modelo numÉrico de calidad de aguas para la ciÉnaga …
TRANSCRIPT

INTRODUCCIÓN
La Ciénaga Grande de Santa Marta (CGSM)forma parte de uno de los ecosistemas lagunares-es-tuarinos más importante de la cuenca del Caribe,tanto desde el punto de vista ecológico como econó-mico. Se localiza sobre la costa norte de Colombia,entre los 10º44’ a 11º00’ N y 74º19’ a 74º31’ W. Lasuperficie del espejo de agua es de 423 km2 y la pro-fundidad promedio es de 1.6 m (Figura 1). Hacia elnorte se comunica con el Mar Caribe por un canal(Boca de la Barra) que se abre a través de la Isla Sa-lamanca, con un ancho aproximado de 280 m. Ladiferencia del nivel del agua en la laguna, entre lasépocas seca y húmeda (inundación), es de 30 a 50cm, pero puede llegar a más de un metro sobre el ce-ro hidrográfico dependiendo del volumen de aguacontinental que entre al sistema. La marea del Cari-be en la región es una combinación diurna y semi-diurna de amplitud irregular pero débil; fluctúa en-tre 20 y 30 cm y raras veces supera los 50 cm. Porel oriente y suroriente, la CGSM limita con el pie-demonte de la Sierra Nevada de Santa Marta de
donde provienen los ríos Frío, Sevilla, Aracataca,Fundación y Ají los cuales desembocan en la Ciéna-ga Grande. Por el occidente, la Ciénaga limita con elcomplejo de ciénagas y canales denominado Pajara-les, con el cual se comunica directamente a través delos caños Grande, Clarín y Tambor. Así mismo, elcomplejo Pajarales esta comunicado con el RíoMagdalena por una serie de caños.
El clima de la región es semiárido, presentan-do cuatro épocas climáticas: seca (diciembre aabril), período lluvioso menor (mayo-junio), secomenor (julio-agosto), y época de lluvias (septiem-bre-noviembre). El área esta influenciada por vien-tos variables en velocidad y duración. En la épocaseca dominan los vientos alisios del norte y nores-te, la velocidad de los cuales varía en el transcursodel día de 1 m·s-1 en las horas de la mañana hasta 9m·s-1 en las horas de la tarde. Desde mayo, cuandoempieza la época de lluvia menor, los vientos se de-bilitan y no poseen una dirección dominante. En laépoca de máximas lluvias soplan los vientos débi-les ecuatoriales de sur o dirección variable.
479INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
Resumen:
En el presente estudio se describe detalladamente la estructura del bloque químico-biológico deun modelo matemático bi-dimensional del ecosistema en un dominio de aguas someras y sus mé-todos de calibración. El modelo incluye las ecuaciones de equilibrio para los componentes si-guientes del ecosistema: fitoplancton, bacterioplancton, zooplancton, materia orgánica muerta,fosfatos, amonio, nitritos, nitratos y oxígeno disuelto. Se presentan los resultados de los cálculosde distribución espacial de los componentes del ecosistema para la Ciénaga Grande de Santa Mar-ta en varias épocas del año.
El objeto del estudio se centra en implementar un modelo de ecosistema para la Ciénaga Grandede Santa Marta (CGSM), con el fin de: 1) producir información multipropósito que sirva como he-rramienta para la toma de decisiones por parte de las autoridades ambientales nacionales o loca-les, y 2) apoyar a los requerimientos de información para proyectos y planeamientos ambientalesdel medio marino, atmosférico y del litoral. Para ello, se realiza el acople de bloques hidrodinámi-cos y químico-biológicos que permiten hacer diagnósticos y pronósticos de las características bió-ticas y abióticas de la Ciénaga, así como simular escenarios de alternativas de soluciones a los pro-blemas ambientales de la misma.
Palabras clave: eutrofización, modelo de calidad de agua, ecosistema costera
Centro de Investigaciones Oceanográficas e Hidrográficas, Escuela Naval de Cadetes Almirante Padilla, Cartagena, Colombia.1 Capitán de Fragata, Biólogo Marino, Msc. Tel.+57+56+694465. E-mail:l [email protected] Ingeniero Oceanólogo, Ph.D. Tel.: +57+56+694465. E-mail: [email protected] Ingeniero Oceanólogo, Ph.D. Tel.: +57+56+694465. E-mail: [email protected]
Artículo recibido el 5 de junio de 2002, recibido en forma revisada el 5 de marzo de 2003 y aceptado para su publicación el 29 de abril de 2003. Puedenser remitidas discusiones sobre el artículo hasta seis meses después de la publicación del mismo siguiendo lo indicado en las “Instrucciones para auto-res”. En el caso de ser aceptadas, éstas serán publicadas conjuntamente con la respuesta de los autores.
MODELO NUMÉRICO DE CALIDAD DE AGUAS PARA LA
CIÉNAGA GRANDE DE SANTA MARTALuis A. Calero1,Yuri S. Tuchkovenko2 y Serguei A. Lonin3

Figura 1. Esquema de localización de la Ciénaga Grande de SantaMarta, del sistema lagunar y los canales de comunicación con elrío Magdalena y el Mar Caribe.
El complejo de la CGSM se caracteriza por suriqueza forestal y faunística; así como por su altaproducción primaria, lo que le permite mantenerimportantes poblaciones (especialmente de peces),que son fundamentales en la economía de los po-bladores de la región.
La construcción en la década de los años 70,de una carretera a lo largo de la costa del mar Cari-be, causó una disminución del intercambio deaguas de la CGSM con el mar. Como consecuenciade lo anterior, se experimentó un aumento de la sa-linidad, mortandad de los manglares, alteracionesen la cadena biótica de este ecosistema y cambiosdel régimen hidroquímico de la CGSM y del Com-plejo Pajarales.
Con el fin de recuperar las características na-turales del ecosistema en general y disminución dela salinidad de las aguas en particular, en el princi-pio de los años 90 algunos de los caños de conexiónentre la CGSM y el río Magdalena fueron profun-dizados. Sin embargo, el resultado deseado no se halogrado, aun cuando la salinidad de las aguas hadisminuido considerablemente. Con el aumento delflujo de agua empezó a llegar mayor cantidad denutrientes, lo que contribuyó a la eutrofización dela Ciénaga.
El los últimos años se presentó una disminu-ción importante de la salinidad, producto del incre-mento de aporte de agua del río Magdalena lo queafectó el desarrollo de los organismos acuáticoscaracterísticos de la Ciénaga, tales cómo las ostrasy camarones. En el complejo cenagoso de Pajara-les, la disminución de la salinidad llevó a un incre-mento en la biomasa de peces de agua dulce.
Por lo tanto, la experiencia demostró queefectuar un manejo ambiental del ecosistema te-niendo en cuenta un solo criterio, en este caso la sa-linidad, no es correcto, por cuanto la complejidadde interrelaciones existentes entre los componentesbióticos y abióticos obliga a analizar otros aspectospara el manejo del ecosistema.
En otras palabras, es necesario un enfoqueecosistémico en la planificación de la protecciónambiental de la CGSM y Complejo Cenagoso dePajarales por lo que los autores desarrollaron unmodelo matemático 2-D para el ecosistema estudia-do. La descripción de la estructura de este modelo,el esquema de su calibración y los resultados de lamodelación, son los objetivos del presente artículo.
El modelo fue desarrollado para el dominio dela CGSM. El intercambio de masas de agua entreeste dominio y el río Magdalena a través del siste-ma de canales y ciénagas del Complejo Pajarales,se calculó con base en un modelo hidráulico de ba-lance, desarrollado en la Universidad Nacional deMedellín. Los datos de este modelo fueron introdu-cidos como las condiciones de contorno para el mo-delo considerado.
MODELO MATEMÁTICO
El modelo matemático del ecosistema de laciénaga esta conformado por dos bloques:
Bloque hidrodinámico: Describe la dinámicadel agua y del transporte de sustancias pasivasy conservativas bajo distintas condiciones hi-dro-meteorológicas, teniendo en cuenta los pa-rámetros morfométricos de la cuenca.
Bloque de eutrofización y del régimen de oxí-geno: Describe los procesos naturales químicobiológicos, responsables por el balance de lamateria y energía en el ecosistema, su niveltrófico y sapróbico. En este bloque se descri-ben los procesos de producción-descomposi-ción de la materia orgánica, utilización de los
480
Luis A. Calero,Yuri S. Tuchkovenko y Serguei A. Lonin
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
·
·

nutrientes por fitoplancton y su regeneraciónpor bacterioplancton, transformación de la ma-teria y energía pasando por la cadena trófica enel componente biótico del ecosistema.
La unión entre estos dos bloques en el mode-lo de calidad de agua, se hace mediante la siguien-te ecuación de transporte de una sustancia no con-servativa C:
donde U y V son componentes por los ejes x ey de la velocidad promediada en la vertical de la co-rriente, calculada en el bloque hidrodinámico; H esprofundidad; t es tiempo; Dh es el coeficiente de di-fusión turbulenta en horizontal; Q0 y QH son flujosde la sustancia a través de la superficie y el fondo; Fson fuentes internas de C, relacionadas con las reac-ciones químico-biológicas en el ecosistema. Los ín-dices ‘x, ‘y y ‘t significan las respectivas derivadas.
En cada paso temporal se resuelve un sistemade ecuaciones de transporte de las sustancias noconservativas. El número de ecuaciones del tipo (1)en este sistema corresponde a la cantidad de loscomponentes simulados del ecosistema. La partederecha de estas ecuaciones contiene funciones noconservativas Fi(Ci, x, y, t) para cada elemento i delecosistema. Estas funciones presentan las sumas al-gebraicas de los términos determinados con base enel balance para los flujos locales de la materia entrelos componentes del modelo, relacionados con di-ferentes reacciones (bio)químicas y las interaccio-nes biológicas:
dCi /dt = Fi,
donde la derivada va a significar la variacióninterna en cada punto local independiente de la va-riación espacial.
Bloque hidrodinámico
Debido a que la profundidad promedio de laCiénaga Grande de Santa Marta es 1.6 metros, ladinámica es descrita de acuerdo a la teoría de aguassomeras. Las ecuaciones básicas del bloque hidro-dinámico del modelo son:
donde H = d + h; d es la profundidad local; h,las elevaciones de agua; f, el parámetro de Coriolis;Pa, la presión atmosférica; g, la gravedad; αo, el vo-lumen específico de agua; Ah, la turbulencia hori-zontal; τsx, y τsy son componentes de la tensión su-perficial del viento; τbx y τby son tensiones del fon-do; E es la proporción de evaporación; R es la pre-cipitación.
Los siguientes términos:
salen de la integración de las ecuaciones bási-cas 3-D para los componentes u, v y w con res-pecto a la coordenada vertical (σ = z/H), del fondo(σ = 0) a la superficie (σ = 1). Estas expresiones,cuando difieren de la unidad, sirven para parame-trizar (analíticamente) la estructura vertical de flu-jo para el modelo 2-D, y se encuentra que no hayninguna necesidad de aumentar (artificialmente) laturbulencia horizontal para describir el efecto dedispersión vertical en cómputos de profundidad-promediados (Rodi, 1993).
Estos parámetros pueden escribirse como:
Para el perfil logarítmico,
donde z0 es un parámetro de rugosidad.
481
MODELO NUMERICO DE CALIDAD DE AGUAS PARA LACIENAGA GRANDE DE SANTA MARTA
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
(1)
(2)
(3)
(4)

Las ecuaciones (1)-(4) se han promediado tambiénpara el ancho del flujo en el caso de los canales an-gostos (Hess, 1989). En otras palabras, cuando eldominio de cálculo es de la escala de malla, enton-ces se utilizan directamente las ecuaciones (1)–(4),y cuando el dominio es de la subgrilla se aplican lasecuaciones integradas por el ancho variable. Estoes necesario para la Ciénaga Grande de Santa Mar-ta debido a la presencia de los cauces estrechos delComplejo Cenagoso de Pajarales.
El sistema (1)-(4) requiere las condiciones decontorno. Por ello, en las fronteras del mar abiertose definen los patrones de los campos oceánicos siel flujo esta entrando al dominio del cálculo; se uti-lizan las condiciones del cálculo en el caso inverso:
donde C* es el patrón oceánico para la sustan-cia C; f(C) es una función de la extrapolación delos nodos del dominio al límite abierto (Orlanski,1976); es la proyección normal del vector ac-tual en la vecindad del límite abierto.
Se ha preestablecido el comportamiento delnivel del mar, determinado por las oscilaciones dela marea con la amplitud (h0) de 20 cm y reflejo deondas largas, se especifica como sigue:
En la entrada de las fuentes de contaminaciónse dan flujosQr de agua y las descargas QC de lassustancias estudiadas:
donde bx y by son los anchos adimensionalesde los canales con respecto al tamaño con respectoal tamaño (∆) de celda de la malla de cálculo.
Las ecuaciones (1)-(4) junto con las condicio-nes del límite (5)-(6) se realizan por el método delas diferencias finitas. Las ecuaciones hidrodiná-micas se han aproximado al esquema de Abbott(Sobey, 1970) y la ecuación de transporte por el es-quema de Flujo Corregido de Transporte (Boris &Book, 1976).
Bloque de eutrofización y del régimen deoxígeno
El funcionamiento del ecosistema de laCGSM se describe teniendo en cuenta en la diná-mica de las siguientes características agregadas a suestado: Bf – fitoplancton; Bb – bacterioplancton; Bz– zooplancton; Borg – materia orgánica muerta(MOM); CPO4 – fosfatos; CNH4 – amonio; CNO2 – ni-tritos; CNO3 – nitratos; CO2 – oxígeno disuelto.
Fitoplancton, Bf , [mgC m-3]
El aumento de biomasa de fitoplancton es elresultado de la fotosíntesis, mientras que su dismi-nución está causada por mortalidad natural, gastosrespiratorios de las células, su precipitación gravi-tacional y el consumo por zooplancton. La produc-ción de fitoplancton se limita por luz (transparenciade agua) y por el contenido de los nutrientes. Estosprocesos se presentan como
Aquí, σf es tasa específica de crecimiento defitoplancton, [h-1], que depende de la luminosidad Iy del contenido de los nutrientes – formas mineralesdel nitrógeno CN y fósforo CPO4; Vf
max es la tasa má-xima posible de crecimiento de fitoplancton, [h-1]; I0es flujo de la radiación solar fotosintética activa através de la superficie del mar, [Wm-2]; Iopt es la lu-minosidad óptima para fotosíntesis, [Wm-2];ckN,ckPO4 son constantes de semi-saturación de la in-tensidad del proceso de utilización de las formasminerales de nitrógeno y fósforo por fitoplancton,[mgl-1]; α es coeficiente integral de extinción de luzcon la profundidad [m-1]; γf es parte de producciónde fitoplancton que se va para abastecimiento de sufuncionamiento (respiración); µf es la tasa especí-fica de mortandad natural de fitoplancton, [h-1]; Wges la velocidad de caída de las células por gravedad,[m h-1]. El tiempo t está en horas.
482
Luis A. Calero,Yuri S. Tuchkovenko y Serguei A. Lonin
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
(5a)
(7)
(7a)
(7b)
(5b)
(6)
(7c)

Bacterioplancton Bb, [mgC m-3]
Así mismo, como el fitoplancton, este elemen-to juega un papel fundamental en el funciona-miento del ecosistema. Su participación está ce-rrando el ciclo de los nutrientes en el ecosistema. Lasbacterias utilizan la materia orgánica autóctona yalóctona como una fuente de materia y energía.Además, como resultado de la actividad metabólicabacteriana, las formas minerales de nitrógeno y fós-foro se regeneran. Para satisfacer a sus requerimien-tos energéticos, las bacterias están oxidando bioquí-micamente tres veces más la materia orgánica muer-ta que se requiere para la construcción de sus célu-las. La producción de bacterias se define por la dis-ponibilidad de la materia orgánica muerta, condicio-nes térmicas y del oxígeno. La pérdida de biomasade bacterias se sucede por la mortandad natural y elconsumo por zooplancton. La respectiva ecuación es
donde Vbmax es tasa máxima posible del crecimien-
to de bacterias, [h-1];
es tasa específica de mortandad
natural de bacterias, [h-1]; Bbmax es biomasa máxi-
ma posible de bacterias, [mgC m-3]; Bkorg es cons-tante de semisaturación de crecimiento, igual a laconcentración del sustrato bajo la cual la tasa espe-cífica real del crecimiento de bacterias es la mitadde la máxima, [mgC m-3]; εing = f(CO2) es el pará-metro de inhibición de los procesos de oxidaciónbioquímica de la materia orgánica muerta y nitrifi-cación por el déficit del contenido de oxígeno en elagua del mar (0 ≤ εing ≤1), que se calcula según laformula: εing = CO2 / (CO2+ CkO2 ) donde CkO2 esconstante de semisaturación del proceso, [mg l-1].
Zooplancton Bz, [mgC m-3]
Se supone que la base alimenticia para el zoo-plancton es el fitoplancton, la materia orgánicamuerta y las bacterias. Se tiene en cuenta también laposibilidad del consumo propio de un grupo de zo-oplancton por el otro. La disminución del contenidode zooplancton se presenta por su respiración y lamortandad natural. La ecuación correspondiente es
Aquí, Vzmax es la tasa específica máxima posi-
ble de crecimiento de zooplancton, [h-1]; γz
es la ta-sa específica de excreciones metabólicas de zoo-plancton, [h-1]; µz es la tasa específica de mortandadnatural de zooplancton, [h-1]; Gj es la tasa de consu-mo de fitoplancton (j=1), bacterioplancton (j=2) ymateria orgánica muerta (j=3) por zooplancton, re-presentando su base alimenticia, [mgC m-3h-1]; ωjson coeficientes de utilización de los alimentos porzooplancton en su crecimiento; ρi y ρk, pi y pk soncoeficientes de selectividad en el consumo de distin-tos tipos de alimentación por el zooplancton; BK esconstante de semisaturación por los alimentos en elproceso de crecimiento de zooplancton, [mgC m-3].
Materia orgánica muerta Borg, [mgC m-3]
Esta materia aparece como el resultado demortandad natural de fitoplancton, bacterias, zoo-plancton y los restos de alimentos no absorbidospor zooplancton. La materia orgánica alóctona pro-viene de los ríos y las cienagas adyacentes. La res-pectiva ecuación para este componente es:
donde θ es coeficiente económico que tiene encuenta los gastos de la energía para el crecimiento;Qorg es el aporte de la orgánica alóctona desdelas fuentes externas, [mgC m-3h-1].
Fósforo de fosfatos, CPO4, [mgP l-1]
La variabilidad de fosfatos en la capa fótica sedefine por la intensidad de su consumo por partedel fitoplancton en el proceso de fotosíntesis y porla regeneración de este elemento en el proceso deoxidación bioquímica de la materia orgánica muer-ta por bacterias. Además, hay que considerar las ex-creciones del fito- y zooplancton durante la respira-ción. Se tiene en cuenta el flujo de fosfatos hacialos sedimentos del fondo, el cual se forma como elresultado de descomposición de la materia orgánicaacumulada en los sedimentos. La ecuación es
483
MODELO NUMERICO DE CALIDAD DE AGUAS PARA LACIENAGA GRANDE DE SANTA MARTA
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
(8)
(11) (9)
(10)
ext

donde βP/C
es coeficiente estequiométrico de tras-paso de mgC a mgP, [mgP mgC-1], para la materiaorgánica muerta Borg; Qorg es la entrada de fosfatosde las fuentes externas, [mgP l-1h-1]; QH
PO4 es tasa decambio de concentración de fosfatos por el inter-cambio de su masa entre el agua y los sedimentosdel fondo, [mgP l-1h-1]; βm3/L = 0.001 es coeficientede traspaso de m3 a litros, [m3l-1]. El primer térmi-no en la parte derecha de (7) describe la regenera-ción de fosfatos en el metabolismo de bacterias Rb,teniendo en cuenta que
Nitrógeno amoniacal, CNH4, [mgN l-1]
Se consume por el fitoplancton en el procesode producción primaria de la materia orgánica y seoxida bajo las condiciones aeróbicas hasta los ni-tratos (la primera fase de nitrificación). Esta formadel nitrógeno mineral es la preferible en la absor-ción por el fitoplancton. La formación del conteni-do del amonio en el medio marino ocurre por lamineralización de la materia orgánica muerta conparticipación de bacterias, por la excreción metabó-lica en la respiración de fitoplancton y zooplanctony por la interacción con los sedimentos del fondo:
donde νΝ1 es tasa específica de la primera fa-se de nitrificación, [h-1]; βN/C es coeficiente este-quiométrico de traspaso de mgC a mgN, [mgNmgC-1], para la materia orgánica muerta Borg;
es la parte del ni-trógeno mineral, consumida por el fitoplancton enla forma amoniacal, donde φes coeficiente de pre-ferencia del consumo del amonio por el fitoplanc-ton con respecto a los nitritos y nitratos; QNH4 es ta-sa de variación de la concentración del amonio porel intercambio con el fondo, [mgN l-1h-1].
Nitrógeno de nitritos, CNO2, [mgN l-1]
Es la forma intermedia del nitrógeno mineralen el proceso de nitrificación. Se supone que la va-riabilidad del contenido de los nitritos en el agua sedefine por la relación entre la primera y segunda fa-se de nitrificación y por su absorción en el fito-plancton en el proceso de fotosíntesis:
Aquí, νΝ2 es tasa específica de la segunda fase de
nitrificación, [h-1]; ; QNO2 es la
entrada de los nitritos desde las fuentes externas,[mgN l-1h-1].
Nitrógeno de nitratos, CNO3, [mgN l-1]
Es el producto final del proceso de nitrifica-ción del nitrógeno mineral y es su forma más esta-ble. Bajo las condiciones aeróbicas en el mar, laconcentración de los nitratos se define por la inten-sidad de su utilización en el fitoplancton en el pro-ceso de fotosíntesis y su acumulación como el re-sultado. Bajo las condiciones anaeróbicas, los ni-tratos se regeneran hasta nitrógeno molecular en elproceso de desnitrificación:
donde ; QNO3 es la entrada de
los nitratos de las fuentes externas, [mgN l-1h-1].
Oxígeno disuelto, CO2, [mg l-1]
La dinámica del oxígeno disuelto en las aguasdel mar se determina por la intensidad del procesode fotosíntesis, el intercambio con la atmósfera, suconsumo en la respiración de bacterias y zooplanc-ton, y también en la nitrificación:
Aquí, βO2/C es coeficiente de traspaso de mgCa mgO2, [mgO2 mgC-1]; βO2/N1, βO2/N2 son equivalen-tes de oxígeno para la primera y segunda fase de ni-trificación, respectivamente, [mgO2 mgN-1]; QO2
bot esel consumo del oxígeno desde una columna deagua con la base unitaria como resultado de los pro-cesos bioquímicos de oxidación y nitrificación enlos sedimentos del fondo, [mgO2 m-2h-1]; QO2
atm es elflujo de oxígeno por el intercambio gaseoso con laatmósfera, [mgO2 m-2h-1].
484
Luis A. Calero,Yuri S. Tuchkovenko y Serguei A. Lonin
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
ext
(12)
(13)
ext
ext
(14)
ext
(15)

La absorción del oxígeno en el fondo, en laprimera aproximación, se puede estimar con ayudade una fórmula empírica del tipo QO2
bot = a(CO2)b
con base en la información sobre el contenido deloxígeno en el agua CO2, donde a y b son coeficien-tes empíricos.
El intercambio de oxígeno con la atmósfera secalcula mediante la siguiente expresión:QO2
atm=ζe,invnt(CO2S - CO2) donde QO2
atm es el flujo deoxígeno, [mg m-2h-1]; nt es coeficiente térmico; nv escoeficiente integral de viento; CO2
S es la concentra-ción de saturación del oxígeno, [mg l-1]; ζi,e son co-eficientes de invasión (evasión), [lm-2h-1].
El bloque químico-biológico del modelo estábasado en la síntesis de los modelos de Fasham etal., (1990), IBSM (1991) y Leonov (1992) para al-canzar la mejor correspondencia entre la compleji-dad de la estructura matemática del modelo, totali-dad y adecuación en la descripción del funciona-miento de distintos componentes del ecosistema yla disponibilidad de los datos de monitoreos ecoló-gicos en la cuenca.
Precalibración
Se efectuó con base en los datos de monitoreoefectuado en la CGSM desde los años 80. Sin em-bargo, solo a partir del año 1993 (después de la re-construcción de los canales entre la Ciénaga y el ríoMagdalena), el monitoreo obtuvo un carácter siste-mático. La lista de observaciones incluyó la determi-nación mensual de los siguientes parámetros: NH4
+,NO2-, NO3-, PO4
3-, O2, SiO3, clorofila “a”, seston or-gánico, transparencia, temperatura y salinidad deagua. Periódicamente fueron efectuadas campañashidro-biológicas para determinación de la producti-vidad de fitoplancton dependiendo de los factoresque lo afecten (nutrientes, transparencia, biomasa defitoplancton), la biomasa de fitoplancton y zooplanc-ton, la cantidad de bacterias. Experimentalmente,fueron determinados los flujos del nitrógeno mineraly del fósforo desde los sedimentos hacia el agua.
Una deficiencia principal en la organizaciónde este monitoreo ecológico fue la ausencia de me-diciones del contenido de la materia orgánica muer-ta en el agua, la cual se considera uno de los com-ponentes básicos del ecosistema. Las estimacionesdel contenido de la materia orgánica muerta a tra-vés de los indicadores de la DBO (demanda bioló-gica de oxígeno), de oxidación de KMnO4 y por elcontenido del detritus en el seston, fueron iniciadassolo a partir del segundo semestre del año 1999.
Otro aspecto importante, a tener en cuenta enla utilización de los datos del monitoreo en la cali-bración del modelo, fue la circunstancia de que lamayor cantidad de mediciones hidrobiológicas fueefectuada antes de la reconstrucción de los canalesy el aumento del nivel de eutrofización de la Ciéna-ga. El análisis de datos del monitoreo testifica quedespués de profundización de los canales en losaños 90, el contenido del nitrógeno mineral en lasaguas de la CGSM aumentó, disminuyendo su pa-pel como factor limitante de la producción prima-ria del fitoplancton (Tabla 1).
Tabla 1. Los valores típicos de los parámetros químico – biológicosdel ecosistema de la CGSM durante distintos periodos de las ob-servaciones y para los años extremos.
En las condiciones actuales, el factor principallimitante es la (baja) transparencia del agua. Laconcentración promedia del clorofila “a” fue de 70mgChl”a”m-3 en los años 80, mientras que en la dé-cada de los 90 aumentó de 200 hasta 250mgChl”a”m-3. El contenido de fosfatos aunque au-mentó de 0.03 mgl-1 hasta 0.09 mgl-1, realmente noafectó a la producción primaria del ecosistema, de-bido a que a partir de los años 80, su concentraciónsuperaba las necesidades del fitoplancton.
Calibración del bloque químico-biológico.
En el área de estudio, el ciclo anual de los pa-rámetros extremos que están perturbando el ecosis-tema (temperatura de agua, cantidad de luz, etc.) sepresenta en forma no pronunciada y es comparablepor amplitud con las variaciones de períodos cortos(perturbaciones de unos días). El balance de losprocesos de producción y descomposición en elecosistema se cumple no en escala de un año comoocurre en las latitudes altas, sino durante un perío-do de unos días. La variabilidad de los componen-tes del ecosistema es mucho menor en comparacióncon la de las latitudes altas.
Teniendo en cuenta lo anterior, la calibracióndel bloque químico-biológico fue efectuada de formaasintótica; es decir, el fin del proceso fue el alcancede un estado estacionario (equilibrado) del ecosiste-ma bajo las condiciones hidrometeorológicas me-
485
MODELO NUMERICO DE CALIDAD DE AGUAS PARA LACIENAGA GRANDE DE SANTA MARTA
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
dias para todo el año. Este estado se caracterizó co-mo un estado estable de oscilaciones diarias con losvalores promedios correspondientes a los datos demediciones.
En la Tabla 2 se presentan las concentracionestípicas de los componentes químico-biológicos delecosistema de la CGSM, obtenidas a través de unpromedio de los datos de monitoreo de 1993-2000para el punto central de la Ciénaga.
La estimación preliminar de los parámetrosdel bloque químico-biológico fue efectuada con ba-se en las fuentes bibliográficas (LGMI, 1979; Par-son et al., 1984; GOIN, 1987; IOAN, 1989; Fas-ham et al., 1990; IBSM, 1991; DHI, 1994) sobrelos valores característicos de los parámetros bajocondiciones semejantes a las observadas en lacuenca estudiada.
La primera fase de calibración del modelo fueefectuada en su versión 0-dimensional. Bajo estaformulación, toda la Ciénaga se considera como unasola celda, el volumen y la profundidad correspon-den al volumen real y a la profundidad media de laCGSM. La variante 0-D permite obtener el balanceneto de los procesos químico-biológicos, sin involu-crar la re-distribución espacial de las sustancias,producida por el bloque hidrodinámico. El objetivoprincipal de la calibración con base en la variante 0-D fue el alcance del ciclo diario de los parámetrosquímico-biológicos, conocidos por sus valores típi-cos desde las mediciones (Tabla 2), además de laconservatividad del balance de los procesos de pro-ducción-descomposición en el ecosistema bajo lascondiciones de luz medias durante un año.
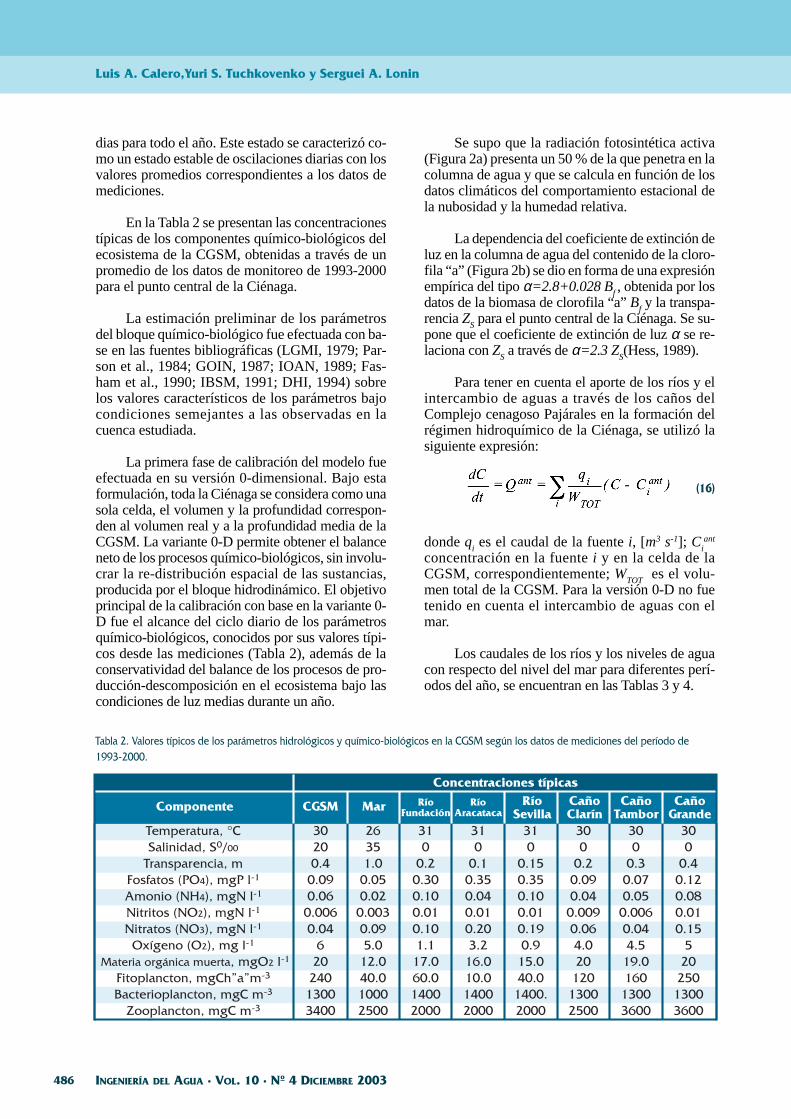
Se supo que la radiación fotosintética activa(Figura 2a) presenta un 50 % de la que penetra en lacolumna de agua y que se calcula en función de losdatos climáticos del comportamiento estacional dela nubosidad y la humedad relativa.
La dependencia del coeficiente de extinción deluz en la columna de agua del contenido de la cloro-fila “a” (Figura 2b) se dio en forma de una expresiónempírica del tipo α=2.8+0.028 Bf , obtenida por losdatos de la biomasa de clorofila “a” Bf y la transpa-rencia ZS para el punto central de la Ciénaga. Se su-pone que el coeficiente de extinción de luz α se re-laciona con ZS a través de α=2.3 ZS(Hess, 1989).
Para tener en cuenta el aporte de los ríos y elintercambio de aguas a través de los caños delComplejo cenagoso Pajárales en la formación delrégimen hidroquímico de la Ciénaga, se utilizó lasiguiente expresión:
donde qi es el caudal de la fuente i, [m3 s-1]; Ciant
concentración en la fuente i y en la celda de laCGSM, correspondientemente; WTOT es el volu-men total de la CGSM. Para la versión 0-D no fuetenido en cuenta el intercambio de aguas con elmar.
Los caudales de los ríos y los niveles de aguacon respecto del nivel del mar para diferentes perí-odos del año, se encuentran en las Tablas 3 y 4.
486
Luis A. Calero,Yuri S. Tuchkovenko y Serguei A. Lonin
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
Tabla 2. Valores típicos de los parámetros hidrológicos y químico-biológicos en la CGSM según los datos de mediciones del período de1993-2000.
(16)
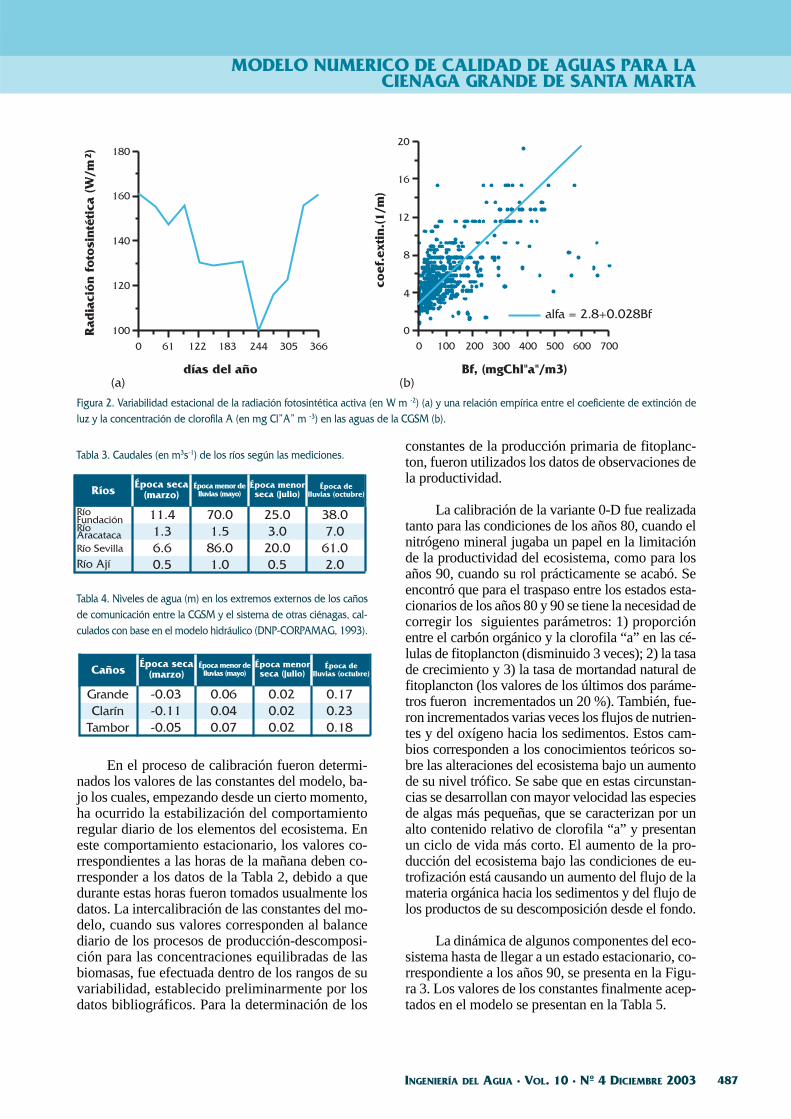
Tabla 3. Caudales (en m3s-1) de los ríos según las mediciones.
Tabla 4. Niveles de agua (m) en los extremos externos de los cañosde comunicación entre la CGSM y el sistema de otras ciénagas, cal-culados con base en el modelo hidráulico (DNP-CORPAMAG, 1993).
En el proceso de calibración fueron determi-nados los valores de las constantes del modelo, ba-jo los cuales, empezando desde un cierto momento,ha ocurrido la estabilización del comportamientoregular diario de los elementos del ecosistema. Eneste comportamiento estacionario, los valores co-rrespondientes a las horas de la mañana deben co-rresponder a los datos de la Tabla 2, debido a quedurante estas horas fueron tomados usualmente losdatos. La intercalibración de las constantes del mo-delo, cuando sus valores corresponden al balancediario de los procesos de producción-descomposi-ción para las concentraciones equilibradas de lasbiomasas, fue efectuada dentro de los rangos de suvariabilidad, establecido preliminarmente por losdatos bibliográficos. Para la determinación de los
constantes de la producción primaria de fitoplanc-ton, fueron utilizados los datos de observaciones dela productividad.
La calibración de la variante 0-D fue realizadatanto para las condiciones de los años 80, cuando elnitrógeno mineral jugaba un papel en la limitaciónde la productividad del ecosistema, como para losaños 90, cuando su rol prácticamente se acabó. Seencontró que para el traspaso entre los estados esta-cionarios de los años 80 y 90 se tiene la necesidad decorregir los siguientes parámetros: 1) proporciónentre el carbón orgánico y la clorofila “a” en las cé-lulas de fitoplancton (disminuido 3 veces); 2) la tasade crecimiento y 3) la tasa de mortandad natural defitoplancton (los valores de los últimos dos paráme-tros fueron incrementados un 20 %). También, fue-ron incrementados varias veces los flujos de nutrien-tes y del oxígeno hacia los sedimentos. Estos cam-bios corresponden a los conocimientos teóricos so-bre las alteraciones del ecosistema bajo un aumentode su nivel trófico. Se sabe que en estas circunstan-cias se desarrollan con mayor velocidad las especiesde algas más pequeñas, que se caracterizan por unalto contenido relativo de clorofila “a” y presentanun ciclo de vida más corto. El aumento de la pro-ducción del ecosistema bajo las condiciones de eu-trofización está causando un aumento del flujo de lamateria orgánica hacia los sedimentos y del flujo delos productos de su descomposición desde el fondo.
La dinámica de algunos componentes del eco-sistema hasta de llegar a un estado estacionario, co-rrespondiente a los años 90, se presenta en la Figu-ra 3. Los valores de los constantes finalmente acep-tados en el modelo se presentan en la Tabla 5.
487
MODELO NUMERICO DE CALIDAD DE AGUAS PARA LACIENAGA GRANDE DE SANTA MARTA
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
Figura 2. Variabilidad estacional de la radiación fotosintética activa (en W m -2) (a) y una relación empírica entre el coeficiente de extinción deluz y la concentración de clorofila A (en mg Cl”A” m -3) en las aguas de la CGSM (b).
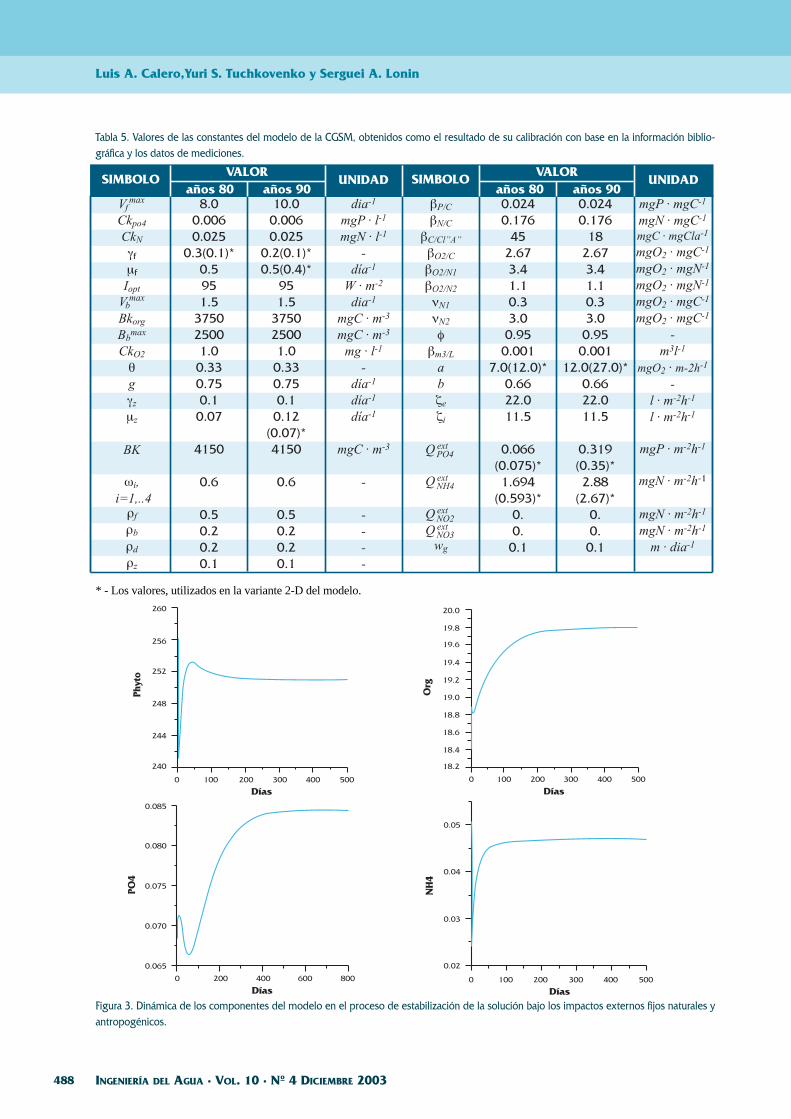
* - Los valores, utilizados en la variante 2-D del modelo.
488
Luis A. Calero,Yuri S. Tuchkovenko y Serguei A. Lonin
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
Tabla 5. Valores de las constantes del modelo de la CGSM, obtenidos como el resultado de su calibración con base en la información biblio-gráfica y los datos de mediciones.
Figura 3. Dinámica de los componentes del modelo en el proceso de estabilización de la solución bajo los impactos externos fijos naturales yantropogénicos.
489
MODELO NUMERICO DE CALIDAD DE AGUAS PARA LACIENAGA GRANDE DE SANTA MARTA
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
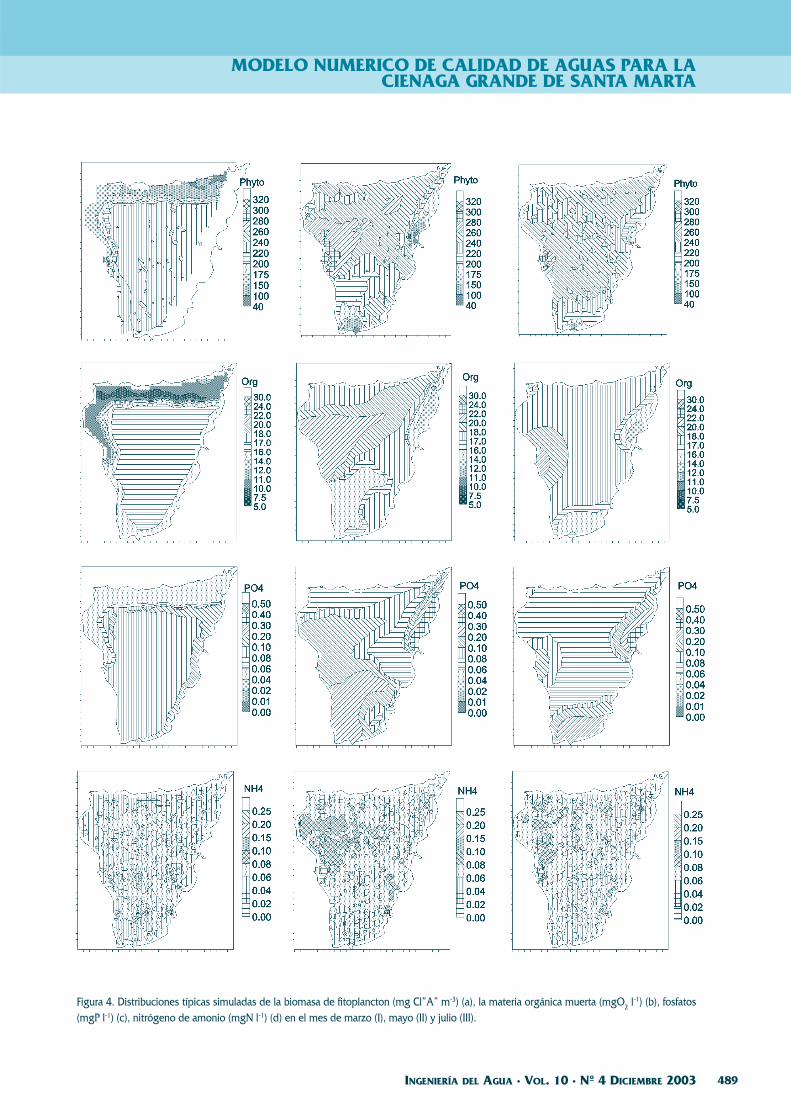
Figura 4. Distribuciones típicas simuladas de la biomasa de fitoplancton (mg Cl”A” m-3) (a), la materia orgánica muerta (mgO2 l-1) (b), fosfatos(mgP l-1) (c), nitrógeno de amonio (mgN l-1) (d) en el mes de marzo (I), mayo (II) y julio (III).
490
Luis A. Calero,Yuri S. Tuchkovenko y Serguei A. Lonin
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003
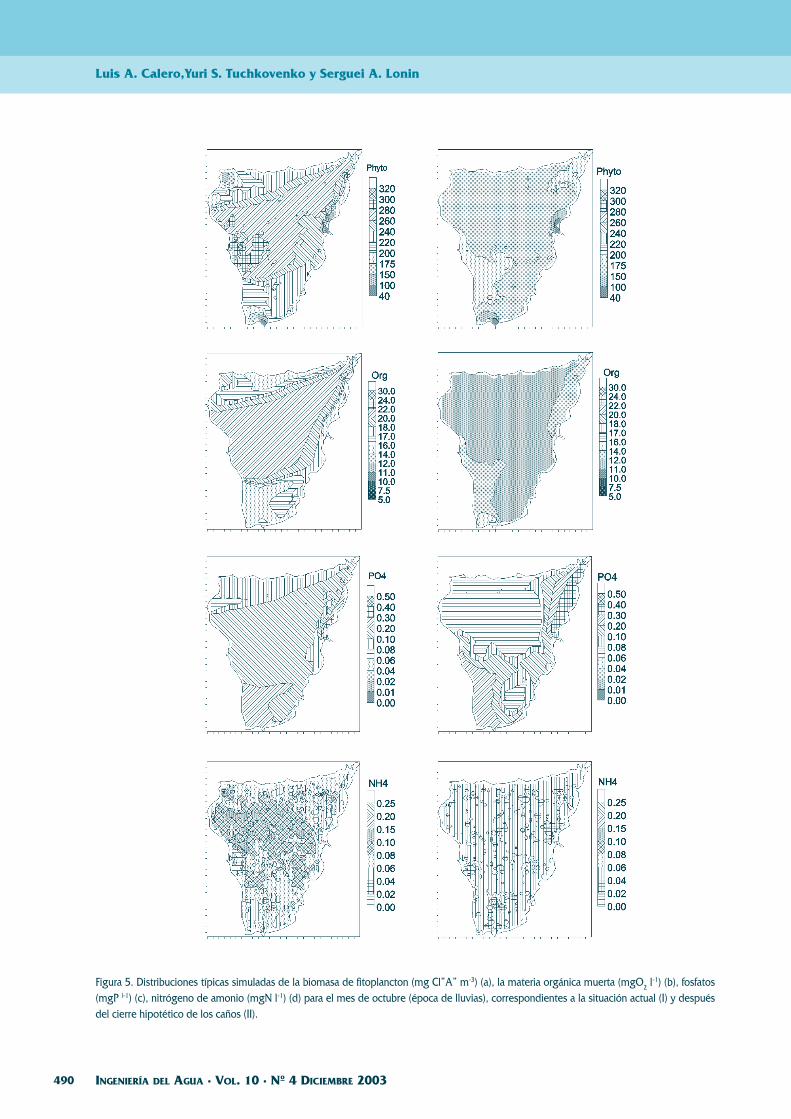
Figura 5. Distribuciones típicas simuladas de la biomasa de fitoplancton (mg Cl”A” m-3) (a), la materia orgánica muerta (mgO2 l-1) (b), fosfatos(mgP l-1) (c), nitrógeno de amonio (mgN l-1) (d) para el mes de octubre (época de lluvias), correspondientes a la situación actual (I) y despuésdel cierre hipotético de los caños (II).

Resultados de cálculo.
Los experimentos numéricos fueron efectuadoscon la versión 2-dimensional, conformado por dosbloques: el hidrodinámico con el transporte de unasustancia conservativa y pasiva y el químico-biológi-co (0-D variante del modelo). Los bloques trabajancon distinta sincronización temporal: el bloque hi-drodinámico con un paso de 20 seg., mientras que elquímico-biológico con 1 hora. Esto está relacionado,por un lado, con diferencias en las escalas temporalesde los procesos, y, por el otro, con un aumento de efi-ciencia de cálculo del modelo. En los experimentosse utilizó una malla espacial de 138 por 156 nodoscon un paso de 200 m. El coeficiente de difusión tur-bulenta en horizontal fue igual a 0.06 m2 s-1, de acuer-do con la fórmula de Okubo (Rodi, 1993). En el mo-mento de unificación de los bloques, fue necesariauna corrección de las constantes del bloque químico-biológico por la apariencia de las dimensiones, ade-más, el intercambio con el mar fue incluido.
Los cálculos fueron efectuados para los cuatromeses del año: marzo, mayo, julio y octubre. La du-ración de cada uno fue de 20 días. Como mostraronlos experimentos con la versión 2-D, este lapso essuficiente para el establecimiento de las distribu-ciones espaciales de los componentes del ecosiste-ma. Los resultados de cálculos para estos meses sepresentan en la Figura 4. En estos resultados se ob-serva que en la época seca la productividad de fito-plancton se limita por un bajo contenido del nitró-geno mineral, debido a que su aporte por los ríos enesta época es mínimo, además que los flujos deagua a través de los caños están dirigidos desde elmar a través de la CGSM a las ciénagas adyacentes.Las aguas marinas, relativamente puras, con un ba-jo contenido de nutrientes y materia orgánica pe-netran en la parte central de la Ciénaga.
Durante la época menor de lluvias, el caudalde los ríos aumenta bruscamente y los flujos en loscaños están dirigidos hacia la CGSM. Debido a quedurante este período del año el nivel de agua en laCiénaga es mas alto que en el mar, el intercambiocon el último prácticamente no juega ninguna im-portancia en el transporte de contaminantes.
Durante el mes de julio, los caudales de los rí-os están cerca a sus valores medios anuales y los ni-veles en la CGSM corresponden más al nivel delmar. Como resultado, el nivel de eutrofización dis-minuye en comparación con lo observado en el ma-yo, pero todavía está superando los valores obteni-dos para la época seca (marzo).
En octubre, el río Magdalena es el principalproveedor de nutrientes y materia orgánica muertaque llega a través del sistema de caños y lagunas.Durante la época de lluvias la contaminación de laCGSM es máxima.
El problema actual de la CGSM es la entradano controlada de una cantidad abundante del aguadulce del río Magdalena durante las épocas lluvio-sas. Este problema se podría solucionar con obrasde ingeniería, consistentes en la construcción de es-clusas, que permitan manejar el aporte del río paralas ciénagas, manteniendo las condiciones del maren la Ciénaga Grande y las condiciones de aguadulce en las demás.
La Figura 5 muestra los resultados de cálculopara la época de lluvias (mayor dulcificación) asu-miendo que los caños Grande, Tambor y Clarín es-tán cerrados. Se ve que en este caso la situaciónecológica se mejora para la CGSM, debido a que elcontenido de la materia orgánica en las aguas y sudulcificación se disminuyen.
CONCLUSIONES
La información del monitoreo de 20 años seutilizó para formulación del modelo ecológico pre-sentado en este artículo para la CGSM, donde se lo-gró establecer las distribuciones espaciales de loscomponentes del ecosistema de la Ciénaga.
Se logró la calibración del bloque químico-biológico llegando a obtener la siguiente secuencia:
Determinación de las constantes que definen las ta-sas específicas de los procesos químico-biológicosy los límites de su variabilidad según las fuentes bi-bliográficas.
Interajuste de las constantes del modelo, el cualabastece el balance de los procesos de producción-descomposición de la materia orgánica bajo los va-lores típicos de los componentes bióticos y abióti-cos del ecosistema. Se hace con base en la variante0-D del modelo.
Corrección de las constantes del bloque químico-biológico en el momento de su unificación con elbloque hidrodinámico en un solo modelo 2-D. Seefectuó comparando los datos de observacionescon los componentes del ecosistema obtenidos pa-ra distintos períodos del año.
491
MODELO NUMERICO DE CALIDAD DE AGUAS PARA LACIENAGA GRANDE DE SANTA MARTA
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003

Las constantes calibradas determinan las tasasde los procesos de transformación de la materia yenergía, donde participan los componentes bióti-cos.
El carácter de la distribución espacial de loselementos del ecosistema se define por los siguien-tes factores: diferencia en los niveles de agua en elmar, en la CGSM y en los caños, además de cam-bios de caudales de los ríos y la intensidad de la ra-diación solar. El intercambio entre la CGSM y elrío Magdalena a través del sistema de las ciénagasafecta más al funcionamiento del ecosistema que elaporte de los ríos.
Las concentraciones más bajas de los elemen-tos del ecosistema se observan durante la época se-ca, cuando las aguas relativamente más amplias en-tran a la CGSM. En los períodos de las lluvias in-tensas, las fuentes de contaminación no solamenteson los ríos, sino también las ciénagas adyacentes,conectadas con el río por el sistema de los caños. Elmes que más se acerca a las condiciones promediasanuales es el mes de julio.
El modelo se convierte en una herramientamuy útil, para la búsqueda y aprobación de alter-nativas de mejoramiento de la calidad de aguas y laconservación de productividad de la cuenca.
REFERENCIAS
Boris, J.P. and Book, D.L. (1976). Flux-corrected transport.III. Minimal-error FCT algorithms. J. Comp. Physics,20, pp. 397-431.
DHI (1994). User Guide and Reference Manual Water Qua-lity Module, release 2-4 MIKE21, Danish HydraulicInstitute.
DNP-CORPAMAG (1993). Plan de recuperación del com-plejo lagunar de la Ciénaga Grande de Santa Marta.Diseño obras hidráulicas. Informe final. Bogota. Co-lombia.
Fasham, M.J.R., Ducklow, H.W., McKelvie, S.M.A. (1990).Nitrogen-based model of plankton dynamics in theoceanic mixed layer. Journal of Marine Research, 48,pp. 591-639.
GOIN-Instituto Oceanográfico Estatal (1987). Modelaciónde los componentes del ecosistema. Problemas de in-vestigación y modelación matemática del ecosistemadel Mar Báltico. vol 3., Rusia, Leningrado, Hidromete-oizdat, 255 p.
Hess, K.W. (1989). MECCA - Programs Documentation.NOAA, Technical Report NESDIS 46, Washington,D.C.
IBSM (1991). Modelación de los procesos de autodepura-ción del agua de las zonas litorales al mar. Ucrania, Se-vastopol, 227 p.
IOAN-Instituto de Oceanología (1989). Modelos de los pro-cesos oceánicos. Rusia, Moscú, 366 p.
Leonov, A.V. (1992). Coupled biotransformation if nitrogenand phosphorous compounds. In: Modelling of WaterResource Systems (Ecological-Economical Aspects).IVP RAN, Moscow, pp. 115-124 (in Russian).
LGMI-Instituto Hidrometeorológico de Leningrado (1979).Modelación de los procesos de transporte y transfor-mación de las sustancias en el mar. Rusia, Leningrado,Hidrometeoizdat, 290 p.
Orlanski, I. (1976). Asimple boundary condition for unboun-ded hyperbolic flows. J. Comput. Phys. 21 (3), pp.251-269.
Parsons, T.R., Takahashi, M. and Hargrave, H. (1984). Bio-logical oceanographic processes. Oxford, New York,Pergamon Press, 324 p.
Rodi, W. (1993). Turbulence Models and Their Applicationin Hydraulics. A State-of-the Art Review, 3rd edition.A.A. Balkema, Rotterdam, 104 p.
Sobey, R.J. (1970). Finite-difference schemes compared forwave-deformation characteristics in mathematical mo-deling of two-dimensional long-wave propagation.U.S. Army Corps of Eng., Coastal Eng. Res. CenterTechn. Mem., 32, 29 p.
492
Luis A. Calero,Yuri S. Tuchkovenko y Serguei A. Lonin
INGENIERÍA DEL AGUA · VOL. 10 · Nº 4 DICIEMBRE 2003