transducción de señales - estudiosecologistas.org ecuador/1b_ref... · • las hormonas...
TRANSCRIPT

TRANSDUCCIÓN DE SEÑALES
• Las células responden a señales o estímulos de su medio ambiente,como pueden ser la luz, nutrientes, o compuestos químicos(hormonas) que indican una situación fisiológica o patológica delorganismo.
• Estos estímulos se perciben y se procesan mediante un procesoconocido como transducción de señales.
• Para ello existen en las células circuitos moleculares formados porreceptores, enzimas, canales y proteínas reguladoras que detectan,amplifican e integran las diversas señales externas.
Rosario A. Muñoz-Clares
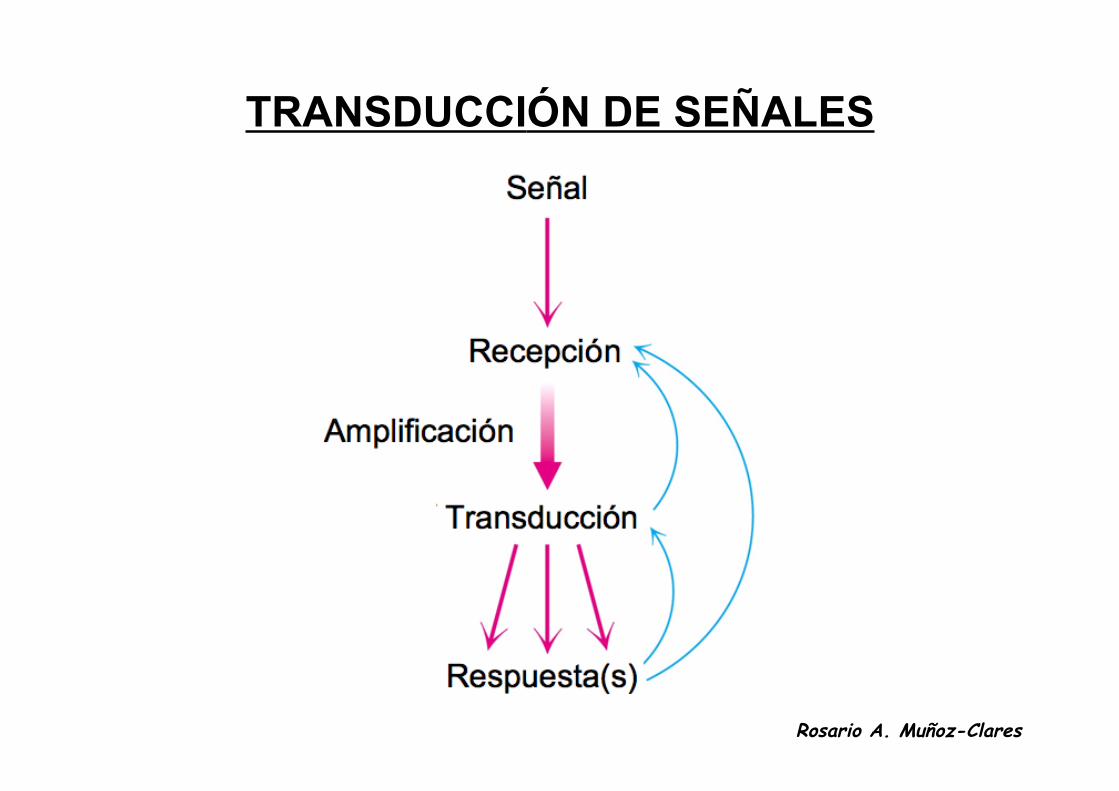
TRANSDUCCIÓN DE SEÑALES
Rosario A. Muñoz-Clares

COMUNICACIÓN INTRACELULAR
• Los seres vivos coordinan sus procesos por medio deseñales químicas complejas.
• Las hormonas son los mensajeros químicos que median lasseñales intra e intercelulares.
• En animales superiores también ejercen esta función losimpulsos electroquímicos transmitidos por el sistemanervioso.
Rosario A. Muñoz-Clares

FUNCIONES DE LAS HORMONAS
• Mantener la homeostasis.– Por ejemplo la insulina y el glucagón.
• Responder a una amplia variedad de estímulos externos.– Por ejemplo la adrenalina y noradrenalina.
• Regular programas cíclicos y de diferenciación y desarrollo.– Por ejemplo las hormonas sexuales.
Rosario A. Muñoz-Clares

TIPOS DE HORMONAS
• ENDOCRINAS– Actúan sobre células distantes del sitio en donde se liberan.
Por ejemplo la insulina y la adrenalina que se sintetizan entejidos periféricos (glándulas endocrinas).
• PARACRINAS– Actúan sobre células próximas al sitio en donde se liberan.
Por ejemplo la interleucina-1 (un factor de crecimientoproteico) liberada por los macrófagos para estimular allinfocito T al que se unen a proliferar y diferenciarse.
• AUTOCRINAS– Actúan sobre las mismas células que las liberan. Por ejemplo
otro factor de crecimiento proteico, la interleucina-2,producida por el linfocito T en respuesta a la interleucina-1.
Rosario A. Muñoz-Clares

HORMONAS ENDOCRINAS
• Se sintetizan en tejidos especializados (glándulas endócrinas).
• Se secretan a la sangre que las transportan a sus lugares deacción.
• Sólo las células que poseen los receptores específicos parauna hormona (células diana) responden a su presencia, auncuando todas las células del cuerpo estén expuestas a lahormona.
Rosario A. Muñoz-Clares

TIPOS DE HORMONAS
• Químicamente la mayoría de las hormonas son:
– POLIPÉPTIDOS
– DERIVADOS DE AMINOÁCIDOS
– ESTEROIDES
• Una excepción importante es el óxido nítrico (NO).
Rosario A. Muñoz-Clares

RUTAS DE TRANSDUCCIÓN DE SEÑALES
• Las hormonas esteroides y tiroideas pueden penetrar a la céluladada su naturaleza química (son moléculas apolares) y se unen areceptores que son transportados al núcleo.
• Pero la mayor parte de las hormonas se unen a receptores de lasuperficie celular y desencadenan cascadas de reaccionesenzimáticas.
• Las tres rutas principales por las que las señales extracelulares setransducen (convierten) en señales intracelulares son:– Las mediadas por los receptores de siete hélices.– Las mediadas por los receptores de tirosín cinasa.– Las mediadas por receptores de acetil colina.
Rosario A. Muñoz-Clares

UNIÓN DEL LIGANDO A LOS RECEPTORES• El primer paso en la transducción de señales es la unión de la
hormona a su receptor.• Los receptores unen a sus ligandos con alta afinidad y
especificidad.• Los receptores, como otras proteínas, unen sus ligandos
correspondientes de acuerdo a la ley de acción de masas.
R + L R-L
KL = [R][L] [R-L]
[R-L] [L]
KL + [L][R]T
=
Rosario A. Muñoz-Clares
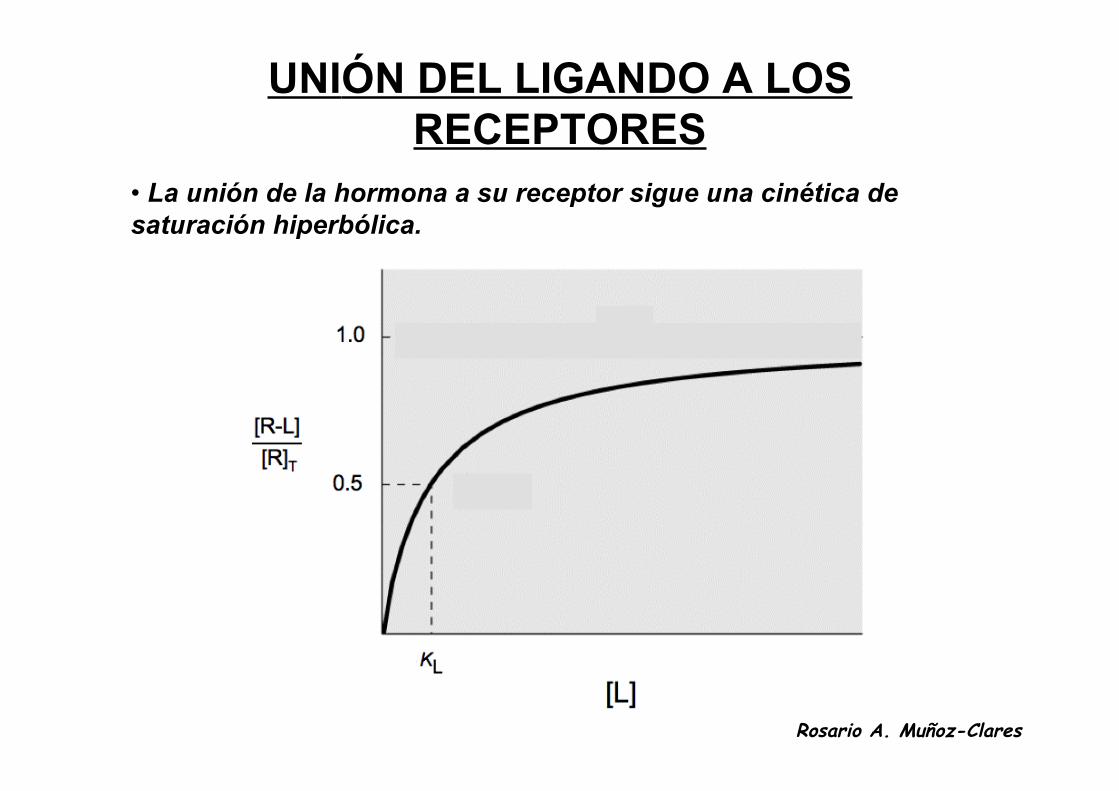
UNIÓN DEL LIGANDO A LOSRECEPTORES
Rosario A. Muñoz-Clares
• La unión de la hormona a su receptor sigue una cinética desaturación hiperbólica.

TIPOS DE RECEPTORES
• Existen cuatro tipos de receptores:
– Receptores esteroideos.
– Receptores de siete hélices (adrenorreceptores).
– Receptores tirosín cinasa (receptores de factores decrecimiento ).
– Receptor de acetil colina (participa en la transmisión delimpulso nervioso).
Rosario A. Muñoz-Clares

RECEPTORES ESTEROIDEOS
• Son proteínas citoplasmáticas y nucleares que cuando unen a lahormona correspondiente se activan como factores detranscripción para proteínas específicas.
• El efecto de esas hormonas se manifiesta en horas más que ensegundos o minutos, porque se deben sintetizar nuevos mRNAsy nuevas proteínas.
Rosario A. Muñoz-Clares

RECEPTORES ESTEROIDEOS
Rosario A. Muñoz-Clares
• Un cambio conformacional que se da cuando se une la hormonapermite que el complejo hormona-receptor se una al DNA.
Hélice 12

RECEPTORES DE SIETE HÉLICES
Rosario A. Muñoz-Clares

RECEPTORES DE SIETE HÉLICES• Son proteínas integrales de membrana con siete hélices
transmembranales y con dominios extracelular e intracelular.
• Están acoplados a las proteínas G heterotriméricas en sudominio intracelular.
• Poseen un sitio de unión para sus ligandos situado hacia elcentro de la bicapa.
• La unión del ligando produce un cambio conformacional en eldominio intracelular que induce a la proteína G a intercambiarGTP por el GDP que tenía unido, lo que desencadena la síntesiso liberación de segundos mensajeros (tales como el cAMP, eldiacil glicerol, el IP3 o el Ca2+) que median la respuesta a la señalhormonal dentro de la célula.
Rosario A. Muñoz-Clares
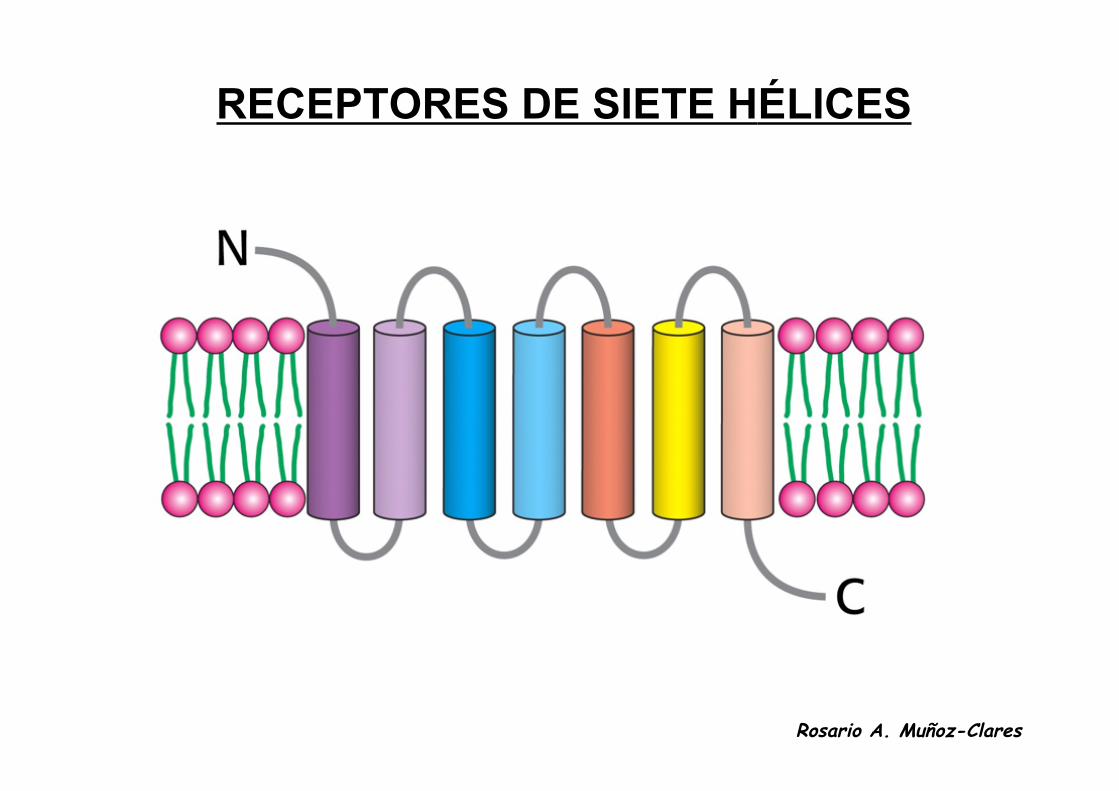
RECEPTORES DE SIETE HÉLICES
Rosario A. Muñoz-Clares

RECEPTORES DE SIETE HÉLICES
Rosario A. Muñoz-Clares

PROTEÍNAS G HETEROTRIMÉRICAS
Rosario A. Muñoz-Clares
• Están formadas por tres subunidades α, β y γ , que están unidas cuando lasubunidad α tiene al GDP unido.• La subunidad α se separa de las otras dos subunidades cuando intercambiaGDP por GTP, y se une entonces a la adenilato ciclasa.
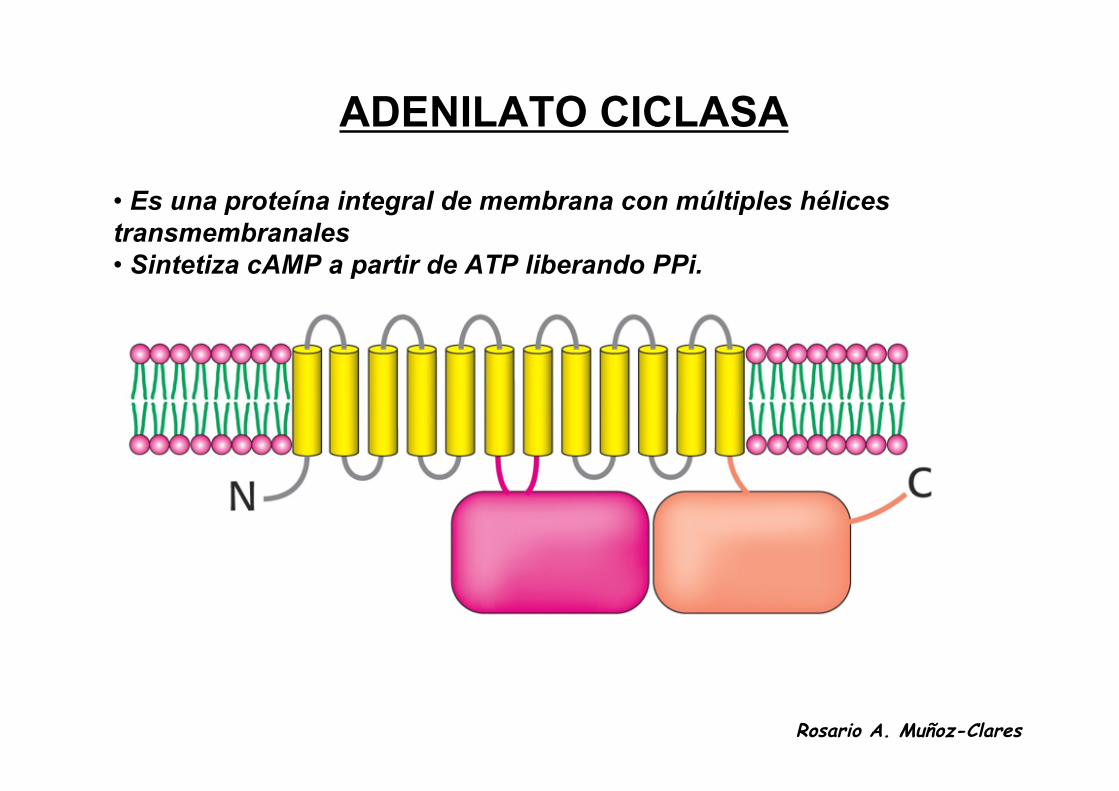
ADENILATO CICLASA
Rosario A. Muñoz-Clares
• Es una proteína integral de membrana con múltiples hélicestransmembranales• Sintetiza cAMP a partir de ATP liberando PPi.

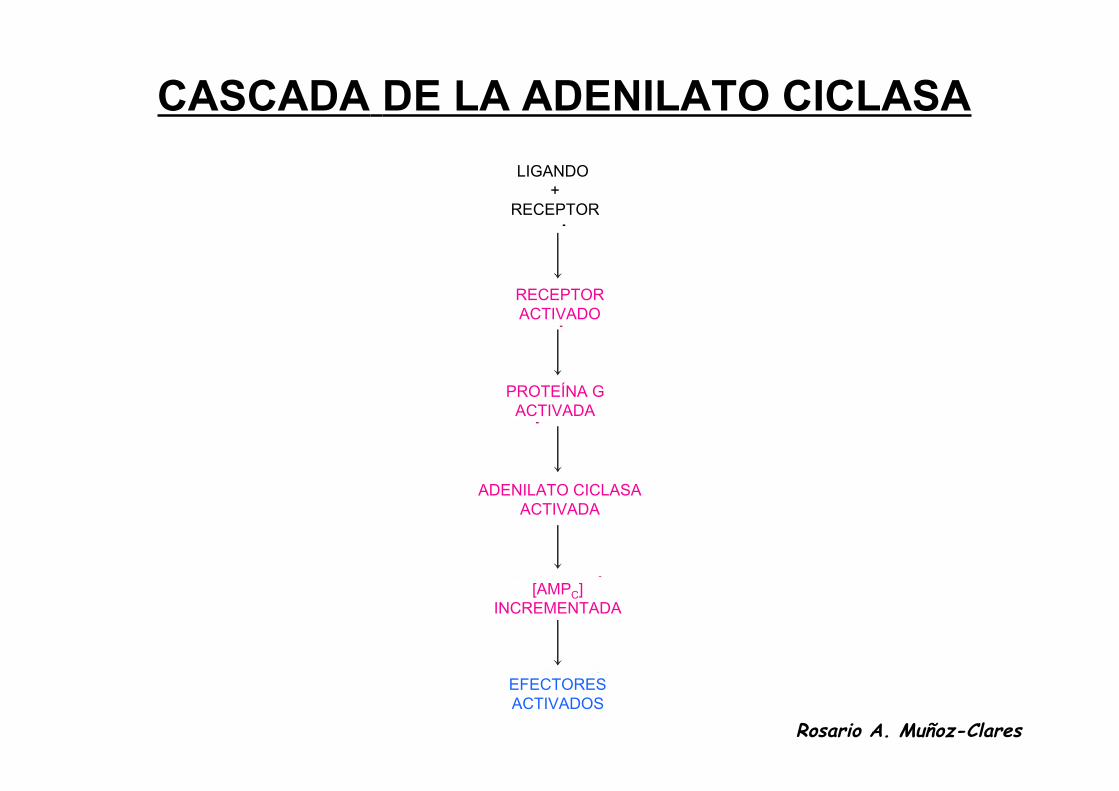
CASCADA DE LA ADENILATO CICLASA
Rosario A. Muñoz-Clares
• La subunidad α activa a la adenilato ciclasa al unírsele. La adenilato ciclasa cataliza la formación de muchas moléculas de cAMP.• Una vez que la subunidad α hidroliza al GTP se separa de la adenilato ciclasa que de esta forma se inactiva.• La subunidad α vuelve a unirse a las subunidades β y γ de la proteína G.

CASCADA DE LA ADENILATO CICLASA
Rosario A. Muñoz-Clares
• La señal original producida por la unión de una sola molécula de la hormona a su receptor se amplifica enormemente porque:
•Un receptor unido a la hormona puede activar a muchas proteínas G.•La adenilato cinasa forma muchas moléculas de cAMP mientras permanece
unida a la subunidad α,

RESPUESTA OLFATORIA
Rosario A. Muñoz-Clares
CASCADA DE LA ADENILATO CICLASA
Rosario A. Muñoz-Clares
LIGANDO +
RECEPTOR
RECEPTORACTIVADO
PROTEÍNA GACTIVADA
ADENILATO CICLASAACTIVADA
[AMPC]INCREMENTADA
EFECTORESACTIVADOS

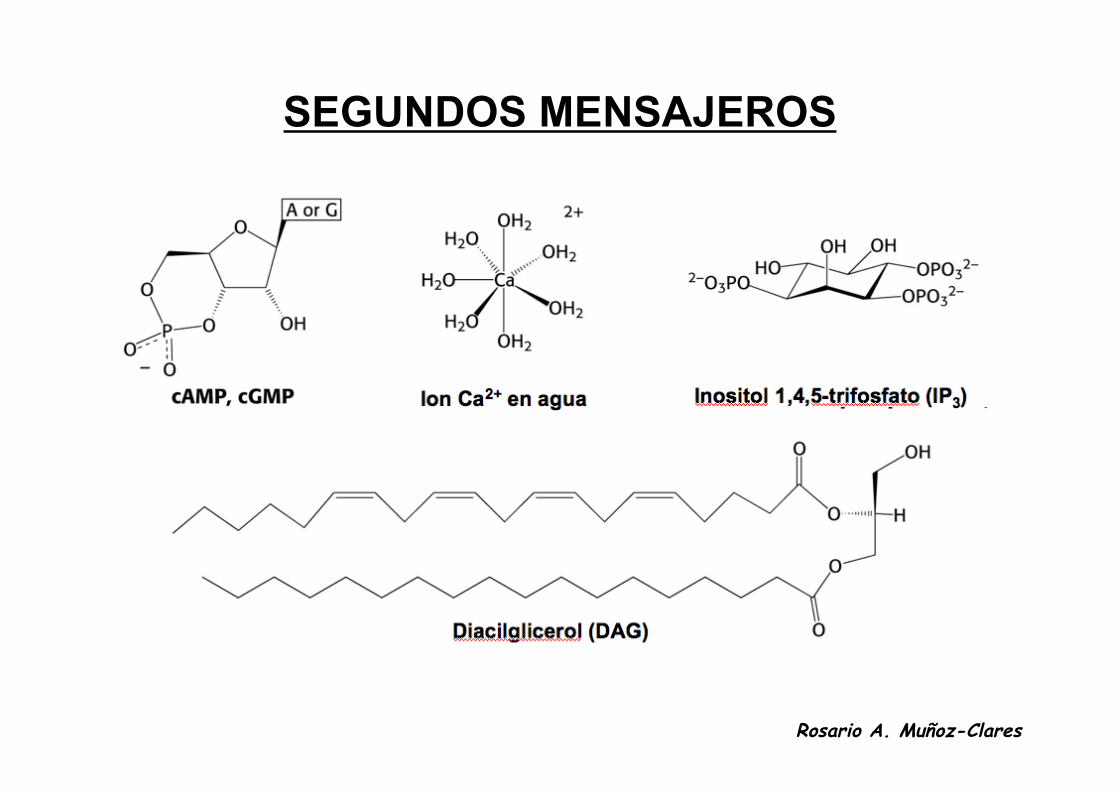
SEGUNDOS MENSAJEROS
• Son señales intracelulares que participan en la transducción de la señalextracelular. Las hormonas son los primeros mensajeros.
• Son iones o moléculas de bajo peso molecular.
• Su concentración cambia en un intervalo amplio y de forma transitoria enrespuesta a la señal extracelular.
• Los más frecuentes son:– el 3',5'-AMP cíclico (cAMP)– 3',5'-GMP cíclico (cGMP)– 1,2-diacilglicerol (DAG)– inositol 1,4,5-trifosfato (IP3)– calcio (Ca2+)
Rosario A. Muñoz-Clares
SEGUNDOS MENSAJEROS
Rosario A. Muñoz-Clares

cAMP
Rosario A. Muñoz-Clares
• El cAMP es un mensajero citosólico que participa en muchos procesos.• Es sintetizado por la adenilato ciclasa a partir de AMP y convertido de nuevo en AMP
por la cAMP fosfodiesterasa. Tiene una vida media muy corta.• Puede activar a la proteín cinasa A (PKA), la que puede fosforilar numeosas proteínas
desencadenando así diversas respuestas.• O puede inducir la apertura de canales catiónicos, lo que desencadena un impulso
nervioso.

PROTEÍN CINASA A (PKA)
Rosario A. Muñoz-Clares
• Posee dos tipos de subunidades, la regulatoria (R) y la catalítica (C) queforman un complejo R2C2 enzimáticamente inactivo.
• La unión del cAMP a las subunidades R disocia al complejo y libera las dossubunidades C activas.

Ca2+ COMO SEGUNDO MENSAJERO
Rosario A. Muñoz-Clares
• El Ca2+ participa como segundo mensajero en numerosas rutas de transducción deseñales, como las involucradas en la visión y el olfato, en la cascada de losfosfoinosítidos y en la regulación de la contracción muscular .
• La células tiene sistemas de transporte para mantener las concentraciones de Ca2+
muy bajas.
• Esto permite que se pueda incrementar mucho y de forma transitoria laconcentración de Ca2+ abriendo canales específicos de este catión, que así puede serusado como segundo mensajero.
• Su capacidad de ser segundo mensajero se debe a que se une fuertemente aproteínas e induce en ellas grandes cambios conformacionales.
• La calmodulina es una proteína que sirve como un sensor de Ca2+ en casi todas lascélulas eucarióticas.
• El complejo Ca2+-calmodulina se une a diversas proteinas (cinasas, bombas)activándolas, lo que media la respuesta a la señal.

CASCADA DE LOS FOSFOINOSÍTIDOS
Rosario A. Muñoz-Clares
• También es iniciada por receptores de siete hélices y mediada por proteínas Gheterotriméricas.
• Los mensajeros intracelulares de esta cascada se producen por la hidrólisis delfosfatidil inositol 4,5-bifosfato (PIP2) que es un fosfolípido de la membranaplasmática.
• La enzima fosfolipasa C cataliza esta hidrólisis, produciendo dos segundosmensajeros: el inositol 1,4,5-trifosfato (IP3) y el diacilglicerol.
• El IP3 abre canales de Ca2+ localizados en las membranas del retículo endoplásmico,permitiendo así la salida del catión de sus lugares de almacenamiento y produciendoun incremento transitorio en la concentración de Ca2+ citosólico.
• El diacilglicerol activa a la proteín cinasa C (PKC) que fosforila a muchas proteínas,las que median la respuesta a la señal inicial.
• La PKC es sólo activa en presencia de Ca2+ y de fosfatidíl serina. El diacilgicerolincrementa la afinidad de la PKC por el Ca2+.
• El IP3 y el diacilglicerol tienen una vida muy corta, son degradados rápidamente.

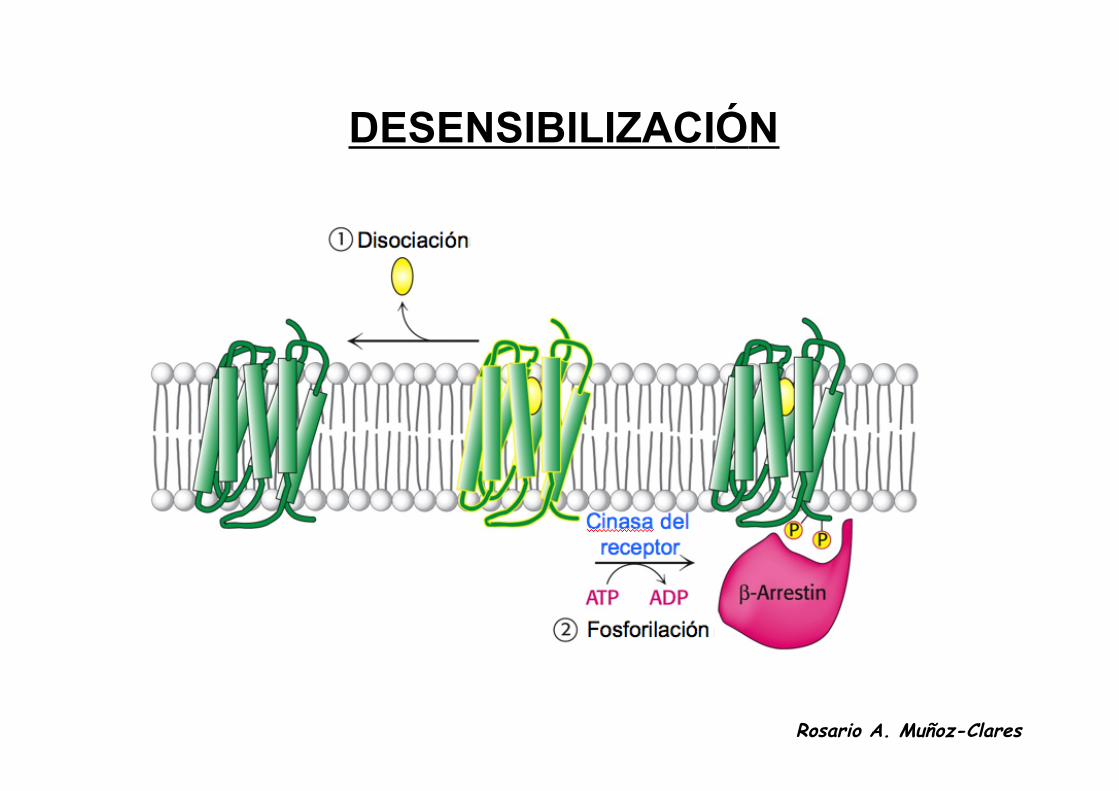
DESENSIBILIZACIÓN
• Los receptores de siete hélices se adaptan a estímulos duraderosreduciendo su respuesta a ellos en un proceso conocido comodesensibilización.
• Consiste en la fosforilación del receptor cuando está unido a lahormona y su internalización en vesículas especializadas.
• Una vez que el estímulo desaparece el receptor es desfosforilado ydevuelto a la superficie celular.
Rosario A. Muñoz-Clares
DESENSIBILIZACIÓN
Rosario A. Muñoz-Clares

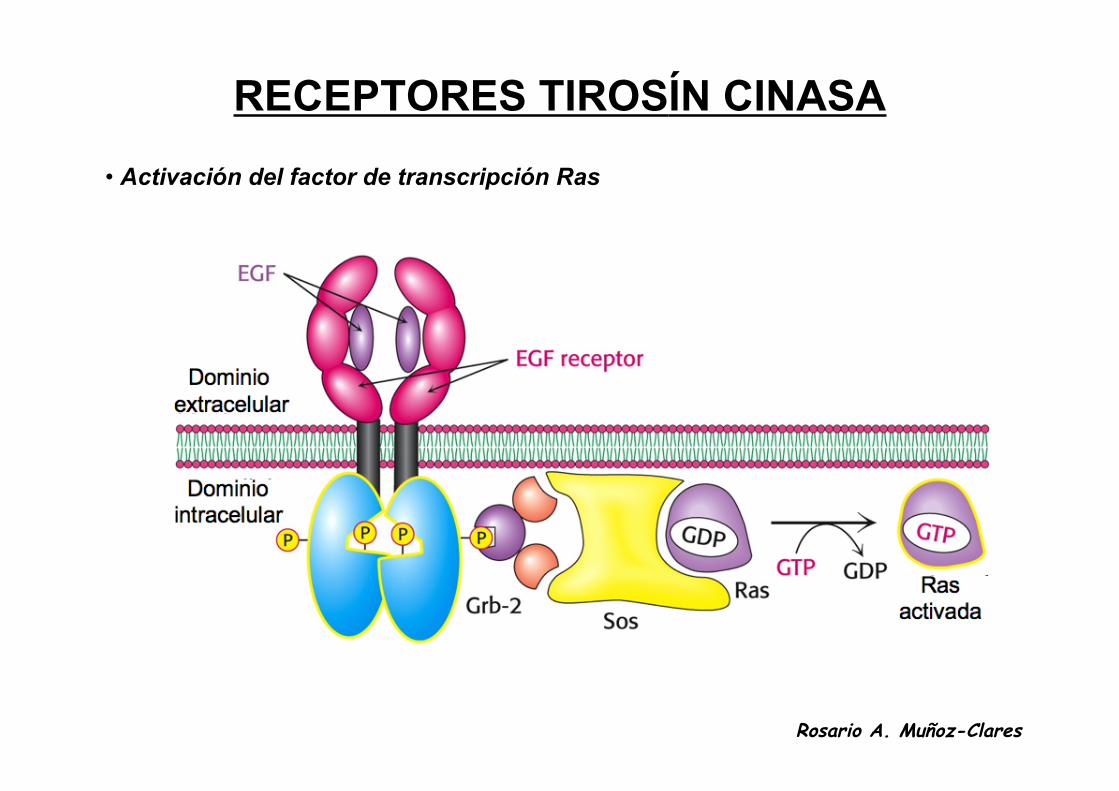
RECEPTORES TIROSÍN CINASA
• Son proteínas integrales de membrana que cuando unen a la hormonacorrespondiente (factores de crecimiento) en su dominio extracelular seactivan como tirosin cinasas o median la activación de tirosín cinasas. Lastirosín cinasas a su vez regulan las actividades de proteínas citoplasmáticasespecíficas que inducen crecimiento celular y diferenciación.
• El dominio catalítico está situado en la parte citosólica de la membrana. Elreceptor se dimeriza y autofosforila al unir a su ligando.
• La autofosforilación incrementa su capacidad de fosforilar a otras proteínasporque las tirosinas fosforiladas son sitios de reconocimiento de estasproteínas.
• Las mutaciones que conducen a una actividad continua de este receptorproducen cánceres.
Rosario A. Muñoz-Clares

RECEPTORES TIROSÍN CINASA
Rosario A. Muñoz-Clares
RECEPTORES TIROSÍN CINASA
Rosario A. Muñoz-Clares
• Activación del factor de transcripción Ras

RECEPTORES DE ACETIL COLINA
• Son canales que atraviesan la membrana y que se abren ocierran cuando unen a un ligando, de manera que inician o paranel flujo de un ión específico, tales como Ca2+ o K+, hacia adentroo afuera de una célula o un organelo.
• Participan en la transmisión del impulso nervioso.
Rosario A. Muñoz-Clares

RECEPTOR DE ACETIL COLINA
Rosario A. Muñoz-Clares
• El receptor de acetil colina es un canal selectivo para cationes regulado porligando que media la transmisión de la señal nerviosa a través de las sinapsis.
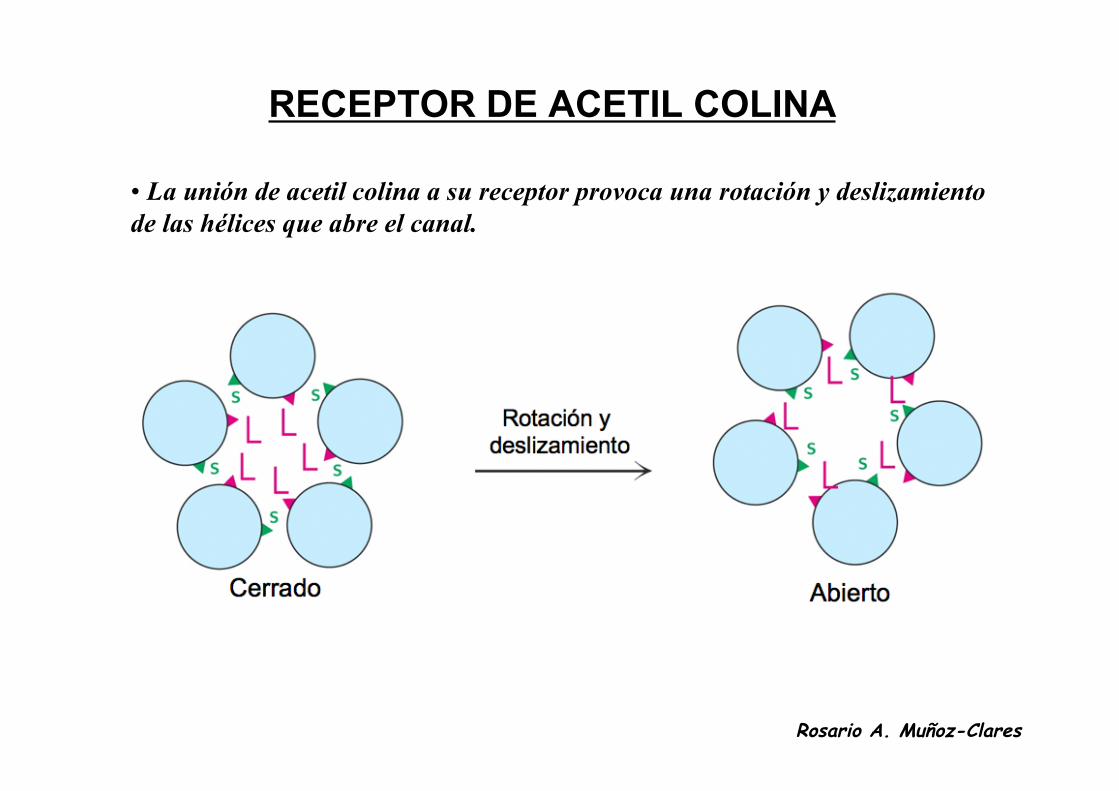
RECEPTOR DE ACETIL COLINA
• La unión de acetil colina a su receptor provoca una rotación y deslizamientode las hélices que abre el canal.
Rosario A. Muñoz-Clares