resumen - sappi.ipn.mxsappi.ipn.mx/cgpi/archivos_anexo/20071131_4651.pdf · captura y la...
TRANSCRIPT

1
INFORME TECNICO FINAL
EFECTO DEL ESTRÉS SOBRE LA PRODUCCIÓN Y SECRECIÓN DE IgA
EN LA MUCOSA NASAL DEL RATÓN
Registro SIP: 20071131
Responsable: Dr Rafael Campos Rodriguez
RESUMEN
El sistema inmunitario de las mucosas (MALT) comprende tejidos
asociados con la superficie de las mucosas respiratoria, digestiva y
genitourinaria, los cuales defienden el organismo de antígenos externos.
El tejido linfoide asociado a nariz (NALT) representa un compartimiento
inductor de la respuesta inmunitaria y se localiza en piso de la nariz de algunos
roedores; mientras que la lámina propia de la mucosa nasal representa al
compartimiento efector. El estrés, provoca un incremento de la secreción de
hormonas glucocorticoides y catecolaminas por activación del eje hipotálamo-
hipófisis-adrenal. Estas hormonas interactúan con receptores de las células
inmunitarias ocasionando cambios en su respuesta. El objetivo de este trabajo
fue determinar los efectos del estrés sobre el número y distribución de las
células plasmáticas IgA+ así como de la secreción de IgA. Se utilizaron
ratones Balb/c machos de 9 semanas de edad, divididos en un grupo control y
dos grupos problema, a los que se aplicó 3 horas de estrés por inmovilización,
durante 4 y 8 días.
La cuantificación de corticosterona en suero mostró un incremento
significativo en los animales estresados durante 4 días.
De cada animal, se obtuvo el NALT y se realizaron tinciones
inmunohistoquímicas para cuantificar células plasmáticas productoras de IgA
La secreción de IgA en el lavado nasal se determinó por ELISA.
Los resultados mostraron que las células plasmáticas IgA+,
disminuyeron significativamente con el estrés de 4 días y se incrementaron a
los 8 días, lo que se correlacionó con los niveles de IgA en el lavado nasal,
mostrando diferencias significativas.

2
Se concluye que el NALT es sensible al esquema de estrés utilizado y
que este afecta la respuesta humoral (células IgA+), posiblemente por la
influencia de las hormonas secretadas durante el estrés.
ANTECEDENTES
SISTEMA INMUNITARIO
Los órganos linfoides se dividen desde un punto de vista funcional en
dos categorías: linfoides primarios o centrales y secundarios o periféricos; y
desde un punto de vista anatómico-estructural en órganos capsulados y no
capsulados.
En los órganos primarios, médula ósea y timo se originan y maduran los
linfocitos B y T hasta convertirse en células competentes, capaces de
reconocer y responder en forma específica a los estímulos antigénicos. Los
órganos linfoides secundarios como los ganglios linfáticos, bazo y tejidos
linfoides asociados a las mucosas (MALT), disponen de linfocitos ya maduros e
inmunológicamente competentes y es en ellos donde se producen las
respuestas inmunitarias frente a estímulos antigénicos.
En las mucosas se encuentra la primera línea de defensa contra antígenos
externos, ya que un gran número de agentes patógenos las utilizan como vía
de entrada. El conjunto de tejidos linfoides organizados y difusos en las
mucosas del organismo, se denominan �tejidos linfoides asociados a las
mucosas� (MALT) los cuales a través de la inmunidad innata y adquirida
mantienen una homeostasis inmunitaria, a lo largo de la gran superficie interior
de las víseras, la cual comprende las mucosas oral, nasal, respiratoria, urinaria
y gastrointestinal. (Tristram GP 2001)
TEJIDO LINFOIDE ASOCIADO A NARIZ
En la mucosa respiratoria, también se encuentra una gran cantidad de
células inmunitarias, las cuales se distribuyen en la lámina propia en forma
difusa, así como en estructuras foliculares no encapsuladas, o constituyendo

3
el tejido linfoide asociado a la nariz (NALT), el cual forma parte de los tejidos
linfoides asociados a las mucosas (MALT). Entre las células del epitelio
respiratorio que recubre el NALT, también se encuentra gran cantidad de
células M (membranosas o con micropliegues), cuya principal función es la
captura y la transitosis de los antígenos desde la porción apical de la célula,
hasta la porción basal, en donde muestran el antígeno a las células
presentadoras de antígenos. (Giannasca PJ y cols 1997). Debido a su
localización anatómica, se considera un análogo del anillo de Waldeyer
humano (Boyaka NP y cols 2000) y funcionalmente se compara con la placa
de Peyer por presentar características inductoras similares. (Kiyono H y
Fukuyama S 2004)
El NALT es un compartimiento inductor y corresponde a la porción de
células linfoides organizadas. En este sitio se inicia la respuesta inmunitaria,
además de ser el sitio de cambio de isotípo de inmunoglobulina IgM+ por IgA+
(Shikina T y cols 2004); una vez terminado este proceso, las células
plasmáticas productoras de IgA, migran hacia la lámina propia sobre la que
descansa el tejido organizado, en este sitio que es la porción efectora, se lleva
acabo la producción de IgA secretora la cual se encuentra presente en el moco
nasal. Esta inmunoglobulina evita que agentes patógenos invadan las mucosas
entre ellas la nasal.
EJE HIPOTALAMO-HIPOFISIS-ADRENAL
El hipotálamo es un centro de integración que recibe y monitorea
información acerca del ambiente, y además coordina las respuestas a través
del sistema nervioso central, autónomo y endocrino. Las principal información
que recibe es la visual, olfativa, auditiva y sensitiva.
Dentro de las funciones del hipotálamo está la de controlar la secreción
de hormonas hipofisiarias, controlar las acciones involuntarias por medio del
SNA (sistema nervioso autónomo) y regular la motivación y los instintos por
medio del sistema límbico. El núcleo paraventricular del hipotálamo secreta la
hormona corticotropina (CRH) dentro del sistema portahipofisiario, la cual
regula subsecuentemente la producción de hormona adrenocorticotrofica
(ACTH) en la glándula hipófisis anterior. La hormona ACTH a su vez actúa en

4
la corteza de la glándula suprarrenal provocando la secreción de cortisol.
Durante los periodos de estrés se produce una sobreestimulacion hipotalámica,
lo cual provoca una hipersecreción hormonal.
El sistema de estrés está constituido por el eje hipotálamo-hipófisis-
adrenal (HPA) y el sistema simpático sistémico, los cuales se encuentran
controlados por señales limbicas, circadianas, neurosensoriales y hormonales.
Las situaciones de estrés, tales como la hipoglicemia, procesos inflamatorios,
fiebre, trauma, ejercicio intenso, cirugía y problemas emocionales pueden
activar este sistema. (Paca´k K y Palkovits M 2001)
MODELOS DE ESTRÉS EN ANIMALES
Hans Selye (Paca´k K y Palkovits M 2001) en 1936, fue el primer
investigador que utilizó el modelo de estrés por inmovilización en ratas, las
cuales manifestaron varios signos que caracterizaron un síndrome ocasionado
por el estrés. La manifestación mas evidente fue la presencia de hipertrofia de
la glándula adrenal, la cual se atribuyó a una sobreestimulacion por las
hormonas ACTH secretadas por la hipófisis anterior. Otra manifestación fue la
presencia de ulceración gástrica, la cual se atribuyó al incremento en la
secreción de ácido clorhídrico por las células parientales en el estómago;
también se encontró involución del sistema timo linfático, la cual se atribuyó a
la implicación del eje hipotálamo-hipófisis-adrenal así como del sistema
nervioso autónomo.
ESTRÉS Y SECRECIÓN DE IgA.
Por otro lado sujetos sometidos a estrés intenso o de larga duración así
como atletas de alto rendimiento sujetos a estrés por ejercicio intenso padecen
de enfermedades respiratorias con mayor frecuencia que los sujetos no
estresados. La razón de esta mayor susceptibilidad se desconoce, pero se
supone esta relaciona con una inhibición de la respuesta inmunitaria. A nivel de
las mucosas, los efectos del estrés se han estudiado solo sobre las
modificaciones de los niveles de IgA en saliva y no existen estudios acerca de
sus efectos a nivel de la mucosa nasal, en particular sobre la producción y
secreción de IgA.

5
OBJETIVO GENERAL
Analizar los efectos del estrés, sobre la cantidad y distribución de las
células productoras de IgA, así como los cambios en la secreción de IgA al
líquido nasal. Así como su efecto sobre síntesis de las citocinas involucradas
en la producción de IgA.
OBJETIVOS ESPECIFICOS 1. Determinas las concentraciones de corticosterona séricas en los ratones
normales y sometidos a estrés.
2. En el NALT de ratones normales y sometidos a estrés, identificar y
cuantificar por inmunohistoquímica las células plasmáticas IgA+
3. En el tejido no NALT (la lámina propia) de ratones normales y estresados,
identificar linfocitos B IgA+.
4. En muestras de lavado nasal de ratones normales y estresados, cuantificar
anticuerpos IgA mediante inmunoensayo enzimático (ELISA).
5. Determinar por RT-PCR la expresión de RNAms para IL-6 e IL-10.
MATERIAL Y METODOS Animales
Se utilizaron ratones Balb/c machos con un peso entre 21-24 g
proporcionados por Harlan México, el manejo de los animales se realizo de
acuerdo a las normas de la Comisión de Bioética de la ESM. Los ratones se
dividieron en tres grupos de 7 animales cada uno; un grupo control, uno con
tratamiento de estrés de 4 días y otro con tratamiento de 8 días.
Tratamiento de estrés. Los animales fueron sometidos a un esquema de estrés por
inmovilización en contenedores cilíndricos apropiados al tamaño del animal (5

6
cm. de largo, 3 cm de alto y 3.5 cm de diámetro) con ventilación adecuada.
Antes y después del periodo de estrés, los animales se mantuvieron en jaulas
durante los días de tratamiento, ingiriendo agua y comida a libre demanda. Los
animales control se mantuvieron en las mismas condiciones.
Los animales problema se sometieron diariamente a 3 horas de estrés,
bajo un horario establecido (8:00-11:00 AM). Para evitar el estado de
adaptación al estrés durante la inmovilización, los ratones se estimularon cada
30 minutos de la manera siguiente: 1.-Agitación de los contenedores durante
10 segundos, iniciando el ciclo a los 10 minutos. 2.-Rotación de los
contenedores durante 10 segundos, iniciando el ciclo a los 20 minutos. 3.-Por
ultimo, se sumergió la cola de los animales en agua fría durante 10 segundos,
iniciando el ciclo a los 30 minutos.
Día Agitación de los
contenedores
Rotación
de los contenedores
Inmersión de la cola en agua
fría
1 x x
2 x x x
3 x x
4 x x x Sacrificar el grupo de ratones controles y estresados durante 4 días 5 x x
6 x x x
7 x x
8 x x x Sacrificar el grupo de ratones estresados durante 8 días
Obtención y procesamiento del material biológico.
Posterior al tratamiento, los animales se anestesiaron profundamente
con vapores de éter, se sangraron por punción cardiaca directa y se
sacrificaron por decapitación. Se procedió a realizar lavado nasal con 2 ml de
solución salina estéril, colocando un catéter en la tráquea, para realizar
infusión retrógrada y colectar el liquido de lavado nasal en tubos Eppendorff,
después se almacenó en congelación para cuantificar posteriormente la sIgA

7
(IgA secretora) mediante ELISA tipo emparedado. Por último se realizaron
curvas de concentración y cuantificación de proteínas por el método de
Bradford.
Para la obtención del NALT, después del lavado nasal se removió la piel
de la cabeza, se retiró el maxilar inferior y los tejidos blandos, como lo
describen (Asanuma y cols, 1997; Heritage y cols, 1998)
El paladar extraído se colocó en contenedores de aluminio de 1 cm³,
sobre una base de medio de inclusión hidrosoluble (Tissue-tek), con el NALT
en posición ventral, éste fue cubierto con una segunda capa de tissue-tek y se
congeló en isopentano. Las muestras fueron almacenadas a �70° C, hasta su
corte en criostato.
Después de extraído el NALT los cráneos fueron fijados por inmersión
en paraformaldehido al 4% por 24 horas. Después de lavado, se
descalcificaron con EDTA al 8% p.H 7.6, realizando un cambio diario durante 8
días. Al término se procesaron para su inclusión en parafina y se obtuvieron
cortes de 7 µm de espesor.
Procesamiento
De las muestras del NALT congeladas se obtuvieron cortes de 7 µm de
espesor, se colocaron en series de portaobjetos previamente tratados con
gelatina la 1% y se fijaron en acetona durante 20 minutos.
Tinción
En los cortes fijados en acetona se realizo la tinción inmunohistoquímica
para identificar por inmunohistoquímica directa, células plasmáticas
productoras de IgA utilizando anticuerpo policlonal de cabra anti-IgA de ratón
conjugado con HRP (Serotec).
Cuantificación de las células.
Las AFC-IgA+ en el NALT se contaron en una área de 30013µm2, en
los cortes de la porción media del tejido, mientras que se utilizó un área
constante de 25050 µm2, para contabilizar las AFC-IgA+ en la lámina propia.
Cuantificación de IgA en líquido nasal por ELISA
Se colocó en cada pozo 100 µg/ml de inmunoglobulinas de conejo anti-
IgA de mieloma de ratón y se incubo durante 18 h a 4° C; después de tres

8
lavados con amortiguador de fosfatos-tween 20 p.H 7.2 (PBS-T) se aplicaron
las muestras de líquido nasal sin diluir incubándose durante 2h a 37° C; al
término, las placas se lavaron 5 veces con PBS-T y 5 veces con PBS,
posteriormente se incubaron durante 2h a 37° C con el conjugado de cabra
anti-IgA de ratón (Serotec) diluido 1:1000 en PBS-T. Finalmente, las placas se
lavaron 3 veces con PBS-T y 3 veces con PBS. Posteriormente se adicionó el
sustrato (Ortofenilendiamina). Después de 15 minutos a temperatura ambiente,
la reacción se detuvo con ácido sulfúrico al 2.5 M se determinó la absorbancia
a una longitud de onda de 490 nm. La curva estándar se hizo utilizando una IgA
de mieloma de ratón (Sigma), el rango de concentración fue de 5 µg a 150 ng.
Aislamiento de RNA total. Cada muestra con un peso entre 50 y 70 mg, fue disuelta en un mL de Trizol reagent (invitrogen) por medio de homogenización de acuerdo a las instrucciones del proveedor. El Trizol fue removido con la adicción de 0.2 mL de cloroformo seguido de la precipitación de RNA total con isopropanol. Los precipitados fueron lavados con etanol al 75 %. La cantidad y la pureza de el RNA fue cuantificado espectrofotometricamente por las lecturas de densidad óptica a 260 y 280 nm de cada muestra; la integridad fue comprobada mediante electroforesis en gel de agarosa. El aislamiento del RNA total de las mucosas almacenadas de los diferentes grupos se basó en el método de guanidina de tiocinato (Chomczynski y Sacchi, 1987) con Tri reagent (Sigma). Como primer paso de este aislamiento fue la homogenización (50-70 mg de tejido/mL) de las muestras de intestino con un homogenizador marca Polytron. Se mantuvo el homogenizado por 5 min a temperatura ambiente para permitir la disociación de complejos de nucleoproteínas, posteriormente se agregó cloroformo 0.2 mL por 1 mL de Tri reagent, tras mezclarse vigorosamente por 15 seg se dejó 15 min a temperatura ambiente, centrifugándose a 12 000 g por 15 minutos a 4ºC. Se obtuvieron tres fases; superficial o fase acuosa (contiene RNA Total), una fase intermedia y una fase inferior orgánica (contiene proteínas); el paso siguiente fue transferir la fase acuosa a un tubo nuevo libre de DNAsas y RNAsas. La precipitación del RNA de la fase acuosa se hizo con isopropanol 0.5 mL /1 mL de Tri reagent usado en la homogenización inicial; ya añadido y mezclado cuidadosamente, el isopropanol se dejó interactuar 10 min y se centrifugó a 12 000 g por 10 min a 4ºC obteniéndose un sedimento blanco, que se lavó con 1 mL de etanol al 75% por cada 1 mL de Tri reagent con la subsiguiente centrifugación a 7500 g por 5 min a 4ºC. Se removió el etanol y se secó brevemente el sedimento (no totalmente para evitar insolubilidad), se disolvió el RNA con agua-DEPC (di-etil pirocarbonato), auxiliado con incubación a 55ºC por 5 min. Todas las muestras de RNA se incubaron con DNasa I (Roche) a una relación 10 U/1mg de RNA total a 37ºC por 30 min. Al término de la incubación se inactivó la enzima mediante 150 µL de fenol:cloroformo: alcohol isoamílico que se agregó a cada una de las muestras y se mezclo vigorosamente por 30 seg, al término de los cuáles se centrifugo la muestra por 2 min a 12000 g a 4ºC. El

9
paso siguiente fue transferir la fase acuosa a un tubo nuevo libre de DNAsas y RNAsas. A la fase acuosa se le agregó 1 mL de etanol al 100% a -20ºC se mezcló cuidadosamente y se incubó por 15 min a -20ºC. Después se centrifugaron las muestras por 2 min a 12000 g a 4ºC obteniéndose un sedimento que se lavó con 1 mL de etanol al 80%, se removió el etanol y se seco brevemente el sedimento y fue disuelto el RNA con agua tratada con DEPC y con ayuda de incubación en un baño a 55ºC, por 5 min. Se determinó la concentración y pureza del RNA por su densidad óptica a 260 nm y 280 nm en un espectrofotómetro Genesys y se guardó a -70 ºC hasta su uso. El RNA se cuantificó con la siguiente fórmula: µgRNA/mL= dilución X As260 X 40 La integridad del RNA se verificó por electroforesis horizontal en gel de agarosa al 0.5%- formaldehído-bromuro de etidio (1 µg/mL). Previo al corrimiento, las muestras (5 mL de RNA total de cada una de ellas) se incuban a 65ºC durante 10 min y posteriormente se colocaron en hielo a 4ºC y se les agregó 1 µL de colorante de corrimiento (formamida al 98%, EDTA 10mM, xilen-cianol al 0.1% y azul de bromofenol al 0.1%), las mezclas se colocaron en el gel de agarosa y formaldehído antes mencionado. El regulador del corrimiento consistió de MOPS 1X. Preparación de cDNA. La reverso transcripción se realizo con el kit �First-strand cDNA Synthesis� (Epicentre) utilizando 0.5 µg de la muestra de RNA total. El cDNA fue cuantificado espectrofotometricamente a una densidad óptica de 260 nm. PCR en tiempo real. El PCR en tiempo real fue usado para medir los niveles de mRNA animales. Nosotros emplemos el equipo Lightcycler y el kit �Lightcycler Taqman entre los grupos de Master� realizado siguiendo las instrucciones del proveedor Roche. mediante el uso de sondas predesarrolladas por el mismo proveedor (pIgR e IgA), tipo Taqman para expresión genética. La transcripción reversa y la reacción en cadena de la polimerasa se hizo en un solo paso con el �Kit Taqman One Step RT - PCR Master Mix Reagents� (Formado por: �Master Mix� sin UNG y �MultiScribe� mas �RNase Inhibitor Mix�), la reacción se llevó a cabo en un volumen final de 10 µL/pozo: 5 µL �Master Mix� sin UNG y 0.25 µL �MultiScribe� mas �RNase Inhibitor Mix�, 1 µL (50 ng de RNA total) RNA total (ajustado según rango dinámico, previamente obtenido), 3.25 µL de agua libre de RNasa y 0.5 µL de sonda de nuestros genes blanco (pIgR e IgA marcados con 6- carboxifluoresceína: FAN) o nuestro control endógeno 18 S (marcado con VIC) control interno (normalizador). Las placas se corrieron utilizando el termociclador en tiempo real ABI 7900, con el siguiente programa: 35 min a 48ºC para la reverso trascripción y 10 min a 95ºC para activar DNA polimerasa, la amplificación se realizó con 45 repeticiones de los dos pasos siguentes: 15 seg de desnaturalización a 95ºC y un paso de 1 min de alineación y extensión a 60ºC.
Análisis estadístico
Utilizando la prueba de ANOVA de una vía, se compararon los grupos
experimentales de cada protocolo con su respectivo testigo. Los datos se

10
analizaron mediante el software Sigma Stat y se graficaron mediante el
software Sigma Plot.
RESULTADOS
Cuantificacion de corticosterona sérica.
Linfocitos B IgA+ en el NALT
En los tres grupos de estudio, las células IgA+ mostraron un patrón de
distribución difuso en todo el NALT, no observándose zonas de concentración
para esta población (Figura 1-A y B).
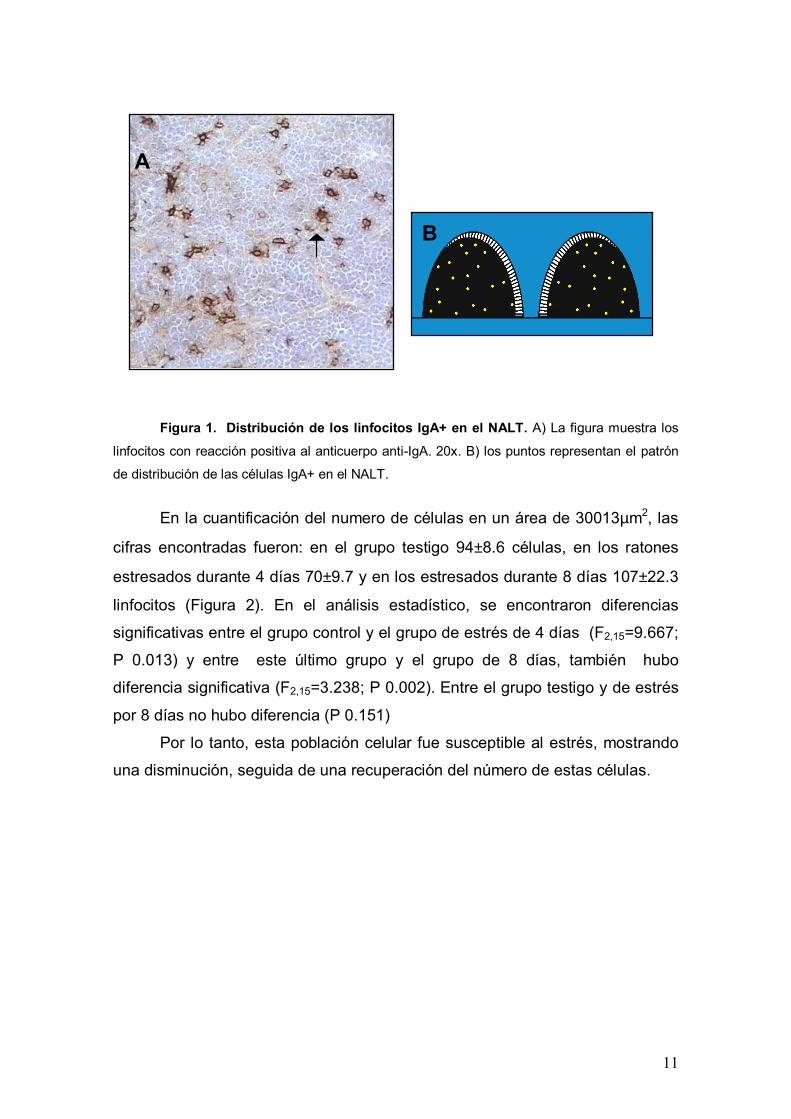
11
Figura 1. Distribución de los linfocitos IgA+ en el NALT. A) La figura muestra los
linfocitos con reacción positiva al anticuerpo anti-IgA. 20x. B) los puntos representan el patrón
de distribución de las células IgA+ en el NALT.
En la cuantificación del numero de células en un área de 30013µm2, las
cifras encontradas fueron: en el grupo testigo 94±8.6 células, en los ratones
estresados durante 4 días 70±9.7 y en los estresados durante 8 días 107±22.3
linfocitos (Figura 2). En el análisis estadístico, se encontraron diferencias
significativas entre el grupo control y el grupo de estrés de 4 días (F2,15=9.667;
P 0.013) y entre este último grupo y el grupo de 8 días, también hubo
diferencia significativa (F2,15=3.238; P 0.002). Entre el grupo testigo y de estrés
por 8 días no hubo diferencia (P 0.151)
Por lo tanto, esta población celular fue susceptible al estrés, mostrando
una disminución, seguida de una recuperación del número de estas células.
A
B

12
4DIAS
Num
ero
de li
nfoc
itos
B Ig
A+
en e
l NA
LT
0
20
40
60
80
100
120
140
160
CONTROL 8 DIAS
P 0.002P 0.013
Figura 2. Cantidad de linfocitos B IgA+ en el NALT. La gráfica compara la cantidad
de células entre los tres grupos de estudio. Con respecto al grupo control, el número de células
disminuyó significativamente con el estrés de 4 días (DF2,15=9.667; P 0.013) y se incremento
con el estrés de 8 días (DF2,15=3.238; P 0.002).
Células plasmáticas IgA+ en el tejido no NALT (lámina propia)
Las células plasmáticas IgA+ se observaron en la lámina propia
entre los acinos mucoserosos de la mucosa revestida por epitelio respiratorio,
predominantemente a nivel de los cornetes (Figura 3). En la mucosa del
tabique y de la región anterior de la nariz las células fueron escasas.

13
Figura 3. Células plasmáticas IgA+ en la lámina propia. Las células plasmáticas se
observan entre los acinos de las glándulas nasal (Flechas). 20X
El conteo de las células se realizó en el área de 25050 µm2 de los
cornetes, encontrando un promedio de 5.3±1.5 células en el grupo testigo, el
grupo de estrés de 4 días mostró 4.6±0.8 células, y el de 8 días 5±0.9 células
(Figura 4). El análisis estadístico no mostró diferencias significativas entre los
tres grupos de estudio (F2,15=0.536; P 0.596), pero aparentemente los animales
estresados durante 4 días mostraron tendencia hacia la disminución del
número de células plasmáticas IgA+.
4 DIAS
Cél
ulas
pla
smat
icas
IgA
+ en
lám
ina
prop
ia
0
2
4
6
8
10
8 DIASCONTROL
Figura 4. Cantidad de células plasmáticas en la lámina propia. La gráfica compara
los dos grupos problema con el testigo. El análisis estadístico, no mostró diferencias
significativas entre los tres grupos de estudio (DF2,15=0.536; P 0.596),
Efecto del estrés en los niveles de IgA+ secretora
En el análisis del inmunoensayo enzimatico (ELISA), los niveles
de sIgA en las muestras nasales del grupo testigo fue de 1.43.±0.10 µg/ml. Los
resultados mostraron una disminucion significativa en los niveles de sIgA con

14
el estrés de 4 días, con cifras de 1.27±0.11 µg /ml. (F2, 24=9.480; P <0.003), en
cambio el grupo tratado durante 8 días presentó cifras de 1.46±0.07 µg /ml, sin
mostrar diferencias significativas con el grupo control (P 0.451). (Figura 5)
Figura 5. Niveles de IgA secretora. La gráfica compara los dos grupos problema con
el testigo, cuantificando los niveles de sIgA en el lavado nasal por ELISA. En el análisis
estadístico, se observó una diferencia significativa entre el grupo control y el de estrés de 4
días (DF2, 24=9.480; P <0.003), así como los grupos problema (P 0.001).
Efecto del estrés por 8 dìas expresión relativa de mRNA para cadenas alfa de IgA y citocinas.
Los resultados de la determinación de a expresión relativa de los diferentes
mRNA en ratones estresados durante 8 días fueron los siguientes: IgA 6.67 +/-
0.57, IL-6 0.81 +/- 0.29, IL-2 2.56 +/- 0.28 e INFγ 4.19 +/- 1.27, IL-12 0.25 +/-
0.04. Por lo que podemos observar que la expresión de la IL-6 no se modifico,
mientras que la expresion de IL-12 se encuentran disminuidas en relación al
control (1.0). Por el contrario, la expresion de IFN-g, IL-2, (Figura 6), asi como
de lisozima, TNF-alfa, TLR-2 , TLR-3, IL-1 e IL-10 se encontro aumentada
(Figura 6 y Figura 7).
4 DIAS
% Ig
A ( µ
g/m
L)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
CONTROL 8 DIAS
P 0.003 P 0.001

15
0
2
4
6
8
10
Control IgA IL-6 IL-12 INF-g IL-2
Niv
eles
Rel
ativ
os d
e m
RN
A
Figura 6. Expresión relativa de mRNA para cadena alfa (IgA) y citocinas en ratones
estresados durante 7 días.
0
2
4
6
8
10
Control Lisoz TNF-a TLR-2 TLR-3 IL-1 IL-10
Niv
eles
Rel
ativ
os m
RN
A
Figura 6. Expresión relativa de mRNA para lisozima (Lisoz), TNF-alfa, TLR-2 , TLR-3,
IL-1 e IL-10 en ratones estresados durante 7 días.

16
DISCUSION
El estrés provocó cambios en el número linfocitos IgA+ en el NALT
El conteo de linfocitos IgA+ por área de tejido en el NALT, mostró una
disminución significativa con el estrés de 4 días, mientras que a los 8 días su
número fue mayor que en el grupo testigo. En la literatura no se encontraron
reportes previos de la cantidad de linfocitos B IgA+ en el NALT ni su relación
con el estrés; Sin embargo los resultados sugieren influencia de las hormonas
del estrés sobre la producción de las citocinas o en la expresión de moléculas
coestimuladoras relacionadas con el cambio de isotipo de IgM a IgA, pues se
sabe que dentro del MALT, en las estructuras que corresponden al
compartimiento inductor, placas de Peyer o en este caso el NALT, ocurre ese
cambio de isotipo, de células IgM+ a IgA+ debido a una transformación de la
cadena µ al isotipo α, regulada por citocinas como el TGF-β. Posteriormente las
células IgA+, migran hacia la lámina propia (porción efectora), en donde se
convierten en blastos IgA+ y células plasmáticas bajo la influencia de IL-5, IL6,
IL10 (Shikina T y cols 2004).
Aunque no se encontraron reportes de la es posible que la disminución en el
número de AFCs IgA en el NALT, se deba a modificaciones en la expresión de
estas moléculas lo cual a su vez modifique el cambio de isotipo de IgM a IgA.
Otra posible explicación es que las células IgA+ se redistribuyan en
otros sitios, como la lámina propia de la mucosa respiratoria, en sangre
periférica u otros órganos linfoides, aunque también se encontró que el
número de AFCs IgA no se modificó significativamente en la lámina propia a
nivel de los cornetes.
Otra explicación de la disminución de las células B IgA+, son los
cambios en la redistribución de las células B (IgM+). Dhabhar FS y cols
mencionan que el estrés agudo provocado durante la mañana ocasionó una
reducción de linfocitos B en sangre periférica (Dhabhar FS y cols 1994). En el
presente trabajo, los ciclos de estrés fueron aplicados en las primeras horas del
día, es posible que estos hayan originado una redistribución de las células IgA+
y con ello su disminución en el NALT, aunque en análisis morfológico no
mostró cambios en la cantidad de linfocitos IgM+.

17
La redistribución de las células linfoides en el MALT está regulada por la
expresión de moléculas de adhesión y quimiocinas. En la mucosa intestinal,
moléculas como TECK/CCL25, MEC/CCL28, MIP-3α/CCL20; y sus receptores
CCR9 y CCR6, pueden regular la migración de las células plasmáticas
productoras de IgA desde los tejidos u órganos linfoides, hacia los sitios de
inflamación y sitios efectores de la mucosa. Aunque no se encontraron
reportes, de cuales son los agentes quimiotacticos que posiblemente participan
en la localización de las AFCs IgA en el NALT, pero es probable que al igual
que en la placa de Peyer existan cambios en la expresión de estos agentes
quimiotacticos que pueden condicionar cambios en la redistribución de estas
células durante el estrés debido al efecto de las hormonas relacionadas con el
mismo. (Lazarus NH, y cols 2003, Kunkel EJ y Butcher EC 2002, Kunkel EJ y
cols 2003, Kunkel EJ y Butcher EC 2003).
Es posible que la alteración de las quimiocinas, así como de moléculas
de adhesión y sus receptores: MAdCAM-1/α4β7, PNAd/L-selectina y VCAM-
1/α4β1, las cuales se han vinculado con la localización de los linfocitos en el
NALT (Csencsits KL y cols 1999), se encuentren disminuidas debido al efecto
del estrés y sus hormonas, al igual de lo que sucede con VCAM-1, selectina-E,
e ICAM-1 las cuales disminuyen en las células endoteliales, después de la
aplicación de glucocorticoides sintéticos, en enfermedades inflamatorias
intestinales (Aziz KE y Wakefield D 1996, Mori N y cols 1999).
El estrés, no modificó la cantidad de células plasmáticas IgA+ en la lámina propia.
El número de células plasmáticas IgA+ en lámina propia se cuantificó a
nivel del tabique nasal, siendo ligeramente menor en los animales estresados,
aunque la diferencia no fue significativa y sugiriendo estar en relación con el
menor número de células IgA+ encontradas en el NALT, con lo cual la cantidad
de células que migran hacia la lámina propia es inferior a lo normal.
Sugiriendo que el mecanismo, seria otra vez la alteración en la expresión de
diversas moléculas, citocinas, quimiocinas y de sus receptores
correspondientes por el estrés y sus hormonas (glucocorticoides). Aunque se
menciona que la lámina propia, no es un sitio preferente de secuestro de

18
linfocitos durante el periodo de estrés, como lo muestran algunos trabajos
(Krynicki M y Olszewski WL 1989, Stefanski V y cols 2003), en donde se
encontró que los linfocitos marcados migran preferentemente hacia la medula
ósea y la piel durante el estrés. Sin embargo, el trafico de estas células hacia
órganos linfoides como el bazo, los nódulos linfoides mesentéricos y
periféricos, fue menor que en el grupo control.
El estrés modificó la cantidad de IgA secretora (sIgA) en el líquido nasal
Los resultados mostraron una disminución significativa de los niveles de
sIgA con el estrés de 4 días y un incremento no significativo en el grupo de
ratones estresados por 8 días. Estos resultados pueden relacionarse con los
cambios tempranos que ocurren durante el estrés agudo, cuya respuesta
revierte tales efectos (periodo de adaptación), ya que agotamiento ocurrido
durante el estrés crónico, provocaría un decremento en el número de células.
(Paca´k K y Palkovits M 2001). En la literatura no se encontraron reportes del
efecto del estrés o de sus hormonas sobre la secreción de sIgA en el moco
nasal, aunque algunos estudios sugieren una relación estrecha entre el estrés,
la inmunidad y la susceptibilidad a infecciones del tracto respiratorio alto
(Cohen S y cols 2001, 2002).
La disminución a los 4 días en la secreción de IgA, se correlaciono con
el menor número de células IgA+ localizadas en el NALT, en donde un factor
determinante, puede ser la sobreactivación del eje HPA durante el estrés. En
saliva por ejemplo, está reportado que durante los periodos de estrés agudo, se
incrementan los niveles de glucocorticoides (Bosch JA y cols 2002, Van Eck M
y cols 1996, Ng V y cols 2003), situación que de repetirse, ejercería efectos
inmunomoduladores sobre las células inmunitarias locales. Como
consecuencia, en el fluido nasal, estas hormonas afectarían el cambio de
isotipo hacia IgA y con ello la cantidad de células plasmáticas en la lámina
propia y la secreción de la inmunoglobulina.
También ha sido propuesta la alteración en el transporte de la sIgA
desde el intersticio glandular hacia la saliva en estrés crónico, lo cual ocasiona
incremento de la susceptibilidad a infecciones en el tracto respiratorio alto
(Graham NM y cols 1998, Jemmott y cols 1983, Phillips AC y cols 2006). Este

19
proceso explicaría la diferencia de células IgA+ en lámina propia y los niveles
de IgA en el fluido nasal. Así, probablemente la deficiencia en el transporte de
sIgA también ocurra a nivel de la mucosa nasal.
Por otro lado el leve incremento de la sIgA en los animales estresados
por 8 días, concuerda con otros reportes, donde el estrés agudo produce
elevación en los niveles de sIgA, además de otros cambios inmediatos en la
composición y la función de la saliva (Bosch JA y cols 1996, Ohira H 2004). En
el presente estudio, la producción de sIgA mostró diferencias en cuanto al
tiempo de aplicación del estrés, lo que se correlacionó con el numero de
linfocitos B IgA+ en el NALT. Por tanto, es posible que el tráfico de estas
células hacia el compartimiento efector, se haya modificado con el estrés.
El estres incremento la expresion del gene para cadenas alpha de la
IgA,, asi como de los genes para las citocinas producidas por celulas del
fneoetipo Th1; interferon gamma e IL-2. Por el contrario, redujo la expresion de
los genes de las citocinas IL12 e IL6.
CONCLUSIONES
1. Se comprueba la hipótesis de influencia del estrés sobre el tejido linfoide
del NALT.
2. Las células IgA+ disminuyeron con estrés por inmovilización de 4 días
así como la secreción de IgA en el fluido nasal, mismas que mostraron
un incremento con el estrés de 8 días, probablemente debido a una
adaptación.
3. El estres incremento la expresion del gene para la cadena alpha por lo
tanto la menor concentracion de IgA en el liquido nasal puede deberse a
menor sintesis de pIgR y por lo tanto del transporte de IgA dimerica..
4. Se sugiere que los efectos del estrés sobre las células productoras de
IgA así como en la secreción de la inmunoglobulina se ejercen a través
de la influencia de las hormonas, catecolaminas y glucocorticoides

20
BIBLIOGRAFIA
Asanuma H, Hodson AT, Iwasaki T, Sato Y, Inaba Y, Aizawa C, Kurata T,
Tamura S. Isolation and characterization of mouse nasal-associated
lymphoid tissue. J Immunol methods. 1997; 202: 123-131.
Aziz KE, Wakefield D. Modulation of endothelial cell expression of ICAM-1,
E-selectin, and VCAM-1 by beta-estradiol, progesterone, and
dexametasone. Cell Immunol 1996; 167: 79-85
Bosch JA, Brand HS, Ligtenberg TJ, Bermond B, Hoogstraten J, Nieuw
Amerongen AV. Psychological stress as a determinant of protein levels and

21
salivary-induced aggregation of Streptococcus gordonii in human whole
saliva. Psychosom Med. 1996; 58: 374-382.
Bosch JA, Ring C, de Geus EJ, Veerman EC, Amerongen AV. Stress and
secretory immunity. Int Rev Neurobiol. 2002; 52: 213-253.
Boyaka PN, Wright PF, Marinaro M, Kiyono H, Johnson JE, Gonzales RA,
Ikizler MR, Werkhaven JA, Jackson RJ, Fujihashi K, Di Fabio S, Staats HF,
and McGhee JR. Human nasopharyngeal-associated limphoreticular
tissues, functional analysis of subepithelial and intraepithelial B and T cells
from adenoids and tonsils. Am J Pathol. 2000; 157: 2023-2035.
Cohen S, Miller GE. Stress, Immunity, and susceptibility to upper respiratory
infection. Psychoneuroimmunology. 2001; 2: 7-18.
Cohen S, Hamrick N, Rodriguez MS, Feldman PJ, Rabin BS, Manuck SB.
Reactivity and vulnerability to stress-associated risk for upper respiratory
illness. Psychosom Med. 2002; 64: 302-310.
Csencsits KL, Jutila MA, Pascual DW. Nasal associated lymphoid tissue:
phenotypic and functional evidence for the primary role of peripheral node
addresin in naive lymphocyte adhesion to high endothelial venules in a
mucosal site. J Immunol. 1999; 163: 1382-1389.
Dhabhar FS, Miller AH, Stein M, Mcewen BS, Spencer RL. Diurnal and
acute stress-induced changes in the distribution of peripheral blood
leukocytes subpoblations. Brain Behav Immun. 1994; 8: 66-79.
Giannasca PJ, Boden JA, Monath TP. Target delivery of antigen to hamster
nasal lymphoid tissue with M-cell-directed lectins. Infec immun. 1997; 65:
4288-4298.

22
Graham NM, Bartholomeusz RC, Taboonpong N, La Brooy JT. Does anxiety
reduce the secretion rate of secretory IgA in saliva?. Med J Aust. 1998; 148:
131-133.
Heritage PL, Brook MA, Underdown BJ, McDermott. Intranasal immunization
with polymer-grafted microparticles activates the nasal-associated lymphoid
tissue and draining lymph nodes. Immunology. 1998; 93: 249-56.
Jemmott JB 3rd, Borysenko JZ, Borysenko M, McClelland DC, Chapman R,
Meyer D, Benson H. Academic stress, power motivation, and decrease in
secretion rate of salivary secretory immunoglobulin J. Lancet. 1983; 1: 1400-
1402.
Kiyono H, Fukuyama S. NALT-versus peyer�s-patch-mediated mucosal
immunity. Nat Rev Immunol. 2004; 4: 699-710.
Krynicki M, Olszewski WL. Influence of thermal stress on lymphocyte
migration pattern in rats. Arch Immunol Ther Exp. 1989; 37: 601-607.
Kunkel EJ, Butcher EC. Chemokines and the tissue-specific migration of
lymphocytes. Immunity. 2002; 16: 1-4.
Kunkel EJ, Campbell DJ, Butcher EC. Chemokines in lymphocyte trafficking
and intestinal immunity. Microcirculation. 2003; 10: 313-323.
Kunkel EJ, Butcher EC. Plasma-Cell homing. Nat Rev Immunol. 2003; 3:
822-829.
Lazarus NH, Kunkel EJ, Johnston B, Wilson E, Youngman KR, Butcher EC.
A common mucosal chemokine (mucosae-associated epithelial
chemokine/CCL28) selectively attracts IgA plasmablasts. J Immunol. 2003;
170: 3799-3805.

23
Mori N, Horie Y, Gerritsen ME, Anderson DC, Granger DN. Anti-
inflammatory drugs and endothelial cell adhesion molecule expression in
murine vascular beds. Gut. 1999; 44: 186-195.
Ng V, Koh D, Chia SE. Examination stress, salivary cortisol, and academic
performance. Psychol Rep. 2003; 93: 1133-1134.
Ohira H. Social support and salivary secretory immunoglobulin A response
in women to stress of making a public speech. Percept Most Skills. 2004;
98: 1241-1250.
Paca´k K. and Palkovits M. Stressor specificity of central neuroendocrine
responses: implications for stress-related disorders. Endocr Rev. 2001: 4;
502�548
Phillips AC, Caroll D, Evans P, Clow A, Hucklebridge F, Der G. Stressful life
events are associated with low secretion rates of immunoglobulin A in saliva
in the middle aged and elderly. Brain Behav Immunol. 2006; 20: 191-197.
Shikina T, Hiroi T, Iwatani K, Ho Jang M, Fukuyama S, Tamura M, Kubo T,
Ishikawa H, Kiyono H. IgA Class Switch ocurrs in the Organized
Nasopharynx, and Gut-Associated Lymphoid Tissue, but not in the diffuse
lámina propia of Airways and Gut. J Immunol. 2004; 172: 6259-6264.
Shikina T, Hiroi T, Iwatani K, Ho Jang M, Fukuyama S, Tamura M, Kubo T,
Ishikawa H, Kiyono H. IgA Class Switch ocurrs in the Organized
Nasopharynx, and Gut-Associated Lymphoid Tissue, but not in the diffuse
lámina propia of Airways and Gut. J Immunol. 2004; 172: 6259-6264.
Tristram GP, Stites DP, Terr AI, Imboden JB. Inmunología Básica y clínica.
10a ed. México: Manual moderno; 2001

24
Van Eck M, Berkof H, Nicholson N, Sulon J. The effects of perceived stress,
traits, mood states, and stressful daily events on salivary cortisol.
Psychosom med. 1996; 58: 447-458