variación geográfica en los venados de cola blanca ...venados de cola blanca, representativa de un...
TRANSCRIPT

Memoria de la Fundación La Salle de Ciencias Naturales 2007, 167: 29-72
Resumen. Se ha propuesto recientemente que los venados de cola blanca néarticos y neotropicalesno son conespecíficos y que los últimos se dividen en varias especies. En el contexto del debate queesta propuesta ha generado, aquí se presenta: a) una comparación de la variabilidad geográfica devenados de cola blanca norteamericanos y venezolanos basada en una nueva unidad para medirvariación clinal, cuyo nombre está dedicado a la memoria del gran biogeógrafo León Croizat; b) unareevaluación taxonómica de las diferencias morfológicas y moleculares entre estos venados; y c)escenarios paleobiogeográficos que, en concordancia con información morfológica, molecular,paleobiológica y tectónica, proponen especiaciones independientes para los venados de los páramosandinos y de Margarita. Los nuevos análisis permiten reafirmar que: 1) los venados de cola blancaneárticos (O. virginianus) y neotropicales no son conespecíficos; 2) el venado de páramo de Mérida(O. lasiotis) es una especie válida aparte de los venados de páramo colombiano (O. goudotii) yecuatoriano (O. ustus); 3) el venado enano de Margarita (O. margaritae) también es una especieválida; y 4) el resto de los Odocoileus venezolanos pertenecen a otra especie válida (O. cariacou) conamplia distribución neotropical. Odocoileus lasiotis y O. margaritae están en peligro de extinción.
Palabras clave. Odocoileus. Páramo. Isla de Margarita. Variación clinal. Biogeografía.
Geographic variation in Venezuelan white-tailed deer (cervidae, Odocoileus), withemphasis on O. Margaritae, the Margarita Island dwarf species
Abstract. It has been recently proposed that nearctic and neotropical white-tail deer are notconspecific, and that the latter are divided into several species. In the context of the debate that sucha proposal has generated, this study presents: a) a comparison of the geographic variability of NorthAmerican and Venezuelan white-tail deer based on a new unit of measurement for clinal variation,whose name is dedicated to the memory of the great biogeographer León Croizat; b) a taxonomicrevaluation of morphological and molecular differences among these deer; and c) paleobiogeographics c e n a rios that, in agreement with morphological, molecular, paleobiological, and te c to n i cinformation, propose independent speciations for deer of the Andean páramos and Margarita Island.The new analyses allow to reaffirm that: 1) nearctic (O. virginianus) and neotropical white-tailed deerare not conspecific; 2) the Mérida páramo deer (O. lasiotis) is a valid species separate from Colombian(O. goudotii) and Ecuadorian (O. ustus) páramo deer; 3) the Margaritan dwarf deer (O. margaritae)is also a valid species; and 4) the remaining Venezuelan Odocoileus belong to another valid species (O.cariacou) with an ample neotropical distribution. Odocoileus lasiotis and O. margaritae areendangered.
Key words. Odocoileus. Páramo. Margarita Island. Clinal variation. Biogeography.
Variación geográfica en los venados de cola blanca(Cervidae, odocoileus) de Venezuela, con énfasis en O.Margaritae, la especie enana de la Isla de Margarita
Jesús Molinari

30 Variación geográfica en venados
Introducción
La historia taxonómica de los venados de cola blanca se inició con las descrip-ciones de Dama virginiana (localidad típica: Virginia, Estados Unidos) en 1780, yCervus capreolus cariacou (localidad típica: Guayana Francesa) en 1784 (Cabrera1961, Smith 1991). Los naturalistas del siglo XIX creían que estos venados estabandivididos en numerosas especies, lo cual se manife stó en la pro l i fe ración dedescripciones taxonómicas (Cabrera 1961, Smith 1991). Aunque durante la primeramitad del siglo XX se propuso considerar a todas las variedades de venado de colablanca una misma especie, Odocoileus virginianus (Lydekker 1915, Hershkovitz 1948),por muchos años más persistió la creencia de que los venados de cola blanca eranvarias especies. Por ejemplo, Méndez-Arocha (1955) argumentó que los representantessuramericanos debían ser reconocidos como una especie aparte de O. virginianus, asaber, O. cariacou. Cabrera y Yepes (1960) llegaron incluso más lejos, reconociendotres especies suramericanas independientes de O. virginianus, a las cuales se referíancomo "cariacús", a saber, O. suacuapara (= cariacou) de tierras bajas, O. columbicus (=goudotii) de los andes colombianos y venezolanos, y O. peruvianus de los andes dePerú y Ecuador. A consecuencia de la influyente obra póstuma de Cabrera (1961),d u ra n te las últimas cuatro décadas del siglo XX el debate sobre la posibleconespecificidad entre los venados de cola blanca norte y suramericanos cedió su lugara la noción, universalmente aceptada, de sólo una especie, O. virginianus, distribuidadesde Canadá hasta Panamá, y desde Colombia hasta las Guayanas y norte de Brasilpor el este, y hasta Bolivia por el sur (Smith 1991, Wilson y Reeder 1993, Tarifa et al.2001, Weber y González 2003). Durante el Pleistoceno, la distribución de Odocoileusen Suramérica fue más amplia que la actual, como lo demuestran restos fósileshallados en el sureste de Brasil (Cartelle 1994). De las 38 subespecies vivientestradicionalmente reconocidas, ocho son suramericanas (Smith 1991). Según Cabrera(1961), las subespecies presentes en Venezuela son O. v. gymnotis, de las tierras bajascontinentales del país, O. v. goudotii, de los páramos del Estado Mérida, y O. v.margaritae, de la Isla de Margarita.
Debido a la escasez de información taxonómicamente relevante, las vicisitudesnomenclaturales de los venados de cola blanca durante el siglo XX obedecieron mása un vaivén de opiniones que a la aparición de nuevos datos (Lydekker 1915, Cabrera1918, 1961, Hershkovitz 1948, Méndez-Arocha 1955, Cabrera y Yepes 1960, Brokx1984). Incluso hoy en día, no hay certeza sobre cuál es el verdadero número de especiesy subespecies en que deben dividirse estos venados, ni sobre los caracteres diagnósticosy límites de distribución de tales taxa.
El debate sobre la conespecificidad entre los venados de cola blanca norte ysuramericanos se basó casi exclusivamente en sólo un carácter externo (la glándulametatarsal, un pliegue de 4 cm de longitud, rodeado por un mechón prominente depelos, en la parte externa de cada una de las patas traseras; Sauer 1984, Quay 1971)siempre grande y funcional en los venados de cola blanca de Estados Unidos, Canadá

31Mem. Fund. La Salle de Cienc. Nat. 167
y norte de México, y siempre atrofiada en los del sur de México, Centro y Suramérica(Méndez-Arocha 1955, Hershkovitz 1958, Quay 1971, Brokx 1972, Baker 1984).Mientras Cabrera estuvo seguro de esta diferencia, fue partidario de reconocerespecies diferentes (Cabrera 1918, Cabrera y Yepes 1960), cuando basándose en infor-mación a la postre errónea, creyó que esta diferencia no existía, cambió de opinión,reconociendo una sola especie (Cabrera 1961).
Por basarse en información errónea e insuficiente, el cambio de opinión deCabrera (1961) siempre mereció reconsideración. En este sentido, Molina y Molinari(1999) aportaron por primera vez datos sobre la frecuencia de caracteres discretoscraneanos y la morfometría mandibular de una muestra relativamente grande devenados de cola blanca, representativa de un área geográfica extensa de Suramérica,como es Venezuela. La comparación de estos datos con información equivalentedisponible para Norteamérica evidenció que el hiato morfológico que separa a losvenados de cola blanca venezolanos y norteamericanos es de gran magnitud, lo cualpermitió proponer la restricción del nombre específico virginianus a los venados decola blanca norteamericanos. Además, la comparación basada en caracteres discretosevidenció que, pese a la menor área geográfica que ocupan, la diferenciación internade los venados venezolanos excede la de los venados norteamericanos, lo cual permitióproponer el reconocimiento, para los venados venezolanos de tres especies plenas:cariacou (sinónimo: gymnotis), lasiotis (endémico de páramos del Estado Mérida,excluido de goudotii), y margaritae. Complementariamente, los datos morfométricosevidenciaron diferencias adicionales entre las tres especies propuestas.
El estudio de Molina y Molinari (1999) fue rápidamente seguido por otro(Moscarella 2001, Moscarella et al. 2003), basado en el análisis de secuencias de laregión de control mitocondrial (D-loop) de venados de cola blanca venezolanos ynorteamericanos. Pese a aportar resultados que, congruentemente con los de Molinay Molinari (1999), indican una notable divergencia filética entre venados de colablanca suramericanos y norteamericanos, Moscarella et al. (2003) continuaronrefiriendo las formas suramericanas a O. virginianus. Para los venados venezolanos enparticular, Moscarella et al. (2003) concluyeron que, por ser su diferenciación genéticainterna inferior al 3%, no hay motivo para reconocer las especies propuestas porMolina y Molinari (1999).
El objetivo del presente trabajo es contrastar, utilizando información adicional ynuevos análisis (entre los cuales se cuentan los basados en una nueva medida de varia-ción clinal), la evidencia y argumentos presentados por Molina y Molinari (1999) yMoscarella et al. (2003). Es de esperar que los resultados obtenidos, además de aclarardudas taxonómicas, estimulen estudios de campo y medidas de conservación encami-nados a conocer mejor y a proteger de la extinción a los venados de páramo yMargarita.

32 Variación geográfica en venados
Materiales y Métodos
Molina y Molinari (1999) revisaron los cráneos y mandíbulas de 140 ejemplaresadultos de venados de cola blanca venezolanos, incluyendo nueve de la Isla deMargarita, registrando dos tipos de información: 1) frecuencia de 13 caracteresdiscretos craneomandibulares, los mismos considerados en un estudio previo sobrevenados de cola blanca norteamericanos (Rees 1969a); y 2) magnitud de 14 variablescontinuas mandibulares, también empleadas en estudios previos sobre venados de colablanca norteamericanos (Rees 1969ab). Los nuevos análisis aquí presentados se basanen análisis no morfométricos (frecuencias de aparición de dichos caracteres discretos)y morfométricos (promedios de dichas variables continuas) de 17 grupos geográficosde venados venezolanos y norteamericanos (Rees 1969a, b; Molina y Molinari 1999).Pa ra estimaciones de masa corp o ral, estos datos fueron comp l e m e n tados coninformación morfométrica de otros estudios (Lavengood et al. 1994, Schaefer y Main1999, Peterson 2003).
Según la nomenclatura tradicional de los venados de cola blanca (Cabrera 1961,Smith 1991), los grupos geográficos considerados (Rees 1969a, b; Molina y Molinari1999) corresponderían a tres subespecies venezolanas (goudotii, gymnotis y margaritae)y ocho norte a m e ricanas (b o re a l i s, c o u e s i, d a c ote n s i s, m a c ro u ru s, m c i l h e n ny i,seminolus, texanus y virginianus).
Análisis de caracteres discretos craneomandibulares
Empleando distancias euclidianas como medida de diferenciación y ligamientopromedio como estrategia de aglomeración, Molina y Molinari (1999) usaron datossobre frecuencias de caracteres discretos craneanos y mandibulares para construir undendrograma que unía, según su similaridad, 7 grupos de venados venezolanos y 10grupos de venados norteamericanos. Contrariamente a lo aseverado por Moscarella etal. (2003), los caracteres discretos como los empleados por Molina y Molinari (1999)son de valor para el estudio de filogenias en cérvidos (p. ej. Bouvrain et al. 1989). Porello, el nuevo análisis aquí presentado de los caracteres discretos de Molina y Molinari(1999) y Rees (1969a) se basa en un tipo de dendrograma, comúnmente empleado encontextos filogenéticos, denominado árbol de "unión de vecinos" (neighbor-joining),también usado por Moscarella et al. (2003) para sus datos genéticos. La construcciónde este dendrograma requirió tres pasos: 1) creación de una matriz de distanciaseuclidianas entre todos los grupos de venados venezolanos y norteamericanos; 2)estandarización de dichas distancias dividiéndolas por su desviación estándar; y 3)ingreso de la matriz de valores estandarizados resultante en el programa MEGAversión 2.1 (Kumar et al. 2001).
Análisis de medidas mandibulares
Al aplicarse a datos morfométricos continuos, el Análisis de ComponentesPrincipales (ACP) permite aislar en el primer eje la varianza debida a diferencias de

33Mem. Fund. La Salle de Cienc. Nat. 167
tamaño, y en los ejes subsiguientes la varianza debida a diferencias de forma (Jolicoeury Mosimann 1960, Bookstein et al. 1985, Schonewald et al. 1985, Rohlf y Bookstein1993, De Marinis 1995, Gutiérrez 2004, Lorenzo et al. 2004). Esto es patente en que,en el primer eje, los pesos de todas las variables son cercanos a +1, mientras que, en losejes del segundo al último, dichos pesos varían ampliamente entre -1 y +1.
Aprovechando esta propiedad del ACP, Molina y Molinari (1999) demostraronque los venados de cola blanca venezolanos poseen una considerable diferenciacióngeográfica en tamaño, forma y dimorfismo sexual de las mandíbulas. Los nuevosanálisis morfométricos de las medidas mandibulares de Rees (1969a, b) y Molina yMolinari (1999) aquí presentados también se valen de un ACP para disectar tamaño yforma, pero se centran en la comparación de venados de cola blanca venezolanos ynorteamericanos, aspecto no considerado por estos autores. Para igualar los tamañosy unidades de las escalas, facilitando así su comparación e interpretación, los puntajesde los ACP aquí graficados aparecen divididos por sus desviaciones estándar.
Estimación de masas corporales
Las masas corporales medias de los venados de cola blanca venezolanos sedesconocen. Mediante un procedimiento similar al empleado por Lowe (1972) para elciervo rojo europeo, fue posible obtener estimados de las masas corporales medias devenados venezolanos y norteamericanos. El procedimiento consistió en efectuar unare gresión lineal, empleando va l o res prev i a m e n te tra n s fo rmados en loga ri t m o snaturales, entre la longitud media del diastema mandibular (eje vertical) y la masacorporal media (eje horizontal) de una especie de cada género de Odocoileinaeamericano, excluyendo a los morfológicamente muy divergentes Alces y Rangifer. Losdatos incluidos en esta regresión (Mendoza et al. 2002) correspondieron a Blastocerosd i ch oto m u s ( d i a stema 8,6 cm; masa 130 kg), Odocoileus virg i n i a n u s ( 7,5; 74 ) ,Hippocamelus bisulcus (7,3; 50), Ozotocerus bezoarticus (5,1; 38), Mazama americana(4,5; 20) y Pudu mephistophiles (2,7; 9).
El valor del coeficiente de correlación fue muy alto (r= 0,971) y mayor que el deanálisis alternativos empleando otras medidas mandibulares en lugar de la longitud deldiastema. La ecuación de regresión fue ln(D)= ln(a) + b ln(M), que despejando lleva aM= (D/a)1/b; donde D= longitud del diastema, M= masa corporal, ln(a)= intersecciónde la recta de regresión con el eje vertical, y b= pendiente de la recta. Con los valoresutilizados ln(a)= 0,1070 y b= 0,4398; por ende a= 1,1130 y 1/b = 2,2735; lo cual lleva aM =(D/1,1130)2,2735 como ecuación final utilizada para estimar masas corporales devenados de cola blanca. Esta ecuación parece especialmente precisa para estimar lamasa corp o ral de machos. Pa ra hembras, la ecuación parece producir unasobrevaluación de la masa que podría alcanzar 15%.
Hay que destacar que la longitud del diastema puede medirse en animales vivos,por lo cual la ecuación obtenida es de interés para trabajos de campo y podríafácilmente perfeccionarse con nuevos datos. Sin embargo, la ecuación no debeaplicarse a Odocoileus hemionus, pues esta especie difiere notablemente de O .virginianus en la longitud relativa del diastema mandibular (Rees 1971).

34 Variación geográfica en venados
Nuevo sistema de unidades para cuantificar variación geográfica
Para permitir la cuantificación precisa de diferenciaciones geográficas, propongouna nueva unidad, denominada croizat (abreviada cr) en honor al gran botánico ybiogeógrafo italo-venezolano León Croizat (1894-1982). Para varias poblaciones, uncroizat pude definirse como "la cantidad de cambio equivalente a la que presentaríandos poblaciones si estuviesen separadas por 1 km de distancia geográfica y difiriesenmorfológicamente en una milésima de la desviación estándar de las distanciaseuclidianas entre todos los pares posibles de poblaciones". Considérese la relación:Cambio morfológico interpoblacional= Distancia morfológica / Distancia geográfica.
Si se expresa el numerador en distancias euclidianas estandarizadas (es decir,distancias euclidianas divididas por su desviación estándar) y el denominador en km, yse multiplica el cociente obtenido por 1000, la cantidad resultante vendrá expresada encroizats. En términos prácticos, esta multiplicación por 1000, además de ahorrardecimales en los va l o res obtenidos (no cabe esperar dife rencias morfo l ó g i c a simportantes entre poblaciones separadas por 1 km), hace que las magnitudes encroizats expresen la cantidad de cambio que se observaría por cada 1000 km deseparación entre poblaciones.
Las distancias euclidianas estandarizadas pueden calcularse para diferenciashereditarias cuantitativas de cualquier tipo, tales como frecuencias de caracteresdiscretos y valores de caracteres continuos. El croizat puede verse como una unidadconceptualmente comparable al darwin (Haldane 1949) y al haldane (Gingerich 1993),con la diferencia principal de que la distancia morfológica se divide por la distanciageográfica en lugar de por el tiempo o el número de generaciones. Aunque ladefinición de croizat presentada se refiere a poblaciones, también podría usarse paracomparar individuos, para lo cual las distancias euclidianas empleadas tendrían queser interindividuales, en lugar de interpoblacionales.
Los valores presentados de cambio morfológico interpoblacional en función de ladistancia geográfica, expresados en croizats, consideran por separado las distanciaseuclidianas entre grupos ge o grá ficos de venados con re s p e c to a: 1) datos nomorfométricos, frecuencias de caracteres discretos; 2) datos morfométricos (tamañomandibular), puntajes en el primer componente del ACP de variables continuasm a n d i b u l a res; y 3) datos morfo m é t ricos (fo rma mandibular), puntajes en loscomponentes a partir del segundo del mismo ACP.
El cálculo de distancias geográficas entre grupos geográficos de venados, con baseen la latitud y longitud de un punto aproximadamente central en el área del mapaocupada por cada uno de dichos grupos, fue posible gracias a la versión HTML delprograma World Distance Calculator de la Fédération Aéronautique Internationale.
Resultados
Caracteres discretos craneomandibulares

35Mem. Fund. La Salle de Cienc. Nat. 167
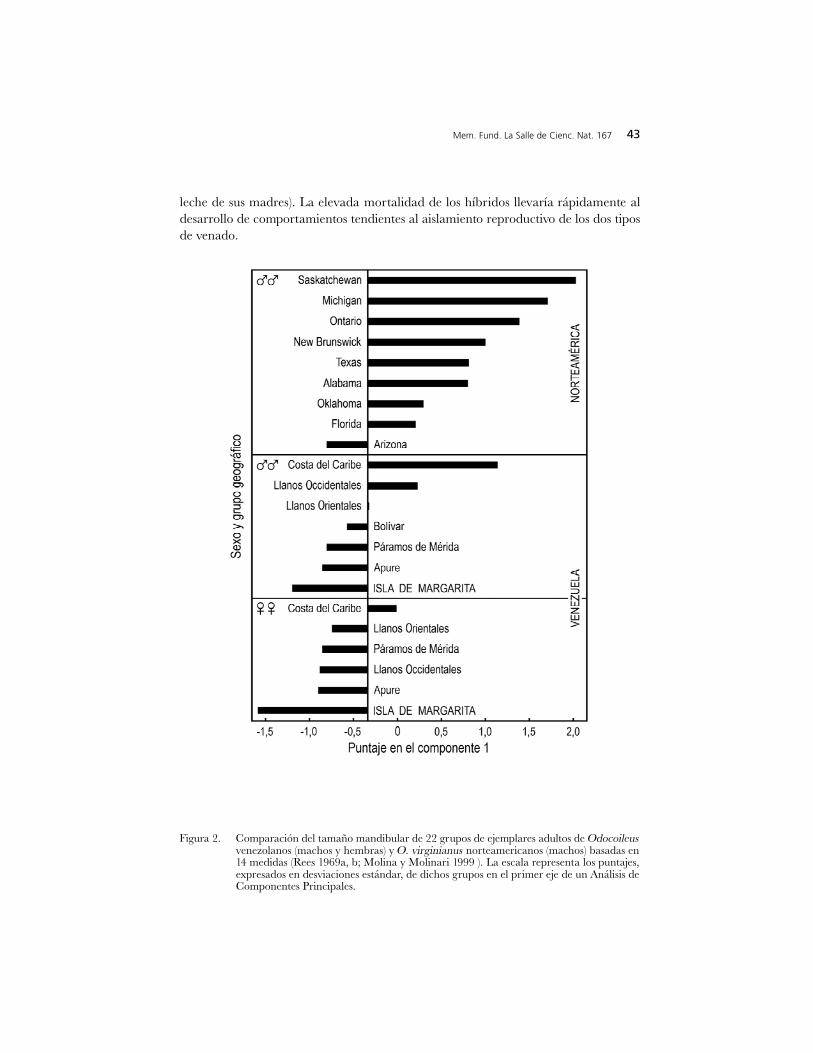
El árbol de unión de vecinos (Figura 1), resume la información sobre frecuenciasde caracteres discretos craneanos y mandibulares en 17 grupos de venados de colablanca venezolanos y norteamericanos (Rees 1969a, Molina y Molinari 1999). Esteárbol corrobora los siguientes aspectos principales (Molina y Molinari 1999): 1) losvenados de cola blanca venezolanos son muy dive rge n tes con re s p e c to a losnorteamericanos; 2) con relación a sus separaciones geográficas, que son muchomenores, los venados de cola blanca venezolanos muestran mayor diferenciación quelos norteamericanos; 3) pese a la corta distancia entre la Isla de Margarita y tierrafirme, la diferenciación del venado insular respecto al resto de sus congéneresvenezolanos excede en magnitud la de los venados de cola blanca norteamericanos deregiones tan distantes como Ontario y Arizona; 4) pese a que los páramos de Méridase encuentran geográficamente cerca de los llanos, la diferenciación del venado de colablanca andino con respecto a sus congéneres llaneros también es considerable; y 5) delos venados de cola blanca de tierras bajas continentales, los más diferenciados son losde la costa. No obstante, es necesario reiterar que la pequeña muestra disponible esuna mezcla de ejemplares pertenecientes a variedades seguramente diferentes, unaoccidental y otra oriental (Molina y Molinari 1999). Además, el árbol aquí presentado(Figura 1) revela un cierto grado de diferenciación craneana del venado de cola blancade Apure (que además destaca por su pelaje claro y tamaño pequeño; Brokx 1984), conrespecto al resto de sus congéneres llaneros, aspecto no evidenciado anteriormente(Molina y Molinari 1999).
Medidas mandibulares y masas corporales
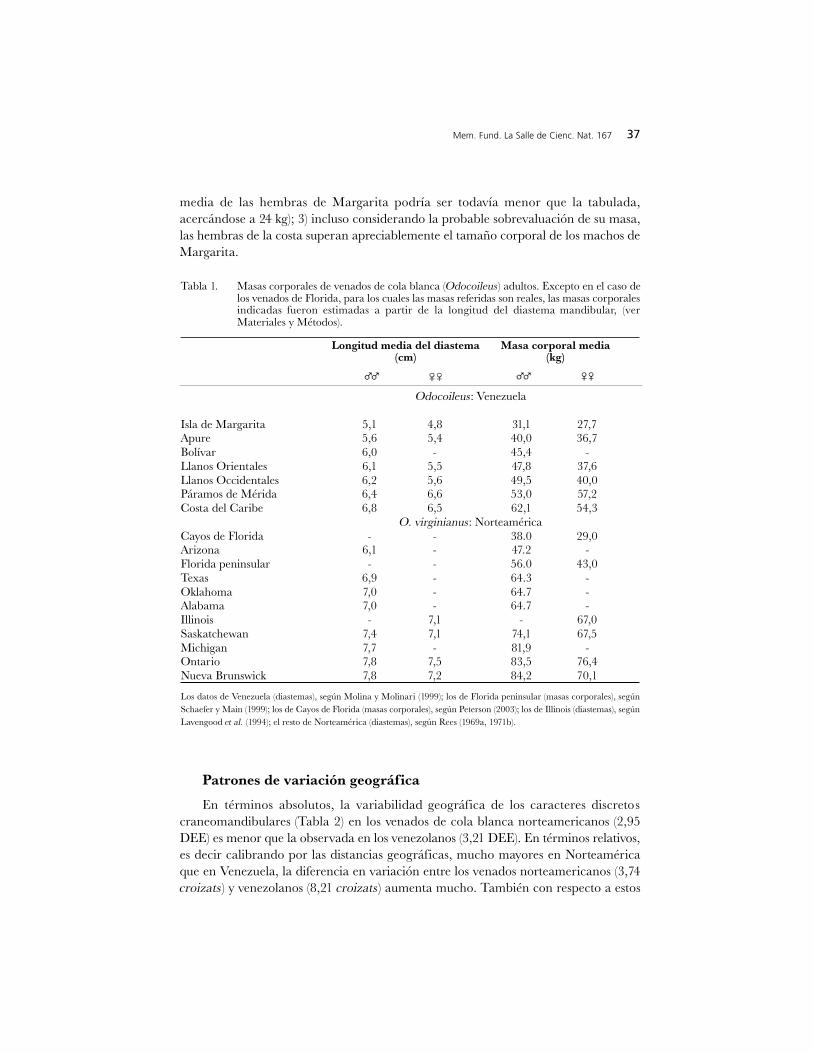
Los valores de masa corporal estimados (Tabla 1), coinciden bien con los valoresde masa reales de los venados de cola blanca norteamericanos (Sauer 1984). Losvenados de la costa del Caribe y de la Isla de Margarita representan, respectivamente,los Odocoileus de mayor y de menor tamaño de Venezuela. La masa estimada de losvenados de la costa es similar a la estimada para los venados de la mitad sur de losEstados Unidos. La masa estimada del venado de Margarita es considerablementemenor que la masa conocida del venado de los Cayos de Florida, el Odocoileus máspequeño de Norteamérica, lo cual es consistente con el hecho de que el cráneo delvenado de Margarita es de menores dimensiones (Osgood 1910, Maffei et al. 1988,Molina y Molinari 1999, Schaefer y Main 1999).
Las diferencias en tamaño cefálico y masa corporal (Figuras 2, 3; Tabla 1)conjuntamente indican que: 1) los machos de la costa superan ampliamente a los deMargarita en tamaño corporal y mandibular, lo último también implica grandesdiferencias en las dimensiones de la cabeza y de las astas; 2) la diferencia en tamañocorporal entre los machos de la costa y las hembras de Margarita es substancialmentemayor que la de estos mismos machos con sus propias hembras (debido a la probablesobrevaluación de las masas corporales de las hembras ya notada; la verdadera masa

36 Variación geográfica en venados
37Mem. Fund. La Salle de Cienc. Nat. 167
media de las hembras de Margarita podría ser todavía menor que la tabulada,acercándose a 24 kg); 3) incluso considerando la probable sobrevaluación de su masa,las hembras de la costa superan apreciablemente el tamaño corporal de los machos deMargarita.
Patrones de variación geográfica
En términos absolutos, la variabilidad geográfica de los caracteres discretoscraneomandibulares (Tabla 2) en los venados de cola blanca norteamericanos (2,95DEE) es menor que la observada en los venezolanos (3,21 DEE). En términos relativos,es decir calibrando por las distancias geográficas, mucho mayores en Norteaméricaque en Venezuela, la diferencia en variación entre los venados norteamericanos (3,74croizats) y venezolanos (8,21 croizats) aumenta mucho. También con respecto a estos
Tabla 1. Masas corporales de venados de cola blanca (Odocoileus) adultos. Excepto en el caso delos venados de Florida, para los cuales las masas referidas son reales, las masas corporalesindicadas fueron estimadas a partir de la longitud del diastema mandibular, (verMateriales y Métodos).
Longitud media del diastema Masa corporal media(cm) (kg)
Odocoileus: Venezuela
Isla de Margarita 5,1 4,8 31,1 27,7Apure 5,6 5,4 40,0 36,7Bolívar 6,0 - 45,4 -Llanos Orientales 6,1 5,5 47,8 37,6Llanos Occidentales 6,2 5,6 49,5 40,0Páramos de Mérida 6,4 6,6 53,0 57,2Costa del Caribe 6,8 6,5 62,1 54,3
O. virginianus: NorteaméricaCayos de Florida - - 38.0 29,0Arizona 6,1 - 47.2 -Florida peninsular - - 56.0 43,0Texas 6,9 - 64.3 -Oklahoma 7,0 - 64.7 -Alabama 7,0 - 64.7 -Illinois - 7,1 - 67,0Saskatchewan 7,4 7,1 74,1 67,5Michigan 7,7 - 81,9 -Ontario 7,8 7,5 83,5 76,4Nueva Brunswick 7,8 7,2 84,2 70,1
Los datos de Venezuela (diastemas), según Molina y Molinari (1999); los de Florida peninsular (masas corporales), según
Schaefer y Main (1999); los de Cayos de Florida (masas corporales), según Peterson (2003); los de Illinois (diastemas), según
Lavengood et al. (1994); el resto de Norteamérica (diastemas), según Rees (1969a, 1971b).

38 Variación geográfica en venados
caracteres (Tabla 2), en los venados norteamericanos se aprecia una elevada yaltamente significativa correlación de la diferenciación morfológica con la distanciageográfica (r= 0,67). En contraste, en los venados venezolanos, el pequeño y nos i g n i fi c a t i vo valor de dicha correlación (r= 0,13) indica que las dispari d a d e sintergrupales carecen de dirección geográfica.
En términos absolutos (Tabla 2), la variación en tamaño mandibular de losvenados norteamericanos (1,53 DEE) es ligeramente mayor que la de los venezolanos(1,38 DEE). En términos relativos, la de estos últimos es mucho mayor (0,85 contra3,97 croizats). En términos absolutos, la variación en forma mandibular de los venadosnorteamericanos (1,64 DEE) es menor que la de los venezolanos (2,30 DEE). Entérminos relativos, la diferencia aumenta mucho (1,03 contra 6,40 croizats). El signopositivo de las correlaciones del tamaño (r= 0,39) y la forma mandibular (r= 0,16) conla distancia ge o grá fica (Tabla 2), indica que los grupos vecinos de ve n a d o sn o rte a m e ricanos tienden a pare c e rse morfo m é t ri c a m e n te. En cambio, el signonegativo (r= -0,33 y -0,19 respectivamente) de dichas correlaciones indica que losgrupos vecinos de venados venezolanos tienden a ser diferentes morfométricamente.
Discusión
Confiabilidad de los datos genéticos disponibles
A los efectos de contrastar nuestros resultados con las conclusiones taxonómicas deMoscarella et al. (2003), es necesario hacer constar que, por ser ejemplares cautivos, 20de los 26 venados venezolanos de los cuales Moscarella et al. (2003) secuenciaron ADNno cumplen el requisito fundamental de tener un origen geográfico seguro para unestudio filogeográfico y taxonómico (Ruedas et al. 2000, Wheeler 2003), 11 de losejemplares dudosos pertenecían a cinco exhibiciones de fauna silvestre del país que nollevan buenos registros sobre sus animales y que mantienen juntos y entrecruzanvenados de diversos orígenes geográficos (Moscarella 2001). Los nueve restantes eranmascotas de habitantes de Margarita, e incluían cuatro venados de tierra firme: unodel vecino Estado Sucre, otro "de Maturín" (un origen imposible, ésta es una ciudad enla cual no hay venados silvestres), y dos del Estado Apure (Moscarella 2001). Elejemplar "de Sucre" fue el único venado de tierra f irme que, por su haplotipo, seagrupó con los de Margarita. Si este ejemplar fuese en realidad de Margarita, lasdistancias genéticas calculadas entre venados insulares y continentales habrían sidomayores.
Distancias genéticas
Según Moscarella et al. (2003) cariacou, margaritae y lasiotis, las tres especiesplenas de Odocoileus venezolanos reconocidas por Molina y Molinari (1999), carecende validez, por lo cual dichos taxa (aunque refiriéndose a lasiotis con el nombregoudotii), deben seguir siendo considerados sólo subespecies de O. virginianus. La

39Mem. Fund. La Salle de Cienc. Nat. 167
única evidencia ofrecida para justificar este retroceso taxonómico consiste en distan-cias genéticas mitocondriales. El razonamiento de Moscarella et al. (2003) tiene doselementos: 1) su propio hallazgo de que las distancias genéticas mitocondriales (D-loop) entre los Odocoileus venezolanos apenas rozan el 3%; y 2) una generalización,basada en ciertos ejemplos (Cronin 1992, Tamate et al. 1998, Nagata et al. 1999,Kuwayama y Ozawa 2000), conforme a la cual lo típico en los cérvidos sería que lasdistancias genéticas mitocondriales entre subespecies de una misma especie alcancen3%. Conjuntamente, ambos elementos indicarían que la variabilidad genética de losOdocoileus venezolanos es insuficiente para dividirlos en varias especies.
Una cuidadosa revisión de la literatura revela que es falso que, en los cérvidos, lasdistancias genéticas mitocondriales entre subespecies típicamente alcancen 3%. Pese asu variabilidad cariotípica y a su distribución en tres continentes, las ocho subespeciesde alce (Alces alces) son prácticamente indistinguibles desde el punto de vistamitocondrial (Cronin 1992, Udina et al. 2002, Hundertmark et al. 2002a, b, 2003).Para las todavía más ampliamente distribuidas especies de Cervus, la distanciagenética promedio entre subespecies a nivel del D-loop es 1,8% (Randi et al. 2001) y lasdistancias genéticas a nivel de citocromo b entre especies perfectamente válidas son aveces tan pequeñas (inferiores a 1-2%), que han sido calificadas de "marginales" (Pitra
Tabla 2. Parámetros cuantitativos de la variabilidad geográfica de venados de cola blancavenezolanos y norteamericanos. DEE = distancia euclidiana estandarizada. Variaciónabsoluta= media de las DEE intergrupales. Variación relativa= media de las DEEdivididas por las distancias geográficas intergrupales. Distancia–distancia= correlaciónentre distancias morfológicas y distancias geográficas que separan a los grupos devenados.
Sólo Sólo Venezuela contraVenezuela Norteamérica Norteamérica
Caracteres cualitativos craneomandibulares
Variación absoluta 3,21 DEE 2,95 DEE NSVariación relativa 8,21 croizats 3,74 croizats < 0,007**Distancia–distancia r = 0,13; NS r = 0,67; < 0.000001 < 0,02*
Tamaño mandibular
Variación absoluta 1,38 DEE 1,53 DEE NSVariación relativa 3,97 croizats 0,85 croizats < 0,002**Distancia–distancia r = –0,33; NS r = 0,39; < 0.02 < 0,02*
Forma mandibular
Variación absoluta 2,30 DEE 1,64 DEE < 0,05**Variación relativa 6,40 croizats 1,03 croizats < 0,0001**Distancia–distancia r = –0,19; NS r = 0,16; NS NS
NS = no significativo.*Coeficientes de correlación comparados mediante la transformación Z de Fisher.**Prueba t de dos colas suponiendo varianzas desiguales.

40 Variación geográfica en venados
et al. 2004). En los muntiacs (Muntiacus) y en los ciervos almizclados (Moschus), lasdistancias mitocondriales entre subespecies suelen ser inferiores a 1-2% y las diferenciasentre especies suelen estar por debajo del 3% (Giao et al. 1998, Su et al. 1999, Wang yLan 2000). Con base en el propio género Odocoileus, en el cual la distancia genéticaentre las especies norteamericanas a nivel de citocromo b puede llegar a ser de sólo1,3%; se ha propuesto que en los cérvidos lo típico podría ser que las especies hermanasse caractericen por divergencias genéticas mutuas muy bajas (Bradley et al. 2003).Ciertamente, la literatura registra algunos ejemplos en los cuales las distanciasgenéticas mitocondriales entre subespecies de cérvidos pueden alcanzar o rebasar el3% (Cronin 1992, González et al. 1998, Nagata et al. 1999, Kuwayama y Ozawa 2000).No obstante, estos ejemplos, y más en particular los citados por Moscarella et al. (2003)para apoyar su generalización, están siendo reevaluados taxonómicamente, lo cual haresultado en que casi todas las "subespecies" implicadas sean ahora consideradasespecies plenas (Polziehn y Strobeck 2002, Ludt et al. 2004, Pitra et al. 2004, Groves2005). Además, estas "subespecies" no deberían haberse usado para calibrar el gradode especiación de los Odocoileus venezolanos ya que las distancias o barrerasgeográficas que las separan son de mucha mayor envergadura que las presentes enVenezuela.
El uso del D-loop como única fuente de información taxonómica (Moscarella etal. 2003) contradice la noción actual de que no existen distancias genéticas "críticas"que perm i tan, por sí mismas, dete rminar la validez del estatus específico osubespecífico de poblaciones animales (Avise y Ball 1990, Sites y Crandall 1997, Naglet al. 2000, Puorto et al. 2001, Polziehn y Strobeck 2002). Para distinguir entre especiesy subespecies, antes que las distancias genéticas, interesa saber si hay suficientediferenciación adaptativa, y aislamiento reproductivo potencial o efectivo (Templeton1989, Crandall et al. 2000, Helbig et al. 2002). La aplicación general de reglas paradefinir especies basadas sólo en distancias genéticas llevaría a negar la existencia degéneros y especies jóvenes diferenciados rápidamente: inclusive con distancias críticasa nivel de D-loop tan bajas como 1-2%, habría que rebajar a "subespecies" simpátricasde una especie única a muchos géneros de pinzones de las Islas Galápagos y de cíclidoshaplocrominos de Africa oriental (Sato et al. 1999, Nagl et al. 2000).
En resumen, la invalidación de las especies reconocidas por Molina y Molinari(1999) valiéndose de una distancia genética "crítica" (Moscarella et al. 2003) es, ademásde cuestionable teóricamente, injustificable con base en lo que se sabe acerca de lavariabilidad genética interespecífica de los cérvidos.
Caracteres discretos craneomandibulares
Para justificar el uso de una distancia genética "crítica" como único criteriotaxonómico válido para los venados de cola blanca venezolanos, Moscarella et al.(2003) adujeron que Molina y Molinari (1999) emplearon datos "morfométricos"sujetos a gran plasticidad fenotípica, por ende irrelevantes para distinguir especies.Moscarella et al. (2003) obviaron que para los cérvidos en general y para Odocoileus

41Mem. Fund. La Salle de Cienc. Nat. 167
en particular es falso que la morfometría craneomandibular esté sujeta a unaplasticidad fenotípica tal que la haga inútil para estudios taxonómicos y filogenéticos(Rees 1969b, Hewison 1997, Meijaard y Groves 2004, Groves 2005), pasando por altoque: 1) las propuestas taxonómicas de Molina y Molinari (1999) no se fundamentaronen datos morfométricos (su análisis de medidas mandibulares tuvo el propósito auxiliarde comparar en tamaño y forma a los grupos ya diferenciados con base en caracteresno morfométricos); 2) los caracteres craneomandibulares con los que explícitamenteMolina y Molinari (1999) justificaron dichas propuestas son de naturaleza discreta ysimilares a los usados con gran éxito por otros autores para construir filogenias deartiodáctilos (Janis y Scott 1987, Gentry y Hooker 1988, Montgelard et al. 1988,Bouvrain et al. 1989, Scott y Janis 1993, Groves 2000, Vrba y Schaller 2000, Webb2000, Geisler 2001, Hassanin y Douzery 2003) y 3) en el caso particular de cérvidos,los estudios basados en caracteres morfológicos discretos como los usados por Molinay Molinari (1999) han llevado a filogenias altamente concordantes con las basadas enADN mitocondrial (Randi et al. 1998, 2001, Groves 2006).
Moscarella et al. (2003) también pasaron por alto que, en conjunto, los caracteresdiscretos empleados por Molina y Molinari (1999) resumen el efecto indirecto denumerosos loci genéticos, por lo cual conjuntamente podrían ser una medida másrepresentativa de diferenciación taxonómica que una corta secuencia mitocondrial, talcomo el D-loop empleado por ellos. Esta es seguramente la explicación de por quédichos caracteres separan a los venados de una manera biogeográficamente lógica(Figura 1).
Debido a que, para fines ta xonómicos, los cara c te res morfológicos sonn e c e s a ri a m e n te comp l e m e n ta rios y a menudo superi o res a los moleculares, loprudente es buscar aspectos coincidentes de ambos tipos de información (Sites yCrandall 1997, Puorto et al. 2001, Polziehn y Strobeck 2002, Zander 2003, Pitra et al.2004, Groves 2005). Aunque hay especies morfológicamente crípticas diagnosticablesgracias a discontinuidades genéticas, también hay especies genéticamente crípticasd i a g n o sticables gracias a discontinuidades morfológicas; incluso hay especiesdifícilmente diagnosticables tanto morfológica como genéticamente, pese a poseeraislamiento reproductivo total (Lee 2000, Mendelson y Shaw 2001).
Medidas mandibulares y masas corporales
La comparación de machos muestra que, la variación geográfica en tamañomandibular de los Odocoileus venezolanos (Figura 2) es considerable aunque entérminos absolutos menor que la de los venados de cola blanca norteamericanos. Seinfiere (Tabla 1) que el venado de Margarita rivaliza por el título de Odocoileusviviente más pequeño con el también diminuto venado de la panameña Isla de Coiba(para una descripción detallada de éste, ver Allen 1904) Por su parte, los venados de lacosta (Figura 2, Tabla 1), son la demostración de que es un error suponer que losvenados de cola blanca sura m e ricanos son siemp re más pequeños que losnorteamericanos (Baker 1984, Smith 1991). La figura 3 permite visualizar la magnitud

42 Variación geográfica en venados
de la diferenciación en tamaño y forma craneomandibular de estos dos Odocoileusvenezolanos extremos. Hay tres motivos por los cuales las diferencias no podríanobedecer a "plasticidad fenotípica," tal como ha sido propuesto (Moscarella et al. 2003):1) dicha plasticidad no debería producir animales tan diferentes en regiones costerastan similares climatológica y florísticamente (Isla de Margarita, La Guaira); 2) lasdiferencias tienden a ser constantes independientemente de si los ejemplares sonsilvestres o criados en cautiverio; y 3) la mejor explicación para el enanismo del venadode Margarita es que es resultado de convergencia adaptativa con otros unguladosinsulares pequeños (Dayan y Simberloff 1999), entre los cuales se incluyen variosOdocoileus (Allen 1904, Brisbin y Lenarz 1984, Maffei et al. 1988, Schaefer y Main1999).
En virtud de su diferenciación morfológica, el venado de Margarita (Tabla 1;Figuras 2, 3) ofrece la oportunidad de formular hipótesis acerca de qué podría sucedersi desapareciese la barrera marina que lo separa de sus congéneres de tierra firme.
En primer lugar, como en los demás ungulados, la masa corporal es determinantepara la segregación de nichos ecológicos en los cérvidos (Jenkins y Wright 1988,Kirchhoff y Larsen 1998, Latham et al. 1999, Domingues y Barbanti 2005). Losvenados de Margarita, por su menor tamaño, deben estar más limitados por la calidaddel alimento, mientras que los de la costa, por su mayor tamaño, principalmente porla cantidad del alimento: los primeros podrían aprovechar parches de follaje de buenacalidad, pero demasiado pequeños para los segundos, mientras que los segundospodrían aprovechar parches extensos de follaje de calidad insuficiente para losprimeros. Es decir, cabría esperar que los venados de Margarita y de la costa, porposeer grandes diferencias de tamaño corporal de origen genético (Figura 3), no seanecológicamente intercambiables, lo cual haría que ambos reuniesen un requisitofundamental para ser considerados especies diferentes (Templeton 1989, Crandall et al.2000).
En segundo lugar, en los cérvidos, la masa corporal y las dimensiones de las astasson decisivas en el éxito sexual de los machos (Clutton-Brock 1987, Markusson yFolstad 1997, Ditchko et al. 2001, Malo et al. 2005, Barto y Bahbouh 2006).Supongamos que las hembras de venado de Margarita aceptan machos de la costa, yque las hembras de venado de la costa aceptan machos de Margarita. En lacompetencia por las hembras de cualquiera de los dos orígenes, los machos de la costa,gracias a su superior tamaño, triunfarían casi siempre sobre los machos de Margarita,impidiéndoles copular. Además, los machos de Margarita tendrían dificultad paracopular con las hembras de la costa debido a que éstas les superan considerablementeen alzada. Esta desigualdad de oportunidades reproductivas haría que la mayoría delos híbridos de primera generación tuviesen un padre de la costa y una madre deMargarita. Si estos híbridos fuesen de tamaño corporal intermedio, podrían estarsujetos a mortalidades elevadas antes de nacer (por alcanzar durante su desarrollo fetalun tamaño excesivo para el sistema reproductor de sus madres) y antes del destete (poralcanzar durante su desarrollo postnatal un tamaño excesivo para mantenerse con la
43Mem. Fund. La Salle de Cienc. Nat. 167
leche de sus madres). La elevada mortalidad de los híbridos llevaría rápidamente aldesarrollo de comportamientos tendientes al aislamiento reproductivo de los dos tiposde venado.
Figura 2. Comparación del tamaño mandibular de 22 grupos de ejemplares adultos de Odocoileusvenezolanos (machos y hembras) y O. virginianus norteamericanos (machos) basadas en14 medidas (Rees 1969a, b; Molina y Molinari 1999 ). La escala representa los puntajes,expresados en desviaciones estándar, de dichos grupos en el primer eje de un Análisis deComponentes Principales.

44 Variación geográfica en venados
En conclusión, las diferencias en tamaño corporal y dimensiones cefálicasobservables entre los Odocoileus venezolanos más divergentes (Figura 3) hacen pensarque, independientemente de las distancias genéticas que los separan, en el presentehabría diferenciación ecológica y aislamiento reproductivo parcial si sus poblaciones sehiciesen simpátricas o parapátricas.
Patrones de variación geográfica
En los venados de cola blanca norteamericanos, el elevado y significativo valor dela correlación (r= 0,67) con la distancia geográfica demuestra que la frecuencia decaracteres discretos craneomandibulares varía de manera fuertemente clinal (Tabla 2).En los venados de cola blanca venezolanos, el casi nulo y no significativo valor de lamisma correlación (r= 0,13) indica que dicha frecuencia, pese a varíar de modo másabrupto que en Norteamérica (8,21 contra 3,74 croizats), no lo hace de manera clinal(Tabla 2). Es necesario notar que el bajo valor de la correlación correspondiente a losvenados venezolanos no podría atribuirse a falta de diferenciación geográfica, pues envariación absoluta (Tabla 2) superan a los venados norteamericanos (3,21 contra 2,95DEE).
Las ideas anteriores se ven reforzadas al considerar los datos morfométricos man-dibulares (Tabla 2) que, por su distinta naturaleza, son una fuente de informaciónindependiente. En los venados norteamericanos, el signo positivo de las correlacionesentre diferenciación morfométrica y distancia geográfica (tamaño, r= 0,39; forma, r=0,16), indica una variación más continua, pues las mayores diferencias aparecen entregrupos geográficos alejados; y la baja variación relativa en tamaño (0,85 croizats) yforma (1,03 croizats) indica una variación más gradual. En contraste, en los venadosvenezolanos, el signo negativo de las correlaciones entre diferenciación morfométricay distancia geográfica (tamaño, r= -0,33; forma, r= -0,19) indica una variación másdiscontinua, pues las mayores diferencias aparecen entre grupos geográficos vecinos; yla elevada variación relativa en tamaño (3,97 croizats) y forma (6,40 croizats) indicauna variación más abrupta. En conjunto, la variación geográfica más continua y gra-dual de los venados norteamericanos apoya la idea de un patrón clinal, mientras quela variación geográfica más discontinua y abrupta de los venados venezolanos apoyala idea contraria.
Al compararlos con sus parientes de las zonas templadas, los vertebrados tropicalesposeen mayores divergencias genéticas intralinaje y cladogénesis más antiguas (p. ej.Da Silva y Patton 1998, Culver et al. 2000, Moritz et al. 2000, Chek et al. 2003,Crawford 2003, Lessa et al. 2003, Martin y McKay 2004). Ello podría deberse a queel menor efecto de los períodos glaciares ha permitido que los procesos locales dediferenciación evolutiva hayan sido más constantes y prolongados en los trópicos(Hewitt 1996, Milá et al. 2000, Lessa et al. 2003). La mayor y menos ordenadavariabilidad geográfica de los venados de cola blanca venezolanos con respecto a losn o rte a m e ricanos (Tabla 2) parece aj u sta rse a este patrón de especiación másproductiva en los trópicos.

45Mem. Fund. La Salle de Cienc. Nat. 167
Es una paradoja que la taxonomía tradicional reconozca 17 subespecies devenados de cola blanca en Estados Unidos y sólo ocho en una área similar enSuramérica (Smith 1991). Lo lógico sería que los taxa suramericanos fuesen másnumerosos y en promedio de mayor jerarquía que los norteamericanos (Figura 1;Tabla 2). Como en otros organismos (Chek et al. 2003), esta paradoja puede explicarsepor el pobre conocimiento biológico que se tiene de los venados de cola blancatropicales, pese a su gran importancia económica (Eisenberg 2000, Weber y González2003). Las estadísticas de México (Mandujano 2004) ilustran la situación: de 276estudios revisados, 189 se centraron en las cinco subespecies de venados de cola blancaneárticas (carminis, couesi, miquihuanensis, texanus, mexicanus) y sólo 76 en las diezsubespecies neot ropicales (a c a p u l c e n s i s, n e l s o n i, o a xa c e n s i s, s i n a l o a e, th o m a s i,toltecus, truei, veraecrucis, yucatanensis). Por supuesto, los estudios sobre los venadosde Centro y Suramérica son mucho menos numerosos y sólo una pequeña fracción deellos ha tocado aspectos taxonómicos.
Figura 3. Vista lateral de los cráneos y denticiones inferiores del holotipo de Odocoileus margaritae(Field Museum of Natural History, Chicago) y de un ejemplar de O. cariacou (U. S.National Museum of Natural History, Washington). El ejemplar de margaritae, unmacho adulto joven, fue colectado cerca de Puerto Viejo (Paraguachoa, Estado NuevaEsparta) en 1910. El ejemplar de cariacou, un macho subadulto, fue colectado cerca deLa Guaira (Estado Vargas) en 1892. Si ambos ejemplares fuesen de la misma edad, ladiferencia de tamaño sería mayor. Dibujos por Sharlene Santana.

46 Variación geográfica en venados
El género Odocoileus todavía no ha sido objeto de una investigación sistemáticaencaminada a unificar los criterios taxonómicos aplicados a las formas de Norte,Centro y Suramérica. Los análisis aquí presentados (Tabla 2) permiten confiar en queuna investigación de esta naturaleza confirmará la mayor y más compleja variabilidadge o grá fica de las fo rmas neot ropicales, apoyando y ampliando las pro p u e sta staxonómicas de Molina y Molinari (1999).
Escenarios paleobiogeográficos: venado de páramo
Excluyendo al venado de alta montaña de la Sierra Nevada de Santa Marta,nominalmente una variedad de Mazama americana (Alberico et al. 2000), hay tresvenados de páramo (Odocoileus) en los andes del norte (Molina y Molinari 1999):lasiotis, cordillera de Mérida en Venezuela; goudotti, cordillera oriental de Colombia;y ustus, cordilleras de Ecuador y extremo sur de Colombia, quizás Perú al norte de ladepresión de Huancabamba. Además, en Perú y Bolivia habita peruvianus, que en lavertiente pacífica no está restringido a alta montaña, pues en los andes centrales, desdela puna hasta la costa, hay continuidad de vegetación abierta adecuada paraOdocoileus.
Cabrera (1961) reconoció a goudotii, ustus y peruvianus, pero sinonimizó a lasiotiscon goudotii debido a que creía que la cordillera de Mérida era sólo una extensión, dealturas continuas, de la cordillera oriental de Colombia (Cabrera 1918). Sin embargo,la f lora y la fauna de los páramos de ambas cordilleras siempre han estado aislados,incluso durante los máximos glaciares, por la depresión del Táchira, una de las máshondas de todos los andes, formada por un corredor de valles de 60 km de anchura y960 m de elevación (Van der Hammen y Cleef 1986). Tomando en cuenta este hechobiogeográfico, Molina y Molinari (1999) revivieron a lasiotis como una formaindependiente de goudotii.
Moscarella et al. (2003) mantuvieron a lasiotis como sinónimo de goudotii. Elloequivale a suponer que ambos venados poseen los mismos caracteres taxonómicos porser recíprocamente monofiléticos. Según estos autores "goudotti" (=goudotii + lasiotis)se diferenció tras iniciarse el Holoceno hace 13000 años, al quedar aislado de losvenados de tierras bajas (O. cariacou, bajo el nombre O. v. gymnotis), debido alascenso altitudinal de la vegetación de páramo. El descenso altitudinal de la vegetaciónde páramo dura n te la última glaciación hizo a los ante c e s o res de "g o u d ot t i"panmícticos con sus congéneres de tierras bajas hasta hace apenas 19000 - 16000 años.
El componente holocénico del escenario de Moscarella et al. (2003), es decir ladiferenciación de una única subespecie de venado de cola blanca en las alturas de lascordilleras de Mérida y oriental de Colombia, implica que la subespecie "goudotii," enuna emigración asincrónica con la del resto de los taxa vege tales y animalescompartidos, tendría que haberse movido de los páramos de una cordillera a los de laotra habiendo ya alcanzado estos ambientes su gran separación actual, que es lamáxima cíclica (Van der Hammen y Cleef 1986). Dicha emigración tendría ademásque haberse logrado contradiciendo la noción de que "goudotii" y "gymnotis" son sólo

47Mem. Fund. La Salle de Cienc. Nat. 167
subespecies de O. virginianus (Moscarella et al. 2003), pues los "goudotii" participantestendrían que haber atravesado la depresión del Táchira sin perder su identidadsubespecífica por entrecruzamiento con los "gymnotis" presentes en el camino.
El componente preholocénico del escenario de Moscarella et al. (2003), es decir lapanmixia entre las poblaciones antecesoras de los venados de páramo y tierras bajas,requiere que, durante la última glaciación, la barrera ecológica que actualmentesepara a estos venados, a saber el "cinturón" de selvas húmedas andinas, se hayareducido y fragmentado enormemente, y que el descenso altitudinal de la vegetaciónde páramo la haya hecho entrar en amplio contacto con la vegetación abierta detierras bajas. Sin embargo, la información paleobotánica no apoya estas nociones.Aunque en el último máximo glaciar la vegetación de páramo descendió de los 4800-3300 m s.n.m. actuales hasta los 3500-2000 m s.n.m., los "cinturones" selváticosandinos no desaparecieron sino que simplemente descendieron de los 3300-1000 ms.n.m. actuales hasta los 2000-800 m s.n.m. (Van der Hammen 1979, Van derHammen y Cleef 1986, Hooghiemstra y Cleef 1995, Hooghiemstra y Van derHammen 2004). Además, por varios ciclos glaciares, el ecotono páramo-selva nubladaha estado por debajo de los 2000 m sólo 5-10% del tiempo (Van der Hammen y Cleef1986), haciendo infrecuentes las oportunidades de introgresión entre venados depáramo y tierras bajas.
En consecuencia, la información paleobotánica en realidad sugiere que las coloni-zaciones de ambientes de páramo por las poblaciones ancestrales de lasiotis, goudotiiy ustus fueron simultáneas e independientes, y que estos tres venados se mantuvieroncompletamente aislados unos de otros y casi completamente aislados de los de tierrasbajas durante varios periodos glaciares, incluyendo el último. Por fortuna, hay pruebasfósiles y arquelógicas que confirman el origen preholocénico de los venados depáramo: 1) en el Ecuador, u st u s fue conte mp o ráneo de la fo rma de tierras bajas "O. salinae"( p ro b a b l e m e n te un sinónimo de O. cari a c o u) al iniciarse el Holoceno (Hoff stet ter 19 5 2 ,1986, Tomiati y Abbazzi 2002); 2) en la altiplanicie de Bogotá (2600 m de eleva c i ó n ) ,g o u d ot i i d i s m i n u yó mucho su imp o rtancia en la dieta de los indígenas cuando el ascensoholocénico del bosque andino hizo desaparecer localmente la ve getación de pára m o( C o rreal 19 9 0 ) .
C o n s i d e rando lo ante ri o r, es oportuno pre s e n tar un nuevo escenario para explicar elo ri gen de los venados de páramo, que a dife rencia del pro p u e sto por Moscarella et al. ( 2 0 0 3 ) ,tome en cuenta la info rmación paleobioge o grá fica disponible. Siguiendo la cronología deM c Kenna y Bell (19 97), este nuevo escenario consta de las etapas siguientes:
A.- Mioceno tardío, hace 5,3-11,2 millones de años: aparición de los Odocoileinae,subfamilia de los Cervidae a la que pertenece Odocoileus, en el registro fósil deEuropa y Asia (McKenna y Bell 1997).
B.- Plioceno te mp rano, hace 5 millones de años: aparición del pri m e rOdocoileinae en Norteamérica (Webb 2000).
C.- Inicio del Plioceno tardío, hace unos 3 millones de años: aparición del género

48 Variación geográfica en venados
Odocoileus en el registro fósil de Norteamérica (Kurtén y Anderson 1980,Webb 2000).
D.- Final del Plioceno tardío, hace unos 2 millones de años: aparición de O.virginianus en el registro fósil de Norteamérica (Kurtén y Anderson 1980).Según Webb (1991, 2000), la diversificación de varios grupos de mamíferossuramericanos de origen neártico, entre ellos los cérvidos, se inició a bajaslatitudes en Norteamérica antes de que se materializara la colonización deSuramérica. En concordancia con esta idea, puede suponerse que en esta etapa,en lo que ahora es el sur de México, debe haber tenido lugar la diferenciaciónde O. cariacou (adaptación a climas cálidos) a partir del ancestro común conO. virginianus y O. hemionus.
E.- Pleistoceno, hace unos 0,5-1,7 millones de años: colonización de Centroamé-rica por Odocoileus (McKenna y Bell 1997), representado por O. cariacou.
F.- Pleistoceno medio, hace unos 0,7-1,2 millones de años: migración de O .cariacou a través del Istmo de Panamá hasta Suramérica (Marshall 1985, Webb1985). Previamente a esta etapa, los andes del norte y las tierras bajas deColombia y Venezuela ya habían adquirido sus configuraciones actuales (Díazde Gamero 1996, Gregory-Wodzicki 2000).
G.- Pleistoceno medio, hace 0,35-1 millón de años: colonización de los ambientesde páramo por O. cariacou. Durante esta etapa, las selvas nubladas de los andesdel norte eran mucho menos diversas que las actuales en formas arbóreas deorigen austral y neártico, por ende preadaptadas a climas fríos, mientras que lasformas arbóreas predominantes, originadas a partir de vegetación de las tierrasbajas circundantes, no estaban todavía bien adaptadas al clima de alta mon-taña. Dada esta composición taxonómica y adaptativamente más pobre, lasselvas nubladas de la etapa tenían un límite altitudinal más bajo, eran menosdensas y en general constituían un tipo de vegetación menos definido que lasactuales (Gentry 1995, Hooghiemstra y Cleef 1995, Webster 1995, Marchant etal. 2002). Por ello, estas selvas fueron una barrera en buena medida franquea-ble, particularmente durante las glaciaciones, para los O. cariacou que,procedentes de tierras bajas, emigraron a los páramos.
H.- Pleistoceno tardío a presente, últimos 350000 años: diferenciación definitiva deO. lasiotis, O. goudotii y O. ustus a partir de las poblaciones de O. cariacou queanteriormente habían colonizado los ambientes de páramo. Durante estaetapa, las selvas nubladas de los andes del norte adquirieron su fisonomíaactual, enriqueciéndose con especies arbóreas de orígenes austral, neártico yautóctono bien adaptadas al clima de alta montaña. Además del registrop a l e o b otánico, como prueba del re c i e n te ori gen de estas selvas, puedemencionarse que la mayoría de sus endemismos vegetales son sólo a nivel deespecie (Gentry 1995, Hooghiemstra y Cleef 1995, Webster 1995, Marchant etal. 2002). El gran desarrollo y la mejor adaptación al frío de las selvas nubladasdurante esta etapa las hizo constituirse en un "cinturón" de vegetación ancho,

49Mem. Fund. La Salle de Cienc. Nat. 167
denso y permanente que desplazó hacia arriba el límite altitudinal inferior delos páramos. Por consiguiente, durante esta etapa, las selvas nubladas no sóloalejaron a los venados de páramo de sus congéneres de tierras bajas, sino quese constituyeron en una barrera difícil de franquear para estos animales, inclusodurante las glaciaciones (cuatro en los últimos 350000 años; Van der Hammeny Hooghiemstra 1997), consolidando así los tres procesos de especiación queindependientemente condujeron a lasiotis, goudotii y ustus.
Lo más parsimonioso es suponer que la progresión que finalmente llevó a laaparición y diversificación de los Odocoileus suramericanos fue de norte a sur, y unavez en Suramérica, de bajas a altas elevaciones. Hershkovitz (1972) creía que losvenados de cola blanca forman una sola especie originaria de América Central, desdedonde se habría dispersado para colonizar Norteamérica y Suramérica. Su escenariocontradice las etapas "D" y "E", e implica el improbable reemplazo de venados bienadaptados a climas templados (O. virginianus) por una forma adaptada a climastropicales (O. cariacou). Tomiati y Abbazzi (2002) postularon que ustus dio origen auna especie de tierras bajas (salinae), lo cual contradice las etapas "G" y "H". Estasautoras no estaban conscientes de que salinae probablemente sea sólo un sinónimo decariacou. Los paleontólogos han sido capaces de diferenciar morfológicamente yreconocer como especies diferentes a venados de páramo y tierras bajas (Hoffstetter1952, 1986, Tomiati y Abbazzi 2002).
Escenarios paleobiogeográficos: venado de Margarita
Desde los puntos de vista tectónico, litosférico y paleoclimático, la regiónnororiental de Venezuela, de la cual fo rma parte la Isla de Marga ri ta, se ha cara c te ri z a d opor una histo ria sumamente intrincada y dinámica, con numerosos y dramáticos cambiosre c i e n tes (More l o ck et al. 1972, Macsotay 1977, Sch u b e rt 1982, Stö ck h e rt et al. 19 94 ,1995, Clay ton et al. 1999, Dean et al. 1999, Pete rson et al. 2000, Weber et al. 2 0 01, Pipery Dean 2002). Moscarella et al. (2003) pro p u s i e ron un escenario para explicar el ori ge ndel venado de Marga ri ta basado sólo en el aspecto paleoclimático. Según este escenari o ,d u ra n te el último máximo gl a c i a r, debido a la unión entre la Isla de Marga ri ta y lapenínsula de Araya ocasionada por el descenso del nivel del mar en unos 120 m (Ru l l1998), la población ante c e s o ra del venado marga ri teño (bajo el nombre O. v. marga ri ta e)debe haber sido continua y panmíctica con las de los O d o c o i l e u s venezolanos de tierra sb ajas continentales (O. cari a c o u, bajo el nombre O. v. gymnot i s)hace apenas19 0 0 0 - 16 0 0 0años. Por ello, m a rga ri ta e se habría di fe renciado dura n te el Holoceno, que comenzó h a c e13000 años, tras quedar aislado en la Isla de Marga ri ta al re gresar el mar a un nive lsimilar al actual. Este escenario supone que la batimetría entre Marga ri ta y Araya ,a c t u a l m e n te cara c te rizada por el casi total predominio de profundidades infe ri o res a5 0 - 100 m, no ha va riado substa n c i a l m e n te desde hace 19000 años.
Debido a que el venado de Margarita es cranealmente el más distintivo de losOdocoileus venezolanos (Tabla 1; Figuras 1, 2, 3), la propuesta de Moscarella et al.

50 Variación geográfica en venados
(2003) para Margarita implica una evolución morfológica inusitadamente rápida.Tomando como referencia el bien fundamentado escenario para los andes presentadoen la sección anterior, la diferenciación de "O. v. margaritae" tendría que habersecompletado en un tiempo casi 30 veces más corto (13000 años) que la del venado depáramo (350000 años). La improbabilidad de semejante diferencia en tasas deevolución invita a proponer un nuevo escenario paleobiogeográfico que, aceptando lanoción de una conexión Margarita-Araya durante el último máximo glaciar, atribuyala notable diferenciación morfológica del venado de Margarita a un aislamiento igualo más antiguo que el del venado de páramo. Este nuevo escenario, que toma en cuentala evidencia tectónica y litosférica ignorada por Moscarella et al. (2003), consta de lasetapas siguientes:
A-F.- Las mismas del escenario propuesto para el venado de páramo.G.- Pleistoceno medio, hace 0,4-1 millón de años: colonización de la Isla de
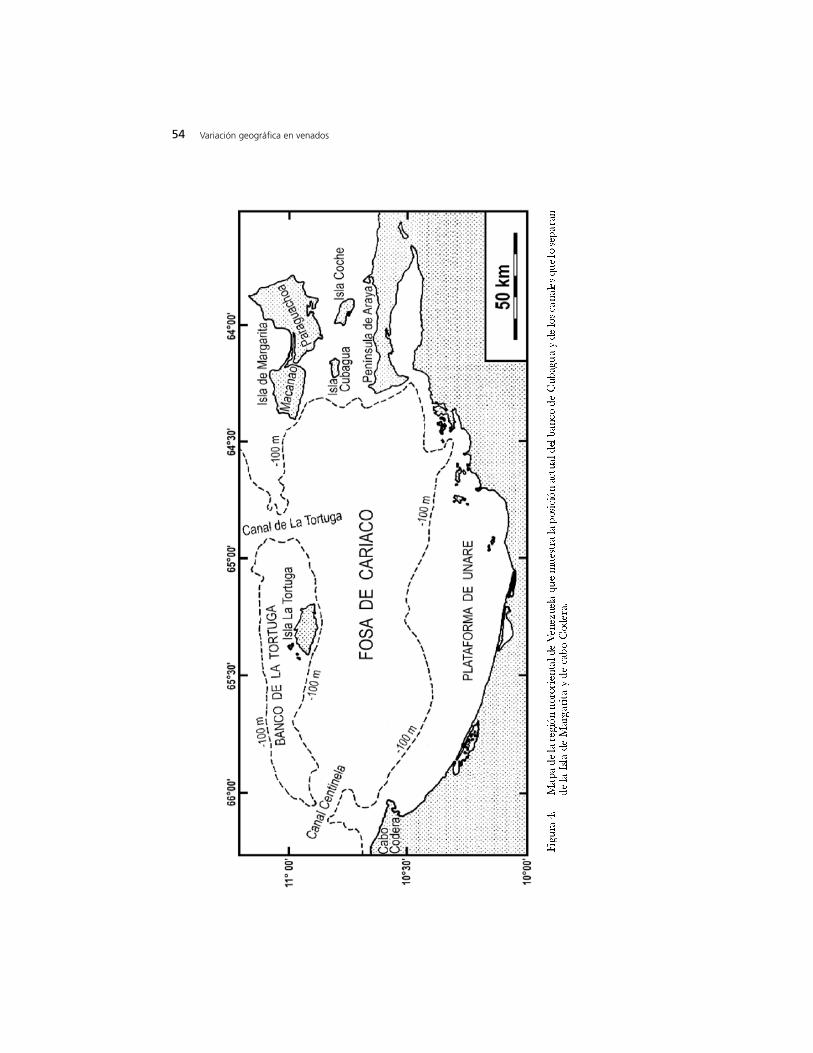
Margarita por O. cariacou ayudado por el descenso del nivel del mar duranteun máximo glaciar. Esta colonización podría haberse producido desde el oeste,partiendo de lo que ahora es el cabo Codera y recorriendo el primitivo bancode Cubagua, hasta llegar a Macanao y posteriormente a Paraguachoa (Figura4). Margarita está formada por dos penínsulas, Paraguachoa (o MargaritaOriental) y Macanao, unidas por un istmo de relieve muy bajo, formado poruna larga y estrecha barra de arena y una albufera con manglares (Figura 4).Esta isla, conjuntamente con la mitad norte de las penínsulas de Araya y Paria,forma parte del borde sur de la placa tectónica del Caribe (Stöckhert et al. 1994,1995). A su vez Margarita, en unión al banco y la isla de La Tortuga, forma unamontaña parcialmente submarina que se extiende hasta cabo Codera (Maloney1966, Figura 4). Suponiendo para la placa del Caribe una velocidad dedesplazamiento hacia el este de ~20 mm/año relativamente a la placa deSuramérica (Weber et al. 2001), Margarita debe haber estado unos 20 km másal oeste hace un millón de años, y unos 8 km más al oeste hace 400000 millonesde años (hay estimados más extremos que permitirían ubicar a Margarita 50km al oeste de su posición actual hace un millón de años; Schubert 1982,Clayton et al. 1999). Si el nivel del mar descendiese 120 m hoy, lo único queimpediría la formación de un puente de tierra entre Margarita y cabo Codera,pasando por el banco de La Tortuga (Figura 4), serían los canales de LaTo rt u ga, cuya profundidad máxima alcanza 135 m, y Centinela, cuyaprofundidad máxima alcanza 146 m (Goñi et al. 2003). Sin embargo, hace 0,4-1 millón de años, no sólo estaba Margarita más próxima a cabo Codera, sinoque la fosa de Cariaco, que tiene una antigüedad de unos dos millones de años,apenas había alcanzado 50-75% de su extensión y profundidad actuales(Schubert 1982), y la isla de La Tortuga y su banco se encontraban más al sur(Maloney y Macsotay 1967). Debido a esta antigua configuración geográfica,los canales La Tortuga y Centinela, si acaso existían, debían ser poco

51Mem. Fund. La Salle de Cienc. Nat. 167
profundos. Por ello, durante las glaciaciones, la ausencia o incipiencia de estoscanales debe haber permitido que hubiera una cre sta monta ñ o s acontinuamente emergida desde cabo Codera hasta Margarita. Desde hace 5millones de años hasta antes del inicio del último máximo glaciar, esta crestadebe haber sido la única conexión de Margarita con tierra firme, pues el maral sur y sureste de la isla era demasiado profundo para permitir la formación depuentes de tierra hacia Araya durante las glaciaciones (Macsotay 1977, Claytonet al. 1999, Aguilera y Rodrigues 2001). La vegetación de esta cresta, cuandoestaba emergida, seguramente se parecía a la de las actuales penínsulas deMacanao y Araya, lo cual podría haber facilitado el poblamiento por losvenados que eventualmente originaron a margaritae.
H.- Pleistoceno tardío hasta bien entrado el último máximo glaciar, hace 17000-400000 millones de años: diferenciación en aislamiento de O. margaritae apartir de las poblaciones de O. cariacou que anteriormente habían colonizadoMargarita. Durante esta etapa no hubo tierras emergidas que conectaran aMargarita con cabo Codera ni con Araya. Por una parte, a medida queMargarita y el banco de La Tortuga se iban acercando a sus posiciones actualesy la fosa de Cariaco se iba expandiendo (Maloney y Macsotay 1967, Schubert1982, Stöckhert et al. 1994, 1995), debe haber llegado un momento en el cuallos canales Centinela y de La Tortuga se formaron y adquirieron suficienteprofundidad para mantener permanentemente desconectada a Macanao decabo Codera. Por la otra, el proceso de reducción de la profundidad entreParaguachoa y Araya, que se fue produciendo a lo largo de la migración deMargarita hacia el este (Macsotay 1977, Clayton et al. 1999), no había avanzadolo suficiente para permitir la formación de puentes de tierra durante losmáximos glaciares. Como prueba de la gran profundidad que llegaron a tenerlas aguas entre Paraguachoa y Araya, puede mencionarse que, durante elMioceno tardío y Plioceno temprano, es decir hace unos 5 millones de añoscuando Margarita debía encontrarse unos 100 km más al oeste, estas aguasestaban habitadas por una rica ictiofauna mesopelágica que dejó fósiles en lossedimentos actualmente emergidos (Formación Cubagua) de Araya, Cubagua yel sur de Paraguachoa (Aguilera y Rodrigues 2001).
I.- Final del último máximo glaciar, hace 17000-13000 años: formación de unistmo entre Paraguachoa y la península de Araya que permitió el contacto entreO. margaritae y O. cariacou. Durante esta breve etapa coincidieron dos facto-res que sinérgicamente propiciaron por primera vez la aparición de dichoistmo. En primer lugar, durante la mayor parte del Pleistoceno superior, hace16000-600000 años, la región formada por Margarita y Araya sufrió unaelevación tectónica permanente, la cual, además de reducir directamente laprofundidad del mar entre Paraguachoa y Araya, también la redujo indirec-tamente al crear ambientes terrestres elevados que por erosión enviabanaluvión al mar (Macsotay 1977). En segundo lugar, el advenimiento del máximo

52 Variación geográfica en venados
glaciar, además de hacer descender el nivel del mar hasta 120 m, dio lugar a unclima árido que propició suelos muy erosionables por la lluvia y el viento,particularmente en laderas. Las lluvias, aunque menos frecuentes que en laactualidad, deben haber sido torrenciales debido al clima árido, arrastrandovastas cantidades de sedimentos hacia el mar. Resumiendo, durante esta breveetapa, surgió un istmo entre Paraguachoa y Margarita porque coincidieron:una gran elevación del lecho marino debida a largos procesos tectónicos ysedimentarios previos; un descenso eustático del nivel del mar; y una elevaciónadicional del lecho marino durante el máximo glaciar debida a una actividadsedimentaria incrementada por la aridez climática (Morelock et al. 1972,Macsotay 1977, Clayton et al. 1999, Dean et al. 1999, Peterson et al. 2000, Pipery Dean 2002). La capa de sedimentos entre Margarita y Araya pudiera tener ungrosor de cientos de metros (Schubert 1982), habiendo sufrido los depósitos másre c i e n tes una imp o rta n te re d i st ribución desde el último máximo gl a c i a r(Morelock et al. 1972). Por este motivo, no es fácil determinar con base en labatimetría actual la forma y amplitud del istmo Paraguachoa-Araya (enrealidad, para ello sería mucho más útil estudiar la probable hidrografía localdurante el último máximo glaciar). Lo que sí se puede decir es que, dada lanaturaleza de los sedimentos submarinos actuales (Morelock et al. 1972) y elclima de la etapa (Clayton et al. 1999, Peterson et al. 2000, Piper y Dean 2002),este istmo debe haber sido una planicie árida, interrumpida por dunas, consuelos salinos de textura gruesa y vegetación muy pobre. Por su baja densidaddebida a estas condiciones ambientales, independientemente del escenario queimaginemos, la población de venados del istmo podría no haber tenido lacapacidad de actuar como un intermediario importante para el flujo genéticoentre Paraguachoa y Araya. Es decir, para margaritae Margarita siempre fueuna isla, ecológicamente durante el último máximo glaciar y geográficamenteel resto del tiempo.
J.- Holoceno, últimos 13000 años: supervivencia de O. margaritae como especiedistinta, pese al breve contacto con O. cariacou durante la etapa anterior.Según lo recién planteado, el venado de Margarita (Figura 3) descendería de lasmismas poblaciones ancestrales que originaron el venado de la costa central deVenezuela (Figura 3). La colonización de Margarita durante el último máximoglaciar por venados de Araya debe haber sido difícil porque ello implicabacompetir con margaritae, cuyo diminuto tamaño corporal es óptimo para unambiente insular, y cuya densidad poblacional debía ser la máxima quepermitía la capacidad de carga de la isla. Los venados de Araya deben haberdejado una huella genética pequeña en Margarita debido a selección en contrade genotipos que determinan su mayor tamaño corporal. En contraste, esprobable que haya habido un flujo genético algo mayor de Margarita a Araya,tal como lo sugiere, si es que no hay un error en la localidad, la secuencia de unvenado de Araya que permitió agruparlo con margaritae (Moscarella et al.

53Mem. Fund. La Salle de Cienc. Nat. 167
2003). El extremo occidental de la península de Araya es ecológicamente casiinsular, lo cual podría dar algunas ventajas a los genotipos heredados demargaritae. Con independencia de si hubo o no introgresión, la sobrevivenciade margaritae como una forma morfológicamente constante y bien diferen-ciada de cariacou, pese al contacto entre ambos durante el último máximoglaciar, prueba que, contrariamente a lo propuesto por Moscarella et al. (2003),se trata de dos especies diferentes.
El escenario para Margarita recién propuesto permite hacer dos predicciones. Laprimera es que algún día será posible encontrar en esta isla fósiles preglaciares demargaritae. La segunda es que la fauna autóctona de la isla está conformada porespecies morfológicamente bien diferenciadas que descienden de inmigrantes llegadosdesde cabo Codera hace cientos de miles de años (p. ej. O. margaritae), y por especiesmorfológicamente poco diferenciadas que descienden de inmigrantes llegados desdeAraya durante el último máximo glaciar (p. ej. el murciélago Pteronotus parnelliifuscus, Gutiérrez 2004).
Tasas de diferenciación
Los escenarios recién propuestos para Los Andes y Margarita son consistentes conla noción cada vez más generalizada de que los procesos de especiación de losvertebrados tropicales típicamente son muy anteriores al último máximo glaciar (DaSilva y Patton 1998, Moritz et al. 2000, Colinvaux et al. 2000, 2001, Chek et al. 2003,Crawford 2003, Lessa et al. 2003, Martin y McKay 2004). Sin embargo, ¿son lostiempos de diferenciación inferidos a partir de distancias genéticas consistentes con losescenarios recién propuestos para los venados de páramo y Margarita? La respuesta esafirmativa. ¿Son estos tiempos consistentes con los escenarios de Moscarella et al.(2003) para dichos venados? La respuesta es negativa.
Con base en sus propias secuencias de ADN mitocondrial (D-loop) de venadosvenezolanos, y pese a escoger como referencia una tasa de mutación alta (5,1% de paresde bases cada 1 millón de años, suponiendo que Odocoileus y Mazama divergieronhace sólo 1,5 millones años), Moscarella (2001) estimó que la separación del venado depáramo con respecto a sus congéneres de tierras bajas sucedió hace "por lo menos"177000 años, y que la separación del venado de Margarita con respecto a suscongéneres de tierra firme sucedió hace "por lo menos" 118000 años. Ambos estimadosmínimos son muy acordes con los escenarios recién propuestos.
Moscarella et al. (2003) no mencionaron tasas de diferenciación. Usar comoreferencia el tiempo de divergencia entre Odocoileus y Mazama (Moscarella 2001)habría negado la propuesta diferenciación de lasiotis y margaritae en sólo 13000 años,la cual llevaría al cálculo de tasas de evolución mitocondrial para los venados de colablanca venezolanos un orden de magnitud mayores que las referidas para cérvidos yotros mamíferos (Brown et al. 1979, Douzery y Randi 1997, Pesole et. al. 2000, Pitra etal. 2004). Por ello, puede concluirse que el marco temporal de los escenariospaleobiogeográficos de Moscarella et al. (2003) es completamente arbitrario.
54 Variación geográfica en venados

55Mem. Fund. La Salle de Cienc. Nat. 167
Conclusiones taxonómicas: uso del nombre "O. virginianus"
La evidencia (presencia/ausencia de la glándula metatarsal) que sirvió para reunira todos los venados de cola blanca en una sola especie es errónea (Molina y Molinari1999). En contraste, la evidencia que sirve para rechazar la conespecificidad de lasformas neotropicales con las neárticas es muy sólida, pudiendo resumirse como sigue:
Moléculas. A nivel aloenzimático (Baccus et al. 1983, Hillestad 1984, Smith et al.1986), la distancia genética entre los venados de cola blanca de Surinam yNorteamérica (D= 0,173) es casi ocho veces mayor que la distancia promedio entresubespecies de venados de cola blanca norteamericanos (D= 0,023) y algo mayor quela distancia entre estos últimos y el ciervo mulo, O. hemionus (D = 0,149). Smith et al.(1986) concluyeron que estas diferencias podrían indicar que los venados de colablanca surinameses y norteamericanos pertenecen a especies diferentes. A nivel de laregión de control del ADN mitocondrial (Polziehn y Strobeck 1998, Moscarella et al.2003), las distancias genéticas entre los venados de cola blanca venezolanos ynorteamericanos (5,6-7,7%) son similares a las que existen entre estos últimos y O.hemionus (7,82%). En los árboles filogenéticos de Moscarella et al. (2003), los venadosde cola blanca norteamericanos se agruparon con el ciervo mulo y no con suscongéneres venezolanos.
Cráneo. En caracteres continuos (Smith et al. 1986), los venados de cola blanca deSurinam y Norteamérica presentan diferencias notables y constantes, teniendo los deSurinam mayor similitud con el venado matacán, Mazama americana. En caracteresdiscretos (Molina y Molinari 1999, Figura 1), los venados de cola blanca de Venezuelay Norteamérica forman dos grupos netamente separados. Las astas de los venados decola blanca neotropicales están menos desarrolladas que las de los neárticos (Baker1984, Villarreal 2002).
Tegumento. Los venados de cola blanca del sur de México y Centroamérica, comolos de Suramérica, se distinguen de los venados de cola blanca neárticos por: el escasoo nulo desarrollo de la glándula metatarsal (Méndez-Arocha 1955, Hershkovitz 1958,Quay 1971, Brokx 1972, Baker 1984); tener colas más cortas que exponen menosblanco cuando son levantadas (Reid 1997); y la ausencia de pelajes estacionalmentediferentes (Baker 1984, Brokx 1984). Los venados de cola blanca neotropicalescomparten las tres características con el también neotropical género Mazama. Losvenados de cola blanca neárticos comparten la primera y tercera característica, y partede la segunda (longitud de la cola) con el ciervo mulo.
Reproducción. Los venados de cola blanca neotropicales producen una cría porparto (Eisenberg 2000). Los venados de cola blanca neárticos y el ciervo muloproducen dos (DeYoung et al. 2002).
La información molecular y craneana claramente indica que hay dos grupose s p e c í f i c a m e n te dife re n tes de venados de cola blanca, uno neártico y ot ro

56 Variación geográfica en venados
suramericano. La información tegumentaria, aunada a la lógica biogeográfica (Ortegay Arita 1998, Morrone y Márquez 2001, Escalante et al. 2004), indica que los venadosde cola blanca del sur de México y Centroamérica pertenecen al grupo suramericano,constituyendo junto con ellos un gran grupo neotropical. La información molecular,tegumentaria y reproductiva sugiere que el venado de cola blanca neártico forma ungrupo con O. hemionus y no con los venados de cola blanca neotropicales, lo cualtambién es bioge o grá fi c a m e n te lógico. Por consiguiente, en su conjunto, lainformación disponible sugiere que Molina y Molinari (1999) estuvieron en lo correctocuando agruparon a todos los venados neotropicales no insulares y de tierras bajas enuna especie plena, O. cariacou (Boddaert 1784), reservando el nombre O. virginianus(Zimmermann 1780) exclusivamente para los venados de cola blanca neárticos.
El uso dado por Molina y Molinari (1999) al nombre O. virginianus dejó sindefinir un problema muy importante, que algún día deberá ser resuelto de maneradefinitiva en México. Este problema consiste en determinar los límites precisos de lasdistribuciones de O. virginianus y O. cariacou, y en determinar si hay alguna intro-gresión entre ambas, tal como sucede con O. virginianus y O. hemionus más al norte(Carr et al. 1986). Al respecto, hay que señalar que los cazadores deportivos reconocendos categorías de venados de cola blanca mexicanos, una compuesta por cincosubespecies más corpulentas y con astas más grandes, por ello muy apreciadas comotrofeos de caza, y otra compuesta por nueve subespecies de menor tamaño y con astasmás pequeñas, por ello de escaso interés cinegético (Villarreal 2002). Lo interesante delasunto es que las subespecies de la primera categoría están distribuidas en la parteneártica de México, por lo cual seguramente son asignables a O. virginianus, mientrasque las subespecies de la segunda categoría están distribuidas en la parte neotropicalde este país, por lo cual seguramente son asignables a O. cariacou. En consecuencia,mientras no se realicen estudios que permitan modificar este arreglo, puede propo-nerse provisionalmente que las subespecies de la primera categoría, a saber: carminis,couesi, miquihuanensis, texanus y quizás mexicanus, sean reunidas bajo el nombreespecífico O. virginianus, y que las subespecies pertenecientes a la segunda categoría,a saber: acapulcensis, nelsoni, oaxacensis, sinaloae, thomasi, toltecus, truei, veraecrucisy yucatanensis, sean reunidas bajo el nombre específico O. cariacou. Puesto queMéxico es el único país en el cual cabe esperar la presencia de venados de cola blancaneárticos y neotropicales, este arreglo puede verse como un paso inicial para definir loslímites de distribución entre O. virginianus y O. cariacou.
En taxonomía existe lo que, con una gran dosis de ironía, se ha dado en llamar elconcepto de "especie inercial", que se usa para describir aquellos casos en los cuales laspoblaciones de dos o más buenas especies se siguen tratando como una sola,independientemente de la información disponible, simplemente porque los zoólogosestán acostumbrados a la idea (Good 1994, Zamudio y Green 1997). Lamenta-blemente, los venados de cola blanca neotropicales han sido víctimas de este concepto,que se traduce en una gran resistencia a separarlos de O. virginianus.

57Mem. Fund. La Salle de Cienc. Nat. 167
Conclusiones taxonómicas: Odocoileus venezolanos
Recapitulando, el no reconocimiento de las formas de los páramos y Margaritacomo especies plenas por Moscarella et al. (2003), en mi opinión resulta de seis errores1) utilizaron muestras de origen poco fiable; 2) en contradicción con la literatura,adujeron que en los cérvidos las distancias genéticas mitocondriales entre subespeciestípicamente alcanzan 3%; 3) pese a que no existen distancias genéticas "críticas" parad i s c e rnir si las poblaciones animales tienen estatus específico o subespecífi c o ,concluyeron que todos los Odocoileus venezolanos son conespecíficos, utilizandocomo único argumento que la distancia genética máxima entre ellos no supera el 3%;4) alegaron falsamente que las conclusiones taxonómicas de Molina y Molinari (1999)se fundamentaron en datos morfométricos y que en los cérvidos los datos de este tiposon irre l eva n tes para fines ta xonómicos debido a plasticidad fe n otípica; 5)desconocieron que los caracteres discretos craneomandibulares en los cuales Molina yMolinari (1999) sí fundamentaron sus conclusiones taxonómicas son similares a losusados con éxito por otros autores para elucidar filogenias de cérvidos y otrosartiodáctilos; y 6) para dar más credibilidad a la idea de que los venados de páramo yM a rga ri ta son sólo "subespecies" poco dife renciadas, pro p u s i e ron escenari o s ,justificados sólo con dos referencias inespecíficas (Vuilleumier 1971, Rull 1998), segúnlos cuales sus diferenciaciones se habrían iniciado hace sólo 13000 años, pese a que lasdistancias genéticas por ellos mismos calculadas sugieren que se trata de procesosmucho más antigüos.
También recapitulando, los nuevos análisis aquí pre s e n tados confi rman yrefuerzan las conclusiones taxonómicas de Molina y Molinari (1999) mediante: 1) lademostración de que la variación geográfica de los Odocoileus venezolanos difiere dela de O. virginianus, no sólo por ser de mayor magnitud, sino también por tener uncarácter no clinal sugerente de especiación más avanzada; 2) la formulación de unp rimer escenario paleobioge o grá fico que, en concordancia con info rm a c i ó nmorfológica, genética, paleontológica y arquelógica, permite concluir que O. lasiotisha estado separado de O. cariacou al menos unos 350000 años; 3) la formulación deun segundo escenario paleobiogeográfico que, en concordancia con informaciónmorfológica, genética y tectónica, permite concluir que O. margaritae ha estadoseparado de O. cariacou unos 400000 años, manteniendo intacta su identidadtaxonómica pese a un breve contacto entre ambos (aislamiento reproductivo yecológico efectivo) hace 17000-13000 años; 4) la generación de estimados de masascorporales, que considerados conjuntamente con las dimensiones cefálicas, permiteninferir que en la actualidad, bajo condiciones naturales, habría dificultad para laintrogresión (aislamiento reproductivo y ecológico potencial) entre O. margaritae y O.cariacou.
Para concluir, es oportuno presentar algunas consideraciones sistemáticas ytaxonómicas sobre las tres especies de venados de cola blanca presentes en Venezuela:

58 Variación geográfica en venados
Odocoileus cariacou
Según Moscarella et al. (2003), cariacou (bajo el nombre O. v. gymnotis) es untaxón polifilético. Lo preciso es decir que cariacou es un taxón parafilético conrespecto a lasiotis y margaritae, con independencia si optamos por considerar a los trestaxa como especies o subespecies. La especiación suele ser un proceso de gemaciónparafilética cuya realidad hay que aceptar (Avise y Wollenberg 1997). Por sus diferen-cias morfológicas, ecológicas y evolutivas cariacou, lasiotis y margaritae merecen, pesea la parafilia del primero, ser considerados especies plenas.
Adaptando la taxonomía tradicional de los Odocoileus de Suramérica (Cabrera1961, Brokx 1984, Smith 1991) al reconocimiento de cariacou, lasiotis y margaritae anivel de especie, se tiene que O. cariacou gymnotis sería el único venado de cola blancapresente en Venezuela fuera del de los páramos de Mérida y de la Isla de Margarita(es de notar que O. goudotii puede estar o haber estado presente en el páramo delTamá, estados Táchira y Apure). Pese a tener el estatus de subespecie única, los O.cariacou gymnotis venezolanos son geográficamente más variables que el conjunto deocho subespecies de O. virginianus consideradas en la figura 1. Este hecho indica quese requieren estudios que investiguen la variación subespecífica de O. cariacouprácticamente desconocida, en toda su área de distribución, que incluye Méxiconeotropical, Centroamérica y tierras bajas de Suramérica al norte y este de Los Andescentrales. Dichos estudios seguramente determinarán que el número de subespecies deO. cariacou es mucho mayor que el actualmente reconocido, y que algunas subespe-cies insulares, en especial curassavicus (Curaçao) y rothschildi (Coiba), deben elevarsea especie. En Venezuela, los venados de la costa y el de Apure son claros candidatos aser reconocidos como subespecies nuevas (Brokx 1984, Molina y Molinari 1999).
Odocoileus lasiotis
La distribución de este venado está restringida a los páramos más elevados de lasierras Nevada y de La Culata, en la parte central de la cordillera de Mérida, en dondeademás se le ha encontrado en restos arquelógicos a 4100 m de altura (Armand 1985).No hay información que permita comprobar si alguna vez la distribución abarcó otrospáramos de esta cordillera.
Seguir considerando a lasiotis sinónimo de goudotii (Moscarella et al. 2003) es unerror biogeográfico ya que ignora la depresión del Táchira. La sinonimización de estosvenados (Cabrera 1961) se basa en una opinión dubitativa del propio Cabrera (1918),quien después de examinar el pelaje de un ejemplar taxidermizado de goudotiiexhibido en Madrid y de leer la descripción del pelaje del holotipo de lasiotis (nuncavio material de esta especie) hecha por Osgood (1914), escribió: "El que no puedodiferenciar es el Odocoileus lasiotis... juzgando por la descripción original de Osgood,si no es lo mismo que columbicus [= goudotii] debe parecérsele mucho... nada deextraño tendría que columbicus llegase por Los Andes de Colombia hasta las

59Mem. Fund. La Salle de Cienc. Nat. 167
montañas del noroeste de Venezuela". Cabrera ilustró con un dibujo las astas y lacabeza del ejemplar de goudotii, pero no examinó su cráneo. El holotipo de goudotiiconsta únicamente de una asta pequeña (Cabrera 1918).
Actualmente, el material de museo de goudotii sigue siendo escaso. Recien-temente, pude examinar los cráneos de un macho (ICN 16161; Figura 5) del páramode Chingaza (Cundinamarca) y una hembra (Inderena 1213, Instituto Alexander vonHumboldt, Colombia) de los páramos del noroeste de Sochaviejo (Boyacá), y la piel deesta hembra y de su cría subadulta. Para fines comparativos, también consideré eldibujo de la cabeza y la descripción del pelaje de un macho de goudotii (Cabrera1918); fotografías de un macho adulto silvestre del páramo de Chingaza; 12 ejemplaresde museo de lasiotis, incluyendo el holotipo (Figura 5, fotografías de la piel), los usadospor Molina y Molinari (1999), y otro recientemente adquirido por la Universidad deLos Andes; observaciones, incluyendo fotografías y videos, de numerosos ejemplaressilvestres de lasiotis; cráneos y pieles de ustus (FMNH 47049, 47050), peruvianus(FMNH 19548, 35219, 35220, 52504, 78421, 78422, 80977, 80978, 80979) y cariacou(muchos de varios museos); y fotografías de peruvianus cautivos.
Figura 5. Vista lateral del cráneo y dentición inferior del holotipo de Odocoileus lasiotis (FieldMuseum of Natural History, Chicago), y del cráneo de un ejemplar de O. goudotii(Instituto de Ciencias Naturales, Universidad Nacional de Colombia, Bogotá). Elejemplar de lasiotis, un macho adulto, fue obtenido en el páramo de los Conejos (EstadoMérida) en 1912. El ejemplar de goudotii, un macho adulto, fue obtenido en el páramode Chingaza (Departamento de Cundinamarca) en 1998. Debe notarse que al cráneo deeste ejemplar le falta, además de las mandíbulas, parte del basicráneo. Dibujos por RogerPuente.

60 Variación geográfica en venados
Las diferencias craneanas entre lasiotis y goudotii son múltiples y saltan a la vista(Figura 5). El rasgo más llamativo de goudotii, apreciable interna o externamente entodos los ejemplares (ICN 16161; Inderena 1213; cabeza ilustrada por Cabrera 1918,macho silvestre del páramo de Chingaza), es un pronunciado hundimiento de la regiónfrontal, justo antes del comienzo del rostro, seguido por una abrupta elevación de lacaja craneana. Esta peculiaridad (Figura 5), ausente en lasiotis, ustus, peruvianus ycariacou, se aprecia también, aunque menos marcadamente, en margaritae (Figura 3),habiendo sido considerada por Osgood (1910) como una característica muy notable deesta especie.
Las diferencias externas también saltan a la vista. El pelaje de lasiotis es muy largoy suelto, al punto de que los ejemplares pueden lucir despeinados a causa del viento oel roce de la vegetación, y tiene una coloración general que podría describirse comogris obscura, equiparable a la de los peruvianus más obscuros, con intensas tonalidadespardas. El pelaje de goudotii, aunque mucho más espeso y largo que el de los muyamarillentos o rojizos venados colombianos de tierras bajas, es claramente más cortoy compacto que el de lasiotis, con una coloración general que podría describirse comogris clara, equiparable a la de ustus, con tonalidades ocráceas.
La gran diferenciación morfológica de lasiotis y goudotii es prueba de que estosvenados, tal como cabe esperar debido a la depresión del Táchira, siempre han sidoalopátricos. Por ello, seguir aplicando el nombre "goudotii" al venado de páramo deMérida (Moscarella et al. 2003) es taxonómica y filogenéticamente incorrecto.
Odocoileus margaritae
El diminuto tamaño corporal (Tabla 1), la divergencia en caracteres craneanos(Figura 1), la forma cefálica, las proporciones dentarias (Figura 3), y la aparienciaexterna (Figura 6), hacen a margaritae un venado bien diferenciado de sus congéneresde tierra f irme. Los rasgos distintivos son muy constantes en todos los ejemplaresdisponibles, independientemente de si proceden de Macanao o Paraguachoa.
Debido al contacto de Margarita con tierra firme durante el último máximoglaciar, cabe esperar que, en las poblaciones vivientes de venados de Margarita(margaritae) y Araya (cariacou), hayan huellas de introgresión mutua ocurrida hace17000-13000 años. La introgresión entre especies congenéricas que han entrado encontacto recientemente o durante el Pleistoceno es un fenómeno bien conocido encérvidos y otros mamíferos (Carr et al. 1986, Groves y Grubb 1987, Goodman et al.1999, Alves et al. 2003, Good et al. 2003, Pitra et al. 2004, Schwartz et al. 2004). Porello, la confirmación de niveles bajos de introgresión entre estos venados durante elúltimo máximo glaciar no podría tomarse como prueba de conespecificidad entremargaritae y caricou, sino más bien lo contrario.
Implicaciones para la conservación
Desde el punto de vista de su conservación, los venados de páramo y Margarita

61Mem. Fund. La Salle de Cienc. Nat. 167
tienen tres tipos de enemigo real o potencial. El primero es la ignorancia taxonómica,el segundo es la caza furtiva y la destrucción de sus hábitat, el tercero lo representanlos animales domésticos y exóticos.
La carencia de buena información taxonómica tiene consecuencias funestas parauna especie en peligro: si su estatus taxonómico es poco conocido o erróneo, la especiepuede no ser objeto de un plan de manejo adecuado, e incluso corre el riesgo ded e s a p a recer comp l eta m e n te sin que se le pre ste atención (Bri to 2004). Losinvestigadores que evalúan el estatus taxonómico de poblaciones animales en peligrollevan una responsabilidad enorme sobre sus hombros, pues la sobrevaloración dedicho estatus puede conducir al desperdicio de recursos disponibles para esfuerzos deconservación, mientras que su subvaloración puede contribuir a que se materialice laextinción. Los venados de páramo y Margarita son especies en peligro crítico que hanrecibido atención de muy pocos estudios taxonómicos (Osgood 1910, 1914, Méndez-Arocha 1955, Molina y Molinari 1999, Moscarella et al. 2003). Lamentablemente,Méndez-Arocha (1955) y Moscarella et al. (2003) subestimaron su diferenciacióntaxonómica. De Méndez-Arocha (1955), a quien puede disculparse por la menorrigurosidad de los métodos de trabajo y la casi nula disponibilidad de material demuseo de su época, puede recordarse que, refiriéndose al venado de Margarita,escribió "tenemos que decir que sus diferencias con los venados de tierra f irmeadyacente son de bien poca importancia, lo cual, incluso, pudiera explicarse por laposibilidad de tráfico de estos animales en cautiverio que muchas veces se realiza". Dehaber examinado Méndez-Arocha los cráneos (Figura 3), seguramente no habríahecho semejante aseveración. Es de esperarse que los datos y argumentos aquípresentados despejen las dudas sembradas por Méndez-Arocha (1955) y Moscarella etal. (2003), contribuyendo así a justificar mejor planes de conservación para los venadosde páramo y Margarita.
La caza furtiva y la destrucción del hábitat son factores negativos de crecienteimportancia que en un futuro cercano podrían atentar contra la supervivencia delvenado de páramo. Ello se debe a la creciente colonización agrícola de ambientes dealta montaña dentro de los parques nacionales Sierra Nevada y Sierra de La Culata.La caza furtiva es la causa de mortalidad más importante para el venado deMargarita. Esta caza es realizada principalmente por algunas familias rurales deMacanao y por los dueños de conucos en Paraguachoa, que se valen para ello dearmas de fuego artesanales y, especialmente, de trampas de lazo (con las cualestambién capturan otros mamíferos). Además, participan en dicha actividad algunosresidentes de Paraguachoa que, equipados con escopetas modernas, se trasladan aMacanao para tal fin. La destrucción del hábitat no es un factor negativo importantepara el venado en Macanao, pero sí lo es en Paraguachoa, debido al crecientedesarrollo urbano y turístico. Las crías de las hembras de venado de Margaritacobradas por los cazadores a menudo terminan como mascotas (Figura 6).
Existe competencia por alimento entre el venado de páramo y los ganados bovinoy equino, aunque éste es el menos importante de los factores negativos para dicho

62 Variación geográfica en venados
venado (M. Molina, com. pers.). Lo que si se conoce bien es que los perros asilvestradosy domésticos son la mayor causa de mortalidad para estos venados, habiendodepredado en cuatro años el equivalente a un quinto de la mayor población actual(menos de 250 individuos) de esta especie que se encuentra en el páramo de Mucubají(Molina y Peñaloza 2002). El ganado es escaso en Margarita, por lo cual actualmenteno es un factor relevante para la supervivencia del venado de la isla. Hay que prestarespecial atención a un eventual desarrollo de la ganadería caprina, por los efectosdestructivos que tendría sobre la cobertura vegetal y la probable competencia con elvenado insular. Existe un importante tráfico de venados de tierra firme hacia la isla(Moscarella 2001, obs. pers.). Incluso algunas familias de Paraguachoa mantienen encautiverio rebaños de estos animales que oscilan entre seis y dos docenas de cabezas.En caso de ser liberados, estos venados "exóticos" representarían un gran peligro parael de Margarita. No hay que olvidar que, debido a la caza furtiva, las poblaciones delvenado nativo de la isla tienen densidades muy inferiores a las que permitirían losambientes naturales en los que sobrevive, lo cual podría dar la oportunidad a losvenados introducidos de establecerse y multiplicarse.
Los venados de páramo y Margarita son especies únicas e irreemplazables. Paralos pocos que han tenido la oportunidad de observarlos, el primero destaca por suhermoso pelaje, que le da una apariencia inconfundible, mientras que el segundo
Figura 6. Odocoileus margaritae en cautiverio. Izquierda, vista completa de una hembra que semantenía cautiva en la península de Macanao en 1994, posteriormente sacrificada porsu dueña para aprovechar su carne. Derecha, detalle de la cabeza de una hembra que semantenía cautiva en la península de Macanao en 2004.

63Mem. Fund. La Salle de Cienc. Nat. 167
destaca por su sorprendente enanismo. Evidentemente, la principal medida a tomarpara salvar de la extinción a estos venados es protegerlos en su ambiente naturalcontrarrestando los factores de mortalidad arriba mencionados. Dados los bajísimosniveles de las poblaciones de ambos, probablemente ha llegado la hora de la cría encautiverio con la idea de obtener ejemplares para repoblación. Con el propósito deasegurar la conservación de la mayor diversidad genética posible, sería ideal que losejemplares fundadores de las poblaciones cautivas procediesen, en el caso del venadode páramo, tanto del páramo de Mucubají como de otros páramos, y en el caso delvenado de Margarita, tanto de Macanao como de Paraguachoa.
Agra d e c i m i e n t o s . El Consejo de Desarrollo Científico Humanístico y Te c n o l ó g i c o(Universidad de Los Andes) y la International Foundation for Science (Estocolmo) aportaronequipo y apoyo financiero a las salidas de campo. Linda Gordon, Bruce Patterson, DaveS chmidt y Paul Velazco env i a ron foto grafías de ejemp l a res de venados ve n e z o l a n o s ,colombianos, ecuatorianos y peruanos depositados en museos de los Estados Unidos. ÁlvaroGarcía Olaechea (Perú) y John Poveda Martínez (Colombia) enviaron fotografías de venadosvivos de sus respectivos países. Leida Valero dibujó el mapa. Eliécer Gutiérrez, Hernán López yJafet Nassar aportaron bibliografía.
Bibliografía.
AGUILERA, O. Y D. RODRIGUES. 2001. An exceptional coastal upwelling fish assemblage in theCaribbean Neogene. Journal of Paleontology 75: 732-742.
ALBERICO, M., A. CADENA, J. HERNÁNDEZ-CAMACHO Y Y. MUÑOZ-SABA. 2000. Mamíferos(Synapsida: Theria) de Colombia. Biota Colombiana 1: 43-75.
ALVES, P. C., N. FERRAND, F. SUCHENTRUNK Y D. J. HARRIS. 2003. Ancient introgression ofLepus timidus mtDNA into L. granatensis and L. europaeus in the Iberian Peninsula.Molecular Phylogenetics and Evolution 27: 70-80.
ALLEN, J. A. 1904. Mammals from southern Mexico and Central and South America. Bulletinof the American Museum of Natural History 20: 29-80.
ARMAND, J. 1985. La Maneta - vestigios esqueléticos de un niño del siglo XVII con marcas deposible antropofagia. Consejo de Publicaciones de la Universidad de Los Andes, Mérida,Venezuela.
AVISE, J. C. Y K. WOLLENBERG. 1997. Phylogenetics and the origin of species. Proceedings of theNational Academy of Sciences of the USA 94: 7748-7755.
AVISE, J. C. Y R. M. BALL. 1990. Principles of genealogical concordance in species concepts andbiological taxonomy. Oxford Survey of Evolutionary Biology 7: 45-67.
BACCUS, R., N. RYMAN, M. H. SMITH, C. REUTERWALL Y D. CAMERON. 1983. Geneticvariability and differentiation of large grazing mammals. Journal of Mammalogy 64: 109-120.
BAKER, R. H. 1984. Origin, classification and distribution. Pp. 1-18. En: L. K. Halls (Ed.),White-tailed deer - ecology and management. Stackpole Books, Harrisburg, Pennsylvania,USA.
BARTO, L. Y R. BAHBOUH. 2006. Antler size and fluctuating asymmetry in red deer (Cervus

64 Variación geográfica en venados
elaphus) stags and probability of becoming a harem holder in rut. Biological Journal of theLinnean Society 87: 59-68.
BOOKSTEIN, F. L., B. C. CHERNOFF, R. L. ELDER, J. M. HUMPHRIES, G. R. SMITH Y R. E.STRAUSS. 1985. Morphometrics in evolutionary biology: the geometry of size and shapechange, with examples from fishes. Academy of Natural Sciences of Philadelphia, SpecialPublication 15: 1-277.
BOUVRAIN G., D. GERAADS Y Y. JEHENNE. 1989. Nouvelles données relatives à la classificationdes Cervidae (Artiodactyla, Mammalia). Zoologischer Anzeiger 223: 82-90.
BRADLEY, R. D., F. C. BRYANT, L. C. BRADLEY, M. L. HAYNIE Y R. J. BAKER. 2003. Implicationsof hybridization in white-tailed deer and mule deer. Southwestern Naturalist 48: 654-660.
BRISBIN, I. L. Y M. S. LENARZ. 1984. Morphological comparisons of insular and mainlandpopulations of southeastern white-tailed deer. Journal of Mammalogy 65: 44-50.
BRITO, D. 2004. Lack of adequate taxonomic knowledge may hinder endemic mammalconservation in the Brazilian Atlantic Forest. Biodiversity and Conservation 13: 2135-2144.
BROKX, P. A. 1972. A study of the biology of the Venezuelan white-tailed deer (Odocoileusvirginianus gymnotis Wiegmann, 1833), with a hypothesis on the origin of South Americancervids. Tesis Doctoral, University of Waterloo, Ontario, Canada. 355 pp.
BROKX, P. A. 1984. South America. Pp. 525-546. En: L. K. Halls (Ed.), White-tailed deer -ecology and management. Stackpole Books, Harrisburg, Pennsylvania, USA.
BROWN, W. M., M. J. GEORGE Y A. C. WILSON. 1979. Rapid evolution of animal mitochondrialDNA. Proceedings of the National Academy of Sciences of the USA 76: 1967-1971.
CABRERA, A. 1918. Sobre los Odocoileus de Colombia. Boletín de la Real Sociedad Española deHistoria Natural 18: 300-307.
CABRERA, A. 1961. Catálogo de los mamíferos de América del Sur. Revista del Museo Argentinode Ciencias Naturales "Bernardino Rivadavia" 4: 309-732.
CABRERA, A. Y J. YEPES. 1960. Mamíferos Sud Americanos. Tomo II, Ediar S.A. Editores,Buenos Aires, Argentina. 160 pp.
CARR, S. M., S. W. BALLINGER, J. N. DERR, L. H. BLANKENSHIP Y J. W. BICKHAM. 1986.Mitochondrial DNA analysis of hybridization between white-tailed deer and mule deer inwest Texas. Proceedings of the National Academy of Sciences of the USA 83: 9576-9580.
CARTELLE, C. 1994. Tempo passado. Mamíferos do Pleistoceno em Minas Gerais. EditoraPalco, ACESITA [Assessoria de Comunicação da Compania Aços Especiais Itabira], BeloHorizonte, Brasil. 131 pp.
CLAYTON, T., R. B. PEARCE Y L. C. PETERSON. 1999. Indirect climatic control of the claymineral composition of Quaternary sediments from the Cariaco basin, northern Venezuela(ODP Site 1002). Marine Geology 161: 191-206.
CLUTTON-BROCK, T. H. 1987. Sexual selection in the Cervidae. Pp. 110-122. En: C. Wemmer,M. (Ed.), Biology and management of the Cerv i d a e. Smithsonian Institution Pre s s ,Washington, USA.
COLINVAUX, P. A., G. IRION, M. E. RÄSÄNEM, M. B. BUSH Y J. A. S. NUNES DE MELLO. 2001.A paradigm to be discarded: geological and paleoecological data falsify the Haffer andPrance refuge hypothesis of Amazonian speciation. Amazoniana 16: 609-646.
COLINVAUX, P. A., P. E. DE OLIVEIRA Y M. B. BUSH. 2000. Amazonian and neotropical plantcommunities on glacial time-scales: the failure of the aridity and refuge hypotheses.Quaternary Science Reviews 19: 141-169.
CORREAL, G. 1990. Evidencias culturales durante el Pleistoceno y Holoceno de Colombia.Revista de Arqueología Americana 1: 69-89.

65Mem. Fund. La Salle de Cienc. Nat. 167
CRANDALL, K. A., O. R. P. BININDA-EMONDS, G. M. MACE Y R. K. WAYNE. 2000. Consideringevolutionary processes in conservation biology. Trends in Ecology and Evolution 15: 290-295.
CRAWFORD, A. J. 2003. Huge populations and old species of Costa Rican and Panamanian dirtfrogs inferred from mitochondrial and nuclear gene sequences. Molecular Ecology 12: 2525-2540.
CRONIN, M. A. 1992. Intraspecific variation in mitochondrial DNA of North American cervids.Journal of Mammalogy 73:70-82.
CULVER, M., W. E. JOHNSON, J. PECON-SLATERY Y S. J. O'BRIEN. 2000. Genomic ancestry ofthe American puma (Puma concolor). Journal of Heredity 91: 186-197.
CHEK, A. A., J. D. AUSTIN Y S. C. LOUGHEED. 2003. Why is there a tropical-temperate disparityin the genetic diversity and taxonomy of species?. Evolutionary Ecology Research 5: 69-77.
DA SILVA, M. N. Y J. L. PATTON. 1998. Molecular phylogeography and the evolution andconservation of Amazonian mammals. Molecular Ecology 7: 475-486.
DAYAN, T. Y D. SIMBERLOFF. 1999. Size patterns among competitors: ecological characterdisplacement and character release in mammals, with special reference to island populations.Mammal Review 28: 99-124.
DE MARINIS, A. M., H. NIKOLOV Y S. GERASIMOV. 1990. Sex identification and sexualdimorphism in the skull of the stone marten, Martes foina (Carnivora, Mustelidae). Hystrix2: 34-46.
DEAN, W. E., D. Z. PIPER Y L. C. PETERSON. 1999. Molybdenum accumulation in Cariaco Basinsediment over the past 24 k.y.; a record of water-column anoxia and climate. Geology 27:507-510.
DEYOUNG, R. W., S. DEMARAIS, R. A. GONZÁLES, R. L. HONEYCUTT Y K L. GEE. 2002.Multiple pate rnity in white - tailed deer (Odocoileus virg i n i a n u s) revealed by DNAmicrosatellites. Journal of Mammalogy 83: 884-892.
DÍAZ DE GAMERO, M. L. 1996. The changing course of the Orinoco River during the Neogene:a review. Palaeogeography, Palaeoclimatology, Palaeoecology 123: 385-402.
DITCHKO, S. S., R. L. LOCHMILLER, R. E. MASTERS, W. R. STARRY Y D. M. LESLIE, JR. 2001.Does f luctuating asymmetry of antlers in white-tailed deer (Odocoileus virginianus) followpatterns predicted for sexually selected traits. Proceedings of The Royal Society of London,Series B, Biological Sciences 268: 891-898.
DOMINGUES, L. Y J. M. BARBANTI. 2005. Gastro-Intestinal transit time in South American deer.Zoo Biology 25: 47-57.
DOUZERY, E. Y E. RANDI. 1997. The mitochondrial control region of Cervidae: evolutionarypatterns and phylogenetic content. Molecular Biology and Evolution 14: 1154-1166.
EISENBERG, J. F. 2000. The contemporary Cervidae of Central and South America. Pp. 189-202. En: E. S. VRBA Y G. B. SCHALLER (Eds.), Antelopes, deer, and relatives-fossil record,behavioral ecology, systematics, and conservation. Yale University Press, New Haven, USA.
ESCALANTE, T., G. RODRÍGUEZ Y J. J. MORRONE. 2004. The diversification of Nearcticmammals in the Mexican transition zone. Biological Journal of the Linnean Society 83: 327-339.
GEISLER, J. H. 2001. New morphological evidence for the phylogeny Artiodactyla, Cetacea, andMesonychidae. American Museum Novitates 3344: 1-53.
GENTRY, A. H. 1995. Patterns of diversity and floristic composition in neotropical montaneforests. Pp. 103-126. En: S. P. Churchill, H. Balslev, E. Forero y J. L. Luteyn (Eds.),Biodiversity and conservation of netropical montane forests. New York Botanical Garden,Nueva York.
GENTRY, A. W. Y J. J. HOOKER. 1988. The phylogeny of Artiodactyla. Pp. 235-271. En: M. J.

66 Variación geográfica en venados
Benton (Ed.), The phylogeny and classification of the tetrapods, Mammals. Volume 2.Clarendon Press, Oxford, Inglaterra.
GIAO, P. M., D. TUOC, V. V. DUNG, E. D. WIKRAMANAYAKE, G. AMATO, P. ARCTANDER Y J. R.MACKINNON. 1998. Description of Muntiacus truongsonensis, a new species of muntjac(Artiodactyla: Muntiacidae) from Central Vietnam, and implications for conservation.Animal Conservation 1: 61-68.
GINGERICH, P. D. 1993. Quantification and comparison of evolutionary rates. American Journalof Science 293A: 453-478.
GONZÁLEZ, S., J. E. MALDONADO, J. A. LEONARD, C. VILÀ, J. M. BARBANTI, M. MERINO, N.BRUM-ZORILLA Y R. K. WAYNE. 1998. Conservation genetics of the endangered Pampas deer(Ozotoceros bezoarticus). Molecular Ecology 7: 47-56.
GOÑI, M. A., H. L. ACEVES, R. C. THUNELL, E. TAPPA, D. BLACK, Y. ASTOR, R. VARELA Y F.MULLER-KARGER. 2003. Biogenic fluxes in the Cariaco Basin: a combined study of sinkingparticulates and underlying sediments. Deep-Sea Research I 50: 781-807.
GO O D, D. A. 19 94 . Species limits in the genus G e rrh o n ot u s ( S qu a m a ta: Anguidae).Herpetological Monographs 8: 180-202.
GOOD, J. M., J. R. DEMBOSKI, D. W. NAGORSEN Y J. SULLIVAN. 2003. Phylogeography andintrogressive hybridization: chipmunks (genus Tamias) in the northern Rocky Mountains.Evolution 57: 1900-1916.
GOODMAN, S. J., N. H. BARTON, G. SWANSON, K. ABERNETHY Y J. M. PEMBERTON. 1999.Introgression through rare hybridization: a genetic study of a hybrid zone between red andsika deer (Genus Cervus) in Argyll, Scotland. Genetics 152: 355-371.
GREGORY-WODZICKI, K. M. 2000. Uplift history of the Central and Northern Andes: a review.Geological Society of America Bulletin 112: 1091-1105.
GROVES, C. P. 2000. Phylogenetic relationships within recent Antilopini (Bovidae). Pp. 223-233.En: E. S. Vrba y G. B. Schaller (Eds.), Antelopes, deer, and relatives - fossil record, behavioralecology, systematics, and conservation. Yale University Press, New Haven, USA.
GROVES, C. P. 2005. The genus Cervus in eastern Eurasia. European Journal of WildlifeResearch 52: 14-22.
GROVES, C. P. Y P. GRUBB. 1987. Relationships of living deer. Pp. 21-59. En: C. M. Wemmer(Ed.), Biology and management of the Cervidae. Smithsonian Institution Press, Washington,USA.
GUTIÉRREZ, E. 2004. Morfometría de los murciélagos de la familia Mormoopidae en Venezuela.Tesis de Licenciatura, Universidad de Los Andes, Mérida, Venezuela. 206 pp.
HALDANE, J. B. S. 1949. Suggestions as to the quantitative measurement of rates of evolution.Evolution 3: 51-56.
HASSANIN, A. Y E. J. P. DOUZERY. 2003. Molecular and morphological phylogenies ofruminantia and the alternative position of the Moschidae. Systematic Biology 52: 206-228.
HELBIG, A. J., A. G. KNOX, D. T. PARKIN, G. SANGSTER Y M. COLLINSON. 2002. Guidelines forassigning species rank. Ibis 144: 518-525.
HERSHKOVITZ, P. 1948. The technical name of the Virginia deer. Proceedings of the BiologicalSociety of Washington 61: 41-48.
HERSHKOVITZ, P. 1958. The metatarsal glands in white-tailed deer and related forms of theNeotropical region. Mammalia 22: 537-546.
HERSHKOVITZ, P. 1972. The recent mammals of the Neotropical Region: a zoogeographic andecological review. Pp. 311-431. En: A. Keast, F. C. Erk y B. Glass (Eds.), Evolution, mammals,and southern continents. State University of New York Press, Albany, Nueva York, USA.
HEWISON, A. J. M. 1997. Evidence for a genetic component of female fecundity in British RoeDeer from studies of cranial morphometrics. Functional ecology 11: 508-517.
HEWITT, G. M. 1996. Some genetic consequences of ice ages and their role in divergence and

67Mem. Fund. La Salle de Cienc. Nat. 167
speciation. Biological Journal of the Linnean Society 58: 247-276.HILLESTAD, H. O. 1984. Stocking and genetic variability of white-tailed deer in the southeastern
United States. Tesis Doctoral, Universidad de Georgia, Athens, Georgia, USA. 121 pp.HOFFSTETTER, R. 1952. Les mammifères pléistocènes de la République de l'Equateur. Mémoires
de la Société Géologique de France (Nouvelle Série) 31: 1-391.HOFFSTETTER, R. 1986. High Andean mammalian faunas during the Plio-Pleistocene. Pp. 218-
245. En: F. Vuilleumier y M. Monasterio (Eds.), High altitude tropical biogeography. OxfordUniversity Press, Nueva York, USA.
HOOGHIEMSTRA, H. Y A. M. CLEEF. 1995. Pleistocene climatic change and environmental andgeneric dynamics in the north Andean montane forest and páramo. Pp. 35-49. En: S. P.Churchill, H. Balslev, E. Forero y J. L. Luteyn (Eds.), Biodiversity and conservation ofnetropical montane forests. New York Botanical Garden, Nueva York.
HOOGHIEMSTRA, H. Y T. VAN DER HAMMEN. 2004. Quaternary Ice-Age dynamics in theColombian Andes: developing an understanding of our legacy. Philosophical Transactions:Biological Sciences 359: 173-181.
HUNDERTMARK, K. J., G. F. SHIELDS R. T. BOWYER Y C. C. SCHWARTZ. 2002a. Geneticrelationships deduced from cytochrome-b sequences among moose. Alces 38: 113-122.
HUNDERTMARK, K. J., G. F. SHIELDS, I. G. UDINA, R. T. BOWYER, A. A. DANILKIN Y C. C.SCHWARTZ. 2002b. Mitochondrial phylogeography of moose (Alces alces): Late Pleistocenedivergence and population expansion. Molecular Phylogenetics and Evolution 22: 375-387.
HUNDERTMARK, K. J., R. T. BOWYER, G. F. SHIELDS Y C. C. SCHWARTZ. 2003. Mitochondrialphylogeography of moose (Alces alces) in North America. Journal of Mammalogy 84: 718-728.
JANIS, C. M. Y K. M. SCOTT. 1987. The interrelationships of higher ruminant families withspecial emphasis on the members of the Cervoidea. American Museum Novitates 2893: 1-85.
JENKINS, K. J. Y R. G. WRIGHT. 1988. Resource partitioning and competition among cervids inthe northern Rocky Mountains. Journal of Applied Ecology 25: 11-24.
JOLICOEUR, P. Y J. E. MOSIMANN. 1960. Size and shape variation in the painted turtle: aprincipal component analysis. Growth 24: 339-354.
KATTAN, G. H., P. FRANCO, V. ROJAS Y G. MORALES. 2004. Biological diversification in acomplex region: a spatial analysis of faunistic diversity and biogeography of the Andes ofColombia. Journal of Biogeography 31: 1829-1839.
KIRCHHOFF, M. D. Y D. N. LARSEN. 1998. Dietary overlap between native Sitka black-tailed deerand introduced elk in Southeast Alaska. Journal of Wildlife Management 62: 236-242.
KUMAR, S., K. TAMURA, I. B. JAKOBSEN Y M. NEI. 2001. MEGA2: Molecular evolutionarygenetics analysis software. Bioinformatics 17: 1244-1245.
KURTÉN, B. Y E. ANDERSON. 1980. Pleistocene mammals of North America. ColumbiaUniversity Press, Nueva York, USA. 442 pp.
KUWAYAMA, R. Y T. OZAWA. 2000. Phylogenetic Relationships among European red deer,wapiti, and sika deer inferred from mitochondrial DNA sequences. Molecular Phylogeneticsand Evolution 15: 115-123.
LATHAM, J., B. W. STAINES Y M. L. GORMAN. 1999. Comparative feeding ecology of red (Cervuselaphus) and roe deer (Capreolus capreolus) in Scottish plantation forests. Journal of Zoology247: 409-418.
LAVENGOOD, J. M., A. WOOLF Y J. L. ROSEBERRY. 1994. Morphological variation in white-taileddeer from Illinois. Transactions of the Illinois Academy of Science 87: 175-185.
LEE, C. E. 2000. Global phylogeography of a cryptic copepod species complex and reproductiveisolation between genetically proximate ''populations''. Evolution 54: 2014-2027.

68 Variación geográfica en venados
LESSA, E. P., J. A. COOK Y J. L. PATTON. 2003. Genetic footprints of demographic expansion inNorth America, but not Amazonia, during the Late Quaternary. Proceedings of the NationalAcademy of Sciences of the USA 100: 10331-10334.
LIZCANO, D. J., V. PIZARRO, J. CAVELIER Y J. CARMONA. 2002. Geographic distribution andpopulation size of the mountain tapir (Tapirus pinchaque). Journal of Biogeography 29: 7-15.
LORENZO, C., L. CUAUTLE Y F. BARRAGAN. 2004. Variación morfométrica a escala temporal enla liebre del Istmo, Lepus flavigularis de México. Anales del Instituto de Biología, SerieZoología 75: 207-228.
LOWE, V. P. W. 1972. Variation in mandible length and body weight of red deer (Cervuselaphus). Journal of Zoology 166: 303-311.
LUDT, C. J., W. SCHROEDER, O. ROTTMANN Y R. KUEHN. 2004. Mitochondrial DNAphylogeography of red deer (Cervus elaphus). Molecular Phylogenetics and Evolution 31:1064-1083.
LYDEKKER, R. 1915. Catalogue of the ungulate mammals in the British Museum (NaturalHistory). Vol IV. Artiodactyla, families Cervidae (deer), Tragulidae (chevrotains), Camelidae(camels and llamas), Suidae (pigs and peccaries), and Hippopotamidae (hippopotamuses).Trustees of the British Museum (Natural History), London, England. 438 pp.
MACSOTAY, O. 1977. Observaciones sobre el neotectonismo cuaternario en el nororientevenezolano. Boletín de Geología 3: 1861-1883.
MA F F E I, M. D., W. D. KL I M ST R A, Y T. J. WI L M E R S. 19 8 8 . C ranial and mandibularcharacteristics of the key deer (Odocoileus virginianus clavium). Journal of Mammalogy 69:403-407.
MALO, A. F., E. R. S. ROLDÁN, J. GARDE, A. J. SOLER Y M. GOMENDIO. 2005. Antlers honestlyadvertise sperm production and quality. Proceedings of The Royal Society of London, SeriesB, Biological Sciences 272: 149-157.
MALONEY, N. J. 1966. Geomorphology of continental margin of Venezuela. Part I. CariacoBasin. Boletín del Instituto Oceanográfico de la Universidad de Oriente 5: 38-53.
MALONEY, N. J. Y O. MACSOTAY. 1967. Geology of La Tortuga Island, Venezuela. AsociaciónVenezolana de Geología, Minería y Petróleo, Boletín Informativo 10: 267-287.
MANDUJANO, S. 2004. Análisis bibliográfico de los estudios de venados en México. ActaZoológica Mexicana 20: 211-251.
MARCHANT, R., A. BOON Y H. HOOGHIEMSTRA. 2002. Pollen-based biome reconstructions forthe past 450000 yr from the Funza-2 core, Colombia: comparisons with model-basedvegetation reconstructions. Palaeogeography, Palaeoclimatology, Palaeoecology 177: 29-45.
MARKUSSON, E. Y I. FOLSTAD. 1997. Reindeer antlers: visual indicators of individual quality?.Oecologia 110: 501-507.
MARSHALL, L. G. 1985. Geochronology and land-mammal biochronology of the TransamericanFaunal interchange. Pp. 49-85. En: F. G. Stehli y S. D. Webb (Eds.), The Great AmericanBiotic Interchange. Plenum Press, Nueva York, USA.
MARTIN, P. R. Y J. K. MCKAY. 2004. Latitudinal variation in genetic divergence of populationsand the potential for future speciation. Evolution 58: 938-945.
MCKENNA, M. C. Y S. K. BELL. 1997. Classification of mammals above the species level.Columbia University Press, Nueva York, USA. 631 pp.
MEIJAARD, E. Y C. P. GROVES. 2004. Morphometrical relationships between South-east Asiandeer (Cervidae, tribe Cervini): evolutionary and biogeographic implications. Journal ofZoology 263: 179-196.
MENDELSON, T. C. Y K. L. SHAW. 2002. Genetic and behavioral components of the crypticspecies boundary between Laupala cerasina and L. kohalensis (Orthoptera: Gryllidae).Genetica 116: 301-310.
MÉNDEZ-AROCHA, J. L. 1955. El nombre científico de los venados caramudos o caramerudosvenezolanos. Memoria de la Sociedad de Ciencias Naturales La Salle 15: 133-136.

69Mem. Fund. La Salle de Cienc. Nat. 167
MENDOZA, M., C. M. JANIS Y P. PALMQVIST. 2002. Characterizing complex craniodentalpatterns related to feeding behaviour in ungulates: a multivariate approach. Journal ofZoology 258: 223-246.
MILÁ, B., D. J. GIRMAN, M. KIMURA Y T. B. SMITH. 2000. Genetic evidence for the effect of apostglacial population expansion on the phylogeography of a North American songbird.Proceedings of The Royal Society of London, Series B, Biological Sciences 267: 1033-1040.
MOLINA, M. Y J. A. PEÑALOZA. 2002. Dog predation on páramo white-tailed deer: the caseMucubají, Sierra Nevada National Park. Memoria de la Fundación La Salle de CienciasNaturales 154: 139-144.
MOLINA, M. Y J. MOLINARI. 1999. Taxonomy of Venezuelan white-tailed deer (Odocoileus,Cervidae, Mammalia), based on cranial and mandibular traits. Canadian Journal of Zoology77: 632-645.
MONTGELARD, C., S. DUCROCQ Y E. DOUZERY. 1998. What is a Suiforme (Artiodactyla)?Contribution of cranioskeletal and mitochondrial DNA Data. Molecular Phylogenetics andEvolution 9: 528-532.
MORELOCK, J., N. J. MALONEY Y W. R. BRYANT. 1972. Structure and sediments of thecontinental shelf of central Venezuela. Boletín del Instituto Oceanográfico de la Universidadde Oriente 11: 127-136.
MORITZ, C., J. L. PATTON, C. J. SCHNEIDER Y T. B. SMITH. 2000. Diversification of rainforestfaunas: an integrated molecular approach. Annual Review of Ecology and Systematics 31:533-63.
MORRONE, J. J. Y J. MÁRQUEZ. 2001. Halffter's Mexican Transition Zone, beetle generalizedtracks, and geographical homology. Journal of Biogeography 28: 635-650.
MOSCARELLA, R. A. 2001. Filogeografía y genética de la conservación del venado caramerudode Venezuela. Tesis de Maestría, Universidad Simón Bolívar, Caracas, Venezuela. 87 pp.
MOSCARELLA, R. A., M. AGUILERA Y A. A. ESCALANTE. 2003. Phylogeography, populationstructure, and implications for conservation of white-tailed deer (Odocoileus virginianus) inVenezuela. Journal of Mammalogy 84: 1300-1315.
NAGATA, J., R. MASUDA, H. B. TAMATE, S. HAMASAKI, K. OCHIAI, M. ASADA, S. TATSUZAWA, K.SUDA, H. TADO Y M. C. YOSHIDA. 1999. Two genetically distinct lineages of the sika deer,Cervus nippon, in Japanese islands: comparison of mitochondrial D-loop region sequences.Molecular Phylogenetics and Evolution 13: 511-519.
NAGL, S., H. TICHY, W. E. MAYER, N. TAKEZAKI, N. TAKAHATA Y J. KLEIN. 2000. The originand age of haplochromine fishes in Lake Victoria, East Africa. Proceedings of The RoyalSociety of London, Series B, Biological Sciences 267: 1049-1061.
ORTEGA, J. Y H. T. ARITA. 1998. The Neotropical-Nearctic limits in Middle America asdetermined by the distribution of bats. Journal of Mammalogy 79: 772-783.
OSGOOD, W. H. 1910. Mammals from the coast and islands of northern South America. FieldMuseum of Natural History, Zoological Series 10: 23-32.
OSGOOD, W. H. 1914. Four new mammals from Venezuela. Field Museum of Natural History,Zoological Series 10: 135-141.
PESOLE, G., C. GISSI, A. DE CHIRICO Y C. SACCONE. 1999. Nucleotide substitution rate ofmammalian mitochondrial genomes. Journal of Molecular Evolution 48: 427-434.
PETERSEON, L. C., G. H. HAUG, R. W. MURRAY, K. M. YARINCIK, J. W. KING, T. J. BRALOWER,K. KAMEO, S. D. RUTHERFORD Y R. B. PEARCE. 2000. Late Quaternary stratigraphy andsedimentation at Site 1002, Cariaco Basin (Venezuela). Pp. 85-99. En: R. M. Leckie, H.Sigurdsson, G. D. Acton y G. Draper (Eds.), Proceedings of the Ocean Drilling Program,Scientific Results. Vol. 165. Texas A & M University, College Station, Texas, USA. 335 pp.
PETERSON, M. N. 2003. Management strategies for endangered Florida Key deer. Tesis deMaestría, Texas A&M University. 143 pp.

70 Variación geográfica en venados
PIPER, D. Z. Y W. E. DEAN. 2002. Trace-element deposition in the Cariaco Basin, Venezuelashelf, under sulfate-reducing conditions - a history of the local hydrography and globalclimate, 20 ka to the present. U.S. Geological Survey Professional Paper 1670: 1-41.
PITRA, C., J. FICKEL, E. MEIJAARD Y P. C. GROVES. 2004. Evolution and phylogeny of old worlddeer. Molecular Phylogenetics and Evolution 33: 880-895.
POLZIEHN, R. O. Y C. STROBECK. 1998. Phylogeny of wapiti, red deer, sika deer, and other NorthAmerican cervids as determined from mitochondrial DNA. Molecular Phylogenetics andEvolution 10: 249-258.
POLZIEHN, R. O. Y C. STROBECK. 2002. A phylogenetic comparison of red deer and wapiti usingmitochondrial DNA. Molecular Phylogenetics and Evolution 22: 342-356.
PUORTO, G., M. G. SALOMALOMÃO, R. D. G. THEAKSTON, R. S. THORPE, D. A. WARRELL Y W.WÜSTER. 2001. Combining mitochondrial DNA sequences and morphological data to inferspecies boundaries: phylogeography of lanceheaded pitvipers in the Brazilian Atlantic forest,and the status of Bothrops pradoi (Squamata: Serpentes: Viperidae). Journal of EvolutionaryBiology 14: 527-538.
QUAY, W. B. 1971. Geographic variation in the metatarsal "gland" of the white-tailed deer(Odocoileus virginianus). Journal of Mammalogy 52: 1-11.
RANDI, E., N. MUCCI, F. CLARO-HERGUETA, A. BONNET Y E. J. P. DOUZERY. 2001. Amitochondrial DNA control region phylogeny of the Cervinae: speciation in Cervus andimplications for Conservation. Animal Conservation 4: 1-11.
RANDI, E., N. MUCCI, M. PIERPAOLI Y E. DOUZERY. 1998. New phylogenetic perspectives on theCervidae (Artiodactyla) are provided by the mitochondrial cytochrome b gene. Proceedingsof The Royal Society of London, Series B, Biological Sciences 265: 793-901.
REES, J. W. 1969a. Morphologic variation in the cranium and mandible of the white-tailed deer(Odocoileus virginianus): a comparative study of geographical and four biological distances.Journal of Morphology 128: 95-112.
REES, J. W. 1969b. Morphologic variation in the mandible of the white-tailed deer (Odocoileusvirginianus): a study of populational skeletal variation by Principal Component andCanonical Analyses. Journal of Morphology 128: 113-130.
REES, J. W. 1971. Discriminatory analysis of divergence in mandibular morphology ofOdocoileus. Journal of Mammalogy 52: 724-731.
REID, F. A. 1997. A field guide to the mammals of Central America and southeast Mexico.Oxford University Press, Oxford, England. 334 pp.
ROHLF, F. J. Y F. L. BOOKSTEIN. 1987. A comment on shearing as a method for "size correction".Systematic Zoology 36: 356-367.
RUEDAS, L. A., J. SALAZAR-BRAVO, J. W. DRAGOO Y T. L. YATES. 2000. The importance of beingearnest: what, if anything, constitutes a "specimen examined". Molecular Phylogenetics andEvolution 17: 129-132.
RULL, V. 1999. Palaeoclimatology and sea-level history in Venezuela. New data, land-seacorrelations, and proposals for future studies in the framework of the IGBP-PAGES Project.Interciencia 24: 92-101.
SATO, A., C. O'HUIGIN, F. FIGUEROA, P. R. GRANT, B. R. GRANT, H. TICHY Y J. KLEIN. 1999.Phylogeny of Darwin's finches as revealed by mtDNA sequences. Proceedings of the NationalAcademy of Sciences of the USA 96: 5101-5106.
SAUER, P. R. 1984. Physical characteristics. Pp. 73-90. En: L. K. Halls (Ed.), White-tailed deer- ecology and management. Stackpole Books, Harrisburg, Pennsylvania, USA.
SCOTT, K. M. Y C. M. JANIS. 1993. Relationships of the Ruminantia (Artiodactyla) and an

71Mem. Fund. La Salle de Cienc. Nat. 167
analysis of the characters used in ruminant taxonomy. Pp. 282-302. En: F. S. Szalay, M. J.Novacek y M. C. McKenna (Eds.), Mammal phylogeny: placentals. Springer-Verlag, NewYork, USA.
SCHAEFER, J. Y M. B. MAIN. 1999. White-tailed deer of Florida. University of Florida,Department of Wildlife Ecology and Conservation 133: 1-19.
SCHONEWALD, C. M., J. W. BAYLESS Y J. SCHONEWALD. 1985. Cranial morphometry of pacificcoast elk (Cervus elaphus). Journal of Mammalogy 66: 68-74.
SCHUBERT C. 1982. Origin of Cariaco basin, southern Caribbean Sea. Marine Geology 47: 345-360.
SCHWARTZ, M. K., K. L. PILGRIM, K. S. MCKELVEY, E. L. LINDQQUIST, J. J. CLAAR, S. LOCHC
Y L. F. RUGIERO. 2004. Hybridization between Canada lynx and bobcats: genetic results andmanagement implications. Conservation Genetics 5: 349-355.
SITES, J. W. JR. Y K. A. Crandall. 1997. Testing species boundaries in biodiversity studies.Conservation Biology 11: 1289-1297.
SKLENÁR, P. Y P. M. JØRGENSEN. 1999. Distribution patterns of páramo plants in Ecuador.Journal of Biogeography 26: 681-691.
SMITH, M. H., W. V. BRANAN, R. L. MARCHINTON, P. E. JOHNS Y M. C. WOOTEN. 1986. Geneticand morphologic comparisons of red brocket, brown brocket, and white-tailed deer. Journalof Mammalogy 67: 103-111.
SMITH, W. P. 1991. Odocoileus virginianus. Mammalian Species 388: 1-13.
STÖCKHERT, B., W. V. MARESCH, A. TOETZ, R. KLUGE, G. KRÜCKHANS, C. K. AISER, V.AGUILAR, T. KLIER, S. LAUPENMÜHLEN, D. PPIEPENBREIER Y I. WIETHE. 1994. Tectonichistory of Isla Margarita, Venezuela - A record of a piece of crust close to an active platemargin. Zentralblatt für Geologie und Paläontologie 1: 485-498.
STÖCKHERT, B., W. V. MARESCH, M. BRIX, C. KAISER, R. KLUGE, G. KRÜCKHANS-LUEDER Y A.TOETZ. 1995. The crustal history of Margarita Island (Venezuela) in detail: A constraint onthe Caribbean plate-tectonic scenario. Geology 23: 787-790.
SU, B., Y. WANG, H. LAN, W. WANG Y Y. ZHANG. 1999. Phylogenetic study of completecytochrome b genes in musk deer (genus Moschus) using museum samples. MolecularPhylogenetics and Evolution 12: 241-249.
TAMATE, H. B., S. TATSUZAWA, K. SUDA, M. IZAWA, T. DOI, K. SUNAGAWA, F. MIYAHIRA Y H.KADO. 1998. Mitochondrial DNA variations in local populations of the Japanese sika deer,Cervus nippon. Journal of Mammalogy 79: 1396-1403.
TARIFA, T., J. RECHBERGER, R. B. WALLACE Y A. NÚÑEZ. 2001.Confirmación de la presencia deOdocoileus virginianus (Artiodactyla, Cervidae) en Bolivia, y datos preliminares sobre suecología y su simpatría con Hippocamelus antisensis. Ecologia en Bolivia 35: 41-49.
TEMPLETON, A. R. 1989. The meaning of species and speciation: a genetic perspective. Pp. 3-27.En: D. Otte y J. A. Endler (Eds.), Speciation and its Consequences. Sinauer, Sunderland,Massachusetts, USA.
TOMIATI, C. Y L. ABBAZZI. 2002. Deer fauna from Pleistocene and Holocene localities ofEcuador (South America). Geobios 35: 631-645.
UDINA, I. G., A. A. DANILKIN Y G. G. BOESKOROV. 2002. Genetic diversity of moose (Alces alcesL.) in Eurasia. Russian Journal of Genetics 38: 951-957.
VAN DER HAMMEN, T. 1979. Historia y tolerancia de ecosistemas parameros. Pp. 55-66. En: M.L. Salgado-Laboriau (Ed.), El medio ambiente páramo. Ediciones del Centro de EstudiosAvanzados del Instituto Venezolano de Investigaciones Científicas, Caracas, Venezuela.
VAN DER HAMMEN, T. Y A. M. CLEEF. 1986. Development of the high Andean paramo flora and

72 Variación geográfica en venados
vegetation. Pp. 153-201. En: F. Vuilleumier y M. Monasterio (Eds.), High altitude tropicalbiogeography. Oxford University Press, Nueva York, USA.
VAN DER HAMMEN, T. Y H. HOOGHIEMSTRA. 1997. Chronostratigraphy and correlation of thePliocene and Quaternary of Colombia. Quaternary International 40: 81-91.
VILLARREAL, O. A. 2002. El Grand-Slam de venado cola blanca mexicano, una alternativasostenible. Archivos de Zootecnia 51: 187-193.
VRBA, E. S. Y G. B. SCHALLER. 2000. Phylogeny of Bovidae based on behavior, glands, skulls,and postcrania. Pp. 203-222. En: E. S. Vrba y G. B. Schaller (Eds.), Antelopes, deer, andrelatives - fossil record, behavioral ecology, systematics, and conservation. Yale UniversityPress, New Haven, USA.
VUILLEUMIER, B. S. 1971. Pleistocene changes in the fauna and flora of South America. Science173: 771-780.
WANG, W. Y H. LANG. 2000. Rapid and parallel chromosomal number reductions in muntjacdeer inferred from mitochondrial DNA phylogeny. Molecular Biology and Evolution 17:1326-1333.
WEBB, S. D. 1985. Late Cenozoic mammal dispersals between the Americas. Pp. 357-386. En:F. G. Stehli y S. D. Webb (Eds.), The great american biotic interchange. Plenum Press, NuevaYork, USA.
WEBB, S. D. 1991. Ecogeography and the Great American Interchange. Paleobiology 17: 266-280.
WEBB, S. D. 2000. Evolutionary history of New World Cervidae. Pp. 38-64. En: E. S. Vrba y G.B. Schaller (Eds.), A n telopes, deer, and re l a t i ves - fossil re c o rd, behav i o ral ecology,systematics, and conservation. Yale University Press, New Haven, USA.
WEBER, J. C., T. H. DIXON, C. DEMETS, W. B. AMBEH, P. JANSMA, G. MATTIOLI, J. SALEH, G.SELLA, R. BILHAM Y O. PÉREZ. 2001. GPS estimate of relative motion between the Caribbeanand South American plates, and geologic implications for Trinidad and Venezuela. Geology29: 75-78.
WEBER, M. Y S. GONZÁLEZ. 2003. Latin American deer diversity and conservation: a review ofstatus and distribution. Ecoscience 10: 443-454.
WEBSTER, G. L. 1995. The panorama of neotropical cloud forests. Pp. 53-77. En: S. P. ChurchillH. Balslev, E. Forero y J. L. Luteyn (Eds.), Biodiversity and conservation of netropicalmontane forests. New York Botanical Garden, Nueva York.
WHEELER, T. A. 2003. The role of voucher specimens in validating faunistic and ecologicalresearch. Biological Survey of Canada (Terrestrial Arthropods), Ottawa, Canada. 21 pp.
WILSON, D. E. Y D. M. REEDER. 1993. Mammal species of the world - a taxonomic andgeographic reference. Smithsonian Institution Press, Washington, USA. 1206 pp.
ZAMUDIO, K. R. Y H. W. GREENE. 1997. Phylogeography of the bushmaster (Lachesis muta:Vi p e ridae): implications for Ne ot ropical bioge o gra p hy, systematics, and conserva t i o n .Biological Journal of the Linnean Society 62: 421-442.
ZANDER, R. H. 2003. Reliable phylogenetic resolution of morphological data can be better thanthat of molecular data. Taxon 52: 109-112.
Recibido: 06 julio 2005Aceptado: 12 junio 2007
Jesús MolinariGrupo de Ecología Animal, Departamento de Biología, Facultad de Ciencias, Universidad de LosAndes, Mérida 5101, Venezuela. [email protected]