trabajos prÁcticos ejercicios adicionales … adicionales tp1... · butia paraguayensis (barb....
TRANSCRIPT

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
1
TRABAJOS PRÁCTICOS
EJERCICIOS ADICIONALES
PRIMERA PARTE Contenido
EJERCICIO 1 – REGISTRO DE CARACTERES MOLECULARES. SITIOS DE RESTRICCION..................... 2 EJERCICIO 2 – NOMENCLATURA BIOLÓGICA III . NOMBRES............................................................ 3 EJERCICIO 3 – ANÁLISIS DE ARTÍCULOS CIENTÍFICOS DE TIPO TAXONÓMICO................................. 3
EJERCICIO 4 – LISTA SINONÍMICA. PANTOMORUS RUIZI................................................................... 3 EJERCICIO 5 – CONSULTA DE NOMENCLATOR ZOOLOGICUS ........................................................... 5 EJERCICIO 6 – CONSULTA DEL ICZN ............................................................................................ 5
EJERCICIO 7 – DECISIONES TAXONÓMICAS. RECONOCIMIENTO DE ESPECIES ................................ 5
EJERCICIO 8 – DECISIONES TAXONÓMICAS. RECONOCIMIENTO DE ESPECIES ................................ 7
EJERCICIO 9 – UTILIZACIÓN DE BASE DE DATOS ............................................................................ 8 EJERCICIO 10 – CONSULTA DE CATÁLOGOS ONLINE....................................................................... 9 EJERCICIO 11 – BÚSQUEDA DE INFORMACIÓN EN INTERNET.......................................................... 9 EJERCICIO 12 – INTERPRETACION FEOGRAMAS ............................................................................. 9 EJERCICIO 13 – ANÁLISIS DE COMPONENTES PRINCIPALES. INTERPRETACIÓN DE RESULTADOS ... 11 EJERCICIO 14 – ANÁLISIS DE COMPONENTES PRINCIPALES. INTERPRETACIÓN DE RESULTADOS ... 14
EJERCICIO 15 – ANÁLISIS DE COMPONENTES PRINCIPALES. INTERPRETACIÓN DE RESULTADOS.. 15

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
2 EJERCICIO 1 – REGISTRO DE CARACTERES MOLECULARES. SITIOS DE RESTRICCION Langor & Sperling (1995) obtuvieron un mapa de restricción (Fig. 8) para cinco especies del género Pissodes ( Coleoptera: Curculionidae), gorgojos muy perjudiciales para las plantaciones de pinos. En el mapa se observan los sitios en que 10 endonucleasas de restricción han cortado los genes mitocondriales de la Citocromo Oxidasa I (COI) y Citocromo Oxidasa II (COII). Las enzimas utilizadas se identificaron mediante los acrónimos A, B, D, E, H, N, R, S, T, y X. Los estados de caracteres para cada especie son sitios presentes o ausentes. Los sitios polimórficos se hallan indicados con líneas interrumpidas (pueden codificarse con un signo de interrogación). El número total de caracteres será igual a la suma de los cortes producidos por todas las enzimas. 1. A partir del mapa de restricción de la figura 8, obtenga una tabla de especies por caracteres. 2. ¿Qué tipo de técnicas emplearía para analizar estos datos? Justifique su respuesta.
Figura 8. Mapa de restricción correspondientes a cinco especies del género Pissodes (Coleoptera: Curculionidae) (Langor & Sperling, 1995). Las 10 enzimas de restricción utilizadas se identifican con letras (A, B, D, E, H, M, R, S, T y X) y los sitios donde cada enzima cortó al ADN mitocondrial (genes COI y COII) se hallan numerados correlativamente.ç

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
3 Referencia LANGOR, D. W. & F. A. H. SPERLING. 1995. Mitochondrial DNA variation and identification of
bark weevils in the Pissodes strobi species group in Western Canada (Coleoptera: Curculionidae). The Canadian Entomologist 127: 895- 911.
EJERCICIO 2 – NOMENCLATURA BIOLÓGICA
Analice los siguientes nombres científicos e indique: 1. A qué reino y categoría taxonómica pertenecen. 2. Qué otra información puede deducirse para cada uno de ellos.
Origanum balearicum Pourret ex Lange (1863) Quedius suturalis (Marsham 1840) Stephens (1912)
Cienfuegosia sulphurea var. genuina Gürke in Mantius Colocasia esculenta (L.) Schott in Schott & Endl. (1832) Helochares abbreviatus (Fabricius, 1801) Cleistes rodea Lindley f. pallida Carnevali Dioclea scabra var. brownii Maxwell Butia paraguayensis (Barb. Rodr.) Becc. (1936) Chalicodoma (Chelostomoides) reflexa Snelling, 1990 Chalicodoma (Chelostomoides) quatridentata (Mitchell) Snelling Culex pipiens pipiens Linneo
EJERCICIO 3 – ANÁLISIS DE ARTÍCULOS CIENTÍFICOS DE TIPO TAXONÓMICO Selecciones tres artículos científicos publicados en revistas periódicas de tipo taxonómico. Podrá consultarlas en biblioteca o a través de Internet. La dirección electrónica de la biblioteca de la Secretaría de Ciencia y Técnica de la Nación es www.biblioteca.secyt.gov.ar. Brinde la cita bibliográfica completa de los trabajos consultados y realice un resumen de los principales aportes taxonómicos que realizó el o los autores (descripción de nuevas especies, nuevas combinaciones, redescripciones, análisis cladísticos, nueva clasificación, etc.). EJERCICIO 4 – LISTA SINONÍMICA. Pantomorus ruizi Morrone & Lanteri (1991) realizaron un estudio taxonómico completo sobre la especie de gorgojos Pantomorus ruizi (Brèthes 1925) (Coleoptera: Curculionidae), la cual presenta un alto grado de variación intra e interpoblacional (Fig. 9). Debido a esta variación la especie ha recibido varios nombres. La lista sinonímica que publicaron dichos autores es la siguiente:

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
4
Figura 9. Variación morfológica hallada en la especie Pantomorus ruizi (Brèthes) (Coleoptera: Curculionidae) (ilustración modificada de Morrone & Lanteri, 1991). Referencia: MORRONE, J. J. & A. A.
LANTERI. 1991(90). Ubicación sistemática y variación intraespecífica de Pantomorus ruizi (Bréthes) (Coleoptera: Curculionidae). Revista de la Sociedad Entomológica Argentina 49(1-4): 17-26.
Pantomorus ruizi (Brèthes 1925) Naupactus subvitatus Fairmaire & Germain, 1861: 7 (non Boheman, 1840); Gemminger & Harold, 1871: 2201 (catálogo); Hustache 1947: 138 (sin. de Asynonychus cervinus); Kuschel, 1949: 15 (sin. de Pantomorus cervinus). Mimographus ruizi Brèthes, 1925: 204; Dalla Torre et al., 1936: 14 (catálogo). Naupactus subvittulus van Emden 1936: 23 (nombre de reemplazo para Naupactus subvittatus Fairmaire & Germain); Kuschel 1949: 15 (sin. de Pantomorus cervinus); Kuschel 1955: 279 (sin. de Naupactus ruizi). Pantomorus ruizi; Buchanan, 1941: 61; Kuschel, 1949: 12; Kuschel, 1952: 236; Morrone, 1983: 15. Naupactus ovelipennis Hustache, 1943 (nomen nudum); Bosq, 1943: 55; Wibmer & O’Brien, 1986: 27 (catálogo). Asynonychus variabilis Hustache, 1947: 139; Kuschel 1949: 12 (sin. de Pantomorus ruizi). Asynonychus variabilis var. intermedius Hustache 1947: 139; Wibmer & O’Brien, 1986: 61 (sin. de N. ruizi) (catálogo). Naupactus ruizi; Kuschel, 1955: 279; Wibmer & O’Brien, 1986: 61 (catálogo). Brinde una explicación acerca de los cambios nomenclaturales expresados en la lista sinonímica.

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
5 EJERCICIO 5 - CONSULTA DE NOMENCLATOR ZOOLOGICUS Durante la realización de un trabajo de revisión sistemática del género Aspidisca se concluye que tres de las especies, por sus características deben ser separadas del género y ubicadas en un nuevo género. Debido a que estas tres especies se caracterizan por su aspecto hialino, se decidió el nombre Hyalina para esta nueva entidad genérica. A través de la consulta de Nomenclator Zoologicus (http://uio.mbl.edu/NomenclatorZoologicus/ ), establezca: 1. ¿Cuántas veces fue utilizado este nombre con anterioridad? Indique quien fue el autor del
nombre, el año de publicación y a qué grupo zoológico pertenece. 2. ¿Es correcto el empleo de este nombre para la nueva entidad? ¿Por qué? 3. ¿Cuál de los nombres mencionados en el punto 1 es el nombre válido? 4. ¿Cuáles son los nombres substitutos de los homónimos junior? ¿Son nombres válidos? EJERCICIO 6 - CONSULTA DEL ICZN II Hydroporus discretus Fairmaire & Brisout de Barneville, 1859 es una especie de coleóptero ampliamente distribuido en Europa y Asia. Este nombre es usado en forma continua desde 1887 y citado en cientos de publicaciones científicas. Al realizar una revisión genérica se concluye que esta especie es sinónimo de Hydroporus neuter Fairmaire & Laboulbène, 1855, nombre no citado con posterioridad a su descripción. De acuerdo al artículo 23.9.3 del ICZN que puede consultarse en http://www.iczn.org/iczn/index.jsp (Chapter 6: Validity of names and nomenclatural acts) indique cual sería el nombre válido del taxón que promueva la estabilidad nomenclatural. Justifique su respuesta.
EJERCICIO 7 - DECISIONES TAXONÓMICAS. RECONOCIMIENT O DE ESPECIES Wagneriella albula y Wagneriella lineata (Coleoptera, Curculionidae) fueron descriptas por Hustache en 1923 (páginas 295 y 297 respectivamente). Lanteri (1982) observó que ambos taxones correspondían a ejemplares macho y hembra de una misma especie. Los machos (W. albula) eran de tamaño más pequeño, con élitros de color casi uniforme, y los primeros artejos antenal y tarsal (pata 1) más engrosados. Las hembras (W. lineata) eran de tamaño mayor, los élitros presentaban un patrón de coloración con líneas oscuras, característico, y los primeros artejos antenal y tarsal no estaban engrosados (Fig. 12). 1. ¿Qué tipo de variación presenta la especie estudiada por Lanteri? 2. ¿Qué decisión nomenclatural corresponde adoptar? Proponga un nombre válido para el taxón y
justifique su respuesta. Referencia:

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
6 LANTERI, A. A. 1982. Estudio taxonómico del género Wagneriella Hustache (Coleoptera:
Curculionidae). Revista de la Sociedad Entomológica Argentina 41(1-4): 61-64.
Figura 12. Caracteres morfológicos de Wagneriella Hustache (Coleoptera: Curculionidae). Hembra (a-j): a, esternito 8; b, ovipositor en vista ventral; c, ovipositor, vista lateral; d, espermateca; e, cabeza, tórax y élitros, vista dorsal; f, idem, vista lateral; g, cabeza, vista frontal; h, antena; i, tarsitos del primer par de patas; j, abdomen, vista ventral. Macho (k-r): k, cabeza, tórax y élitros, vista dorsal; l, vista lateral; m, cabeza, vista frontal; n, antena; o, tarsitos del primer par de patas; p, abdomen; q, aedeagus, vista ventral; r, idem, vista lateral.

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
7 EJERCICIO 8 – DECISIONES TAXONÓMICAS. RECONOCIMIENT O DE ESPECIES Para la especie de tucura Metaleptea brevicornis (Linneo) (Orthoptera: Acridoidea) se habían descripto dos subespecies, M. brevicornis brevicornis (Linneo), distribuida en América del Norte y América Central, y M. brevicornis adspersa (Blanchard), distribuida en América del Sur (Fig. 14), sobre la base de diferencias en sus caracteres morfométricos. Nuevos estudios realizados por Donato & Cigliano (2000), demostraron que dichos taxones presentaban además, una notable diferenciación tanto en la genitalia masculina (externa e interna), como en la genitalia femenina, y que existía correlación entre ambos tipos de caracteres. 1. ¿Cree Usted que las nuevas evidencias reunidas por Donato & Cigliano, podrían afectar el
estatus de los taxones de Metaleptea brevicornis? 2. ¿Qué decisión taxonómica y nomenclatural adoptaría? 3. ¿Los nuevos caracteres observados por los autores, le sugieren algún tipo de aislamiento
reproductivo común en insectos? 4. Brinde el nombre válido de, o de los taxones en cuestión.
Figura 14. Metaleptea brevicornis (Linneo): a, macho, vista lateral; b- e, genitalia de una de las subespecies estudiadas; f-i genitalia de otra de las subespecies. b, f: genitalia masculina externa; c, d, g, h, genitalia masculina interna; e, i: genitalia femenina. Referencia:

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
8 DONATO, M. & M. M. CIGLIANO. 2000. Revision of the genus Metaleptea Brunner von
Wattenwyl (Orthoptera; Acrididae; Hyalopterygini). Transactions of the American Entomological Society 126:145-173.
EJERCICIO 9 - UTILIZACIÓN DE BASE DE DATOS Timi et al. (1999) describen la fauna de trematodes (Platyhelminthes) parásitos conocidos para la anchoíta (Engraulis anchoita) de Argentina y Uruguay. Como resultado de este trabajo los autores reconocieron tres especies. Dos desarrollan su fase adulta en la anchoita: Parahemiurus merus (Linton, 1910) (parásita del estómago de la anchoíta) y Lecithochirium microstomum Chandler, 1935 (parásita de la vejiga natatoria). La tercera forma es un estadio larvario (metacercaria) identificada a nivel genérico (Cardiocephaloides sp. ) que se desarrolla en los ojos de la anchoita y que su fase adulta se encuentra en otro hospedador.
Lecithochirium sp.
1. Con anterioridad al estudio realizado por Timi et al. (1999), Parahemiurus merus se conocía para otras especies del género Engraulis: E. australis; E. capensis; E. encrasicolus, E. japonicus, E. mordax y E. ringens. A partir de la información disponible en www.fishbase.org indique: • ¿Para qué océanos se conoce esta especie de parásito? • ¿Todas las especies de Engraulis mencionadas son tratadas como válidas en la base de datos? Justifique. 2. Lecithochirium microstomum se conoce también para las siguientes especies de peces: Trichiurus lepturus, Micropogonias furnieri, Oligoplites palometa; Oligoplites saliens e, Isopisthus parvipinnis. • ¿Cuáles de estas especies se distribuyen en el Océano Atlántico Sudoccidental y cuáles pueden ser encontradas específicamente en el Mar Argentino? 3. No se conoce la forma adulta de la metacercaria de Cardiocephaloides sp. hallada en la anchoita. Sin embargo es conocido que las especies del género parasitan especies de aves del género Spheniscus. • A partir de la distribución de Engraulis anchoíta y de la información suministrada en la base de datos http://www.bsc-eoc.org/avibase indique qué especie de Spheniscus podría ser el hospedador definitivo de esta especie parásita.
Referencia: TIMI, J.T.; S.R. MARTORELLI & N.H. SARDELLA. 1999. Digenetic trematodes parasitic on
Engraulis anchoíta (Pisces: Engraulidae) from Argentina and Uruguay. Folia Parasitologica 46: 132-138.

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
9 EJERCICIO 10 - CONSULTA DE CATÁLOGOS ONLINE Los siguientes nombres corresponden a especies de arañas de la familia Lycosidae descriptas por Mello-Leitão y sus holotipos se encuentran depositados en el Museo de La Plata.
1. Lycosa beltran Mello-Leitão, 1942 2. Lycosa bezzii Mello-Leitão, 1944 3. Lycosa bridarollii Mello-Leitão, 1945 4. Lycosa chacoensis Mello-Leitão, 1942 5. Lycosa cosquin Mello-Leitão, 1941 6. Lycosa hylton-scottae Mello-Leitão, 1941 7. Lycosa leucophthalma Mello-Leitão, 1940 8. Lycosa nigrosecta Mello-Leitão, 1940 9. Lycosa planithoracis Mello-Leitão, 1938 10. Lycosa passerina Mello-Leitão, 1941
Indique como son consideradas en el Catálogo Mundial de Arañas elaborado por N. I. Platnick (The World Spider Catalog, Version 8.5, http://research.amnh.org/entomology/spiders/catalog/INTRO1.html ) EJERCICIO 11 - BÚSQUEDA DE INFORMACIÓN EN INTERNET A través de una búsqueda en internet averigüe que Instituciones reúnen colecciones biológicas y que grupos están representados. Algunas direcciones electrónicas sugeridas son: 1. Museo de La Plata (http://www.fcnym.unlp.edu.ar/ ) 2. The natural History Museum, Londres, Gran Brataña (http://www.nhm.ac.uk/research-curation/collections/departmental-collections/index.html ) 3. Smithsonian Institution, Washington D.C., EEUU (http://www.mnh.si.edu/rc/ )
EJERCICIO 12 – INTERPRETACION FEOGRAMAS La especie de tucura Trimerotropis pallidipennis (Orthoptera: Acrididae) se distribuye en toda América. En algunas poblaciones sudamericanas de zonas montañosas se observaron inversiones cromosómicas pericéntricas, presentes en determinadas frecuencias; las poblaciones norteamericanas, en cambio, no presentan dichas inversiones (son monomórficas). Matrajt et al. (1996) llevaron a cabo un estudio cromosómico de poblaciones de la Argentina, distribuidas a lo largo de varios gradientes altitudinales, con el fin de establecer si el polimorfismo cromosómico observado se correlaciona con alguna variable ambiental (mayor altitud y/o de temperaturas más bajas). El cariotipo de los machos de T. pallidipennis consiste en 23 cromosomas (22 + X0), los pares 1-3 son grandes, 4-8 medianos y 9-11 pequeños. Los cromosomas grandes son submetacéntricos (SM), el cromosoma X es metacéntrico (M), y los restantes son acrocéntricos (A). Las inversiones pericéntricas se presentan en los pares de cromosomas medianos y como consecuencia de ellas la
Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
10 forma acrocéntrica básica cambia en submetacéntrica (SM1, SM2, SM3 y SM4), metacéntrica (M) o acrocéntrica invertida (AI). Los valores de frecuencia de aparición de estos reordenamientos en poblaciones de Mendoza, se pueden observar en la tabla IV, en que 0 significa ausencia de la morfología cromosómica indicada en la segunda columna, y 1, que todos los individuos de dicha población presentan dicha morfología. En las figura 17 se representan los fenogramas obtenidos al analizar la matriz de frecuencias cromosómicas de la tabla IV y una matriz de datos de frecuencias de alozimas (Matrajt et al. 1996; Confalonieri et al., 1998). 1. ¿Considera Usted que los fenogramas basados en datos de frecuencias cromosómicas y
frecuencias de alozimas postulan agrupamientos similares? 2. ¿Los agrupamientos observados se correlacionan con algún parámetro ambiental? En la figura 18 se ilustra el fenograma resultante de un análisis de frecuencias cromosómicas que incluye cuatro de las poblaciones mendocinas previamente estudiadas (todas menos Chosmes y Cacheuta) y otras dos poblaciones, de Quilmes (Tucumán, 2150 m) y de Bariloche (Río Negro, 863 m). Esta última localidad y Puente del Inca son las que registran temperaturas mínimas más bajas, entre todas las muestras estudiadas. 3. La nueva información sobre temperaturas mínimas registradas ¿afecta la interpretación de los
datos realizada previamente por Usted? Evalúe los efectos de la altura y las temperaturas mínimas en el surgimiento de reordenamiento cromosómicos.
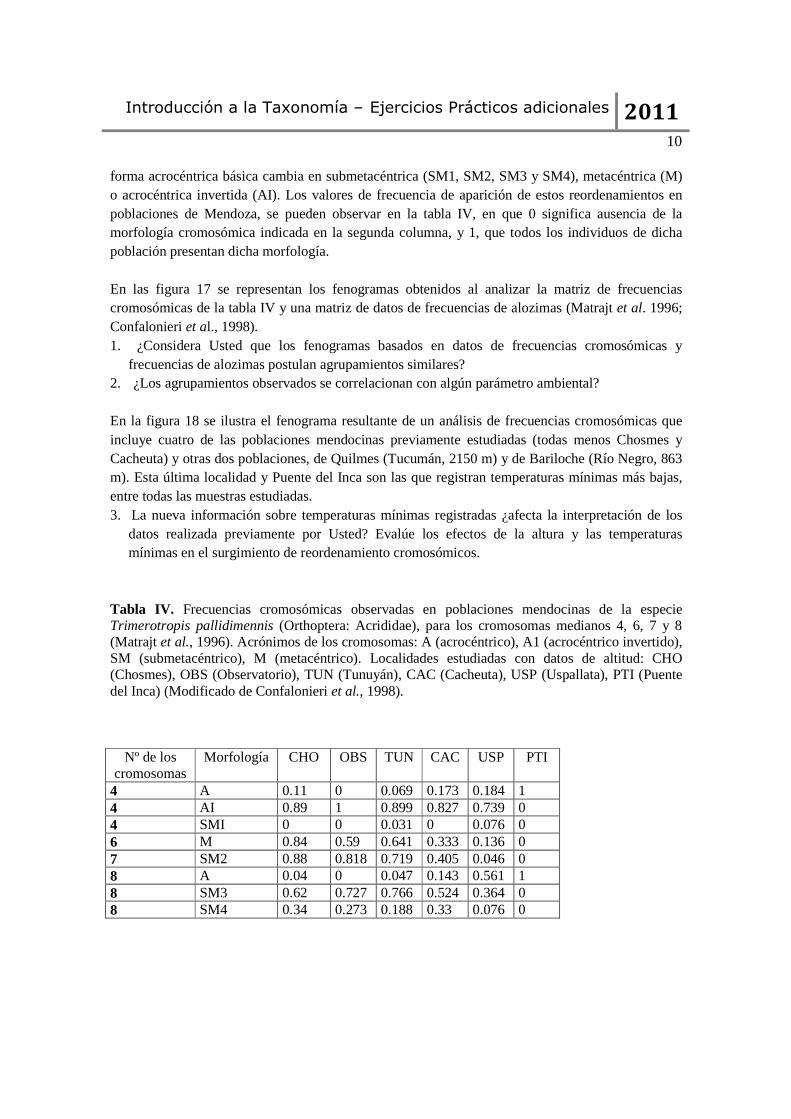
Tabla IV. Frecuencias cromosómicas observadas en poblaciones mendocinas de la especie Trimerotropis pallidimennis (Orthoptera: Acrididae), para los cromosomas medianos 4, 6, 7 y 8 (Matrajt et al., 1996). Acrónimos de los cromosomas: A (acrocéntrico), A1 (acrocéntrico invertido), SM (submetacéntrico), M (metacéntrico). Localidades estudiadas con datos de altitud: CHO (Chosmes), OBS (Observatorio), TUN (Tunuyán), CAC (Cacheuta), USP (Uspallata), PTI (Puente del Inca) (Modificado de Confalonieri et al., 1998).
Nº de los
cromosomas Morfología CHO OBS TUN CAC USP PTI
4 A 0.11 0 0.069 0.173 0.184 1 4 AI 0.89 1 0.899 0.827 0.739 0 4 SMI 0 0 0.031 0 0.076 0 6 M 0.84 0.59 0.641 0.333 0.136 0 7 SM2 0.88 0.818 0.719 0.405 0.046 0 8 A 0.04 0 0.047 0.143 0.561 1 8 SM3 0.62 0.727 0.766 0.524 0.364 0 8 SM4 0.34 0.273 0.188 0.33 0.076 0

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
11 Figura 17. Fenograma UPGMA de poblaciones de la especie T. pallidipennis (Orthoptera: Acrididae) basado en los datos de frecuencias cromosómicas de la tabla IV y en frecuencias de alozimas. Para cada localidad se brindan los datos de altitud. (Modificado de Matrajt et al., 1996).
Figura 18. Fenograma UPGMA basado en datos de frecuencias cromosómicas de poblaciones de la especie T. pallidipennis (Orthoptera: Acrididae). Incluye cuatro localidades de Mendoza (= Tabla IV) y dos localidades de otras provincias, Quilmes (Tucumán) y Bariloche (Río Negro) (Modificado de Confalonieri et al., 1998). Referencias: CONFALONIERI, V. A., A. S. SEQUEIRA, L. TODARO & J. C. VILARDI. 1998. Mitochondrial
DNA and phylogeography of the grasshopper Trimerotropis pallidipennis in relation to clinal distribution of chromosome polymorphisms. Heredity 81: 444- 452.
MATRAJT, M., V. CONFALONIERI & J. VILARDI. 1996. Parallel adaptive patterns of allozyme and inversion polymorphisms on an ecological gradient. Heredity 76: 246- 354.
EJERCICIO 13 – ANÁLISIS DE COMPONENTES PRINCIPALES. INTERPRETACIÓN DE RESULTADOS En el sudoeste de la provincia de Buenos Aires se hallan representadas dos subespecies de abejas, Apis mellifera mellifera (abeja germánica) y Apis mellifera ligustica (abeja italiana) (Hymenoptera: Apidae), y existen además ejemplares con características intermedias entre ambas subespecies. Las abejas italianas presentan por lo general menor grado de agresividad y mayor productividad, es por
1,2 1 0,8 0,6 0,4 0,2 0
Puente del Inca 2720 mBariloche 863 mQuilmes 2150 mUspallata 1830 mTunuyán 900 mObservatorio 827 m
1,5 1 0,5 0Chosmes 554 mObservatorio 827 mTunuyán 900 mCacheuta 1237 mUspallata 1830 mPuente del Inca 2720 m
0,1 0,08 0,05 0,03 0Chosmes 554 mObservatorio 827 mTunuyán 900 mCacheuta 1237 mUspallata 1830 mPuente del Inca 2720 m
Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
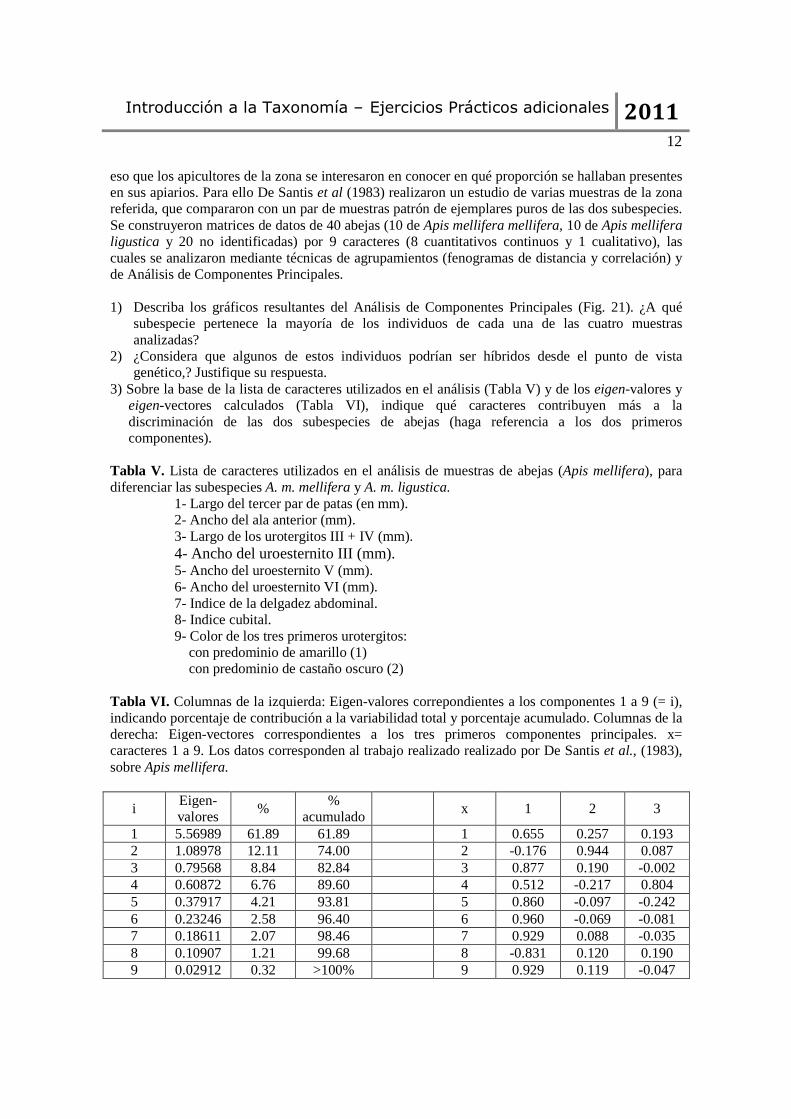
12 eso que los apicultores de la zona se interesaron en conocer en qué proporción se hallaban presentes en sus apiarios. Para ello De Santis et al (1983) realizaron un estudio de varias muestras de la zona referida, que compararon con un par de muestras patrón de ejemplares puros de las dos subespecies. Se construyeron matrices de datos de 40 abejas (10 de Apis mellifera mellifera, 10 de Apis mellifera ligustica y 20 no identificadas) por 9 caracteres (8 cuantitativos continuos y 1 cualitativo), las cuales se analizaron mediante técnicas de agrupamientos (fenogramas de distancia y correlación) y de Análisis de Componentes Principales. 1) Describa los gráficos resultantes del Análisis de Componentes Principales (Fig. 21). ¿A qué
subespecie pertenece la mayoría de los individuos de cada una de las cuatro muestras analizadas?
2) ¿Considera que algunos de estos individuos podrían ser híbridos desde el punto de vista genético,? Justifique su respuesta.
3) Sobre la base de la lista de caracteres utilizados en el análisis (Tabla V) y de los eigen-valores y eigen-vectores calculados (Tabla VI), indique qué caracteres contribuyen más a la discriminación de las dos subespecies de abejas (haga referencia a los dos primeros componentes).
Tabla V. Lista de caracteres utilizados en el análisis de muestras de abejas (Apis mellifera), para diferenciar las subespecies A. m. mellifera y A. m. ligustica.
1- Largo del tercer par de patas (en mm). 2- Ancho del ala anterior (mm). 3- Largo de los urotergitos III + IV (mm). 4- Ancho del uroesternito III (mm). 5- Ancho del uroesternito V (mm). 6- Ancho del uroesternito VI (mm). 7- Indice de la delgadez abdominal. 8- Indice cubital. 9- Color de los tres primeros urotergitos: con predominio de amarillo (1) con predominio de castaño oscuro (2)
Tabla VI. Columnas de la izquierda: Eigen-valores correpondientes a los componentes 1 a 9 (= i), indicando porcentaje de contribución a la variabilidad total y porcentaje acumulado. Columnas de la derecha: Eigen-vectores correspondientes a los tres primeros componentes principales. x= caracteres 1 a 9. Los datos corresponden al trabajo realizado realizado por De Santis et al., (1983), sobre Apis mellifera.
i Eigen-valores
% %
acumulado x 1 2 3
1 5.56989 61.89 61.89 1 0.655 0.257 0.193 2 1.08978 12.11 74.00 2 -0.176 0.944 0.087 3 0.79568 8.84 82.84 3 0.877 0.190 -0.002 4 0.60872 6.76 89.60 4 0.512 -0.217 0.804 5 0.37917 4.21 93.81 5 0.860 -0.097 -0.242 6 0.23246 2.58 96.40 6 0.960 -0.069 -0.081 7 0.18611 2.07 98.46 7 0.929 0.088 -0.035 8 0.10907 1.21 99.68 8 -0.831 0.120 0.190 9 0.02912 0.32 >100% 9 0.929 0.119 -0.047

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
13
Figura 21. Ilustración de los gráficos resultantes de la técnica de Análisis de Componentes Principales (PCA), aplicado a muestras de abejas (Apis mellifera) de cuatro apiarios (a, b, c, d). Los individuos de la muestra problema se numeraron del 1 al 20 (círculos negros), los de Apis mellifera mellifera del 21 al 30 (círculos blancos) y los de Apis mellifera ligustica del 31 al 40 (cuadrados blancos) (modificado de De Santis et al., 1983).

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
14 Referencia: DE SANTIS, L., A. BOLOGNESE, L. CORNEJO, J. V. CRISCI, N. DIAZ, A. A. LANTERI & J. VIDAL SARMIENTO. 1983. Estudio taxonómico de dos subespecies de Apis mellifera (A. m. mellifera y A. m. ligustica) en proceso de hibridación, mediante el empleo de técnicas numéricas. Revista del Museo de La Plata (N. S. Zool.) 13: 45-63. EJERCICIO 14 – ANÁLISIS DE COMPONENTES PRINCIPALES. INTERPRETACIÓN DE RESULTADOS En las figuras 22 y 23 se ilustran los resultados de la aplicación de la técnica de Análisis de Componentes Principales a individuos pertenecientes al grupo de Asynonychus durius (Coleoptera: Curculionidae) (Lanteri et al., 1987). La figura 22 representa los componentes I y II, y la figura 23, los componentes II y III. 1. ¿Qué diferencia observa entre ambos gráficos? ¿Cuál de ellos resulta más discriminativo para
las especies del grupo de A. durius? ¿Cuál de ellas se encuentra más diferenciada de las restantes?
Figura 22. Gráfico bidimensional correspondiente a un Análisis de Componentes Principales, aplicado al estudio de las especies del grupo de Asynonychus durius (modificado de Lanteri et al., 1987). El componente I (abcisa) acumula el 41.42% de la variación y el II (ordenada), el 13.37 %. Las unidades de estudio son individuos. Asynonychus durius (círculos blancos); A. santafecinus (cuadrados blancos); A. viridipallens (triángulos blancos); A. tessellatus (círculos negros); A. pallidus morfotipo I (triángulos negros); A. pallidus morfotipo II (cuadrados negros).

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
15
Figura 23. Ídem Figura 22. Representación de los componentes II (abscisa), que acumula el 13.37 % de la variación y el componente III (ordenada), que acumula el 9.55%.
Referencia: LANTERI, A. A., N. B. DÍAZ, M. S. LOIÁCONO & M. DEL C. COSCARÓN. 1987. Aplicación
de técnicas numéricas al estudio sistemático del grupo de Asynonychus durius (Germar) (Coleoptera: Curculionidae). Entomological. Arbaiten Museu Frey, 35/36: 171-198.
EJERCICIO 15 - ANÁLISIS DE COMPONENTES PRINCIPALES. INTERPRETACIÓN DE RESULTADOS La especie de tucura Metaleptea brevicornis (Orthoptera: Acridoidea) tiene una amplia distribución en las Américas. Dentro de este taxón se reconocían dos subespecies: M. brevicornis brevicornis, distribuida en la costa este de América del Norte hasta Costa Rica, y M. brevicornis adspersa, distribuida desde Panamá hasta el centro de la Argentina. Posteriormente estas subespecies fueron elevadas a la categoría de especies sobre la base de diferencias en las estructuras genitales (Donato & Cigliano, 2000). Como parte de este trabajo los autores realizaron un análisis multivariado, a fin de establecer si cada una de las especies presentaba variación clinal en el tamaño corporal, ya que diferentes autores sostenían que la variación morfométrica observada en dichos taxones se correspondía con un gradiente ambiental, geográfico. Examinaron especímenes de ambas especies y construyeron dos matrices separadas, una de machos y otra de hembras, ya que existe dimorfismo sexual en el tamaño, y reunir los dos sexos en una misma matriz podría inducir a error. Los individuos estudiados proceden de las localidades que se enumeran en la Tabla VII (las localidades del trabajo original de Donato & Cigliano se agruparon por países y se redujo el número de las mismas para simplificar la ejercitación). 1. De acuerdo al gráfico correspondiente a un análisis de componentes principales (fig. 26) aplicado
al estudio de las especies de Metaleptea. Cosidera que las dos especies se hallan claramente separadas en el análisis?
2. ¿La variación observada es continua o se forman grupos bien diferenciados dentro de cada especie?

Introducción a la Taxonomía – Ejercicios Prácticos adicionales 2011
16 3. Considera usted que hay variación geográfica clinal en estas especies? Justifique su respuesta.
Tabla VII. Datos de procedencia de los ejemplares hembra de Metaleptea brevicornis y M. adspersa.
Acrónimo Localidad/País Acrónimo Localidad/País Acrónimo Localidad/País Ala Alabama (EE.UU) Hon Honduras Pan Panamá Arg Argentina Illi Illinois (EE.UU) Par Paraguay Bol Bolivia La Lousiana (EE.UU) Per Perú Bra Brasil Ma Maryland (EE.UU) Su Surinam Del Delaware (EE.UU) Mex Méjico Ten Tennesse (EE.UU) Ec Ecuador Mich Michigan (EE.UU) Te Texas (EE.UU) Flo Florida Mis Missouri (EE.UU) Va Virginia (EE.UU) Ga Georgia (EE.UU) Nic Nicaragua U Uruguay Gu Guatemala NJ New Jersey (EE.UU)
Figura 26. Gráfico bidimensional correspondiente a un Análisis de Componentes Principales, aplicado al estudio de las especies de Metaleptea (modificado de Donato & Cigliano, 2000). El componente I (abcisa) acumula el 49.5% de la variación y el II (ordenada), el 23.30 %. Las unidades de estudio son individuos hembra. Referencia: DONATO, M. & M. M. CIGLIANO. 2000. Revision of the Genus Melaneptera Brunner von
Wattenwyl (Orthoptera; Acrididae; Hyalopterygini). Transactions of the American Entomological Society 126:145-173.