membrana (i parte)membrana (i parte) prof. iván...
TRANSCRIPT

Prof. Iván RebolledoMembrana (I parte)Membrana (I parte)Membrana (I parte)Introducción
La estructura y función de las células dependen de las membranas, las cuales no sólo separan el interior de la célula de su medio ambiente sino también definen los compartimentos internos de las células eucarióticas, incluyendo el núcleo y los organeloscitoplasmáticos.
Todas las membranas comparten una estructura común : una bicapade fosfolípidos. A esta bicapa se asocian proteínas y carbohidratos. Las proteínas participan en los procesos de transporte y los carbohidratos en procesos de reconocimiento celular.
En 1935, Hugh Davson y James Danielli propusieron que la membrana plasmática era una bicapa de lípidos sobre la cual quedaban adheridos una capa de proteínas globulares en ambas interfases lípido-agua. Ellos fueron los primeros en intentar explicar las propiedades fisiológicas de la membrana sobre bases estructurales.
El modelo de Davson-Danielli es un modelo laminar, es decir, sus componentes lipídicos y proteicos se encuentran dispuestos en láminas o capas continuas. Estas láminas forman un plano en dos dimensiones de proteínas separadas por un plano de lípidos : un emparedado lipo-proteico. De esta manera, las propiedades hidrofílicasde la membrana se explicaban por las cubiertas proteicas asociadas a los residuos fosfatos de los fosfolípidos y las propiedades hidrofóbicas a su centro lipídico.
Modelos molecularesde la membrana
A. Modelo de Davson-Danielli
Los principales problemas que se plantearon al inicio de los años 30 eran los relacionados con la permeabilidad y las tensión superficial de las membranas naturales. Si la membrana era una bicapa lipídica, ¿cómo es que podía ser atravesada por el agua? y ¿cómo disminuía la bicapa lipídica la tensión superficial frente al medio acuoso?.

Membrana celular (I parte)
El modelo de Robertson considera a la membrana como compuesta de dos líneas electrónicamente densas de 20 Å de espesor con un espaciointerlínea de 35 Å, lo que hace un espesor total de la membrana de 75 Å. Aunque las dos líneas densaseran de 20 Å de espesor podíanobservarse diferencias entre lascapas externa e interna, es decir, había una asimetría con respecto a sus mitades externa e interna.
B. Modelo de unidad de membrana ode Robertson
En los años 50, la resolución del ME ya permitía observar la composición estructural celular, utilizando tetróxido de osmio (OsO4) como fijador. Las membranas celulares se veían como líneas gruesas muy teñidas de negro. Una década después, David Robertsonutilizando permanganato de potasio (KMnO4) como fijador encontró que las líneas gruesas en realidad eran estructuras trilaminares. Todas las membranas que él examinó(animales, vegetales, microrganis-mos) mostraban la misma estructura trilaminar. Esto deducía que la imagen no era un artefacto. Además, la misma imagen aparecía aunque se cambiara de fijador. Así, Robertsonintrodujo el concepto de “unidad de membrana” para describir una estructura que estaba presente en membranas procaróticas y eucarióticas. La pregunta aquí es: ¿qué se tiñe de
negro en la membrana? Cuando se tiñieron membranas a las que se extrajeron los lípidos, se mantenía la estructura trilaminar y cuando se teñían membranas lipídicas sin proteínas la observación de la estructura trilaminar no se había alterado. Estas conclusiones sugieren que las proteínas y las terminaciones hidrofílicas de las moléculas lipídicas se tiñen con el osmio y se convierten en una línea electrónicamente densa al ME:

Membrana celular (I parte)
Por esto, la revisión crítica de este modelo está basada en las siguientes observaciones: (a) en cortes muy finos se aprecia la existencia de puentes que atraviesan la bicapalipídica (ver figura más abajo); (b) por criofractura se reveló la existencia de numerosas partículas en el plano de clivaje de la membrana; y (c) la aplicación de fijadores que evitan la extracción de proteínas ofrece un aspecto granular a la membrana.
Las variaciones de grosor que pueden existir entre las membranas pueden ser explicadas por la relación proteína : lípido. Ejemplos:
Membrana Prot:Lipi
Mielina 0.25
Membrana hepatocito 1.00
Membrana célula. intestinal 4.60
Membrana externa mitocondr 1.20
Membrana interna mitocondr 3.60
Láminas cloroplasto 0.80
Estas diferencias en la relación proteína : lípido puede correlacio-narse con las funciones particulares que desempeñan cada una de estas membranas. Por ejemplo, la membrana interna de la mitocondria posee gran cantidad de proteínas enzimáticas que intervienen en el metabolismo respiratorio y producción de ATP. La membrana presente en las vainas de mielina contienen altas concentraciones de lípidos para lograr un aislamiento eléctrico que permita una eficiente conduc-tibilidad nerviosa.
En la actualidad, el modelo de la unidad de membrana representa una acentuada simplificación de la ultraestructura de la membrana y no toma en cuenta las proteínas que atraviesan dicha membrana.
C. Modelo del mosaico fluído
El modelo del mosaico fluído es el que hasta ahora tiene una aceptación general y enfatiza las características dinámicas de la organización de la membrana, ya que incluye la movilidad e interacción entre las proteínas y los lípidos en la membrana.

Membrana celular (I parte)
de movimientos:a) rotacional: en torno a un eje perpendicular a la superficie de la membrana,b) lateral: desplazamiento en el plano de la membrana, yc) transversal: pasar de una cara de la bicapa a la otra, promovido por una proteína translocadora denomi-nada flipasa.
Las proteínas también experimen-tan movimientos: la rotacional y la lateral, nunca transversal.
Este modelo contempla 2 categorías grandes de proteínas: las integrales y las periféricas. Las proteínas integrales o intrínsecas son de difícil extracción, ya que están mantenidas firmemente en la bicapa por interacciones hidrofó-bicas y/o hidrofílicas. Como se muestra en el esquema, algunas de ellas atraviesan toda la membrana (transmembranales), en cambio otras están asociadas a la capa externa o interna de la bicapa.
Como se mencionó en la sección de proteínas, las características químicas de los radicales laterales de los aminoácidos determinan la estructura terciara de la proteína. Así, las porciones de la proteína que están incluidas en la bicapa lipídicatendrán sus radicales hidrofóbicosorientados hacia las cadenas hidrocarbonadas de los lípidos y los radicales hidrofílicos se orientan
Este modelo descrito en 1972 por S.J. Singer y G.L. Nicolson, considera a la doble capa de lípidos como el esqueleto aglutinante de la membrana, en la cual se encuentran incluidos las proteínas que interactúan con los lípidos y tanto los lípidos como las proteínas pueden realizar movimientos de traslación dentro de la bicapa. Esta posibilidad de movimiento presupone que entre las moléculas de lípidos y proteínas deben existir interacciones débiles, principal-mente de carácter hidrofóbica.
Como puede observarse en el dibujo de arriba, una bicapa de lípidos es la base fundamental de la membrana, en ella se encuentran dispersas varias proteínas, algunas atravesando toda la membrana pero otras solo ocupan una mitad de la misma.
El estado fluido de la membrana implica la movilidad de sus componentes. Recordamos el tema de los lípidos que poseen tres tipos

Membrana celular (I parte)
Las actividades diferentes de los distintos tipos de membranas parecen ser el reflejo de las diferentes clases de proteínas presentes en la membrana o en alguna región de ella. Además las diferencias en el espesor de la membrana pueden estar relacionado con la presencia de cantidades importantes de ciertas proteínas. Por ejemplo, en membranas gruesas predominan las proteínas periféricas en cambio, en las membranas delgadas predominan las proteínas integrales.
hacia el exterior o interior de la membrana. La proteína transmem-branal dispondrá sus radicales hidrofóbicos en la sección que cursa por la membrana y sus radicales hidrofílicos se orientan hacia el interior y al exterior de la membrana, como se muestra en la figura de abajo.
C. Análisis por criofractura
La aplicación de la técnica de criofractura al estudio de las membranas ha aportado muchos conocimientos sobre la estructura de las membranas y ha apoyado el modelo de mosaico fluido.
La técnica de la criofractura se inicia congelando el tejido. Para evitar la formación de cristales de hielo que pudieran destruir las estructuras celulares, la muestra se infiltra con glicerol y se congela rápidamente en freón líquido (o nitrógeno líquido). En corto tiempo el tejido queda congelado y duro. En un borde del mismo se golpea con una cuchilla muy fría, producién-dose un plano de fractura que abarca toda la muestra y que corresponde a regiones de menor unión molecular.
Las proteínas periféricas o extrínsecas son de fácil extracción, ya que están asociadas a la membrana por enlaces electro-estáticos débiles, ya sea a las cabezas hidrofílicas de las moléculas lipídicas o a las porciones hidrofílicas de las proteínas integrales que sobresalen fuera de la membrana. Estas proteínas no llegan a formar una capa continua sobre los lípidos.

Membrana celular (I parte)
Así, pueden aparecer partículas de diferentes tamaños en una o en ambas caras, deduciendo la presencia de proteínas en dichas mitades de la membrana.
En el caso de las membranas (plasmáticas e intracitoplasmáticas) el plano de fractura se produce entre las 2 capas monomoleculares de la bicapa lipídica, es decir, entre la hoja interna y externa de la membrana.
Luego, sobre estas superficies se deposita una fina capa de un metal pesado a fin de producir una réplica la cual pueda observarse al ME. La imagen producida muestra una superficie llena de partículas pequeñas que representan compo-nentes moleculares de la membrana, que por lo menos se extienden hasta la mitad del grosor de la membrana. (ver figura de abajo).
Doble capa de lípidos
Desde los tiempos de Irving Langmuir (1800) se iniciaron los estudios tendientes a averiguar la estructura química de la membrana Langmuir encontró que los lípidos forman capas (monocapas) sobre una superficie hidrofílica, ordenán-dose los grupos hidrofóbicos hacia el aire y los hidrofílicos hacia el agua.
Como el plano de fractura pasa por el centro de la doble capa de lípidos, podrán mostrarse 2 tipos de caras: la cara P de la fractura o cara protoplasmática y la cara E o cara ectoplasmática.

Membrana celular (I parte)
Luego, Gorter y Grendel en 1926 hicieron estudios cuantitativos sobre los lípidos existentes en la membrana de eritrocitos y otras células, encontrando siempre una relación de superficie igual a dos, lo que sugería la existencia de una doble capa de lípidos. Concluyeron que los grupos polares (hidrofílicos) de los lípidos estaban orientados hacia el exterior y los grupos no polares (hidrofóbicos) hacia el interior de la membrana. Sus experimentos y conclusiones ya establecieron las propiedades anfipáticas de las moléculas de lípidos.
Componentes químicos
Las membranas plasmáticas e intracitoplasmáticas están constitui-das por lípidos, carbohidratos y proteínas, en diferentes propor-ciones que varían según los diferentes tipos celulares, como puede observarse en la siguiente tabla.
Proteínas % Lípidos %
Eritrocito hum. 60 39Hepatocito rata 59 40Mielina SNC 20 79Mitocondrias 70 29
bicapa
Lípidos
Las principales moléculas lipídicasencontradas en la membrana son: fosfolípidos, colesterol y galacto-lípidos en proporciones variables según la célula y organelo. A pHneutro, los fosfolípidos pueden ser ácidos o neutros. La gran mayoría (80-90 %) son neutros, dentro de los cuales se destacan la fosfatidilcolina, la fosfatidiletanolamina y esfingo-mielina. El restante 10-20 % son fosfolípidos ácidos que general-mente se encuentran asociados a las proteínas de la membrana, dentro de los cuales mencionamos la fosfatidilserina y fosfatidilglicerol.
En conclusión: la organización de los lípidos en doble capa, la formación espontánea de la bicapa y la cohesión de las moléculas dentro de la bicapa, determinan que las membranas sean estructuras contínuas, sin rupturas, solo desplazándose para acomodar las moléculas de proteínas. Además, son estructura deformables que se adaptan a diferentes procesos celulares: locomoción, reproducción, citosis, etc.

Membrana celular (I parte)
Por el contrario, cuando la temperatura es baja, el colesterol interactúa con los ácidos grasos promoviendo la fluidez. Este comportamiento es vital en las membranas de los animales que hibernan.
Un hecho interesante sobre el colesterol es que está ausente de las membranas de los procariontes y también de la membrana interna de las mitocondrias: una relación muy reveladora para precisar el origen de las mitocondrias (ver capítulo Mitocondria)
Por último, el colesterol se requiere para el crecimiento y diversas funciones celulares, pero sus altos contenidos contribuyen al desarrollo de enfermedades cardiovasculares.
Los gliceroesfingolípidos son minoritarios en las membranas, aunque hay excepciones: 10 % en los eritrocitos, 25% en la vaina de mielina y 30 % en la membrana de las microvellosidades de las células absortivas del epitelio intestinal.
Un fosfolípido importante en los procesos de señalización celular es el fosfatidilinositol, que solo se encuentra en la mitad interna de la membrana. Este lípido suele encontrarse asociado con la fosfatidilserina y ambos están cargados negativamente, lo que confiere a la mitad interna de la membrana su característica carga negativa (ver página 61 de Lípidos).
Un componente lipídico muy importante en la membrana es el colesterol. Como se mostró en el capítulo de lípidos, esta molécula posee un extremo polar muy reducido y uno apolar muy amplio. Este último extremo le permite ubicarse entre los extremos hidrofóbicos de la bicapa lipídica. Dependiendo de la temperatura, el colesterol tiene efectos sobre la fluidez de la membrana. A temperaturas altas, el colesterol interfiere con el movimiento de las cadenas de ácidos grasos de los fosfolípidos de la capa externa de la membrana, aumentando la rigidez y haciendo que sea menos permeable a pequeñas moléculas.
Carbohidratos
Al igual que los lípidos, su distribución en la membrana es asimétrica, ya que los glucolípidos y glucoproteínas sólo se encuentran en la capa externa de la membrana. Así, las células están cubiertas por una capa de carbohidratos, que suele llamarse glucocálix, el cual es muy evidente en la superficie de las células que conforman el epitelio del intestino. Los oligosacáridos del glucocálix tienen doble función: por un lado sirven de protección a las células y, por otro lado, desempeñan

Membrana celular (I parte)
Como se explicó antes, las proteínas se clasifican en extrínsicaso intrínsicas, llamadas también periféricas o integrales, respectiva-mente.
Las proteínas periféricas represen-tan el 30 % de las proteínas de membrana, pueden aislarse con tratamientos químicos suaves y como ejemplo están la espectrina de los eritrocitos y el citocromo C en la mitocondria. En estas proteínas, los aminoácidos hidrofílicos se ubican en la superficie y los hidrofóbicos se ocultan en su interior.
Las proteínas intrínsicas represen-tan el 70 % de las proteínas de membrana, para aislarlas se requiere de tratamientos drásticos y están asociados a carbohidratos y lípidos. En estas proteínas los aminoácidos hidrofóbicos están inmersos en la capa lipídica y los hidrofílicos se exponen hacia las superficies exterior y citoplasmática. La porción hidrofóbica tiende a rodearse de un tipo determinado de lípidos, llamados lípidos anulares cuyas funciones son restringir su difusión lateral, actuar como cofactor de su función y evitar escape de solutos por dicha interfaz.
Algunas de estas proteínas intrínsicas se ubican hacia la superficie citosólica y otras hacia el exterior. Esta ubicación demuestra la disposición asimétrica de las proteínas en la membrana.
funciones de reconocimiento celular, como por ejemplo, los leucocitos (glóbulos blancos) en ocasiones deben abandonar los vasos sanguíneos, las células de los vasos llamadas células endotelialesdeben reconocer a los leucocitos para dejarlos atravesar la pared de los vasos. La membrana de las células endoteliales poseen una proteína llamada selectina que es capaz de reconocer los carbohidra-tos de las glucoproteínas en la membrana de los leucocitos.
Proteínas
Ya que los lípidos son los elementos estructurales fundamen-tales de la membrana, las proteínas constituyen las moléculas que desempeñan las funciones especí-ficas de las membranas, tales como transporte, endocitosis, reconoci-miento, etc.

Membrana celular (I parte)
Los carbohidratos de la glucoforinadeterminan los antígenos de los grupos sanguíneos. El sistema ABO son cadenas de oligosacáridos ya explicados en el capítulo de Carbohidratos (página 50).
Otro punto interesante es que en los carbohidratos de la glucoforinapredomina el ácido siálico (ácido N-actilneuramínico), cuya pérdida progresiva a través de la circulación durante los 120 días de sobrevida, va convirtiendo al eritrocito en célula extraña, la cual será eliminada por los macrófagos existentes en el bazo.
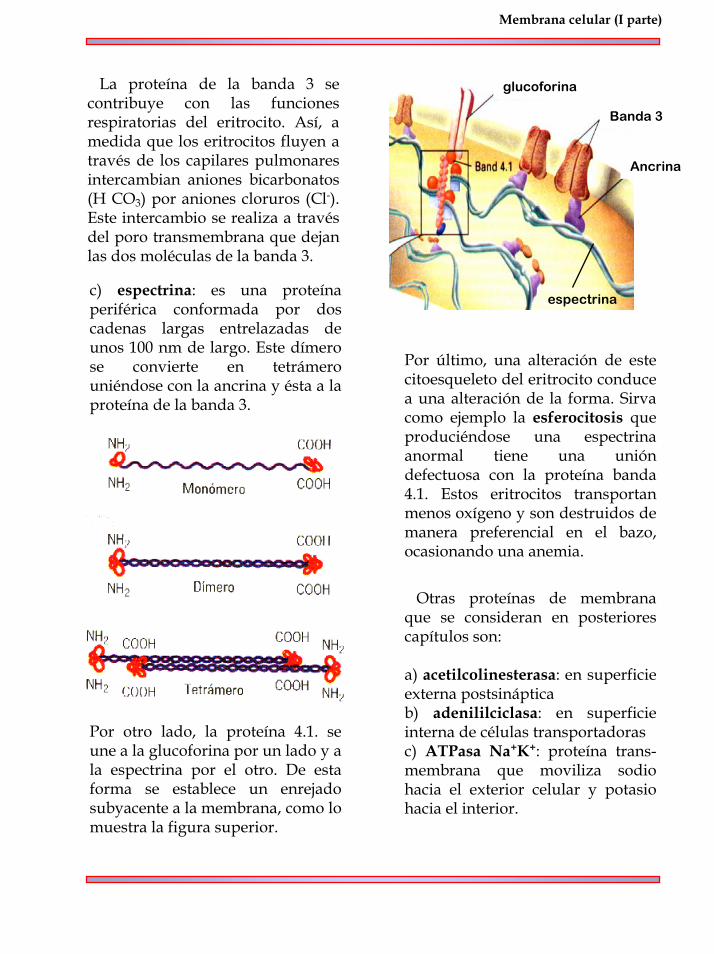
b) proteína de la banda 3: se llama así por su posición en una columna de gel de poliaacrilamida. Es una proteína transmembrana con pocos carbohidratos hacia el exterior.
Una forma de estudiar las proteínas en la membrana ha sido la utilización de eritrocitos, que al ser puestos en soluciones hipotónicas, se hinchan y revientan, quedando trozos de membrana. Una pregunta previa: ¿cómo saber en este trozo cuál superficie es la citosólica y cuál la exterior?
Algunas de las proteínas caracteri-zadas en estos trozos son:a) glucoforina: es una proteína transmembrana, cuya secuencia de 131 aminoácidos forman una simple cadena polipeptídica. La porción amino terminal extracelular es hidrofílica proyectándose hacia el exterior. Unidas a este dominio existen unas 16 cadenas de oligosacáridos, los cuales están unidos a la serina o treonina. La porción intramembranal corres-ponde a la secuencia lineal de 23 aminoácidos dispuestas en alfa hélice con propiedades hidrofóbicas. La porción carboxilo terminal es intracelular.
Como se observa en la figura superior, está conformada por dos unidades (un dímero), cada uno con una larga cadena que atraviesa 12 veces la membrana. Los extremos N y C terminales se ubican en el lado citosólico.
Membrana celular (I parte)
La proteína de la banda 3 se contribuye con las funciones respiratorias del eritrocito. Así, a medida que los eritrocitos fluyen a través de los capilares pulmonares intercambian aniones bicarbonatos (H CO3) por aniones cloruros (Cl-). Este intercambio se realiza a través del poro transmembrana que dejan las dos moléculas de la banda 3.
c) espectrina: es una proteína periférica conformada por dos cadenas largas entrelazadas de unos 100 nm de largo. Este dímero se convierte en tetrámero uniéndose con la ancrina y ésta a la proteína de la banda 3.
glucoforina
Ancrina
Banda 3
espectrina
Por último, una alteración de este citoesqueleto del eritrocito conduce a una alteración de la forma. Sirva como ejemplo la esferocitosis que produciéndose una espectrinaanormal tiene una unión defectuosa con la proteína banda 4.1. Estos eritrocitos transportan menos oxígeno y son destruidos de manera preferencial en el bazo, ocasionando una anemia.
Otras proteínas de membrana que se consideran en posteriores capítulos son:
a) acetilcolinesterasa: en superficie externa postsinápticab) adenililciclasa: en superficie interna de células transportadorasc) ATPasa Na+K+: proteína trans-membrana que moviliza sodio hacia el exterior celular y potasio hacia el interior.
Por otro lado, la proteína 4.1. se une a la glucoforina por un lado y a la espectrina por el otro. De esta forma se establece un enrejado subyacente a la membrana, como lo muestra la figura superior.

Membrana celular (I parte)
1. Anotar 3 evidencias de la asimetría mostrada por las moléculas constituyentes de la membrana :
a) __________________________________________________b) __________________________________________________c) __________________________________________________
2. Nombrar los movimientos que presentan las siguientes moléculas en la membranaa) fosfatidiletanolamina ________________________________________b) transportador de glucosa ________________________________________
3. Anote 2 razones moleculares por las cuales se dice que la membrana plasmática es asimétrica:a) ____________________________________________________________b) ____________________________________________________________
4. ¿A qué se refiere el modelo de la “unidad de membrana” ? ____________________________________________________________________________________________________________________________
5. Si Ud. quisiera obtener una membrana que permanezca fluída en un ambiente con calor y otra fluía con ambiente frío, ¿qué moléculas utilizaría en cada caso?
a) calor ________________________ b) frío _________________________
6. ¿Cuáles serían las consecuencias de una mutación en las siguientes proteínas (anote solo una para cada proteína)
a) glucoforina ___________________________________________________b) proteína banda 3 ______________________________________________c) ATPasa Na-K _________________________________________________
7. ¿Cuál es la función del colesterol en la membrana? ______________________________________________________________________________________________________________________________