los regadíos así como establecer un nuevo planteamiento sobre la conveniencia o no de la construcciónde nuevos canales de drenajes o la de nuevos embalses en cabecera de los ríos (presas del Alamo y Aciscar, ambas enproyecto). Otras obras tales como el nuevo trazado de la carretera N-340 (Cádiz-Barcelona), el levantamiento de nuevostendidos eléctricos o la proliferación excesiva de aerogeneradores, parecen incidir también de una manera importante enel futuro de la zona.
Desde el punto de vista agrícola, entendemos que el uso abusivo de plaguicidas, herbicidas y de fertilizantes procedentesde las zonas sometidas a una explotación no ganadera, constituirían en cierta medida la gran amenaza para los futurosecosistemas. A estas habría que sumarles la construcción de drenajes artificiales a través de canales y acequias queimpedirían la acumulación del agua, ya que el posible uso de dicho recurso no ha de revestir grandes inconvenientes dadasu disponibilidad e intenso manejo que de el se realiza.
VI. CONCLUSIONES
A pesar de la existencia de estudios que cuantificaron la viabilidad y coste económico de la restauración de algunas zonasde La Janda, y tras los 10 años trascurridos desde el inicio de nuestros trabajos, aún no se han alcanzado o ejecutados ningunade las propuestas de restauración realizadas por entonces.
No obstante este grupo de investigación continúa actualizando sus conocimientos sobre el funcionamiento de estosecosistemas palustres, a pesar de no observar un reconocimiento de éstos por parte de la Administración. Finalmenteentendemos que la reciente constitución de la Agencia Andaluza del Agua y de la Confederación Hidrográfica delMediterráneo Andaluz una vez que asuman las competencias de las cuencas del Guadalete y del Barbate serán básicos paradar el impulso definitivo a la restauración de los humedales de La Janda.
VII. BIBLIOGRAFÍABERNALDO DE QUIROS, J.L. "Excursión ornitológica a La Janda (Marzo-Abril 1920)". Bol. Real Soc. Esp. His. Nat. XX: 236-248. 1920.BORJA, F. Cuaternario reciente, holoceno y periodos históricos del SW de Andalucía. Paleogeografía de medios litorales y fluvio-litorales de los últimos 30.000
años. Tesis doctoral. Fac. Geografía. Univ. Sevilla. pp. 469. 1992.CABRÉ, J. y E. Hernández-Pacheco. "Avance al estudio de las pinturas prehistóricas del extremo sur de España". Trabajos de la comisión de investigaciones
paleontológicas y prehistóricas, 3: pp. 35. 1914.CANO, M.D.; J.M. Recio, y F. Diaz del Olmo. "Soil formation in alluvial terrace of river Guadalquivir. The evidence of small Mendoza lakes (Andalusia.Spain)".Catena 20: pp. 63-71. 1993.CASTRO, J.C.; J.M RECIO. “Sobre el estado actual de la laguna de La Janda y su posible regeneración parcial”. In: Reunión sobre Medio Ambiente en Andalucía
I : pp. 61-66. Córdoba, 1990.CASTRO, J.C.; M.D. Cano, M.L. Torres, y J.M. Recio. “Recuperación parcial de la laguna de La Janda: acciones concretas en Los Derramaderos y Tapatana. (Cádiz,sur de España)”. In Jornadas Técnicas Internacionales: Bases Ecológicas para la Restauración de Humedales en la Cuenca Mediterránea. Public. Consejería deMedio Ambiente. Junta de Andalucía. Huelva. 1993.CASTRO, J.C.; I. Perevonznik, J.M. Recio. “Tirsificación y acciones antrópicas en la depresión de La Janda (Cádiz)”. In: VII Coloquio de Geografía Rural I : pp.145-152. Córdoba, 1994.CASTRO, J.C.; M.A. Dueñas, y J.M. Recio. "Bases ecológicas para el restablecimiento de humedales en la cuenca del río Barbate: la laguna de La Janda (Cádiz)".
Janda 2: pp. 137-156. 1996.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 184/276
Almoraima, 35, 2007
184
CHAPMAN, A., W. Buck. Unexplored Spain. 1910. Edición facsimil Incafo. 1978.CONSEJERIA DE MEDIO AMBIENTE. Bases ecológicas para la restauración de humedales en la cuenca mediterránea. Public. Consejería de Medio Ambiente.Junta de Andalucía. pp. 348. Sevilla. 1995.CONSEJERÍA DE MEDIO AMBIENTE. Proyecto: Restauración de Humedales de La Janda, Cádiz. Documento de actuación. Del. Prov. Cádiz. pp. 5. 2005.DIAZ DEL OLMO, F. y J.M. Recio. “Tierras Negras y cambio climático en Andalucía: edafogénesis durante el fín del Pleistoceno y el Holoceno”. In: WorkshopUmweltueranderungen in Holozän und aktuelle dynamik in Westandalusien. Eischtätt. Alemania. 1994.DUEÑAS LÓPEZ, M.A. & J.M. Recio Espejo. Bases ecológicas para la restauración de los humedales de la Janda (Cádiz, España) . Serv. Public. Univ.de Córdoba.pp. 475. 2000.FINLAYSON, J.C. Birds of the Straits of Gibraltar . T & A.D. Poyser. pp. 534. London. 1992.HERNÁNDEZ-PACHECO, E. "Las tierras negras del extremo sur de España y sus yacimientos paleolíticos". Trab. del Museo Nac. de CC.NN. de Madrid. Secc.Geología 13: pp. 3-24. 1915.IBARRA, P. Naturaleza y hombre en el sur del Campo de Gibraltar: un análisis paisajístico integrado . Public. Consejería de Medio Ambiente. Junta de Andalucía.pp. 490. Sevilla. 1993.IBARRA, P. “El entorno paisajístico de la depresión de La Janda”. In: Jornadas de Campo en la Depresión de La Janda. AEQUA-GAC I : pp. 40-57. 1995.
IRBY, L. The ornithology of the straits of Gibraltar . 2nd Ed. Taylor & Francis. London. 1895.LUQUE, L., C. Zazo, J.M. Recio, M.A. Dueñas, J.L. Goy, J. Lario, González-Hernández, F., Dabrio, C.J. y González-Delgado, A. "Evolución sedimentaria de la lagunade La Janda (Cádiz) durante el Holoceno". Cuaternario y Geomorfología, vol.13 (3-4): pp. 43-50. 1999.MAS, M. "La conservación del arte rupestre en las sierras del Campo de Gibraltar. Un primer diagnóstico". Espacio, Tiempo y Forma, serie I, Prehist. y Arqueol.7: pp. 93-128. 1994.MAS, M.; G. Torra, S. Ripoll, B. Gavilan, J.C. Vera, y J.F. Jorda. “El poblamiento prehistór ico en las sierras próximas a la antigua laguna de La Janda”. In : Jornadasde Campo en la Depresión de La Janda. AEQUA-GAC I : pp. 92-104. 1995.MOSQUERA, M.A.J. y J.C. Finlayson. “Ecología de las comunidades de aves de La Janda”. Jornadas de Campo en la Depresión de La Janda. AEQUA-GAC I : pp.4-15. 1995.RECIO, J.M., J.C. Castro y A. Santiago. Jornadas de Campo en la Depresión de La Janda. AEQUA-GAC. pp. 149. 1995.RECIO, J.M., J.C. Castro, y M.A. Dueñas. “Agricultura y medio ambiente en el sur de la Península Ibérica. El caso de la laguna de La Janda”. In: Arbeitstreffen
Landnutzungswandel und Umweltveränderungen in Spanien. Freiburg i. Br. 1996.RECIO, J.M., M.A Dueñas, y J.C. Castro. "La laguna de La Janda (Cádiz, España): bases para su restauración ecológica". Ecología, 11: pp. 125-142. 1997a.RECIO, J.M., M.A. Dueñas, y J.C. Castro. "Reseña histórica del proceso de desecación de la laguna de La Janda (Cádiz)". Almoraima, 18: pp. 95-111. 1997b.SAAVEDRA, E. La geografía de España del Edrisí . Imp. Fortanet. pp. 89. Madrid. 1881.
VERNER, W. My life among the wild birds in Spain. John Bale, Sons & Danielsson Ltd. London. 1909.VERNER, W. "Shotting, Historical notes". Gibraltar Chroniques XII : pp. 73-79. 1912.YATES, G. K. Bird life in two deltas . Faber & Faber. London. 1945.ZAZO, C., G. Obejero. "Niveles marinos cuaternarios en el litoral de la provincia de Cádiz". Trabajos sobre neógeno-cuaternario 5: pp. 141-145. 1976.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 185/276
Comunicaciones
185
ABSTRACT
The lesser kestrel (Falco naumanni), an endangered bird, breeds on the cliffs of the Rock of Gibraltar, at North Front. Aselsewhere throughout its range, this species has shown a considerable decline at Gibraltar throughout the twentieth century.We discuss probable reasons for this decline. We also discuss possible conservation measures and outline an action planfor the conservation of the lesser kestrel in Gibraltar.
Keywords: Lesser kestrel, Falco naumanni, Gibraltar, Decline, Conservation, Action Plan.
RESUMEN
El cernícalo primilla (Falco naumanni), un ave amenazada, anida en los acantilados de la cara norte del peñon de Gibraltar.Como en otras partes, esta especie conoció un considerable descenso en su población a lo largo del siglo XX. Consideramoslas posibles causas de estos cambios en población así como posibles medidas de conservación y desarollamos un plan deacción para conservar la especie en Gibraltar.
DECLINE OF THE LESSER KESTREL(FALCO NAUMANNI ) IN GIBRALTAR:POSSIBLE CAUSES, CONSERVATION AND ACTION PLAN
Keith J. Bensusan / John E. Cortés

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 186/276
Almoraima, 35, 2007
186
INTRODUCTION
The lesser kestrel (Falco naumanni) is a migratory falcon that breeds across the Palearctic, from Portugal to Manchuriain China, mainly between latitudes of 30º-55º (Cramp et al. 1980; Hollom et al. 1988; Tucker & Heath 1994). The specieswinters across almost the whole of subsaharan Africa, avoiding only the west african rainforests and the deserts of the hornof Africa in the north-east of the continent. However, most individuals winter in southern Africa, from Zimbabwe to SouthAfrica (Cade 1982; Tucker & Heath 1994). In addition, some of the populations of southern Iberia and north Africa aresedentary (Cramp et al. 1980). The lesser kestrel is a globally threatened species (Herediaet al. 1996; BirdLife International2000; Liven-Schulman et al. 2004) which is classified as Vulnerable by BirdLife International (Collar et al. 1994; Tucker& Heath 1994; BirdLife International/European Bird Census Council 2000).
Tucker & Heath (1994) cite factors such as food-loss due to agricultural intensification, pesticides, loss of foraging habitatsand loss of nest-sites as important factors that have led to the species’ decrease. Liven-Schulman et al. (2004) recentlysuggested that, in Israel, the main causes of reduction in lesser kestrel numbers are increased distance between breedingand hunting sites, and the use of pesticides in city parks and lawns. Parks and lawns cannot perhaps be considered to forman important global component of lesser kestrel foraging ground, but since pesticides are also applied readily to agriculturalareas this factor must still be considered important.
The Iberian peninsula is of particular importance to the lesser kestrel’s European status in that, with between 5.167-8.210pairs (BirdLife International/European Bird Census Council 2000), 44%-45% of european lesser kestrels breed in Iberia.Gibraltar’s lesser kestrel colony forms a very tiny component of that proportion; only five to ten pairs were given asbreeding on the Rock in 2000 (BirdLife International/European Bird Census Council 2000). The lesser kestrel has showna large decline in Gibraltar, which has mirrored that witnessed all over the species’ range. The species is protected by law
in Gibraltar under schedule 1 of the ‘Nature Protection Act, 1991’ (L/N 11 of 1991). This paper discusses probable reasonsfor the decline of the species of the Rock. We also outline possible conservation measures and recommend an action planfor the conservation of the lesser kestrel in Gibraltar which complements that proposed by Perez (2006).
METHODS & RESULTS
Data on population size has been gathered using older historical records (Rait-Kerr 1935; Cortés et al. 1980) and morerecent data taken from GONHS Bird Reports (Holliday 1995; Garcia 2001-2005) or GONHS records. In addition, Reid(1873) recorded that ‘the Lesser Kestrel breeds in numbers on the Rock, especially at the North Front…’. Similarly, Irby(1895) recorded ‘vast numbers’ breeding on the north face of the Rock, although he does not specify the exact number of
pairs. It is interesting to note that, in addition to breeding records, Cortés et al. (1980) mention that at the time that theirwork was published, some birds stayed to winter on the Rock, with numbers occasionally reaching 50 individuals. Thisis certainly no longer the case. Figure 1 summarises the number of pairs of lesser kestrel breeding at Gibraltar from 1935to the present. The number of breeding pairs at Gibraltar seems to have increased somewhat recently, although this apparentincrease could possibly be the result of more intensive surveying.
The increase in size of the town of La Linea de la Concepción in neighbouring Spain has certainly resulted in loss of foraginghabitat for Gibraltar’s lesser kestrels. These birds could, until about 15 years ago, be seen hunting at the Neutral Groundon a frequent basis (shown in figure 2). However, this area of open, lightly grazed ground became gradually smaller untilit disappeared altogether a few years ago. This has forced Gibraltar’s lesser kestrels to forage beyond the northern limitof La Linea, an increasingly urbanised area. This means that distance between the colony and it’s foraging grounds has

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 187/276
Comunicaciones
187
grown from 1-2km after the creation of the runway on the isthmus to over 5km at present. All of Gibraltar’s lesser kestrelsare now forced to hunt on the slopes of the Sierra Carbonera behind La Linea de la Concepción, and further north behindLa Alcaidesa ( pers. obs.; figure 2). Although much of Sierra Carbonera is property of the spanish military, rapid urbandevelopment is taking place on the lower eastern slopes of the sierra, around the area known as Santa Margarita. Likewise,the area of La Alcaidesa is likely to be threatened by development eventually. This places a new pressure on the lesserkestrels’ foraging ground.
DISCUSSION
Causes of Decline
Liven-Schulman et al. (2004) highlight increased distance between feeding grounds and nest-sites as a cause of decline inlesser kestrel populations. This is most likely to have impacted lesser kestrels breeding at Gibraltar. One further aspect isthe possible impact that feral pigeons (Columba livia domest.) may have on lesser kestrel colonies. These have now takensome of the nest-sites at North Front that were formally occupied by pairs of lesser kestrels. The medieval castle thatencapsulates Vieja Castellar harbours an until-recently thriving colony of lesser kestrels on its walls. The feeding groundsof this colony lie in the immediate vicinity of the colony, so that increased foraging distance from the colony cannot beregarded as a threat. Furthermore, the area in which these birds forage is composed of wild, grassy areas and Cistus matorral,an area in which no pesticides are applied. However, a reduction in breeding pairs has recently occurred at this site, andthis has coincided with an increase in nesting feral pigeons ( pers. obs.), apparently introduced at the site a few years ago.Aggression towards lesser kestrels from feral pigeons has been reported from this site (P. Acolina, pers. comm.).
Figure 1.Number of pairs of lesser kestrel breeding at Gibraltar from 1935 to the present. The apparent increase in recent years could be the product of more intensive sampling.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 188/276
Almoraima, 35, 2007
188
Conservation
The removal of feral pigeons from sites in and around North Front is highly desirable if we are to maximise the numberof potential nest holes for lesser kestrels. Tucker & Heath (1994) comment that the conservation of foraging habitat aroundexisting colonies must receive the highest priority and that careful land-use planning should ensure the avoidance of building developments in such areas. It is acknowledged that most of Gibraltar’s lesser kestrel’s traditional foraginggrounds have already been destroyed through urbanisation, and that in any case practically all of this land was/is on Spanishsoil and therefore out of the Gibraltar Government’s control.
One possibility for the conservation of this species in Gibraltar is that they begin to use the newly restored habitat on the greatsand slopes of the east side of the Rock. This area has recently been restored to its original habitat (Cortés et al. 2000). However,since there is an absence of grazers in the area, large sections of the great sand slopes are dominated by tall grasses. In Crauin France, a 50% reduction in the number of grazing sheep has resulted in taller and denser vegetation, reducing grasshopperpopulations and rendering them less accessible. Lesser kestrels in the area have decreased as a result (Tucker and Heath 1994).Non-intensive grazing of the site, as recommended by Tucker & Heath (1994), would be an important step towards the creationof a new foraging habitat for lesser kestrels on the Rock. Floral and invertebrate diversity would also benefit from such ameasure. Since erosion is a factor to consider with regard to the Sand Slopes, large grazers such as ungulates are unsuitable.Rabbits (Oryctolagus cunniculus) dig extensive burrows which might threaten the stability of the slopes. A smaller grazer thatis native to the hinterland, such a small, grazing rodent might be the best option. The Mediterranean pine vole ( Pitymysduodecimcostatus) is a candidate. Further investigations into the likely impact of voles at this habitat would be required beforesuch an introduction takes place. Large invertebrates that are found, or were found on the isthmus and talus slopes could bereintroduced to the great sand slopes, as these would provide food for foraging lesser kestrels.
Legislation
At present, the lesser kestrel is protected under schedule 1 of the ‘Nature Protection Act, 1991’ (L/N 11 OF 1991), whichprotects all bird species. Schedule 3 of the Act affords further protection to endemic and restricted-range taxa. This shouldbe extended to include globally threatened species such as the lesser kestrel. Several international conventions have beenratified by the United Kingdom on behalf of Gibraltar. These include the EC Habitats Directive (92/43/EEC), the EC BirdsDirective (79/409/EEC) and the Bonn Convention (Heath & Evans 2000). The Bonn Convention aims to protect migratoryspecies across the whole of their range. The EC Birds Directive aims to protect all birds within European territories, withparticular emphasis on Annex 1 species such as the lesser kestrel. The cliff at North Front lies on the northern boundaryof the Upper Rock Nature Reserve (L/N 51 of 1993). The cliffs of the Rock also receive protection as part of a Natura 2000site under the EC Habitats Directive, due in part to the presence of breeding lesser kestrels.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 189/276
Comunicaciones
189
Figure 2. Map of the northern end of the Rock of Gibraltar and the spanishhinterland, showing nesting site and past and present feeding sites of lesserkestrels breeding on the Rock. The white ellipse shows the location of the colonyof lesser kestrels on the north face of the Rock. The black ellipses show past(dashed line) and present (solid line) feeding grounds.
ACTION PLAN
This action plan is based on that given by Heredia et al.(1996), but alterations have been made to take specificaspects of Gibraltar’s lesser kestrel colony and theenvironment in which they live into account.
1. Aims
1.1. In the short term, to maintain the lesser kestrel breedingcolony at its present (2007) level or larger.
1.2. In the medium to long-term, to increase the population
size of the lesser kestrel in Gibraltar to a level at which it nolonger qualifies as a threatened species. For both of theseaims, habitat restorations, and breeding and reintroduction
programmes, are important prerequisites.
2. Objectives
2.1. Grazing levels on grassland – low density grazingshould be promoted in foraging areas of lesser kestrels inorder to allow for a high diversity of plants and invertebrates,and so that invertebrates are accessible. This seemsparticularly important on the great sand slopes (should an
introduction at this site take place), as large stands of tallgrasses dominate this site at present.
2.2. Promote the full legal protection of the species and important sites – It is recommended that the lesser kestrel beelevated to schedule 3 of the ‘Nature Protection Act, 1991’.
2.3. All range states should be encouraged to produce anational action plan for the Lesser Kestrel – as such, thisaction plan should be adopted by the Gibraltar Government’sMinistry for the Environment as the means with which toconserve the lesser kestrel in Gibraltar.
3. Species and Habitat Protection
3.1. Promote the designation of protected areas for the Lesser Kestrel – Landowners should be encouraged to manage theirland in a way that is beneficial to the well-being of our lesser kestrel colony, and to implement regular conservationmanagement. Landowners in Gibraltar are the Government of Gibraltar (sand slopes).
3.2. Protection of the colony from disturbance – accidental and deliberate disturbance of the colony should not occur.Industrial works taking place below the cliff face should take into account that a colony of a species of global concern existsat North Front. Similarly, tours of the tunnels within the Rock should take into account that nests are sometimes locatedclose to Jock’s Battery. Information should be supplied to all parties using the cliffs or the area immediately below theseat North Front, particularly during the breeding season.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 190/276
Almoraima, 35, 2007
190
4. Monitoring and Research
4.1. Surveys - breeding surveys should continue to take place at the lesser kestrel colony at North Front every year in orderto obtain an accurate picture of the species’ status in Gibraltar. This should continue to include counting the number of pairsat the colony, recording the location of each nest on the cliff and counting the number of fledglings at each nest.
4.2. Reintroduction and Recolonisation - since lesser kestrels can be bred and reared by the GONHS Raptor RehabilitationUnit, a possibility would be the establishment of a colony, using birds reared by GONHS, on the cliffs of the east side of the Rock. Habitat management would need to be considered before this takes place. A purpose-built wall with nest holesor strategically-placed nest boxes should also be considered. Any reintroduction or reinforcement/supplementationprogrammes should be carried out under the guidelines given by the IUCN/SSC Re-introduction Specialist Group (1998).Surveys, as described in 4.1., should be carried out on any newly formed colonies.
5. Public awareness
5.1. Raise awareness on the importance of lesser kestrel feeding habitats and colonies - Public awareness campaigns onthe plight of the lesser kestrel should be carried out, to include authorities, schools and the media. Being a large, attractiveanimal, the lesser kestrel should be used as a flagship species for the conservation and management of habitats. This shouldinclude breeding sites and foraging sites.
REFERENCESBirdLife International. Threatened Birds of the World. Barcelona and Cambridge, UK: Lynx Edicions and BirdLife International. 2000.BirdLife International/European Bird Census Council. European bird populations: estimates and trends. Cambridge, UK: BirdLife International (BirdLifeConservation Series No. 10). 2000.
CADE, T.J. The Falcons of the World. London, UK, 1982: Collins.CHEYLAN, G. "Le Faucon crécerellette (Falco naumanni) en France: statut actuel et regression". Faune de Provence (CEEP), 12: 45-49. 1991.COLLAR, N.J., Crosby, M.J. & Stattersfield, A.J. Birds to watch 2: the world list of threatened birds. Cambridge, UK, 1994: BirdLife International (BirdLifeConservation Series No.4).CORTES, J.E., Finlayson, J.C., Garcia, E.F.J. & Mosquera, M. The Birds of Gibraltar . Gibraltar, 1980: Gibraltar Books.CORTES, J.E., Linares, L. & Shaw, E. "The restoration of the vegetation of the east slopes of the Rock of Gibraltar: background and first steps". Almoraima, 23: 65-86. 2000.CRAMP, S., Simmons, K.E.L., Gillmor, R., Hollom, P.A.D, Hudson, R., Nicholson, E.M., Ogilvie, M.A., Olney, P.J.S., Roselaar , C.S., Voous, K.H., Wallace, D.I.M.& Wattel, J. Handbook of the birds of Europe, the Middle East and North Africa: The Birds of the Western Palearctic . Oxford, UK, 1980: Oxford University Press.GARCIA, E.F.J. The Systematic List 2001. Gibraltar Bird Report , 1: 8-27. 2001.GARCIA, E.F.J. The Systematic List 2002. Gibraltar Bird Report , 2: 11-36. 2002.GARCIA, E.F.J. The Systematic List 2003. Gibraltar Bird Report , 3: 13-39. 2003.GARCIA, E.F.J. The Systematic List 2004. Gibraltar Bird Report , 4: 13-39. 2004.GARCIA, E.F.J. The Systematic List 2005. Gibraltar Bird Report , 5: 13-39. 2005.GIBRALTAR GAZETTE . "Nature Protection Act", First Supplement to the Gibraltar Gazette No. 2608, 9th May 1991 Gibraltar (L/N 11 of 1991). 1991.GIBRALTAR GAZETTE . "Nature Conservation Area (Upper Rock) Designation Order", First Supplement to the Gibraltar Gazette No. 2717, 25th March 1993Gibraltar (L/N 51 of 1993). 1993.
HEATH, F. & Evans, M.I. eds. Important Bird Areas of Europe, Priority Sites for Conservation Vol. 2, Southern Europe. Cambridge, UK, 2000: BirdLife International(BirdLife Conservation Series no. 8).HEREDIA, B., Rose, L. & Painter, M. Globally threatened birds in Europe: Action plans. Germany: Council of Europe. 1996.HOLLIDAY, S.T. "Report on the birds of the Strait of Gibraltar: 1989". Alectoris, 9: 1-44. 1995.HOLLOM, P.A.D., Porter, R.F., Christiensen, S. & Willis, I. Birds of the Middle East and North Africa. Calton, UK, 1988: T. & A.D. Poyser.IRBY, L.H. The Ornithology of the Straits of Gibraltar , Second Edition. London, UK, 1895: Taylor& Francis.IUCN/SSC Re-introduction Specialist Group. IUCN Guidelines for Re-introductions. Cambridge, UK, 1998: IUCN.LIVEN-SCHULMAN, I., Leshem, Y., Alon, D. & Yom-Tov, Y. "Causes of population declines of the Lesser Kestrel Falco naumanni in Israel". Ibis, 146: 145-152. 2004.PARR, S. & Naveso, M. Interim report on Lesser Kestrel research in central Turkey . Unpublished. 1994.PEREZ, C.E. Biodiversity Action Plan, Gibraltar: Planning for Nature. Gibraltar: Gibraltar Ornithological and Natural History Society (GONHS). 2006.RAIT-KERR, H. "The birds of Gibraltar". Articles in the Gibraltar Chronicle. 1935.REID, P.S.G. Stray notes on ornithology. A record of ornithological observations in Gibraltar, Bermuda, Great Britain and Ireland, New York, South Africa, Natal,the Canary Islands & Spain. Unpublished. 1871-1890.TUCKER, G.M. & Heath, M.F. Birds in Europe: their conservation status. Cambridge, UK, 1994: BirdLife International (BirdLife Conservation Series No. 3).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 191/276
Comunicaciones
191
1. INTRODUCCIÓNCualquier cazador, gestor de coto, naturalista o, simplemente, aficionado al campo, podría plantearse la pregunta, trasrecorrer las sierras gaditanas…, "si el corzo es la especie típica de aquí, ¿por qué hay tan pocos? ¿Cuántos podría haberen esta finca?".
Creemos que esta es una de las preguntas básicas que un buen gestor cinegético se debe plantear, pero, ¿cómo establecemoscuántos corzos pueden existir en una finca concreta?
Dedicar tiempo a estas cuestiones no es banal, ya que el corzo (Capreolus capreolus) se considera como especie de especialinterés en los planes de ordenación de la Comarca, y ya existen algunos estudios que tratan de responder a estas preguntas.Algunos de estos son:
- Plan Comarcal de Ordenación Cinegética del P.N. de los Alcornocales. Uno de los objetivos prioritarios es restaurar laspoblaciones de corzo, considerándola como la especie de mayor interés y más genuina del parque natural (Sánchez, 2005)
- Evaluación de las poblaciones de corzo en Andalucía, bajo el marco del Plan Andaluz de Caza Mayor, puesto en marchapor la Consejería de Medio Ambiente. Este estudio muestra un incremento de las poblaciones de este pequeño unguladoen los últimos años en Cádiz, que parece estar relacionado con la disminución de las poblaciones de ciervo, así comocon las mejoras realizadas en el hábitat.
- PORN y PRUG del Parque Natural Los Alcornocales. Ambos documentos normativos consideran al corzo como laespecie cinegética más adecuada en este parque al ser el herbívoro con un impacto menor sobre la vegetación frente aotras especies cinegéticas más dañinas para la regeneración del alcornocal, y encontrar aquí una de sus distribucionesmundiales más meridionales.
ESTABLECIMIENTO DE UNADENSIDAD ÓPTIMA PARA EL CORZO
EN UNA FINCA DEL PARQUE NATURALDE LOS ALCORNOCALES
Alejandro Vega Contreras / Ricardo Zamora Diaz / Departamento de Ingeniería Forestal ETSIAM, Córdoba
Eduardo Briones Villa / Biólogo, Ayuntamiento de los Barrios

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 192/276
Almoraima, 35, 2007
192
- El corzo en Los Alcornocales, Mapa de calidad de hábitats. Publicación desarrollada dentro del Programa de MedidasCompensatorias con motivo de la construcción de la Autovía Jerez-Los Barrios. Aquí se evidencian relaciones entre laabundancia de corzos y las características concretas de los distintos hábitats representativos del Parque Natural LosAlcornocales (Braza y otros, 2004).
- Estación de referencia del corzo andaluz. Centro de coordinación, certificación y divulgación del corzo andaluz comoun ecotipo de elevado interés en el ecosistema mediterráneo, así como una pieza de caza valiosa y singular. (Oliverosy S. José, 2005)
Es decir, como además confirma la creciente afición de los cazadores por la especie, el corzo de Los Alcornocales(Capreolus capreolus) se ha convertido en una especie con un gran atractivo cinegético y muy valioso desde el punto devista ecológico.
OBJETIVOS Y METODOLOGÍA
Establecemos como objetivo conocer, para una adecuada gestión, cuál puede ser la población óptima, en equilibrio con elmedio, en una finca concreta. Para ello aplicamos los últimos estudios científicos, modificándolos en lo necesario para suaplicación en la zona concreta de estudio, y utilizamos herramientas informáticas, como los sistemas de informacióngeográfica, para mejorar tanto el procesamiento de la información como la presentación de los resultados.
Con la realización de este trabajo, se pretende (y creemos que se consigue) calcular una densidad óptima para la poblaciónde corzos en la finca Hoyo de Don Pedro, propiedad del Ayuntamiento de Los Barrios, incluida dentro del parque natural,así como establecer las medidas necesarias, siguiendo las directrices del parque natural, para que esta población sea establesegún los usos en esta finca.
Para ello:
- Analizamos la información existente sobre Hoyo de Don Pedro.
- Creamos un mapa de coberturas del suelo, de acuerdo a los últimos estudios.
- Estimamos la densidad actual de corzo en la finca.
- Calculamos el número total de corzos que podría existir en la finca en el estado actual.
Establecemos las directrices sobre los tratamientos silvícolas y otras mejoras para aumentar la población de corzos ymejorar su gestión.
CARACTERÍSTICAS DEL ÁREA DE ESTUDIO
La finca Hoyo de don Pedro pertenece al Ayuntamiento de Los Barrios, provincia de Cádiz, y está incluida dentro del ParqueNatural Los Alcornocales, perteneciendo a la formación orográfica de sierra La Palma.
Posee una extensión de 714,43 Ha Su altitud va desde los 76 m en su base a los 583 m en su cima, con una media de 347 m.
Posee pendientes medias de 21% y máximas de 52%. La orientación principal es SE-E.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 193/276
Comunicaciones
193
La precipitación media es de 1.150 mm, con una distribución bastante irregular, siendo el invierno la estación más lluviosa,seguida del otoño.
Las temperaturas son suaves durante todo el año, con medias mensuales que van de 4,6ºC en diciembre a 24,3ºC en agosto.
Destacan sus vaguadas con dirección oeste-este, lo que provoca que este monte esté expuesto a los vientos de levante. Esteviento, como se sabe, procedente del Mediterráneo, viene cargado de humedad y durante el verano atenúa la sequía estivalal condensar, por efecto de la altitud, la humedad atmosférica, originando nieblas y precipitaciones horizontales importantes.
Presenta tres gargantas principales por las cuales discurre agua durante todo el año, aunque con caudal algo irregular(garganta de Benharas, del Prior, y del Capitán o Botafuegos).
DESARROLLO DEL MÉTODO
A pesar de que hay estudios científicos que nos indican cómo se puede calcular el número de corzos en el parque natural(Braza y otros, 2004), las adaptaciones para poder aplicar estos, en una zona concreta del parque, se basarán en elconocimiento del medio y en los fundamentos de los estudios realizados.
El mapa de coberturas que creamos para Hoyo de Don Pedro se basa en la clasificación propuesta por Braza (2004), basadoen grupos de vegetación, por composición botánica, del parque.
Para establecer el número actual de corzos en la finca, y según recomienda el estudio Proyecto de Seguimiento dePoblaciones Cinegéticas (Consejería de Medio Ambiente de la Junta de Andalucía, 1999-2004), el método utilizado es elde “estima de abundancia por transectos de indicios”.
Para poder aprovechar los datos obtenidos en años anteriores, se contó con la colaboración de la misma persona encargadadurante estos años de recoger los datos.
Zonificación del monte
Para definir el mapa de coberturas de la finca nos apoyamos en datos de campo obtenidos desde la primavera de 2004 hastael inicio de la misma en 2005.
El trabajo se basó en fotografía digital y visitas continuadas a campo, para definir aún más los rodales. Con esta información,en gabinete, y mediante el programa Arc-View, terminamos de definir el mapa y calculamos las áreas de cada tipo de hábitat.
Para Hoyo de Don Pedro definimos los siguientes tipos de hábitat:
QuejigalÁreas con más del 75% de quejigo andaluz o moruno (Quercus canariensis).
Constituye una de las formaciones forestales más singulares del parque, formando un bosque marcescente rico enespecies epifitas y lianas (principalmente hiedra: Hedera helix).
Ocupa las zonas más húmedas, en el fondo de los valles, gargantas y las umbrías. La presencia de helechos (Polystichumsetiferum) denota un buen estado de conservación.
El sotobosque está compuesto por espino, durillo, agracejo, acebo, brezo cucharero y escobón.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 194/276
Almoraima, 35, 2007
194
Lo más común es que aparezcan masas adultas con el suelo despejado, con una gruesa capa de hojarasca y especiesherbáceas mesofíticas (de sitios húmedos) como ficaria, vinca, gladiolo y cebolla albarrana.
Se trata de una de las zonas más apetecibles para el corzo, especialmente cuando el estado de madurez de la masa permitela presencia de sotobosque.
Alcornocal
Masas con más del 75% de alcornoque (Quercus suber ).
Es el tipo de bosque más abundante en la zona, crece sobre sustratos ácidos, en localidades con precipitaciones superioresa los 700 mm anuales.
El sotobosque puede estar formado por especies muy diversas, dependiendo de la exposición (solana-umbría), laposición topográfica y la intensidad de perturbaciones (incendios, desmontes).
Algunas de estas son: madroños, brezo blanco, helechos, durillos (en umbrías) y jaguarzo, brezo común o brecina,escobón blanco, jerguen, labiérnagos, brezo de escobas, en solanas.
Estas zonas son bastante buenas para el corzo aunque no llegan a alcanzar el potencial del quejigal, debido a la menorpalatabilidad de las especies presentes en este hábitat, por su menor humedad.
Ojaranzal
Constituye la vegetación más original del parque (bosques relícticos de laurisilva bien conservados). Se desarrollan entramos altos de gargantas y en altiplanicies con suelos hidromorfos, o bajo régimen constante de nieblas.
Este microclima de humedad permanente y pobreza en nutrientes del sustrato de arenisca han favorecido la existenciade esta comunidad vegetal peculiar y diversa.
Aparecen asociados al ojaranzo ( Rhododendrum ponticum) especies como avellanillo, laurel y unas 30 especies dehelechos.
Nos encontramos en el hábitat con un mayor potencial para sustentar poblaciones de corzo.
Herriza
Con este término local se denominan los brezales densos y de escaso porte, generalmente desprovistos de cubierta arbórea.Se desarrollan sobre suelos poco evolucionados, pobres en nutrientes, de acidez elevada y con niveles elevados de aluminiosoluble. Aparecen, principalmente, en las cumbres y crestas rocosas de las formaciones de areniscas oligo-miocénicas de
las sierras del Aljibe (Ojeda y otros, 2002), mayoritariamente incluidas en el Parque Natural Los Alcornocales.
Las plantas leñosas dominantes en estas comunidades son la robledilla (Quercus lusitanica), brezos (Calluna vulgaris,Erica australis y E. scoparia), jaras (Cistus populifolius, Halimium alyssoides) y las aulagas (Genista spp., Stauracanthusboivinii).
A pesar de no aparecer normalmente con cubierta arbórea alguna, es de alto valor para el corzo.
Acebuchal
Masas con mas del 75% de pies de acebuche (Olea europaea ssp. oleaster ), suele estar entremezclado con alcornocal,o aparecen únicamente acompañados de herbazales, como en el caso que nos ocupa.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 195/276
Comunicaciones
195
Se presentan sobre suelos margosos en la base de las sierras silíceas (como Hoyo de Don Pedro).
No suelen ocupar grandes extensiones, pero su presencia para el corzo es interesante.
Estos serán, en principio, los tipos de hábitat que entran a valorar la población de corzos en Hoyo de Don Pedro; por elcontrario, los siguientes tipos de vegetación serán tomados como áreas a descontar del total de la superficie de la finca.Aunque pensamos que es simplificar la situación real, para poder aplicar el modelo predictivo, dichas áreas no intervendrán.
Pinar
Masas adultas, con más de 4 m de altura y más del 75% de Fracción de Cabida Cubierta (FCC) de pies de Pinus spp.Casi todos son Pinus pinaster (negral), excepto un pequeño rodal de P.pinea (piñonero).
Son masas que, por sí solas, sin subpiso arbustivo, no son demasiado atractivas para el corzo, pero que a medida quela FCC disminuye y aumenta la insolación en el suelo se van transformando en atractivas para este, (la variedad deespecies en la herriza aumenta, así como su densidad, [Correa y otros, 2000]), aunque no llegan a tener el potencial deotros hábitats como el alcornocal.
Regenerado de pinar
Zonas donde se ha establecido un regenerado natural de Pinus pinaster , principalmente tras el incendio del año 1997.
Aunque en el mapa general de la finca no se ha hecho diferenciación alguna dentro de este tipo, sería adecuadodiferenciar dos tipos de regenerado de pinar:
- Regenerado con FCC superior al 75% y altura media superior a 2,5 m. Se le asigna un valor potencial para la
población de corzo de 0.- Regenerado con FCC inferior al 75% y altura media inferior a 2,5 m. Aparece mezclado con herriza, por lo que
sería posible asignarle un valor potencial para la población de corzo mayor de 0.
Eucaliptal
Zonas con mas del 50% de eucalipto, cuyo destino, según lo previsto por los gestores del monte, es la eliminación ysustitución por otra especie autóctona adecuada a la estación, pero de las cuales no tenemos datos de la capacidadsustentadora para poblaciones de corzo, aunque suponemos será baja.
Se le asigna valor 0 hasta que la nueva especie esté asentada y el eucalipto eliminado por completo.
Pastizal
Se definen como tal los herbazales de suelos margosos de las bases de las sierras silíceas.
Podríamos decir, por las especies que aparecen, que estos son pastizales de bujeo, con leguminosas anuales, comotréboles y carretones (Trifolium spp., Medicago spp.) en zonas menos pastoreadas, y cardos, tagarninas y esparraguerasen zonas con mayor presión ganadera.
El valor potencial para el corzo dependerá de la distancia a masas arbóreas o arbustivas y de qué tipo sean éstas, ya queel corzo utiliza los herbazales siempre que estén intercalados con estratos superiores que le proporcionen refugio.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 196/276
Almoraima, 35, 2007
196
Improductivos:
Como improductivos definimos aquellas zonas compuestas por afloramientos rocosos, caminos y pequeños cercadoscon carácter científico.
Son zonas con valor 0 para el corzo.
Bosque de ribera
Se desarrolla en el tramo intermedio-bajo de las gargantas. En Hoyo de Don Pedro solo se puede decir que aparece enel tramo inferior de la garganta del Capitán.
Las especies principales son alisos ( Alnus glutinosa), sauces (Salix pedicellata, S. atrocinerea), arraclán ( Rhamnus
frangula)..., especies típicas de la parte baja de los canutos gaditanos.
Estimación de la densidad actual
Debido, principalmente, a que el corzo en esta comarca es una especie especialmente esquiva (aquí se le llama “el duendedel bosque”) y a la dificultad del terreno para la observación directa, el método seleccionado para la estimación de ladensidad actual es el de transectos de indicios.
El método consiste en colocar transectos en cada tipo de hábitat a muestrear.
Estos transectos son líneas rectas de 500 m trazadas en el terreno, en las cuales, cada 100 m, comenzando en el inicio delrecorrido, se coloca una unidad básica de muestreo denominada “placeta”. Esta consiste en una superficie circular de 5mde radio donde se rastrearán los indicios de presencia del corzo, principalmente sus excrementos.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 197/276
Comunicaciones
197
Los transectos se revisan cada mes para evitar, si se quisiera aumentar la frecuencia de la revisión, cambiar las costumbresdel corzo y, asimismo, procurar que no se pierdan los rastros por efecto de la climatología. Una vez registrados, losexcrementos contados se eliminan para no influir en próximas revisiones.
Para obtener la densidad se utiliza la siguiente fórmula, teniendo en cuenta que estas densidades serán por tipo de hábitatmuestreado, por lo que, posteriormente, la densidad final se obtendrá por ponderación de las densidades parciales.
D = Nº grupos de excrementos / Ha
Nº días entre revisiones x tasa de defecación diaria
En Hoyo de Don Pedro
Durante los años comprendidos entre 2000 y 2004 se hanrevisado cuatro transectos en las unidades quejigal, alcornocaly herriza.
Tras calcular la media de todos los datos de cada transecto yponderarlas con la superficie de cada hábitat muestreado,encontramos que en la finca existe una densidad media de6.887 corzos/100 Ha, lo que significa una población aproxi-mada de 34 corzos en la finca.
Estimación de la densidad potencial
Bajo esta denominación entendemos el tamaño de la pobla-ción de corzos que puede sustentar esta finca (en su particularlocalización), con esta vegetación, con las particulares condi-ciones de usos y en equilibrio con el medio.
Nos basaremos en la ecuación de predicción definida porBraza y otros (2004), en El corzo en los alcornocales, mapa decalidad de hábitats.
Antes de aplicar esta ecuación hay una serie de condicionantesque hay que tener en cuenta:
En el diseño de los coeficientes de abundancia de corzo según hábitat, Braza y otros (2004) analizaron las siguientes
variables: Vegetación, altitud, tipos de suelo, pH del suelo y distancias a vías principales.No todas estas variables son cuantificadas finalmente en la ecuación de predicción. Tras el estudio, dichos autoresdemostraron que con tan solo altitud y vegetación se puede construir un modelo matemático significativo que indique lacantidad de corzos.
La ecuación de predicción está diseñada para el conjunto del parque, por lo que las peculiaridades de cada finca deben sertenidas en cuenta en cada aplicación concreta.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 198/276
Almoraima, 35, 2007
198
Si no fuese así, la ecuación de predicción, por lo general, en zonas eminentemente corceras, puede infravalorar la población,(Vega, “en realización”).
En Hoyo de Don Pedro, si se aplica la ecuación de predicción, se tiene:
Nº de corzos en una finca = (CE alcornocal - C altitud) x sup. alcornocal + (CE quejigal - C altitud) x sup. quejigal +(CE quejigal basófilo) x sup. quejigal basófilo + (CE herriza - C altitud) x sup. herriza +(CE acebuchal - C altitud) x sup. acebuchal + (CE ojaranzal - C altitud) x sup. ojaranzal
CE alcornocal = 10,28 Series Superficie (ha)CE quejigal = 16,30 Alcornocal 168'12CE quejigal basofilo = 13,24 Q. moruno 70'15
CE herriza = 12,37 Herriza 257'09CE acebuchal = 10,13 Acebuchal 5'69CE ojaranzal = 20,25 Ojaranzal 3'92
C altitud = 0,017x altitud media de la finca = 0,017x347 = 5,899
Nº corzos = (10,28 – 5,899) x 1,6812 + (16,30 - 5,899) x 0,7015 +(12,30 – 5,899) x 2,57 + (10,13 - 5,899) x 0,0569 +(20,25 - 5,899) x 0,0392 = 32 corzos
Como podemos observar, los datos obtenidos de la revisión de los transectos de indicios (34 corzos) se asemejan bastantea los cálculos de la ecuación de predicción (32 corzos), lo que indica un ajuste matemático muy bueno, pero si aceptamosestos datos como definitivos estaríamos aceptando que las condiciones en Hoyo de Don Pedro serían las de la media delparque (en cuanto a climatología, usos de la finca, etc.), lo cual no es cierto, (Vega, “en realización”).
Por tal motivo, nos planteamos adecuar la ecuación de predicción a Hoyo de Don Pedro, sin restar su validez general. Atal efecto, nos basamos en los fundamentos de todas las variables estudiadas para definir esta ecuación. Tras un análisispudimos ver que:
- La base de la abundancia de corzo, el elemento que más influye sin estar presente, es el agua “natural”. Esto quedademostrado tanto en el coeficiente de altitud (más penalización mientras más altitud, al haber menores caudales a mayoraltura, Braza y otros, [“Los corzos, pues, muestran una preferencia por las altitudes bajas independientemente de lavegetación, probablemente debido a que en estas zonas bajas es donde se encuentran los lugares mas húmedos yprotegidos.”]; como en la vegetación (mayores densidades máximas en hábitats más húmedos: ojaranzal, quejigal,…).
- Aunque no entra directamente en la valoración, la distancia a vías principales, es decir, las perturbaciones producidaspor el hombre, sí pueden ser considerables, por lo que nosotros estimamos que a cotas demasiado bajas (donde el hombretiene más actividad, movimiento con maquinaria, de ganado,…) la penalización debe incrementarse progresivamentehacia abajo también, a partir de una cota determinada.
- En zonas con una alta precipitación horizontal (como es nuestro caso), la cobertura arbórea es de vital importancia. Lacaptación de esta es mayor con una buena cobertura.
Teniendo en cuenta estos fundamentos, y volviendo a observar el mapa de coberturas diseñado especialmente para estetrabajo, podremos darnos cuenta que existe una franja intermedia altitudinal que no tiene sentido que tenga penalización,ya que las condiciones son las idóneas para que las densidades que se puedan dar sean las máximas diseñadas para cadahábitat, (Vega, “en realización”).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 199/276
Comunicaciones
199
Pasamos, pues, a aplicar la ecuación de predicción introduciendo modificaciones para adecuar el estudio a Hoyo de Don Pedro:
Modificaciones de la ecuación de predicción para Hoyo de Don Pedro (Vega, “en realización”):
Coeficiente C de altitud.
Nuestra propuesta consiste en sustituir en su cálculo la altura media por un factor, función de una franja intermedia en lacual no existe penalización por altura.
Este factor lo que hace es aplicar el coeficiente penalizador definido por Braza y otros (2004) fuera de esta franja intermediaóptima. El valor del nuevo factor se halla ponderando las superficies, tanto por encima de la cota superior óptima, comopor debajo de la cota inferior óptima, que tiene la finca.
Esta franja intermedia óptima la hemos establecido entre las cotas de 76m y 400m, ya que es entre estas cotas dondeencontramos corrientes de agua durante todo el año, así como una cubierta arbórea y arbustiva, densa y de calidad para elcorzo (ojaranzal y quejigal).
La ecuación tendría ahora esta expresión general para el factor de altitud:
0,017 (P1(/ Cota superior finca – Cota”óptimo superior”/) + P2(/Cota inferior finca – Cota”óptimo inferior”/)) + P3
Siendo:
P1: porcentaje de superficie de la finca por encima de la cota “óptimo superior”.
P2: porcentaje de superficie de la finca por debajo de la cota “óptimo inferior”.
P3: porcentaje de superficie de la finca entre cota “óptimo superior e inferior”. Al ser la franja óptima, P3 no está penalizado,o, lo que es lo mismo, su factor de penalización es el mínimo, es decir, 1.
En cuanto a la vegetación, se han hecho las siguientes modificaciones:
Acebuchal
Como acebuchal definimos aquellas zonas propiamente de acebuchal (masas arbóreas adultas cerradas), más la zona depastizal influidas por la cobertura arbórea que este acebuchal proporciona.
Herriza incluida en regenerado de P. pinaster .
Se incluye en la superficie de herriza del mapa de coberturas un 70% de la superficie de regenerado de P. pinaster con FCCinferior al 75% y altura media inferior a 2,5 m.
Cálculos propuestos
P1 = 32,25% Cota superior finca = 583mCota “óptimo superior” = 400m
P2 = 4,48% C altitud = 0,017mCota inferior finca = 76m
P3 = 62,26% Cota “optimo inferior” = 150m
C altitud ponderado = 0,017(/583 – 400/ x 0,3325 + /76-150/ x 0,0448) + 0,6026 = 1,693

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 200/276
Almoraima, 35, 2007
200
Nº de corzos = (10,28 - 1,693) x 1,681 + (16,3 - 1,693) x 0,7015 + (12,37 - 1,693) x 2,805 + (10,13 - 1,693) x 0,093 + (20,25- 1,693) x 0,0392 = 56 corzos.
Contrastada esta cifra con expertos conocedores de la finca, parece que se ajusta mejor al valor potencial de poblacióncorcera de Hoyo de Don Pedro.
DISCUSIÓN
La ecuación de predicción modificada y la abundancia estimada por los transectos indican que es posible aumentar lapoblación de corzos actual de Hoyo de Don Pedro, pasando de una densidad media actual de 6,88 corzos / 100 Ha (34 corzos)a una densidad media potencial de 10,73 corzos / 100 Ha (56 corzos).
Las densidades máximas que se pueden dar en esta finca, manteniendo un equilibrio entre la población de corzos y lavegetación son:
Densidad en alcornocal = 8,58 corzos / 100 Ha
Densidad en quejigal = 14,6 corzos / 100 Ha
Densidad en herriza = 10,67 corzos / 100 Ha
Densidad en acebuchal = 8,44 corzos / 100 Ha
Densidad en ojaranzal = 18,56 corzos / 100 Ha
El hábitat menos aprovechado por la población actual es la herriza, por lo que manteniendo la clasificación actual (sin
intentar cambiar la herriza por otro hábitat mas favorable para el corzo), las mejoras deben ir enfocadas sobre esta superficiey sobre el área de regenerado de P. pinaster (mezclado con herriza).
Se hace necesario establecer una serie de mejoras en la finca para que la población de corzos por si sola alcance su óptimo.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 201/276
Comunicaciones
201
PROPUESTA DE INTERVENCION Y DE MEJORAS
Tratamientos sobre la vegetación
Aprovechando el mapa de coberturas diseñado, y con un mapa de pendientes de la finca, se pueden seleccionar lassuperficies a tratar. Proponemos las siguientes actuaciones:
- Aclareos de la superficie clasificada como regenerado de P. pinaster , respetando especies de interés. Se pueden admitirdos grados de intensidad: baja intensidad, en zonas con una pendiente mayor del 25%, y media intensidad, en zonas conpendiente menor del 25%.
- Creación de pequeñas zonas de pastizal y de rebrotes leñosos (roza entre dos tierras y a 0,5m, de forma alternativa), enzonas de regenerado de P. pinaster y de herriza, con pendiente menor del 15%, asegurando la presencia de manchasde cobertura arbórea cercana y aisladas del tránsito, tanto de vehículos como de personas.
- Repoblaciones pequeñas de alta densidad (bosquetes) en zonas de acumulación de suelo por escorrentías superiores, quehayan sido afectadas en años anteriores por incendios, con protección de los vientos de levante, y en zonas clasificadas comoherriza o regenerado de P. pinaster . Estas se harán con alcornoque y con quejigo andaluz en zonas especialmente húmedas.
Todos estos tratamientos se harán adecuando las fechas al ciclo biológico del corzo, especialmente la creación de zonasde pastizal y los aclareos, para reducir, en lo posible, el estrés a la especie. Lo ideal es que dichos tratamientos selvícolasse integraran en un plan de ordenación del monte.
Zonas de baja pendiente ideales para repoblaciones y creación de praderas, sobre herriza (azul) y regenerado de pinaster (verde).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 202/276
Almoraima, 35, 2007
202
Mejoras en la finca
- Mejora de la guardería (mayor formación y reducción de la zona de vigilancia).
- Mallado de la parte inferior de la finca con cercado ganadero y eliminación de cercados antiguos sin posibilidad dereutilización.
- Establecimiento de tres bebederos-comederos, no como cebaderos, sino para suministrar desparasitantes, sobre todo enverano y para realizar conteos-avistamientos con facilidad. Se colocarán en masas arbóreas adultas, con pocosotobosque y lo más alejados posible de las corrientes de aguas naturales.
- Establecimiento de áreas preventivas de defensa contra incendios forestales, que además de su función principal contraincendios forestales facilite en alguna zona el rececho al corzo, así como el aumento de pastizales naturales y rebrotes.
- Mejora de la accesibilidad a la finca, siempre que venga acompañada de una mejor vigilancia.
- Control de predadores/competidores. Éste se centrara principalmente en mantener las densidades actuales de ciervo enla finca ( muy por debajo de la media en el parque natural debido a las especiales características de la misma ), y disminuiren lo posible las de zorro y cochino asilvestrado.
BIBLIOGRAFÍABRAZA, F., San José, C. y López, M. El corzo en Los Alcornocales: mapa de calidad de hábitats. Estación Biológica de Doñana-Sevilla y Consejería de Obras Públicasy Transporte, Gestión de Infraestructuras de Andalucía, S.A. 90 pp. 2004.CONSEJERÍA DE MEDIO AMBIENTE. Informe sobre las poblaciones de ciervo y corzo en Andalucía. Junta de Andalucía, Sevilla. 2002. 18 pp.CONSEJERÍA DE MEDIO AMBIENTE. Plan de Ordenación Cinegética del Parque Natural Los Alcornocales. Junta de Andalucía, Sevilla. 2002.
CONSEJERÍA DE MEDIO AMBIENTE. 2004. Informe sobre las poblaciones de ciervo y corzo en Andalucía. Junta de Andalucía, Sevilla. 2002. 9 pp.CORREA, R. M., Sánchez-Castilla, E., y Ojeda, F. “Efecto de los pinares de repoblación sobre la biodiversidad y estructura de las herrizas en el Parque Natural deLos Alcornocales”. Almoraima, 23: pp. 183-188. 2000.OJEDA, F. “La Herriza: La cenicienta del Parque Natural”. Almoraima, 27: pp. 145-148. 2002.OLIVEROS, F., y San José, C. “La estación de referencia del corzo andaluz”. Almoraima, 31: pp. 181-187. 2005.SÁNCHEZ GARCÍA, J. M. “La Ordenación Cinegét ica Comarcal Del P.N. Los Alcornocales: Resultados tras doce años de gestión”. Almoraima, 31: pp. 169-179. 2005.VEGA, A. Estudio de sustentabilidad de las poblaciones de corzo (Capreolus capreolus) en el monte Hoyo de D. Pedro, término municipal Los Barrios (Cádiz).T.P.F.C. ETSIAM. Universidad de Córdoba. (en realización).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 203/276
Comunicaciones
203
RESUMEN
Se ha tratado de determinar la presencia y distribución de las especies de mamíferos carnívoros en el término municipalde Los Barrios. Para ello se han utilizado cinco métodos de obtención de información; muestreos directos (aguardos yrecorridos en vehículo, diurnos y nocturnos), muestreos indirectos (transectos para determinación de indicios, huellas yseñales), fototrampeo, encuestas y bibliografía.
Se han detectado siete especies de carnívoros mediante los tres primeros métodos; zorro ( Vulpes vulpes), tejón ( Melesmeles), meloncillo ( Herpestes ichneumon), nutria ( Lutra lutra), garduña ( Martes foina), jineta (Genetta genetta) y
comadreja ( Mustela nivalis). Se ha conseguido información actual o histórica de otras cuatro especies mediante los dosúltimos; gato montés (Felis silvestris), turón ( Mustela putorius), lince ( Lynx pardinus), y lobo (Canis lupus).
El zorro, la jineta y el meloncillo son las especies que aparecen mejor distribuidas y que presentan densidades más altas,aunque hay otras especies como la nutria y el tejón que, siendo menos esperable, también alcanzan distribuciones ampliasy densidades elevadas.
DISTRIBUCIÓN Y ESTATUS DELOS MAMÍFEROS CARNÍVOROSEN EL TÉRMINO MUNICIPAL DELOS BARRIOS (CÁDIZ)
F. J. Gómez Chicano / Capataz Forestal
E. Briones Villa / Delegación de Medio Ambiente, Ayuntamiento de Los Barrios
A. Muñoz Brenes / Delegación Provincial de Cádiz, Consejería de Medio Ambiente

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 204/276
Almoraima, 35, 2007
204
INTRODUCCIÓN
Los mamíferos carnívoros ocupan la cúspide de las cadenas tróficas por lo que desarrollan un papel fundamental en el mediodonde viven controlando las poblaciones de sus presas habituales, siendo necesarios en la organización y funcionamientode los ecosistemas (Bueno 1996). A pesar de esta importancia para el mantenimiento del equilibrio ecológico y de estartodos los carnívoros ibéricos (excepto el zorro) protegidos por la ley son escasos los estudios sobre su distribución,herramienta imprescindible para abordar los problemas de conservación (Maurer 1994).
En la provincia de Cádiz solo existen dos aproximaciones a su distribución aún incompletas realizadas por la SociedadGaditana de Historia Natural (SGHN) y la Sociedad Española para la Conservación y el Estudio de los Mamíferos (SECEM)en forma de atlas de especies reproductoras de mamíferos terrestres en cuadrículas de 10 Km. Dentro del Parque NaturalLos Alcornocales se han citado un total de ocho especies de mamíferos carnívoros; meloncillo, jineta, zorro, comadreja,gato montes, garduña, nutria y tejón (Belmonte et al. 2003).
El interés de los autores de este artículo por conocer las especies de mamíferos carnívoros y su distribución en el municipiode Los Barrios ha llevado a la realización de este trabajo, aportando así una herramienta para su conservación. De este modo,abordando la conservación de las distintas especies de carnívoros es posible abarcar también la conservación de unafracción importante de los ecosistemas en los que estos viven (Barea-Azcón 2004).
ÁREA DE ESTUDIO
El término municipal de Los Barrios con sus 356 kilómetros cuadrados, se encuentra ubicado en el sur del Parque NaturalLos Alcornocales. La geología de las sierras corresponde a terrenos de areniscas silíceas o del Aljibe, intercalados con capasde margo-arcillas en los pies de sierra y los bujeos. La máxima cota se encuentra en el pico Cruz de Romero con 781 m.de la sierra del Niño. Atraviesan el término dos importantes ríos, de las Cañas o Palmones que alimenta el embalse de CharcoRedondo y del Tiradero o Raudal. La temperatura media anual es de 17º C. Las precipitaciones medias anuales oscilan entre763 mm. y 1180 mm. a consecuencia del relieve montañoso (Torres 1995).
El paisaje es de relieves suaves en su mayoría, cubriendo el alcornocal (Quercus suber ) con monte bajo todas las partesmedias y altas de las sierras y los pastizales y acebuchales (Olea sylvestris) las partes bajas. Las choperas (Populus alba)y alisedas ( Alnus glutinosa) con sotobosques de zarzas ( Rubus ulmifolius) y adelfas ( Nerium oleander ) acompañan loscauces de los ríos. En los terrenos de cultivos abandonados aparecen manchas continuas de jergen ( Calycotome villosa)y lentisco (Pistacea lentiscus) salpicados de acebuchal. También coexisten en algunos montes públicos repoblaciones depino piñonero (Pinus pinea) y pino negral (Pinus pinaster ) junto al alcornocal y quejigal (Quercus canariensis).
El municipio de Los Barrios tiene una actividad forestal muy importante siendo los principales aprovechamientos laextracción del corcho, la recolección de la piña y la corta de leña, junto a una intensa actividad cinegética y ganadera. Estostipos de usos han permitido desde siempre la coexistencia de los mamíferos carnívoros con las actividades humanas en elecosistema, excepto en épocas en las que la persecución de estos para el aprovechamiento de las pieles o por competenciacinegética estuvo permitida.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 205/276
Comunicaciones
205
MATERIAL Y MÉTODOSPara la obtención de datos sobre presencia y distribución de las distintas especies de carnívoros se ha recurrido a cincométodos; muestreos directos, muestreos indirectos, fototrampeo, encuestas y bibliografía.
Muestreos directosSe incluyen aguardos diurnos y nocturnos en zonas especialmente aptas para la observación de individuos, tales comoabrevaderos y cruces de veredas, así como recorridos en vehículo todoterreno por carriles. Para los recorridos nocturnosen coche se utilizó el foco cinegético como método de iluminación. En todas las variedades de muestreos directos realizadosse anotaron las especies observadas y la localización de las observaciones.
Muestreos indirectos
Para la obtención de indicios de la presencia de las distintas especies se diseñaron un total de 10 transectos de 500 m. delongitud para recorrer a pié en los que se registraron huellas, excrementos, restos de presas, arañazos etc. Cada recorridose realizó dos veces anotando tipo de indicio, localización y especie a la que pertenecía. Los recorridos se diseñaron demanera estratificada (Tellería 1986).
En el apartado de Anexo, se encuentran los planos con la disposición y localización de los transectos realizados en los cotosde caza mayor y en el de caza menor.
FototrampeoEl fototrampeo es una técnica basada en la colocación de una cámara fotográfica que funciona al ser pisada una planchapor el animal, de forma que este queda capturado instantáneamente en una fotografía (Wemmer 1996). La ubicación delas cámaras se hizo de forma aleatoria y no siguiendo una distribución homogénea en el espacio (Llimona y Camps 2002),
Figura 1. Paisaje del término municipal de Los Barrios, área de estudio.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 206/276
Almoraima, 35, 2007
206
sino mas bien tras haber localizado letrinas, pasos frecuentados o lugares donde se les atraía mediante cebos de olor o presasvivas. El material fotográfico utilizado ha sido carretes de 27 exposiciones y 100 asas pasándose los negativos a formatodigital para su posterior tratamiento. El número de cámaras instaladas han sido de cinco en siete puntos de muestreodiferentes, funcionando durante todo el periodo de estudio.
EncuestasMétodo consistente en una batería de encuestas realizadas a los guardas de caza de los cotos cinegéticos del términomunicipal en las cuales se ha requerido información sobre las especies observadas, año de observación y tipo deavistamiento (ejemplares vivo o muertos). Este método, a pesar de ciertos factores que influyen en la detección de lasespecies, aporta valiosa información sobre la distribución general de las especies (Yela y Calvo 2004). El número deencuestas realizado ha sido de 36.
Bibliografía
Se ha revisado toda la bibliografía existente susceptible de contener información acerca de presencia y distribución demamíferos carnívoros en el área de estudio. Para ello se ha realizado una búsqueda bibliográfica de literatura científica ycinegética extrayendo citas de interés sobre algunas especies, desechándose aquellas en las que no especifica el origen delos datos.
Toda la información recogida con los cinco métodos ha sido informatizada en forma de base de datos con todas lasobservaciones desglosadas por especie, localización, tipo de muestreo etc. y posteriormente volcada sobre la cartografíadel término municipal (Base cartográfica del Ayuntamiento de Los Barrios. Escala 1:10.000). Esta información seencuentra bien representada en los mapas de distribución en el Anexo.
Figura 2. Fototrampero situado en uno de los siete puntos de muestreo.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 207/276
Comunicaciones
207
RESULTADOS Y DISCUSIÓN
Se han generado un total de 220 observaciones de carnívoros siendo citadas un total de 11 especies. Hay cuatro especiesde carnívoros para las cuales todas las citas corresponden solo a encuestas y/o bibliografía; lince, lobo, gato montés y turón.El resto de especies descritas son; meloncillo, jineta , zorro, comadreja, garduña, nutria y tejón. De éstas es la jineta la máscitada con 48 observaciones y la garduña la que menos con nueve. En la siguiente tabla se ofrecen todas las observacionesdesglosadas según la procedencia de los datos.
ESPECIE Método directo Método indirecto Fototrampeo Encuestas Bibliografía TOTALZorro 3 12 7 11 0 33Tejón 3 12 1 13 0 29Meloncillo 9 6 10 11 0 36
Nutria 1 13 0 12 0 26Garduña 7 2 0 0 0 9Gineta 5 9 5 29 0 48Turón 0 0 0 3 0 3Comadreja 1 0 0 22 0 23Gato mont. 0 0 0 7 0 7Lince 0 0 0 1 3 4Lobo 0 0 0 1 1 2TOTAL 29 54 23 110 4 220
Según los patrones de distribución las especies pueden separarse en dos grupos, el de las especies de carnívoros condistribución amplia en el área de estudio (zorro, meloncillo, jineta, tejón, nutria y comadreja) y el de especies condistribución localizada a determinados ambientes forestales (garduña). Por último, en cuanto a las especies de las que solo
tenemos datos sobre la base de encuestas y/o bibliografía, los registros sobre el lince y el lobo son extraídos en su mayoríade la bibliografía. En el caso del gato montes todas las observaciones provienen de encuestas y en cuanto al turón, aparececitado aunque su presencia y distribución es aún una incógnita en la provincia.
A partir de los transectos realizados a pié para la localización de indicios (huellas y excrementos) se ha calculado el IndiceKilométrico de Abundancia (IKA) para cada especie detectada. Dicho índice es una estima de abundancia que permitedefinir el estado de una población en términos comparativos. Su cálculo es el cociente entre el número de indicios contadosy el número de kilómetros de recorrido (Tellería 1986). En la tabla siguiente se ofrecen los valores del IKA obtenidos paracada especie de la que se han obtenido registros durante el muestreo, ordenados de forma decreciente.
ESPECIE IKAZorro 2,8Meloncillo 0,8Tejón 0,6Nutria 0,4Jineta 0,3

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 208/276
Almoraima, 35, 2007
208
DISTRIBUCIÓN Y ESTATUS DE LAS ESPECIES
1. Zorro. Especie cosmopolita y de carácter oportunista es una de las que concentra un mayor número de observaciones,distribuyéndose a lo largo de toda el área de estudio. De hábitos crepusculares y nocturnos, aunque también se le puedever cazando durante el día, usa habitualmente veredas y pistas forestales para sus desplazamientos, marcándolos conexcrementos que deposita en el suelo. Aunque parece que alcanza mayores densidades en zonas abiertas donde abundael conejo, se ha comprobado su presencia en todos los ecosistemas del área de estudio, alcornocal, acebuchal, pastizal,zonas de ribera y áreas antropizadas, incluso basureros. Es probablemente capaz de aprovechar el espacio dejado porotras especies debido a su mayor tolerancia a las alteraciones del hábitat (Reig et al. 1985).
2. Meloncillo. Probablemente la especie más fácil de observar (debido a sus hábitos diurnos) ya que permanece activodesde el amanecer hasta el atardecer disminuyendo solo su actividad en las horas centrales del día. Es frecuente verloscazando en parejas. Ha sido la especie más fotografiada obteniéndose hasta 12 instantáneas de un mismo individuo. Sedistribuye por casi todo el término alcanzando mayores densidades en zonas de monte bajo con pastizal y acebuchaldonde abunda el conejo. También se han encontrado indicios de su presencia en zonas muy próximas a núcleos urbanos.
3. Jineta. Especie que aglutina el mayor número de observaciones. De actividad nocturna ocupa hábitats muy variados,desde pinares, alcornocales y quejigales con alta disponibilidad de árboles huecos o grandes piedras hasta acebuchalescon monte bajo donde se han encontrado letrinas usadas por varios individuos en tejados de casas abandonadas. Debidoa este carácter nocturno es difícil de observar y muchos de los registros de la especie proceden de animales capturadosen trampas para zorros a través de las encuestas. Las letrinas situadas en rocas accesibles son otra fuente de datos acercade su presencia. Se distribuye por la mayoría de la superficie estudiada, probablemente debido a que las temperaturassuaves de la zona favorezcan la termofilia que presenta la especie (Virgós et al. 2001). Es abundante en zonas con alta
densidad de sus presas favoritas, conejos, roedores y aves de pequeño y mediano tamaño. En la mayoría de losexcrementos encontrados aparecen restos de micromamíferos e insectos.
4. Tejón. Especie que no suele alcanzar grandes densidades en las zonas donde habita (Revilla et al. 2000) y también dehábitos fundamentalmente nocturnos, por lo que suele pasar desapercibido con facilidad, hemos encontrado sin embargocubiles muy próximos a viviendas rurales. Se distribuye por amplias zonas del municipio estando presente en la mayoríadel área del Parque Natural y fincas de caza menor con abundancia de sus presas, conejos, micromamíferos, roedores, yfrutos silvestres y cultivados. También habita zonas de vegas y huertas cultivadas con bosquetes de acebuches yalcornoques. Es habitual descubrir sus letrinas cerca de las bocas de los cubiles, que se suelen asentar sobre terrenos blandoso bajo grandes piedras. Por su corpulencia también se encuentran los pelos del lomo adheridos a los espinos de lasalambradas.
5. Garduña. Poco habitual en la zona de estudio, aunque es una especie muy difícil de observar. Existen datos antiguosde capturas ocasionales muy cotizadas en el municipio. Los datos actuales sobre su presencia son de individuostrampeados en jaulas para zorros, algún avistamiento y excrementos, ya que tienen la costumbre de dejarlos sobrecaminos y veredas y en letrinas muy características sobre rocas. Aunque no debe presentar grandes densidades en ningúncaso, se han definido zonas en las que su presencia es estable y estas coinciden con bosques de árboles viejos y grandespiedras donde encuentra seguridad para instalar su cubil. Adapta su dieta a la disponibilidad de presas (Gil-Sánchez1999), aunque las más habituales son los pequeños roedores, aves y frutos.
6. Comadreja. Es el carnívoro de menor tamaño de cuantos se distribuyen en la zona de estudio, sus hábitos diurnos y sucarácter confiado le hacen relativamente observable. Sin embargo, sus indicios pasan con frecuencia desapercibidos yson más difíciles de localizar que los de otras especies de mayor tamaño (King 1989). Se han registrado citas dispersas

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 209/276
Comunicaciones
209
Figura 3. Zorro (Vulpes vulpes). Figura 4. Meloncillo ( Herpestes ichneumon ).
Figura 5. Jineta (Genetta genetta). Figura 6. Huella de Tejón.
Figura 7. Garduña ( Martes foina). Figura 8. Comadreja ( Mustela nivalis ).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 210/276
Almoraima, 35, 2007
210
por todo el territorio aunque es seguro que la especie se encuentra mejor distribuida, ocupando hábitats de alcornocaly acebuchal con monte bajo y zonas de huertas próximas a núcleos urbanos donde encuentra refugio en los muros depiedra. Se hacen necesarios pues muestreos intensivos en el futuro para definir el estado actual de sus poblaciones. Sualimentación es variada, incluyendo conejos que caza en sus madrigueras (Blanco 1998).
7. Nutria. Especie ligada a hábitats fluviales y lacustres, está presente en el área en prácticamente todos los ríos, embalsesy pantanetas. El hecho de haber sido detectada en pequeños embalses alejados de grandes cauces implica su capacidadpara atravesar amplias zonas secas. Su detectabilidad es alta debido a la facilidad para encontrar sus excrementos sobrelas piedras y troncos de las orillas de las masas de agua. Su dieta se basa en peces, cangrejos de río, mamíferos de pequeñotamaño e insectos. Aunque en el pasado fue perseguida ya que su piel era apreciada, actualmente parece que su poblaciónestá en expansión habiéndose obtenido registros incluso en hábitats costeros. Procesos de recolonización similares hansido descritos en numerosas poblaciones peninsulares (Ruiz-Olmo y Delibes 1998).
8. Turón.Carnívoro ligado a cauces fluviales con vegetación y abundante y alta densidad de conejos y roedores de pequeñoy mediano tamaño. Su elevada especificidad a los hábitats riparios (Barea-Azcón et al. 2004) junto con su carácterdiscreto hacen que su presencia y distribución continúe siendo un enigma en el Parque Natural de Los Alcornocales yen la provincia de Cádiz. Todas las citas provienen de animales trampeados entre 1970 y 1980 y la información se ha
conseguido a través de las encuestas. Una de dichas citas corresponde al término de Tarifa pero dada la proximidad alos límites del área de estudio ha sido incluida. Probablemente el turón es una de las especies que requiere un mayoresfuerzo de muestreo en el futuro para determinar el estado actual de sus poblaciones.
9. Gato montés. Se han recogido seis observaciones de la especie y todas provienen de encuestas a guardas de fincas. Dosde ellas se sitúan geográficamente fuera del área de estudio aunque nuevamente se han incluido por su proximidad. Lascuatro del término corresponden a animales abatidos o muertos en cepos hace más de 30 años. No existen evidenciasde la presencia actual de la especie, aunque si de poblaciones próximas geográficamente por lo que el área de estudiosería cuando menos susceptible de una futura colonización. En cualquier caso, dado que su distribución coincide agrandes rasgos con la de su presa habitual en los ecosistemas mediterráneos que es el conejo (Gil-Sánchez et al. 2003)será la dinámica poblacional de éste el que probablemente determine su presencia.
Figura 9. Nutria ( Lutra lutra ). Figura 10. Hábitat de la nutria.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 211/276
Comunicaciones
211
10. Lobo. Su presencia en la zona en el pasado está atestiguada por una referencia bibliográfica (Montesdeoca 2004) y el datode una encuesta. En la primera se describe como fue abatido el último lobo de Zanona, en la cañada de Juan Azorero, enla primera década del siglo XX. En la segunda, más interesante por ser mucho más reciente, se cita un lobo abatido enLa Almoraima en el año 1991, que era conocido por la guardería de la finca por atacar crías de muflones y gamos.
11. Lince. Se han recopilado cuatro citas bibliográficas, dos del área de estudio y otras dos fuera del término que por suimportancia y proximidad se han decidido incluir.
AÑO CITA LOCALIDAD1998 Capturado Cañada del Valle - Medina Sidonia1980 Abatido Zanona - Los Barrios1978 Avistamiento La Almoraima - Castellar de la Frontera
196? Capturado Ojén - Los Barrios
CONCLUSIONES
Hay tres especies de carnívoros, jineta, zorro y meloncillo que presentan una amplia distribución en el área de estudio yun gran número de observaciones en los distintos tipos de muestreos realizados. Aunque el zorro es probablemente laespecie que presenta una mayor distribución y densidad en el área de estudio, la jineta aparece con un mayor número deobservaciones totales. Esto es debido al gran número de datos sobre presencia de jinetas que se obtuvieron mediante lasencuestas, la mayoría de ellos procedentes de individuos capturados en jaulas frente a la menor cantidad de zorros que sonatrapados mediante este sistema de trampeo. En el caso del meloncillo, su grado de distribución y su densidad aparecenreflejados en la totalidad de los tipos de muestreo.
La nutria es una especie que también presenta un alto número de observaciones aunque obviamente su distribución estácondicionada por la disponibilidad de masas de agua. Un caso complejo es el del tejón, cuya densidad debe ser menor que lasespecies citadas anteriormente debido a su ecología y sin embargo posee un IKA mayor con respecto a alguna de estas. Estopuede deberse al alto grado de detectabilidad de sus indicios debido a su tamaño corporal y peso. Es el caso contrario a lacomadreja que probablemente esté mejor distribuida de lo que se infiere de la información obtenida dado su pequeño tamaño.
En los casos del lobo y del lince las citas bibliográficas y encuestas arrojan informaciones antiguas de presencia del lobo yotras más recientes de linces que probablemente se correspondan con casos conocidos de individuos erráticos o en dispersión.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 212/276
Almoraima, 35, 2007
212
ANEXOS
Figura 11. Transeptos de carnívoros en el coto de caza mayor "El Rincón" CA:10444 y "Los Garlitos" CA:10442.
ANEXO I

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 213/276
Comunicaciones
213
Figura 12. Transeptos de carnívoros en el coto de caza menor "La Gineta", CA:10920.
ANEXO II

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 214/276
Almoraima, 35, 2007
214
ANEXO III

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 215/276
Comunicaciones
215
ANEXO IV

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 216/276
Almoraima, 35, 2007
216
BIBLIOGRAFÍABAREA-AZCON, J.M., E. Ballesteros-Duperon, Moleon, J.M. Gil- Sánchez, E. Virgos, y M. Chirosa. "Distribución de los mamíferos en la provincia de Granada".
Acta Granatense , 3: 43-53. 2004.BELMONTE, J., J.J. Ceballos, I. Sánchez y J.M. Soria. "Avance del Atlas de los mamíferos terrestres de la provincia de Cádiz (excepto quirópteros)". Revista dela Sociedad Gaditana de Historia Natural, Vol. III: 7-16. 2003.BLANCO, J.C. Mamíferos de España . Ed. Geoplaneta. 1998.BUENO, F. "Importancia ecológica de los carnívoros".Pp.171-182. En: R. García-Perea, R.Baquero, R. Fernández-Salvador y J. Gibert (eds.) Carnívoros;evololución, ecología y conservación. CSIC-MNCN.SECEM. Madrid. 1996.GIL-SÁNCHEZ, J.M. "Dieta de la garduña ( Martes foina ) en una localidad de las Sierras Subbéticas de Granada (SE de España)". Doñana Acta Vertebrata , 23(1):83-90. 1996.GIL- SÁNCHEZ, J.M., G. Valenzuela, y J, F. Sánchez. "Iberian Wildcat (Felis silvestris tartessia) predation on rabit: functional response and age selection". ActaTheriologica, 44 (4): 421-428. 1999.KING, C. Wealsels Stoats. Comstock Cornell. 1989.LLIMONA, F. y D. Camps. "Uso del espacio en la gineta en dos sectores del Parc de Collserola". Quercus 179. 2004.
MAURER, B.A. Geographical population analysis: tools for the analysis of population. Oxford; Boston. Blackwell Scientific Population. 1994.MONTESDEOCA, J. Revista de la Sociedad de caza La Jineta. 2004.NAVES, J., A. Fernández, J.F. Gaona,y C. Nores. "Uso de cámaras automáticas para la recogida de información fáustica". Doñana Acta Vertebrata, 23(2). 189-199. 1996.REIG, S., L. Cuesta, y F. Palacios. "The impact of human activities on the food habits of Red Fox and Wolf in Old Castille, Spain". Rev. Ecol. (Terre Vie), 40: 151-155. 1985.REVILLA, E., D. Palomares y Delibes. "Defining key habitats for low density populations of Eurasian badger in Mediterranean environments" . BiologicalConservation, 95: 269-277. 2000.RUIZ-OLMO, J., M. Delibes. La Nutria en España ante el horizonte del año 2000 . SECEM. 1998.TELLERIA, J.L. Manual para el censo de vertebrados terrestres. Editorial Raíces. 1986.VIRGOS, E, T. Romero, J. G. Mangas, y J. L. Teleira. "Factors determining “gaps” in the distribution of a small carnivores, the common genet (Genetta genetta),in central Spain". Canadian journal of Zoology, 79(9): 1544-1551. 2001.YELO, N.D. y J.F. Calvo. "Aproximación a la distribución y estatus de los mamíferos carnívoros en la región de Murcia". Galemys, 16(2). 21-37. 2004.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 217/276
Comunicaciones
217
RESUMEN
Spartina densiflora es una gramínea procedente de Sudamérica que está invadiendo numerosas marismas mareales en elgolfo de Cádiz provocando problemas muy graves para la conservación de estos ecosistemas, como la pérdida debiodiversidad. En el Campo de Gibraltar Spartina densiflora se localiza en las desembocaduras de los ríos Vega y Jara yen el estuario del río Palmones. Conocer el estado de la invasión en estas zonas es de especial relevancia a la hora de optar
entre diferentes técnicas de erradicación o control. Mientras en el río Palmones sería factible la eliminación mecánica delos clones de Spartina densiflora, en el caso de los estuarios de los ríos Vega y Jara el empleo de la ganadería como medidade bio-control parece la estrategia más aconsejable. No obstante, la amplia gama de efectos que la presión de la herbivoríaprovoca sobre las comunidades vegetales y, en concreto, sobre la evolución de especies invasoras hace necesario llevar acabo estudios detallados sobre la utilidad de la ganadería como herramienta de gestión en materia de especies invasoras.
EL EMPLEO DE TÉCNICAS DE CONTROL
Y ERRADICACIÓN EN LA LUCHACONTRA ESPECIES INVASORAS:CONVENIENCIA DE LA HERBIVORÍA COMO MÉTODO DE
BIOCONTROL DE LA INVASIÓN DE SPARTINA DENSIFLORA
BRONGN. EN EL CAMPO DE GIBRALTAR
Enrique Mateos Naranjo / Susana Redondo Gómez / Jesús M. Castillo / M. Enrique FigueroaDepartamento de Biología Vegetal y Ecología. Facultad de Biología. Universidad de Sevilla

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 218/276
Almoraima, 35, 2007
218
INTRODUCCIÓN
En el contexto actual de la globalización de las relaciones económicas y mercantiles, la invasión de especies exóticas escada día un problema más grave para la conservación de los ecosistemas. Las marismas mareales han sido descritas comouna de las áreas más afectadas por la introducción de estas especies invasoras debido a su carácter de transición entreecosistemas de diferente naturaleza y a la cercanía de importantes puertos marítimos donde llegan muchas de ellas, en lasaguas de lastre o en el exterior del casco de embarcaciones mercantes y de recreo. La entrada de especies exóticas en lasmarismas puede ocasionar numerosos efectos negativos, tales como: la competencia con la flora nativa (Kittelson y Boyd,1997), la alteración de la cantidad y naturaleza del detritus (Adam, 1990), la modificación del hábitat de la fauna autóctona(Partridge, 1987) y la alteración de la dinámica sedimentaria de los estuarios (Long y Mason, 1983).
Spartina densiflora es una gramínea procedente de Sudamérica (Mobberley, 1956) que está invadiendo actualmente elnorte de África (Nieva, 1996) y América (Kittelson y Boyd, 1997) y el suroeste de Europa (golfo de Cádiz; Castillo et al.,2000) (figura 1). En el golfo de Cádiz, S. densiflora aparece en varios estuarios donde presenta diferente grado de invasión,siendo en el estuario conjunto de los ríos Tinto y Odiel (Huelva) donde muestra un mayor desarrollo, tanto en el espacioocupado como en la diversidad de hábitats invadidos (Nieva, 1996).
La invasión de S. densiflora es uno de los principales problemas de conservación a los que se enfrentan las marismasandaluzas atlánticas en el siglo XXI. La biodiversidad de las marismas andaluzas del golfo de Cádiz se ha vistoespecialmente mermada debido a su efecto (Luqueet al., 2003). Spartina densifloradesplaza por competencia interespecíficaa gran parte de las especies autóctonas, pues muestra una capacidad competitiva muy elevada debido a su forma decrecimiento (clonal en falange; Figueroa y Castellanos, 1998) y a que presenta un rango ecológico muy amplio (Kittelsony Boyd, 1997). Estas capacidades han dado lugar a la formación de comunidades prácticamente mono-específicas
conocidas popularmente como ‘mares de Spartina’ (Rubio, 1985).
Se han estudiado, mediante experimentos de campo y laboratorio, diferentes aspectos ecológicos de S. densiflora en el golfode Cádiz (Castillo et al., 2000; Castillo et al., 2003; Castillo et al., 2004; Figueroa y Castellanos, 1998; Nieva et al., 1996).Sin embargo aún quedan muchas incógnitas sobre el papel que esta especie desempeña en las marismas que invade. En vistade la gravedad de la situación actual como consecuencia de la invasión de S. densiflora sería necesario el desarrollo deconocimientos teóricos y aplicados dirigidos a luchar eficazmente contra la invasión en las marismas andaluzas. En estacomunicación se analiza la adecuación de las diferentes técnicas de control y erradicación existentes para el caso concreto deS. densiflora en el Campo de Gibraltar, centrándonos principalmente en el papel de la hervivoría como método de biocontrol.
ESTADO DE LA INVASIÓN DE SPARTINA DENSIFLORA EN EL CAMPO DE GIBRALTARDentro del ámbito geográfico que nos ocupa (Campo de Gibraltar) S. densiflora se localiza en el río Palmones (bahía deAlgeciras) y en las desembocaduras de los ríos Vega y Jara (Tarifa). En estas dos localidades el estado de sus poblacionesvaría en función de su extensión e importancia con respecto a otras especies.
Las poblaciones de S. densiflora en el río Palmones se caracterizaron por la presencia de clones dispersos de aspectoraquítico, escasa cobertura, y situados sobre sustrato arenoso.
Por otro lado, la presencia de S. densiflora en la desembocadura de los ríos Vega y Jara fue mucho mayor, con coberturaspróximas al 80% en las zonas más cercanas al cauce principal de los ríos. En los huecos dejados por la especie invasorase encontraron individuos de Halimione portulacoides y de los géneros Sarcocornia y Juncus. La cobertura se redujo de

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 219/276
Comunicaciones
219
forma gradual con la distancia al cauce hasta quedar limitada a clones aislados. Los clones de S. densiflora de los ríos Vegay Jara, a diferencia de los del género Juncus, presentaron cierto grado de ramoneo por parte del ganado vacuno,especialmente aquellos que se alejaban más del cauce principal del río (Castillo, “obs. pers.”).
MEDIDAS DE CONTROL O ERRADICACIÓN. EFECTO DE LA HERBIVORÍA SOBRE S. DENSIFLORA
El desarrollo de técnicas de control o erradicación debe estar apoyado tanto en el conocimiento de la especie invasora haciala que van destinadas como en las características fisicoquímicas del medio donde se vayan a aplicar, con el fin de que seantécnicas lo más específicas posibles, y evitar así la interferencia sobre otras especies y/o procesos naturales.
Existe una enorme controversia sobre la conveniencia del uso de técnicas de control o erradicación en la lucha contra las
especies invasoras. En gran medida el empleo de unas u otras técnicas depende del estado de la invasión y de la capacidadde movilización de los medios técnicos y económicos necesarios. En la mayoría de los casos las invasiones pasandesapercibidas o son detectadas cuando ocupan áreas extensas, condiciones en las que la erradicación resulta económicay técnicamente desfavorable. A pesar de esta circunstancia la bibliografía recoge ejemplos de erradicaciones efectivas,como es el caso de Carpobrotus edulis en la isla de Menorca y Pistia stratiotes en Doñana (Fraga et al., 2004; Ortega et al., 2004). Si la erradicación no es viable, las medidas de control pasan a cobrar mayor importancia. El fundamento de estasmedidas, se basa en llevar la especie invasora a niveles poblacionales aceptables para la conservación del ecosistema.
En la bibliografía existe numerosa información sobre el uso de técnicas de tipo químico (herbicidas) para la erradicación o elcontrol de especies del género Spartina (Hedge et al., 2003; Patten, 2002; Paveglio et al., 1996). Aunque en muchos casos losresultados obtenidos con el uso de herbicidas han sido satisfactorios, el empleo de productos químicos en la lucha contra S.
Figura 1. Distribución mundial de las poblaciones de Spartina densiflora.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 220/276
Almoraima, 35, 2007
220
densiflora en un espacio natural como el Parque Natural del Estrecho no es aconsejable, ya que existe un gran desconocimientosobre la eficacia de estos productos en esta especie, y los efectos perjudiciales derivados de esta actividad, tanto en el medio comoen los propios organismos de la zona, incluyendo al hombre. Esta circunstancia obliga a la búsqueda de técnicas de lucha contrala invasión más respetuosas con el medioambiente. Además, hay que resaltar la alta carga económica que supone el uso deherbicidas tanto por la síntesis del producto en sí como por el equipo necesario para su aplicación y la recurrencia de ésta, quedebería ser durante varios años debido a la posibilidad de que exista un banco de semillas.
Otro tipo de técnicas serían las de tipo mecánico (para la erradicación o el control), las cuales consistirían en la retiradade forma manual o mediante máquinas de todos los clones de la especie invasora. Esta técnica gozaría de mayor aceptaciónsocial que el uso de productos químicos, y en cierto modo podría ser muy útil para la lucha de S. densiflora en aquellas áreasdonde esta presente una escasa cobertura, como es el caso del río Palmones. El gran inconveniente de dicha técnica radica
en el gasto económico derivado del pago de los salarios de los trabajadores y del equipamiento, sobre todo en zonas dondela extensión ocupada por la especie invasora es mayor (Ríos Jara y Vega). Además de las dificultades técnicas derivadasde la inestabilidad del terreno asociada a los ecosistemas litorales, las cuales dificultarían los trabajos manuales y la entradade maquinaria pesada.
Por último, citaremos las técnicas de tipo biológico conocidas como de biocontrol. Su fundamento consiste en laintroducción de organismos vivos cuyas actividades interfieran sobre el desarrollo normal de las poblaciones de las especiesinvasoras. La problemática de esta técnica reside en que normalmente no consigue la eliminación total de la especieinvasora, sólo una reducción hasta niveles no perjudiciales para el ecosistema. En el caso del control de S. densiflora porpresión ganadera el éxito dependerá del estado de la invasión y de las características de la zona (en el caso de fangos noconsolidados no es posible el acceso de ganado). En zonas de fangos consolidados con escasa influencia mareal donde la
invasión se encuentre en estadios iniciales la actividad del ganado puede llegar a eliminarla. En el caso de poblacionesconsolidadas, con gran altura de tallos, donde la cobertura de S. densiflora alcance el 100% la acción del ganado tendríaun éxito limitado. En situaciones intermedias, donde la cobertura es inferior la eliminación de biomasa aérea por ramoneopodría frenar el crecimiento en falange de S. densiflora, dejando huecos susceptibles de ser ocupados por otras especiesdiferentes a la invasora, aumentando con ello la diversidad de la zona. En este último caso, la ganadería podría adquirir unagran trascendencia como herramienta de gestión del Parque Natural del Estrecho para la lucha contra S. densiflora.
A pesar de la importancia que tienen las técnicas de control biológico como la herbivoría en la lucha contra la invasión deespecies exóticas en diferentes ecosistemas, los estudios de este tipo en el género Spartina son muy escasos, ya que de lassiete especies consideradas como invasoras dentro del género Spartina (S. alterniflora, S. anglica, S. densiflora, S.maritima, S. patens, S. pectinata y S. townsendii; Daehler y Strong, 1996), sólo en el caso de S. alterniflora (Bortolus,2003), S. densiflora (Daehler, 1995) y S. patens (Hester, 1994; Ford, 1998) se han acometido estudios para conocer el efectode la herbivoría sobre sus poblaciones.
Para conocer los posibles efectos de la actividad ganadera en la evolución de S. densiflora en el Campo de Gibraltar,previamente hay que analizar la propia interacción planta – herbívoro, ya que esta actividad podría actuar en beneficio oen contra de la invasión, pues los efectos sobre las especies vegetales son muy variados y dependen del tipo de herbívoro,de la intensidad de la herbivoría, de la antigüedad del ramoneo en la zona, del clima y del tipo de suelo (Hobbs et al., 1992;Milchunas et al., 1993).
En el caso de pequeños mamíferos e insectos los efectos van dirigidos principalmente a las semillas, tanto durante supermanencia en la planta progenitora como en los bancos de semillas. También tienen cierto efecto sobre las inflorescenciasy el establecimiento de las plántulas, suponiendo una reducción de la abundancia de éstas en futuras generaciones (Louda

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 221/276
Comunicaciones
221
et al., 1995). Con respecto a la influencia por parte de grandes herbívoros como puede ser el ganado, los efectos sobre lavegetación son muy complejos, ya que, aparte de la gran reducción de biomasa vegetal que promueven, tienen efectosdirectos e indirectos sobre el crecimiento de la vegetación, la dispersión de las semillas, la reproducción y la fenología(Belsky 1986; Collins 1987; Bortolus et al., 2003). Estudios previos muestran que los herbívoros más generalistas puedenprovocar una reducción de especies de plantas superior a la que inducen los especialistas (Maron et al., 2001).
En relación a la intensidad de la herbivoría numerosos trabajos consideran que un pastoreo moderado por herbívorosnaturales mantiene una alta diversidad como consecuencia del consumo de especies dominantes (Perevolotsky y Seligman,1998). Del mismo modo una alta presión herbívora puede llevar a una reducción de la diversidad (Milchunas y Lauenroth,1993; Ungar, 1998).
El efecto de la antigüedad de la actividad de ramoneo ha sido estudiado principalmente en pastizales con larga historia depastoreo, donde se ha visto que la diversidad es máxima en presencia de los herbívoros y, en general, no se observa lainvasión de especies exóticas ni la pérdida de especies nativas aún en casos de alta carga ganadera (Milchunas et al., 1988,Noy-Meir et al., 1989).
Por último, tanto el clima como la composición del suelo se han descrito como factores que tienen una importanterepercusión sobre el efecto que la herbivoría puede tener sobre las comunidades vegetales (Milchunas et al., 1988).Condiciones ambientales desfavorables pueden agravar los efectos derivados de la herbivoría pudiendo acabar con laeliminación de especies menos tolerantes. Se han llevado a cabo estudios que demuestran como variaciones locales en losfactores abióticos en la marisma son causantes de variaciones en los efectos de la herbivoría sobre las comunidadesvegetales (Gough et al., 1998).
En vista de los numerosos factores a considerar a la hora de valorar el efecto de la herbivoría sobre la vegetación y en quémedida ésta resulta efectiva contra la lucha de especies invasoras, urge analizar concretamente el efecto de la ganaderíasobre la invasión de S. densiflora en el Parque Natural del Estrecho. En este sentido el Grupo de Investigación Ecología,Citogenética y Recursos Naturales de la Universidad de Sevilla se propone realizar el seguimiento de una serie de parcelasde exclusión, donde se impida la herbivoría, y compararlas con datos de parcelas control donde tenga lugar el ramoneo.Los resultados derivados de este estudio permitirán valorar la conveniencia de la herbivoría como herramienta de gestiónen materia de lucha contra especies invasoras en los distintos espacios naturales, sobre todo en aquellos que presenten unacierta tradición ganadera.
BIBLIOGRAFÍAADAM, P. Saltmarsh Ecology. Cambridge University Press, Cambridge, 1990, pp 461.
BELSKY, A.J. “Does herbivory benefit plants? a review of the evidence”, American Naturalist , 127(1986), pp 870-892.BORTOLUS, A., Laterra, P y Iribarne, O. “Crab-mediated phenotypic changes in Spartina densiflora brong”, Estuarine, Coastal and Shelf Science, 59 (2003), pp.97-107.CASTILLO, J.M., Fernández-Baco. L. Castellanos. E.M. Luque. C.J. Figueroa. M.E y Davy. A.J. “Lower limits of Spartina densiflora and S. maritima in aMediterranean salt marsh determinated by different ecophysiological tolerances”, J. Ecol. 88 (2000), pp 801-812.CASTILLO, J.M., Rubio-Casal. A.E. Figueroa. M.E y Nieva. F.J. “Intratussock tiller distribution and biomass of Spartina densiflora Brongn. in an invaded saltmarsh”, Lagascalia, 23 (2003), pp. 61-73.CASTILLO, J.M., Rubio-Casal. A.E. Redondo. S. Álvarez López, A.A. Luque T. Luque. C. Nieva. F.J. Castellanos. E.M y Figueroa. M.E.. “Short-term responsesto salinity of an invasive cordgrass”. Biological Invasions . 7 (2004), pp. 29-35.COLLINS, S.L. “Interaction of disturbances in tallgrass prairie: a field experiment”, Ecology, 68 (1987), pp. 1243-1250.CRAIN, C.M., Silliman. B.R. Bertness. S.L y Bertness. M. D. “Physical and Biotic drivers of plant distribution across estuarine salinity gradients”,Ecology, 85 (2004),pp. 2539-2549.DAEHLER, C.C. y Strong, D.R. “Status, prediction and prevention of introduced cordgrass Spartina spp”. Invasions in Pacific estuaries, USA, BiologicalConservation, 78 (1996), pp. 51-58.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 222/276
Almoraima, 35, 2007
222
DAEHLER, C.C. y Strong. D.R. “Status, prediction and prevention of introduced cordgrass Spartina spp. Invasions in Pacific estuaries, USA”. BiologicalConservation, 78 (1996), pp. 51-58.FRAGA, P., Estaún, I. Alarcón. A. Cots. R. y Da Cunha, G. Eradication of Carpobrotus in Minorca. Internacional. Workshop “Invasive Plants in the MediterraneanType Regions of the World”, 33, Mèze (Hèrault), France. (2004)FIGUEROA, M.E. y Castellanos. E.M.. “Vertical structure of Spartina maritima and Spartina densiflora in Mediterranean marshes”. En: Plant form and vegetationstructure (eds M.J.A. Werger, P.J.M. van der Aart, H.J. During & J.T.A. Verhoeven), (1988), pp. 105-108. SPB Academic Publishing, The Hague, Holanda.FORD, M.A. y Grace. J.B. “ The interactive effects of vertebrate herbivory and fire on a coastal marsh in Louisiana, the Pearl River, Wetlands, 18 (1998), pp. 1-8.GOUGH, L. y Grace. J.B. “Herbivore effects on plant species density at varying productivity levels”, Ecology, 79 (1998), pp. 1586-1594.GOUGH, L. y Grace. J.B. “Effects of floofing, salinity and herbivory”, Oecologia, 177 (1998), pp. 527-535.HEDGE, P., Kriwoken. L.K y Patten. K. “A review of Spartina management inWashington State, US”, Journal of aquatic plant management , 41(2003), pp. 82-90.KITTELSON, P.M. y Boyd. M.J. “Mechanisms of expansion for an introduced species of cordgrass, Spartina densiflora, in Humboldt Bay, California”, Estuaries,20 (1997), pp. 770-778.HOBBS, R.J. y Huenneke. L.F. “Disturbance, diversity, and invasion: implication for conservation”, Conservation Biology, 6 (1992), pp. 324-337.LONG, S.P. y Mason. C.F.. Saltmarhs Ecology, Blackie, Glasgow. 1983.
LOUDA, S.M. y Potvin. M. A.. “Effect of inflorescensefeeding insects on the demography and lifetime fitness of a native plant” Ecology, 76 (1995), pp. 229-245.LUQUE, C.J., Castellanos, E.M. Castillo. J.M y Figueroa. M.E. Diversidad en marismas mareales mediterráneas I. Cambios de la diversidad específica . En:Diversidad Biológica y Biodiversidad. Madrid. 2003.MARON, J.L. y Vilà. M.. “When do herbivores affect plant invasion? Evidence for the natural enemies and biotic resistance hypotheses”, Oikos, 95 (2001), pp. 361-373.MILCHUNAS, D.G., Sala. O.E y Lauenrotn. W.K. “A generalized model of the effects of grazing by large herbivores on grassland community structure”, American
Naturalist , 132 (1988), pp. 87-106.MILCHUNAS, D.G. y Lauenrotn. W.K. “A quantitative assessment of the effects of grazing on vegetation and soils over a global range of environments”, Ecological
Monographs, 63 (1993), pp. 327-366.MOBBERLEY. D.G. “Taxonomy and distribution of the genus Spartina”, Iowa State College J. Sc. 30 (1956), pp. 471-574.NIEVA. F.J. 1996. Aspectos ecológicos en Spartina densiflora Brongn , Tesis Doctoral. Universidad de Sevilla, España.NOY-MEIR, I., Gutman. M y Kaplan. Y. “Responses of Mediterranean grassland plants to grazing and protection”, Journal of Ecology , 77(1989), pp. 290-310.ORTEGA, F., Dana. E.D y García. P. Control of Pistia stratiotes near Doñana Natural Park (SW Spain), Workshop “Invasive Plants in the Mediterranean Type Regionsof the World”, 33, Mèze (Hèrault), France. (2004).PARTRIDGE. T.R. “Spartina in New Zealand”, New Zealand Journal of Botany , 25 (1987), pp. 567-575.PATTEN. K. “Smooth cordgrass (Spartina alterniflora) control with imazapyr”, Weed technology, 16 (2002), pp. 826-832.
PAVEGLIO, F.L., Kilbride. K.M. Grue. C.E. Simenstad. C.A. y Fresh. K.L. “Use of Rodeo(R) and X-77(R) spreader to control smooth cordgrass (Spar tina alterniflora)in a southwestern Washington estuary.1. Environmental fate”, Environmental toxicology and chemistry, 15 (1996), pp. 961-968.PEREVOLOTSKY, A. & Seligman, N.G. “Role of grazing in mediterranean rangeland ecosystems”, BioScience, 48 (1998), pp. 1007-1017.PETTIT, N.E., Froend. R.H y Ladd P.G. “Grazing in remnant woodland vegetation: changes in species composition and life form groups”, Journal of VegetationScience, 6 (1995), pp. 121-130.RUBIO, J.C. 1985. Ecología de las Marismas del Odiel, Tesis doctoral, Universidad de Sevilla, España.TAYLOR, K.L. y Grace. J.B. “The effects of vertebrate herbivory on plant community structure in the coastal marshes of the Pearl river, Louisiana, USA”,Wetlans, 15 (1995), pp. 68-73.UNGAR. I.A. “Are biotic factor significant in influencing the distribution of halophytes in saline habitats?”, Botanical review, 64 (1998), pp. 176-199.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 223/276
Comunicaciones
223
RESUMEN
La realización de proyectos de ordenación en el Parque Natural Los Alcornocales (Cádiz), pone de manifiesto, a través delos resultados de los inventarios de la vegetación a escala monte, el decaimiento del alcornocal como consecuencia de surelativo envejecimiento y de la presencia de abundantes daños, así como por la ausencia o escasez más o menos generalizadatanto de pies menores como de regeneración. También se analizan las cargas ganaderas y cinegéticas, detectando laincompatibilidad de las elevadas cargas conjuntas actuales con los logros de regeneración natural. Finalmente, se proponen
algunos de los objetivos que faciliten la persistencia y estabilidad de los ecosistemas forestales.Palabras clave: inventario, decaimiento, herbívoros, Quercus suber
1. INTRODUCCIÓN
Las formaciones de alcornoque en general y los alcornocales de la provincia de Cádiz en particular tienen una enormeimportancia desde diferentes puntos de vista: por un lado, por la elevada cuantía de la superficie arbolada por dicha especie,que además genera una importante cuantía de productos directos (corcho, ganado, caza y otros menores: hongos, leñas,brezo, turismo, miel, etc); por otro, por la importante biodiversidad de los ecosistemas (además de alcornocales, aparecenquejigales, acebuchales, pastizales, matorrales, y en menor medida melojares, acebedas y otras, sin olvidar las importantes
especies de pteridofitas); también destaca desde el punto de vista social por la cuantía de jornales generados y por laintegración que implica con la comunidad rural.
Durante los últimos años se han acometido diferentes estudios del alcornocal en localizaciones concretas del Parque NaturalLos Alcornocales, destacando los realizados en los montes de Cortes de la Frontera y Ronda (Torres, 1995) y otrosrealizados en las inmediaciones de la sierra del Aljibe (Jurado, 2002). En ambos estudios se han realizado inventarios sobremasas de alcornocal y de quejigal, fundamentalmente, caracterizando los ecosistemas mediante los valores de densidad,área basimétrica, regeneración, etc.
En el presente trabajo se analizan esas mismas características en diferentes comarcas del parque, que cuenta con más de170.000 hectáreas, a partir de los valores obtenidos de los inventarios de los proyectos de ordenación redactados en losúltimos años.
CARACTERIZACIÓN SILVOPASTORALDE LOS ALCORNOCALES DELA PROVINCIA DE CÁDIZ
Luis Linares García / Ingeniero de Montes

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 224/276
Almoraima, 35, 2007
224
2. MATERIAL Y MÉTODOS
2.1. Formaciones vegetales
En la figura 1 aparece la distribución de las principales formaciones presentes en los montes ordenados; existe unaconsiderable variedad de formaciones vegetales y tipos fisionómicos que han sido resumidos para una mejor comprensión.La heterogeneidad de la estructura y aún más la de la distribución es una de las características fundamentales.
Por último, aunque no cartografiados por separado e incluidos en el quejigal, aparecen los canutos, formaciones muy particularesencajadas en los arroyos que presentan una elevada biodiversidad, con amplia presencia de taxones catalogados del ámbitomacaronésico y paleoártico ( Ilex aquifolium, Culcita macrocarpa, Vandenboschia speciosa, Pteris incompleta, etc).
2.1.1. Algunas notas sobre la gestión del alcornocal en el pasado
Durante gran parte del siglo XIX los montes de la Comarca estuvieron sometidos al hacha y al diente de los ganados,originando múltiples montes bajos; cuando el corcho adquirió valor económico, a finales del siglo XIX y principios delXX, los alcornocales se orientaron a la producción de corcho, potenciando al alcornoque frente al quejigo y el acebuche,manteniendo altas cargas pastantes ganaderas (entonces de cabra), a las que se añadieron las del ciervo una vezreintroducido (hacia 1960) y expandido, continuando con las rozas de matorral a hecho (erosión y pérdida de suelo, pérdidade microclima por disminución de la cobertura arbórea, etc), quemas pastorales en herrizas, las podas para carboneo, condaños reiterados por los descorches mal ejecutados o fuera de tiempo o ambas; en definitiva, un excesivo número de factoresconfluyentes achacables a más de un siglo de continua explotación que no contempló, salvo contadas excepciones, larenovación del arbolado. Además, desde 1960 se produce cierto alejamiento del hombre del medio que propicia una ineficazlucha contra enfermedades y plagas, favorecidas por el envejecimiento del arbolado.
Por otro lado, tradicionalmente, exceptuando casos concretos (Majadas de Ronda, Robledal y la Sauceda), muchasordenaciones sólo han regulado el aprovechamiento corchero, relegando al olvido la ordenación selvícola. Todo estecúmulo de factores ha propiciado el envejecimiento del alcornocal, la disminución de la cantidad de corcho producido yla disminución de su calidad. Se perdió de vista el hecho de que sin árboles no hay corcho, y que debe prevalecer laordenación selvícola por encima de la gestión de los descorches, que no son más que consecuencia de lo primero.
2.2. Método de toma de datos
2.2.1. Estudio de las formaciones vegetales
Durante los últimos años se han realizado en montes localizados por todo el Parque Natural Los Alcornocales inventariospor muestreo estadístico sistemático; la superficie de algunos de esos montes, la superficie inventariada y su desglose por
formaciones vegetales, así como el número de parcelas, su radio y el lado de malla aparecen en la tabla 1.En cada parcela se han medido los pies mayores (perímetro ≥ 40 cm), los pies menores (perímetro < 40 cm y altura superiora 1,30 metros) y la regeneración viable (altura inferior a 1,30 m); se han medido todas las especies presentes, diferenciandoen el alcornoque entre descorchados y bornizos; también se han anotado alturas de descorche de todos los alcornoques, así como los daños y la intensidad de los mismos en los árboles tipo (cuatro por parcela). Se ha completado el estadillo de campocon datos de las especies y las características del matorral (altura, cubierta, etc), especies más mordisqueadas, tratamientosselvícolas ejecutados, etc.
La solicitud del error del número de pies por hectárea, con una probabilidad fiducial del 95%, ha sido en general inferior al 20%.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 225/276
Comunicaciones
225
2.2.2. Estudio de las cargas ganaderas y la gestión actual
La carga ganadera media es variable, ya que depende sobre todo de la oferta alimenticia herbácea, es decir, del porcentajeque representa en el monte la superficie de unos bujeos altamente productivos y, sobre todo, de las subvenciones de la PACy de la suplementación; no obstante, dicha carga oscila entre las 0,10 -0,25 UGM (a la que sumar la carga de la fauna),fundamentalmente bovino de retinta, utilizada como vientre para cubrir con sementales de aptitud cárnica de limusina yen menor medida con charolesa, fleckvieh, rubia de Aquitania y la propia retinta. También es importante el ganado de lidia.
Para comprender el pastoreo en los montes es necesario reconocer el binomio bujeo-sierra, es decir, el pastoreo requierede la existencia de formaciones herbáceas y leñosas –frutescentes y arboladas–. En efecto, septiembre-octubre marcan elinicio del aprovechamiento en las zonas arboladas, al abrigo de las lluvias, evitando en gran medida el pisoteo y degradaciónde los fácilmente encharcables bujeos que en esa época marcan un máximo productivo relativo; al terminar la montaneray acercarse la primavera, el ganado va buscando nuevamente los bujeos, abandonando las superficies boscosas durante elestío, precisamente cuando más daño podría causar a la vegetación. Se explica así que el pastoreo es continuo en losdiferentes lotes de pastos en que quedan divididas las fincas; es habitual también la reserva en pie de algunos lotes parasu consumo durante el estío. Existen prados de siega, pero son escasos. Las mejoras realizadas consisten en la eliminaciónde la vegetación invasora (Calicotome villosa, Teline linifolia, Genista monspessulana, etc) y abonado con superfosfato.
El periodo de cubriciones va del 24 de diciembre al 24 de junio, concentrándose el 60% de los partos en noviembre-diciembre. La suplementación es práctica habitual en los meses de invierno, durante aproximadamente 2-3 meses al año,en una cuantía de 3 U.F/U.G.M3. El rebaño tipo estaría integrado por un 71% de vacas nodrizas, 15% de añojas, 10% deeralas y 4% de sementales. En montes bien gestionados paren alrededor del 80-85% de las vacas nodrizas.
2.2.3. Estudio de las cargas cinegéticas y la gestión actual
El Parque se caracteriza por una escasa presencia de cerramientos cinegéticos, razón por la que el ciervo, la especie másabundante, se desplaza de unos montes a otros en una trasterminancia que busca los bujeos en primavera y la bellota eninvierno. Por esta razón son complejos los inventarios y también la gestión de la actividad cinegética.
Figura 1. Distribución porcentual de las formaciones vegetales predominantes en el Parque Natural Los Alcornocales.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 226/276
Almoraima, 35, 2007
226
Aunque hay unas cuantas fincas con poblaciones de corzo elevadas (medias de 8-9 corzos/Km 2, hasta 25 corzos/Km2) ymínima densidad de ciervo (menos de tres individuos/Km2), lo habitual es la presencia de varias especies cinegéticas:ciervo, corzo, gamo, muflón, puntualmente jabalí en fincas cercadas y cochino asilvestrado, híbrido de cerdo y jabalí.
En general, tanto en montes públicos como privados existe una excesiva densidad de ciervo (Cervus elaphus), con unamedia hipotética estimada por la Consejería de Medio Ambiente de 20-22 ciervos/Km2; paradójicamente en los recientescerramientos en regeneración se están contabilizando entre 40-50 ciervos/Km2, densidad que limita una mayor presenciade corzo en montes con características ecológicas adecuadas, a la vez que impide la regeneración de las especies arbóreasy de las arbustivas de alto rango ecológico (pérdida de diversidad genética). El gamo y el muflón presentan una densidadmedia baja, mientras que son puntualmente altas las poblaciones de cochino asilvestrado, que consume el recurso bellota,que es posible origen de peste porcina y que está enfrentado con el corzo por su predación sobre los corcinos a pesar delas medidas adoptadas para su erradicación.
La calidad de los trofeos de ciervo es baja, en parte porque la edad media de las capturas –generalmente monterías yganchos– es baja, de 2-3 años y porque si bien la cantidad de comida es aparentemente suficiente, no lo es su calidad.
3. RESULTADOS Y DISCUSIÓN
3.1.1 Resultados dasométricos
En la figura 2 aparece una distribución numérica del número de pies de alcornoque por clases perimétricas en un cuarteltipo, con estructura prácticamente idéntica a nivel de cantón: se trataría de una masa con aparente representación de todaslas clases diamétricas por lo que en parte podría asimilarse a una masa irregular incompleta; sin embargo, no es así, ya queen parte el diferente tamaño obedece en muchas ocasiones a otros factores que no son la edad (diferente calidad del suelo,exposición, pendiente, etc) y porque se observa un marcado déficit de las clases perimétricas inferiores, uno de los gravesproblemas que se encuentran en la mayor parte de los montes; de hecho, existe cierta semirregularización del alcornoqueen el conjunto del parque.
Término Superficie Superficie Sup. Formaciones vegetales (Ha) Nº Radio Lado demunicipal total (Ha) inventariada Qs Qc-Qs Qs-Os Masa mixta Pastizal Matorral parcelas (m) malla
(Ha) arbolado arboladoZanona Los Barrios 2.573,2 1.251,7 1.251,7 203 16 250El Pedregoso Tarifa 3.972,9 2.135,1 1.065,4 376,4 225,1 468,2 104 18 400El Alisoso Medina 2.146,1 813,8 813,8 158 16 230El Sanguinar Jimena 258,4 233,9 201,8 31,9 39 18 240Las Corzas Algeciras 328,7 224,9 182,1 42,8 100 16 150Fantasma Jerez 801,9 639,4 431,9 180,9 - 26,6 107 14 250Dehesa Ojén Los Barios 1.880,0 1.880,0 Inventario pie a pie de toda la superficie - - -La Lapa Jimena 815,8 575,04 500,52 27,54 42,6 3,36 31 18 400Los Chorros Los Barrios 410,3 224,6 98,1 31,6 57,9 37,0 29 18 240Las Navas de G. Los Barrios 348,2 189,0 103,4 46,5 27,8 11,3 35 18 190La Zorrilla Los Barrios 976,4 421,2 323,7 137 18 160
Mata Ruiz Grazalema 527,9 362,5 362,5 47 16 275El Ahijón Alcalá 564,1 459,7 217,7 88,2 112,2 41,6 39 18 350Moracha Alcalá 886,2 758,9 549,7 161,9 47,3 207 15 190
TOTAL 16.490,10 10.169,74 1.236
Tabla 1. Montes inventariados, superficies y datos del inventario

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 227/276
Comunicaciones
227
En la figura 3 aparece la distribución del número de pies por hectárea de las diferentes especies inventariadas en un montetipo, poniéndose de manifiesto que tanto el acebuche como el quejigo presentan poblaciones de aparente menor edad que
el alcornoque. Además, el informe selvícola confirma que la elevada densidad de bornizo es tan solo aparente y obedecea la presencia de monte bajo de bornizos en superficies relativamente escasas pero que son capaces de desvirtuar losresultados medios del inventario.
Por último, el análisis detallado de los resultados de la tabla 2 y el trabajo de campo realizado ponen de manifiesto lossiguientes aspectos relativos a las diferentes especies principales:
Quercus suber
- Decaimiento del alcornoque detonado por las sequías y los golpes de calor, agravado por descorches extemporáneos–fuera de plazo, con lluvia, con levante seco–, descorches en años muy secos, sobre arbolado envejecido, etc. No todoes la famosa seca, existen multitud de causas involucradas.
- En general, densidad defectiva para el óptimo aprovechamiento corchero: el número de pies mayores es insuficiente paraun objetivo de producción corchera preferente que fuera además compatible con el uso ganadero y cinegético.
- No obstante, la densidad es muy variable por montes de diferentes comarcas, encontrando alcornocales mejorconservados en Jerez, norte de Alcalá y Jimena de la Frontera, sierra de Ojén y en peor estado en Tarifa, Los Barrios,etc. Los principales factores involucrados tienen que ver con la exposición, altitud y tipo de suelo, relacionado ademáscon el anterior porque las areniscas puras aparecen en las cotas más elevadas.
- También es defectiva el área basimétrica, sobre todo si se contrasta con los óptimos 22-25 m2 /Ha (Torres, 1995), si bienparece detectarse la imposibilidad de que la media de la variable alcance valores tan elevados en el conjunto del parque;es decir, hay montes cuya calidad de estación les permite alcanzar esos valores, pero son poco significativos en elconjunto (Dehesa de Ojén, Zanona, El Pedregoso, Moracha).
Figura 2. Distribución de pies/Ha de alcornoque por clases perimétricas (El Gamir).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 228/276
Almoraima, 35, 2007
228
Figura 3. Distribución de clases perimétricas de varias especies.
- Marcado envejecimiento del alcornoque, con una distribución diamétrica según campana de Gauss desplazada hacia lasclases superiores (ver figura 2), con escaso porcentaje de las clases artificiales de edad inferiores; aunque se observaen la tabla 2 que hay montes con una elevada densidad de bornizos, ésta es más bien aparente, ya que en realidad existenpequeñas superficies con densidad incluso superior a 5.000 pies/Ha que distorsionan los valores medios; en muchasocasiones los bornizos son monte bajo.
- Ausencia relativa de pies menores y regeneración, salvo contadas excepciones, irregularmente distribuida, impedida, entreotros factores (envejecimiento, descorches, decaimiento, pérdida de calidad del suelo, etc) por una acusada herbivoría.
- De hecho, se pone de manifiesto que los montes que soportan actualmente, o lo han hecho hasta el último lustro, unaelevada carga ganadera y/o cinegética presentan los valores absolutos de pies menores y de regeneración más bajos(Tarifa, El Sanguinar, Los Chorros, etc). Sólo al abrigo de matorrales densos se consigue cierta regeneración.
Número de pies mayores / Ha Área basimétrica (m2 / Ha) Pies menores / Ha Regenerado / Ha Otras variablesDescorchadosBornizos Otras TOTAL Q. suber AB otras TOTAL Q. suber Otras Otras Q. suber Otras SD ID
especies especies arbóreas arbóreas (m2/Ha)Zanona 134,6 104,1 24,3 263,0 18,5 0,9 19,4 42,0 59,0 - - 308,8 19,8El Alisoso 116,7 87,9 14,3 218,9 12,0 1,0 13,0 41,7 71,0 - - 192,6 20,1Garcisobaco 75,9 34,6 21,1 131,6 7,7 1,7 9,4 8,4 4,9 37,9 0 0 130,0 19,6
El Sanguinar 76,1 16,4 3,5 96,0 10,3 0,2 10,5 9,1 10,3 0 0 207,0 21,7Las Corzas 57,8 46,1 33,2 137,1 10,2 1,7 11,9 32,7 33,6 70,1 - - 149,0 17,3Fantasma 90,7 46,4 97,6 234,7 15,7 8,8 24,5 26,5 136,45 94,8 0 0 318,0 22,7Dehesa Ojén 121 47,0 105 273,0 14,1 8,2 22,3 - - - - 210,3 -El Pedregoso 106,0 102,0 72,0 280,0 19,1 5,2 24,3 53,4 32,0 31,1 12,5 19,8 257,4 17,5La Lapa 84,2 54,2 18,7 157,1 10,3 1,9 12,2 32,2 23,3 5,2 30,7 252,1 31,9Los Chorros 61,7 13,9 83,6 159,2 11,0 5,0 16,0 7,1 31,5 0 6,8 249,8 23,5Las Navas de G 73,0 117,8 29,2 220,0 12,8 3,3 16,1 30,9 49,9 51,7 0 0 157,5 18,1La Zorrilla 40,0 62,5 42,0 144,5 8,6 2,4 11,0 45,2 51,8 - - 136,2 23,4Mata Ruiz 92,1 30,2 77,0 196,9 12,5 6,4 18,9 9,3 40,7 0 2,1 20,9 198,0 16,0El Ahijón 47,1 29,0 45,6 121,7 5,2 3,4 8,6 51,9 24,7 1,0 18,6 7,8 79,0 19,0Moracha 128,8 30,7 28,6 188,1 18,1 6,0 24,1 12,3 0,3 6,9 0 0 482,6 27,8MEDIA 81,6 51,4 43,5 176,4 11,6 3,5 15,1 26,8 38,0 36,7 2,4 5,4 208,0 18,7
Tabla 2. Variables dasométricas.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 229/276
Comunicaciones
229
- En cualquier caso, el número de pies menores y de regeneración viable debe valorarse considerando aspectos tales comoel estado sanitario del arbolado, la densidad de la masa, la edad media de la misma, la fracción de cabida cubierta y otros;tal es el caso de montes como La Mata Ruiz y Moracha, con elevada cubierta que impide en gran medida, junto con losfitófagos, la instalación de la regeneración natural.
- El recurso corcho disminuye con el paso de los años; a la ausencia generalizada de bornizos se suma la casi total ausencia
de segunderos; la intensidad de descorche es relativamente normal al tratarse de masas no adehesadas (Montero & Grau,1986), si bien destaca por elevada la del monte La Lapa, debido sobre todo a una temprana puesta en producción delarbolado, con área basimétrica del árbol reducida.
- El valor de la superficie de descorche medio es de 208 m2 /Ha, que representa aproximadamente 40 Qcc/Ha (1 Qcc,quintal castellano, equivale a 46 Kg); dicho valor resulta excepcional en Moracha, monte que produce 85,6 Qcc/Ha.
Quercus canariensis
- Existen densos rodales de latizales que provienen del cese de las talas o podas, hacia 1960, asociado a bajas cargas deciervo, que se reintrodujo en esa década (Fantasma, Dehesa de Ojén, La Zorrilla).
- Sin embargo, ante cargas pastantes altas como las actuales la regeneración viable es escasa o inexistente en la mayoría
de los montes.- En cualquier caso, el número de pies menores y de regeneración viable frente al de pies mayores es muy superior al del
alcornoque y en los últimos años el quejigo ha ocupado, sobre todo en vaguadas y umbrías, el terreno perdido antañoen favor del alcornoque.
Olea sylvestris
- Resiste muy bien el pastoreo en zonas con cierta protección del matorral; estado de pujanza generalizado, con unarelación de pies menores y de regeneración frente a pies mayores muy superior a la del quejigo y por supuesto a la delalcornoque.
- Los acebuchales de Cádiz tienen una gran extensión e importancia ecológica y paisajística.
Figura 4. Nº de pies por clases perimétricas de varias especies.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 230/276
Almoraima, 35, 2007
230
Otras especies arbóreas y arbustivas
- Las especies tales como el aliso, fresno, álamos, madroño, agracejos, etc, alcanzan valores significativamente disparesentre montes, con valores mínimos en los excesivamente desbrozados (Moracha) o muy pastoreados (La Mata Ruíz) ymuy altos en aquellos sin ciervo (Dehesa de Ojén).
Especies catalogadas
En general se recuperan, apareciendo excelentes masas de acebo, rodales de laurel, de avellanillo, diversos pteridofitos,etc, destacando por la elevada densidad la comarca de la sierra de Ojén. Como caso particular destaca el monte Dehesa deOjén, con la cuantía de especies catalogadas que aparecen en la figura 4.
En definitiva, la persistencia del alcornocal está comprometida en amplias superficies si las condiciones macroclimáticascontinúan como hasta ahora, con la precipitación mal repartida intra e interanualmente (la precipitación del año 2005 hasido una tercera parte de la media), paulatina pérdida de microestación por pérdida de suelo, pérdida de arbolado y desombra y excesiva insolación que dificulta la regeneración, ya de por sí muy difícil por el exceso de carga de fitófagos.
4. CONCLUSIONES
4.1. Objetivos de ajuste de las cargas pastantes
El mejor mecanismo de restauración de la cubierta vegetal y de mejora de la calidad de los trofeos es el ajuste de cargaspastantes, en este caso la reducción de cargas pastantes de ganado y fauna cinegética, con la complejidad que deriva delcarácter abierto de los cotos. En efecto, las especies leñosas más palatables son consumidas por la ganadería y la caza,constituyendo en el caso del ciervo un importante recurso en determinadas épocas del año, superando como media el 20%de la dieta (Facultad de Veterinaria de Córdoba; 1990), valor que se estima similar para la vaca.
El análisis de cargas realizado en los proyectos de ordenación pone de manifiesto el sobrepastoreo observado en las especiesmás o menos palatables. Aunque la oferta alimenticia –en cantidad y calidad– no es el único factor limitante en un pastadero(agua, espacio, tranquilidad, etc) y la carga admisible no puede establecerse exclusivamente a través de su producción,siendo más lógico ir ajustando la carga de un modo empírico a partir del análisis de los signos de degradación, estabilizacióny progresión que pueden encontrarse en las diferentes áreas de pastoreo, qué duda cabe que debe aventurarse un valor decarga pastante a través de la disponibilidad forrajera de los diferentes ecosistemas, que garanticen la regeneración de lasespecies leñosas, y el consumo medio del ganado y de las especies cinegéticas.
El coste económico de cada unidad forrajera consumida es muy alto, por incidir tan severamente sobre la vegetación y elsuelo; se deben reducir las cargas de ganado controlable (bovino, sobre todo) y de ciervo, fomentando la restauración y laexpansión del corzo, limitada según la calidad de estación de cada zona. El proyecto de ordenación cinegética del ParqueNatural Los Alcornocales establece una densidad óptima de 15 ciervos/Km2 en montes con ganado y de 20 ciervos/Km2
en aquellos sin ganado; el gamo es especie cuya densidad se pretende controlar y son especies a erradicar el muflón y elcochino asilvestrado. El corzo aún puede incrementar su densidad, máxime con las mejoras realizadas (siembras, pequeñasrozas, captación de manantiales, abrevaderos, suplementación, etc), especie acorde a los objetivos de restauración de lacubierta vegetal por la selectividad de su dieta y su escasa capacidad de ingesta. Así, son muchos los montes en los que seestá reduciendo la carga ganadera o de ciervo.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 231/276
Comunicaciones
231
4.2. Objetivos de ayudas artificiales y naturales a la regeneración
Además de rebajar la carga pastante que propicie cierta regeneración continua y extendida, es necesario acometer larestauración de superficies relativamente amplias dado el considerable retraso acumulado en los logros de regeneración.Hasta el año 2000 no existían cerramientos con malla cinegética; ahora los hay tanto en montes públicos como privados,con superficie variable entre las 40-300 hectáreas (normalmente 100 Ha) y, si bien no son la mejor de las soluciones porla fragmentación del territorio que implican, es la única posibilidad actual.
Algunos criterios en la elección de superficies a regenerar son los siguientes:
- Superficies con Fcc menor del 40%, de edad avanzada, con huecos que permitan la instalación de los regenerados–naturales o artificiales–: suelos con la calidad y la capacidad ecológicas suficientes para el desarrollo de la
vegetación arbórea (microestación favorable).- Que no se dificulten las monterías o lo haga en la menor escala posible, con el objetivo de mantener una intensidad de
caza aceptable.
- Permeables al corzo e impermeables al ciervo, realizando batidas previas al cerramiento completo y recechos posterioressobre ciervo y cochino asilvestrado.
Por otro lado, se realizan ayudas a la regeneración natural con rozas selectivas bajo copas y en su periferia, y plantación ysiembra con alcornoque y quejigo, con estirpes de la zona y preparación mecanizada del terreno cuando sea posible (rasos),no considerando en principio la implantación de arbustivas. El tubo protector para las semillas, que germinan mejor y seprotegen frente a aves y roedores; plantas sin tubo en general (sólo en zonas con fuertes levantes, insolación, estrés hídrico).
REFERENCIAS BIBLIOGRÁFICASTORRES, E., 1995. Estudio de los principales problemas selvícolas de los alcornocales del Macizo del Aljibe (Cádiz y Málaga). Tesis Doctoral. E.T.S.I. Montes. Madrid.JURADO, V., 2002. Los bosques de las Sierras del Aljibe y del Campo de Gibraltar (Cádiz-Málaga). Ecología, transformaciones históricas y gestión forestal.Consejería de Medio Ambiente. Junta de Andalucía. Sevilla.Estudio 94 S.L.TORRES, E., 1995. Estudio de los principales problemas selvícolas de los alcornocales del Macizo del Aljibe (Cádiz y Málaga). Tesis Doctoral. E.T.S.I. Montes. Madrid.MONTERO, G. y J.M. Grau. 1986. El coeficiente y la intensidad de descorche. Ventajas e inconvenientes de su aplicación. I Conresso Florestal Nacional de Portugal. Lisboa.FACULTAD DE VETERINARIA DE CÓRDOBA; 1990. Departamento de Producción Animal. Aprovechamiento racional del área de interés cinegético de Zanona,Cádiz. Córdoba.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 232/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 233/276
Comunicaciones
233
RESUMEN
Las marismas y estuarios desempeñan funciones ecológicas de gran interés, prestando numerosos servicios ambientales.
Sin embargo, en muchos casos son grandes desconocidos por diversos factores. Con este trabajo se quiere dar a conoceralgunos de los recursos naturales existentes en dos espacios naturales del ámbito geográfico del campo de Gibraltar,concretamente la vegetación del Paraje Natural de las Marismas del Río Palmones y del Paraje Natural del Estuario del RíoGuadiaro. Se ha elaborado una cartografía de detalle de la vegetación de ambos espacios, y la información asociada ha sidoincorporada a una base de datos para ser utilizada por los gestores, con el fin de llevar a cabo una mejor gestión activa deestos espacios naturales.
CARTOGRAFÍA Y DESCRIPCIÓN DELA VEGETACIÓN DEL PARAJE NATURAL
DE LAS MARISMAS DEL RÍO PALMONESY DEL PARAJE NATURAL DELESTUARIO DEL RÍO GUADIARO
Carlos J. Luque Palomo / Eloy M. Castellanos VerdugoDpto. Biología Ambiental y Salud Pública. Facultad de Ciencias Experimentales. Universidad de Huelva
Antonio A. Álvarez López / Jerónimo Muñoz González Alfredo E. Rubio Casal / Enrique Figueroa ClementeDpto. Biología Vegetal y Ecología. Facultad de Biología. Universidad de Sevilla
Javier Quijada MuñozRed de Información Ambiental de Andalucía. Asesoría técnica de la Consejería de Medio Ambiente de la Junta de Andalucía. Sevilla

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 234/276
Almoraima, 35, 2007
234
1. INTRODUCCIÓN
La Consejería de Medio Ambiente de la Junta de Andalucía (CMA) y la Universidad de Sevilla firmaron en el año 2002un acuerdo para llevar a cabo un proyecto de investigación denominado “Cartografía y evaluación de la flora y vegetaciónhalófita y de los ecosistemas de marismas que se encuentren dentro de la Red de Espacios Naturales Protegidos deAndalucía (RENPA)”, financiado con fondos europeos. En dicho proyecto se han estudiado las comunidades vegetales dela mayor parte de las marismas y saladares de la costa andaluza incluidos en Espacios Naturales Protegidos, así como otrosespacios propuestos como Lugares de Interés Comunitario.
El principal objetivo de este estudio consistió en levantar la información básica de la comunidad vegetal de estos espacios,para incorporarla a una base de datos, acompañada de su base cartográfica a escala de detalle.
1.1. LocalizaciónLos espacios incluidos en este proyecto, en el contexto geográfico del Campo de Gibraltar, fueron el Paraje NaturalMarismas del Río Palmones y el Paraje Natural del Estuario del Río Guadiaro.
El primero se localiza al norte de la bahía de Algeciras (Cádiz), donde se forma un estuario en la desembocadura del ríodel mismo nombre. Cubre una superficie aproximada de 53 Ha, encontrándose la mayor parte del espacio natural situadaen una isla, dividida en dos mitades por un canal mareal que lo atraviesa de norte a sur. Este paraje natural engloba variosecosistemas, y entre ellos destacan las marismas, especialmente por la superficie ocupada, además de su interés ecológico.Son marismas mareales, con un fuerte componente halófito. La comunidad vegetal predominante está formada por especiesde la familia quenopodiácea, junto a grandes extensiones de juncales. Estas marismas se pueden considerar como mixtasdesde el punto de vista de la oscilación mareal, pues no alcanzan una amplitud tan extensa como las marismas del Golfo
de Cádiz, con oscilaciones próximas a los cuatro metros durante las mareas equinocciales, ni tampoco se comportan comolas marismas mediterráneas, con oscilaciones en torno a los 50 cm.
Por otro lado, el Paraje Natural del Estuario del Río Guadiaro se localiza al este de la costa gaditana, muy próximo a lafrontera con la provincia de Málaga. Aproximadamente ocupa 31 Ha en la margen derecha de la desembocadura del ríoGuadiaro. Este delta ha estado sometido a numerosos y grandes cambios, tanto naturales como antrópicos, a lo largo desu historia. En este paraje natural sobresale especialmente la superficie ocupada por vegetación palustre, dominada porcarrizos. Igualmente destaca una amplia zona, localizada al norte del espacio natural, formada por una revegetación, a modode parque, con unas especies de matorral mediterráneo y otras ornamentales. Este paraje no llega a formar una marisma,en su sentido estricto, debido a la escasa oscilación mareal del mar Mediterráneo, que no logra adentrarse e inundarsuficientemente el estuario. Por tanto, el componente halófilo en la vegetación es residual.
2. MATERIAL Y MÉTODOS
Este trabajo se llevó a cabo en sucesivas fases. En primer lugar se elaboró una cartografía previa de los polígonos concaracterísticas funcionales y paisajísticas más o menos homogéneas, mediante fotointerpretación a escala 1:2.500 deimágenes ortorrectificadas del vuelo del olivar de 1998 (fotogramas aéreos verticales en B/N). El conjunto de polígonoscon el mismo patrón y mismas características florísticas determina una unidad, y a cada unidad se le asignó un códigoprovisional. Es necesario resaltar que los límites de los espacios naturales aquí incluidos no corresponden exactamente conlos legalmente establecidos por la ley 2/89 de Inventario de Espacios Naturales Protegidos de Andalucía.
Posteriormente se comprobó in situ la correspondencia con las unidades especificadas, corroborando, corrigiendo o generandonuevas unidades que no habían sido fotointerpretadas. Se visitó prácticamente la totalidad de ambos espacios naturales para

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 235/276
Comunicaciones
235
definir con mayor precisión los límites y contenidos de las distintas unidades vegetales. Paralelamente, se llevó a cabo unadescripción fisionómica y de la composición florística de cada uno de los nuevos polígonos y unidades exploradas. Para ellose realizaron diversos muestreos de campo, distribuidos por todo el ámbito de estudio, y que reflejara la variabilidad realexistente. Los taxones se determinaron siguiendo a Valdés et al. (1987) y VVAA (1980-2002). Los muestreos realizados selocalizaron mediante GPS y fueron de tres tipos: A) Muestreos de cobertura lineal, denominados transectos por la CMA. Estemétodo fue descrito por Canfield (1941) como de intercepción lineal y que, en nuestro caso, se realizó con una longitud de20 m sobre la vegetación. El número total de transectos realizados en el Paraje Natural Marismas del Río Palmones ascendióa un total de tres, mientras que en el Paraje Natural del Estuario del Río Guadiaro no se hicieron transectos por el tipo devegetación existente. B) Muestreos de inventarios: se utilizó el método Braun-Blanquet, siguiendo prescripciones de la CMA,asignando a cada una de las comunidades su sintaxón correspondiente, siguiendo a Rivas-Martínez et al. (2001 y 2002). Lasuperficie de las unidades de muestreo varió en función del tipo de formación, aunque el tamaño habitual fue de 25m2. A cada
una de las especies presentes se le asignó una cobertura, utilizando el índice de abundancia-dominancia o índice de cobertura(tabla 1). El número total de inventarios fue de nueve en el Parque Natural Marismas del Río Palmones, y trece en el ParqueNatural del Estuario del Río Guadiaro. C) Notas de campo: utilizados para registrar especies poco habituales en unadeterminada comunidad, o especies de especial interés. En total se realizaron dos muestreos en el Parque Natural del Estuariodel Río Guadiaro y cuatro en el Paraje Natural Marismas del Río Palmones.
Por último, tras finalizar las labores de campo, se realizó un trabajo de gabinete para la comprobación y determinación delas unidades vegetales existentes, realizando la cartografía definitiva de las distintas unidades vegetales, y presentadasfinalmente a escala 1:5.000. Esta cartografía se asoció a una completa base de datos que describía las característicasbiogeográficas, florísticas, geobotánicas, etc. de las distintas unidades definidas (figura 1).
Figura 1. Relaciones establecidas en las bases de datos asociadas a la cartografía.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 236/276
Almoraima, 35, 2007
236
3. BIOGEOGRAFÍA Y BIOCLIMATOLOGÍA
Jerárquicamente, y siguiendo a Rivas-Martínez (1987), la fitogeografía de ambos parajes naturales se encuadra de lasiguiente forma: Reino Holártico > Región Mediterránea > Subregión Mediterránea occidental > SuperprovinciaMediterráneo-Iberoatlántica > Provincia Gaditano-Onubo-Algarviense > Sector Gaditano > Subsector Algecireño.
Según su bioclimatología, y siguiendo a Rivas-Martínez (1987), ambos parajes naturales quedan así definidos: RegiónMediterránea > Piso Termomediterráneo. Dentro de este piso, nuestra zona de estudio queda enmarcada dentro delhorizonte termomediterráneo superior. En estos espacios naturales, el ombroclima corresponde con el subhúmedo, porestar su pluviometría entre los 600 y 1.000 mm, que determina este tipo.
4. VEGETACIÓN
4.1. Vegetación potencial
La vegetación característica de ambos parajes naturales es de tipo azonal, debido a que uno o varios parámetros físico-químicos,como sucede en nuestro caso, principalmente con los gradientes de humedad, salinidad y nivel de inundación mareal se elevaen un biotopo muy por encima de los niveles de tolerancia media. En estos casos, las comunidades vegetales capaces decolonizarlos se hacen superespecializadas, pobres florísticamente y cosmopolitas. Esta vegetación azonal no va ligadaespecíficamente a una región o zona climática, sino que responde a situaciones ambientales excepcionales (Gómez et al., 1995).
Dentro del Paraje Natural de las Marismas del Río Palmones, existen dos geomacroseries de vegetación azonal: geomacroseriede saladares y salinas, y geomacroserie de dunas y arenales costeros. En la primera, las comunidades de marismas están
dominadas por unos pocos taxones de la familia Chenopodiaceae adaptados a condiciones ambientales muy extremas, comoson la salinidad y la inundación periódica. Constituidos por arbustos y subarbustos halófilos de medio o bajo porte,encuadrados en la clase SARCOCORNIETEA FRUTICOSAE Br.-Bl. & Tüxen ex A. & Bolòs 1950, principalmente.
En cuanto a la geomacroserie de dunas y arenales costeros, la comunidad vegetal está dominada por especies terófitas,vivaces (caméfitas, hemicriptófitas y criptófitas), así como por fanerófitas cespitosas, y unas pocas especies de fanerófitasescaposas adaptadas a vivir en arenales costeros y dunas. Se encuentran sometidas a la influencia directa del aire marino,y forman parte de los ecosistemas dunares litorales. Esta comunidad está estructurada por diversos tipos de vegetaciónherbácea o leñosa que tiende a evolucionar hacia otro tipo de vegetación arbustiva densa, que representa la etapa madurao final de la sucesión (Rivas-Martínez et al., 1980). La única comunidad presente es la clase AMMOPHILETEA Br.-Bl.& Tüxen 1943 em. Br.-Bl. 1952, dominada por especies vivaces principalmente.
DESCRIPCIÓN CÓDIGO1 Individuos que cubran más del 75% de la parcela 52 Individuos que cubran entre un 50 y un 75% 43 Individuos que cubran entre un 25 y un 50% 34 Individuos que cubran entre un 5 y un 25% 25 Abundante pero con valor de cobertura bajo o bien pocos individuos y pequeña cobertura 16 Individuos raros o únicos con pequeña cobertura r7 Pocos individuos y pequeña cobertura +8 Cobertura equivalente a +. Individuo fuera del inventario (+)9 Cobertura equivalente a 1. Individuo fuera del inventario (1)10 Cobertura equivalente a 2. Individuo fuera del inventario (2)
Tabla 1. Intervalos de cobertura y códigos asignados en los inventarios.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 237/276
Comunicaciones
237
En cuanto al Paraje Natural del Estuario del Río Guadiaro, la vegetación azonal existente corresponde con la “Geoserieedafohigrófila termomediterránea jerezana, onubense litoral y algarviense litoral”, por su relación con la ribera-cauce del río.También, como vegetación azonal, aparece dentro de este espacio natural, una comunidad vegetal ligada a la geomacroseriede dunas y arenales costeros, con presencia de la clase descrita anteriormente, así como la clase CAKILETEA MARITIMAE Tüxen & Preising ex Br.-Bl. & Tüxen 1952. Ambas clases están dominadas por especies vivaces principalmente.
4.2. Unidades cartografiadas
4.2.A. Paraje Natural Marismas del Río Palmones (figura 2)En la tabla 2 se indican las unidades, el número de polígonos, y la superficie total, media, máxima y mínima de las distintasunidades y polígonos que la conforman.
4.2.A.1. Zonas sin vegetación. Las zonas sin vegetación fanerógama ocupan casi la mitad de la superficie total del paraje(47,4%). En este apartado, se incluyen zonas de fangos y arenas desnudas, así como masas de agua.
Unidad 1: Fangos desnudos intermareales. Esta unidad recoge las franjas intermareales asociadas a los bordes de lared de drenaje que quedan al descubierto durante las bajamares. En general, son fangos limo-arcillosos muy húmedosde reciente deposición, por lo que son poco consistentes. Normalmente, no existe vegetación fanerógama asociada, porpresentar numerosas horas de inundación mareal.
Figura 2. Cartografía con las unidades detectadas en el Paraje Natural Marismas del Río Palmones.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 238/276
Almoraima, 35, 2007
238
Unidad 13: Caños mareales. Incluye las zonas frecuentemente cubiertas por agua en la intrincada red de drenaje quefluye por toda la marisma. El área ocupada por la red de drenaje está en torno al 33,1% de su superficie total. Por estared es por donde circula el flujo y reflujo mareal, distribuyendo la materia y energía por todo el estuario.
Unidad 14: Arenas desnudas. Zonas con sustrato arenoso desprovista de vegetación. Corresponde con la zona de playa,en la bocana del estuario, y está formado por un polígono de 0,95 Ha.
4.2.A.2. Matorral sin arbolado. El matorral sin arbolado es la comunidad vegetal más extendida por la marisma, debidoa la ausencia de estrato arbóreo natural en nuestros ecosistemas de marismas. Se encuentra formado principalmente poralmajos ( Arthrocnemum macrostachyum y Sarcocornia fruticosa), especies perennes predominantes en las marismasatlánticas andaluzas.
Unidad 3: Comunidad de Sarcocornia fruticosa, acompañada por otras quenopodiáceas y juncos (Cistancho
phelipaeae-Sarcocornietum fruticosae). En esta unidad, el suelo desnudo está prácticamente ausente. La comunidadque incluye esta unidad está dominada por Sarcocornia fruticosa, que cubre grandes extensiones de marisma media, ycon una abundancia bastante elevada, alcanzando un porte medio en torno a los 35 cm. A veces, esta especie llega aconstituir formaciones prácticamente monoespecíficas. También aparecen, con escasa abundancia, otras quenopodiáceasarbustivas como Halimione portulacoides y Arthrocnemum macrostachyum. El estrato herbáceo no supera el 25% decobertura, y la única especie herbácea registrada fue Juncus maritimus, un geófito rizomatoso bastante abundante enel paraje natural. Sólo existe una asociación en esta unidad, Cistancho phelipaeae-Sarcocornietum fruticosae. Laextensión de esta unidad es alta, 8,94 Ha, lo que supone cerca del 16,9% de la superficie total de este espacio natural.
Unidad 5: Sarcocornia fruticosa acompañada por Halimione portulacoides (Cistancho phelipaeae-Sarcocornietum
fruticosae). Esta comunidad es típica de zonas de marisma media, distribuyéndose ampliamente por todo el parajenatural. Constituida principalmente por quenopodiáceas arbustivas, con presencia de herbáceas y suelo desnudo concobertura inferior al 25%. Los arbustos no suelen alcanzar los 45 cm de altura. Se caracteriza por la presencia de
Unidad A B C D E1 Fangos desnudos intermareales 5 6.63 1.33 5.67 440.92 Pradera de Zostera noltii 5 1.51 0.30 0.83 525.63 Comunidad de Sarcocornia fruticosa , acompañada por otras
quenopodiáceas y juncos 4 8.94 2.24 5.42 597.14 Pastizal cont inuo de Juncus maritimus 5 1.57 0.31 0.74 375.95 Sarcocornia fruticosa acompañada por Halimione portulacoides 19 9.33 0.49 4.89 59.26 Bordes de caños con juncales y quenopodiáceas 12 1.24 0.10 0.30 136.27 Poza con escasa vegetación 10 0.073 0.007 0.015 23.68 Vegetación de marisma media-alta acompañada de pastizal no halófilo 2 0.15 0.072 0.076 690.99 Juncal con Halimione portulacoides acompañado por otras halófilas 7 3.41 0.49 2.73 214.910 Pastizal nitrófilo en suelo de marisma 2 1.03 0.52 0.83 2046.4
11 Mar de Spartina densiflora 1 0.050 - - -12 Pastizal de Spartina maritima con quenopodiáceas acompañantes 3 0.26 0.087 0.23 34.413 Caños mareales 1 17.52 - - -14 Arenas desnudas 1 0.95 - - -15 Arenas con colonizadoras primarias 1 0.33 - - -
A = Frecuencia; B = Superficie total (Ha); C = Superficie media (Ha); D = Polígono máxima superficie (Ha); E = Polígono mínimasuperficie (m2).
Tabla 2. Unidades del Paraje Natural Marismas del Río Palmones.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 239/276
Comunicaciones
239
Sarcocornia fruticosa y Halimione portulacoides. Junto a éstas aparecen dos herbáceas, Scirpus maritimus subspmaritimus y Juncus maritimus, ambos geófitos rizomatosos. La comunidad predominante es la asociación Cistancho
phelipaeae-Sarcocornietum fruticosae. Es la unidad con vegetación más extensa del espacio natural, 9,33 Ha, lo quesupone el 15% de la superficie total.
Unidad 8: Vegetación de marisma media-alta acompañada de pastizal no halófilo ( Inulo crithmoidis-
Arthrocnemetum macrostachyi). Formada por grandes extensiones de Arthrocnemum macrostachyum, especiecaracterística de la alianza Arthrocnemion macrostachyi, y de la asociación Inulo crithmoidis-Arthrocnemetummacrostachyi. Esta comunidad se encuentra en zonas de marisma media-alta y, por tanto, con menor influencia mareal.También aparecen otras especies menos abundantes, como Halimione portulacoides y Sarcocornia fruticosa. Esteestrato arbustivo ocupa el 70% de la cobertura vegetal de la unidad. Además, el estrato herbáceo cubre aproximadamente
un 30%, siendo Oxalis pes-caprae la especie más abundante. La altura media de la vegetación es de 45 cm. Esta unidadcubre sólo 0,15 Ha, distribuidas en dos polígonos.
4.2.A.3. Vegetación acuática. En este apartado se incluyen las comunidades caracterizadas por permanecer más de lamitad del día cubiertas por las aguas mareales.
Unidad 2: Pradera de Zostera noltii ( Zosteretum noltii). Esta unidad se caracteriza por la presencia de la fanerógamamarina Zostera noltii, especie característica de la asociación Zosteretum noltii. Su hábitat lo constituyen fangosintermareales cubiertos por las aguas del estuario durante periodos prolongados. Estas condiciones son tan restrictivasque impiden la colonización de otras fanerógamas. La cobertura de esta especie puede alcanzar el 95% de la unidad. Launidad está formada por cinco polígonos, y ocupa una superficie total de 1,51 Ha. Un hecho de gran interés es que esteespacio natural es el límite oriental de la distribución de fanerógama marina Zostera noltii en las marismas de Andalucía
estudiadas en este proyecto.
4.2.A.4. Pastizales. En esta agrupación se recogen varias unidades caracterizadas por la elevada cobertura relativa de suestrato herbáceo. Los pastizales muestran diferentes características, como son los halófilos, típicos de marismas saladas;
juncales, formados por geófitos junciformes; pastizales colonizadores de arenas, etc.
Unidad 4: Pastizal continuo de Juncus maritimus ( Juncetalia maritimi). Está formada por un pastizal continuo de Juncus maritimus, que ocupa el 99% de la cobertura vegetal. Junto a este geófito rizomatoso solo aparece Sarcocornia fruticosa, y la altura media de la comunidad es de 80 cm. La comunidad predominante es Juncetalia maritimi. Su hábitatcaracterístico corresponde con zonas cercanas a caños pequeños o áreas encharcadas. Aparecen cinco polígonosdistribuidos por todo el paraje natural, sumando un total de 1,57 Ha.
Unidad 6: Bordes de caños con juncales y quenopodiáceas ( Juncetalia maritimi). En esta unidad se ha registradouna relativamente elevada diversidad específica, con dos especies de herbáceas y cinco arbustivas. La cobertura de lavegetación es del 100%. La especie más abundante es Juncus maritimus, que ocupa en torno al 70% de la coberturavegetal, completando ésta, Sarcocornia fruticosa y Halimione portulacoides. La altura de la comunidad vegetal es de70 cm. Otras especies menos abundantes son Inula crithmoides, Arthrocnemum macrostachyum, Suaeda maritimasubsp. maritima y Sarcocornia perennis subsp. perennis. Esta comunidad dominante es el orden Juncetalia maritimi,seguida de la asociación Cistancho phelipaeae-Sarcocornietum fruticosae. Se distribuye por todo el paraje, cubriendouna superficie de 1,24 Ha.
Unidad 7: Poza con escasa vegetación ( Juncetalia maritimi). Las pozas son pequeñas depresiones de escasaprofundidad (en torno a 20-30 cm) encharcadas temporalmente. Sus diez polígonos ocupan una superficie total de 0,073

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 240/276
Almoraima, 35, 2007
240
Ha. Se localizan principalmente en la marisma media-alta de la zona central del paraje. La escasa vegetación asociadaa esta unidad se encuentra rodeando el perímetro de las pozas, siendo Juncus maritimus la única especie registrada enestos bordes.
Unidad 9: Juncal con Halimione portulacoides acompañado por otras halófilas ( Juncetalia maritimi). La coberturavegetal en esta unidad es del 100%, entre el estrato herbáceo y arbustivo. La altura media de la vegetación se acerca alos 50 cm. Las especies más abundantes son Juncus maritimus y Halimione portulacoides. Además de estas dos especies,también se aparecen Sarcocornia fruticosa, en el estrato arbustivo, y la compuesta terófita Cotula coronopifolia, en elherbáceo. Como acompañantes arbustivas se encuentran tres especies de caméfitos de la familia Plumbaginaceae,
Limonium angustifolium, L. ovalifolium y L. ferulaceum. Entre las herbáceas presentes aparecen diversas gramíneas ySuaeda maritima subsp. maritima. El orden Juncetalia maritimi y la asociación Cistancho phelipaeae-Sarcocornietum
fruticosae dominan la comunidad. Esta unidad está repartida en siete polígonos en el este y oeste del paraje, cubriendoun total de 3,41 Ha.
Unidad 10: Pastizal nitrófilo en suelo de marisma ( Fumarion wirtgenii-agrariae). Coincide con un área dondeposiblemente existió alguna pequeña instalación o asentamiento. Por esto, se desarrolla una vegetación de grancomponente ruderal y nitrófilo sobre suelo de marisma, con el estrato herbáceo como elemento principal. La coberturade la vegetación es del 100%, y la altura media de unos 30 cm. Es la unidad de mayor riqueza específica del paraje natural.Las especies más abundantes son Oxalis pes-caprae y algunas gramíneas. También aparecen Phragmites australis,geófito rizomatoso de altura superior a los dos metros, y la compuesta Dittrichia viscosa, asociada a zonas degradadaso transformadas. Las demás especies presentan una menor abundancia. Esta comunidad queda integrada dentro de laalianza Fumarion wirtgenii-agrariae, dominada por anuales nitrófilas y ruderales de invierno termomediterráneo.
Distribuida en dos polígonos, al noroeste y suroeste de la marisma.Unidad 11: Comunidad de Spartina densiflora (Spartinetum densiflorae). Esta unidad alberga la única población dela gramínea Spartina densiflora registrada en las marismas del río Palmones. Se localiza al este del Paraje, cercana ala bocana del estuario. Ocupa unos 500 m2 de superficie. Son pastizales muy densos y continuos, donde la cobertura dela vegetación alcanza el 100%, y con una altura media de 180 cm. Ésta es la especie dominante, un geófito rizomatoso,es característica de la asociación Spartinetum densiflorae. Las únicas especies acompañantes son Sarcocornia fruticosay Juncus maritimus.
Unidad 12: Pastizal de Spartina maritima con quenopodiáceas acompañantes (Spartinetum maritimae). Esta unidadincluye una de las especies pioneras o colonizadoras primarias de los fangos intermareales. La cobertura vegetal de launidad es del 100%, y la especie predominante es la gramínea geófita rizomatosa Spartina maritima. La altura media de
la comunidad es de unos 25 cm. También aparecen dos quenopodiáceas, Sarcocornia perennis subsp. perennis ySarcocornia fruticosa, aunque con escasa cobertura. La única asociación presente en esta unidad es Spartinetum maritimae.Se han localizado tres poblaciones de Spartina maritima al este del paraje, cuya superficie total cubre 0,26 Ha.
Unidad 15: Arenas con colonizadoras primarias (Otanthus-ammophiletum arundinaceae). Comunidad de coloni-zadoras de arenas costeras, especies pioneras importantes en la formación de dunas costeras. La cobertura vegetal esinferior al 25%, y su altura media alcanza los 20 cm. Las especies registradas son la gramínea Ammophila arenariasubsp. arundinacea, acompañada por Eryngium maritimum, Polygonum maritimum y Cakile maritima subsp. maritima.Esta unidad se localiza al este del paraje, junto a la zona de playa. Esta comunidad corresponde con la asociaciónOtanthus-ammophiletum arundinaceae.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 241/276
Comunicaciones
241
4.2.B. Paraje Natural del Estuario del Río Guadiaro(figura 3)
En la tabla 3 se indican las unidades, el número de polígonos,y la superficie total, media, máxima y mínima de las distintasunidades y polígonos que la conforman.
4.2.B.1. Zonas sin vegetación. Ocupan una superficie de9,12 Ha, lo que supone un 28,7% del paraje.
Unidad 11: Arenas desnudas. Zonas con sustrato areno-so de procedencia mareal desprovista de vegetación. Si-
tuada en la playa y desembocadura del río, ocupa unasuperficie de 4,16 Ha.
Unidad 12: Pasarelas de madera. Existen dos pasarelasde maderas, instaladas para el uso público, situadas una alnorte y otra al sur.
Unidad 14: Lagunas. Las lagunas litorales son masas deagua dulce o salobre, de superficie superior a variasdecenas de metros cuadrados, y sin influencia mareal. Sonfragmentos del antiguo cauce del río Guadiaro. Son doslagunas situadas al sur y oeste del espacio natural.
Unidad 15: Zona residencial. Incluye parte de las vivien-das localizadas en el límite suroeste.
Unidad 16: Desembocadura del Río Guadiaro. Se tratade parte del cauce del río Guadiaro.
4.2.B.2. Matorral con arbolado: Incluye las unidades con predominio del estrato arbustivo y presencia de arbolado. Cubreuna superficie de 6,38 Ha, lo que representa el 20,1% del paraje.
Unidad 9: Matorral denso con arbolado aclarado y carrizo. Este espacio ha sido sometido a un proceso derevegetación, y su aspecto recuerda más a un parque que a una zona natural. Se caracteriza por la elevada presencia deespecies de matorral mediterráneo. La cobertura de la vegetación es del 100% y la altura media alcanza los 350 cm. La
especie predominante es Pistacia lentiscus (lentisco), con cobertura del 70%, y que puede llegar a desarrollar portearbóreo. También aparecen con elevadas coberturas Rubus ulmifolius, y Olea europea (acebuche). Otras acompañantesson Phragmites australis, Arundo donax, Tamarix gallica, Polygonum equisetiforme, Retama monosperma, Oxalis pes-caprae, etc. Se localiza al norte del paraje natural, y ocupa 4,56 Ha, lo que supone un 14,34% del área total.
Unidad 18: Matorral denso con arbolado acompañado de pastizal. Está dominada por un matorral bien desarrolladoacompañado de arbolado. La vegetación ocupa el 100% de la cobertura, y su altura media alcanza los 5 m. En el matorraldestacan Pistacia lentiscus y Rubus ulmifolius. En el estrato arbóreo dominada Olea europea, que alcanza el 50% decobertura vegetal, junto a Tamarix gallica y Cupressus sempervirens. El estrato herbáceo, si bien está presente, cubreuna escasa superficie. Esta unidad se localiza al noroeste del paraje.
Figura 3. Cartografía con las unidades detectadas en el Paraje Natural delEstuario del Río Guadiaro.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 242/276
Almoraima, 35, 2007
242
4.2.B.3. Vegetación acuática. Se incluyen las unidades constituidas por comunidades dominadas por especies típicamente
acuáticas. Estas comunidades se caracterizan por encontrarse permanentemente inundadas, ya sea en el cauce principal oen zonas interiores.
Unidad 7: Charcas eutrofizadas con vegetación acuática ( Ruppion maritimae). Se trata de una masa de aguatemporal ubicada en una depresión del terreno, antiguo cauce cegado del río, y con una profundidad de escasoscentímetros, y por tanto, tiene una gran dependencia de las precipitaciones o crecidas del río. Habitada por la fanerógamaacuática Ruppia maritima, característica de la alianza Ruppion maritimae. Se localiza en una charca con una superficiede 0,24 Ha al este del paraje natural.
4.2.B.4. Vegetación palustre
Unidad 1: Carrizal-cañaveral ( Phragmition australis). Esta unidad se caracteriza por contener formaciones mixtasde Phragmites australis y Arundo donax. La cobertura vegetal de esta unidad es del 100% y su altura supera los dosmetros. La alianza Phragmition australis domina la comunidad. Las únicas especies acompañantes observadas son lasherbáceas anuales Salsola kali y Parietaria judaica. Esta unidad aparece al oeste del paraje y rodea el antiguo brazo delrío Guadiaro antes de su colmatación.
Unidad 2: Cañaveral ( Phragmition australis). Caracterizada por ser formaciones monoespecíficas de Arundo donax.La cobertura vegetal de esta unidad es del 100% y su altura alcanza los cuatro metros. No existen especies acompañantes.La alianza Phragmition australis domina la comunidad. Aparece en polígonos de poca superficie, y suele estar asociadaa zonas donde el suelo posee un grado de humedad alto.
4.2.B.5. Pastizales. Se incluyen todas las unidades donde el estrato herbáceo tiene una importante presencia, independien-temente de su ecología. Ocupa 12,38 Ha, es decir, el 38,9% del paraje natural.
Unidad A B C D E1 Carrizal-cañaveral 3 1.58 0.52 0.87 11362 Cañaveral 6 0.28 0.04 0.12 1493 Pastizal nitrófilo con vegetación palustre y matorral disperso 1 2.33 - - -4 Pastizal de Cynodon dactylon con vegetación palustre 2 6.39 3.19 6.32 7045 Pastizal degradado acompañado de matorral 1 0.03 - - -6 Arenas con colonizadoras primarias 3 1.41 0.47 1.22 2357 Charcas eutrofizadas con vegetación acuática 1 0.24 - - -8 Tarajal con vegetación riparia 2 0.57 0.28 0.42 14409 Matorral denso con arbolado aclarado y carrizo 5 4.56 0.91 1.66 257710 Plantaciones de Cortadeira selloana 4 1.38 0.35 0.92 69511 Arenas desnudas 4 4.16 1.04 3.62 24612 Pasarelas de madera 2 0.30 0.15 0.016 135213 Cinturón de Juncus con vegetación hidrófila acompañante 6 0.84 0.14 0.40 14114 Lagunas 2 0.63 0.32 0.33 301815 Zona residencial 1 0.34 - - -16 Desembocadura del Río Guadiaro 1 3.7 - - -17 Tarajal en arenas 3 1.23 0.41 0.75 92918 Matorral denso con arbolado acompañado de pastizal 2 1.82 0.91 0.97 845
A = Frecuencia; B = Superficie total (Ha); C = Superficie media (Ha); D = Polígono máxima superficie (Ha); E = Polígono mínimasuperficie (m2).
Tabla 3. Unidades del Paraje Natural del Estuario del Río Guadiaro.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 243/276
Comunicaciones
243
Unidad 3: Pastizal nitrófilo con vegetación palustre y matorral disperso ( Fumarion wirtgenii-agrariae). Secaracteriza por la presencia de un pastizal bien desarrollado, dominado por Oxalis pes-caprae, especie asociada a zonasdegradadas y/o transformadas, y que alcanza el 60% de la cobertura vegetal. Además, aparecen otras herbáceas comoPolygonum aviculare, Cortaderia selloana, Anagallis arvensis, Euphorbia terracina, Medicago polymorpha subsp.
polymorpha, etc. También se encuentran en esta unidad las hidrófilas Phragmites australis y Juncus acutus, querepresentan el 35% de la cobertura de la vegetación. La altura media de esta comunidad está en torno a un metro. Entreotras especies acompañantes destacan Pistacia lentiscus, con una cobertura significativa y dispuesta en las zonas mássecas y elevadas topográficamente, además de Dittrichia viscosa, Rubus ulmifolius y Asparagus aphyllus.Fitosociológicamente, domina la alianza Fumarion wirtgenii-agrariae.
Unidad 4: Pastizal de Cynodon dactylon con vegetación palustre (Trifolio fragiferi-Cynodontion). Está formada por
un pastizal denso donde la cobertura vegetal es del 100% y su altura media de 70 cm. Está dominada por la gramíneaCynodon dactylon, que ocupa más del 75% de esta cobertura. Además, existe un estrato abundante de vegetación palustrede Juncus inflexus y Phragmites australis con coberturas superiores al 40% entre ambas. Esta unidad se localiza en la mitadsur del espacio natural, ocupando un antiguo cauce del río Guadiaro. Se trata de la unidad más extensa del paraje natural,pues cubre 6,39 Ha, que representa el 20,10%. La alianza Trifolio fragiferi-Cynodontion domina la unidad.
Unidad 5: Pastizal degradado acompañado de matorral ( Polygono arenastri-Poetalia annuae). Se encuentrasituada en la orilla del río Guadiaro y próxima a su desembocadura, ocupando 0,03 Ha. Se trata del único lugar del parajedonde se ha registrado una especie halófila, concretamente Inula crithmoides, aunque con escasa cobertura. Predominangramíneas y compuestas anuales comunes de zonas degradadas. En esta comunidad destaca el orden Polygonoarenastri-Poetalia annuae, con las especies características Polygono aviculare y Plantago coronopus, perteneciente a
la clase Polygono-Poetea annuae (comunidad de anuales efímeras pioneras y nitrófilas antropozoogénicas).Unidad 6: Arenas con colonizadoras primarias ( Ammophiletea). Caracterizada por la presencia de pioneras de arenaslitorales, donde la cobertura vegetal es inferior del 6%, y la altura media en torno a cinco centimetros. Se localiza al surdel paraje, junto a la zona de playa. En el inventario realizado, sólo se han observado tres especies, Polygonummaritimum, Cakile maritima subsp. maritima y Salsola kali.
Unidad 10: Plantaciones de Cortadeira selloana. Esta unidad se caracteriza por la elevada presencia de una especieornamental exótica, la gramínea Cortaderia selloana (plumero de la Pampa). La cobertura vegetal es del 100%, y estaespecie ocupa la mayor parte de ella. Las otras dos especies presentes son Pistacia lentiscus y Rubus ulmifolius. La alturamedia de la vegetación es de cuatro metros y domina el estrato herbáceo al que pertenece la citada gramínea.
Unidad 13: Cinturón de Juncus sp. con vegetación hidrófila acompañante ( Phragmition australis). Esta unidad secaracteriza por formar cinturones de juncos alrededor de las masas de agua, antiguos cauces del río Guadiaro. Lacobertura vegetal de la unidad del 100% y su altura media supera el metro. El estrato herbáceo es el único presente enesta unidad, en la cual aparecen tres especies, Juncus inflexus, Phragmites australis y Scirpus maritimus. La alianzaPhragmition australis domina en esta unidad.
4.2.B.6. Bosques. Agrupa las unidades dominadas por el estrato arbóreo. En estos casos son especies consideradasnormalmente arbustivas, que alcanzan dicho porte arbóreo. Ocupa 1,80 Ha, lo que representa el 5,66% del paraje natural.
Unidad 8: Tarajal con vegetación riparia (Tamaricetum gallicae). En el bosque de tarajes, la cobertura vegetal esdel 100 %, alcanzando una altura media de hasta cinco metros. El estrato arbóreo es el predominante, en el que Tamarixgallica (taraje) posee coberturas entre el 40% y el 90%. En este estrato también puede encontrarse ocasionalmente

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 244/276
Almoraima, 35, 2007
244
Fraxinus angustifolia. El estrato arbustivo es el segundo en dominancia, con coberturas entre el 25% y el 50%, siendo Rubus ulmifolius la especie más abundante, junta a Pistacia lentiscus. El estrato herbáceo es el menos abundante, concoberturas inferiores al 25%. Phragmites australis es la especie acompañante más abundante de este estrato, junto a
Juncus inflexus. Esta unidad se localiza al norte del paraje, formando un pequeño bosque en galería sobre un cauce deagua. La asociación Tamaricetum gallicae domina la comunidad. Además, aparecen dos comunidades secundarias, losespinares mixtos representados por la alianza Pruno-Rubion ulmifolii, y la alianza Phragmition australis, representandocomunidades anfibias de helófitos de escaso porte.
Unidad 17: Tarajal en arenas (Tamaricetum gallicae). Este bosque de tarajes se diferencia de la anterior unidad,principalmente por situarse sobre sustrato arenoso, además de la composición de algunas especies y de su abundancia.La cobertura vegetal es del 100%, alcanzado una altura media de hasta cinco metros. El estrato arbóreo está caracterizado
por Tamarix gallica, con coberturas superiores al 75%. El segundo estrato en dominancia es el herbáceo, que representaentre el 25% y el 40% de la cobertura vegetal, destacando Phragmites australis. Por último, el estrato arbustivo noalcanza el 25% de la cobertura de la vegetación. La especie más abundante de este estrato es Dittrichia viscosa y, además,aparecen Rubus ulmifolius y Pistacia lentiscus. Esta comunidad se encuentra dispersa en tres núcleos, dos de ellossituados al noreste y otro al sureste del Paraje. La asociación Tamaricetum gallicae domina la comunidad, donde aparecesecundariamente, la alianza Phragmition australis.
INTERÉS DEL TRABAJO
Toda esta información se integrará en la Red de Información Ambiental de Andalucía de la CMA de la Junta de Andalucía.
De este modo, se podrá obtener una rápida información adecuada, selectiva, relevante, precisa y fiable de estos ParajesNaturales, al disfrutar de una completa base de información que ayudará a llevar a cabo una correcta planificación yordenación de los recursos, y ejecutar una correcta política de gestión activa en nuestros Espacios Naturales Protegidos porparte de la Consejería de Medio Ambiente.
BIBLIOGRAFÍACANFIELD, R.: “Application of the line interception method in sampling range vegetation”. J. Forestry , 39 (1941), pp. 388-394.GÓMEZ, F., J.F. Mota, J. Peñas, J. Cabello y F. Valle: “Vegetación del Parque Natural de la Sierras Subbéticas”. Reconocimiento biofísico de Espacios NaturalesProtegidos: Parque Natural Sierras Subbéticas. Moreira, J.M. (coord.). Consejería de Medio Ambiente. Junta de Andalucía. 1995.RIVAS-MARTÍNEZ, S., M. Costa, S. Castroviejo y E. Valdés: “Vegetación de Doñana (Huelva, España)”. Dpto. Botánica. Universidad Complutense. Lazaroa, vol.2. (1980). Madrid.RIVAS-MARTÍNEZ, S.: Mapa de series de vegetación de España. ICONA. Madrid. 1987.
RIVAS-MARTÍNEZ, S., T.E. Díaz, F. Fernández-González, J. Izco, J. Loidi, M. Lousa y A. Penas: “Addenda to the syntaxonomical checklist of 2001”. ItineraGeobotanica, vol.14 (I) y (II). (2002).RIVAS-MARTÍNEZ, S., F. Fernández-González, J. Loidi, M. Lousa y A. Penas: “Syntaxonomical checklist of vascular plant communities of Spain and Portugal toassociation level”. Itinera Geobotanica , vol.14. (2001).VALDÉS, B; S. Talavera y E. Fernández-Galiano: Flora Vascular de Andalucía Occidental. Ketres editora, S.A. Barcelona. 1987.VVAA (1980-2002). Flora Ibérica. Castroviejo, S. (Coord.). Vols. 1-8, 14.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 245/276
Comunicaciones
245
RESUMEN
En el presente trabajo se ha realizado un análisis morfológico del medio físico del Parque Natural Los Alcornocales. Paraello se ha utilizado como fuente de información básica un modelo digital de elevaciones, a partir del cual se han estudiadodistintas características topográficas, como la pendiente, el relieve local y la curvatura del terreno. El análisis integrado deestos parámetros mediante SIG ha permitido la clasificación morfológica automática del terreno.
Palabras clave: Sistema de información geográfica; Modelo digital de elevaciones; Cartografía geomorfológico; Parque
Natural Los Alcornocales.
ABSTRACT
A morphological analysis of the physical way of the Natural Park Los Alcornocales has been carried out in this paper. Withthis goal, a digital elevation model has been used as the main source of basic information, so that different topographiccharacteristics have been studied, as the slope, the local relief and the curvature of the surface. The integrated analysis of these parameters by GIS has allowed the morphologic automatic classification of the area.
Keywords: Geographical information system; Digital elevation model; Geomorphological mapping; Los AlcornocalesNatural Park.
APLICACIÓN DE UN SISTEMA DEINFORMACIÓN GEOGRÁFICA ALANÁLISIS DEL MEDIO FÍSICO EN EL
PARQUE NATURAL LOS ALCORNOCALES.APROXIMACIÓN A UNA CARTOGRAFÍAGEOMORFOLÓGICA A PARTIR DEUN MODELO DIGITAL DE ELEVACIONES
Lorena Martínez-Zavala / Antonio Jordán López / Patricia Illana Sanmiguel

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 246/276
Almoraima, 35, 2007
246
INTRODUCCIÓN
Hasta hace relativamente poco tiempo, los estudiosgeomorfológicos estaban basados en el análisis de caracterís-ticas del relieve a partir de mapas topográficos o en la medidadirecta de determinados parámetros en el campo. En laactualidad, el uso de los Sistemas de Información Geográfica(SIG) permite el estudio detallado de un gran número devariables del medio físico.
El conocimiento profundo de las características topográficasdel medio constituye un punto de partida para el estudio de lossuelos debido a su influencia sobre la sedimentación y arras-tre de partículas, nutrientes, el movimiento del agua superfi-cial y muchos otros aspectos importantes (Jenny, 1941;Gerrard, 1981). Sin embargo, la topografía del terreno noinfluye sólo sobre los procesos edáficos, sino que la variedadde formas puede reflejar cambios menos perceptibles en elsustrato, la insolación, la disponibilidad de agua o la vegeta-ción (Schaub & Jerold, 1999; Lark, 1999; Ruiz et al., 1999;Martínez-Zavala, 2001; Bellinfante et al., 2002).
Un modelo digital del terreno constituye una herramienta básica
en el estudio de los factores del medio físico y de su interrelación.A partir de una estructura básica, como un modelo digital deelevaciones (MDE), es posible construir otros modelos deriva-
dos mediante cálculos matemáticos más o menos simples (Doyle, 1978). La determinación automática de las formas del terrenoelaborada a partir de la modelización de las características del terreno permite un conocimiento profundo del medio físico yconstituye una herramienta rápida y eficaz en la gestión y planificación de los espacios naturales protegidos.
El Parque Natural Los Alcornocales (figura 1) se encuentra localizado en la provincia de Cádiz, aproximadamente entrelas coordenadas 36º50’-36º00’ N y 5º50’-5º20’ W. El parque incluye las sierras del Aljibe y de Algeciras, constituidas porel afloramiento de estratos de arenisca silícea alternados con otros sustratos de naturaleza caliza o margosa.
El clima es de tipo mediterráneo, con una temperatura media que oscila entre 15 y 18ºC. La precipitación anual es variable,
ya que existe un gradiente de precipitación muy acusado que crece desde la costa hasta las sierras, y oscila entre 660 y 1.400mm (Jordán y Bellinfante, 2000).
En el área de estudio, Jordán (2000) estableció cinco sistemas de paisaje principales:
1. Las sierras del Campo de Gibraltar y del Aljibe, que constituyen un conjunto de morfogénesis estructural denudativa,formado por afloramientos de areniscas silíceas (Oligoceno-Mioceno) fuertemente plegados. Aunque no se trata desierras muy elevadas (la máxima elevación es la de El Aljibe, con 1.092 metros sobre el nivel del mar), la erosióndiferencial de los estratos consolidados y no consolidados ha generado un relieve extraordinariamente irregular (conpendientes superiores al 70%). La presencia de capas de margas y arcillas intercaladas entre la arenisca favorece losprocesos de deslizamiento, especialmente tras períodos de intensas lluvias.
Figura 1. Área de estudio.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 247/276
Comunicaciones
247
2. Los valles e intercalaciones que cruzan las sierras del Aljibe y Algeciras están constituidos por materiales carbonatadoscomo calcarenitas y margas. Los afloramientos de mayor extensión se localizan en las proximidades de Jimena de laFrontera, al norte de la sierra del Niño y en una banda alargada cruzada por el curso bajo del arroyo de la Miel (Algeciras).También son frecuentes los conjuntos alomados formados por arcillas de Facinas, principalmente en el sector oeste delárea de estudio (en Tarifa o en los valles del Almodóvar y el Tiradero). En general se trata de terrenos de laderassuavemente inclinadas y de poca elevación.
3. Complejo de colinas margo-areniscosas de Jimena de la Frontera. Este sistema coincide básicamente con los valles delGuadiaro y el Genal. Se trata de un sistema de morfogénesis estructural denudativa que presenta un relieve variado ypendientes que oscilan entre el 0 y el 46%. El sustrato litológico está constituido por afloramientos alternantes, y a vecespoco diferenciados, de areniscas micáceas y margas.
4. Sistema de arcillas y margas de Tarifa y Tahivilla. Otros afloramientos del Flysch margoso-areniscoso micáceo deAlgeciras se observan también en puntos de Tarifa y Tahivilla, con una topografía y fisiografía similar, aunquepredomina el sustrato arcilloso. Las lomas y colinas son de escasa elevación, pero la pendiente puede ser elevada.
5. Dentro del Parque Natural Los Alcornocales existen diversos afloramientos de calcarenitas y biocalcarenitas.Principalmente, estos afloramientos se sitúan sobre algunos puntos al sur de la porción occidental de sierra Bermeja,en Jimena de la Frontera. Se trata de materiales que datan del Eoceno y Paleoceno (Terciario), y Cretácico superior. Lapendiente media de las distintas unidades es muy diversa, de hasta el 50%. Como consecuencia, el paisaje es variado,con zonas de fisiografía suave, lomas, colinas y cerros.
OBJETIVOSEl objetivo principal de este trabajo es la realización de un modelo digital del terreno que describa las característicasgeomorfológicas del Parque Natural Los Alcornocales. Este objetivo implica los siguientes pasos:
- Realización de un modelo digital de elevaciones.
- Realización de modelos derivados (pendiente, curvatura y relieve local).
- Realización de un modelo digital morfológico del terreno.
- Descripción de las principales geoformas del área de estudio.
METODOLOGÍA GENERALEl método utilizado en la diferenciación de las formas del terreno está basado en los procedimientos de clasificaciónautomática utilizados por Hammond (1964), Dikau et al. (1991) e Iriagaray et al. (1997), con algunas modificacionessugeridas por Martínez-Zavala et al. (2005).
Como fuente básica de información se ha utilizado un modelo digital de elevaciones con una resolución de 100 x 100 m, apartir del cual se ha obtenido información acerca de otras características del terreno, como la pendiente, la curvatura de lasuperficie o el relieve.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 248/276
Almoraima, 35, 2007
248
Modelo digital de elevaciones
Un modelo digital de elevaciones (MDE) se define como una estructura numérica que representa la distribución espacialde la altitud de la superficie del terreno. De esta forma, un MDE puede describirse mediante la siguiente ecuación(Felicísimo, 1994):
z = ζ (x, y)
donde:
- z es la elevación del punto situado en las coordenadas x e y.
- ζ es la función que relaciona la variable z con su localización geográfica.
El MDE fue construido utilizando un método de interpolación de elementos finitos. Para ello se digitalizaron las curvasde nivel a partir de mapas topográficos (E 1:50.000), así como algunos puntos de elevación conocida (puntos geodésicos,picos, accidentes geográficos, etc.). Una vez realizada la interpolación se comprobó la exactitud del modelo mediante lacomparación de la red hidrográfica real con la generada a partir del MDE.
La unidad mínima de información corresponde a una celda de 100 x 100 m de superficie. Esta precisión se considerasuficiente para conseguir los objetivos formulados inicialmente.
Gráficamente, el MDE puede representarse codificando las altitudes en tonos de gris, de forma que los tonos más claroscorresponden a las zonas más elevadas y las zonas más oscuras a los puntos de menor altitud.
Modelo digital de pendientes
La pendiente puede calcularse a partir del MDE, como la máxima tasa de cambio de cada celda respecto a sus vecinas. Pararealizar el cálculo de la pendiente es necesario definir en primer lugar el gradiente topográfico. En un MDE se puede describirla elevación en el entorno de un punto problema como un plano de ajuste, cuya expresión matemática sería la siguiente:
z = a00 + a10 · x + a01 · y
donde los coeficientes a10y a01
corresponden a las derivadas de la elevación con respecto a los ejes X e Y, respectivamente:
a10
= ∂ z ;a01
= ∂ z∂ x ∂ y
Estos coeficientes pueden representarse mediante los vectores V #y W #, definidos por las componentes para los ejes X, Y yZ. El producto vectorial entre estos dos vectores es otro vector, P#, perpendicular a ambos y al plano que los contiene, es
decir, a la superficie del terreno.∂ x 0 – ∂ z · ∂ y – ∂ z / ∂ z – a
10
P#= V #· W #= 0 · ∂ y = – ∂ x · ∂ z = – ∂ x / ∂ y = – a 01
∂ z ∂ z – ∂ x · ∂ y 1 1
La pendiente en un punto del terreno se define como el ángulo existente entre el vector gradiente en ese punto, P#, y el ejeZ (vertical). El eje Z puede expresarse también como un vector, H #. El ángulo (α) entre estos dos vectores puede calcularsede la siguiente manera:
α = cos–1 P#· H # = 1 = tan–1 √ a210
+ a201
mod (P#) · mod ( H #) √ a210 + a2
01 + 1

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 249/276
Comunicaciones
249
donde mod(P#) es el módulo del vector P#, y mod( H #) el del vector H #.
La pendiente de cada celda ha sido calculada a partir de un plano ajustado a los valores de elevación de un grupo de 3x3celdas centrado en ella (Burrough, 1986). Debido a que esta expresión proporciona el valor de la pendiente en radianes,es necesario transformarlo a unidades porcentuales.
La pendiente se clasificó en rangos según el criterio que se muestra en la tabla 1.
Modelo digital de curvatura
Un modelo digital de curvatura del terreno (MDC) puede usarse para describir características relativas al drenaje, lo quepuede permitir el análisis de aspectos relacionados con la acumulación de escorrentía y la disponibilidad de agua(Zeverbergen y Thorne, 1987; Moore et al., 1991). El MDC indica el grado de concavidad/convexidad de la celda, de modoque un valor negativo indica convexidad y un valor positivo concavidad. Un valor igual o muy próximo a cero significaque la superficie es plana (figura 2).
La curvatura puede definirse como la tasa de cambio de la pendiente, y depende, por lo tanto, de la derivada de segundo
grado de la elevación (Felicísimo, 1994). De este modo, podemos decir que la curvatura del terreno equivale a la suma delas derivadas parciales de segundo orden respecto a los ejes de coordenadas ( x e y):
C = ∂2 z + ∂2 z∂ 2 z2 ∂ 2 y2
El MDC, por lo tanto, puede calcularse a partir de cada celda del MDE original, mediante una aproximación polinómicaa una superficie de ajuste de segundo grado:
z = a00
+ a10
· x + a01
· y + a11
· x · y + a20
· x2 + a02
· y2
donde las derivadas primeras de z respecto de x e y son:
∂ z= a10 + 2 · a20 · x + a11 · y∂ x
∂ z = a01 + 2 · a02 · y + a11 · x∂ y
Y, por lo tanto:
∂2 z = 2 · a20∂ 2 x
∂ 2 z = 2 · a02∂ 2 y
CLASE RANGO DE PENDIENTE INCLINACIÓN DE LAS LADERAS1 0-3% Muy suave2 3-8% Suave3 8-21% Ondulado4 21-41% Escarpado
5 41-50% Muy escarpadoTabla 1. Clases de pendiente.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 250/276
Almoraima, 35, 2007
250
CLASE RANGO DE ELEVACIÓN RELIEVE LOCAL1 0-30 m Relieve plano o casi plano2 30-90 m Relieve localizado3 90-150 m Relieve moderado4 150-300 m Relieve fuerte5 300 – 900 m Relieve montañoso6 >900 m Relieve montañoso muy alto
Tabla 2. Clases de relieve local.
De esta manera, la curvatura en cada celda del MDT puede calcularse de la siguiente manera:
C = 2(a20
+ a02
)
Según la expresión, valores negativos indican concavidad (cubetas de sedimentación, valles, etc.) mientras que valorespositivos indican convexidad (macizos, domos, cumbres, partes altas de las laderas, etc.).
Relieve local del terreno
El estudio del relieve local se ha realizado directamente a partir del MDE. Para ello, se ha delimitado una ventana cuadradade 1.000 m de lado correspondiente a cada celda (es decir, una ventana que incluye a 100 celdillas). A continuación, se hadeterminado el rango de elevación en cada ventana, asignando ese valor a la celda central (figura 3). A partir de este rangode elevación se ha clasificado el relieve local, utilizando el criterio de la tabla 2.
Modelo digital de geoformas
Las formas del terreno son el resultado de los procesos de modelado que actúan sobre la superficie. El modelado es, porlo tanto, función de los distintos agentes geomorfológicos que tienen lugar en la génesis del paisaje. El clima, la geología,la vegetación o el suelo son algunos de estos factores. Teniendo esto en cuenta, Hammond (1964) llevó a cabo unaclasificación general de las formas del terreno en Estados Unidos, diferenciando cinco tipos principales de relieve: llanuras,mesetas, llanuras con colinas o montañas, colinas/montañas abiertas y colinas/montañas. A su vez, cada uno de estos tiposprincipales fue dividido en clases y subclases en función de la pendiente, el relieve local y el tipo de perfil (curvatura/ convexidad de la superficie).
A su vez, y basándose en los trabajos ya citados de Hammond, Dikau et al. (1991) realizaron la clasificación automáticadel terreno en áreas de Estados Unidos. Iriagaray et al. (1997) y Martínez-Zavala et al. (2005) llevaron a cabo estudiossimilares. La misma metodología ha sido aplicada en el presente trabajo, aunque con algunas modificaciones.
Figura 3. Determinación del relieve local en la ventana de 10.000 m2.Figura 2. Concavidad/convexidad del terreno.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 251/276
Comunicaciones
251
La clasificación automática de formas principales del terreno se ha llevado a cabo a partir de la pendiente, el relieve localy la posición relativa. Para producir resultados homogéneos y comparables a los obtenidos por Hammond y Dikau, lapendiente se ha considerado “suave” por debajo del 4% (tabla 2). En la metodología utilizada por Dikau et al. (1991) elfactor pendiente se describía según el porcentaje de la superficie con pendiente suave, y se establecía a partir de una ventanade 1.000 m de lado, generalizándose a cada celdilla de 40.000 m2. En este trabajo se ha utilizado como unidad básica unaceldilla de 10.000 m2, lo que permite obtener resultados con una precisión más elevada.
Para ello, a cada celdilla del MDT se le ha asignado un código compuesto por tres dígitos, correspondientes a cada uno de lostres factores mencionados, tal y como se muestra en la tabla 3.
RESULTADOS Y DISCUSIÓN
El modelo morfológico del terreno permitió diferenciar cuatro tipos principales de formas: “colinas y montañas”,“mesetas”, “llanuras”, y “valles” (figura 4).
Colinas y montañas
Las laderas corresponden a zonas de fuerte pendiente, de forma generalmente cóncava y relieve de moderado a fuerte. Laactuación de los procesos erosivos es intensa, aunque la génesis de la forma esta condicionada principalmente por lapresencia de estratos rocosos resistentes. La morfología de las laderas se halla caracterizada por una marcada red de drenajeencajada entre los estratos de arenisca. Las principales laderas montañosas con alto relieve se distribuyen sobre las sierrasde Ojén, el Niño y de Enmedio.
FORMAS DEL TERRENO CLASES CÓDIGO (ABC)Valles Valles 111 112Llanuras Llanuras suaves con relieve localizado 121 122
Llanuras irregulares de bajo relieve 211 212Llanuras irregulares de relieve moderado 221 222
Mesetas Mesetas con relieve moderado 131 231Mesetas con relieve considerable 141 241Mesetas con alto relieve 151 251
Llanuras con colinas o montañas Llanuras con colinas 132 232Llanuras con colinas altas 142 242Llanuras con montañas bajas 152 252
Colinas y montañas abiertas Colinas abiertas bajas 321 322Colinas abiertas moderadas 331 332
Colinas abiertas altas 341 342Montañas abiertas bajas 351 352Colinas y montañas Colinas muy bajas 412
Colinas bajas 422Colinas moderadas 432Colinas altas 441 442Montañas bajas 451 452
Tabla 3. Clasificación de las formas del terreno. A: pendiente; B: relieve local; C: curvatura.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 252/276
Almoraima, 35, 2007
252
En algunos casos en que el sustrato está formado por rocascarbonatadas, como la sierra de las Cabras, la forma delterreno también está condicionada por los procesos de diso-lución kárstica.
La zona media de las laderas de las sierras, donde la pendientees más suave y el relieve local es moderado o alto, se haagrupado en una clase de colinas y montañas abiertas. Estoincluye, además, algunas formaciones de lomas. Las lomasconstituyen zonas de relieve más o menos accidentado, comoresultado de la disección del terreno por una serie de talwegs.
Se distinguen de otro tipo de formas de relieve acusado por suproyección vertical alargada y por la presencia de dos vertien-tes principales con una dirección predominante.
En el parque, las lomas corresponden a las sierras del Aljibe,debido a su morfogénesis. Las sierras de arenisca estánformadas por afloramientos de estratos rocosos con una graninfluencia estructural, que originan formas alargadas y orien-tadas según la dirección de los plegamientos tectónicos. Deesta manera, en el sector central y norte las sierras siguen unadirección norte-sur (sierra del Aljibe), mientras que en el
sector sur y el Campo de Gibraltar siguen una direccióndominante noroeste-sureste (sierra de Luna, sierra de Ojén osierra de Montecoche).
Mesetas
Las mesetas constituyen porciones de terreno relativamente planas y elevadas. Normalmente están limitadas al menos en uno desus lados por un escarpe abrupto hacia zonas más bajas. Su origen se debe a elevaciones tectónicas de terreno plano y acontinuación subdivididas por la incisión de profundas gargantas o valles. La superficie topográfica recuerda una forma tabularo suavemente ondulada por la erosión (Zinck, 1988-89).
Llanuras
En este conjunto se han incluido zonas con formas de perfil también cóncavo, de pendiente algo más acusada que en el casoanterior (3-8 %) y relieve plano o localizado. En este caso aparecen ya procesos erosivos de baja intensidad.
Se trata de formas asociadas al contacto entre los valles y las bandas de piedemonte que constituyen las faldas de las sierras.
En la parte culminante de las sierras el terreno pasa a formas determinadas por la intersección de dos laderas. Cuando elperfil del terreno es redondeado o tabular se trata de una zona amplia y fácilmente delimitable. En este caso, la pendientees generalmente baja. La diferencia entre este tipo de formas y los valles resulta del relieve más acusado y, frecuentemente,de su forma algo más convexa.
Algunos ejemplos de este tipo de formas son los llanos del Juncal (en la sierra de Luna), las crestas de la sierra de la Palma,de Ojén o de Montecoche.
Figura 4. Mapa de formas del terreno. Map of landforms.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 253/276
Comunicaciones
253
Valles
Los valles constituyen formas suavizadas y cóncavas con relieve poco acusado y pendiente muy baja (0-3%) donde losprocesos de sedimentación predominan frente a los procesos erosivos. En este conjunto se incluyen zonas de vega, fondosde valles, depresiones, llanuras aluviales y cubetas de sedimentación. Constituyen áreas más o menos extensas deacumulación de sedimentos procedentes de las aportaciones fluviales y la escorrentía de las sierras vecinas. Las unidadesde mayor extensión de este tipo son las constituidas por la cuenca baja del Barbate y los terrenos asociados a la cubeta desedimentación de La Janda al oeste del parque, así como el área de Castellar de la Frontera, donde se encuentra el embalsedel Guadarranque o la cuenca del embalse de Charco Redondo en el límite oriental.
CONCLUSIONESMediante un método jerárquico de clasificación se ha dividido el Parque Natural Los Alcornocales en cuatro clases deformas geomorfológicas principales. La superficie incluida en cada clase está caracterizada por una serie de característicasgeomorfológicas que la distinguen de las áreas vecinas.
Los resultados muestran que el método utilizado permite identificar con fiabilidad las principales formaciones del terreno,y con mucho mayor detalle, un número elevado de subclases. Como objetivo de futuras investigaciones, el uso combinadode imágenes de satélite y de cartografía geológica preexistente a una escala adecuada, puede permitir una mayor precisiónen la identificación de las geoformas, en el levantamiento de nueva información, en el análisis físico de cuencashidrográficas, o en el caso de la realización de un mapa de suelos, de riesgos geológicos o de erosión.
La clasificación de las formas del terreno propuesta en el presente trabajo refleja las principales diferencias físicas yestructurales del área estudiada. Sin embargo, los resultados obtenidos pueden ser mejorados. El uso de un MDE másdetallado podría permitir la identificación de microformas que no pueden percibirse con la resolución empleada, comoformas fluviales que aparecen como superficies planas.
REFERENCIASBELLINFANTE, N., Jordán, A., Anaya, M., y Martínez-Zavala, L. Propuesta metodológica para la realización de un modelo de distribución potencial de usos
forestales basado en parámetros geomorfoedáficos, climáticos y topográficos (Memoria del proyecto, no publicada), Sevilla, Universidad de Sevilla – Consejeríade Medio Ambiente (Junta de Andalucía), 2002.BURROUGH, P. A. Principles of Geographical Information Systems for land resources assesment , Nueva York, Oxford University Press, 1986.DIKAU, R., & Schmidt, J. “Georeliefklassifikation”; En: Schneider-Sliwa, R., Schaub, D., y Gerold, G.: Angewandte landschaftsökologie , Stuttgart (Alemania),Springer, 1999, pp. 217-244.DIKAU, R., Brabb, E. E., y Mark, R. “Landform classification of New Mexico by computer”, US Geological Survey, Open File Report 91/634, 1991.DOYLE, F. J. “Digital terrain models: an overview”, Photogrammetric Engineering and Remote Sensing, 44/12 (1978), pp. 1481-1485.FELICÍSIMO, A. M. Modelos digitales del terreno. Introducción y aplicaciones a las ciencias ambientales, Oviedo, Ediciones PENTALFA, 1994.GERRARD, A. J. Soils and landforms. An integration of Geomorphology and Pedology, Londres, George Allen & Unwin, 1981.HAMMOND, E. H. Classes of landsurface form in the forty eight states USA, Washington, Annual Association of American Geographers v54, 1964.IRIAGARAY, C., Fernández, T., y Chacón, J. “Aplicación de un sistema de información geográfica al análisis del medio físico en el sector de Rute (Córdoba)”,Cuaternario y Geomorfología, 11/1-2 (1997), pp. 99-112.JENNY, H. Factors of soil formation, New York, McGraw Hill, 1941.JORDÁN, A., y Bellinfante, N. “Cartografía de la erosividad de la lluvia estimada a partir de datos pluviométricos mensuales en el Campo de Gibraltar (Cádiz)”,Edafología, 7-3 (2000), pp. 83-92.JORDÁN, A. El medio físico del Campo de Gibraltar: unidades geomorfoedáficas y riesgo de erosión (Tesis Doctoral), Sevilla, Universidad de Sevilla, 2000.LARK, R. M. “Soil-landform relationships at within-field scales: an investigation using continuous classification”, Geoderma, 92 (1999), pp. 141-165.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 254/276
Almoraima, 35, 2007
254
MARTÍNEZ-ZAVALA, L., Jordán, A., Anaya, M., Gómez, I.A., y Bellinfante, N. “Clasificación Automática de Elementos Geomorfológicos en la Cuenca del RíoTepalcatepec (México) a Partir de un Modelo Digital de Elevaciones. Clasificación Automática de Formas del Terreno”, Cuaternario y Geomorfología. Vol. 19. Núm.3-4 (2005), pp. 49-61.MARTÍNEZ-ZAVALA, L. Análisis territorial de la comarca del Andévalo occidental: una aproximación desde el medio físico (Tesis Doctoral). Sevilla, Universidadde Sevilla, 2001.MOORE, I. D., Grayson, R. B., y Landson, A. R. “Digital terrain modelling: a review of hydrological, geomorphological and biological applications”, Hydrologicalprocesses, 5 (1991), pp. 3-30.RUIZ, L. A., Pardo, J. E., Porres de la Haza, M. J., Fernández Sarra, A. “Relaciones entre las características de modelos digitales de elevación y texturas aplicadas al análisisde la vegetación”. En: Casta, O., Quintanilla, A. (eds.): Teledetección. Avances y aplicaciones, Albacete, Asociación Española de Teledetección, 1999, pp. 76-79.ZEVERBERGEN, L. W., y Thorne, C. R. “Quantitative analysis of land surface topography”, Earth surface processes and landforms, 12 (1987), pp. 47-56.ZINCK, J. A. Physiography and soils – Soil Survey courses, Enschede (Holanda), ITC, 1988-89.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 255/276
Comunicaciones
255
RESUMEN
Limonium emarginatum es un endemismo del Estrecho cuya distribución está limitada a once poblaciones. La separaciónespacial existente entre las poblaciones podría llevarlas a un aislamiento reproductivo y a una deriva genética que puedehacer peligrar su supervivencia. Esta situación hace necesario el desarrollo de estrategias que permitan su recuperación enalgunos enclaves. En este sentido este trabajo realiza un estudio de los factores bióticos y abióticos en diferentes zonas delEstrecho con el fin de caracterizar el hábitat idóneo para realizar trasplantes de Limonium emarginatum.
INTRODUCCIÓN
Limonium emarginatum Kuntz. Wild. Es una especie catalogada como “vulnerable” en el Libro Rojo de la FloraAmenazada de Andalucía. Las poblaciones de este endemismo del Campo de Gibraltar, Ceuta y Marruecos se localizanen once zonas diferentes a ambas orillas del Estrecho. Su distribución es dispar, diferenciándose poblaciones biendesarrolladas en el medio con otras de una extensión muy reducida en el espacio.
La heterogeneidad de su distribución y la separación espacial entre las poblaciones podría llevarlas a un aislamientoreproductivo y a una deriva genética que fuera incapaz de responder ante las nuevas situaciones que pudieran plantearsedebido a nuevos factores como los determinados por el cambio climático (Figueroa et al., 2003).
Una de las medidas correctoras del aislamiento espacial y genético de las poblaciones consistiría en el desarrollo de nuevaspoblaciones que sirvan de puente entre las demás.
CREACIÓN DE UNA NUEVA POBLACIÓNDE LA ESPECIE PROTEGIDA LIMONIUM EMARGINATUM ENEL LITORAL DELESTRECHO DE GIBRALTAR
Orlando Garzón / Jesús M. Castillo / Susana Redondo / M. Enrique FigueroaDepartamento de Biología Vegetal y Ecología. Facultad de Biología. Universidad de Sevilla

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 256/276
Almoraima, 35, 2007
256
El objetivo de este estudio es analizar las diferentes condiciones tanto abióticas como bióticas con el fin de determinar lazonas de trasplante idóneas para el establecimiento de una nueva población de L. emarginatum.
La creación de nuevas poblaciones de L. emarginatum en zonas desocupadas situadas entre poblaciones silvestrespreexistentes podría facilitar el intercambio genético entre poblaciones y podría contribuir a la conservación de esteendemismo del estrecho de Gibraltar (Asensi, 1984; Sánchez García, 2000).
Por tanto, resulta de interés desarrollar una metodología capaz de generar nuevas poblaciones mediante trasplantes en zonascosteras favorables para L. emarginatum.
MATERIAL Y MÉTODOS
Se pusieron a germinar semillas de L. emarginatum recolectadas en septiembre de 2004 en Punta Carnero (Algeciras). Lassemillas se colocaron sobre papel de filtro, en placas de petri de 5 cm de diámetro, cuatro réplicas, de 25 semillas cada unay se regaron con agua destilada. Las placas se introdujeron en un germinador (ASL Aparatos Científicos M-92004, Madrid)donde se mantuvieron con un régimen de 10 horas de luz (25º C, 400-700 nm, 35 mol m-2 s-1) y 14 horas de oscuridad (5ºC) durante 30 días (Keiffer y Ungar, 1997). Se eligió este régimen de temperaturas para simular las temperaturas que seregistran durante el otoño en el estrecho de Gibraltar.
Las semillas germinadas se fueron transfiriendo a macetas. Finalmente se obtuvieron 20 ejemplares que crecieron en perlitadurante cinco meses en un invernadero con temperatura controlada, entre 21-25ºC expuestas a la radiación solar. Conposterioridad se pasaron a macetas con sustrato extraído de la zona del trasplante.
Con objeto de seleccionar la zona de trasplante más idónea se estudiaron las características abióticas del medio, siendo lasvariables estudiadas: temperatura, dirección y frecuencia de viento, humedad relativa y precipitaciones. Estos datos secomplementaron con los aportados por la estación de Tarifa, del Instituto Nacional de Meteorología durante los años 2004-2005, poniendo especial énfasis en los datos obtenidos en los meses de verano donde las condiciones meteorológicas semanifiestan más estresantes para las plantas debido al déficit hídrico.
Por otro lado, se analizó la distribución geográfica de las especies y su orientación espacial. De igual modo se estudió lafrecuencia y dirección del viento en el Estrecho, y cómo éste afecta a la temperatura y humedad relativa presentes. Además,se realizaron estudios a pequeña escala, determinando si en una misma zona con diferente orientación al viento, existencambios de temperatura apreciables tanto en cala Parra (Algeciras) como en la Isla de las Palomas (Tarifa).
Otro factor que se tuvo en cuenta fue la influencia de la geología sobre el desarrollo de las poblaciones y si la geomorfología
afecta al desarrollo de las plantas. En el primer punto, se han tratado todas las poblaciones y en el segundo se ha analizadoel contenido hídrico relativo sobre distintos núcleos de la población en la isla de las Palomas, diferenciándose núcleos deroca y de arena. Para ello se han tomado cinco muestras por cada núcleo, con porciones del mesófilo de las hojas. Lasmediciones de peso seco se realizaban in situ. A continuación se introducían en botes herméticos de agua destilada parasu hidratación, con un mínimo de 24 horas, en cámara frigorífica. De nuevo se pesaban las muestras, ahora hidratadas. Unavez concluida esta operación se secaban en una mufa durante 48 horas hasta la total deshidratación, para pesar la materiaorgánica. Con estos datos podemos calcular el estado de hidratación de las hojas, y por tanto, obtener una aproximaciónsobre el estrés hídrico que puede sufrir la planta.
Se realizó un inventario de las plantas presentes en distintas zonas con el fin de conocer su estado de transformación.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 257/276
Comunicaciones
257
Figura 1. Climograma de Tarifa. Datos correspondientes al periodo de tiempo1971-1999 recogidos de la estación meteorológica de Tarifa (Cádiz). Altitud:41m. Latitud: 36º 00´ 53´´ N. Longitud 005º 35´ 52´´ W.
Figura 2. Datos suministrados por el INM estación de Tarifa, Cádiz. Altitud 41m.Latitud: 36°00’53'’N. Longitud: 05°35’52'’W. Período 1971-1999.
RESULTADOS
Naturaleza geológica
En el estudio ecológico que realizamos sobre L. emarginatum, se apreciaba una indiferencia edáfica para este endemismo.Se le podía encontrar sobre sustrato ácido (Unidad flysch de Algeciras) como básico (calcarenitas biogénicas del Cretácicoen Tarifa. Gutiérrez Mas, 1991). En cuanto que un tipo de sustrato permita un mejor desarrollo de la población, se haobservado que en la costa europea, la mayor distribución se da en las poblaciones de Gibraltar, Isla de las Palomas y PuntaCamarinal, de sustrato básico, mientras que en la costa africana, encontramos una distribución amplia en Ceuta con sustratopredominante ácido.
Condiciones ambientales (temperatura, humedad relativa, dirección del viento)
Se han tomado los datos correspondientes a la estación del INM en Tarifa. Esta estación se encuentra en el punto másseptentrional de la costa Europea y sus valores pueden ser tomados como representativos para todo el arco del estrecho deGibraltar. En el gráfico siguiente aparece el climograma de Tarifa.
Valores promedio climáticos para Tarifa:
Temperatura media 17,1ºCPrecipitación anual 603mmHumedad relativa media 79%Número medio mensual de horas de sol 2538h.
Tabla 1. Datos climáticos correspondientes al período 1971-1999 recogidos de la estación meteorológica de Tarifa (Cádiz).Altitud: 41m. Latitud: 36º 00´ 53´´ N. Longitud 005º 35´ 52´´ W.
La rosa de los vientos anual de Tarifa nos pone de manifiesto la polaridad de la dirección del viento, en sus componenteseste y oeste. Los vientos de componente este alcanzan mayores velocidades en la zona del estrecho.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 258/276
Almoraima, 35, 2007
258
En cuanto a la frecuencia, los vientos de componente este sonlos más abundantes con una frecuencia promedio en torno al43,28%. Los vientos de componente oeste tienen una frecuen-cia del 26,13%, los de componente suroeste con un 5,84% ylos noroeste con una frecuencia del 8,72% (figura 3).
Orientación geográfica de las poblaciones
La orientación de las plantas en cada población sigue unapauta constante en Gibraltar (este y sureste), Punta Carnero-cala Parra (este), punta chorlito (este y sureste), Guadalmesí (este), Isla de las Palomas (en todas direcciones con predomi-nio en dirección este o sureste), Calamocarro, Benzú y Cho-rrillo (este) y Castillejo (este). En Punta Camarinal y PuntaPaloma el predominio se da en dirección oeste.
Por tanto, en la zona del Estrecho, se aprecia una tendencia auna mayor distribución de las plantas de L. emarginatum conorientación hacia el este.
Importancia de los microhábitats
Con un mayor detenimiento hemos estudiado las condicionesabióticas de la Isla de las Palomas (Tarifa), al ser la población
que se encuentra más desarrollada. En la Isla de las Palomashemos encontrado una asimetría en la distribución de L.emarginatum, observándose una mayor abundancia en el esteque en el oeste de la isla. La presencia de los vientos puedetener un papel destacado en esta situación (Garzón et al.,2003) y hemos realizado una serie de mediciones de tempe-ratura continuas a lo largo del mes de agosto de 2005, que sepresentan en la figura 4.
Se aprecian unas oscilaciones de la temperatura de modosignificativo en las máximas, entre las situadas al este y las
situadas al oeste. Las diferencias térmicas a la misma hora yentre los dos sensores han llegado a marcar 6,8ºC de diferen-cia entre las temperaturas máxima de las zonas dispuestas aleste y al oeste de la Isla de las Palomas.
Las fluctuaciones térmicas en las zonas de levante y ponientecon respecto al tipo de viento vienen reflejadas en la figura 5.
El análisis de cluster nos determina dos agrupaciones dedatos. Se corresponden los días en los que el viento dominan-te. El grupo de la izquierda se correspondería con viento deloeste y el de la derecha con viento del este.
Figura 3. Distribución de los valores promedio de viento a lo largo del año enTarifa. Datos elaborados a partir de los suministrados por el INM estación deTarifa, Cádiz. Altitud 41m. Latitud: 36°00’53'’N. Longitud: 05°35’52'’W.Período 1971-1999.
Figura 4. Distribución de temperaturas máximas y mínimas en las zonas delevante y poniente de la Isla de las Palomas, a lo largo del mes de agosto de 2005.Mediciones realizadas con sensores Tinytag Ultra H°C/%RH, en intervalos deuna hora.
Figura 5. Análisis de cluster. En el eje de ordenadas el grado de semejanza de losvalores de la matriz (temperatura máxima y mínima en la zona este y oeste de laIsla de las Palomas). En el eje de abscisas los días del mes de agosto de 2005.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 259/276
Comunicaciones
259
En la población de la Isla de las Palomas hemos dispuesto parael seguimiento de la población de siete núcleos que difierenen cuanto al sustrato y su orientación. Los núcleos n1 y n2están dispuestos al este de la isla. Los núcleos n3, n4 y n5 estándispuestos al sur y oeste de la isla. Dentro de los núcleos n1y n2, se han dispuesto dos zonas, que se corresponden consustrato suelto (arena) o no (roca).
La humedad relativa medida en el verano de 2004 nos muestralas siguientes curvas en función de la prevalencia del viento:
El viento de levante aporta una mayor humedad en la zona delevante, y de igual manera se puede decir del viento deponiente en su zona. Comparando ambos vientos, el deponiente aporta mayor humedad en esta época del año a lazona de levante de la isla que viceversa. Las plantas que seencuentran en el este de la isla van a recibir los efectosbenéficos de la humedad del viento tanto de levante como deponiente, mientras que las plantas situadas al oeste de la isla,tendrán un aporte de humedad por el viento de poniente, y unmenor aporte del viento de levante. Estas observaciones secorresponden con la distribución presente en la isla de las
Palomas de L. emarginatum, con una mayor distribución enel este de la isla (Garzón et al., 2004).
De la observación de las poblaciones se obtiene que suelencolonizar aquellas zonas que se encuentran expuestas alviento, especialmente de levante. En cala Parra, cercana aPunta Carnero, se encuentra una pequeña población situadaen el cabo de la Cala. Se realizó la medición de la temperaturaen el cabo comentado y en la zona más convexa de la cala, auna altura de unos 4 m sobre el nivel del mar. Los resultadosobtenidos se muestran en la figura 7, donde las temperaturasmáximas difieren en las dos zonas. En la zona expuesta a laacción del viento la temperatura máxima se mantiene cons-tante en torno a los 18ºC, mientras que en el interior de la calase llegan a alcanzar valores de 35ºC.
Estado de agregación del sustrato
Dentro de la Isla de las Palomas, L. emarginatum se desarrollaen sustrato disgregado, arena (a) y en sustrato duro, roca (r).En la zona este de la isla, hemos señalado cuatro núcleos,denominados n1r, n1a, n2r y n2a. Los denominados n1r y n2rse caracterizan porque el sustrato rocoso está horadado por lo
Figura 6. Datos obtenidos de las mediciones realizadas durante nueve díasentre junio y septiembre de 2004, con alternancia de vientos de levante yponiente en la Isla de las Palomas en Tarifa.
Figura 7. Valores de temperatura máxima y mínima en Cala Parra. Los valoresde temperatura han sido tomados entre el 16 y el 22 de enero de 2005, en dos zonasde cala Parra correspondientes con la zona convexa y el cabo más al sur. Datostomados con sensores Tinytag Ultra H°C/%RH, en intervalos de una hora.
Figura 8. Contenido hídrico relativo de los siete núcleos de la población de laIsla de las Palomas. Datos tomados el 24/9/04.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 260/276
Almoraima, 35, 2007
260
que sería el embrión de pequeñas marmitas, que presentan elaspecto de maceteros donde se localizan las plantas. Elsustrato dentro de estas oquedades estaría constituido pormateriales disgregados. N1a y n2a serían zonas de sustratoarenoso. La determinación del contenido hídrico relativopuede darnos una idea del estado fisiológico de las plantas. Aun menor contenido hídrico, mayor estrés hídrico padecería laplanta. La comparación de datos de plantas cercanasespacialmente, pero en diferentes medios, puede aportarinformación relevante para el presente trabajo.
Las mediciones se realizaron en septiembre, por ser el mesúltimo del verano, y donde se podrán manifestar en mayormedida los efectos del estrés hídrico.
De los núcleos tratados conviene comparar los de las pobla-ciones de levante n1 y n2, tanto en arena como en roca. Seobserva que las diferencias en el contenido hídrico sonsignificativas entre los núcleos n1r y n1a. Entre los núcleosn2r y n2 la ligera diferencia observada no es significativa. Losdatos sugieren que el sustrato de roca va a permitir una mayorasimilación de agua por parte de las plantas.
Zona elegida
Atendiendo a la distribución de las poblaciones de L.emarginatum, se observa una discontinuidad entre las mis-mas. Esta dispersión puede tener un efecto negativo en cuantoal aislamiento genético se refiere. Se establecen dos zonas enla costa europea que pueden ser susceptibles de estudio decara a una reintroducción, la correspondiente a la costaalgecireña, comprendida entre Punta Carnero y Guadalmesí,y la que abarca la costa de Tarifa, entre la Isla de las Palomasy Punta Paloma- Punta Camarinal. Dado que entre Isla de las
Palomas y Punta Camarinal, dos de las poblaciones conmayor número de individuos, sólo existe un núcleo muyreducido en Punta Paloma hemos considerado de interésiniciar en ese tramo el trasplante de nuevos individuos, demodo que pudiera servir de punto de conexión entre las dosgrandes poblaciones citadas.
Hemos comparado las temperaturas máxima y mínima en lazona elegida con las temperaturas máxima y mínima de la Islade las Palomas, tanto en su vertiente este como oeste. Eltiempo de medición ha sido de 28 días en el mes de agosto de
Figura 9. Promedio de temperaturas máximas y mínimas en la Isla de lasPalomas y Punta Paloma (n=34). Cada núcleo se nombre con la zona demedición seguida por un guión con la dirección del viento. Los días compren-didos entre el 4 de agosto y el 18 de septiembre de 2005. Datos obtenidos consensores, en intervalos de una hora.
Figura 10. Fragmento rocoso de la Unidad de Bolonia que se introduce en elmar, expuesto a la acción del viento del este (levante) y del oeste (poniente).
Figura 11. Imagen de detalle de la zona elegida donde se aprecia la unidadflysch de Bolonia con alternancia de estratos de diferente consistencia.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 261/276
Comunicaciones
261
2005 para la Isla de las Palomas y de 15 días para la zona de Punta Paloma, diferenciando los datos obtenidos en todos loscasos según el viento predominante del este o del oeste.
En la posible nueva población de Punta Paloma, las temperaturas oscilan entre 24,6ºC y 21,4ºC con vientos del este, y29,6ºC y 21,3ºC con viento del oeste. En la Isla de las Palomas, zona este, las temperaturas oscilan entre 24,5ºC y 20,7ºCcon vientos de levante, y 28,2ºC y 21,2ºC con vientos de poniente. En la zona oeste las temperaturas oscilan entre 31,1ºCy 20,3ºC con vientos de levante y 26,9ºC y 20,4ºC con vientos del oeste (figura 9).
El análisis de los resultados obtenidos nos permite apreciar una mayor semejanza de los datos de la zona de Punta Palomay la vertiente este de la Isla de las Palomas en Tarifa.
Especies presentes en el promontorio
Se ha observado la presencia de Chritmum maritimum, Plantago coronopus, Sporobolus pungens, Lotus creticus, Sonchustenerrimus, Lobularia marítima, Plantago lagopus, Helichrysum picardii (Valdés B., 1987)
Como aspectos más destacables, citar la presencia en zonas cercanas de Juniperus phoenica subsp. turbinata. Y de un modomás distante la presencia de una mancha de Carpobrotus edulis, especie invasora sudafricana.
En las siguientes imágenes se observa la zona elegida y el promontorio que sería objeto del trasplante de nuevos individuos(figura 10).
Presencia de diaclasas y deslizamientos en la vertiente este del promontorio (figura 11).
En las imágenes inferiores se puede observar la disposición aislada del promontorio y su condición de cercanía al mar,expuesto a la acción de los vientos de componente este y oeste mayoritarios en la zona (figura 12).
Figuras 12. (A) Imágenes hacia el este, (B) sur y (C) oeste desde el promontorio elegido.
A B C

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 262/276
Almoraima, 35, 2007
262
DISCUSIÓN
La dispersión de las poblaciones de L. emarginatum puede llevar este endemismo a un aislamiento genético que limite sucapacidad de respuesta ante cambios en el medio y con ello la persistencia de sus poblaciones. La variación de las condicionesambientales como consecuencia del cambio climático y la acción antrópica en zonas que inicialmente poseían un grandesarrollo del endemismo, hace necesario el desarrollo de una metodología adecuada que permita la reintroducción de nuevasplantas, bien en zonas nuevas bien en poblaciones en mal estado, en aras a la biología de la conservación de la especie.
El factor geológico no parece incidir en el desarrollo del endemismo, que presenta una gran capacidad de adaptación adistintos sustratos. La zona elegida responde a unas características geológicas similares a las de Punta Paloma o PuntaCamarinal, donde se localizan las poblaciones de L. emarginatum de mayor desarrollo, y por tanto, no debiera sufrir ningúncondicionamiento por esta causa.
Respecto al régimen climático, se consideran estables las condiciones para buena parte del Estrecho, en especial en las zonasmás septentrionales, que abarcan la Isla de las Palomas y Punta Paloma (Tarifa). Dada la incidencia de los vientos en elEstrecho, y el carácter dominante de los vientos de componente este y oeste en la zona, las fluctuaciones de temperatura segúnel viento dominante parecían de interés. Estas fluctuaciones han determinado que los vientos de componente este (levante),más abundantes en la zona, determinarían un mejor desarrollo de plantas expuestas al este (frecuencia anual de los vientos,E:43,28%, W:26,3%). Como hemos observado en las mediciones realizadas en cala Parra (Algeciras), la exposición directaa la acción del viento es benigna como elemento atenuante de las elevadas temperaturas estivales.
Estas diferencias térmicas tienen su correspondencia con la humedad relativa resultante, mientras que los vientos decomponente oeste aportan un grado de humedad parecido para las orientaciones este y oeste de las poblaciones, los vientos
de componente este aportan una humedad elevada a las zonas de orientación este y de un claro valor inferior para laorientación oeste. Este factor, en condiciones de estrés hídrico como el correspondiente a la época estival, determinaría laorientación de los trasplantes de nuevos individuos. La mayoría de las poblaciones naturales están orientadas hacia el este.
La geomorfología del promontorio es determinante. Por los datos obtenidos en le medida del contenido hídrico relativo,las plantas que estén situadas sobre las diaclasas o en las oquedades de las marmitas, dispondrán de un mayor aporte hídrico.Por tanto este factor debe ser tenido en cuenta a la hora de efectuar los trasplantes.
Del inventario de especies presentes se deriva una dinámica similar a la encontrada en Punta Camarinal, donde las especiesmás representativas son Helichrysum picardii y Juniperus phoenica subsp. turbinata (Garzón et al., 2004). La presenciade una mancha de Carpobrotus edulis en una zona cercana al promontorio se concibe como una amenaza para la poblaciónpor lo que debería ser eliminada, en este sentido la Consejería de Medio Ambiente está realizando una campaña de
erradicación en las costas de Huelva y Cádiz.Para realizar los trasplantes, se decidió agrupar los nuevos individuos en grupos de cinco y en diferentes localizaciones,con objeto de corroborar las hipótesis de partida de que: 1) los organismos dispuestos hacia levante se desarrollan mejor,y por otro lado que 2) los individuos dispuestos a lo largo de las diaclasas o entre los estratos de arenisca tendrán un mejordesarrollo. Los trasplantes seguirían las siguientes localizaciones:
- Fracturas de las diaclasas en dirección este (cinco ejemplares).
- Al abrigo del plano de arenisca, dirección este (cinco ejemplares).
- Sobre las margas en dirección este (cinco ejemplares).
- Fracturas de las diaclasas en dirección oeste (cinco ejemplares).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 263/276
Comunicaciones
263
La posición topográfica ha sido identificada como el factor ambiental que determina principalmente la composición de lascomunidades vegetales en acantilados costeros (Kingston y Waldren, 2003). Todas las poblaciones de L. emarginatum sesituan en zonas expuestas a la acción del viento y del mar. El nivel de la pleamar, localizado en la base del promontoriodeterminaría las características ambientales descritas en otras poblaciones (Garzón et al., 2003) de elevada salinidad debidoal spray marino, elevada humedad relativa y fuerte acción del viento (Adam, 1990).
La época idónea para llevar a cabo el trasplante de los plantones de Limonium emarginatum es en época de lluvias, quepermitirán inicialmente una mayor supervivencia de los ejemplares trasplantados.
Deseamos que la colaboración de la dirección del Parque Natural del Estrecho para esta experiencia piloto, sea secundadapor la ciudadanía, consciente de que la gestión conservadora del medio es algo que nos atañe a todos.
BIBLIOGRAFÍAADAM, P. Salt marsch Ecology. Cambridge University Press. Cambridge, UK. 1990.ASENSI MARFIL, A. Limonietum emarginati (Chritmo-Limonion ) nueva asociación para los sectores gaditano y tingitano. Documents Phytosociologiques. Vol.VIII.5: pp. 455-465. 1984.CHAPMAN, V.J. Coastal Vegetation. Segunda edición. Pergamon Press. 1976.FIGUEROA CLEMENTE, M. E., S. Redondo , T. Luque, J. M. Castillo, A. E. Rubio-Casal, A. A. Álvarez López, E. E. Castellanos, C. J. Luque, F. J. Jimenez Nieva,O. Garzón Gómez. Diversidad genética y diversidad ecológica. MA medioambiente 2003/42 . Humedales. Junta de Andalucía, Consejería de Medio Ambiente. 2003.GARZÓN, O., J. M. Castillo, M. E. Figueroa. "Descripción ecológica de las poblaciones de Limonium emarginatum en el Estrecho de Gibraltar". VI Jornadas de Flora,Fauna y Ecología del Campo de Gibraltar. Almoraima 31. 2003.GARZÓN, O., J. M. Castillo, M. E. Figueroa. "Aproximación ecológica al estado de conservación de las poblaciones de Limonium emarginatum , un endemismo delEstrecho de Gibraltar". Sociedad Gaditana de Historia Natural. Volumen IV. 2004.GUTIÉRREZ MAS, J. M. et al. Introducción a la geología de la provincia de Cádiz. Servicio de publicaciones de la Universidad de Cádiz. 1991.KEIFFER, C. H., I. A. Ungar. "The effect of extended exposure to hypersaline conditions on the germination of five inland halophyte species". American Journal
of Botany 84, pp. 104-111. 1997.KINGSTON, N. y S. Waldren. "The plant communities and environmental gradients of Pitcairn Island: The significance of invasive species and the need forconservation management". ANNALS OF BOTANY 92 (1): pp. 31-40. 2003.SÁNCHEZ GARCÍA, I. Flora amenazada del litoral gaditano. Junta de Andalucía, Consejería de Medio Ambiente. pp. 120-121. 2000.VALDÉS, B. et al. Flora Vascular de Andalucía Occidental. Vols. 1 a 3. Ketres editora S.A. 1987.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 264/276

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 265/276
Comunicaciones
265
EL CABLE
En octubre de 1997 se instaló la primera interconexión eléctrica entre España y Marruecos a través de las aguas del Estrechode Gibraltar. Con una capacidad de 700 MW esta instalación constituía el primer eslabón del anillo eléctrico circumediterraneo.Actualmente dicha capacidad es claramente insuficiente, saturándose la línea en las horas punta de consumo eléctrico. Elproyecto de refuerzo dotará a la interconexión de dos líneas en paralelo con un cable de reserva compartido por ambas,aumentando la capacidad hasta los 1.400 MW.
El proyecto de la segunda interconexión consiste en el tendido de tres cables con una parte subterránea y una partesubmarina de unos 29 Km aproximadamente. En tierra se ampliarán las subestaciones de Tarifa y de Fardioua en Marruecos.
TRAMITACIONES AMBIENTALES
Las consultas iniciales al estudio de impacto ambiental comenzaron en diciembre de 2001 y terminaron en abril de 2002con la emisión de un informe al Ministerio de Medio Ambiente. Posteriormente se realizó el estudio de impacto ambiental,en colaboración con la Consejería de Medio Ambiente de la Junta de Andalucía, donde se definían ya medidas preventivasy correctoras del impacto.
La obra atraviesa espacios pertenecientes a la Red NATURA 2000, como son el Paraje Natural Playa de los Lances y elParque Natural del Estrecho, y en virtud del artículo 6 de la Directiva 92/43/CEE de Hábitats, deben proponerse medidascompensatorias al impacto de las obras.
Red Eléctrica de España contactó con la Fundación MIGRES para el diseño y la posterior ejecución de las medidascompensatorias.
MEDIDAS COMPENSATORIAS
DEL PROYECTO REMO IIDE RED ELÉCTRICA DE ESPAÑA
Manuela de Lucas Castellanos / Fundación MIGRES

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 266/276
Almoraima, 35, 2007
266
MEDIDAS COMPENSATORIAS
Las medidas compensatorias, a diferencia de las medidas preventivas y correctoras del impacto, no deben ir dirigidas afrenar las consecuencias directas que la construcción de una infraestructura produce, sino a paliar el posible efecto difusoque ésta puede tener en el ecosistema. En este sentido, los proyectos recomendados en el presente documento no serestringen a compensar exclusivamente a aquellos hábitats y especies afectados por la construcción y ulterior funciona-miento de la infraestructura, si no que van dirigidos a fortalecer los elementos más sensibles del ecosistema con el objetivode reducir su fragilidad.
La filosofía que ha guiado el proceso de definición de las medidas compensatorias ha sido la de involucrar, desde el primer
momento, a la sociedad gaditana en general y a la campogibraltareña en particular. Para ello, se realizó una consulta entreorganismos de investigación, organizaciones no gubernamentales de carácter ambientalista y particulares con el objeto deconocer aquellas actuaciones de conservación o lagunas de conocimiento consideradas prioritarias. Los organismoscontactados fueron:
- Verdemar (Ecologistas en Acción).
- Instituto de Estudios Campogibraltareños.
- Agaden Cádiz.
- Grupo Ornitológico del Estrecho (GOES).
- Colectivo Ornitológico Cigüeña Negra (COCN).
- Circé.
- Sociedad Gaditana de Historia Natural.
- Gibraltar Ornithological and Natural History Society (GONHS).
- Asociación Palestra.
- ORNITOUR.
- Estación Biológica de Doñana.
- Consejería de Medio Ambiente.
- Diputación Provincial de Cádiz.
- Universidad de Cádiz.
- Asociación Eólica de Tarifa.- Sociedad Española de Ornitología (SEO).
- EGMASA.
- Asociación Grandes Industrias del Campo de Gibraltar (AGY).
- Subdelegación del Gobierno del Campo de Gibraltar.
- Universidad de Sevilla.
- Asociación Naturaleza y Turismo Rural en el Estrecho.
- Ecologistas en Acción.
- Ayuntamiento de Tarifa.
- Ayuntamiento de Algeciras.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 267/276
Comunicaciones
267
- Cofradía de Pescadores.
Se recibieron cerca de 80 sugerencias de actuaciones de muy diversa índole, desde recomendaciones generales hastaproyectos extremadamente concretos. A estas sugerencias se añadieron aquellos proyectos que el equipo técnico, tomandocomo base su experiencia profesional en la zona, consideró de realización prioritaria.
Se realizó una selección de los proyectos que se adaptaban mejor al concepto de medida compensatoria y que fueronconsiderados prioritarios atendiendo a las necesidades de conservación de la zona afectada. Estos proyectos se agruparonen tres grandes grupos:
- Conservación.
- Investigación.
- Difusión.
Dentro del capítulo de conservación están los siguientes proyectos:
- Actividades para evaluar la influencia de las actividades subacuáticas en los fondos marinos.
Objetivo: búsqueda y establecimiento de puntos de inmersión alternativos, marcaje de zonas susceptibles de albergararrecifes artificiales en zonas y equipamiento de nuevos puntos de inmersión para disminuir el impacto directo.
- Protección y regeneración de áreas litorales con flora amenazada.
Objetivo: eliminación de especies exóticas invasoras y creación de microreservas en zonas con poblaciones de especialinterés.
- Actuaciones para la conservación de ríos y arroyos costeros de la mitad meridional de la provincia de Cádiz.
Objetivo: caracterización de las cuencas y de las comunidades faunísticas de los ríos y arroyos estacionales entre los ríosSalado y Guadiaro, seguimiento de la población del salinete del río de La Vega, protección de pozas estivales yconstrucción de abrevaderos, y construcción de depuradoras en núcleos rurales.
- Gestión y manejo de terrenos de importancia ecológica.
Objetivo: adquisición o mantenimiento de calidad ecológica en zonas importantes.
Dentro del capítulo de investigación, cabe destacar los siguientes proyectos:
- Estudio de la biota intermareal y fanerógamas marinas del parque natural del estrecho.
Objetivo: evaluar el estado de las praderas de faneróganas marinas en el estrecho.
- Estudio ictiológico e inventario de las comunidades asociadas en la zona intermareal rocosa del Parque Natural delEstrecho.
Objetivo: caracterización espacio/temporal de poblaciones ícticas, descripción de las estructuras de las comunidades,de las interacciones entre poblaciones y determinación del estado de conservación de la zona intermareal.
- Estudio de la pesquería del voraz (Pagellus bogaraveo) del Estrecho de Gibraltar.
Objetivo: diferenciación de stocks por métodos genéticos, campañas de marcado de población adulta, y asignaciónespacial del esfuerzo pesquero.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 268/276
Almoraima, 35, 2007
268
- Estudio de la migración del atún rojo atlántico, Thunnus thynnus, en relación con la maduración sexual y la puesta enel mar Mediterráneo.
Objetivo: conocimiento espaco-temporal de la maduración y puesta a lo largo de la migración reproductiva, y valoraciónde la capacidad reproductora.
En el bloque de difusión:
- Co-financiación de documentales de alta calidad sobre el fenómeno de la migración.
Objetivo: poner en valor el fenómeno de las migraciones y difundirlo en amplios ámbitos.
- Estudio de la viabilidad del uso ecológico y cultural de la Isla de las Palomas (Tarifa).
Objetivo: conocer actuaciones futuras en la Isla mediante la utilización sostenible de sus recursos.
- Análisis socioeconómico de la industria de la observación de cetáceos en Tarifa.
Objetivo: conocer la potencialidad del sector y poder mejorar el servicio maximizando el recurso y minimizando losefectos negativos.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 269/276
Comunicaciones
269
J. M. Guerra García / A. Ruiz Tabares / J. C. García GómezLaboratorio de Biología Marina. Departamento de Fisiología y Zoología. Facultad de Biología. Universidad de Sevilla
SEGUIMIENTO DE LAS COMUNIDADESINTERMAREALES DELA ISLA DE TARIFA (CÁDIZ):EDUCACIÓN AMBIENTAL Y CONSERVACIÓN
RESUMEN
En el presente trabajo se incluyen los resultados correspondientes al estudio de las comunidades intermareales de la Islade Tarifa, desarrollado durante los años 2003, 2004 y 2005 en el contexto de las prácticas de la asignatura de BiologíaMarina de la licenciatura de Biología de la Universidad de Sevilla. Se utilizaron análisis estadísticos univariantes ymultivariantes (ordenación y clasificación) para establecer las diferencias espacio-temporales en la abundancia de lasespecies de algas y de los principales grupos de fauna asociada. Las algas dominantes fueron Corallina elongata, Fucusspiralis, Gelidium sesquipedale, Jania rubens, Laurencia pinnatifida y Ulva rigida, y los crustáceos (especialmentegammáridos, caprélidos e isópodos) dominaron sobre moluscos, poliquetos y equinodermos. No se encontraron diferenciassignificativas en las comunidades a lo largo de los tres años de estudio, y la Isla de Tarifa sigue manteniendo un estado deconservación excelente. Sería aconsejable que el seguimiento de las comunidades intermareales se llevase a cabo todos los
años como parte de los programas de vigilancia y monitorización del Parque Natural del Estrecho. En este sentido, la laborde alumnos y voluntarios resultaría fundamental en las campañas de muestreo y trabajo de laboratorio, de modo quetérminos como formación docente, educación ambiental y voluntariado discurran paralelos a los de seguimiento y controlde nuestros ecosistemas marinos.
Palabras clave: Isla de las Palomas, Tarifa, intermareal, algas, macrofauna, educación ambiental.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 270/276
Almoraima, 35, 2007
270
INTRODUCCIÓN
Recientemente, la Consejería de Medio Ambiente de la Junta de Andalucía ha financiado un proyecto para otorgar al frentelitoral comprendido entre los términos municipales de Algeciras y Tarifa la figura de protección de Parque Natural. Se tratadel Parque Natural del Estrecho, declarado Parque Natural en marzo de 2003 con la finalidad de proteger unos valores naturalesy culturales excepcionales, y también para impulsar la economía local y mejorar la calidad de vida de sus habitantes. Es unespacio marítimo-terrestre, con superficies equivalentes de mar y tierra, que se extiende a lo largo de 54 kilómetros de costa,entre el cabo de Gracia (límite occidental) y punta de San García (extremo oriental). En el ámbito marino del Parque Naturaldel Estrecho destaca la Isla de Tarifa (también denominada Isla de las Palomas), que acoge comunidades marinas muybiodiversas y estructuradas. El Plan de Ordenación de los Recursos Naturales (PORN) considera a la Isla de Tarifa como zonade Reserva (A) catalogándola de espacio marino de extraordinario interés en el que la normativa limita todos los usos y
actividades que puedan significar una alteración de las condiciones ambientales (BOJA, 2003).Guerra-García y García-Gómez (2000) hicieron una revisión de los distintos grupos animales marinos presentes en la Isla deTarifa, recopilando la bibliografía existente y Guerra-García et al. (2000, 2001a) llevaron a cabo un estudio en la isla paracaracterizar la comunidades de algas de la zona intermareal. La isla disfruta de una situación biogeográfica privilegiada yaque se sitúa en la región del estrecho de Gibraltar, lugar de confluencia de aguas mediterráneas y atlánticas, lo que contribuyea incrementar la biodiversidad marina, que se mantiene en un estado de excelente conservación gracias a las restricciones deacceso impuestas por el destacamento militar de artillería de costa que ha tenido su base en la isla durante los últimos años.
Además de ser un excelente escenario para estudios marinos debido al buen estado de conservación y riqueza biológica,la Isla de las Palomas es un lugar idóneo para llevar a cabo labores docentes y de educación ambiental, sobre todo en lazona intermareal. Al respecto, el laboratorio de Biología Marina de la Universidad de Sevilla, viene impartiendo en la isla
desde hace varios años las prácticas de la asignatura “Biología Marina” de quinto curso de la licenciatura de Biología.Durante la práctica se muestra a los alumnos como se distribuyen las especies en la zona intermareal, y se realizan muestreosen los distintos cinturones para estudiar en el laboratorio las algas y la fauna epífita (crustáceos, moluscos, poliquetos,equinodermos que viven asociados a las algas). Con esta práctica se pretende, por una parte, que los alumnos aprendana reconocer y a identificar las especies de la zona intermareal, y por otra, que participen en un programa de seguimientoy monitorización destinado a controlar las comunidades intermareales de la Isla de Tarifa a lo largo del tiempo. Estosestudios son fundamentales como herramientas para la gestión y conservación de la isla, ya que permiten generar seriestemporales de datos que pueden poner de manifiesto la regresión de especies, bien por causas naturales o antropogénicas.En este sentido, el laboratorio de Biología Marina desarrolla actualmente en colaboración con la Consejería de MedioAmbiente de la Junta de Andalucía proyectos de vigilancia ambiental destinados a controlar el estado de las comunidadesde sustrato blando y sustrato rocoso del litoral andaluz (Guerra-García et al., 2001b).
En el presente trabajo se exponen los resultados correspondientes al estudio de las comunidades intermareales de la Islade las Palomas desarrollado durante los años 2003, 2004 y 2005 en el contexto de las prácticas de la asignatura de BiologíaMarina de la licenciatura de Biología de la Universidad de Sevilla. Se aportan datos cualitativos y cuantitativos deabundancia de las principales especies de algas y de los grupos de macrofauna dominante.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 271/276
Comunicaciones
271
Figura 1. Mapa del Parque Natural del Estrecho mostrando los ámbitos marino (blanco) y terrestre (negro).
MATERIAL Y MÉTODOS
Área de estudioLa Isla de Tarifa está situada en el estrecho de Gibraltar, en el término municipal de Tarifa (figura 1), constituyendo el puntomás meridional de Europa. Tiene una superficie de 21 Ha y presenta 2 Km de costa. El sustrato está constituido por margasy areniscas de flysh. Su localización con influencia mediterránea y atlántica y las restricciones de acceso, contribuyen aincrementar la biodiversidad marina en la isla (Guerra-García y García-Gómez, 2000), haciendo que ésta sea una de laszonas más interesantes y con mayor grado de protección del Parque del Estrecho (BOJA, 1993).
Muestreo y procesado de las muestrasEl intermareal de la Isla de Tarifa seleccionado para el estudio está localizado en la zona de poniente. Se escogió esta zona
intermareal por ser la más diversa, como han mostrado estudios previos (Guerra-García et al., 2000). Tras una exploracióndetallada del intermareal, se delimitaron visualmente los distintos cinturones de algas. Aunque en la zona podíandiferenciarse hasta siete cinturones (Guerra-García et al., 2000, 2001a), algunos de estos eran muy parecidos entre sí y seoptó por considerar únicamente los cinco cinturones que podían diferenciarse con mayor claridad. En cada uno de loscinturones, se dispusieron al azar tres cuadrículas de 20 x 20 cm2 y se embolsaron todas las algas presentes en el cuadradodejando la roca totalmente descubierta tras raspar con una espátula. Los muestreos se llevaron a cabo durante los mesesde marzo, abril o mayo de 2003, 2004 y 2005, siempre en bajamar viva, considerándose los cinturones algales desde el nivelcero de marea (cinturón 1) hasta la zona más alta del intermareal (cinturón 5).
Las muestras se fijaron en formol al 4% y se llevaron al laboratorio donde se procedió a la separación e identificación delos organismos. Las algas dominantes se identificaron hasta nivel de especie y a cada una de ellas se le asignó

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 272/276
Almoraima, 35, 2007
272
cualitativamente un valor de cobertura relativo comprendidoentre 0 y 3 [0=ausente, 1=presente (0.1-20 %), 2=abundante(20-60 %), 3=muy abundante (60-100 %)]. En el caso de lamacrofauna asociada, se cuantificó el número de poliquetos,moluscos, crustáceos y equinodermos de cada muestra.
Análisis estadísticosPara cada uno de los cinco cinturones se calculó la media y elerror estándar de las abundancias de los distintos grupos deorganismos, y de los índices de diversidad de Shannon-Wiener(Shannon y Weaver, 1963) y de equitatividad de Pielou (Pielou,
1984). Por otra parte, se llevaron a cabo análisis multivariantesde ordenación y clasificación. Con la matriz cualitativa de algasy con la matriz cuantitativa de macrofauna asociada se realiza-ron sendos análisis de Cluster empleándose el algoritmo deagrupación UPGMA y el índice de similaridad de Bray-Curtis.Para la comparación entre la ordenación de los cinturones apartir de las algas y de la macrofauna asociada, se utilizó elanálisis Canónico de Correspondencias (CCA) y para verificarla significación estadística del análisis se aplicó el test deMonte Carlo. Para los análisis estadísticos se utilizaron losprogramas PRIMER (Clarke y Gorley, 2001) y PC-ORD(McCune y Mefford, 1997).
RESULTADOS Y DISCUSIÓN
Las algas dominantes en el transecto y sus valores de abundancia cualitativa se representan en la tabla 1. Además de loscorrespondientes a los años 2003, 2004 y 2005, se han incluido también los datos obtenidos en 1998 durante el estudio deGuerra-García et al. (2000). El primer cinturón, junto al nivel cero de marea, estuvo dominado por Gelidium sesquipedaleen todos los muestreos. El segundo cinturón estuvo dominado por Gymnogongrus patens en 1998 mientras que en el restode años fue Corallina elongata la especie dominante. Jania rubens fue la especie con mayor cobertura en el tercer nivel,
junto con Lithophyllum lichenoides, Laurencia pinnatifida, Corallina elongata y Caulacanthus ustulatus. El cuarto nivel
Figura 3. Contribución de cada grupo de macrofauna asociada a la abundancia total del intermareal.
Figura 2. Análisis de Cluster elaborados con la matriz de algas y la matriz de
macrofauna asociada. Se han utilizado todas las réplicas (a,b,c) de cadacinturón (1-5) los tres años muestreados (2003, 2004 y 2005).

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 273/276
Comunicaciones
273
estuvo dominado por algas verdes, Ulva rigida yChaetomorpha spp. y el quinto cinturón estuvo claramentedominado por Fucus spiralis en todos los muestreos. No seaprecian cambios importantes en la composición de algas a lolargo del tiempo, si bien parece existir una regresión deespecies como Gigartina acicularis, Gymnogongrus patens,
Laurencia pinnatifida y Valonia utricularis, y un incrementoen la cobertura de Ulva rigida. Sería interesante prestarespecial atención a este ligero aumento en la proporción de U.rigida, ya que, generalmente el incremento en el porcentaje
de algas verdes tiende a asociarse con procesos de eutrofización(Niell y Pazó, 1978).
El análisis de Cluster elaborado a partir de los valores cuali-tativos de cobertura de las algas (figura 2), mostró una claraseparación entre los cinturones de algas, independientementedel año muestreado. Las réplicas correspondientes a losdistintos cinturones forman subgrupos diferentes. Los cintu-rones 1 y 5 difieren claramente del resto, mientras que entrelos cinturones 2, 3 y 4 hay una mayor similitud. En cualquiercaso, el análisis refleja que las muestras se separan fundamen-talmente en función de la posición que ocupan en el transectoy que no existen diferencias claras entre los tres añosmuestreados (2003, 2004 y 2005). En el cluster obtenido apartir de las abundancias de los distintos grupos de macrofauna,la separación entre cinturones no es tan clara y solamentealgunas réplicas de los cinturones 1 y 5 parecen separarseclaramente del resto. Esto es debido a que en este caso, laidentificación ha sido a nivel de grandes grupos taxonómicosy no a nivel de especie. Posiblemente si los organismos seidentificaran hasta las categorías de género o especie, laseparación entre los cinturones sería más clara.
Cuando se representaron los porcentajes totales (sin conside-rar cinturones) de cada grupo de organismos de la faunaasociada (figura 3), los crustáceos fueron claramente domi-nantes en número de individuos en los años 2003 y 2004,mientras que en 2005 dominaron los moluscos. Parece existiruna tendencia temporal hacia el incremento del número demoluscos, especialmente gasterópodos y bivalvos, en detri-mento de los crustáceos (figura 3). Los crustáceos dominan-tes fueron los gammáridos, seguidos de caprélidos e isópodos. Figura 4. Valores medios y error estándar de la media de la abundancia (nº
organismos/0.04m2), diversidad de Shannon-Weaver (H’) y equitatividad (J)del total de fauna asociada en cada uno de los cinco cinturones del transecto.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 274/276
Almoraima, 35, 2007
274
Figura 5. Valores medios y error estándar de la media de la abundancia (nºorganismos/0.04m2) de cada uno de los grupos taxonómicos de macrofaunaasociada considerados.
Si consideramos los cinturones por separado, los mayoresvalores de abundancia se registraron en los niveles 2 y 3(figura 4). Los niveles más altos (4 y 5) pasan más tiempoemergidos, y por tanto son menos los organismos que puedensoportar la desecación. El nivel 1 por su parte, aunque es elque está más tiempo sumergido y, por tanto, con mayoresniveles de humedad, está más expuesto a la batida de las olascreándose unas condiciones de estrés negativas para muchasespecies. En el caso de la diversidad se registró un patróngeneralizado de disminución conforme nos aproximamos a
los niveles más altos, mientras que la equitatividad de grupostaxonómicos fue más baja en las estaciones intermedias,debido a los picos de abundancia. La figura 5 muestra comolas abundancias de los distintos grupos dominantes (moluscos,crustáceos y poliquetos) es mayor en los cinturones interme-dios 2 y 3. El patrón obtenido en 2004 y 2005 es muy similar,pero en 2003 se registró una abundancia considerable decrustáceos en el nivel 1 de Gelidium sesquipedale. En cual-quier caso, las abundancias decrecen significativamente con-forme nos aproximamos a los cinturones más elevados domi-nados por el alga Fucus spiralis.
El análisis CCA (figura 6) muestra las relaciones existentesentre los grupos de la macrofauna y las especies de algas. Eleje 1 absorbió el 32% de la varianza total y el eje 2 el 25%. Lasespecies Laurencia pinnatifida, Caulacanthus ustulatus,
Lithophyllum lichenoides y Valonia utricularis,correlacionaron significativamente con el primer eje, siendolas especies que más influyeron en la composición y abundan-cia de la fauna epífita. Las algas Corallina elongata,Chaetomorpha spp. y Gelidium sesquipedale fueron tambiénimportantes en la distribución de la macrofauna asociada ya
que correlacionaron significativamente con el segundo eje.El presente estudio supone el punto de partida para futurosestudios más específicos en la zona intermareal de la Isla delas Palomas. Es conveniente que este seguimiento anualbasado en las especies de algas más importantes y en losprincipales grupos de macrofauna asociada, siga llevándose acabo todos los años como parte de los programas de vigilanciay monitorización del Parque Natural del Estrecho. Algosimilar podría también realizarse en otros puntos del parquepara elaborar una red de seguimiento y control de todos los

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 275/276
Comunicaciones
275
Figura 6. Representación gráfica del análisis de componentes principales(CCA) elaborado a partir de la matriz de algas y de la matriz de fauna asociada.
Figura 7. Alumnos de la asignatura de Biología Marina de la Facultad deBiología, en la isla de Tarifa, muestreando en la zona intermareal (arriba) yrealizando buceo con equipo ligero (abajo).
intermareales del parque natural, ya que estas zonasmediolitorales son muy frágiles y se encuentran muy deterio-radas en la mayor parte de la costa andaluza debido a lafacilidad de acceso y la consiguiente presión humana (excur-sionismo, marisqueo incontrolado, etc.) En este sentido lalabor de alumnos y voluntarios sería muy útil en la ejecuciónde las campañas de muestreo (figura 7) y separación deorganismos. Términos como formación docente, educaciónambiental y voluntariado deben discurrir paralelos a los deseguimiento y control para lograr objetivos comunes destina-
dos a la conservación de nuestros ecosistemas marinos.
BIBLIOGRAFÍABOJA, 2003. Boletín Oficial de la Junta de Andalucía. Decreto 308/2002 por el quese aprueba el Plan de Ordenación de Recursos Naturales del Frente LitoralAlgeciras-Tarifa (BOJA nº18, 28 enero 2003, pp. 1849-1935) y Decreto 57/2003de declaración del Parque Natural del Estrecho (BOJA nº54, 20 marzo 2003, pp.5925-5944)CLARKE, K.R. & R.N. Gorley, . 2001. Primer (Plymouth Routines in MultivariateEcological Research) v5: User Manual /Tutorial. PRIMER-E Ltd., Plymouth, pp. 91.GUERRA-GARCÍA, J.M. & J.C. García-Gómez, 2000. La fauna submarina de laIsla de las Palomas (Tarifa, Cádiz). Temas de Flora, Fauna y Ecología del Campode Gibraltar. Cuadernos del Instituto II , Campo de Gibraltar. pp. 7-17.
GUERRA-GARCÍA, J.M., J.E. Sánchez-Moyano, J. CORZO , S. MORENO & J.C.García-Gómez, 2000. Descripción de las comunidades de algas de la Isla de lasPalomas (Tarifa) y de otros enclaves del sur de España. Almoraima, 23: pp. 189-194.GUERRA-GARCÍA, J.M., J.E. Sánchez-Moyano, J.Corzo, C.M. López de laCuadra, S. Moreno & J.C. García-Gómez, 2001a. El macrofitobentos mediolitoralde la Isla de las Palomas (Tarifa, sur de España), un enclave sujeto a estudio técnicopara su conservación. Actas de las I Jornadas Internacionales sobre reservasmarinas. Ed. Ministerio de Agricultura, Pesca y Alimentación, pp. 383-391.GUERRA-GARCÍA, J.M., I. García-Asencio, & J.E. Sánchez-Moyano, 2001b."Parvipalpus onubensis, a new species (Crustacea: Amphipoda: Caprellidea) fromthe Atlantic coast of Southern Spain". Scientia Marina 65(4): pp. 333-339.McCUNE, B. & M.J. Mefford, 1997. PC-ORD. Multivar iate Analysis of EcologicalData. MJM Software Design, Gleneden Beach, pp. 77.NIEL, F.X. & PAZÓ, J.P. 1978. "Incidencia de vertidos industriales en laestructura de poblaciones intermareales. II. Distribución de la biomasa y de la
diversidad específica de comunidades de macrófitos de facies rocosa". Investiga-ciones Pesqueras 42(2): pp. 213-239.PIELOU, E.C. 1984. The interpretation of ecological data: a primer on classificationand ordination. John Wiley an Sons, New York, pp. 263.SHANNON, C.E. & W. Weaver, 1963. The mathematicl theory of communication.University of Illinois Press, Urbana, Illinois, pp. 117.

8/9/2019 Almoraima 35
http://slidepdf.com/reader/full/almoraima-35 276/276