nerva-mecanismo de adquisiciÓn de hierro...
TRANSCRIPT

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 1
NERVA-MECANISMO
DE ADQUISICIÓN DE HIERRO MINERAL
1. NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
1.1. Niveles de Hierro en el Cuerpo Humano
1.2. El Hierro en la Sangre
2. ASIMILACIÓN DE HIERRO MINERAL POR EL TORRENTE SANGUINEO
2.1. Hipótesis
2.2. Información
2.2.1. Introducción
2.2.2. Sistemas de los organismos superiores para limitar la concentración de
hierro libre
2.2.2.1. Las transferrinas
2.2.2.2. La ferritina
2.2.2.3. El grupo hemo
2.2.3. Mecanismos bacterianos de captación de hierro
2.2.3.1. Captación directa a de hierro
3. PROYECTO DE INVESTIGACIÓN
3.1. Determinación del contenido de hierro en el polvo de pirita
3.2. Determinación de ferritina, transferrina y hierro en muestras sanguíneas
3.2.1. Método turbidimétrico para la determinación de ferritina
3.2.2. Test inmunoturbidimétrico en suero
3.2.3. Prueba fotométrica colorimétrica para el hierro con factor aclarante de
lípidos (LCF)

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 2
1. NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
La Neisseria meningitidis, es una β proteobacteria, que causa meningitis bacteriana, una
infección del líquido cerebroespinal y las membranas de la médula espinal, con una alta
movilidad y transmisión y virulencia difícil de predecir, y por tanto de prevenir.
La práctica médica habitual asocia la transmisión de la enfermedad a contactos
personales en el espacio de un metro de la garganta del portador donde el microorganismo es
viable o activo. Sin embargo, la dificultad encontrada en limitar su contagio aplicando rutinas
de tratamiento al entorno cercano al afectado, y la diseminación espacial del microorganismo
y la enfermedad en los sujetos y en el ámbito geográfico, nos hacen cuestionar esta hipótesis
en busca de soluciones al problema de la transmisión de la enfermedad.
Efectivamente, la distribución en la tierra de la meningitis alcanza a todos los ámbitos
del planeta, algunas epidemias como la del África Subsahariana de una lado a otro del
continente, hacen difícil de creer en un único mecanismo de transmisión “persona a persona”.
Las epidemias de meningitis ocurren en zonas ambientalmente especiales, Molesworth et al
presentan evidencia de que la relación entre el medio ambiente y la localización de las
epidemias es cuantificable y proponer un modelo de predicción de áreas de riesgo para
epidemias de meningitis basado en variables ambientales (0).
En 1987 Caugant et al (1) demuestran la expansión intercontinental de Neisseria
Meningitidis (clones del complejo ET-S) en los brotes de la enfermedad de meningitis de
aquella temperada en Europa, Sudáfrica, Latinoamérica y EE.UU. Se describen brotes en el
Ártico (2) y en lugares tan aislados que obligan a plantear diversos medios de transmisión
para alcanzar tan diferentes localizaciones.
Pero es que en el mismo sujeto, se refieren las típicas invasiones de Neisseria
Meningitidis en el tramo intranasal (3) (4) y (5), y en todas las demás aberturas del cuerpo
humano…, oídos (6), vagina y ano (7), lo que sugiere una movilidad más allá de la referida
comúnmente.
La clasificación de seguridad del Center of Diseases Control de los Estados Unidos,
(CDC), para los trabajadores de laboratorio de análisis microbiológicos, establece un Nivel 2
para su correcto manejo, correspondiente a la alta virulencia de la Neisseria meningitidis, y a
su limitada movilidad, limitada, pero no nula movilidad (8).

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 3
Relacionados con la movilidad de la Neisseria meningitidis están los crecientes
problemas en la aviación civil transoceánica, vuelos de larga duración (9) y de los sistemas de
ventilación forzada y el polvo que generan (10). Esto lleva a estudiar la dispersión de la
bacteria en aerosoles (10) con todo tipo de otras especies microbiológicas, ligadas a partículas
de polvo de toda clase. El papel del polvo en la aparición de brotes epidémicos en el África
Subsahariana es especialmente interesante debido a la relación del aumento del mismo en las
sequías de los años 70 y 80 (del pasado siglo XX) y los brotes de la epidemia (0).
Griffis (11) estudia en un modelo epidemiológico “la lenta, silenciosa, fecal-oral,
transmisión de la meningitis…”, para concluir que la transmisión por aerosol del
meningococo determina la magnitud de una epidemia…
Neisseria meningitidis y gonorrhoeae han desarrollado un mecanismo perfectamente
adaptado a su vida dentro del organismo humano. Ambas especies pueden colonizar a su
huésped, sus mucosas, sin generar ninguna manifestación clínica detectable, convirtiéndose en
portadores, y permaneciendo así dentro de la población. Cuando se desarrolla la infección
pueden sobrevivir a las respuestas inflamatorias del anfitrión (12).
A los estudios citados se le suman los que demuestran que el número de portadores de la
bacteria que no han desarrollado la enfermedad, no tiene relación estadística alguna con el
número de afectados y la morbilidad en los brotes de la enfermedad (13). Can et al. (14)
“muestran en el análisis de un brote en una escuela primaria en Londres que una prolongada
secuencia de casos de infección meningocócica puede ocurrir con una tasas bajas de
transmisión y de portadores, en una población sin pérdida desproporcionada de inmunidad
específica” (cita textual traducida), sugiriendo un episodio de colonización prolongada, con
una transmisión infrecuente del organismo niño a niño en la escuela. Todavía más complicado
de resolver, y sorprendente, es la verificación realizada por Kyungcheol Ti et al. (15) de que
la reducción de la flora normal de la cavidad nasofaríngea por el tratamiento con antibióticos
no afecta a la colonización de la misma por la Neisseria meningitidis.
En los brotes de meningitis relacionados en el norte de Grecia a principios del siglo
XXI, (16) se observa que el número de portadores en la población autóctona es grande, que la
relación de los mismos no aumenta debido a los fuertes flujos migratorios de los países
vecinos, y que no hay entre ellos crecimiento de portadores de las cepas responsables del
incremento de los afectados en la zona por la enfermedad.
Todo este conocimiento nos obliga a estudiar otras vías de transmisión de la
enfermedad, además de las ya citadas de persona a persona.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 4
Para ello lo primero que hacemos es estudiar las características de estos
microorganismos, su microbiología, bioquímica, sus nichos ecológicos, en definitiva su modo
de vida para así CONOCERLOS, de acuerdo a la descripción de Microbiology, Prescott,
Harley, Klein (17).
De acuerdo a la descripción de Microbiology Prescott, Harley, Klein, (17) las Beta
Proteobacteria:
1. Se solapan metabólicamente con las α-proteobacteria, pero suelen usar sustancias que
provienen de la descomposición de materia orgánica en zonas anaerobias.
2. Considerable diversidad metabólica, dependiendo de los diferentes órdenes a los que
pertenezcan los microorganismos.
3. Orden Neisseriales.
a. Los miembros de este orden son no móviles, aerobios, cocos gram-negativos,
que en general aparecen en parejas con las paredes adyacentes rebajadas.
b. Pueden tener cápsulas y pilosidades.
c. Son quimiorganotróficos, oxidasas positivos, y casi siempre catalasas
positivos.
La Neisseria gonorrheae, causa la sífilis y la Neisseria meningitidis es uno de los
agentes causantes de la meningitis.
Puede deducirse que las Neisserias se adhieren a superficies bien irrigadas de sangre y
húmedas, y en las mucosas que protegen los accesos exteriores al cuerpo humano, vagina,
nariz y faringe. Ya hemos visto que también entran por oído (6) y ano (7). Se describen
meningitis en primates, pero no en otras especies animales (18).
La respiración aerobia de la Neisseria, y su posibilidad de alimentarse con materia
orgánica en descomposición anaerobia nos lleva a estudiar sus mecanismos de respiración
ligados a los equilibrios óxido-reducción de diferentes especies químicas presentes en el
medio.
Aparecen desde hace tiempo estudios de aumento de afectados por meningitis ligados a
aumentos de cantidad de hierro en el medio, tanto debido a bacterias (19) como a hongos (20).
El equilibrio redox del Fe permite la “respiración” aerobia de las Neisserias en zonas
anaerobias o de poco oxígeno y así el desarrollo de la enfermedad ligado al metabolismo del
hierro.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 5
La descripción de la dependencia de la Neisseria Meningitidis respecto del hierro es
conocida desde hace años (21), (22), (23). A su vez también se describe profundamente el
aumento de la virulencia en diferentes cepas y una mayor capacidad de infección de las
mismas relacionado con mayores cantidades de hierro en el medio (22), (24), (25).
Por ello los huéspedes desarrollan un completo sistema de contención del hierro frente
a la invasión (18) microbiana, posicionando proteínas de una gran capacidad de enlace con el
metal, para secuestrarlos en sitios estratégicos para impedir dicha invasión:
a. Disminuyendo los niveles de hierro en los fluidos corporales (sangre), tejidos
enfermos y células invadidas.
b. Sintetizando inmunoglobulinas para conseguir la adquisición de antígenos de los
microbios.
Los mecanismos bioquímicos responsables de la adquisición de hierro y de la virulencia
y capacidad de infección están siendo descritos en trabajos recientes (26), (27) y (28).
Pero la especial virulencia de esta enfermedad, y la dificultad de desentrañar los
mecanismos de su transmisión están ligados a una microbiología absolutamente adaptada de
la Neisseria meningitidis, a la invasión de su huésped y a protegerse de su sistema inmunitario
(12).
Dos características diferenciales de la Neisseria meningitidis son claramente esenciales
para la invasión del huésped:
a. La composición del polisacárido capsular de la membrana que permite su
supervivencia en fluidos extracelulares y
b. Tipo IV Pili (TFP), que son estructuras multiméricas esenciales para la adhesión de
las bacterias capsuladas virulantes a las células del huésped (29) en un mecanismo
de modificaciones cito esqueléticas con formación de profusiones asociadas con la
entrada de la Neisseria meningitidis dentro de las células endoteliales (30), (31)
Esta capacidad de modificación superficial ofrece un mecanismo adaptativo formidable
para modular el tropismo de los tejidos, la superación de las respuestas inmunes del huésped,
y la supervivencia en un ambiente cambiante y hostil en la célula invadida (12) (31).

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 6
Cada fibra de Pili se compone de una pequeña subunidad de proteína de pili (pil E) (32)
que es ensamblada dentro una estructura α helicoidal para formar el citado pili (33). Además
de los pilis se describen otros dispositivos que favorecen la adherencia de la Neisseria a las
células del anfitrión (12), (29).
Los mecanismos de reconocimiento de las células a invadir por los pilis están ligados a
una proteína de la membrana celular del huésped, la CD46, envuelta en la regulación de la
activación de su sistema complementario de defensa frente a infecciones. Es un paso crítico
en el establecimiento de la enfermedad en el huésped, promoviendo el paso a las meninges de
las bacterias invasoras (34).
Para estudiar la influencia del hierro en la meningitis, la Neisseria, la virulencia de las
cepas y la infección, seguiremos el siguiente esquema:
1. Química del hierro. Niveles del hierro en el cuerpo humano.
2. El hierro en la sangre
2.1. Desde el hierro libre al hierro ligado a proteínas de la sangre.
3. Adquisición de hierro en el huésped por Microorganismos.
3.1. Mecanismos de adquisición por Neisseria meningitidis
4. Relación de los niveles en Neisseria meningitidis, desarrollo de la enfermedad de
adquisición de hierro mineral (partículas de calcineros de pirita) en la sangre.
5. ¿Cómo resolver el puzzle de la interacción Neisseria meningitidis-células del huésped?
5.1. Interfase mineral/microorganismo.
5.2. La situación en Nerva
6. Propuesta del Nerva Mecanismo
7. Trabajo experimental. Confirmación del Nerva Mecanismo
7.1. Material y métodos
7.2. Resultados y discusión
7.3. Conclusiones
8. Transmisión de la enfermedad en Nerva
8.1. Ruta principal
8.2. Factores de riesgo directo relacionados con el hierro. Atmósfera, agua, suelo.
8.3. Factores de riesgo añadido. Afección al sistema respiratorio.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 7
9. Organismos causantes de meningitis.
9.1. Bacterias
9.1.1. Neisseria meningitidis. Principales serogrupos
9.2. Vírus
9.3. Otros
9.3.1. Meningitis amebiana. Neigleria fowleri.
10. Consecuencias. Propuestas de Actuación
10.1. Actuaciones Sanitarias
10.1.1. Nivel General
10.1.2. Actuaciones en Nerva
10.2. Actuaciones Ambientales
10.2.1. Nivel General
10.2.2. Actuaciones en Nerva
10.2.2.1. Comunidad Autónoma
10.2.2.2. Local
10.2.2.3. Familiar
10.2.2.4. Personal
10.3. Actuaciones de I+D y Universidad
10.3.1. Ingeniería de la Sanidad Ambiental.
1.1. NIVELES DE HIERRO EN EL CUERPO HUMANO
El hierro tiene dos formas oxidadas: Fe2+ y Fe3+, además de la capacidad de formar
compuestos complejos, en una configuración octahédrica, con 6 moléculas que actúan como
ligandos. Las proteínas de la sangre transferrín, ferritinas, etc, son estructuras de este tipo de
complejos de hierro.
Los cationes Fe2+ y Fe3+ precipitan como hidróxidos en medio básico, mientras a pH
ácidos estarían disueltos, Fe2+ y Fe3+ como hierro libre. La rápida oxidación de Fe2+ a Fe3+ en
ambientes aerobios, hace que su existencia sea mayor en ambientes reductores. Por otra parte
el Fe3+ tiene tendencia a precipitar como hidróxido en medios no ácidos.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 8
Estas dos consideraciones llevan a que en nuestro entorno cercano, aerobio y a pH
próximo a la neutralidad, el hierro aparece como Fe(OH)3 que al estar precipitado , no es de
fácil acceso para los organismos vivos que lo necesitan, aunque en pequeñas cantidades. Así
se han desarrollado microorganismos de todo tipo que consiguen la energía en los equilibrios
redox del hierro, y lo hacen biodisponible introduciéndolo en la materia viva.
El cuerpo humano contiene entre 3 y 5 gramos de hierro, (en las mujeres un poco menos
cantidad en general), y de estos, dos terceras partes en células rojas del sistema circulatorio,
como hemoglobina y un 15-25% se almacena como ferritina y homosiderina. El hierro
restante va en los músculos como mioglobina (8%) en los citocromos y en enzimas que lo
llevan como hierro complejado en su molécula.
El hierro presente en el transferrín de la sangre solo asciende a 3 mg, pero el
intercambio diario a su través es diez veces mayor. El papel del transferrín es fundamental en
la distribución del hierro en el organismo humano, por su capacidad de tomar/dejar hierro del
entorno (35).
1.2. EL HIERRO EN LA SANGRE
El equilibrio del hierro es tan sutil que su transporte en el cuerpo humano a través de la
sangre está totalmente regulado, para evitar mínimas cantidades de falta o exceso, ambas
situaciones con dramáticos efectos en el sujeto.
El control de los niveles de hierro en sangre se realiza por diferentes proteínas, que lo
complejan a partir del hierro libre, tanto para la realización de sus funciones celulares, como
para dificultar su acceso a potenciales microorganismos invasores.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 9
La entrada de hierro en las células, puede esquematizarse así (36):
PROTEINAS DE HIERRO EN LA SANGRE:
TRANSFERRIN: (o transferrina) (37)
El hígado segrega una proteína, la betaglobulina, apotransferrina, que tiene una elevada
afinidad por el Fe3+, con el que forma el hierro-transferrín, la transferrina.
Las transferrinas varían desde la serotransferrina, que se encuentra en el plasma
sanguíneo, la lactotransferrina, que se encuentra en la mayoría de los fluidos extracelulares,
como la leche, lágrimas, saliva, o fluidos gastrointestinales, a la ovatransferrina, presente en
huevos.
Las transferrinas llevan a cabo diferentes funciones, mientras la serotransferrina
transporta el hierro por la sangre, la ovotransferrina y la lactotransferrina son complejantes del
hierro, por lo que actúan como agentes antimicrobianos al disminuir su disponibilidad.
En la web TRANSFERRINS (38) se encuentra una descripción de las moléculas,
configuraciones, secuencias y diferentes tipos que aparecen en distintas especies vivas.
Fe
En la Dieta
En Sangre
TRANSFERRINA
FERRITINA
Otras FERROPROTEINAS (incluyendo Grupo
Hemo)
FUENTES POTENCIALES DE HIERRO PARA PATÓGENOS

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 10
FERRITINA (37)
Se encarga del almacenamiento del hierro en el interior de las células, de manera que se
encuentre disponible cuando el metabolismo celular lo necesite. En su acopio evita la
posibilidad, ligada al Fe3+ en el medio, de formación de radicales libres en el medio.
Se han encontrado funciones de las ferritinas distintas de las conocidas de acumulación,
y así aparecen en organismos unicelulares, y en células pobres en hierro como las secretoras
de insulina. Posiblemente sean reguladoras de la síntesis de proteínas, y en insectos y
moluscos pueden funcionar como moléculas trasportadoras de hierro. También se ha descrito
su papel en la regulación de la proliferación celular (35).
EL GRUPO HEMO (36)
Las hemoproteínas portan la mayor cantidad de hierro en el organismo:
• Hemoglobina, la más importante, encargada del transporte del oxígeno, puede llegar a
tener hasta el 65% del hierro total del cuerpo humano.
• Los citocromos. Se encuentran en las mitocondrias y en el retículo endoplasmático,
participan en las reacciones redox, de transferencia de energía.
• Las catalasas y peroxidasas. Son las enzimas encargadas de la protección celular frente
a oxidantes enérgicos, como el peróxido de hidrógeno, formador de radicales libres,
tóxicos para la mayoría de las células.
MECANISMOS DE ADQUISIÓN/CESIÓN DE HIERRO POR TRANSFERRÍN
La capacidad demostrada del transferrín para adquirir/ceder hierro al medio, le confiere
un papel clave en la relación huésped-microorganismo invasor, tanto por la posibilidad de
retirarle el hierro, adquiriéndolo del medio conjuntamente con un anión de carbonato, a pH
neutro, y por su capacidad para cederlo al medio a pH ácido, y así hacerlo biodisponible al
invasor.
El transferrín humano (serum-transferrín) presenta una estructura con dos dominios
bilobulares, en una doble conformación abierta/cerrada (38). Los lóbulos están unidos por un
enlace peptídico, y se subdividen en dos dominios del mismo tamaño.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 11
El equilibrio de adquisición/cesión de hierro donde cada transferrín puede complejar
dos moléculas de Fe3+ puede esquematizarse así:
Apo-Trf (Apotransferrín) + Fe3+ ↔ Fe3+.Trf (Hierro.transferrín)
(Sin hierro) (Nolotransferrin)
Conformación abierta Conformación cerrada
La orientación de los lóbulos, dominios y los ejes de rotación son diferentes para
cada tipo de transferrín. En los tres tipos de transferrín la estructura se estabiliza por enlaces
disulfuro, de los que hasta ahora se dispone de información limitada, pero parecen ser
fundamentales en la estabilidad del complejo de hierro.
Para cargar el Fe3+ al apotransferrín se utiliza un quelato férrico, usualmente nitrilo
acetato férrico, NTA.Fe3+
Apo.Trf + NTA.Fe3+ ↔ Trf.Fe3+.NTA
que en presencia de exceso de bicarbonato, el NTA es reemplazado por CO3= , en un
equilibrio reversible, que facilita el paso de hierro del medio al trasferrín y viceversa.
Trf.Fe3+.NTA + CO3= ↔ Trf.Fe3+.CO3
= + NTA
Minzani et al (39) demuestra que la estructura conformacional abierta del
apotransferrín y un intermedio de hierro-absorbido son similares, y casi indistinguibles.
Después de la entrada del CO3=, aparece un estado intermedio inestable, abierto,
Trf.Fe3+CO3=, para acabar cerrándose a una estructura estable Trf.Fe3+CO3
=. Hirose (38)
concluye que es el intermedio con el hierro absorbido el paso central que permite el equilibrio
de entrada/salida del metal a la proteína.
Baker et al. (40) concluye que existe una serie de ligandos muy favorables para
complejar Fe3+ con la inclusión de un anión externo, con la contrapartida de ofrecer la
posibilidad de comenzar los procesos de cesión al medio, confirmando los primeros estudios
de Aisen et al (41)

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 12
La influencia del pH en estas reacciones ofrece las claves de la entrada del hierro
en la sangre, facilitando el salto de la Neisseria Meningitidis a su interior, al hacerlo
biodisponible para los microorganismos patógenos, dando lugar a la infección consiguiente
(34).
Lee el al. (42) demuestran la evidencia de que por cada Fe3+ que entra en el
Apotransferrín se pierden tres H+.
ApoTrf + Fe3+ ↔ Fe.Trf + 3H+
Luego el equilibrio está desplazado hacia la salida del hierro en medio ácido, y hacia su
entrada en el apotransferrín en medios menos ácidos. De acuerdo a Hirose (38) tiene su
óptimo a pHs en el entorno de la neutralidad, pH=7. Aunque puede variar si el anión de
cambio es diferente al carbonato (43). Egan et al (44) refieren la recogida de hierro en el
endosoma de la célula a pH = 5.5.
NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 13
2. ASIMILACIÓN DE HIERRO MINERAL POR EL TORRENTE SANGUINEO
2.1. HIPÓTESIS
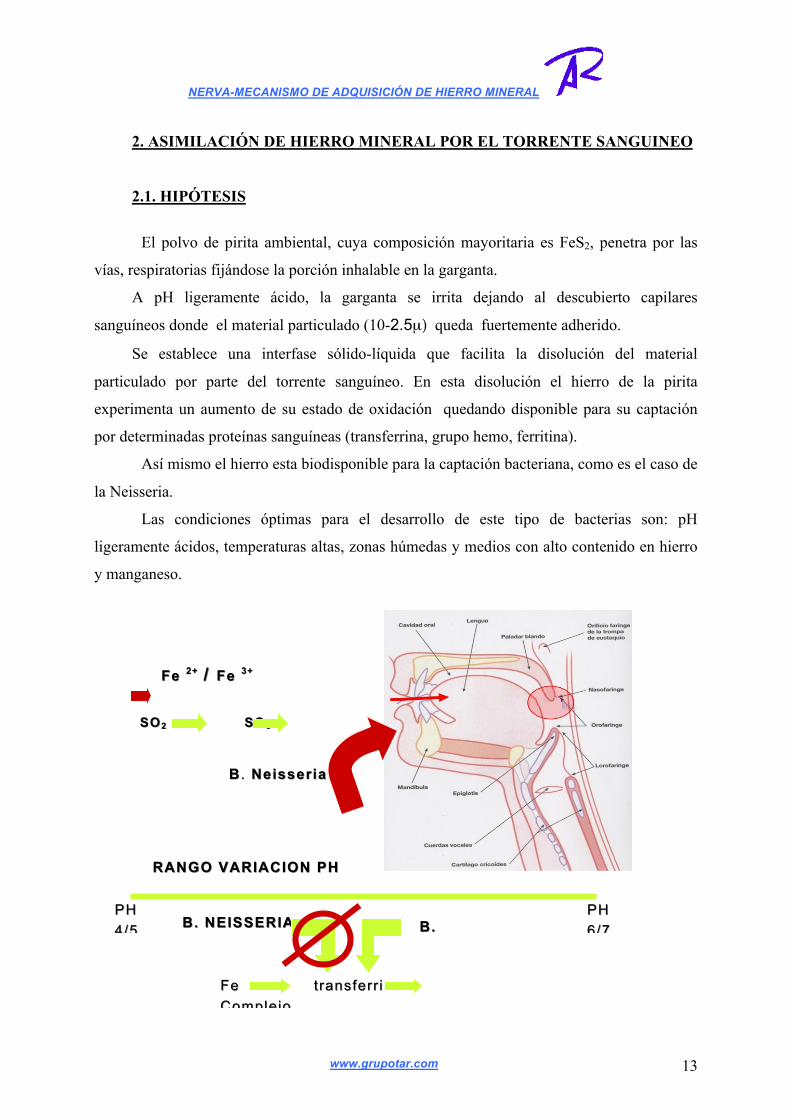
El polvo de pirita ambiental, cuya composición mayoritaria es FeS2, penetra por las
vías, respiratorias fijándose la porción inhalable en la garganta.
A pH ligeramente ácido, la garganta se irrita dejando al descubierto capilares
sanguíneos donde el material particulado (10-2.5µ) queda fuertemente adherido.
Se establece una interfase sólido-líquida que facilita la disolución del material
particulado por parte del torrente sanguíneo. En esta disolución el hierro de la pirita
experimenta un aumento de su estado de oxidación quedando disponible para su captación
por determinadas proteínas sanguíneas (transferrina, grupo hemo, ferritina).
Así mismo el hierro esta biodisponible para la captación bacteriana, como es el caso de
la Neisseria.
Las condiciones óptimas para el desarrollo de este tipo de bacterias son: pH
ligeramente ácidos, temperaturas altas, zonas húmedas y medios con alto contenido en hierro
y manganeso.
Fe Fe 2+2+ / / Fe Fe 3+3+
SOSO 2222 SOSO 3 3 HH 22SOSO 44
BB . . NeisseriaNeisseria
RANGO VARIACION PHRANGO VARIACION PH
PH PH 4/54/5
PH PH 6/76/7
Fe Fe transferr ina transferr ina Complejo Complejo FeFe --TransTrans
B. NEISSERIAB. NEISSERIA B. B. NEISSERIANEISSERIA

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 14
2.2. INFORMACIÓN
2.2.1. Introducción
Los cationes metálicos como el Fe 2+, Zn 2+, Cu 2+, Mn 2+, Co 2+ 0 Cd 2+, son
oligoelementos esenciales para el crecimiento de todos los organismos. Muchos de ellos
participan como cofactores en procesos enzimáticos o forman parte de estructuras anatómicas
celulares. Sin embargo, una elevada concentración de estos elementos puede tener efectos
deletéreos o incluso letales sobre las células, ya sea por inhibición de ciertos procesos
metabólicos (como por ejemplo la respiración anaeróbica) o por acumulación de radicales
libres generados en los procesos de reducción en los que participan. Por ello es fundamental
que las células no sobrepasen la concentración intracelular a partir de la cual un determinado
catión metálico resulta ser tóxico. Con esta finalidad, los organismos han ido desarrollando
mecanismos específicos de captación así como sistemas de excreción. Obviamente, todos
estos procesos deben estar perfectamente coordinados entre sí, y con las condiciones del
entorno, por lo que, no sólo implican proteínas que realizan funciones concretas, sino también
reguladores que controlan la expresión de éstas según las necesidades celulares.
Por otra parte, y desde la óptica de los organismos superiores, la necesidad de la
adquisición de dichos cationes por parte de los microorganismos puede ser utilizada como
mecanismo de defensa inespecífico frente al desarrollo de procesos infectivos.
Nos centraremos en tres de estos elementos esenciales: el hierro, el zinc y el manganeso,
que son, en este orden y hasta el momento, los más estudiados y de los que se dispone más
información sobre los sistemas relacionados con su transporte, acumulación y regulación.
2.2.2. Sistemas de los organismos superiores para limitar la concentración de
hierro libre
De todos los cationes metálicos mencionados, es con toda seguridad el metabolismo del
hierro el que más se conoce, tanto en lo que hace referencia a sus aspectos bioquímicos, como
a su transporte y almacenamiento.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 15
De hecho, el hierro es el elemento más controlado dentro del huésped, prácticamente no
se halla en forma libre, gracias a la existencia de una serie de moléculas encargadas de
retenerlo, tanto para facilitar sus diferentes funciones celulares en el organismo como para
dificultar que los microorganismos dispongan de él.
La cantidad de hierro total presente en el organismo se encuentra mayoritariamente
formando parte del grupo hemo, dando lugar a hemoproteínas como la hemoglobina (que
representaría un (65% del hierro total), la mioglobina (que representaría un 4%) diversos
compuestos que controlarían los procesos de oxidación intracelular alrededor del 0.1% se
encontraría en forma de transferrina en el plasma sanguíneo y del orden del 15 al 30% se
hallaría almacenado en forma, principalmente, de ferritina o a veces también como
hemosiderina, en las células reticuloendoteliales y en los hepatocitos.
A continuación se describirán brevemente las principales características de dichas
proteínas.
2.2.2.1. Las transferrinas
Los organismos superiores incorporan el hierro a través de la dieta. Este elemento se
absorbe a su paso por el intestino delgado. Básicamente, el hígado secreta, en la bilis,
cantidades moderadas de una globulina beta denominada apotransferrina que presenta una
elevada afinidad por este ión.
La combinación de ambos da lugar a la transferrina, que es reconocida por receptores
específicos de las células de la mucosa intestinal, siendo trasvasada al torrente sanguíneo.
Las transferrinas son un grupo ubicuo de proteínas, presentes en todos los vertebrados, y
que presentan como característica esencial una elevada afinidad por Fe 3+ .Existen distintas
variantes, todas con estructura análoga, como por ejemplo la serotransferrina (que se
encuentra en el plasma), la lactoferrina lactotransferrina (que se halla en la mayoría de los
fluidos extracelulares como la leche, las lágrimas, saliva o fluidos gastrointestinales) o la
ovotransferrina o conalbúmina (presente en los huevos) Las diversas transferrinas pueden
llevar a cabo diferentes funciones. Así, mientras la serotranferrina se encarga básicamente del
transporte de Fe3+ vía plasma,la ovotransferrina y la lactotransferrina son quelantes de hierro
actuando al disminuir la disponibilidad de hierro libre, como agentes antimicrobianos. La cesión del hierro por parte de las transferrinas es un proceso dependiente de pH
y de la presencia de receptores específicos.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 16
Sin embargo, la característica más destacable de este grupo de proteínas es que, a
pesar de presentar una gran afinidad por el hierro, prácticamente nunca se encuentran
saturadas por este metal. De hecho, en las moléculas de transferrina tan solo un tercio de los
posibles puntos de unión a Fe 3+ de las transferrinas que circulan por sangre está ocupado.
Esta característica permite que, si existe un aumento repentino de la concentración de hierro
libre en el medio, éste pueda ser captado rápidamente, manteniéndose así siempre una
concentración de hierro libre muy escasa. Este mecanismo de secuestro juega un papel
importante dentro de los mecanismos de defensa del huésped frente a posibles infecciones. De
hecho, se sabe que el suministro de hierro a pacientes con infecciones puede empeorar su
patología. De acuerdo con ello, estudios recientes han demostrado que la inoculación de
hierro exógeno, potencia las infecciones experimentales de mieroorganismos como
Aeromonas spp., Clostridium spp., Corynebacterium spp., Escherichia col¡, Klebsiella spp.,
Salmonella spp., Listeria spp., Neisseria spp., Pasteurella spp., Pseudomonas spp.,
Staphylocoocus spp., Vibrio, spp., Yérsinia spp., Mycobacterium avium y Mycobacterium
tuberculosis (De Voss et al. (1999), Achenbach et al. (1997)
2.2.2.2. La ferritina
La ferritina se encarga del almacenamiento del hierro e, igual que pasaba con la
transferrina, es una proteína presente en muchos organismos (animales, plantas, hongos y
bacterias). La ferritina almacena hierro en el interior de las células, de forma que éste se
encuentra disponible, en forma soluble, para usarse cuando el metabolismo celular lo requiera.
Pero además la ferritina también tiene un efecto protector sobre las células, pues almacena el
hierro en una forma inocua evitando que tome partido en reacciones que podrían dar lugar a la
formación de radicales libres.
2.2.2.3. El grupo hemo
La mayor cantidad del hierro presente en el organismo está asociado al grupo prostético
hemo, que forma parte de las denominadas hemoproteínas. Entre ellas se encuentran:
La hemoglobina, que es la más importante y abundante de todas ellas.
Encargada del transporte de oxígeno, y puede llegar a contener un 65% del hierro total del organismo.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 17
Los citocromos (citocromo c oxidasa, b/b6, b5, c554, f, cd, etc.), que se encuentran en las mitocondrias y en el retículo endoplasmático, y que participan en las reacciones de transferencia de electrones.
Las catalasas y peroxidasas, que son enzimas encargadas de la protección
celular frente a las especies químicas potencialmente dañinas generadas en las reacciones de oxidación en las que participa, entre otros sustratos, el peróxido de hidrógeno.
Desde un punto de vista químico, el grupo hemo forma parte del conjunto de
compuestos que recibe el nombre de porfirinas.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 18
2.2.3. Mecanismos bacterianos de captación de hierro
A pH neutro la concentración de Fe 3+ viene marcada por que la concentración de
Fe 3+ en solución sea extremadamente baja. Se estima que la concentración mínima de
hierro para el crecimiento bacteriano, óptimo es de, en general, 1 mM . Por lo tanto, la
escasa disponibilidad de hierro hace que este elemento actúe como factor limitante para
el desarrollo de la mayoría de microorganismos. Debido a su importancia en el
metabolismo, los microorganismos han desarrollado una serie de mecanismos para la
captación eficiente de dicho metal. En especial los microorganismos patógenos, ya que
para ellos, la escasez de hierro en el medio se ve acentuada por la presencia, como
hemos citado anteriormente, de proteínas específicas sintetizadas por el huésped
encargadas de la adquisición, transporte y almacenamiento de dicho elemento. De esta
forma se establece una competencia entre los mecanismos del patógeno y los del
huésped.
2.2.3.1. Captación directa a de hierro
Algunos microorganismos anaeróbicos, como Clostridium perfrigens, son capaces
de captar hierro creando a su alrededor un mieroambiente de pH más ácido. En este
ambiente reductor, se favorece la aparición de especies químicas más solubles,
formadas por Fe2+ además de facilitar la liberación de este metal por parte de proteínas
del huésped como la transferrina o la lactoferrina (Neilands 1995).
3. PROYECTO DE INVESTIGACIÓN
3.1. Determinación del contenido de hierro en el polvo de pirita
Las muestras recepcionadas en laboratorio procedían de la cantera próxima fueron
sometidas a análisis cualitativo y cuantitativo con el fin de identificar la presencia de
hierro y la concentración del mismo.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 19
Análisis cualitativo.
Tomamos 0.025g de polvo de pirita que fue previamente triturado hasta un
tamaño del orden 10-0.25µ. Se procedió a la disolución de la muestra en presencia de
ác. Sulfúrico concentrado. Con ello se oxidó el hierro de la pirita a ión férrico. Para la
identificación del ión férrico empleamos el ensayo del tiocianato potásico, que forma
un complejo rojo (que se asemeja al vino tinto) con dicho ión.
Análisis cuantitativo.
Partimos de dos muestras diferentes de polvo de pirita, una de aspecto anaranjado
y otra marrón-rojizo. Se prepararon muestras de 0.025g de cada una de ellas por
triplicado, que fueron tratadas con 11µL de ácido sulfúrico concentrado. Llevándose
finalmente hasta un volumen 50 µL. Empleamos un espectrofotómetro UV-V para la
determinación de la concentración de hierro en las muestras.
Para ello se preparo patrones de concentración 1, 5, 15, 25, 30 ppm a partir de una
disolución de hierro comercial en ácido nítrico hexahidratado. Para la formación de
complejo se añadió 1 µL de KSCN concentrado.
3.2. Determinación de ferritina, transferrina y hierro en muestras sanguíneas
3.2.1. Método turbidimétrico para la determinación de ferritina
Principio del método
El método está basado en la reacción de la ferritina de la muestra con unas
párticulas de látex a las que se ha ligado anticuerpos anti-ferritina. LA determinación es
turbidimétrica y puede ser medida en un analizador gracias a la relación entre la
absorbancia y concentración permite una calibración con un rango de medición entre 0
y 500µg/l.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 20
Procedimiento.
Se tomaron las muestras y se separaron en distinto viales para adecuar cada una a
unas condiciones determinadas:
• Muestra de suero sanguíneo con heparina. • Muestra de suero sanguíneo con heparina y pirita • Muestra de suero sanguíneo con heparina, pirita y ácido sulfúrico. • Heparina y pirita.
Esta última como referencia ya que en algunos casos se ha podido constatar la
absorbancia del anticoagulante del reactivo.
Se preparó un blanco con 300µL de patrón y 200µL de reactivo látex, un Standard
con 300µL de tampón 200µL de látex y 60µL de Standard y las muestras de forma
idéntica al Standard.
Tras su incubación en un baño termostatizado a 37ºC se midieron en el
espectrofotómetro.
3.2.2. Test inmunoturbidimétrico en suero
Principio del método.
La transferrina presente en la muestra reacciona específicamente con un anti-
transferrina humano. La turbidez surgida debido a la formación de un inmunocomplejo
antígeno-anticuerpo se mide a 340nm, siendo proporcional a la concentración de
transferían en la muestra.
Procedimiento
Se tomaron las muestras y se separaron en distinto viales para adecuar cada una a
unas condiciones determinadas:
• Muestra de suero sanguíneo con heparina. • Muestra de suero sanguíneo con heparina y pirita • Muestra de suero sanguíneo con heparina, pirita y ácido sulfúrico. • Heparina y pirita.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 21
Esta última como referencia ya que en algunos casos se ha podido constatar la
absorbancia del anticoagulante del reactivo.
Se preparo un blanco con 800µL de reactivo, un standard con 800µL de reactivo
y 5µL de stándard y de forma idéntica al standard se trataron las muestras.
Tras su incubación durante 10minutos en un baño termostático se lee la
absorbancia a 340nm.
3.2.3. Prueba fotométrica colorimétrica para el hierro con factor aclarante de
lípidos (LCF)
Principio del método
El Fe3+ reacciona con el cromazurol B y cetitrilmetilbromuro de amonio (CTMA)
para formar un complejo ternario coloreado con una máxima absorbancia a 623nm. La
intensidad del color producido es directamente proporcional a la concentración de hierro
en la muestra.
La prueba también puede ser usada en la combinación con el equipo TIBC para
determinar la capacidad total de fijación de hierro.
Procedimiento
Se tomaron las muestras y se separaron en distinto viales para adecuar cada una a
unas condiciones determinadas:
• Muestra de suero sanguíneo con heparina. • Muestra de suero sanguíneo con heparina y pirita • Muestra de suero sanguíneo con heparina, pirita y ácido sulfúrico. • Heparina y pirita.
Esta última como referencia ya que en algunos casos se ha podido constatar la
absorbancia del anticoagulante del reactivo.
Se preparo un blanco con 50µL de agua destilada y 1000µL de RGT, y Standard
con 50µL de STD y 1000µL de agua destilada y de forma idéntica al standard se
trataron las muestras.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 22
Tras la lectura de las absorbancias se procedió al cálculo de concentración según
la siguiente fórmula:
Resultados
Los resultados no fueron concluyentes debido a la formación de coagulaos en las
muestras, que dieron unas mediciones erróneas y a la falta de muestras.
ΔAmuestra
ΔASTD C = 100*

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 23
BIBLIOGRAFÍA
(0). Environmental Risk and meningitis Epidemics in África.
Anna M. Molesworkth, Luís E. Cuevas, Stepehn J. CONNOR, Andrew P. Morsse and
Madeleine C. Thomson.
Liverpool School of Tropical Medicine, Liverpool, United Kingdom and University of
Liverpool, Liverpool, United Kingdom.
Emerg Infect Dis (serial online) 2003 oct. Available from: URL:
http://www.cdc.gov/ncido/EID/vol9no10/03/0182.htm
(1). Intercontinental spread of Neisseria meningitidis clones of the ET-5 complez.
Caugant DA, Froholm LO, Bovre K, Holten E, Fras CE, Mocca LF, Zollinger WD,
Selander RK. Dept. of Biology, University of Rochester, NY.
Antonie Van Leeuwenhoek. 1987;53(6):389-94
(2). Carriage of Neisseria meningitidis in a semi-isolated arctic community.
Holten E, Bratlid D, Bovre K.
Scand J Infect Dis. 1978;10(1):36-40
(3). Intranasal infection of infant mice with Neisseria meningitidis.
Fiona G. Mackinnon, Andrew R. Gorringe, Simon G.P. Funnell and Andrew Robinson.
Biologics Division, PHLS Centre for Applied Microbiology and Research, Porton
Down, Salisbury SP4 OJG, U.K.
Neisseria Meningitidis. Microbial Patogénesis 1992; 12; 415-420.
(4). Interaction of Neisseria meningitidis with human nasopharyngeal mucosa:
attachment and entry into columna epithelial cells.
Stephens DS, Hoffman LH, McGee ZA.
The Journal of infectious diseases. 1983 Sep;148(3):369-76.
(5). Attachment of Neisseria meningitidis to human mucosal surfaces: influence of pili
and type or receptor cell.
Stephens DS, McGee ZA.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 24
The Journal of Infectious Diseases. 1981 Apr; 143(3):525-32.
(6). Acute bacterial meningitis as a complicarion o otitis media and related mortality
factors.
Geyik MF, Kokoglu OF, Hosoglu S, Ayaz C.
Department of Infectious Diseases and Clinical microbiology, dicle University Hospital,
Diyarbakir, Turkey. [email protected].
Yonsei Med J. 2002 Oct; 43(5):573-8.
(7). Nasopharyngeal, vaginal and anal carriage of Neisseria Meningitidis in Nigeria.
Gignani HC, Uganabo JA.
J Común Dis. 1989 Mar; 21 (1):41-5.
(8). Laboratoy-Acquired Meningococcal Disease-United Status, 2000.
MMWR. Weekly. February 22, 2002/51(07);141-4.
(9). Esposure to patients with meningococcal disease on aircrafts—United States, 1999-
2001.
MMWR Morb Motal Wkly Rep. 2001 Jun 15; 50(23):485-9.
(10). Airborne microorganism monitoring: a comparison of servera methods, including
a new direct counting technique.
Hysek J, Fisar Z, Zizka Z, Kofronova O, Binek B.
Institute of Landscape Ecology, Czechoslovak Academy of Sciences, Prague.
Zentralbl Mikrobiol. 1991;146(6):435-43.
(11). Epidemia meningococcal disease: síntesis of a hypothetical
inmmunoepidemiologic model.
Griffis JM.
Rev Infect Dis. 1982 Jan-Feb;4(1):159-72.
(12). Host Cell Invasion By Pathogenic Neisseriae.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 25
Christoph Dehio, Scott D. Gray-wen, and Thomas f. Meyer. (Dept. Medical Genetics &
Microbiology, University of Toronto, Toronto, Ontario M5S 1A8, Canada.)
Subcellular Biochemistry, volume 33: Bacterial Invasion into Eukaryotic Cell, edited by
Oelschlaeger and hacker. Kluwer Academic/Plenum Publichers, New York, 2000.
(13). Characteristics of the meningococcal carrier state in organiz collectives and its role
in the development of generalized form meningococcal infection.
Efimox EI, Khokhlov DT.
Zh Mikrobiol Epidemiol Immunobiol. 1985 Sep;(9):64-7.
(14). Neisseria Meningitidis in a primary school.
Cann KJ, Rogers TR, Jones DM, Noah ND, Burns C.
Department of Medical Microbiology, Charing Cross and Westminster Medical School,
London.
Archives of Disease in Childhood. 1987 Nov;62(11):1113-7.
(15). Nasopharyngeal carrieage of Neisseria Meningitidis in general population and
meningococcal disease.
Ichhpujani RL, Mohan R, Grover SS, Joshi PR, Kumari S.
Departamen of Microbiology, N.I.C.D., Delhi.
J Commun Dis. 1990 Dec;22(4):264-8.
(16). Carriage of Neisseria Meningitidis and Neisseria lactamica in northern Greece.
Kremastinou J, Tzanakaki G, Levidiotou S, Markou F, Themeli E, Voyiatzi A, Psoma
E, Theodoridou M, Blackwell CC.
National meningitis Reference Laboratoy, National School of Public He 196 alexandras
Ave., Athens, Greece.
FEMS Immunol Med Microbiol. 2003 Oct 24;39(1):23-9.
(17). Bacteria: The Proteobacteria.
Prescott, Harley, Klein.
Prescott’s Microbiology, 4/e.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 26
(18). Iron Loading and disease Surveillance.
Eugene D. Weinberg
Indiana university, Bloomington, Indiana, USA.
Past Issue. Vol.5, No. 3 May-June
(19). The iron overload syndrome in patients with severe bacterial inflammatory
diseases and convalescents.
Soboleva MK, Gavalov SM.
Gematol Transfuziol. 1993 Mar;38(3):21-4.
(20). Iron overload exacerbates experimental meningoencephalitis by Cryptococcus
neoformans.
Barluzzi R, Saleppico s, Nocentini A, Boelaert JR, Neglia R, Bistoni I, Blasi E.
Departament of Experimental medicine and Biochemical Sciences, University of
Perugia, perugia, Italy.
J Neuroimmunol. 2002 Nov; 132(1-2):140-6.
(21). Assimilation of iron by pathogenic Neisseria spp.
Yancey RJ, Finkelstein RA.
Infect Immun. 1981 May;32(2):592-9.
(22). Iron as a replacement for mucin in the establishment of meningococcal infection in
mice.
Calver GA, Kenny CP, Lavergne G.
Can J Microbiol. 1976 Jun;22(6):832-8.
(23). Iron acquisition by Neisseria Meningitidis in vitro.
Archibald FS, De Voe IW.
Infect Immun. 1980 Feb ;27(2) :322-34.
(24). Intranasal infecction of infant mice with Neisseria Meningitidi
Mackinnon FG, Gorringe AR, Funnell SG, Robinson A.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 27
Microb Pathog. 1992 Jun;12(6):415-20.
(25). Iron acquisition systems in the pathogenic Neisseria.
Anthony B. Schryvers & Igor Stojilikovic
Molecular Microbiology. Vol 32 Issue 6 page 1117-June 1999
(26). Identification of iron-activvated and-repressed Fur-dependent genes by
transcriptome analysis of Neisseria Meningitidis group B
Renata Grifantini, Shite Sebastian, Elisabetta Frigimelica, Monia Draghi, Erika
Bartolini, Alessandro Muzzi, Rino Rappuoli, Guido Grandi and Caroline Attardo
Genco.
Chiron SpA, siena, Italy and Department of Medicine, Section of infectious Diseases,
and Department of Microbiology, Boston University School of Medicine, Boston, MA
02118.
Communicated by Stanley Falkow, Stanford University, Stanford, CA, May 19, 2003,
(received for review January 6, 2003)
Department of Medicine, Section of infectious diseases, Boston University School of
Medicine, 650 Albany Street, Boston, Ma 02118. E-mail: [email protected]
Proc. Natl Acad Sci USA. 2003 August 5; 100(6): 9542-9547
(27). The Neisseria meningitidis haemoglobin receptor: its role in iron utilization and
virulence.
Stojiljkovic I, Hwa V, de Saint Martin L, O’Gaora P, Nassif X, Heffr F, So M.
Department of Molecular Microbiology and Immunology, Oregon health Sciences
University, Portland 97201, USA.
Molecular Microbiology. 1995 Feb;15(3):531-41.
(28). Mechanisms of iron acquisition by the human pathogens Neisseria meningitidis
and Neisseria Gonorrhoeae.
Rohde KH, Dyer DW.
Department of Microbiology and Immunology, University of Oklahoma health Sciences
Center, 975 NE 10th Stret, Oklahoma City, Oklahoma 73104, USA.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 28
Front Biosci. 2003 Sep 1; 8:d1186-218.
(29). Interactions of pathogenic Neisseria with host cells. Is it possible to assemble the
puzzle.
Xavier Nassif, Céline Pujol, Philippe Morand and Emmanuel Eugene.
INSERM U411, Faculté de Médecine Necker-Enfants Malades, université René
Descartes, Paris, France.
Molecular Microbiology, 1999 32(6), 1124-1132.
(30). Microvilli-like structures are associated with the internalization of virulent
capsulated Neisseria Meningitidis into vascular endothelial cells.
Eugene E, Hoffmann I, Pujol C, Couranud PO, Bourdoulous S, Nassif
INSERM U411, Faculte de Medecine Necker-Enfants Malades, University Rene
Descartes, Paris, France. CNRS UPR 415, Institud Cochin de Genetique Moleculaire,
Paris, France.
R1(39). Host cel invasion by pathogenic Neisseriae.
Dehio C., Gray-Owen SD., Meyer TF..
Dept. Infektionsbiologie, Max-Planck-Institut fur Biologie, Tubiengen, Germany.
Subcell Biochem. 2000; 33:61-96
J Cell Sci. 2002 Mar 15;115(Pts 6):1231-41.
(31). The effect of variations in the expression of pili on the interaction of Neisseria
Meningitidis with human nasopharyngeal epithelium.
Rayner CF, Dewar A, Moxon ER, Virji M, Wilson R.
Department of Thoracic Medicine, Royal Brompton National Heart and Lung Institute,
London, United Kingdom.
J Infect Dis. 1995 Jan;171(1):113-21.
(32). Pilus genes of Neisseria Gonorrhoeae: Chromosomal Organization and DNA
Sequence.
Proc. Nat. Acad. SCi. USA 81.6110-6114.
Meyer T.F, Billyard E, Haas R, Storzbach S and So. M.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 29
Proc. Natl. Acad. Sci. USA. Vol. 81, pp. 6110-6114, Octubre 1984 Genetics.
(33). Structure of fibre forming protein pilin at 2.6 A resolution.
Nature 378: 33-38
Parge H.E, forest K.T., Hickey M.J, Christensen D.A, Getzoff E.D and Tainer J.A. 1995
(34). CD46 in Meningococcal Disease
Linda Johansson, Anne Rytkönen and Ann-Beth Jonsson. Microbiology and Tumor
Biology Center, Nobels väg 16, Box 280, Karolinska Institutet, SE-171 77 Stock-holm,
Sweden.
Peter Bergman and Birgitta Agerberth. Department of Medical Biochemistry and
Biophysics
Helena Källström. Department of Cell and Molecular Biology
Tomas Hökfelt. Department of Neuroscience, Karolinska Institutet, SE-171 77
Stockholm, Sweden.
Barbara Albiger. Swedish institute for Infectious Disease Control, SE-171 82 Solna,
Sweden
Roberto Cattaneo. Molecular Medicine Program, Mayo Clinic, Guggenheim 1838, 200
First Street SW, Rochester, MN 55905, USA.
Science, 2003 Jul 18;301(5631):373-5.
(35). The ferritins: molecular properties, iron storage function and cellular regulation
Paule M. Harrison, Krebs Institute, Departament of Molecular Biology and
Biotechonology, University of Sheffeld, Firth Court, Western Bank, Sheffield, UK.
Paolo Arosio, Unit of Protein Engineering, DIBIT, San Raffaele Scientific Institute,
Milano and Cattedra di Chimica, University of Brescia, Italy.
BBA (Biochimica et Biophysica Acta) 1275 (1996) 161-203. ELSEVIER.
(36). Iron metabolism in pathogenic bacteria.
Ratledge D., Dover LG.
Departament of Biological Sciences, University of Hull, HU6 7RX.
Annu Rev Microbiol. 2000;54:881-941.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 30
(37). Importancia de los metaloreguladores FUR y ZUR en la virulencia de Salmonella
typhimurium.
Susana Campoy Sánchez.
Universidad Autónoma de Barcelona. Departamento de Genética y Microbiología.2002
(38). The Structural Mechanism for Iron Uptake and Release by Transferrins.
Massaki Hirose.
The research Institute for Food Science, Kyoto University, Uji, Kyoto 611-0011, Japan
Biosci. Biotechnol. Biochem. 64(7), 1328-1336. 2000
(39). Alternative Structural State of Transferrin. The Crystallographic analysis of iron-
loaded but domain-opened. Ovotransdferrin N-lobe.
Kimihiko Mizutani, Honami Yamashita, Hirofumi Kurokawa, Bunzo Mikami and
Masaaki Hirose.
From the Research Institute for Food Science, Kyoto University, Uji, Kyoto 6110011,
Japan.
J Biol chem., vol 274, Issue 15, 10190-10194, April 9, 1999.
(40). Dealing with iron: Common structural principles in proteins that transport iron and
heme
Heather M. Baker, Bryan F. Anderson and Edward N. Baker.
School of Biological Sciences and department of Chemistry, University of Auckland,
Private Bag 92-019, Auckland, New Zealand; and Institute of Molecular Biosciences,
Massey University, Palmerston North, New Zealand.
Publisched online before print Marche 17, 2003,
(41). Stoichiometric and site Characteristics of the Binding of Iron to Human
Transferrin
Philip Aisen, Adela Leibman and Jay Zweier.
From the departament of Biophysics and Medicine, Albert Einstein College of
medicine, Bronx, New York 10461.

NERVA-MECANISMO DE ADQUISICIÓN DE HIERRO MINERAL
www.grupotar.com 31
The Journal of Biological Chemistry. Vol. 253. Nº 6, Issue of March 25, pp. 1930-1937,
1978.
(42). The pH-induced release of iron from transferrin investigated with a continuum
electrostatic method.
Lee DA., Goodfellow JM.
Department of Crystallografhy, Birkbeck College, University of London, United
Kingdom.
Biophys J. 1998 Jun;74(6): 2747-59.
(43). Anion exchange in human serum transferrin N-lobe: a model study with variant
His249Ala.
He QY, Woodworth RC, Chasteen ND.
Department of Chemistry, University of New Hampshire, Durham, NH 03824, USA.
J Biol Inorg Chem. 2003 Jul;8(6):635-43. Epub 2003 May.