localización de los mrna de los dos tipos de somatolactina
TRANSCRIPT

Publicación Previa al VIII Congreso Nacional de Acuicultura
(Santander, mayo 2001)
Localización de los mRNA de los dostipos de somatolactina en hipófisis de dorada
Herrero-Turrión, M.J.; Velasco, A.; Rodríguez, R.; Aijón, J. y Lara, J.Instituto de Neurociencias de Castilla y León (INCyL), Univ. de Salamanca, Facultad deMedicina, Campus Unamuno 37007 Salamanca (España)
Resumen
Se ha analizado por hibridación in situ (ISH) la expresión de mRNA de los dos tipos de somatolactina(SL1 y SL2) descritas hasta el momento en hipófisis de dorada (Sparus aurata). El método de ISH noradiactivo que optimizamos en este trabajo utiliza como sondas oligonucleótidos marcados en su extremo5’ con biotina o digoxigenina, lo que permite realizar ISH simples y dobles de muy alta especificidad ysensibilidad. Las distintas oligosondas de SLs empleadas presentan una fuerte señal fundamentalmente enla pars intermedia (PI) de la glándula pituitaria. Este estudio detecta por primera vez células quecoexpresan ambas formas. Además, en el estudio de tres grupos de tallas diferentes, producidas por elcrecimiento asincrónico de la dorada en cultivo industrial, no hemos apreciado diferencias cualitativas enlos niveles de expresión de la familia de genes GH/PRL.
Palabras clave: Somatolactina, familia GH/PRL, RNAm, hibridación in situ, Sparus aurata
Abstract
Localization of mRNA of the two types of somatolactin in the pituitary of gilthead sea breamThe expression of mRNA of the two types of somatolactin (SL1 and SL2) which have been found, up tothe present, in the pituitary of gilthead sea bream (Sparus aurata) has been analysed by in situhybridization (ISH). The method of non-radioactive ISH which we optimise in this study usesoligonucleotides labelled in extreme 5’ with biotin or dioxygenin as probes, which allows simple anddouble ISH of high specificity and sensitivity to be performed. The distinct probes of SL’s used present astrong signal fundamentally in the pars intermedia (PI) of the pituitary gland. This study detects cellswhich co-express both forms for the first time. Moreover, in the study of three groups of different sizes,produced by asynchronic growth of the bream in industrial cultivation, we did not find qualitativedifferences in the levels of expression of the GH/PRL gene family.
Keywords: Somatolactin, GH/PRL family, mRNA, in situ hybridization, Sparus aurata
Introducción
Se ha propuesto que las hormonas pertenecientes a la superfamilia GH de proteínasprovienen de un gen ancestral común y se desarrollaron por procesos de amplificacióny divergencia (Nicoll et al. 1986; Takayama et al. 1991b). Además, estos genes sontranscritos por un mismo factor de transcripción, Pit-1 (Ono et al. 1994). Lasomatolactina (SL) es una hormona de la familia GH/PRL, fue aislada por primeravez en el bacalao atlántico (Gadus morhua) (Rand-Weaver et al. 1991a), y se produceprincipalmente en la pars intermedia (PI) de la hipófisis de la mayoría de los peces.
La SL ha sido caracterizada exclusivamente en peces, por métodos como: aislamientode la proteína e identificación de su secuencia aminoacídica, clonación de su cDNA odeterminación de la propia secuencia genómica (Tabla 1).
Tabla 1. Especies de peces en los que se ha caracterizado las SLs. El asterisco (*) indica que aunque todavía no se hapublicado la secuencia aminoacídica está accesible en las bases de datos del Gen Bank; s/n indica que esta secuencia

aminoacídica no está incluída en las bases de datos.
Nº acceso Especie GenBank Referencia
ACTINOPTERIGIOTELEÓSTEOS
Percomorpha (Pleuronectiforme)
hhSL Hippoglossus hippoglossus L02117 Iraqi et al. 1993
poSL Paralichthys olivaceus M33695 Ono et al. 1990
ssSL Solea senegalensis U06753 Pendón et al. 1994
tmSL Tetraodon miurus AF253066 Rand-Weaver et al. *
Percomorpha (Perciforme)
saSL1 Sparus aurata P54863 Astola et al. 1996
saSL2 Sparus aurata P79894 Cavari et al. 2000
sgSL Signatus guttatus AB026186 Ayson et al. *
soSL Sciaenops ocellatus AF062520 Zhu et al. 1999
dlSL Dicentrarchus labrax AJ277390 Company et al. 2000
Percomorpha (Scorpaeniforme)
clSL Cyclopterus lumpus L02118 Iraqi et al. 1993
Salmonidae
okSL Oncorhynchus keta D10640 Takayama et al. 1991a, b
omSL Oncorhynchus mykiss s/n Yang et al. 1997
Otros teleósteos
gmSL Gadus morhua D10639 Takayama et al. 1991a, b
aaSL Anguilla anguilla U63884 May et al. 1997
caSL Carassius auratus U72940 Cheng et al. 1997
ipSL Ictalurus punctatus AF267991 Tang 1992
CONDRÓSTEO
atSL Acipenser transmontanus AB017200 Amemiya et al. 1999
SARCOPTERIGIODIPNOI
paSL Protopterus annectens AB017766 Amemiya et al. 1999
En todas las especies examinadas, a excepción de los salmónidos, la SL se puedeencontrar en dos formas, glicosilada y no glicosilada. Además, la alineación de lassecuencias aminoacídicas de las SLs caracterizadas demuestra que está altamenteconservada a lo largo de la filogenia y que posee unas características comunessignificativas (Fig.1).
Figura 1 Alineamiento de las secuencias aminoacídicas de las SLs caracterizadas (ver tabla 1)[ClustalW]. Los caracteres (-) se insertan para adaptar el alineamiento. La secuencia consenso común a
todas las SLs se marca en rojo, con los residuos de Cisteina y el sitio de N-glicosilación subrayados. Loscuatro dominios conservados de las SLs y los péptidos señal se marcan en azul.
hSL MNMMTVKQGVWA ALLWPYLLAASIPLD CKDEQGSFSACPSIS QEKLLDRVIQHAELI YRVSEESCSMFEEMF VPFPLRLQRNQAGYpoSL MNMMTVKQQGVWA ALLWPYLLTASIPLD CKEEQGSLSRCPSIS QEKLLDRVIQHAELI YRVSEESCSMFEEMF VPFPLRLQRNQAGY
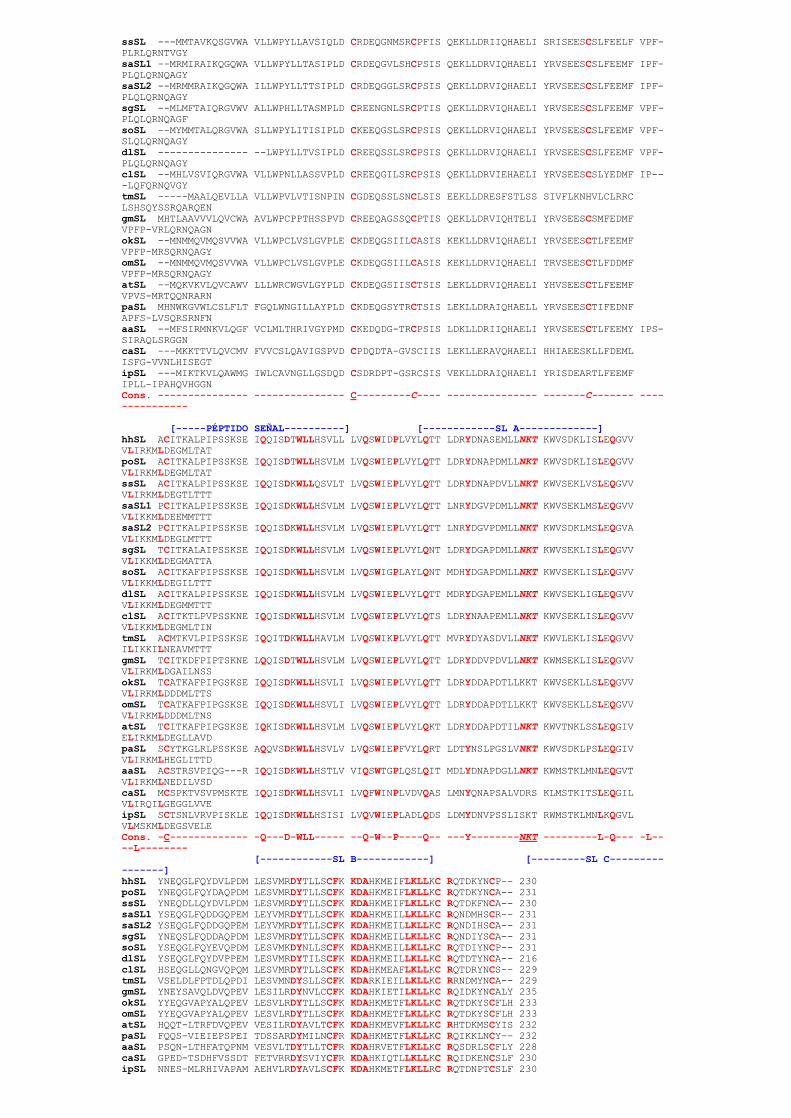
ssSL MMTAVKQSGVWA VLLWPYLLAVSIQLD CRDEQGNMSRCPFIS QEKLLDRIIQHAELI SRISEESCSLFEELF VPFPLRLQRNTVGYsaSL1 MRMIRAIKQGQWA VLLWPYLLTASIPLD CRDEQGVLSHCPSIS QEKLLDRVIQHAELI YRVSEESCSLFEEMF IPFPLQLQRNQAGYsaSL2 MRMMRAIKQGQWA ILLWPYLLTTSIPLD CRDEQGGLSRCPSIS QEKLLDRVIQHAELI YRVSEESCSLFEEMF IPFPLQLQRNQAGYsgSL MLMFTAIQRGVWV ALLWPHLLTASMPLD CREENGNLSRCPTIS QEKLLDRVIQHAELI YRVSEESCSLFEEMF VPFPLQLQRNQAGFsoSL MYMMTALQRGVWA SLLWPYLITISIPLD CKEEQGSLSRCPSIS QEKLLDRVIQHAELI YRVSEESCSLFEEMF VPFSLQLQRNQAGYdlSL LWPYLLTVSIPLD CREEQSSLSRCPSIS QEKLLDRVIQHAELI YRVSEESCSLFEEMF VPFPLQLQRNQAGYclSL MHLVSVIQRGVWA VLLWPNLLASSVPLD CREEQGILSRCPSIS QEKLLDRVIEHAELI YRVSEESCSLYEDMF IPLQFQRNQVGYtmSL MAALQEVLLA VLLWPVLVTISNPIN CGDEQSSLSNCLSIS EEKLLDRESFSTLSS SIVFLKNHVLCLRRCLSHSQYSSRQARQENgmSL MHTLAAVVVLQVCWA AVLWPCPPTHSSPVD CREEQAGSSQCPTIS QEKLLDRVIQHTELI YRVSEESCSMFEDMFVPFPVRLQRNQAGNokSL MNMMQVMQSVVWA VLLWPCLVSLGVPLE CKDEQGSIILCASIS KEKLLDRVIQHAELI YRVSEESCTLFEEMFVPFPMRSQRNQAGYomSL MNMMQVMQSVVWA VLLWPCLVSLGVPLE CKDEQGSIILCASIS KEKLLDRVIQHAELI TRVSEESCTLFDDMFVPFPMRSQRNQAGYatSL MQKVKVLQVCAWV LLLWRCWGVLGYPLD CKDEQGSIISCTSIS LEKLLDRVIQHAELI YHVSEESCTLFEEMFVPVSMRTQQNRARNpaSL MHNWKGVWLCSLFLT FGQLWNGILLAYPLD CKDEQGSYTRCTSIS LEKLLDRAIQHAELL YRVSEESCTIFEDNFAPFSLVSQRSRNFNaaSL MFSIRMNKVLQGF VCLMLTHRIVGYPMD CKEDQDGTRCPSIS LDKLLDRIIQHAELI YRVSEESCTLFEEMY IPSSIRAQLSRGGNcaSL MKKTTVLQVCMV FVVCSLQAVIGSPVD CPDQDTAGVSCIIS LEKLLERAVQHAELI HHIAEESKLLFDEMLISFGVVNLHISEGTipSL MIKTKVLQAWMG IWLCAVNGLLGSDQD CSDRDPTGSRCSIS VEKLLDRAIQHAELI YRISDEARTLFEEMFIPLLIPAHQVHGGNCons. CC C
[PÉPTIDO SEÑAL] [SL A] hhSL ACITKALPIPSSKSE IQQISDTWLLHSVLL LVQSWIDPLVYLQTT LDRYDNASEMLLNKT KWVSDKLISLEQGVVVLIRKMLDEGMLTATpoSL ACITKALPIPSSKSE IQQISDTWLLHSVLM LVQSWIEPLVYLQTT LDRYDNAPDMLLNKT KWVSDKLISLEQGVVVLIRKMLDEGMLTATssSL ACITKALPIPSSKSE IQQISDKWLLQSVLT LVQSWIEPLVYLQTT LDRYDNAPDVLLNKT KWVSEKLVSLEQGVVVLIRKMLDEGTLTTTsaSL1 PCITKALPIPSSKSE IQQISDKWLLHSVLM LVQSWIEPLVYLQTT LNRYDGVPDMLLNKT KWVSEKLMSLEQGVVVLIKKMLDEEMMTTTsaSL2 PCITKALPIPSSKSE IQQISDKWLLHSVLM LVQSWIEPLVYLQTT LNRYDGVPDMLLNKT KWVSDKLMSLEQGVAVLIKKMLDEGLMTTTsgSL TCITKALAIPSSKSE IQQISDKWLLHSVLM LVQSWIEPLVYLQNT LDRYDGAPDMLLNKT KWVSEKLISLEQGVVVLIKKMLDEGMATTAsoSL ACITKAFPIPSSKSE IQQISDKWLLHSVLM LVQSWIGPLAYLQNT MDHYDGAPDMLLNKT KWVSEKLISLEQGVVVLIKKMLDEGILTTTdlSL ACITKALPIPSSKSE IQQISDKWLLHSVLM LVQSWIEPLVYLQTT MDRYDGAPEMLLNKT KWVSEKLIGLEQGVVVLIKKMLDEGMMTTTclSL ACITKTLPVPSSKNE IQQISDKWLLHSVLM LVQSWIEPLVYLQTS LDRYNAAPEMLLNKT KWVSEKLISLEQGVVVLIKKMLDEGMLTINtmSL ACMTKVLPIPSSKSE IQQITDKWLLHAVLM LVQSWIKPLVYLQTT MVRYDYASDVLLNKT KWVLEKLISLEQGVVILIKKILNEAVMTTTgmSL TCITKDFPIPTSKNE LQQISDTWLLHSVLM LVQSWIEPLVYLQTT LDRYDDVPDVLLNKT KWMSEKLISLEQGVVVLIRKMLDGAILNSSokSL TCATKAFPIPGSKSE IQQISDKWLLHSVLI LVQSWIEPLVYLQTT LDRYDDAPDTLLKKT KWVSEKLLSLEQGVVVLIRKMLDDDMLTTSomSL TCATKAFPIPGSKSE IQQISDKWLLHSVLI LVQSWIEPLVYLQTT LDRYDDAPDTLLKKT KWVSEKLLSLEQGVVVLIRKMLDDDMLTNSatSL TCITKAFPIPGSKSE IQKISDKWLLHSVLM LVQSWIEPLVYLQKT LDRYDDAPDTILNKT KWVTNKLSSLEQGIVELIRKMLDEGLLAVDpaSL SCYTKGLRLPSSKSE AQQVSDKWLLHSVLV LVQSWIEPFVYLQRT LDTYNSLPGSLVNKT KWVSDKLPSLEQGIVVLIRKMLHEGLITTDaaSL ACSTRSVPIQGR IQQISDKWLLHSTLV VIQSWTGPLQSLQIT MDLYDNAPDGLLNKT KWMSTKLMNLEQGVTVLIRKMLNEDILVSDcaSL MCSPKTVSVPMSKTE IQQISDKWLLHSVLI LVQFWINPLVDVQAS LMNYQNAPSALVDRS KLMSTKITSLEQGILVLIRQILGEGGLVVEipSL SCTSNLVRVPISKLE IQQISDKWLLHSISI LVQVWIEPLADLQDS LDMYDNVPSSLISKT RWMSTKLMNLKQGVLVLMSKMLDEGSVELECons. C QDWLL QWPQ YNKT LQ LL [SL B] [SL C] hhSL YNEQGLFQYDVLPDM LESVMRDYTLLSCFK KDAHKMEIFLKLLKC RQTDKYNCP 230poSL YNEQGLFQYDAQPDM LESVMRDYTLLSCFK KDAHKMEIFLKLLKC RQTDKYNCA 231ssSL YNEQDLLQYDVLPDM LESVMRDYTLLSCFK KDAHKMEIFLKLLKC RQTDKFNCA 230saSL1 YSEQGLFQDDGQPEM LEYVMRDYTLLSCFK KDAHKMEILLKLLKC RQNDMHSCR 231saSL2 YSEQGLFQDDGQPEM LEYVMRDYTLLSCFK KDAHKMEILLKLLKC RQNDIHSCA 231sgSL YNEQSLFQDDAQPDM LESVMRDYTLLSCFK KDAHKMEILLKLLKC RQNDIYSCA 231soSL YSEQGLFQYEVQPDM LESVMKDYNLLSCFK KDAHKMEILLKLLKC RQTDIYNCP 231dlSL YSEQGLFQYDVPPEM LESVMRDYTILSCFK KDAHKMEILLKLLKC RQTDTYNCA 216clSL HSEQGLLQNGVQPQM LESVMRDYTLLSCFK KDAHKMEAFLKLLKC RQTDRYNCS 229tmSL VSELDLFPTDLQPDI LESVMNDYSLLSCFK KDARKIEILLKLLKC RRNDMYNCA 229gmSL YNEYSAVQLDVQPEV LESILRDYNVLCCFK KDAHKIETILKLLKC RQIDKYNCALY 235okSL YYEQGVAPYALQPEV LESVLRDYTLLSCFK KDAHKMETFLKLLKC RQTDKYSCFLH 233omSL YYEQGVAPYALQPEV LESVLRDYTLLSCFK KDAHKMETFLKLLKC RQTDKYSCFLH 233atSL HQQTLTRFDVQPEV VESILRDYAVLTCFK KDAHKMEVFLKLLKC RHTDKMSCYIS 232paSL FQQSVIEIEPSPEI TDSSARDYMILNCFR KDAHKMETFLKLLKC RQIKKLNCY 232aaSL PSQNLTHFATQPNM VESVLTDYTLLTCFR KDAHRVETFLKLLKC RQSDRLSCFLY 228caSL GPEDTSDHFVSSDT FETVRRDYSVIYCFR KDAHKIQTLLKLLKC RQIDKENCSLF 230ipSL NNESMLRHIVAPAM AEHVLRDYAVLSCFK KDAHKMETFLKLLRC RQTDNPTCSLF 230

Cons. DYCF KDALKLLC RC [SL D]
La SL ha sido implicada en una gran variedad de procesos fisiológicos, como lamaduración sexual o, en general, algunos aspectos de la reproducción (Planas et al.1992; Rand-Weaver et al. 1992, 1995; Rand-Weaver & Swanson 1993; Olivereau &Rand-Weaver 1994a, b; Mousa & Mousa 2000). Otros procesos en los que puedejugar un papel importante son: la respuesta al estres en la que se activan las célulassomatolactogénicas (Rand-Weaver et al. 1993; Johnson et al. 1997), la regulaciónácido-base y de los niveles de calcio (Olivereau et al. 1981; Wendelaar Bonga et al.1984, 1986; Kakizawa et al. 1993, 1995a, b; Kakizawa et al. 1996), el metabolismo delos ácidos grasos y del fosfato (Kaneko et al. 1993a; Lu et al. 1995; Company et al.2000) y la adaptación a los fondos oscuros en las especies no salmonidas (Van Eys1980; Olivereau et al. 1980; Ball & Batten, 1981; Rand-Weaver et al. 1993; Zhu &Thomas 1995, 1996, 1997, 1998; Kakizawa et al. 1995a; Zhu et al. 1999). En general,podemos considerar que la función principal que lleva a cabo la familia de hormonasGH/PRL/SL en los teleósteos es la regulación de diferentes iónes.
Recientemente, Cavari et al. (2000) han identificado, por primera vez en un pez, unasegunda forma de SL en la dorada (Sparus aurata, Linnaeus 1758). Esta última, sedenomina SL2 a diferencia de la SL, única forma caracterizada en las demás especiesde peces, o SL1 en el caso de S. aurata. La SL2 se diferencia solamente en onceaminoácidos de la forma SL1, clonada por Astola et al. (1995). De estos onceaminoácidos, siete son substituidos por otros aminoácidos semejantes en cuanto a supolaridad. Por lo tanto, dadas las semejanzas tan marcadas entre SL1 y SL2, la técnicade ISH nos ofrece un instrumento ideal para estudiar la distribución de estas proteínasen la hipófisis de la dorada.
Material y métodos
Preparación de la muestra
Hemos utilizado ejemplares juveniles (de 87, 188 y 346 días) de S. auratade tres tamaños diferentes para cada edad, provenientes de una mismafreza (febrero 2000) de la piscifactoria MARESA (Ayamonte, Huelva).La selección de los tres grupos, teniendo en cuenta el crecimientoasincrónico de esta especie en cultivo industrial, se efectúo por sucesivostamizados en determinados periodos de tiempo según las necesidades delcultivo industrial, estableciendo unas tallas y pesos para cada uno de elloscon valor estadístico significativo para su posterior análisis comparativo.Todas las soluciones empleadas en este trabajo fueron tratadas condietilpirocarbonato que previene la degradación del RNA. Al menos trespeces de cada grupo, previamente anestesiados, fueron decapitados y susencéfalos fijados por perfusión previa o por inmersión de las muestras,según el tamaño del animal, en una solución de paraformaldehido entampón fosfato salino pH 7,3 (PBS). Después de la fijación, las hipófisisfueron disecadas, se lavaron varias veces en tampón fosfato pH 7,3 (PB)y se crioprotegieron en sacarosa/PBS, antes de encastrarlas en OCT(Tissue-TekR). Secciones de criostato de 8-10 m, transversales ysagitales, se montaron en portaobjetos pretratados con poli-L-lisina(Menzel-Glaxe) y se almacenaron a -20 ºC hasta su uso. Todas losprocedimientos utilizados en la manipulación de los animales cumplen lanormativa de la Directiva de la Comisión Europea (86/609/EEC) y laLegislación Española (BOE 67/8509-12, 1998).
Diseño de oligonucleótidos

Como sondas utilizamos oligonucleótidos de 24, 30 y 31 nucleótidosmarcados en su extremo 5’ con digoxigenina (1) o biotina (2). El análisisde los alineamientos de secuencias nucleotídicas (datos no mostrados) yaminoacídicas (Fig. 2) de la familia de proteínas GH/PRL/SL de S.aurata, determinó un diseño de las oligosondas en las regiones proteícasalrededor de los dominios D de estas proteínas y en sus zonas notraducidas en los extremos 3’ de los cDNAs. Las oligosondas "anti-sentido" utilizadas en este estudio son: SaSL1a(1) y SaSL1c(2)
5’tcgacagctgtgcatgtcattttg3’ y SaSL1d(2)5’caggtttaattaggcgagttaagccactgc3’ complementarias al cDNA de SL1(673-696 nt y 1281-1311 nt respectivamente, Cavari et al. 1995);SaSL2a(2) 5’tgcacagctgtgtatgtcattctg3’ y SaSL2c(1) y SaSL2d(2)5’ggtggagagagggcaaagcattacacctac3’ complementarias al cDNA deSL2 (688-711 nt y 875-904 nt, Cavari et al. 2000). Mientrás, lasoligosondas "anti-sentido" diseñadas para la hormona de crecimiento(GH) y la prolactina (PRL) son SaGHa(1) y SaGHc(2),5’tctcagcgactcgtcggtgcccagactttg3’ complementarias al cDNA de GH(533-562 nt, Cavari et al. 1994; Funkenstein et al. 1991) y SaPRLa(2)5’ggaaatgttgtcctcgccgatgtcattggac3’ complementaria al cDNA de laPRL (534-564 nt, Santos et al. 1999). Como controles utilizamosoligosondas "sentido" de la misma secuencia que los cDNAs de S. auratade SL1 (SaSL1b (1)), SL2 (SaSL2b (2)), GH (SaGHb (1)) y PRL(SaPRLb(2)).
Figura 2. Alineamiento de las secuencias aminoacídicas de la familia de proteínas GH/PRL/SL deSparus aurata [ClustalW]. Las caracteres (-) se insertan para adaptar el alineamiento. La secuencia
consenso se marca en rojo y los residuos de Cisteina y el sitio de N-glicosilación se muestransubrayados. Los cuatro dominios conservados se marcan en azul y los péptido señal aparecen
subrayados en cada secuencia proteíca.SL1 MRMIRAIKQGQWAVL LWPYLLTASIPLDCR DEQGVLSHCPSISQE KLLDRVIQHAELIYR VSEESCSLFEEMFIPFPLQLQRNQAGYPC 89SL2 MRMMRAIKQGQWAIL LWPYLLTTSIPLDCR DEQGGLSRCPSISQE KLLDRVIQHAELIYR VSEESCSLFEEMFIPFPLQLQRNQAGYPC 89GH MDRVVLMLS VMSLGVSSQPITDGQ RLFSIAVSRVQHLHL LAQRLFSDFESSLQTEDQRQLNKIFLQDFC 69PRL MAHRETNGSKLFI TVLCMVAACSAVPIN DLLDRASQRSDMLHS LSTTLTKDLSNHVPPVGWTMMPRPPLC 70Consenso C C L C C [DOMINIO A]SL1 ITKALPIPSSKSEIQ QISDKWLLHSVLMLV QSWIEPLVYLQTTLN RYDGVPDMLLNKTK WVSEKLMSLEQGVVVLIKKMLDEEMMTTTY 178SL2 ITKALPIPSSKSEIQ QISDKWLLHSVLMLV QSWIEPLVYLQTTLN RYDGVPDMLLNKTK WVSDKLMSLEQGVAVLIKKMLDEGLMTTTY 178GH NSDYIISPIDKHETQ RSSVLKLLSISYRLV ESWEFPSRSLS GGSAPRNQ ISPKLSELKTGIHLLIRANEDGAEIFPDS 147PRL HTSSLQTPNDKEQAL QLSESDLMSLARSLL QAWQDPLVDLSNSAN SLLHPSQSSISNKIR ELQEHSKSLGDGLDILSGKMGPAAQAISSL 160ConsensoPK SLL WPL NKT L [DOMINIO B] [DOMINIO C]SL1 SEQGLFQDDGQPEML EYVMRDYTLLSC FKKDAHKMEILLKLL KCRQNDMHSCR 231SL2 SEQGLFQDDGQPEML EYVMRDYTLLSC FKKDAHKMEILLKLL KCRQNDIHSCA 231GH SALQLAPYGNYYQSL GTDESLRRTYELLAC FKKDMHKVETYLTVA KCRLSPEANCTL 204PRL PYRGSNDIGEDNISK LTNFHFLLSC FRRDSHKIDSFLKVL RCRAAKVQPEMC 212Consenso LLC FDHKL CRC [DOMINIO D]
Hibridación in situ (ISH).
Para la realización de las ISHs hemos empleado el método descrito porHerrero-Turrión et al. (2001) que brevemente a continuación se describe.Se ha comprobado que tratando el tejido previamente a la hibridación condiferentes agentes químicos mejoran los resultados, puesto que evitan lasseñales de fondo. Con este objetivo, realizamos una postfijación en 4 %paraformaldehido en PBS durante 10 minutos a temperatura ambiente (22

ºC), varios lavados en PBS y una acetilación en una solución de 1,5 %Trietanolamina (TEA), 150 mM NaCl y 0,25 % anhídrido acético durante10 minutos a temperatura ambiente. Tres lavados en PBS, deshidrataciónen una batería de etanoles y secado de las secciones preparan laprehibridación. Ésta, se realizó en una cámara húmeda durante dos horasa 45 ºC en una mezcla de hibridación (MH) compuesta de: 4X citratosódico salino (SSC: 600 mM NaCl, 50 mM citrato sódico pH 7,0), 10%sulfato dextrano, 1X Denhardt’s (50 X: 5 g ficoll type 400, 5 gpolivinilpirroldina y 5 g seroalbumina bovina (BSA) fracción V), 50 %formamida deionizada, 200 g/ml de DNA de esperma de salmóndesnaturalizado, 20 unit/ml de RNAt de levadura y PB. La estimación delas temperaturas de hibridación (Tm) para cada oligosonda sedeterminaron experimentalmente y son similares a las calculadasteóricamente (Heath et al. 1996). La hibridación se realizó durante toda lanoche a 45 ºC en cámara húmeda con la MH y la correspondienteoligosonda a una concentración de 15-20 ng/ml. Después, las seccionesfueron lavadas en condiciones de alta astringencia: un lavado en 0,1 %dodecil sulfato sódico (SDS), 2X SSC durante 30 minutos a temperaturaambiente y dos lavados en 0,1 % SDS, 0,1X SSC durante 30 minutos a45 ºC y a temperatura ambiente respectivamente. A continuación, lassecciones en las que hibridamos las oligosondas marcadas con ladigoxigenina se incubaron en una solución con 100 mM Tris-HCl, 150mM NaCl2, 1 % BSA y 2 % suero normal de oveja durante 1 hora atemperatura ambiente. Esta solución fue reemplazada por una idéntica ala que se adicionaron anticuerpos anti-digoxigenina conjugados confosfatasa alcalina (PA, Boehringer Mannheim) a una dilución 1:500durante al menos 90 minutos a temperatura ambiente. Por otra parte, lassecciones en las que hibridamos las oligosondas marcadas con biotina sepreincubaron en 200 mM PBS con Triton X100 al 0,2 % (PBS-Tx)durante 30 minutos a temperatura ambiente, se incubaron con la moléculaestreptavidina conjugada con peroxidasa (P, 1 g/ml, Jackson-ImmunoResearch) durante 3 horas a temperatura ambiente. Ambos tiposde secciones fueron lavados en PBS (3 X 5 min) e incluidas en lostampones de revelado. Para las que contienen PA, la mezcla de reveladoestá constituida por una solución de 100 mM Tris-HCl pH 9,5, 100 mMNaCl, 50 mM MgCl2 se le adicionaron 20 l/ml de la solución 18,75mg/ml NBT (4-nitro blue tetrazolium chloride) y 9,4 mg/ml BCIP (5-bromo-4-cloro-3-indol-fosfato) hasta la visualización óptima de losprecipitados azul oscuro. Por su parte, las que contienen P se incubaronen una mezcla de revelado compuesta por una solución de 200 mM Tris-HCl pH 7,5, 0,02 % de 3,3’-diaminobenzidina (DAB) y 0,003 % H2O2hasta la visualización nítida de los precipitados de color marrón. Lasreacciones colorimétricas se bloquearon con varios lavados en aguadestilada y las secciones fueron montadas en una solución de glicerol. Enalgunos casos, para mejorar la visualización de las señales, las seccionesse contratiñeron en hematoxilina férrica.
Se han realizado varios controles de las ISH: (1) secciones incubadas enla MH sin la oligosonda correspondiente, (2) secciones incubadas con laMH conteniendo el oligonucleótido "con sentido" correspondiente, (3)secciones hibridadas con las respectivos oligosondas "anti-sentido", perono incubadas con el anti-digoxigenina-PA o estreptavidina-P quecorrespondería en cada caso. Todos estos controles fueron negativos.
Doble Hibridación in situ.
Se llevaron a cabo dos tipos de marcajes utilizando esta técnica: (a) unahibridación con oligosonda marcada con digoxigenina, una posterior con

la correspondiente marcada con biotina, o viceversa, e incubacionesconsecutivas con anticuerpos anti-digoxigenina-PA y estreptavidina-P;(b) empleando dos oligosondas marcadas con biotina en indistinto orden,pero primero una hibridación con una de las oligosondas e incubacióncon estreptavidina-P y, posteriormente la segunda hibridación, con la otrasonda e incubación con estreptavidina-PA. En ambos casos, las seccionesfueron procesadas con DAB, analizadas, lavadas con PBS, procesadascon NBT/BCIP y otra vez analizadas.
Resultados y discusión
Hasta el momento la localización de células productoras de SL en diferentes especiesde peces se ha determinado fundamentalmente por estudios inmunohistoquímicos(IHC) (Rand-Weaver et al. 1991b; Kaneko et al. 1993b; Olivereau & Rand-Weaver1994a, b; Zhu & Thomas 1995; Dores et al. 1996; García-Hernández et al. 1996;Vissio et al. 1996, 1997; Rendón et al. 1997; Villaplana et al. 1997, 2000; Johnson etal. 1997; Saga et al. 1999; Mousa & Mousa 1999; Salam et al. 2000). De este modo,las células inmunoreactivas a SL (ir-SL) se han localizado principalmente en la PI dela hipófisis, bordeando el tejido neurohipofisario. Estas células son las quehistológicamente se definen como positivas al ácido periódico reactivo de Schiff, aexcepción de los salmónidos en los que se definen como cromofóbicas. La presenciade células ir-SL ha sido descrita en numerosas especies (Tabla 1), suponiéndose suuniversalidad en todos los teleósteos, aunque hay especies como Gymothoraxmeleagris en la que no se ha detectado mediante estudios IHC esta proteína (Dores etal. 1996).
Por consiguiente, dadas las limitaciones del método IHC y la reciente clonación en S.aurata de un segundo tipo de SL, con muy pocas diferencias respecto al primero(Cavari et al. 2000), el empleo de la ISH se hace necesario para analizar su patrón dedistribución. Para ello, en primer lugar optimizamos las condiciones de la ISH paraesta especie y la detección específica de sus mRNAs. La utilización de oligosondas noradioactivas, marcadas en su extremo 5’ con biotina o digoxigenina, se ha evidenciadocomo un método con alta especificidad y sensibilidad. La especificidad de lasoligosondas se ha demostrado al utilizar dos o tres oligonucleótidos que soncomplementarios al mismo transcrito de mRNA. Así, para el caso de las SLs,diseñamos oligosondas complementarias a los cDNAs, tanto en su zona traducidacomo en la no traducida. Las primeras, presentan un grado de similitud muy alto, del84 % (SaSL1a ó SaSL1c, y SaSL2a), y a pesar de ello, y en condiciones de altaastringencia, hemos demostrado que son específicas (Fig. 3). En este aspecto, otrosestudios han empleado como sondas oligonucleótidos con una similud más alta, del92 %, y demuestran que son altamente especificos en condiciones de alta astringencia(Sommer et al. 1990).
Figura 3. Secciones medio-sagitales de la hipófisis de animales juveniles de S. aurata de346 días. a: Señales de ISH de oligosonda para SL1 (SaSL1a). b: Señales de ISH de
oligosonda para SL1 (SaSL1d). Barra de escala, 200 mm. c: Señales de ISH de oligosondapara SL2 (SaSL2c). d: Señales de ISH de oligosonda para SL2 (SaSL2d). Barra de escalas,
100 mm. El asterisco (*) marca las células de SL presentes en la PPD.

Hasta ahora, el estudio de células que expresan la SL por ISH se describeexclusivamente en dos trabajos. En el primero de ellos (Kaneko et al. 1993c), estatécnica se utilizó para demostrar la expresión génica de la SL de la trucha arco iris enlas células que rodean al tejido neurohipofisario que penetra en la PI. Los mismosautores demostraron, en estudios ultrastructurales, que las moléculas de SL sonbiosintetizadas y almacenadas en los gránulos secretores de estas células. Por otraparte, Kakizawa et al. (1993), realizaron un estudio fisiológico en esta misma expecie,cuantificando los niveles de mRNA de la SL en ambientes hipocalcemicos ehipercalcemicos, determinando la posible función hipercalcémica de esta hormona.
Las dobles ISH de secciones hipofisarias de S. aurata han demostrado que ambostipos de SLs se coexpresan en las mismas células (somatolactogénicas), situándoseprincipalmente en la PI de la hipófisis, formando un cordón de células alrededor de laneurohipófisis (NH), como en otras especies de teleósteos (Rand-Weaver et al. 1991b;Kaneko et al. 1993c; Parhar & Iwata 1994; Zhu & Thomas 1995, 1996). Además, haycélulas SLs-ir aisladas o formando agrupaciones en porciones más internas de lapropia PI (Fig. 4). También, aunque con expresión más débil, se encuentran célulaspositivas a las SLs en S. aurata en grupos celulares aislados principalmente en la zonaventral de la proximal pars distalis (PPD) (Fig. 3d; Fig. 5a), como en distintasespecies del género Oncorhynchus y en Seriola dumerilii (Rand-Weaver et al. 1991b;Olivereau & Rand-Weaver 1994a; García-Hernández et al. 1996), lo que sugierediferentes actividades según el patrón de distribución dentro de la propia hipófisis. Laproximidad de las células somatolactogénicas al tejido neural, mostrando a menudoprocesos celulares que contactan con las ramas neurohipofisarias, sugiere que éstasestán reguladas por substancias neurohipotalámicas.
Figura 4. Sección transversal de la PI hipofisaria de animales juveniles de S. aurata de 188días. a: Señales de ISH para SL2. b: Doble ISH (SL2 y SL1) de la sección mostrada en a. c:
Detalle de a. d: Detalle de c. Barra de escalas, a y c: 100 mm, b y d. 50 mm.
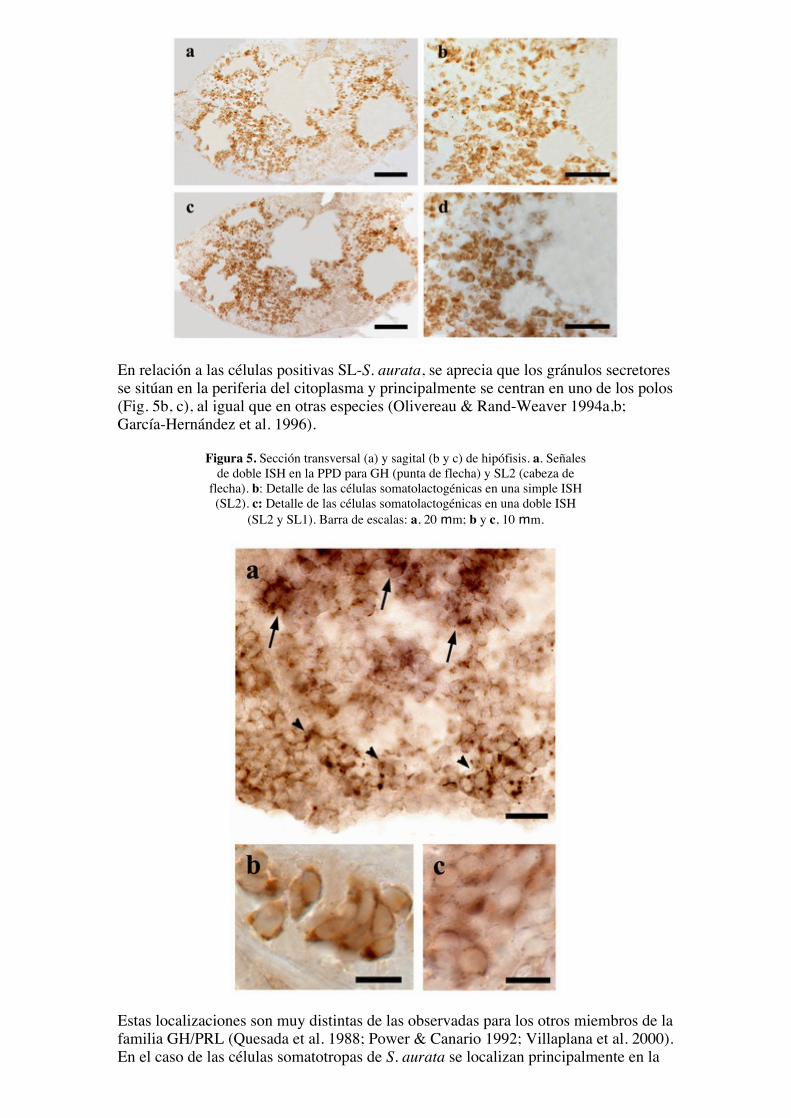
En relación a las células positivas SL-S. aurata, se aprecia que los gránulos secretoresse sitúan en la periferia del citoplasma y principalmente se centran en uno de los polos(Fig. 5b, c), al igual que en otras especies (Olivereau & Rand-Weaver 1994a,b;García-Hernández et al. 1996).
Figura 5. Sección transversal (a) y sagital (b y c) de hipófisis. a. Señalesde doble ISH en la PPD para GH (punta de flecha) y SL2 (cabeza de
flecha). b: Detalle de las células somatolactogénicas en una simple ISH(SL2). c: Detalle de las células somatolactogénicas en una doble ISH
(SL2 y SL1). Barra de escalas: a, 20 mm; b y c, 10 mm.
Estas localizaciones son muy distintas de las observadas para los otros miembros de lafamilia GH/PRL (Quesada et al. 1988; Power & Canario 1992; Villaplana et al. 2000).En el caso de las células somatotropas de S. aurata se localizan principalmente en la
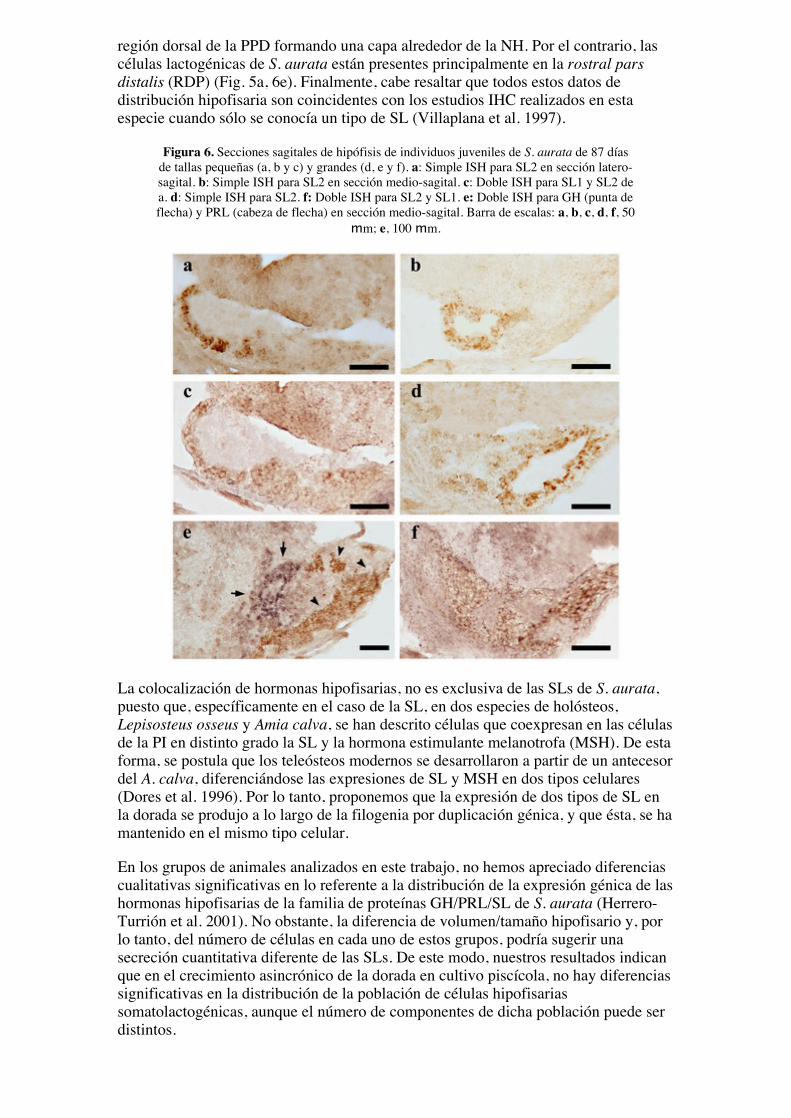
región dorsal de la PPD formando una capa alrededor de la NH. Por el contrario, lascélulas lactogénicas de S. aurata están presentes principalmente en la rostral parsdistalis (RDP) (Fig. 5a, 6e). Finalmente, cabe resaltar que todos estos datos dedistribución hipofisaria son coincidentes con los estudios IHC realizados en estaespecie cuando sólo se conocía un tipo de SL (Villaplana et al. 1997).
Figura 6. Secciones sagitales de hipófisis de individuos juveniles de S. aurata de 87 díasde tallas pequeñas (a, b y c) y grandes (d, e y f). a: Simple ISH para SL2 en sección latero-sagital. b: Simple ISH para SL2 en sección medio-sagital. c: Doble ISH para SL1 y SL2 dea. d: Simple ISH para SL2. f: Doble ISH para SL2 y SL1. e: Doble ISH para GH (punta deflecha) y PRL (cabeza de flecha) en sección medio-sagital. Barra de escalas: a, b, c, d, f, 50
mm; e, 100 mm.
La colocalización de hormonas hipofisarias, no es exclusiva de las SLs de S. aurata,puesto que, específicamente en el caso de la SL, en dos especies de holósteos,Lepisosteus osseus y Amia calva, se han descrito células que coexpresan en las célulasde la PI en distinto grado la SL y la hormona estimulante melanotrofa (MSH). De estaforma, se postula que los teleósteos modernos se desarrollaron a partir de un antecesordel A. calva, diferenciándose las expresiones de SL y MSH en dos tipos celulares(Dores et al. 1996). Por lo tanto, proponemos que la expresión de dos tipos de SL enla dorada se produjo a lo largo de la filogenia por duplicación génica, y que ésta, se hamantenido en el mismo tipo celular.
En los grupos de animales analizados en este trabajo, no hemos apreciado diferenciascualitativas significativas en lo referente a la distribución de la expresión génica de lashormonas hipofisarias de la familia de proteínas GH/PRL/SL de S. aurata (Herrero-Turrión et al. 2001). No obstante, la diferencia de volumen/tamaño hipofisario y, porlo tanto, del número de células en cada uno de estos grupos, podría sugerir unasecreción cuantitativa diferente de las SLs. De este modo, nuestros resultados indicanque en el crecimiento asincrónico de la dorada en cultivo piscícola, no hay diferenciassignificativas en la distribución de la población de células hipofisariassomatolactogénicas, aunque el número de componentes de dicha población puede serdistintos.

Conclusiones
1. Hemos optimizado las condiciones de la ISH, para la determinación de laexpresión génica de una familia de hormonas hipofisarias con grandessimilitudes estructurales.
2. Es la primera vez que se encuentra que los dos tipos de somatolactinas de ladorada se expresan en las mismas células, es decir, están colocalizadas.
3. No hay una diferencia apreciable en la distribución de célulassomatolactogénicas entre los animales de dorada, de crecimiento asincrónico,provenientes de una misma freza.
Agradecimientos
Este trabajo ha sido financiado por la Junta de Castilla y León y F.S.E. (Sa 30/99 y Sa103/01) y FEDER-CICYT (1FD97-1739).
Bibliografía
Amemiya, Y., Sogabe, Y., Nozaki, M., Takahashi, A. y Kawauchi, H.(1999). Somatolactin in the white sturgeon and African lungfish and itsevolutionary significance. Gen Comp Endocrinol114, 181-90.Astola, A., Pendon, C., Ortiz, M. and Valdivia, M. M. (1995). Cloning andexpression of somatolactin, a pituitary hormone related to growth hormone andprolactin from gilthead seabream, Sparus aurata. Gen Comp Endocrinol104(3), 330-6.Ayson, F. G., de Jesus, E. G., Amemiya, Y., Moriyama, S., Hirano, T. yKawauchi, H. (1999). Isolation and cDNA cloning of somatolactin inrabbitfish (Siganus guttatus). Gen Comp Endocrinol 115, 292-300.Ball, J. N. and Batten, T. F. (1981). Pituitary and melanophore responses tobackground in Poecilia latipinna (teleostei): Role of the pars intermedia PAScells. Gen Comp Endocrinol 44, 233-248.Cavari, B., Le Bail, P. Y., Levavi-Sivan, B., Melamed, P., Kawauchi, H. yFunkenstein, B. (1994). Isolation of the growth hormone and in vitrotranslation of the mRNA isolated from pituitaries of the gilthead seabreamSparus aurata. Gen Comp Endocrinol 95, 621-29.Cavari, B., Noso, T. y Kawauchi, H. (1995). Somatolactin, a novel pituitaryprotein: isolation and characterization from Sparus aurata. Mol Mar BiolBiotechnol. 4(2), 117-22.Cavari, B., Funkenstein, B. and Kawauchi, H. (2000). Cloning of a secondvariant of somatolactin-encoding cDNA from the gilthead seabream Sparusaurata. Fish Physiology and Biochemistry 22 (2), 145-150.Cheng, K. W., Chan, Y. H., Chen, Y. D., Yu, K. L. and Chan, K. M. (1997).Sequence of a cDNA clone encoding a novel somatolactin in goldfish,Carassius y. Biochem Biophys Res Commun. 232, 282-7.Company, R., Calduch-Giner, J. A., Mingarro, M. y Pérez-Sánchez, J.(2000). cDNA cloning and sequence of European sea bass (Dicentrarchuslabrax) somatolactin. Comparative Biochemistry and Physiology Part B 127,183-192.Dores, R. M., Hoffman, N. E., Chilcutt-Ruth, T., Lancha, A., Brown, C.,Marra, L. y Youson, J. (1996). A comparative analysis of somatolactin-ralatedimmunoreactivity in the pituitaries of four neopterygian fishes and onechondrostean fish: an immunohistochemical study. Gen Comp Endocrinol. 102,79-87.

Funkenstein, B., Chen, T. T., Powers, D. A. y Cavari, B. (1991). Cloning andsequencing of the gilthead seabream (Sparus aurata) growth hormone-encodingcDNA. Gene. 103, 243-7.García-Hernández, M. P., García-Ayala, A., Elbal, M. T. y Agulleiro, B.(1996). The adenohypophysis of Mediterranean yellowtail, Seriola dumerilii(Risso, 1810): an immunocytochemical study. Tissue Cell 28(5), 577-85.Heath, P. R., Sanders, M. W. y Pearson, R. C. A. (1996). In situhybridization wiyh 35S- labelled oligonucleotides. In In situ hybridizationtechniques for the brain, Z. Henderson, ed. (Univ. Oxford: Methods inNeurosciences).Herrero-Turrión, M.J.; Velasco, A.; Rodríguez, R.; Aijón, J. y Lara, J.(2001). Métodos no radioactivos de hibridación in situ, simple y doble, para GHy PRL de dorada. VIII Congreso Nacional de Acuicultura.Iraqi, F., Gong, Z. y Hew, C. L. (1993). Isolation and characterization ofsomatolactin genes from cold water marine teleost lumpfish (Cyclopteruslumpus) and habilut (Hippoglossus hippoglossus). Mol. Mar. Biol. Biotechnol.2, 96-103.Johnson, L. L., Norberg, B., Willis, M. L., Zebroski, H. and Swanson, P.(1997). Isolation, characterization, and radioimmunoassay of Atlantic halibutsomatolactin and plasma levels during stress and reproduction in flatfish. GenComp Endocrinol 105, 194-209.Kakizawa, S., Kaneko, T., Hasegawa, S. and Hirano, T. (1993). Activationof somatolactin cells in the pituitary of the rainbow trout Oncorhynchus mykissby low environmental calcium. Gen Comp Endocrinol. 91, 298-306.Kakizawa, S., Kaneko, T., Hasegawa, S. and Hirano, T. (1995a). Effects offeeding, fasting, background adaptation, acute stress, and exhaustive exerciseon the plasma somatolactin concentrations in rainbow trout. Gen CompEndocrinol. 98, 137-46.Kakizawa, S., Kaneko, T., Ogasawara, T. y Hirano, T. (1995b). Changes inplasma somatolactin levels during spawing migration of chum salmon. FishPhysiology Biochemistry 14, 93-101.Kakizawa, S., Kaneko, T. y Hirano, T. (1996). Elevation of plasmasomatolactin concentrations during acidosis in rainbow trout. J Exp Biology199, 1043-1051.Kaneko, T. y Hirano, T. (1993a). Role of prolactin and somatolactin incalcium regulation in fish. J Exp Biology 184, 31-45.Kaneko, T., Kakizawa, S. y Yada, T. (1993b). Pituitary of "cobalt" variant ofrainbow trout separated from the hypothalamus lack most pars intermedia andneurohypophysial tissue. Gen Comp Endocrinol. 92, 31-40.Kaneko, T., Kakizawa, S., Yada, T. y Hirano, T. (1993c). Gene expressionand intracellular localization of somatolactin in the pituitary of rainbow trout.Cell Tissue Res. 272, 11-16.Lu, M., Swanson, P. y Renfro, J. L. (1995). Effect of somatolactin and relatedhormones on transport by flounder renal tubule primary cultures. Am. J.Physiol. 268, R577-R582.May, D., Todd, C. M. y Rand-Weaver, M. (1997). cDNA cloning of eel(Anguilla anguilla) somatolactin. Gene 188, 63-7.Mousa, S. A. y Mousa, M. A. (1999). Immunocytochemical and histologicalstudies on the hypophyseal-gonadal system in the freshwater nile tilapia,Oreochromis niloticus (L.), during sexual maturation and spawning in differenthabitats. J Exp Zool 284, 343-54.Mousa, M. A. y Mousa, S. A. (2000). Implication of somatolactin in theregulation of sexual maturation and spawning of Mugil cephalus. J Exp Zool287, 62-73.Nicoll, C. S., Mayer, G. L., y Rusell S. M. (1986). Structural features ofprolactins and growth hormones that can be related to their biologicalproperties. Endocr. Rev. 7, 169-203.

Olivereau, M., Aimar, C. y Olivereau, J. M. (1980). Response of the teleostpituitary (goldfish, eel) to deionized water. Cell Tissue Res. 208, 389-404.Olivereau, M., Olivereau, J. M. y Aimar, C. (1981). Specific effect ofcalcium ions on the calcium sensivite cells of the pars intermedia in thegoldfish. Cell Tissue Res. 214, 23-31.Olivereau, M. y Rand-Weaver, M. (1994a). Immunocytochemical study ofthe somatolactin cells in the pituitary of Pacific salmon, Oncorhynchus nerca,and O. keta at some stages of the reproductive cycle. Gen Comp Endocrinol.93, 28-35.Olivereau, M. y Rand-Weaver, M. (1994b). Inmunoreactive somatolactincells in the pituitary young , migrating, spawing and spent chinook salmon,Oncorhynchus tshawytscha. Gen Comp Endocrinol. 13, 141-151.Ono, M., Takayama, Y., Rand-Weaver, M., Sakata, S., Yasunaga, T., Noso,T. y Kawauchi, H. (1990). cDNA cloning of somatolactin, a pituitary proteinrelated to growth hormone and prolactin. Proc Natl Acad Sci U S A. 87(11),4330-4.Ono, M., Harigai, T., Kaneko, T., Sato, Y., Ihara, S. y Kawauchi, H. (1994).Pit-1/GH factor-1 involvement in the gene expression of somatolactin. MolEndocrinol. 8, 109-15.Parhar, I. S. y Iwata, M. (1994). Gonadotropin releasing hormone (GnRH)neurons project to growth hormone and somatolactin cells in the steelhead trout.Histochemistry 102(3), 195-203.Pendón, C., Martinez-Barbera, J. P., Perez-Sanchez, J., Rodriguez, R. B.,Grenett, H. y Valdivia, M. M. (1994). Cloning of the sole (Solea senegalensis)growth hormone-encoding cDNA. Gene 145, 237-40.Planas, J. V., Swanson, P., Rand-Weaver, M. y Dickhoff, W. W. (1992).Somatolactin stimulates in vitro gonadal steroidogenesis in coho salmon,Oncorhynchus kisutch. Gen Comp Endocrinol. 87, 1-5.Power, D.M. and Canario, A.V. (1992). Immunocytochemistry ofsomatotrophs, gonadotrophs, prolactin and adrenocorticotropin cells in larvalsea bream (Sparus auratus) pituitaries. Cell Tissue Res. 269(2):341-346.Quesada, J., Lozano, M. T., Ortega, A. y Agulleiro, B. (1988).Immunocytochemical and ultrastructural characterization of the cell types in theadenohypophysis of Sparus aurata L. (teleost). Gen Comp Endocrinol 72(2),209-25.Rand-Weaver, M., Noso, T., Muramoto, K. y Kawauchi, H. (1991a).Isolation and characterization of somatolactin, a new protein related to growthhormone and prolactin from Atlantic cod (Gadus morhua) pituitary glands.Biochemistry 30(6), 1509-15.Rand-Weaver, M., Baker, B. J. y Kawauchi, H. (1991b). Cellularlocalization of somatolactin in the pars intermedia of some teleost fishes. CellTissue Res. 263, 207-215.Rand-Weaver, M., Swanson, P., Kawauchi, H. y Dickhoff, W. W. (1992).Somatolactin, a novel pituitary protein: purification and plasma levels duringreproductive maturation of coho salmon. J Endocrinol. 133(3), 393-403.Rand-Weaver, M., Pottinger, T. G. y Sumpter, J. P. (1993). Plasmasomatolactin concentrations in salmonid fish are elevated by stress. J.Endocrinol. 138, 509-515.Rand-Weaver, M. y Swanson, P. (1993). Plasma somatolactin levels in cohosalmon (Oncorhyncus kitsutch) during smoltification and sexual maturation.Fish Physiology Biochemistry 11, 175-182.Rand-Weaver, M., Pottinger, T. G. y Sumpter, J. P. (1995). Pronouncedseasonal rhythms in plasma somatolactin levels in rainbow trout. J Endocrinol146, 113-9.Rendón, C., Rodríguez-Gómez, F. J., Muñoz-Cueto, J. A., Piñuela, C. ySarasquete, C. (1997). An immunocytochemical study of pituitary cells of theSenegalese sole, Solea senegalensis (Kaup 1858). Histochem J. 29(11-12), 813-

22.Saga, T., Yamaki, K., Doi, Y. y Yoshizuka, M. (1999). Chronological studyof the appearance of adenohypophysial cells in the ayu (Plecoglossus altivelis).Anat Embryol 200(5), 469-75.Salam, M. A., Ando, H., Ban, M., Ueda, H. y Urano, A. (2000).Immunocytochemical analysis of pituitary cells in pre-spawing chum salmon.Zoological Science 17, 805-819.Santos, C. R., Brinca, L., Ingleton, P. M. y Power, D. M. (1999). Cloning,expression, and tissue localisation of prolactin in adult sea bream (Sparusaurata). Gen Comp Endocrinol 114(1), 57-66.Sommer, B., Keinanen, K., Verdoorn, T. A., Wisden, W., Burnashev, N.,Herb, A., Kohler, M., Takagi, T., Sakmann, B. y Seeburg, P. H. (1990). Flipand flop: a cell-specific functional switch in glutamate-operated channels of theCNS. Science 249, 1580-5.Takayama, Y., Ono, M., Rand-Weaver, M. y Kawauchi, H. (1991a). Greaterconservation of somatolactin a presumed pituitary hormone of the growthhormone/prolactin family than of growth hormone in teleost fish. Gen CompEndocrinol. 83, 366-374.Takayama, Y., Rand-Weaver, M., Kawauchi, H. y Ono, M. (1991b). Genestructure of chum salmon somatolactin, a presumed pituitary hormone of thegrowth hormone/prolactin family. Mol Endocrinol. 5, 778-86.Tang, Y. (1992). A study on the catfish (Ictalurus punctatus) growth hormonegene family: structures of growth hormone and prolactin genes andsomatolactin cDNA, their evolutionary implication and expression in thepituitary gland. Thesis. University of Maryland.Van Eys G. J. (1980). Structural changes in the pars intermedia of the cichlidteleost Sarotherodon mossambicus as a result of background adaptation andillumination. II. The PAS-positive cells. Cell and Tissue Res. 210, 171-9.Villaplana, M., García Ayala, A., García Hernández, M. P. y Agulleiro, B.(1997). Ontogeny of immunoreactive somatolactin cells in the pituitary ofgilthead sea bream (Sparus aurata L., Teleostei). Anat Embryol. 196(3), 227-34.Villaplana, M., García-Ayala, A., Chaves-Pozo, E. y Agulleiro, B. (2000).Identification of mammosomatotropes, growth hormone cells and prolactincells in the pituitary gland of the gilthead sea bream (Sparus aurata L.,Teleostei) using light immunocytochemical methods: an ontogenic study. Anat.Embryol. 202, 421-429.Vissio, P. G., Paz, D. A. y Maggese, C. (1996). The adenohypophysis of theswamp eel, Synbranchus marmoratus, an immunocytochemical analysis.Biocell 20, 155-61.Vissio, P. G., Somoza, G. M., Maggese, M. C., Paz, D. A. y Strussmann, C.A. (1997). Structure and cell type distribution in the pituitary gland of pejerreyOdontesthes bonariensis. Fisheries Science 63, 64-68.Wendelaar Bonga, S. E., Van der Meij, J. C., Van der Krabben, W. A. yFlik, G. (1984). The effect of water acidification on prolactin cells and parsintermedia PAS-positive cells in the teleost fish Oreochromis (formerlySarotherodon) mossambicus and Carassius auratus. Cell Tissue Res. 238, 601-9.Wendelaar Bonga, S. E., Van der Meij, J. C. y Flik, G. (1986). Response ofPAS-positive cells of the pituitary pars intermedia cells in the teleost Carassiusauratus to acid water. Cell Tissue Res. 243, 609-617.Yang, B. Y., Arab, M. y Chen, T. T. (1997). Cloning and characterization ofrainbow trout (Oncorhynchus mykiss) somatolactin cDNA and its expression inpituitary and nonpituitary tissue. Gen Comp Endocrinol. 106, 271-280.Zhu, Y. y Thomas, P. (1995). Red drum somatolactin: development of ahomologous radioimmunoassay and plasma levels after exposure to stressors orvarious backgrounds. Gen Comp Endocrinol 99, 275-288.

Zhu, Y. y Thomas, P. (1996). Elevations of somatolactin in plasma andpituitaries and increased alpha-MSH cell activity in red drum exposed to blackbackground and decreased illumination. Gen Comp Endocrinol 101, 21-31.Zhu, Y. y Thomas, P. (1997). Effects of somatolactin on melanosomeaggregation in the melanophores of red drum (Sciaenops ocellatus) scales. GenComp Endocrinol 105, 127-33.Zhu, Y. y Thomas, P. (1998). Effects of light on plasma somatolactin levels inred drum (Sciaenops ocellatus). Gen Comp Endocrinol 111, 76-82.Zhu, Y., Yoshiura, Y., Kikuchi, K., Aida, K. y Thomas, P. (1999). Cloningand phylogenetic relationship of red drum somatolactin cDNA and effects oflight on pituitary somatolactin mRNA expression. Gen Comp Endocrinol. 113,69-79.
Artículo publicado en la Revista AquaTIC nº 13 (especial VIII Congreso Nacional de Acuicultura), mayo 2001