tejido muscular
TRANSCRIPT

Tejido muscular. Prof. Tomás Atauje Calderón
Histología
Obstetricia – Ciclo III

Tejido muscular
• Permite los movimientos del cuerpo y la reducción del diámetro de las vísceras y los vasos. Ello es posible porque posee células alargadas que son capaces de acortarse.
• Además, se asocian entre sí de manera tal que componen láminas o haces, los cuales se disponen de forma paralela a fin de poder actuar de manera coordinada. Debido a su gran longitud, estas células se llaman también fibras musculares.
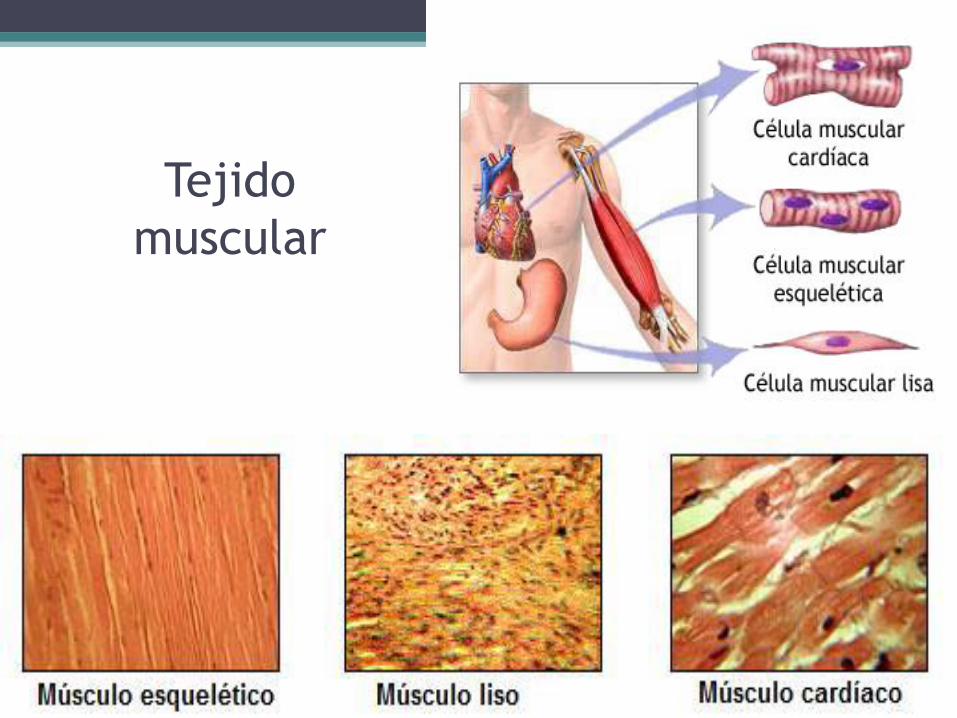
• El tejido muscular puede dividirse en tres tipos:
▫ Estriado voluntario ▫ Estriado cardiaco ▫ Liso
Tejido
muscular

Tejido muscular estriado voluntario
• También llamado tejido muscular esquelético. • Parte esencial de cada uno de los músculos del esqueleto; que están
compuestos de tejido muscular estriado voluntario y tejido conectivo. En este último transcurren los vasos y los nervios destinados a las células musculares y además se forman los tendones.
• Los músculos no solo hacen posible la locomoción sino cualquier movimiento voluntario del cuerpo; existen músculos estriados que no pertenecen al esqueleto sino a vísceras, como los de la faringe y la parte superior del esófago o el diafragma.
• En ocasiones, los músculos esqueléticos se mueven involuntariamente como respuesta a estímulos que producen movimientos reflejos. Ejemplo: Músculos del miembro inferior (reflejo rotuliano) y músculos de los párpados.
*Vistos al microscopio óptico, los músculos esqueléticos presentan bandas
transversales oscuras alternadas con bandas claras, por lo cual se los llama músculos estriados.

Células musculares estriadas
esqueléticas • Miden hasta 30 cm de largo y aunque en la mayoría de células el ancho es de 10 a 100
μm, a veces alcanzan los 500 μm.
• Son cilíndricas y de ancho uniforme, excepto en los extremos, donde se adelgazan. Además, son multinucleadas y todos los núcleos se localizan por debajo de la membrana plasmática, la cual está rodeada por la lámina externa (capa de material secretado por la propia célula).
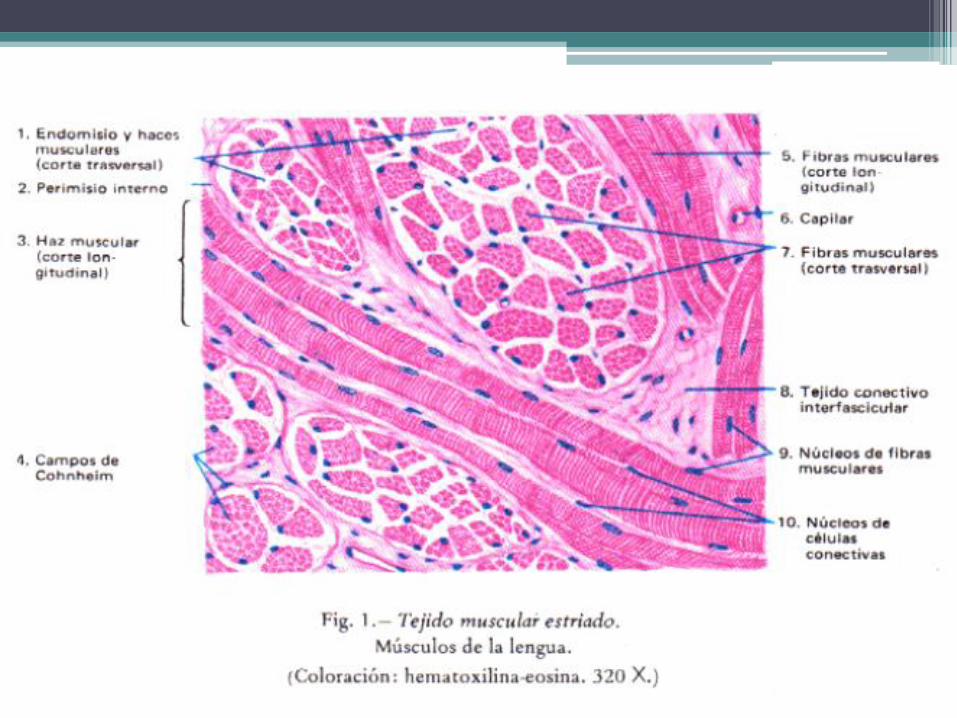
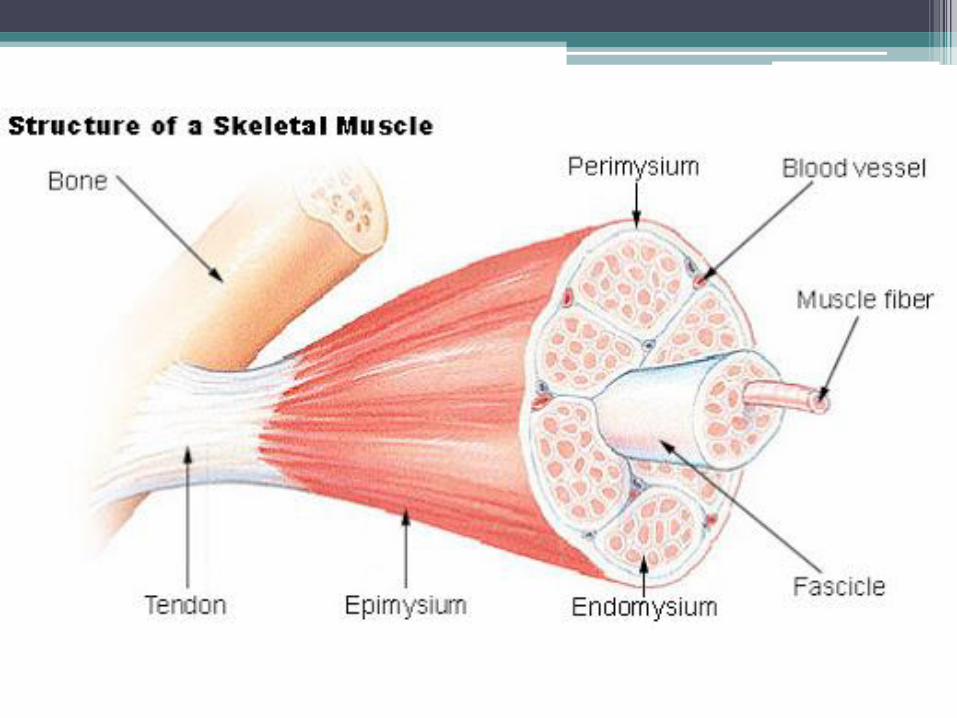
• En cada músculo las células musculares estriadas se disponen paralelamente una al lado de la otra, formando agregados longitudinales que se conocen con el nombre de fascículos. Algunos músculos poseen un solo fascículo, pero la mayoría están integrados por varios, los cuales siguen el eje mayor del músculo.
• Cada célula muscular esta rodeada por el endomisio, que es una red de fibras reticulares situada en torno de la lámina externa. En cambio, cada fascículo está rodeado por una capa de tejido conectivo denso, denominado perimisio. A su vez, el conjunto de fascículos (el músculo) está envuelto par una capa de tejido conectivo mas denso que el perimisio, llamado epimisio.
• Los vasos sanguíneos del músculo ingresan por el epimisio, se ramifican en el perimisio y emiten capilares continuos que transcurren por el endomisio, en torno de las células musculares. Los nervios tienen un recorrido similar.

Tipos de células musculares estriadas
esqueléticas • Fibras rojas: Son pequeñas y finas. Su color se debe a que están rodeadas por
abundantes capilares sanguíneos y a que contienen gran cantidad de mioglobina. En las fibras rojas, la energía se genera mediante procesos oxidativos, motivo por el cual poseen abundantes mitocondrias. Si bien las fibras rojas se contraen más lentamente que las fibras blancas e intermedias, pueden hacerlo en forma repetida debido a que son muy resistentes a la fatiga.
• Fibras blancas: Son más grandes que las fibras rojas y las fibras intermedias. Gran parte de la energía que consumen deriva de la glucólisis anaerobia; motivo por el cual sus mitocondrias son relativamente escasas, lo mismo que los capilares que rodean a las células. Si bien se contraen rápidamente, se fatigan con facilidad.
• Fibras intermedias: Poseen características tintoriales, morfológicas y funcionales que se hallan entre las de las fibras rojas y las fibras blancas.
• Casi todos los músculos estriados voluntarios poseen los tres tipos de fibras.
• La intensidad de la fuerza mecánica que genera un músculo cuando se contrae depende del número y del tamaño de sus fibras; y ya que son las más grandes, las fibras blancas son las que aportan mayor fuerza relativa.

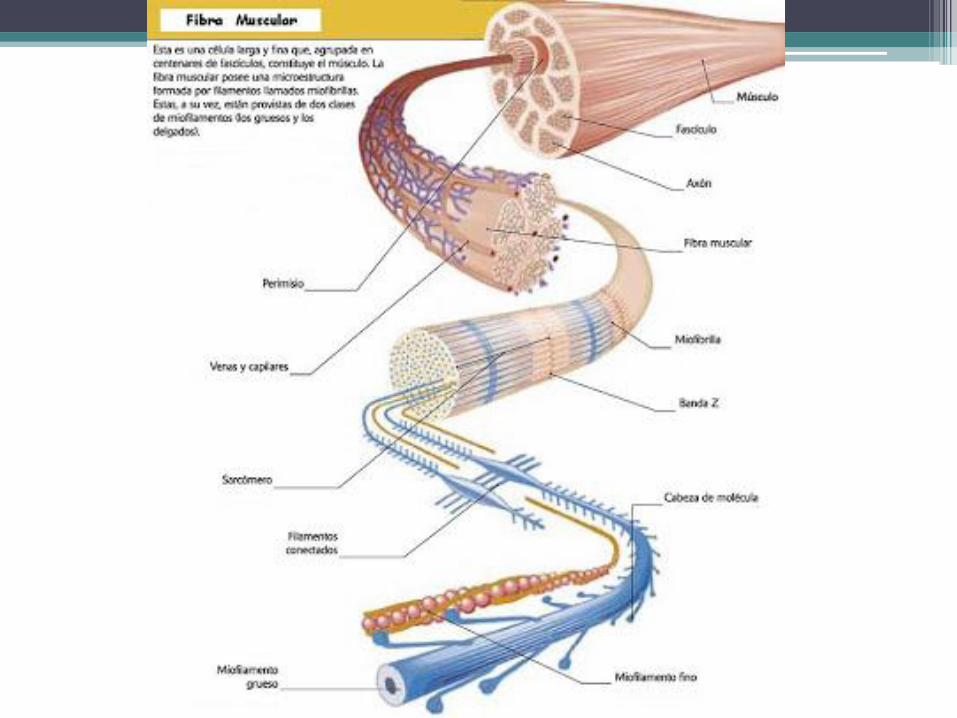
Ultraestructura de las células
musculares esqueléticas • Las células musculares estriadas voluntarias poseen una organización muy compleja y es
uno de los ejemplos más salientes de adaptación morfofuncional y de conversión de energía química en trabajo mecánico del organismo. El diseño de las células musculares estriadas es tan eficiente que algunas son capaces de contraerse y de relajarse mas de 100 veces por segundo y de producir un trabajo mil veces superior a su peso.
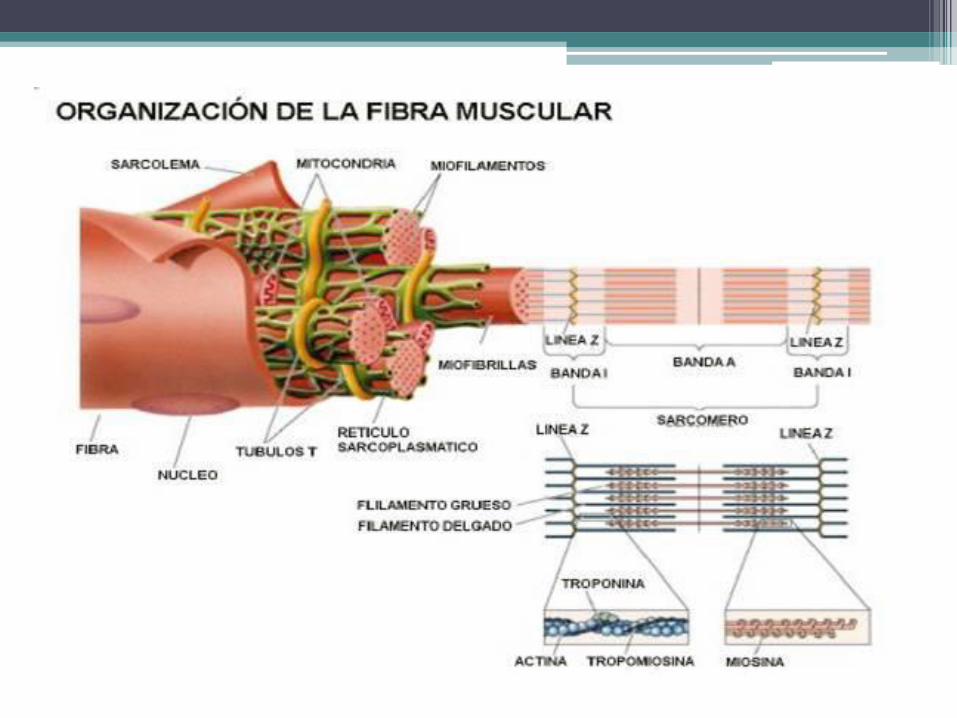
• Membrana plasmática: También llamada sarcolema, está rodeada por la lámina
externa. En toda su extensión desarrolla unas invaginaciones tubulares que ingresan perpendicularmente en el citoplasma y que están separadas entre sí por distancias más a menos regulares. Estas invaginaciones se denominan túbulos T y sus luces se comunican con el espacio extracelular.
• Citoesqueleto: Los componentes del citoesqueleto que se vinculan con la actividad
mecánica de las células musculares estriadas están representados por unas estructuras regulares y estables llamadas miofibrillas, adaptadas para acortarse durante las contracciones y alargarse en los períodos de reposo.
• En general, las miofibrillas son tan largas como las propias células musculares y se disponen paralelamente una al lado de la otra, siguiendo el eje longitudinal de las células. Cada miofibrilla mide 1 a 2 μm de diámetro y su número depende del grosor de la fibra muscular.

Ultraestructura de las células
musculares esqueléticas • La miofibrilla está compuesta por una sucesión de unidades contráctiles denominadas
sarcómeros, los cuales miden alrededor de 2 μm de longitud y tienen un ancho semejante al de la miofibrilla.
• En el límite entre un sarcómero y otro hay una estructura electrodensa, el disco Z. Se localiza en el centro de una región de poca densidad, la banda I, llamada así porque es isótropa. Como está dividida por el disco Z, la banda I se compone de dos hemibandas, una de las cuales pertenece a un sarcómero y la otra al sarcómero vecino.
• En cada sarcómero, el espacio que separa a las dos hemibandas I está ocupado por la banda A, que es mucho más densa que las hemibandas I. Su nombre se debe a que es anisótropa, es decir, birrefringente. En la parte media de la banda A se distingue una zona de poca densidad, la banda H, cuyo centro presenta la línea M, que posee una densidad parecida a la de la banda A.
• Las distintas bandas del sarcómero resaltan de la superposición periódica. De los filamentos de actina con los otros componentes del citoesqueleto. Además, como a lo ancho de toda la célula cada banda se encuentra en registro con bandas de su misma clase pertenecientes a las demás miofibrillas, en conjunto generan franjas alternantes de diferentes densidades y grosores, que son las que le confieren la designación de estriado a esta clase de músculo
• En la estructura básica del sarcómero, se observan filamentos de actina que parten de los discos Z y fibras gruesas entre dichos fiIamentos. Los cortes transversales seriados del sarcómero revelan que las hemibandas I contienen únicamente filamentos de actina; que la zona central de la banda A posee sólo fibras gruesas, y que las zonas de la banda A cercanas a las hemibandas I están compuestas por ambos elementos. En estas zonas, cada filamento de actina se halla rodeado por tres fibras gruesas, y cada una de estas, por seis filamentos de actina, de modo que el número de filamentos duplica al de las fibras.

Ultraestructura de las células
musculares esqueléticas • Retículo endoplasmático: Recibe el nombre de retículo sarcoplasmático. Posee una
organización muy distinta de la de los demás tipos celulares y se asocia con los túbulos T del sarcolema. Esta organización se debe a que desempeña funciones esenciales durante la contracción y la relajación muscular.
• Los túbulos T corren transversalmente entre las miofibrillas, en los límites entre las hemibandas I y la banda A de los sarcómeros, de modo que cada sarcómero se vincula con dos túbulos T. Cada túbulo T está acompañado por dos conductos del retículo sarcoplasmático, de diámetro bastante mayor que el del túbulo, llamados cisternas terminales.
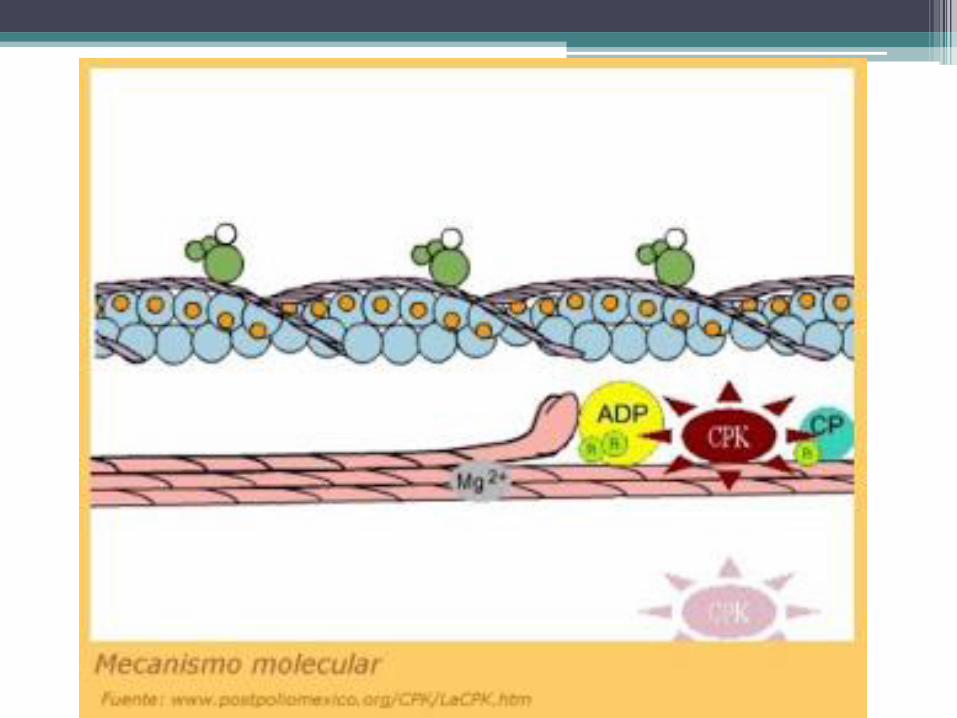
• En el músculo relajado, las cisternas terminales almacenan iones calcio. Los cuales están unidos a una proteína llamada calsecuestrina. La contracción muscular se desencadena cuando el calcio se separa de esta proteína, sale del retículo sarcoplasmático y pasa al citosol. El calcio cruza la membrana del retículo sarcoplasmático a través de un canal iónico dependiente de voltaje, que se abre cuando dicha membrana se despolariza. Al cesar la contracción, el calcio retorna al retículo sarcoplasmático a través de una permeasa activa que consume energía cedida por el ATP.
• Mitocondrias: También llamadas sarcosomas; se distribuyen entre las miofibrillas y por debajo del sarcolema. Son más abundantes en las fibras rojas e intermedias que en las fibras blancas.
• Inclusiones: El citosol contiene gotas de lípidos y gránulos de glucógeno. Las primeras prevalecen en las fibras rojas y los segundos en las fibras blancas.

Inervación del músculo esquelético • Las fibras nerviosas motoras que inervan a las células musculares estriadas voluntarias se
componen de axones pertenecientes a neuronas localizadas en el mesencéfalo, la protuberancia, el bulbo y las astas anteriores de la medula espinal.
• Cada axón se ramifica en el endomisio de una fibra muscular y los extremos de las ultimas ramas presentan una dilatación llamada terminal sináptico, que establece contacto con un área circunscrita del sarcolema, denominada placa motora terminal. Esta estructura de contacto se conoce como sinapsis neuromuscular.
• A nivel de la placa motora, la célula muscular presenta una concavidad poco profunda y su sarcolema se halla separado de la membrana plasmática del terminal sináptico por un espacio muy pequeño que se llama espacio sináptico primario. Además, el sarcolema emite hacia el citoplasma una serie de invaginaciones denominadas espacios sinápticos secundarios. Tanto el espacio sináptico primario como los secundarios están ocupados por una lámina basal continua que deriva de la lámina externa de la célula muscular.
• A nivel de la concavidad de la placa motora, el sarcolema recibe el nombre de membrana postsináptica, mientras que por su parte la membrana plasmática del terminal sináptico, separada del sarcolema por el espacio sináptico primario, se llama membrana presináptica.
• Cerca de esta, en el interior del terminal sináptico, se encuentran numerosas vesículas de alrededor de 50 nm de diámetro, conocidas como vesículas sinápticas, que contienen acetilcolina, neurotransmisor que induce la contracción de la célula muscular voluntaria.
• La capa de mielina que rodea al axón finaliza antes del nacimiento de las ramas destinadas a las células musculares, por lo que esas ramas solo están cubiertas por la célula de Schwann.

Contracción del músculo esquelético • La célula muscular estriada voluntaria se contrae cuando es inducida por el axón de la
neurona motora. El proceso se inicia al generarse un potencial de acción en la neurona. que recorre el axón y llega al terminal sináptico. La llegada del potencial de acción abre los canales de calcio dependientes de voltaje presentes en la membrana plasmática del terminal sináptico, lo cual permite que el calcio del medio extracelular ingrese en el terminal.
• El calcio desencadena la exocitosis de las vesículas sinápticas, cuya acetilcolina se libera en el espacio sináptico primario. De inmediato, la acetilcolina difunde hacia los espacios sinápticos secundarios y se combina con un receptor específico de la membrana postsináptica. Este receptor actúa como un canal de sodio dependiente de ligando (el ligando es la acetilcolina) y su apertura permite que el sodio extracelular ingrese en la célula muscular., lo cual despolariza a la membrana postsináptica y desencadena un potencial de acción que se propaga por todo el sarcolema.
• El sarcolema de los túbulos T posee una proteína transmembranosa que está en contacto con canales de calcio dependientes de voltaje presentes en la membrana de las cisternas terminales del retículo sarcoplasmático. Cuando esa proteína es alcanzada por el potencial de acción que recorre el sarcolema cambia de forma, interactúa con los canales de calcio de las cisternas terminales y los abre, lo cual permite que el calcio almacenado en el retículo sarcoplasmático pase al citosol.
• Una vez en el citosol, el calcio se combina con la troponina C del sarcómero. El complejo calcio-troponina C se une a la troponina I, cuya función es mantener a la tropomiosina junto al filamento de actina para que este no pueda tomar contacto con las cabezas de la fibra gruesa. Debido a que la unión de la troponina I con el complejo calcio-troponina C inhibe a la troponina I, la tropomiosina se libera de ella y se separa del filamento de actina, lo cual les permite a las cabezas de la fibra gruesa conectarse con dicho filamento.
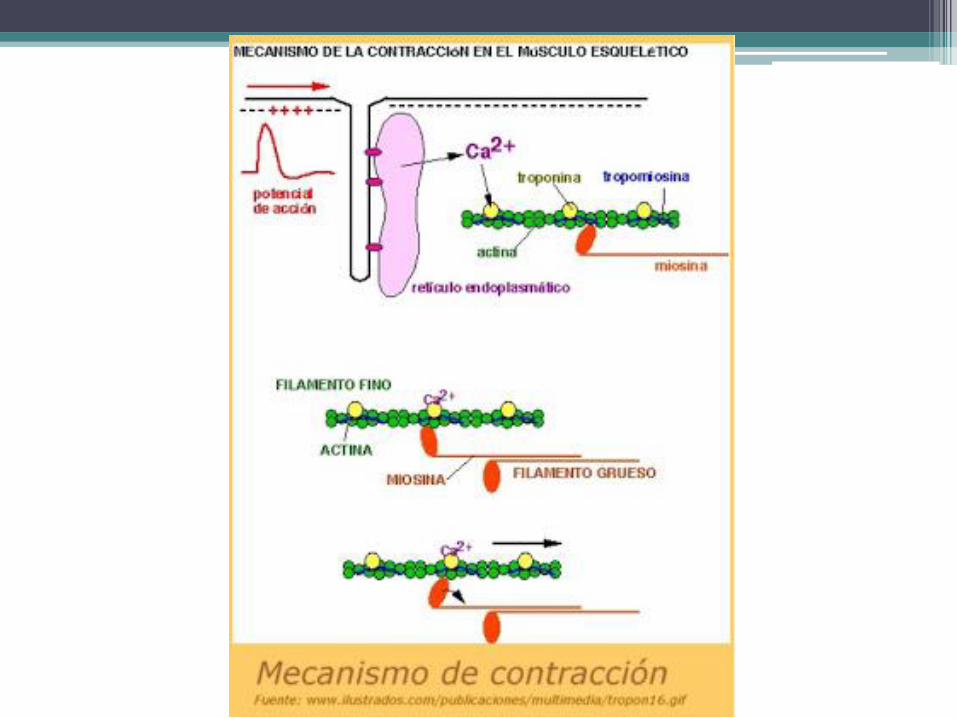

Contracción del músculo esquelético
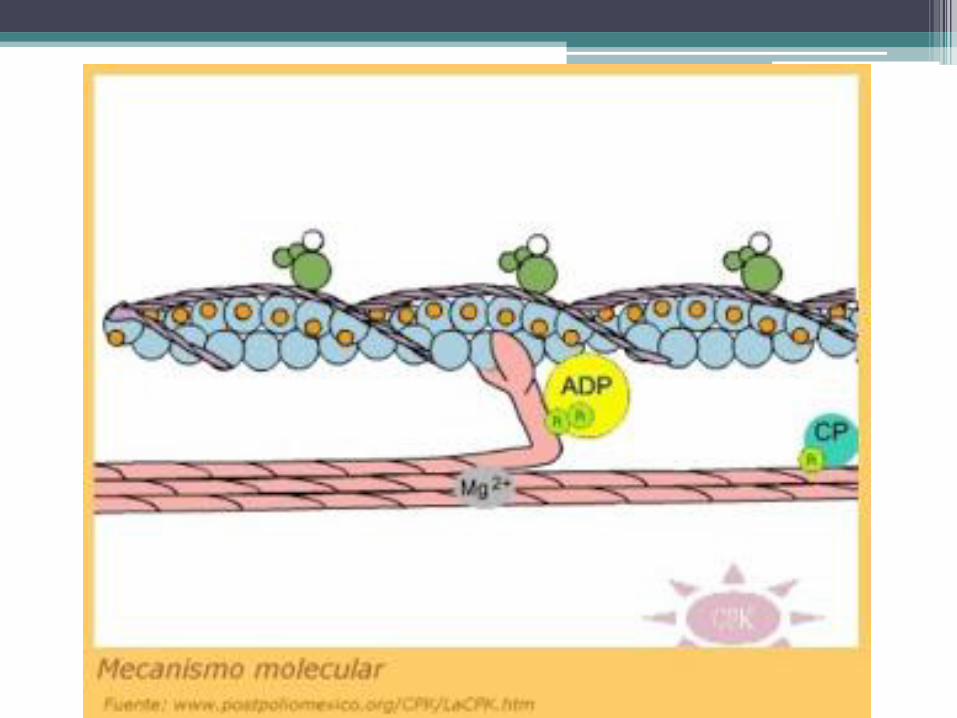
• La contracción se debe a que las cabezas de las fibras gruesas se deslizan sobre los filamentos de actina en dirección de los discos Z, hecho que los acerca mutuamente y comprime el tramo elástico de las titinas. La energía consumida por estos procesos se libera del ATP mediante una ATPasa presente en las cabezas de las fibras gruesas.
• Cuando la contracción cesa, las cabezas de las fibras gruesas se separan de los filamentos de actina. Por consiguiente, el tramo elástico de las titinas se elonga pasivamente y recupera su longitud de reposo, lo cual provoca la separación de los discos Z y el alargamiento de los sarcómeros.
• El cese de la contracción se produce cuando el calcio regresa al retículo sarcoplasmático y desaparece del citosol. Esta función la realizan las bombas de calcio presentes en la membrana del retículo. La ausencia de calcio en el citosol hace que la tropomiosina retorne a su posición de reposo, lo cual impide el contacto de las cabezas de las fibras gruesas con los filamentos de actina.
• La relajación muscular se debe a que la acetilcolina se desprende de su receptor (situado en la membrana postsináptica) y es hidrolizada mediante la acetilcolinesterasa, enzima presente en el espacio sináptico.



Tejido muscular estriado cardiaco
• Compone el miocardio, que es la pared muscular del corazón. Consta de células cilíndricas mucho más cortas y delgadas que las del tejido muscular estriado voluntario, pues miden alrededor de 90 μm de largo por 15 μm de diámetro. No obstante, debido a que las células cardíacas no actúan como unidades independientes y se unen por sus puntas, forman fibras que funcionan de manera semejante a las fibras de los músculos voluntarios.
• Las células cardíacas, al igual que las células esqueléticas, también presentan bandas transversales oscuras alternadas con bandas claras, pero poseen un solo núcleo central alargado, en lugar de muchos núcleos periféricos. Además, algunas células cardiacas tuercen su dirección y se unen con las células cercanas.
• Con tinciones especiales, los cortes longitudinales muestran también unas líneas oscuras transversales. Se llaman discos intercalares y revelan los lugares donde las células se unen por sus puntas.
• Las células musculares cardíacas están separadas por tabiques muy delgados de tejido conectivo laxo, por los que transcurren capilares sanguíneos y fibras nerviosas.
• A diferencia de las células musculares voluntarias, las cardíacas carecen de células satélites, de modo que si se lesionan no pueden ser reemplazadas.


Ultraestructura de las células
musculares cardiacas • Poseen una sola miofibrilla (no varias), que ocupa casi todo el diámetro celular y es "invadida"
desde la periferia por "tabiques" incompletos, integrados por mitocondrias, túbulos y sáculos del retículo sarcoplasmático, túbulos T procedentes del sarcolema, gotas de lípidos, gránulos de glucógeno y gránulos de lipofuscina (pigmento que aumenta con la edad).
• Mitocondrias: Son muy numerosas y poseen crestas apretadas, largas e irregulares. Las gotas de lípidos y los gránulos de glucógeno se hallan cerca de las mitocondrias, a las que proveen. Respectivamente, de ácidos grasos y de glucosa para que generen energía química en forma de ATP. Dado que para esa función las mitocondrias demandan gran cantidad de oxígeno, este se acumula en moléculas de mioglobina, las cuales abundan en el citosol.
• La posición central del núcleo interfiere en el trayecto de los sarcómeros que corren por el eje central de la miofibrilla, que al divergir crean dos espacios triangulares junto al núcleo. Ambos contienen mitocondrias, gotas de lípidos y gránulos de glucógeno. En uno de ellos reside el complejo de Golgi, que es relativamente pequeño.
• El sarcolema emite túbulos T mucho más anchos que los de las células musculares estriadas voluntarias. Son tan grandes que permiten que la lámina externa que rodea al sarcolema ingrese en éstos y tapice sus membranas. En las células cardíacas, los túbulos T corren junto a los discos Z de la miofibrilla solitaria, no entre las hemibandas I y la banda A.

Ultraestructura de las células
musculares cardiacas • El retículo sarcoplasmático está compuesto por numerosos túbulos
longitudinales anastomosados, los cuales de tanto en tanto emiten unos sáculos pequeños que se orientan de dos maneras: algunos corren en dirección longitudinal y se apoyan sobre la cara interna del sarcolema, mientras que otros corren en dirección transversal y se asocian a los túbulos T . Los sáculos transversales equivalen a las cisternas terminales de las células musculares estriadas voluntarias, aunque no forman tríadas.
• Los discos intercalares corresponden a las uniones entre las puntas de las células
cardíacas. Son escaleriformes y sus tramos transversales se localizan en medio de dos hemibandas I, por lo que equivalen a discos Z. Además, en esos tramos las membranas celulares enfrentadas se interdigitan, lo cual incrementa considerablemente la adhesión.
• Las membranas celulares de los discos intercalares se conectan entre sí mediante los siguientes tipos de uniones: ▫ Desmosomas: Soportan gran parte de las tracciones generadas al contraerse las células
cardíacas. ▫ Fascias adhesivas: Refuerzan a los desmosomas, pues son áreas de unión que poseen
elementos análogos a los de los cinturones adhesivos de los epitelios. ▫ Uniones comunicantes: A través de los cuales los potenciales eléctricos de acción se
propagan de una célula a otra a fin de que estas sincronicen sus contracciones.


Inervación del músculo cardiaco
• La inervación del tejido cardíaco es provista por fibras simpáticas y parasimpáticas del sistema nervioso autónomo.
• Si bien no desencadenan las contracciones cardíacas, regulan su frecuencia e intensidad.

Contracción del músculo cardiaco • Los sarcómeros de la miofibrilla de la célula muscular cardíaca son
idénticos a los de las miofibrillas de la célula muscular voluntaria y también se contraen cuando se eleva la concentración de calcio en el citosol. Sin embargo, en la célula cardíaca, el calcio proviene tanto del retículo sarcoplasmático como de la matriz extracelular.
• Además de ser independientes de la voluntad, las células musculares
cardíacas no necesitan de nervios para contraerse. Así, sus contracciones se clasifican como miógenas o intrínsecas, pues se desencadenan por la despolarización espontánea del sarcolema.
• Esta despolarización se transfiere a la membrana de los sáculos longitudinales y transversales del retículo sarcoplasmático (a los sáculos transversales llega a través de los túbulos T) y abre los canales de calcio del sarcolema y de la membrana del retículo sarcoplasmático, lo que permite que entren en el citosol iones calcio procedentes de la matriz extracelular y del retículo.
• A la inversa, durante la repolarización se activan las bombas de calcio de ambas membranas y los iones retornan a sus lugares de origen.

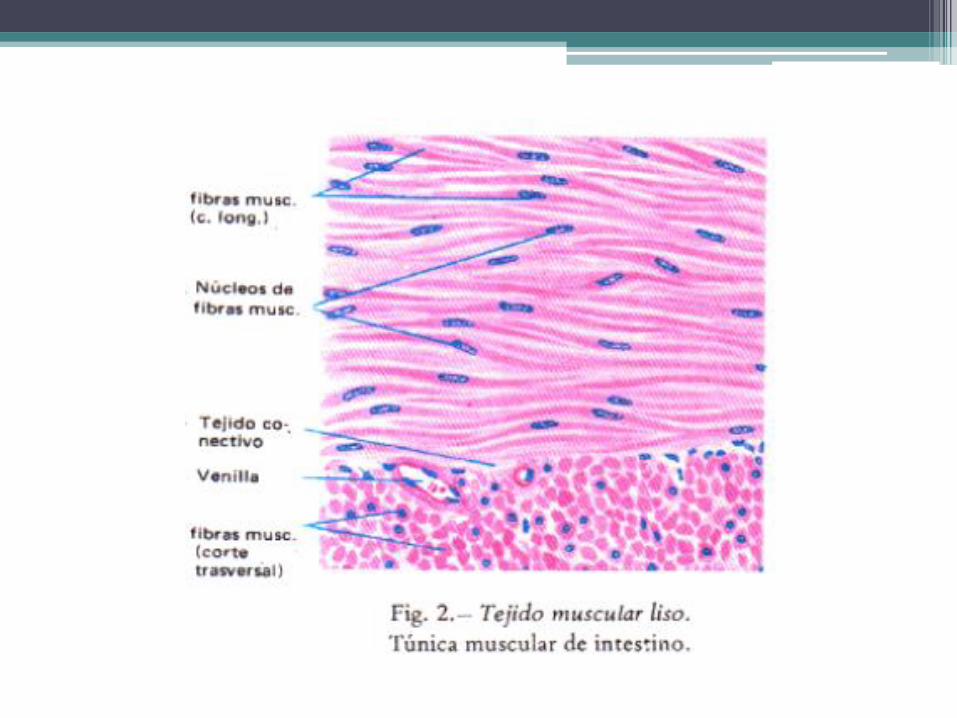
Tejido muscular liso • Las células del tejido muscular liso son fusiformes, se contraen de
modo involuntario y se les dice lisas porque no presentan las bandas claras y oscuras que caracterizan a las células musculares estriadas. Se encuentran en la mayor parte de los órganos de los sistemas digestivo, respiratorio, urinario y reproductor, en los vasos sanguíneos, en la piel, en el bazo y en el ojo.
• La longitud de las células musculares lisas varía en los distintos órganos. Por ejemplo, las de los vasos sanguíneos mas pequeños miden solo 25 μm, las del intestino miden en promedio 200 μm y las del útero grávido pueden alcanzar los 500 μm.
• La parte central de la célula es la mas ancha y aloja al núcleo de forma alargada. Las mitocondrias, la mayor parte del retículo endoplasmático y el complejo de Golgi se localizan en el citoplasma perinuclear, que contiene abundantes ribosomas libres y gránulos de glucógeno. El resto del citoplasma está reservado para los componentes del citoesqueleto, los cuales no forman miofibrillas y poseen una organización mucho más sencilla, además son capaces de generar contracciones acordes con las exigencias mecánicas de los órganos.

Tejido muscular liso
• Como en las células musculares estriadas, entre los filamentos de actina se intercalan fibras de miosina II, cuyo deslizamiento sobre los filamentos produce las contracciones. Debe señalarse que estas fibras de miosina II son mucho menos gruesas que las de los sarcómeros de las células musculares estriadas.
• En torno de la membrana plasmática de la célula muscular lisa existe una lámina externa que la separa del tejido conectivo circundante y que es producida por la propia célula. Además, la membrana plasmática desarrolla invaginaciones pequeñas que invaden el citoplasma superficial, llamadas caveolas. Dado que la membrana de estas posee canales y bombas de calcio, se cree que las caveolas desempeñan funciones similares a las de los túbulos T de las células musculares estriadas.
• Si bien en algunos órganos existen células musculares lisas aisladas, por lo general se asocian y componen láminas o fascículos de distinto espesor. En estas estructuras, la parte ancha de cada célula se sitúa junto a las partes delgadas de las células vecinas.

Inervación del músculo liso • Tejido muscular liso visceral: Sus contracciones son espontáneas, rítmicas y
coordinadas; esto último porque los impulsos contráctiles se propagan de una célula muscular a otra a través de las uniones comunicantes presentes en las membranas plasmáticas de las células contiguas. Solo unas pocas células musculares lisas son inervadas por nervios del sistema nervioso autónomo.
• Se trata de fibras simpáticas y parasimpáticas, cuyas acciones inductivas son mediadas por los neurotransmisores noradrenalina y acetilcolina, respectivamente, de efectos generalmente opuestos. Debido a que la contracción muscular es espontánea, estas fibras nerviosas regulan solamente la frecuencia y la intensidad de las contracciones, que también son moduladas por ciertos factores mecánicos y por algunas sustancias como la oxitocina (músculo uterino), la serotonina, la somatostatina y la motilina (músculo intestinal).
• Tejido muscular liso multiunitario: Sus células carecen de uniones comunicantes y se
contraen cuando son inducidas por las fibras simpáticas y parasimpáticas que las inervan. Estas fibras nerviosas se ramifican a medida que se profundizan en el tejido muscular, y cada rama terminal recorre un sector de la membrana plasmática de una de las células musculares, con la cual establece sinapsis sucesivas, denominadas por ese motivo "en passant" (de paso). Las uniones neuromusculares son más sencillas que las del músculo estriado voluntario, y la distancia entre la membrana del terminal sináptico y la membrana de la célula muscular lisa es mayor. Varias estructuras poseen tejido muscular liso multiunitario, por ejemplo: el cuerpo ciliar del ojo y algunas arterias.
• Tejido muscular liso mixto: Presenta características de los tejidos musculares visceral y
multiunitario.

Contracción del músculo liso • Sea espontáneamente o al ser inducida por un terminal sináptico, cuando la
membrana plasmática se despolariza, el calcio extracelular (depositado en las caveolas) ingresa en el citosol de la célula muscular lisa. Además, se suma calcio procedente del retículo endoplasmático.
• El calcio hace contraer a la célula, para lo cual se une a la calmodulina,
que es una proteína citosólica similar a la troponina C. El complejo calcio-calmodulina activa a una quinasa, cuya función es tomar iones fosforo del ATP y fosforilar a las fibras de miosina II. Así, las cabezas de las fibras de miosina II interactúan con los filamentos de actina y se deslizan sobre ellos, al igual que en los sarcómeros de las células musculares estriadas.
• Sólo se contraen las partes periféricas de la célula, pues en la parte central
están los filamentos de desmina, que no se acortan. Ello protege al núcleo y a los organoides de las compresiones mecánicas derivadas de las contracciones.