instituto nacional de pesquisas da amazÔnia-inpa …...de várzea abrigam grande diversidade de...
TRANSCRIPT

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA-INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA DE ÁGUA DOCE E
PESCA INTERIOR- BADPI
O ciclo de vida de Neoechinorhynchus (Neoechinorhynchus) buttnerae Golvan, 1956
(Eoacanthocephala: Neoechinorhynchidae) parasito de Colossoma macropomum
(Cuvier, 1818) (Characiformes: Characidae) da Amazônia brasileira.
Felipe de Sousa Lourenço
Manaus – Amazonas
Junho de 2017

Felipe de Sousa Lourenço
O ciclo de vida de Neoechinorhynchus (Neoechinorhynchus) buttnerae Golvan, 1956
(Eoacanthocephala: Neoechinorhynchidae) parasito do Colossoma macropomum
(Cuvier, 1818) (Characiformes: Characidae) da Amazônia brasileira
Orientador: José Celso de Oliveira Malta, Dr
Manaus – Amazonas
Junho de 2017
______________________________________________________________________
Fonte financiadora: CNPq (Conselho Nacional de Desenvolvimento Científico e
Tecnológico).
Dissertação de mestrado apresentada ao Instituto
Nacional de Pesquisas da Amazônia como parte dos
requisitos para a obtenção do título de Mestre em
Ciências Biológicas, área de concentração em Biologia
de Água Doce e Pesca Interior.

iii
SINOPSE
L892 Lourenço, Felipe de Sousa
O ciclo de vida de Neoechinorhynchus (Neoechinorhynchus) buttnerae
Golvan, 1956 (Eoacanthocephala: Neoechinorhynchidae) parasito do
Colossoma macropomum (Cuvier, 1818) (Characiformes: Characidae) da
Amazônia brasileira /Felipe de Sousa Lourenço. --- Manaus: [s.n.], 2017.
xiii, 45 f.: il.
Dissertação (Mestrado) --- INPA, Manaus, 2017.
Orientador: José Celso de Oliveira Malta
Área de concentração: Biologia de Água Doce e Pesca Interior
1. Parasita. 2. Acantocephala. 3. Larva. 4. Characiforme. 5. Piscicultura. I.
Título.
CDD 578.65
Sinopse: Foi estudado o ciclode vida de Neoechinorhynchus (Neoechinorhynchus) buttnerae
Golvan, 1956 (Eoacanthocephala: Neoechinorhynchidae) parasito do Colossoma
macropomum (Cuvier, 1818) (Characiformes: Characidae), através da descrição morfológica
de seus estágios larvais e identificação do seu hospedeiro intermediário. Foram coletadas
amostras de plâncton para determinação dos hospedeiro intermediário e amostras de peixe
para identificação de infestação por N. (N.) buttnerae. O hospedeiro intermediário
encontrado é uma espécie de Ostracoda. Todos os peixes coletados estavam parasitados por
N. (N.) buttnerae. Através de experimento laboratorial, foram encontradas 11 fases larvais do
parasita, todos dentro de uma única espécie de Ostracoda, cultivados em laboratório. As
fases larvais foram descritas e comparadas a larvas de outras espécies de
Neoechinorhynchidae com seus ciclos de vida descritos na literatura.
Palavras-Chaves: Acantocephala; Larva; Characiformes; Ostracoda; Piscicultura;
Zooplâncton; Amazônia.

iv
Agradecimentos
Aos meus pais José Nestor e Francisneide de Sousa por todo o apoio durante minha
graduação e mestrado, pelos puxões de orelha pra largar o videojogo e trabalhar e por
todo o cuidado e carinho;
Ao Instituto Nacional de Pesquisas da Amazônia e ao Programa em Biologia de Água
Doce e Pesca Interior pela infraestrutura e oportunidade concedidas;
À CNPq (Conselho Nacional de Desenvolvimento Cientifico e Tecnológico) pela
concessão da bolsa de estudo;
Ao Dr. José Celso de Oliveira Malta, pela confiança depositada em mim, pela paciência
e pelos ensinamentos;
Ao Jeffson Nobre, pelos ensinamentos e correções, ajuda a conseguir e montar o
material de campo e pelos contatos;
Ao seu Edilson Araújo (Responsável Técnico do Laboratório de Parasitologia de Peixes
– LPP) pelas dicas dadas a mim no laboratório para facilitar meu trabalho, e pelo
empenho em deixar o laboratório sempre organizado;
Aos meus colegas de laboratório pela descontração, ajuda e todas as dicas e conversas
sobre meu trabalho que ajudaram a torná-lo mais fácil;
Às minhas ajudantes de campo, Ana Paula, Jéssica Tavares, Adria Costa e Anne Ezaguy
por terem aceitado acordar cedo e me acompanharem na estrada e ajudarem a coletar
minhas amostras;
À minha namorada Anne Ezaguy por todo carinho e dedicação e ajuda com os desenhos
que sem ela eu não teria conseguido fazer;
Ao seu Rashid, por disponibilizar sua propriedade para fazer as coletas e aos peixes
utilizados no trabalho;
À equipe do Programa de Pós-graduação do Badpi, Dr. Edinaldo Nelson ex-
coordenador e Drª Cláudia de Deus atual coordenadora;
Aos professores das disciplinas do Badpi pelo conhecimento transmitido;

v
Ao Laboratório de Microscopia do INPA, por ajudar nas medidas e fotografias deste
trabalho;
Aos meus colegas de turma do Badpi/2015, por todos os momentos de descontração,
diversão e pelas conversas onde todos se ajudaram em seus trabalhos;
E a todos que contribuíram com meu trabalho de uma forma ou de outra.

vi
SUMÁRIO
Resumo...........................................................................................................................8
Abstract...........................................................................................................................9
1. Introdução...................................................................................................................10
2. Objetivo Geral............................................................................................................15
3. Objetivos específicos..................................................................................................15
4. Material e Métodos.....................................................................................................15
5. Resultados...................................................................................................................22
6. Discussão....................................................................................................................31
Refências........................................................................................................................39

vii
LISTA DE FIGURAS E TABELAS
Figura 1: Viveiro utilizado para as coletas de zooplâncton e
peixes...............................................................................................................................16
Figura 2: Juvenil de Colossoma macropomum................................................................18
Figura 3: Espécie de Ostracoda hospedeiro intermediário de Neoechinorhynchus
buttnerae..........................................................................................................................22
Figura 4: Ostracodas infectados por juvenis de N. (N.) buttnerae: A: Ostracoda com
juvenil dentro (circulo vermelho). B: Ostracoda com juvenil saindo. C: Juvenil fora do
Ostracoda. D: Larva de Fase 9 fora do Ostracoda...........................................................22
Figura 5: Ovo e larvas de Neoechinorhynchus buttnerae (Fase 1 a 5). A: Ovo. B:
Acântor eclodido (Fase 1). C: Acantela (Fase 2). D: Acantela (Fase 3). E: Acantela
(Fase 4). F: Acantela (Fase 5). Abreviações: CE, Camada externa; CI, Camada interna;
MF, Membrana de fertilização; En, Entoblasto; NVT, Núcleo Ventral do Tronco; NA,
Núcleo Apical; Re, Receptáculo da Probóscide, COS, Células dos Órgãos Sexuais; NS,
Núcleo Subcuticular; Lem, Lemnisco.............................................................................25
Figura 6: Larvas de Neoechinorhynchus buttnerae (Fase 6 a 9). A: Fase 6. B: Fase 7. C:
Fase 8. D: Fase 9. Abreviações: Pro, Probóscide; Lem, Lemnisco; COS, Células dos
órgãos sexuais; RC, Reservatório de Cemento; GC, Glândula de Cemento; Te,
Testículos.........................................................................................................................27
Figura 7: Juvenis de Neoechinorhynchus buttnerae........................................................29
Figura 8: Desenho esquemático representando o ciclo de vida de Neoechinorhynchus
buttnerae..........................................................................................................................30
Tabela 1. Medidas das espécies do gênero Neoechinorhynchus Stiles & Hassall, 1905
com ciclo de vida descrito (medidas de ovo, acântor e acantela em µm e de juvenis e
adultos em mm)...............................................................................................................31

8
Resumo
Foi descrito o ciclo de vida de Neoechinorhynchus (Neoechinorhynchus) buttnerae,
parasita de Colossoma macropomum, baseado na descrição morfológica de seus
estágios larvais e identificação de seu hospedeiro intermediário. Foram coletadas
amostras de plâncton de um viveiro escavado no município Rio Preto da Eva,
Amazonas, Brasil. Foram analisados exemplares de C. macropomum parasitados para
coleta dos adultos de N (N.) buttnerae e avaliar o parasitismo e os índices parasitários.
Todos os peixes necropsiados estavam parasitados N. (N.) buttnerae. O hospedeiro
intermediário de N (N.) buttnerae foi uma espécie de Ostracoda. Para a descrição dos
estágios larvais de N. (N.) buttnerae foi utilizado método experimental em laboratório.
Foi encontrado um total de onze estágios larvais, uma fase de acântor, e dez fases de
acantela e o 11º estágio o infectante. Tentativas de se cultivar o Ostracoda até o
desenvolvimento da larva em estágio infectante falharam, sendo cultivado até no
máximo 26 dias, onde foram observadas as larvas em estágio juvenil. Todos os estágios
foram encontrados em uma única espécie de Ostracoda. Os estágios larvais foram
descritos morfologicamente e comparados com outras espécies da família
Neoechinorhynchidae com ciclos de vida já descritos.
Palavras-Chaves: Acantocephala; Larva; Characiformes; Ostracoda; Piscicultura;
Zooplâncton; Amazônia.

9
Abstract
The life cycle of Neoechinorhynchus (Neoechinorhynchus) buttnerae, parasite of
Colossoma macropomum, was described based on morphological descriptions of its
larvae’s development and the identification of its intermediate host. Samples of
plankton were collected in a pond excavated in Rio Preto da Eva, Amazon State, Brazil.
Parasitized C. macropomum were captured to collect N (N.) buttnerae adults and to
evaluate parasitism and parasite indices. All of the sampled fish were heavily infected
with N. (N.) buttnerae. The intermediate host was a species of Ostracoda. For the
description of the larvae development, a laboratory experimental procedure was used. A
total of eleven larval stages, one acanthus phase, and ten acanthella phases were found
and the 11th stage was the infecting one. Attempts to cultivate Ostracoda until the
development of the infecting stage larva have failed. Being cultivated until the
maximum of 26 days, where they were observed the larvae in juvenile stage. All the
larvae stages were found in a single Ostracoda species. The larval stages were described
morphologically and compared to other species of the Neoechinorhynchidae family with
previously described life cycles.
Key-words: Acantocephala; Larvae; Characiformes; Ostracoda; Fish farm; Infestation;
Amazon.

10
1. Introdução
A área de várzea localizada às margens dos grandes rios de água branca da
Amazônia, chega em média a 48 km de extensão (Lowe-McConnell 1975). Podendo
chegar a 100 km de extensão em algumas áreas (Junk 1970). Estas águas brancas e áreas
de várzea abrigam grande diversidade de fauna e flora (Marlier 1967; Junk 1970). Essa
fauna é mantida graças à variação sazonal do nível da água dos rios que inunda a várzea
e floresta, fertilizando a água e a terra com os nutrientes sedimentados (Junk et al.
1989).
Essa inundação garante que diversos peixes utilizem alimentos provenientes da
floresta inundada (Goulding 1980) ou da matéria orgânica derivada de algas da várzea
(Forsberg et al. 1993). Este complexo de águas ricas em sedimentos e de inundação
sazonal garante uma fonte de alimentos que sustenta grande diversidade de peixes que
chega a 3.000 espécies (Sioli 1976; 1984). Dentre estas espécies está o Colossoma
macropomum (Cuvier, 1818).
Colossoma macropomum é uma das espécies de peixes mais importantes da
ictiofauna amazônica sendo conhecido popularmente como tambaqui. Distribui-se
amplamente nas bacias dos rios Orinoco e Amazonas, vivendo em lagos e áreas
marginais alagadas associadas às calhas dos rios principais. Pode alcançar mais de um
metro de comprimento e atingir 30 kg e é considerado o segundo maior peixe de
escamas da bacia amazônica (Goulding e Carvalho 1982).
Possui uma combinação única de dentes do tipo molariforme, adaptados para
esmagamento de sementes duras e numerosos filamentos branquiais alongados que são
empregados para filtrar o zooplâncton. Destaca-se também o seu opérculo alongado que
suporta abas carnudas grandes. Os apêndices operculares ampliados ajudam a aumentar
o fluxo de água, com o zooplâncton através das brânquias (Goulding e Carvalho 1982).
É uma das espécies de caracídeos que realizam migração nos rios em grandes
cardumes, sendo importante para a pesca (Goulding 1979; 1980; 2013). Durante a cheia
os cardumes se desfazem, e os peixes se dirigem para a floresta alagada onde se
alimentam (Goulding 1980). Quando a água desce a maior parte dos peixes foge para os
canais dos rios, e outros vão para lagos de várzea (Goulding e Carvalho 1982).
Na floresta alagada, os adultos se alimentam principalmente de frutos e
sementes, seguidos por zooplâncton e arroz-bravo Oriza sp. (Carvalho 1981). No

11
período de seca, a maior parte da alimentação é zooplâncton, principalmente de espécies
de Cladocera. Os jovens imaturos de C. macropomum alimentam-se principalmente de
zooplâncton, tendo frutos secos e sementes como complemento (Carvalho 1981;
Goulding e Carvalho 1982).
O tambaqui é a maior espécie dos Characiformes, ocorrendo na parte
setentrional da América do Sul. Devido à sobre pesca de seus estoques naturais ele
integra a lista de espécies protegidas e sua pesca está proibida durante o período de
reprodução, chamado regionalmente de defeso (novembro a março). Esta medida visa
ajudar na recuperação dos estoques (Araujo-Lima e Goulding 1998). Entretanto a
demanda por esta espécie está aumentando, gerando um crescente interesse pela
produção de peixes em cativeiro (Varella et al. 2003; Morais et al. 2009).
Colossoma macropomum é a espécie mais produzida na piscicultura do
Amazonas com quatorze mil toneladas, equivalente à renda total anual de R$ 100
milhões (IBGE 2015). Sua produção em cativeiro é importante devido à sobre-pesca dos
estoques naturais, o que acarretou numa diminuição do desembarque pesqueiro no
estado do Amazonas (Batista e Petrere 2003).
O tambaqui tem bom desempenho zootécnico, como o peso que pode chegar até
2,2 kg em apenas um ano e meio de produção, quando alimentado duas vezes ao dia
(Mello et al. 2015), e apresenta um melhor crescimento em viveiro escavado quando
comparado ao sistema de canal de igarapé (Arbeláez-Rojas et al. 2002). Seus juvenis
apresentam uma maior resiliência para transporte, quando comparados a juvenis de
outras espécies (Gomes et al. 2003).
Outro motivo para o aumento de sua produção é o custo da alimentação, que
pode ser reduzido utilizando proteínas vegetais alternativas ao milho, como a farinha de
babaçu ou a crueira de mandioca, mantendo a mesma eficácia das rações comerciais
(Lopes et al. 2010; Pereira Junior et al. 2013).
Em tanques de piscicultura é necessário fazer a fertilização para aumentar a
quantidade de fitoplâncton e consequentemente de zooplâncton e outros invertebrados,
importantes para a alimentação complementar da maior parte das espécies de peixes
cultivadas em cativeiro (Ostrensky e Boeger 1998). A composição dessa fauna
planctônica pode variar em períodos de seca e de chuva (Macedo e Sipaúba-Tavares
2005).

12
Dentre os táxons encontrados em viveiro de C. macropomum estão em maior
quantidade: Copepoda, Cladocera e Rotifera, e em menor quantidade, larvas de Diptera,
Trichoptera, Ephemeroptera, Conchostracoda e Ostracoda (Costa 2013). Os Ostracoda
são crustáceos aquáticos com uma carapaça bivalve, e possuem de 0,5 mm a 2 mm na
fase adulta. São bentônicos, mas possuem capacidade natatória podendo chegar à
superfície. Possuem reprodução sexuada, assexuada ou mista (Horne et al. 2002).
Os Ostracoda assim como outros invertebrados aquáticos são importantes
hospedeiros intermediários de parasitas de peixes e outros vertebrados de ambiente
aquático. Estes seres são responsáveis por completar os ciclos de vida de diversas
espécies de parasitas de peixes (Kennedy, 2006).
Os peixes são os vertebrados mais parasitados, sendo seus parasitas incluídos em
vários filos e com um grande número de espécies já descritas (Malta 1984). São
conhecidas aproximadamente 10.000 espécies de parasitas, sendo 4.200 ectoparasitas de
peixes, possuindo distribuição mundial e afetando todas as espécies de peixes, em
praticamente todos os corpos d’água (Eiras 1994).
São citados para o C. macropomum espécies parasitas de nove táxons:
Prokariotae (Monera); Fungi; Ciliophora; Sarcomastigophora; Myxozoa;
Platyhelminthes, (Monogenoidea, Digenea e Cestoda); Nematoda; Acanthocephala e
Arthropoda (Copepoda, Branchiura e Isopoda) (Malta e Varella 1983; 2000; Malta et al.
2001; Fischer et al. 2003; Thatcher 2006).
O filo Acantocephala é monofilético e é um dos dois únicos filos conhecidos que
as espécies são exclusivamente parasitas, sem nenhuma fase de vida livre. Este grupo
possui pouca variação de ciclo de vida, e de estruturas corporais, tornando o trabalho
dos sistematas muito difícil. Todas as espécies de Acantocephala possuem um ovo de
vida livre com o acântor, um artrópode é hospedeiro intermediário para a fase de
acantela e cistacanto, um vertebrado é o hospedeiro definitivo (Kennedy 2006).
Quando comparadas as espécies de outros filos parasitas, as espécies de
Acantocephala apresentam uma uniformidade morfológica, baixa diversidade e nunca
são livres natantes. Mesmo assim elas são consideradas um grupo com alto sucesso
evolutivo, pois parasitam espécies de todas as classes de vertebrados, em todos os
continentes e biomas. Diferente das espécies dos filos Nematoda e Platyhelminthes que
têm grande sucesso evolutivo e grande diversidade de espécies (Kennedy 2006).

13
Populações e comunidades de parasitas em geral são importantes indicadores de
estresse ambiental, estrutura de cadeia alimentar e biodiversidade (Marcogliese 2005).
Por existirem poucos estudos em relação ao ciclo de vida de acantocéfalos, é difícil
utilizá-los como marcadores regionais ou globais, mas eles podem ser usados
efetivamente como marcadores locais (Kennedy 2006).
Neoechinorhynchus (Neoechinorhynchus) rutili (Mueller, 1780) foi usado com
sucesso para determinar a origem geográfica de Salmo trutta Linnaeus, 1758. A
infecção acontecia apenas nos peixes provenientes do riacho que desaguava em um
pequeno lago, enquanto os peixes estocados no lago não estavam parasitados (Kennedy
e Burrough 1978).
Os acantocéfalos podem ser usados para conhecer qual o alimento do peixe. Os
altos índices de infestação de Acanthocephalus lucii (Mueller, 1780) em Anguilla
anguilla (Linnaeus, 1758) indicou que se alimentavam de Asellus aquaticus (Linnaeus,
1758) (Kennedy e Moriarty 2002).
As espécies de Acantocephala podem ser utilizadas como indicadoras de
poluição orgânica. Os seus hospedeiros intermediários sofrem declínio em águas
poluídas, o que acarreta numa menor intensidade de parasitismo nos hospedeiros
definitivos (Kussat 1969; Overstreet e Howse 1977; Vidal-Martinez et al. 2003).
Para se realizar estes estudos é necessário conhecer os ciclos de vida destas
espécies, principalmente conhecer seus hospedeiros intermediários. São conhecidos para
a família Neochinorhynchidae nove ciclos de vida descritos, sendo seis deles do gênero
Neoechinorhynchus Stiles & Hassall, 1905 (Al-Sady 2009).
Nove espécies de Neochinorhynchidae têm os ciclos de vida descritos.
Paulisentis missouriensis Keppner, 1974 (Keppner 1974); P. fractus Van Cleave &
Bangham, 1949 (Cable e Dill 1967); Octospiniferoides chandleri Bullock, 1957
(DeMont e Corkum 1982); Neoechinorhynchus (Neoechinorhynchus) cristatus Lynch,
1936 (Uglem 1972); N. saginatus Van Cleave e Bangham, 1949 (Uglem e Larson
1969); N. (N.) iraqensis Amin et al. 2001 (Al-Sady 2009); N. (N.) emydis (Leidy, 1851)
(Hopp 1954); N. (N.) cylindratus (Van Cleave, 1913) (Ward 1940) e N. (N.) rutili
(Mueller, 1780) (Merritt e Pratt 1964).
As espécies de Acantocephala são altamente específicas aos hospedeiros
definitivos e muito pouco específicas para seus hospedeiros intermediários e paratênicos

14
(Kennedy 2006). Elas são menos específicas que outros grupos de helmintos, e
apresentam mais hospedeiros intermediários e paratênicos do que as espécies parasitas
de outros filos (Poulin e Mouillot 2003). A variedade de hospedeiros depende mais de
suas semelhanças ecológicas do que de sua proximidade taxonômica (Poulin 2005).
Neoechinorhynchus (Neoechinorhynchus) buttnerae pertence à família
Neoechinorhynchidae, e é específico do C. macropomum. Possui uma probóscide
pequena em relação ao corpo, esférica com aproximadamente 0,30 mm de diâmetro.
Possui três tipos de ganchos, sendo os ganchos grandes superiores, médios e pequenos
inferiores. Possui receptáculo inserido na união do pescoço com a probóscide, e o
gânglio nervoso se localiza na parte inferior dessa estrutura (Golvan 1956).
A probóscide na região anterior do corpo é utilizada para a fixação no intestino
do hospedeiro. Dependendo do tamanho da probóscide, do tamanho dos espinhos e da
quantidade de indivíduos, esses parasitos podem causar danos superficiais, profundos
ou lesões mais graves causando hemorragias. As espécies de Acanthocephala
representam ameaça ao hospedeiro, por interferirem na estrutura responsável pela
absorção de nutrientes (Thatcher 2006).
As espécies de Acanthocephala são mundialmente conhecidas por não causarem
danos em criações de peixes por terem alta especificidade e por serem heteroxenas. Na
Amazônia, as criações de C. macropomum vêm sofrendo sérios problemas com o N.
(N.) buttnerae. Nestes ambientes confinados o ciclo de vida N. (N.) buttnerae têm se
completado com a presença dos hospedeiros intermediários e definitivos ocasionando
uma explosão nas populações de N. (N.) buttnerae.
Em alguns casos ocorre infestação maciça por esses parasitas, o que acaba
obstruindo a luz do intestino. Essas condições causam alterações na morfologia externa
dos jovens de C. macropomum e indivíduos parasitados crescem menos que os
saudáveis, com achatamento da região dorsal anterior do corpo, e cor mais pálida (Malta
et al. 2001).
Estas infestações maciças causam alterações histopatológicas no intestino,
afetando a mucosa, submucosa e tecido muscular, provocando inflamações e
granulomas (Jerônimo et al. 2017; Matos et al. 2017). Nos casos de penetração no
tecido muscular podem ocorrer metaplasias, com substituição do tecido muscular por

15
tecido conjuntivo frouxo, com presença de macrófagos, linfócitos, eosinófilos e
fibroblastos (Matos et al. 2017).
Um estudo detalhado sobre o ciclo de vida de N. (N.) buttnerae foi realizado ao
longo desse projeto através de amostragem no hospedeiro e seu ambiente de cultivo. Ao
mesmo tempo, um sistema de aquários foi planejado para abrigar o hospedeiro
infectado, o qual proporcionou a observação de novos dados que auxiliaram na
descrição do ciclo de vida de N. (N.) buttnerae.
Os períodos de incubação da larva, o local de fixação no hospedeiro e
características biométricas de formas larvais foram detalhados. As novas informações
auxiliarão na manutenção do estado de saúde do C. macropomum através do controle de
N. (N.) buttnerae nos tanques de criação do C. macropomum.
2. Objetivo Geral
Conhecer o ciclo de vida de Neoechinorhynchus (Neoechinorhynchus) buttnerae
através de detalhes de sua biologia taxonômica e reprodutiva.
3. Objetivos específicos
• Descrever a morfologia do ovo de N. (N.) buttnerae e suas adaptações;
• Descrever a morfologia do acântor dentro do ovo de N. (N.) buttnerae;
• Conhecer, identificar os hospedeiros intermediários de N. (N.) buttnerae;
• Descrever a morfologia da acantela de N. (N.) buttnerae;
• Descrever a morfologia do cistacanto de N. (N.) buttnerae;
• Avaliar se as larvas de N. (N.) buttnerae utilizam hospedeiros paratênicos;
• Definir o desenvolvimento das larvas acântor até a fase de acantela infectante,
sua ocorrência e tempo de vida;
• Verificar associações entre o hábito alimentar do hospedeiro e o ciclo de vida de
N. (N.) buttnerae.
4. Material e métodos
Aquisição dos peixes

16
Foi localizada uma propriedade particular no Município de Rio Preto da Eva no
km 116 da rodovia estadual AM-010 onde cultivam C. macropomum. O proprietário
disponibilizou dez peixes criados em viveiro escavado (Figura 1), que foram utilizados
neste trabalho.
Os C. macropomum de diferentes tamanhos foram capturados com rede de
arrasto. Depois foram colocados em baldes de água com Eugenol a 0,5 mL/L para
eutanásia e levados conservados em gelo até o Laboratório de Parasitologia de Peixes
(LPP) do INPA para necrópsia.
Figura 1: Viveiro da propriedade localizada no Município de Rio Preto da Eva
no km 116 da rodovia estadual AM-010, onde foram feitas as coletas de zooplâncton e
de Colossoma macropomum.
Eutanásia e necrópsia dos Colossoma macropomum
Os peixes foram eutanasiados conforme as “Diretrizes da Prática de Eutanásia
do CONCEA” de 2013 e aprovado pela “Comissão de Ética no Uso de Animais”
(CEUA) do INPA Processo 038/2016. Seguindo as normas, os peixes foram colocados
em um recipiente contendo água com óleo de cravo (Eugenol) na concentração de 0,5

17
ml por litro. Os peixes ficaram expostos até cessarem os batimentos operculares, a
coluna vertebral foi seccionada 20 minutos após esse evento.
Em seguida foram pesados e medidos (comprimento padrão). A necrópsia foi
feita de acordo com o “Roteiro de Necropsias do LPP”. Cada peixe foi examinado
externamente, a procura de qualquer anomalia, que possa caracterizar se uma infestação
maciça está afetando a saúde do peixe (Figura 2).
A cavidade abdominal foi aberta, utilizando uma fina tesoura, iniciando a
perfuração atrás da região urogenital, e cortando até a região das nadadeiras peitorais,
abrindo em direção às vísceras nesses dois pontos anteriores, para fazer uma abertura do
abdômen. Esta abertura foi feita para observar as vísceras.
O intestino foi examinado à procura de anomalias, como posições anormais,
inchaços, descolorações, cistos de parasitas, lesões e outros sinais patológicos que
podem ser causados por Acantocephala. O canal alimentar foi retirado inteiramente,
fazendo um corte transversal no esôfago e corte no reto próximo ao ânus.
O complexo inteiro foi colocado em uma placa, para separação cuidadosa dos
órgãos. Baço e fígado foram separados e o canal alimentar separado em seções:
estômago e esôfago, região pilórica e intestino e colocados em placas de Petri cobertas
com água destilada. O intestino foi dissecado cuidadosamente para a verificação da
presença de N. (N.) buttnerae.

18
Figura 2: Juvenil de Colossoma macropomum (Cuvier, 1818) hospedeiro definitivo de
Neoechinorhynchus (Neoechinorhynchus) buttnerae Golvan, 1956.
Coleta do zooplâncton para identificação dos hospedeiros intermediários
Arrastos horizontais com redes de plâncton de 20, 60 e 200 µm de malha foram
feitos nas águas do viveiro onde foram coletados os peixes parasitados. Os arrastos
horizontais foram feitos em toda camada superficial da água para coleta do zooplâncton.
Arrastos de fundo foram feitos para coleta de bentos.
Uma parte deste material foi fixada em formol 5%, outra em álcool 70% e uma
terceira foi mantida viva em aquários com água dos tanques e com aeração. Sob
microscópio estereoscópico os organismos foram examinados, identificados e as formas
larvais de N. (N.) buttnerae procuradas, coletadas e identificadas.
A água coletada nos tanques foi examinada sob microscópio estereoscópico à
procura de artrópodes parasitados por larvas acantela de N. (N.) buttnerae. Espécies de
vida livre de Rotifera; Copepoda (Calanoida e Cyclopoida); Cladocera; Ostracoda;
Annelida; Insecta (larvas de Diptera, de Hemiptera da família Notonectidae, de
Odonata, de Coleoptera). Os espécimens foram examinados e dissecados sob
microscópio estereoscópico.
Especial atenção foi dada aos espécimens de Ostracoda, por serem os principais
hospedeiros intermediários da família Neoechinorhynchidae (Hopp 1954; Merritt e Pratt
1964; DeMont e Corkum 1982). Além de exemplares de Copepoda e Cladocera este
último por ser a principal fonte de zooplâncton na alimentação de C. macropomum na
natureza (Goulding e Carvalho 1982).
Coleta dos Neoechinorhynchus buttnerae adultos, dos ovos e das fases larvais
Os N. (N.) buttnerae foram retirados vivos do intestino com o auxílio de
estiletes, pinceis finos e pinças para evitar que a probóscide se rompesse e ficasse presa
na parede intestinal. Em seguida foram colocados em placas de Petri em solução salina
a 0,65% e mantidos em refrigerador por aproximadamente 24 horas para que a
probóscide ficasse totalmente evaginada. Posteriormente foram fixados em A.F.A. e
conservados em etanol 70° GL (Amato et al. 1991).

19
O estudo anatômico dos N. (N.) buttnerae foi realizado de acordo com Amin
(1969). Para os estudos morfológicos os N. (N.) buttnerae foram corados com carmim
alcoólico clorídrico (Eiras et al. 2006). Algumas fêmeas de N. (N.) buttnerae foram
usadas para extrair os ovos, que foram retirados por ruptura da porção posterior do
corpo, lavados em água de torneira, centrifugados e lavados novamente. Os indivíduos
adultos e os ovos foram medidos, fotografados, desenhados, alguns foram fixados e
depositados na coleção de invertebrados do INPA.
Triagem e identificação dos hospedeiros intermediários
Os hospedeiros intermediários que estavam parasitados foram identificados até
menor nível taxonômico possível, fotografados e desenhados. Os que não estavam
parasitados foram colocados em placas de Petri. Posteriormente foram selecionados 720
espécimes do hospedeiro intermediário.
Os 720 hospedeiros intermediários foram colocados em placas de Petri grandes e
deixados 24 h sem alimento, depois que os ovos de N. (N.) buttnerae foram colocados
nas placas, com o auxilio de pipetas, eles foram ingeridos pelos hospedeiros
intermediários. Depois de duas horas, os hospedeiros intermediários foram transferidos
para 10 placas de cultura de células com 12 fossos cada uma, com no máximo 10 ml de
água em cada fosso e três indivíduos. Foram dissecados diariamente sob microscópio
estereoscópico os hospedeiros intermediários, e foi acompanhado o desenvolvimento
das larvas de N. (N.) buttnerae dentro do hospedeiro intermediário, sob microscópio
ótico.
Não foi disponibilizada nenhuma aeração, mas utilizou-se uma tampa de acrílico
para reduzir evaporação. Foi disponibilizado alimento esporadicamente, consistindo de
folhas secas trituradas à mão, assim como restos de ração triturada.
Para observação da larva acântor eclodida do ovo, foi feita uma eclosão
artificial, que consiste em deixar a lâmina temporária com os ovos secar, e reidratar
imediatamente, neste processo a membrana exterior do ovo é rompida e a larva acântor
é liberada no meio aquoso (Uglem 1972).

20
Cálculo dos índices parasitários
Os índices parasitários como a Prevalência (P); Intensidade (I); Intensidade
média de Infecção (IMI) e a Abundância (A) foram calculados e analisados de acordo
com Bush et al. (1997).
Prevalência (%): relação entre o número de hospedeiros parasitados divididos pelo
número de hospedeiros examinados, multiplicados por 100:
P =NP
NE X 100
Onde:
NP = número de peixes parasitados por uma determinada espécie de parasita.
NE = número total de peixes examinados.
A comunidade parasitária componente foi classificada como: espécies centrais
(espécies com prevalências superiores a 66%); espécies secundárias (prevalências entre
33 e 66%) e espécies satélites (prevalências abaixo de 33%) (Bush et al., 1997).
Intensidade: expresso como variação numérica (número de N. (N.) buttnerae
encontrados):
Intensidade média: mede a quantidade média de parasitismo entre os hospedeiros
parasitados:
IMI =Nsp1
NPsp1
Onde:
Nsp1 = número de N. (N.) buttnerae.
NPsp1 = número de peixes parasitados por N. (N.) buttnerae.
Abundância: número médio de N. (N.) buttnerae encontrados para o total de
hospedeiros examinados:

21
AM =NTP
NPE
Onde:
NTP = número total de N. (N.) buttnerae.
NPE = número total de peixes examinados (parasitados e não parasitados) na amostra.
Imagens, desenhos e medidas taxonômicas
Os desenhos foram feitos a partir de montagens totais de exemplares em lâminas
permanentes e provisórias em câmara clara acoplada a microscópio de luz com contraste
de fase Olympus BH-2 e Zeiss Axioscope 2 plus e digitalizados por meio de uma mesa
digitalizadora Kanvus Life 127 - TB-LIFE 127. As fotomicrografias dos espécimes
menores foram feitas em um microscópio óptico Zeiss e, para os maiores foi utilizado
estereomicroscópio Zeiss com câmera digital acoplada (Axio Cam MRc 3 e Axio Cam
ICc 5, respectivamente).
Todas as pranchas são originais, todas as medidas e escalas foram feitas com o
auxílio do software Bel micro e Zen 2-Lite. As medidas foram apresentadas em
micrometros (μm) ou milímetros (mm) com médias apresentadas entre parênteses.
Quando isso não ocorrer será indicada a unidade correspondente.
O material testemunho desse trabalho foi depositado na Coleção de
Invertebrados não Insecta do INPA, em Manaus, Protocolo de recebimento: INPA 23,
Seção: ACANTHOCEPHALA.
5. Resultados
A prevalência de N. (N.) buttnerae em C. macropomum foi de 100%. A
intensidade média 378,4, a amplitude máxima 921 e a mínima 107. O mesmo valor de
378,4 foi encontrado para abundância. O peso médio dos peixes foi de 146,73 g.
As larvas de N. (N.) buttnerae foram encontradas em uma única espécie de
Ostracoda, que foi coletada no viveiro amostrado, dentre as 3 espécies do grupo
encontradas no viveiro (Figura 3). O hospedeiro intermediário foi identificado ate o

22
nível de Superfamilia, sendo um Cypridoidea. Foram encontradas nos indivíduos
coletados no viveiro duas fases larvais de N. buttnerae no que aparenta ser uma única
espécie de Ostracoda que foram fixadas em álcool e formol (Figura 4).
Figura 3: Espécie de Ostracoda coletada no viveiro de criação de Colossoma macropomum (Cuvier, 1818)
hospedeira intermediária de Neoechinorhynchus (Neoechinorhynchus) buttnerae Golvan, 1956.

23
Figura 4: Ostracodas infectados por juvenis de Neoechinorhynchus (Neoechinorhynchus) buttnerae
Golvan, 1956 parasito de Colossoma macropomum (Cuvier, 1818): A: Ostracoda com juvenil dentro
(Circulo vermelho). B: Ostracoda com juvenil saindo. C: Juvenil fora do Ostracoda. D: Larva Acantela
fora do Ostracoda.
Os ovos (Figura 5A) de N. (N.) buttnerae têm forma ovoide, e possuem três
camadas recobrindo a larva acântor. A primeira camada externa é fina e transparente
assim como a de fertilização, enquanto que a camada interna é coberta de grânulos
refratores nas laterais. O comprimento médio do ovo é de 0,036 mm e 0,026 mm de
largura, enquanto que o da larva é de 0,028 mm de comprimento por 0,015 mm de
largura.
Fase 1, a larva acântor eclodida artificialmente (Figura 5B), não apresentava
ganchos laterais ou apicais comuns a outras espécies do gênero Neoechinorhynchus, e
apresentava um volume ligeiramente maior ao acântor de dentro do ovo. Média de
0,030 mm de comprimento e 0,019 mm de largura. Apresentava alguns vacúolos no seu
interior, mas não foi possível determinar qual a região posterior e anterior da larva.
Fase 2, com 4 dias desde a ingestão dos ovos, a larva (Figura 5C) aumentou o
tamanho, com 0,088 mm de comprimento e 0,073 mm de largura. Nesta fase a larva
tinha forma arredondada, ausência de ganchos laterais e aglomerado de núcleos na
região central do corpo, o que caracteriza o início da fase de acantela.
Fase 3, com 6 dias após a ingestão dos ovos a acantela (Figura 5D) estava
ligeiramente maior, com 0,151 mm de comprimento e 0,111 mm de largura, com 4
núcleos gigantes da hipoderme ao redor da massa nuclear central, que ainda não havia
sido diferenciada. Os núcleos da região anterior que irão formar o lemnisco se
encontravam próximos a região apical.
Fase 4, com 9 dias desde a ingestão dos ovos, a acantela (Figura 5E) passou a ter
um formato alongado, com 0,201 mm de comprimento por 0,057 mm de largura. Os
núcleos apicais da massa nuclear central se diferenciaram no receptáculo e na
probóscide primordial, os núcleos gigantes apicais se diferenciaram para formar um
lemnisco primordial. A massa nuclear da região posterior começou a se separar para
formar futuramente os órgãos genitais.
Fase 5, com 13 dias de desenvolvimento, a acantela (Figura 5F) media 0,410
mm de comprimento e 0,091 mm de largura, a probóscide apesar de invaginada,
começava a ficar mais evidente, o lemnisco se encontra em formação. O receptáculo da

24
probóscide estava quase formado. Os órgãos sexuais começaram a tomar forma a partir
da massa nuclear do entoblasto.
Figura 5: Ovo e larvas de Neoechinorhynchus (Neoechinorhynchus) buttnerae Golvan, 1956 (Fase 1 a 5).
A: Ovo. B: Acântor eclodido (Fase 1). C: Fase 2 (Acantela). D: Fase 3 (Acantela). E: Fase 4 (Acantela).

25
F: Fase 5 (Acantela). Abreviações: CE = Camada externa; CI = Camada interna; MF = Membrana de
fertilização; En = Entoblasto; NVT = núcleo ventral do tronco; NA = núcleo apical; Re = Receptáculo da
probóscide: COS = Células dos órgãos sexuais; NS = núcleo subcuticular; Lem = Lemnisco.
Fase 6, com 15 dias de desenvolvimento larval, a probóscide da acantela
(Figura 6A) estava invaginada e pode-se notar os ganchos dentro do receptáculo. O
comprimento da larva era de 0,463 mm e 0,102 mm de largura. Na região posterior foi
possível observar uma estrutura semelhante a uma glândula de cemento, indicando que
se trata de um macho, podendo nessa fase ser definido o sexo.
Fase 7, com 17 dias de desenvolvimento, a acantela (Figura 6B) mede 0,673 mm
de comprimento e 0,143 de largura, sem muita diferença morfológica evidente. A
probóscide continua invaginada, sem parecer ter a capacidade de sair do receptáculo
nessa fase, o lemnisco está bem visível. Os órgãos sexuais estão mais bem formados,
apesar de ser difícil a diferenciação.
Fase 8, com 20 dias da infecção dos ostracodas a acantela (Figura 6C) se
encontra com comprimento de 0,820 mm e 0,153mm de largura, e está tão parecida com
os adultos que pode ser confundida com a fase juvenil. A probóscide está evaginada,
sendo possível medir os ganchos superiores, que são muito maiores que os ganchos
inferiores nessa fase. Os órgãos sexuais já estão bem distintos e pode-se distinguir
claramente macho e fêmea.
Fase 9, com 23 dias após a ingestão dos ovos a acantela (Figura 6D) mede 0,803
mm de comprimento e 0,314 mm de largura. Apesar de estarem com um comprimento
menor, todas as larvas encontradas nesta fase, estavam encolhidas, e não foi possível
deixá-las esticadas, como colocando em água na geladeira, ou esticando
mecanicamente. Esta fase se assemelha muito com a fase juvenil infectante, mas ainda
não forma o cisto.

26
Figura 6: larvas de Neoechinorhynchus (Neoechinorhynchus) buttnerae (Fase 6 a 9). A: Fase 6
(Acantela). B: Fase 7 (Acantela). C: Fase 8 (Acantela). D: Fase 9 (Acantela). Abreviações: Pro =
Probóscide; Lem = Lemnisco; COS = Células dos órgãos sexuais; RC = Reservatório de Cemento; GC =
Glândula de Cemento; Te = Testículos.

27
Fase juvenil (Figura 7A e B) com 26 dias da ingestão dos ovos, mede 1,250 mm
de comprimento e 0,222 mm de largura. A probóscide se encontra totalmente formada
com capacidade de sair da bainha totalmente, e com dimensão de ganchos compatível
com a descrição dos adultos. Apesar de ser considerado um juvenil, nesta fase ainda não
existe o cisto de infecção.
Fase de juvenil infectante, chamada de cistacanto (Figura 7C) não pode ser
observada no material do experimento de infecção dos Ostracoda, mas foram
encontradas no local de coleta dentro dos crustáceos. Mediam 1,417 mm de
comprimento e 0,385 mm de largura, ganchos bem definidos semelhantes aos dos
adultos e uma cápsula que recobre o corpo, quando este está enrolado no hospedeiro
intermediário, esta é considerada a fase larval final, que é caracterizada pela formação
dessa cápsula.
Com estas descrições de todas as fases larvais, pode-se notar que o
Neoechinorhynchus buttnerae utiliza apenas um hospedeiro intermediário para concluir
seu ciclo de vida. Seu ciclo de vida completo pode ser observado na figura 8, que ilustra
como ocorre o ciclo completo deste Acantocephala. Os dados de medidas de
comprimento estão na Tabela1, em comparação com outras espécies do gênero
Neoechinorhynchus.
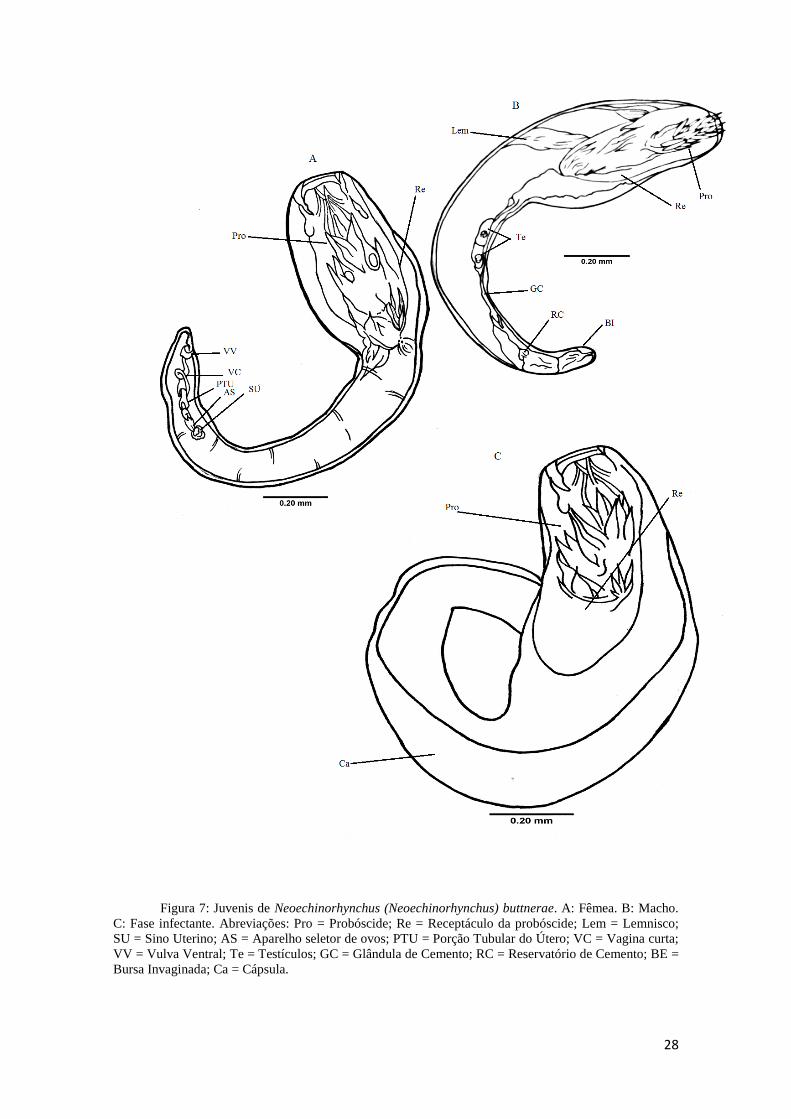
28
Figura 7: Juvenis de Neoechinorhynchus (Neoechinorhynchus) buttnerae. A: Fêmea. B: Macho.
C: Fase infectante. Abreviações: Pro = Probóscide; Re = Receptáculo da probóscide; Lem = Lemnisco;
SU = Sino Uterino; AS = Aparelho seletor de ovos; PTU = Porção Tubular do Útero; VC = Vagina curta;
VV = Vulva Ventral; Te = Testículos; GC = Glândula de Cemento; RC = Reservatório de Cemento; BE =
Bursa Invaginada; Ca = Cápsula.
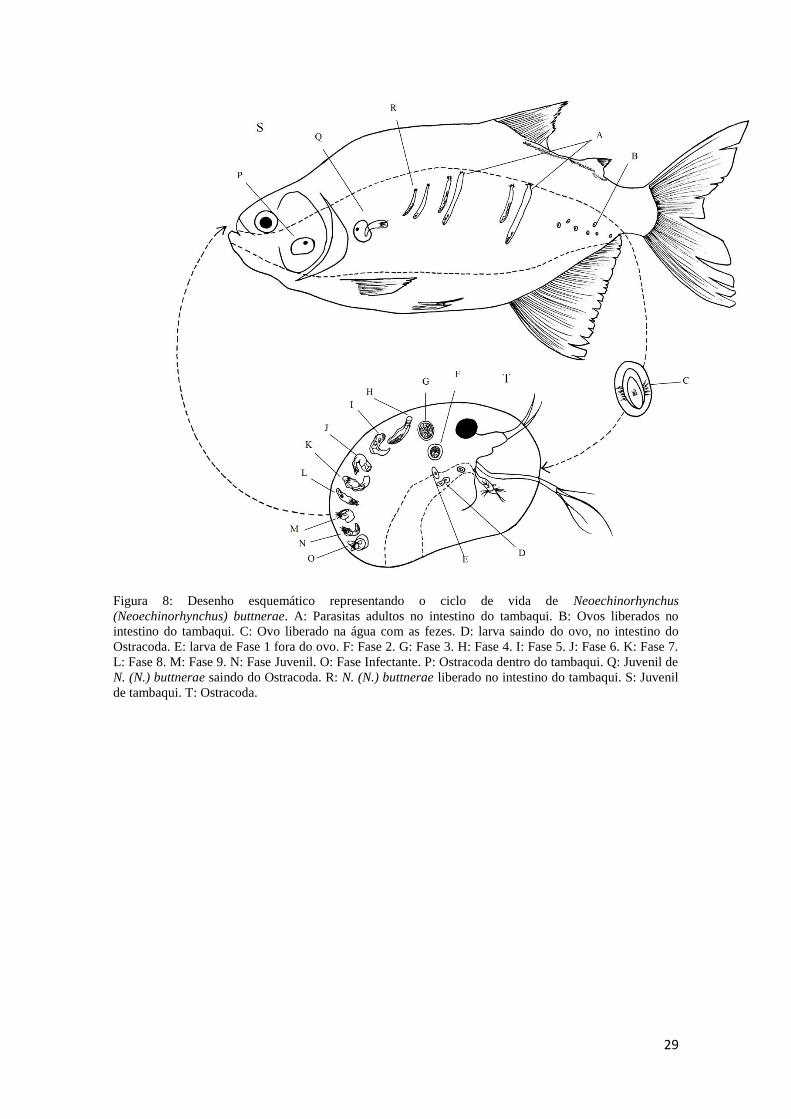
29
Figura 8: Desenho esquemático representando o ciclo de vida de Neoechinorhynchus
(Neoechinorhynchus) buttnerae. A: Parasitas adultos no intestino do tambaqui. B: Ovos liberados no
intestino do tambaqui. C: Ovo liberado na água com as fezes. D: larva saindo do ovo, no intestino do
Ostracoda. E: larva de Fase 1 fora do ovo. F: Fase 2. G: Fase 3. H: Fase 4. I: Fase 5. J: Fase 6. K: Fase 7.
L: Fase 8. M: Fase 9. N: Fase Juvenil. O: Fase Infectante. P: Ostracoda dentro do tambaqui. Q: Juvenil de
N. (N.) buttnerae saindo do Ostracoda. R: N. (N.) buttnerae liberado no intestino do tambaqui. S: Juvenil
de tambaqui. T: Ostracoda.

30
Tabela 1. Medidas dos ovos e fases larvais das espécies do gênero Neoechinorhynchus Stiles &
Hassall, 1905 com ciclo de vida descrito ( Comprimento x largura, medidas de ovo, acântor e
acantela em µm e de juvenis e adultos em mm).
N. (N.)
cristatus
N. (N.)
iraqensis
N.
saginatus
N. (N.)
emydis
N. (N.)
cylindratus
N. (N.)
rutili
N. (N.)
buttnerae
Ovos 56x27 40x18 45x18 25x22 57x28 27x17 36x26
Acantor no
ovo
42x14 39x15 22x11 24x9 28x15
Acantor
livre
62x15 45x18 68x41 33x14 62x26 40x17 30x19
Acantela 1 76x42 55x40 101x73 33x24 53x34 40x23 88x73
Acantela 2 85x72 189x95 47x31 100x79 50x19 151x111
Acantela 3 150x90 256x99 57x48 100x79 177x95 201x57
Acantela 4 280x93 306x99 66x52 790x90 305x146 410x91
Acantela 5 500 600 128x105 650x150 463x102
Acantela 6 700 1100 288x83 673x143
Acantela 7 1000 380x47 820x153
Acantela 8 550x69 803x314
Acantela 9 420x70 1250x222
Acantela 10 380x100
Acantela 11 450x100
Acantela 12 710
Juvenil 1.1 1.1 1.44 2.48 x 0.31 0.4x1 1.41x0,38
Fêmea
adulta
53.8 x
0.8
16.2 x 1 9.1 x 0.52 1.28 30x1.5
Macho
adulto
20.6 x
0.71
10.35 x
0.96
5.5 x 0.49 1.28 22x1
Tempo
(dias)
20 13 16 21 28 48-57 30-32
6. Discussão
Foi estudada a parasitofauna de C. macropomum criados em barragem de
igarapé de terra firme no Município de Iranduba (21 km de Manaus), na fase de
alevinagem e nenhum N. (N.) buttnerae foi encontrado. Os peixes utilizados tinham o
peso médio inicial de 3,85 g. (Alves 2000).
No Estado de Rondônia foi estudada a fauna parasitaria de C. macropomum
criados em tanques escavados utilizados para engorda e pesque-pague. Os peixes
pesavam em média 907 g. Nenhum N. (N.) buttnerae foi encontrado (Godoi et al. 2012).
No Estado do Amapá, C. macropomum criados oito tanques também não foi encontrado
nenhum N. (N.) buttnerae (Dias et al. 2015).

31
A fauna parasitaria foi avaliada em C. macropomum criados em tanques-rede no
Estado do Amapá e não foi encontrada nenhuma espécie de parasitas intestinais. Os
peixes pesavam 2 kg em média (Santos et al. 2013). No Estado do Amazonas, próximo
a Manaus, a fauna parasitaria de C. macropomum criados em tanque-rede foi analisada,
e nenhum N. (N.) buttnerae foi encontrado (Morais et al. 2009).
A fauna parasitaria de 105 C. macropomum, criados em tanque-rede, foi
analisada. Provenientes do município de Iranduba próximo a Manaus. Apenas um N.
(N.) buttnerae foi encontrado (Varella et al. 2003). Neste trabalho os C. macropomum
criados em tanques escavados na terra as infestações por N. (N.) buttnerae foram muito
altas, a prevalência foi de 100%. A ausência de N. (N.) buttnerae nos outros trabalhos
está relacionada à ausência do hospedeiro intermediário no ambiente, sem o qual, N.
(N.) buttnerae não pode completar o seu ciclo de vida.
No rio Amazonas e Solimões foram feitas capturas de C. macropomum na região
de Tefé/Coari estado do Amazonas e de Santarém no Estado do Pará, para o estudo de
sua fauna de parasitas em ambiente natural. A prevalência foi de 78% e a intensidade
média 98, nos peixes da região de Tefé/Coari. E de 55% e 16 respectivamente nos
peixes de Santarém (Fischer et al. 2003).
Em uma fazenda de criação de peixes foram adquiridos 25.000 C. macropomum
com 1 a 3 cm de comprimento total e tinham 30 dias de vida. Após 60 dias foram
colocados em quatro barragens com cerca de 1 hectare cada. Após cinco meses os C.
macropomum os peixes pararam de comer e começaram a morrer. Os peixes foram
analisados e estavam com uma alta infestação de N. (N.) buttnerae. A prevalência era de
100%, a intensidade variou de 30 a 406 e a intensidade média de 125,6 acantocéfalos
por peixe. Não foi possível debelar a infestação e todos os peixes tiveram que ser
sacrificados (Malta et al. 2001).
Em coletas realizadas em tanques escavados, no estado de Rondônia e
Amazonas, para estudo de histopatologia de N. (N.) buttnerae em C. macropomum, foi
encontrada prevalência de 100% nos dois locais e intensidade média de 262 em
Rondônia e 476 no Amazonas e peso dos peixes de 30 g a 2 kg (Jerônimo et al. 2017).
Outro estudo de histopatologia com mesma espécie em viveiro escavado próximo à
Manaus encontrou prevalência de 100% em juvenis com média de 57 g. com
intensidade média de 240, 4 (Matos et al. 2017).

32
Neste trabalho a prevalência foi de 100%, como aconteceu em outros C.
macropomum criados em tanques escavados ou em barragens onde ocorreram
infestações maciças por N. (N.) buttnerae. Em C. macropomum capturados no ambiente
natural a prevalência e a intensidade média foram menores de 78% e 98 para peixes do
rio Solimões e de 55% e 16 para os do rio Amazonas Fischer et al. (2003).
Nos C. macropomum criados em tanques escavados ou em barragens foram
criadas as condições ideais para a explosão numérica das populações de N. (N.)
buttnerae. Nos tanques havia uma grande concentração de C. macropomum, os
hospedeiros definitivos, e estes foram adquiridos parasitados com os adultos de N. (N.)
buttnerae. Nos tanques onde foram colocados os peixes existiam populações da espécie
de Ostracoda que é a hospedeira intermediária. E nos últimos 20 anos esta história de
parasitismo vêm se repetindo nas criações de C. macropomum.
Paulisentis missouriensis é um Acantocephala da família Neoechinorhynchidae,
tem como hospedeiro definitivo Semotilus atromaculatus (Mitchill, 1818). O seu
hospedeiro intermediário é o Copepoda Cyclops vernalis Fischer, 1853 (Keppner 1974).
Paulisentis fractus também utiliza a mesma espécie de peixe como hospedeiro
definitivo, mas utiliza um Copepoda de outra espécie como único hospedeiro
intermediário Tropocyclops prasinus (Fischer, 1860) (Cable e Dill 1967).
Octospiniferoides chandleri, também da família Neoechinorhynchidae, tem
como hospedeiro definitivo Gambusia affinis (Baird e Girard, 1853). Seus hospedeiros
intermediários são duas espécies de Ostracoda, Cypridopsis vidua (Müller, 1776) e
Physocypria pustulosa (Sharpe, 1897) (DeMont e Corkum 1982). Neoechinorhynchus
(Neoechinorhynchus) cristatus parasita de Catostomus macrocheilus Girard, 1856
utiliza como hospedeiro intermediário um Ostracoda Cypridopsis helvetica Kaufmann,
1900 (Uglem 1972).
Neoechinorhynchus saginatus parasita S. atromaculatus, utiliza o Ostracoda C.
vidua como hospedeiro intermediário (Uglem e Larson 1969). Neoechinorhynchus
(Neoechinorhynchus) emydis parasita da tartaruga Graptemys geographica (LeSueur,
1817), necessita de dois hospedeiros intermediários, o Ostracoda Cypria maculata Hoff,
1942 como primeiro hospedeiro intermediário e o molusco Campeloma rufum
(Haldeman, 1841) como segundo hospedeiro intermediário (Hopp 1954).

33
Neoechinorhynchus (Neoechinorhynchus) cylindratus tem como hospedeiro
definitivo Huro salmoides (Lacepède, 1802), e possui dois hospedeiros intermediários.
O primeiro hospedeiro intermediário, o Ostracoda Cypria globula Furtos, 1933 e como
segundo hospedeiro intermediário o peixe Lepomis pallidus (Ward 1940).
Neoechinorhynchus (Neoechinorhynchus) rutili encontrado na forma adulta em espécies
de peixes de catorze famílias, utiliza o Ostracoda Cypria turneri Hoff, 1942 como
hospedeiro intermediário (Merritt e Pratt 1964).
Neoechinorhynchus (Neoechinorhynchus) iraqensis parasito de Liza abu
(Heckel, 1843) e outros peixes de água doce do Iraque. O hospedeiro intermediário é o
Copepoda Cyclops hyalinus Rehberg, 1880 (Al-Sady 2009).
O uso de espécies de Ostracoda como hospedeiro intermediário é comum para as
espécies do gênero Neoechinorhynchus Stiles & Hassall, 1905. Das sete espécies do
gênero com ciclo de vida descrito, apenas uma utiliza uma espécie de Copepoda como
hospedeiro intermediário, o N. (N.) iraqensis (Al-Sady 2009). Ostracodas são crustáceos
conhecidos por serem bentônicos ou pelágicos, o que pode diminuir a predação por
peixes filtradores como o tambaqui.
Neste trabalho o hospedeiro intermediário de N. (N.) buttnerae parasito do C.
macropomum foi uma espécie de Ostracoda. Como C. macropomum se alimenta, em
uma parte do ano diretamente de Ostracoda no ambiente natural (Goulding e Carvalho
1982) e, como nos tanques escavados e barragens sempre tem esta espécie de
Ostracoda, que é o hospedeiro intermediário, a ação antrópica facilitou o fechamento do
ciclo de vida nas fazendas de criação de C. macropomum na Amazônia.
Outras espécies do gênero Neoechinorhynchus utilizam a estratégia de dois
hospedeiros intermediários por conta dos hábitos alimentares de seus hospedeiros
definitivos. Neoechinorhynchus (Neoechinorhynchus) emydis Van Cleave, 1919
parasito de Graptemys geographica Lesueur, 1817, uma tartaruga de água doce norte-
americana, precisa utilizar um segundo hospedeiro intermediário, no caso um caramujo,
que se alimenta de ostracodas (primeiro hospedeiro intermediário) (Hopp 1954).
As larvas acântor de N. (N.) cristatus eclodidas artificialmente, não tinham
ganchos e espinhos laterais. Mas quando elas eclodiam naturalmente apresentavam estas
ornamentações (Uglem 1972). Neste trabalho não foi possível observar a larva acântor

34
de N. (N.) buttnerae eclodida naturalmente, só foi possível observar as eclodidas
artificialmente, por isso a ausência dos ganchos e espinhos laterais.
A acantela de fase 2 e 3, com 4 e 6 dias respectivamente, de N. (N.) buttnerae é
mais similar à de N. saginatus relatada por Uglem e Larson (1969) quando comparada à
outras espécies. Ambas possuem um formato mais romboidal como primeira fase
acantela e um entoblasto mais amplo comparado com o da fase de acântor, com
diferenciação dos chamados núcleos gigantes que formam o lemnisco e os órgãos
reprodutores.
Quanto à próxima fase 4 com 9 dias de desenvolvimento se assemelha bastante
com a acantela de 10 dias de N. saginatus. Onde acontece a separação do entoblasto e a
formação do pseudoceloma, junto com o receptáculo da probóscide, onde fica inserido a
massa celular que dá origem ao gânglio cerebral (Uglem e Larson 1969).
Na larva de fase 5, de 13 dias, além do receptáculo e do gânglio cerebral,
encontra-se formado o lemnisco, e outras células na região anterior que formarão a
probóscide e órgãos relacionados. Esta fase se assemelha a larva de 28 dias de N. (N.)
cylindratus com o receptáculo formado e células geradoras da probóscide no ápice da
região anterior (Merritt e Pratt 1964).
A larva de fase 6 com 15 dias de desenvolvimento se assemelha à de N. rutili
com 38 dias no que se refere a formação da evaginação de uma bursa, o que indica ser
um macho. Em Neoechinorhynchus (N.) rutili foram observados os testículos e as
glândulas de cemento nesta fase (Merritt e Pratt 1964). Em N. (N.) buttnerae eles não
foram observados ainda nessa fase.
Na fase 7 com 17 dias de desenvolvimento, pode-se notar um lemnisco mais
desenvolvido, e a probóscide parcialmente formada com ganchos aparentes. Em outras
espécies como N. (N.) iraqensis a probóscide já estava diferenciada com 8 a 10 dias de
desenvolvimento (Al-Sady 2009).
Com 20 dias de desenvolvimento a larva de fase 8, encontra-se com o aparelho
reprodutor parcialmente formado, e com a probóscide com capacidade de contração. Em
P. fractus a fase semelhante a essa se dá com 7 dias de desenvolvimento larval, onde os
órgãos sexuais de machos e fêmeas já são bem distinguíveis (Cable e Dill 1967).
Na fase 9 com 23 dias de desenvolvimento, a larva encontra-se com probóscide
totalmente formada e com tamanho reduzido. Em N. (N.) emydis também ocorre uma

35
ligeira diminuição do comprimento da larva de 14 e de 16 dias de desenvolvimento. Isto
foi atribuído pelos autores à formação do sistema muscular que permite à larva se
contrair (Hopp 1954).
A fase juvenil com 26 dias é muito semelhante ao adulto, com os órgãos sexuais
já formados, pronto para desenvolver o cisto que recobre o corpo. Em outras espécies
como N. (N.) emydis, essa fase se dá com 21 dias de desenvolvimento (Hopp 1954), 12
dias em N. (N.) iraqensis (Al-Sady 2009), 14 dias em N. saginatus (Uglem e Larson
1969).
Em N. (N.) emydis a membrana cobre todo o corpo do juvenil infectante
enrolado no caramujo (Hopp 1954). No caso de N. (N.) cylindratus, o juvenil também se
encontra enrolado dentro da membrana ou cisto e os autores descrevem o cisto como de
cor branca, sendo a esta membrana formada por células do fígado modificadas. Este
cisto é encontrado no fígado do segundo hospedeiro intermediário (Ward 1940).
A fase infectante de N. (N.) buttnerae não foi encontrada no experimento onde a
espécie de Ostracoda ingeriu os ovos. Mas foi encontrada nas amostras de plâncton
coletadas na água do tanque escavado. A membrana que recobre o corpo do juvenil
recobre apenas uma parte do corpo, sendo diferente das duas citadas no parágrafo
anterior. As outras espécies de Neoechinorhynchus com larvas descritas, não
apresentam membrana na fase infectante. Assim como N. (N.) buttnerae estas espécies
possuem apenas um hospedeiro intermediário.
O tempo encontrado para se alcançar a fase infectante de N. (N.) buttnerae foi
um pouco mais longo que de outras espécies do mesmo gênero, com exceção de N. (N.)
rutili que levou de 48 a 57 dias (Merritt e Pratt 1964). Neoechinorhynchus
(Neoechinorhynchus) iraqensis levou de 13 a 14 dias (Al-Sady 2009), 21 dias para N.
emydis (Hopp 1954), 16 dias para N. saginatus (Uglem e Larson 1969) e N. (N.)
cristatus 20 dias (Uglem 1972). Todas estas espécies utilizaram uma espécie de
Ostracoda como primeiro hospedeiro intermediário, com exceção de N. iraqensis.
Esta diferença de duração no desenvolvimento da larva pode estar relacionado
com o tamanho delas. As fases de acantela de N. (N.) buttnerae são consideravelmente
maiores que as larvas de outras espécies do mesmo gênero, o que está em proporção ao
tempo de desenvolvimento também, como visto na tabela 1. Isto não vale por exemplo
para N. (N.) rutili, que as larvas são menores que as de N. (N.) buttnerae, mas possui um

36
tempo de desenvolvimento maior 48 a 57 dias, enquanto o N. (N.) buttnerae
aproximadamente 30 a 35 dias.
As fêmeas do gênero Neoechinorhynchus na fase adulta tendem a ser maiores
que os machos, como mostra a tabela 1. A fêmea adulta de maior comprimento do
gênero foi N. (N.) iraqensis, com 53 mm, seguida pela fêmea de N. (N.) buttnerae com
30 mm. A fase adulta de N. (N.) iraqensis é consideravelmente maior que a das outras
espécies (Al-Sady 2009), mas em contrapartida, suas larvas são menores que as de N.
(N.) buttnerae, mesmo sendo maiores que as das outras espécies.
Quanto às fases larvais de N. (N.) cristatus, suas medidas não estão incluidas na
tabela. Seu desenvolvimento larval é semelhante ao de N. rutilli. Com exceção ao seu
tempo de desenvolvimento que leva 20 dias para chegar à fase infectante, enquanto o N.
(N.) rutilli leva de 48 a 57 dias (Uglem 1972).
A espécie do gênero com mais fases larvais descritas é a de N. (N.) emydis, com
doze fases de acantela descritas, mais o acantor e juvenil (Hopp 1954). Mas estas larvas
são notavelmente menores que as outras espécies, especialmente de N. (N.) buttnerae.
Seu tempo de desenvolvimento também é relativamente curto, com 21 (Hopp 1954).
Octospiniferoides chandleri pode influenciar comportamento fotofílico no
hospedeiro intermediário o Ostracoda C. vidua, tornando-o mais susceptível à predação.
O Ostracoda P. pustulosa infectado com N. (N.) cylindratus, a maioria dos indivíduos
parasitados se encontram na superfície, sendo facilmente capturados com rede de
plâncton na superfície, enquanto que apenas 1,83% dos P. pustulosa encontrados no
fundo estavam parasitados (DeMont e Corkum 1982).
Espécies de Amphipoda também sofrem influência no seu comportamento
quando hospedeiras intermediárias de Acantocephala. Gammarus pulex (Linnaeus,
1758), infectado por Pomphorhynchus laevsi (Müller, 1776) e Polymorphus minutus
(Zeder, 1800) tem seu comportamento alterado, apresentando uma maior afinidade com
a luz, sendo este comportamento chamado de fotofílico (Cézilly et al. 2000).
Foi testada a predação de Amphipoda Gammarus lacustris Sars, 1863 pelo pato
selvagem do Canadá. Nos experimentos, notaram que os patos se interessavam mais
pelos Amphipoda que estavam infectados por cistacanto de Polymorphus paradoxos
Connell and Corner, 1957, pois estes tinham comportamento fotofílico e de evasão
alterado, além de apresentarem cor alaranjada (Bethel e Holmes 1977).

37
Estas alterações comportamentais favorecem a predação dos hospedeiros
intermediários, pois o comportamento é alterado de tal forma que facilita os mesmos
serem predados (Moore 1984). Apesar de serem bentônicos, foi observado neste
trabalho em aquário que alguns indivíduos nadavam próximos à superfície. O C.
macropomum como peixe filtrador, pode comer mais facilmente um Ostracoda
infectado com juvenil infectante de N. (N.) buttnerae presente no plâncton.
Os Ostracoda são conhecidos por se reproduzirem tanto sexuadamente, quanto
assexuadamente, ou seja, as fêmeas podem ser partenogenéticas e colocar ovos, mesmo
sem a presença de machos no ambiente. A reprodução sexuada de Ostracoda gera ovos
de resistência que podem durar meses, ou até décadas em solo seco e serem eclodidos
com as condições certas, como reidratação e luz (Horne et al. 2002).
Estes ovos podem ser resistentes a estações como seca ou inverno, podendo
sobreviver por muito tempo. Este mecanismo permitiu que os Cypridoidea, principal
(Super-familia de Ostracoda de água Doce), tivessem sucesso evolutivo no período
quaternário, com maior distribuição e riqueza em ambientes não-marinhos (Horne e
Martens 1998).
O fato de microcrustáceos terem essa adaptação os torna bastante resistentes em
ambientes que costumam passar por secas no ambiente natural. Em tanques de
piscicultura o produtor precisa secar o viveiro após a despesca, para que haja oxidação
da matéria orgânica no fundo, evitando assim que o excesso desse material provoque
uma diminuição na concentração de oxigênio do tanque (Ostrensky e Boeger 1998).
Depois de aproximadamente duas semanas o produtor os enche novamente, e
os ovos de resistência que tenham ficado no sedimento eclodem, assim os Ostracoda e
outros microcrustáceos voltam a povoar esse tanque. Isso quer dizer que as populações
do hospedeiro intermediário continuam no viveiro, mesmo após o mesmo ter sido
esvaziado.
As técnicas utilizadas para se desinfectar um viveiro quando seco consistem
basicamente em uso de cal virgem (CaO) ou cal hidratada (Ca(OH)2, que matam
microosganismos com o aumento da temperatura e pH, ou apenas com aumento de pH,
respectivamente (Ostrenksky e Boeger 1998). Esta técnica é amplamente utilizada pelos
piscicultores no Brasil e não tem se mostrado efetiva para os hospedeiros intermediários
de N. (N.) buttnerae.

38
Quanto à forma de proliferação do hospedeiro intermediário em tanques que
antes não o tinham, pode-se considerar o uso do sedimento transportado para a
construção do tanque. Os ovos de resistência podem resistir por períodos relativamente
longos e serem eclodirem em outros lugares onde haja luz e água, no caso, novos
viveiros.
Outra forma pode ser o transporte de alevinos ou juvenis, de tanques
contaminados para outros não contaminados, pois além dos peixes estarem parasitados
por N. (N.) buttnerae, a água utilizada no transporte pode estar carregando o seu
hospedeiro intermediário. Neste trabalho, a água utilizada para transportar os peixes do
viveiro para o laboratório foi analisada, e constatou-se a presença dos hospedeiros
intermediários nesta água.
Bibliografia citada
Alves, J.M. 2000. Monitoramento da parasitofauna de alevinos de tambaqui,
Colossoma macropomum (Cuvier, 1818), em barragens de igarapé de terra firme
no município de Iranduba, - AM. Dissertação de Mestrado, Instituto Nacional de
Pesquisas da Amazônia, Manaus, Amazonas. 71p.
Al-Sady, R.S. 2009. The Life Cycle and Larval Development of Neoechinorhynchus
iraqensis. Al-Haitham Journal for Pure and Applied Science, 22: 9-14.
Amato, J.F.R.; Boeger, W.A.; Amato, S.B. 1991. Protocolos para laboratório coleta e
processamento de parasitas do pescado. Imprensa Universitária, Universidade
Federal do Rio de Janeiro, Rio de Janeiro, Brasil. 81p.
Amin, O.M. 1969. Helminthes Fauna of Suckers (Catostomidae) of the Gila River
System, Arizona. II. Five Parasites from Catostomus spp. American Midland
Naturalist, 82: 429-443.
Araújo-Lima, C.A.R.M.; Goulding, M. 1998. Os frutos do tambaqui: ecologia,
conservação e cultivo na Amazônia. Tefé: Sociedade Civil Mamirauá; Brasília:
CNPq, 186p.
Arbeláez-Rojas, G.A.; Fracalossi, D.M.; Fim, J.D.I. 2002. Body composition of
tambaqui, Colossoma macropomum, and matrinxã, Brycon cephalus, when raised

39
in intensive (igarapé channel) and semi-intensive (pond) culture systems. Revista
Brasileira de Zootecnia, 31: 1059-1069.
Batista V.S.; Petrere M. Jr. 2003 Characterization of the commercial fish production
landed at Manaus, Amazonas State, Brazil. Acta Amazonica. 33: 53–66.
Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. 1997. Parasitology meets
ecology on its own terms: Margolis et al. 1982. Revisited. Journal of
Parasitology, 83: 575-583.
Cable, R.M.; Dill, W.T. 1967. The morphology and life history of Paulisentis fractus
Van Cleave & Bangham, 1949 (Acanthocephala: Neoechinorhynchidae)." The
Journal of Parasitology, 53: 810-817.
Carvalho, M.L. 1981. Alimentação do tambaqui jovem (Colossoma macropomum
Cuvier, 1818) e sua relação com a comunidade zooplanctônica do lago Grande-
Manaquiri, Solimões-AM. Dissertação de Mestrado, Instituto Nacional de
Pesquisas da Amazônia/Fundação Universidade do Amazonas, Manaus,
Amazonas. 90p.
Cézilly, F.; Grégoire, A.; Bertin, A. 2000. Conflict between co-occurring manipulative
parasites? An experimental study of the joint influence of two acanthocephalan
parasites on the behaviour of Gammarus pulex. Parasitology, 120, 625-630.
Costa, J.I.D. 2013. Avaliação econômica e participação do plâncton no cultivo de
tambaqui em viveiros com diferentes densidades de estocagem. Dissertação de
Mestrado, Universidade Estadual Paulista, Jaboticabal, São Paulo. 80p.
DeMont, D.J.; Corkum, K.C. 1982. The life cycle of Octospiniferoides chandleri
Bullock, 1957 (Acanthocephala: Neoechinorhynchidae) with some observations
on parasite-induced, photophilic behavior in ostracods. The Journal of
Parasitology, 125-130.
Dias, M.K.R.; Neves, L.R.; Marinho, R.G.B.; Tavares-Dias, M. 2015. Parasitic
infections in tambaqui from eight fish farms in Northern Brazil. Arquivo
Brasileiro de Medicina Veterinária e Zootecnia, 67, 1070-1076.
Eiras, J.C. 1994. Elementos de Parasitologia. Fundação Eng. Antônio de Almeida,
Porto, Portugal. 339p.

40
Eiras, J.C.; Takemoto, R.M.; Pavanelli, G.C. 2006. Métodos de estudo e técnicas
laboratoriais em parasitologia de peixes. 2 ed. Eduem, Maringá, Brasil. 199
Fischer, C.; Malta, J.C.O.; Varella, A.M.B. 2003. The fauna of parasites of the
tambaqui, Colossoma macropomum (Cuvier, 1818) (Characiformes: Characidae)
from middle Solimões River and lower Amazonas River and their potential as
biological indicators. Acta Amazonica, 33: 651-662.
Forsberg, B.R.; Araujo-Lima, C.A.R.M.; Martinelli, L.A.; Victoria, R.L.; Bonassi, J.A.
1993. Autotrophic carbon sources for fish of the central Amazon. Ecology, 74:
644-652.
Godoi, M.M.I.D.M.; Engracia, V.; Lizama, M.D.L.A.P.; Takemoto, R.M. 2012.
Parasite-host relationship between the tambaqui (Colossoma macropomum Cuvier
1818) and ectoparasites, collected from fish farms in the City of Rolim de Moura,
State of Rondônia, Western Amazon, Brazil. Acta Amazonica, 42: 515-524.
Golvan, Y.J. 1956. Acanthocéphales d'Amazonie. Redescription d'Oligacanthorhynchus
iheringi Travassos, 1916 et description de Neoechinorhynchus buttnerae n. sp.
(Neoacanthocephala-Neoechinorhynchidae). Annales de Parasitologie, 31: 500-
524.
Gomes, L.C.; Roubach, R.; Araujo‐Lima, C.A.; Chippari‐Gomes, A.R.; Lopes, N.P.;
Urbinati, E.C. 2003. Effect of fish density during transportation on stress and
mortality of juvenile tambaqui Colossoma macropomum. Journal of the World
Aquaculture Society, 34: 76-84.
Goulding M. 1979. Ecologia da Pesca do Rio Madeira. CNPq-INPA, Manaus, Brazil.
172 pp
Goulding, M. 1980. The fishes and the forest: explorations in Amazonian natural
history. University of California Press, Berkeley, USA. 280p.
Goulding, M. 2013. Man and fisheries on an Amazon frontier (Vol. 4). Springer Science
& Business Media. Dordrecht, Netherlands. 137p.
Goulding, M.; Carvalho, M.L. 1982. Life history and management of the tambaqui
(Colossoma macropomum, Characidae) an important Amazonian food
fish. Revista Brasileira de Zoologia, 1: 107-133.

41
Hopp, W.B. 1954. Studies on the morphology and life cycle of Neoechinorhynchus
emydis (Leidy), an acanthocephalan parasite of the map turtle, Graptemys
geographica (Le Sueur). The Journal of parasitology, 40: 284-299.
Horne, D.J.; Cohen, A.; Martens, K. 2002. Taxonomy, morphology and biology of
Quaternary and living Ostracoda. The Ostracoda: applications in Quaternary
research, 131: 5-36.
Horne, D.J.; Martens, K. 1998. An assessment of the importance of resting eggs for the
evolutionary success of Mesozoic non-marine cypridoidean Ostracoda
(Crustacea). Archiv für Hydrobiologie, 52, 549-561.
IBGE, 2015. Censo Agropecuário, Estados, Amazonas
(http://www.ibge.gov.br/estadosat/temas.php?sigla=am&tema=pecuaria2015).
Acessed on 24/02/2017.
Jerônimo, G.T.; de Pádua, S.B.; de Andrade Belo, M.A.; Chagas, E.C.; Taboga, S. R.;
Maciel, P. O.; Martins, M. L. 2017. Neoechinorhynchus buttnerae
(Acanthocephala) infection in farmed Colossoma macropomum: A pathological
approach. Aquaculture, 469, 124-127.
Junk, W.J. 1970. Investigations on the ecology and production-biology of the “floating
meadows” (Paspalo-Echinochloetum) on the Middle Amazon. Amazoniana, 2:
449-495.
Junk, W.J.; Bayley, P.B.; Sparks, R.E. 1989. The flood pulse concept in river-floodplain
systems. Canadian special publication of fisheries and aquatic sciences, 106:
110-127.
Kennedy, C.R.; C. Moriarty. 2002. Long-term stability in the richness and structure of
helminth communities in eels, Anguilla anguilla, in Lough Derg, River Shannon,
Ireland. Journal of helminthology 76: 315-322.
Kennedy, C.R.; Burrough, R.J. 1978. Parasites of trout and perch in Malham Tarn. Field
Stud, 4: 617-619.
Kennedy, C.R. 2006. Ecology of the Acanthocephala. Cambridge University Press. New
York, USA. 249p.

42
Keppner, E.J. 1974. "The life history of Paulisentis missouriensis n.
sp.(Acanthocephala: Neoechinorhynchidae) from the creek chub Semotilus
atromaculatus." Transactions of the American Microscopical Society,93: 89-100.
Kussat, R.H. 1969. A comparison of aquatic communities in the Bow River above and
below sources of domestic and industrial wastes from the city of
Calgary. Canadian Fisheries and Cultures, 40: 3-31.
Lopes, J.M.; Pascoal, L.A.F.; Silva Filho, F.P.D.; Santos, I.B.; Watanabe, P.H.; Araujo,
D.M.; Pinto, D.C; Oliveira, P.D.S. 2010. Farelo de babaçu em dietas para
tambaqui. Revista Brasileira de Saúde e Produção Animal, 11: 519-526.
Lowe-McConnell, R.H. 1975. Fish communities in tropical freshwaters. In Fish
communities in tropical freshwaters. Longman.
Macedo, C.F.; Sipaúba-Tavares, L.H. 2005. Comunidade planctônica em viveiros de
criação de peixes em disposição sequencial. Boletim do Instituto de Pesca, 31, 21-
27.
Malta, J.C.O. 1984. Os peixes de um lago de várzea da Amazônia Central (Lago
Janauacá, Rio Solimões) e suas relações com os crustáceos ectoparasitas
(Branchiura: Argulidae). Acta Amazonica, 14: 355-372.
Malta, J.D.O.; Gomes, A.L.S.; de Andrade, S.M.S.; Varella, A.M.B. 2001. Massive
infestation by Neoechinorhynchus buttnerae Golvan, 1956 (Eoacanthocephala:
Neochinorhynchidae) in young" tambaquis" Colossoma macropomum (Cuvier,
1818) cultured in the Central Amazon. Acta Amazonica, 31: 133-143.
Malta, J.C.O.; Varella, A.M.B. 1983. Os argulídeos (Crustacea: Argulidae) da
Amazônia Brasileira. Aspectos da ecologia de Dolops striata (Bouvier, 1899) e D.
carvalhoi Lemos de Castro, 1949. Acta Amazonica, 13: 299-306.
Malta, J.C.O.; Varella, A.M.B. 2000. Argulus chicomendesi sp. n. (Crustacea:
Argulidae) parasita de peixes da Amazônia brasileira. Acta Amazonica, 30: 481-
498.
Marcogliese, D.J. 2005. Parasites of the superorganism: are they indicators of
ecosystem health? International journal for parasitology, 35: 705-716.
Marlier, G. 1967. Ecological studies on some lakes of the Amazon valley.
Amazoniana, 1: 91-115.

43
de Matos, L.V.; de Oliveira, M.I.B.; Gomes, A.L.S.; da Silva, G.S. 2017. Morphological
and histochemical changes associated with massive infection by
Neoechinorhynchus buttnerae (Acanthocephala: Neoechinorhynchidae) in the
farmed freshwater fish Colossoma macropomum Cuvier, 1818 from the Amazon
State, Brazil. Parasitology Research, 1-9.
Mello, F.D.; Oliveira, C.A.; Ribeiro, R.P.; Resende, E.K.; Povh, J.A.; Fornari, D.C.;
Barreto, RV.; Mcmanus, C e Streit Jr, D. 2015. Growth curve by Gompertz
nonlinear regression model in female and males in tambaqui (Colossoma
macropomum). Anais da Academia Brasileira de Ciências, 87, 2309-2315.
Merritt, S.V.; Pratt, I. 1964. The life history of Neoechinorhynchus rutili and its
development in the intermediate host (Acanthocephala:
Neoechinorhynchidae). The Journal of parasitology, 50: 394-400.
Moore, J. 1984. Altered Behavioral Responses in Intermediate Hosts--An
Acanthoceptalan Parasite Strategy. The American Naturalist, 123, 572-577.
Morais, A.M.; Varella, A.M.B.; Villacorta-Correa, M.A.; Malta, J.C.O. 2009. Fauna de
parasitos em juvenis de tambaqui Colossoma macropomum (Cuvier, 1818)
(Characidae: Serrasalminae) criados em tanques-rede em lago de várzea da
Amazônia central. Biologia Geral e Experimental. 9:14-23.
Ostrensky, A.; Boeger, W. A. 1998. Piscicultura: fundamentos e técnicas de manejo.
Guaíba: Agropecuária, 211 p.
Overstreet, R.M.; Howse, H.D. 1977. Some parasites and diseases of estuarine fishes in
polluted habitats of Mississippi. Annals of the New York Academy of
Sciences, 298: 427-462.
Pereira-Junior, G.P.; de Oliveira Pereira, E.M.; Pereira-Filho, M.; de Sousa Barbosa, P.;
Shimoda, E.; Brandão, L.V. 2013. Desempenho produtivo de juvenis de tambaqui
(Colossoma macropomum Cuvier, 1818) alimentados com rações contendo
farinha de crueira de mandioca (Manihot esculenta, Crantz) em substituição ao
milho (Zea mays). Acta Amazonica, 43: 217-226.
Poulin, R. 2005. Relative infection levels and taxonomic distances among the host
species used by a parasite: insights into parasite specialization. Parasitology, 130:
109-115.

44
Poulin, R.; Mouillot, D. 2003. Parasite specialization from a phylogenetic perspective: a
new index of host specificity. Parasitology, 126: 473-480.
Santos, E.F.; Tavares-Dias, M.; Pinheiro, D.A.; Neves, L.R.; Marinho, R.D.G.B.; Dias,
M.K.R. 2013. Parasitic fauna of tambaqui Colossoma macropomum (Characidae)
farmed in cages in the State of Amapá, eastern Amazon. Acta Amazonica, 43,
105-111.
Sioli, H. 1976. A limnologia na região Amazônica brasileira. p. 153-165. In: Anais do I
encontro nacional sobre limnologia, piscicultura e pesca continental. Fundação
João Pinheiro, Diretoria de Meio Ambiente, Centro de Recursos Naturais. Belo
Horizonte, Brasil.
Sioli, H. 1984. The Amazon and its main affluents: hydrography, morphology of the
river courses, and river types. p. 127-165, In: The Amazon. Springer, Dordrecht,
Netherlands.
Thatcher, V.E. 2006. Aquatic Biodiversity in Latin America: Amazon Fish Parasites.
2nd edition, Pensoft Publishers, Praga, Bulgaria, 508p.
Uglem, G.L.; Larson, O.R. 1969. The life history and larval development of
Neoechinorhynchus saginatus Van Cleave and Bangham, 1949 (Acanthocephala:
Neoechinorhynchidae). The Journal of Parasitology, 55: 1212-1217.
Uglem, Gary L. 1972. The life cycle of Neoechinorhynchus cristatus Lynch, 1936
(Acanthocephala) with notes on the hatching of eggs. The Journal of parasitology,
58: 1071-1074.
Varella, A.M.B.; Peiro, S.N.; Malta, J.C.O.; Lourenço J. N. P. 2003. Monitoramento da
parasitofauna de Colossoma macropomum (Cuvier, 1818) (Osteichthyes:
Characidae) cultivados em tanques-rede em um lago de várzea na Amazônia,
Brasil, p. 95-105. In: Urbinati, E.C.; Cyrino, J.E.P. (Eds.). Anais do Simpósio
Brasileiro de Aqüicultura, Jaboticabal, São Paulo.
Vidal-Martínez, V.M.; Aguirre-Macedo, M.L.; Noreña-Barroso, E.; Gold-Bouchot, G.;
Caballero-Pinzón, P. I. 2003. Potential interactions between metazoan parasites of
the Mayan catfish Ariopsis assimilis and chemical pollution in Chetumal Bay,
Mexico. Journal of Helminthology, 77: 173-184.

45
Ward, H.L. 1940 "Studies on the life history of Neoechinorhynchus cylindratus (Van
Cleave, 1913) (Acanthocephala)." Transactions of the American Microscopical
Society 59: 327-347.