modulaciÓn de la respuesta …digital.csic.es/bitstream/10261/87714/1/tesis adegared.pdf · en...
TRANSCRIPT



Departamento de Bioloxía animal, Bioloxía vexetal e EcoloxíaUniversidade da Coruña
MODULACIÓN DE LA RESPUESTA ANTIDEPREDATORIA ENLA RANA VERDE IBÉRICA (PELOPHYLAX PEREZI): USO DE SUSTANCIAS DE ALARMA Y APRENDIZAJE
Memoria presentada por la Licenciada en Biología Adega Gonzalo Martínezpara optar al título de Doctor en Biología por la Universidade da Coruña
Adega Gonzalo Martínez
Jose Martín RuedaMuseo Nacional de
Ciencias Naturales (CSIC)
Pilar López MartínezMuseo Nacional de
Ciencias Naturales (CSIC)
Directores:
Tutor:
Pedro Galán RegaladoUniversidade da Coruña
Coruña, Marzo de 2009


A mis padresA Alba


Índice
Introducción general 7
Capitulo V
Capitulo IV
Capitulo II
Capitulo III
Capitulo I
Conclusiones 101
Agradecimientos 1
Los renacuajos de rana verde ibérica pueden aprendera reconocer nuevos depredadores mediante lasseñales de alarma de individuos coespecíficos
Aprendizaje, memoria y olvido aparente de señalesquímicas procedentes de nuevos depredadores enrenacuajos de rana verde
Olvido adaptativo en renacuajos de rana verde: laInhibición latente y el Aprendizaje de lo irrelevantepueden evitar falsos reconocimientos de depredadores
Evaluación quimiosensorial del riesgo de depredaciónen renacuajos de rana verde Ibérica: Diferencias entreel aprendizaje de depredadores mediante sustanciasde alarma o sustancias de estrés
Las sustancias químicas de depredadores, pero no lasde individuos coespecíficos influencian la seleción decharcas en individuos metamórficos de rana verde
29
45
59
73
89


AGRAdeciMienTOS


... y he aquí la parte de la tesis que más trabajo me ha costado escribir.Precisamente porque esta tradición “literaria” será lo más leído, y porque seguro queme olvidaré de alguien y lo lamentaré eternamente
Hace poco más de seis años mi vida cambió por completo. Sin lugar a dudas la elabo-ración de una Tesis Doctoral es mucho más que un trabajo normal, es un “modo devida”, un desafío personal al que llegas sin tener apenas conocimiento de cómo fun-cionan las cosas y en el que te emocionas con todos los pequeños descubrimientosque haces día a día. En primer lugar, desearía agradecer profundamente al que fuesu impulsor: el doctor José Guitián, profesor de ecología y mi autentico “tutor encu-bierto” durante los últimos años de carrera. Él fue el que decidió que debía volar lejosde aquella universidad en busca de lo mejor que pudiera encontrar y el que me“obligó” a ir a un curso de postgrado del CSIC que cambiaría mi vida.
“El curso del Ventorrillo” fue la mejor manera de abrirle los ojos a esta licenciada ne-ófita y fue precisamente en él donde conocí a las personas más importantes en la re-alización de esta tesis. Mis directores: Jose Martín y Pilar López, dos investigadoresexcepcionales en los que siempre he encontrado apoyo y ayuda y que me han dado lalibertad y seguridad necesarias para elegir el tema de esta tesis, plantear mis expe-rimentos y desarrollar mis ideas. Ellos me acogieron con los brazos abiertos en sudespacho y en sus vidas, han soportado mis estupideces y me han guiado todos estosaños de aprendizaje tanto profesional como personal.
También quisiera agradecérselo de forma especial a Pedro Galán, por compartir con-migo su gran conocimiento zoológico, por preocuparse de avisar de los plazos buro-cráticos que tanto odio y por acogerme con una sonrisa cada vez que aparecía porsu despacho, como siempre, sin avisar.
Mención especial a mis compañeros de equipo: Liesbeth, cabeza bien amueblada ypies en el suelo; Luisa, risueña y alegre como pocas; Marianne y sus ideas extrañas;Pedro Moreira y su obsesión por los diseños experimentales perfectísimamente per-fectos; Kevin (y Rita), el guiri feliz. Emilio, tranquilidad y calma. A Nuria por las char-las, la paciencia y su seguridad, por compartir sus conocimientos sobre tortugas y,sobre todo, parte de su vida conmigo: Ya sabes, cuando vuelvas a Badajoz a por tor-tugas ¡Cuenta conmigo! A Carlos, por ayudarme en todo lo posible y más, por acom-pañarme al campo, ayudarme a capturar especimenes, por su apoyo, por lasconversaciones interminables sobre como funciona el mundo en general y los bichosen particular, por las noches en vela y por las teorías absurdas pero bien fundamen-tadas que nos montábamos.
Al Dr. W. Cooper por acogerme en Indiana, tener paciencia con mi pésimo inglés yanimarme a hacer todo aquello que quisiera en la vida. Al Dr. Chivers por incluirmeen su laboratorio y tener paciencia cuando no entendía su pésimo inglés.

A todos aquellos que han pasado por El Ventorrillo (becarios, voluntarios, ayudantesesporádicos, postdoctorales y científicos titulares) y han compartido conversacionessesudas, risas en el césped, momentos de agobio, cervecitas vespertinas y salidas noc-turnas. A Nino, una de las mejores personas que he conocido en este mundo, siempredispuesto a ayudar y a invitar a cerveza.
Por supuesto, al Museo de Ciencias Naturales y a la Estación biológica del Ventorrilloen cuyas instalaciones pude realizar todos los experimentos de la tesis.
A Nocko, amigo internetero que me abrió su Flickr para que eligiera la foto que másme gustara para la portada.
No puedo olvidar a mi familia porque, por suerte o por desgracia, soy como soy gra-cias a ellos y son, sin duda alguna, las personas más importantes en mi vida. Mis pa-dres que, aunque no les gustó que emigrara, me apoyaron siempre en mis decisionesy en “esas cosas raras que haces con los bichos”. Mi hermana, a la que he tenido lasuerte de redescubrir y que se ha convertido en un pilar fundamental en mi vida.
Y ¿que decir de toda esa gente que ha estado ahí desde el principio o que he conocidoen estos años? Esas personas que han vivido conmigo la realización de esta tesis doc-toral, con sus altos y bajos. Paula la señora del Avante, Patricia la escritora de éxito,Mariluz la abogada agresiva…que han aguantado mis ausencias, mis apariciones ydesapariciones repentinas, mis cambios de planes porque los bichos nunca descan-san… y aún así me siguen llamando para preguntarme que qué tal me va y quecuándo vuelvo a Santiago; Anita, chocolate y Alicia, galleta (caramelo!!)…que me hanacogido en su piso de Madrid, me han sacado y han cuidado de mi mucho más de loque me merecía, grandes amigas con las que tengo la enorme suerte de vivir. Sandra,mi física bohemia, con la que tengo conversaciones telefónicas de varias horas cada6 meses y que me ha demostrado que siempre se puede contar con ella; Max el rusopedante, que me acogió cuando más lo necesitaba. La gente de Internet, que siempreme ha animado en los momentos bajos y que me han hecho reír muchísimo, quien meiba a decir a mí que conocería personas tan fantásticas.
Ó meu neno, por compartir comigo toda a sua ilusión por vivir, pase o que pasesempre nos quedará Portugal.



inTROducción


La depredación se considera como uno de los procesos claves en la regulación delas poblaciones animales, así como uno de los factores selectivos más ampliamentedistribuidos en la naturaleza (Darwin 1859). En términos generales, puede definirsecomo la interacción en la que un organismo captura y consume una fracción significa-tiva de la biomasa de otro organismo (Abrams 2000). La depredación ejerce unaimportante influencia sobre los organismos a escala individual, ecológica y evolutiva,afectando tanto al comportamiento como a la morfología, la dinámica poblacional,la estructura de las comunidades o las estrategias vitales de las especies (Sih 1992).Por lo tanto, el riesgo de depredación puede ser considerado como una de las mayoresfuerzas selectivas en la evolución, favoreciendo la aparición de complejas estrategiasantidepredatorias tanto morfológicas como comportamentales (Lima y Dill 1990;Lima 1998).
Un evento depredador puede ser descrito como un ciclo con diferentes fases que,en total, componen un episodio de depredación: búsqueda, encuentro, detección,identificación, ataque, captura y consumo de la presa (Endler 1991). Por supuesto, laspresas intentarán interrumpir este ciclo de eventos tan pronto como sea posible,mediante elaboradas adaptaciones defensivas que pueden manifestarse a través demodificaciones de su comportamiento, morfología o historia vital (Kats y Dill 1998).Sin embargo, muchas de estas adaptaciones son costosas para las presas (Werner yAnholdt 1996; Wisenden 2000; Werner y Peacor 2003). Por ejemplo, en el desarrollode estructuras defensivas un organismo puede consumir energía y recursos quepodrían haber sido invertidos en crecimiento o en reproducción. Asimismo, un cambiode hábitat o de los patrones de actividad en presencia de los depredadores puededisminuir la tasa de alimentación o la tasa de encuentros con potenciales parejas (Sih1980; Lima y Dill 1990). Además, la presión depredadora puede obligar a los animalesa utilizar hábitats subóptimos (Gotceitas y Godin 1991) o a retrasar cambios ontogé-nicos de nicho (Olson et al. 1995). La modificación de la historia vital para evitar elriesgo de depredación también conlleva costes que pueden reflejarse en el acceso alos sucesivos estadios del ciclo vital en condiciones que comprometan la supervivenciade los individuos. Por ejemplo, el acortamiento de una fase del ciclo vital para escaparde la depredación puede suponer para el individuo el acceso al siguiente estadio conun menor desarrollo y por tanto una mayor vulnerabilidad frente a otros depredadoreso frente a las variaciones ambientales (Warkentin 1995; 1999; 2000).
Detección de depredadores y valoración del riesgo de depredación
Las señales acústicas, visuales, táctiles o químicas pueden aportar la informaciónnecesaria para identificar a un potencial depredador, y pueden ser utilizadas en con-junto o por separado, dependiendo del ambiente físico en el que se muevan las presas.Independientemente del canal de información utilizado, la intensidad de la respuesta
“La vida es un proceso de
adquisición de conocimientos”
Konrad Lorenz
9

antidepredatoria de un individuo debería ajustarse al riesgo de depredación al queestá sometido (Helfman 1989). Sin embargo, la elaboración y mantenimiento decomportamientos antidepredatorios resulta costosa, ya que los individuos se enfrentana situaciones de compromiso en términos de tiempo y energía que pueden compro-meter su éxito biológico (Helfman 1989). De este modo, el desarrollo de mecanismospara evaluar correctamente el riesgo de depredación, actuar de acuerdo con el nivelde riesgo y minimizar los costes asociados, aportará ventajas evolutivas a las especiespresa. Así, las distintas especies han desarrollado diferentes formas de detectar yevaluar el riesgo de depredación en función de las características del ambiente en elque viven, así como distintas estrategias para minimizarlo en función de la presióndepredadora a la que son sometidas. Esta hipótesis se conoce como la “threat-sensitive
predator avoidance hypotesis” (hipótesis de la respuesta dependiente del nivel de riesgode depredación, Helfman 1989).
Señales químicas utilizadas en la detección de depredadores en medios
acuáticos
En aguas turbias o en medios complejos, con abundante vegetación, las presascapaces de detectar las señales químicas que indican la presencia de los depredadorestendrán más posibilidades de escapar que aquellas que utilizan únicamente el canalvisual (Kats y Dill 1998). Muchos trabajos previos han mostrado que un gran númerode organismos acuáticos modifican su morfología, comportamiento y/o característicasdel ciclo vital en presencia de señales químicas asociadas a la depredación, reduciendocon ello el riesgo de ser capturados (revisiones en Kats y Dill 1998; Brönmark y Hans-son 2000). Estas señales químicas proporcionan una manera muy precisa de evaluarel riesgo de depredación.
En las interacciones depredador-presa existen básicamente cuatro fuentes de sus-tancias químicas que previenen a las presas de la proximidad de un depredador: (1)las sustancias liberadas por el propio depredador (denominadas cairomonas según ladefinición de Dodson et al. 1994; Brönmark y Hansson 2000), (2) las sustancias emi-tidas por una presa acosada y estresada pero no dañada (señales de estrés o “distur-bance cues”, Chivers y Smith 1998), (3) las señales de alarma relacionadas consustancias emitidas por una presa cuando está siendo consumida por el depredador(sustancias de alarma o “alarm cues”, Chivers y Smith 1998) y (4) aquellas relacionadascon los productos de excreción emitidos por el depredador y asociadas al consumo deuna presa (Laurila et al. 1997; Chivers y Mirza 2001).
El reconocimiento de cualquiera de estos tipos de señales químicas representa unaventaja para las presas, puesto que permite la evaluación del riesgo de depredacióndesde la seguridad de un refugio, y la detección de un depredador en medios que difi-culten la localización visual por ausencia de luz (por ej. medios turbios o profundos) odebido a una alta complejidad estructural (por ej. con abundante vegetación), inclusodespués de que el depredador haya abandonado el área (Dodson et al. 1994; Kats yDill 1998).
10

Las cairomonas son señales químicas emitidas por los depredadores y utilizadas por laspresas en beneficio propio para detectar su presencia. Por tanto, no reportan ningún beneficio aldepredador (Weldon 1980; Larsson y Dodson 1993; Weber 2003). Las presas han evolucionadopara reconocer estas sustancias emitidas por los depredadores y utilizarlas en su propio beneficio,ya que indican la cercanía de un potencial depredador (Hokit y Blaustein 1995). Puesto que laemisión de señales químicas por parte del depredador produce un beneficio general en otrocontexto (por ej. en el reconocimiento de posibles parejas reproductoras), y dado que es imposiblesu total supresión (como es el caso de los productos de excreción), la emisión de cairomonas debeser consecuentemente ajustada por el depredador, reduciendo en lo posible su utilización. La ha-bilidad de las presas para reconocer depredadores a través de las cairomonas crea un escenariocoevolutivo que empuja a los depredadores a decidir entre minimizar la emisión de esas sustanciasquímicas o modificarlas, al mismo tiempo que las presas se ven empujadas a aumentar su sensi-bilidad a las señales existentes o desarrollar un reconocimiento de las nuevas señales.
Las señales de estrés (“disturbance cues”) son sustancias químicas liberadas por las presasque han sido acosadas, pero no capturadas, por un depredador. Este tipo de señales ha sido muypoco estudiado, sin embargo, los experimentos realizados en este campo sugieren un uso amplia-mente extendido (Hazlett 1985; 1989; 1990a; 1990b; Wisenden et al. 1995; Kiesecker et al. 1999).En algunas especies de anfibios, el amonio (NH4+) puede formar parte de esta señal. Varios estu-dios demuestran que individuos sometidos a estrés depredatorio excretan mayor cantidad relativade este compuesto, al mismo tiempo que otros individuos sin estrés depredatorio, aumentan elnúmero de comportamientos antidepredatorios al ser expuestos a bajas concentraciones de amo-nio (Hazlett 1990a; Kiesecker et al. 1999).
Las sustancias de alarma o feromonas de alarma (“alarm cues”, Blum 1969;Smith 1992) sonsustancias químicas liberadas por presas heridas por el depredador, y que producen una respuestaantidepredatoria en individuos coespecíficos. Las adaptaciones defensivas desencadenadas porlas sustancias de alarma han sido documentadas en muchos grupos animales, incluyendo insectosterrestres (Blum 1985; Teerling et al. 1993; Manzoli-Palma et al. 1998), insectos acuáticos (Huryny Chivers 1999), crustáceos (Hazlett 1994), arácnidos (Machado et al. 2002), gasterópodos (Atemay Stenzler 1977; Ichinose 2002), peces (Von Frisch 1938; Pfeiffer 1977; Mathis y Smith 1993a),anfibios (Chivers et al. 1996a) y mamíferos (Rottmann y Snowdon 1972).
La dieta del depredador influye también en el reconocimiento por parte de la presa, de lassustancias químicas que éste emite. Varios estudios han demostrado que los organismos presaexhiben respuestas antidepredatorias ante señales químicas de depredadores que recientementehan sido alimentados con individuos coespecíficos, pero no ante depredadores que han sidoalimentados con una dieta diferente (Chivers y Smith 1998). Estas respuestas antidepredatoriashan sido documentadas en larvas de insectos (Chivers et al. 1996b), anfibios (Wilson y Lefcort1993; Laurila et al. 1998; Chivers y Mirza 2001), gasterópodos (Jacobsen y Stabell 1999), yanémonas marinas (Howe y Harris 1978). En la mayoría de los casos las señales químicas libera-das en los productos de excreción de los depredadores han sido directamente asociadas con lassustancias de alarma de las presas (Howe y Harris 1978; Mathis y Smith 1993b; Stabell y Lwin1997). Así, las sustancias de alarma emitidas por la presa al ser depredada, son liberadas por eldepredador en las heces, la orina o los productos respiratorios (Gelowitz et al. 1993) y reconocidaspor las demás presas.
Aprendizaje de depredadores
A lo largo de sus vidas, las presas se encuentran con multitud de otros animales,algunos de los cuales pueden ser depredadores potenciales. Pero, ¿cómo pueden saberlos individuos qué especies son potencialmente peligrosas y cuáles no? Las respuestasa las señales de los depredadores pueden estar fijadas genéticamente, y por tanto serinnatas, o pueden también haber sido adquiridas por medio del aprendizaje. Muchosvertebrados han demostrado poseer una capacidad innata para reconocer a sus
11

depredadores; por ej. mamíferos (Barros et al. 2002), aves (Veen et al. 2000; Goth2001), anfibios (Kats et al. 1988; Kiesecker y Blaustein 1997) y peces (Berejikian et al.2003; Hawkins et al. 2004). Sin embargo, muchos otros estudios llevados ha cabo enestos mismos grupos de animales, han demostrado que el reconocimiento de depre-dadores es también el resultado de un aprendizaje; mamíferos (McLean et al. 2000;Griffin et al. 2001), aves (Maloney y McLean 1995; McLean et al. 1999), anfibios (Sem-litsch y Ryer 1992; Woody y Mathis 1998; Mirza et al. 2006; Gonzalo et al. 2007), ypeces (Chivers y Smith 1994; Ferrari et al. 2005).
La habilidad de aprender a reconocer nuevos depredadores tiene implicacionessumamente importantes para la supervivencia (Mirza y Chivers 2001). En general, ladepredación es muy variable a través del tiempo y el espacio (Hileman y Brodie 1994;Sih et al. 2000; Mathis et al. 2003), con lo que la habilidad para aprender a reconocernuevos depredadores puede resultar más ventajosa que el reconocimiento innato, enaquellas ocasiones en que las comunidades de depredadores no permanezcanconstantes durante largos periodos de tiempo.
El papel del hábitat en el desarrollo del aprendizaje
Los depredadores o el grado del riesgo de depredación al que se enfrenta un indi-viduo pueden variar a medida que éste crece (Brönmark y Miner 1992), debido a cam-bios en sus preferencias de hábitat (Werner y Gilliam 1984), o simplemente a cambiosprovocados por el paso de las estaciones en las condiciones bióticas (por ej. variaciónestacional de los depredadores), en las abióticas (por ej. desecación de las masas deagua que habitan), o en ambas (Gilliam y Fraser 2001). De cualquier modo, la forma yla frecuencia de la depredación pueden cambiar de año en año, de estación en estación,de día en día; y no sólo temporalmente, sino también en el espacio (entre microhábi-tats). Como resultado de estas variaciones, muchas veces resulta inviable para las pre-sas adoptar la estrategia de evitar a todos los depredadores potenciales, ya que estecomportamiento supondría costes excesivos para el individuo, como por ejemplo, unapeligrosa reducción de su eficacia alimenticia y supervivencia (Lawler 1989; Skelly1992). En este contexto inestable, los individuos capaces de tomar decisionesantidepredatorias muy ajustadas y basadas en experiencias recientes (es decir,individuos capaces de aprender y recordar) presentarían una ventaja adaptativa(Brown y Chivers 2005). Así, las poblaciones expuestas a un riesgo de depredaciónvariable en el tiempo, pueden haber evolucionado hacia el uso de una respuestaaprendida, que les permitiera balancear los compromisos (“trade-offs”) entre respues-tas antidepredatorias y otros beneficios alimenticios o reproductivos.
De igual modo, en las poblaciones caracterizadas por una presión depredadora ele-vada o por una diversidad reducida de depredadores, se puede predecir la existenciade un riesgo de depredación poco variable en el tiempo. De este modo, las presasreconocerán a un nuevo depredador de manera innata, simplemente porque se tratauna especie nueva. Esta estrategia evolutiva es conocida como neofobia (Breden et al.1987). La neofobia puede ser adaptativa si la presión depredadora es muy alta, o si elnúmero de especies depredadoras es pequeño, o si concurren ambas circunstancias.
12

Señales químicas y aprendizaje de depredadores por organismos acuáticos
Estudios realizados en diferentes especies han demostrado que las sustancias dealarma liberadas por los individuos al ser atacados por un depredador, resultan fun-damentales en el proceso de aprendizaje de reconocimiento de nuevos depredadorespotenciales por individuos coespecíficos (revisión en Chivers y Smith 1998; Brown yChivers 2005). Por ejemplo, Chivers y Smith (1994a) demostraron que el piscardoamericano, Pimephales promelas, es capaz de aprender a reconocer el olor de undepredador cuando éste le ha sido previamente presentado junto con sustancias dealarma liberadas por coespecíficos. El aprendizaje se produce hasta el punto de queun solo evento de condicionamiento es suficiente para desencadenar una respuestaantidepredatoria y de que esta respuesta se mantiene incluso dos meses después delcondicionamiento. Numerosos investigadores han examinado el papel de esta sustanciade alarma en P. promelas (Wisenden y Harter 2001; Ferrari et al. 2005; Ferrari y Chivers2006a) y en otros peces, como los zebra danios (Brachydanio rerio: Suboski et al. 1990),la trucha arcoiris (Oncorhynchus mykiss: Brown y Smith 1998), la trucha de arroyo(Salvelinus fontinalis: Mirza y Chivers 2000), el salmón Chinook (Oncorhynchus
tshawytscha: Berejikian et al. 1999) y el espinoso de arroyo (Culaea inconstans: Chiverset al. 1995). Este tipo de aprendizaje ha sido asimismo documentado en planarias(Wisenden y Millard 2001), libélulas (Wisenden et al. 1997), caracoles (Rochette et al.1998), cangrejos de río (Hazlett 2003) y en tres especie de anfibios: los tritonesNotophthalmus viridescens (Woody y Mathis 1998), los renacuajos del sapo Bufo
americanus (Mirza et al. 2006) y los renacuajos del la rana verde ibérica Pelophylax pe-
rezi (Gonzalo et al. 2007; presente tesis). En todos estos casos, las presas fueron capa-ces de aprender a reconocer el olor de un depredador después de una única exposicióna la mezcla de la sustancia de alarma de individuos coespecíficos con sustanciasquímicas de un depredador desconocido hasta ese momento.
Aprendizaje de depredadores: condicionamiento clásico
Las conclusiones de la mayoría de los experimentos sobre aprendizaje de recono-cimiento de nuevos depredadores por parte de las presas dejan claro que se trata deun aprendizaje asociativo, que puede ser considerado un ejemplo de condicionamientoclásico Pavloviano (Blair et al. 2001; Maren 2001; Schafe et al. 2001). En el condicio-namiento clásico, el sujeto aprende que existe una relación causal entre dos estímulos,el estímulo incondicionado o natural (EI), el cual produce una respuesta incondicionadafuerte y consistente, y un estímulo condicionado (EC), el cual, de forma aislada no pro-duce una respuesta similar a la del EI, pero que, debido a que se presenta en asociacióncon éste último, llega a desencadenar por sí solo una respuesta condicionada de igualo similar efecto. En el aprendizaje de nuevos depredadores, un estímulo desconocidopor la presa, que no produce respuesta al inicio (estímulo neutro), es presentado juntoa un indicador de alto riesgo de depredación (EI), ante el cual se desencadena unarespuesta antidepredatoria. Tras el condicionamiento, la sola presencia del ahora EC(antes neutro) desencadena en la presa una respuesta antidepredatoria similar a laoriginada por el EI (Davis 2000).
13

Aprendizaje y memoria
Aunque numerosos estudios han investigado el aprendizaje, muy pocos le han pres-tado atención a la memoria, es decir a la capacidad de retener aquello que ya ha sidoaprendido. Los procesos de aprendizaje y memoria se encuentran muy ligados, ya queel aprendizaje produce pocos beneficios si la información aprendida en un momentodado no es almacenada o memorizada. Sin embargo, los procesos de aprendizaje y lamemoria son distintos. Aprender consiste esencialmente en la adquisición de nuevainformación, mientras que la memoria tiene otros componentes, como la retención dela información adquirida o la posibilidad de que ésta sea modificada por nuevas expe-riencias (Shettleworth 1998). Estudios llevados a cabo en piscardos europeos (Phoxi-
nus phoxinus) o americanos (P. promelas) han demostrado que los individuos queaprenden mediante condicionamiento a reconocer a un depredador nuevo, son capacesde memorizar sus señales y de responder a las mismas, durante un periodo de tiempoque varía entre unos días hasta varias semanas o meses después (Magurran 1989;Chivers y Smith 1994a, 1994b; Brown y Smith 1998; Mirza y Chivers 2000). Además,Hazlett et al. (2002), trabajando con diferentes especies de cangrejos de río, demos-traron la existencia de diferencias en la retención de lo aprendido en base a las distintasestrategias vitales de cada especie. Así, las especies invasoras recuerdan durante mástiempo los depredadores potenciales con los que fueron condicionados, que las espe-cies no invasoras. Esto parece indicar que la habilidad para aprender y memorizardepredadores potenciales podría ser especialmente importante para la supervivenciade aquellas especies que pueden encontrarse con una gran variedad de depredadoreso que viven en ambientes muy variables. Sin embargo, las investigaciones dirigidas acuantificar la duración de la memoria o el ritmo de pérdida al que ésta se halla some-tida, son mucho menos comunes que los estudios acerca de la adquisición de nuevainformación (Shettleworth 1998).
Mecanismos inhibidores del aprendizaje
La capacidad de aprender (y memorizar) depredadores potenciales puede resultarde suma importancia para la supervivencia de los individuos, ya que les permite ajustarsu comportamiento al riesgo de depredación de cada momento. Sin embargo, existe elpeligro de que respondan con un comportamiento antidepredatorio a un estímulo cual-quiera, simplemente porque en algún momento haya aparecido en conjunción con in-dicadores de alto riesgo de depredación (sustancias de alarma). Por lo tanto, este tipode aprendizaje, basado en la presentación de dos estímulos de manera conjunta, puedeoriginar respuestas hacia estímulos irrelevantes desde el punto de vista de la depre-dación. Por ejemplo, Magurran (1989) demostró que los piscardos europeos (Phoxinus
phoxinus) podían llegar a reconocer como depredador al inofensivo pez dorado. Otrostrabajos muestran resultados similares (Chivers y Smith 1994a; Yunquer et al. 1999).Este tipo de respuestas a estímulos que no son peligrosos puede interferir con un com-portamiento ajustado a la realidad del hábitat, y por tanto representa un coste para elindividuo (Brown y Chivers 2005). Sin embargo, existen mecanismos que pueden evitarel mantenimiento de estas asociaciones. Los más importantes son la pérdida de memo-
ria, la inhibición latente y el aprendizaje de lo irrelevante.
14

En algunas especies se puede apreciar que, una vez hecha la asociación, se produceuna disminución de la respuesta o una perdida de memoria con el tiempo, hasta llegara la extinción del comportamiento antidepredatorio (cangrejos de río: Hazlett et al.2002; peces: Ferrari y Chivers 2006b; anfibios: presente tesis, capítulo 3). Esta extin-ción, lejos de ser un fallo de la memoria, puede ser una estrategia para tratar coninformación confusa. Según Turner et al. (2005) los individuos se basan en las expe-riencias más recientes para seleccionar el tipo de comportamiento antidepredatoriomás adecuado a cada circunstancia. Así, a medida que el momento del encuentro conel depredador se aleja en el tiempo, y si no existe ningún nuevo refuerzo del condicio-namiento (un nuevo encuentro con ambos estímulos asociados), los individuos puedenllegar a olvidar la asociación creada y no perseverar en comportamientos antidepre-datorios ante estímulos de especies no depredadoras que viven en el mismo hábitat.
La inhibición latente (“latent inhibition”) de una respuesta aprendida ocurre cuandoel individuo ha sido expuesto previamente a un estímulo neutro antes de encontrarsecon este estímulo en combinación con el estímulo incondicionado, lo que inhibe laformación de la asociación (Ferguson et al. 2001). Así, la presencia de las señales deun depredador sin la presencia de la sustancia de alarma inhibe un futuro aprendizaje,el cual ocurriría, normalmente, cuando ambas sustancias asociadas. Acquistapace etal. (2003) con cangrejos de río y Ferrari y Chivers (2006b) con piscardos americanos,demostraron que los individuos expuestos repetidamente al olor de un depredadorpotencial son incapaces de aprender a escapar de este olor cuando es presentado conposterioridad en conjunción con sustancia de alarma de individuos coespecíficos. Estemecanismo podría estar encaminado a evitar el aprendizaje de información irrelevantey a evitar reaccionar ante especies no depredadoras con las que comparten hábitat.
El aprendizaje de lo irrelevante (“learned irrelevance”) también provoca un falloaparente en la formación de asociaciones entre estímulos. Según Bennet et al. (2000)este fenómeno ocurre cuando la exposición aleatoria a dos estímulos (el estímuloneutro y el incondicionado) impide la formación de la asociación cuando ambosestímulos se presentan conjuntamente. Hazlet (2003), en un experimento en el quesometió a cangrejos de río al olor de un pez depredador y a sustancia de alarma de in-dividuos coespecíficos en orden aleatorio antes de presentarles los dos estímulos jun-tos, demostró que los cangrejos eran incapaces de realizar la asociación de ambosestímulos y fallaban a la hora de reconocer al pez como depredador potencial. Sin em-bargo, al igual que el anterior mecanismo, el aprendizaje de lo irrelevante podría estarencaminado a evitar reacciones innecesarias ante especies no depredadoras con lasque los individuos comparten su hábitat.
Tanto en la inhibición latente como en el aprendizaje de lo irrelevante, la detecciónde un estímulo procedente de un potencial depredador por parte de una presa no sehalla unida temporalmente a estímulos que indiquen un aumento de la presión dedepredación, por lo que, cuando los dos estímulos son detectados al mismo tiempo, laformación de la asociación, y por tanto el aprendizaje, son inhibidos.
15

Aprendizaje en anfibios
Aunque existen multitud de trabajos sobre aprendizaje, muy pocos se han llevadoa cabo en anfibios. Además de la presente tesis, sólo dos trabajos han descrito la capa-cidad de aprender a reconocer depredadores nuevos en otros anfibios. Woody y Mathis(1998) demostraron que los tritones adultos de la especie Notophthalmus viridiscens
eran capaces de asociar sustancia de alarma con olores de un depredador desconocido.Lo mismo han demostrado Mirza et al. (2006) para los renacuajos del sapo americano(Bufo americanus). Estas dos especies, al igual que la rana verde ibérica (Pelophylax
perezi, sobre la que trata la presente tesis), pasan largos periodos de su vida en faseacuática y se encuentran ampliamente distribuidas en hábitats muy diversos, lo queocasiona su convivencia con una gran variedad de depredadores que pueden ir cam-biando rápidamente. El aprendizaje puede ser fundamental en especies presa que vivenen hábitats donde las especies depredadoras varían en el tiempo o en hábitats en losque pueden aparecer nuevos depredadores, y en aquellas especies que viven y se re-producen en diferentes tipos de hábitats, puesto que pueden encontrarse con un rangomuy amplio y diverso de depredadores. Así, la capacidad de aprender el riesgo queposeen nuevas especies depredadoras puede resultar muy ventajoso para los renacua-jos o los tritones. Incluso, en algunas ocasiones, podría ser más ventajoso que lacapacidad de reconocer de manera innata distintos tipos de depredadores (Gonzalo etal. 2007). Los resultados obtenidos hasta ahora sugieren que las especies de anfibioscon capacidad para aprender a reconocer nuevos depredadores acuáticos son aquellasque pasan mucho tiempo en el agua (como ocurre con los tritones), o aquellas especiesde anuros cuyos renacuajos pasan por largos periodos de crecimiento antes demetamorfosearse (Gonzalo et al. 2007).
¿Aprendizaje contra depredadores exóticos?
Actualmente, los depredadores introducidos están considerados como una de lascausas más importantes del declive de las especies, y de la pérdida de biodiversidad, alo largo de todo el mundo (Vitousek et al. 1997). Una revisión reciente (Salo et al. 2007)sugiere que los depredadores exóticos suponen impactos más severos en las pobla-ciones de presas nativas que los depredadores nativos. Así, en las comunidades dondelas presas han coexistido con los depredadores por largos periodos de tiempo evolu-tivo, estás han podido responder a la presión depredadora desarrollando comporta-mientos o morfologías que reducen las oportunidades de encontrarse con losdepredadores o aumentan las oportunidades de escapar de ellos al detectarlos (Limay Dill 1990). En contraste con esto, las comunidades de presas con depredadoresintroducidos no suelen tener experiencia con respecto a éstos y, por tanto, no poseencomportamientos de escape específicos para hacer frente a los nuevos depredadores.Esta inexperiencia de las presas puede facilitar en gran manera la eficacia en la cazade los depredadores introducidos, en comparación con la que presentan losdepredadores nativos.
Existen numerosos estudios en los que se ha documentado un declive en las pobla-ciones de anfibios nativos tras la introducción de especies de depredadores exóticos(por ej. Gamradt y Kats 1996; Goodsell y Kats 1999; Gillespie 2001, revisión en Kats y
16

Ferrer, 2003). Sin embargo, otros estudios han mostrado la ausencia de tales efectos(Kiesecker y Blaustein 1997). Según Chivers et al. (2001), esta falta de efectos negativospuede deberse, en parte, al éxito de las ranas al reconocer y escapar de los depredado-res introducidos. En el experimento de Chivers et al. (2001), individuos juveniles deRana aurora fueron capaces de reconocer señales químicas de la rana toro (Rana ca-
tesbiana), una especie depredadora introducida. No obstante, no pudo determinarsesi este reconocimiento por parte de los juveniles procedía de un proceso de aprendizajeo si estaba genéticamente determinado, puesto que los juveniles provenían de una po-blación sintópica con la rana toro. Aunque la capacidad de aprendizaje de nuevos de-predadores ha sido muy estudiada, se podría esperar que éste mecanismo fuera usadopor especies que utilizan las sustancias de alarma como indicadores de peligro poten-cial. Futuros estudios indicarán qué especies son capaces de aprender y cuales no. Éstasúltimas serán las especies más amenazadas por la introducción de nuevas especies dedepredadores.
Especie de Estudio
La rana verde Ibérica (Pelophylax perezi) es una especie endémica de la Península Ibérica yel sur de Francia, que se encuentra dentro del complejo sistema hibridogenético de las ranas ver-des europeas (Garcia-París 1997).
Es una especie muy abundante y estrictamente acuática (Lizana et al. 1989) que ocupa todotipo de ambientes (Malkmus, 1982; Meijide et al., 1994; Lizana et al. 1995; Malkmus 1997), en-contrándose tanto en medios lóticos (ríos, arroyos, ramblas, acequias), donde selecciona zonascon escasa corriente, como lénticos (charcas, balsas agrícolas, marjales, zanjas, embalses) (Graciay Pleguezuelos 1990; Pollo et al. 1998; Egea-Serrano et al. 2005). Existe cierto grado de segrega-ción espacial entre individuos adultos y juveniles ya que los subadultos ocupan charcastemporales de escasa profundidad y arroyos con corriente rápida, donde no suelen encontrarseadultos (Lizana et al. 1989). Por otra parte, se trata de una especie con escasos requerimientosecológicos (Llorente et al. 2002) y alta plasticidad ecológica, lo que la convierte en colonizadoratemprana de ambientes perturbados por acciones tales como minería o incendios (Sánchez yRubio 1996; Galán 1997a), así como de cuerpos de agua recientemente creados (Malkmus 1982).
La actividad reproductora es tardía y tiene lugar entre abril y finales de verano (Bea et al.1994), las larvas eclosionan a los 5-8 días de la puesta, y al cabo de de 3-4 días comienzan a nadarlibremente (Vidal 1966; Gonzalez de la Vega 1988) predominantemente en el fondo y en la co-lumna de agua (Díaz-Paniagua 1987). El estadio larvario es relativamente largo, teniendo lugarla metamorfosis a las 8-12 semanas de la puesta (Gonzalez de la Vega 1988), aunque existennumerosas larvas invernantes procedentes de puestas tardías que triplican el tamaño para elmismo estadio larvario y se metamorfosean al año siguiente (Hernandez y Seva 1986).
Las larvas son típicos habitantes del fondo de los cuerpos de agua (Díaz-Paniagua 1985).Dado el tardío período reproductor de la especie, ocupan masas de agua con característicaspropias del verano: menores dimensiones, menor cobertura vegetal, mayor temperatura ymenores concentraciones de oxígeno (Díaz-Paniagua 1983; Díaz-Paniagua 1988). Casi siempreocupan aguas relativamente profundas y permanentes (García-París 1989), aunque puedendesarrollarse también en charcas temporales. Son depredadas por anfibios (Pleurodeles waltl)(Santos et al. 1986), reptiles (Natrix maura, Mauremys leprosa, Emys orbicularis) (Santos yLlorente 1998; Santos et al. 2000; Gómez-Mestre y Keller 2003) y crustáceos (Cruz y Rebelo 2005;datos propios). La introducción de especies exóticas, como peces (Gambusia hobrooki, Carasius
auratus) o crustáceos (Procambarus clarkii), en cuerpos de agua donde la especie estaba presenteha conducido a la regresión de diversas poblaciones en la Península Ibérica (Galán 1997b;Martínez-Solano et al. 2003; Rodríguez et al. 2005).
17


Objetivos de la presente tesis:
El objetivo general de la presente tesis es estudiar y clarificar como actúan los me-canismos de aprendizaje (definido como el cambio en el patrón de comportamientoindividual basándose en la experiencia) para permitir a las presas enfrentarse acualquier peligro presente en el ambiente (incluso aquellos en principio novedosos odesconocidos). En particular, se quiere profundizar en la manera en que la presa ad-quiere información sobre la identidad y la peligrosidad de los depredadores y cómoajusta continuamente sus respuestas ante los riesgos existentes y la información reci-bida del medio.
La especie elegida para la realización de la tesis, la rana verde ibérica, presenta unascaracterísticas ideales para el estudio del aprendizaje como una respuesta adaptativaante la depredación: es un anfibio estrictamente acuático, con ubicuidad en el hábitat,una alta plasticidad ecológica y un largo periodo larvario. Estas características hacenque la detección de depredadores por señales químicas sea un mecanismo muy ven-tajoso para esta especie, y que sea proclive a presentar mecanismos de aprendizaje yaque se encuentran en ambientes altamente variables y con una gran diversidad depotenciales depredadores que varían a lo largo del tiempo y de los hábitats. En estatesis se explorará la capacidad de esta rana para tomar decisiones ajustadas de escapede acuerdo a sus experiencias recientes.
Capítulo 1: La hipótesis de la respuesta dependiente del nivel de riesgo de depredación(“threat-sensitive predator avoidance hypotesis”) nos dice que la intensidad de larespuesta antidepredatoria de un individuo debería ajustarse al riesgo de depredaciónal que está sometido (Helfman 1989). Así, las presas deberían evaluar el riesgoexistente y comportarse de un modo flexible dependiendo de su intensidad. En estecapítulo se examina la capacidad de los renacuajos de rana verde ibérica para evaluarel riesgo que supone un depredador conocido y actuar en consecuencia a su nivel depeligrosidad, al mismo tiempo que se comprueba la capacidad de aprendizaje queposeen estos renacuajos al presentárseles un posible nuevo depredador en conjuncióncon indicadores de alto riesgo de depredación.
Capítulo 2: El aprendizaje es el primer paso de la adquisición de memoria, que consisteen retener lo aprendido y la posibilidad de modificar la información gracias a nuevasexperiencias. La habilidad de memorizar nuevos depredadores puede ser muyventajosa para la supervivencia, sin embargo responder a todos las señales químicasporque accidentalmente han aparecido una vez mezcladas con señales de alarma puedeser muy costoso (Lima y Dill 1990). En este capítulo se examina la capacidad de reten-ción de la información que poseen los renacuajos de la rana verde ibérica y si la pérdidade la memoria es un mecanismo efectivo para evitar desencadenar comportamientosantidepredadores no adaptativos en presencia de especies no peligrosas con las quecomparten el hábitat.
Capítulo 3: Aparte de la pérdida de la memoria, existen dos mecanismos particular-mente relevantes para evitar situaciones en las que una señal química sea reconociday recordada como peligrosa. Ambos se basan en la detección previa de la señal sin estarunida a indicadores de alarma. En este capítulo se analiza la presencia en los renacuajos
19

de estos dos mecanismos: la inhibición latente y el aprendizaje de lo irrelevante y cómoson capaces de inhibir los procesos de aprendizaje de nuevos depredadores.
Capítulo 4: Existen dos sustancias químicas relacionadas con el riesgo de depredaciónemitidas por las presas, las sustancias de alarma y las de estrés. Tradicionalmentelas sustancias de alarma han sido consideradas indicadores de alto riesgo paralos individuos coespecíficos mientras que las sistancias de estrés conllevan un riesgomenor. En este capítulo examinamos si ambas sustancias provocan una reacción anti-depredatoria ajustada a su nivel de riesgo y si existen diferencias en el aprendizajemediado por ambas sustancias.
Capítulo 5: En los anfibios, la metamorfosis conlleva un cambio de requerimientosecológicos y una gran transformación tanto corporal como fisiológica. Los anfibiosmetamorfoseados generalmente utilizan señales visuales o auditivas para reconocera los depredadores, pero al escapar saltan y se esconden en el agua. En el medio acua-tico un individuo metamorfico capaz de retener la capacidad de los renacuajos paradetectar señales químicas sería muy ventajoso ya que le permitiria evaluar posiblesriesgos. Es este capítulo se examina la capacidad de individuos recién metamorfosea-dos para seleccionar un hábitat en función de las sustancias químicas presentes en él,ya sean de depredadores o de individuos coespecíficos.
20

REfEREnCiAS
Abrams PA (2000) The evolution of pre-dator-prey interactions: theory andevidence. Annu Rev Ecol Syst31:79–105
Acquistapace P, Hazlett BA, Gherardi F(2003) Unsuccessful predation andlearning of predator cues by cray-fish. J Crust Biol 23:364–370
Atema J, Stenzler D (1977) Alarm subs-tance of the marine mud snailNassarius obsoletus: biological cha-racterization and possible evolu-tion. J Chem Ecol 3:173–187
Barros M, Boere V, Mello EL Jr, Tomaz C(2002) Reactions to potential pre-dators in captive-born marmosets(Callithrix penicillata). Int J Prima-tol 23:443–454
Bea A, Montori A, Pascual X (1994) Her-petofauna dels Aiguamolls de l’Em-pordà. En: Els sistemas naturalsdels Aiguamolls de l’Empordà. Tre-balls de la Institució Catalana d’-Historia Natural 13:359–407
Bennet CH, Wills SJ, Oaakeshott SM, Mac-kintosh NJ (2000) Is the contextspecificity of latent inhibition a suf-ficient explanation of learned irre-levance? Q J Exp Psych B, CompPhysiol Psychol 53B:239–253
Berejikian BA, Smith RJF, Tezak EP,Schroder SL, Knudsen CM (1999)Chemical alarm signals and com-plex hatchery rearing hábitats af-fect antipredator behaviour andsurvival of chinook salmon (On-
corhynchus tshawytscha) juveniles.Can J Fish Aquat Sci 56:830–838
Blair HT, Schafe GE, Bauer EP, RodriguesSM, LeDoux JE (2001) Synapticplasticity in the lateral amygdala: Acellular hypothesis of fear conditio-ning. Learn Mem 8:229–242
Blum MS (1969) Alarm pheromones.Annu Rev Entomol 14:57–80
Blum MS (1985) Alarm pheromones.Kerkut, G. A. y Gilbert, L. I. (eds),Comparative insect physiology, bio-chemistry, and pharmacology. Vol.9. Pergamon Press, pp:193–224
Breden F, Scott M, Michel E (1987) Gene-tic differentiation for anti-predatorbehaviour in the Trinidadianguppy, Poecilia reticulate. AnimBehav 35:618–620
Brönmark C, Miner JG (1992) Predator-induced phenotypic change in bodymorphology in the crucian carp.Science 258:1348–1350
Brönmark C, Hansson L (2000) Chemicalcommunication in aquatic systems:an introduction. Oikos 88:103–109
Brown GE, Chivers DP (2005) Learningas an adaptive response to preda-tion. In: Ecology of predator/preyinteractions (Barbosa P, Castella-nos I eds). Oxford University Press,Oxford, pp:34–54
Brown GE, Smith RJF (1998) Acquiredpredator recognition in juvenilerainbow trout (Oncorhynchus
mykiss): Conditioning hatchery-re-ared fish to recognize chemicalcues of a predator. Can J Fish AquatSci. 55:611–617
Chivers DP, Mirza RS (2001) The impor-tance of predator-diet cues in theresponses of larval woodfrogs tofish and invertebrate predators. JChem Ecol 27:45–51
Chivers DP, Smith RJF (1994a) Fatheadminnows, Pimephales promelas, ac-quire predator recognition whenalarm substance is associated withthe sight of unfamiliar fish. AnimBehav 48:597–605
Chivers DP, Smith RJF (1994b) The roleof experience and chemical alarmsignaling in predator recognitionby fathead minnows, Pimephales
promelas. J Fish Biol 44:273–285
21

Chivers DP, Smith RJF (1998) Chemicalalarm signalling in aquatic preda-tor-prey systems: a review andprospectus. Écoscience 5:338–352
Chivers DP, Kiesecker JM, Anderson MT(1996a) Avoidance response of aterrestrial salamander (Ambystoma
macrodactylum) to chemical alarmcues. J Chem Ecol 22:1709–1716
Chivers DP, Wisenden BD, Smith RJF(1996b) Damselfly larvae learn torecognize predators from chemicalcues in the predator’s diet. AnimBehav 52:315–320
Chivers DP, Brown GE, Smith RJF (1995)Acquired recognition of chemicalstimuli from pike, Esox lucius, bybrook sticklebacks, Culaea incons-
tans (Osteichthyes, Gasterostei-dae). Ethology 99:234–242
Cruz MJ, Rebelo R (2005) Vulnerabilitiyof Southwest Iberian amphibians toan introduced crayfish, Procamba-
rus clarkii. Amphibia-Reptilia26:293–303
Davis M (2000) The role of the amygdalain conditioned and unconditionedfear and anxiety. In The amygdala.A functional analysis (Aggleton JPed) Oxford UP. NY pp:213–287
Darwin CR (1859) El origen de las espe-cies. Alianza Editorial S. A.
Díaz-Paniagua C (1983) Influencia de lascaracterísticas del medio acuáticosobre las poblaciones de larvas deanfibios en la Reserva Biológica deDoñana (Huelva, España). Doñana,Acta Vertebrata 10:41–53
Díaz-Paniagua C (1987) Estudio en cau-tividad de la actividad alimenticiade las larvas de siete especies deanuros. Rev Esp Herpetol 2:189–197
Díaz-Paniagua C (1988) Temporal segre-gation in larval amphibian commu-nities in temporary ponds at alocality in SW Spain. Amphibia-Reptilia 9:15–26
Dodson SI, Crowl TA, Peckarsky BL, KatsLB, Covich AP, Culp JM (1994) Non-visual communication in freshwa-ter benthos: an overview. J N AmBenthol Soc 13:268–282
Egea-Serrano A, Oliva-Paterna FJ, MiñanoP, Verdiell D, de Maya JA, Andreu A,Tejedo M, Torralva M (2005) Actua-lización de la distribución de losanfibios de la Región de Murcia (SEPenínsula Ibérica). Anales de Biolo-gía 27:53–62
Endler JA (1991) Interactions betweenpredators and prey. En: Behaviou-ral ecology. Third edition (Krebs JR,Davies NB eds) Blackwell Publis-hers. pp:169–196
Ferguson HJ, Cobey S, Smith BH (2001)Sensitivity to change in reward isheritable in the honeybee, Apis me-
llifera. Anim Behav 61:527–534Ferrari MCO, Trowell JJ, Brown GE, Chi-
vers DP (2005) The role of leaningin the development of threat-sensi-tive predator avoidance in fatheadminnows. Anim Behav 70:777–784
Ferrari MCO, Chivers DP (2006a) Lear-ning threat-sensitive predator avoi-dance: how do fathead minnowsincorporate conflicting informa-tion? Anim Behav 71:19–26
Ferrari MCO, Chivers DP (2006b) Therole of latent inhibition in acquiredpredator recognition by fatheadminnows. Can J Zool 84:505–509
Galán P (1997) Colonization of spoil ben-ches of an opencast lignite mine innorthwest Spain by amphibiansand reptiles. Biol Conserv 79:187–195
Galán P (1997b) Declive de poblacionesde anfibios en dos embalses de LaCoruña (Noroeste de España) porintroducción de especies exóticas.Bol Asoc Herpetol Esp 8:38–40
22

Gamradt SC, Kats LB (1996) Effect of in-troduced crayfish and mosquito-fish on California newts. ConservBiol 10:1155–1162
García-París M (1989) Guía para la iden-tificación de los renacuajos españo-les. Quercus 38:16–23
García-París M (1997) Rana perezi Seo-ane, 1885. (Gasc JP Ed).Atlas of am-phibians and reptiles in Europe.Muséum National d’Histoire Natu-relle, Paris. pp:152–153
Gelowitz CM, Mathis A, Smith RJF (1993)Chemosensory recognition of nor-thern pike (Esox lucius) by brookstickleback (Culaea inconstans):population differences and the in-fluence of predator diet. Behaviour127:106–118
Gillespie GR (2001) The role of introdu-ced trout in the decline of the spot-ted tree frog (Litoria spencieri).South-eastern Australia. Biol Con-serv 100:187–198
Gilliam JF, Fraser DF (2001) Movementin corridors: enhancement bypre-dation threat, disturbance andhabitat structure. Ecology 82:124–135
Gómez-Mestre I, Keller C (2003) Experi-mental assessement of turtle pre-dation on larval anurans. Copeia2003:349–356
Gonzalez de la Vega JP (1988) Anfibios yreptiles de la provincia de Huelva.Ertisa, Huelva.
Gonzalo A, López P, Martín J (2007) Ibe-rian green frog tadpoles may learnto recognize novel predators fromchemical alarm cues of conspeci-fics. Anim Behav 74:447–453
Goodsell, JA, Kats LB (1999) Effect of in-troduced mosquitofish on Pacifictreefrogs and the role of alternativeprey. Conserv Biol 13:921–924
Gotceitas V, Godin JGJ (1991) Foragingunder the risk of predation in juve-nile Atlantic salmon (Salmo salar
L.): effects of social status and hun-ger. Behav Ecol Sociobiol 29:255–261
Goth A (2001) Innate predator-recogni-tion in Australian brush-turkey(Alectura lathami, Megapodiidae)hatchlings. Behaviour 138:117–136
Gracia P, Pleguezuelos JM (1990) Distri-bución de los anfibios en la provin-cia de Granada (SE PenínsulaIbérica). Anales de Biología 16:71–84
Griffin A.S, Evans CS, Blumstein DT(2001) Learning specificity in ac-quired predator recognition. AnimBehav 62:577–589
Hazlett BA (1985) Disturbance phero-mones in the crayfish Orconectes vi-
rilis. J Chem Ecol 11:1695–1711Hazlett BA (1989) Additional sources of
disturbance pheromone affectingthe crayfish Orconectes virilis. JChem Ecol 15:381–385
Hazlett BA (1990a) Source and nature ofdisturbance-chemical system incrayfish. J Chem Ecol 16:2263–2275
Hazlett BA (1990b) Disturbance phero-mone in the hermit crab Calcinus
lavimanus (Randall, 1840). Crusta-ceana 58:314–316
Hazlett BA (1994) Alarm responses inthe crayfish Orconectes virilis andOrconectes propinquus. J Chem Ecol20:1525–1535
Hazlett BA (2003) Predator recognitionand learned irrelevance in the cray-fish Orconectes virilis. Ethology109:765–780
Hazlett BA, Acquistapace P, Gherardi F(2002) Differences in memory ca-pabilities in invasive and nativecrayfish. J Crustacean Biol 22:439–448
23

Hawkins LA, Magurran AE, Armstrong JD(2004) Innate predator recognitionin newly-hatched Atlantic salmon.Behaviour 141:1249–1262
Helfman GS (1989) Threat-sensitive pre-dator avoidance in damselfish-trumpetfish interactions. BehavEcol Sociobiol 24:47–58
Hernandez A, Seva E (1986) Datos preli-minares sobre la alimentación derana común (Rana perezi) en laprovincia de Alicante. Ayudas a lainvestigación 1984 3:37–46
Hileman KS, Brodie ED Jr (1994) Survivalstrategies of the salamander Des-
mognathus ochrophaeus: interac-tion of predator-avoidance andanti-predator mechanisms. AnimBehav 47:1–6
Hokit DG, Blaustein AR (1995). Predatoravoidance and alarm-response be-haviour in kin-discriminating tad-poles (Rana cascadae). Ethology101:280–290
Howe NR, Harris LG (1978) Transfer ofthe sea anemone pheromone, an-thopleurine by the nudibranch Aeo-
lida papillosa. J Chem Ecol4:551–556
Huryn AD, Chivers DP (1999) Contras-ting behavioral responses by detri-tivorous and predatory mayflies tochemicals released by injured cons-pecifics and their predators. J ChemEcol 25:2729–2740
Ichinose K (2002) Influence of age andbody size on alarm responses infreshwater snail Pomacea canalicu-
lata. J Chem Ecol 28:2017–2028Jacobsen HP, Stabell OB (1999) Predator-
induced alarm responses in thecommon periwinkle, Littorina litto-
rea: dependence on season, lightcondition, and chemical labelling ofpredators. Mar Biol 134:551–557
Kats LB, Dill LM (1998) The scent ofdeath: chemosensory assessmentof predation risk by prey animals.
Écoscience 5:361–394Kats LB, Ferrer RP (2003) Alien preda-
tors and amphibian declines: re-view of two decades of science andthe transition to conservation. Di-vers Distrib 9:99–110
Kats LB, Petranka JW, Sih A (1988) Anti-predator defenses and the persis-tence of amphibian larvae withfishes. Ecology 69:1865–1870
Kiesecker JM, Blaustein AR (1997) Popu-lation differences in responses ofred-legged frogs (Rana aurora) tointroduced bullfrogs. Ecology78:1752–1760
Kiesecker JM, Chivers DP, Marco A, Quil-chano C, Anderson MT, BlausteinAR (1999) Identification of a dis-turbance signal in larval red-leggedfrogs, Rana aurora. Anim Behav57:1295–1300
Larsson P, Dodson SI (1993) Chemicalcommunication in planktonic ani-mals. Arch Hydrobiol 129:129–155
Laurila A, Kujasalo J, Ranta E (1997) Di-ferent antipredator behaviour intwo anuran tadpoles: efects of pre-dator diet. Behav Ecol Sociobiol40:329–336
Laurila A, Kujasalo J, Ranta E (1998) Pre-dator-induced changes in life his-tory in two anuran tadpoles: effectsof predator diet. Oikos 83:307–317
Lawer SP (1989) Behavioural responsesto predator and predation risk infour species of larval anurans. AnimBehav 38:1039–1047
Lima SL (1998) Stress and decision ma-king under the risk of predation:recent developments from beha-vioral, reproductive, and ecologicalperspectives. Advances in the studyof behaviour, vol. 27: Stress and be-haviour (Møller AP, Milinski M, Sla-ter PJB eds) Academic Press.pp:215–290
24

Lima SL, Dill LM (1990) Behavioral deci-sions made under the risk of preda-tion: a review and prospectus. CanJ Zool 68:619–640
Lizana M, Arco C, del Morales JJ, Bosch J,Cejudo D, López FJ, Gutiérrez J,Martín R (1995) Atlas provisionalde la herpetofauna en el SistemaCentral segoviano. Rev Esp Herpe-tol 9:113–132
Lizana M, Ciudad MJ, Pérez-Mellado V(1989) Actividad, reproducción yuso del espacio en una comunidadde anfibios. Treballs de la SocietatCatalana d´Ictiologia i Herpetología2:92–127
Llorente GA, Montori A, Carretero MA,Santos X (2002) Rana perezi. (Ple-guezuelos JM, Márquez R, Lizana MEds). Atlas y Libro Rojo de los anfi-bios y reptiles de España. DirecciónGeneral de la Conservación de laNaturaleza-Asociación Herpetoló-gica Española. Madrid. pp:26–128
Machado G, Bonato V, Oliveira PS (2002)Alarm communication: a new func-tion for the scent-gland secretion inharvestmen (Arachnida: Opiliones)Naturwissenschaften 89:357–360
Magurran AE (1989) Acquired recogni-tion of predator odour in the Euro-pean minnow (Phoxinus phoxinus).Ethology 82:216–223
Malkmus R (1982) Beitrag zur Verbrei-tung der Amphibien und Reptilienin Portugal. Salamandra 18:218–299
Malkmus R (1997) Verbreitung und Bio-topwahl des Iberischen Wasser-frosches Rana perezi Seoane 1885auf Madeira, den Azoren und denKanaren. Nachr naturwiss Mus As-chaffenburg 104:65–69
Maloney RF, McLean IG (1995) Historicaland experimental learned predatorrecognition in free-living New Zea-land robins. Anim Behav 50:1193–1201
Manzoli-Palma MF, Gobbi N, Palma MS(1998) Alarm pheromones and theinfluence of pupal odour on the ag-gressiveness of Polybia paulista
(Ihering) (Hymenoptera: Vespi-dae). J Venom Anim Toxines 4:61–69
Maren S (2001) Neurobiology of Pavlo-vian fear conditioning. Annu RevNeurosci 24:897–931
Martínez-Solano I, Barbadillo LJ, LapeñaM (2003) Effect of introduced fishon amphibian species richness anddensities at a montane assemblagein the Sierra de Neila, Spain. Herpe-tol J 13:176–173
Mathis A, Smith RJF (1993a) Intraspecificand cross-superorder responses tochemical alarm signals by brookstickleback. Ecology 74:2395–2404
Mathis A, Smith RJF (1993b) Fatheadminnows, Pimephales promelas,learn to recognize northern pike.Esox lucius, as predators on thebasis of chemical stimuli from min-nows in the pike’s diet. Anim Behav46:645–656
Mathis A, Murray KL, Hickman CR (2003)Do experience and body size play arole in responses of larval ringedsalamanders, Ambystoma annula-
tum, to predator kairomones? La-boratory and field assays. Ethology109:159–170
McLean IG, Holzer C, Studholme BJS(1999) Teaching predator-recogni-tion to a naive bird: Implicationsfor management. Biol Conserv87:123–130
McLean IG, Schmitt MT, Jarmam PJ, Dun-can C, Wynne CDL (2000) Learningfor life: training marsupials to re-cognize introduced predators. Be-haviour 137:1361–1376
Meijide M, Meijide F, Arribas O (1994)Atlas herpetológico de la provinciade Soria. Rev Esp Herpetol 8:45–58
25

Mirza RS, Chivers DP (2000) Predator-training enhances survival of brooktrout: evidence from laboratoryand field enclosure studies. Can JZool 78:2198–2208
Mirza RS, Ferrari MCO, Kiesecker JM, Chi-vers DP (2006) Responses ofAmerican toad tadpoles to preda-tion cues: behavioural responsethresholds, threat-sensitivity andacquired predation recognition. Be-haviour 143:877–889
Olson MH, Mittelbach GG, Osenberg CW(1995) Competition between pre-dator and prey: Resource-basedmechanisms and implications fromstage-structures dynamics. Ecology76:1758–1771
Pfeiffer W (1977) The distribution offright reaction and alarm substancecell in fishes. Copeia 1977:653–665
Pollo JC, Velasco-Marcos JC, González-Sanchez N (1998) La fauna herpe-tológica del espacio natural de losArribes del Duero. Bol. Asoc Herpe-tol Esp 9:4–10
Rochette R, Arsenault DJ, Justome B,Himmelman JH (1998) Chemically-mediated predator recognition le-arning in a marine gastropod.Ecoscience 5:353–360
Rodríguez CF, Bécares E, Fernández-Aláez M, Fernández-Aláez C (2005)Loss of diversity and degradation ofwetlands as a result of introducingexotic crayfish. Biol Inv 7:75–85
Rottmann SJ, Snowdon CT (1972) De-monstration and analysis of analarm pheromone in mice. J CompPhysiol Psychol 81:483–490
Salo P, Korpimäki E, Banks PB, Nords-tröm M, Dickman CR (2007) Alienpredators are more dangerous thannative predators to prey popula-tions. Proc R Soc B 274:1237–1243
Sánchez J, Rubio JL (1996) Atlas prelimi-nar de los anfibios y reptiles de lasSierras Prebéticas albacetenses. Al-
Basit 38:5–30Santos FJ, Salvador A, García C (1986)
Dieta de las larvas de Pleurodeles
waltl y Triturus marmoratus (Am-phibia: Salamandridae) en simpa-tría en dos charcas temporales deLeón. Rev Esp Herpetol 1:293–313
Santos X, González-Solís J, Llorente GA(2000) Variation in the diet of tehviperine snake, Natrix maura, inreltion to prey availability. Eco-graphy 23:185–192
Santos X, Llorente GA (1998) Sexual andsize-related differences in the dietof the snake Natrix maura from theEbro Delta (Spain). Herpetol J8:161–165
Schafe GE, Nader K, Blair HT, LeDoux JE(2001) Memory consolidation ofPavlovian fear conditioning: A ce-llular and molecular perspective.Trends Neurosci 24:540–546
Semlitsch RD, Ryer HU (1992) Modifica-tion of anti-predator behaviour intadpoles by environmental condi-tioning. J Anim Ecol 61:353–360
Shettleworth SJ (1998) Cognition, evolu-tion and behaviour. Oxford-Univer-sity Press, New York
Sih A. (1980) Optimal behavior: can fora-gers balance two conflicting de-mands? Science 210:1041–1043
Sih A (1987) Predators and prey lifes-tyles: an evolutionary and ecologi-cal overview. Predation: direct andindirect impacts on aquatic com-munities (Kerfoot WC Sih A eds)University Press of New England.pp:203–224
Sih A (1992) Integrative approaches tothe study of predation: generalthoughts and a case study on sun-fish and salamander system. AnnZool Fennici 29:183–198
Sih A, Ziemba R, Harding KC (2000) Newinsights on how temporal variationin predation risk shapes prey beha-viour. Trends Ecol Evol 15:3–4
26

Smith RJF (1992) Alarm signals in fishes.Rev Fish Biol Fish 2:33–63
Stabell OB, Lwin MS (1997) Predator-in-duced phenotypic changes in cru-cian carp are caused by chemicalsignals from conspecifics. Env BiolFishes 49:145–149
Suboski MD, Bain S, Carty AE, McQuoidLM, Seelen MI, Seifert M (1990)Alarm reaction in acquisition andsocial transmission of simulatedpredator recognition by zebradanio fish (Brachydanio rerio). JComp Psych 104:101–112
Teerling CR, Pierce HD Jr, Bordan JH et al.(1993) Identification and bioacti-vity of alarm pheromone in thewestern flower trips, Frankliniella
accidentalis. J Chem Ecol 19:681–697
Turner AM, Turner SE, Lappi HM (2005)Learning, memory and predatoravoidance by freshwater snails: ef-fects of experience on predator re-cognition and defensive strategy.Anim Behav 72:1443–1450
Veen T, Richardson DS, Blaakmeer K,Komdeur J (2000) Experimentalevidence for innate predator recog-nition in the Seychelles warbler.Proc Roy Soc Lond B 267:2253–2258
Vidal A (1966) Estudio biológico de lasislas Pitiusas: Anfibios. Publicacio-nes del Instituto de Biología Apli-cada 40:81–112
Vitousek PM, Mooney HA, Lubchenco J,Melillo JM (1997) Human domina-tion of Earth’s ecosystems. Science277:494–499
Von Frisch K (1938) Zur psychologie desfische-schwarmes. Naturwissens-chaften 26:601–606
Warkentin KM (1995) Adaptive plasti-city in hatching age: a response topredation risk trade-offs. Proc NatlAcad Sci Unit States Am 92:3507–3510
Warkentin KM (1999) Effects of hatchingage on development and hatchlingmorphology in the red-eyed tree-frog, Agalychnis callidryas. Biol JLinn Soc 68:443–470
Warkentin KM (2000) Wasp predationand wasp-induced hatching of red-eyed treefrog. Anim Behav 60:503–510
Weber A. (2003) More than one ‘fish kai-romone’? Perch and sticklebackkairomones affect Daphnia life his-tory traits differently. Hydrobiolo-gia 498:143–150
Weldon PJ (1980) In defense of “kairo-mone” as a class of chemical relea-sing stimuli. J Chem Ecol6:719–725
Werner EE, Anholt BR (1996) Predator-induced behavioural indirect ef-fects: consequences to competitiveinteractions in anuran larvae. Eco-logy 77:157–169
Werner EE, Gilliam JF (1984) Theontogenetic niche and speciesinteractions in size-structured po-pulations. Annu Rev Ecol Syst15:393–425
Werner EE, Peacor SD (2003) A reviewof trait-mediated indirect interac-tions. Ecology 84:1083–1100
Wilson DJ, Lefcort H (1993) The effect ofpredator diet on the alarm res-ponse of red-legged frog Rana au-
rora, tadpoles. Anim Behav46:1017–1019
Wisenden BD (2000) Scents of danger:the evolution of olfactory orna-mentation in chemically-mediatedpredator-prey interactions. Animalsignals: signaling and signal designin animal communication. (Es-pmark Y, Amundsen T, RosenqvistG eds) Tapir Academic Press,Trondheim, Norway. pp:365–386
27

Wisenden BD, Chivers DP, SmithRJF (1995) Early warning in thepredation sequence: a disturbancepheromone in Iowa darters (Ethe-
ostoma exile). J Chem Ecol21:1469–1480
Wisenden BD, Chivers DP, Smith RJF(1997) Learned recognition of pre-dation risk by Enallagma damselfly
larvae (Odonata, Zygoptera) on thebasis of chemical cues. J Chem Ecol23:137–151
Wisenden BD, Harter KR (2001) Motion,not shape, facilitates association ofpredation risk with novel objectsby fathead minnows (Pimephales
promelas). Ethology 107:357–364
Wisenden BD, Millard MC (2001) Aqua-tic flatworms use chemical cuesfrom injured conspecifics to assesspredation risk and to associate riskwith novel cues. Anim Behav62:761–766
Woody DR, Mathis A (1998) Acquired re-cognition of chemical stimuli froman unfamiliar predator: Associativelearning by adult newts,Notophthalmus viridescens. Copeia1998:1027–1031
28

cAPÍTuLO i
LOS RENACUAJOS DE RANA VERDE IBÉRICA
PUEDEN APRENDER A RECONOCER NUEVOS
DEPREDADORES MEDIANTE LAS SEñALES DE
ALARMA DE INDIVIDUOS COESPECíFICOS
IBERIAN GREEN FROG TADPOLES MAY LEARNTO RECOGNIZE NOVEL PREDATORS FROMCHEMICAL ALARM CUES OF CONSPECIFICS


En este trabajo hemos examinado la habilidad de los renacuajosde rana verde ibérica (Pelophylax perezi) para evaluar la magnitud delriesgo de depredación y ajustar su comportamiento utilizando señalesquímicas de una serpiente depredadora, cuando esta señal química se lespresentaba sola o mezclada con sustancia de alarma de individuos coes-pecíficos. Así, Los renacuajos expuestos a las señales de alarma junto conlas señales químicas de la serpiente redujeron sus movimientos conside-rablemente más que cuando se les presentaban solo las señales químicasde la serpiente. También examinamos si los renacuajos eran capaces deasociar señales químicas nuevas y desconocidas para ellos (es decir, deun pez exótico no peligroso) con un riesgo de depredación después de laexposición simultánea de estas con sustancia de alarma de individuos co-específicos. Los renacuajos expuestos al las señales químicas del pezinofensivo en conjunto con la sustancia de alarma redujeron sus nivelesde actividad, y posteriormente también redujeron su actividad en unaexposición posterior sólo a las señales químicas del pez, de una manerasimilar a como la redujeron en presencia de las señales químicas dela serpiente. Por lo tanto, los renacuajos aprendieron a reconocer lasseñales químicas del pez como peligrosas cuando previamente habíansido asociadas con sustancia de alarma de individuos coespecíficos. Lahabilidad de aprender a reconocer depredadores puede ser particular-mente ventajosa para organismos que viven en ambientes con una altadiversidad de depredadores, incluso especies exóticas introducidas quepueden llegar a afectar a su supervivencia.
Este cápitulo se corresponde con: Gonzalo A, Lopez P, Martín J (2007) Iberian green frog tadpoles maylearn to recognize novel predators from chemical alarm cues of conspecifics. Anim Behav 74:447–453


IberIan green frog tadpoles may learn to recognIze novel
predators from chemIcal alarm cues of conspecIfIcs
Many antipredator adaptations are induced by the prey’s ability to recognise chemical
cues from predators and to act according to the threat level posed by that predator.
However, predator recognition often requires learning by prey individuals. We tested the
ability of Iberian green frog (Pelophylax perezi) tadpoles to assess the magnitude of
predation risk and adjust their behaviour by using perceived cues from a predatory snake,
when this stimulus was found alone or associated with chemical alarm cues from
conspecific tadpoles. Tadpoles exposed to alarm cues and the predatory snake scent
together reduced their movement rates to a greater extent than when the snake scent
was found alone, and reduced movement even more in the subsequent exposure to the
predator snake scent alone. We also tested whether tadpoles were able to associate novel
chemical cues (i.e. from an exotic non-predatory fish) with predation risk after a simul-
taneous exposure with conspecific alarm cues. Tadpoles exposed to non-predatory fish
cues and alarm cues together reduced their activity levels, and reduced activity in the
subsequent exposure to the fish cue alone, in a similar way as they reduced movement in
presence of predatory snake cues. Therefore, tadpoles learnt to perceive the fish cues as
risky when these were previously associated with alarm cues. Predator recognition
learning ability may be particularly advantageous for organisms whose environment
may have a wide range of types of predators, even new exotic introduced species of
predators that can affect the survival of prey.
IntroductIon
An important component of antipre-dator behaviour is the ability to detectand recognize predators (Lima and Dill1990). Many antipredator adaptationsare induced or mediated by the prey’sability to recognise chemical cues frompredators (Kats and Dill 1998). Manyaquatic animals, including some inverte-brates, fishes and amphibians, use che-mical cues to assess predation risk (e.g.Von Frisch 1938; Petranka et al. 1987;Dodson et al. 1994; Kiesecker et al. 1996;Chivers and Smith 1998). Chemical cuesmay arise from the predators, but oftenthey may be released by prey animalswhen they are captured by a predator(i.e. alarm cues) (Chivers and Smith1998; Kats and Dill 1998), which serve asa reliable and imminent indicator of risk
of risk for conspecifics (Chivers andSmith 1998).
Prey animals can reduce the probabi-lity of being captured by a predator byaltering their behaviour after detectingcues that indicate increased predationrisk (reviewed by Kats and Dill 1998).Thus, a higher responsiveness to preda-tor or alarm cues may increase prey sur-vival probabilities (Downes 2002).However, according to the threat-sensi-tive hypothesis (Helfman 1989), naturalselection should favour individuals thattake action appropriate to the magnitudeof threat, which would require an accu-rate discrimination of the current level ofrisk that each predator poses. Thus, theresponse of prey to predators may becontext dependent (Maerz et al. 2001).
33

For example, many prey species onlyrespond to chemical cues of a predatorwhen the predator is fed a diet that con-tains conspecifics (e.g. Mathis and Smith1993; Wilson and Lefcort 1993; Chiverset al. 1996).
In some cases, predator recognitionrequires learning by prey individuals(e.g. fishes: Mathis and Smith 1993; dam-selfly larvae: Chivers et al. 1996; crayfish:Hazlett 2003). Individuals must expe-rience simultaneously a predator cue anda danger cue, such as an alarm chemicalcue released by a crushed conspecific,before the predator cue is considered asa danger signal. The ability to acquire re-cognition of predation risk is of obviousfitness benefit. Chemical alarm signalshave been shown to be important in faci-litating learned recognition of predationrisk by prey animals such as fishes (Göz1941, Magurran 1989, Chivers and Smith1994, 1998; Mathis and Smith 1993; Lar-son and McCormick 2005) or adult newts(Woody and Mathis 1998). Releaser-induced recognition learning involvesthe simultaneous exposure to aversivestimulus and a neutral stimulus causinglearned aversion to the neutral stimuli(Yunker et al. 1999). The result of this le-arning mechanism is acquired predatorrecognition in which predator naïve indi-viduals show appropriate antipredatorbehaviour to the cue of a potential pre-dator even though they have had no di-rect exposure to the predator. Severalauthors have demonstrated such acqui-red predator recognition in fishes by pai-ring alarm cues with the visual orchemical cues of a predator (e.g. Chiversand Smith 1994, 1995; Larson andMcCormick 2005). Therefore, the abilityof prey to learn to recognize a novelpredator should minimize the prey’s riskof capture.
Predator recognition abilities of preyanimals may have implications for cu-rrent conservation issues. For example,
although the effects of introduced preda-tors on native species are complex(Kiesecker and Blaustein 1997, 1998),many amphibian populations havedeclined after the introduction of exoticpredator species (Kiesecker and Blaus-tein 1998; Kupferburg 1997). Tadpoleshave evolved a number of behaviouraland morphological adaptations to sur-vive and coexist with their naturalpredators, but those antidepredator me-chanisms that exist for native predatorsmay not be sufficient to allow coexis-tence with introduced predator species(Gamradt and Kats 1996). Thus, severalstudies have found that amphibian popu-lations were affected by new introducedpredators (Goodsell and Kats 1999;Adams 1999; Kiesecker and Blaustein1997; Knapp and Matthews 2000;Murray et al. 2004), but not all amphi-bian species were negatively affected(Hecnar and M´Closkey 1997). Recently,Bosch et al. (2006) have found that Rana
iberica tadpoles could detect chemicalcues from both native and exotic troutspecies and reacted by decreasing theiractivity, although the response towardnative predators was stronger than theresponse toward exotic trout. The au-thors suggested that these antipredatorbehavioural responses were inefficientagainst the introduced trout, but did notreveal the origin of this antipredator be-haviour in response to exotic predators.We hypothesised that tadpoles respon-ses might be elicited because exotic troutreleased chemicals cues similar to thosereleased by native trout, but it alsoremains possible that R. iberica tadpoleswere able to learn recognition of newpotential predators.
In this study, we examined whethertadpoles of the Iberian green frogs (P.
perezi) can use chemical cues of preda-tors and/or alarm substances releasedfrom conspecifics to adjust their beha-viour in response to the perceived
34

predation risk. Furthermore, we aimedto determine whether tadpoles can learnto associate chemical cues to which tad-poles can not be genetically predisposed(e.g. those from non-predatory exotic fishspecies) with predation risk throughtheir association with the simultaneouspresence of alarm cues of conspecifics.
methodsstudy animals
We collected 63 Iberian green frogtadpoles (SVL, Mean±SE=1.3±1.2 cm,Gosner’s stage: 24; see Gosner 1960) bynetting during July 2005 at several smallponds in Collado Mediano (Madrid,central Spain). Tadpoles were housedindividually at “El Ventorrillo” FieldStation, 10 km from the capture area, inplastic aquaria (18 x 25 cm and 10 cmhigh) with water at ambient temperatureand under a natural photoperiod. Theywere fed every two days with commer-cial fish flakes.
We also captured in a larger pond atthe same locality two Viperine snakes,Natrix maura, to be used as native preda-tor scent donors. This snake is predomi-nantly aquatic and mainly feeds onamphibians, both larvae and adults, andfishes (Haley and Davies 1986; Braña1998). Snakes were housed individuallyin plastic cages (36 x 25 cm and 13 cmhigh) containing sawdust and tree barkfor cover and a pond with water (10 cmdiameter). The snakes’ cages were placedin a different room than the tadpoles’aquaria to avoid contact with the scentand visual stimuli before they were tes-ted. To avoid potential confoundingeffects of the diet on the results, all sna-kes were fed small pieces of commercialfreshwater fishes, obtained from a fish-market, for 3 weeks before collectingtheir chemical stimuli.
We obtained from a commercialdealer non predatory zebra danio fish
(Brachyodanio rerio) to be used as sourceof neutral scent. Before and after the ex-periment finished, fishes were maintai-ned in a large filtered aquarium andregularly fed with commercial fish flakes.
All the animals were healthy duringthe trials, all maintained or increasedtheir original body mass, and all tadpolesmetamorphosed into subadult frogs.These frogs and the snakes were retur-ned to their exact capture site. The expe-riments were performed under licensefrom the “Consejería de Medioambientede la Comunidad de Madrid” (the Envi-ronmental Agency of the local Govern-ment of Madrid). Procedures areconformed to recommended guidelinesfor use of live Amphibians in laboratoryresearch (ASIH 2004).
preparation of chemical stimuli
Alarm cues of tadpoles were preparedfrom three tadpoles (SVL, Mean±SE=1.2±0.1 cm). They were cold anes-thetised by placing at 4 °C for 20 min,inducing them deep hypothermia, and,then, euthanasied with a quick blow tothe head to avoid suffering (ASIH 2004).We did not use a chemical anaesthetic,because these chemicals may interferewith natural tadpoles’ chemical cues insubsequent trials. The extract was thenprepared by putting these dead tadpolesin a clean disposable plastic dish, andmacerating them in 600 mL of distilledwater. The stimulus water was then filte-red through absorbent paper to removesolid particles, and immediately frozenin 10 mL portions until used (Woody andMathis 1998).
The snake scent was prepared byplacing the snakes individually in cages(36 x 25 cm and 13 cm high) containing500 mL of clean water and left overnight.Then, we extracted and mixed the water,and frozen it in 10 mL portions until use.Clean water was collected from a nearby
35

high mountain spring that did not housefrogs nor fishes nor snakes.
The neutral stimulus was prepared byplacing zebra danio fishes in groups ofthree into a 3 L aquariumthree into a 3 Laquarium with clean water for 3 days.These aquaria were aerated but not filte-red. Fishes were not fed during this shortperiod to avoid contaminating waterwith food odour. Thereafter, water wasdrawn from the aquaria and frozen in 10mL portions until its use in experiments.Fishes were returned and fed in theirhome large aquaria. We prepared controlwater in an identical manner but withoutplacing fishes or snakes in the aquaria(Woody and Mathis 1998).
experimental design
We planned an experiment withsequential determined trials to conditionthe tadpoles, and tried to determinewhether frog tadpoles were able toassess predation risk and to learn recog-nition of novel predators. Between trialswe allowed the tadpoles to rest for oneday to avoid stress. We randomly distri-buted the tadpoles in four differenttreatments of 15 tadpoles each, and con-ducted the experiment in two differentseries. In each trial single individualtadpoles were tested separately. Thus,different trials were considered as repli-cates of each treatment. In the first series(Series 1), on Day 1 (’response to controlclean water alone’) individual tadpoles(n=30) from two treatments (‘control’and ‘experimental’) were tested withclean water. The objective of this trialwas to determine the basal activity levelsof tadpoles in a predator-free environ-ment, and to use this number as a controlfor the effect of predator and alarm che-mical cues in further trials. On Day 3 (‘in-itial response to chemical cues’)individual tadpoles from the ‘control’treatment were exposed to the snake
predator chemical cues alone mixed withclean water. Individual tadpoles from the‘experimental’ treatment were exposedto both the scent of the predatory snakeand conspecific chemical alarm cues,thus, simulating the cues from a preda-tory snake that was eating a conspecifictadpole. This allowed us to measure theeffect of the alarm cues on tadpole beha-viour. The objective of this trial was todetermine whether frog tadpoles wouldreact to the paired presentation of cons-pecific alarm cues and predator chemicalcues. On Day 5 (‘conditioned response tothe predator stimulus’) individual tadpo-les from the two treatments were expo-sed to the predatory snake chemical cuesalone mixed with clean water. The objec-tive of this trial was to determine whe-ther tadpoles were able to adjust theirbehaviour accordingly to the predationrisk perceived in the previous trial. Wepredicted that tadpoles from the experi-mental treatment would have a greaterfright reaction than tadpoles from thecontrol treatment because the previoussimultaneous presentation of predatorand alarm cues would indicate that thepredator was more dangerous than whenthe predator cues were presented alone.
In the second series (Series 2), weused different individual tadpoles(n=30), to avoid previous experience. Wefollowed the same procedure as in pre-vious series; Day 1 (‘response to controlclean water alone’), Day 3 (‘initial res-ponse to chemical cues’), and Day 5(‘conditioned response to the non-preda-tory cues’). However, we used non-pre-datory fish scent instead of predatorysnake scent. The objective of this experi-ment was to determine whether tadpoleswould react to the paired presentation ofconspecific alarm cues and neutral che-mical cues from non-predatory fish (i.e.cues not previously associated withdanger). Such paired stimuli might beexperienced by a predator naïve prey
36

exposed simultaneously to conspecificalarm cues and the scent of a predator, orto chemical cues from a predator thatwas eating a conspecific. We aimed to de-termine whether tadpoles were able tolearn to recognize and associate the neu-tral cue with danger after the previousexposure.after the previous exposure.We predicted than only tadpoles of theexperimental treatment (i.e. conditionedwith conspecific alarm cue and the neu-tral cue) would respond with a fright re-action to the neutral cue alone. The twoseries were carried out in parallel, andobservations were carried out blind.
Tadpoles were tested individually ingrey, U-shaped, gutters (101 x 11.4 cmand 6.4 cm high) sealed at both ends withplastic caps. We marked the internal partof the gutters with four crossing linesthat created five subdivisions of equalsurface. We made rectangular releasecages (21x7.6 cm and 6.4 cm high) by se-wing to gether clear, perforated plasticnormally used for needlepoint (2 mmsquare holes), which were placed in themiddle of the central subdivisions. Wefilled each gutter with 3 L of clean waterfrom a mountain spring at 20 °C, and pla-ced clear plastic over each trough oneither side of the cage to isolate thesystem from air movements in the tes-ting room (see Rohr and Madison 2001).
We made different test solutions, 20mL each (2 ice aliquots), using combina-tions of clean water or water with alarmcues, and water with snake or fish scent,and a control treatment of clean wateralone. We assigned test solutions to oneend of each trough (right or left) by stra-tified randomization, and assigned 20mL of clean water (2 ice aliquots) to theopposite end.
We placed a single tadpole coveredwith a release cage in each gutter, andwaited 5 min for habituation. Then wedeposited the test solution ices and webegan trials by slowly lifting the cages
above each tadpole 5 min after we depo-sited the test solution ices aliquots (i.e.after the ices aliquots had entirelythawed). We subsequently stood asmotionless as possible recording from ahidden point the quadrant that eachtadpole occupied at 1 min intervals for30 min. We calculated levels of activityfrom the number of lines crossed by eachtadpole during the observation period(Rohr and Madison 2001). Diffusion ofchemicals in still water may be a slowprocess. However, all individual tadpolesused in the experiment were observed atleast once in all of the subdivisions of thegutter, so we were confident that all tad-poles were really exposed to the chemi-cal stimuli. Moreover, tadpoles oftenshowed episodes of fast swimmingwhich should contribute to diffuse che-micals in water.
Levels of activity (number of linescrossed) were log10 transformed andthen tested by general linear modelling(GLM) (Grafen and Hails 2002). We usedday of the ‘trial’ as a within variable, and‘predator’ (i.e. snake vs. fish) and ‘alarm’(i.e. control treatment with absence ofalarm cues vs. experimental treatmentwith presence of alarm cues) as catego-rical between variables. We included theinteractions between variables in themodel to test for the effects of the diffe-rent treatments (with or without alarmcues) depending on the type of predatorcues and the day of the trial. Subsequentpost hoc multiple comparisons weremade using Tukey’s pairwise compari-sons (Sokal and Rohlf 1995).
results
On average, tadpoles reduced theiractivity more with predatory snakechemical cues (Series 1) than with non-predatory fish chemical cues (Series 2)(‘predator’ effect; Table 1; Fig. 1). Also,control tadpoles were more active than
37

experimental tadpoles conditioned withconspecific alarm cues (‘alarm’ effect),and there were significant differencesbetween the three days of the experi-ments (‘trial’ effect); all tadpoles beingmore active the first day than the rest ofthe days (Table 1). However, all thetwo-way interactions between factorswere significant.
In Series 1 (Fig, 1a), activity levels onDay 1 (water alone) did not differ bet-ween control and experimental tadpoles(Tukey’s tests, P=0.60). On Day 3 (preda-tory snake chemical cues with or withoutalarm cues) tadpoles from the two treat-ments decreased their activity in relationto the previous day (P=0.001 in bothcases), but experimental tadpoles expo-sed to snake cues combined with alarmcues decreased significantly more theiractivity than control tadpoles exposed tosnake cues alone (P<0.0001). Finally, onDay 5 (snake cues alone) the activity ofcontrol tadpoles did not change with res-pect to their activity on Day 3 (P=0.90),but activity increased significantly inexperimental tadpoles (P=0.0001),
although it did not reach the activity le-vels of the control treatment on this Day5 (P=0.0001).
In Series 2 (Fig. 1b), activity levels onDay 1 (water alone) did not differ bet-ween the two treatments of tadpoles(Tukey’s tests, P=0.99). On Day 3 (fishchemical cues with or without alarmcues), activity decreased significantlyonly in experimental tadpoles exposed tofish cues combined with alarm cues(P=0.0001), but it did not change signifi-cantly in control tadpoles exposed to fishchemical cues alone (P=0.60). On Day 5(fish cues alone), activity of control tad-poles did not change with respect to theprevious days 1 and 3 (P=0.90), whereasactivity of experimental tadpoles increa-sed significantly with respect to the pre-vious day 3 (P=0.0001), but withoutreaching the activity levels of control tad-poles (P=0.009). In addition, there was ano significant three-way interaction bet-ween the factors and the trials, whichindicated that tadpoles reacted in a simi-lar way in the two series of the experi-ment, and showed that the alarm cue
table 1. Results of a GLM testing the effects of ‘predator’ type (snake vs. fish) and presence orabsence of ‘alarm’ chemical cues (between effects) on the activity levels of Iberian green frogtadpoles in the different days of the ‘trials’ (within effect) (see methods).
38
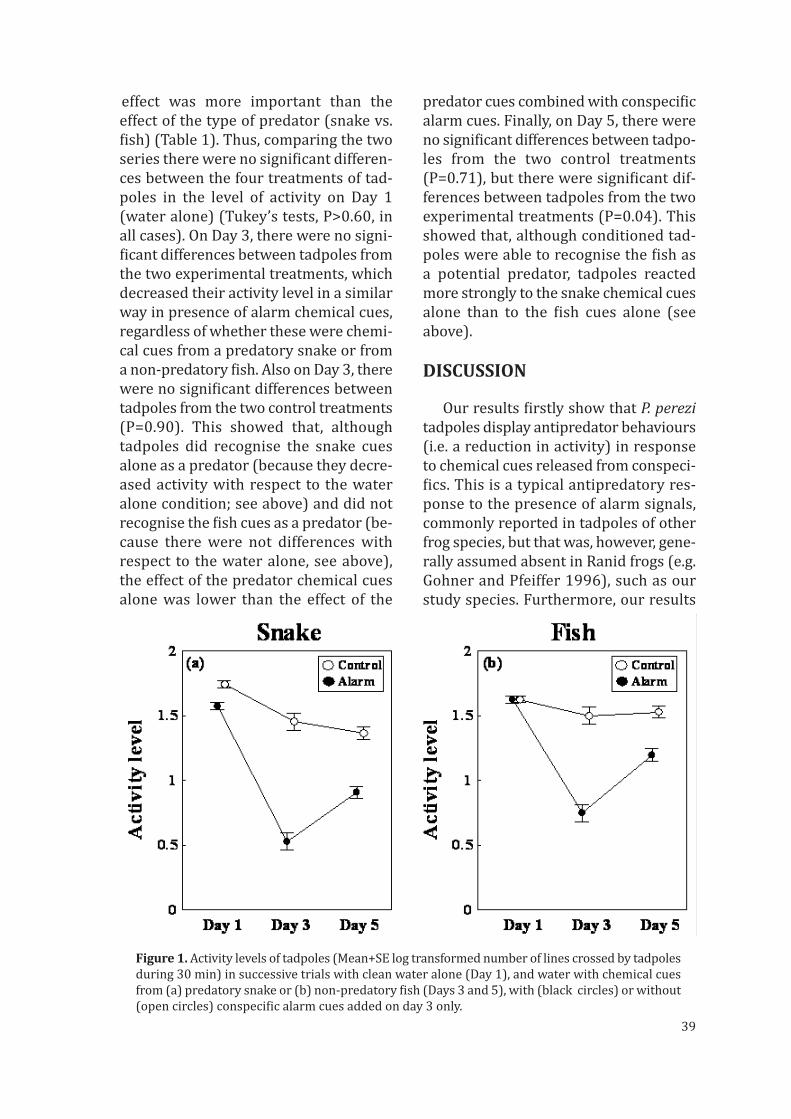
effect was more important than theeffect of the type of predator (snake vs.fish) (Table 1). Thus, comparing the twoseries there were no significant differen-ces between the four treatments of tad-poles in the level of activity on Day 1(water alone) (Tukey’s tests, P>0.60, inall cases). On Day 3, there were no signi-ficant differences between tadpoles fromthe two experimental treatments, whichdecreased their activity level in a similarway in presence of alarm chemical cues,regardless of whether these were chemi-cal cues from a predatory snake or froma non-predatory fish. Also on Day 3, therewere no significant differences betweentadpoles from the two control treatments(P=0.90). This showed that, althoughtadpoles did recognise the snake cuesalone as a predator (because they decre-ased activity with respect to the wateralone condition; see above) and did notrecognise the fish cues as a predator (be-cause there were not differences withrespect to the water alone, see above),the effect of the predator chemical cuesalone was lower than the effect of the
predator cues combined with conspecificalarm cues. Finally, on Day 5, there wereno significant differences between tadpo-les from the two control treatments(P=0.71), but there were significant dif-ferences between tadpoles from the twoexperimental treatments (P=0.04). Thisshowed that, although conditioned tad-poles were able to recognise the fish asa potential predator, tadpoles reactedmore strongly to the snake chemical cuesalone than to the fish cues alone (seeabove).
dIscussIon
Our results firstly show that P. perezi
tadpoles display antipredator behaviours(i.e. a reduction in activity) in responseto chemical cues released from conspeci-fics. This is a typical antipredatory res-ponse to the presence of alarm signals,commonly reported in tadpoles of otherfrog species, but that was, however, gene-rally assumed absent in Ranid frogs (e.g.Gohner and Pfeiffer 1996), such as ourstudy species. Furthermore, our results
figure 1. Activity levels of tadpoles (Mean+SE log transformed number of lines crossed by tadpolesduring 30 min) in successive trials with clean water alone (Day 1), and water with chemical cuesfrom (a) predatory snake or (b) non-predatory fish (Days 3 and 5), with (black circles) or without(open circles) conspecific alarm cues added on day 3 only.
39

indicated that P. perezi tadpoles wereable to modify their antipredatory beha-viour according to their previous expe-rience with chemical cues of thepredator. At the end of the Series 1, con-ditioned experimental tadpoles decrea-sed their activity more than controltadpoles when exposed to snake chemi-cal cues alone, after the previous presen-tation of snake chemical cues inconjunction with alarm cues. This solu-tion mix could simulate the chemicalcues released by a predator that was ea-ting a conspecific tadpole, and, thus, atadpole can attribute to that predator ahigher risk than to a potential predatorbut that was not actually attacking tad-poles. Thus, tadpoles seemed to assessthe magnitude of predation risk and ad-justed their behaviour by using perceivedcues that vary according to the simulatedpredator diet.
The results suggested that theresponse of frog tadpoles represented aform of threat-sensitivity (Helfman1989). According to the threat-sensitivityhypothesis, prey species should behaveflexibly towards a varying degree of pre-dator threat and, consequently, leavemore time for other activities when thethreat is low (Helfman 1989). The be-havioural response elicited by the pre-datory snake in control tadpoles wasweaker than that elicited by the samesnake in experimental tadpoles. This sug-gested that snakes were not perceived tobe very dangerous predators unless thetadpoles had been previously exposed tosnakes that “had eaten conspecifics” (i.e.mix of alarm cues plus snake chemicalcues). Thus, alarm cues marked snakes,and allowed tadpoles to recognize snakesas dangerous predator in future encoun-ters regardless of the snake’s recent diet.In our case, this snake species preysmainly on adult (or metamorphosed)frogs, although it can also preys on tad-poles in lower proportions (Braña 1998).
Thus, it is possible that tadpoles percei-ved this snake species not to be a verydangerous predator, unless they assessedthat this snake was actually preying ontadpoles.
Furthermore, tadpoles were able tobecome conditioned to recognize a noveland non-dangerous chemical cue asdangerous through associations of thiscue with conspecific chemical alarmcues. Thus, experimental tadpoles wereconditioned to recognize B. rerio (a non-predatory fish) as a predator after thesimultaneous exposure to conspecificalarm cues. As in the previous experi-ment, when we simulated a predatoreating tadpoles, the alarm cues markedthat “novel predator” as dangerous, andfish were recognized as predators insubsequent exposures.
Fish are well known for their ability toacquire recognition of novel stimuli asdangerous. A single, simultaneous expo-sure to conspecific alarm cues and anovel stimulus transfers risk to the novelstimulus whether or not it is a novel che-mical cue (e.g. Göz 1941; Chivers andSmith 1995; Suboski et al. 1990, reviewby Smith 1992, 1997). Fish learned to re-cognize and avoid predator chemicalcues after a single simultaneous encoun-ter with predator and conspecific alarmcues. The ability to acquire recognition ofnovel predators has also been found inplatyhelminths (Wisenden and Millard2001), crayfish (Hazlett 2003) and dam-selflies (Chivers et al. 1996). In amphi-bians we are only aware of one paperwhich found this ability in adults newtsNotophthalmus viridiscens (Woody andMathis 1998). These authors found thatnewts that spent all of their adult life inwater were able to associate chemicalalarm cues with chemicals from an unfa-miliar predator. Our results showed thatgreen frog tadpoles were able to learn asdangerous a chemical cue from a nonpredator fish. Therefore, this learning
40

mechanism may be especially importantto the survivorship of prey species thatare likely to find a high variety of preda-tors while they are in the aquatic phase.To learn to recognize novel predators alsomay be of particular importance to preyspecies that live in habitats where the pre-dator species vary across seasons orwhere new species of predators could ap-pear (such as introduced predators). Preyspecies that live and breed in differentkinds of aquatic habitats (such as greenfrogs) may experience a wide range oftypes of predators.Having the capacity oflearning about the actual danger of newspecies could be very advantageous fortheir tadpoles, even more than having agenetically determined capacity to recog-nize diverse types of predators.
Some studies have documented thedecline of native frogs following theintroduction of exotic predator species(e.g. Gamradt and Kats 1996; Goodsell andKats 1999; Gillespie 2001), but otherstudies reported a lack of effects (Kiesec-ker and Blaustein 1997). According toChivers et al. (2001) this may be in partdue to the success of these frogs in recog-nizing and avoiding introduced predators.In the experiment of Chivers et al. (2001),juvenile treefrogs were able to recognizechemicals from an introduced predatoryspecies (bullfrog), but it was not determi-ned whether treefrogs learnt that bull-frogs were a threat or whether therecognition was genetically determined,because treefrogs came from a populationsyntopic with bullfrogs. Thus, moststudies about new predator-speciesrecognition have ignored the mechanismswhich allow the recognition of a novelchemical stimulus as a potential predator.In our study, we found that this mecha-nism may be the association between theconspecific alarm cues with an unknownstimulus. Therefore, we could expect thatthis mechanism was also used in otherspecies which use alarm cues.
Our results suggested that learning torecognize novels predators could be apossible mechanism to face up exoticintroduced predators. The effects of intro-duced predators on native species arecomplex (Kiesecker and Blaustein 1997,1998). The ability of prey to recognize anintroduced predator should minimize theprey’s risk of capture (Kiesecker andBlaustein 1997; Chivers et al. 2001).However, recognition of the predator doesnot by itself imply that there will not besignificant predator effects, or the res-ponse elicited could be inefficient againstthe introduced species (Bosh et al. 2001).Our results showed that, althoughconditioned tadpoles were able to recog-nize the fish as a potential predator,tadpoles reacted more strongly to thesnake chemical cues than to the fish cues.This result may imply that tadpoles weremore sensitive to native predators than tonovel introduced ones. Recognition ofsnakes may be, thus, genetically determi-ned, but also it is likely that tadpoles mayrequire more frequent exposures to thenovel fish chemical cue combined withalarm cues to elicit a fright response simi-lar to that elicited from native predatorysnakes (reinforcement). Nevertheless,since we only tested one native and onenovel predator species, the differentresponses to snake and fish might bepredator-class or -species specific, and, forexample, the response to a novel snakemight be as great as for the native snakepredator. Further experiments that repli-cate predator types (i.e. multiple noveland native predators) are needed toexplain these differences.
Our knowledge of amphibian capacityfor learning recognition of new predatorsis poor. But, it seems an important mecha-nism to avoid new or introduced preda-tors. The results of Woody and Mathis(1998) and our own data suggest thatamphibian species with a higher capacityof learning recognition of new aquatic
41

predators may be those that spend a lotof time in the water, such as aquaticnewts or anuran species whose tadpoleshave long periods of growth before me-tamorphosis (e.g. Iberian green frogs).Further experiments are needed to as-certain which species are able to learnabout new predators, and which speciescan not, as these would be the speciesmost at risk from introduced predatorspecies.
references
Adams MJ (1999) Correlated factors in amphi-bian decline: exotic species and habitatchange in western Washington. J WildlManag 63:1162–1171
ASIH (2004) Guidelines for use of live Amphi-bians and Reptiles in field and laboratoryresearch. 2nd Ed. Herpetological AnimalCare and Use Committee (HACC) of theAmerican Society of Ichthyologists andHerpetologists. Lawrence, Kansas.
Braña F (1998) Natrix maura (Linnaeus, 1758).In: Fauna Ibérica, Vol. 10 (Ramos MA ed)Museo Nacional de Ciencias Naturales,Consejo Superior de Investigaciones Cien-tíficas Madrid pp. 440–454
Bosch J, Rincón PA, Boyero L, Martínez-Solano I(2006) Effects of introduced salmonids ona montane population of Iberian frogs.Conserv Biol 20:180–189
Chivers DP, Smith RJF (1994) The role of expe-rience and chemical alarm signalling inpredator recognition by fathead minnows,Pimephales promelas. J Fish Biol 44:273–285
Chivers DP, Smith RJF (1995) Free-living min-nows rapidly learn to recognize pike aspredators. J Fish Biol 46:949–954
Chivers DP, Smith RJF (1998) Chemical alarm sig-nalling in aquatic predator/prey interac-tions: a review and prospectus Ecoscience.5:338–352
Chivers DP, Wisenden BD, Smith RJF (1996) Dam-selfly larvae learn to recognize predatorsfrom chemical cues in the predator’s diet.Anim Behav 52:315–320
Chivers DP, Wildy EL, Kiesecker JM, Blaustein AR(2001) Avoidance response of juvenile pa-cific tree frogs to chemical cues of introdu-ced predatory Bullfrogs. J Chem Ecol27:1667–1676
Dodson S I, Crowl TA, Peckarsky BL, Kats LB, Co-vich AP, Culp JM (1994) Non-visual com-munication in freshwater benthos: anoverview. J N Am Bentho. Soc 13:268–282
Downes SJ (2002) Does responsiveness to preda-tor scents affect lizard survivorship?Behav Ecol Sociobiol 52:38–42
Gamradt SC, Kats LB (1996) Effect of introducedcrayfish and mosquitofish on Californianewts. Conserv Biol 10:1155–1162
Gillespie GR (2001) The role of introduced troutin the decline of the spotted tree frog (Li-
toria spencieri) in South-eastern Australia.Biol Conserv 100:187–198
Gohner M, Pfeiffer W (1996) The distribution ofthe Mauthner axons in fish and amphi-bians and its relation to the fright reactionin Ostariophysi and Anura. Rev Suisse Zool103:859–891
Goodsell JA, Kats LB (1999) Effect of introducedmosquitofish on Pacific treefrogs and therole of alternative prey. Conserv Biol13:921–924
Gosner KL (1960) A simplified table for staginganuran embryos and larvae with notes onidentification. Herpetologica 16:183–190
Göz H (1941) Über den art-und individualgeruchbei fischen. Zeitschrift für VergleichendePhysiologie. 29:1–45
Grafen A, Hails R (2002) Modern statistics for thelife sciences. Oxford: Oxford UniversityPress.
Haley A, Davies PMC (1986) Diet and foraging be-haviour of Natrix maura. Herpetological J1:53–61
Hazlett BA (2003) Predator recognition and lear-ned irrelevance in the crayfish Orconectes
virilis. Ethology 109:765–780Hecnar SJ, M'Closkey RT (1997) The effects of
predatory fish on amphibian species rich-ness and distribution. Biol Conserv79:123–131
Helfman GS (1989) Threat-sensitive predatoravoidance in damselfish-trumpetfishinteractions. Behav Eco. Sociobiol 24:47–58
Kats LB, Dill LM (1998) The scent of death: che-mosensory assessment of predation byprey animals. Ecoscience 5:361–394
Kiesecker JM, Blaustein AR (1997) Populationdifferences in responses of red-leggedfrogs (Rana aurora) to introduced bull-frogs. Ecology 78:1752–1760
42

Kiesecker JM, Blaustein AR (1998) Effects of in-troduced bullfrogs and smallmouth basson the microhabitat use, growth and sur-vival of native red-legged frogs. ConservBiol 12:776–787
Kiesecker JM, Chivers DP, Blaustein AR (1996)The use of chemical cues in predator re-cognition by western toad (Bufo boreas)tadpoles. Anim Behav 52:1237–1245
Knapp RA, Matthews KR (2000) Non-native fishintroductions and the decline of the moun-tain yellow-legged frog from within pro-tected areas. Conserv Biol 14:428–438
Kupferberg SJ (1997) Bullfrog (Rana catesbeiana)invasion of a California river: the role oflarval competition. Ecology 78:1736–1751
Larson JK, McCormick MI (2005) The role of che-mical alarm signals in facilitating learnedrecognition of novel chemical cues in acoral reef fish. Anim Behav 69:51–57
Lima SL, Dill LM (1990) Behavioral decisionsmade under the risk of predation: a reviewand prospectus. Can J Zool 68:619–640
Maerz JC, Panebianco NL, Madison DM (2001) Ef-fects of predator chemical cues and beha-vioral biorhythms on foraging activity ofterrestrial salamanders. J Chem Ecol27:1333–1344
Magurran AE (1989) Acquired recognition ofpredator odour in the European minnow(Phoxinus phoxinus). Ethology. 82:216–223
Mathis A, Smith RJF (1993) Fathead minnows,Pimephales promelas, learn to recognizenorthern pike, Esox lucius, as predators onthe basis of chemical stimuli from min-nows in the pike’s diet. Anim Behav46:645–656
Murray DL, Roth JD, Wirsing AJ (2004) Predationrisk avoidance by terrestrial amphibians:the role of prey experience and vulnerabi-lity to native and exotic predators. Etho-logy 110:635–647
Petranka JW, Kats LB, Sih A (1987) Predator-preyinteractions among fish and larval amphi-bians: use of chemical cues to detect pre-datory fish. Anim Behav 35:420–425
Rohr JR, Madison DM (2001) A chemically me-diated trade-off between predation riskand mate search in newts. Anim Behav62:863–869Rev
Smith RJF (1992) Alarm signals in fishes. FishBiol Fish 2:33–63
Smith RJF 1997 Avoiding and deterring preda-tors. In: Behavioural Ecology of TeleostFishes (Ed. Godin JGJ)Oxford UniversityPress. Oxford pp 163–190
Sokal RR, Rohlf FJ (1995) Biometry. 3rd edn. NewYork:W. H. Freeman.
Suboski MD, Brian S, Carty AE, McQuoid LM, Se-elen MI, Seifert M (1990) Alarm reactionin acquisition and social transmission ofsimulated-predator recognition by zebradanio fish (Brachyodanio rerio). J CompPsychol 104:101–112
Von Frisch K (1938) Zur psychologie des fische-schwarmes.Naturwissenschaften 26:601–606
Wilson DJ, Lefcort H (1993) The effect of preda-tor diet on the alarm response of red-legged frog, Rana aurora tadpoles. AnimBehav 46:1017–1019
Wisenden BD, Millard MC (2001) Aquatic flat-worms use chemical cues from injuredconspecifics to assess predation risk andto associate risk with novel cues. AnimBehav 62:761–766
Woody DR, Mathis A (1998) Acquired recogni-tion of chemical stimuli from an unfamiliarpredator: associative learning by adultnewts, Notophthalmus viridiscens. Copeia1998:1027–1031
Yunker WK, Wein DE, Wisenden BD (1999) Con-ditioned alarm behavior in fathead min-nows (Pimephales promelas) resultingfrom association of chemical alarm phero-mone with a nonbiological visual stimulus.J Chem Ecol 25:2677–2686
43


cAPÍTuLO ii
APRENDIZAJE, MEMORIA Y OLVIDO APARENTE
DE SEñALES qUíMICAS PROCEDENTES DE NUEVOS
DEPREDADORES EN RENACUAJOS DE RANA VERDE
LEARNING, MEMORIZING AND APPARENT FOR-GETTING OF CHEMICAL CUES FROM NEw PRE-DATORS BY IBERIAN GREEN FROG TADPOLES


Muchas adaptaciones antidepredatorias son provocadas por lahabilidad de las presas para reconocer señales químicas de los depreda-dores y actuar de acuerdo al nivel de riesgo que posee ese depredador.Sin embargo, en algunos casos, el reconocimiento de los depredadoresrequiere que las presas pasen por un proceso de aprendizaje. Losrenacuajos de rana verde ibérica (Pelophylax perezi) poseen la habilidadde aprender a reconocer nuevos depredadores potenciales. En este tra-bajo hemos examinado las capacidades de retención de lo aprendido (me-moria) de estos renacuajos. Los renacuajos fueron condicionados conseñales químicas de un pez exótico inofensivo en conjunto con sustanciasde alarma de individuos coespecíficos y se examinó si eran capaces de re-tener la respuesta condicionada (una reducción en el nivel de actividad).Encontramos que los renacuajos condicionados reducían sus niveles deactividad en las subsecuentes exposiciones sólo a las señales químicasdel pez. Los renacuajos fueron capaces de recordar esta asociación y deresponder consecuentemente por lo menos hasta nueve días después delcondicionamiento. La habilidad para aprender a reconocer y memorizardepredadores potenciales puede ser de especial importancia para la su-pervivencia de las presas que pueden llegar a encontrarse con una altavariedad de depredadores. Sin embargo, después de esos nueve días, enausencia de refuerzo, hay una falta de repuesta a las señales químicasdel pez. Esta ausencia de respuesta puede ser explicada si los renacuajosse comportan de acuerdo a la hipótesis de la respuesta dependiente delriesgo de depredación, y el riesgo percibido en la señal química aprendidadisminuye con el tiempo. Pero también puede ser debida a un proceso deaparente pérdida de memoria para evitar respuestas no adaptativas aseñales químicas de especies no peligrosas que aleatoriamente aparecie-ran mezcladas con la sustancia de alarma. Así, este estudio demuestraque los renacuajos de rana verde Ibérica, en ausencia de refuerzo, recuer-dan las señales químicas de un depredador aprendido solo por un tiempolimitado, lo que puede ser adaptativo en un contexto de percepción deamenaza.
Este cápitulo se corresponde con: Gonzalo A, López P, Martín J (2009) Learning, memorizing andapparent forgetting of chemical cues from new predators by Iberian green frog tadpoles. Anim Cognit
(in press).


LearnIng, memorIzIng and apparent forgettIng
of chemIcaL cues from new predators
by IberIan green frog tadpoLes
Many antipredator adaptations are induced by the prey’s ability to recognize chemical
cues from predators. However, predator recognition often requires learning by prey
individuals. Iberian green frog tadpoles (Pelophylax perezi) have the ability to learn new
potential predators. Here, we tested the memory capabilities of Iberian green frog
tadpoles. We conditioned tadpoles with chemicals cues from a nonpredatory fish in
conjunction with conspecific alarm cues, and examined whether tadpoles retained their
conditioned response (reduction of activity level). We found that conditioned tadpoles
reduced their activity levels in subsequent exposures to the nonpredatory fish cues alone.
Tadpoles were able to remember this association and reduced movement rate at least for
nine days after. The ability to learn and memorize potential predators may be especially
important for the survivorship of prey species that are likely to find a high variety of
predators. However, after those nine days there was a lack of response to the non-preda-
tory fish cues alone in absence of reinforcement. This could be explained if tadpoles
behave according to the threat sensitive predator avoidance hypothesis, and the perceived
risk to the learning cue diminished over time, or it could be due to an apparent forgetting
process to avoid nonadaptative responses to chemical cues of nondangerous species that
were randomly paired with alarm cues. Thus, this study demonstrates that green frog
tadpoles in the absence of reinforcement remember the chemical cues of a learned
predator only for a limited time which may be adaptative in a threat-sensitive context.
IntroductIon
Predators often induce shifts in preybehavior and antipredatory behavior isoften mediated by the prey’s ability torecognize chemical cues from predatorsand to react according to the threat levelposed by that predator (Lima and Dill1990; Lima 1998). Failure to respond toa potential predator may be fatal. Howe-ver, unnecessary antipredatory behaviormay have direct energetic costs as well ascosts associated with reduced opportu-nity to feed or reproduce (Lima and Dill1990). Sensory information obtainedabout a predator may assist an organismin assessing the potential risk accuratelyand, therefore, reduce these costs (Limaand Dill 1990; Chivers and Smith 1998).Experience with predation cues is an
important element in the development ofantipredator behavior in a wide range ofvertebrates (Von Frisch 1938; Petrankaet al. 1987; Kiesecker al. 1996; Chiversand Smith 1998) as well as invertebrates(Dodson et al. 1994; Jacobsen and Stabell2004; see review in Kats and Dill 1998).Therefore, learning may conceivably af-fect several aspects of antipredator beha-vior and experience may enhance theability to recognize predators.
In aquatic environments, reception ofchemical cues released by predators andinjured prey is an important sensorymode by which most prey gather infor-mation about the threat of predation(Dodson et al. 1994; Chivers and Smith1998; Kats and Dill 1998). There are a
49

variety of chemical cues associated withpredation, some from the predator(kairomones) and other cues released byinjured prey (i.e., alarm cues; Chivers andSmith 1998; Kats and Dill 1998). Chemi-cal alarm cues are important in facilita-ting learned recognition of predationrisk by prey animals such as many fishes(Göz 1941; Magurran 1989; Mathis andSmith 1993; Chivers and Smith 1994a,1998; Larson and McCormick 2005),adult newts (Woody and Mathis 1998) orfrog and toad tadpoles (Mirza et al 2006;Gonzalo et al. 2007). Releaser inducedrecognition learning involves the simul-taneous exposure to an aversive stimulusand a neutral stimulus causing learnedaversion to the neutral stimuli (Yunker etal. 1999). The result of this learningmechanism is acquired predator recog-nition in which predator näıve indivi-duals show appropriate antipredatorbehavior to the cue of a potential preda-tor even though they have had no directexposure to the predator. Several authorshave showed such acquired predator re-cognition by pairing alarm cues with thevisual or chemical cues of a predator(e.g., Chivers and Smith 1994a; Larsonand McCormick 2005; Gonzalo et al.2007).
Although numerous studies haveinvestigated learning, less attention hasbeen directed at memory. Learning andmemory are linked; there is little point tolearning if the information cannot berecalled and remembered. However, theprocesses are distinct, and there are dif-ferences between them. Learning is es-sentially the acquisition of memory,whereas memory has other composites,such as retention and the potential forinterference (Shettleworth 1998).European and fathead minnows, afterlearning predator recognition by condi-tioning them with alarm cues, retain thememory of the potential predator andrespond to their signals a few days to
several weeks after (Magurran 1989; Chi-vers and Smith 1994a, 1994b; Brownand Smith 1998; Mirza and Chivers2000). Also, different species of crayfishpresent different times of retention of thelearning response (Hazlett et al. 2002).Research directed at quantifying me-mory duration, how rates of forgettingprogress or what factors cause variationin forgetting rates is far less commonthan studies investigating the acquisitionof information (Shettleworth 1998).Some studies in the past have interpretedfailure to continue to respond to certainstimuli as memory "failure” with poten-tially negative fitness consequences. Ho-wever, since adaptive forgetting wasproposed (Kraemer and Golding 1997),very few studies have explored this topic(Brown et al 2002, Golub and Brown2003, Hawkins et al. 2007).
Amphibians can learn that unknowncues are dangerous when these unk-nown cues are mixed with conspecificalarm cues (Mirza et al. 2006; Gonzalo etal. 2007). Even cues from nonpredatoryspecies can be learned as dangerous(Gonzalo et al. 2007). However, the po-tential retention of this learning associa-tion in amphibians remains unexplored.The aim of this study was to examine theretention in the near future of a conditio-ned response to a “new” predator by Ibe-rian green frog tadpoles (Pelophylax
perezi), and discuss the possible adapta-tive significance of the lack of thisresponse in a threat-sensitive context.
Iberian green frogs live and breed indifferent kinds of aquatic habitats (Gar-cía-París 2000) with a wide range oftypes of predators. Also, tadpoles ofthese frogs have long periods of growthbefore metamorphosis (García-París2000), and their predator species varyacross seasons. A previous study showedthat Iberian green frog tadpoles that hadbeen exposed to non-predatory fish che-mical cues mixed with conspecific alarm
50

cues responded two day later to the“new” predator (non-predatory fish)cues alone with a reduction of activity(Gonzalo et al. 2007). Thus learning andmemory could be especially importantmechanisms for the survivorship of tad-poles and for accurately assessing therisk posed by a predator. Then, to test theduration of the response to a “new” pre-dator in the near future, we conditionedIberian green frog tadpoles with chemi-cals cues from a nonpredatory fish (i.e.cues not previously associated with dan-ger) in conjunction with conspecificalarm cues, and examined whethertadpoles retained their conditionedresponse.
methodsstudy animals
We collected 185 Iberian green frogtadpoles (SLV, Mean+SE=5.4±0.2 cm;Gosner’s stage=25; see Gosner 1960) bynetting during July of 2007 at severalsmall ponds in Collado Mediano (Madrid,Central Spain). Tadpoles were housed ingroups of five tadpoles at “El Ventorrillo”Field Station, 10 km from the capturearea, in plastic aquaria (49x29 cm and 25cm high) with 5 L of water at ambienttemperature and under a natural photo-period. They were fed every day withcommercial fish flakes.
We obtained from a commercialdealer nonpredatory zebra danio fish(Brachyodanio rerio) to be used as sourceof neutral scent. Before and after the endof the experiment, fishes were maintai-ned in a large filtered aquarium and re-gularly fed with commercial fish flakes.All the animals were healthy during thetrials; all maintained or increased theiroriginal body mass.
preparation of chemical stimuli
Alarm cues of tadpoles were prepa-
red from three tadpoles (SLV, Mean±SE=4.2±0.1 cm). They were cold anestheti-zed by placing at 4 °C for 20 min, indu-cing deep hypothermia, and, then wereeuthanasied with a quick blow to thehead to avoid suffering (ASIH 2004). Wedid not use a chemical anaesthetic be-cause these chemicals may interfere withnatural tadpoles’ chemical cues in sub-sequent trials. The extract was then pre-pared by putting these dead tadpoles ina clean disposable plastic dish, and ma-cerating them in 3000 mL of distilledwater. The stimulus water was then filte-red through absorbent paper to removesolid particles, and immediately frozen in10 mL portions until used (Woody andMathis 1998).
The fish stimulus was prepared byplacing ten zebra danio fishes into a 10 Laquarium with clean water for threedays. These aquaria were aerated but notfiltered. Fishes were not fed during thisshort period to avoid contaminatingwater with food odor. Thereafter, waterwas drawn from the aquaria and frozenin 10 mL portions until its use in experi-ments. Fishes were returned and fed intheir home aquaria. We prepared controlwater in an identical manner but withoutplacing fish in the aquaria (Woody andMathis 1998).
experimental design
We randomly assigned each group offive tadpoles to two different treatments(control group, N=90; or experimentalgroup, N=90). On the first day of the ex-periment tadpoles from the ‘control’treatment were exposed to the fishchemical cues alone mixed with cleanwater. At the same time, tadpoles fromthe ‘experimental’ treatment were expo-sed to both the scent of the fish and cons-pecific chemical alarm cues, thus,simulating the cues from a predatory fishthat was eating a conspecific tadpole. A
51

previous study showed that tadpolesconditioned with these mix of stimuli(fish and alarm cues) were two days laterable to recognize the fish chemical cuesalone as coming from a predator (Gon-zalo et al. 2007). Thus, experimental tad-poles were considered as conditioned.The conditioning events (control and ex-perimental) were carried on in the tad-poles’ home plastic aquaria (49x29 cmand 25 cm high) to avoid stress due totransfer from one aquaria to another. Wemade different conditioning solutions, 20mL each (2 ice aliquots), using combina-tions of clean water with fish chemicalcues, and alarm cues with fish chemicalcues. After ice aliquots were thawed wepipetted conditioning solutions in thecentre of the aquaria.
To test for the duration of the res-ponse to this predator recognition, ondays 3, 6, 9, 12, 15 or 18 after the initialconditioning, different individuals fromthe two treatments were tested with thefish chemical cues alone in clean water.15 individuals from the two groups weretested at the same time in parallel andobservations were carried out blind tothe tadpole treatment. Each day, we tes-ted 15 individuals from each of the twogroups (control and experimental).These tadpoles were chosen randomlyfrom the several groups we had. After thetrial, tadpoles were kept separately andnot used in subsequent trials. New indi-vidual tadpoles were used in each trial toensure that we were just testing the ca-pacity of retention of the learning asso-ciation. We could not use the sameindividual tadpoles in more than one testbecause successive exposures withoutreinforcement could lead to learning theinnocuousness of the “non-dangerous”predator (Hazlett 2003) or habituationto the predator cues, so that the resultsof the experiment would not reflect du-ration of the response alone.
Tadpoles were tested individually ingrey, U-shaped gutters (101x11 cm and6 cm high) sealed at both ends with plas-tic caps (see Rohr and Madison 2001 fordetailed descriptions). We marked theinternal part of the gutters with fourcrossing lines that created five subdivi-sions of equal surface. We filled eachgutter with 3 L of clean water (20 °C)from a mountain spring, which did notcontain fish. We placed clear plastic overeach trough on either side of the cageto isolate the system from air movementsin the testing room (see Rohr and Madi-son 2001). Each trial lasted 1 h andconsisted of a 30 min pre-stimulusperiod and a 30 min post- stimulusperiod separated by a stimulus introduc-tion. We assigned test solutions (10 mLof fish scent) to one end of each trough(right or left) by stratified randomiza-tion, and assigned 10 mL of clean waterto the opposite end. We placed a singletadpole in each gutter, and waited 5 minfor acclimatation. Then, we started the‘pre-stimulus’ test (30 min). Thereafter,we added the test solution and, then,immediately started the ‘post-stimulus’period (30 min). During both the pre-and the post-stimulus periods we recor-ded from a blind the quadrant that eachtadpole occupied at 1 min intervals for30 min. We calculated the number oflines crossed for all periods and this wasconsidered as an index of general activity(Rohr and Madison 2001; Gonzalo et al.2007). Diffusion of chemicals in stillwater may be a slow process. However,all individual tadpoles used in the expe-riment were observed at least once in allof the subdivisions of the gutter, so wewere confident that all tadpoles werereally exposed to the chemical stimuli.Moreover, tadpoles often showedepisodes of fast swimming which shouldcontribute to diffuse chemicals in water.
52

data analyses
For each trial we calculated levels ofactivity as the difference between thenumbers of line crossed between the preand post-stimulus periods. Positive va-lues indicate increased movement follo-wing addition of the stimulus; negativevalues indicate decreased activity. Datawere log transformed to ensure norma-lity (Shapiro–Wilk’s tests, P>0.05 in allcases) and homogeneity of variances (Le-vene's tests, P>0.05 in all cases), and thentested by general linear modeling (GLM;Grafen and Hails 2002). We used ‘day’ ofthe trial (i.e., munber of days since the in-itial conditioning event) and ‘conditio-ning’ (i.e., control treatment conditioningwith the absence of alarm cues vs expe-rimental treatment conditioning with thepresence of alarm cues) as categoricalvariables. We included the interactionsbetween variables in the model to test forthe effects of the different treatments(conditioned with or without alarmcues) depending on the day of the trial.Subsequent post hoc multiple compari-sons were made using Tukey’s pairwisecomparisons (Sokal and Rohlf 1995).
resuLts
On average, control tadpoles weremore active than experimental tadpolesconditioned with conspecific alarm cue(‘conditioning’ effect: F1,168=31.05,P<0.0001; Fig. 1). Also, there were signi-ficant differences between the differentdays of the trials (‘days’ effect:F5,168=6.45, P<0.002). Therefore, overallactivity of tadpoles increased over timesince the initial conditioning event. Ho-wever, the interaction between factorswas significant (F5,168=3.76, P=0.002;Fig. 1). Thus, three days after the initialconditioning event experimental tadpo-les, previously exposed to a combinationof alarm cues and fish odor; significantly
decreased activity in comparison withcontrol tadpoles (Tukey’s test: P=0.02),the same difference was noted six(P=0.001) and nine days (P=0.003) afterthe conditioning event. However, on daystwelve, fifteen and eighteen there wereno significant differences in activity bet-ween experimental and control tadpoles(P≥0.90 in all cases). Although, therewere no significant differences in activitylevel between days for the control tadpo-les (P≥0.05 in all cases), there weresignificant differences between the expe-rimental tadpoles depending of the day;activity of experimental tadpoles did notdiffer on days three, six, and nine (P≥0.90in all cases), but activity was significantlylower on these days than on days fifteenand eighteen (P<0.02), which were notsignificantly different between them(P=0.90). Beginning on day twelve andeach test day thereafter, activity of expe-rimental tadpoles was not significantlydifferent from activity of experimentaltadpoles (P≥0.20 in all cases).
dIscussIon
Our results showed that three, six andnine days after exposure to chemical cuesfrom the potential predatory fish, theexperimental Iberian green frog tadpolesdisplayed antipredator behaviors (i.e., areduction in activity) in response tothese chemical cues, suggesting that tad-poles still remembered the learned cueassociation nine days after exposure.Twelve days after the initial conditioning,mean activity of experimental tadpoles inresponse to fish chemical cues was hig-her but variance was also high, sugges-ting that some of the experimentaltadpoles, but not others, still were able toreact to the fish as a predator. In contrast,at fifteen and eighteen days post-expo-sure, experimental tadpoles did not showany antipredator behavior, and behavedas the control tadpoles. Several studies
53
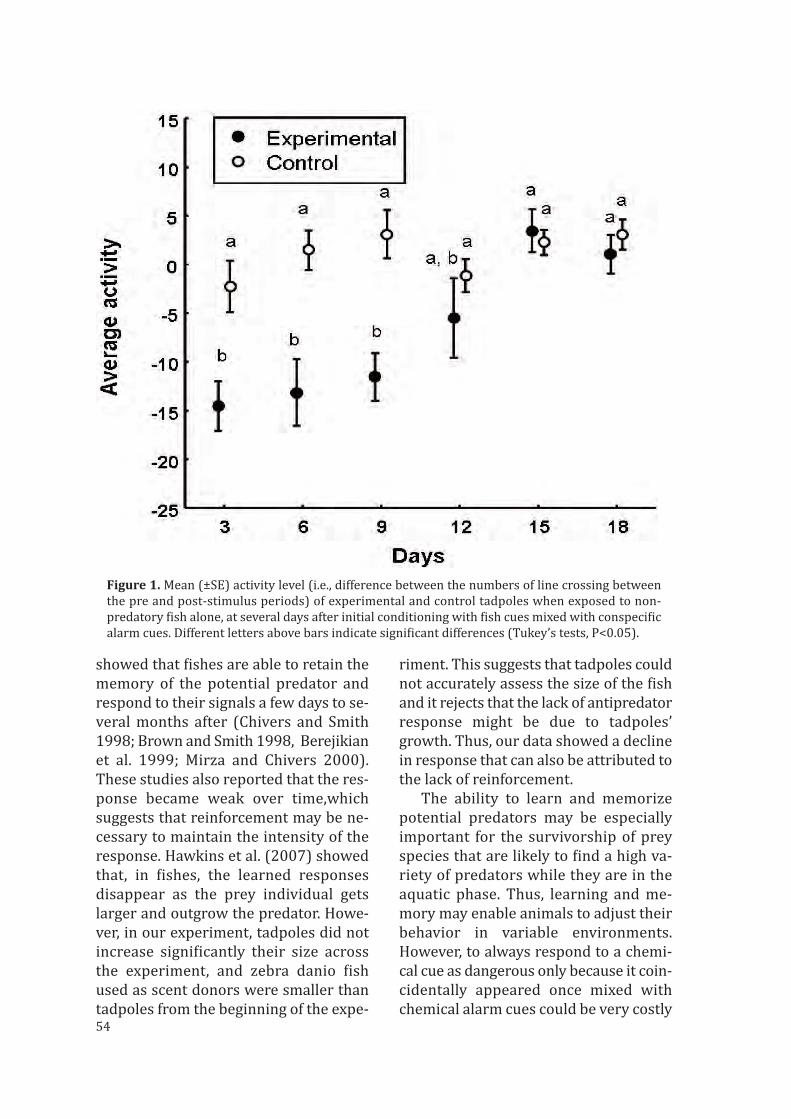
showed that fishes are able to retain thememory of the potential predator andrespond to their signals a few days to se-veral months after (Chivers and Smith1998; Brown and Smith 1998, Berejikianet al. 1999; Mirza and Chivers 2000).These studies also reported that the res-ponse became weak over time,whichsuggests that reinforcement may be ne-cessary to maintain the intensity of theresponse. Hawkins et al. (2007) showedthat, in fishes, the learned responsesdisappear as the prey individual getslarger and outgrow the predator. Howe-ver, in our experiment, tadpoles did notincrease significantly their size acrossthe experiment, and zebra danio fishused as scent donors were smaller thantadpoles from the beginning of the expe-
riment. This suggests that tadpoles couldnot accurately assess the size of the fishand it rejects that the lack of antipredatorresponse might be due to tadpoles’growth. Thus, our data showed a declinein response that can also be attributed tothe lack of reinforcement.
The ability to learn and memorizepotential predators may be especiallyimportant for the survivorship of preyspecies that are likely to find a high va-riety of predators while they are in theaquatic phase. Thus, learning and me-mory may enable animals to adjust theirbehavior in variable environments.However, to always respond to a chemi-cal cue as dangerous only because it coin-cidentally appeared once mixed withchemical alarm cues could be very costly
figure 1. Mean (±SE) activity level (i.e., difference between the numbers of line crossing betweenthe pre and post-stimulus periods) of experimental and control tadpoles when exposed to non-predatory fish alone, at several days after initial conditioning with fish cues mixed with conspecificalarm cues. Different letters above bars indicate significant differences (Tukey’s tests, P<0.05).
54

(Lima and Dill 1990). In those circums-tances, the longer the time intervalbetween successive contacts with a par-ticular cue labeled as “dangerous”, themore likely that it was not actually dan-gerous. Thus, the lack of antipredatorresponse, far from a failed memory, couldbe a powerful strategy for dealing withconflicting information. In another expe-riment (Gonzalo et al. 2007) Iberiangreen frog tadpoles behaved according tothe threat sensitive predator avoidancehypothesis (Helfman 1989) when theywere confronted with a native predatorsnake (to which they innately react, Gon-zalo et al. unpublished data). Thus, sna-kes were not perceived to be verydangerous predators unless the tadpo-les had been previously exposed tosnakes that ‘had eaten conspecifics’ (i.e.mix of alarm cues plus snake chemicalcues). According to the threat sensitivepredator avoidance hypothesis, prey spe-cies should behave flexibly towards avarying degree of predator threat and,consequently, leave more time for otheractivities when the threat is low (Helf-man 1989). If tadpoles reacted very we-akly to a natural and familiar predatorthat is not actually attacking tadpoles, itis possible that they were assessing riskbased on predator diet and adaptivelybalancing the costs and benefits of pre-dator avoidance. In the present study, wegave to tadpoles incomplete or unreliableinformation regarding a new predatoridentity, allowing them only to “smell” anew potential predator. As tadpoles didnot have other additional information onthe actual risk of this particular predator,the selection of an antipredator behaviorshould be informed by recent experien-ces (Turner et al. 2005, Ferrari andChivers 2006). Thus, as time goes byfrom the encounter with the potentialpredator, the perceived risk would dimi-nish over time from the exposure to theconditioning event and fish percived as
low risk predators due to lack of reinfor-cement. It would also allow tadpoles toshow avoidance behaviors only in theface of active predators and high preda-tion risk, and to reduce costs associatedwith unnecessary antipredatory beha-vior. This hypothesis is supported by arange of studies examining memory thathave revealed that, even after apparentforgetting, a latent (residual) memorypersists and can be revealed by facilita-ted acquisition in a subsequent learningevent (Plotkin and Oakley 1975; Matzelet al. 1992; Monk et al. 1996; Nicholsonet al. 2003; Philips et al. 2006). Thus,a memory may outlast its behavioralexpression.
However, Philips et al. (2006) alsofound that in water snails, latent memoryto attacks with electric shocks decayed atfour days. Thus, the lack of antipredatorresponse could also be due to a realforgetting phenomenon (Philips et al.2006). Traditionally, forgetting was con-sidered a failing of memory, but over thepast decades researchers have movedtowards the idea that the ability to forgetmay be advantageous (Kraemer andGolding 1997). Learning and memoryallow animals to adjust their behavior toadapt to changeable environments andthus cope with a degree of unpredictabi-lity (Shettleworth 1998). In such envi-ronments, animals that use learning andmemory to hone their behavior will haveadvantages over other more behaviorallyfixed individuals. However, animalscontinually receive information abouttheir environment and must filter thisinformation to focus on those aspectsmost important to survival (Dukas2002). Thus, prey are often confrontedwith multiple types of potential preda-tors, and a response caused by a coinci-dental pairing of unrelated cues inducingantipredator behavior could prove verycostly to prey species. Forgetting proces-ses that help map an animal’s behavior to
55

the instabilities inherent in a changingenvironment could, thus, contribute tosurvival (Hendersen 1985). In naturalconditions, tadpoles have to encounter awide range of mixed cues (e.g., alarmcues, predator cues, nonpredator cues).As we see in the present study, to elicit anantipredator response to predator cuesduring the first days, even overestima-ting the potential risk level, could beadaptative because tadpoles had recentpassed through a conditioning event ofbiological importance (learning a newdangerous predator). However, as timepasses and tadpoles do not face up thesame ‘predator that had eaten conspeci-fics’ (i.e., the mix of fish cues with alarmcue), the possibility of the two cues acci-dentally appearing together increases.Therefore, to lose the tencency to res-pond to the previous association couldbe adaptative, because it may preventtadpoles to do not persevere with mala-daptive antipredator behaviors towardsnon-dangerous species that live in thesame habitat.
references
ASIH (2004) Guidelines for use of live Amphi-bians and Reptiles in field and laboratoryresearch. Herpetological Animal Care andUse Committee (HACC) of the AmericanSociety of Ichthyologists and Herpetolo-gists, Lawrence, Kansas
Berejikian BA, Smith RJF, Tezak EP, Schroder SL,Knudsen CM (1999) Chemical alarm sig-nals and complex hatchery rearing habi-tats affect antipredator behavior andsurvival of chinook salmon (Oncorhynchus
tshawytscha) juveniles. Can J Fish Aq Sci56:830–838
Brown GE, Smith RJF (1998) Acquired predatorrecognition in juvenile rainbow trout(Oncorhynchus mykiss): conditioning hat-chery-reared fish to recognize chemicalcues of a predator. Can J Fish Aq Sci55:611–617
Brown GE, Gershaneck DL, Plata DL, Golub JL(2002) Ontogenetic changes in response toheterospecific alarm cues by juvenile lar-gemouth bass are phenotypically plastic.
Behaviour 139:913–927Chivers DP, Smith RJF (1994a) Fathead minnows,
Pimephales promelas, acquire predator re-cognition when alarm substance is asso-ciated with the sight of unfamiliar fish.Anim Behav 48:597–605
Chivers DP, Smith RJF (1994b) The role of espe-rience and chemical alarm signalling inpredator recognition by fathead minnows,Pimephales promelas. J Fish Biol 44:273–285
Chivers DP, Smith RJF (1998) Chemical alarm sig-nalling in aquatic predator/prey interac-tions: a review and prospectus. Ecoscience5:338–352
Dodson SI, Crowl TA, Peckarsky BL, Kats LB,Covich AP, Culp JM (1994) Non-visual com-munication in freshwater benthos: anoverview. J Natl Am Benthol Soc 13:268 –282
Dukas R (2002) Behavioural and ecological con-sequences of limited attention. Phil TransR Soc Lond B 357:1539–547
Ferrari MCO, Chivers DP (2006) Learning threat-sensitive predator avoidance: how dofathead minnows incorporate conflictinginformation? Anim Behav 71:19–26
García-París M (2000) Rana perezi. In: Ramos MA(ed) Amphibia, Lissamphhibia. Fauna Ibe-rica vol 24. Museo Nacional de CienciasNaturales (CSIC), Madrid, pp 467–478
Gonzalo A, López P, Martín J (2007) Iberian greenfrog tadpoles may learn to recognize novelpredators from chemical alarm cues ofconspecifics. Anim Behav 74:447–453
Gosner KL (1960) A simplified table for staginganuran embryos and larvae with notes onidentification. Herpetologica, 16:183–190
Göz H (1941) Über den art-und individualgeruchbei fischen. Z Verg Physiol 29:1–45
Golub JL, Brown GE (2003). Are all signals thesame?: Ontogenetic changes in the res-ponse to conspecific and heterospecificalarm signals by juvenile green sunfish(Lepomis cyanellus). Behav Ecol Soc54:113–118
Grafen A, Hails R (2002) Modern statistics for thelife sciences. Oxford University Press,Oxford
Hawkins LA, Magurran AE, Armstrong JD (2007)Ontogenetic learning of predator recogni-tion in hatchery-reared Atlantic salmon,Salmo salar. Anim Behav 75:1663–1671
Hazlett BA, Acquistapace P, Gherardi F (2002)Differences in memory capabilities in in-vasive and native crayfish. J CrustaceanBiol 22:439–448
56

Hazlett BA (2003) Predator recognition and lear-ned irrelevance in the crayfish Orconectes
virilis. Ethology 109:765–780Helfman GS (1989) Threat-sensitive predator
avoidance in damselfish-trumpetfish inter-actions. Behav Ecol Sociobiol 24:47–58
Hendersen RW (1985) Fearful memories: Themotivational significance of forgetting. In:Brush FR, Overmier JB (eds) Affect, condi-tioning, and cognition: Essays on the de-terminants of behaviour. Erlbaum,Hillsdale, New Jersey, pp 43-53
Jacobsen HP, Stabell O-B (2004) Antipredator be-haviour mediated by chemical cues: therole of conspecific alarm signalling andpredator labelling in the avoidance res-ponse of a marine gastropod. Oikos104:43-50
Kats LB, Dill LM (1998) The scent of death: che-mosensory assessment of predation byprey animals. Ecoscience 5:361–394
Kiesecker JM, Chivers DP, Blaustein AR (1996)The use of chemical cues in predator re-cognition by western toad (Bufo boreas)tadpoles. Anim Behav, 52:1237–1245
Kraemer PJ, Golding JM (1997) Adaptive forget-ting in animals. Psychon B Rev 4:480–491
Larson JK, McCormick MI (2005) The role of che-mical alarm signals in facilitating learnedrecognition of novel chemical cues in acoral reef fish. Anim Behav 69:51–57
Lima SL (1998) Nonlethal effects in the ecologyof predator-prey interactions: what are theecological effects of anti-predator deci-sion-making? Bioscience 48:25–34
Lima SL, Dill LM (1990) Behavioral decisionsmade under the risk of predation: a re-view and prospectus. Can J Zool 68:619–640
Magurran AE (1989) Acquired recognition of pre-dator odour in the European minnow(Phoxinus phoxinus). Ethology 82:216–223
Mathis A, Smith RJF (1993) Fathead minnows, Pi-
mephales promelas, learn to recognize nor-thern pike, Esox lucius, as predators on thebasis of chemical stimuli from minnows inthe pike’s diet. Anim Behav 46:645–656
Matzel LD, Collin C, Alkon DL (1992) Biophysicaland behavioral correlates of memory sto-rage, degradation, and reactivation. BehavNeurosci 106:954–963
Mirza RS, Chivers DP (2000) Predator recognitiontraining enhances survival of brook trout:evidence from laboratory and field-enclo-sure studies. Can J Zool 78: 2198–2208
Mirza RS, Ferrari MCO, Kiesecker JM, Chivers DP(2006) Responses of American toad tadpo-les to predation cues: behavioural res-ponse thresholds, threat-sensitivity andacquired predation recognition. Behaviour143:877–889
Monk CM, Gunderson VM, Grant KS, Mechling JL(1996) A demonstration of the memory sa-vings effect in infant monkeys. Dev Psychol32:1051–1055
Nicholson DA, Sweet JA, Freeman Jr JH (2003)Long-term retention of the classically con-ditioned eyeblink response in rats. BehavNeurosci 117:871–875
Petranka JW, Kats LB, Sih A (1987) Predator-preyinteractions among fish and larval amphi-bians: use of chemical cues to detect pre-datory fish. Anim Behav 35:420–425
Philips GT, Tzvetkova EI, Marinesco S, Carew TJ(2006) Latent memory for sensitization inAplysia. Learn Mem 13:224–229
Plotkin HC, Oakley DA (1975) Backward condi-tioning in the rabbit (Oryctolagus cunicu-
lus). J Comp Physiol Psychol 88:586–590Rohr JR, Madison DM (2001) A chemically media-
ted trade-off between predation risk andmate search in newts. Anim Behav62:863–869
Shettleworth SJ (1998) Cognition, evolution andbehaviour. Oxford University Press, NewYork
Sokal RR, Rohlf FJ (1995) Biometry. WH Freeman,New York
Turner AM, Turner SE, Lappi HM (2005) Lear-ning, memory and predator avoidance byfreshwater snails: effects of experience onpredator recognition and defensive stra-tegy. Anim Behav 72:1443–1450
Von Frisch K (1938) Zur psychologie desfische-schwarmes. Naturwissenschaften26:601–606
Woody DR, Mathis A (1998) Acquired recognitionof chemical stimuli from an unfamiliarpredator: associative learning by adultnewts, Notophthalmus viridescens. Copeia1998:1027–1031
Yunker WK, Wein DE, Wisenden BD (1999) Con-ditioned alarm behavior in fathead min-nows (Pimephales promelas) resultingfrom association of chemical alarm phero-mone with a nonbiological visual stimulus.J Chem Ecol 25:2677–2686
57


cAPÍTuLO iii
OLVIDO ADAPTATIVO EN RENACUAJOS DE RANA
VERDE: LA INHIBICIÓN LATENTE Y EL APRENDI-ZAJE DE LO IRRELEVANTE PUEDEN EVITAR FALSOS
RECONOCIMIENTOS DE DEPREDADORES
ADAPTATIVE FORGETTING IN FROGTADPOLES: LATENT INHIBITION ANDLEARNING IRRELEVANCE MAY AVOIDPREDATOR MISIDENTIFICATION


El reconocimiento de los depredadores a menudo necesita unproceso de aprendizaje por parte de las presas. Los renacuajos de ranaverde ibérica (Pelophylax perezi) tienen la habilidad de aprender a re-conocer nuevos depredadores potenciales cuando encuentran sus señalesquímicas asociadas con sustancias de alarma de individuos coespecíficos.Pero, una respuesta antidepredadora desencadenada por una asociaciónaleatoria de señales químicas no relacionadas puede ser muy costosapara las presas. En este trabajo hemos estudiado dos fenómenos (la in-hibición latente y el aprendizaje de lo irrelevante) que pueden ayudar alos renacuajos a evitar las respuestas no adaptativas hacia señales quí-micas de especies no peligrosas que pueden aparecer aleatoriamenteasociadas a señales de alarma. Nuestros resultados demuestran que,cuando los renacuajos de rana verde Ibérica son sometidos a un patrónaleatorio de señales de alarma y sustancias químicas de un nuevo de-predador, durante los cuatro días anteriores o posteriores a la detecciónsimultánea de ambos, no se forma ninguna asociación de aprendizaje.Estos resultados muestran la existencia de un efecto de “aprendizaje delo irrelevante” en los patrones de aprendizaje de los renacuajos de estaespecie. Además, los renacuajos claramente inhibieron la formación deuna asociación entre las sustancias químicas de un nuevo depredador yla señal de alto riesgo de depredación después de un periodo de cuatrodías durante el que fueron expuestos sólo a las señales químicas del de-predador. Este resultado muestra la existencia de un efecto de “inhibiciónlatente” en el aprendizaje de sustancias relacionadas con un aumentodel riesgo de depredación. Ambos mecanismos (aprendizaje de lo irrele-vante e inhibición latente) lejos de ser considerados un fallo en el reco-nocimiento de depredadores, pueden ser vistos como vías adaptativaspara enfrentarse con información conflictiva, y como estrategias paraevitar el aprendizaje de información irrelevante que desencadenencostosas respuestas antidepredatorias ante estímulos de individuos nodepredadores.
Este cápitulo se corresponde con: Gonzalo A, López P, Martín J. Adaptative forgetting in frog tadpoles:latent inhibition and learning irrelevance may avoid predator misidentification. Anim Behav(Submited manuscript).


AdAptAtIve forgettIng In frog tAdpoles: lAtent
InhIbItIon And leArnIng IrrelevAnce mAy
AvoId predAtor mIsIdentIfIcAtIon
Predator recognition often requires learning by prey individuals. Iberian green frog
tadpoles (Pelophylax perezi) have the ability to learn new potential predators when their
chemical cues are finding paired with conspecific alarm cues. But, a response caused by
a random pairing of unrelated cues inducing antipredator behavior to non-predator
stimuli could be very costly to prey species. Here we study two phenomena (latent
inhibition and learning irrelevance) that could help tadpoles to avoid nonadaptative
responses to chemical cues of non-dangerous species that sometimes may appear
randomly pairing with alarm cues. Our results demonstrated that when Green frog
tadpoles experienced a random pattern of alarm cues and predator cues over the four
days before or after the simultaneous detection of these two cues, no learned association
was formed. These results showed the existence of an effect of learned irrelevance on
learning in these frog tadpole species. Also, tadpoles clearly inhibited the formation of a
learning association between predator and alarm cues after a four days period during
which they had been exposed to the predator cues alone. This result showed the existence
of an effect of latent inhibition on learning about cues related to increased predation risk
in tadpoles. Both learned irrelevance and latent inhibition far away to be considered as
a fail in predator recognition, can be rather seen as adaptive ways for dealing with
conflicting information, and as strategies to avoid learning irrelevant information and
costly antipredatory responses to non-predatory stimuli.
IntroductIon
An important component of antipre-
dator behaviour is the ability to detect
and recognize predators (Lima and Dill
1990). Many antipredator adaptations
are induced or mediated by the prey’s
ability to recognise chemical cues from
predators (Kats and Dill 1998). Many
aquatic animals, including some inverte-
brates, fishes and amphibians, use che-
mical cues to assess predation risk (e.g.
Von Frisch 1938; Petranka et al. 1987;
Dodson et al. 1994; Kiesecker et al. 1996;
Chivers and Smith 1998). Chemical cues
may arise from the predators, but chemi-
cals may also be often released by prey
when they are captured by a predator
(i.e. alarm cues) (Chivers and Smith
1998; Kats and Dill 1998), which serve as
a reliable and imminent indicator of risk
for conspecifics (Chivers and Smith
1998).
In some cases, chemically based pre-
dator recognition requires learning by
prey individuals (e.g. fishes: Mathis and
Smith 1993; damselfly larvae: Chivers et
al. 1996; crayfish: Hazlett 2003; amphi-
bians: Mirza et al. 2006, Gonzalo et al.
2007). Prey must be exposed simultane-
ously to a predator cue and a danger cue,
such as an alarm cue released by a crus-
hed conspecific, before the predator cue
is considered as a danger signal. Howe-
ver, in natural conditions, prey do not
find the predator and alarm cues paired
alone, but they also find a mix of many
several cues of a wide range (e.g. alarm63

cues, predator cues, non-predator cues).
So, prey are often confronted with cues
from multiple types of potential preda-
tors but also with cues from non-preda-
tor species, and a response caused by a
random pairing of any unrelated cues in-
ducing unnecessary antipredator beha-
vior to non-predatory stimuli could
prove to be very costly to prey species
(Belden et al. 2000). Although numerous
studies have investigated learning of pre-
dators by prey (see above), less attention
has been directed to mechanisms for
avoiding or forgetting the erroneous
learning of non-dangerous species as
dangerous. These latter forgetting me-
chanisms are, however, very important
because they helped to prevent prey to
do not persevere with mal adaptive anti-
predator behaviours towards non-preda-
tor species.
In classical conditioning learning,
the subject learns about the relationship
between two stimuli, an unconditioned
stimulus, which produces a strong, con-
sistent, overt unconditioned response,
and a conditioned stimulus, which on it-
self produces either no overt response
or a weak response usually unrelated to
the response that eventually will be
learned. (Kamprath and Wotjak 2004).
However, the rate at which a subject ac-
quires conditioned responses to paired
presentations of conditioned stimulus
and unconditioned stimulus can be affec-
ted by prior exposure to those individual
stimuli (Allen et al. 2002). There are two
phenomena that might be particularly
relevant to the situations that prey pro-
bably experience in nature, one is ‘latent
inhibition’, where the previous exposure
to a stimulus (without any reinforce-
ment) results in a reduction of the
strength of a learned association that
could be formed later (Ferguson et al.
2001). The other phenomenon is ‘lear-
ned irrelevance’ (Bennet et al. 2000), in
which the random exposure to two cues
altered later learning that would follow
from exposure to the two paired cues.
Thus, if both of these phenomena also
occur in the predator learning process,
the detection of a cue from a potential
predator that was not temporally linked
to conspecific alarm cues indicating
increased predation risk might later
inhibit the formation of the association
and learning that would occur normally
when the two cues appear together.
However, as far as we know very little
work has being made to demonstrate the
existence of these phenomena in the pre-
dator learning processes. Hazlett (2003)
showed learned irrelevance in virile
crayfish (Orconectes virilis). When cray-
fish are given uncorrelated presentations
of conspecific alarm cues and the cues of
a novel predator, associative learning
fails to occur. Acquistapace et al. (2003)
demonstrated latent inhibition in cray-
fish. Pre- exposure to cues of a novel pre-
dator for two hours during three
consecutive days prevents associative le-
arning of those cues with high risk. Also
Ferrari and Chivers (2006a) demonstra-
ted the existence of latent inhibition in fa-
thead minnows. Minnows pre-exposed to
novel predator cues for one hour on five
consecutive days fail in the posterior as-
sociative learning of the predator.
Amphibian tadpoles can learn to
recognize as dangerous unknown chemi-
cal cues when these have been pre-
viously presented mixed with conspecific
alarm cues (Mirza et al. 2006; Gonzalo et
al. 2007). Even cues from non-predatory
species can also been learned as dange-
rous if they have been previoulsy asso-
ciated to alarm cues (Gonzalo et al.
2007). However, the existence of mecha-
nisms able to prevent learning associa-
tion in amphibians remains unexplored.
In this study, we investigated whether
latent inhibition and learned irrelevance
may inhibit the associative learning of
predator cues in tadpoles of the Iberian
64

green frogs (Pelophylax perezi). This spe-
cies lives and breeds in different kinds of
aquatic habitats (García-París 2000) that
contains a wide range of types of preda-
tors and non predators organisms. Also,
Iberian green frogs have long periods of
growth before metamorphosis (García-
París 2000), and their predator species
vary across seasons. Thus, accurate lear-
ning of actual predator cues should be
important for P. perezi tadpoles. Previous
experiments showed that Iberian green
frog tadpoles are able to learn potential
predators with the presentation of the
chemical cue mixed with alarm cues
(Gonzalo et al. 2007), but in natural con-
ditions frog tadpoles have to face up with
a wide range of mixed cues (e.g., alarm
cues, predator cues, nonpredator cues),
and the learning inhibition mechanisms
could be an adaptative way to avoid lear-
ning irrelevant information. So, in this
experiment we exposed Iberian green
frog tadpoles to a random patron of non-
predator and alarm chemical cues, or to
nonpredator cues alone for four days,
before the conditioning and examined
whether tadpoles recognized the non
predatory fish cues as dangerous.
methodsstudy animals
During August 2006 We collected 180
Iberian green frog tadpoles (SVL,
MEAN±SE=5.2±0.3 cm, Gosner’s stage:
24; see Gosner 1960) by netting at seve-
ral small ponds in Collado Mediano (Ma-
drid, central Spain). Tadpoles were
randomly housed in groups of five indi-
viduals at “El Ventorrillo” Field Station,
10 km from the capture area, in plastic
aquaria (18 x 25 cm and 10 cm high)
with 5 L. of water at ambient tempera-
ture and under a natural photoperiod.
Tadpoles were fed every day with com-
mercial fish flakes.
We obtained from a commercial
dealer non predatory zebra danio fish
(Brachyodanio rerio) to be used as source
of neutral scent. Before and after the
experiment finished, fishes were main-
tained in a large filtered aquarium and
regularly fed with commercial fish flakes.
All the animals were healthy during
the trials, all maintained or increased
their original body mass. The tadpoles
were returned to their exact capture site.
The experiments were performed under
license from the “Consejería de Me-
dioambiente de la Comunidad de Ma-
drid” (the Environmental Agency of the
local Government of Madrid). Procedures
are conformed to recommended guideli-
nes for use of live Amphibians in labora-
tory research (ASIH 2004).
preparation of chemical stimuli
Alarm cues of tadpoles were prepa-
red from three tadpoles (SVL, Mean±SE=
4.2±0.1 cm). They were cold anestheti-
sed by placing at 4 °C for 20 min, indu-
cing them deep hypothermia, and, then,
euthanatized with a quick blow to the
head to avoid suffering (ASIH 2004). We
did not use a chemical anaesthetic, be-
cause these chemicals may interfere with
natural tadpoles’ chemical cues in subse-
quent trials. The extract was then prepa-
red by putting these dead tadpoles in a
clean disposable plastic dish, and mace-
rating them in 3000 mL of distilled water.
The stimulus water was then filtered
through absorbent paper to remove solid
particles, and immediately frozen in
10 mL portions until used (Woody and
Mathis 1998).
The fish stimulus was prepared by
placing ten zebra danio fishes into a 10 L
aquarium with clean water for three
days. These aquaria were aerated but not
filtered. Fishes were not fed during this
short period to avoid contaminating
water with food odour. Thereafter, water
was drawn from the aquaria and frozen
65

in 10 mL portions until its use in experi-
ments. Fishes were returned and fed in
their home large aquaria.
general methodology
Tadpoles were first exposed, in their
own aquarium, over a period of five days
to different sequences of exposure to a
novel chemical cue (zebra danio fish),
conspecific alarm cue, or both types of
cues mixed (pre-exposure period). The
particular sequences and durations are
described below for each experimental
treatment.
Following this pre-exposure period,
tadpoles were tested individually in an
observation aquaria consisting of grey,
U-shaped gutters (101x11 cm and 6 cm
high) sealed at both ends with plastic
caps. We marked the internal part of the
gutters with four crossing lines that
created five subdivisions of equal sur-
face. We filled each gutter with 3 L of
clean water from a mountain spring,
which not contained fish no tadpoles, at
20ºC. We placed clear plastic over each
trough on either side of the cage to iso-
late the system from air movements in
the testing room (see Rohr and Madison
2001). We assigned test solutions (clean
water or zebra danio fish cues) to one
end of each trough (right or left) by stra-
tified randomization, and assigned 20
ml of clean water (2 ice aliquots) to the
opposite end.
In all treatments, we placed a single
tadpole covered with a release cage in
each gutter, and waited 5 min for habi-
tuation. Then we deposited the test solu-
tion ices (zebra fish for experimental
tadpoles or clean water for control tad-
poles) and we began trials by slowly lif-
ting the cages above each tadpole 5 min
after we deposited the test solution ices
aliquots (i.e. after the ices aliquots had
entirely thawed). We subsequently stood
as motionless as possible recording from
a hidden point the quadrant that each
tadpole occupied at 1 min intervals for
30 min. We calculated levels of activity
from the number of lines crossed by each
tadpole during the observation period
(Rohr and Madison 2001, Gonzalo et al
2007). Diffusion of chemicals in still
water may be a slow process. However,
all individual tadpoles used in the expe-
riment were observed at least once in all
of the subdivisions of the gutter, so we
were confident that all tadpoles were re-
ally exposed to the chemical stimuli.
Moreover, tadpoles often showed episo-
des of fast swimming which should
contribute to diffuse chemicals in water.
experimental design
We randomly assigned tadpoles to six
treatments of 30 individuals each (15
control and 15 experimental). Thus, each
individual tadpole only participated in
one treatment either as control or expe-
rimental. In each treatment, tadpoles
followed a different pre-exposure se-
quence and combination of exposure to
chemical cues of zebrafish and/or cons-
pecific alarm cues (see below). In short,
treatment 1 was a control of basal acti-
vity of tadpoles when exposed to alarm
cues alone. Treatments 2 and 3 were con-
trols of predator learning (treatment 2
for testing the existence of classical
conditioning learning, and treatment 3
for testing whether older fish cues
still allowed tadpoles learning preda-
tors.). Treatments 4 and 5 tested the
existence of learning irrelevance. Treat-
ment 6 tested the existence of latent
inhibition.
Treatment 1: Exposure to alarm cues
alone (basal activity)
This experiment was performed to
test whether the exposure in consecutive
days to alarm cues alone altered the
66

behavioral pattern of activity of tadpoles.
This was a control to test whether expo-
sure to alarm cues alone had any effect
on tadpoles’ general activity, indepen-
dently of the presence of other cues, and
whether this effect was maintained
throughout time. All 30 tadpoles within
this treatment (maintained in six diffe-
rent aquaria) were exposed to alarm
cues alone on days 2 and 4 and to clean
water on days 1 and 3, On day 6, tadpoles
were individually placed in the observa-
tion aquaria (see general methodology
above) and tested randomly with either
zebrafish cues (experimental group,
N=15) or clean water (control group,
N=15).
Treatment 2: Classical conditioning
Green frog tadpoles are able to learn
new cues as dangerous (Gonzalo et al.
2006), so this treatment was designed to
confirm whether tadpoles were able to
learn association of fish cues with high
predation risk. Also, we used this
treatment as a control of tadpole beha-
vior after learning, which could be com-
pared with potential learning occurring
in treatments 4, 5 and 6. All tadpoles
(N=30; maintained in six different aqua-
ria) were trained by exposing them si-
multaneously to zebrafish cues and
alarm cues on days 2 and 4, and to clean
water on days 1 or 3. On day 6, tadpoles
were placed individually in the observa-
tion aquaria (see above) and tested ran-
domly with either zebrafish cues
(experimental, N=15) or clean water
(control, N=15).
Treatment 3: Sequential exposure (wi-
thout removal) followed by simultaneous
exposure
This treatment simulated a natural
situation where tadpoles lived constantly
exposed to the chemical cues of a simu-
lated predator (zebrafish) that someti-
mes ate conspecifics, which released
alarm cues. During the first four days, all
30 tadpoles within this group (maintai-
ned in six different aquaria) were trained
by exposing them on alternate days to ze-
brafish cues alone followed the next day
with alarm cues alone. We did not change
water of aquaria between days, to maxi-
mize time that zebrafish cues were inside
the aquaria. On day 5, all tadpoles were
exposed to the simultaneous presenta-
tion of zebrafish cues and alarm cues for
2 h. On day 6, tadpoles were individually
placed in the observation aquaria (see
above) and tested randomly with either
zebrafish cues (experimental) or clean
water (control).
Treatment 4: Sequential exposure (with
removal) followed by simultaneous expo-
sure
This treatment was designed to test
whether tadpoles showed the pheno-
mena of learning irrelevance. We simu-
lated a natural situation where tadpoles
lived with a non predator (zebrafish),
which chemical cues appeared indepen-
dently of predation events. During the
first four days, all 30 tadpoles within this
group (maintained in six different aqua-
ria) were trained with presentation on
alternate days of zebrafish cues alone fo-
llowed the next day with alarm cues
alone. Zebrafish cues were placed in the
training aquarium for only 2 h., and the-
reafter we siphoned about 85% of the
water from the training container (as in
Hazlet 2003) and immediately replaced
it with clean water (at room temperature
to avoid temperature shock). Water was
removed to avoid the permanence of
high concentration of zebrafish cues in
the water, thus avoiding the mix of alarm
and zebrafish cues. Exposure to fish cues
was performed on either days 1 and 3 or
days 2 and 4 and alarm cues were added
67

on the alternate days. On day 5, all tadpo-
les were exposed simultaneously to ze-
brafish and alarm cues together for 2 h.
On day 6, tadpoles were individually pla-
ced in the observation aquaria (see
above) and tested randomly with either
zebrafish cues (experimental, N=15) or
clean water (control, N=15).
Treatment 5: Simultaneous exposure
followed by sequential exposure (with
removal)
This experiment was also designed to
test whether tadpoles present the pheno-
mena of learning irrelevance. However, in
nature, a series of independent exposu-
res to two types of cues (predator and
alarm) could occur either before or after
the simultaneous detection of those cues.
Thus, tadpoles were trained in a similar
way than in the treatment 4, but in this
experiment, on day 1, all tadpoles (N=30)
were first exposed simultaneously to ze-
brafish cues and alarm cues for 2 h (with
draining of 85% of water after the expo-
sure to remove the zebrafish cues)(Haz-
let 2003). Then all tadpoles were
exposed to zebrafish cues alone (with
posterior draining of 85% of water) on
either days 2 and 4 or on days 3 and 5
and to alarm cues alone on alternate
days. On day 6, tadpoles were indivi-
dually placed in the observation aquaria
(see above) and tested randomly with ei-
ther zebrafish cues (experimental, N=15)
or clean water (control, N=15).
Treatment 6: Exposure to fish cues alone
followed by simultaneous exposure
This experiment was designed to test
whether tadpoles present the pheno-
mena of latent inhibition. We simulated
a natural situation where tadpoles lived
with a predator (zebrafish), but where
there were not predation events during
several days, but only until the last day of
training. All tadpoles (N=30) were expo-
sed to zebrafish cues alone on days 2 and
4 (without water removal after the expo-
sure) and to clean water on days 1 and 3.
On day 5, all tadpoles were simultane-
ously exposed to zebrafish cues and to
alarm cues together for 2 h. On day 6,
tadpoles were tested individually (see
above) with either zebrafish cues (expe-
rimental, N=15) or clean water (control,
N=15).
data analyses
Each treatment was analyzed separa-
tely. We used one way ANOVAs to test for
differences in activity level of tadpoles on
day 6 (number of lines crossed in the
observation aquaria; log transformed)
between control (clean water) and expe-
rimental (zebrafish cues) groups of tad-
poles (Sokal and Rohlf 1995).
results
Effects of alarm cues alone
In treatment 1,: there were no signi-
ficant differences in activity on day 6 bet-
ween control and experimental tadpoles
(‘treatment’ effect: F1.28=0.41, P=0.50).
Thus, the previous exposure to alarm
cues alone did not have effects on the ac-
tivity of tadpoles when they later found
fish cues alone (Fig. 1).
Classical conditioned learning and dura-
tion of fish chemical cues
In treatment 2, there were significant
differences in activity on day 6 between
control and experimental tadpoles (‘tre-
atment’ effect: F1.28=9.99, P=0.003;).
Thus, tadpoles showed evidence of the
persistence of a learned association bet-
ween zebra fish cues and elevated preda-
tion risk by the significant later reduction
of activity in response to zebra fish cues
68
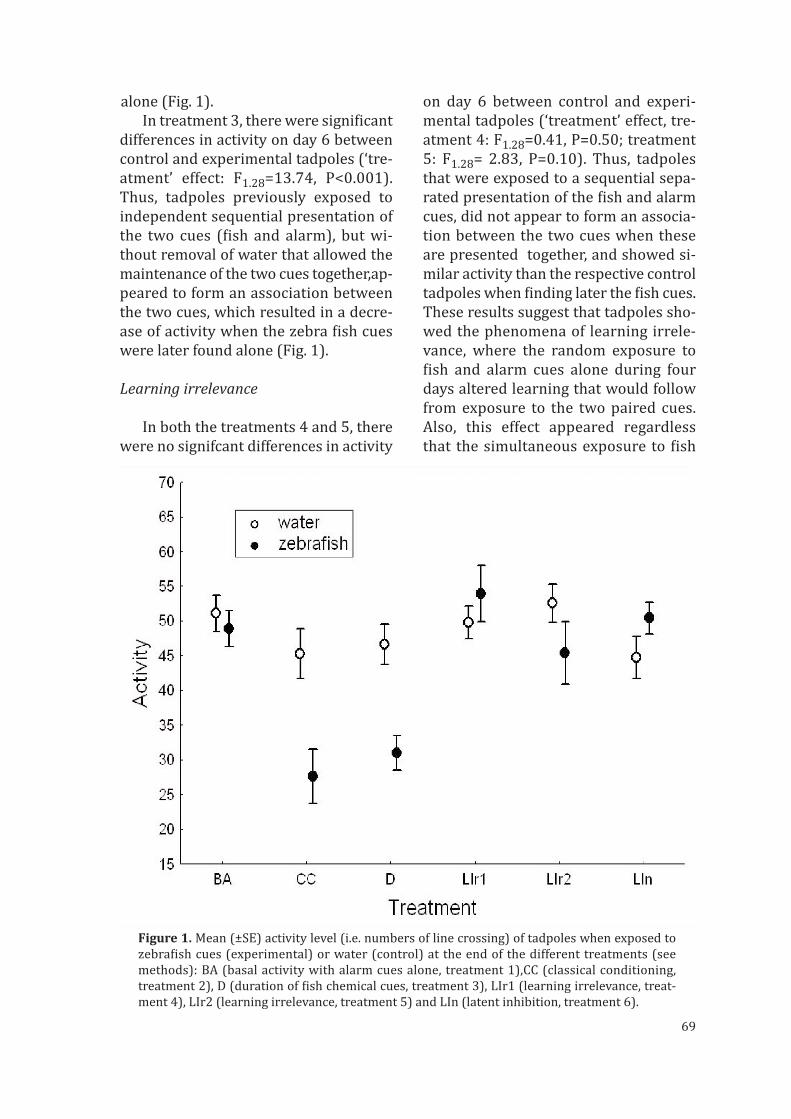
alone (Fig. 1).
In treatment 3, there were significant
differences in activity on day 6 between
control and experimental tadpoles (‘tre-
atment’ effect: F1.28=13.74, P<0.001).
Thus, tadpoles previously exposed to
independent sequential presentation of
the two cues (fish and alarm), but wi-
thout removal of water that allowed the
maintenance of the two cues together,ap-
peared to form an association between
the two cues, which resulted in a decre-
ase of activity when the zebra fish cues
were later found alone (Fig. 1).
Learning irrelevance
In both the treatments 4 and 5, there
were no signifcant differences in activity
on day 6 between control and experi-
mental tadpoles (‘treatment’ effect, tre-
atment 4: F1.28=0.41, P=0.50; treatment
5: F1.28= 2.83, P=0.10). Thus, tadpoles
that were exposed to a sequential sepa-
rated presentation of the fish and alarm
cues, did not appear to form an associa-
tion between the two cues when these
are presented together, and showed si-
milar activity than the respective control
tadpoles when finding later the fish cues.
These results suggest that tadpoles sho-
wed the phenomena of learning irrele-
vance, where the random exposure to
fish and alarm cues alone during four
days altered learning that would follow
from exposure to the two paired cues.
Also, this effect appeared regardless
that the simultaneous exposure to fish
figure 1. Mean (±SE) activity level (i.e. numbers of line crossing) of tadpoles when exposed to
zebrafish cues (experimental) or water (control) at the end of the different treatments (see
methods): BA (basal activity with alarm cues alone, treatment 1),CC (classical conditioning,
treatment 2), D (duration of fish chemical cues, treatment 3), LIr1 (learning irrelevance, treat-
ment 4), LIr2 (learning irrelevance, treatment 5) and LIn (latent inhibition, treatment 6).
69

and alarm cues occurred before (treat-
ment 5) or after (treatment 4) the inde-
pendent presentation of the cues alone
(Fig. 1).
Latent inhibition
In treatment 6, there were no signifi-
cant differences in activity on day 6
between control and experimental tad-
poles (‘treatment’ effect: F1.28=2.71,
P=0.10). Thus, tadpoles did not show an
association of zebrafish cues with preda-
tion risk when they were exposed to ze-
brafish cues alone for four days before
the simultaneous presentation of the ze-
brafish cues mixed with alarm cues. This
result suggested that tadpoles showed
the phenomena of latent inhibition
where the previous frequent exposure to
the fish stimulus alone (without any rein-
forcement with alarm cues) resulted in a
reduction of the strength of the learned
association that could be formed later
with the simultaneous presentation of
fish and alarm cues (Fig. 1).
dIscussIon
The learning ability in predator re-
cognition has been demonstrated in a
wide variety of taxa (review in Chivers
and Smith 1998). In almost every case,
the experiment protocol was an easy
classical conditioning experiment, in
which a single paired presentation of a
predator odour and alarm cues induces
an antipredator behaviour in a post ex-
posure to the predator odour alone. But,
in nature, animals continually receive
information about their environment
and must filter this information to focus
on those aspects most important to sur-
vival (Dukas 2002). Our experiments cle-
arly demonstrate (as in Hazlet 2003) that
the temporal pattern of exposure to pre-
dator and alarm cues completely deter-
mines whether a prey organism will
form a learned association with inputs
from a successful predation event, which
can be used when the predator cues will
be later detected alone.
Predator cues can persist in the ha-
bitat for a long time (Hazlett 2003). Fish
cues persist for more than one day
(Hazlett 2003; present study) whereas
the duration of alarm cues is shorter; Fe-
rrari et al. (2008) showed than in natural
conditions wood frog (Rana sylvatica)
alarm cues suffer a quickly degradation
in natural conditions, but nothing is
know about the duration in laboratory
conditions (as in our experiment). Only
Hazlet (1999) tested the persistence of
crayfish (Orconectes virilis) alarm cues in
artificial conditions, and concluded that
alarm cues persists at least 6 hours. Our
study showed than tadpoles can form an
association between predator cues and
alarm cues even when they were presen-
ted in different days. Thus, one day old
fish cues were effective in eliciting the
learning association in tadpoles.
In other laboratory experiment, clas-
sically conditioned green frog tadpoles
reduced their activity in response to the
fish chemical cues, and this response
could last at least nine days without
further reinforcement (Gonzalo et al. in
press). However, in nature, tadpoles are
not exposed to the presence of a single
event of paired cues, but to a mix of
different types of cues. So, the ability of
learning to recognize a predator could be
more complex than just detecting two
paired cues simultaneously. Avoiding
erroneous learning could help animals to
avoid nonadaptative responses to chemi-
cal cues of non-dangerous species that
sometimes may appear randomly pairing
with alarm cues.
Learned irrelevance occurs when the
random presentation of the two cues
indicates to the prey that the two cues
are not causally linked. Hazlett (2003)
demonstrated the existence of this
70

phenomenon in crayfish. Likewise, in our
study, tadpoles failed to form an associa-
tion between an unconditioned stimulus
and a novel cue when they had been,
previously or posteriorly, exposed to the
two cues separately. These results sho-
wed than tadpoles are able to learn that
the two types of cues are no causally con-
nected if the two cues are found indepen-
dently before or after they are found
simultaneously. Therefore, our results
demonstrate for the first time the exis-
tence of the process of learned irrele-
vance in frog tadpoles.
Latent inhibition occurs when a
previous exposure to a neutral stimulus
indicates to the animal that this cue is
not linked with others with biological
significance. Latent inhibition has been
demonstrated in crayfish (Acquistapace
et al. 2003) and in fathead minnows (Fe-
rrari and Chivers, 2006a). Likewise, in
our experiment tadpoles also failed to
form the association when they were
previously exposed to the unconditioned
fish stimulus alone for four days before
the unconditioned (fish) and the condi-
tioned (alarm) cues were presented
together. Therefore, our results demons-
trate for the first time the existence of the
effect of latent inhibition in frog tadpo-
les.
The ability to learn potential preda-
tors may be especially important for the
survivorship of prey species that are li-
kely to find a high variety of predators
while they are in the aquatic phase. Thus,
learning may enable animals to adjust
their behaviour in variable environ-
ments. However, to always respond to a
chemical cue as dangerous only because
it coincidentally appeared once mixed
with chemical alarm cues could be very
costly (Lima and Dill 1990; Belden et al.
2000). Thus, both learned irrelevance
and latent inhibition far away to be con-
sidered as a fail in predator recognition,
can be rather seen as adaptive ways for
dealing with conflicting information and
as strategies to avoid learning irrelevant
information. Nevertheless, these two
processes could also limit the ability of
prey to learn new potential predators
(Ferrari and Chivers 2006a), for example,
in the case of the Iberian green frog tad-
poles, this mechanism could interfere in
the learning of chemicals from those
predators that usually capture other
species but opportunistically attack also
tadpoles.
Both learning irrelevance and latent
inhibition seem to produce the same
effect, but typically the exposure to both
conditioned and unconditioned stimuli
un-correlated with each other retards
subsequent learning more than the expo-
sure to one type of stimulus alone (Ben-
net et al 1995). So learning irrelevance
seems to be even more disruptive than
latent inhibition (Bennet et al 1995).
Ferrari and Chivers (2006b) showed that
recently information plays a major role
in elicit antipredator responses with
more or less intensity in fathead min-
nows when they are exposed to predator
cues. So, if the information is always up-
dated, it is likely that tadpoles that lear-
ned irrelevance of the fish cue, or that
showed latent inhibition to the fish cue,
would be able to learn later that the fish
cue is a predator cue if they were reinfor-
ced repeatedly with alarm cues. Future
work should tell us if both processes
affect in a different way this possible
posterior learning ability, and if learning
irrelevance retards more than latent
inhibition the posterior associative
learning.
references
Acquistapace P, Hazlett BA, Gherardi F (2003)
Unsuccessful predation and learning of
predator cues by crayfish. J Crust Biol
23:364–370
71

Allen MT, Chelius L, Masand V, Gluck MA, Myers
CE, Schnirman G (2002) A Comparison of
Latent Inhibition and Learned Irrelevance
Pre-Exposure Effects in Rabbit and
Human Eyeblink Conditioning. Integr
Physiol Behav Sci37:188–214
ASIH (2004) Guidelines for use of live Amphi-
bians and Reptiles in field and laboratory
research. Lawrence, Kansas: Herpetologi-
cal Animal Care and Use Committee
(HACC) of the American Society of Ichth-
yologists and Herpetologists
Belden LK, Wildy EL, Hatch AC, Blaustein AR
(2000) Juvenile western toads, Bufo bo-
reas, avoid chemical cues of snakes fed ju-
venile, but not larval, conspecifics. Anim
Behav 59:871–875
Bennett C, Maldonado A, Mackintosh N (1995)
Learned irrelevance is not the sum of ex-
posure to CS and US. Q J Exp Psychol B,
Comp Physiol Psychol 48B:117–128
Bennet CH, Wills SJ, Oaakeshott SM, Mackintosh
NJ (2000) Is the context specificity of la-
tent inhibition a sufficient explanation of
learned irrelevance?. Q J Exp Psychol B,
Comp Physiol Psychol 53B:239–253
Chivers DP, Smith RJF (1998) Chemical alarm
signalling in aquatic predator–prey
systems: a review and prospectus. Ecos-
cience 5:338–352
Chivers DP, Wisenden BD, Smith RJF (1996)
Damselfly larvae learn to recognize preda-
tors from chemical cues in the predator’s
diet. Anim Behav 52:315–320
Dodson SI, Crowl TA, Peckarsky BL, Kats LB, Co-
vich AP, Culp JM (1994) Nonvisual commu-
nication in fresh-water benthos: an
overview. J N Am Benthol Soc 13:268–282
Dukas R (2002) Behavioural and ecological con-
sequences of limited attention. Phil Trans
R Soc Lond B. 357:1539–547
Ferguson HJ, Cobey S, Smith BH (2001) Sensiti-
vity to change in reward is heritable in the
honeybee, Apis mellifera. Anim Behav
61:527–534
Ferrari MCO, Chivers DP (2006a) The role of la-
tent inhibition in acquired predator recog-
nition by fathead minnows. Can J Zool
84:505–509
Ferrari MCO, Chivers DP (2006b) The role of le-
arning in the development of threat-sensi-
tive predator avoidance: how do fathead
minnows incorporate conflicting informa-
tion? Anim Behav 71:19–26
Ferrari MCO, Messier F, Chivers DP (2008) Degra-
dation of chemical alarm cues under natu-
ral conditions: risk assessment by larval
woodfrogs. Chemoecology 17:263–266
García-París M (2000) Rana perezi. In: Ramos MA
(ed.) Amphibia, Lissamphhibia. Fauna Ibe-
rica vol 24. Museo Nacional de Ciencias Na-
turales (CSIC), Madrid, pp: 467–478
Gonzalo A, López P, Martín J (2007) Iberian green
frog tadpoles may learn to recognize novel
predators from chemical alarm cues of
conspecifics. Anim Behav 74:447-453
Gonzalo A, López P, Martín J (in press) Learning,
memorizing and apparent forgetting of
chemical cues from new predators by Ibe-
rian green frog tadpoles. Anim Cogn
Gosner KL (1960) A simplified table for staging
anuran embryos and larvae with notes on
identification. Herpetologica 16:183–190
Hazlett BA (1999) Responses to multiple chemi-
cal cues by the crayfish Orconectes virilis.
Behaviour 136:161–177
Hazlett BA (2003) Predator recognition and lear-
ned irrelevance in the crayfish Orconectes
virilis. Ethology 109:765–780
Kats LB, Dill LM, (1998) The scent of death: che-
mosensory assessment of predation risk by
prey animals. Ecoscience 5:361–394
Kiesecker JM, Chivers DP, Blaustein AR (1996)
The use of chemical cues in predator recog-
nition by western toad tadpoles. Anim
Behav 52:1237–1245
Lima SL, Dill LM (1990) Behavioral decisions
made under the risk of predation: a review
and prospectus. Can J Zool 68:619–640
Mirza RS, Ferrari MCO, Kiesecker JM, Chivers DP
(2006) Responses of American toad tadpo-
les to predation cues: behavioural response
thresholds, threat-sensitivity and acquired
predation recognition. Behaviour 143:877–
889
Mathis A, Smith RJF (1993) Fathead minnows,
Pimephales promelas, learn to recognize
northern pike, Esox lucius, as predators on
the basis of chemical stimuli from minnows
in the pike’s diet. Anim Behav 46:645–656
Petranka JW, Kats LB, Sih A (1987) Predator–prey
interactions among fish and larval amphi-
bians: Use of chemical cues to detect pre-
datory fish. Anim Behav 35:420–425
Rohr JR, Madison DM (2001) A chemically media-
ted trade-off between predation risk and
mate search in newts. Anim Behav 62:863–
869
Sokal RR, Rohlf FJ (1995) Biometry. Freeman, NY.
Von Frisch K (1938) Zur psychologie des fische-
schwarmes. Naturwiss 26:601–606
Woody DR, Mathis A (1998) Acquired recognition
of chemical stimuli from an unfamiliar
predator: associative learning by adult
newts, Notophthalmus viridescens. Copeia
1998:1027–1031
72

cAPÍTuLO iV
EVALUACIÓN qUIMIOSENSORIAL DEL RIESGO
DE DEPREDACIÓN EN RENACUAJOS DE RANA
VERDE IBÉRICA: DIFERENCIAS ENTRE EL APREN-DIZAJE DE DEPREDADORES MEDIANTE SUSTAN-CIAS DE ALARMA O SUSTANCIAS DE ESTRÉS
CHEMOSENSORY ASSESSMENT OF PREDATIONRISk BY IBERIAN GREEN FROG TADPOLES: DIF-FERENCES wHEN LEARNING NEw PREDATORSFROM ALARM OR DISTURBANCE CUES


En los ambientes acuáticos muchas presas confían en la informa-ción quimiosensorial proveniente de individuos coespecíficos heridos(sustancia de alarma) o estresados (sustancias de estrés) para evaluarel riesgo de depredación. Las sustancias de alarma son consideradascomo una señal de alto riesgo mientras que las señales de estrés debenser percibidas como de bajo riesgo. Estas señales químicas también pue-den ser utilizadas por los individuos para aprender a nuevos depredado-res potenciales. En este trabajo hemos examinado si los renacuajosmuestran una respuesta antidepredadora cuando se les presentan seña-les de alarma o señales de estrés, y si son capaces de aprender a reconocery memorizar señales químicas de nuevos depredadores asociándolos asustancia de alarma o de estrés. Los resultados muestran que los rena-cuajos redujeron su actividad en presencia de las señales de estrés, peroen menor grado que con las sustancias de alarma de individuos coespe-cíficos. Además, los renacuajos aprendieron nuevos depredadores graciasa las asociaciones con sustancias de alarma o de estrés. Sin embargo, elperiodo de retención del la asociación fue mucho más corto para las sus-tancia de estrés que para las sustancias de alarma. De este modo, los re-nacuajos de rana verde ibérica se comportan de acuerdo a la hipótesisde la respuesta dependiente del riesgo de depredación cuando valoran elriesgo de depredación asociado aun depredador dependiendo si el pro-ceso de aprendizaje ha sido debido a sustancia de alarma o una sustanciade estrés.
Este cápitulo se corresponde con: Gonzalo A, López P, Martín J. Chemosensory assessment of predationrisk by Iberian green frog tadpoles: differences when learning new predators from alarm or distur-bance cues . Behav Ecol Sociobiol (Submited manuscript).


chemosensory assessment of predatIon rIsk by IberIan
green frog tadpoles: dIfferences when learnIng new
predators from alarm or dIsturbance cues
In aquatic environments many prey rely on chemosensory information from injured
(alarm cues) or stressed conspecifics (disturbance cues) to assess predation risk. Alarm
cues are considered as a sign of higher risk than disturbance cues. These cues could be
used by prey to learn potential new predators. Here we tested whether Iberian green frog
tadpoles (Pelophylax perezi) exhibited antipredator responses to alarm and disturbance
cues of conspecifics, and whether tadpoles were able to learn and memorize cues of new
predators that had been previously associated with alarm or disturbance cues. Tadpoles
reduced their activity in presence of disturbance cues, but weakly when compared with
their response to alarm cues. Also, tadpoles learned new predators thanks to their
association with alarm or disturbance cues. However, the period of retention of the
learning association was shorter for disturbance than for alarm cues. Tadpoles seemed
to behave according to the threat sensitive predator avoidance hypothesis when assessing
risk level of a predator depending on the learning process had been with disturbance
or alarm cues.
IntroductIon
In aquatic habitats diverse prey
organisms rely on chemosensory infor-
mation to assess local predation risk
(Smith 1992; Chivers and Smith 1998).
Aquatic environments are ideal for the
solution and dispersal of chemicals cues,
which are especially useful in turbid
water and habitats highly structured or
for species with poor developed visual
senses (Wisenden 2000). Antipredatory
or defensive responses occur to chemi-
cals cues released by predators but also
to chemicals cues released by other prey
(i.e. alarm signals) (Kats and Dill 1998).
Chivers and Smith (1998) divided
chemical alarm signaling systems into
two general categories based on the
point in the predation sequence when
cues are emitted. Damage-released alarm
cues are those chemicals released by
prey animals only upon being captured
by a predator. In contrast, disturbance
cues are chemicals that are released by
senders that have been disturbed or
stressed but not captured by a predator.
Damage-released alarm cues have been
found in a variety of aquatic animals (e.g.
fish: Smith 1992; Chivers and Smith
1994; amphibians: Pfeiffer 1966; Hews
and Blaustein 1985; Wilson and Lefcort
1993; insects: Sih 1986; crustacea:
Hazlett 1994; sea anemones: Howe and
Sheikh 1975; echinoderms: Lawrence
1991; gastropods: Appleton and Palmer
1988; review in Chivers and Smith
1998).
Chemical disturbance cues also have
been found in aquatic animals (crayfish:77

Hazlett 1985, 1989, 1990a; hermit crabs:
Hazlett 1990b; fishes: Wisenden et al.
1995; Jordão and Volpato 2000; Mirza
and Chivers 2002; amphibian tadpoles:
Kiesecker et al. 1999; Bryer et al 2001),
but they have received less attention
than alarm cues. Iowa darters, Etheos-
toma exile, increase antipredator beha-
vior when exposed to chemical cues of
disturbed conspecifics (Smith 1979; Wi-
senden et al. 1995). Both crayfish, Orco-
nectes virilis, and hermit crabs, Calcinus
lavimanus, increase antipredator beha-
vior when exposed to chemical cues re-
leased by disturbed conspecifics (Hazlett
1985, 1990b). In addition, O. virilis res-
ponds to disturbance cues produced by
numerous heterospecifics, including
other crayfish species, leeches, newts
and fish (Hazlett 1985, 1989, 1990a).
This suggests that disturbance cues have
a composition pretty common across dif-
ferent taxa.
Kiesecker et al. (1999) showed that
red-legged frog tadpoles release ammo-
nium (NH4+) upon being disturbed by a
predator, and that conspecific tadpoles
respond to the ammonium by increasing
antipredator behaviors. Also, red-legged
frog tadpoles respond with antipredator
behavior upon detecting a pulse of
commercial ammonium. So, Kiesecker et
al. (1999) argued that the disturbance
cue may be ammonium, excreted from
the gills or urine during periods of incre-
ased metabolic activity that is required
for effective escape or when prey are
exposed to some stressful situation such
as the presence of a predator. This rele-
ase of disturbance cues may not be
intentional but may represent a normal
physiological process to which other
individuals have become sensitive
(Mirza and Chivers 2002).
The release and detection of alarm
and disturbance cues have important im-
plications for predator–prey interactions.
Prey animals that detect alarm or distur-
bance cues have an early warning of the
presence of a predator and may be able
to avoid an encounter by leaving the area
(Jordão 2004) or by reducing move-
ments (Mirza and Chivers 2002) and
becoming cryptic. Early detection of a
predator’s presence will allow prey to
increase vigilance, which will probably
result in an improved chance of survival
should the encounter escalate to an
attack (Hews 1988; Mathis and Smith
1993).
However, according to the threat sen-
sitive predator avoidance hypothesis
(Helfman 1989), natural selection should
favor individuals that take action appro-
priate to the magnitude of threat, which
would require an accurate discrimina-
tion of the current level of risk that each
predator poses. Thus, the response of
prey to alarm cues should be stronger
than the response to disturbance cues, as
disturbance cues could be perceived as
lower level risk than alarm cues (Chivers
and Smith 1998). Disturbance cues are
thought to be low-level indicators of risk
to which prey animals respond with
antipredator behavior, but the detection
of disturbance cues can provide a survi-
val benefit during an encounter with a
predator (Mirza and Chivers 2002).
Disturbance cues are perceived as
low-level indicators of threat but they
might still facilitate learning to recognize
potential predators (Chivers and Smith
1998). Mirza and Chivers (2002) showed
that damage-released alarm cues could
facilitate learning recognition of preda-
tors. Recently, Ferrari et al. (2008)
showed than juvenile rainbow trout use
the disturbance cues as a warning cue,
but trouts are not able to use disturbance
cues to learn new predators. To our
knowledge there were no studies about
the capacity of amphibians to retain the
memory of a new predator learned by
distressed cues. As disturbance cues
are low level indicators of threat we
78

hypothesized that, if tadpoles are able to
learn new predators by associating them
to disturbance cues, tadpoles should
react to the new predator chemical cues
more weakly, and retain the memory of
this predator for less time, than tadpoles
that have learnt the predator by associa-
ting it to alarm cues.
In our experiments, we first tested
whether Iberian green frog tadpoles
(Pelophylax perezi) exhibited an antipre-
dator response, and which was the
magnitude of this response, to damage-
released alarm cues from injured cons-
pecifics and to chemical cues from
disturbed conspecifics. Moreover, an ad-
ditional experiment examined whether
tadpoles were able to learn to associate
chemical cues from non-predatory exotic
fish species (to which tadpoles can not be
genetically predisposed) with predation
risk through their association with the si-
multaneous presence of alarm cues of
conspecifics, or cues of disturbed cons-
pecifics, and whether each type of cue re-
sulted in different periods of retention of
the learning association.
methodsstudy animals
Iberian green frog tadpoles (SVL,
Mean±SE=5.4±0.2 cm, Gosner’s stage:
25; see Gosner 1960) were collected by
netting at several small ponds in Collado
Mediano (Madrid, central Spain). Tadpo-
les were housed in groups of five at “El
Ventorrillo” Field Station, 10 km from the
capture area, in plastic aquaria (49 x 29
cm and 25 cm high) with 5 L. of water at
ambient temperature and under a natu-
ral photoperiod. They were fed every day
with commercial fish flakes.
We obtained from a commercial dea-
ler non predatory zebra danio fish
(Brachyodanio rerio) to be used as source
of neutral scent. Before and after the
experiment finished, fishes were main-
tained in a large filtered aquarium and
regularly fed with commercial fish flakes.
All the animals were healthy during
the trials, all maintained or increased
their original body mass. The experi-
ments were performed under license
from the “Consejería de Medioambiente
de la Comunidad de Madrid” (the Envi-
ronmental Agency of the local Govern-
ment of Madrid). Procedures are
conformed to recommended guidelines
for use of live Amphibians in laboratory
research (ASIH 2004).
preparation of chemical stimuli
Alarm cues of tadpoles were
prepared from three tadpoles (SVL,
Mean±SE=4.2±0.1 cm). They were cold
anesthetized by placing at 4 °C for 20
min, inducing them deep hypothermia,
and, then, euthanasied with a quick blow
to the head to avoid suffering (ASIH
2004). We did not use a chemical anaes-
thetic, because these chemicals may in-
terfere with natural tadpoles’ chemical
cues in subsequent trials. The extract
was then prepared by putting these dead
tadpoles in a clean disposable plastic
dish, and macerating them in 3000 mL of
distilled water. The stimulus water was
then filtered through absorbent paper to
remove solid particles, and immediately
frozen in 10 mL portions until used
(Woody and Mathis 1998).
Disturbance cues of tadpoles were
prepared from 20 tadpoles that were pla-
ced into a 200 mL aquarium and stressed
by simulating a predator attack with
a wooden bird model for 30 s. The pre-
dator attacks consisted in moving the
wooden model around the aquarium
simulating ten trays to catch the tadpoles
with the beak. Care was taken not to
touch or damage any of the tadpoles.
As we did not know the period of degra-
dation of the disturbance cues, we prepa-
red them immediately before each
79

experiment, and quickly used them to
avoid degradation. Ten different tadpoles
were used each time and then kept sepa-
rately, all 20 tadpoles used (10 for expe-
riment) behaved normally 1 h after being
stressed.
The fish chemical stimulus was prepa-
red by placing ten zebra danio fishes into
a 10 L aquarium with clean water for
three days. This aquarium was aerated
but not filtered. Fishes were not fed du-
ring this short period to avoid contami-
nating water with food odor. Thereafter,
water was drawn from the aquaria and
frozen in 10 mL portions until its use in
experiments. Fishes were returned and
fed in their home large aquaria. We pre-
pared control water in an identical man-
ner but without placing fish in the
aquaria (Woody and Mathis 1998).
Experiment 1: responses of tadpoles to
alarm and disturbance cues
To know whether Iberian green frog
tadpoles were able to detect disturbance
cues we planed an experiment were they
were exposed to either (1) chemical cues
from conspecifics disturbed with the
wooden bird model, (2) alarm cues from
conspecifics, or (3) clean water as a
control of basic activity levels.
Tadpoles were tested individually in
grey, U-shaped gutters (101 x 11.4 cm
and 6.4 cm high) sealed at both ends with
plastic caps. We marked the internal part
of the gutters with four crossing lines
that created five subdivisions of equal
surface. We filled each gutter with 3 L of
clean water at 20ºC that was obtained
from a mountain spring that not contai-
ned fish. We placed clear plastic over
each trough on either side of the cage to
isolate the system from air movements in
the testing room (see Rohr and Madison
2001). Each trial lasted 1 h and consisted
of a 30 min pre-stimulus period and a 30
min post- stimulus period separated by a
stimulus introduction. We assigned 10
mL of test solutions (water, alarm cues or
disturbance cues) to one end of each
trough (right or left) by stratified rando-
mization. We placed a single tadpole in
each gutter, and waited 5 min for habi-
tuation. Then, we started the ‘pre-stimu-
lus’ test (30 min). Thereafter, we added
the test solution and, immediately after
we started the ‘post-stimulus’ period (30
min). During both the pre- and the post-
stimulus periods we recorded from a
blind the quadrant that each tadpole oc-
cupied at 1 min intervals for 30 min. We
calculated levels of activity from the
number of lines crossed by each tadpole
during the observation period (Rohr and
Madison 2001, Gonzalo el al. 2007). We
used 45 tadpoles, 15 assigned randomly
to each treatment. Tadpoles only were
tested once and then discarded for the
next experiment. All individual tadpoles
used in the experiment were observed at
least once in all of the subdivisions of the
gutter, so we were confident that all
tadpoles were really exposed to the
chemical stimuli.
For each trial we calculated levels of
activity as the difference between the
numbers of line crossing between the
pre- and post-stimulus periods. Positive
values indicated increased movement
following addition of the stimulus; nega-
tive values indicate decreased activity.
Data were log transformed and then tes-
ted with a one way ANOVA, with the tre-
atment as a categorical between variable.
Subsequent post hoc multiple compari-
sons were made using Tukey’s pairwise
comparisons (Sokal and Rohlf 1995).
Experiment 2: retention of the learning as-
sociation
We designed this experiment to deter-
mine whether tadpoles were able to
learn to recognize novel predators by as-
sociating the predator chemical cues
80

with disturbance cues from conspecifics,
and to know, in case that learning occu-
rred, the duration of the memory of this
predator recognition.
We randomly assigned twelve groups
of five tadpoles to three different
treatments (‘control’, ‘alarm’ and‘distur-
bance’). On the fist day of the experiment
tadpoles from the ‘control’ treatment
were exposed to the fish chemical cues
alone mixed with clean water. Simultane-
ously tadpoles from the ‘alarm’ treat-
ment were exposed to both the scent of
the fish and conspecific chemical alarm
cues, thus, simulating the cues from a
predatory fish that was eating a conspe-
cific tadpole. Similarly, tadpoles from the
‘disturbance’ treatment were exposed to
both the scent of the fish and conspecific
disturbance cues, thus, simulating the
cues from a predatory fish that failed an
attack to conspecific tadpoles but stres-
sed them. Previous studies showed that
tadpoles conditioned with the mix of fish
and alarm cues were two days later able
to recognize the fish chemical cues alone
as coming from a predator (Gonzalo et al.
2007) and were able to remember the
predators al least nine days after being
conditioning with alarm cues (Gonzalo el
al. in press). Thus tadpoles from the
alarm group were considered as condi-
tioned.
To test for the duration of the memory
of predator recognition, on days 1, 3, 6 or
9 after the initial conditioning, different
individual tadpoles from the three treat-
ments were tested with the fish chemical
cues alone in clean water. The experi-
ment was carrying on using the same
experimental procedure than in the pre-
vious Experiment 1. All tadpoles from the
three groups were tested with 10 mL of
fish scent. Each trial lasted 1 h and con-
sisted of a 30 min pre-stimulus period
and a 30 min post-stimulus period sepa-
rated by a stimulus introduction. Indivi-
dual form the three groups were tested
in parallel and observations were carried
out blind. Each day, we tested 15 indivi-
duals from each of the three groups
(control, alarm and disturbed). These
tadpoles were chosen randomly from the
several groups we had, and, after the trial
were kept separately and not used in
subsequent trials. New individual tadpo-
les were used in each trial to ensure that
we were just testing the capacity of re-
tention of the learning association. We
did not use the same individual tadpoles
in more than one test because they could
progressively learn the irrelevance of
the “non-dangerous” predator (Hazlett
2003) or habituate to the predation pres-
sure, so that the results might not reflect
duration of memory alone and will be
unclear.
For each trial, we calculated levels of
activity as the difference between the
numbers of line crossing between the
pre- and post-stimulus periods. Positive
values indicated increased movement fo-
llowing addition of the stimulus; negative
values indicated decreased activity. Data
were log transformed and then tested by
general linear modeling (GLM; Grafen
and Hails 2002). We used ‘day’ of the trial
(i.e. days from the initial conditioning
event) and ‘conditioning treatment’ (i.e.
control vs. alarm vs. disturbed) as cate-
gorical between variables. We included
the interactions between variables in the
model to test for the effects of the diffe-
rent treatments depending on the day of
the trial. Subsequent post hoc multiple
comparisons were made using Tukey’s
pairwise comparisons (Sokal and Rohlf
1995).
results
Experiment 1: responses of tadpoles to
alarm and disturbance cues
There were significant differences
among treatments in changes in average
81
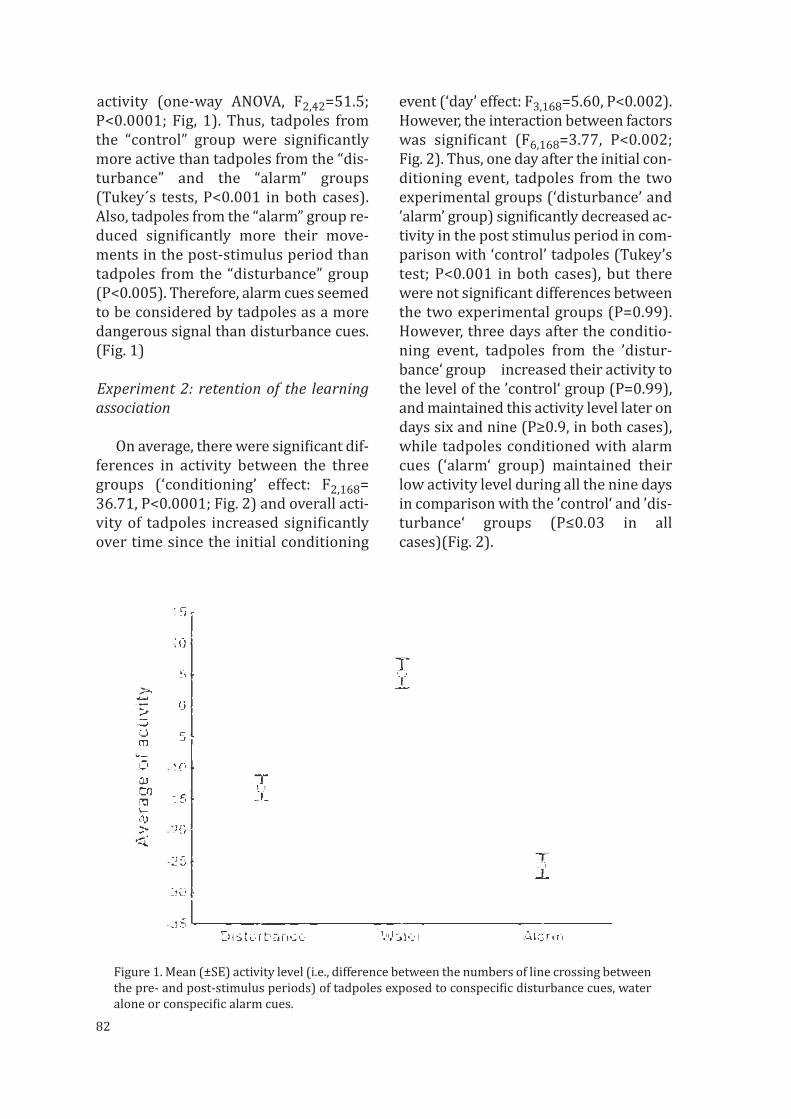
activity (one-way ANOVA, F2,42=51.5;
P<0.0001; Fig, 1). Thus, tadpoles from
the “control” group were significantly
more active than tadpoles from the “dis-
turbance” and the “alarm” groups
(Tukey´s tests, P<0.001 in both cases).
Also, tadpoles from the “alarm” group re-
duced significantly more their move-
ments in the post-stimulus period than
tadpoles from the “disturbance” group
(P<0.005). Therefore, alarm cues seemed
to be considered by tadpoles as a more
dangerous signal than disturbance cues.
(Fig. 1)
Experiment 2: retention of the learning
association
On average, there were significant dif-
ferences in activity between the three
groups (‘conditioning’ effect: F2,168=
36.71, P<0.0001; Fig. 2) and overall acti-
vity of tadpoles increased significantly
over time since the initial conditioning
event (‘day’ effect: F3,168=5.60, P<0.002).
However, the interaction between factors
was significant (F6,168=3.77, P<0.002;
Fig. 2). Thus, one day after the initial con-
ditioning event, tadpoles from the two
experimental groups (‘disturbance’ and
’alarm’ group) significantly decreased ac-
tivity in the post stimulus period in com-
parison with ‘control’ tadpoles (Tukey’s
test; P<0.001 in both cases), but there
were not significant differences between
the two experimental groups (P=0.99).
However, three days after the conditio-
ning event, tadpoles from the ’distur-
bance‘ group increased their activity to
the level of the ’control‘ group (P=0.99),
and maintained this activity level later on
days six and nine (P≥0.9, in both cases),
while tadpoles conditioned with alarm
cues (‘alarm‘ group) maintained their
low activity level during all the nine days
in comparison with the ’control‘ and ’dis-
turbance‘ groups (P≤0.03 in all
cases)(Fig. 2).
Figure 1. Mean (±SE) activity level (i.e., difference between the numbers of line crossing between
the pre- and post-stimulus periods) of tadpoles exposed to conspecific disturbance cues, water
alone or conspecific alarm cues.
82

dIscussIon
The results of the first experiment
show that Iberian green frog tadpoles
display antipredator behaviors (i.e. a
reduction in activity) in response to
chemical cues released from disturbed
conspecifics. This is a typical antipreda-
tory response reported in Iberian green
frog tadpoles in the presence of alarm
cues (Gonzalo et al. 2007). But the results
also showed that reduction of activity
levels is stronger in response to alarm
cues than to disturbance cues. According
to the threat sensitive predator avoi-
dance hypothesis,hypothesis, prey spe-
cies should behave flexibly towards a
varying degree of predator threat and,
consequently, leave more time for other
activities when the threat is low (Helf-
man 1989). If tadpoles react weakly to
disturbance cues, it is possible that tad-
poles are assessing risk based on the na-
ture of the chemicals cues and adaptively
balancing the costs and benefits of pre-
dator avoidance. Often the intensity of an
animal’s antipredator response reflects
the level of threat posed by the predator
(Helfman 1989; Chivers et al. 2001).
When it comes to chemical cues, the type
of cue that an animal detects may be used
to mediate the intensity of the antipreda-
tor response. The threat-sensitive preda-
tor avoidance hypothesis predicts that as
the threat of chemical cue decreases, the
intensity of the antipredator response
will likewise decrease (Helfman 1989).
Disturbance cues are thought to be low
level indicators of risk whereas alarm
cues are high level indicators of risk
(Chivers and Smith 1998), so is logical
that, if tadpoles can asses acutely the risk
level, they react differently to predators
labeled with different types of chemical
cues, as we have found in our experi-
ment. Also, another explanation might be
that the presence of disturbance cues
figure 2. Mean (±SE) activity level (i.e., difference between the numbers of line crossing between
the pre- and post-stimulus periods) of experimental and control tadpoles when exposed to
non-predatory fish alone, several days after the initial conditioning with fish cues mixed with
conspecific alarm cues, or fish cues mixed with conspecific disturbance cues.
83

could put the tadpoles in a state of heigh-
tened vigilance, which could result in a
decrease of activity similar to the anti-
predatory typical response (Ferrari et al
2008). Different concentration of both
types of cues could be an alternative ex-
planation for the differences in activity
reduction, but in natural habitats, the
concentration of alarm cues when a tad-
pole is trapped by a predator should be
higher than the concentration of distur-
bance cues released by a tadpole when is
stressed by a predator. Therefore, diffe-
rences in reduction of activity are more
likely a consequence of the different
nature of the two cues rather than a
consequence of the different cues’
concentration.
The ability to learn and memorize
potential predators may be especially
important for the survivorship of prey
species that are likely to find a high va-
riety of predators while they are in the
aquatic phase (Gonzalo et al. 2007). The
results of the second experiment showed
that tadpoles remember the fish chemi-
cal cues for nine days if they had been
conditioning with alarm cues. Tadpoles
also reacted to the fish chemicals if they
had been conditioning with disturbance
cues, which implies that tadpoles are
able to learn and remember al least for
one day the potential predator labeled
with the disturbance cues. However, tad-
poles had apparently forgotten quickly
the disturbance learned association, be-
cause in the next days they behaved as
control tadpoles, and did not reduce their
activity.
In natural conditions, tadpoles conti-
nually receive information about their
environment and must filter this infor-
mation to focus on those aspects most
important to survival (Dukas 2002). So,
tadpoles have to face up with a wide
range of mixed chemicals signals, (e.g.,
alarm cues, disturbance cues, predator
chemical cues, nonpredator cues). Howe-
ver, prey animals often do not have com-
plete information about their environ-
ment, and can make less accurate
estimations of predation risk. This could
lead them to either over- or underesti-
mate risk (van der Veen 2002). In our
experiments we gave tadpoles incom-
plete or unreliable information regarding
a new predator identity, allowing them
only to “smell” a new potential predator
with different levels of threat. As we see,
to elicit an antipredator response to pre-
dator chemicals cues during the first day
after the conditioning event, even overes-
timating the potential risk level, could be
adaptative because tadpoles had passed
recently through a conditioning event of
biological importance (learning a new
potential predator). But the differences
of signaling risk level between the two
types of cues (disturbance vs. alarm)
were clear when we tested the tadpoles’
memory to the learned association. As
tadpoles did not have other additional in-
formation on the actual risk of this parti-
cular predator, the selection of an
antipredator behavior should be infor-
med by recent experiences (Turner et al.
2005; Ferrari and Chivers 2006). Thus, a
predator labeled with alarm cues is con-
sidered more dangerous than a predator
labeled with disturbance cues, and, the-
refore, more prone to be remembered
and to unleash antipredator behaviors in
the tadpoles over the time. In contrast to
our results, juvenile trouts are not able to
learn chemicals from new predators
from disturbance cues (Ferrari et al.
2008). This difference could be explained
by the differences in the time of exposure
to the predator chemicals before the
conditioning event. We found activity
reduction in tadpoles 24 h after the con-
ditioning, but responses had dissapeared
after 72 h, However, Ferrari et al. (2008)
did not test fishes until 48 h after their
conditioning. Thus, if prey are able to
retain the association between predator
84

cues and disturbed cues only for a few
hours, activity reduction of fishes might
be undetected after 48 h. Also, it is likely
that the learning capacities of tadpoles
differ from those of fishes.
Although the disturbance cues have
not been well studied as the alarm cues,
their involvement in tadpoles decision
making over time is evident. Mirza and
Chivers (2002) demonstrated that juve-
nile brook charr enhance survival during
encounters with predators, if brock charr
were previously conditioned with distur-
bance cues. Ferrari et al. (2008) argued
that this result could be due to the effect
of disturbance cues in the increase of
vigilance. But, as the brook charr were
tested few hours after the conditioning
they coud retain the association between
the two cues. Similarly, our data show
that tadpoles are able to learn new pre-
dators, only for few hours, thanks to dis-
turbance cues and to react to the
chemical cues of those predators the next
day after the conditioning. However, tad-
poles do not retain the association as
long as they do when they have learned
predators by an association with alarm
cues.
In a previous experiment, Iberian
Green frog tadpoles behaved flexibly
towards a varying degree of predator
threat, such that some snakes normally
perceived as a low dangerous predator
were perceived as a very dangerous one
if tadpoles had been previously exposed
to snake chemical cues mixed with alarm
cues (Gonzalo et al. 2007). Our current
data support the idea that Iberian Green
frog tadpoles behaved according to
the threat sensitive predator avoidance
hypothesis when they faced up a preda-
tor labeled with disturbance cues or
alarm cues. Alarm cues alone were per-
ceived as a sign of high risk whereas
disturbance cues elicited a weaker reac-
tion. Also, our results indicated that
Iberian Green frog tadpoles are able to
modify their antipredatory behavior ac-
cording to their previous experience
with chemical cues of the predator.
Thereby, fish were perceived to be very
dangerous predators over the time if the
tadpoles had been previously exposed to
fish cues mixed with alarm cues (i.e.
chemical cues from a predator that was
eating a conspecific) but the same fish
species was perceived as a dangerous
predator only for one day if tadpoles had
been exposed to fish cues mixed with
disturbance cues (i.e., chemical cues from
a potential predator that frightened
some conspecifics). Our goal here is
to show that there is a clear large diffe-
rence in long time effects of two types of
chemical cues with different degree of
threat, one of them (alarm cues) used to
memorize predators and other (distur-
bed cues) used to be awareness at first,
but not to memorize a predator.
references
Appleton RD, Palmer AR (1988) Water-borne sti-
muli released by predatory crabs and
damaged prey induce more predator-re-
sistant shells in a marine gastropod. Proc
Natl Acad Sci USA. 85:4387–4391
ASIH (2004) Guidelines for use of live Amphi-
bians and Reptiles in field and laboratory
research. Herpetological Animal Care and
Use Committee (HACC) of the American
Society of Ichthyologists and Herpetolo-
gists, Lawrence, Kansas
Bryer PJ, Mirza RS, Chivers DP (2001) Chemosen-
sory assessment of predation risk by slimy
sculpins (Cottus cognatus): responses to
alarm, disturbance, and predator cues.
J Chem Ecol 27:533–546
Chivers DP, Smith RJF (1994) Fathead minnows,
Pimephales promelas, acquire predator re-
cognition when alarm substance is asso-
ciated with the sight of unfamiliar fish.
Anim Behav 48:597–605
Chivers DP, Smith RJF (1998) Chemical alarm sig-
naling in aquatic predator/prey interac-
tions: a review and prospectus. Ecoscience
5:338–352
85

Chivers DP, Mirza RS, Bryer PJ, Kiesecker JM
(2001) Threat-sensitive predator avoi-
dance by slimy sculpins: understanding
the role of visual versus chemical informa-
tion. Can J Zool 79:867–873
Dukas R (2002) Behavioural and ecological con-
sequences of limited attention. Phil Trans
R Soc Lond B 357:1539–547
Ferrari MCO, Chivers DP (2006) Learning threat-
sensitive predator avoidance: how do fa-
thead minnows incorporate conflicting
information? Anim Behav 71:19–26
Ferrari MCO, Vavrek MA, Elvidge CK, Fridman B,
Chivers DP, Brown GE (2008) Sensory
complementation and the acquisition of
predator recognition by salmonid fishes.
Behav Ecol Sociobiol 63:113–121
Gonzalo A, López P, Martín J (2007) Iberian green
frog tadpoles may learn to recognize novel
predators from chemical alarm cues of
conspecifics. Anim Behav 74:447–453
Gonzalo A, López P, Martín J (in press) Learning,
memorizing and apparent forgetting of
chemical cues from new predators by
Iberian green frog tadpoles. Anim Cogn
Gosner KL (1960) A simplified table for staging
anuran embryos and larvae with notes on
identification. Herpetologica 16:183–190
Grafen A, Hails R (2002) Modern statistics for the
life sciences. Oxford University Press,
Oxford
Hazlett BA (1985) Disturbance pheromones in
the crayfish Orconectes virilis. J Chem Ecol
11:1695–1711
Hazlett BA (1989) Additional sources of distur-
bance pheromone affecting the crayfish Or-
conectes virilis. J Chem Ecol 15:381–385
Hazlett BA (1990a) Disturbance pheromone in
the hermit crab Calcinus laevimanus (Ran-
dolf, 1840). Crustaceana 58:314–316
Hazlett BA (1990b) Source and nature of distur-
bance chemical system in crayfish. J Chem
Ecol 20:2263–2275
Hazlett BA (1994) Alarm responses in the cray-
fish, Orconectes virilis and Orconectes pro-
pinquus. J Chem Ecol 20:1525–1535
Hazlett BA (2003) Predator recognition and lear-
ned irrelevance in the crayfish Orconectes
virilis. Ethology 109:765-780
Helfman GS (1989) Threat-sensitive predator
avoidance in damselfish–trumpetfish inter-
actions. Behav Ecol Sociobiol 24: 47–58
Hews DK (1988) Alarm response in larval wes-
tern toads, Bufo boreas: release of larval
chemicals by a natural predator and its ef-
fect on predator capture efficiency. Anim
Behav 36:125–133
Hews DK, Blaustein AR (1985) An investigation
of the alarm response in Bufo boreas and
Rana cascadae tadpoles. Behav Neural Biol
43:47–57
Howe NR, Sheikh TM (1975) Anthopleurine: A
sea anemone alarm pheromone. Science
189:386–388
Jordão LC (2004) Disturbance chemical cues de-
termine changes in spatial occupation by
the convict cichlid Archocentrus nigrofas-
ciatus Behav Process 67:453–459
Jordão LC, Volpato GL (2000) Chemical transfer
of warning information in non-injured fish.
Behaviour 137:681–690
Kats LB, Dill LM (1998) The scent of death: che-
mosensory assessment of predation risk by
prey animals. Ecoscience 5:361–394
Kiesecker JM, Chivers DP., Marco A, Quilchano C,
Anderson MT, Blaustein AR (1999) Identi-
fication of a disturbance signal in larval
red-legged frogs, Rana aurora. Anim Behav
57:1295–1300
Lawrence JM (1991) A chemical alarm response
in Pycnopodia helianthoides (Echinoder-
mata: Asteroidea). Marin Behav Physiol
19:39–44
Mathis A, Smith RJF (1993) Fathead minnows, Pi-
mephales promelas, learn to recognize nor-
thern pike, Esox lucius, as predators on the
basis of chemical stimuli from minnows in
the pike’s diet. Anim Behav 46:645–656
Mirza RS, Chivers DP (2002) Behavioural respon-
ses to conspecific disturbance chemicals
enhance survival of juvenile brook charr,
Salvelinus fontinalis, during encounters
with predators. Behaviour 139:1099–1110
Pfeiffer W (1966) Die verbreitung der schreckre-
aktion bei kaulquappen und die herkunft
des schreckstoffes. Z Vergl Physiol 52:79–
98
Rohr JR, Madison DM (2001) A chemically media-
ted trade-off between predation risk and
mate search in newts. Anim Behav 62:863–
869
Sih A (1986) Antipredator responses and the
perception of danger by mosquito larvae.
Ecology 67:434–441
Smith RJF (1979) Alarm reaction of Iowa and
johnny darters (Etheostoma, Percidae, Pis-
ces) to chemicals from injured conspecifics.
Can J Zool 57:1278–1282
Smith RJF (1992) Alarm signals in fishes. Rev Fish
Biol Fish 2:33–63
Sokal RR, Rohlf FJ (1995) Biometry. WH Freeman,
New York
86

Turner AM, Turner SE, Lappi HM (2005) Learning,
memory and predator avoidance by fres-
hwater snails: effects of experience on pre-
dator recognition and defensive strategy.
Anim Behav 72:1443-1450
van der Veen IT (2002) Seeing is believing: infor-
mation about predators influences yello-
whammer behavior. Behav Ecol Sociobiol
51:466–471
Wilson DJ, Lefort H (1993) The effect of predator
diet on the alarm response of red-legged
frog, Rana aurora, tadpoles. Anim Behav
46:1017–1019
Wisenden BD (2000) Olfactory assessment of pre-
dation risk in the aquatic environment. Phil
Trans R Soc Lond B 355:1205–1208
Wisenden BD, Chivers DP, Smith RJF (1995) Early
warning of risk in the predation sequence: A
disturbance pheromone in Iowa darters
(Etheostoma exile). J Chem Ecol 21:1469–
1480
Woody DR, Mathis A (1998) Acquired recognition
of chemical stimuli from an unfamilia
predator: Associative learning by adult
newts, Notophthalmus viridescens. Copeia
1998:1027–1031
87


cAPÍTuLO V
LAS SUSTANCIAS qUíMICAS DE DEPREDADORES,PERO NO LAS DE INDIVIDUOS COESPECíFICOS
INFLUENCIAN LA SELECIÓN DE CHARCAS EN
INDIVIDUOS METAMÓRFICOS DE RANA VERDE
PREDATOR, BUT NOT CONSPECIFIC, CHEMICALCUES INFLUENCE POND SELECTION BYRECENTLY METAMORPHOSED GREEN FROGS


En los anfibios, los adultos y las larvas tienen requerimientosecológicos diferentes, lo que puede forzar a los individuos reciénmetamorfoseados a dispersarse. La presencia de señales químicas deindividuos coespecíficos y depredadores puede proporcionarles informa-ción sobre las cualidades del hábitat, lo que puede influir en las decisionesde asentamiento de los juveniles. En este trabajo hemos examinado en ellaboratorio si la selección de una charca por individuos recién metamor-foseados de rana verde ibérica (Pelophylax perezi) está influida por lapresencia, en el agua, de señales químicas de individuos coespecíficos y/ode depredadores. Los resultados sugieren que las ranas verdes Ibéricasson capaces de detectar la presencia de señales químicas de serpientesdepredadoras en el agua, y así evitar esas charcas. Sin embargo, las ranasno mostraron ni atracción ni rechazo a charcas con sustancias químicasde individuos coespecíficos. Así, los juveniles recién metamorfoseados,deben seleccionar sus territorios post-metamórficos basándose en la faltade riesgo de depredación, y posiblemente en características del hábitat,pero no en la presencia de otros coespecíficos.
Este cápitulo se corresponde con: Gonzalo A, Cabido C, Galán P, López P, Martín J (2006) Predator, butnot conspecific, chemical cues influence pond selection by recently metamorphosed Iberian green frogs,Rana perezi. Can J Zool 84:1295-1299


Predator, but not consPecIfIc, chemIcal
cues Influence Pond selectIon by recently
metamorPhosed IberIan green frogs
In amphibians, adults and larvae have different ecological requirements, which could
force recently metamorphosed individuals to disperse. The presence of chemical cues of
conspecifics and predators could provide information about the habitat quality, which
might influence the juveniles’ settlement decisions. We examined in the laboratory
whether pond choice by recently metamorphosed Iberian green frogs (Pelophylax perezi)
is influenced by the presence of chemical cues from conspecifics and/or from predators
in the water. Our results suggest that frogs were able to detect the presence of chemical
cues of snake predators in the water, and that avoided entering such ponds. However,
frogs did not show either attraction or avoidance to ponds with conspecifics chemical
cues. Thus, juvenile frogs may select their post-metamorphic territories relying on the
lack of predation risk, and possibly on some habitat features, but not on the presence
of conspecifics.
IntroductIon
In amphibians, changes in life style
are especially obvious in the transition
from aquatic larvae to terrestrial postme-
tamorphic and adult stages (Duellman
and Trueb 1986). Adults and larvae often
have different ecological requirements,
which could force recently metamorpho-
sed individuals to disperse when they
complete their growth as larvae. During
the transient phase of dispersal is very
important an appropriate selection of the
future home range (Clobert et al. 2001).
In amphibians, the distance of dispersion
is generally small and juveniles may
occupy home ranges in vacant spots
among adults or in peripheral locations
(Zug et al. 2001). Thus, recently meta-
morphosed juveniles may need to search
for peripheral potential home ranges,
where is important to evaluate habitat
quality and predation risk.
In selecting a good home range where
to establish, a dispersing individual have
to considerate several factors like
physicalones (e.g., temperature, humi-
dity), and also the presence of conspeci-
fics or predators (Stamps and Tanaka
1981; Clobert et al. 2001). Some amphi-
bians may detect conspecific chemical
cues (e.g., Jaeger et al. 1986; Aragon et al.
2000; Waldman and Bishop 2004). Cons-
pecific cues could provide information
about habitat quality, because they may
reflect the presence of food, appropiate
environmental conditions, and low pre-
dation risk (Woody and Mathis 1997). In-
dividuals may, then, settle in clusters
because they are attracted to conspeci-
fics, rather than habitat features (Stamps
1988; Graves et al. 1993; Muller et al.
1997; Quinn and Graves 1999). Never-
theless, the presence of conspecifics
might not be desirable when it may in-
crease competition for food or potential
mates (Griffiths et al. 1991).
Additionally, the detection of preda-
tors is important. Prey often respond to
the threat of predation by decreasing the
93

frequency or efficiency of behaviour
associated with other activities (Lima
and Dill 1990). Minimizing the negative
effects of such trade-offs requires prey to
discriminate threat levels and to adjust
their behaviour accordingly. Animals that
fail to respond appropriately to predator
stimuli have a decreased probability of
survival. In aquatic vertebrates, both vi-
sual (Karplus and Algom 1981) and che-
mical cues (Chivers and Smith 1993) may
be important for predator recognition.
However, chemical detection of preda-
tors is clearly advantageous to prey when
the use of other sensory mechanisms is
difficulted by environmental characteris-
tics, such as turbid water or abundant ve-
getation (Kats and Dill 1998). Whereas
many frog tadpoles deetct predators via
chemical cues, from predators or conspe-
cific alarm cues (Kats and Dill 1998),
adult frogs mainly use vision and hearing
to detect predators, which could be ad-
vantageous when a terrestrial predator
approaches. However, many frogs escape
by jumping to the water (Williams et al.
2000). Thus, tadpoles’ ability to detect
chemicals of predators could be advan-
tageous, especially when assessing a po-
tential habitat. Many studies have tested
the mechanisms of predator chemical de-
tection in amphibian larvae (e.g. Pe-
tranka and Hayes 1998; Petranka et al.
1987; Manteifel 1995; Griffiths at al.
1998; see review in Kats and Dill 1998).
However, only a few studies have exami-
ned the prevalence of this mechanism in
metamorphosed frogs or toads (Heinen
1994; Flowers and Graves 1997; Schley
and Griffiths 1998; Belden et al. 2000;
Chivers et al. 2001).
In this paper, we examined in the
laboratory whether pond choice by
recently metamorphosed Iberian green
frogs (P. perezi) is influenced by the pre-
sence of chemical cues from conspecifics
and predators in the water. We hypothe-
sized that 1) if frogs would be attracted
to territories near conspecifics, they
should select ponds containing conspeci-
fic cues, and 2) frogs should avoid ponds
containing predator cues.
methodsstudy animals
During August 2003, we collected 20
recently metamorphosed green frogs
(SVL, Mean±SE=20.7±0.2 mm) at several
small ponds in Collado Mediano (Madrid,
central Spain). In this habitat, frogs had
the opportunity to choose among small
nearby ponds, that contained abundant
aquatic vegetation. Frogs were maintai-
ned under two experimental conditions
at “El Ventorrillo” field station (5 km
from the capture site). Twelve frogs
were housed individually in aquaria
(18x25x10 cm), with water at ambient
temperature and under a natural photo-
period. The other eight frogs were kept
in groups in two aquaria (20x20x15 cm)
to be used as donors of conspecifics
scent. We fed frogs with tenebrio larvae
(Tenebrio molitor) twice a week.
We also captured in nearby larger
ponds two viperine snakes, Natrix
maura, to be used as predator scent do-
nors. This snake is predominantly aqua-
tic and feeds mainly on insects,
amphibians, both larvae and adults, and
fishes (Haley and Davies 1986; Braña
1998). Snakes were housed individually
in cages (36x25x13 cm) containing saw-
dust and tree barks for cover and a pond
with water. The snake’s cages were pla-
ced in rooms separate from the frogs to
avoid contact with the scent and visual
stimuli by frogs. We kept the snakes two
weeks in captivity, where they were fed
small pieces of commercial freshwater
fishes obtained from a fish-market. We
did not feed snakes with frogs to avoid
the potential effect of frogs responding to
alarm cues of conspecifics rather than to
snake’s chemical cues alone (Chivers and
94

Mirza 2001). All animals seemed healthy
during the trials, and we returned them
to their exact capture sites at the end of
trials.
experimental design
We conducted the experiments in
glass terraria (50x40x50 cm). Two plas-
tic rectangular trays (18x40x2 cm) filled
with water were placed at the opposite
ends of the terrarium. Between the trays
we placed a plate (14x40 cm) of Pores-
pan. Thus, we simulated two ponds sepa-
rated by a central dry area.
We planned a repeated measures
design in which each frog was tested in
each of three treatments in a randomized
sequence, but participated in only one
test per day. In the ‘control’ treatment,
the two ponds were filled with clean
water (coming from a nearby clean
mountain stream that did not house
frogs nor snakes) In the ‘conspecifics’
treatment, we used clean water in one
pond and water with conspecific frog
chemical stimuli in the other pond, to
test for attraction or avoidance to cues of
conspecifics. In the ‘predator snake’ tre-
atment, we used clean water in one pond
and water containing viperine snake che-
mical stimuli in the other pond, to test for
possible recognition and avoidance of
water used by potential predators. We
randomly altered the spatial position of
the ponds in each trial. We took the water
with conspecific’s chemical stimuli from
the aquaria where groups of frogs had
been maintained for at least three days.
The water with snake’s chemical stimuli
was prepared by placing the snakes in
aquaria with clean water for 48 h. On the
day of testing, we removed water from
each tank to fill the ponds (200 ml each)
in the respective choice tests, 15 min
prior to the start of each test.
Tests were made between 11:00 and
13:00 h when frogs were fully active. We
gently placed a single juvenile frog cove-
red with a small glass cage in the centre
of experimental aquarium, and waited 5
min for habituation. Each test began
when we lifted the cage, releasing the
frog. We subsequently stood as motion-
less as possible recording frogs’ behavior
from a hidden point 4 m away from the
aquaria. Preliminary trials showed that
frog behavior did not differ bewteen si-
tuations when there was a motionless
observer and trials filmed with video, wi-
thout any observer.in the room. Typically,
frogs stayed a few minutes in the middle
of the aquarium, and then jumped to the
water of one pond, and occasionally
changed from one pond to the other.
We noted the location of the frog each
5 min, the first pond election, and the
number of times that the frog changed
between ponds. If a frog was located in
any of the two ponds, it was designated
as having chosen that pond, whereas if it
was located out of the water, it was desig-
nated as having made no choice. We also
recorded the ‘total time’ that the frog
spent in each pond, and calculated the
‘relative time’ spent in each pond as the
time spent in a particular pond/total
time spent in any pond. Each trial lasted
2 h, after which the aquarium, the trays
and the Porespan were carefully washed
with unodorus detergent and drained to
avoid odour contamination.
data analyses
We determined the frog’s preferred
pond by calculating in which pond the
frog spent greater than 50 % of its time
(excluding time spent in the no-choice
area). To assess whether frogs chose one
of the ponds, we calculated the number
of frogs that spent greater than 50 % of
their time in a particular stimulus pond
and compared it to an expected binomial
distribution assuming frequencies to be
equiprobable on each pond (for a similar
95

procedure, see Chivers et al. 1997; Ara-
gón et al. 2000). To compare the ‘total
time’ and ‘relative time’ spent in the pond
with chemical cues, and the number of
the number of changes between ponds
across treatments, we used nonparame-
tric Friedman’s two-way ANOVA. To in-
clude the control treatment in these
analyses we used the time spent in one
of the two ponds with clean water selec-
ted at random. Pairwise comparisons of
means were conducted using nonpara-
metric multiple comparison procedures
described in Sokal and Rohlf (1995).
results
In their first choice, frogs did not
significantly select any pond in any of the
treatments; based on the null hypothesis
that the likelihood of entering one pond
was equal for any of the ponds (clean
water or water with chemical cues), the
probability that one type of pond was
first chosen was not significantly diffe-
rent from the expected in any of the three
treatments (two-tailed binomial tests,
clean/chemicals, control: 5/7, P=0.77;
conspecifics: 4/8, P=0.39; snake: 7/5,
P=0.77).
Total time spent in the pond with
water with chemical cues differed signi-
ficantly between treatments (Friedman’s
ANOVA, χ22,12=9.30, P<0.01) (Table 1).
Thus, frogs spent significantly less time
in the pond with chemical cues in the
snake treatment than in the other treat-
ments (non-parametric multiple compa-
risons, P<0.02, in both cases), but there
were non significant differences between
the conspecifics and the control treat-
ments (P=0.18). Similarly, relative time
spent in the pond with water with
chemical cues differed significantly
between treatments (Friedman’s ANOVA,
χ22,12=13.31, P<0.002) (Table 1). Thus,
frogs spent significantly relatively less
time in the pond with scent in the snake
treatment than in the other treatments
(non-parametric multiple comparisons,
P<0.02 in both cases), but there were
non significant differences between the
conspecifics and the control treatment
(P=0.48). The number of changes bet-
ween ponds did not differ significantly
among treatments (Friedman’s ANOVA,
χ22,12=3.80, P=0.15) (Table 1).
When we estimated the pond selec-
ted by frogs from the proportion of time
spent in each pond, frogs in the control
and conspecifics treatments did not sig-
nificantly select any pond (Table 1). Ho-
wever, in the snake treatment, frogs
clearly avoided ponds containing water
with viperine snake chemical cues. Simi-
lar results arose when we considered the
relative time spent in each pond to assess
the pond chosen (Table 1).
dIscussIon
Our results suggest that recently
metamorphosed Iberian green frogs
were able to detect the presence of che-
mical cues of snake predators in the
water, and that they left the pond as soon
as they detected predator cues. Visual
cues are important in eliciting an anti-
predator response in anurans (e.g. Wi-
lliams et al. 2000), but the use of more
than one sensory stimulus could be ad-
vantageous in habitat selection. The de-
tection of chemical cues may inform
about the continuous presence of a pre-
dator in a certain area even if it is hidden
(Kats and Dill 1998), whereas visual
cues, only inform about the current pre-
sence of the predator. Thus, when frogs
select a pond, the combined use of visual
and chemical detection could be more
advantageous than the use of visual cues
alone. Although visual detection could be
useful to avoid active foraging predators,
chemical cues are more useful in detec-
ting ambush predators, especially in
complex habitats where visual detection96
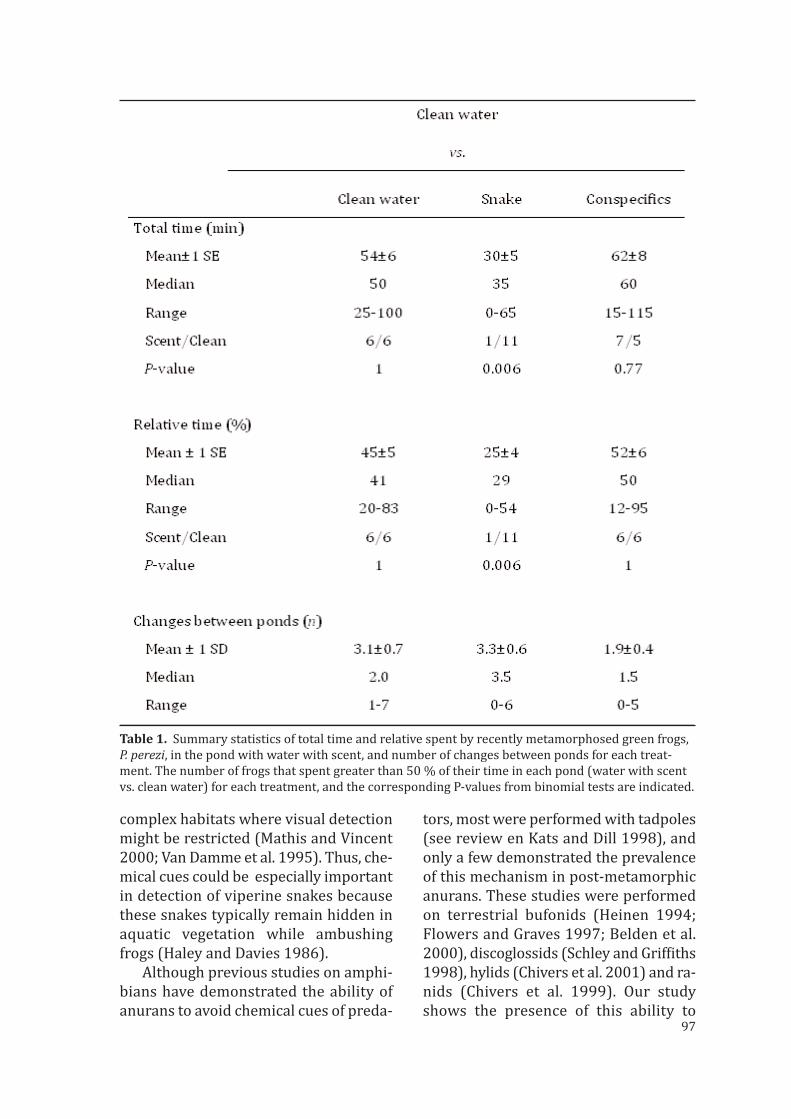
complex habitats where visual detection
might be restricted (Mathis and Vincent
2000; Van Damme et al. 1995). Thus, che-
mical cues could be especially important
in detection of viperine snakes because
these snakes typically remain hidden in
aquatic vegetation while ambushing
frogs (Haley and Davies 1986).
Although previous studies on amphi-
bians have demonstrated the ability of
anurans to avoid chemical cues of preda-
tors, most were performed with tadpoles
(see review en Kats and Dill 1998), and
only a few demonstrated the prevalence
of this mechanism in post-metamorphic
anurans. These studies were performed
on terrestrial bufonids (Heinen 1994;
Flowers and Graves 1997; Belden et al.
2000), discoglossids (Schley and Griffiths
1998), hylids (Chivers et al. 2001) and ra-
nids (Chivers et al. 1999). Our study
shows the presence of this ability to
table 1. Summary statistics of total time and relative spent by recently metamorphosed green frogs,
P. perezi, in the pond with water with scent, and number of changes between ponds for each treat-
ment. The number of frogs that spent greater than 50 % of their time in each pond (water with scent
vs. clean water) for each treatment, and the corresponding P-values from binomial tests are indicated.
97

detect snake chemicals in recently meta-
morphosed juveniles of a ranid, and sug-
gests its importance in habitat selection.
This mechanism may be presented in
other anuran groups because anurans
have well developed vomeronasal organs
(Scalia 1976).
Metamorph Iberian green frogs are
often forced to disperse from breeding
ponds, which obviously provide suitable
habitat for the adults, to small nearby
swallow temporal ponds (Lizana et al.
1989), because adults compete for the
better places (Schneider and Steinwarz
1990), with larger adults being found in
central positions in the pond. Thus, there
is some degree of spatial segregation
between adults and juveniles (Lizana et
al. 1989). Moreover, adult frogs eat any
prey smaller than themselves, including
metamorphs (personal observation). Ju-
veniles can either select territories with
similar body size conspecific presence
(Stamps 1988; Graves et al. 1993), or
avoid them (Léna and Fraipont 1998).
The detection of conspecific chemicals
could be advantageous in selecting a ha-
bitat either because they provide infor-
mation about the habitat quality (Woody
and Mathis 1997), or because they reflect
a high potential local competition for
food (Griffiths et al. 1991). The detection
of conspecifics chemical cues has been
found common in many newts and sala-
manders (e.g. Jaeger et al. 1986; Aragon
et al. 2000) but more rarely in anurans
(Waldman and Bishop 2004). Our results
suggest that recently metamorphosed
Iberian green frogs do not show attrac-
tion or avoidance to ponds with conspe-
cifics chemicals. Thus, Iberian green
frogs do not seem able to detect chemical
cues of conspecifics. However, it remains
possible that the frogs would detect
conspecifics and be attracted to low den-
sities of conspecifics, but not to high den-
sities of conspecifics (as might indicate
water with concentrated chemical cues
from the small aquaria containing the
four conspecifics used as donor in our ex-
periment), because competition may be
too high. If this was the case, then it is
possible that the lack of response to
conspecific cues might be a density issue.
Alternatively, the presence of conspeci-
fics might not be important when selec-
ting a pond. Food could be available to all
individuals in all ponds and not be a res-
tricted factor for juveniles, or not be as
important as other factors. Adult male
Iberian green frogs defend small territo-
ries against other males, but only during
the mating season (Schneider and Stein-
warz 1990). Also, because this frog oc-
curs in relatively dense populations, the
localization of ponds with low popula-
tion densities of conspecifics might be
difficult.
We conclude that post-metamorphic
Iberian green frogs seemed to select
ponds relying on the potential threat of
predation by water snakes, whereas
conspecific cues did not seem to in-
fluence their decisions.
references
Aragón P, López P, Martín J (2000) Conspecific
chemical cues influence pond selection
by male newts Triturus boscai. Copeia
2000:874–878
Belden LK, Wildy EL, Hatch AC, Blaustein AR
(2000) Juvenile western toads, Bufo boreas,
avoid chemical cues of snakes fed juvenile,
but not larval, conspecifis. Anim Behav
59:871–875
Braña F (1998) Natrix maura (Linnaeus, 1758).
In Fauna Ibérica, vol. 10. Edited by M.A.
Ramos. Museo Nacional de Ciencias Natu-
rales, CSIC. Madrid. pp. 440–454
Chivers DP, Smith RJF (1993) The role of olfaction
in chemosensory-based predator recogni-
tion in the fathead minnow, Pimephales
promelas. J Chem Ecol 19:623–633
Chivers DP, Mirza RS (2001) Predator diet cues
and the assessment of predation risk by
aquatic vertebrates: a review and prospec-
tus. In Chemical signals in vertebrates, vol.
9. Edited by D.A. Marchlewska-Koj, J.J. Lepri
and D. Müller-Schwarze. Plenum, New
York. pp 277–28498

Chivers DP, Wildy EL, Blaustein AR (1997) Easter
long-toed salamanders (Ambystoma ma-
crodactylum columbianum) larvae recog-
nize cannibalistic conspecifics. Ethology
103:187–197
Chivers DP, Kiesecker JM, Wildy EL, Belden LK,
Kats L, Blaustein AR (1999) Avoidance
response of post-metamorphic anurans to
cues of injured conspecifics and predators.
J Herpetol 33:472–476
Chivers DP, Wildy EL, Kiesecker JM, Blaustein AR
(2001) Avoidance response of juvenile pa-
cific tree frogs to chemical cues of introdu-
ced predatory bullfrogs. J Chem Ecol
27:1667–1676
Clobert J, Danchin E, Nichols JD, Dhondt AA
(2001) Dispersal. Oxford University Press,
Oxford
Duellman WE, Trueb L (1986) Biology of Amphi-
bians. McGraw-Hill, New York
Flowers MA, Graves BM (1997) Juvenile toads
avoid chemical cues from snake predators.
Anim Behav 53:641–646
Graves BM, Summers CH, Olmstead KL (1993)
Sensory mediation of aggregation among
postmetamorphic Bufo cognatus. J Herpe-
tol 27:315–319
Griffiths RA, Edgar PW, Wong ALC (1991) Site-
specific competition in tadpoles: growth
inhibition and growth retrieval in natter-
jack toads, Bufo calamita. J Anim Ecol
60:1065–1076
Griffiths RA, Schley L, Sharp PE, Dennis JL, Roman
A (1998) Behavioural responses of mallor-
can midwife toad tadpoles to natural and
unnatural snake predators. Anim Behav
55:207–214
Haley A, Davies PMC (1986) Diet and foraging be-
haviour of Natrix maura. Herpetol J 1:53–
61
Heinen JT (1994) Antipredator behaviour of
newly metamorphosed american toads
(Bufo a. americanus), and mechanisms
of hunting by eastern garter snakes
(Thamnophis s. sirtalis). Herpetologica
50:137–145
Jaeger RG, Goy JM, Tarver M, Márquez C (1986)
Salamander territoriality: pheromonal
markers as advertisement by males. Anim
Behav 34:860–864
Karplus I, Algom D (1981) Visual cues for preda-
tor face recognition by reef fishes. Z
Tierpsychol 55:343–364
Kats LB, Dill LM (1998) The scent of death: che-
mosensory assessment of predation risk
by prey animals. Ecoscience 5:361–394
Léna JP, Fraipont M (1998) Kin recognition in the
common lizard. Behav Ecol Sociobiol
42:341–347
Lima SL, Dill LM (1990) Behavioral decisions
made under the risk of predation: a re-
view and prospectus. Can J Zool 68:619–
640
Lizana M, Ciudad MJ, Perez-Mellado V (1989) Ac-
tividad, reproducción y uso del espacio en
una comunidad de anfibios. Treballs Soc
Cat Ictiol Herpetol 2:92–127
Manteifel Y (1995) Chemically-mediated avoi-
dance of predators by Rana temporaria
tadpoles. J Herpetol 29:461–463
Mathis A, Vincent F (2000) Differential use of vi-
sual and chemical cues in predator
recognition and threat-sensitive predator-
avoidance responses by larval newts
(Notophthalmus viridescens). Can J Zool
78:1646–1652
Muller KL, Stamps JA, Krishnan VV, Willits NH
(1997) The effects of conspecific attrac-
tion and habitat quality on habitat selec-
tion in territorial birds (Troglodytes
aedon). Am Nat 150:650–661
Petranka JW, Hayes L (1998) Chemically media-
ted avoidance of a predatory odonate
(Anax junius) by American toad (Bufo
americanus) and wood frog (Rana sylva-
tica) tadpoles. Behav Ecol Sociobiol
42:263–271
Petranka JW, Kats LB, Sih A (1987) Predator prey
interactions among fish and larval amphi-
bians: use of chemical cues to detect pre-
datory fish. Anim Behav 35:420–425
Quinn VS, Graves BM (1999) Space use in res-
ponse to conspecifics by the red-backed
salamander (Plethodon cinereus, Pletho-
dontidae, Caudata). Ethology 105:993–
1002
Scalia F (1976) Olfaction. In Frog neurobiology.
A handbook. Edited by R. Llinas and W.
Prech. Springer-Verlag, Berlin. pp. 213–
233
Schley L, Griffiths RA (1998) Midwife toads (Aly-
tes muletensis) avoid chemical cues from
snakes (Natrix maura). J Herpetol 32:572–
574
Schneider H, Steinwarz D (1990) Mating call and
territorial calls of the Spanish lake frog,
Rana perezi (Ranidae, Amphibia). Zoolo-
gischer-Anzeiger 225:265–277
Sokal RR, Rohlf FJ (1995) Biometry, 3rd. ed. W.H.
Freeman, New York
Stamps JA (1988) Conspecific attraction and ag-
gregation in territorial species. Am Nat
131:329–347
99

Stamps JA, Tanaka S (1981) The relationship bet-
ween food and social behaviour in juvenile
lizards. Copeia 1981:422–434
Van Damme R, Bauwens D, Thoen C, Vanderstig-
helen D, Verheyen, RF (1995) Responses of
naive lizards to predator chemical cues. J
Herpetol 29:38–43
Waldman B, Bishop PJ (2004) Chemical commu-
nication in an archaic anuran amphibian.
Behav Ecol 15:88–93
Williams CR, Brodie Jr. ED, Tyler MJ, Walker SJ
(2000) Antipredator mechanisms of Aus-
tralian frogs. J Herpetol 34:431–443
Woody DR, Mathis A (1997) Avoidance of areas
labelled with chemical stimuli from dama-
ged conspecifics by adult newts, Notoph-
thalmus viridescens, in a natural habitat. J
Herpetol 31:316–318
Zug GR, Vitt LJ, Caldwell JP (2001) Herpetology:
an introductory biology of Amphibians
and Reptiles, 2nd ed. Academic Press, San
100

cOncLuSiOneS


1. Dependiendo de experiencias previas, los renacuajos de rana verde ibéricason capaces de discriminar el nivel de riesgo que posee el depredador y ajus-tar su comportamiento en función de éste, de acuerdo con la hipótesis de larespuesta dependiente del nivel de riesgo de depredación.
2. Los renacuajos de rana verde ibérica aprenden a reconocer una señal químicanueva y no peligrosa como peligrosa al asociarla con sustancias de alarmade individuos coespecíficos. Este mecanismo de aprendizaje puede ser espe-cialmente importante para la supervivencia de especies que se encuentrancon una gran variedad de depredadores durante su fase acuática o con dife-rentes especies depredadoras dependiendo de las estaciones.
3. Los renacuajos de rana verde ibérica son capaces de asociar una sustanciaquímica con un riesgo de depredación y reaccionar ante ella por lo menosdurante nueve días.
4. A partir de los nueve días del aprendizaje se produce una perdida de la res-puesta, lo que sugiere una necesidad de refuerzo para mantener la asocia-ción y la intensidad de la respuesta.
5. La perdida de la respuesta con el tiempo y la aparente perdida de memoriapueden ser una estrategia evolutiva encaminada a evitar comportamientosantidepredatorios costosos en respuesta a sustancias químicas de especiesno peligrosas que accidentalmente aparecieron emparejadas con sustanciade alarma de coespecíficos
6. Los renacuajos de rana verde ibérica presentan los mecanismos de aprendi-zaje de lo irrelevante y de inhibición latente que en determinadas circustan-cias les imposibilita realizar asociaciones entre un incremento del riesgo dedepredación, indicado por la sustancia de alarma de coespecíficos, y sustan-cias químicas de un supuesto depredador.
7. Ambos mecanismos (aprendizaje de lo irrelevante e inhibición latente) sonvías adaptativas para enfrentarse con información conflictiva, y estrategiaspara evitar el aprendizaje de información irrelevante que desencadenen cos-tosas respuestas antidepredatorias ante estímulos de especies no depreda-doras.
8. Los renacuajos de rana verde ibérica, reaccionan ante sustancias de estrésde individuos coespecíficos en menor medida que ante sustancias de alarmade coespecíficos, comportándose de acuerdo con la hipótesis de la respuestadependiente del nivel de riesgo de depredación, así el tipo de sustancia de-tectada es usada para determinar la intensidad de la respuesta antidepre-datoria desencadenada.

9. Así mismo, los renacuajos de rana verde ibérica se comportan de acuerdo ala hipótesis de la respuesta dependiente del nivel de riesgo de depredacióncuando valoran el riesgo de depredación asociado a un depredador depen-diendo si el proceso de aprendizaje ha sido debido a sustancias de alarma oa sustancias de estrés.
10. Los renacuajos son capaces de recordar al menos nueve días a los depreda-dores potenciales marcados con sustancias de alarma, pero sólo durante undía a los marcados con sustancia de estrés. Así, la sustancia de alarma esutilizada para memorizar y recordar depredadores, pero la sustancia deestrés no, al implicar menor riesgo.
11. Los individuos metamórficos de rana verde ibérica son capaces de reconocerlas señales químicas de los depredadores y evaluar la seguridad del hábitatterrestre en base a ellas.
12. En contraste, las señales químicas de individuos coespecíficos no parece serdetectadas o no son utilizadas como método de evaluación de la calidad delhábitat.
13. El reconocimiento del peligro que posee un depredador varía en función deexperiencias anteriores de los renacuajos y se modifica según estas experien-cias. En los renacuajos de rana verde ibérica, el reconocimiento y la memoriade depredadores no es algo fijo, sino que continuamente se actualiza lainformación recibida y actúan en base a ella.
14. En todas sus decisiones antidepredatorias, los renacuajos se comportan deacuerdo con la hipótesis de la respuesta dependiente del nivel de riesgo dedepredación que implica tener un comportamiento flexible en la respuestaantidepredatoria para adaptarse ajustadamente al riesgo de depredaciónexistente en cada momento.
15. La habilidad de aprender a reconocer a un depredador es más compleja quesimplemente la asociación de dos sustancias químicas simultáneas. Existenmecanismos en el aprendizaje para evitar asociaciones incorrectas que pue-dan llevar a los renacuajos a desencadenar respuestas hacia sustanciasquímicas de especies no peligrosas con las que comparten el hábitat.



La presente tesis estudia e intenta clarificar cómo actúan los
mecanismos de aprendizaje (definido como el cambio en el patrón
de comportamiento individual basándose en la experiencia) para
permitir a las presas enfrentarse a cualquier peligro presente en el
ambiente (incluso aquellos en principio novedosos o desconocidos).
En particular, se quiere profundizar en la manera en que la presa
adquiere información sobre la identidad y la peligrosidad de los
depredadores y cómo ajusta continuamente sus respuestas ante los
riesgos existentes y la información recibida del medio.
En los renacuajos de rana verde ibérica han evolucionado
mecanismos de aprendizaje de depredadores mediante sustancias
emitidas por individuos conespecíficos. Sin embargo, la habilidad de
aprender a reconocer a un depredador es más compleja que simple-
mente la asociación de dos sustancias químicas simultáneas. El
reconocimiento del peligro que posee un depredador varía en función
de experiencias anteriores de los renacuajos y se modifica según estas
experiencias. En los renacuajos de rana verde ibérica, el reconoci-
miento y la memoria de depredadores no es algo fijo, si no que conti-
nuamente se actualiza la información recibida y se actúa en base a
ella. Así los renacuajos se comportan de una manera flexible ajustando
sus comportamientos antidepredatorios al riesgo de depredación
existente en cada momento, siempre de acuerdo con la hipótesis de
la respuesta dependiente del nivel de riesgo de depredación.
Departamento de Ecología Evolutiva
Museo Nacional de ciencias NaturalesDepartamento de Bioloxía animal, Bioloxía vexetal e Ecoloxía
Universidade da Coruña