universidad de caldas vicerrectorÍa de … · locales realizadas por los agricultores (lobo et...
TRANSCRIPT

UNIVERSIDAD DE CALDAS VICERRECTORÍA DE INVESTIGACIONES Y POSTGRADOS
Guía para la presentación de
Proyectos de investigación, innovación y creación
CONTENIDO
1. Información general
Título del proyecto: Caracterización de aislamientos de Botrytis cinerea Pers. e identificación de genes
análogos de resistencia en diferentes genotipos de mora de castilla (Rubus glaucus Benth)
Nombre de los Grupos de Investigación:
Grupo de Investigación en Biodiversidad y Biotecnología (Universidad Tecnológica de Pereira)
Nombre del Grupo: Grupo de Investigación en Biodiversidad y Biotecnología Facultad/Departamento: Ciencias Ambientales
Clasificación _A__
Nombre del Grupo: Facultad/Departamento:
Clasificación _____
Nombre: Facultad/Departamento:
Clasificación _____
Nombre: Facultad/Departamento:
Clasificación _____
Tipo de proyecto de I&D: Investigación Básica: Investigación Aplicada: Creación: Innovación Tecnológica
1:
Tipo de innovación: Innovación tecnológica de producto Innovación tecnológica de proceso Innovación organizacional
Área Estrátegica del Plan de Desarrollo
Biotecnología
Artes, Cultura y Humanidades Problemática Social
Salud
Ambiental
No corresponde a ninguna de las áreas estratégicas
INTEGRANTES DEL EQUIPO DE INVESTIGACIÓN (indicar cuál es el investigador responsable)
NOMBRE/TIPO DE VINCULACIÓN EN LA UNIVERSIDAD DE CALDAS
DEPARTAMENTO o PROGRAMA
ÁREA DE CONOCIMIENTO (según anexo 1)
1 Se refiere a aquellos proyectos que tienen como objetivo el desarrollo de nuevos productos o procesos, así como las
modificaciones tecnológicas importantes en productos o procesos
X
X

Liliana Isaza Valencia (Responsable) Facultad de Ciencias
Ambientales UTP Otra
Marta Leonor Marulanda (Director) Facultad de Ciencias
Ambientales UTP Ciencias Naturales
Carlos Felipe Barrera Sánchez (Codirector) Facultad de Ciencias
AgropecuariasUniversidad de Caldas
Agronomía, Veterinaria y afines
Lugar de Ejecución del Proyecto: (Municipio/Departamento) Ciudad: Pereira Departamento: Risaralda
Presupuesto
Valor total del proyecto: $ 188.767.760 Fuentes de financiación: Sistema General de Regalías
Valor solicitado en esta convocatoria: $
Duración total (meses): 30
2. Resumen ejecutivo
La mora de castilla (Rubus glaucus, Benth) en Colombia es una de las frutas más
importantes y con mayor proyección comercial. Es cultivada por pequeños y medianos
agricultores, constituyendo una de las principales fuentes de ingresos, empleo rural y de
alternativa agroindustrial (Botero y Franco, 2007). Entre los factores que afectan la
producción de la mora sobresalen el ataque de enfermedades y la escasa disponibilidad
de información para agilizar en cada caso su control.
Dentro de las enfermedades más limitantes e importantes del cultivo se encuentra el
moho gris producido por el hongo Botrytis cinerea (Dashwood y Fox, 1998; Molina et ál.
2004). Esta enfermedad es considerada una de más comunes y serias del género
Rubus y de otros pequeños frutos como fresas, uvas, frambuesa a nivel mundial (Molina
et al. 2004). En Colombia, se han reportado pérdidas de frutos de mora entre 50% y
76%, a causa del moho gris (Tamayo et al., 2002)
El Grupo de Investigación en Biodiversidad y Biotecnología de la Universidad
Tecnológica de Pereira, mediante investigación científica y participativa ha venido
adelantando investigaciones tendientes a la consecución de materiales de siembra de
mora de excelente calidad fitosanitaria y productiva que les permitan a los agricultores
ser más competitivos en el mercado. Esta propuesta de investigación pretende realizar

la caracterización morfológica, patogénica y genética de aislamientos de Botrytis
cinerea Pers. en cultivos de Rubus glaucus Benth en Colombia y la búsqueda de
posible resistencia a la enfermedad en diferentes genotipos de mora de castilla
previamente caracterizados por el Grupo de Investigación y está vinculada al programa
“Mejoramiento de la calidad del material de siembra para la competitividad de la cadena
productiva de mora”.
3. Conformación y trayectoria del equipo de investigadores
El Grupo de Investigaciones en Biodiversidad y Biotecnología (COL0000719), inició
actividades en 1993, como parte de la Facultad de Ciencias Ambientales de la
Universidad Tecnológica de Pereira, a través de proyectos de investigación y de
postgrado en biotecnología, biología molecular, agroecología y biodiversidad. Las líneas
de investigación responden a la ubicación geográfica y ecológica de la región, al
compromiso social de la Universidad y a la búsqueda de opciones agrícolas y de
manejo de la vida silvestre que procuren combinar producción y conservación,
biodiversidad y necesidades humanas.
Líneas de Investigación:
Biología molecular
Cultivo de tejidos
Mejoramiento genético
Fitopatología
Bosques, agro ecosistemas y biodiversidad
Humedales naturales
En los últimos cinco años el grupo de investigación en Biodiversidad y Biotecnología ha
centrado su investigación en especies cultivadas y forestales de interés comercial,
tales como nogal cafetero (Cordia alliodora (Ruíz & Pavón), guayacán rosado (Tabebuia

rosea (Bertol D.C), mora de castilla (Rubus glaucus Benth) y algunas especies y
cultivares del género Heliconia.
El grupo presenta una extensa producción académica, con la publicación de más de 60
artículos en revistas indexadas nacionales e internacionales, libros y capítulos de libros.
Además de dos patentes de invención. Actualmente, el grupo se encuentra
desarrollando junto con tres grupos de investigación de la Universidad Tecnológica de
Pereira y un Grupo de investigación de la Universidad Libre el proyecto:
“DESARROLLO DE CAPACIDADES CIENTÍFICAS Y TECNOLÓGICAS EN
BIOTECNOLOGÍA APLICADAS A LOS SECTORES DE LA SALUD Y LA
AGROINDUSTRIA EN EL DEPARTAMENTO DE RISARALDA”, financiado con recursos
del Sistema General de Regalías, vigencia 2014 – 2019.
4. Descripción del proyecto
4.1 Planteamiento de la pregunta o problema de investigación y su
justificación
La mora (Rubus glaucus, Benth) en Colombia es una de las frutas más importantes y
con mayor proyección comercial. Es cultivada por pequeños y medianos agricultores,
constituyendo una de las principales fuentes de ingresos, empleo rural y de alternativa
agroindustrial (Botero y Franco, 2007). El cultivo de la mora de castilla se ha
incrementado considerablemente en los últimos años en Colombia, debido al aumento
de la demanda de la fruta tanto para el consumo humano y agroindustrial de alimentos,
como para la exportación, de esta manera se constituye en una alternativa de
producción, generadora de ingresos para los pequeños y grandes productores (de la
Rotta, 2001).
A pesar de su reconocida importancia en la generación de ingresos para los
productores, este cultivo ha tenido poco desarrollo tecnológico, la calidad y
productividad presentan alta variabilidad, debido principalmente a la falta de variedades

reconocidas y a la falta de material de siembra de calidad genética y fitosanitaria. Hasta
el momento la siembra de esta especie se realiza a través de empleo de selecciones
locales realizadas por los agricultores (Lobo et al., 2002). Entre los factores que afectan
la producción sobresalen el ataque de enfermedades y la escasa disponibilidad de
información para agilizar en cada caso su control. En forma general se menciona la
existencia de un patógeno, pero no se ofrece la información que señale su importancia
epidemiológica ni las alternativas de solución (de la Rotta, 2001). Dentro de las
enfermedades más limitantes e importantes del cultivo se encuentra el moho gris
producido por el hongo Botrytis cinerea (Dashwood y Fox, 1998; Molina et al. 2004).
Esta enfermedad es considerada una de más comunes y serias del género Rubus y de
otros pequeños frutos como fresas, uvas, frambuesa a nivel mundial (Molina et al.
2004). Tamayo et al., 2002, reportaron pérdidas de frutos de mora entre 50% y 76% en
Colombia, a causa del moho gris.
La incidencia de este tipo de agentes fitopatógenos ha llevado a una aplicación
excesiva de fungicidas lo cual genera fenómenos de resistencia en hongos como B.
cinerea (Medina, et al., 2009). Además del problema mencionado, actualmente los
consumidores muestran una preferencia por productos agrícolas libres de compuestos
químicos debido a los efectos secundarios de estos sobre la salud humana y el medio
ambiente (Calvo et al., 2012).
Uno de los objetivos propuestos por la Cadena Agroalimentaria de la Mora es realizar la
evaluación de materiales de mora frente al comportamiento de problemas fitosanitarios.
En este contexto, se presenta como una opción viable la evaluación de la diversidad de
Botrytis cinerea en Colombia, así como, la caracterización y evaluación del
comportamiento de materiales de mora de castilla frente al ataque de este fitopatógeno
en procura de la búsqueda de materiales promisorios que permitan aumentar la calidad
y la productividad de este importante cultivo.
Esta propuesta hace parte de un conjunto de investigaciones llevadas a cabo por la
Universidad Tecnológica de Pereira, dentro del marco del programa “Mejoramiento de la

calidad del material de siembra para la competitividad de la cadena productiva de mora”
financiado por el Sistema General de Regalías.
4.2 Marco Teórico
4.2.1 Generalidades del cultivo de la mora
El cultivo de la mora es originario de los andes americanos, pertenece a la familia
Rosaceae, género Rubus, que cuenta con alrededor de 400 especies distribuidas en
América, Europa, África y Asia se constituye en uno de los géneros de mayor diversidad
genética del reino vegetal (Alvarado, 2002). Es una planta arbustiva perenne con tallos
rastreros, sus hojas son trifoliadas y alternas, su propagación se puede realizar por
semilla, por acodo, por estaca, in-vitro, entre otras (Bernal y Díaz, 2006). En Colombia
la principal especie de mora cultivada es R. glaucus, conocida como mora de castilla
(Correa, 2002), cuyo origen comprende las zonas tropicales altas de América.
El desarrollo óptimo del cultivo de la mora se encuentra entre los 1.900 y 2.400
m.s.n.m., con temperaturas entre 14°C y 19°C, humedad relativa del 65% al 80%, y una
precipitación anual de 1.200 mm a 1.700 mm. Los suelos deben ser francos, con buena
cantidad de materia orgánica, y adecuado drenaje que evite encharcamientos (Franco,
2002).
En Colombia existen variaciones fenotípicas de esta especie en cuanto a tamaño, color
y calidad del fruto, que posiblemente se produjo por una selección practicada en
antiguas épocas a partir de plantas silvestres (Córdoba y Londoño 1996). Este cultivo
está distribuido en el país desde el Putumayo hasta el Magdalena Medio. Otras
variedades cultivadas en el país son la mora negra, Rubus bogotensis HBK, la mora de
páramo, Rubus giganteus, la mora pequeña, R. megalococus y la mora grande Rubus
nubigenus.

Colombia cuenta con 7.007 hectáreas plantadas de mora, de las cuales 4.922 están en
edad productiva. Se presentan rendimientos promedio de 15 toneladas por hectárea, lo
que significa una producción de 73.856 toneladas anuales. En el departamento de
Risaralda el área sembrada en mora es de aproximadamente de 400 hectáreas, con
una producción de 2.137 toneladas y un rendimiento de 5,3 toneladas por hectárea
(DANE, 2011).
Este cultivo es muy importante para la región, ya que requiere de mucha mano de obra,
ofertando empleo en la mayoría de los casos a todo el grupo familiar; además la mora
es un fruto con gran demanda en los mercados nacionales e internacionales, es uno de
los cultivos de fruta que despierta mayor interés en Colombia dada su buena
comercialización como fruta fresca y como materia prima en la industria alimenticia para
la elaboración de jugos, mermeladas o concentrados de pulpa. Esto ha permitido un
incremento relativamente rápido del cultivo (Medina, et al., 2009).
4.2.2 Botrytis cinerea como factor limitante del cultivo de la mora
Entre los factores que afectan la producción sobresalen el ataque de enfermedades y la
escasa disponibilidad de información para agilizar en cada caso su control. El moho gris
o pudrición del fruto causado por Botrytis cinerea Pers, es considerada una de las
enfermedades más comunes y serias del género Rubus. El género Botrytis constituye
un grupo de hongos fitopatógenos ampliamente conocido, en el que podemos encontrar
especies que parasitan a una sola especie vegetal, como B. tulipae, B. squamosa o B.
fabae, patógenos de tulipán, cebolla y haba respectivamente, y una especie,
denominada B. cinerea, capaz de infectar al menos 235 especies de plantas distintas
(Elad, 1997), causando la enfermedad conocida en Colombia como “Moho Gris” o
“Botrytis”. B. cinerea es un hongo fitopatógeno que causa importantes pérdidas
económicas en un amplio rango de cultivos de importancia agrícola (Antonov et al.,
1997). Este patógeno puede atacar a los cultivos en cualquier estado de su desarrollo e
infectar cualquier parte de la planta, colonizando hojas, tallos, flores, tubérculos y frutos
(Elad, 1997; Muckenschnabel et al., 2002), por lo que los ataques reportados por este

hongo afectan no sólo el rendimiento, sino además la calidad de los cultivos,
provocando deterioros de gran magnitud incluso durante las etapas de transporte y
almacenaje de los productos agrícolas (Elad, 1988; Hammer et al., 1990; Berrie, 1994;
Rosslenbroich y Stuebler, 2000).
El control de esta enfermedad se efectúa mediante el uso de fungicidas como Captan,
Benomil, Clorotalonil y Botrycidas de última generación; sin embargo, debido a su modo
de acción (inhibición de la síntesis de Ergosterol, del crecimiento micelial y de la
germinación), han ocasionado el desarrollo de resistencia por parte del hongo, haciendo
ineficaces las aplicaciones (Stehmann y De Waard, 1996; Rosslenbroich et al., 2000),
además su acumulación en la fruta es tóxica para personas y animales (EPA, 2005),
afectando el posicionamiento de la fruta en el mercado internacional.
Dado que cada vez son mayores las objeciones de orden fitosanitario que se plantean
sobre el uso excesivo de plaguicidas de síntesis, debido al riesgo que representan para
la salud humana, la preocupación por sus posibles efectos nocivos adquiere especial
importancia, si tenemos en cuenta que gran parte de la fruta cosechada es consumida
en fresco y es muy corta la proximidad temporal entre la aplicación de los pesticidas y
su consumo. Dado el peligro que representan los residuos de pesticidas para la salud
humana, los consumidores en el mundo están exigiendo que el uso de productos
químicos en la producción agrícola se reduzca significativamente (Zapata et al., 2013),
se hace necesario entonces, buscar herramientas para generar alternativas
tecnológicas que contribuyan al manejo integrado de la enfermedad (Saldarriaga, 2005).
El conocimiento y la caracterización de la diversidad de Botrytis cinerea en Colombia,
así como, la evaluación del comportamiento de materiales de mora de castilla frente al
ataque de este fitopatógeno permitirán la búsqueda y selección de materiales
promisorios, resistentes o tolerantes al ataque del hongo, que permitan aumentar la
calidad y la productividad de este importante cultivo.
4.2.3 Agente causal y ciclo de vida de Botrytis cinerea

Botrytis cinerea es la forma imperfecta o asexual del hongo Botryotina fuckeliana de
Bary (Whetzel) que pertenece a la división Ascomycota, clase Discomycetes, orden
Helotiales y familia Sclerotiaceae (Castaño, 2015). Aunque normalmente se designe a
los hongos por el estado sexual o teleomorfo, en este caso, el estado anamorfo, B.
cinerea es aceptado, siendo reconocido en todo el mundo por micólogos y fitopatólogos
(Benito, 2010).
La forma teleomorfa del hongo, B. fuckeliana, fue descrita por Bary en 1886 como un
apotecio pediceliar de color marrón que nace sobre un esclerocio. La parte fértil está
formada por ascas entremezcladas con hifas de longitud similar e incolora. Las ascas
son cilíndricas, planas en la parte superior y contienen 8 ascosporas elípticas, incoloras
(Benito, 2010), los apotecios son pedicelados producidos a partir de momias en el suelo
o esclerocios (Castaño, 2015).
La forma anamorfa, B. cinerea, pertenece a la división Deuteromycota, clase
Hyphomycetes, orden Moniliales y familia Moniliaceae, cuyos conidióforos y conidios
son hialinos (Castaño, 2015). El patógeno produce un micelio gris largo y abundante,
conidióforos ramificados con células apicales redondeadas en racimos, incoloras o
grises, unicelulares y presencia de conidios ovoides. Los conidióforos y los racimos de
conidias se asemejan a un racimo de uvas. Las conidias son liberadas fácilmente en
tiempo húmedo y son transportadas por el aire. El hongo produce frecuentemente
esclerocios negros, duros y de forma irregular (Agrios, 2005).
Botrytis sobrevive al invierno en desechos de plantas como micelio o como esclerocios
(Figura 1). El hongo no parece infectar las semillas, pero se puede transmitir con
semillas contaminadas con esclerocios o con trozos de restos de plantas infectadas con

el hongo. El hongo requiere una temperatura entre 18 y 23 °C, tiempo húmedo para
obtener mejor crecimiento, esporulación, liberación de esporas, germinación y
establecimiento de la infección. El patógeno es activo a bajas temperaturas y causa
considerables pérdidas en cultivos. Las esporas penetran los tejidos a través de heridas
o aberturas naturales y producen micelio en los pétalos de las flores viejas, follaje seco,
bulbos muertos y así sucesivamente. Los esclerocios de Botrytis usualmente germinan
produciendo micelio que puede infectar directamente, en algunos casos los esclerocios
germinan produciendo apotecios y ascosporas (Agrios, 2005).
Figura 1. Ciclo de vida de Botrytis cinerea (Tomado de Agrios, 2005)
4.2.4 Interacción patogénica planta-Botrytis
Las plantas constantemente interactúan con una gran variedad de microorganismos
tales como virus, bacterias, hongos, nematodos e insectos. Las poblaciones silvestres

pueden generalmente enfrentar este tipo de patógenos potenciales. Sin embargo,
cuando los cultivos genéticamente uniformes cubren áreas extensas, pueden sufrir
graves daños por patógenos virulentos, y por lo tanto, se requieren medidas costosas
de protección para reducir las pérdidas en la productividad. Desde hace tiempo se
conoce que moléculas localizadas en la superficie de las células del patógeno o
secretadas por las mismas activan las reacciones de defensa en las plantas. Estas
moléculas pueden ser de diferente naturaleza química (carbohidratos, lípidos, y
proteínas) (García y Lozoya, 2004).
B. cinerea infecta principalmente a los órganos fructíferos de la planta (flores y frutos),
pero también es capaz de originar pudriciones en los tallos, ahogamientos de plántulas,
manchas foliares y pudriciones de tubérculos, bulbos y raíces (Agrios, 1997). Como se
puede observar, existen grandes diferencias entre los huéspedes a los que puede
infectar, lo que hace pensar que B. cinerea posee un mecanismo de infección complejo.
De forma general, el ciclo de infección (Prins et al., 2000) puede considerarse dividido
en varias etapas:
1. Adhesión de los conidios sobre la superficie del huésped.
2. Germinación de los conidios si las condiciones son favorables.
3. Penetración del tejido vegetal, bien a través de heridas o de aberturas naturales,
mediante la participación de distintas enzimas, la utilización de procesos enzimáticos, la
excreción de toxinas o combinación de estas.
4. Muerte de las células adyacentes al punto de penetración, dando lugar a la formación
de una lesión primaria.
5. En algunos casos, una fase de latencia, durante la cual parece que los mecanismos
de la planta logran controlar al patógeno, que permanece localizado en las áreas
necrosadas de las primeras lesiones.

6. Una vez vencidas las defensas de la planta, se inicia la diseminación en el tejido
vegetal circundante, originando la colonización y la maceración.
7. Esporulación del hongo sobre el tejido macerado, produciéndose una nueva
generación de conidios que están listos para ser dispersados e iniciar un nuevo ciclo de
infección.
En el proceso infectivo de Botrytis cinerea Pers. existen una serie de factores que
inciden en la patogenicidad del hongo. Un factor de patogenicidad, puede tomarse
como cualquier agente que contribuya a la penetración, invasión, colonización y
maceración de un tejido vegetal vivo (Benito, et al., 2000). En la etapa de penetración
de la epidermis, el hongo desarrolla estructuras de tipo apresorio que le ayudarán a
establecerse (Clark y Lobeer, 1976). Paralelamente, se activarán una serie de procesos
enzimáticos destacándose la participación de enzimas hidrolíticas extracelulares como
endo y exo poligalacturonasas, pectin metil esterasas, pectin liasas, las que están
implicadas en la degradación de la pared celular vegetal, o proteasas (Johnston y
Williamson, 1992; Reignault, 1994; Movahedi et al.,1990b; Movahedi y Heale, 1995;
Sasaki y Nagayama, 1994). En este sentido, desde hace algunos años se conoce a
Botrytis cinerea, por ser un hongo necrotrófico que posee la destreza de matar a las
células de su huésped, para iniciar su ciclo de vida. Lo anterior, sugiere que el patógeno
produce toxinas que le ayudarían en su colonización pero a su vez en la planta, se
desarrollarían productos génicos de tipo tóxico que limitaría el crecimiento de B. cinerea
lo cual consideraría distintos mecanismos y proteínas que responden frente a la
infección producida por este hongo. Durante el transcurso de la interacción entre
Botrytis cinerea y el huésped, la planta intenta enérgicamente evitar la invasión y el
proceso de la enfermedad activando una multitud de rutas de defensa, como la
producción de metabolitos antifúngicos y proteínas involucradas en patogénesis (van
Baarlen et al., 2004). Dentro de ellas, se encuentran proteínas con respuesta a
patógenos (proteínas PR) y enzimas hidrolíticas (quitinasas y 1,3-ß- glucanasas), las
que están a su vez relacionadas con hormonas endógenas de plantas como son ácido

salicílico (SA) y ácido jasmónico (JA). Estos aumentan en presencia de patógenos
como B. cinerea induciendo separadamente a un grupo de genes que codifican para
proteínas antimicrobianas.
4.2.5 Genes Análogos de Resistencia
La resistencia de las plantas está estrechamente correlacionada con la capacidad de
respuesta de una planta hacia un organismo potencialmente patógeno, los mecanismos
de defensa de las plantas se pueden dividir en: a. Mecanismos preexistentes o
constitutivos, formados por las características estructurales de la pared celular y la
presencia de compuestos químicos depositados en el tejido epidérmico y b.
mecanismos de defensa inducibles formados por la síntesis de novo de compuestos
químicos antimicrobianos cuya síntesis se activa después de un intento de invasión al
tejido vegetal por un patógeno (García y Lozoya, 2004). Como requisito para que una
planta sea capaz de reconocer de manera eficiente a un patógeno, y por lo tanto de
activar las respuestas de defensa apropiadas, se sugiere que hay receptores
localizados en la membrana plasmática de las células vegetales encargados de activar
las respuestas de defensa (Staskawicz., 1995)
Tras el reconocimiento de los genes R, de resistencia de la planta y de los factores de
avirulencia (Avr) del patógeno, se desencadena la rápida activación de la respuesta
defensiva de la planta. Este modelo denominado gen a gen, fue propuesto por Flor
(1971) y ha sido confirmado posteriormente en numerosas interacciones planta-
patógeno (Innes, 1998). La caracterización molecular de varios genes de resistencia de
diversas especies de plantas, con especificidad para una amplia variedad de
patógenos, ha develado que estos comúnmente codifican para proteínas de estructura
similar aunque altamente polimórficos (Dangl y Jones, 2001).
En los últimos años, se ha abordado la clonación de numerosos genes de avirulencia a
partir de diversos patógenos, así como genes de resistencia procedentes de diversas
especies vegetales. La clonación de los genes R revela la existencia de un número

limitado de motivos estructurales (Hammond- Kosack y Parker, 2003). Estos motivos
incluyen dominios posiblemente implicados en el reconocimiento o en la interacción con
los factores de avirulencia y dominios de señalización. Este limitado repertorio de
motivos sugiere que tanto el reconocimiento específico del patógeno por las proteínas R
como los procesos subsiguientes están mecanísticamente conservados (Gil, M.J.,
2005).
Según Ellis y Jones en 1998, un dominio es una secuencia de aminoácidos de una
proteína con una función particular y de acuerdo al dominio común que comparten las
proteínas que son codificadas por los genes R se agrupan en cinco clases diferentes:
a. Genes R que codifican para proteínas con dominio actividad de proteína de
serina/treonina. El Gen R representativo de este grupo es un gen Pto de la planta de
tomate que confiere resistencia a la bacteria Pseudomona syringae pv. tomato y
codifica para una proteína con actividad de cinasa serina/treonina.
b. Genes R que codifican para proteínas con dominios repetidos ricos en leucina
(RRL). Dentro de este grupo se incluye a los genes Xa21 del arroz (Oryza sativa L.),
que confieren resistencia a la bacteria Xanthomonas oryzae ex Ishiyama pv. oryzae
raza 6 y los genes Cf de tomate que confieren resistencia en contra del hongo
fitopatógeno Cladosporium fulvum Cooke (Ellis et al., 2000). Estos genes de resistencia
se caracterizan porque las proteínas que codifican contienen una región RRL
extracelular y un dominio transmembranal, pero carecen propiamente de una región
intracelular. La región RRL contiene leucinas y otros aminoácidos hidrofóbicos en
intervalos regulares, puede también contener prolinas y asparaginas espaciadas
regularmente (Kobe y Deisenhofer, 1995). El dominio RRL de las proteínas R podría ser
el dominio de unión para la proteína AVR del patógeno. Alternativamente, el dominio
RRL podría facilitar la interacción entre las proteínas R con otras proteínas participantes
en la activación de la resistencia.

c. Genes R que codifican para proteínas con dominio de unión para nucleótidos.
Muchos genes de resistencia que codifican para regiones RRL también codifican para
secuencias de aminoácidos con una alta similitud para dominios de unión a nucleótido
(DUN). La presencia de dominios DUN en algunas proteínas R (los productos de la
codificación de los genes R) sugiere que la unión a nucleótidos de trifosfato es esencial
para el funcionamiento de estas proteínas. Los genes con dominios DUN-RRL son
abundantes en las plantas.
d. Genes R que codifican para proteínas con dominio de cierre de leucina. Dentro
de la subclase de proteínas R que poseen dominios DUN-RRL, hay otros subgrupos de
proteínas (RPS1, RPM1 y Prf) que confieren resistencia en contra de P. syringae, y
todas poseen un probable dominio de cierre de leucina en su extremo amino terminal.
Estos dominios son conocidos por su papel durante la homo y heterodimerización de
factores de transcripción eucarióticos. Estas regiones pueden además establecer
interacciones entre proteínas con otras funciones. No se conoce actualmente su función
en las proteínas R, pero se está investigando si intervienen en la homodimerización con
otras proteínas durante la activación de la resistencia (Martin, 1999).
e. Genes R que codifican para proteínas con dominios semejantes a los receptores
Toll/Interleucina-1 (TIR) citoplásmicos de células animales. Las proteínas N y L6 forman
un segundo subgrupo de las proteínas con dominios DUN-RRL. Estos dominios se
localizan en el extremo amino terminal de las proteínas R, y tienen una alta similitud con
dominios de señalización citoplásmica de la proteína Toll de Drosophila y los receptores
para interleucina- 1 (IL-1) de mamíferos (Lawrence et al., 1995; Whitman et al., 1994).
La existencia de este tipo de dominio en proteínas R sugiere que podrían activar las
respuestas de defensa por un mecanismo similar al utilizado por las proteínas Toll e IL-
1. Por ejemplo, se ha observado que una de las funciones comunes, tanto de Toll como
de las proteínas N y L6, es activar la producción de especies reactivas de oxígeno
(Hammond-Kosack y Jones, 1996).

El control de la enfermedad Moho gris en mora, producida por el hongo fitopatógeno
Botrytis cinerea, es ineficiente a pesar del uso de productos químicos y de las prácticas
culturales; se hace necesario entonces, buscar herramientas para generar alternativas
tecnológicas que contribuyan al manejo integrado de la enfermedad (Saldarriaga, 2005).
El conocimiento y la caracterización de la diversidad de Botrytis cinerea en Colombia,
así como, la evaluación del comportamiento de materiales de mora de castilla frente al
ataque de este fitopatógeno permitirán la búsqueda e identificación de materiales
promisorios, resistentes o tolerantes al ataque del hongo, que permitan mejorar la
calidad y la productividad de este importante cultivo.
4.3 Objetivos
4.3.1 Objetivo General
Caracterizar morfológica, patogénica y genéticamente el agente
fitopatógeno Botrytis cinerea e identificar genes análogos de resistencia en
diferentes genotipos de mora de castilla.
4.3.2 Objetivos Específicos
Caracterizar morfológica y molecularmente aislados de Botrytis cinerea
procedentes de diferentes zonas productoras de mora en Colombia.
Seleccionar los aislamientos más patogénicos para realizar pruebas de
patogenicidad en nueve genotipos de mora previamente caracterizados.
Realizar un análisis mediante genes análogos de resistencia (RGA) que
permitan la aproximación a posibles fuentes de resistencia en varios
genotipos de mora.
4.4. Metodología propuesta

4.4.1. Para el objetivo específico 1: Caracterizar morfológica y molecularmente
aislados de Botrytis cinerea procedentes de diferentes zonas productoras de mora en
Colombia.
4.4.1.1. Muestreos
Se realizarán muestreos de órganos afectados (botones florales y/o frutos) en las zonas
moricultoras más representativas del país (Antioquia, Santander, Tolima, Valle del
Cauca, Caldas, Quindío, Risaralda y Cundinamarca). La localización de los puntos de
muestreo se realizará conforme al modelo de distribución al azar. Los muestreos de los
hongos permitirán evaluar la diversidad genética y morfológica de los mismos. Estos
resultados se correlacionarán con la capacidad patogénica de los diferentes
aislamientos frente a diferentes cultivares de mora.
Los aislamientos serán conservados en papel filtro y/o liofilizados para asegurar que
conserven su viabilidad para pruebas posteriores.
4.4.1.2. Caracterización morfológica de Botrytis cinerea
Se tomarán secciones de frutos y/o botones florales que presenten mayor esporulación,
de al menos 20 aislamientos de hongos colectados en diferentes zonas productoras de
mora del país. En el Laboratorio de Biotecnología Vegetal de la Facultad de Ciencias
Ambientales de la Universidad Tecnológica de Pereira, serán transferidos a cajas Petri
en medio Papa-Dextrosa-Agar (PDA) e incubados a 23 ºC en oscuridad. Posteriormente
se realizarán cultivos monospóricos para contar con un material puro, en donde las
condiciones genéticas, bioquímicas y fisiológicas sean más homogéneas (Castellanos
et al., 2011). Las características morfológicas se evaluarán en cinco cajas de Petri con
el medio de cultivo antes mencionado, sembrando en el centro de cada caja una espora
del hongo y se utilizarán cinco cajas de Petri por cada aislamiento. Se hará la

evaluación diaria de: diámetro de crecimiento de micelio, distribución de esclerocios y
coloración del micelio.
Crecimiento diario del micelio:
El diámetro de crecimiento diario se realizará desde que se deposita la espora en el
centro de las cajas hasta que el hongo completa su crecimiento al llenar la caja. Para
esto se trazarán dos rectas que se cruzan en el centro de la caja, de esa manera, por
cada placa se tomarán dos mediciones (mm), una horizontal y otra vertical. Con los
datos obtenidos se sacarán promedios diarios del diámetro de la colonia para
determinar la tasa de crecimiento lineal mediante un análisis de regresión (r) para cada
aislamiento. Para determinar diferencias estadísticas entre los valores de crecimiento
diario de los aislamientos se realizará un análisis de varianza (ANAVA), y Prueba de
Tukey (P=0.05%). Se utilizará el SAS Statistical Software, 1989).
Distribución de Esclerocios:
Para cada aislamiento monospórico pasados 30 días aproximadamente, se tomarán
datos de la distribución, coloración, forma, tamaño y profundidad de los esclerocios y
con base a estas características se establecerán los siguientes tipos de distribución
(Tanovic et al., 2009):
E1: Esclerocios encontrados en su mayoría en el centro.
E2: Esclerocios grandes y dispersos.
E3: Esclerocios pequeños numerosos y dispersos.
E4: Sin esclerocios.
El tamaño de los esclerocios se determinará midiendo largo y ancho (mm) de un mínimo
de 50 esclerocios tomados en forma aleatoria de cada aislamiento, utilizando el
microscopio de luz.
Color de micelio:

Será evaluado el día 30, después de la siembra, a través de una carta de color RAL
esta evaluación consistirá en ir comparando el micelio de las cepas con la tabla de
color, una vez recogidos estos datos, se le otorgará a cada color un número según la
intensidad de éste (Sanhueza, 2012 y Villalobos, 2006).
4.4.1.3. Caracterización genética
Se realizará utilizando el protocolo de obtención de ADN genómico DNeasy 96 plant Kit
ref 69181. Para determinar la integridad del ADN cromosomal y la ausencia de ARN, se
realizará una electroforesis en gel de agarosa al 1% con tinción con bromuro de etidio
por 1 hora a 95 voltios, en buffer Tris Borato EDTA (TBE) 1X. Para visualizar el ADN
obtenido los geles serán dispuestos en un transiluminador Biorad Molecular Imager®
Gel Doc™.
Posteriormente, se realizará la verificación genética de Botrytis cinerea, con primers
específicos diseñados para esta especie, para descartar la contaminación con otro
hongo. La caracterización genética con marcadores moleculares microsatélites se hará
a través de la amplificación por PCR de las muestras de ADN de cada uno de los
aislamientos. Serán utilizados nueve primers microsatélites reportados previamente por
Fournier et al., 2002 (Tabla 1).
Tabla 1. Primers microsatelites reportados para Botrytis cinerea
Nombre del Primer Secuencia
Bc1 F:AGGGAGGGTATGAGTGTGTA
R:TTGAGGAGGTGGAAGTTGTA
Bc2 F:CATACACGTATTTCTTCCAA
R:TTTACGAGTGTTTTTGTTAG
Bc3 F:GGATGAATCAGTTGTTTGTG
R:CACCTAGGTATTTCCTGGTA
Bc4 F:CATCTTCTGGGAACGCACAT
R:ATCCACCCCCAAACGATTGT

Bc5 F:CGTTTTCCAGCATTTCAAGT
R:CATCTCATATTCGTTCCTCA
Bc6 F:ACTAGATTCGAGATTCAGTT
R:AAGGTGGTATGAGCGGTTTA
Bc7 F:CCAGTTTCGAGGAGGTCCAC
R:GCCTTAGCGGATGTGAGGTA
Bc9 F:CTCGTCATAACCACGCAGAT
R:GCAAGGTCTCGATGTCGATC
Bc10 F:TCCTCTTCCCTCCCATCAAC
R:GGATCTGCGTGGTTATGACG
Los análisis estadísticos se realizarán con el programa GENAlEX versión 6.2 (Peakell y
Smouse, 2006) se evaluarán los siguientes parámetros estadísticos: medidas de
variabilidad genética, medidas de diversidad genética, medidas de distancia genética.
4.4.2. Para el objetivo específico 2: Seleccionar los aislamientos más patogénicos
para realizar pruebas de patogenicidad en nueve genotipos de mora previamente
caracterizados.
4.4.2.1. Caracterización patogénica
Para las pruebas de patogenicidad se realizará un diseño completamente al azar con al
menos 20 tratamientos (correspondientes a los aislamientos del hongo) y cuatro
repeticiones por tratamiento, cada repetición constará de cinco frutos sanos y en el
mismo estado fenológico. Se realizará inoculando frutos maduros sanos de mora de
castilla (ecotipo San Antonio, que se ha demostrado presenta mayor susceptibilidad al
ataque de hongos, Comunicación personal M. Sc. Alegría Saldarriaga, Corpoica La
Selva).
Los frutos serán desinfectados previamente con hipoclorito de sodio y lavados con agua
destilada estéril. Para su inoculación se utilizará una concentración de 1.2x106

conidias/ml, utilizando para ello la cámara de Neubauer y se empleará el método de
aspersión de frutos. Se conservarán en cámaras húmedas y se realizarán
observaciones cada 24 horas.
Las evaluaciones se realizarán teniendo en cuenta la siguiente escala: 1. Sin síntomas
(fruto sano), 2. Síntomas leves (poca presencia de micelio), 3. Síntomas severos
(abundante presencia de micelio < del 60%) y 4. Afectación total del fruto (muerte o
presencia del micelio > del 75%). Así se seleccionarán dos aislamientos más
patogénicos (los que en menor tiempo alcancen el grado 4 de afectación), los cuales
serán utilizados en las pruebas de patogenicidad sobre 9 accesiones de mora de
castilla previamente caracterizadas genéticamente por el Grupo de Investigación en
Biodiversidad y Biotecnología, para establecer si en alguno de ellos se presenta
tolerancia o posible resistencia.
Evaluación de posible tolerancia
Las pruebas de patogenicidad en las nueve accesiones de mora de castilla se
realizarán siguiendo la metodología antes descrita. Como testigo susceptible se incluirá
el ecotipo San Antonio.
Se realizará un diseño factorial (2 x 10) teniendo en cuenta los genotipos de mora y los
aislamientos del hongo. El criterio para establecer una posible tolerancia al ataque de B.
cinérea será en aquella accesión o accesiones donde haya más del 80% de frutos
sanos (Grado 1) o con síntomas leves (Grado 2) en el menor tiempo, comparado con el
testigo susceptible.
El o los genotipos de mora preseleccionados por su posible tolerancia o resistencia a
B. cinérea serán considerados como material promisorio sujeto a una posterior
investigación en campo y bajo condiciones de inóculo natural antes de estar a
disposición de los agricultores.

4.4.3. Para el objetivo específico 3: Realizar un análisis mediante genes análogos de
resistencia (RGA) que permitan la aproximación a posibles fuentes de resistencia a la
enfermedad
El ADN genómico será usado para la amplificación por PCR usando Genes Análogos
de Resistencia (RGA). Los primers degenerados usados en la amplificación de la PCR y
que han sido reportados en la Familia Rosaceae se muestran en la tabla 2:
Tabla 2. Genes Análogos de Resistencia reportados en la familia Rosaceae
Nombre de primer Secuencia*
F1 GGAGTCGGTAAAACAACAATAG F2 GGDGTDGGNAARACWAC F3 GGDGTDGGNAARACSAC R1 GAGGGCTAAAGGAAGGCC R2 IARIGCIARIGGIARNCC R3 AGIGCHAGNGGNAGNCC R4 AGNGCHAGNGGYAANCC R5 AGNGCYAANGGYAANCC R6 AANGCHAGNGGYAANCC K2F ATCNACATCATCNAGHACRA K2R TYGTKCTNGATGATGTNGA BP2f GGNGGDGTDGGSAARAC BP2r GCTAGTGGCAMNCCWCC
DegRos-f MDTKSBDRRRSSBDTTTWHRMM DegRos-r RKDYWYDHMWHRDWKBWBMWK OLE1121 GGWATGGGWGGWRTHGGWAARACHAC OLE1122 ARNWYYTTVARDGCVARWGGVARWCC
P-loop GAATTCGGNGGNAAGACAAC
* Secuencia codificada de acuerdo con la Unidad Internacional de Bioquímica: D(A, G o T); R (A o G); W
(A o T); Y (C o R); S (G o C); H (A, C o T); M (A o C); K (G o T); N (A, G, T o C); I Inosina (Fuente Xu, et al., 2005, Samuelian, et al., 2008, Afanador et al., 2015)
Se realizará una amplificación directa entre los motivos reportados previamentes por
Xu, et al., 2005, Samuelian, et al., 2008, Afanador et al., 2015 en la familia Rosaceae.
Las accesiones de mora con amplificación positiva determinarán posiblemente la
presencia de genes de resistencia. Para su confirmación se procederá a la
secuenciación de los productos amplificados, posteriormente las secuencias serán
analizadas mediante BLAST en bases de datos públicas, las secuencias que
pertenezcan a los mismos genes serán visualizadas en cluster originados por análisis
Clustal W. Resultado que será contrastado con las evaluaciones de patogenicidad
realizada con los aislamientos más patogénicos. En caso de tener resultados positivos
en la tolerancia o resistencia de las accesiones de mora al ataque de Botrytis cinerea,
este material vegetal servirá de insumo en futuras investigaciones y se promoverá entre
los moricultores, disminuyendo la presión sobre el medio ambiente y aumentado las
posibilidades de comercialización a nacional e internacional.
4.5 Resultados esperados2
A continuación se relacionan los resultados a obtener por cada uno de los objetivos
específicos:
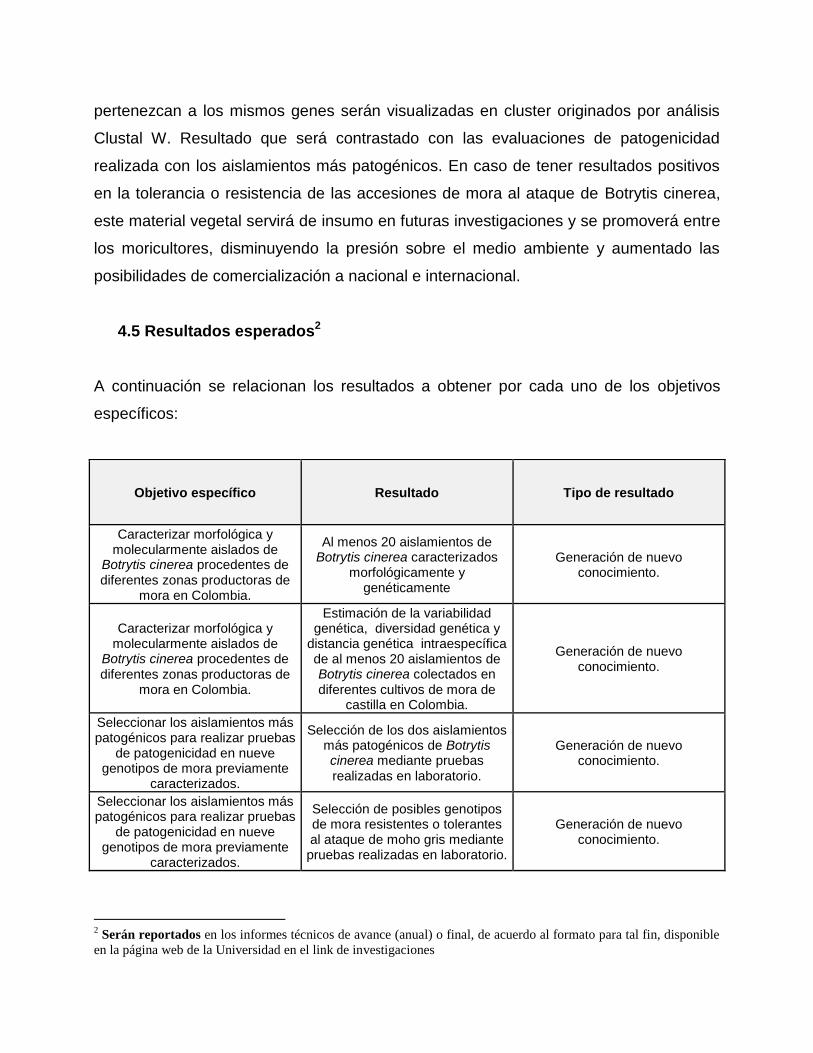
Objetivo específico Resultado Tipo de resultado
Caracterizar morfológica y molecularmente aislados de
Botrytis cinerea procedentes de diferentes zonas productoras de
mora en Colombia.
Al menos 20 aislamientos de Botrytis cinerea caracterizados
morfológicamente y genéticamente
Generación de nuevo conocimiento.
Caracterizar morfológica y molecularmente aislados de
Botrytis cinerea procedentes de diferentes zonas productoras de
mora en Colombia.
Estimación de la variabilidad genética, diversidad genética y
distancia genética intraespecífica de al menos 20 aislamientos de Botrytis cinerea colectados en diferentes cultivos de mora de
castilla en Colombia.
Generación de nuevo conocimiento.
Seleccionar los aislamientos más patogénicos para realizar pruebas
de patogenicidad en nueve genotipos de mora previamente
caracterizados.
Selección de los dos aislamientos más patogénicos de Botrytis cinerea mediante pruebas realizadas en laboratorio.
Generación de nuevo conocimiento.
Seleccionar los aislamientos más patogénicos para realizar pruebas
de patogenicidad en nueve genotipos de mora previamente
caracterizados.
Selección de posibles genotipos de mora resistentes o tolerantes al ataque de moho gris mediante pruebas realizadas en laboratorio.
Generación de nuevo conocimiento.
2 Serán reportados en los informes técnicos de avance (anual) o final, de acuerdo al formato para tal fin, disponible
en la página web de la Universidad en el link de investigaciones
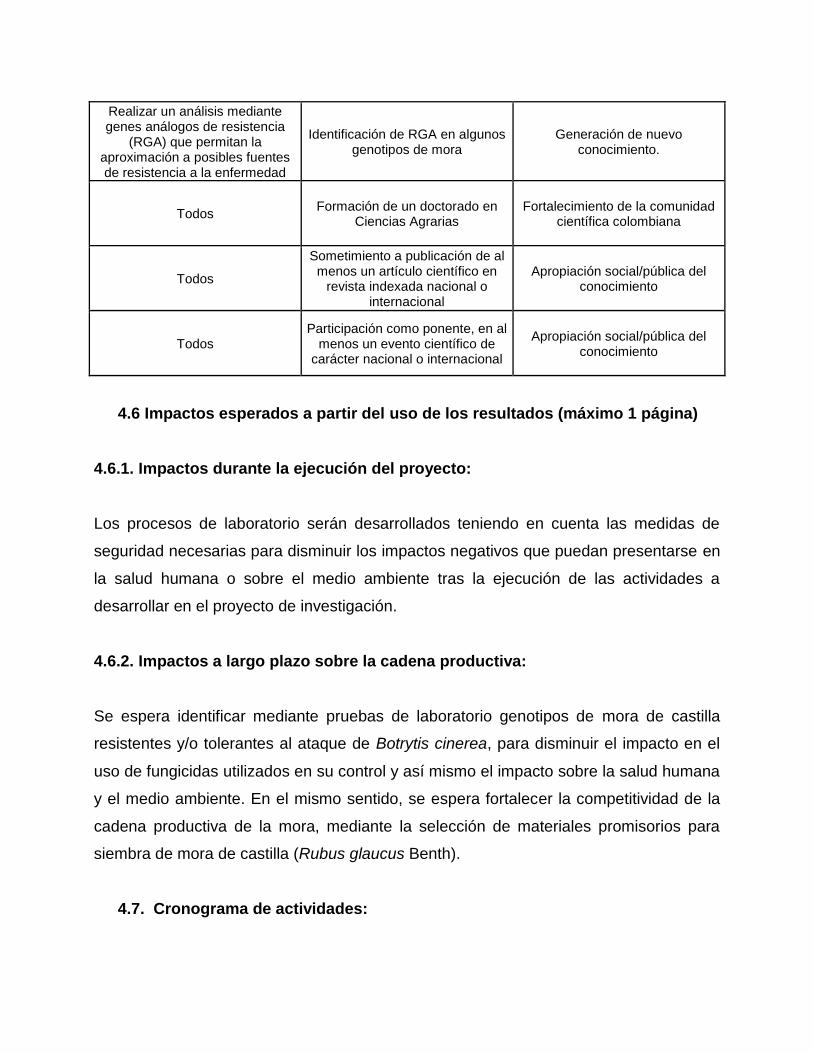
Realizar un análisis mediante genes análogos de resistencia
(RGA) que permitan la aproximación a posibles fuentes de resistencia a la enfermedad
Identificación de RGA en algunos genotipos de mora
Generación de nuevo conocimiento.
Todos Formación de un doctorado en
Ciencias Agrarias Fortalecimiento de la comunidad
científica colombiana
Todos
Sometimiento a publicación de al menos un artículo científico en
revista indexada nacional o internacional
Apropiación social/pública del conocimiento
Todos Participación como ponente, en al
menos un evento científico de carácter nacional o internacional
Apropiación social/pública del conocimiento
4.6 Impactos esperados a partir del uso de los resultados (máximo 1 página)
4.6.1. Impactos durante la ejecución del proyecto:
Los procesos de laboratorio serán desarrollados teniendo en cuenta las medidas de
seguridad necesarias para disminuir los impactos negativos que puedan presentarse en
la salud humana o sobre el medio ambiente tras la ejecución de las actividades a
desarrollar en el proyecto de investigación.
4.6.2. Impactos a largo plazo sobre la cadena productiva:
Se espera identificar mediante pruebas de laboratorio genotipos de mora de castilla
resistentes y/o tolerantes al ataque de Botrytis cinerea, para disminuir el impacto en el
uso de fungicidas utilizados en su control y así mismo el impacto sobre la salud humana
y el medio ambiente. En el mismo sentido, se espera fortalecer la competitividad de la
cadena productiva de la mora, mediante la selección de materiales promisorios para
siembra de mora de castilla (Rubus glaucus Benth).
4.7. Cronograma de actividades:
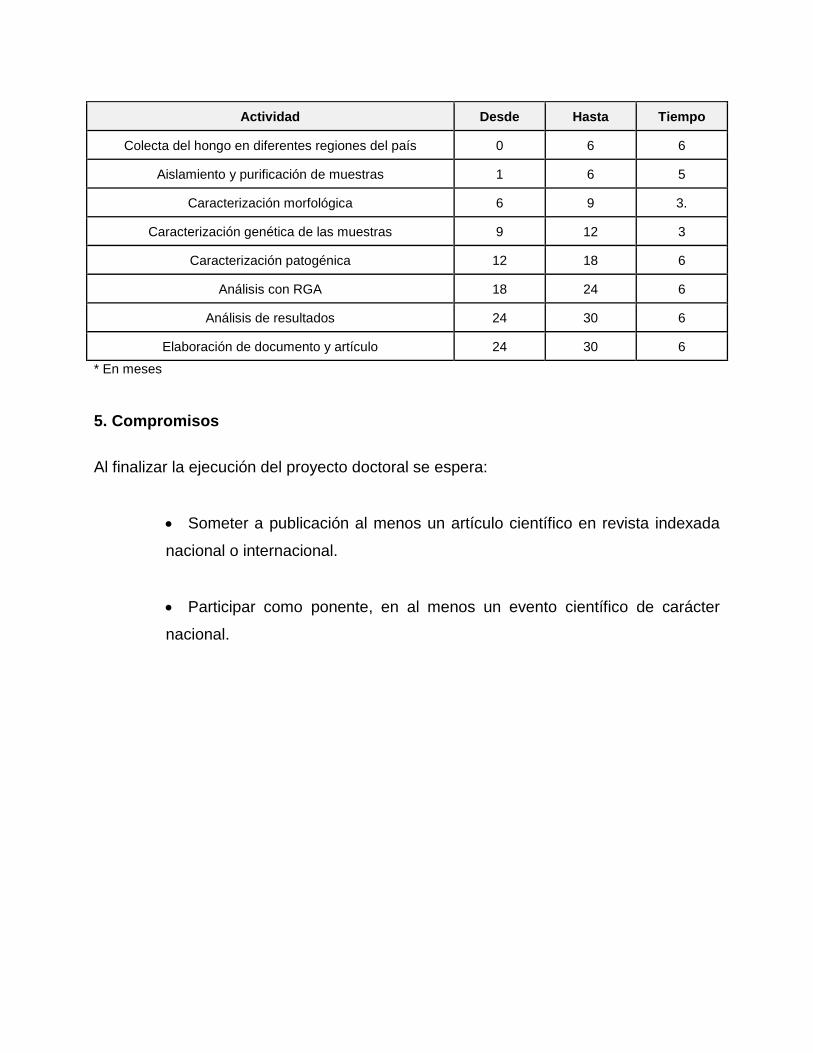
Actividad Desde Hasta Tiempo
Colecta del hongo en diferentes regiones del país 0 6 6
Aislamiento y purificación de muestras 1 6 5
Caracterización morfológica 6 9 3.
Caracterización genética de las muestras 9 12 3
Caracterización patogénica 12 18 6
Análisis con RGA 18 24 6
Análisis de resultados 24 30 6
Elaboración de documento y artículo 24 30 6
* En meses
5. Compromisos
Al finalizar la ejecución del proyecto doctoral se espera:
Someter a publicación al menos un artículo científico en revista indexada
nacional o internacional.
Participar como ponente, en al menos un evento científico de carácter
nacional.

6. Bibliografía
Afanador, L., Mejía, F., González, A., Alvarez, E. 2015. Identifying and Analyzing the
Diversity of Resistance Gene Analogs in Colombian Rubus Genotypes. Plant Disease.
99: 7. 994-1001.
Agrios, G.N. 1997. Plant Pathology. Academic Press, San Diego, California. Fourth
Edition. P. 3,8, 184, 252.
Agrios G.N. 2015. Plant Pathology 5th Edition. San Diego, California. Academic Press
Alvarado A., L. 2002. Mora (Rubus). Consejo Nacional de Producción (CNP). Boletín
Quincenal. Disponible desde 19 Junio / 2002 En:
http://www.cnp.go.cr/php_mysql/admin/KTML/ uploads/files/ boletines/
mora_junio_2002.
Antonov, A., Stewart, A. and Walter, M. 1997. Inhibition of conidium germination and
mycelial growth of Botrytis cinerea by natural products. Proc. 50th. N.Z. Plant Protection
Conf. 159-164.
Benito, D. 2000. Factores de Patogenicidad de Botrytis cinérea Revista Iberoamericana
Micol. 2000; 17: 543 – 546.
Benito, D. 2010. Analisis y Caracterización de genes de Botrytis cinerea, cuya expresión
se induce in planta en la interacción B. cinérea – Tomate. Centro Hispano Luso de
Investigaciónes Agrarias. Salamanca.
Bernal, J. Díaz, J. 2006. Alternativas para el manejo integrado del cultivo de la mora
(Rubus glaucus B) En: seminario, frutales de clima frio moderado. Memorias del IV

Seminario Nacional de Frutales de Clima Frío Moderado. Corpoica, Universidad
Pontificia Bolivariana. Medellín P.75-86
Berrie, A. 1994. The importance of Botrytis cinerea as a storage rot of apple cv. Cox and
pear cv. Conference. Norweg. J. Agric. Sci. S17: 383-389. Horticult. Sci. 115: 102-107.
Botero, O., M. J. y Franco, G. 2007. Identificación y caracterización de la Mancha
necrótica de las hojas de mora (Rubus glaucus Benth), producida por Pseudomonas, en
el municipio de Trujillo (Valle). Revista Corpoica. Ciencia y Tecnología Agropecuaria.
Bogotá, Colombia, 8 (2), 22-25.
Calvo, J.A., Rivera, G., Orozco, S., y Orozco, R. 2012. Aislamiento y evaluación in vitro
de antagonista de Botrytis cinerea en Mora. Agronomía mesoamericana, 23 (2), 225-
231.
Castaño, J. 2015. Principios básicos de hongos Fitopatogenos. Universidad de Caldas,
Centro Editorial. Primera edición.
Castellanos, G., Jara, C. y Mosquera, G. 2011. Guías Prácticas de Laboratorio para el
Manejo de Patógenos del Frijol. Centro Internacional de Agricultura Tropical
Clark, C. y Lobeer, J. 1976. Comparative histopathology of Botrytis squamosa and
Botrytis cinerea on onions leaves. Phytopathology . 66: 1279-1289.
Córdoba, O; Londoño, J. 1996. Evaluación de seis materiales de mora Rubus spp, en
condiciones de clima frío moderado. Trabajo de Grado (Ing. Agr) Universidad Nacional
de Colombia. Medellín.
Correa, L. 2002. Blackberry: A New Crop Option to Brazil. En Ciencia Rural, Santa
Maria, 32: 151 158.

Dangl, J.L, y Jones, J.D.G. 2001. Plant pathogens and integrated defence responses to
infection. Nature 411, 826 – 833.
Dashwood.,E. P y Fox, R. A. 1998. Infection of flowers and fruits of raspberry by Botrytis
cinerea. Plant Pathol., 37, 423-430.
De Bary, A. 1886. Ueber einige Sclerotinien and Sclerotienkrankheiten. Bot. Zeitung 44,
377–474.
De La Rotta; M.C. 2001. Enfermedades de la Mora de Castilla. Instituto Colombiano
Agropecuario, ICA. Bogotá, 3-36.
Departamento Administrativo Nacional de Estadística. DANE. 2011. Resultados
encuesta nacional agropecuaria ENA. Colombia.
Elad, Y. 1988. Involvement of ethylene in the pathogenicity of Botrytis cinerea Pers. on
rose and carnation flowers and the possibility of control. Ann. Appl. Biol. 113: 589-598.
Elad, Y. 1997. Effect of filtration of solar light on the production of conidia by field
isolates of Botrytis cinerea and on several diseases of greenhouse-grown vegetables.
Crop Protection 16, 635-642.
Ellis, J., Doods, P., y Pryor, T. 2000. Structure, function and evolution of plant disease
resistence genes. Current Opinion in Plant Biology 3:278-284.
Ellis J y Jones D. 1998 Structure and function of proteins controlling strain-specific
pathogen resistance in plants. Curr Opin Plant Biol 1:288-293.
EPA. 2005. Pesticide Registration Status of the United States of America. [En línea].
http:// www.cfpub.epa.gov/appref/rereg/status.cfm?show=rereg#c. [Consultada: febrero
de 2015].

Flor, H.H. 1947. Host parasite interactions in flax rust. Its genetics and other implications
Phytopatology 45: 680-685.
Fournier, E., Giraud, T., Loiseau, A., Vautrin, D., Estoup, A., Solignac, M., Cornuet, J. M.
and Brygoo, Y. 2002. Characterization of nine polymorphic microsatellite loci in the
fungus Botrytis cinerea (Ascomycota). Molecular Ecology Notes, 2: 253–255.
Franco, G. 2002. El Cultivo de La Mora. El cultivo de la mora. Corporación Colombiana
de Investigación Agropecuaria, CORPOICA Regional 4. Rionegro – Antioquia. 102 p.
Franco, G. y Giraldo, C. 1998. El cultivo de la mora. Corporación Colombiana de
Investigación Agropecuaria, CORPOICA Regional 9. Programa Nacional de
Transferencia de Tecnología Agropecuaria, PRONATTA. 130 p.
García, E. y Lozoya, E. 2004. Genes de Resistencia a Enfermedades en Plantas.
Revista Mexicana de Fitopatología, vol. 22, núm. 3, diciembre, 20pp. 414-422 Sociedad
Mexicana de Fitopatología, A.C. Texcoco, México.
Gil, M.J, Coego A., Mauch-Mani, B., Jorda, L., y Vera, P. 2005. The Arabidopsis csb3
mutant reveals a regulatory link between salicylic acid-mediated disease resistance and
the methyl-erythritol 4-phosphate pathway. Plant J. 44: 155–166. [PubMed]
Hammer, P.E., Yang, S.F., Reid, M.S. y Marois, J. 1990. Postharvest control of Botrytis
cinerea infections on cut roses using fungistatic storage atmospheres. J. Am. Soc. Hort.
Sci. 115:102-107.
Hammond-Kosack, K., and Jones, J.D.G. 1996. Resistance gene-dependent plant
defense responses. The Plant Cell 8:1773-1791.

Hammond – Kosack. K.E y Parker J.E. 2003. Deciphering plant – pathogen
communication. Fresh perspectives for molecular resistance breeding. Curr. Opin.
Biotech., 14: 177 – 193.
Johnston D.J., Williamson B. 1992. Purification and characterization of four
polygalacturonases from Botrytis cinerea. Mycol Res. 1992; 96: 342- 349.
Kobe, B. y Deisenhofer, J. A. 1995. Structural basis of the interactions between leucine-
rich repeats and protein ligands. Nature. 374: 183-186.
Lawrence, G.J., Finnegan, E.J., Ayliffe, M.A., and Ellis, J.G. 1995. The L6 gene for flax
rust resistance is related to the Arabidopsis bacterial-resistance gene Rps2 and the
tobacco viral resistance gene-N. The Plant Cell 7: 1195-1206.
Lobo, M., Medina, C.I., Delgado, O.A., Zuluaga, M. L., Cardona, M., y Osorio, A.
(2002). Recursos genéticos de frutales andinos en el sistema de bancos de
germoplasma del estado colombiano. En: Memorias IV Seminario Nacional de Frutales
de Clima Frío Moderado. CORPOICA, Universidad Pontificia Bolivariana, C.D.T.F.,
Medellín, Colombia, 43-48.
Martin, G.B. 1999. Functional analysis of plant disease resistance genes and their
downstream effectors. Current Opinion in Plant Biology 2:273-279.
Medina, C., Cristancho, D., y Uribe, D. 2009. Respuesta fisiológica y capacidad
antagonista de aislamientos filosféricos de levaduras obtenidos en cultivos de mora
(Rubus glaucus). Act biol. Colom, 14 (3): 179-196.
Moffett, P., Farnham, G., Peart, J. and Baulcombe, D.C. 2002. Interaction between
domains of a plant NBS-LRR protein in disease resistance-related cell death. EMBO
Journal 21: 4511-4519.

Molina, G. S., De la Rotta, M. C., y Torres, E. 2004. Incidencia de infecciones
quiescentes de Botrytis cinerea en flores y frutos de mora de castilla (Rubus glaucus
Benth). Agronomía Colombiana. 22 (2): 101-109.
Movahedi, S., Heale, J. 1990 b. The roles of aspartic proteinase and endo-pectin lyase
enzymes in the primary stages of infection and pathogenesis of various host tissues by
different isolates of Botrytis cinerea Pers ex. Pers. Physiol Mol Plant Pathol. 36: 303-
324.
Movahedi, S., Heale, J. 1995. Purification and characterization of an aspartic proteinase
secreted by Botrytis cinerea Pers ex. Pers in culture and in infected carrots. Genet. 248:
547-552.
Muckenschnabel, I., Goodman, B. A., Williamson, B., Lyon, G. and Deighton, N. 2002.
Infection of leaves of Arabidopsis thaliana by Botrytis cinerea: changes in ascorbic acid,
free radicals and lipid peroxidation products. J. Exp. Bot. 53 (367): 207-214.
Peakall, R., & Smouse, P.E. 2006. Genalex 6: genetic analysis in Excel. Population
genetic software for teaching and research. Molecular Ecology Notes, 6, 288–295.
Prins, T.W. Wagemakers, L.,Schouten, A., y VanKan, J.A.L. 2000. Cloning and
characterization of a glutathiones – transferase homologue from the plant pathogenic
fungus Botrytis cinerea. Molecular Plant. Pathology 1, 169 – 178.
Reignault, P., Mercier, M., Bompeix, G., Boccara, M. 1994. Pectin methylesterase from
Botrytis cinerea: physiological, biochemical and inmunochemical studies. Microbiology.
140: 3249-3255.
Rosslenbroich, H. y Stuebler, D. 2000. Botrytis cinerea –History of Chemical Control and
Novel Fungicides for its Management. Crop Protection. 19, 557-561.

Rosslenbroich, H. y Stuebler, D. 2000. Botrytis cinerea – history of chemical control and
novel fungicides for its management. Crop Protection, 19, 8-10, 557-561
Saldarriaga, A. 2005. Caracterización del agente causante de la antracnosis en tomate
de árbol, manzano y mora. Tesis de Ingeniería agronómica. Facultad de Ciencias
Agropecuarias. Universidad de Caldas, Manizales, Colombia, 192.
Samuelian, s., Baldo, A., Pattison, J., Weber, C. 2008. Isolation and linkage mapping of
NBS-LRR resistance gene analogs in red raspberry (Rubus idaeus L.) and classification
among 270 Rosaceae NBS-LRR genes. Tree Genetics and Genomes. 4: 881-896.
Sanhueza G. (2012) Determinación de resistencia de diez aislamientos de Botrytis
cinerea Pers. ex Fr.,obtenidos de frutos de arándano (Vaccinium corymbosum L.), a
cinco fungicidas. Universidad Austral de chile. Facultad de ciencias agrarias.
Sasaki, I., Nagayama, H. 1994. b-glucosidase from Botrytis cinerea: its relation to the
pathogenicity of the fungus. Biosci Biotech Biochem; 58: 616-620.
Staskawicz, B. J.; Ausubel, F. M.; Baker, B. J.; Ellis, J. G. and Jones, J. D. G. 1995.
Molecular genetics of plant disease resistance. Science 268:661–667
Stehman, C. & De Waard, M. 1996. Sensitivity of Population of Botrytis cinerea to
Triazoles, Benomyl and Vinclozolin. European Journal of Plant Pathology. 102: 171-180.
Tamayo, M.; Bernardo, G. & Becerra, D. 2002. Enfermedades de la mora de Castilla
(Rubus glaucus, Benth) en Antioquia. En: Boletín Bimestral Ascofi Informa, 28, 16-19.
Tanovic, B., Delibašic,G., Milivojevic, J., Nikolic, M. 2009. Characterization of Botrytis
cinerea Isolates from small fruits and grapevine in Serbia. Arch. Biol. Sci., Belgrade, 61
(3), 419-429.

Van Baarlen, P., Legendre, L. And van Kan, J. A. L. 2004. Plant defense compouds
against Botrytis infection. In Botrytis: Biology, Pathology and Control (Y. Eland, B.
Williamson, P. Tudzynski and N. Delen, eds), pp 143-161. Kluwer Academic Press,
Dordrech, The Netherlands .
Villalobos, C. 2006. Características morfológicas y patogénicas de aislamientos de
Botrytis cinerea Pers. ex Fries desde arándano (Vaccinium corymbosum L. y Vaccinium
virgatum Ait.) resistentes a benomilo . Universidad Austral de chile. Facultad de ciencias
agrarias.
Whitman, S., Dinesh-Kumar, S.P., Choi, D., Hehl, R., Coor, C., and Baker, B. 1994. The
product of the tobacco mosaic virus resistance gene N: similarity to Toll and the
interleukin-1 receptor. Cell 78:1101-1115.
Xu Qiang, Wen Xiaopeng, Xiuxin Deng. 2005. Isolation of TIR and nonTIR NBS–LRR
resistance gene analogues and identification of molecular markers linked to a powdery
mildew resistance locus in chestnut rose (Rosa roxburghii Tratt) Theor Appl Genet . 111:
819–830.
Zapata, J. A., Cotes, A. M., Uribe, L. A., Díaz, A., Villamizar, L., Gómez, M. I.,
Saldarriaga, A., Álvarez, R., & Gómez, E. 2013. Desarrollo de prototipos de
bioplaguicida a base de Rhodotorula glutinis LvCo7 para el control de Botrytis cinerea
en cultivos de mora. Bogotá (Colombia): CORPOICA, 80.
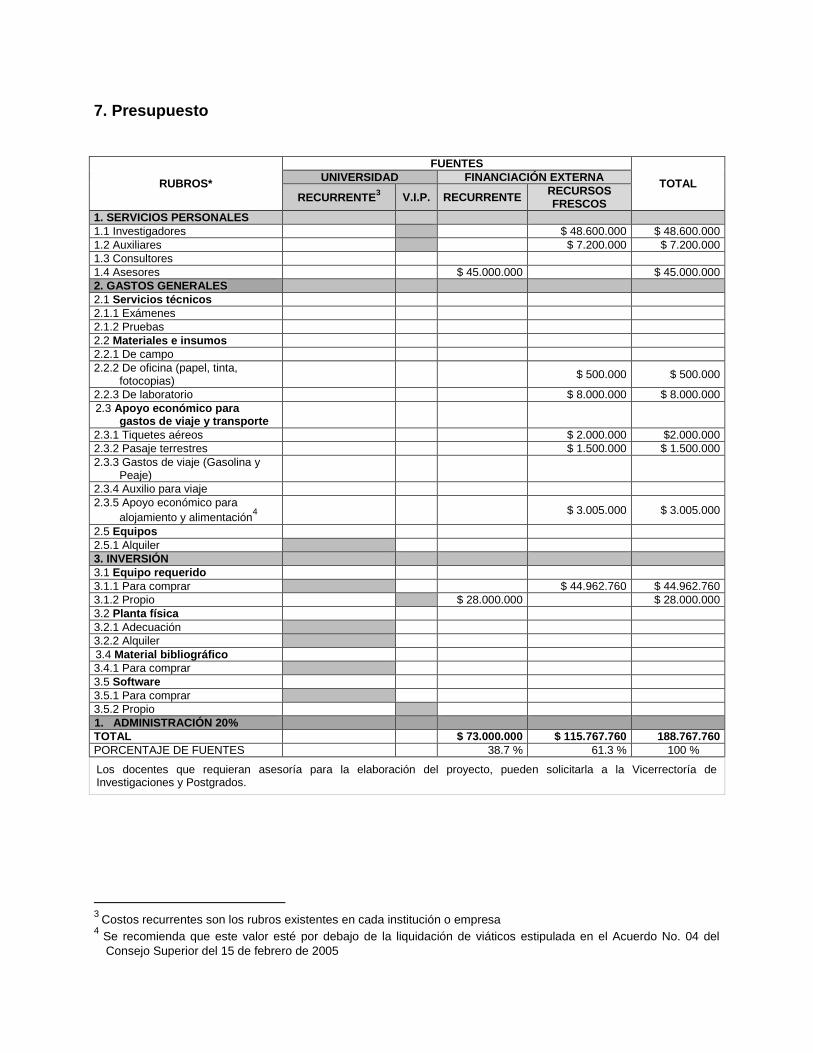
7. Presupuesto
RUBROS*
FUENTES
TOTAL UNIVERSIDAD FINANCIACIÓN EXTERNA
RECURRENTE3 V.I.P. RECURRENTE
RECURSOS FRESCOS
1. SERVICIOS PERSONALES
1.1 Investigadores $ 48.600.000 $ 48.600.000
1.2 Auxiliares $ 7.200.000 $ 7.200.000
1.3 Consultores
1.4 Asesores $ 45.000.000 $ 45.000.000
2. GASTOS GENERALES
2.1 Servicios técnicos
2.1.1 Exámenes
2.1.2 Pruebas
2.2 Materiales e insumos
2.2.1 De campo
2.2.2 De oficina (papel, tinta, fotocopias)
$ 500.000 $ 500.000
2.2.3 De laboratorio $ 8.000.000 $ 8.000.000
2.3 Apoyo económico para gastos de viaje y transporte
2.3.1 Tiquetes aéreos $ 2.000.000 $2.000.000
2.3.2 Pasaje terrestres $ 1.500.000 $ 1.500.000
2.3.3 Gastos de viaje (Gasolina y Peaje)
2.3.4 Auxilio para viaje
2.3.5 Apoyo económico para
alojamiento y alimentación4
$ 3.005.000 $ 3.005.000
2.5 Equipos
2.5.1 Alquiler
3. INVERSIÓN
3.1 Equipo requerido
3.1.1 Para comprar $ 44.962.760 $ 44.962.760
3.1.2 Propio $ 28.000.000 $ 28.000.000
3.2 Planta física
3.2.1 Adecuación
3.2.2 Alquiler
3.4 Material bibliográfico
3.4.1 Para comprar
3.5 Software
3.5.1 Para comprar
3.5.2 Propio
1. ADMINISTRACIÓN 20%
TOTAL $ 73.000.000 $ 115.767.760 188.767.760
PORCENTAJE DE FUENTES 38.7 % 61.3 % 100 %
Los docentes que requieran asesoría para la elaboración del proyecto, pueden solicitarla a la Vicerrectoría de Investigaciones y Postgrados.
3
Costos recurrentes son los rubros existentes en cada institución o empresa 4
Se recomienda que este valor esté por debajo de la liquidación de viáticos estipulada en el Acuerdo No. 04 del
Consejo Superior del 15 de febrero de 2005
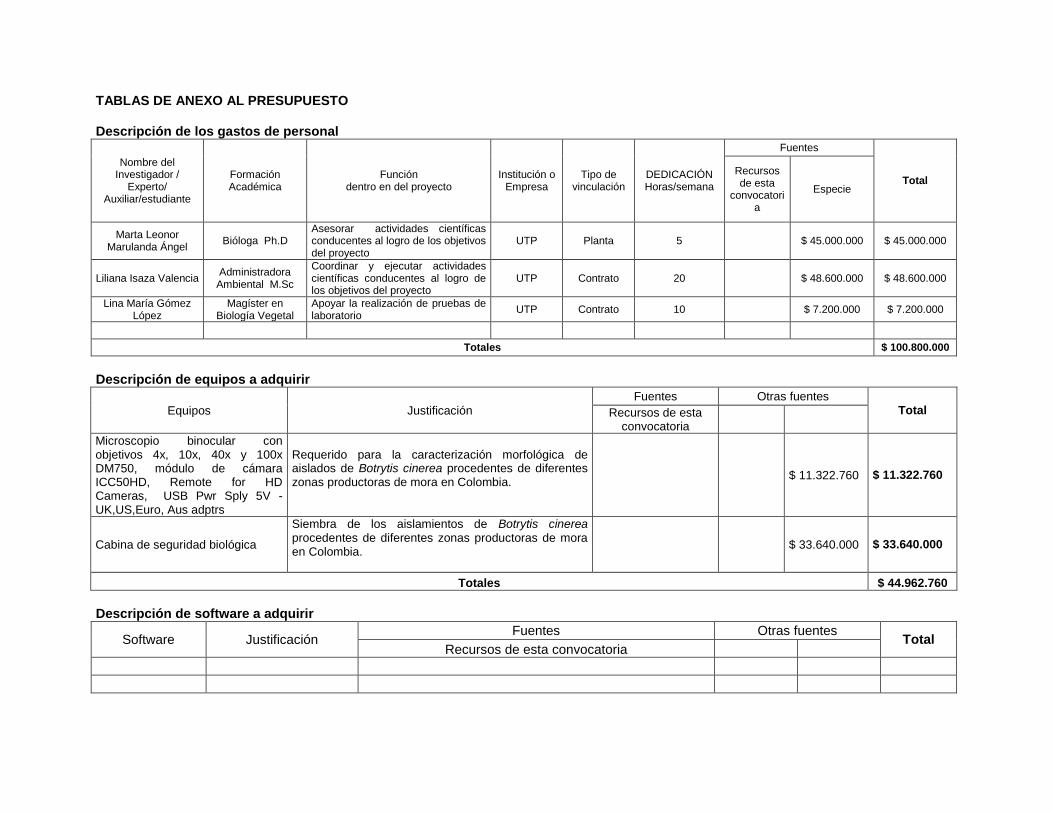
TABLAS DE ANEXO AL PRESUPUESTO Descripción de los gastos de personal
Nombre del Investigador /
Experto/ Auxiliar/estudiante
Formación Académica
Función dentro en del proyecto
Institución o Empresa
Tipo de vinculación
DEDICACIÓN Horas/semana
Fuentes
Total Recursos de esta
convocatoria
Especie
Marta Leonor Marulanda Ángel
Bióloga Ph.D Asesorar actividades científicas conducentes al logro de los objetivos del proyecto
UTP Planta 5 $ 45.000.000 $ 45.000.000
Liliana Isaza Valencia Administradora Ambiental M.Sc
Coordinar y ejecutar actividades científicas conducentes al logro de los objetivos del proyecto
UTP Contrato 20 $ 48.600.000 $ 48.600.000
Lina María Gómez López
Magíster en Biología Vegetal
Apoyar la realización de pruebas de laboratorio
UTP Contrato 10 $ 7.200.000 $ 7.200.000
Totales $ 100.800.000
Descripción de equipos a adquirir
Equipos Justificación
Fuentes Otras fuentes Total Recursos de esta
convocatoria
Microscopio binocular con objetivos 4x, 10x, 40x y 100x DM750, módulo de cámara ICC50HD, Remote for HD Cameras, USB Pwr Sply 5V - UK,US,Euro, Aus adptrs
Requerido para la caracterización morfológica de aislados de Botrytis cinerea procedentes de diferentes zonas productoras de mora en Colombia.
$ 11.322.760 $ 11.322.760
Cabina de seguridad biológica
Siembra de los aislamientos de Botrytis cinerea procedentes de diferentes zonas productoras de mora en Colombia.
$ 33.640.000 $ 33.640.000
Totales $ 44.962.760
Descripción de software a adquirir
Software Justificación Fuentes Otras fuentes
Total Recursos de esta convocatoria
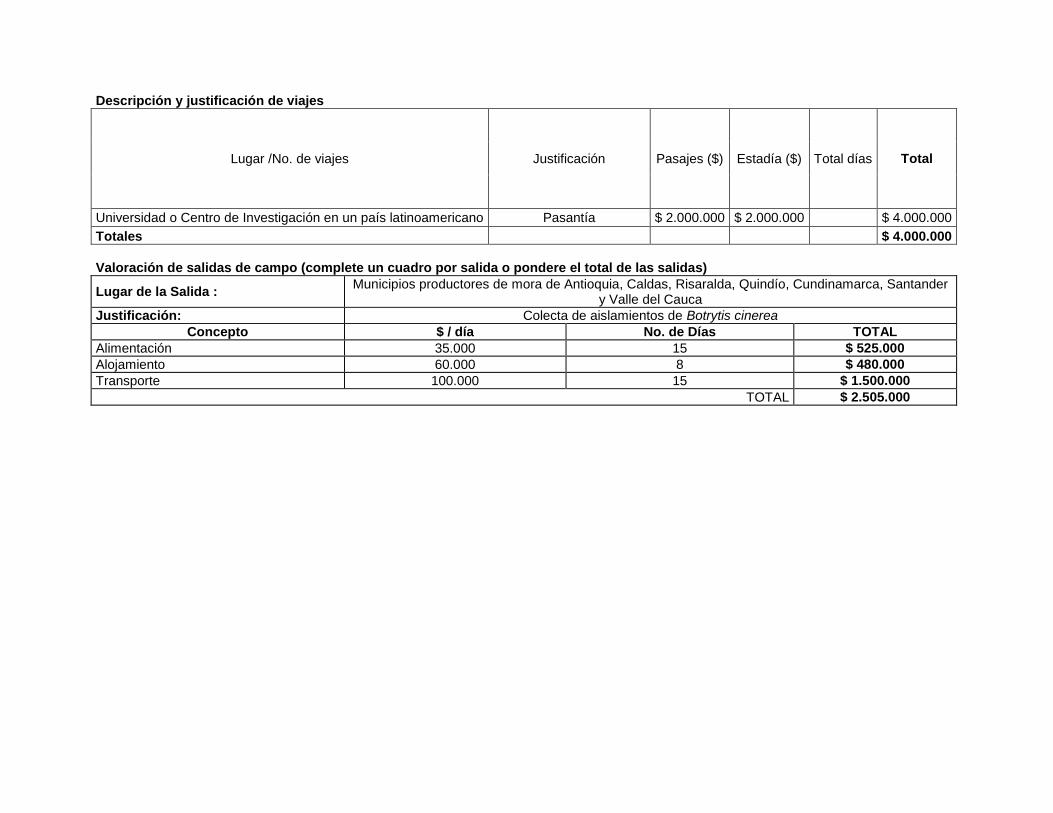
Descripción y justificación de viajes
Lugar /No. de viajes Justificación Pasajes ($) Estadía ($) Total días Total
Universidad o Centro de Investigación en un país latinoamericano Pasantía $ 2.000.000 $ 2.000.000 $ 4.000.000
Totales $ 4.000.000
Valoración de salidas de campo (complete un cuadro por salida o pondere el total de las salidas)
Lugar de la Salida : Municipios productores de mora de Antioquia, Caldas, Risaralda, Quindío, Cundinamarca, Santander
y Valle del Cauca
Justificación: Colecta de aislamientos de Botrytis cinerea
Concepto $ / día No. de Días TOTAL
Alimentación 35.000 15 $ 525.000
Alojamiento 60.000 8 $ 480.000
Transporte 100.000 15 $ 1.500.000
TOTAL $ 2.505.000