u .2otzflr u - biblioteca complutensebiblioteca.ucm.es/tesis/19911996/x/3/x3023901.pdf · u...
TRANSCRIPT

.2otzflrUNIVERSIDAD COMPLUTENSEDE MADRID
FACULTAD DE CIENCIAS BIOLOGICAS
UNIVERSIDAD COMPLUTENSE
INMUNOCOMPLEJOSLA INFECCION POR
CIRCULANTES Y MECANISMOS AUTOINMUNES ENEL VIRUS DE LA INMUNODEFICIENCIA HUMANA
TESIS DOCTORAL
ESPERANZAFEIJOOBLANCO
1994
uuuuuuuuuuuuuu
uuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu

‘U.
Fundación Jiménez DinCLíNICA DE NUESTRA SEÑORA DE LA CONCEPCION
Avda. de los Reyes Católicos. 2 (Ciudad Universitaria)28040 - MADRID
DñaRosaMaría García Delgado, Doctora en CienciasBiológicasy JefeAsociado
del Departamentode Inmunologíade la FundaciónJiménezDíaz
CERTIFICA: que la presente Tesis Doctoral titulada :“Inmunocomplejos
circulantesy mecanismosautoinmunesen la infección por el virus de la
inmunodeticienciahumana”ha sido realizadabajo mi dirección por Esperanza
Feijoó Blancoy reunelascondicionesnecesariaspara optaral Gradode Doctor
en CienciasBiológicas.
Y paraque conste,firmo el presentecertificadoen Madrid, a 29 deJulio de 1994
TELE FO N O544 16 00.5441549 10 07 . 549
uuuuuuuuueuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuueuuuuuuuuuuuu
A-
A,
oc,

uuuuuuuuuuuuuuuuuuuuuuuuueuuuuuuuuu Ami padre,de quiensiempreguardaréun
entrañablerecuerdo.eu Ami madre,por su fortalezay dedicaciénu a sushijos.u• A mis hermanos,por su cariñosoapoyo.uuuuuuuuuuuuuuuuuu

uuuuuuuu Una mañanade Julio de 1994:ueu• Ya Casi he terminado de escribir mi tesis por eso me gustaría expresar mi• agradecimientoa todasaquellaspersonasque,de algún modo,han influido en su realización.u• A Rosa,porhabermeayudadoa descifrarmis resultados,perdidosy desordenadosenu miles de cuadernos,a los queella se refería como malditos,con un gran sentidodel humor.
ioraciaspor todo!.u Al Dr. Ortiz, de quien siemprerecordarésu gran calidad humanay sus clases deu Inmunología; le agradezcosu ayuda durante todo este tiempo que he trabajado en elu Departamento.u• A M~ Angeles Muñoz, por sucolaboraciónen la resoluciónde algunosde los análisisu realizadosen estatesis.
uu Al Dr. Fernández-Guerrero,por su ayudaen la valoraciónclín¡ca de los pacientes.u A Gemma,con quienhe recorridoprácticamentetodoel surde Inglaterrabajoun viento
conocido en aquellaszonascomo galeray una lluvia interminable;a Pepe,a quien siempreu recordaréal oír un ritmo brasileño;a Lorena,por habermeenseñadola cultura bolivianay au Carmen,por todoslos momentosquecompartimosen el laboratorio, quiero agradecerlessu• gran amistady suapoyo incondicionalcomoamigos.uu A Val, por habermehechopartícipede susclasesen los colegiosy haberdespertadomiu interéspor la enseñanza.
A Dolores,por todaslas dosisde “optimina’ y sentidodel humor queme ha ofrecidou duranteestos últimos meses,de quien esperoque escribaunagran tesissobrealguna de las
u miles de hipótesisque se plantea.u• A Ana por mostrarun gran companerismoen todaslas horasde trabajoque hemosu compartido.uu A Eduardo,por haberrealizadotodaslas determinacionesde Inmunoglobulinasqueheu necesitadoa pesarde sus quejaspor el poco volumen de mis muestras,y a todoslos que• compartimosel espaciodel laboratorio6, especialmenteaAurora,porrevisarlasprimerashojasu que escribíde mi tesis,a Pablo,Ana del Amo, Susana,M2 Joséy a Pilar.u
Por último, a todos mis compañerosdel Departamentopor su disposiciónen todou momentoa resolver alguna de mis dudas.Todosestos añosquehe pasadoen el laboratorio• formaránpartede misrecuerdosquecon el pasodel tiemposedesdibujaránal evocarlos,comou escribióPablo Nerudaal describirsusmemoriasen su libro “Confieso quehe vivido”, pero de• los quesiempreme acordarécon grancariño.
uuuuuuuuu

uuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu• Esta tesis ha sido realizada en el Dpto. deu Inmunologíade la FundaciónJiménezDíaz,bajou la d¡rección de la Dra. GarcíaDelgado, con una• ayuda recibida por el Fondo de Tnvestigacfones
Sanitarias(FIS 90/227).
uuuuuuuuuuuuu

uuuuu• Indice
uu Abreviaturas
u 1.- INTRODUCCION 1uu 1.1.- BIOLOGíA MOLECULAR DEL VIRUS 1-11V. 1u 1.1.1.-Filogeniay estructura. 2u 1.1.2.- Organizacióngenómica. 3• 1.1.3.- Interaccióncon la moléculaCD4. 4u 1.1.4.- Ciclo biológico. 6u
1.2.- CARACTERíSTICAS CLíNICAS DE LA INFECCION. 8u 1.3.- ALTERACIONES DEL SISTEMA INMUNE EN EL SiDA. 12
1.3.1.- Disminuciónde linfocitos T CD4+. 12u 1.3.2.- Alteración de los linfocitos B. 15• 1.3.3.- Alteración de los Monocitos. 16• 1.3.4.- Alteraciónde las célulasNK 17uu 1.4.- CONTROL INMUNOLOGICO DEL HTV. 17u 1.4.1.- Respuestahumoral. 17
1.4.2.- Respuestacelular. 18u 1.5.- IMPLICACION DE LOS INMUNOCOMPLEJOSCIRCULANTES
EN LA INFECCION. 19uu• 2.- OBJETIVOSY PLAN DE TRABAJO. 22uuu 3.- MATERIAL Y METODOS. 24
u 3.1.- MATERIAL 24uu 3.2.- METODOS. 25uu 3.2.1.- Análtsis inmunológicosrealizadosparael diagnósticode la infección• y seguimientode los pacientesinfectados. 25uu 3.2.2.- Aislamientode Inmunocomplejoscirculantesdel suerodeu pacientesinfectados:Niveles y concentración. 25u
3.2.3.- Composición:Detecciónde antígenop24 y anticuerposanti-HIV. 26
3.2.4.- Disociaciónde los Inmunocomplejos. 27u• 3.2.5.- Determinaciónde actividad anti-CD4en los Inmunocomplejospor:uu 3.2.5.6.- Unión a unamoléculaCD4 recombinantesoluble. 27u• 3.2.5.7.- Unión a la moléculaCD4 expresadaen extractosde
• membranade una línea linfoblásricaT humanaCD4+. 28
uuuuuu

euuuu• 3.2.6.- Unión de los Inmunocomplejosa linfocitos de sangreperiférica. 31
uu 3.2.7.- Inhibición de la proliferacióniii vitro de linfocitos de sangreperiféricau por Inmunocomplejos. 32
u 3.2.8.- Infección fi vitro de linfocitos de sangreperiféricapor Inmunocomplejos. 34uu 3.2.8.1.- Cuantificaciónde nivelesde antígenop24 en el sobrenadantede los• cultivos celulares. 34uu 3.2.8.2.-Detecciónde ADN proviral integradoen el genomacelularpor• la reacciónen cadenade la polimerasa(PCR). 35uu 3.2.9.- Estudiode la capacidadcitotóxica de linfocitos T CD8+ de pacientes
infectadosfrente a linfocitos T CD4+ medianteensayosde liberaciónde Cr~’. 40
• 3.2.10.- MétodosestadístIcos. 42uuu 4.- RESULTADOSY DISCUSION. 43u• 4.1.-PAPEL PATOGENICODE LOS ICC EN LA INFECCIONPOR MIV. 43u
4.1.1.- Asociaciónentrela progresiónde la infección y las siguientesu característicasde los Inmunocomplejos:
u 4.1.1.1.-Frecuenciade niveleselevadosen suerode pacientesinfectados. 43• 4.1.1.2.-Composición. 46• 4.1.1.3.- Actividad anti-CD4. 54uu 4.1.2.- Potendaciónde la infección por Inmunocomplejos. 65uu 4.1.2.>.- Unión de los ICC a Unfocitosde sangreperiférica. 65u 4.1.2.2.-Implicación de los ICC en la inhibición del crecimientocelular. 71
u 4.1.3.- Capacidadinfectiva de los Inmunocomplejos. 74uu• 4.2.- PRESENCIA DE LINFOCITOS T CD8+ EN PACIENTES INFECTADOS• CAPACES DE LISAR LINFOCITOS T CD4+ ESTIMULADOS CON PHA. 78uu• 5.- DISCUSION FINAL. 86u
5.1.- IMPLICACION DE LOS INMUNOCOMPLESOS EN LA INFECCION. 86u 5.2.- MECANISMOS AUTOINMUNES IMPLICADOS EN LA DEPLECIONu DE LINFOCITOS T CD4+. 90uu• 6.- CONCLUSIONES. 92uu 7.- BIBLIOGRAFíA. 93uuuuuu

euuuuu Abreviaturasuuu AcM Anticuerpo monoclonalu ADN Acido desoxirribonucleico
ADCC Citotox¡cidadcelulardependientede anticuerposu ARN Acido ribonucleico• BSA Albúmina de suerobovinou CDC Centrosde Controlde EnfermedadesdeAtlanta• CLH-Gly Acido clorhídrico-Glicinau CMV Citomegalovirusu CPM Cuentaspor minutou DMSO DimetilsulfóxidodNTP Desoxirribonucleótidostrifosfatou EBV Virus de EpsteinBarru FITC Fluoresceína
• FLS Tamañocelularu HIV Virus de la Inmunodeficienc¡aHumanau HTLV-1 Virus linfotrópicoT humanotipo 1• ICC Inmunocomplejoscirculantesu IL-2 Interleuquina2
u LRFL Intensidadde fluorescenciarojau L9OLS Complejidadcelularu LGFL Intensidadde fluorescenciaverdeu LTR Secuenciasde repeticiónlargasterminales• MI-TC Sistemaprincipal de Histocompatibilidad• NK Células“Natural Killer”• NP-40 Nonidet-p40• OPO Orto-fenildiamina
u PCR Reacciónen cadenade la polimerasaPEL Linfocitos de sangreperiféricaPES Solución salina
u PGL Linfadenopatíageneralizadapersistenteu PHA Fitohemaglutininau PMSF Fenil metil sulfonil fluorurou PE Ficoeritrina• PO Peroxidasau SIDA Síndromede InmunodeficienciaAdquiridau SIV Virus de la inmunodeficienciaen simiosu STF Suerode ternerafetal
SDS Dodecilsulfato sódicou TCR Receptorde linfocitos Tu TRIS Hidroximetil-amino-metanouuuuuuuuuuuuuu

euuuuuuuuuuuuuuuuuuueuuuuuuuuuuuuuuuueuuuuuuuu 1.- INTRODUCCIONuuu
uuuuuu

uuuuuu• 1.- BIOLOGíA MOLECULAR DEL VIRUS DE LA INMUNODEFICIENCIA HUMANA,uuu Fue a finales de 1981 cuandose publicó el primer casode SIDA, a pesarde queen
• aquel momento no se sabía la etiología de esta enfermedad(1). Se trataba de un jovenu
homosexual americanocon neumoníaquepresentabaun estadogravede inmunodeficiencia.
• A partir de esteaño seempezarona observarmáscasosde jóvenes, no solamente
u americanossino también europeos,pertenecientesa dos gruposde riesgo, homosexualesyu• drogadictos,quepadecíanenfermedadesinfecciosaso neoplasiasmuy poco frecuentesen la
• poblaciónnormal,comoSarcomade Kaposi,Tuberculosiso NeumoníaporPnewnocystiscarinhl.
u La evaluacióninmunológicainicial de estospacientesdemostróunamarcadadeficienciauu de la función celular y una pérdidaselectivade linfocitos T CD4+. Comoresultadode estau inmunodeficiencia,estos pacienteseran víctimas de patógenosque podrían ser fadilmente
u controladospor un SistemaInmunenormal. Esteestado,en su forma másavanzada,ha sidou• denominadoSíndromede InmunodeficienciaHumanaAdquirida (SIDA).
uFueron dos investigadores,Robert C. Galio y Luc Montagnier, quienesen 1983u
u demostraronpor primera vez queel agenteque inducía un estadode inmunodeficienciaen el• hombreera un retrovirus(2), denominadoactualmentevirus de la InmunodeficienciaHumanau
(HIV). Se tratabaentoncesde una enfermedaddevastadoracausadapor una clasede agentesu infecciosos, los retrovirus, detectadosno hace mucho tiempo en seres humanoscomou responsablesdeciertaspatologías.uuu El primer retrovirushumanosedescubrióen 1980 y fue denominadoVirus Linfotrópico
• T humanode tipo 1 (HTLV-T) porquese demostróque podía infectar linfocitos T humanos.u
Estevirus causaun tipo de leucemia,conocidacomo Leucemiade células Ten adultos (LAT).
• endémicade algunasáreasdel Japón,Africa y el Caribe, aunqueactualmentese ha extendido
u a otras regiones del mundo. Dos años despuésse descubrió el HTLV-II, implicadou• probablementeen algunoscasosde Leucemiasde célulasT, Leucemiade las célulaspeludasyu algunoslinfomas crcinicos.
u En un principio se pensóqueel agenteetiológico del SIDA podía ser un retrovirusuu relacionadoconel HTLV-I, perocuandoseaislóy se estudiaronsuscaracterísticasmorfológicas• y su genoma,se demostróque estabamásrelacionadocon un grupo de virus citopáticosno
u transformantesdenominadosLentivirus (3,4).uuuu 1uuuuu

uuuuu• El descubrimientodel HIV, como agentequecausabael SIDA, facilitó la apariciónde• una prueba para determinar la presencia de anticuerposfrente al MIV. Los resultadosuu consecuentesa este análisis revelaronque se trataba de una infección con un periodo de• latencia muy largo y queel númerode personasinfectadasera mucho mayor queel de lasU personasquepresentabanun Síndromede Inmunodeficiencia.Esta diferenciaimplicaba queu
existía un estadode pandemia.• La mayoríade las personasinfectadaseran homosexuales,drogadictos,o bien, niños
U nacidosde madresseropositivas.Estasobservacionessirvieron para definir las tresvías deuu transmisióndel virus, sexual intravenosay materno-fetal.u En la población del Oestede Africa, en 1985, se aisló un nuevovirus relacionadocon
U el primer virus de la InmunodeficienciaHumanaaislado,HIV-l, al quese le denominóHIV-2.uEstenuevovirus estabatambiénasociadoa un Síndromede inmunodeficienciasimilar alSIDA,
• aunquemenosagresivo(5).uu
1.1.1.- Filogenia y estructura.e
El origen filogenéticodel virus esdificil de determinardebidoa la gran variación queu• existeentre las secuenciasde nucleótidosde las cepasvirales estudiadashastael momento,
u procedentesde pacientes infectados de diferentes países, y a la carencia de datosu
epidemiológicosanterioresa los primeroscasosde SIDA diagnosticados.u Con los datosevolutivosqueexistenhastaahora,en los quese hancomparadodistintasU secuenciasdel ARN de retroviruscaracterísticosde losprimates,sehasugeridoqueambostipos
de virus, el HIV-l y el HIV-2, podríanhaberevolucionadode un antecesorcomúnhace40 años,u• relacionadocon el Virus de la Inmunodeficienciaen simios, SIV (6).
• Por otro lado, estudiosrealizadospara determinaranalogíasentre las proteínasdeu distintosvirus, basadosen la detecciónde reaccionescruzadasde los anticuerposdel suerode
u los organismosinfectados,hanapoyadola hipótesisde la relacióndel MIV con el virus que
a causaun Síndromede Inmunodeficienciaen los simios (SIV). De hecho,el SIVm,c, responsableuu de una forma de inmunodeficienciay encefalopatíaen los monospresentaunahomología deU un 75% en la secuenciade nucleátidosconel virus HIV-2 (7) y en ambosvirus la organización
u de los genesestructuralesy reguladoresesvirtualmenteidéntica.u
Actualmentesecreequeel HIV-1 se originó en Africa Centraly quesu granexpansiónU pudo habersido debida a la aparicióndel virus en gruposde alto riesgode transmisióncomo
U la poblaciónafricana,los homosexualesy los drogadictosde Europay EstadosUnidos.euu
2
uuuuu

uuuuuu Por microscopiaelectrónicade alta resoluciónse pudo describirla forma casiesférica
• del virión, cuyo diámetrooscila entrelos soy tUI nm. Actualmentese conoceperfectamenteu
la estructuradelvirus, constituidapor unacubiertaexterna,o envoltura,formadaporunadoble• capa de moléculas lipídicas, procedentede las membranasde las células infectadas,y una
U nucleocápsideen el interior. En la bicapa lipídica estánancladasproteínasde origenhumanou
como moléculasdel complejoprincipal de histocompatibilidadde clase1 y de claseII, y lasu glicoproteinasdel virus gpl2O y gp4l, situadasen el exterior e inmersaen la membrana
• respectivamente(8,9). Estasproteínasdela envolturajueganun papeldecisivoen la infecciónu
de suscélulasdiana, los linfocitos T CD4+, ya queestánimplicadasen la entradadel virus a• estascélulas.Debajode la cubiertadel virus seencuentraunacapade proteínamatricial,pl7,u
querodeaal núcleo o cápside,cuyaforma se asemejaa un conotruncadoy hueco.La cápsideestácompuestapor unaproteína,denominadap24, queprotegeel materialgenéticodel virus,
• dosmoléculasde ARN de una longitudde 9.200 nucleótidos,las enzimastranscriptasainversa,
u integrasa,proteasay endonucleasa,u• y las proteínaspó y p7.(Fig. 1.1). 72 -. -. - —
uuuuuuuu• 0Vu• -
•• Fig.1.1.- Estructuradel HTV. §4-.• Uv’ru ~U 1.1.2.- Organización genómica. ‘%—.>
• A pesar dequela estructuradel genomadelHIV-1 es la típíc í queapareceen todoslosu• retrovirus,en cuantoa la presenciade dos elementosde repeticiónlargos en cadaextremoy
U tres genesestructurales,su organizacióngenómicapareceser única entreestegrupo. Esta
u peculiaridadsedebea que,ademásde los genescaracterísticosestructuralesgagpo! y ern-’, losu• cualescodifican las proteínasdel coreo nucleocápside.la transcriptasainversay las proteínas
• de la envoltura respectivamente,presentaotros genesespecíficos(uit, rey, uzef vif vpr, vpu)u (HIV-1) o (qn> (HIV-2). implicadosen distintasfunciones.(Fig. 1.2).
uuu 3uuuuu

uuuuuu ‘CI ‘ev
u Eu 1 vp& ‘o’
¡ E EKi
uuuu Fig. 1.2.- Genomadel HTV. La escalanuméricainferior representael tamañode los• genesen Kiiobases.uu
Al inducir mutacionesdirigidaso delecionesen el ARN delvirus, seha demostradoque• los genes&zt, rey y nef jueganun papelimportanteen la regulaciónde la expresiónviral. Losuu genes&zt y rey, son genestransactivadoresquecodificanproteínasesencialesparala regulación• transcripcionaly post-transcripcional(10).u El gennefcodificaa unaproteínaasociadaa la membranade27 Kd que funcionacomouu un reguladornegativode la transcripción(11). En el ensamblajede las proteínasde los nuevos• virionesestá implicadaunaproteínade 23 Kd codificadapor el genvhf, cuyamutaciónno afectau a la replicaciónviral sino quealtera la infectividad de los nuevosvirus (12).
u El gen vpr codifica un proteína reconocidapor anticuerposdel suerode personasu• infectadas,pero su función es aúndesconocida(13). Respectoal genvpu se ha comprobado
• codificaunaproteínaaisladaen muchasvariantesdel HIV-1, perono del HIV-2 ni delSIV, queuu pareceestar implicada en el procesode salidade virionesde las célulasinfectadasal exterior• (14).u 1.l.3. Interaccióncon la moléculaCD4.uu• El virus interaccionacon su célula diana mediantela unión de la glicoproteinade lau envolturaviral, gpl2O, con la molécula C04, expresadaen la superficie de los linfocitos Tu• cooperadores(15). Estamoléculaestá presente,en concentracionesaltas,en la superficie de• estasubpoblaciónlinfocitaria, pero tambiénapareceen unamenorproporción en monocitos,u
macrófagosy célulasdendríticas(16) y por experimentosiii varo, seha demostradoqueel111Vu tambiénpuedeinfectar estascélulas.Ante la capacidaddel virus de infectar macrófagosseha
• postuladola hipótesisdequeestascélulaspodríanactuarcomoreservoriosdelvirus, yaqueésteu
no presentaun efectocitopático sobreellas,y su infección explicaríala presenciadel virus en
u otrasregionesdel organismo,entreeUasel cerebro,dondepodíaserel responsablede algunas
u de las patologíasneurológicasqueaparecenen algunospacientesseropositivos(17).uuu 4uuuuuu

uuuuu• Al utilizar una forma soluble de la moléculaCD4 se determinóque la afinidad que
u muestrantanto el MIV-] como el HTV-2, por su receptor,es muy elevada,del ordende lfr9Mu• (18) y conanticuerposmonoclonalesdirigidos frentea la moléculade CD4 sedemostróquela
• unión entreeJvirus y la céluladianaseproducíaporla interacciónde la proteínagpI2Ocon unau regiónde la moléculaCD4, Vi, localizadaen el dominio terminal (19).uu• Estudiosmásespecíficoshandemostradoquela interacciónparecelocalizadaentrelos
u residuos413 y 456 de la proteínagpl2O. Estefragmentoincluye tresregionesdiscontinuasqueu• sesituaríanen unazonamuy próxima en el espaciodebidoal plegamientode la proteína.De
• este modo, se establecenmúltiples interaccionescon ciertasregionesde la molécula CD4,u localizadasentre los residuos37-53, en la zona variable VI, y otra posible región en V2u• (20).(Fig. 1.3).uu Fíg. 1.3.- Esquemade la estructura de la moléculauu CD4, una glicoproteína de 433• aminoácidos con cuatro dominios
u extracelulares. La región entre losu• aminoácidos 16 y 159, comprendelos
• Domain principalessitios de unión con la proteínau 3
• gp 120 del HIV y los Anticuerposu monoclonalesOKT4a, Leu3ay OKT4f.uuuuuuu Respectoa la zonade interaccióncon CD4 en la proteínagp 120, seha demostradoqueu la regiónvariableV3 de estaproteínaes críticaparaquese produzcala infección. Mutaciones
u en estaregión modifican el tropismocelular del virus, generandoseviriones con una reducidau• afinidad,y alterantambiénla capacidadde formación de sincitios in vitro (21, 22).
• Aunqueanteriormenteno habíaevidenciasde unaalteraciónen la regiónV3 de gp 120
u durante la entrada del virus, algunas observacionessugirieron que esta región podíau• interaccionarcon unamoléculaaccesoriade la superficiecelularconactividad proteasa.Como
• modelodeestasmoléculassepropusounaproteínade la superficiecelularsimilar ala tripsina,uu denominadaTL2 (23).uuu 5uuuuu
84
16
HIV VP120 blndíngOKT4AL,u 3AOKT4F
¶30
1 59
303
Domain2
DomaIn
408 ser415 sar
433COOH
Donisin4

uuuuu• A pesarde la variabilidad del virus, en la región V3 de gpl2O de diferentescepasde
u 111V y SIV se handetectadorepeticionesde dipeptidosaltamenteconservadas,RP, KP y GPu• (24). Estasregionesestánimplicadasen la especificidadquepresentala dipeptidil peptidasaIV
• (DPPIV), denominadaCD26 (25).
• Dehecho,recientementesehademostrado,medianteensayosexperimentales,queCD26u• actuacomocofactorde CD4 debidoasu interacciónconla región V3 de la proteínagpl2O. En
• estos ensayosse utilizó un Anticuerpomonoclonal(AcM 1F7), dirigido frente a CD26, que
inhibió, en un 89%, la entradadel HIV a las célulasdiana(26).uTambién se ha observadoqueproteínasde la superficiecelular puedenunirsea gp4l
• y como consecuenciade este reconocimientose ha sugeridoquesu unión condicionaríala
u entradadel virus, aunquesu papelen la infección aúnno estáclaro (27).uu• 1.1.4.- Ciclo biológico.
u El ciclo biológico comienzacuandounapartículaviral seune a la superficieexternadeu• su célula diana. La unión se producepor Ja interacciónde la molécula CD4 de un linfocito T
• conla proteínagp 120 delvirus (28). Iii viti-o seha demostradoquedespuésdel reconocimientou
de la célula diana,el HIV no penetraen el interiorde la célula por un procesode endocitosis
u mediadopor su receptor.Este mecanismoes utilizado por otros retrovirus que reconocen
u moléculasespecíficasde la superficiecelular.En el casodel1-11V, la entradase produceporunu• proceso independientede la acidificación de los endosomas(29). Además, mutaciones
u provocadasen la región interna de CD4, supuestamenteresponsablede la internalizacióndel
u virus, no inhiben la infección (30).u
Estosargumentosfueron decisivosparasugerirun mecanismode entradadel virus de
• la Inmunocieficienciahumanadiferente en el que, despuésde la interaccióngpl2O-CD4,seu produceuna fusión de la membranadel virus y de la célula. En esteprocesose creetambién
• quela protefnagp4l del virus juegaun papelesencialy seha sugeridoun modeloen el queuna
u vezqueseha producidola interacciónde gp 120con CD4 seproduceun cambioconformacional
u en la proteínagpl2O quepermitequeel extremohidrofóbicoaminoterminalde laproteínagp4lu• se inserteen la membranacelular, iniciandoel procesode fusión (31).
• Su ciclo biológico recuerdaal ciclo característicode todos los retrovirus porquese
u puedendistinguirlas mismasetapasen suciclo biológico.La primeramitad delciclo comprendeuu cuatro fases:uniónal receptor,entrada,transcripcióninversae integración,y la segundamitad,
U cinco: transcripción,procesamientodel ARNm, síntesis de proteínas,ensamblajede lasu partículasvíricasy procesode gemación(Fig. 1.4).
uu
6uuuuu

uu•• (o >0• \\ ,/‘
u onu,u‘7
• ,0 ou 1
— Geron,,c RNa•DNAu —,Q Nondidu DNA
e U¡’n’ej,We~ 00$•ue• Fig. 1.4.- Ciclo celulardel MIV ¾>
uu
Cuandose producela fusión del virus con la membranacelular, la nucleocápsideviralU se introduceen el interior de la célula y en el citoplasmatiene lugar la liberacióndel ARN y
u su transcripcióna unadoblecadenade ADN. La transcripciónla lleva a cabola transcriptasauu inversadel virus, complejo enzimático.característicosolo de los retrovirus.formadopor tres• enzimas,unapolimerasa,una ribonucleasay una integrasa.u
Aunquela mayoríade las moléculasde ADN transcritaspermanecenen el citoplasma• de la célula hu¿sped,algunasde estasmoléculasson transportadashacia el núcleo, dondese
• circularizane integranen el genomacelular.En esteprocesointervienela integrasadelvirus.u A partir de esemomento,el ADN vírico integrado,denominadoprovirus, seduplicará
u conjuntamentecon los genesde la célula cada vez que ésta se divida. Esto significa que de
• alguna forma, la eliminación del virus del organismosuponela destrucciónde las célulasu infectadasy que una personainfectada, tarde o temprano,puede sufrir las complicaciones
u clínicas asociadasal SIDA, debidoa la capacidadde estevirus de permaneceren un estadou latenteduranteun largo periodode tiempouu Comoconsecuenciade la capacidaddel virus parapermaneceren un estadolatente,lau producción de nuevaspartículasvirales ocurre esporádicamentey solo en algunascélulas
u infectadas.Iii viti-o seha demostradoque la replicacióndel ADN-proviral tiene lugardespuésu
de la activación de la célula, ya que la estimulación de células infectadascon mitógenos,
• antígenoso poruna exposiciónalogénica,conducea la generaciónde nuevosvirionescapaces
de producir un efectocitopáticoen las células de estoscultivos (32). En relacióncon estosu• resultados experimentales,se ha observadoque una infección productiva en linfocitos
u mantenidosen cultivosestá favorecidaal estimularsu proliferacióncon Interleuquina2 (IL-2)u
y Fítohemaglutinina(PHA) (33).uue 7uuuuu

uuuuu• La transcripcióndelADN-proviralcomienzacuandociertosnucleátidosde lassecuencias
u - LTR son activadospor la unión de enzimasnucleares,como NF-kB, producidasdurantelau
activaciónde lascélulasinmunitariasparafomentarsu propiatranscripción(34). Sonproteínas
U reguladoraspresentesen prácticamentetodaslascélulashumanasque intensificanla actividadu
transcripcionalde muchosgenes.Otros mecanismos,observadosin viti-o, que inducenuna• replicaciónviral. estánrelacionadoscon la infecciónpor otrosvirus, comoAdenovirus,Herpes
• simple(35). o el virus de la HepatitisB. Esteúltimo, estimulala infecciónen cultivos mediante
u la unión de la proteínaX, un trans-activadortranscripcional,a las regionesLTR del virus (36).u• Algunasdelasmoléculasde ARN transcritasformaránel materialgenéticodeunanueva
• generaciónde virus y otrasactuaránde mensajeros,traduciéndoseen las distintasproteínasyu enzimasespecíficasdel virus. El ensamblajede las nuevaspartículasvíricas tiene lugar a nivel
• de la membranacelulary en la maduraciónde los nuevosvirionesestádirectamenteimplicado
• el procesode gemación,en el cual, durantela salida de los viriones, partede la membranau celularse utiliza paraformar la envolturade los nuevosvirus.
u• 1.2.- CARACTERISTICAS CLINICAS DE LA INFECCION PORHIN’.u
La infeccióninicial en un individuo puedeserclínicamentesilenteo presentarun cuadro• transitoriode fiebre, linfadenopatía,farinízitis o manifestacionesneurológicas(37). Estecuadro
u generalmenteremiteal mismo tiempoquesedetectauna respuestade anticuerposen el suerou
del paciente.
• Posteriormentela infección puedeentraren una fase latente o persistircon unabaja
• replicaciónviral duranteaños,antesde queel SistemaInmuneestélo suficientementedañadou• paraqueaparezcaunasintomatología.
• Entrelasalteracionesneurológicasmásfrecuentesobservadasen lospacientesinfectadosu
por HIV destacala demenciainducidapor el propiovirus. En estospacientesse ha observado• unamarcadadisfuncióncognitivay una memoriadeteriorada.Mediantecultivoscelularesseha
U sugeridoqueel virus puedereplicarseen el cerebro,particularmenteen célulasmicrogliales.
u Este tropismo que muestrapodría explicar estecuadroneurológico (38). Aunque tambiénu• existenevidenciasde quemecanismosindirectos,comola producciónde factoresneurotóxicos,
• podríanestarimplicadosen algunasde las alteracionesneurológicasobservadasen la infecciónu ,. -
(39). Una delascaracterísticasclínicasmasllamativasen los pacientesinfectadosesla apariciónu de infeccionesoportunistasy tumoresdespuésde quesu SistemaInmuneha sido severamente
• comprometido(40,41). Estasmanifestacionesclínicas son las que definen un Síndromedeu
InmunodeficienciaAdquirida en estospacientes.uu• 8uuuuu

uuuuu• Una de las infeccionesoportunistasmásfrecuenteses una neumoníaproducidaporu
Pneurnocystiscarinil, cuya frecuenciaen pacientesconSIDA essuperiora la quesedetectaen• pacientescon otro tipo de inmunodeficienciacelularT. Aunqueel tratamientocon antibióticos
U puededisminuir la mortalidaddebida a esta infección en estospacientes(42>, existen otrasu
infecciones problemáticas,producidaspor Toxopk¡smagondii, C¡yptococcusneofomzanso
• c’ytornegalovirus,responsables,en algunasocasiones,de alteracionesen el SistemaNervioso
• Central (SNC), que requierenuna terapiamáscompleja.
Algunosagentesantiviralesutilizadosparael tratamientode algunasinfecciones,comou• el DUPO (1,3-dihidro-2-propoximetilguanina),suministrado a pacientes infectados por
u Cytomegalovirus,puedencausaruna gran citotoxicidad en la médula óseay de esta manerau• puedencontribuir al agravamientode su estadode inmunodeficiencia(43).
U Otras infeccionesfrecuentesson las producidaspor C~ptosporidiumo Mycobacterium
u av¡um-intracelulare,asociadasa un cuadro de diarrea severa y una enfermedadpulmonaru• respectivamente.
• Aunquemuchasde las infeccionesquepadecenestospacientespuedensertratadas,a
medida que empeorasu estado de inmunodeficiencia,aparecenotras infecciones.Estasufrecuentementecausansu muerte (44), incluso aunquealgunas de las infecciones estén
• producidaspor patógenosde baja virulencia.
Las infeccionesque padecenson el resultadode la interacciónentre las alteracionesu• inmunológicasdel pacienteinfectadoy el ambientemicrobiano en el quevive. De estemodo,
• la frecuenciarelativade algunasinfeccionesen los pacientescon SIDA varía en las distintasu partesdel mundo.Por ejemplo,en Africa, la neumoníaproducidapor Pneunzocystiscarinil es
• muy poco frecuente,mientrasqueenfermedadesgastrointestinalese infeccionescausadaspor
u Alycobacteriumtuberculosisy Q>ptococcusneofonnans son más frecuentes(45). Otro ejemplou
seríala infección por Isosporabelil en Haití.
• Ademásdepadecerinfeccionesoportunistas,producidasporvirus, bacteriaso parásitos,
u algunospacientesinfectadosdesarrollantumorescomo el Sarcomade Kaposi, linfomas B deu
alto grado o carcinomasde células escamosas(46). El Sarcomade Kaposi es una alteración
• proliferativa multicéntrica del endotelio linfático, que aparececon mayor frecuenciaen los
u pacienteshomosexualescon SIDA. Esta diferenciacon respectoa otros pacientescon SIDA,u• pertenecientesa otros grupos de riesgo, es un rasgo epidemiológico que aún permanece
U inexplicable. Se ha relacionado con varios cofactores, especulándoseque podría estar
u relacionadocon la infección por CMV, e> uso de nitritos, drogas. factoresgenéticos,e inclusou• con otrasenfermedadescíe transmisiónsexual.
uuu 9uuuuu

uuuuu• Se ha sugeridoque las lesionesdel Sarcomade Kaposipodríansercausadasen parteu
por factores solubles producidospor células endoteliales.Estas células estaríana su vezsometidasa unaestimulaciónpor factoresdecrecimientosintetizadospor célulasT infectadas
• por retrovirus(47).
También son frecuenteslos Linfomas B de alto grado, la mayoría extranodularesy deu
localizaciónen el SistemaNervioso Central(48).
uLos pacientesinfectadospuedendesarrollartambiénunavariadaseriede alteracionesu
• hematológicas,comotrombocitopenia(49), u otrascitopenias,algunasde las cualespodríanser
• el resultado,al menosen parte,de la producciónde anticuerposcitotóxicos (50).uu• Respectoa la infecciónpor MIV en niños, ésta difiere de la infección queapareceen• adultosy generalmentesecaracterizaporun gravedefectohumoral,responsablede unamayor
incidenciade infeccionesbacterianasen los niños infectados(51).uu• La última revisión del Sistemade Clasificación de la Infección por MIV, realizadaau
finales de Diciembre, en 1993, por los Centros del Control de EnfermedadesAmericanos• (CDC), incluye trescategoríasclínicasen la infección (52):
eu
* CategoríaA: Infección asintomática,primoinfección o POL.u Incluye uno o varios de los criterios enumeradosa continuaciónen un adulto o
• adolescente(~ 13 años)infectadopor HIV:uu• - Infección MIV asintomática.u
- Linfadenopatíapersistentegeneralizada(PGL).
- Primoinfecciónsintomática.u• * CategoríaB: Infección sintomática,excluidaslas categoríasA o C.u• Manifestacionesclínicasen un adultoo adolescenteinfectadopor HIV, no incluidasen
• la categoríaC, y que respondenal menosa unade las condicionessiguientes:uu• - Estar ligadasal 1-11V o ser indicativasde un déficit inmunitario.
• - Tenerunaevolución clínica o requerirun tratamientoquesevea complicadou• por la infección por MIV.uu• 10uuueu

uueuu• Algunasdelas enfermedadesqueformanpartede estacategoríase citanacontinuación:
e - Angiomatosisbacilar.u• - Candidiasisorofaríngea.
• - Candidiasisvulvovaginalpersistente.e
- Displasiadel cuello uterino,carcinomacervicalin situ.• - Síntomasgeneralescomo fiebre (38,5~ C) o diarreasevera.
U - Leucoplasiavellosa de la lengua.u
- Herpeszósterrecidivante.
• - Púrpuratrombocitopénicaidiopática.
• - Neuropatíaperiférica.uu• * CategoríaC: Enfermedadesindicativasde SIDA.
u Cuandouna personainfectada presentaalguna de las enfermedadesincluidasen lau• siguientelista, quedaclasificadadefinitivamenteen la categoríaC:
U - Infeccionesbacterianasmúltiples en niños > 13 años.
• - Candidiasisde los brónquios.tráqueao pulmones.u
- Candidiasisesofágica.
• - Cáncer invasivo del cuello del útero.
• - Coccidioidomicosisdiseminadao extrapulmonar.u• - Criptococosisextrapulmonar.
U - Criptosporidiosisintestinal con diarrea.
e - Enfermedadpor citomegalovirus,no del hígado,bazoo ganglios.u• - Retinitis por citoniegalovirus(con pérdida de visión).
• - Encefalopatíarelacionadacon el HIV.e
- Herpessimple: úlceracrónica,bronquitis, neumonitis.
• - 1-listoplasmosisdiseminadao extrapulmonar.
U - Isosporidiasisintestinalcon diarrea.e
- Sarcomade Kaposi.• - Neumoníaintersticial linfoide en niños.
• - Linfoma de Burkitt.
e - Linfoma inmunoblástico.u• - Linfoma primario del cerebro.
- Infección diseminadao extrapulmonarpor Mycobacteriurnavium.u
- Infección pulmonarporMycobacteriurntuberculosis.
uu• 11euuuu

euuuu• - Infección extrapulmonarpor Mycobacterium tuberculosb.
• - Neumoníapor Pneurnocyvtiscarinil.u
- Neumoníarecurrente.
• - Leucoencefalopatíamultifocal progresiva.u
- Septicemiarecurrentepor Salmonella(no tifoidea).• - Toxoplasmosiscerebral.
• - Síndromeconsuntivodebidoal 111V.eu• Cadauna de las trescategoríasha sido subdividida en otrastres, Al, A2, A3, Bí, 82,• 83. Cl, C2 y C3, segúnel númerode linfocitos CD4+ circulantes:
- 1. (=500) células¡ plu• - 2. (200-499) ‘ 1”• -3.(=200) ‘ 1”u Estaúltima revisión ha incluido comocasosde SIDA todosaqueuossujetosinfectados
• quepresentenmenosde 200 linfocitosT CD4+ ¡pl. Esteúltimo criterio no hasido aúnaceptado
por los paiseseuropeos,en los quepara diagnosticarun casode SIDA es necesarioqueele
pacientepresentealgún cuadroclínico asociadoa estesíndromede inmunodeficiencia(53).
u• 1.3.- ALTERACIONES DEL SISTEMA INMUNE EN EL SII)A.uu• 1.3.1.- Disminución de Linfocitos T CI)4+ en la infección por HIN’,
ue
Actualmenteno hay ninguna dudaacercade queel rasgomáscaracterísticodel SIDA• es la reduccióndel númerode linfocitos T CD4±circulantes.Sin embargo,aún no existeun
acuerdorespectoa cómo el 111V conducea la destrucciónde estascélulas, críticasparaelefuncionamientodel SistemaInmune.
• La explicaciónmássimpleparacomprenderla pérdidade estoslinfocitos seríael propio
efectocitopáticodel virus, en el quese destruyenlas células infectadaspor la generaciónyu• liberación de las nuevaspartículasviricas (54). Además, también se ha demostradoque la
• acumulacióndeADN viral en el citoplasmade unacélula infectadaconducea la muertecelulare (55).Otromecanismoimplicadoen la eliminaciónde los linfocitos T4, observadoin viti-o, sería
• la formación de sincitios, células gigantesmultinucleadas,formadaspor la unión de células
• infectadas,conantígenosviralesen susuperficie,y no infectadas,mediantela interacciónde lae• proteínagp 120 viral y la moléculaCD4 (56).
uu• 12uueuu

uuuuu• Porotro lado,la respuestainmunecelular, mediadaporLinfocitos T citotóxicos,CTLs,
• como mecanismodel organismoparacontrolar la infección, tambiénconduciríaa unamuerteu
directa de los linfocitos T CD4±infectados.De hecho, in vitro, se ha observadola lisis de
• célulasquepresentanproteínasvirales ensu superficiepor CTLs específicos(57). Por lo tanto,
estoslinfocitos citotóxicos, ademásde controlar la infecciónviral, es posible quejueguenunu• papelpatogénico,al contribuir en la destrucciónde estascélulasclavesparael funcionamiento
• del SistemaInmune.uu• Una de lasparadojasmásllamativasde la infección,esquesolamentesepuededetectar,• por técnicasde hibridación iii situ, una pequeñafracción de células infectadas.En etapasue precocesde la infección solo se observa que uno de cada 10.000 linfocitos T CD4+ está• infectadoy que, incluso, en pacientescuyainfecciónseencuentraen etapasmuy avanzadas,con• linfocitos T CD4+ en su circulación sanguíneamuy bajos, la proporción de sus linfocitos Tu
mantenidosen cultivo que estáinfectadaes muy pequeña.En estoscultivos, uno de cada40• linfocitos aproximadamentees capaz de producir nuevosviriones(58).
• Estasobservacioneshan hechopensara muchosinvestigadoresque estosmecanismosu citopáticosdirectos,descritosanteriormente,no son suficientesparaexplicarla disminucióntan
• drásticade linfocitos T CD4+ en el SIDA y. basándoseen datosexperimentales,hanpropuesto
• otros mecanismosindirectos que podrían estar implicados en la disminución de estau• subpoblacióncelular.
uu Uno de estosmecanismosimplica la destrucciónde linfocitos T CD4+ que presentanu
la proteínagpl2O unida al receptorCD4. Estoslinfocitos puedenunir, procesary presentar
• gpl2O solubleasociadaa antígenosDR, y por ello podríanserpotencialescélulas dianaspara
u linfocitos citotóxicosespecíficosde gpl2O, restringidospormoléculasdel MHC declaseII. Estau• hipótesisestábasadaen la observaciónin viti-o de donesT citotóxicoscapacesde Usarcélulas
• T CD4+ quepasivamentehanadsorbidogpl2O en susuperficie(59).Teniendoen cuentaestose experimentosy queiii vivo se ha detectadola proteínagpl2O libre en circulación,seha sugerido
u queestacitotoxicidad específicade gpl2O puedeserun mecanismosustancialimplicado en la
• disminuciónde los linfocitos T4.eu• En estesentido,tampocosepodríaexcluir queanticuerposanti-HIV puedanmediarla• lisis porComplementoo porcélulascitotóxicas,de célulasCD4+ no infectadasquepresentene• gp120de una forma pasivaen su superficie.uu• 13ueuuu

De forma similar, también se ha descritoque anticuerposque reconocenel sitio de
unión a CD4 en la proteínagpl2O podríanprovocarla generaciónde anticuerposanti-idiotipo
que presentasenla imageninterna de eseepítopo. Estosanticuerposseríanentoncescapaces
de imitar la proteínagpl2O e interaccionarcon la moléculaCD4 expresadaen linfocitos T,
induciendounaentradade Ca~2y unaalteraciónen la función de estascélulas (60).
FiQura 1.5.- Generaciónde una reactividadanti-CD4 en sujetosseropositivos.Anticuerpos
dirigidos frenteal idiotipo de anticuerposespecíficosque reconocenel sitio de
unión a CD4 en la proteínagp 120 podrían interaccionarcon la molécula CD4
nativa, expresadaen la superficiede célulasT.
Determinacionesexperimentalesactualeshansugeridola apoptosiscomootro potencial
mecanismopara explicar la disminución de linfocitos T CD4+. La muerteprogramadade la
célulaes un mecanismosuicida fisiológico, esencialen muchosprocesos,comola maduración
de las células del SistemaInmune. Este procesoaparececoncretamenteen las células T
intratímicasy es esencialen la destrucciónde clonescelularesautoreactivos(61).
En la activacióniii viti-o de célulasT, de un individuo infectadoasintomático,inducida
poranticuerposfrentea sureceptor,TCR, sepudoobservarestefenómenode apoptosisen una
fracción de células T CD4+ (62). Tambiénse ha demostradoesta muerte celular inducida
despuésde unaactivacióncon complejosde moléculasdel MHC de claseII y superantígenos
bacterianos(63). La apoptosisde linfocitos T4 tambiénseha observadoen cultivos estimulados
de linfocitos de sangre periférica que presentanen su superficie gpl2O unida a CD4 y
anticuerposespecíficosde gpl2O (64).
14
b,nding MIt
gp $20
0
uuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu

uuuuu• Ante la demostraciónde quelas proteínasdel 111V, gpl2O y gp4l, tienencaracterísticas
• similaresa losantígenosde histocompatibilidad,yaquecompartenalgunasregioneshomólogas,u
seha sugeridoqueun mecanismoautoinmunepodría estarimplicado en la disminuciónde los
• linfocitos T CD4+. Como consecuenciade esta homología podrían generarseentonces
• anticuerposdirigidos frente a estasproteínasque fuesen capacesde atacary alterara losu• antígenosde histocompatibilidad,deteriorandola función inmunitaria (65).
uu A partede estosmecanismosindirectos,existenotrosmecanismosdirectos,observadosu• recientemente,en los que el virus estaríadirectamenteimplicado en el dañocelular. Uno de
u ellosestábasadoen la observaciónde queel 111V pareceafectarpreferentementealas células
u T de memoria,implicadasen la respuestafrentea una segundaexposiciónantigénica(66).eTambién se ha determinadoqueexisteunaalteraciónen la respuestacelularen la que
u podría estar implicado directamenteel virus, ya queen la respuestade célulasT duranteel
estadoasintomáticode la infección apareceuna depleccióno anergiade la poblaciónThl,u• relacionadacon la inmunidadcelular, con mayor frecuenciaquede la poblaciónTh2, asociada
• a una inmunidad humoral. Estaalteraciónha sido observadatambién en algunasinfeccionesu producidaspor parásitos(67). Estefenómenopodría ayudara explicar laactivaciónpersistente
• de las células8 durantela infección.
U Por último, tambiénse ha sugeridoqueprecursoresde los linfocitos Tenel timo podríanu
ser infectados,y quesu infecciónproduciríaunaalteraciónen la prolíferacióny diferenciación• de la poblaciónde célulasT CD4+ maduras(68).
• Todosestosmecanismosestaríanimplicadosenla disminuciónde los linfocitos T CD4+,u cuya alteraciónafectaríaa toda la homeostasisde las poblacionescelulares.
u• 1.3.2.- Alteración de los linfocitos B en la infección.uuu Aunqueinicialmentetodaslos estudiossobrela inmunopatogénesisde la infección por
HIV estabandirigidos haciael estudiodel estadode inmunodeficienciacelularque seproducíau• en los pacientesinfectados,al pocotiempo deconocerseesteSíndromede Inmunodeficiencia,
u se observótambién que estospacientespresentabanuna alteraciónen su SistemaInmune
u humoral.u• Algunasde las patologíaso procesosobservadosen estospacientes,asociadosa una
• respuestahumoral alterada,son hipergammaglobulinemia,presenciade células B activadasu circulantes,Inmunocomplejoscirculantes(69), autoanticuerposy paraproteínas(70, 71).
uuu 15
uuuuu

uuuuu• A pesarde queexisteunahiperactivaciónde las célulasB, mediantecultivoscelulares
• se ha comprobadoque los pacientescon SIDA presentanuna incapacidadmuy marcadaenu• generaruna respuestade anticuerposfrente a nuevos antígenos(48). Esta incapacidadde• reacciónes el principal problemaque apareceen niños con SIDA, quienestodavía no han
• desarrolladoanticuerposfrente a la mayor parte de los patógenosambientales,y comou
consecuenciade esta incapacidadde respuesta,en ellos aparecenfrecuentementeinfecciones
u bacterianas(72). Se hanpropuestoalgunosmecanismos,basadosen estudiosexperimentales,
u quepodríancontribuir a la hiperactividadde las célulasB en los individuosinfectadospor 111V.u
Porejemplo,la presenciadelvirus,o glicoproteinasviralespurificadasen cultivos,puedeinducir
• unaestimulaciónpoliclonal de las célulasB (73). Otro argumentoqueapoyaestahipótesises
la detecciónen la proteínagpl2O de unasecuenciahomóloga a la neuroleuquina,factor queuestimulalas célulasB (74).
• Entre los factorespropuestosparaexplicar la estimulaciónde célulasE en el SIDA seu
incluyetambiénla infecciónde un grannúmerode célulasB porel virus de Epstein-Barr,EBV• (75). Las células E transformadaspor estevirus puedenser también infectadaspor MIV en
• algunascondicionesexperimentalesy permitir unareplicaciónactivadel virus sin serdestruidas,
de modoquepodríanactuarde reservorioparael HIV en el organismo(76).u• Por otro lado, los pacientescon SIDA desarrollancon frecuencialinfomas E de alto
• grado,comoel Linfoma de Burkitt. Estoslinfomaspuedendesencadenarsepor unapérdidadelu
control de las célulasT sobrelas célulasB, una estimulaciónantigénicacrónicao un aumento• de células E infectadaspor EBV. En relación a este último hallazgo, se ha encontradoen
• algunoslinfomas desarrolladosen pacientescon SIDA, antígenosdel EEV (77).uu• 1.3.3.- Alteración de Monocitos en la infección.
U A pesarde que los linfocitos T CD4+ son las principalescélulasdianadel 111V, otrasu
células como célulasE transformadaspor EBV, monocitos,célulasde la médulaósea,célulasu endoteliales,célulasglialesy célulasepitelialesde la vagina puedensertambiéninfectadas(78,
79, 80, 81). El MIV puedepermanecerlatentedentrode los monocitosy en estesentido,estasucélulaspuedenactuarcomoreservorios,permitiendola distribucióndel virus a otroslugaresdel
• organismo,como el SistemaNervioso Central. evitando que se lleve a cabo una respuesta
inmuneefectiva (82). La vía de entradaen estascélulaspuedeestarmediadapor el receptoru• CD4 (83), perose ha sugeridounasegundavía de entradaa travésde la unión de complejos
• anticuerpo-virusa los receptoresde la fracción constantede las inmunoglobulinas,FeR,u expresadosen los monocitos(84).
uu
16uuuuu

uuuuu• Aunque se han observadoalteracionesen la función de los monocitos, como una
• quimiotaxisdefectiva o una disminuciónen la producciónde anión superóxido(O;), en losu• pacientescon SIDA (85), estasalteracionesno puedenserexplicadastotalmentepor un efecto
• directodel virus ya quesolo seobservaunapequeñafracción de los monocitosinfectada.Esta
• alteraciónpodríasermásbien,unaconsecuenciade una interaccióndefectuosaconlas célulasu
T o unadeficienciaen factoresimplicadosen la estimulaciónde monocitos.
uu 1.3.4.- Alteración de las células NK en el SIDA.uu• Ademásde las alteracionesdescritasanteriormenterespectoa la funciónde lascélulas
u T, B y Monocitos,se handetectadootrasalteraciones,en la respuestainmune mediadaporu• células, en pacientescon SIDA. Por ejemplo, la función de las células Natural Riller está
• disminuidaen estospacientes(86). In vitro seha demostradoqueestaactividadpuedeelevarse
al añadiral mediode cultivo Interleuquina2 (IL-2), lo cual sugierequeestosdefectospodríanu• sersecundariosa la alteraciónde lascélulasT CD4+.
u1.4.- CONTROL INMUNOLOGICO DEL HIV.u
• 1.4.1.- Respuesta humoral.
• Al estudiar la respuestahumoral en los pacientesinfectadospor 1-11V frente a lasu
distintasproteínasvirales se hanobservadodiferenciascuantitativasquehan sido estudiadas
• para determinarsu asociacióncon la progresiónde la infección hacia un estado de SIDA.
• Particularmentese ha demostradoque la respuestade anticuerposfrente a las proteínasde lau
envoltura,gp 120 y gp4l, se mantienedurantetodo el cursode la infección (87), mientrasque
• la especificidadfrentea la proteínade la nucleocápside,p24, disminuyeen lospacientesquese
u encuentranen un estadioterminal (88).uUn análisisde la frecuenciade alteracionesde variosparámetrosinmunoldgicosen
• sangre periférica de un grupo de 200 pacientesinfectados, remitidos al laboratorio de
U Inmunologia de la FundaciónJiménezDíaz, reveló que la disminuciónde linfocitos T CD4+,uel aumentode IgA, la ausenciade anticuerposanti-gagy la presenciadeantígenosvirales(p24),
• aparecíanconunafrecuenciasignificativamentemayoren los sujetosclínicamenteafectadosqueu en los asintomáticos.Estavaloraciónconjuntaarrojó un índicede riesgorelativode 25,5 (89).u• Sin embargo,asícomo los anticuerposanti-envpermanecenestablesen la infección, no todos
• los pacientesconalto riesgode desarrollarel SIDA carecende anticuerposanti-p24,y no todosu
los sujetosasintomáticosmuestranniveles elevadosde estosanticuerpos(90).uu• 17uuuuu

uuuuu• No existetodavíaningunaexplicaciónantela diferenterespuestahumoralfrente a las
• distintasproteínasviralesqueseobservaa lo largo de la infección,aunqueseha sugeridoqueu
la respuestafrentea las proteínasgpl2O y gp41 seaunarespuestaE Timo-independientey que• en la respuestafrente a las proteínasp24 y pYl esténimplicadoslos linfocitos T cooperadores.
• Tambiénseha publicado,al estudiarla asociaciónentreel estadoclínico delpaciente,u• y la presenciay evolución de anticuerposfrente a otrasproteínasdel virus, quela pérdidade
u anticuerposfrentea la proteínapi7 estáasociadaa la progresiónde la enfermedad(91) y que
• la apariciónde anticuerposdirigidosfrente a proteínasreguladoras,como p27, codificadaporu
el gennef, tienelugar al principio de la seroconversión(92).
uDurante la infección, en la respuestahumoral, se genera una gran variedad deu
• anticuerposfrente a las proteínasde la envolturadel virus, sin embargo,solo una fracción de
• esos anticuerpos son capacesde neutralizar la infectividad del virus (93). El epítopo
U inmunodominanteen la proteínagpl2O pareceestarlocalizadoen una regiónconservadaen elu• extremoCOOH terminal (94). Por el contrario el sitio de unión a CD4 en la proteínagpl2O
U está en un hueco que pareceser un sitio antigénico inaccesiblepara la producción deu
anticuerpos.También se ha propuestoque la formación de anticuerposneutralizantespodría• estarinhibida por un procesode autotolerancia,ya queexisteunasimilitud entrelas proteínas
• de la envolturadel HIV y estructurasdel MHC queseunena CD4 (95).uu• 1.4.2.- Respuesta celular.uu
Al contrariode la gran diversidadde estudiosque se han realizadosobrela respuestau deanticuerposen los pacientesinfectadospor HIV, la documentaciónde respuestasespecíficas
u de células T ha sido, de alguna manera, más difícil y más tardía. No obstante,si se hau
observado,desdequeempezarona publicarselos primeroscasosde SIDA, queduranteelcurso
• de la infecciónapareceuna intensarespuestade célulasT citotóxicasy queéstadisminuyeen
u los pacientescon SIDA (96)uTambiénseha observadoquelascélulasT CD8+ de pacientesconSIDA mantienenuna
• función citolítica normal a pesarde haberperdidosu respuestacitotóxica específicafrenteal
1-11V (97).u• Durante estos últimos años se han identificado respuestasespecificasde células T
• citotóxicas,CTLs, frentea proteinasestructuralesdel virus queformanpartede la envolturao
u de la nucleocápside(98,99),e incluso frente a la transcríptasainversa (100).uuu
18uuuuu

uueuu• Tambiénsehandemostradootrosmecanismoscitotóxicos mediadospor célulasTen los
que no existe una restricción por moléculas de clase T del Complejo Principal deu
Histocompatibilidad(MHC), en los queestaríanimplicadosfactoressolubles(101).
• La respuestacelularjuega un papel muy importanteen el control de la infección. Sin
• embargo,al desarrollarseun estadodeSIDA seproduceunarápidacaídade la función inmune
asociadaa un aumentode la replicaciónviral (102).u• En los estadiosmásavanzadosde la infección seproduceun aumentode los niveles deu
antígenop24 en el suerode los pacientes,cuyos niveles, al principio de la infección, son• generalmenteindetectables.Esteaumentoy la pérdidade anticuerposanti-p24en el suero,
• aunqueéstospodríanestarformandopartede Inmunocomplejos(103-105),suponeunaintensau
replicaciónviral. Esta podría debersea la disminución del control inmunológico del virus,• aunquetambiénseha propuestoquepodríaserel resultadode la apariciónde variantesvirales
• másagresivasen el mismo individuo (106). (Fig. 1.6).uu Infodon Doath• 4, MIno<or no $y,T,ptoms
• E1 DOC
• E
CD4PBL••
0-• Anlibodios lo HIV
uHIV-spociflc CR
uArilibodIes lo 1-11V p24
Vire mía• A. A,x~nA~
u ______/‘
• 4-8 weeks Up 1o12 years 2-a yeafs
uu• Fig. 1.6.- Esquemadel curso de la infección en un pacienteque desarrollael SiDA en• función de lasalteracionesde su SistemaInmunológico.
1,5.- IMPLICACION DE LOS ¡CC EN LA INFECCION.uu• Despuésde lasprimerasdescripcionesacercade unagran alteraciónde lascélulasB en
sujetosinfectadospor HfV, se sugirió que los linfocitos B podrían estar implicados en lau• patogénesisdel Síndromede InmunodeficienciaHumana(46), y actualmente,la posibilidadde
• queestascélulasparticipenen la alteracióndelSistemaInmunedurantela infecciónescadadía
másrelevante.uuu
19
uuuuu

la viti-o se ha demostradola síntesisespontáneade Inmunoglobulinaspor célulasB de
sujetos seropositivos(48). Sin embargo, en esta actividad tan intensade las células E, de
naturalezapoliclonal, la respuestafrente al HIV constituyeel principal componente(69).
La respuestaespecíficafrente al virus y el total de la síntesisde IgG muestranuna
correlación significativa, ya que del 20 al 50% de las Inmunoglobulinasproducidasestán
dirigidasfrente a antígenosvirales (107).
Todosestosdatossoportanla hipótesisde queunafuerte respuestahaciaantígenosdel
¡-¡IV, másqueunaactivaciónpoliclonal indiscriminada,es el resultadodela hiperactividadque
apareceen las célulasB de sujetosseropositivos.
Un reflejo de esta alteraciónde los linfocitos E en la infección es la presenciaen el
suerode muchossujetosinfectadosde elevadosniveles de Inmunocomplejoscirculantes(¡CC)
(108).
Los Inmunocomplejosse forman ja vivo cuando anticuerposespecíficosse unen a
determinadosantígenoscomo consecuenciade una respuestainmune. Estoscomplejosson
normalmenteeliminadosde la circulaciónpor célulasquepresentanreceptoresde la fracción
constantede las inmunoglobulinas.FcR. las cualesestándistribuidaspor todo nuestroSistema
linfoide.
Sin embargoen muchasenfermedadesautoinmunes,infecciosasy neoplásicas,los
inmunocomplejosseacumulany causandañospatológicos,comoinflamación,inmunosupresión
y trombocitopenia.
En la infección por ¡-¡IV seha descrito,quealgunasveceslos inmunocomplejospueden
contribuir al desarrollo de ciertasmanifestacionesclínicas que aparecenen los pacientes
infectados,como trombocitopeniapúrpura autoinmuneo glomerulonefritis (109, líO) y otras
alteracionescomo altralgias,mialgiaso vasculitis (III).
A pesarde quesepuededemostrarla presenciade inmunocomplejoscirculantesen un
porcentajeelevadode pacientesinfectadospor HTV (112), no todoslos pacientescon niveles
elevadosde inmunocomplejospresentanunapatologíadebidaa su presencia.De estaforma,
seha sugeridoquela infecciónporHIV, ademásde predisponeral pacientea padecersucesivas
enfermedadesinfecciosas,producidaspor otrosagentesinfecciosos,asícomo el desarrollode
enfermedadesneoplásicas,puede inducir la aparición de fenómenosautoinmunes(113).
Respectoa este último fenómenose ha postuladoqueanticuerposanti-HTV pueden
dañarcélulasno infectadaspor reaccionescruzadasconconstituyentesde la superficiecelular.
20
uuuuueueeeuueuuuuueeuueueuuuueeuuuueuuueuuuuueuuuueeeuuuuu

uuuuu• Este fenómenode mimicidad molecularpodría actualmenteconstituir la basede la
• generaciónde un mecanismoautoinmuneen pacientesseropositivos(114). A favor de estau• hipótesis se han documentadodeterminantesno polimórficos de moléculasdel Complejo
• Principal de Histocompatibilidad(MHC) de clase 11 que muestrancierta homología conu
proteínasvirales, presenciade anticuerposanti-DR en el suerode sujetosinfectadosy alguna• homología de epitoposviralescon moléculasde clase1, aunqueanticuerposdirigidos frente a
• estosantígenosno hansido aúndescritos(115, 116, 117).
u Estudiosrealizadosacercade la composicióndelos inmunocomplejos,hanreveladoquee• presentan isotipos específicosde Inmunoglobulinasy que la presencia de IgA en los
• inmunocomplejosestáasociadacon la progresióndel estadoclínico del paciente(118).u Algunos investigadoreshan demostrado la presencia de antígenos virales en
• inmunocomplejosy hanobservado,en distintosgruposde pacientesinfectadospor 111V, quela
• disminuciónde anticuerposanti-p24está asociadaa la aparicióny detecciónen el suerodeu
antígenop24. Estosautoreshan sugeridoque la elevadaproducciónde antígenop24 en los
• estadiosterminaiesde Ja infección, conduciríaa la formación de compJejosque reduciríanla
• detecciónde anticuerposfrentea p24 en el suerode estospacientes(119).ue Sinembargo,otros investigadoreshanobservadoen pacientestratadosconAZT, en los
• queseha inducidounasupresiónde la antigenemia,queéstano va acompañadade un aumento
• de anticuerposanti-p24,con lo cual, hansugeridoque la disminuciónde anticuerposfrente ae• p24 podríadebersesimplementea una respuestahumoralalterada(120).
• Tambiénseha sugeridoquela presenciade inmunocomplejoscirculantesen el suerode
upacientesinfectadospodría estarimplicada en la disminuciónde receptoresC3b, observadaeneritrocitosde algunospacientes(121).
• Estasobservacioneshan dirigido a algunosinvestigadoresa mostrarcierta inquietudue respectoal dañoinmunológicoquepodríancausarlos inmunocomplejoscirculantesdurantela
infección.
U Ante la observaciónhi’i viti-o de que complejosde anticuerpo-viruspodían inducir la
u infecciónde monocitosy macrófagos,mediantela entradadel virus por mediacióndel receptoreu paraInmunoglobulinas(122),algunosinvestigadoressostienenquelosinmunocomplejospodrían
• incrementarla infectividad del HIV.u Todoslosresultadosexperimentalesdescritosanteriormentehansidoconsideradospara
• reflexionarsobre el papel de la respuestacelular B en la infección por 111V, bien por su
• implicaciónen el control de la expansióndelvirus, o bien porsupapelpatogénicoal participaru
en el agravamientode la infección (123).
uu• 21uueue

ueeuueuuuuuuuueueueuueeeeeueeeeeeueeeuuueeeu• 2.- OBJETIVOSY PLAN DE TRABAJOueeueuuueuueee

uuuueuuu
La progresiónde la infección por el virus de la InmunodeficienciaHumanahacia un• estado de SIDA varía mucho en los pacientesinfectados.Algunos pacientesdesarrollanel
síndromecompletoen pocosmesesmientrasqueotrospermanecenasintomáticosduranteune• largo períodode tiempo.
uU Los factoresqueinfluyen en la progresióndel Síndromede InmunodeficienciaHumanaU• adquiridano seconocenmuybienaunquesi se handescritovarios factoresinmunológicosque
• se asociancon un peor pronósticodel sujeto infectado,como la disminucióndel número de
linfocitosT CD4+ en sangreperiféricao la disminuciónde anticuerposfrentea la proteínaviralu• p24, que indican un grado de infección bastanteavanzado.
uu
Actualmenteseestánestudiandodiferentesparámetrosinmunológicoscuya evolución• permitadistinguir entrelos sujetosinfectadosquienesvañ a desarrollarJa enfermedaden un
• plazo máso menosbreve, lo cual seríaenormementeimportantea la hora de aplicar actualesu medidasterapéuticas,dadoqueéstasconllevanriesgoscolateralesy suponenun elevadocoste.
ueuu• OBJETIVOS
ue
Teniendo en cuenta estas consideraciones,los objetivos de esta tesis fueron los• siguientes:
eu
1.- Estudio de> significado patológico de los inmunocomplejoscirculantesen la• infección por el virus de la Inmunodeficiencia Humana, dada la elevada
prevalenciade niveleselevadosen el suerode pacientesinfectados.ueuuu• 2.- Estudiode unarespuestacitotóxicafrentea linfocitos T CD4+ de sujetossanos
• mediadapor célulasT CDS+ de pacientesinfectados.eeeu• 22uueeu

uuuuu• PLAN DE TRABAJOuu
1.- Para determinarel papel patogénicode los inmunocomplejoscirculantesen la• infección, asícomo su implicaciónen la progresiónhacia un estadode SIDA, serealizaronlos
• siguientesestudios:uu• 1.1.- Determinacióndenivelesde inmunocomplejosen elsuerodepacientesseropositivosporu
precipitaciónconSulfatoamónico1,04 M.u• 1.2.- Análisis de la composiciónde los inmunocomplejosen función de su contenidoenu
antígenosvirales y de anticuerposespecíficosfrente a las distintasproteínasdelvirus• medianteensayosinmunoenzimáticosy WesternBlot.
uu
1.3.- Actividad anti-CD4 en los inmunocomplejos.Esta actividad fue valorada medianteu ensayosinmunoenzimáticos,en función de la capacidadde los inmunocomplejos
U circulantesde unirse a una molécula CD4 recombinantesoluble o a esta moléculau
expresadaen la superficiecelular de una línea linfoblásticaT humanaCD4+.
uU 1.4.- Potenciaciónde la infección.Parainferir si los inmunocomplejospuedeninducir un dañou• inmunológico in vivo, se estudiósu capacidadde unión a la superficie de linfocitos T• CD4+ por citometría de flujo y su capacidadde alterar Ja función de esta población
• celular, inhibiendosu proliferación en cultivosestimuladoscon Fitohemaglutinina.uu• 1.5.- Capacidadinfectiva de los inmunocomplejos.Esta valoración se realizó medianteu
ensayosin viti-o, en cultivos de linfocitos de sangre periférica estimuladoscon• Fitohemaglutininaen presenciade inmunocomplejos,en los que se cuantificó la
• presenciade antígenop24 en el sobrenadantede los cultivosy se analizó la presencia
de ADN proviral en el genomade las célulaspor técnicasde reacciónen cadenade lau• polimerasa(PCR).
uu 2.- La determinaciónde una respuestaen la infección mediadapor células T sobreu• linfocitos T CD4+ no infectados,sevaloró estudiandola capacidadcitotóxica de linfocitos T
• CD8+,aisladosde sangreperiféricadepacientesseropositivos,sobrecélulasT CD4+ de sujetosu
normales,estimuladascon PHA y marcadascon CA’.
uu• 23uueuu

uuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu• 3.- MATERIAL Y METODOSuuuuuuuuuuu

uuuuuuu 3.1.- MATERIAL
uuu Todas las muestrasde Suero, Inmunocomplejoscirculantesy linfocitos de sangre
uperiféricaseobtuvieronde pacientesinfectadospor elVirus de la InmunodeficienciaHumana.
Estospacientesfueronremitidosal Dpto. de Inmunologíade la FundaciónJiménezDíaz
u para su control inmunológico, desde distintos servicios de la Fundación Jiménez Díaz,u
principalmentedesdeel Departamentode Infecciosas,en dondese realizó un reconocimiento• clínico exhaustivode estospacientes.
uLos pacientesfueron incluidos en las distintascategoríasde la infección, A, B y C,e
• definidaspor los Centrosde Control de Enfermedadesde Atlanta. En la última revisión de
• 1993, el nuevo sistema de clasificación se añadierontres condicionesclínicas más para elu
diagnósticode SIDA: tuberculosispulmonar,neumoníarecurrentey cáncercervical invasivo.u Ademásse ha ampliadoel diagnósticode SIDA a todasaquellaspersonasinfectadasque
• presentenmenosde 200 linfocitos CD4+/ pl en sangre(52).u
Respectoa esta última revisión, merecela penadestacarqueen paíseseuropeosse
• decidióquelospacientesseropositivosasintomáticosno podíanserdiagnosticadosen un estado
• de SIDA solamentepor sus niveles de linfocitosT CD4+ (124).uu• Las muestrasutilizadascomo controlesse obtuvieronde pacientesseronegativosque
• presentabanotraspatologíaso de sujetossanos.u
Los cultivoscelularesse realizarona partir de linfocitos de sangreperiféricade sujetos• sanosy de célulasde una línea linfoblásticaT humanaCD4±,denominadaJM, cedidapor el
Dpto. de Inmunologiadel Hospital de La Princesa.uuuuuuuuuuuuueu• 24uueuu

uuuuu• 3.2.- METODOS
e 3.2.1. Análisis inmunológicos realizados para el diagnóstico y seguimiento de losu• pacientes infectados.
uEn esteestudiose efectuaronlas siguientesvaloraciones:u
• Serología:
• - Presenciade Anticuerposespecíficosanti-T-TIV y AntígenoHIV (p24) en suero,u
por ensayos inmunoenzimáticos (Abbott recombinant T-TIV-1/HIV-2 3rd• generationy Abbott HTV Ag-1).
• - Detecciónde anticuerposfrente a proteínasespecíficasde la envolturay lau nucleocápside (Abbott, envacor HIV-l. ETA), utilizado como test de
• confirmación
uu
Estudio inmunológico del paciente:• - Niveles de linfocitos CD2+, CD4+, CDS+ y CDI9+ en sangreperiférica,por
• Citometría de flujo (Epics C, Coulter). Para su determinaciónse utilizarone• anticuerposmonoclonales específicospara identificar estas subpoblaciones
• linfocitarias (Coulter).
• - Pruebascutáneasfrente a antígenosdel Tétanos,Difteria, Estreptococo(Grupou• C9), Tuberculina, Glicerina, Cándida (albicans), Trichophyton, y Proteus
u (mirabiiis). (División Merieux).
uu
El seguimientoclínico de estospacientesfue llevado a caboen el Dpto. de Infecciosas• de la FundaciónJiménezDíaz,cuyasHistoriasClínicas hansido de gran ayuda paraevaluar
todoslos resultadosde estaTesis.uu• 3.2.2.- Aislamiento de inmunocomplejos circulantes del suero de los pacientes infectados:uu• Para el aislamientode los inmunocomplejosse utilizó una técnica puestaa punto en• nuestrolaboratoriobasadaen la precipitaciónde loscomplejosdel suerocon Sulfato amónico
u 1,04 M.e• * Determinación de niveles de inmunocomplejos circulantes en Suero
• a. Previamentese centrifugaron las muestrasde suero a 11000 rpm durante5u minutosen una microcentrífuga(¡-lercaus,Sepatech).
uu• 25uuuuu

uuuuuu b. En tubosde cristal de 8 ml semezclaron:3,5 ml de Solución salinaPH 7.2, 0,5
ml de Sueroy 1,4 ml de SulfatoAmónico 4M.u• c. Posteriormentesemantuvieronlos tubos1 horaen un bañoa 37Q C y sedejaron
• 30 minutosa temperaturaambiente.
e. Despuéssecentrifugarondurante20 minutosa 1400 g. ar ambiente.u• f. Se decantóel sobrenadante,dejandoescurrir el tubo boca abajo de 5 a 10
• minutosy secandoel interior de la pareddel tubocon papelde filtro, sin tocar
el sedimento.u• g. Los inmunocomplejosse disolvieronen 3 ml de PBScon 20 pl de NaOH iN.
• h. La concentraciónde proteínasfue calculada a partir de la densidadópticau
obtenidadel precipitadoredisuelto,medida en un espectrofotómetro(Beckman
• DU-5) a 280 nm frentea un blancode soluciónsalina - NaOH IN. Seconsideró
• 1,4 el coeficientede extinción, media aproximadadel coeficientede extinciónu quemuestranlasInmunoglobulinashumanas.Los resultadossedieronen pg/ml.
u• En la valoración de niveles de inmunocomplejoscirculantesen el suerode personasu
sanasse obtuvoun valor medio de 36,05 ±22,8 ~ig/ml, con un valor máximo de 81,6 pg/ml.• Los nivelessuperioresa 200 pg¡ml fueronconsideradoselevadosa efectosclínicos.uu
* Concentración de los inmunocomplejos:• Para concentrar los inmunocomplelosse siguieron Jos primeros pasos descritos
anteriormente,desdea) hastag). Los inmunocomplejosseresuspendierondespuésen 1,5 mlude Solución salinay 0,5 ml de Sulfatoamónico4 M. Posteriomentese dejarontoda la nochea
• ia ambiente.Al día siguientesecentrifugarondurante20 minutosa 1400 g.
Por último sedecantaronlos sobrenadantesy los inmunocomplejosseresuspenderieronu• en 0,5 ml de Solución salinay 4 pl de NaOH IN. Estasmuestrasse guardarona -20~ C.
uu
3.2.2.- Composición de los ICC: Detección de antígeno p24 y anticuerpos anti-HIV.u• El análisisde la composiciónde los inmunocomplejoscirculantesse realizó en función
u dela presenciade anticuerposanti-HIV y deantígenop24 medianteensayosinmunoenzimáticosu• (ABBOTr RecombinantHIV-l/HIV-2 3rd generationEIA y ABBOTT HTVAg-1).
• Los distintos anticuerposfrente al HIV presentesen los inmunocomplejoscirculantesu
se detectaronpor Western-Blot(New Lav-l3lot 1, PASTEURDiagnostie)
uu• 26uuuuu

uuuuu• La cuantificaciónde las InmunoglobulinasIgG, TgM e IgA en los inmunocomplejosse
u determinópor nefelometría(Array, Beckman).Estamisma técnica se utilizó paravalorar lauconcentraciónde inmunoglobulinasen el suerode estospacientes.
u• 3.2.4.- Disociación de los inmunocomplejos:uu• La técnicautilizadaparadisociarlos ¡CC estábasadaen el métododescritopor Kesten
u et al. (1988) (124). 200 pl de las muestrasde ¡CC fueronsometidosa un tratamientoácidoconu• 40 pl de CIH-Gly 1M (pH 2) (Panreac),en presenciade 0,05% de SDS,durante10 minutosa
• 370 C. Posteriormentelas muestrasfueron neutralizadascon 30 pl de TRIS IM (Sigma)y
resuspendidasen un volumende 300 pl de PBS (pH 7,2).uu• 3.2.5.- Determinación de Actividad anti-C04 en los ¡CC.uu• La determinaciónde unaactividadanti-CD4en los inmunocomplejossevaloró en base• a la capacidadde los inmunocomplejosde unirsea una moléculaC04 recombinantesoluble,ou
bien, a estamolécula expresadaen la superficiecelular.Ambasvaloracionesse llevaron a cabo• medianteensayosinmunoenzimáticos.
uu
3.2.5.6.- Unión de los inmunocomplejos a una molécula CD4 recombinante soluble:u• En esteanálisissevaloró la presenciade actividadanti-CD4de losinmunocomplejosporu
su capacidadde unirse a una molécula CD4 soluble recombinante(1.600 U/mí, T Ceil• Science,INC.)fijada en placasde poliestirenode fondo plano.
u .- Descripcióndel ensayoinmunoenzimático:u• a. Para obteneruna concentraciónde 16 U/pocillo de la moléculaCD4 soluble
• recombinantediluida en solución salina (1600 U/mí), se añadieron 100u pl/pocillo en placasde alta adherencia,de 96 pocillos(Costar).
• b. Las placasse mantuvieron2 horasa 370 C y 12 horasa 40C.
• c. Posteriormentese lavaron3 vecescon soluciónsalinay Tween-200,05% (Difco
laboratorios).u• d. Se dispensaronlOO pl de inmunocomplejos(30-40pg/ml) en cada pociflo y las
• placasse incubarondurante 1 hora a 370 C y 24 horasa 42 C.u e. Despuésse lavaron 3 vecescon solución salina.
eu• 27
uuuuu

uueuu• f. 100 pl de Anticuerpo monoclonalde cabra frente a ¡gO humana(1/1000),
• conjugadoconperoxidasa(GAHu-PO,Sera-lab),se incubarondurante1 hora au
372 C, lavándoselas placasdespuésconsolución salina.• g. Se añadieron100 pl de solución de revelado,H20, con OPD ( o-fenildiamina,
• Abbott) durante20 minutos.u
h. La reacciónse detuvocon 50 pl de Acido sulfúrico IN (50H2, 96% Panreac).
• i. La lectura de absorbanciasse realizó a 492 nm (A~2), utilizando un lector de
• placasde micropocillos(SLT 210, Kontron).u
Cálculo de resultados:
• La valoración de la presencia o ausencia de Actividad anti-CD4 en losu
inmunocomplejosse determinóaplicandola ecuaciónsiguiente:% Activ. anti-CD4 = ( A492 Muestra ¡A492 Blanco ) x 100
• Se consideróestaactividadcomopositivacuandofue =262%,ya queen el grupocontrolu
analizado,formadopor 10 inmunocomplejosno relacionados,aisladosdel suerode 6 pacientes• conArtritis reumatoidey 4 con Hepatitis8, se obtuvo un valor medio de 132 ± 65 %.
uu 3.2.5.7.- Unión de los ¡CC a la molécula CD4 expresada en membranas celulares:uu• La presenciade actividadanti-C04en los inmunocomplejoscirculantessevaloró en esteu
casopor la unión de los inmunocomplejosa la moléculaCD4 expresadaen la superficiecelular.• Paraello seutilizó un ensayoinmunoenzimáticodescritopor Silvestriset al. (125)con
• algunasmodificaciones,en el que se utilizó paraobtenerel extractode membranas,célulasdeu una línea linfoblástica1 humanaCD4+.
u• * Mantenimiento de la línea linfobléstica T humana CD4+:u
Los cultivosde la línea celularse realizaroncon Medio RPM? 1640 suplementadocon• Suerode ternerafetal al 10%,cuyo complementohabíasido inactivadopor calentamientoa 56~
C durante30 minutos,L-Glutamina2mM, 50 U/ml de penicilinay 50 pg/ml de estreptomicina.uAl añadirestosreactivosseutilizaronfiltros de0,22 pm (Millex-GS). La preparacióndel
• mediodecultivo asícomoel manejode loscultivos, fueronrealizadosbajocondicionesestériles,
u en unaCampanade flujo horizontal (Telstar).u• Lascélulassemantuvieronen frascosesterilizados(Costar),en un volumende 10 ml de
• medio completo, en una atmósfera de CO2 al 5%, a 372C. Para el seguimiento de la
u proliferacióncelularse utilizó un Microscopiode contrastede fases(OlympusIM).
uu
28uuuuu

euuuu
A medida que la concentracióncelular aumentaba,superandouna concentraciónde• 5x10’ céiuias/lOmJ, se iban haciendopasessucesivosa nuevosrecipientes.realizándoseuna
• dilución 1/2 con medio decultivo completo.u
En la congelaciónde las células se utilizó Medio RPM? suplementadocon DMSO
• (Sigma)al 20% y STFal 40%.
• En criotubosde 1,8 ml (Nunk) secongelabande 2 a 4 x 10’ célulasen 0,5 ml del Mediou
de congelacióny 0,5 ml del Medio RPM!. Primero se guardabanen un congeladora -40~Cy
• posteriormentese almacenabanen contenedoresconNitrógeno líquido.
• La descongelaciónde las célulasse hacíaen un bañoa 37~C y previamenteal cultivo seulavaban tresvecespor centrifugacionessucesivas,a 300 g, con medio RPMI. La viabilidad
• celular se determinópor exclusióndel coloranteAzul tripan.
U La expresiónde CD4 en la línea linfoblásticase confirmó por citometría de flujo, conu• un anticuerpomonoclonalespecíficode CD4, conjugadoconFicoeritrina (Dako).
• Fenotipo de la línea celular: C04+ (82,9%)uuuuuuuuuuuuuu• Preparación del ensayo inmunoenzimético:
• * Obtención del extracto de membrana:u
Lascélulasde la línea celularlinfoblásticaT humana(2x10’) seresuspendieronen 1 ml• de soluciónsalina(pH 7.2) con NP-40 al 0,5%, PMSF2 mM eInhibidor de Tripsina 1 U/mí,
a obtenido de clara de huevode gallina, y se incubarondurante 1 hora, en hielo, con agitaciónu• ocasional.
• Posteriormentesecentrifugarona 400 g durante>5 minutosa 40 C, en unaCentrífuga
Modelo CS-GR de Beckman, para eliminar el núcleo. A continuación se sometieronau• ultracentrifugación(100.000g) durante30 minutos,a 40 C, en unaCentrífugaModelo12-21 de
• Beckman,paraeliminar la fracción correspondienteal citoesqueleto.uu
29uueuuu
LRFL

euuuu• Determinaciónde la concentraciónde proteínas en las muestrasde extractosde
• membrana
:
uDespuésde resuspenderel extractode membranaen 1 ml de soluciónsalinasevaloró
• la concentraciónde proteínasde estasolución porcolorimetría(Métodode BIO-RAD). basado
• en el cambioqueseproduceen la absorbanciamáximade unasoluciónácidadelcoloranteAzulu CoomassieG-250 cuandose unea proteínas.
• Esta absorbanciavaria de 465 nm a 595 nm. El cambio de color del coloranteque se
• produceestárelacionadocon la concentraciónde proteínasde la muestra.u
La lecturase efectuóa 595 nm en un espectrofotómetromodelo DU-5 de Beckman.
• Paraconstruirla curva patróny calcularla concentraciónde proteínasde la muestraobtenida
del extractode membranasserealizarondilucionesseriadasdeunamuestrainicial deAlbúminau
bovina (1 mg/mí), (ESA, SIGMA). La concentraciónde proteínasobtenidaen los extractosde
• membranavariabaen cadaaislamiento,pero en todoslos casosseobtuvo unaconcentración=
u a 0,180mg/ml al partir de unaconcentracióncelularde 2x10’ células/ml.u• Los extractosde membranaseguardaronen alícuotasde 500 pl a -80~ C.
u• * Ensayo inmunoenzimético:uu• Previamentesevaloró la concentraciónóptima de proteínasdel extractode membrana
u unido al fondo de los pocillos de placasde alta adherenciaen la que se podía detectarlau• moléculaCD4. Estavaloraciónse realizó utilizando un ensayoinmunoenzimáticoen el quese
• comprobóla presenciade esta molécula con un anticuerpomonoclonalde ratón anti-CD4
humanoconjugadoconperoxídasa(CelíFree),y en el quese observóqueunaconcentracióndeu• 3 pg/pocillo de las proteínasdel extractode membranaera suficienteparadetectarunaunión
• específicadel anticuerpoanti-CD4.uu• Parala preparaciónde las placasde altaadherenciade fondo planocon el extractode
• membrana(3 pg/pocillo), se dispensaron200 pI del extracto diluido en una solución conu
carbonatosódicopH 9,3. La placa se incubó a 40 C durantetoda la noche.Posteriormentese
• hicieron tres lavados con PBS-Tween-200,05% (Difco) y se bloqueó con PBS-BSA 3%,
manteniendola placa a 370 C durante1 hora, y despuésa 40 C durantetoda la noche.eLa presenciade CD4 en los extractosde membranafijados a la placa se confirmó
• medianteun ensayoinmunoenzimáticocon anticuerposespecíficos(anti-CD4humanode ratóne
purificado,T Celí Sciences)y (anti-IgO ratón conjugadocon peroxidasa,Sera-Lab).uu
30uuuue

eueuu• Una vez fijado el extracto de membranaen la placa se dispensaron100 pl de los
• inmunocomplejos(200 pg/ml) porpocillo, porduplicado,incubandola placadurantetreshorasu• a temperaturaambiente.
• Despuésdelavarla placa seañadieron100 pl del anticuerpoconjugado«-¡gO bu-PO
• (l/5x104) (Sera-Lab)a cadapocillo, dejandola placa a 37Q C durante1 hora.u
Pararevelarla reacciónenzimáticaseutilizó soluciónde sustrato,H202con OPD como
• cromógeno(Abbott), 100 pl/pocillo, durante20 minutos.
La reacciónenzimáticase paró con50 pl de 504H2 1 N (Panreac)y la lecturase llevóu• a cabo a una longitud de onda de 492 nm en un espectrofotómetrode placas (Kontrol
• Analytical, STL-210), frente a un blancode solución salina.ee• Cálculo de los resultados:• % Actividad anti-CD4= ( A492 Muestra/A~ Blanco) x lOOue• Como control de la unión de los inmunocomplejosa CD4 se utilizó un anticuerpo
• monoclonal anti-CD4 (T-Cell Sciences) en ensayos inmunoenzimáticosde competiciónu
indirectos.e• 3.2.6.- Unión de los inmunocomplejos a linfocitos de sangre periférica:eu• La unión de Jos inniunocompiejosa Ja superficie de linfocitos de sangreperiférica se
• estudiópor Citometriade flujo. En esteensayoindirectosevaloróla inhibición queseproducíau• en el marcajecon a-CD4-FITC (DAKO) de linfocitos de sangreperiférica incubadoscon
• diferentesmuestrasde inmunocomplejos.Paraello se utilizó el siguienteprotocolo:
uu• t Aislamiento de linfocitos de sangre periférica en un gradiente de Ficolí:u
a) 5 ml de sangreperiféricacon heparina1% (Rovi) de sujetossanossediluyeronu• con 5 ml de solución salina,pH 7.2.
• b) En tubosde 10 ml con2,5 ml de Ficolí (Lymphoprep,Nicomed)seañadieron5
ml de la sangrediluida evitando que se mezclasecon el Ficoil. Estos seu• centrifugarona 400 g, durante20 minutos.
• c) Del gradienteque se forma se recogió con una pipeta Pasteurla capa deu
linfocitos quese localiza entreel Ficolí y el plasma.uu• 31uueuu

eeuee• d) Las célulasse lavaronconsolución salinamediantesucesivascentrifugacionesa
• 500 g, durante7 minutos.u• e) El recuentodecélulasserealizóen unaplacade NeubauerconAzul Tripan para
• ver tambiénla viabilidad de estascélulas, en un Microscopioóptico (Nikon), yu
ajustarla suspensióncelulara unaconcentraciónde 10’ células/mlen medio decultivo RPMI 1640 (Bio-Wittaker) o Solución isotónica de Cloruro Sódicoal
• 0,9% (Braun Medical).ee
* fletección de ICC sobre linrocitos de sangre periférica:eu a) Cultivo de linfocitos de sangreperiférica, 106/200 ¡sí, durante2 horas,a 370, enu• presenciade lOO pl de inmunocomplejos,concentradosaproximadamentea 200
• ¡sg/ml.e
b) Lavadode las célulascon solución salinaen unaMicrofuga (Rereaus).
• c) Marcajecon 20 pl de anti-CD4-FTTC(Dako) durante20 minutosa 40 C.
• d) Lavadoy resuspensiónde la célulasen 300 jil de PES.
e) Determinaciónde la fluorescenciaen un CitómetroEPICS-C,COULTER.ee• Como control se consideróen cada ensayoel % de célulasmarcadascon anti-CD4dee
PBLs queno habíansido incubadosconinmunocomplejos.El valor obtenidoen todoslos casos
• estabaen un rangoentreun 30 y 37%.
• Paraconfirmarla unión de loscomplejosa la superficiecelularseutilizó un anticuerpoemonoclonalfrentea IgG humanaconjugadoconfluoresceína(Dako).
ue 3.2.7.- Inhibición de la prolireración de linfocitos de sangre periférica poru• inmunocomplejos, en cultivos:
uParadeterminarla capacidadde los inmunocomplejosde inhibir el crecimientocelulare
• se hicieron cultivos in viti-o de linfocitos de sangreperiféricade sujetosnormales,estimulados
• conPHA (1 mg/mí), enpresenciadediferentesinmunocomplejosaisladosdel suerodepacientes
infectadospor MIV.e• Como control se utilizaron inmunocomplejosaislados del suero de pacientes no
• infectadosconpatologíasautoinmunes,comoArtritis Reumatoide,o enfermedadesinfecciosas,e comoHepatitisB, en las quesedetectanconfrecuenciaelevadosnivelesde inmunocomplejos.
ue• 32ueuue

euee
con * Ensayo de inhibición de la proliferacién de linfocitos de sangre periférica estimulados
a en presencia de inmunocomplejos.ee• Paravalorar si la presenciade ¡CC en los cultivosde linfocitos estimuladoscon PHA
u influía en su proliferaciónseaislaronlinfocitos de sangreperiférica de sujetosnormales.e
El aislamientode linfocitosserealizómedianteun gradientedecentrifugaciónconFicoil
• (NycomedPharmacía).Una vezpurificadosse ajustaronaunaconcentraciónde lx 106 enmediou
completo RPMI-1640 (Flow), suplementadocon 10% de Suero de ternerafetal (Flow), L-• Glutamina2mM (Flow), 50 U/ml de penicilina (Fiow) y 50 ¡sg/ml de estreptomicina(Flow).
• 100¡sí de estasuspensiónse transfirierona placasde 96 pocillosde fondoplano(Costar)u quese mantuvieronen cultivo a 370 C, en unaatmósferade CO2 al 5%, durante72 horas.
• Loscultivosseestimularoncon10 pl de Fitohemaglutinina(1 mg/mí) (Gibco),dosisque
• previamente se había determinado como óptima. y se realizaron en presencia dee
inmunocomplejos(200 ¡sg/mI) aisladosdel suerode pacientesinfectadospor H¡V y sujetosno
• infectadoscon niveles elevadosde ¡CC en suero.
• La proliferacióncelular sedeterminépor la incorporaciónde Timidina H3 en el DNA
de las células.Paraello seañadióa cadapocillo 1 pCi de Timidina H~ (Amershan)durantelase• últimas 18 horas.
• Las células se recogieron utilizando un colector de células automático (Skatron,e• Noruega).
• La precipitacióndel DNA de estascélulasse»evóa cabocon J~l2O destiJadasobrepapel
de filtro. Una vez que los filtros correspondientesa los distintospocillosde la placa se secaron,e• secolocaronen viales minipoli-O (Beckman)con 2m1 de Líquido de centelleo(Fórmula989,
• Dupont) paraefectuarsu lecturaen un Contadorde centelleoBeta (BeckmanLS 2800).uu
* Resultados:• La valoración de la inhibición del crecimientocelular por ¡CC se determinépor la
disminuciónde incorporaciónde Timidina H3 en el DNA de las células,en un contadorBetau• (cpm).
• El porcentajede inhibición del crecimientocelular por los ¡CC se calculé aplicandola
u siguientefórmula:uuU % ¡nh.= lOO - ((cpm PHA - cpm ICC)/cpm PHA) x 100euue• 33ueeue

uueuu• cpm PHA= Timidina H~ incorporadapor el ADN de las célulasestimuladascon PHA.
uu• cpm ICC= Timidina H~ incorporadapor el ADN de las célulasestimuladascon PHA, en• presenciade inmunocomplejos.
eu
Se consideróun porcentajede inhibición significativo cuandoésteera =22%, ya que• ICC no relacionados,de pacientesseronegativos,presentabanun porcentajede inhibición del
U crecimientocelular inferior a un 22%.uu• 3.2.8.- Infección in vitro de linfocitos de sangre periférica por ICC:eeu Primeroseaislaronlinfocitos de sangreperiféricade sujetossanosporun gradientede• densidadconFicoil. 2x10’ linfocitos/ml seincubaronenplacasde24 pocillos(Costar)conmedioe
RPMI (Flow), suplementadocon suerode ternerafetal al 10% (Flow), penicilina 50 U/mí,• estreptomicina50 ¡sg/ml y L-Glutamina2 mM (Flow).
uu
Lascélulasse activaroncon20 pl de PI-JA (lmg/ml) (Gibco) durante72 horas,en una• atmósferade CO2, a 370 C. A los tresdíasde cultivo seeliminó partedel sobrenadantede los
• cultivos y las células se incubaroncon los diferentes inmunocomplejosaislados, a unae
concentraciónde 200 ¡sg/mí, resuspendidosen medio de cultivo RPMI.
• Para mantenerlos cultivos durantemás de una semanase añadió al medio IL-2
• recombinante(100 U/mi, Amersham).eA los dos, cuatro, seisy diez díasde cultivo se recogieronlos sobrenadantesde los
• distintoscultivos paramedir la concentraciónde antígenop24.
La viabilidad celular a lo largo del cultivo se valoró por exclusióndel coloranteAzulu• tripan.
eu
3.2.8.1.- Cuantificación de niveles de antígeno p24 en el sobrenadante de los cultivos• celulares.
uLa determinaciónde niveles de antígenop
24 tanto en el sueroy los inmunocomplejose• de los pacientesinfectados,así comoen los sobrenadantesde los cultivoscelulares,se realizó
U por un ensayoinmunoenzimático(Abbott).euuee 34uueeu

3.2.8.2.- Detección de ADN proviral integrado en el genoma celular por técnicas de PCR.
La deteccióndeHIV- 1 en ADN genómicosupusoenprimer lugarla extraccióndeADN
y, en segundolugar, la amplificaciónde dossegmentosdiana,correspondientesa las regiones
envypol del ADN proviral. mediantela reacciónen cadenade la polimerasa(PCR).
1) Extraccióndel ADN:
El ADN fue aislado de linfocitos de sangreperiférica cultivados en presenciade
diferentesinmunocomplejosdepacientesinfectados.La extracciónseefectuóporprecipitación
salina, técnicadescritapor Miller, 5. A. et al., en 1988, con algunasmodificaciones.
Los linfocitos de cada cultivo se resuspendieronen 1 ml de solución de lisis (STMT),
compuestapor sacarosa0.32 M, Tris-CIH 1 mM, pH 7.5, MgCI2 5 mM y Tritón X-¡00 al 1%.
Posteriormentesecentrifugarona 1500g durante10 minutosy el precipitadoobtenido,
correspondientea los núcleoscelulares,seresuspendióde nuevoen 0,5mIde lasoluciónSTMT,
volviéndosea centrifugaren las mismascondiciones.
El ADN se liberó de los núcleosmedianteun tratamientode 18 horasa 370 C, con300
¡sg/mideproteinasaK (Sigma),en unasoluciónde CINa 75 mM, EDTA 24 mM y SDSal 0.5%.
Las proteínasse eliminaronmedianteprecipitacióncon CINa saturadoy centrifugacióna 1500
g, de 15 a 30 minutos.
El sobrenadanteobtenidose mezclósuavementeconun volumenigual de isopropanol,
hastaobtenerunamasacompactade ADN, transferidaa un tubo tipo eppendorf,con 1 ml de
etanolal 70%.Se centrifugóa 3000 g durante2 minutosy despuésde eliminarel sobrenadante,
el precipitadode ADN sesecóal vacíodurante10 segundos.Posteriormenteseresuspendióen
100 pl deTampónElectrolito (Tris-CHI 0.01 M, pH 7.5 y EDTA 0.001 M, pH 8), calentándose
a 500 C paradisolver el ADN, y procediéndoseacontinuacióna su cuantificación.
2) Cuantificación del ADN:
Paracalcularla cantidadde ADN extraída,semidió la absorbanciadevhias diluciones
de la muestraa 260 nm, en un espectrofotómetroUV/V DU-5 (Beckman).La concentración
se obtieneal utilizar la siguientereacción:
ADN ¡sg/ml = A260 (inversade la dilución) x 50
Es aconsejabletambién medir la Absorbanciaa 280 nm, ya quesiemprequeel índice
A2~ / A280 seamuy diferentea 1.5, esposiblequeexistaunacontaminaciónconproteínas.
35
u

3) Electroforesis en geles de agarosa:
Para comprobar que el ADN extraído no estlí degradado. la muestra se somete a una
electroforesis en geles de qarosa al 1%. Previamente el azar se disuelve con calor y agitación.
en el tampón electrolito TAE (Tris-acetato 0.04 M, EDTA 0.001 M). Una vez que la solución
alcíinza el punto de ebullickín, se deja enfriar por debajo de 00” C y se añade una concentración
final de 0.5 pg/ml de bromuro de etidio (Sigma), que permite visualizar el ADN una vez llevada
a cabo la electroforesis.
El gel se deja polimerizar en frío y posteriormente se introduce en la cubeta de
electroforesis que contiene solución TAE.
Las muestras de ADN se diluyen en un volumen igual de la solución TAE y se dejan
migar en el gel a 60 V durante 30 minutos, en una cubeta horizontal (BRL), Una vez que ha
terminado LI rlectrof«resis. el ADN se ohserw en un visualizador de geles con luz ultravioleta
incorp«rad;l, Fotodyne (lherlaho).
4) Amplilicariím del ADN por la reaccih en cadena de la polimerasa (PCR)
El ADN de los linfocitos fue amplificado para su estudio, mediante la reacción en
cadena de Ia enzima Tq ADN polimerasa (Perkin Elmer cetus, Norwalk, CT), tkcnica que
permite. aprovechando la naturaleza termoestable de esta enzima, la expansi6n exponencial del
segmento :I amplificar. mediante la repetición de tres pasos hlísicos (desnaturalización de la
muestra. alineamiento dc los cehxfores o iniciadores y ex~ensii>n del ADN) (136).
En este caso los fragmentos a amplificar, para poder inferir la presencia de ADN
proviral integrado en el genorna celular, pertenecían a las regiones erw y [XI/ del HIV-l.
Un problema de la alta sensibilidad de la PCR es que una mínima contaminación de la
muestra o de los reactivos por la secuencia diana de ADN se amplificaría y podría dar lugar,
a la hora de interpt-rtar los resultados para el diagrxístico de la infección, a falsos positivos.
Para evitar cualquier posibilidad de contaminacicin. la preparación de las muestras de
PCR se realizan en una cabina de flujo laminar y nunca se utilizan en el mismo sitio de trabajo
pkmidos o productos de PCR, utilizándose las precauciones recomendadas por Knok et al.
(127).
Previamente. para comprobar que el lisado obtenido se podía amplificar se hicieron
PCRs con iniciadores específicos de la B-glohina humana.
36

uuuuu• ParadetectarADN viral se utilizaron oligonucleótidosespecíficosparaamplificar las
regionespot(JAI7/JA2O)y env (JAI3/JA 16). La reacciónencadenadela polimerasaserealizóu
en dos reacciones,la primera con un par de oligonucleótidos externos (JA13/1A16) y
u (JAI7/JA2O)y la segundaconun par de oligonucleótidosinternos(1A18/JAI9)y (JA14/3A15)
• (NestedPCR).u• A) REGION ENV gp4l
U ¡a PCR (primers: JA13 JAI6)
• Condicionesde reacción:u- TermocicladorPerkin Elmer9600
• - Volumenfinal 50 ¡sI:
u - 5 pl IOx PCR buffer (Tris 10 mM final pH 8.3)u - 14 ¡st MgCI2 25 mM (7 mM final)U - 0,25 ¡sí dNTP 10 mM (200 ¡sM)• - 1,25 ¡sí de cadaprimer a 10 pmoles/pl• - 29 pl H20 DEPC• - 2,5 U Taq pol (Perkin Elmer)• Añadir 2 ¡sg de ADN genómicoen lO pl
Cubrir con aceitemineral ligeroCic>os:
-95v C 5 minutos- 24 ciclos: 950 C 20 s
450 C 20su 720 C 20s (+ 2s. por ciclo)u - 720 C 5 minutosu• Y PCR (primers: JAI4 JAIS)
uCondicionesde reacción:
- 5 ¡sí del productode la primeraamplificación.- 4 pI lOx PCR buffer (Tris 10 mM final pH 8,3)
u - 8 pI MgCI2 25 mM (7 mM final)u - 0,25 ¡sí dNTP 10 mM (200 ¡sM)• - 1,25 ¡sí de cada primer a 10 pmoles/¡slu - 30,25 ¡sí 1120 DEPCu - 2,5 U Taq pol (Perkin Elmer)u - Cubrir con aceitemineral ligero• - Ciclos:
~950 C 5 minutos-24 ciclos: 950 C 20s
500 C 20 su 720 C 20s (+ 2s. por ciclo)uuuuuu• 37uuuuu

uuuuuu• B) REO¡ONPOL
uía PCR (primers:JA17 JA2O)
Condicionesde reacción:uu - TermocicladorPerkin Elmer 9600u• - Volumen final 50 ¡sí:
u• - 5 ¡sI lOx PCRbuffer (Tris 10 mM final pH 8,3)• - 14 ¡sí MgCI
2 25 mM (7 mM final)• - 0,25 ¡sí dNTP 10 mM (200 ¡sM)
- 1,25 pl de cadaprimer a 10 pmoles/¡sl- 29 ¡sí H20 DEPC
- 2,5 U Taq pol (Perkin Elmer)• - Añadir 2 ¡sg de ADN genómicoen 10 ¡sI• - Cubrir conaceitemineral ligero• - Ciclos:• -95~ C 5 minutos• -24 ciclos: 950 C 20s
410 C 20 s• 720 C 20s (+ 2s.por ciclo)
2~ PCR (primers:JAI8 JAI9)uCondicionesde reacción:
u• - 5 ¡sí del productode la primeraamplificación.u - 4 ¡sí lOx PCRbuffer (Tris lO mM final pH 8,3)
• - 7,5 ¡sI MgCI, 25 mM (3 mM final)• - 0,25 ¡sí dNTP lO mM• - 1,25 pl de cada primer a 10 pmoles/pI
- 30,25 ¡sI H20 DEPC- 2,5 U Taq pol (Perkin Elmer)- Cubrir con aceitemineral Ligero- Ciclos:
• 950 C 5 minutos• -24 ciclos: 950 C 20 su 4l~ C 20s• 72~ C 20s (+ 2s. por ciclo)• ~720 C 20s. (+ 2s. porciclo)
uu• En todos los experimentosse pusieron controles negativos, sin ADN, sometidos
e exactamenteal mismo proceso,paradetectaralgunaposiblecontaminación.uuuuu• 38uuuuu

5) Análisis de los productos amplificados:
Paracomprobarque la amplificaciónha sido correctase analizóel 20% del producto
amplificado, en estecaso 10 ¡sí del 20 amplificado, en gelesde agarosaal 1.5%en el tampón
electrolitoTAE, previamentedescrito.La tinción serealizócon bromurode etidio (0.5 ¡sg/mí).
Paracompararel tamañodelproductoamplificadoseutilizaronmarcadoresde bajopeso
molecular(OX 174 digeridocon HaeTil, Stratagene).
Los productosamplificadostienen172 pb (regiónenv,correspondientea gp4l) y 129 pb
(regiónpol).
Descripción de los primers utilizados:
PRIMER
JA 13JA 14JAISJAI6
JA>?JA 18JA 19JA2O
SECUENCIAS(5’-3’)
TrCCTTGG GT~CT~G G GAG C
GCAGCAGGAAOCACTATOGGCCAGGACTCYrGCCTGGAGCAGGTATCITTCCACAGCCAG
TACAGGAGCAGATGATACAG
GGAAACCAAAAATGATAGGG
ATTATGTTGACAGGTGTAGG
CCTGG CYf~AATTVfACY~G O
GENES Y LOCALTZACION
env (8004-8023)env (8022-8041)env (8194-8l75)env (8209-8190)
Po1 (2431-2450)pol (2481-2500)pol (2610-2591)pol (2697-2678)
39
uuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuueuuu

uuuuu• 3.2.9.- Estudio de la capacidad citotóxica de linfocitos T CDS+ de pacientes Infectados
frente a linfocitos T CD4+ mediante ensayos de liberación de C¿’.uu• EstosensayosfueronutilizadosparadeterminarJa presenciaen pacentesinfectadosporu
H¡V de célulasT citotóxicascapacesde reconocerdeterminantesantigénicosen la superficie• de linfocitos no infectados.
uComocélulas diana se utilizaron linfocitosde sangreperiféricade individuosnormalesu
u y en algunos casosde pacientesinfectados,estimuladoscon PRA (¡mg/mí, BioWittaker).
u Durantesietedíassemantuvieronen mediode cultivo RPMI 1640 (Gibco) suplementadoconuun 10% de Sil, L-Glutamina2 mM, 50 U/ml de penicilina, y 50 ¡sg/ml de estreptomicina,a
• 37~ C, en unaatmósferade CO2 al 5%.
u El aislamientode PBLs se realizóenun gradientedecentrifugaciónconFicoll-Hypaque.uu• Como células efectoras se seleccionaronlinfocitos T CD8÷de PBLs aisladosdee
pacientesinfectados,medianteseparacióninmunomagnética(DynabeadsM-450 CD8).Las células fueron aisladasde sangreperiférica medianteesferasinmunomagnéticas
• (Dynabeads.Dynal), cubiertascon un anticuerpomonoclonalfrente a CD8. Posteriomentese
u aislaronporaplicaciónde un imán (Separadorde partículasmagnéticas,Dynal). Se lavarontresu• vecescon PBS frío conun 2% (y/y) de STFy sesepararonde las esferascon un anticuerpo
• anti-Fab’ (DetachaBead,Dynal).
u Posteriormentese comprobópor Citometria de flujo que la población seleccionadau• estabaconstituidaen su mayorpartepor linfocitos T CDS+ (89%), encontrándoseen todaslas
• muestrasanalizadasunacontaminaciónmínimapor célulasNK =1%.u Por último las célulasfueron congeladascon medio RPMI, 20% de DM50 y 40% de
• STFen criotubosde 2ml y conservadasen nitrógenolíquido hastasu utilización.
uu
* Determinación de la respuesta citotóxica:• La cuantificaciónde la respuestacitotóxica mediadapor linfocitos T CD8+ se realizó
• medianteensayosde liberaciónde Cr Si, en los quesesiguieronlos siguientespasos:uuu 1.- Marcajede las célulasdiana(0,5 x 10’), resuspendidasen loo ¡sí de STF,con 100 pCi
• de51CrO
4Na2,New EngJandNuclear, en tubosde 12 ml cónicos(Costar),duranteunau
hora en un baño a 370C.
uu• 40
uuuuu

uuuuu• Paraeliminarel excesode cromo las célulasse lavan4 vecescon Medio RPMI
con STFal 5% por sucesivascentrifugacionesa 1700 rpm.u
Posteriormentesecuentanlascélulas,sehaceunaviabilidad y secompruebala
• radiactividadincorporada.Parael ensayodecitotoxicidadseutilizan 2000 célulasdiana,
U con una incorporaciónde cromo de aproximadamente500 cpm.eEn el casode los linfocitos estarelación fue la queseobteníanormalmenteen
• cadaensayo,peroen el casode las célulasde la línealinfoblástica, la incorporaciónde
U cromo fue menor y por lo tanto seutilizó un mayor númerode célulasdiana, 4000 au• 5000,paraobtenerun marcajeigual.
uU t- Dispersiónen placasde 96 pocillos con el fondo en forma de V (Costar) de lasu• células diana, entre 2000 y 4000 células en lOO ¡sí de Medio RPMI y de las células
• efectorasen distintasconcentraciones,en un volumen de 0,1 mí, para tenerdistintas
relacionescélula efectora- célula diana(E:D): (10:1), (5:1), (2:1) y (1:1). Cadaensayou• sevaloré por triplicado.
• Paraque las célulasquedenlo suficientementepróximasse centrífugala placau a 80 g durante5 minutos.
• La placa de citotoxicidad se incubadurante4 horasa 37Q C, en una atmósfera
• de CO2 al 5%. Al cabo de ese tiempo se recoge el sobrenadante(100 ¡sí) parau determinarel porcentajede Cr
51 liberado en tubosde plástico.La lectura se efectuó en
• un contadorgamma(Cobra U, lvisa).
uu
Cálculo de los resultados:• El porcentajede lisis específicase calcula paracadapoblaciónde linfocitos citotóxicos,
• de cada pacienteinfectadopor HIV estudiado,con la siguientefórmula:uu• [(Lib. exp.cpm - Lib. esp.cpm)/(Lib. máx. cpm - Lib. esp.cpm) ] x 100uu• Liberaciónexperimental:
U Liberaciónde Cr5’ por las célulasdiana incubadascon las célulasefectoras.
u Liberaciónespontánea:u• Liberaciónde Cr5’ por las célulasdianaincubadascon medio de cultivo.
U Liberaciónmaxíma:
Liberaciónde C¿1de las célulasdiana incubadascon CLH 0,1 N.uuu• 41
uuuuu

uuuue
* Inhibición de la respuesta citotóxica:uu
Paradeterminarel fenotipode lascélulasefectorasy suespecificidadfrentea lascélulas• diana,se utilizaron anticuerposmonoclonalesfrente a CD8 y CD4 (Dako) respectivamente.
• Previamentea los ensayosde citotoxicidad, las célulasefectorasse incubaroncon 10u• ¡sg/ml de Anticuerpoanti-CD8purificado,durante30 mm, a 40 C.
• Paradeterminarun procesode bloqueoen la lisis celular, partede las células diana
• utilizadasen los ensayos,se incubaroncon 10 ¡sg/ml de Anticuerpoanti-CD4, durante1 hora,u
a 370 C, durantela fasedel marcajecon Cr~’.
u• 3.2.10.- Métodos estadísticos.uu• La comparaciónde los resultadosobtenidosen los diferentesgrupos de pacientes
u infectados por el virus de la inmunodeficienciahumana, establecidospara su estudiou• comparativo,se realizó mediantela aplicaciónde distintaspruebasde significación.
• Seaplicóla pruebade Studentparaidentificarla diferenciade las mediasde dosgrupos.
Paraestudiarla asociacióno independenciade dosvariablescualitativasdentrodeunae• muestra,se utilizó la pruebade chi-cuadrado,de acuerdoconla forma descritapor Bancroftet
• al. (128),utilizando la correcciónde Yatescuandofue necesario.u Por último, paraver la fuerzadeasociaciónentredosvariablesserealizóunacorrelación
• lineal.
• En todoslos análisisse consideróestadisticamentesignificativo un valor de p menor auu 0,05.
uuuuuueuuuuuuueuu
42uuuuu

uuueuuuuueuuuuuuuuueeueeuuueuuueueuueueuueeu
4.- RESULTADOS Y DISCUSIONuuuueueeuuu

eeeueeuuueueueuuuueuuuuuuuuueueuuueuuueueue• 4.1.- PAPEL PATOGENICO DE LOS INMUNOCOMPLEJOS CIRCULANTES• EN LA INFECC¡ON POR 111V.
uueeeeuuueueue

uuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu• 4.1.1.- Asociación entre la progresión de la infección y las siguientes característ¡cas deU los Inmunocomplejos:u• 4.1.1.1.-Frecuenciade niveleselevadosen el suerode pacientesinfectados.u
4.1.1.2.-Composición.
u 4.1.1.3.-Actividad anti-CD4.uuuuuuuuuuuu

uuuuu• 4.1.1.1.- Frecuencia de niveles elevados de Inmunocomplejos circulantes en el suero de
• pacientes infectados por HIV.uu• En un primer estudio,paravalorar el significado patológico de los inmunocomplejos
• circulantesen la infección por el virus de la InmunodeficienciaHumana,se analizaronlosu
niveles de ¡nmunocomplejoscirculantesen el suerode 69 pacientesinfectados.• La determinaciónde los niveles de inmunocomplejosse llevó a cabopor precipitación
con Sulfato amónico1,04 M.u
En función de las trescategoríasdefinidasen la infecciónpor H¡V por los Centrosdel
• Control de Enfermedadesde Atlanta, el 45% de los pacientesfue incluido en la categoríaA,u
ya queseencontrabanen un estadoasintomático,mientrasqueel 21% y el 34% se incluyeron• en las categoríasB y C respectivamente,teniendoen cuentasu cuadroclínico.
• En cadacategoríase distinguierontresniveles segúnel númerode linfocitos T CD4±u quepresentabanlos pacientesen sangreperiférica,siendoun númeromayorque500células/¡sl
• equivalentea un nivel 1, un recuentoentre500 y 200, un nivel 2, y unosniveles de linfocitos
• menoresa 200/pl,un nivel 3. Siguiendoestebaremo,un 23% de lospacientesse incluyeronenu un nivel 1, un 52% en un nivel 2 y un 25% en un nivel 3.uu En total, el porcentajede pacientesen cada grupofue de un 16% (Al), un 23% (A2),
• un 6% (A3), un 4% (BI), un 10% (B2), un 7% (B3), un 3% (Cl), un 19% (C2) y un 12% (C3).u
Los nivelesde ¡CCen el suerode estospacientesfueronelevados,encontrándoseen un
• rango de 114 a 4.100 ¡sg/mL
• La mediade los niveles de inmunocomplejosobtenidaen los tresgruposde pacientesu incluidosen las categoríasclínicasA, B y C fue de 1.074 ±95& 947 + 474 y 791 ±646 ¡sg/ml
• respectivamente.
• Estosresultadosindican queno existeuna diferenciasignificativa entrelos niveles deu• inmunocomplejosquepresentanlos pacientesy su estadioclínico (t= 1,23 / p=O,22).
• Del mismo modo, tampocoexisteuna diferenciasignificativa entre la media de los
e niveles de inmunocomplejoscorrespondientesa los tresgruposde la infección realizadosenufunción del númerode linfocitos T CD4+/¡sl quepresentabanlos pacientes,745 ±766 (1); 976
U ±625 (2) y 1.096 ± 1.040 (3), (t= 1,10 / p=O,28).uu• Niveles elevadosde inmunocomplejos(> 500 ¡sg/mí)sedetectaronen 50 pacientes,loU cual suponeun 72% de los casosestudiados.24 de estospacientes(48%) permanecíanen un
u estadoasintomático(A), mientrasqueel restode lospacientes(52%)presentabaalgúnsíntomau• clínico quelo incluía en unacategoría8 o C (n=26).
ue 43uuueu

uuuuuu Al compararel estadoclínico de los pacientes,en función de su clasificaciónen una
ucategoríaA, B ó C de la infección, con la presenciade niveles elevadosde inmunocomplejoscirculantes en suero, superioresa 500 ¡sgml, observamosque no existía una diferencia
• significativa (p > 0.05),ya queniveles elevadosde inmunocomplejossepodíanencontrarconu igual frecuenciaen el grupo de pacientesasintomáticosy en aquellosformadospor pacientes
• incluidos en unacategoríaB ó C de la infección.
• Sin embargo,si seobservóunadiferenciasignificativa(p c 0.05)conrespectoalnúmero
u de linfocitos T CD4+ que presentabanen circulaciónestospacientes,ya que un 79% de losu• pacientesquepresentabanniveleselevadosde inmunocomplejos,>500¡sg/mí,teníanun número
• de linfocitos T CD4+ menor a 500 células/pl, mientrasque el 21% de estos pacientesu
presentabannivelesde linfocitos en sangreperiféricasuperioresa 500 células/pl(TABLA 1.¡).u• Relación entre la presencia de niveles elevados de Inniunocomplejos circulantes ( > 500u ¡sg/mí) y la detección de antígeno p24 en el suero de pacientes infectados por HIN.ue• En un grupo de 42 pacientescon niveles elevadosde inmunocomplejos,se hizo unu
estudioparadeterminarsi esteparámetroestabao no relacionadocon la presenciade antígeno
• p24 en suero. En este análisis se encontróque la mayoría de los pacientescon niveles de
• inmunocomplejoselevados(n=30),un 72% de loscasos,no presentabaniveles significativosdeu• antígenop24 en suero.
• En 14 casosde estegrupo de pacientes,en los queno se detectóantígenoen suero,lo
cual suponeun 46% de los casosanalizados,sí sepudo detectarésteen los inmunocomplejosu• despuésde disociarlosa pH ácido.
• Estoúltimo implica queen algunoscasosde la infección se podría estarencubriendola
presenciade antígenosviralesmediantela formacióndecomplejosantígeno-anticuerpo,siendo
• de estaforma indetectablesen el suero.
uuuuuuuuuueeu• 44uuuuu

TABLA 1.1.- Asociación entrela presenciadepacientesinfectadospor HIV, suCD4+ en sangreperiférica.
Pacientesn= 69
Estado Clínico
An=31
B/Cn = 38
p ¡
niveles elevadosde ¡CC (>500 ¡sg/mI) enestadioclínico y sus niveles de linfocitos T
Niveles ¡CC> 500 ¡sg/ml
24 26 X2=0,31
pO,S7
8 42 X2=3,9
p<O,05Niveles ¡CC 7 12 8 11c 500 ¡sg/ml
45
Niveles
> 500/pln= 16
lin. T4
~c500/pln = 53
p
uuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu

auuuu• 4.1.1.2.-Análisis de la composición de los ¡CC.uu• En un segundoestudio,en el quese analizóla composiciónde los inmunocomplejos• correspondientesa 141 pacientes,se observóque la clasede inmunoglobulina¡gO estabau
presenteen el 80% de loscasosanalizados,mientrasqueIgM sedetectóen un 15%,eIgá solo• en el5%. La mediade las concentracionesde estasinmunoglobulinasen los inmunocomplejos
• fue de 6,3 ±5,9 (¡gO), 2,9 ± 2,1 (¡gM) y 1,4 ±0,2 mg/dl (IgA).u
Seobservóqueno existíaunacorrelaciónentrelos nivelesde inmunocomplejosy losde
• las inmunoglobulinas¡gG, ¡gM e ¡gA en el suerode los pacientes(r= 0,4. r= 0,03 y r= 0,2),
• ni entrelos niveles de IgO en los inmunocomplejosy en el suero(r= 0.5).uu• Posteriormentesedeterminóla composiciónde los inmunocomplejosen función de la
u presenciade anticuerposfrentealvirus de la InmunodeficienciaHumanay deantígenosvirales,u
fundamentalmentep24.Esteestudiose llevó a caboen un grupode 66 pacientes,incluidospor
u suestadoclínico y susnivelesde linfocitos C04+ en los siguientesgrupos:Al (9), A2 (16), A3
u (4), 81(2), B2 (6), 83 (6), CI (2), C2 (12) y C3 (9). En esteanálisisfuerondefinidoscuatrouvariantesen los Inmunocomplejoscirculantes: (Ac+Ag±),(Ac+Ag-), (Ac-Ag+) y (Ac-Ag-).
u• * Análisis de la composición de los ¡CC en relación con la progresión de la infección.uu• Al teneren cuentasolamenteuna de las dos variablessiguientes,presenciao no deu
anticuerposanti-HTV o de antígenop24 en los mnmunocomplejos.no se observóninguna• diferenciasignificativa entreunacomposicióndeterminadade los inmunocomplejosy el estado
• clínico en el quese encontrabanlos pacientes,asintomáticoso incluidosen unacategoríaE ó
C (p > 0,05) (Tabla 2.1).u• Sin embargo,si se observóunadiferenciasignificativa entrela presenciao ausenciaen
• los inmunocomplejosdeanticuerposfrenteal virus o antígenop24 conrespectoa los nivelesdeu
linfocitos T CD4+ que presentabanestos pacientes,observándoseque la presenciaen los
• inmunocomplejosde antígenop24 era mayoren pacientescon niveles de linfocitos inferiores
• a 500/¡sl, (p < 0,05), (Tabla 2.11).u
La frecuenciacon la queaparecíanlasdistintasvariantesde inmunocomplejos,definidas
• en función de su composición,se analizóen distintos grupos de pacientes,establecidosen
• función del estadoclínico de los pacientes,asintomático o sintomático, incluidos en lasu• categoríasE ó C de la infección, o bien, teniendoen cuentalos niveles de linfocitos T CD4+
• quepresentabanen sangreperiférica,superioresa500/¡sl,entre200y 500/¡slo inferioresa 500/pl.u• 46uuuuu

euuuu• En este análisis se observó que los inmunocomplejosen los que se detectaronu
anticuerposanti-H¡V y antígenop24, (Ac+Ag+), aparecíanconmayor frecuenciaen pacientes• queseencontrabanen un estadiomásavanzadoC (46%), mientrasqueinmunocomplejosAc+
• Ag- eranmásfrecuentesen pacientesasintomáticos(50%>. LosinmunocomplejosAc- Ag+ se
observaronconunafrecuenciamayor en pacientesincluidos en unacategoríaA (50%) (Tablau• 2.111).
• Con respectoal estadoinmunocomprometidode los pacientes,seobservóque¡CC conu
unacomposición(Ac+Ag+) solo aparecíanen pacientesconunosnivelesde linfocitosT CD4+
u circulantesmenoresa 500/¡sl (Tabla 2.1V).
• Segúnestosdatos la presenciaconjuntade anticuerposfrente al HIV y antígenop24u
en los inmunocomplejospodría significar un mayor riesgode progresiónen la infecciónhacia
• un estadode SIDA y la presenciade estetipo de inmunocomplejosen la circulación podría
• considerarsecomo un parámetrode mal pronóstico.uuu Tambiénse realizóun estudiocomparativoen 59 de estospacientesparadeterminarsi
u la presenciade antígenop24 en los inmunocomplejoso en el sueroestabaasociadaal gradodeuinfección o con los niveles de linfocitos T CD4+ quepresentabanestospacientes.
u En esteanálisisseobservóque no existíaunaasociaciónentrela presenciao nó en losu
inmunocomplejosy en el suerode antígenop24 conel estadoclínico del paciente,ya que las• variantesestudiadas(AgICC+/AgS+), (Ag¡CC-/AgS+), (AgICC+/AgS-) y (AgICC-/AgS-),
• aparecíancon igual frecuenciaen pacientesincluidosen una categoríaA o en las categoríasE
u ó C (Tabla 2V).u• Pero si se observóque la presenciade antígenop24 tanto en el suerocomo en los
• inmunocomplejoscirculantesera másfrecuenteen aquellospacientescon nivelés de linfocitosu T CD4+ en circulacióninferioresa 500/¡sl (Tabla 2.VT).
u• * Análisis de la especificidad de los anticuerpos frente a las distintas proteínas viralesu
observada en ICC de pacientes infectados por HIV.u• Las diferentesespecificidadesde los anticuerposfrente al T-I¡V presentesen aquellosu• inmunocomplejosen los que se había determinadopreviamenteanticuerposanti-HIV por• ensayosinmunoenzimáticosse analizaronpor WesternBlott.
Estavaloraciónse llevó a caboen 17 pacientes,incluidosen las siguientescategoríasdeu
la infección : A (8), E (4) y C (5), de los cuales,11 presentabanun númerode linfocitos T
• CD4+ menora 200/pl.u• 47ueeuu

aueuu• Lo mássignificativode esteestudioesquese observaronanticuerposfrentea gplGOen
• todaslas muestrasde inmunocomplejosanalizadasy quela presenciade anticuerposfrente au
la proteína gp4l sólo pudo ser detectadaen inmunocomplejosde pacientescon alguna• manifestaciónclínica, incluidos en unacategoríaB ó C de la infección (Tabla2.VII).
• En esteanálisistambiénseobservóquela frecuenciade anticuerposfrenteala proteínau• p24 era mayoren aquellospacientescon nivelesde linfocitos T CD4+ en circulacióninferiores
• a 200 células/¡sl(Tabla 2.VIII).
Estosresultadossugierenque en la infección por ¡-(IV sepodría valorar por WesternuBlot la aparicióno disminuciónde anticuerposconunadeterminadaespecificidadfrente a las
• distintasproteínasdel virus como parámetrode cierto pronósticoen la infección.uuuuuuuuueuuuuuuuuuuuuuuuuuuuuuuuuu
48uuuuu

TABLA 2.1.- Asociaciónentrela presenciade Anticuemosfrente al HIV o Antígenop24
en los inmunocomplejosanalizadosy el estadoclínico quemuestranlos pacientes.
Composición
ICC
Categoría
A (n=29)
Categorías
B/C (n=37) p
Ac+ 11 16 X2= 0,03
p = 0,85Ac- 18 21
Ag+ 8 ¡3 X2= 0,14
p = 0~69Ag- 21 24
TABLA 2.¡I.- Asociación entrela presenciade Anticuerposanti-HIV o Antígenop24
en los inmunocomplejosy los nivelesde linfocitos T CD4+ quemuestranlos pacientes.
4+ <500/¡sl~
p
Composición
ICC
CD4+ >500/¡sl~CO
n= 13
Ac+ 3 24 X2= 1,3
p 0,25Ac- lO 29
Ag+ 1 20 X24,8
p< 0,05Ag- 12 23
49
auuuuuuuuu
uuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu

TABLA 2.¡T¡.- Frecuenciade los distintostiposde inmunocomplejosdefinidosen funciónde su
composiciónen los tresgruposde pacientesanalizados,establecidospor el estadioclínico
quemuestranlos pacientes.
¡CC Cat.A I~aLB Cat.C
Ac+ Ag+ 31% 23% 46%
Ac+ Ag- 50% 35% 15%
Ac- Ag+ 50% 12% 38%
Ac- Ag- 45% 16% 39%
TABLA 2.¡V.- Frecuenciade los distintostiposde inmunocomplejosdefinidosen función desu
composiciónen los tresgruposde pacientesanalizados,establecidospor los nivelesde linfocitos
T CD4+ quepresentanlos pacientesen sangreperiférica.
Composición
¡CC
CD4/¡sí
> 500
CD4/¡sl
200-500
CD4/¡sl
< 200
Ac+ Ag+ 0% 69% 31%
Ac+ Ag- 21% 50% 29%
Ac- Ag+ 12% 62% 26%
Ac- Ag- 29% 42% 29%
50
eueuuuuuuuuuuuuuuuueuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu

TABLA 2.V.- Relación entrela presenciade Antígenop24 en los ¡CCy/o en
el suerodepacientesinfectadospor HIV y su estadoclínico.
Ag ¡CC Ag suero Cat. A Cat. B/C
+ + 8% 9%
+ - 23% 24%
- + 19% 18%
- 50% 49%
TABLA 2.VI.- Relaciónentrela presenciade Antígeno p24 en los ICC y/o en
el suerode pacientesinfectadospor HTV y susniveles de linfocitos T CD4+.
Ag
ICC
Ag
suero
CD4/¡sl
>500
CD4/¡sl
<500
+ + 0% 11%
+ - 17% 25%
- + 33% 15%
- - 50% 49%
51
uuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu
uuuu
uuuuuuuu

TABLA 2.VIT.- Frecuenciade las distintasespecificidadesdelos anticuerposfrentea las
diferentes proteínas del virus observada en inmunocomplejosde
pacientesinfectadospor HIV, clasificadosen dosgruposen función de
la categoríade la infección en la que fueron incluidos.
Ac HIV Cat.A Cat. B/C
cc-gplGO 100% 100%
a-izpl2O 75% 89%
a-p68 25% 44%
u-p55 37% 55%
«-p52 37% 44%
«-gp4l 0% 44%
«-p4O 25% 33%
«-p34 12% 22%
a-p24 50% 55%
«-p18 12% 11%
52
euueuuueuuuuueueuuuuuuueuuuuuueuuuuuuuuuuuuuueeueueuuuuuuuu

TABLA 2.VII¡.- Frecuenciade las distintasespecificidadesdelos anticuerposfrentea las
diferentes proteínas del virus observadaen inmunocomplejos de
pacientesinfectadospor HTV, clasificadosen dos gruposen funciónde
los nivelesde linfocitos T CD4+ quepresentabanen circulación.
Ac HTV
CD4/¡sl
> 200
CD4/¡sI
< 200
a-gp160 100% 100%
a-gpI2O 72% 100%
a-p68 27% 50%
a-p55 45% 50%
u-pS2 45% 33%
a-gp4l 27% 16%
a-p4O 27% 33%
«-p34 18% 16%
a-p24 45% 66%
«-pl8 9% 16%
53
euuuueuuuuuuuuuuuuuuuueuuuuuuuuuuuuuuuuuuuuuuu
uuuuuuuuuu

eeuuu• 4.1.1.3.-Detección de actividad anti-CD4 en Inmunocomplejos circulantes aislados del
u suero de pacientes infectados por HIN.eu• Actualmenteexisten muchos indicios para pensarquedurantela infecciónpor HIVe
puedeaparecerunarespuestade origen autoinmune.No solamenteesta hipótesisesta basadaen la observaciónen algunospacientesde
• enfermedadesen lasquepuedenestarimplicadosfenómenosautoinmunes,como unapúrpurau
trombocitopénicaidiopática. sino también por algunosdatos inmunológicosdetectadosen• pacientesinfectados(112).
• Recientementese ha descrito la presencia de complejos gpl2O/anti-gpl2O en
u inmunocomplejoscirculantesaisladosdel suerode pacientesinfectadospor HIV, capacesdeu• unirse a la molécula CD4 expresadaen linfocitos de sangreperiférica, que podrían estar
U implicadosen potenciarla infección (129).ue• En nuestrotrabajo nos propusimosdeterminarsi en las muestrasque teníamosde
• inmunocomplejoscorrespondientesa pacientesinfectados,existíaunaactividadanti-CD4. Paraa
determinaresta actividad se valoró la unión de los inmunocomplejosa una molécula CD4• recombinantesolubleo bien,a estamoléculaexpresadaen extractosde membranadeunalínea
• linfoblásticaT humanaCD4+. Ambosestudiosse realizaronpor ensayosinmunoenzimáticos.uu• Valoración de la unión de los Inmunocnmplejos a una molécula CD4 recombinante
e soluble.u
Esteestudiofue llevado a caboen un grupo de 74 pacientes.Siguiendola clasificación
U utilizadapor los Centrosde Controlde EnfermedadesdeAtlanta, los pacientesfueronincluidos
u en las siguientescategoríasde la infección: 34 (A), 14 (B), y 26 (C).uu Dentrodecadacategoríasehandefinidotresnivelesen funcióndel númerodelinfocitos• T CD4+ quemuestranlos pacientesen sangreperiférica.Siguiendoestecriterio, los pacientesu se incluyeron en un nivel 1(17), un 2 (37) y un 3 (20).uu• La determinaciónde actividad anti-CD4 en los inmunocomplejossevaloró en funciónu de su capacidadde unirse a una molécula CD4 recombinantesoluble fijada en placas deuu poliestireno.Esta unión se reveló en ensayosinmunoenzimáticoscon un anticuerpoanti-IgGU humanaconjugadoconperoxidasay los resultadosobtenidosendensidadesópticassereflejaronu
en porcentajesde actividad anti-CD4(Tabla 3.1).uuu 54uuuuu

En este estudio se determinóque 12 pacientespresentabaninmunocomplejoscon
actividad anti-CD4(Figura 3.1), lo cual suponeun 16% de los casos.
Aunque la presenciade esta actividad anti-CD4 en los inmunocomplejosno está
relacionadacon el grado de la infección, ya quefue detectadacon unasimilar frecuenciaen
inmunocomplejosaisladosdel suerode pacientesasintomáticos(47%)y de aqueliosincluidos
por su estadioclínico en una categoríaB ó C de la infección (53%), la mayoría de estos
pacientes(n=8), presentabaniveles de linfocitos T CD4+ en sangreperiférica inferiores a
500/pl.
La existenciade esta actividad anti-CD4en los inmunocomplejosfue observadaen un
porcentajerelativamentebajo de los pacientesanalizadosy tan solo en un caso se pudo
diagnosticarcomo patologíaautoinmune,una púrpuratrombocitopénicaidiopática.Peroaún
así,no sepuededescartarla hipótesisdequeestaactividad tengaalgún significadopatogénico
en la infecciónde algunospacientes.
De hecho, en un caso, en el que se pudo valorar la evolución de la infección del
paciente,se observóun cambioen la actividadanti-CD4de sus inmunocomplejos.En estecaso
se analizaron distintas muestrasde inmunocomplejosrecogidasa lo largo de dos años,
detectándoseun cambioen la actividadanti-CD4de los inmunocomplejosaisladosa finalesdel
segundoaño. Estecambio coincidió con una progresión de la infección, así como con una
disminuciónde los niveles de linfocitos T CD4+ en sangreperiféricadel paciente(Figura3.2).
Con estosresultadosno se podría utilizar de una forma generalizadala presenciade
actividad anti-CD4 en los inmunocomplejoscomo un parámetrode mal pronóstico de la
infección, aunquesi se podría teneren cuentaen aquelloscasosen los quese observeuna
disminuciónde linfocitos TCD4+,ya quelos inmunocomplejospodríanestarimplicadoseneste
proceso.
Esta actividad frente a la molécula CD4, detectadaen los inmunocomplejos,podría
debersea la presenciade antígenogpl2O o de anticuerposantiidiotipo dirigidos frente a
anticuerposanti-gpl2O en los inmunocomplejos.Esta última idea podría apoyarseen cierta
manera,en la observaciónde queen el 28% de los inmunocomplejosque presentabanesta
actividad no se detectóantígenop24 formandopartede los inmunocomplejos,lo cual podría
reflejarla no existenciade otrosantígenosviralescomoconsecuenciade la ausenciadeunafase
replicativadel virus en la infección.
55
eeuuuuuuuuU
uuU
uUuuuuuuuueeeuuuuuuUuuuuuuuuuuuuuuuuuuuuuuuuu

TABLA S.l.- Datos referentes a los inmunocomplejosutilizados en la determinación deactividad anti-CD4.
¡CC % Act. «004 CLINICA 0D4/gI
-310 Neg. 158% 02 239
1-315 Pos. 731% 82 385
-327 Neg. 113% Al 901
1-349 Neg. 254% 03 154
1-285 Neg. 259% A2 315
1-301 Neg. 204% El 629
-303 Neg. 114% 03 63
-371 Neg. 253% 03 32
1-372 Pos. 272% A2 396
-373 Neg. 193% A3 266
1-374 Neg. 71% A2 448
1-375 Neg. 124% Al 648
1-388 Pos. 812% 02 252
.359 Pos. 375% A2 396
-358 Pos. 312% 82 378
1-355 Neg. 172% A2 290
(-356 Neg. 229% A3 176
1-361 Neg. 73% 02 365
3-362 Neg. 85% Cl 507
-364 Neg. 144% Al 640
1-365 Neg. 175% 02 288
1-366 Ncg. 190% 02 312
1-387 Neg. 80% 03 120
1-376 Neg. 96% 03 99
¡CC% Act. «CD4CD4/¡sl
(Identificaciónde la muestra)(% de actividad anti-CD4)(Linfocitos T CD4+ circulantes)
56
U
U
uueeuueueueuuuU
eeuuuueuU
uU
uu
ueuU
uueeuuuuuu
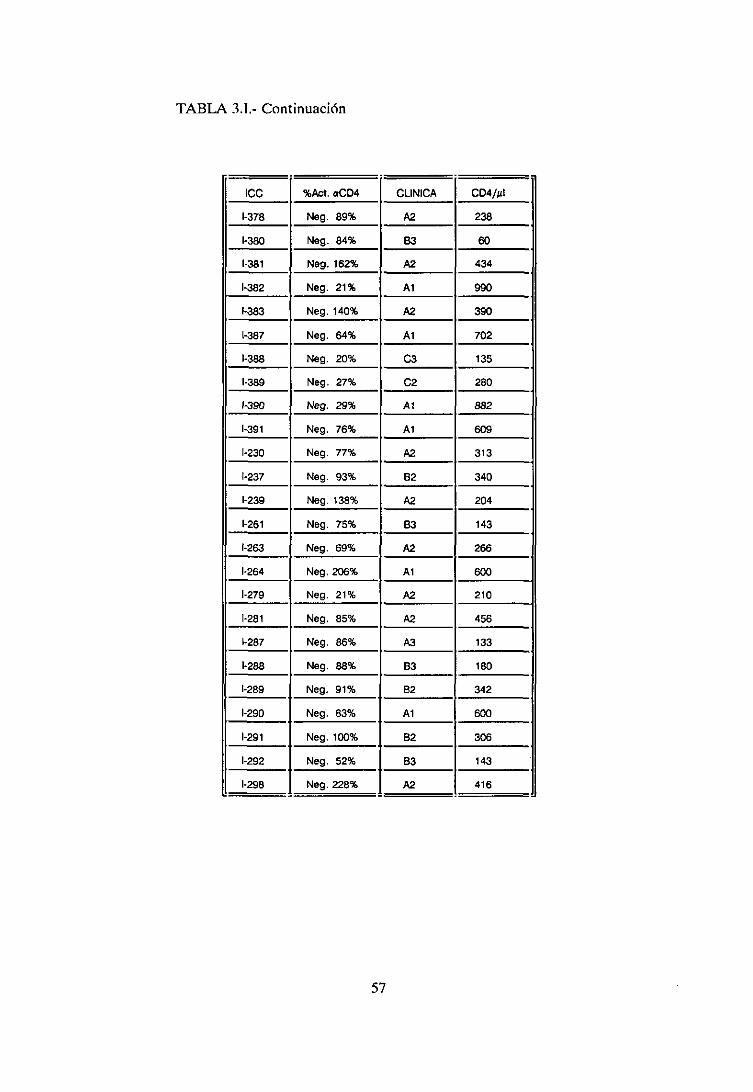
CC %Act. «004 CLíNICA CD4/gl
1-378 Neg. 89% A2 238
-380 Neg. 84% RS 60
1-381 Neg. 162% A2 434
1-382 Neg. 21% Al 990
1-383 Neg. 140% A2 390
-387 Neg. 84% Al 702
1-388 Neg. 20% 03 135
1-389 Neg. 27% 02 280
1-390 Nog. 29% Al 882
1-391 Neg. 76% Al 609
1-230 Neg. 77% A2 313
1-237 Neg. 93% 62 340
1-239 Neg. 138% A2 204
1-261 Neg. 75% RS 143
1-263 Neg. 89% A2 266
1-264 Neg. 208% Al 600
1-279 Neg. 21% A2 210
1-281 Neg. 85% A2 456
1-287 Neg. 88% AS 133
1-288 Neg. 88% RS 180
1-289 Neg. 91% 62 342
1-290 Neg. 83% Al 800
1-291 Nog. 100% 62 306
1-292 Neg. 52% 63 143
1-298 Neg. 228% A2 416
57
eeueueuuueu
TABLA 3.1.- Continuación
uueuuuueuuueeeuuuuuueeuuuuuueeuuu

TABLA 3.1.-Continuación
00 %Act. «004 CLíNICA 004/al
1-304 Neg. 83% A2 304
1-305 Neg. 50% 02 225
1-306 Neg. 68% CI 962
-307 Neg. 128% B2 224
1-308 Neg. 102% 02 364
1-309 Neg. 90% 02 247
-311 Neg. 92% Rl 760
1-313 Neg. 122% 02 336
1-314 Neg. 120% A2 227
1-317 Neg. 97% 02 440
-328 Pos. 328% Al 799
.331 Neg. 124% A2 227
-335 Neg. 115% 02 323
1-338 Pos. 367% B3 143
-338 Pos. 269% 03 24
1-339 Pos. 377% Al 779
1-340 Pos. 489% Al 583
1-341 Neg. 195% 03 154
1-342 Pos. 439% Al 595
1-343 Neg.241% 03 39
¡.344 Pos. 305% B3 120
1-348 Neg. 213% 03 102
-350 Neg. 54% AS 276
1-352 Neg. 99% 02 208
¡.354 Neg. 79% 02 290
58
uU
ueeuuueuuuueuuuuuuuuauueueuuueuuueuuueeuueuueeuuuueuuuueu

TABLA 3.¡I.- Frecuenciade inmunocomplejosconactividadanti-rCD4en pacientesinfectadospor HIV, divididosen funciónde suestadioclínico y susniveles de linfocitos TCD4+ en sangreperiférica.
FIGURA 3.1Actividad anti-rCD4 en inmunocamplejos
de pacientes infectadas por HIV
% Actividad antl-rCD4
810731
439489
36 69 376 8 306
312
IZUZEIZ Li Mt
272
1 2 3 4 6 6 7 8 9 10 11 12
lOO con una actividad anti-r004 262%Actividad snti-rOD4 en lnmunocompleJosde pacientes no Infectados • 132~ 66%
59
aeeeeeuueuuU
uuuuuueeuee
1000
800
600
400
200
o
ueuueuueeuuuuueuuuuueueeeuueUueueuuu
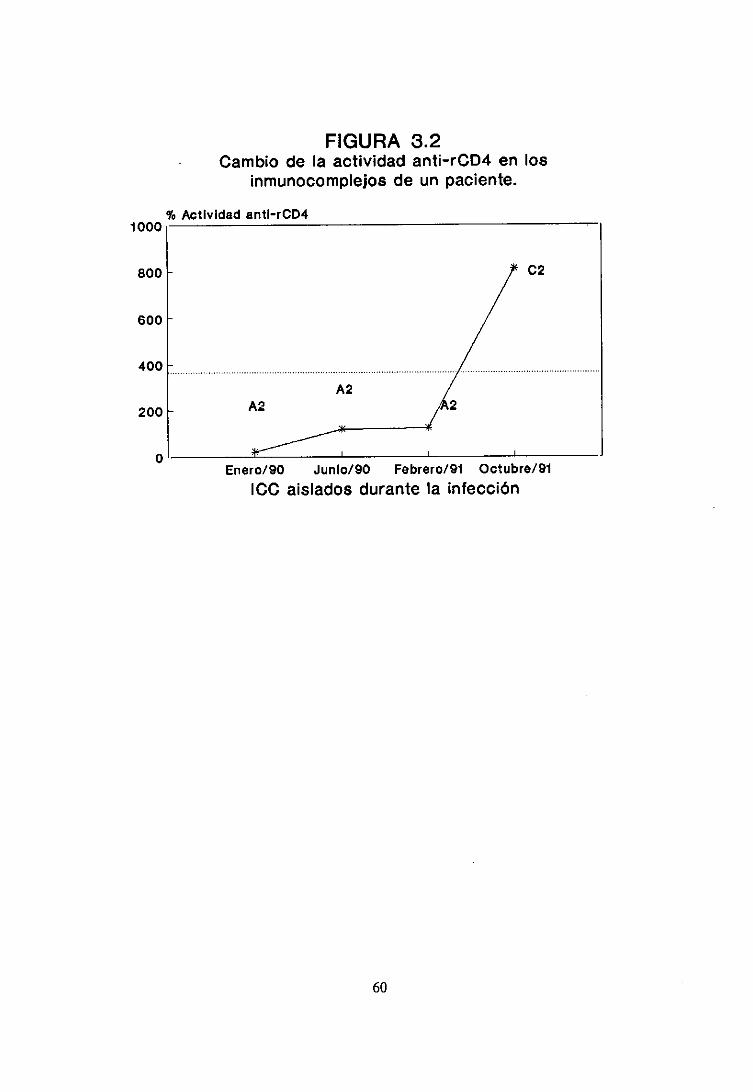
eeeeeeuu FIGURA 3.2e
Cambio de la actividad anti-rGD4 en los• inmunocomplejos de un paciente.
e % Actividad anti-rCD4• 1000uu
800uu 600eu 400uu• 200ue 0uueuuueeuueuuuueuueueeeuueUu
60uuUee
Enero/90 Junio/90 Febrero/91 Octubre/91lOO aislados durante la infección

ueuuu• * Unión de los Inmunocomplejos circulantes a la molécula CD4 presente en extractos
• de membrana de una línea linI’oblástica T humana CD4+.uu• Paradeterminarsi los inmunocomplejosaisladosdel suerodepacientesinfectadoseran
• tambiéncapacesde unirsea la moléculaCD4 expresadaen la superficiecelular se utilizaronu
extractosde membranade una línealinfoblástica T humanaCD4+.
• Al igual queenelestudioanterior,en el quesevaloró la unión delosinmunocomplejos
a unamoléculaCD4 recombinantesoluble,la unión de los inmunocomplejosa lamoléculaC04u• presenteen los extractosde membrana,fijadosen placasdepoliestireno.sellevó a caboconun
• anticuerpoanti-IgG humanaconjugadocon peroxidasa.
Los resultadosen densidadesópticas se transformarontambién en porcentajesdeu• actividadanti-CD4,considerándoseuna actividad positiva cuandofue =210%,ya queen el
• grupocontrol analizado,formadopor 11 muestrasde inmunocomplejosno relacionados,aislados
del suerode pacientesconArtritis reumatoide,se detectóun % de actividad mediade 110 ±u• 50%.
• El estudioseUevó a cabocon muestrasde inmunocomplejosaisladasdel suerode 33e pacientesinfectados,incluidos en función de su estadoclínico, en unacategoríaA (57%), E
• (18%)y C (25%). En basea susniveles de linfocitos T CD4+ seagruparonen tresniveles: 1
• (32%), 2 (48%> y 3 (20%).uEn un 28% de los inmunocomplejosse detectóesta actividad anti-CD4, conun valor
• medio de 318,7± 123%, mientrasque en el 72% de los inmunocomplejosno fue detectada.
u Aunque no se reflejó una relación entre la presenciade actividad anti-CD4 en los
U inmunocomplejosy el estadoclínico de los pacientes,ya queun 57% de los inmunocomplejosu• con actividad anti-CD4correspondíaa pacientesasintomáticosy un 43% a aquellosincluidos
u en unacategoríaB ó C, seobservóqueestosinmunocomplejospertenecíanmayoritariamenteu• (85%) a pacientescon nivelesde linfocitos inferioresa 500/pl (Tabla 3.111).
• Sin embargo,al compararla media de los niveles de linfocitos T CD4+, (349 ± 180u
células/pl), de pacientescuyos inmunocomplejosteníanunaactividad anti-CD4, conla media• obtenida,(424 ±269células/pl),en aquellospacientesconinmunocomplejossin estaactividad,
• seobservóqueno existíaunadiferenciasignificativa (p>O,OS).uu• En un posterioranálisis,realizadoen trespacientesinfectados,cuyos inmunocomplejos• presentabanunaactividadanti-CD4, se comprobóqueestaactividad también aparecíaen elu
sueroy en los inmunocomplejosdisociados.uuu 61uuuuu

euuuu• Por último, en la valoración de la unión de los inmunocomplejosa la molécula CD4
Uexpresadaen extractosde membrana,serealizarondospruebasde control,en lasqueseanalizóla disminución de la actividad anti-CD4 en los inmunocomplejosal realizar, por un lado,
• diluciones seriadasde los inmunocomplejos(Figura 3.3), y por otro lado, por el efectou
competitivode la unión de un anticuerpomonoclonalanti-CD4humanode ratóna la molécula
• CD4 (Figura3.4).uuuuuuueuueuuuueeueuuuuuuueeueuuueeuUueu• 62
uuuuu

TABLA 3.¡II.-
II
0.3
0.25
0.2
0.16
0.1
0.06
Frecuenciade inmunocomplejoscon actividad anti-C04 en pacientesinfectadospor H¡V, divididos en función de su estadioclínico y susnivelesde linfocitos T CD4+ en sangreperiférica.
uAct. anti-CD4 > 500/¡sl < 500/pl
15% 85%
FIGURA 3.3Disminución de la actividad anti-0D4
detectada en uno de los inmunocomplejos
o-o.0.271
0.257
1/200 1/300 1/500Distintas diluolones
1/1000
63
Estado Clínico Niveles lin.CD4
uuuueuuuueuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu

FIGURA 3.4Inhibición de la actividad anti-OD4
por anticuerpos frente a la molécula 0D4
D.O.
DIlucIón 1/200 de la muestra
~ ico W ícc ~ Ac. antl-004
Inhibición debida a la unión de unAc. monaclanal anti-0D4 a la molécula
0D4, expresada en extractos de membrana.
64
0.4
0.3
0.2
0.1
o
uuuueuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu
1/200

Uuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu• 4.1.2.- Potenciación de la infección por los Inmunocomplejos.u
4.1.2.1.- Unión de los ICC a linfocitos de sangreperiférica.u
4.1.2.2.-¡mplicaciónde los ICC en la inhibición del crecimientocelular.
uuuuuuuu

euuuuU 4.1.2.1.- Unión de Inmunocomplejos circulantes a linfocitos de sangre periférica.uuu Una vezcomprobadala capacidaddelos inmunocomplejoscirculantes,aisladosdelsuero
• de pacientesinfectados,paraunirsea una moléculaCli» recombinanresolubleo expresadaenu
extractosde membrana,seestudióla capacidadde los inmunocomplejosde unirsea la molécula• nativa, expresadaen linfocitos T CD4+ de sangreperiférica.
uu
La unión de los inmunocomplejosa la superficiede estoslinfocitos podría inducir una• respuestacitotóxica mediadapor célulasT dependientede anticuerpos(ADCC), observadain
u vitro por Lyerly et al. (59) con anticuerposcitotóxicosfrentea antígenosviralesexpresadosenu
la membranacelular. Esto supondríael agravamientodel estadode inmunodeficienciadel
• pacienteinfectado.
La valoración de la unión de los inmunocomplejosa CD4 se llevó a cabo en doceu• inmunocomplejosde sujetos seropositivos,en los que previamentese había detectadouna
• actividadanti-CD4 por su capacidadde unión a una moléculaCD4 soluble recombinante,y
e cinco inmunocomplejosaisladosdel suerode individuos no infectados,utilizadoscomocontrol.uEl análisisde la capacidadde estosinmunocomplejosde unirsea linfocitos de sangre
• periférica de sujetosnormalesse realizó indirectamente,por citometría de flujo, valorandola
inhibición de la unión de un anticuerpomonoclonalanti-CD4conjugadoconfluoresceína,(Acmu• anti-CD4-FITC),a linfocitos de sangreperiférica de sujetossanos.
U De estegrupo de inmunocomplejos,tan solo tres de ellos inhibieron de una forma
significativa,superiora un 50 % de inhibición, el marcajede linfocitos de sangreperiféricaconuu esteanticuerpomonoclonalfrente a CD4, en un 60, 62 y 90 % respectivamente(Figura 4.1).
• Estos inmunocomplejosfueron aislados del suero de dos pacientesasintomáticos,u incluidosen unacategoríaA2 de la infección,y de un pacienteen un estadiomásavanzadoC2
u (Tabla 4.1).
Estosresultadossugierenquela unión de los inmunocomplejosa la moléculaCD4varíau
en función de la forma en que se encuentreesta molécula, bien en su forma soluble y
U recombinante,o expresadaen extractosde membrana,o bien,en lasuperficiecelular,formaqueu
reflejaríamejor unasituaciónin vivo.En la figura 4.2 semuestrandosde los histogramasobtenidospor citometríade flujo en
• los queseapreciala inhibición del marcajede linfocitos de sangreperiféricaconun anticuerpo
monoclonal anti-CD4-FJTC cuando estas células fueron previamente incubadas cone• inmunocomplejosaisladosdel suerode un pacienteinfectadopor HIV (1.
uu• 65
euuuu

euuuuU La unión de los inmunocomplejosa la superficiecelular se comprobómedianteunu
ensayodirectoen el queseutilizó un anticuerpomonoclonalfrentea la InmunoglobulmnaIgO• humana,conjugadocon fluoresceína.
• En estos ensayosse observóparalelamenteuna disminución del marcajecon unu
anticuerpoanti-CD4-PEen linfocitos de sangreperiférica y un aumentodel marcajecon un
• anticuerpoanti-IgG-hu-FITC(Figuras4.3 y 4.4).
uu
La unión de los inmunocomplejosa la superficiede linfocitos T CD4+ podría estaru mediadapor anticuerposanti-idiotipo frentea anticuerposanti-gpl2O, aunqueno se puede
u descartarque estaunión seadebidaa la presenciade antígenogp 120 en los inmunocomplejos.uEn los tresinmunocomplejosde detectóla presenciade la proteínap24 del virus y en
• dosde ellos, sedeterminóla presenciade anticuerposanti-gpI2Opor WesternBlot (Tabla4.1).uuuueuuuuuuuuuuuuuuuuuuuuuuueuuuu 66
uuuuu

TABLA 4.¡.- Datosreferentesa losinmunocomplejosen los queseconfirmó sucapacidaddeunión a linfocitos de sangreperiféricapor citometríade flujo.
¡CCICC
% de Inh. delmarcajede PBLscon a-CD4-F¡TC
% Act.a-rCD4
¡CC¡sg/ml
Ag¡CCpg/ml 1_Ac-HIV
50 a-gpl2O
Cat. CD4/pl
1 60% POS. 557 A2 396
2 62% POS. 255 54 Neg. A2 238
3 90% POS. 855 5 a-gpl2O C2 300
ICC (¡sg/mí)Ag ICCCat.CD4/pl
(Nivelesde Inmunocomplejoscirculantesen suero)(Presenciade antígenop24 en los inmunocomplejos)(Categoríade la Infección por HIV)(NiveLesde linfocitos T C04+ circulantes)
FIGURAInhibición del marcaje
ant¡—0D4-FITC en PBLs
100
80
60
40
20
o
4.1con anticuerposincubados con lOO
% Inhibición90
62
— 2012V] 16HH~H4í3 í~-J~L3- i,,,,rfl,i,,j,~mmm
1 2 3 4 6 6 7 8 9 10lnmunocomplejos con actividad anti-rOD4
% de inhibición no signifIcativo < 26%
*
12
67
euueUuuuueuuuueuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu

FIGURA 4.2.- Inhibición del marcajecon un anticuerpomonoclonalanti-CD4-F¡TC,en
un 60%, en linfocitos de sangreperiférica de sujetos no infectados,
previamenteincubadoscon inmunocomplejosaisladosdel suerode un
pacienteinfectado(1
(A)
(E)
LGFL
(A) Marcaje de linfocitos de sangreperiféricacon anti-CD4-F¡TC: (30%).
(B) Marcaje de linfocitos de sangre periférica con anti-CD4-FITC,
previamenteincubadoscon inmunocomplejosaisladosdel suerode un
pacienteinfectado(1: (12%).
68
eUueuueeuuuuueuuuuueueuuuueueuueeuuuuuueuueuuuuuuuuueeueueu
LGFL

FIGURA 4.3Alteración del marcaje con anti-0D4-PE y
anti-lgGhu-FITC en PBLS por lOO
Frecuencia (%)
control (Pala) m PBLa + icc (1)
Disminución y aumento del marcaje con Acanti-004-PE y antl-lgGhu-FlTc en PBLa
incubados con ico de un pacIente HlV~
69
403630262016106o
Anti-C04-PE Anti-lgG4u-FiTcAc. monoclonal utilizado en el marcaje
euuuUuueuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuuu

FIGURA 4.4.-
(A)
(C)
Alteración del marcajecon un anticuerpoanti-CD4-PEy anti-TgG-hu-
FITC en linfocitos de sangre periférica previamente incubadoscon
inmunocomplejosaisladosdel suerode un pacienteinfectado(1
(B)
(D)
(A) Marcaje de linfocitos de sangreperiféricacon anti-CD4-PE:(32%).
(8) Marcaje de linfocitos de sangreperiférica previamenteincubadoscon
inmunocomplejosaisladosdel suerode un pacienteinfectado (1 con -
anti-CD4-PE:(15%).
(C) Marcaje de linfocitos de sangreperiféricacon anti-lgG-hu-FITC:(4%).
(D) Marcaje de linfocitos de sangreperiférica previamenteincubadoscon
inmunocomplejosaisladosdel suerode un pacienteinfectado (1 con
anti-IgG hu-FITC: (¡4%).
70
eueuuuuuuuuuuuuuuuuuuuuuuuuuueuuuuuuuuuuuuuuuuuuuuuuuuuuuuu

4.1.2,2.- Inhibición del crecimiento celular por Inmunocomplejos circulantes.
La capacidadde los Inmunocomplejoscirculantes, aisladosdel suero de pacientes
infectadospor el virus de la InmunodeficienciaHumana,de alterar el crecimientocelular, se
valoró en cultivos iii vio-o. En estoscultivosse determinóla inhibición de la proliferaciónde
linfocitos de sangreperiféricaestimuladoscon PRA.
La inhibiciónproducidapor la presenciaen el mediodecultivo delos inmunocomplejos
se midió a las 72 horas mediantela detecciónde una disminución de la incorporación de
Timidina H3 en el ADN de los linfocitos.
En esteestudiose determinóel efectode la presenciaen el medio de cultivo de siete
inmunocomplejos(Z43, Z39, Z23, Z53, Z42, Z44y Z45), aisladosdel sueropacientesinfectados
por HIV, incluidos por su estadioclínico en distintascategoríasde la infección.
Con el propósitode compararel efectode los inmunocomplejosasociadosa la infección
por HIV con otros inmunocomplejosobservadosen pacientescon otras enfermedades,se
utilizaron ademáscinco inmunocomplejosno relacionados,aisladosdel suerocíepacientescon
patologíasdiferentes,HepatitisE (N7 y NIO) y artritis reumatoide(N20, N30 y N56).
Paradeterminarla inhibición de la proliferación celularsecalculó el % de Inhibición
de la proliferación que suponía la presenciaen el medio de cultivo de cada muestrade
inmunocomplejos,en funciónde la disminuciónde timidina H3 incorporadapor el ADN de los
linfocitos con respectoal control (Linfocitos estimuladoscon PI-lA sin la presenciaen elmedio
de cultivo de inmunocomplejos).
Comoresultadode estecálculo, en los ensayosde proliferacióncelularseconfirmó que
3 de los 7 inmunocomplejosanalizados,Z39, Z53 y Z42, fueron capacesde afectar el
crecimientocelular, inhibiendo de forma significativa la proliferaciónde linfocitos de sangre
periféricaestimuladosconPHA. La presenciade estostresinmunocomplejosen los mediosde
cultivo supuso un % de inhibición de la proliferación celular de un 52, 40 y 69%
respectivamente,mientrasque la media del % de inhibición de la proliferaciónobtenidocon
inmunocomplejosasociadosaotraspatologíasfue de un 10,5 ±7,1 % (Figura4.5).
Los tres inmunocomplejosque inhibieron la proliferación fueronaisladosdel suerode
pacientesinfectadosque por su estadoclínico fueron incluidos en las categoríasC3, A3 y C3
respectivamente(TABLA 4.11).
71
euUuuUeuUuuuuuuUeuuUuuuuuuuuuuUuuuuuuueuueuueuU
uuuuueuuuuuu

euuuU• El mecanismoporelcual los inmunocomplejospuedenafectarelcrecimientocelularno
lo hemosanalizadoen estetrabajoperoen éste,quizáspodría estarimplicada la unión de losu• inmunocomplejosa la moléculaCD4.
• Respectoa esteproceso,algunosinvestigadores,comoBanda et al. (64), hansugeridou
un modelo en el que la interacción de la molécula CD4 expresadaen los linfocitos T
• cooperadoresy complejosgpl2O-anti-gp120podíadesencadenarun fenómenodeapoptosisyhan
e propuestoqueestefenómenopodríaestarimplicadoen deplecióndelinfocitosT CD4+ duranteu
la infección.
uuueueuuUuueueueuuuueuuuueUuuuuuuuuue• 72uuuuu
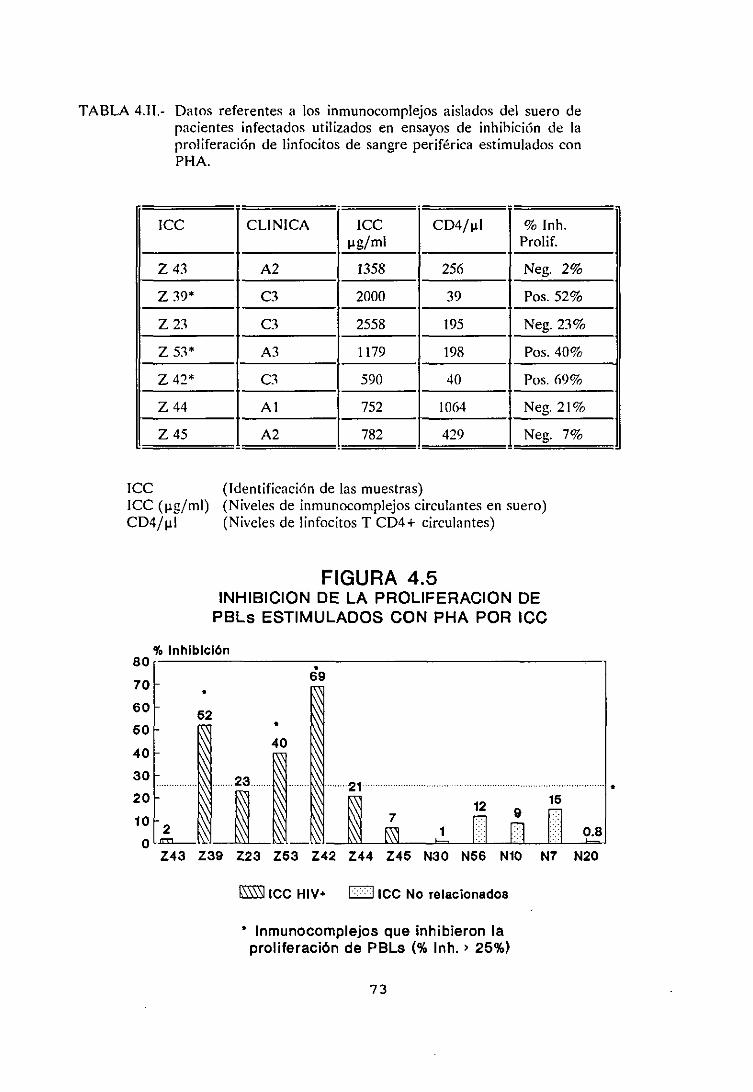
TABLA 4.11.- Datos referentesa los inmunocomplejosaisladosdel suerodepacientesinfectadosutilizados en ensayosde inhibición de laproliferaciónde linfocitosPHA.
de sangreperiféricaestimuladoscon
ICC CLíNICA ¡CC¡sg/ml
CD4/pl % Inh.Prolif.
Z 43 A2 1358 256 Neg. 2%
Z 39* C3 2000 39 Pos.52%
Z 23 C3 2558 195 Neg. 23%
Z53* A3 1179 198 Pos.40%
Z 42~ C3 590 40 Pos.69%
Z44 Al 752 1064 Neg.21%
Z 45 A2 782 429 Neg. 7%
ICC (¡sg/mí)(Identificaciónde las muestras)(Niveles de inmunocomplejoscirculantesen suero)(Nivelesde linfocitos T CD4+ circulantes)
FIGURA 45INHIBICION DE LA PROLIFERACION DE
PBLs ESTIMULADOS CON FHA POR lOO
% inhibición
* Inmunacomplejos que inhibieron la
proliferacián de PBLs (% InI,. > 26%>
73
UuUUUUuUuUUuuUUuUuU ¡CC
CD4/¡s 1
80
70
60
60
40
30
20
10
o
UUuUUuUUUuUUUuUuuuuUUuuuuuUuuuu
Z43 Z39 Z23 ZGS Z42 Z44 Z46 NSO N66 NIO N7 N20
:cc HiV~ m icc No relacionados

UUUUUUUUUeuUUuUUUeeUUUeUUUUUeeuuuUUUuUeUUUUu• 4.1.3.- Capacidad infectiva de los inmunocomplejos circulantes.
UUUUUUUuuUuUUU

auUUUU
u La presenciade partículasvirales formandopartede los mnmunocomplejoscirculantesu• fue puesta en evidenciapor Bolognesi et al. en un trabajo publicado en 1989, en el que se
U postuló la infecciónde monocitosy macrófagosvía complejosde anticuerposy virus (122).U
Estosexperimentosin vio-o sugierenque los inmunocomplejospodríantenerun papel
U importante en la patogénesisde la infección, en cuanto a que pueden incrementar la
u infectividad.uUU La capacidadde los inmunocomplejosde comportarsecomo complejosinfectivos fue
analizadaen estetrabajoen cultivosde linfocitos de sangreperiféricaestimuladosconPHA yu• en presenciade distintos inmunocomplejos.
U Paraesteanálisissedeterminóla presenciade antígenop24 en los sobrenadantesde los
cultivos por un ensayoinmunoenzimático.U
Los cultivosse realizaronen presenciade diferentesimunocomplejosaisladosdel suero
U de pacientesinfectadosy fueronmantenidosduranteun máximo de dos semanascon PHA ee
¡nterleuquina-2recombinante.• Los inmunocomplejoscorrespondíana pacientesincluidosen funcióndesuestadoclínico
• en las siguientescategoríasde la infección: Al (14%), A2 (5%), BI (14%), 82 (5%), Cl (5%),
C2 (19%) y C3 (38%).U• De los 25 inmunocomplejosanalizados,siete indujeron la síntesisde antígenop24 en
• cultivos de linfocitos de sujetossanos,obteniéndosedistintasconcentraciones,en un rango deU 14 y 25 pg/ml. a los 6 diasde cultivo (Figura5.1). Cinco de estosinmunocomplejospertenecíanU• a pacientesincluidosen unacategoríaC de la infección.UUU La viabilidad celular se cuantificó por exclusióndel azul tripan al inicio del cultivo y a• los 4,6,8 y 10 díasde cultivo, obteniéndoseun 90% de viabilidad a los 4 días, un 80% a los 6 y
U 8 díasy un 50% a los 10 díasde cultivo.UUU Para comprobarque la síntesisde antígenop24, detectadaen el sobrenadantede los
u cultivos de linfocitos de sangre periférica realizados en presencia de diferentesUU inmunocomplejos,estabaasociadaa una infección de los linfocitos, se analizóla presenciade
U ADN viral integradoen el genomade estascélulas.UUUUUu 74UuUUU

UuUuUU En esteanálisisse amplificaron dosfragmentosdiana del ADN viral, correspondientes
a lasregionesenv(gp4l) y poí, conoligonucícótidosespecíficos,mediantela reacciónen cadenauU de la enzimaTaq ADN polimerasa.U Esteensayose llevó a cabo en tres de los cultivosde linfocitos realizadosen presencia
e de inmunocomplejos,detectándoseen dos de los casosADN del virus integradoen el genomaU• de estascélulas(Figura 5.2).
UU A pesarde que solo en uno de los cultivos se obtuvo síntesisde antígenop24, esteu• último ensayoapoya la hipótesisde que en Inmunocomplejosde pacientesinfectadospor el
U virus de la ¡nmunodeficienciaHumanapuedenexistir partículasvirales íntegrascapacesdeu
infectar linfocitos T CD4+.• Esta capacidadinfectiva de los inmunocomplejoscirculantescorroboraríael papel
U patogénicoque tendríanlos inmunocomplejoscirculantesen la infección.uuaUUUUU flGURA 5.1• p24 en el sobrenadante de cultivos de• PBLs realizados en presencia de iCC
U p24 (pg/mi)U 30
U• 26
U• 20
Uu 15
u louU 6
uu 1 (03) 2 (Al> 3 (02) 4 (CS> 6 (B1) 6 (C2> 7 (03>
U• Inmunocomplejas de pacientes incluidos• en distintas categorías de la infección
UuUUUU 75uUUUU
26
16 19714



UuuuuuuuUUuuuUuUUuuUuuuuuuuuUUUUuuuUuuuuuuuUU
4.2.- PRESENCIADE LINFOCITOS T CDS+ EN PACIENTES INFECTADOSe CAPACESDE LISAR LINFOCITOS T CD4+ ESTIMULADOS CON PHA.uuuUeUuuuUUuu

uUuuuu
Ante algunoshallazgosserológicos,observadosen pacientesinfectadospor el virus deu
la inmunodeficienciahumana,comolapresenciaen suerodeautoanticuerposfrentea antígenos
U linfocitarios (130), se ha sugeridoque durantela infección podría generarseun fenómeno
uU
Los anticuerposobservadosen la infección por ¡-¡IV que se han descritohasta el
• momento están dirigidos frente a moléculas de clase II del Complejo Mayor deu
Histocompatibilidad(95), la moléculaCD4 (131), e inclusofrentea la proteínabásicade mielinau (132).
U Teniendoen cuentaestosdatosseha sugeridocomohipótesisparaexplicarla depleción
u de linfocitos T CD4+ queotros mecanismos,ademásdel propio efectocitopáticodel virus,u• como un mecanismoautoinmunefrente al SistemaInmuney probablementefrente al tejido
• nervioso (133-111),podrían también estar implicados en la disminución de esta poblaciónU
celular.
• Estarespuestaautoinmunepodría también estarmediadapor linfocitos T y para su
U estudiodecidimosanalizarla respuestacitotóxicade linfocitosT CD8+ de pacientesinfectadosU por el virus de la inmunodeficienciahumanafrentea linfocitosT CD4+ de sujetossanos.Este
• análisisse llevó a cabocon ensayosde liberaciónde Cg’.
uu
Comocélulasdianase utilizaron linfocitos de sangreperiféricade pacientesinfectadosu y de personassanas,activadoscon PRA durantesiete díasy marcadoscon Cr ~ así como
• célulasde una línea linfoblásticaT humanaCD4+.uU• Como células efectoras se utilizaron linfocitos T CD8+ seleccionadosde sangreu
periféricade pacientesinfectadosy sanosmedianteesferasinmunomagnéticascubiertasconun• anticuerpomonoclonalanti-CDS. El fenotipode estascélulasse analizóporcitometríade flulo,
• comprobándosequeexpresabanCD8 en su superficie.UU• Característicasde las célulasefectorasseleccionadas:Uu
Pureza: =89% CD8+• Rendimiento: 75-80%
• Viabilidad: =97%U
Contaminación por células NK: = 1%uUu 78uUUUU

uuUuUU En primerlugar serealizóun estudioparacompararla respuestacitotóxicafrentea una• línealinfoblásticaT humanaCD4+, mediadapor linfocitos de pacientesinfectadosy personas
sanas.Esta respuestase analizóen sietepacientesincluidospor su estadoclínico en distintasu• categoríasde la infección, A (2), B (2) y C (3), y cinco individuos sanos.
U De los ensayosde liberación de CI-~’ se infirió que la respuestacitotóxica mediadapor
u los linfocitos de los pacientesinfectadosfrente a la líneatumoral era menorquela observadauU en el grupocontrol de sujetossanos(p<O,OS).La mediadel porcentajede lisis correspondienteu acadagrupo fue de 10,8 ±6,9%y 21,4 ±8,9% respectivamente(Figura 6.1).
U Esta observación,acercade una disminución de la respuestacitotóxica en pacientesuu seropositivosrespectoa la capacidadcitotóxicanaturalfrenteal desarrollode tumores,mediadaU por célulasNK, tambiénha sido demostradaconotraslíneastumoralespor otros investigadoresU (134).UUU En segundolugar se estudió la respuestacitotóxica mediadapor células T C08+ de
sujetosinfectadosfrentea célulasT CD4 + procedentesde sujetossanoso infectados,utilizandoUu como control célulasT C08+ de sujetossanosy siguiendoel siguienteesquemade trabajo:uU DISTINTOS ENSAYOS DE OITOTOXIOIDADuu
CÓI. efectoras (E) 1 CéI. diana (D)
UUuUuuUUuu _________________________
uu En los ensayosde citotoxicidad del tipo A y E, solamenteserealizarondospruebascone• linfocitos T CD8+ de un pacienteinfectado(ZGI) y de un sujetosano(NIO), debidoa queun
• óptimo marcajecon C¿’ de las célulasdiana, en estecasolinfocitos de sangreperiférica delu pacienteinfectado,requeríaunaprevia estimulaciónconPHA. Esta estimulacióninduciría la
u activación del provirus en las células infectadas,lo cual dificultaría manteneruna razonable
viabilidad en los cultivos, necesariaparael marcajeconcromo de los linfocitos.uUU 79uuuUuU
E E
Lin. T 0DB. HiV~ Lin. T GD8. HiV- ¡
Aj D IB loLin. T 0D4. HlV~ Lin. T 0D4. HiV-
D D
u

uuUUUU Ademásdeestadificultad, en aquellospacientesinfectadosquemuestrenun estadode
U inmunodeficienciamuy avanzado,seríadifícil obtenerun número suficientede linfocitos Tu• CD4+ en la poblaciónde las célulasefectotas.
• En estaspruebasse observóque los linfocitos T CDS+ del pacienteinfectadoeranUU capacesde lisar linfocitos autólogos(% lisis = 29%),mientrasque no sedetectóningún tipo• de respuestacitotóxica por partedel sujetosano.Uu
Sin embargo,dadoquenuestrointerésestabadirigido, no al estudiode ~TLS específicosU frente a antígenosvirales, sino al estudio de la presenciade linfocitos T CD8+ frente a
linfocitos T CD4+ no infectados,se realizaronlos ensayosde citotoxicidad del tipo C y D.U
Como células diana se utilizaron linfocitos de sangreperiférica de sujetos sanos,
U estimuladoscon PHA y marcadosconU Corno célulasefectorasse seleccionaronlinfocitos T CD8+ de sangreperiféricade 30UU pacientesinfectadosincluidos en unacategoríaAl (3), A2 (3), A3 (1), BI (2), B2 (4), B3 (6),• Cl (1), C2 (3) y C3 (7).U Comogrupocontrolse utilizaron linfocitosT COS+ de 10 sujetossanos.Su citotoxicidaduU frentea linfocitos no infectadosfue negativa,obteniéndoseun porcentajede lisis en todoslos• casosinferior al 5%.uuU En esteestudioseobservóquecuatrodelos pacientesinfectadosseleccionados,un 14%U del total, presentabanlinfocitos T CDS+ capacesde lisar linfocitos estimuladoscon PHA deu
sujetossanos.Estospacientesseencontrabanen estadiosavanzadosde la infección,B2, C3, C3• y C2 respectivamente(Figura 6.2).
U El porcentajede lisis obtenidoen estoscasosfue de un 47% (Z22), 12% (Z43), 22%u
(Z50) y 18% (Z52) (Figura 6.3). La relacióncélulaefectora- célula diana,(E:D). en estoscasos• fue (4:1), (20:1), (3:1) y (25:1) respectivamente.
uu
En estosensayosdecitotoxicidadsecomprobóquela respuestacitotóxicaestabadirigidaU frente a la molécula C04, ya que la lisis fue bloqueadaen todoslos casoscon un anticuerpo
monoclonalespecíficode CD4. Por otro lado, se confirmó queesta lisis estabamediadaporuU linfocitos T CD8+ debido a que su actividad citotóxica se podía inhibir con un anticuerpou monoclonalfrentea CDS (Figura 6.4).U Tambiénsecomprobóque estarespuestacitotóxica disminuíaen funciónde la relaciónU• célula efectora/ célula diana(E:D) (Figura 6.5).uUu 80UUUuu

UuUuuU Por último, se pudo confirmar que esta lisis no estabadirigida frente a moléculasdeU
clase¡1 delComplejomayorde Hístocompatibilidad,ya queen ensayosparalelosseobservóqueU no era inhibida con anticuerposanti-DR. En la Figura 6.6 semuestraesteensayoen uno de
U los casos(Z43).UuU A pesarde que los cuatro pacientes,en los que se detectóesta respuestacitotóxicaU frente a célulasT CD4+ no infectadas,estabanen estadiosmuy avanzadosde la infección yu• presentabannivelesde linfocitos T C04+ en circulaciónmenoresa 400 células/pl,no sepuedeU considerarqueestarespuestaestéasociadaaun estadioavanzadode la infecciónya queno en
u todoslos pacientesestudiados,incluidos en unacategoríaC, se detectóestarespuesta(Tablau• 6.1).
U Se trataríamásbien,de un mecanismopatogénicomásqueapareceen algunospacientes
u infectadospor el virus de la InmunodeficienciaHumana, que podría estar implicado en lauU depleciónde linfocitos T CD4+ en estoscasos.UuuuUuUUuuUuUUuUuuUUuUuuUUUuUU 81UuUUU

FIGURA 6.1Respuesta citotóxica frente a unalínea linfoblástica T humana 0D4’
Pacientes HIV~ (n-7) Grupo control (~.s)Disminución de la respuesta citotóxica
en pacientesinfectados por HiV
FIGURA 6.2Incidencia de la respuesta citotóxica
frente a células T OD4~ no infectadas
50
40
30
20
lo
o
UUUUUUU
40
30 -
% lisis
20 -
UUuUuUUuUUUUUuUuUU
X-2t4~9
X- 10.8+6
10-
o
%lisis
uuuuuuUuuuUUuuUuuUuUUUUUeUUUUUu
Al A2 AS BI B2 BS Cl C2 03Pacientes (n.30) incluidos en distintas
categorías de la Infección
82
-w

FIGURA 6.3Respuesta citotóxica frente a PBLspor células T OD8~ de 6 pacientes
% lIsis celular
FIGURA 6.4Lisis inhibida por anticuerpos frente
aOD4yOD8
Lisis total EZ P.c. anti-cD4 P.c. antl-C08
83
uUuUUUuUUuUUUUuUeUUUuuUUUUuUU
60
so
40
30
20
10
oZ22 Z43 ZGO Z52 Z63 ZSS
células 1 CD8~ (Electoras>• % de lisie correspondiente a células T
CD8~ de 10 suietos sanos < 5%
% lisis60
60
4030
20
10
o
uuUUUUUUuUUUuUUuUuUUUUUuUUUUUU
Z22 ZSO Z52LinfooIlos T COS.

FIGURA 6.5Disminución de la lisis en función dela relación cél. efectora 1 cél. diana
FIGURA 6.6Lisis bloqueada por distintos
anticuerpos monoclonales
Z43
1 1
10
hlIII
Linfocitos T cD8+
Lisis total
~ Aa. antí-DA
m Ao. Aa. anti-CDS
m Ao.
84
% lisis
12
Z4310
14
12
10 -
a
6-
4
2
o
UUuUuUUUuUUUUUUuUuUeUuUUuUUUUUUUUUUUUUUuUUUUUUUUuuUUUU
2
(E:D)
de ¡Isis
1214
1210
8
6
4
2
a
6
uUuU
q

TABLA 6.1.- Datosreferentesal estadode los pacientesinfectados,cuyoslinfocitos T CD8+fueronutilizadoscomocélulasefectorasen los ensayosde citotoxicidadcelular.
Pao. Un. C044-/gl CLINICA % Lisis
ZiS 162 B3 Neg.
222 338 82 Pos. 47%
Z38 165 Neg.
237 315 82 Neg.
Z40 195 83 Neg.
741 114 AS Neg.
Z42 472 A2 Neg.
243 7 03 Pos. 12%
Z44 290 A2 Neg.
245 <200 83 Neg.
Z46 40 03 Neg.
749 570 Cl Neg.
250 27 C3 Pos. 22%
251 240 82 Neg.
252 217 02 Pos. 18%
253 810 Al Neg.
Z54 170 83 Neg.
ZSS 532 Al Neg.
258 304 02 Neg.
257 448 81 Neg.
258 90 83 Neg.
259 363 A2 Neg.
760 <200 03 Neg.
781 258 02 Neg.
262 <200 03 Neg.
283 266 B2 Neg.
Z64 15 03 Neg.
Z65 40 03 Neg.
Z66 569 81 Neg.
ZICO 644 Al Neg.
85
uUUuuUUUuuuuUUUUuuuuuuUUuuuuuUUuuuuuuUuUuuuUUUUuUUUUuuuUuUu

UuuUuuuUUuuUuuuUuuUuuuuUUUuuUuuuUUuUuuuu ~.- DISCUSION FINALuU 5.1.- Implicación de los inmunocomplejosen la infección.UU 5.2.- Mecanismosautoinmunesimplicados en la depleción de linfocitos Tu CD4+.
UuuuuuuuuuUuuU

UuuUUU Con los resultadosobtenidossevaloraronlos dosobjetivosplanteadosen estatesis.EnUu primer lugar, determinarla implicaciónde los inmunocomplejoscirculantesen la infeccióny enU segundolugar, analizarla posibleparticipaciónde un mecanismoautoinmuneen la depleciónu de linfocitos T C04+.UU• 5.1.- IMPLICACTON DE LOS INMUNOCOMPLEJOSEN LA INFECCIONuu
Aunqueel periodode latenciaprevio al SIDA alcanzaun términomediode 10 añosunau vez quese ha producidola infección, éstevaríade una forma muy drásticade unosenfermosU a otros (135)uU Los factoresqueinfluyen en la progresiónde la infección hacia un estadode SíndromeU de Inmunodeficienciano se conocenmuybien, a pesarde quesisehandescritovariosfactores
u inmunológicosqueseasocianconun peorpronósticodel sujetoinfectado,comola disminuciónudel númerode linfocitosT CD4+ en sangreperiféricao la disminucióndeanticuerposanti-p24,
• que indican un gradode infección avanzado(89).UU• La elevadaprevalenciade altos niveles de inmunocomplejosen el suerode pacientesu infectados,remitidosal Departamentode Inmunologíade la FundaciónJiménezDíaz, fue un
dato decisivoparavalorarsu posibleimplicación en la infección.uUU Niveleselevadosdeinmunocomplejosenpacientesseropositivos,determinadosmedianteu
ensayosde unión a Clq, asícomo defectosen el aclaramíentode éstosporel sistemaretículo-U endotelial, fueroninicialmentedocumentadosen 1985 (112,136).uu
En estetrabajo seobservaronniveles elevadosde inmunocomplejos(> 500 ¡sg/mí) en• el 72% de los pacientesanalizados,sin que la mayoría padecieseuna patologíaasociadaa
u Inmunocomplejos.uU La presenciade niveles elevadosno estabaasociadaal estadioclínico del paciente,enU cuantoa su relacióncon unade lastrescategoríasde la infección,perosi existíaunadiferencia
significativa (p<O,05)alcompararel gradode inmunodeficienciade los pacientes,yaquede unuu grupo de 53 pacientescon niveles de linfocitos T CD4+ en circulación inferiores a 500u células/¡sl. 42 de ellos presentabanniveles de inmunocomplejoselevados,superioresa 500U
¡sg/mí,mientrasquesolo 11 de ellos teníannivelesde inmunocomplejosinferioresa 500 ¡sg/ml.UUuU 86
uUuuu

UuUuUU Al analizarla composiciónde los inmunocomplejos,en función de la presenciao nédeU se observóunadiferenciasignificativa entrela detecciónde antígenop24 en
antígenop24, noU los inmunocomplejosy el estadoclínico en el que se encontrabanlos pacientes(p>O,OS).U Sin embargo, si se observóuna diferenciasignificativa con respectoa Jos niveles de
u linfocitos en sangreperiféricaquepresentabanlos pacientes,observándoseque la presenciaenU• los inmunocomplejosde antígenop24 es másfrecuenteen pacientescon niveles de linfocitos
U circulantesinferioresa 500/pl (p’c 0,05).UU Respectoa la presenciade antígenop24 en los inmunocomplejos,Lange, et al.,u observaronqueal principio de la infección elantígenop24 circulantepodía estarpresentesoloU formando parte de los inmunocomplejos,y que en estadios más avanzados,ademásdeuU detectarseantígenop24 en los inmunocomplejostambiénse podíadetectarlibre en el suero• (103).En nuestroestudioseobservóquedurantetodoelcursode la infecciónsepuededetectar
U antígenop24 formandoparte de los inmunocomplejoscirculantes,ya que fue encontradoenu• inmunocomplejosaislados del suero de pacientesincluidos en diferentescategoríasde laU infección. En algunospacientes,solo se detectóantígenop24 en los inmunocomplejosy no en
• el suero.u
Estos resultadosindican que en algunoscasosde la infección los inmunocomplejos
U podríanestarencubriendola presenciade antígenosviralesen circulación,alno serdetectados
U en suero,de tal forma queencubriríanuna fasereplicativadel virus.uUU Los inmunocomplejosen los quesimultáneamentese detectaronanticuerposfrente al
virus y antígenop24, (Ac+Ag+), aparecíanconmayorfrecuenciaen aquellospacientesqueporu• su estadoclínico fueron incluidosen unacategoríaC de la infección (46%)y solamentefueron
• detectadosen pacientescon nivelesde linfocitos T CD4+ en circulacióninferioresa 500/pl.U La presenciade estos inmunocomplejos,Ac+Ag+, indicaría un mayor riesgo deu• progresiónen la infecciónhacía un estadode SIDA.uuU Al valorarlasdistintasespecificidadesde los anticuerposfrentea lasdiferentesproteínasU delvirus,presentesen losinmunocomplejos,sedeterminóqueanticuerposanti-gpl2OaparecíanU en el 100% de los casosanalizados,mientrasquela frecuenciade anticuerposfrentea lasotrasUU proteínasdel virus era mucho menor, observándoseque anticuerposfrente a gp4l solo• aparecíanen inmunocomplejosde pacientescon algún síntoma clínico, incluidos en una
u categoría8 6 C de Ja infección.uuuuU 87uuuuu

uUUUUU Ante la observaciónen el suerode pacientesinfectadosde anticuemosfrente a laU moléculaCD4 porMuller et al. (137)y otrosautoanticuerposdescritosfrentea otrasmoléculasu• linfocitarias, como antígenosde clase11 del Complejo mayor de Histocompatibilidad(95), seU postulóla posiblerelevanciade un fenómenoautoinmuneen la infección por ¡-¡TV.U De acuerdocon estasobservaciones,seanalizóla presenciade actividad anti-CD4enU• inmunocomplejoscirculantesde pacientesinfectados.U La actividadanti-CD4en los inmunocomplejoscirculantesfue valoradaen función deu
su capacidaddeunirsea unamoléculaCD4 solublerecombinanteo aestamoléculaexpresada• en extractosde membranade unalínea linfoblásticaT CD4+ humana.Respectoal primer tipo
U de valoraciónseobtuvoqueun 16% de los inmunocomplejosmostrabaunaactividadanti-CD4.U Aunqueestaactividad fue observadaen un porcentajemuy bajo,y en los estudiosde
• significación (Chi-cuadrado)no se observóuna diferencia significativa entre el grado de
U infección y la presenciade actividad anti-CD4 en los inmunocomplejosaisladosde estosu• pacientes,no podemosdescartarla hipótesisde queéstostenganalgún significadopatogénicoU en algunos pacientes,ya que un 72% de los inmunocomplejosen los que se detectóesta
U actividad correspondíana pacientescon nivelesde linfocitos circulantesinferioresa 500/pl.UU La capacidadde unión de los inmunocomplejosa la molécula CD4 expresadaenU extractosde membranacelularfue observadaen un porcentajemayorde los pacientes,un 28%.U
Aunqueno se reflejó una relación entreel estadoclínico de los pacientesy la presenciadeU actividad anti-CD4 en los inmunocomplejos, si se observó que la mayoría de losU inmunocomplejosen los quesedetectóestaactividad(85%)pertenecíana pacientesconniveles
u de linfocitos en sangreperiférica inferioresa 500/¡sl.u• La unión de los inmunocomplejosa la molécula CD4 podría estar mediada por la
U presenciaen los inmunocomplejosde antígenogp 120 o bien, de anticuerposantiidiotipo, esU
decir,anticuerposdirigidos frentea anticuerposespecíficosde la proteínagpl2O del virus. Esta• última hipótesisfue sugeridapor Kowalsky,et al., al demostrarla presenciade autoanticuerpos
• frentea la moléculaCD4 en el suerode algunospacientesinfectados(130).uUU Ante la observacióniuz vio-o de inmunocomplejoscapacesde unirsea unamoléculaCD4U solublerecombinanteo expresadaen extractosde membrana,se estudió la capacidadde losuU inmunocomplejosde unirsea la superficiede linfocitos de sangreperiférica.u De los docecasosen los queseobservóunaactividadfrente a la moléculaCD4 soluble
U recombinante,solo tresfueroncapacesde unirsea la moléculaCD4 expresadaen la superficieUU celular.Uu• 88UuuUU

UUUU
La diferentecapacidadde los inmunocomplejosde unirsea la moléculaCD4, en función
u del estadoen queseencuentre,bienen unaforma recombinantesoluble,expresadaen extractosu• de membranas,o bien, en la superficiecelular,podría serdebida a la diferenteexposiciónde
u epítoposespecíficosde la molécula, quedependeríade la estructuraen quese encuentrelaU molécula máso menosdesnaturalizada.UU• La unión de los inmunocomplejosa la molécula CD4 y la hipótesis postuladaporU Morrow et al, sobrela presenciade partículasviralesformandopartede los inmunocomplejosu• (138), son dos datos quesuponenque los inmunocomplejoscirculantespuedenproducir en
• algunoscasosun agravamientode la infección al sercapacesde infectar nuevoslinfocitos TUU CD4+.u La detecciónde antígenop24 en los sobrenadantesde linfocitos de sangreperiféricade• sujetossanosestimuladoscon PHA, cultivadosen presenciade inmunocomplejosaisladosdelu
suerode pacientesinfectadospor H¡V, confirmaríala capacidadde los inmunocomplejosde• potenciarla infección. Estacapacidaddc los inmunocomplejosya fue demostrada¡a vio-o por
U Robinsonet al., quienesobservaronla infecciónde macrófagosmediadapor inmunocomplejosUU (84).• Estos resultados,junto a la detecciónpor la reacción en cadenade la Taq ADN
U polimerasade ADN viral integradoen el genomade los linfocitos, fueron decisivos parauU adjudicarunacapacidadinfectiva a los inmunocomplejos.U La confirmación de ADN vírico integrado en el genomacelular se realizó previa
extraccióndelADN delos linfocitoscultivadosen presenciade inmunocomplejosy amplificaciónU• de dos fragmentos diana del ADN viral, correspondientesa las regiones env y pol, con
U oligonucleátidosespecíficosy la enzimaTaq AON polimerasa.
Estaobservaciónindicaquelosinmunocomplejospuedenestarimplicadosenla infecciónu• y que in vko, podrían contribuir a la disminución de linfocitos T CD4+ al ser capacesde
U infectarnuevascélulas.U
Al igual que en algunasinfecciones,como las producidaspor Herpesvirus,reovirus y
• flavivirus, en la infección por el virus de la inmunodeficienciahumana los anticuerpos
• producidosademásde tenerun papelprotector,puedenserconsiderados,algunasveces,como
U anticuerpospotenciadoresde la infección (122).uU La capacidadde los inmunocomplejosde inhibir elcrecimientode linfocitos de sangreU periférica estimuladoscon PIlA fue observadaen un 33% de los casosanalizados.La vía poru
la cual los inmunocomplejosbloquearonel crecimientocelular no ha sido analizadaen esteUu• 89uUUuU

UuUUuU trabajo,pero en otros trabajos,algunosinvestigadoreshansugeridoun modelo en el cual, laU interacciónde complejosgpl2O-anti-gpl2Oconla moléculaCD4 generaríaunaseñala nivel deu
membranaque induciría un fenómenode apoptosis(64).u
Todos los resultadosanterioresindican que los inmunocomplejoscirculantesparecenUU tenerun papelpatogénicoen la infección.Estepapelsuponequeademásde la función básicaU de defensade nuestro organismo frente a microorganismosinfecciosos que realizan los
inmunocomplejos,la presenciade éstosen el sueropuedetenertambiénun efectoperjudicial.U
Normalmentelos inmunocomplejosse formanL’z vivo cuandolosantígenosseunena los
U anticuerposcomoconsecuenciadedistintasreaccionesinmunes.Estoscomplejossoneliminados
u de la circulación mediantecélulas quepresentanreceptoresde la fracción constantede lasU• inmunoglobulinas(RFc) en susuperficie,las cualesseencuentranampliamentedistribuidasen
U nuestroSistemalinfático.
Sin embargo,en muchasenfermedadesautoinmunes,infecciosaso neoplásicas,losu• inmunocomplejosse acumulany causangravesalteracionespatológicas,como inflamación,
U inmunosupresióno trombocitopenía.uU• 5.2.- MECANISMOS AUTOINMUNES IMPLICADOS EN LA DEPLECTON DEU LINFOCITOS T CD4+.Uuu Los mecanismos patogénicos por los cuales la infección por el virus de la
U InmunodeficienciaHumanaconducea la depleciónde linfocitos T CD4+ y al desarrollode unu
Síndromede Inmunodeficienciaadquirida, no estánsuficientementeesclarecidos.
U La idea de queotrosmecanismos,ademásdel propioefectocitopáticodelvirus, esténu
implicadosen la destruccióndel SistemaInmunede los pacientesinfectados,estápresenteenlas hipótesisquepostulanmuchosinvestigadorespara explicar la disminucióntan drásticade
• linfocitos T CD4+ quese produceen estossujetos.uU• Actualmenteexistenalgunasevidenciasquesugierenun papelde la autoinmunidadenU la patogénesisde la infecciónporel virus de la inmunodeficienciahumana.Sepiensaqueexisteu un procesoautoreactivoimplicado en la inducciónde un desequilibriodel SistemaInmunedeu• los pacientesinfectados,aunquetodavíano hayun acuerdoacercadel papelde estefenómeno
U en la patogénesisdel SIDA.UUuUu 90uuUuu

UUUuu• Ante la detecciónen el suerode pacientesinfectadosde autoanticuerposfrente a
U moléculaslinfocitarias (95-137), se analizósi una respuestamediadapor linfocitos T CD8+U
frentea linfocitosT CD4+ no infectadospodríaexistir o generarsedurantela infecciónpor elu virus de la Inmunodeficienciahumana.UUU La respuestamediada por linfocitos T citotóxicos constituye un componentemuyU importantede la respuestadenuestroorganismofrentea muchasdelas infeccionesvirales(139)U
y la disminuciónde una respuestacitotóxica específicafrente al HIV, mediadapor CTLs,asociadaa la progresiónde la infección(140),sugierequeestarespuestapodríatenerun papel
U importanteen la inhibición de la replicaciónviral.uuU En los ensayosde citotoxicidad realizadoscon linfocitos T CD8+, aisladosde sangre• periféricade pacientesinfectados,se estudióla capacidadde estascélulasde lisar linfocitos noU infectadosde sujetosnormales,estimuladoscon PI-JA. En esteanálisisseobservóqueexistíanu• linfocitoscitotóxicoscapacesde lisarlinfocitos T CD4+ en un 14% delos pacientesestudiados.
• La especificidadde esta respuestafue comprobadainhibiendo la lisis con anticuerposUU anti-CD4. Estosresultadosindicanqueposiblementeestarespuestaestétambiénimplicada enu la patogénesisde la infección de estospacientes.UuU Esta respuestacitotóxica frente a linfocitos T C04+ no infectadossugiereque en la• infección por HIV podría generarseun fenómenoautoinmune,que estaríaimplicado en la
• depleciónde linfocitos T CD4+.u
Una respuestaautoinmunefrentea linfocitos no infectadosmediadapor linfocitos T
U citotóxicos fue observadapor Zarling et al., en algunos pacientesinfectados(141). EstosU investigadoresobservaronqueestarespuestadirigida frentea algunamoléculade la superficieUU delinfocitos no aparecíaen monosinfectadosy propusieronqueestarespuestacitotóxicapodría• estarimplicada en la patogénesisde la infección.uuuUeuuuuUuU
91uUuUe

UuuuueuuUuUUuUuUuuUuUuUuuuuuuUUuuUuUuUuuuUUUU 6.- CONCLUSIONESuuUuUuuuuuuuuu

UUUuuUUUUuUU
1.- Los Inmunocomplejoscirculantesparecentenerun papelpatogénicoen la infecciónU por el virus dela inmunodeficienciahumana,teniendoen cuentaqueniveleselevadosen suero,uU superioresa 500 ¡sg/mí,y la presenciadeantígenop24 formandopartede los InmunocomplejosU se asocian con una progresión de la infección hacia un estado de SIDA y que losU inmunocomplejossoncapacesde unirsein vio-o a la moléculaCD4, de inhibir la proliferaciónU delinfocitos de sangreperiféricaestimuladosconFHA y de potenciarla infeccióndelinfocitose• de sujetossanosmantenidosen cultivo.UuUUU 2.- En algunospacientesinfectadosapareceunarespuestacitotóxicafrentea linfocitosu
T CD4+ de sujetossanosmediadapor linfocitos T CD84-. Estehallazgo seriaunaevidenciaU másdequeotrosprocesos,verosimilmentedenaturalezaautoinmune.podríanestarimplicados,U ademásdel efectocitopáticodel virus, en la depleciónde linfocitos T CD4+.UuuUuUUUUUuUuuUUUuueUuUU 92UUuUU

UUUuuUuUuUUUUUuUUUuuUUUuUUuUuUUUUUuUUuuUuUU
7.- BIBLIOGRAFIAuuueuUuuuuUuuUUu

UUUUUU 1. Gottlreb,M., Schroff,R., Schanker,H. yWeismas,J.PneumocystiscarinU pneumoniaandu mucosalcandidiadisin previouslyhealthyhomosexualmen.N. Engl. J. Med.,305: 1425-
U 1430. (1981).UU 2. Gallo, R. C., Salahuddin,5. Z., Popovic,M., Shearer,O. M., Kaplan,M., Haynes,B. F.U yPalker,T. J. Frequentdetectionand isolationof cytopathicretrovirus(HTLV-Ifl) fromU patientswith AIDS andat risk of AIDS. Science224: 500-503.(1983).U 3. Gonda,M. A.,Wong-Staal,F., GaUo, R. C., Clements,J. E., Narayan,O. y Gilden, R. V.U Sequencehomology and morphologic similarity of HTLV-1l1 and Visna virus, aU pathogeniclentivirus. Science,227: 173-177.(1985).
UU 4. Chiu, 1., Yaniv, A. y Dahlberg,J. E. Nucleotide sequenceevidencefor relationshipofU AIDS retrovirusto lentivirus. Nature,317: 366-8. (1985).
u 5. Clavel, F., Guetard,D. y Brun-Vezinet,F. Isolation of a new humanretrovirusfromWest African patientswith AJOS. Science,233: 343-6. (1986).
UU 6. Smith, T. F., Srinivasan,A., Schochetman,O. y Marcus,lv!. The ph¡logenetichistory ofU immunodeficiencyviruses.Nature,333: 573-5. (1988).uu 7. Chakrabarti,L., Guyader,M. y Alizon, M. Sequenceof símíanímmunodeficiencyvirusU from macaqueand its relationshipto otherhumanandsimian retrovirus.Nature,328:U 543-7. (1987).U 8. Robey,W., Safai, B., Oroszlan,5., Arthur, L., Gonda,M., Gallo, R. C. y Fischinger,P.U T. Characterizationof envelopeandcorestructuralgeneproductsof HTLV-III with seraU from AIDS patients.Science,228: 593-5. (1985).UU 9. Alían, J. 5., Coligan, J., Oroopman,J. y Essex,M. A new HTLV-II1/LAV encoded• antigendetectedby antibodiesfrom AJDS patients.Sejence,230: 810-15. (1985).UU 10. Fisher,A. O., Feinberg,M. 8. y Josheph,A. F. The transactivatorgeneof HTLV-III isU esentialfor virus replication.Nature,320: 367-371.(1986).
e 11. Nafees,A. y Venkatesan,5. Nef proteinof HIV-1 is a transcripcionalrepresorof HW-1LTR. Science,241: 1481-5. (1988).
UU 12. Sodroski,J., Goh, W. C. y Rosen,C. Replicativeandcytopathicpotencial of HTLV-U III/LAV with sorgenedeletiones.Science,231:1549-53.(1986).UU 13. Wong-Staal,F., Chanda,P. y Ohrayeb,3. HumanImmunodeficiencyvirus: the eighthu gene.Res.Hum. Retovirus,3: 33-9.(1987).U
14. Cohen,E. A., Terwilliger, E. F., Sodroski,J. O. y Haseltine,W. A. Identification of aU protein encodedby the vpu geneof HIV-l. Nature, 334: 532-4. (1988).uU 15. McDougal,J. 5., Klatzmann,D. R. y Maddon,P. J. CD4-gp120 interactions.Immunol.,
• 3: 552-8. (1991).UuUUu 93uUUuU

UuUUUU 16. Patterson,5. y Knight, 5. Susceptibiiityof humanperipheralblood dendritic cells toU infection by humanimmunodeficirncyvirus. 3. Gen. Virol., 68: 1177-1183.(1987).u
17. Gyorkey, F., Melnick, 3. L., Sivikovics, 3. C., et al. RetrovirusresemblingHTLV inmacrophagesof patientswith AIDS. Lanceti, 106. (1985).
u 18. Deen,K.C., McDougal,J. 5., e Inacker,R. A solubleform of CD4 protein inhibits AIDSU virus infection. Nature,331: 84-6. (1988).UU 19. Spickett, G. y Dalgleish, A. C. Cellular immunology of HIV infection. Clin. Exp.U Immunol., 71: 1-7. (1988).
UU 20. Clayton, L. K., Hussey,R. E., y Steinbrich,R. Substitutionof murine for humanCD4U residuesidentifie aminoacidscritical for 141V-gp120binding.Nature,335: 363-6. (1988).U 21. Goudsmith,3., Debouck,C., Meloen, R. H., Smit, L. Bakker, M., Asher, D. M., et al.U Humanimmunodeficiencyvirus type1 neutralizationepitopewithconservedarchitectureU elicits early type-specificantibodiesin experimentallyinfectedchimpanzees.ProcNatíU Acad Sci USA 85: 4478. (1988).U• 22. Ivey-Hoyle, M., Culp, 3. 5., Chaikin, M. A., Hellmig, E. D., Matthews, T. J., et al.U Envelopeglycoproteinsfrom biologicail diverseisolates of MIV havewidely different
affinities for CD4. Proc.Natí. Acad. Sci. USA, 88: 512-16.(1991).U 23. Qureshí,N. M., Coy, D. 14., Garry, R., Henderson,L. A. Characterizationof a putativeU cellularreceptorfor HIV- 1 transmembraneglycoproteinusingsyntheticpeptides.AIDSU 4: 553. (1993).uU 24. Callebaut,C., Krust, B., Jacotot,E. y Hovanessian,A. T ceil activation antigen,CD26,U as a cofartorfor entryof MIV in CD4+ Celis. Science,262: 2045-2050.(1993).U• 25. Stephens,P. E., Clements,O., Yarranton,O. T., y Moore, 3. A chink in HIV’s armour?.U Nature,343: 219. (1990).
U 26. Barnett, 5. W., Quiroga, M., Werner,A. Dina, D. y Lev>’, 3. A. DistinguishingfeaturesU of an infectious moleculardone of the highly divergent and noncytopathichumanU virus type2 IJCL strein. 3. Virol. 67: 1006. (1990).immunodeficiency
U 27. Morimoto, C. 3., Torimoto, Y., Levinson, O., Rudd,C. E., Schrieber,M., Dang,N. H.,U et al. 1F7, a novel ceil surfacemolecule, involved in helper function of C04 cells. 3.U Immunol., 143: 3430-9.(1989).
U• 28. Dalgleish,A., Beverly,P.,Claphan,P., et al.The CD4 antigenis an essentialcomponent
of the receptorfor the AIDS retrovirus.Nature,312: 763-766.(1984).
U 29. Stein,8. 5., Oowda,5. D., Lifson, 3., et al. pH-IndependentMIV entry into CD4-positiveU T cells via virus envelopefusion to the plasmamembrane.Ceil, 49: 659-668.(1987).U• 30. Bedinger. P., Moriarty, A., von Borstle, R., et al. Internalization of the humanU immunedeficiencyvirus doesnot require the cytoplasmicdomainof CD4. Nature,334:• 162-5. (1988).UUuUUuuU

eUUUUU 31. Kowalski, M., Potz, J., Basiripour, L., et al. Funtional regions of the envelopeU glycoproteinof humanimmunodeficiencyvirus type 1. Science,237: 1351-1355.(1987).U 32. Fisher, A. O., Ratner, L., Mitsuya, H., et al. Infectiousmutants of HTLV-III with
u changesin the 3’region and markedlyreducedcytopathiceffects.Science,223: 655-9.U (1986).U• 33. Luciw, P., Cheng-Mayer, C. y Lev>’, J. A. Mutational analysis of the humanU immunodeficiencyvirus: the orf-B region down-regulatesvirus replication.Proc.Acad.U Sci. USA, 84: 1434-8.(1987).UU 34. Gendelman,H.E., Phrlps, W., Feigenbaum,L., et al. Transactivationof the humanU immunodeficiencyvirus long terminal repeatsequencesby DNA virus.Nature,325: 67-U 70. (1986).U 35. Jones,K.A., Kadonga,J.T., Luciw, P.A. y Tijain, R. Activation of the AIDS retrovirusU promoterby ceDulartranscr¡ptionfactor, SP!.Science,231: 755-9. (1986).UU 36. Nabel, O. y Baltimore, O. An inducible transcription factor activatesexpressionofU immunodeficiencyvirus in T celís.Nature,326: 711-13.(1987).uU 37. Cooper,O. A., GoId, J.,Maclean,P., Donovan,B., Finlayson,R., Barnes,T., et al.Acute
AIDS retrovirusinfection. Definition of clinical iilnessassociatedwith seroconversion.U Lancet, i: 537-540. (1985).
u 38. Koeng, 5., Gendelman,14. E., Orenstein,3. M., DalCanto,M. C., Pezeshkpour,O. H.,U et al. Detectionof AJOSvirus in macrophagesin brain tissuefrom AIDS patientswithU encephalopathy.Science,223: 1089-1093.(1986).UU 39. Price, R. W., Brew, B., Sidtis, 3., Rosenblum,M., Scheck,A., y Clear>’, P. The brain inU AIOS: CentralNervousSystemHTV-infection andAIDS dementiacomplex.Science,
239: 586-592.(1988).U 40. Masur,Il., Michelis, M. A., Greene,3. B., Onarato,1., Holzman, R. 8., Wormser,O., et
al. An outbreak of community-acquiredPneunzocystiscarinil Pneumonia: InitialU manifestationof cellular immunedysfunction.N. Engí.3. Med., 305: 1431-1438.(1981).UU 41. Ziegler,3. L., Beckstead,3. A., Volberding,P. A., Abrams,0. 1., Levine, A. M., Lukes,U R. J.,et al.Non-Hodgkin’slymphomain 90 homosexualmen.Relationshipto generalizedU lymphadenopathyand theacquiredimmunodeficiencysyndrome.N. Eng. 3. Med.,311:U 565-570.(1984).u
42. Aliegra, C. J.,Chabner,8. A., Tuazon,C. U., Ogata-Aratiki,O., Baird, 8., Drake,3. C.,Simmons,3. T., et al. Trimetrexatefor the treatmentof Pnewnocystiscarinhlpheumonia
U in patientswith the acquiredimmunodeficiencysyndrome.N. Engl. 1. Med., 317: 987-U 984. (1987).UU 43. CollaborativeDHPO treatmentstudy group: treatmentof seriouscytomegalovirusU infectionwith 9-(1,3-dihidroxi-2-propoximetil)guaninain patientswith AIOS. N. Engl.• J. Med.,314: 801-805.(1986).UUUUeUUeU

UeUUUU 44. Fauci, A. 5., Macher, A. M., Longo, D. L., Lane, 14., Rook, A. H., Masur, 14., yU Gelmann, E. P. Acquired immunodeficiency syndrome: Epidemiologic, clinical,• immunologic,and therapeuticconsiderations.Ann. Inter. Med., 100: 92-106.(1984).
U 45. Kovacs, 3. A., y Masus, 14. Oportunistic infections. In: ATOS Etiolo~’, Diagnosis,u Treatment,andPrevention,2nded.,editedby DeVita, V., Helíman,5., y Rosenberg,5.,U Pp. 199-266.Lippincott, Philadelphia.(1988).UU 46. Stahl, R. E., Friedman-Kien. A., Durbin, R., Marmor, M., y Zolla-Pazner, 5.U Immunologicabnormalitiesin homosexualmen: Relationshipto ICaposi’ssarcoma.Am.U 3. Med., 73: 171-178.(1982).U
47. Ensoli, B., Biberfeld, P.,Nakamur,5., Salahuddin,5. Z., Wong-Staal,F. y Gallo, R. C.Possiblerole of growthfactorsand cytokinesin pathogenesisof Kaposi’s sarcoma.In:Abstractsof the IV InternationalConferenceon AIDS, Book 2, June12-16,Stockholm,
u Sweeden,p. 137. (1988).UU 48. Lane,14. C., Masur,H., Edgar,L. C.,Whalen,O., Rook,A. y Fauci,A. 5. AbnormalitiesU of B-cell activation and immunoregulation in patients with the acquiredU mmunodeficiencysyndrome.N. Engí. 3. Med., 309: 453-458.(1983).UU 49. Donahue,R. E., Johnson,M. M., Zon, L. 1., Clark, 5. C.,y Groopman,3. E. Suppression
of invitro hematopojesisfollowing humanimmunodeficiencyvirus infection.Nature,326:e 200-203.(1987).U 50. Stricker, R. B., Abrams,D. 1., Corash,L., y Shuman,M. A. TargetplateletantigeninU homosexualmen with immune thrombocytopenia.N. Engl. 3. Med., 313: 1375-1380.U (1985).UU 51. Bernstein,L. 3., Krieger, B. Z., Novick, B., Sicklick, M. 3., y Rubinstein,A. Bacterial• infection in the acquiredimmunodeficiencysyndromein children. Pediatr.Infect,Dis.,U 4: 472-475.(1985).U 52. Centersfor DiseaseControland PreventionU. 5. 1993 revisedclassificationsystemforU HIV infection and expandedsurveillancecasedefinition for AIDS amongadolescentsU and adults.MMWR. Morb. Mortal. Wkly. Rep., 41 (RR-17): 1-19. (1992).UU 53. Acelle-Park,R. A., Stroobant,A., Smith, E., Haikala, O., Koch, M. A., et al. EuropeanU AIDS definition. Lancet,339:671. (1992).Uu 54. Fauci, A. 5. íhe human ¡mmunodeficiencyvirus: Infectivity and mechanismsofU pathogenesis.Science,238: 617-622.(1988).u
55. Weller, 5. K., Soy,A. E., y Temin, 14. M. Correlationbetweencelí killing and massivesecond-roundsuperinfectionby membersof somesubgroupsof avian leukosisvirus. 3.Virol., 33: 494-506.(1980).
U 56. Lifson, 3. D., Reyes,O. R., McOrath, M. 5., Stein, B. 5., y Engleman,E. O. AIDS• retrovirusinducedcytophatology:Giantcell formationandinvolvementof CD4 antigen.• Science,232: 1123-1127. (1986).UUU
96UuUuu

UuUUUU 57. Walker, E. D., Chakrabarti,S., Moss, B., et al. HIV-specific cytotoxicT lymphocytesinU seropositiveindividuals.Nature,328: 345-348. (1987).U 58. Gatner,S., Markovits, P., Kaplan, M. H.. Gallo, R. C. y Popovic, M. The role ofU mononuclearphagocytesin HTLV-II/LAV infection. Science,233: 215-219. (1986).u
59. Lanzavecchia,A., Roosnek,E., Gregor>’, T., Berman,P., y Abrignani, 5. T cell canU presentantigenssuch as 141V gp120 targettedto their own surfacemolecules.Nature,U 334: 530-532.(1988)uU 60. Kornfeld, 14., Cruikshank,W. W., Pyle, S. W., Berman,J. 5. y Center.D. Lymphocyte• activationby HIV-1 envelopeglycoprotein.Nature,335: 445-8. (1988).U
61. Cohen,J. J.,Duke,R. C., Fadok,V., y Sellius, K. 5. Apoptosisandprogramedcelí deathU in immunity. Ann. Rey. Immunol., 10:267-93.(1992).
• 62. Meyaard,L., Otto, 5. A., Jonker, R. R., Mijnster, M. 1., Keet, R. P:, y Miedema, F.U Programmeddeathof T cells in HIV-1 infection. Science,257: 217. (1992).U• 63. Groux, 14., Torpier, O., Monte, D., Mouton, Y.. et al. Activation-induceddeath by• apoptosisin CD4+ T celis from humanimmunodeficiencyvirus-infectedasymptomattc• individuals.J. Exp. Med., 175: 331-40. (1992).
U 64. Banda,N. K., Bernier,1., Kurahara,D. K., Kurrle, R., Haigwood,N., Sekaly, R. P., etU al. Crosslinking CD4 by human immunodeficiencyvirus gpl2O primes T cells forU activation-inducedapoptosis.J. Exp. Med., 176: 1099. (1992).uU 65. Dalgleish,A. O., Wilson, 5., Gompels,M., Ludlam,C., Gazzard,E., Coates,A. M., et al.U T-cell receptorvariable gene productsand early HIV-1 infection. Lancet, 339: 824.U (1992).u• 66. Schnittman,5. M., Lane,14. C., Greenhause,3., Justement,J.5., Baseler,M. y Fauci,A.
5. Preferentialinfection of CD4+ memor>’ T celís by humanimmunodeficiencyvirus• type 1: evidencefor a role in the selectiveT-cell funetionaldefectsobservedin infected
individuals.Proc.Natí. Acad. Sci. USA., 87: 6058.(1990).
U 67. Bukrinsky, M. 1., Stanwick, T. L., Dempsey, M. P., Stevenson,M. QuiescentTU lymphocytesasaninduciblevirus reservoirin I-IIV-1 infection.Science,254: 423. (1992).UU 68. Germain,R. N. Antigen processingand CD4+ T celí depletionin AIDS. Cell 54: 441-U 444. (1988)u• 69. Yarchoan,R., Redfiel, R. R.,yBroder, 5. Mechanimsof B ceil activationin patientswithU acquiredimmunodeficiencysyndromeandrelateddisorders.Contributionof antibod>’-
producing B celís, of Epstein-Barrvirus-infected B cells, and of immunoglobulinproduction inducedby human T cdl lymphotropic virus type 11I/lymphadenopathy-
U associatedvirus. 3. Clin. Invest.,78: 439-447. (1986).uU 70. Stricker, R. B., Mchugh, T. M., Moody. O. 3., Morrow, W. 3., Stites,O. P., et al. AnU AIOS-relatedcytotoxicautoantibodyreactswith a specificantigenon stimulatedCD4+
U cells. Nature,327: 710-713.(1987).uuu 97uuuUu

uUUUuU 71. Heriot, K., Hallquist, A. E., y Tomar, R. 14. Paraproteinemiain patientswith acquired
U immunodeficiencysindrome(AIDS) or lymphadenopathysyndrome(LAS). Clin. Chem.,U 31: 1224-1226. (1985).U 72. Pahwa, R., Good, R. A., y Pahwa, 5. Prematurity, hypogammaglobulinemia,and
u neurophathologywith humanimmunodeficiencyvirus (141V) infection.Proc.Natí.Acad.U Sci. USA, 84: 3826-3830.(1987).UU 73. Schnittman,5. M., Lane,14. C., Higgins,5. E., Folks, T., y Faucí,A. 5. Diret polyclonalU activationof humanB lymphocytesby the acquiredimmunodeficiencysyndromevirus.U Science,233: 1084-1086.(1986).U• 74. Gurney,M. E., Heinrich, 5. P., Lee, M. R., y Yin, ¡-1. MolecularcíoningandexpressionU of neuroleukin,a neurotropicfactor for spinalandsensoryneurons.Science,234: 566-U 574. (1986).U 75. Eírx, D. L., Redfield, R. R., y Tosato,O. Defective regulation of Epstein-BarrvirusU infection in patientswith the acquiredimmunodeficiencysyndrome(ATDS) or AIDS-U relateddisorders.N. Engí. 3. Med., 314: 874-879.(1986).UU 76. Montagnier, L., Gruest,3., Chamaret,5., Daugnet,C., Aider, C., Guetard,D., et al.U Adaptation of lymphadenopathy-associatedvirus (LAV) to replication in EBV-• transformedB lymphoblastoidcell lines. Science,225: 63-66. (1984).U 77. Groopman;3. E., Suilivan, 3. L., Mulder, C., Oinsburg,D., Orkin, 5. H., O’Hara, C.,U Falchuk,K., et al. Pathogenesisof B cell lymphoma in patientswith AIDS. Blood, 67:
u 612-615.(1986).UU 78. Ho, D. D., Rota,, T. R., y Hirsch, M. 5. Infection of monocyte/macrophagesby humanU 1 lymphotropicvirus type III. 3. Clin. Invest., 77: 1712-1715.(1986).UU 79. Folks, T. M., Kessler,5., Cottler-Fox, M., Justement,it, Orenstein,J., y Fauci, A. 5.U Infectionand replication of humanimmunodeficiencyvirus-1 (HIV-1) in purified CD4U negativeprecursorceUsfrom normalbonemarrow.In: Abstractsof the IV InternationalU Conferenceon AIDS, Book 1. June12-16, Stockholm,Sweeden,p 180. (1988).U 80. Nelson,J.A., Wiley, C. A., Reynolds-Kohler,C., Reese,C. E., Margaretten,W., y Lev>’,U 3. Human immunodeficiencyvirus detectedin bowel epithelium from patientswithU gastrointestinalsymptoms.Lancet, 1: 259-262. (1988).UU 81. Pomerantz,R. 1., de la Monte,5., Donegan,5. P., Rota,T. R., Vogt, M., Craven,D. E.,U et al. Humanimmunodeficiencyvirus (141V) infection of theuterinecerv¡x. Ann. ínter.U Med., 108: 321-327.(1988).u
82. Koenig, 5., Qendelman,14. E., Orenstein,3. M., Dal Canto, M. C., Pezeshkpour,O.H.Yungbluth, M., et al. Detectionof AIDS virus in macrophagesin brain tissuefrom
U AIDS patientswith encephalopathy.Science,223: 1089-1093.(1986).uuUuUu• 98UUuuU

UuUuu• 83. Moscicki, R. A., Amento, E. P., Krane, 3. T., y Colvin, R. B. Modulation of surfaceU antigensof a humanmonocytecelí line, U937, during incubationwith T lymphocyte-U conditioned medium: detection of T4 antigen and its presenceon normal blood• monocytes.3. Immunol., 131:743-748.(1983).
u84. Robinson,W. E., Montefiori,D.C., y Mitchel, W. M. Antibody-dependentenhancement
of humanimmunodeficiencyvirus type 1 infection. Lancet, 1: 790-794.(1988).
u 85. Prince,14. E., Moody, D. 3., Shubin,B. J.. y Fahey,J. L. Defectivemonocytefunction inU acquiredimmunodeficiencysyndrome(AIDS): Evidencefrom a monocyte-dependentT-U ceD proliferativesystem.3. Clin. lmmunol., 5: 21-25. (1985).uU 86. Rook,A. 14., Masur,U., Lane,14. C., Frederick,W., Kasahara,T., Macher,A. M., et al.• Interleukin-2enhancesthedepressednaturalkilier andcytomegalovirus-specificcytotoxic
activitiesof lymphocytesfrom patientswith the acquiredimmunedeficiencysyndrome.3. Clin. Invest., 72: 398- 403. (1983).
u 87. McDougal, J. 8., Kennedy,M. 8., Nicholson, 3. et al. Antibody responseto humanu immunodeficiencyvirus in homosexualmen: Relationof antibodyspecificity, titre, andU isotypeto clinical status,severityof immunodeficiency,anddeseaseprogression.J. Clin.U Invest., 80: 316-24.(1987).Uu 88. Lange, J. M., Paul, D. A., Huisman, M. O., et al. PersistentMIV antigenaemiaand• decline of 141V core antibodiesassociatedwith transition to AIDS. Br. Mcd. J. 293:• 1459-62. (1986).
u 89. Sanz,F. 3., García, R. y Ortiz, F. Evaluacióninmunológicade la infecciónpor el virusu de la InmunodeficienciaHumana.Inmunología,Vol. 10, 3: 24-29. (1991).u• 90. Simmonds,P., Lainson, F., Cuthbert, R., et al. MIV antigen and antibod>’ detection:• Variableresponsesto infection in theEdinburghhaemophiliaccohort.Br. Mcd. 3. 296:U 593-8. (1988).u• 91. Lange,3. M., de Wolf, F., Krone,W.3., et al. Declineof antibod>’ reactivity to outerviralU core protein pl7 is an earlier serological marker in deseaseprogressionin human• immunodeficiencyvirus infection thananti-p24decline.AIDS, 1:155-9.(1987).
u 92. Ranki, A., VaDe, 5., Krohn, M, et al. Long latencyprecedessero-conversionin sexualí>’u transmitted141V infection. Lancet,u: 589-92.(1987).u• 93. Weiss,R. A., Clapham,P. R., et al. Neutralizationof humanT-lymphotropicvirus type• III by seraof AIDS andAIDS-risk patients.Nature,316: 69-72. (1985).u• 94. Palker,T. J., Matthews,T. 3., Clark, M. E., Cianciolo, 0. 3., Randalí,R. R., Langlois,A.• 3., et al. A conservedregion at the COOH terminusof humanimmunodeficiencyvirus• gpl2O envelopeglycoproteincontainsan immunodominantepitope.Proc.Natí.Acad.Sci.
U USA, 84: 2479-2483.(1987).
u 95. Oolding, 14., Rober, F. A., Gates,F. T., Linder, W., Beining, P. R., Hoffman, T., yGolding, E. Identification of homologousregionsin humanimmunodeficiencyvirus 1
U gp4l andhumanMMC class11131 domain.3. Exp. Mcd., 167: 914-923.(1988).uuu 99uuUUu

UuUuuU 96. Wahren,B., Morfeldt-Manson,L., Biberfeld, G.,Moberg,L., Sónnerborg,A., Ljungman,U P., et al. Characteristicsof the specific celí-mediatedimmune responsein human• immunodeficiencyvirus infection.J. Virol., 61: 20 17-2023.(1987).uu 97. Pantaleo,O., De María, A., Koenig, 5., Butini, L., Moss, B., Baseler,M., et al. CD8+lymphocytesof patientswith AIDS maintainnormalbroadcytolytic functiondespitetheu lossof humanimmunodeficiencyvirus-specificcytotoxicity. Proc.Natí.Acad.Sci. USA,U 87: 4818-4822.(1990).u• 98. Koenig, 5., Earl, P., Powell, D., Pantaleo,O., Merli, 5., Moss, B., y Fauci,A. 5. Oroup-U specific, MHC class 1 restricted cytotoxic responsesto HIV-l envelopeproteins byu clonedperipheralblood T cellsfrom an HIV-l infectedindividual. Proc.Natí. Acad.ScL• USA, 85: 8638. (1988).
u99. Johnson,R. P., Trocha,A., Yang,L., Mazzara,O., Panicali,D., Buchanan,T, y Walker,
U B. HIV-1 gag-specificcytotoxic T lymphocytes recognizemultiple highly conservedepitopes:fine specificityof the gag-spscificresponsesby using unstimulatedperipheralblood mononuclearcelis andcloned effectorcelís.1. Immunol., 147: 1512. (1991).u
• 100. Walker, B. D., Flexner,C., Paradis,T., Fuller, T. C., Hirsch, M., Schooley,R., y Moss,• B. HIV-1 reversetranscriptaseis a target for cytotoxic T lymphocytes in infectedu individuals.Science,240: 64. (1988).u• 101. Brinchmann,J., Gaudernack,O., y Vartdal, F. CD8+ celís inhibit MIV replication in• naturally infectedCD4+ Tcells.Evidencefor a solubleinhibitor.3. Immunol., 144: 2961-
2966. (1990).
U 102. Lev>’, 3. A. Mysteriesof 141V: Challengesfor therapyandprevention.Nature,333: 519-
u 522. (1988).uU 103. Lange,J. M., Paul,D. A., De Wolf, F., Coutinho, R. A., Ooudsmit,3., et al. Viral geneu expression, antibod>’ production and immune complex formation in human• immunodeficiencyvirus infection. AIDS, 1:15-20.(1987).u• 104. Allain, Y. P., Laurian,Y., Paul, D., Verroust,F.. Leuther,M., Gazengel,M., et al. Long-U term evaluation of MIV antigen and antibodies to p24 and gp4l in patientswith
hemophilia.Potencialclinical importance.N. Engl. 3. Med., 317: 1114-1121.(1987).
u 105. Franchini,O., Robert-Guroff,M., Aldovini, A., Kan, N. C., y Wong-Staal,F. SpectrumU of naturalantibodies againstfive HTLV-1II antigensin infected individuals:• Correlationof antibodyprevalencewith clinical status.Blood, 69: 437-441.(1987).uu 106. Chent-Mayer,C., Seto,D., Tateno,M., y Lev>’, 3. Biologic featuresof MIV thatcorrelateU with virulencein the host. Science,240: 80-82. (1988).u
107. Amadori,A., Zamarchi,R., Ciminale, V., et al. HIV-1-specific B celí activation.A majorconstituentof spontaneousB ceil activation during HIV-1 infection. Y. Immunol. 143:
U 2146-2152.(1989).UUUu• 100
uuuuu

UUUuuU 108. Pahwa,S.. Quilop. M. T., Lange, M., Pahwa, R. N., y Grieco, M. U. Defective B-• lymphocytefunction in homosexualmen in relation to the acquiredimmunodeficiency• syndrome.Ann. Intern. Med., 101: 757-763.(1984).
uU 109. Morris, L., Distenfeld,A., Amorosi, E., y Karpatkin,5. Autoimmunethrombocytopenia
purpurain homosexualmen.Ann. Int. Med., 96: 714. (¡982).U 110. Walsh,C. M., Nardi, M. A., y Karpatkin, 5. Qn the mechanismof thrombocytopenicU purpurain sexuallyactivehomosexualmen.N. Engí. J. Med., 311: 635-639.(1984).uu 111. Kopelman. R. G. y Zolla-Pazner,5. Associationof human immunodeficiencyvirus• infection andautoimmunephenomena.Am. 1. Med., 84: 82-8. (1988).
U112. McDougal, 3. 5., Hubbard,M., Nicholson,J., Jones,8., Holman, R. C., Roberts,3., et al.
Immunecomplexesin the acquired immunodeficiencysyndrome(AIDS): reslationshipto deseasemanifestation,risk group,and immunologicdefect.3. Clin. Immunol, 5:130-8.
u (1985).U• 113. Sonnabend,3., Witkin, S.S., y Purtilo, D. T. Acquired immunodeficiencysyndrome,U opportunisticinfections,andmalignanciesin malehomosexuals.A hipothesisof etiologicU factors in pathogenesis.JAMA, 249 (17):2370-4.(1983).U• 114. Oldstone, M. E. A. Molecular mimicry and autoimmunedisease.Celí, 50: 819-820.• (1987).
U 115. Andrieu, 1. M., Even, P., y Venet, A. AIOS and relatedsymdromesas a viral-inducedu autoimmunediseaseof tbe immune system: an ant¡-MHC 11 disorder.TherapeuticU implications. AIDS Res.,2:163. (1986).uU 116. Golding, 14., Shearer, G. M., 1-FIlman, K., et al. Common epitope in humanu immunodeficiencyvirus (141V) 1-0P41 and HLA class II elicits immunosuppressive• autoantibodiescapable of contributing to immune dysfuntion in 141V 1- infected
U individuals.3. Clin. Tnvest.83: 1430-1435.(1989).uU 117. Auffray, C. y Novotny, J. Speculationson sequencehomologiesbetweenthe fibronectin
cell-attachmentsite, major histocompatibility antigens,and a putative ATDS viruspolypeptide.Hum. Immunol., 15: 381-390.(1986).
• 118. Procaccia,5., Lazzarin,A., Colucci,A., Gasparini,A., Forcellini, P.y Lanzanova,D. IgM,U IgO and IgA rheumatoidfactorsand circulating immune complexesin patientswithu AIDS andAIDS-relatedcomplexwith serologicalahnormalities.Clin. Exp.Immunol.,67:U 236-244.(1987).U• 119. Lange, 3. M., Paul, D. A., De Wolf. F., Coutinho, R. A. y Goudsmit, 3. Viral gene• expressíon. antibod>’ production and immunecomplex formation in human
U immunodeficiencyvirus infection. A1DS, 1:15-20.(1987).
u 120. Weber, 3., Cheingsong-Popov,R. y Panagiotide,C. MIV core proteins in AIDS• pathogenesis.In UCLA Symposiumon humanRetrovirus,New York, (1989).UuUUU 101uUUuu

UUUuUU 121. Tausk, F. A., McCutchan,J. A., Spechko,P., Scjreiber,R. D., y Gigli, 1. AlteredU erytrocyteC3b receptorexpression,immunecomplexes,andcomplementactivation inu homosexualmen in varying risk groups for acquiredimmunedeficiency syndrome.J.• Clin. lnvest.,78: 977-982.(1986).
u122. Bolognesi,D. AIDS. Do antibodiesenhancethe infection of cellsby MIV?. Nature,340:
U 431-2. (1989).
u 123. Amadori, A y Chieco-Bianch,L. B-cell activation and 141V infection: deeds and• misdeeds.Immunol. Toda>’, Vol.11, 374. (1990).u• 124. Kestens.L., Hoofd, O., Gigase, P. L., Deleys. R., et al. HIV antigen detection iii
U circulating immunecomplexes.J. Virol. Methods,31: 67-76. (1991).u
125. Silvestris,F., Azzolini, C. y Dammascco,F. Differential isotypeexpressionandbindingpropertiesof T celí-reactiveantibodies in Muman ImmunodeficiencyVirus (141V)Infection. J. Clin. Immunol., 2:107-115.(1992).
UU 126. Mullis, K. B., Faloona, F. A. Specific synthesisof DNA in vitro via a polymerase-U catalyzedchain reaction.MethodsEnzymol., 155: 335-350.(1987).u• 127. Kwok, S., KeUogg, D. E., McKinney, N., Spasic,D.. Goda, L., Levenson,C., y Sninsky,U J. J. Effects of primer-templatemismatcheson the polymerasechain reaction:human• immunodeficiencyvirus type 1 model studies.Nucí. Acids. Res., 18: 999-1005.(1990).U 128. Bancrof, 14. Introduction to Biostatistics.New York.
u Eds. Hoeber-Harper.(1975).uu 129. Amadori, A., de Silvestro,O., Zamarchi, R., Veronese,M. L., Mazza, M. R., Schiavo,U O., et al. CD4 epitopemasking by gpl2O/anti-gpl2Oantíbod>’ complexes:A potencial• mechanismfor CD4+ ceIl funtion Down-Regulationin ATOS patients..1. Tmmunol.,148:U 2709-2716.(1992).U• 130. Kowalski, M., Blair, A., Basiripour. L., Lu, Y, Blohm, O, Haseltine,W y Sodroski,J.
Antibodies to CD4 in individuals infectedwith humanimmunodeficiencyvirus type 1.Proc. Natí. Acad. Sci. USA, 86: 3346. (1989).
U 131. Thiriat, C., Goudsmith,3., Schellekens,P., Barin, F., Zagur>’, O., et al. Antibodies toU sCD4 in HIV-l infected individuals.AJOS, 2: 345. (1988).uU 132. Mathiesen,T., Sonnerborg,A., y Wahren,B. Detectionof antibodiesagainstmyelinbasicU protein and increasedlevel of MIV IgO antibodiesand MIV antigenafter stabilizationu of ímmunecomplexesin seraandCFS of MIV infectedpatients.Viral Immunol., 2: 1.• (1989).
U133. Ziegler,J., y Stites,D. O. Hypothes¡s:AIOS is an autoimmunedeseasedirectedat the
immune system and triggered by a Iymphotropic retrovirus. Clin. Immunol.uU Immunopathol.,41: 305. (1986).UuuUU 102uUuUU

UUuUu• 134. Brenner, B. O., Oryllis, C., Oornitsky, M., y Wainberg, M. A. Changesin natural• immunity during the courseof HIV-l infection. Clin. Exp. Immunol., 93(2): 142-8.• (1993).
U 135. Fauci,A. 5. Immunopathogenicmechanismsin humanimmunodeficiencyvirus (MIV)
u infection. Ann. ínter.Med., 114: 678-93.(1991).uU 136. Bender,B. 5., Quinn, T. C., y Spivak,3. L. Homosexualmenwith thombocytopeniahaveU impaired reticuloendothelialsystem Fc receptor-specificclearence.Blood, 70: 392-5.• (1987).u• 137. Miller, C., Kukel, 5., y Bayer, R. Relationshipof antibodiesagainstC04+ T ceUsin
HIV-infected patients to markersof activation and progression:autoantibodiesare• closely associatedwith C04 cell depletion.Immunology,79: 248-254.(1993).
U 138. Morrow, W. 3., Wharton,M., Stricker, R. B. y Lay>’, 3. A. Circulatingimmunecomplexes
u in patientswith acquired-immunedeficiencysyndromecontain the AIDS-associated• retrovirus.Clin. Immunol. Immunopathol.,40:515-524.(1986).uU 139. Tsubota, H., Lord, C. 1., Watkins, O., Morimoto, C. and Letvin. A cytotoxic Tu lymphocyteinhibits AIOS virus replication in peripheralblood lymphocytes.3. Exp.• Med., 169: 1421. (1989).u
140. Hoffenbach,A., Langlade-Demoyen,P., Dadaglio, O., Vijnier, E., Michel, F., Mayaud,U C., Autran, B., y Plata, F. Unusualí>’ high frequenciesof HIV-specific cytotoxic T
lymphocytesin humans..1. Immunol., 142: 452. (1989).• 141. Zarling, J. F., Ledbetter,3. A., Sias, J., Fultz, P., Eichberg, J., Gjeset,O., y Moran, P.U HIV-infectedhumans,but not chimpanzees,havecirculatingcytotoxicT lymphocytesthatU lyseuninfectedCD4+ celís. J. Immunol., 144: 2992-98. (1990).UUUuUuuuuuUuUUuuUUuuU
103UUuUU