la ecofisiologÍa en el Ámbito forestal - dialnet · guntarse sobre los controles que el ambiente...
TRANSCRIPT

LA ECOFISIOLOGÍA EN EL ÁMBITO FORESTAL
José A. Pardos Carrión
E. T. S. de Ingenieros de Montes. UPM. Ciudad Universitaria s/n. 28040-MADRID (España). Correoelectrónico: [email protected].
Resumen
Tras una alusión a la importancia de la Ecofisiología como interfase entre Fisiología Vegetal yEcología se muestran algunos resultados de trabajos en los que se evaluaron parámetros fisiológicosy anatómicos de varias especies arbóreas, con objeto de resaltar el papel que puede desempeñar laEcofisiología, esclareciendo cuestiones y aportando información al técnico en la gestión de los eco-sistemas forestales y la repoblación forestal. En el apartado “Uso del agua e intercambio gaseoso” secomparan diversos parámetros hídricos y del flujo de savia en hayas y robles en un rodal mixto,subrayándose sus diferencias. También se muestra el uso de la discriminación isotópica de carbonocomo herramienta en la caracterización del comportamiento y uso del agua en plantaciones de euca-lipto. Bajo el epígrafe “Cubierta arbórea y microclima” se contemplan las respuestas de brinzales dehaya plantados bajo el dosel de un fustal de repoblación de pino silvestre, resaltando la influenciaque la intensidad de una clara experimental ejecutada en el pinar tuvo sobre las hayas a través de lavariación provocada en la irradiancia que recibían y el agua de que disponían en términos de pará-metros hídricos y de intercambio gaseoso. En un tercer apartado titulado “Variación inter e intraes-pecífica” se subraya la influencia de la especie y de taxones de rango inferior sobre el crecimiento yparámetros fisiológicos en trabajos llevados a cabo en una plantación experimental de pinos carras-co y piñonero; y, en condiciones semicontroladas, de varias procedencias de pino rodeno y de clo-nes de eucalipto (E. globulus). La respuestas de parámetros foliares de varias especies de frondosasque comparten estación, y la del pino canario sometido a condiciones variables de disponibilidad deluz y agua, sirven de ejemplo en el apartado sobre “Desarrollo y plasticidad fenotípica”.
Palabras clave: Gestión forestal, Ecofisiología aplicada, Uso del agua, Respuestas fisiológicas
11ISSN: 1575-2410© 2005 Sociedad Española de Ciencias Forestales
Cuad. Soc. Esp. Cienc. For. 20: 11-23 (2004) «Actas de la I Reunión sobre Ecología, Ecofisiología y Suelos Forestales»
INTRODUCCIÓN
La actividad forestal dedica buena parte desus miras y esfuerzos a la conservación y gestiónde los montes, cuyos diversos usos se ejercitan enun marco ecológico, que los condiciona y debepresidirlos, y en el que los vegetales son actoresprincipales. En su actuación, las plantas interac-cionan con el ambiente y el estudio de esa inter-acción constituye el ámbito de la Ecofisiología.Creo, por tanto, acertado la incorporación de tal
disciplina, con nombre propio, en el grupo prime-ramente llamado de Ecología y Suelos Forestalesde la SECF, para constituir el grupo "Ecología,Ecofisiología y Suelos Forestales".
Los procesos vitales, los mecanismos enellos implicados, la postulación de las funcionesque permiten hacer predicciones de los fenóme-nos vitales, constituyen objeto de la FisiologíaVegetal. Esos mecanismos fisiológicos se dan enun ambiente determinado y la Ecofisiología sesitúa en la interfase Fisiología-Ecología para pre-

guntarse sobre los controles que el ambiente ejer-ce sobre el crecimiento, la supervivencia, lareproducción, la competencia, abundancia y dis-tribución geográfica, para ayudarnos a entenderla significación funcional y la evolución de loscaracteres de las plantas. La Ecología "plantealas preguntas y la Fisiología suministra las herra-mientas para su contestación". Se trata, por tanto,de una ciencia experimental que aplica metodo-logías comunes con la Fisiología, aunque requie-re también la participación de otras propias de laBioquímica, Biología Molecular y Biofísica, alas que hay que añadir los aspectos aplicados, ennuestro caso vinculados al ámbito forestal y alconjunto de materias integradas en las CienciasForestales y disciplinas coadyuvantes.
Las interacciones con los factores bióticos yabióticos pueden considerarse filtros que deter-minan la presencia de una especie en una esta-ción determinada, que se unen a los filtros his-tóricos que han permitido su llegada, tras lacual se reduce la presencia inicial, que sólomantienen aquellas especies cuya eficacia bio-lógica les permite llevar a cabo con éxito lasfunciones vitales (LAMBERS et al., 1998). Y ello,con cambios en el clima y en el suelo, que seunen a los derivados de las acciones antropogé-nicas, las cuales a su vez contribuyen a la alte-ración de los primeros.
En el ámbito forestal, la gestión selvícola delas masas arbóreas implica acciones técnicas encondiciones de habitación no siempre óptimaspara las plantas. Ello conduce a estrés conreducción de la tasa de los procesos fisiológicos,lo que se traduce en menor crecimiento y otrasalteraciones del desarrollo. La evitación y latolerancia constituyen formas de respuesta antesituaciones de estrés y los términos de endureci-miento, aclimatación y adaptación se usan, condiferente escala temporal y espacial, también dematices en su aplicación, para designar respues-tas al estrés (LEVITT, 1980). A ellos se une elconcepto de plasticidad fenotípica que amplía ytrasciende al de interacción genotipo x ambiente(SULTAN, 2000).
Ante todas estas situaciones, se producen enlas plantas cambios complejos que exigen la ela-boración y aplicación de modelos ecológicosque permitan predecir comportamientos de lasplantas, valorar costes y beneficios (los trade-
offs) para las mismas y dirigir actuaciones de losgestores forestales (DIXON et al., 1990).
En lo que sigue se muestra un abanico deresultados, en su mayor parte ya publicados,todavía lejos de la configuración de modeloalguno, pero que revisten interés porque cubrenalgunos aspectos ecofisiológicos de especiesarbóreas de interés forestal en España y paracuya obtención se han puesto en juego técnicasde uso común en Fisiología Vegetal. Con ello sepretende resaltar el papel de la Ecofisiología enel ámbito de la SECF y poner de manifiesto elinterés de sus aportaciones para el técnico fores-tal español, las cuales pueden ser de gran utilidaden la toma de decisiones inherentes a la gestiónde las masas arboladas y a la planificación de larepoblación de terrenos con vocación forestal. Sedistinguen cuatro apartados: “Uso del agua eintercambio gaseoso”, “Cubierta arbórea ymicroclima”, “Variación inter e intraespecífica”y “Desarrollo y plasticidad fenotípica”.
USO DEL AGUA E INTERCAMBIOGASEOSO
La existencia de una vía común para la entra-da y salida de gases y vapor de agua por difusiónestomática enfrenta a las plantas, especialmenteen condiciones de estrés hídrico, con el dilema deoptimizar la nutrición primaria de carbono con lamenor pérdida de agua por transpiración. De ahíla interconexión entre las variables y parámetrosimplicados en las relaciones hídricas (potencialhídrico y sus componentes, déficit de la presiónde vapor, conductancia estomática al vapor deagua, conductividad hidráulica, señales hormo-nales del status hídrico del suelo) y los relaciona-dos con la absorción y fijación de carbono (irra-diancia, conductancia estomática, concentracióninterna, bioquímica de la fijación), factores todosque inciden directa o indirectamente en la efi-ciencia en el uso del agua y del carbono en laplanta (LANDSBERG & GOWER, 1997).
En un rodal mixto de haya y roble en elHayedo de Montejo, si bien ambas especiesmostraron un patrón similar en la evolucióndiurna del potencial hídrico, en el haya a mediamañana se producía un descenso más acusado ymantenido por más tiempo (Figura 1, izda.).
12
JOSÉ A. PARDOS CARRIÓN «La Ecofisiología en el ámbito forestal»

Valores más altos del potencial al amanecer enroble al final del estío, con bajo contenido enagua del suelo, reflejan una mejor recuperacióna la alta demanda evaporativa y limitación esti-val de agua, que evidencia una menor sensibili-dad a la sequía que el haya. Ambas especiesmostraron también diferencias en la conductan-cia estomática al vapor de agua (Figura 1, dcha.)y en la tasa de fotosíntesis (no mostrado) amediodía con valores mayores en roble, en todaslas fechas de medición.
En el mismo rodal mixto se puso de manifies-to el diferente comportamiento de haya y roble enla evolución diaria de la velocidad de la saviamedida por el método de flujo de calor constante(GRANIER, 1994), algunos de cuyos valores semuestran en la figura 2. El flujo de savia se redu-jo al disminuir la eficiencia hidráulica.
En tres plantaciones experimentales deEucalyptus globulus, con árboles de cinco asiete años de edad, pertenecientes a dos familiasde dos procedencias y varios clones selectospertenecientes a ENCE en Huelva (LasHerrumbres, Valdeoscuro y San Sebastián), se
evaluó la supervivencia y el crecimiento en diá-metro y se correlacionó con los valores de dis-criminación isotópica del carbono (∆13C) medi-dos en muestras tomadas del tronco a la alturanormal (SUN et al., 1996). Se encontró una altacorrelación positiva entre el valor de ∆ y lasupervivencia (Figura 3, izda.), con los valoresmás bajos en la procedencia Geeveston (9.226)de lugar de origen mas húmedo, menos adapta-da al clima mediterráneo y más eficiente entranspiración. También el crecimiento en diáme-tro guardaba buenas correlaciones con ∆ en lastres estaciones (Figura 3,dcha). La disponibili-dad de agua en las Herrumbres fue menor debi-do a la existencia de una capa impermeable dearcilla no presente en las otras dos estaciones.Asimismo, el diferente sistema radical de losclones (más superficial) frente a las plantas desemilla podría relacionarse con las diferenciasen la eficiencia transpiratoria estimada a travésde ∆. Estos resultados sugieren un fuerte efectode la disponibilidad de agua en los valores de ∆que resultó un buen indicador del agua disponi-ble por la planta.
13
Cuad. Soc. Esp. Cienc. For. 20: 11-23 (2005) «Actas de la I Reunión sobre Ecología, Ecofisiología y Suelos Forestales»
Figura 1. Patrones diarios de potencial hídrico (izda) y conductancia estomática al vapor de agua (dcha) en haya yroble albar a lo largo del periodo vegetativo (tomado de ARANDA et al., 2000)
-3
-2
-1
0 6:00 9:00 12:00 15:00 18:00 21:00
- 3
- 2
- 1
0 6:00 9:00 12:00 15:00 18:00 21:00
6:00 9:00 12:00 15:00 18:00 21:00
-3
-2
-1
0 6:00 9:00 12:00 15:00 18:00 21:00
- 3
- 2
- 1
0 6:00 9:00 12:00 15:00 18:00 21:00
-3
-2
-1
0 6:00 9:00 12:00 15:00 18:00 21:00
- 3
- 2
- 1
0 6:00 9:00 12:00 15:00 18:00 21:00
16 June 1994 16 June 1994
3 August 1994 3 August 1994
7 September 1994 7 September 1994
F.sylvatica
F.sylvatica
F.sylvatica Q. petraea
Q. petraea
Q. petraea
Pote
ncia
l hí
dric
o (M
Pa)
5
4
3
2
1
4
3
2
1
0
06:00 9:00 12:00 15:00 18:00 21:00
6:00 9:00 12:00 15:00 18:00 21:00 6:00 9:00 12:00 15:00 18:00 21:00
6:00 9:00 12:00 15:00 18:00 21:00 6:00 9:00 12:00 15:00 18:00 21:00
5
4
3
2
1
0
6
5
4
3
2
1
0
6
5
4
3
2
1
0
6
5
4
3
2
1
0
F. sylvati ca
F. sylvati ca
F. sylvati ca
Q.petraea
Q.petraea
Q.petraea
15 June 1994
2 August 1994 2 August 1994
2 August 1994
6 September 19946 September 1994
Conductancia

CUBIERTA ARBÓREA Y MICROCLIMA:PLANTACIÓN DE HAYAS EN UN PINAR
En la interacción medio-plantas, los rodalesarbóreos ejercen una marcada influencia sobreel microclima creándose un sistema interactivoen el que cualquier cambio en alguno de suscomponentes conduce a un ajuste en el resto. Lacubierta arbórea, receptora de la radiación solarreduce la cantidad de radiación que transmite asu través y modifica parcialmente su espectro, elPAR es menor bajo cubierta, más acusadamenteen frondosas y más rico en longitudes de ondadel rojo lejano, lo cual va a condicionar la pre-sencia y, en su caso, desarrollo de las especiessusceptibles de germinar bajo la misma. El doselarbóreo modifica también el régimen térmicorespecto al existente en suelos desnudos, con unaumento de temperatura en invierno y un des-censo en verano, circunstancias que afectaránasimismo a la regeneración de la masa arbórea ya la presencia y competencia con especies acce-sorias (AUSSENAC, 2001). El dosel arbóreoactuará asimismo como pantalla interceptora delas precipitaciones, su densidad será factorinfluyente en la evaporación potencial; la arqui-tectura de la copa, la morfología y anatomía delas hojas afectarán no sólo el status hídrico delarbolado y parámetros fisiológicos relacionadoscon el mismo, sino también condicionarán ladisponibilidad de agua para el regenerado; ytodo ello implicará adaptaciones fisiológicas alas modificaciones microclimáticas.
La eliminación parcial del arbolado median-te clareos, claras y entresacas, deberá por tantopracticarse sobre bases funcionales teniendo pre-
sente el comportamiento de los árboles y los con-dicionamientos ambientales. Estos tratamientosno sólo aumentan los niveles de radiación, cuali-tativa y cuantitativamente, lo que tiene un efectodirecto sobre la fotosíntesis, sino que mejoran ladisponibilidad de agua y reducen el estrés hídri-co en el estío en los árboles que se dejan. Todoello, unido a la más rápida mineralización delhumus, liberándose nitrógeno, derivada de lamayor iluminación y disponibilidad de agua,conduce al aumento de la tasa de crecimiento endiámetro y aumento de la biomasa leñosa produ-cida (KOZLOWSKY et al., 1991). Todo lo cual ade-más de repercutir en el comportamiento del posi-ble regenerado de la especie principal, revisteimportancia en las masas mixtas dada la diferen-te capacidad de desarrollo de las especies partici-pantes y la competencia por los recursos deriva-da de la misma; y tiene, si cabe, aun mayor tras-cendencia si el objetivo pretendido es introducirotra especie más noble bajo el dosel arbóreo deuna conífera y acabar formando una masa mixta.
Bajo esta perspectiva, a continuación semuestran algunos de los resultados obtenidos enuna plantación de brinzales de haya realizada enla proximidad del Hayedo de Montejo bajo eldosel de un pinar adulto, de repoblación de Pinussylvestris, bajo diferentes condiciones de irradia-ción tras ser sometido a una clara sistemática, enrelación con mediciones de crecimiento, paráme-tros hídricos y de intercambio gaseoso, con el finde analizar el grado de influencia de su intensidaden la supervivencia y desarrollo de los brinzales.
Las hayas se plantaron a 2.5 m x 2.5 m en unpinar de 45 años con 55 m2 ha-1 de área basimé-trica y a los cuatro años se eliminaron filas de
14
JOSÉ A. PARDOS CARRIÓN «La Ecofisiología en el ámbito forestal»
Figura 2. Evolución diaria del consumo de agua referido a todo el árbol, un día de estío, en tres hayas y tres robles,con diferente grado de codominancia- dominancia, (dcha: Fs, hayas, Qpe, robles) y relación flujo de savia y conduc-tividad hidráulica (tomado de ARANDA et al. 2004)
0 . 0
1 . 0
2 . 0
3 . 0
6:00 9:00 12:00 15:00 18:00 21:00 0:00
Qpe1 Qpe2 Qpe3
0 . 0
1 . 0
2 . 0
3 . 0
6:00 9:00 12:00 15:00 18:00 21:00 0:00
Qpe1 Qpe2
0 . 0
1 . 0
2 . 0
3 . 0
6:00 9:00 12:00 15:00 18:00 21:00 0:00
Fs1 Fs2 Fs3
0 . 0
1 . 0
2 . 0
3 . 0
6:00 9:00 12:00 15:00 18:00 21:00 0:00
Fs1 Fs2
0 . 0
1 . 0
2 . 0
3 . 0
6:00 9:00 12:00 15:00 18: 00 21:00 0:00
Fs1 Fs2 Fs3
0.0
0.5
1.0
1.5
2.0
2.5
0.0 0.5 1.0 1.5
Beech Sessile oak
Fdmed = 1.83 Khsf R2 = 0.85
Sessile oak (spring)
Fdm
id (
dm-3
dm
-2 h
-1)
Khsf (dm -3 dm-2 h-1 MPa -1)

pinos manteniendo un 50% y dejando cuatrosituaciones de espaciamiento (Figura 4) (véaseARANDA et al., 2001). Se realizaron fotografíashemisféricas del dosel de pinos y con un softwa-re Hemiview se calcularon los factores de luzdirecta, difusa y global. Se llevaron a cabomediciones morfológicas y anatómicas durantedos años, encontrándose relaciones significati-vas con el factor de iluminación global, cuyoaumento condujo al de la masa foliar específica.El primer año, el aumento en esta estuvo másrelacionado con el grosor de la lámina foliar(Figura 5) mientras que en el segundo la signifi-cación se expresó con el del parénquima enempalizada (no mostrado); asimismo la mayorradiación produjo más densidad estomática. Losresultados muestran la capacidad de los brinza-les de haya de aclimatarse a la condición demayor iluminación provocada por la aberturadel dosel del pinar que se manifiestan en cam-bios en la anatomía de la hoja.
La clara realizada afectó también a la dispo-nibilidad de agua para las hayas plantadas y, enconsecuencia, a los parámetros hídricos. En lascuatro situaciones planteadas, descendió elpotencial hídrico al amanecer a lo largo delperiodo vegetativo en los dos años de medición.La clara más intensa practicada en el pinar (T2)condujo a los valores mayores de potencialhídrico de base (Tabla 1), mientras que los valo-res más altos del potencial osmótico a plena tur-gencia (Figura 6), se obtuvieron en las hayasbajo pinar no aclarado, con una clara relación
negativa entre dicho potencial y la concentra-ción de osmolitos (datos no mostrados). El ajus-te osmótico, como mecanismo de tolerancia a lasequía, fue limitado bajo condiciones de sombra(pinar no aclarado), restringiéndose la aclimata-ción de las hayas a la sequía en tal situación,pudiendo incluso ponerse en peligro la supervi-vencia de las plantas en años extremadamentesecos. Resulta evidente la compleja interacciónentre sequía y sombra sobre el potencial de acli-matación de las hayas y se pone de manifiestoque, si bien la presencia de un dosel protector esnecesario en los primeros años, transcurridadicha etapa la reducción del dosel significamenor intercepción de la lluvia y menor consu-mo de agua, lo que favorece, especialmente enverano, la disponibilidad de recursos a las hayas.
La clara condujo a la postre a un mayor cre-cimiento en altura y diámetro de las hayas(Figura 7), en paralelo con un aumento de lafotosíntesis neta a saturación de luz (Asat) y unmáximo en la conductancia estomática al vaporde agua (Figura 8). Asimismo la masa especifi-ca foliar y el contenido en nitrógeno referido amasa aumentaron en el rodal aclarado con unarelación positiva entre (Asat) y el contenido ennitrógeno referido a superficie foliar (datos nomostrados). La ausencia de significación para lalimitación no estomática de A se confirmó por lafalta de descenso de Fv/Fm, consecuencia de lafotoinhibición tras la abertura del dosel depinos; y en el rodal aclarado se encontró unvalor mayor de la eficiencia cuántica máxima de
15
Cuad. Soc. Esp. Cienc. For. 20: 11-23 (2005) «Actas de la I Reunión sobre Ecología, Ecofisiología y Suelos Forestales»
Figura 3. Relación entre discriminación isotópica de carbono y supervivencia en Valdeoscuro (izda) y diámetro nor-mal en Las Herrumbres (dcha) en Jeerland, 2, familia 84 y Geeveston, 9, familia 226 y varios clones seleccionados(tomado de PITA et al., 2001)
Las Herrumbres
y = 0,0624x + 16,151
R2 = 0,6996
16,4
16,6
16,8
17
17,2
17,4
17,6
10 15 20 25
diámetro normal cc (cm)
brinzales
413.7
334.1
115.3
115.2
Valdeoscuro
y = 0,0144x + 16,527R2 = 0,9512
16,9
17
17,1
17,2
17,3
17,4
17,5
17,6
17,7
0 20 40 60 80 100
supervivencia(%)
(per
mil)
2.849.226334.1
115.2
∆

los centros de PSII abiertos (Fv/Fm). Estosresultados (no mostrados, véase ARANDA et al,2004) subrayan la eficacia de reducir la cubierta(COLLET et al., 2001) y evidencian que la reduc-ción de la competencia por luz y agua favorecelos parámetros de intercambio gaseoso y lascondiciones de crecimiento de las hayas, lo quepermitirá la transformación con éxito del pinaren una masa mixta de frondosa y conífera.
VARIACIÓN INTER E INTRAESPECÍFICA
La información obtenida en los ensayos tra-dicionales en campo, en términos de supervi-vencia y crecimiento, a nivel de especie, proce-dencia y, en ocasiones, familia y genotipo, aun-que muy útil para la valoración y posterior elec-ción del material de uso en repoblación forestal,requiere esperas estimadas en un tercio a lamitad del turno de la especie. Con el fin de acor-tar dicho tiempo y predecir un comportamientofuturo, aun con las siempre problemáticas corre-laciones genotipo x edad, se ponen en juegoparámetros morfológicos, nutricionales, hídri-cos y de intercambio gaseoso entre otros, medi-dos en plántulas de corta edad que se someten aunas condiciones que simulan alguno de los fac-tores ecológicos a que van a estar sometidas en
campo y que pueden servir para vaticinar com-portamientos futuros.
Se presentan resultados de las respuestas devarias especies a diferentes situaciones ambien-tales, en términos de crecimiento y parámetrosfisiológicos en pino carrasco y pino piñonero enuna misma plantación; en seis procedencias dePinus pinaster bajo diferentes regímenes de dis-ponibilidad de agua en condiciones parcialmentecontroladas; finalmente, la variación entre clonesde eucalipto en situación de limitación de agua.
La forestación de tierras marginales deantiguo uso agrícola plantea la necesidad deconocer bajo una perspectiva ecofisiológica lacapacidad de adecuación de las especies utiliza-bles. Con tal fin, en Arganda (Madrid) en unterreno de cultivo agrícola de secano, bajo unclima mesomediterráneo seco, con un suelo ran-ker aluvial, de textura franca, ligeramente bási-co, con escasa materia orgánica y contenidoaceptable en nutrientes para coníferas, se instalóuna plantación experimental de Pinus hale-pensis y P. pinea. El mayor crecimiento del pinocarrasco se justifica por su mayor eficacia enprimavera, con tasas de fotosíntesis y transpira-ción mas altas, que propiciaron un tercer ciclode crecimiento al final de esta estación, frente ala existencia de un solo ciclo anual en el piñone-ro; y, si bien ambas especies se adaptaron bien a
16
JOSÉ A. PARDOS CARRIÓN «La Ecofisiología en el ámbito forestal»
Tabla 1. Potencial hídrico de base en hayas (C, control;T, tratamiento con alta irradiancia)
Figura 4. Distribución de la plantación de hayas (toma-do de ARANDA et al., 2001)
Figura 5. Grosor de la lámina foliar
7
8
9
10
11
12
13
a
ab
b
a
a
b b1998 1999
C T1 T2 T3 C T1 T2 T3
hayas
pinos seedlingC
C
E
S W
NT1T1
T2
T2
T3
T3
1998 1999Tratamiento 10 jun 29 jul 2 sept 11 junio 29 julio 9 sept
C -0.16 ± 0.02 a -0.79 ± 0.05 b -1.51 ± 0.16 a -0.21 ± 0.03 a -1.38 ± 0.28 b -1.27 ± 0.21 bT2 -0.14 ± 0.02 a -0.50 ± 0.09 a -1.29 ± 0.17 a -0.17 ± 0.03 a -0.46 ± 0.06 a -0.37 ± 0.04 a

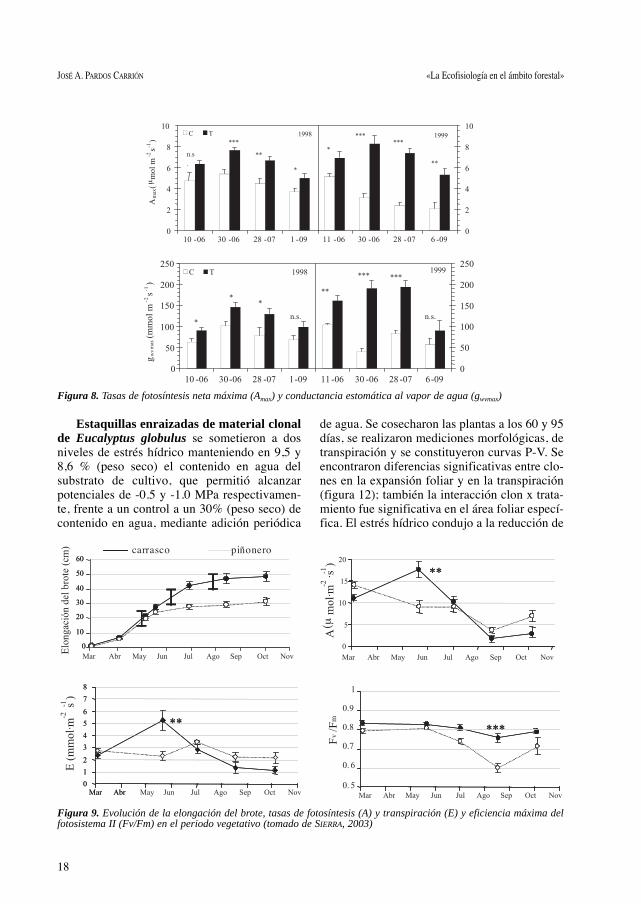
la sequía estival, lo hizo con más éxito el pinocarrasco. En la figura 9 se muestran la evolucióndel crecimiento, las tasas de fotosíntesis y trans-piración y la eficiencia máxima del fotosistemaII (Fv/Fm), cuyas diferencias justifican unaparte del diferente temperamento de ambasespecies y ratifican la necesidad de una adecua-da elección de especie en función de los condi-cionantes ecológicos del lugar de plantación.
Las diferencias intraespecíficas, a nivel deprocedencias, pueden ser de igual importanciaque las interespecíficas. Varias procedenciasibéricas y de Landas (Francia) de Pinuspinaster se sometieren a riego a capacidad decampo, cada una o dos semanas, poniéndose enjuego variables y parámetros de crecimiento,hídricos y de intercambio gaseoso. Cabe subra-yar que las diferencias más acusadas entre pro-cedencias se encontraron bajo limitación deagua (R2) y en pleno verano, como se muestraen la figura para el potencial osmótico a turgen-cia plena, tiempo en que descendió en algunasprocedencias (Oria y Arenas) 0.4 MPa (figura10), en contraste con las plantas con buen sumi-nistro de agua, en las que no se produjo ajusteosmótico alguno. El descenso en los potenciales
osmóticos a plena turgencia y pérdida de turgen-cia al alcanzar el pleno estío, así como el delmódulo de elasticidad máximo y el de la rela-ción peso turgente a peso seco, parámetros deri-vados de las curvas P-V, se produjeron en para-lelo al descenso del potencial osmótico(FERNÁNDEZ et al., 1999).
En un experimento con las mismas proce-dencias y condiciones experimentales se encon-traron diferencias en parámetros de intercambiogaseoso entre regímenes de agua. Las tasas deintercambio gaseoso en respuesta al potencialhídrico sólo descendieron en conjunto parapotenciales inferiores a -13 MPa, como se mues-tra en la figura 11 para la fotosíntesis, sin dife-rencias apreciables entre procedencias. Las pro-cedencias Ibéricas, en contraste con Landas,mostraron tendencia a economizar agua al finalde la primavera evidenciando una adaptación alestío seco de sus localidades de origen. Las dife-rencias de crecimiento entre procedencias no seexpresaron en los valores de la tasa de fotosínte-sis, dependiente de otros factores no controladosy podrían explicarse por diferencias en el usodel agua y diferente productividad de nitrógeno(datos no mostrados).
17
Cuad. Soc. Esp. Cienc. For. 20: 11-23 (2005) «Actas de la I Reunión sobre Ecología, Ecofisiología y Suelos Forestales»
Figura 6. Variación del potencial osmótico Ψπ100 (tomado de ARANDA et al., 2001)
Figura 7. Crecimiento tras la clara (tomado de ARANDA et al., 2004)
20
40
60
80
100
120C T2
Hei
ght (
cm)
0
3
6
9
12
0
1998 1999
C T2
Stem
dia
met
er (
mm
)
18
JOSÉ A. PARDOS CARRIÓN «La Ecofisiología en el ámbito forestal»
Estaquillas enraizadas de material clonalde Eucalyptus globulus se sometieron a dosniveles de estrés hídrico manteniendo en 9,5 y8,6 % (peso seco) el contenido en agua delsubstrato de cultivo, que permitió alcanzarpotenciales de -0.5 y -1.0 MPa respectivamen-te, frente a un control a un 30% (peso seco) decontenido en agua, mediante adición periódica
de agua. Se cosecharon las plantas a los 60 y 95días, se realizaron mediciones morfológicas, detranspiración y se constituyeron curvas P-V. Seencontraron diferencias significativas entre clo-nes en la expansión foliar y en la transpiración(figura 12); también la interacción clon x trata-miento fue significativa en el área foliar especí-fica. El estrés hídrico condujo a la reducción de
Figura 8. Tasas de fotosíntesis neta máxima (Amax) y conductancia estomática al vapor de agua (gwvmax)
0
2
4
6
8
10
11 -06 30 -06 28 -07 6 -09
1999
0
2
4
6
8
10
10 -06 30 -06 28 -07 1 -09
C T 1998
Am
ax (
µm
ol m
-2 s
-1 )
n.s.
**
*
* ***
*** ***
**
0
50
100
150
200
250
11 -06 30 -06 28 -07 6-09 0
50
100
150
200
250
10 -06 30-06 28 -07 1-09
gw
vm
ax (
mm
ol m
-2 s
-1 )
C T 1998 1999
*
* *
n.s. n.s.
**
*** ***
0
10
20
30
40
50
60
Elo
ngac
ión
del b
rote
(cm
)
0
10
20
30
40
50
60
Mar Abr May Jun Jul Ago Sep Oct Nov
20
A (µ
mol
·m-2
·s-1)
**
0
5
10
15
Mar Abr May Jun Jul Ago Sep Oct Nov
**
0
1
2
3
4
5
6
7
8
Mar Abr May Jun Jul Ago Sep Oct Nov
-2s-1
**
0
1
2
3
4
5
6
7
8
Mar Abr
E (
mm
ol·m
)
**
0.7
0.8
0.9
1
0. 6
0. 5
***
Fv/F
m
Mar Abr May Jun Jul Ago Sep Oct Nov
***
carrasco piñonero
Figura 9. Evolución de la elongación del brote, tasas de fotosíntesis (A) y transpiración (E) y eficiencia máxima delfotosistema II (Fv/Fm) en el periodo vegetativo (tomado de SIERRA, 2003)

los potenciales osmóticos a pérdida de turgen-cia y turgencia plena y aumentó el contenidohídrico relativo a pérdida de turgencia y elmódulo de elasticidad máximo (no mostrado).Las diferencias entre clones respecto a relacio-nes hídricas sólo fueron significativas en el tra-tamiento de sequía más moderada. Se apreciaun cierto ordenamiento clonal en los parámetroshídricos en respuesta al estrés hídrico (Figura13) consistente con las diferencias encontradasen la expansión foliar y, también, con las obser-vaciones hechas en campo.
La construcción de curvas P-V permitiódeterminar los principales parámetros hídricos(tabla con sus valores en PITA Y PARDOS, 2001)
Dada la importancia de la cavitación duran-te el estrés hídrico (TOGNETTI et al., 1998), conuna metodología de cultivo semejante, en clo-nes de Eucalyptus globulus sometidos a estrés
se midió la conductividad hidráulica y otrosparámetros (evolución de la superficie foliar,evapotranspiración por planta y, potencial hídri-co de base).
La conductividad hidráulica se midió en seg-mentos de tallo escindidos de plantas sometidasa diverso grado de estrés, calculando la pérdidade conductividad hidráulica, y ajustándose des-pués una sigmoide exponencial a la curva devulnerabilidad para los diferentes valores depotencial hídrico que experimentaron las plan-tas. La sequía redujo la evapotranspiración hastaun 70-80% respecto a las plantas con buen sumi-nistro de agua y también el crecimiento de lashojas, con una relación significativa entreambos parámetros (Figura 14). Los clones ensa-yados perdieron hasta un 70% de conductividadhidráulica para valores de potencial hídricomenos negativos de -1 MPa y se alcanzaron pér-
19
Cuad. Soc. Esp. Cienc. For. 20: 11-23 (2005) «Actas de la I Reunión sobre Ecología, Ecofisiología y Suelos Forestales»
Figura 10. Variación estacional de π100 en plantas bajoestrés hídrico, con ajuste osmótico
Figura 11. Tasa de fotosíntesis respecto a ψ (tomado deFERNÁNDEZ et al., 1999 y 2000)
Figura 12. Relación entre tasa de transpiración y la superficie de hoja media de siete clones de E. globulus, en plan-tas con un contenido en agua del sustrato del 30% (C), y 8.6% (S2)
Control. 2ªcosecha
y = 0,0495x + 7,5078
R2 = 0,8347
5
15
25
35
45
55
0 500 1000 1500
área foliar (cm 2 planta -1)
Tra
nspi
raci
ón (
g di
a-1
)
115.2
115.3
131.2
131.3
161.3
334.1
413.7
S2. 2ªcosecha
y = 0,0216x + 5,8089
R2 = 0,8135
5
10
15
20
25
30
35
0 500 1000
área foliar (cm 2 planta -1)
Tra
nspi
raci
ón (
g di
a-1
)
115.2
115.3
131.2
131.3
161.3
334.1
413.7

didas del 85 % sin producirse caída de hojas Elcoeficiente de la pendiente de la curva varióapreciablemente entre los clones mas y menosvulnerables (véase PITA et al., 2003).
DESARROLLO Y PLASTICIDADFENOTÍPICA
Tema de gran interés actual, se ejemplificacon la respuesta a la irradiancia en la anatomía
foliar de ocho frondosas y con el comportamien-to del pino canario.
La realización de fotografías hemisféricas yel cálculo de GSF (global site factor) como sub-rrogado de la irradiancia relativa y la determina-ción de caracteres morfoanatómicos, permitióestablecer relaciones entre las ocho especies(haya, roble albar, melojo, acebo, serbal, cere-zo y espino) presentes en el Hayedo de Montejoy los parámetros medidos. Se puso de manifiestola existencia de un mismo patrón para la relación
20
JOSÉ A. PARDOS CARRIÓN «La Ecofisiología en el ámbito forestal»
Figura 13. Clasificación clonal de acuerdo con el ajuste osmótico (OA, MPa) y el módulo de elasticidad (εmax, MPa)medido en plantas estresadas los días 49-59, e incremento del tamaño de la hoja media(∆LM) entre días 60 y 95
6.95 5.93 5.33 5.08 4.96 3.36 3.16
0.04
0.13
0.31
0.41
0
100
200
300
400
500
600
700
800
LM
(mm
2 h
oja
-1)
ε (MPa)
AO
(MP
a)
∆
115.
3
413.
7
161.
313
1.2
131.
311
5.2
334.
1
Figura 14. Relación entre superficie foliar (d.41) y transpiración de la planta (2 semanas después de la imposición dela sequía (d.40-44). Símbolos en blanco, bien regados (tomado de PITA et al., 2003)
y = 30,115Ln(x) - 131,97R2 = 0,8269
10
15
20
25
30
35
40
45
50
55
60
65
100 200 300 400 500area foliar(cm2 planta -1)
evap
otra
nspi
raci
ón (
g pl
anta
-1 d
ia-1
)
115.7
115.18
115.16
115.21
115.7
115.18
115.16
115.21c
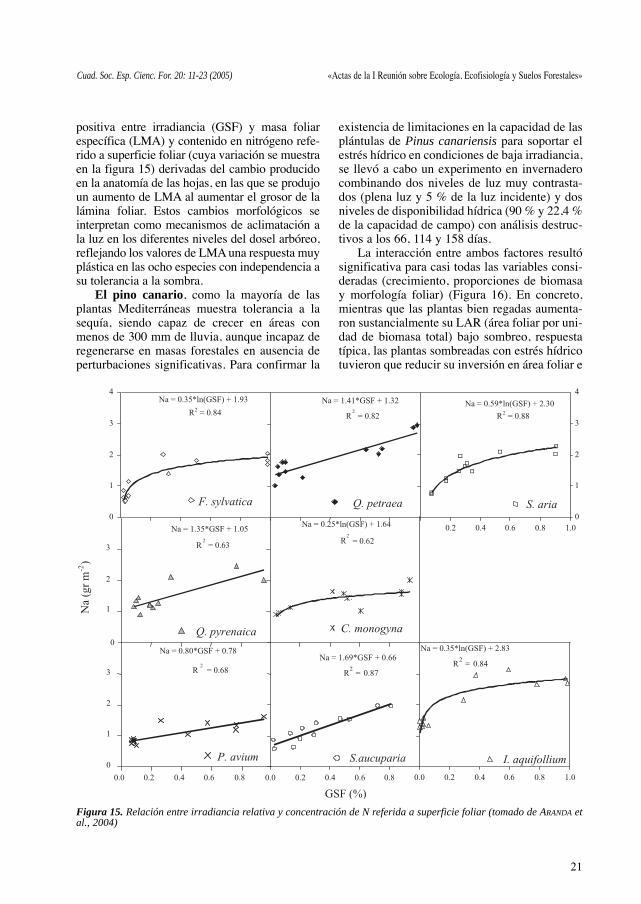
positiva entre irradiancia (GSF) y masa foliarespecífica (LMA) y contenido en nitrógeno refe-rido a superficie foliar (cuya variación se muestraen la figura 15) derivadas del cambio producidoen la anatomía de las hojas, en las que se produjoun aumento de LMA al aumentar el grosor de lalámina foliar. Estos cambios morfológicos seinterpretan como mecanismos de aclimatación ala luz en los diferentes niveles del dosel arbóreo,reflejando los valores de LMA una respuesta muyplástica en las ocho especies con independencia asu tolerancia a la sombra.
El pino canario, como la mayoría de lasplantas Mediterráneas muestra tolerancia a lasequía, siendo capaz de crecer en áreas conmenos de 300 mm de lluvia, aunque incapaz deregenerarse en masas forestales en ausencia deperturbaciones significativas. Para confirmar la
existencia de limitaciones en la capacidad de lasplántulas de Pinus canariensis para soportar elestrés hídrico en condiciones de baja irradiancia,se llevó a cabo un experimento en invernaderocombinando dos niveles de luz muy contrasta-dos (plena luz y 5 % de la luz incidente) y dosniveles de disponibilidad hídrica (90 % y 22,4 %de la capacidad de campo) con análisis destruc-tivos a los 66, 114 y 158 días.
La interacción entre ambos factores resultósignificativa para casi todas las variables consi-deradas (crecimiento, proporciones de biomasay morfología foliar) (Figura 16). En concreto,mientras que las plantas bien regadas aumenta-ron sustancialmente su LAR (área foliar por uni-dad de biomasa total) bajo sombreo, respuestatípica, las plantas sombreadas con estrés hídricotuvieron que reducir su inversión en área foliar e
21
Cuad. Soc. Esp. Cienc. For. 20: 11-23 (2005) «Actas de la I Reunión sobre Ecología, Ecofisiología y Suelos Forestales»
Figura 15. Relación entre irradiancia relativa y concentración de N referida a superficie foliar (tomado de ARANDA etal., 2004)
0
1
2
3
4
0 0.2 0.4 0.6 0.8 1.0
Na = 0.35*ln(GSF) + 1.93 R 2 = 0.84
Na = 1.41*GSF + 1.32
R 2 = 0.82
Na = 0.59*ln(GSF) + 2.30
R2 = 0.88
1
2
3
4
Na
(gr
m-2
)
Na = 1.35*GSF + 1.05
R 2
= 0.63
1
2
3
Na = 0.25*ln(GSF) + 1.64
R 2
= 0.62
Na = 0.80*GSF + 0.78
R 2
= 0.68
0
1
2
3
0.0 0.2 0.4 0.6 0.8 0.0
Na = 1.69*GSF + 0.66
R 2 = 0.87
0.2 0.4 0.6 0.8
0 Na = 0.35*ln(GSF) + 2.83
R 2
= 0.84
0.0 0.2 0.4 0.6 0.8 1.0
I. aquifollium
GSF (%)
Q. petraea F. sylvatica
Q. pyrenaica C. monogyna
S.aucuparia P. avium
S. aria

incrementar su proporción de biomasa radical(RMF). La poca viabilidad de este fenotipo seconfirmó por la extrema reducción del alarga-miento del tallo e incluso la muerte de algunasplántulas (CLIMENT et al., 2004).
Estos resultados confirman que la sequíapuede acentuar los efectos negativos de la som-bra en la regeneración del pino canario en con-diciones secas, las cuales predominan en granparte del área natural de la especie.
Agradecimientos
Agradezco la ayuda prestada para la confec-ción de este trabajo a Pilar Pita, Ismael Aranda,José Mª Climent, Jaime Puértolas y JesúsAlonso.
BIBLIOGRAFÍA
ARANDA, I.; GIL, L. & PARDOS, J.A.; 2000. Waterrelations and gas exchange in Fagus sylvati-ca and Quercus petraea in a mixed stand at
their southern limit of distribution inEurope. Trees 14: 344-352.
ARANDA, I.; BERGASA, L.F.; GIL, L. & PARDOS,J.A.; 2001. Effects of relative irradiance onthe leaf structure of Fagus sylvatica see-dlings planted in the understory of a Pinussylvestris stand after thinning. Ann. For. Sci.58: 673-680.
ARANDA, I. ; GIL, L. & PARDOS, J.A.; 2001.Effects of thinning in a Pinus sylvestrisstand on foliar water relations of Fagussylvatica seedlings planted within the pine-wood. Trees 15: 358-364.
ARANDA, I.; GIL, L. & PARDOS, J.A.; 2004.Improvement of growth conditions and gasexchange of Fagus sylvatica seedlings plan-ted below a recently thinned Pinus sylvse-tris stand. Trees 18: 211-220.
ARANDA, I.; PARDO, F.; GIL, L. & PARDOS, J.A.,2004. Anatomical basis of the change in leafmass per area and the investment with relati-ve irradiance within the canopy of eight tem-perate tree species. Acta Oecol. 25: 187-195.
AUSSENAC, G.; 2001. Interactions between foreststands and microclimate: Ecophysiological
22
JOSÉ A. PARDOS CARRIÓN «La Ecofisiología en el ámbito forestal»
Figura 16. Interacción entre factores (triángulos: 90 % capacidad de campo; cuadrados: 22,4 % de la cc.) La res-puesta de plantas sombreadas y bien iluminadas es aparentemente opuesta (ver análisis alométrico). Además, la ampli-tud de la diferencia entre ambos niveles de riego es siempre más acusada bajo sombreo. SLA: área foliar específica;LAR: área foliar por unidad de biomasa total; LMF y RMF, proporciones de biomasa foliar y radical, respectivamen-te (tomado de CLIMENT et al., 2004)
0,20
0,40
0,60
0,80
LM
F
0,09
0,14
0,19
0,24
SLA
(cm
2/m
g)
SLA
(cm
2/m
g)
0,05
0,06
0,08
0,09
0,20
0,25
0,30
0,35
0,40
RM
F
Luz Sombra Luz Sombra
Luz Sombra Luz Sombra

aspects and consequences for silviculture.Ann. For. Sci. 57: 287-301.
CLIMENT, J.M.; ALONSO, J.; PARDOS, J.A.; GIL, L.& ARANDA, I.; 2004. Irradiance affects deve-lopmental plasticity for contrasting wateravailability in Canary Island pine.Simposium Creta.
COLLET, C.; LANTER, O. & PARDOS, M.; 2001.Effect of canopy opening on the morphologyand anatomy of naturally regenerated beechseedlings. Ann. For. Sci., 58: 127-134.
FERNANDEZ, M.; GIL, L. & PARDOS, J.A.; 1999.Response of Pinus pinaster provenances atearly age to watere supply. I Water relationparameters. Ann. For. Sci. 56: 179-187.
FERNÁNDEZ, M.; GIL, L. & PARDOS, J.A.; 2000.Effects of water supply on gas exchange inPinus pinaster Ait provenances during theirfirst growing season. Ann. For. Sci. 57: 9-16.
FITTER, A.H. & HAY, R.; 2002. EnviromentalPhysiology of Plants. 3rd edit., AcademicPress, 367 pp.
GRANIER, A. et al.; 1994. Axial and radial waterflow in the trunks of oak trees: a quantitati-ve and qualitative analysis. Tree Physiol.14:1383-1396.
KOZLOWKI, T.T. & PALLARDY, S.G.; 1998.Physiology of Woody Plants. AcademyPress, pp 411.
LAMBERS, H.; STUART, F. & PONS, T.L.; 1998.Plant Physiological Ecology. Springer, 540pp.
LANDSBERG, J.J. & GOWER, S.T.; 1997. Applica-tions of Physiological Ecology to ForestManagement. Academic Press, 354 pp.
LEVITT, J.; 1980. Responses of Plants toEnvironmental Stresses. Vol. 1, AcademicPress, 497 pp.
PITA, P.; SORIA, F.; CAÑAS, I.; TOVAL, G. & PARDOS,J.A.; 2001. Carbon isotope discrimination andits relationship to drought resistance underfield conditions in genotypes of Eucalyptusgobulus. Forest Ecol. Manag. 141: 211-221.
PITA, P. & PARDOS, J.A.; 2001. Growth, leaf mor-phology, water use and tissue water relationsof Eucalyptus globulus clones in response towater deficit. Tree Physiol. 21, 599-607.
PITA, P.; GASCÓ, A. & PARDOS, J.A.; 2003.Xylem cavitation, leaf growth and leaf waterpotential in Eucalyptus globulus clonesunder well-watered and drought conditions.Funct. Plant Biol. 30: 891-899.
SULTAN, S.E.; 2000. Phenotipic plasticity forplant development, function and life-history.Trends Plant. Sci. 5: 537-542.
SUN, Z.J. et al.; 1996. Stable carbon isotopes asindicators of increased water use efficiencyin white spruce (Picea glauca) seedlings.Plant Cell Environ. 19, 887-894.
TOGNETTI, R.; LONGOBUCCO, A. & RASCHI, A.;1998. Vulnerability of xylem to embolism inrelation to plant hydraulic resistance inQuercus pubescens and Quercus ilex co-occurring in a Mediterranean coppice standin central Italy. New Phytol. 139: 437-447.
23
Cuad. Soc. Esp. Cienc. For. 20: 11-23 (2005) «Actas de la I Reunión sobre Ecología, Ecofisiología y Suelos Forestales»