histologÍa de hoja y comparaciÓn de epidermis...
TRANSCRIPT

HISTOLOGÍA DE HOJA Y COMPARACIÓN DE EPIDERMIS FOLIAR DE
Gaultheria anastomosans PRESENTE EN BOSQUE DE NIEBLA Y PÁRAMO DEL
PARQUE ECOLÓGICO MATARREDONDA, BOGOTÁ (COLOMBIA)
PAULA NATALIA AMARANTA NEIRA MORENO
Grupo de Investigación de Ecología y Conservación de Plantas de Colombia
(GIECPC)
UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS
FACULTAD DE CIENCIAS Y EDUCACIÓN
PROYECTO CURRICULAR DE LICENCIATURA EN BIOLOGÍA
BOGOTÁ D.C.
2018

2
HISTOLOGÍA DE HOJA Y COMPARACIÓN DE EPIDERMIS FOLIAR DE
Gaultheria anastomosans PRESENTE EN BOSQUE DE NIEBLA Y PÁRAMO DEL
PARQUE ECOLÓGICO MATARREDONDA, BOGOTÁ (COLOMBIA).
PAULA NATALIA AMARANTA NEIRA MORENO
PROPUESTA DE TRABAJO DE GRADO BAJO LA MODALIDAD
INVESTIGACIÓN – INNOVACIÓN PARA OPTAR POR EL TÍTULO DE
LICENCIADA EN BIOLOGÍA
Director
GERMÁN ANTONIO NIÑO GALEANO
Codirectora
MERY HELEN TÍJARO OREJUELA
UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS
FACULTAD DE CIENCIAS Y EDUCACIÓN
PROYECTO CURRICULAR DE LICENCIATURA EN BIOLOGÍA
BOGOTÁ D.C.
2018

3
Dedicatoria
A Patricia, Alejandro, Luisa, Susana, Tatiana, Martín, Emilia, Sonia y Jorge;
porque la familia es lo más importante.
Agradecimientos
A mi directora Mery Helen Tíjaro Orejuela por siempre creer en mí e influenciarme a
realizar proyectos. Gracias a ella por llevarme tan lejos.
A mi familia; mis papas Alejandro y Patricia, mis hermanas Luisa, Susana y Tatiana, mi
abuela Sonia, mis sobrinos Martín y Emilia y a mi pareja Jorge; fuente de toda inspiración
para siempre dar lo mejor, gracias por su apoyo en todo este proceso y por creer siempre en
mi a ciegas. Por ellos y para ellos todo lo que hago.
A la profesora Carmen Helena Moreno por abrirnos las puertas de su grupo de investigación
y convidarnos todos sus equipos, gracias a eso realizamos este proyecto y los anteriores.
A Lizdey Cárdenas por entrenarme en el arte de la histología vegetal. Al director del Grupo
de Investigación en Ecología y Conservación de Plantas de Colombia (GIECPC), German
Niño, por enseñarme y ayudarme en todo este proyecto. Al profesor Oscar Mahecha por
colaborarme y brindarme su conocimiento sin restricción alguna. A María Paula Lemus,
Helena Erazo, Sebastián Vélez, Camilo Forero, Io Natali Araujo y David Aroca por siempre
colaborarme, apoyarme, animarme y estar cuando las fuerzas se agotan. Al Parque Ecológico
de Matarredonda por abrirnos las puertas y dejarnos entrar al magnifico mundo de los
Páramos.
Y por último a la Universidad Distrital Francisco José de Caldas y a cada uno de los
profesores que contribuyeron en mi correcta formación como Licenciada en biología.

4
TABLA DE CONTENIDO
1. INTRODUCCIÓN…..…………………………………………………………..……5
2. DESCRIPCIÓN DEL PROBLEMA...………………………………………………..7
2.1.Preguntas problema……………………………………………………………….7
2.2.Planteamiento del problema………………………………………………………7
3. JUSTIFICACIÓN……………………...……………………………………………...8
4. OBJETIVOS…………..………………………………………………………………9
4.1.Objetivo general…………………………………………………………………..9
4.2.Objetivos específicos……………………………………………………………...9
5. ESTADO DEL ARTE……………………………………………………………….10
5.1.Antecedentes…………………………………………………………………….10
5.2.Marco espacial…………………………………………………………………...11
5.2.1. Parque ecológico Matarredonda…………………………………………11
5.2.2. Complejo de páramos de Cruz Verde – Sumapaz……………………….12
5.3.Marco teórico…………………………………………………………………….13
5.3.1. Gaultheria anastomosans...……………………………………………...13
5.3.2. Páramo…………………………………………………………………...13
5.3.3. Bosque de niebla…………………………………………………………14
5.3.4. Plasticidad………………………………………………………………..15
5.3.5. Tejidos vegetales………………………………………………………...15
5.3.5.1.Tejidos meristemáticos………………………………………………15
5.3.5.2.Parénquima…………………………………………………………..16
5.3.5.3.Tejidos de sostén…………………………………………………….16
5.3.5.4.Tejidos vasculares o conductores……………………………………17
5.3.5.5.Tejidos protectores…………………………………………………..17
6. METODOLOGÍA……………………...…………………………………………….19
6.1.Fase de campo…………………………………………………………………...19
6.2.Fase de laboratorio………………………………………………………………20
6.3.Fase de medición………………………………………………………………...24
7. RESULTADOS…………………...…………………………………………………25
7.1.Descripción de tejidos…………………………………………………………...25
7.2.Comparación estadística de epidermis…………………………………………..32
8. DISCUSIÓN…………………………..…………………………………………….37
9. CONCLUSIONES………...…………………………………………………………43
10. RECOMENDACIONES……………….……………………………………………44
11. BIBLIOGRAFÍA……………………………………………...……………………..45

5
1. INTRODUCCIÓN
Colombia y algunos países de Latinoamérica son privilegiados por la presencia de páramos
ya que son zonas con una riqueza natural monumental y además de esto ofrecen a nuestro
país diferentes servicios ambientales como el hídrico del cual la población se beneficia; hoy
en día con temas como el cambio climático, los estudios realizados en estas áreas comienzan
a tener una mayor importancia, puesto que pueden informan sobre el estado de estos
ecosistemas y promueven soluciones de problemáticas ambientales las cuales pueden tener
un impacto correcto en la conservación de los páramos, es por esto que se decidió trabajar en
el complejo de páramos de Cruz verde – Sumapaz ubicado al suroeste del departamento de
Cundinamarca y al noroeste del Meta, específicamente en el Parque ecológico de
Matarredonda, el cual hace parte de este complejo de páramos y ocupa parte del oriente de
Bogotá, estas zonas debido a la cercanía con la ciudad y a diferentes factores como los
ambientales se ven intervenidas negativamente, debido a esto surge la necesidad de
investigación en este tipo de áreas donde se intenta promover la importancia de la
conservación de la diversa riqueza natural y de los diferentes recursos ambientales que presta
y que son fuente de vida.
Esta investigación se basó en el estudio de la planta Gaultheria anastomosans la cual al ser
un arbusto presente en diversos hábitats, se puede deducir que en los páramos, donde tiene
una presencia dominante, ha generado mecanismos que le permiten una mayor adaptación
frente a plantas que son exclusivas de un área específica, conduciendo a estas a desarrollar
modificaciones a niveles fenotípicos, ya sea limitando o aumentando el crecimiento de
tejidos y órganos vegetales, por ello responden de manera más efectiva a factores
ambientales, esta capacidad de las plantas se asume desde su plasticidad, generando
condiciones para su supervivencia (Reyes, 2001).
Las características de los tejidos de las plantas se deben ver teniendo en cuenta el paso de la
historia ocurrida hace unos 450 a 500 millones de años, en el paleozoico medio, cuando las
plantas conquistaron la tierra. El medio terrestre ofrece ventajas respecto al medio acuático:
más horas y más intensidad de luz, y mayor circulación libre de CO2, sin embargo, las
plantas tienen que solventar nuevas dificultades, casi todas relacionadas con la obtención y
retención de agua, con el mantenimiento de un porte erguido en el aire y también con la

6
dispersión de las semillas en medios aéreos. Para ello las plantas se hacen más complejas:
agrupan sus células y las especializan para formar tejidos con funciones especializadas que
son capaces de hacer frente a estas nuevas dificultades (Megías et al., 2017); estos tejidos,
así mismo, también son capaces de reestructurarse para responder de manera más eficaz al
estrés ambiental.
Este estudio se enfocó en la histología vegetal debido a que es una rama que poco se ha
implementado en la flora diversa de nuestro país y teniendo en cuenta la experiencia de este
trabajo y de algunas referencias bibliografías se ha convertido en una estrategia óptima para
determinar implicaciones fisiológicas, ecológicas y hasta genéticas de todos los organismos.

7
2. DESCRIPCIÓN DEL PROBLEMA
2.1. Planteamiento del problema
Aunque algunos estudios se han realizado en variedad de especies de la Familia Ericaceae,
en Gaultheria anastomosans aún no se ha evidenciado una investigación profunda, causando
que ciertas características internas de la planta sean desconocidas y haciendo aún más
ignorada la histología de sus órganos o de sus procesos de plasticidad desarrollados en
ecosistemas estratégicos. Sin embargo, esta planta al encontrarse de manera dominante en
ecosistemas de páramo y bosque de niebla se hace característica de estas zonas, por lo cual el
estudio de esta y de toda la diversidad que se encuentra allí adquiere una importancia
relevante, ya que estos ecosistemas son características de pocos países y al ser Colombia uno
de los privilegiados en poseerlas, encontrándolas en un complejo de páramos muy extenso,
hace que la conservación de esta área y de esta especie sea trascendental debido a su
constante amenaza por diferentes factores como la intervención del hombre o el cambio
climático.
El complejo de páramos de Cruz Verde – Sumapaz, donde se llevó a cabo esta investigación,
es un páramo golpeado por la actividad humana como; la minería, la ganadería y actividades
antrópicas en general (agricultura, tala de árboles) y es hasta hace poco que allí se han
tomado iniciativas en pro de la conservación, por lo tanto, el Parque Ecológico de
Matarredonda que se ha dedicado al conocimiento de la biodiversidad y al ecoturismo,
deciden aprovechar esta iniciativa y abrir sus puertas a la investigación de la flora de estas
zonas para seguir aportando razones que muestren la importancia y la conservación que
deben poseer los páramos.
2.2. Preguntas Problema
¿Cuál es la organización a nivel histológico de la hoja de Gaultheria anastomosans?
¿Existen diferencias significativas entre la epidermis de la hoja de Gaultheria anastomosans
en bosque de niebla y páramo, que permitan determinar la plasticidad de la planta?

8
3. JUSTIFICACIÓN
La especie Gaultheria anastomosans es un arbusto que se considera típico de los ecosistemas
de páramo; gracias a la histología se puede evidenciar a un nivel más específico los
diferentes tejidos que la componen y así mismo, aquellas modificaciones en sus estructuras
internas que va creando al estar presente en ecosistemas con condiciones tan singulares.
Debido a esto se considera la histología como una estrategia óptima para observar el estado
interno en el cual se encuentran las especies que habitan estos ecosistemas y al ser Colombia
el segundo país con más hectáreas de páramos del continente americano, estas
investigaciones adquieren una importancia fundamental, ya que son promotoras de
mecanismos para poder preservarlas a pesar de la intervención del hombre y el cambio
climático.

9
4. OBJETIVOS
4.1. Objetivo general.
Describir la histología de la hoja y comparar histológicamente mediante pruebas estadísticas
la epidermis de Gaultheria anastomosans en bosque de niebla y páramo, ubicada en el
Parque Ecológico Matarredonda, Bogotá D.C., Colombia.
4.2. Objetivos Específicos.
Describir histológicamente los tejidos presentes en la hoja de Gaultheria
anastomosans.
Comparar la epidermis de la hoja de Gaultheria anastomosans de bosque de niebla y
páramo mediante métodos estadísticos.
Estimar la plasticidad de la planta desde las diferencias y/o similitudes que se
encuentren en epidermis de la hoja de Gaultheria anastomosans, como respuesta a
factores ambientales.

10
5. ESTADO DEL ARTE
5.1. Antecedentes:
Entre las investigaciones realizadas sobre la especie Gaultheria anastomosans se encuentran
dos trabajos de tesis, uno que realice en conjunto con Garzón (2017) titulado “ESTUDIO
HISTOLÓGICO DE TALLO DE Gaultheria anastomosans PRESENTE EN PÁRAMO Y
BOSQUE DE NIEBLA” donde se hace una comparación de tejidos del tallo de Gaultheria
anastomosans en dos ecosistemas: páramo y bosque de niebla, obteniendo como resultado
diferencias significativas en las células del xilema y el tejido medular como respuesta a la
presión de factores ambientales; y así mismo, uno de Cárdenas (2015) “COMPARACIÓN
HISTOLÓGICA FOLIAR DE Gaultheria anastomosans (L. f.) Kunth DE LA FAMILIA
ERICACEAE PRESENTE EN BOSQUE DE NIEBLA Y PÁRAMO DEL PARQUE
ECOLÓGICO MATARREDONDA VÍA BOGOTÁ- CHOACHÍ” donde se realiza una
estandarización de la metodología para realizar cortes histológicos en hoja y una
comparación superficial de los tejidos de hoja de Gaultheria anastomosans en dos
ecosistemas: páramo y bosque de niebla.
Por otra parte, Laskowsk (2000) realiza un estudio titulado “CARACTERÍSTICAS
ANATÓMICAS DE LA HOJA Y EL TALLO DEL SEMERUCO (Malpighia emarginata
DC) CULTIVADO EN DOS LOCALIDADES DEL ESTADO DE LARA” donde también
hace una comparación de la especie que crece en dos huertos que poseen manejo hortícola,
precipitación y temperatura promedio anual desigual, para mostrar que dicha planta adquiere
una estrategia de xeromorfismo en el xilema como respuesta a condiciones ambientales.
Molina et al., (1997) realizan un libro “CERROS, HUMEDALES Y ÁREAS RURALES” en
el cual describen la diversidad de fauna y flora de estas áreas, donde se encuentra el género
Gaultheria y algunas especies de este como características de estas zonas y de montes de
tierra fría.
Lozano & Schnetter (1976) realizan el trabajo “ESTUDIOS ECOLÓGICOS EN EL
PÁRAMO DE CRUZ VERDE, COLOMBIA II. LAS COMUNIDADES VEGETALES”,

11
donde se expone la diversidad del páramo y se indica la presencia de Ericáceas como
especies dominantes en estos ecosistemas.
Además, se pueden encontrar algunos trabajos como bitácoras, guías o inventarios en los
cuales se nombra la especie en diferentes zonas y se realiza una descripción anatómica de
esta, sin embargo, no se evidencia un análisis más extenso, estos son: “Bitácora de Flora”
realizada por Marín & Parra (2015); guía de campo “Flora Ilustrada del Páramo de
Chingaza” realizada por Madriñan (2010); “Aspectos florísticos y fitogeográficos de
Pamplona” elaborado por Sánchez & Gelviz (2004); “La flora del alto Sumapaz (cordillera
oriental, Colombia) realizado por Franco & Betancourt (1999); “Inventario Florístico del
Páramo de un Frontino (Urrao-Antioquia)” realizado por Sánchez (1998) de la Universidad
Nacional; guía de las “100 Plantas Útiles del Páramo de Rabanal” elaborado por Flórez y
otros.
5.2. Marco geográfico:
5.2.1. Parque Ecológico de Matarredonda
El Parque Ecológico Matarredonda se encuentra en la vereda El Verjón, haciendo parte del
complejo de páramos de Cruz Verde - Sumapaz, ubicado al Oriente del Departamento de
Cundinamarca, sobre la cordillera Oriental, entre los municipios de Choachí, Ubaque y
Bogotá (CAR, 2004). Posee una extensión de 1800 hectáreas, con alturas entre 3.300 hasta
los 3.500 m.s.n.m. Alberga diferentes ecosistemas, como bosques altoandinos primario y
secundario, donde habitan una gran variedad de fauna y flora (Morales, Et al., 2007).
El Parque cuenta con ocho caminos totalmente reglamentados para hacer caminatas de
carácter ecológico, de sensibilización e inducción en pro del ambiente; en los recorridos se
puede apreciar diferentes tipos de vegetación, ecosistemas, grandes recursos de
enriquecimiento natural e importantes nacimientos de cuencas hidrográficas que riegan la
capital y algunas poblaciones de Cundinamarca, el Meta y la Orinoquía (Fundación
Humedales de Bogotá, 2016).

12
Los páramos están privilegiadamente ubicados entre los 3.100 m.s.n.m. y los 4.000 m.s.n.m.,
razón por la cual su incidencia de luz es significativa para el óptimo desarrollo de las
comunidades vegetales. Además, sus temperaturas bajas producen una lenta evaporación,
reteniendo el agua y convirtiéndose en fuentes primordiales de este recurso (Ortíz & Reyes
Bonilla, 2009). En el subpáramo predomina vegetación de tipo arbustivo, matorrales del
género Gaultheria. Así mismo también se encuentra el bosque de niebla, también llamado
bosque muy húmedo montano (bmh-MB), ubicado entre los 2.000 m.s.n.m. y los 3.000
m.s.n.m. (CORTOLIMA, 2007).
Imagen 1. Mapa del lugar de estudio. Google Earth, todos los derechos reservados. (Garzón, 2017)
5.2.2. Complejo de páramos de cruz verde – Sumapaz
Es considerado uno de los páramos más grandes del mundo con una extensión de 266.750 ha,
este se encuentra al suroeste del departamento de Cundinamarca y al noroeste del Meta,
incluida una pequeña parte del norte del Huila. Comprende un sector de 25 municipios,
dentro de los cuales los de mayor área son: Bogotá, D. C., San Luis de Cubarral, La Uribe,
Guamal y Gutiérrez. El complejo de Cruz Verde - Sumapaz se sitúa entre los 3.250 y 4.230

13
m de altitud y comprende los páramos de Choachí, Cruz Verde, Las Ánimas, Monserrate,
Andabobos, El Cajón, El Cedral, Sumapaz, El Nevado, Nuevo Mundo, Pasquilla, Usme,
Chisacá y Las Mercedes, entre otros (Morales et al., 2007).
El complejo posee una importancia hidrológica para la región, ya que en él están una
multitud de lagunas naturales, más de 20 reconocidas por el IDEAM, así mismo, se
encuentran los embalses de Chisacá y La Regadera, que abastecen al Distrito Capital
(Morales et al., 2007).
Este complejo de páramos se considera como área prioritaria dentro de los ecosistemas
estratégicos de páramos. Cleef en 1997, lo describió como el segundo centro biogeográfico
de la cordillera Oriental, después del Cocuy. Un estudio de la CAR y la Universidad
Nacional (2004) afirma que los páramos en territorio de la CAR mejor documentados desde
el punto de vista florístico son los de Sumapaz, con registros de 635 especies de 280 géneros
y 80 familias (Morales et al., 2007).
5.3. Marco teórico:
5.3.1. Gaultheria anastomosans
La especie Gaultheria anastomosans corresponde a plantas de tipo arbustivo, con altura que
va de 1 m a 1.5 m de altura, sus ramas son delgadas, muy resistentes y cubiertas de
numerosos pelos; sus hojas miden aproximadamente 5 cm de largo por 3 cm de ancho,
adquieren una forma ovalada, alargada y con nervaduras muy marcadas por el envés de la
misma, en cuanto a su inflorescencia, esta se encuentra en racimos provistos de flores de
forma acampanada de color blanco. Según Kausel (1949), las especies de Gaultheria por lo
general son plantas heliófilas (dependientes del sol). Según Mabberley (1987) y Stevens
(2001) tienen una tendencia a crecer en suelos ácidos, como lo son la mayoría de los
representantes de esta familia (Teillier & Escobar, 2013, pág. 137). Posee un metabolismo
fotosintético tipo C3. La distribución de estos arbustos es muy amplia, hallando registros de
su presencia en todos los continentes, y en diversos ecosistemas, razón por la cual se
considera con una tasa de adaptación alta.

14
5.3.2. Páramo
Según Morales y otros, (2007) los páramos se encuentran en Colombia por encima de los
3.000 metros de altitud, así mismo sus temperaturas bajas producen una lenta evaporación
produciendo una retención de agua convirtiéndolos en la fuente constante de este recurso el
cual es primordial para la supervivencia de los seres vivos. Debido a su altura su incidencia
de luz es significativa para el óptimo desarrollo de las comunidades vegetales. (Ortíz &
Reyes Bonilla, 2009).
Desde una perspectiva general, los páramos de Colombia se asimilan a áreas altas, frías,
húmedas, nubladas y con vegetación abierta hasta arbustiva, dentro de la que se destacan los
emblemáticos frailejones. Las familias más ricas a nivel de flora aproximadamente son
Asteraceae (100 géneros y 710 especies), Orchidaceae (57 géneros y 580 especies), Poaceae
(40 géneros y 150 especies), Melastomataceae (12 géneros y 110 especies) y Bromeliaceae
(7 géneros, 100 especies). Los géneros con mayor número de especies son Epidendrum (105
especies), Espeletia (80 especies), Pleurothallis (80 especies), Diplostephium (75 especies),
Miconia (65 especies), Hypericum (55 especies), Monticalia (55 especies) y Baccharis (55
especies) (Morales et al., 2007).
5.3.3. Bosque de niebla
El bosque de niebla, también llamado bosque muy húmedo montano (bmh-MB), está
ubicado entre los 2.000 m.s.n.m. y los 3.000 m.s.n.m. (CORTOLIMA, 2007). Se consideran
como sitios boscosos donde el aire, proveniente de regiones bajas, húmedas y cálidas, se
condensa para producir regularmente nubosidad que permanece la mayor parte del tiempo.
Debido a las características de estos ecosistemas toman una gran importancia ya que sirven
de hábitat o sustento para muchas especies y por tener la capacidad de almacenar agua que
capturan de las nubes y la neblina (Armenteras et al., 2007).
En los bosques de niebla de Colombia predomina vegetación de tipo arbustivo, se estima un
total de 2.341 registros biológicos, los cuales corresponden a 1.657 especies, en su mayoría
pertenecientes a las familias botánicas Rubiaceae, Melastomataceae, Ericaceae,
Gesneriaceae, Asteraceae, Araceae, Piperaceae, Orchidaceae, Bromeliaceae y Solanaceae.

15
Así mismo, especies endémicas como las Ericáceas (Cavendishia albopicata, Diogenesia
antioquiensis, Macleania penduliflora), especies emblemáticas de la zona andina, como la
palma de cera (Ceroxylon quinduense) y los pinos nativos colombianos (Podocarpus y
Prumnopitys) (Armenteras et al., 2007).
5.3.4. Plasticidad
Las plantas al estar presentes en un mismo lugar soportan las condiciones ambientales
desfavorables como la falta de agua, altas y bajas temperaturas o escasez de nutrientes de
este, haciendo que a lo largo de su evolución desarrollen mecanismos que les permitan
tolerar y hacer frente a estas. Así pues, este mecanismo se denomina: plasticidad; esta ha sido
confundida con flexibilidad, estabilidad y regulación interna, llegando a la conclusión de que
sólo se trata de homologías. Es conocido que la flexibilidad es la característica por la cual un
organismo puede crecer y reproducirse en cualquier ambiente por variación de su forma o
manteniendo una forma constante; estabilidad indica una condición en la cual no ocurren
muchos cambios; y regulación interna es la tendencia de un sistema a mantener sus
características morfológicas y fisiológicas constantes. La plasticidad es un mecanismo que
favorece a las especies ante condiciones heterogéneas, lo que hace que éstas respondan
positivamente a la selección natural, definida como la preservación de las variaciones útiles y
la eliminación de las nocivas; en términos generales es la descendencia con modificación
(Reyes et al., 2001).
5.3.5. Tejidos Vegetales
Según Paniagua et al., (2002) un tejido es un grupo de células de origen, estructura y
funciones comunes, que atendiendo a su función se pueden clasificar en meristemos,
parénquima, de sostén, vasculares o conductores, protectores y secretores.
5.3.5.1. Tejidos meristemáticos
Tanto los vegetales como los animales proceden de una única célula o cigoto que se ira
dividiendo para formar el embrión, caracterizado por un conjunto de células escasamente
diferenciadas que sufren divisiones mitóticas. Posteriormente las células originadas se van

16
diferenciando para dar lugar a los diferentes tejidos del organismo animal o vegetal. En los
vegetales existen zonas denominadas meristemos las cuales están formadas de células
capaces de hacer crecer al organismo no solo aumentando el volumen de sus órganos sino
multiplicando el número de estos. Estos meristemos se consideran tejidos embrionarios que
estarán en la planta durante toda su vida, causantes del crecimiento permanente del individuo
vegetal gracias a la capacidad de división y diferenciación (Paniagua et al., 2002).
5.3.5.2. Parénquima
Es considerado como un tejido de relleno, ocupando espacios entre otros tejidos más
diferenciados; los tejidos parenquimatosos adquieren funciones como la fotosíntesis,
elaboración y almacenamiento de sustancias, secreción, excreción, entre otras (Paniagua et
al., 2002).
Entre los tipos de parénquima podemos encontrar: 1. Parénquima asimilador o clorofílico en
este se realizara la fijación del carbono mediante la fotosíntesis, por lo que se localizara
debajo de la epidermis, este tendrá un desarrollo mayor en las hojas donde puede ser de dos
formas: a. Parénquima en empalizada: constituido por células prismáticas y b. Parénquima
lagunar: formado por células más redondeadas y con espacios intercelulares amplios; 2.
Parénquima de reserva: encargado del almacenamiento de diversas sustancias la más
frecuente y abundante es el almidón que se encuentra en amiloplastos; 3. Parénquima
aerífero: lo poseen plantas acuáticas o que crecen en lugares encharcados donde se forman
cordones celulares donde se conducirá el aire; 4. Parénquima acuífero: se encuentra en
plantas de climas secos, sus tejidos poseen parénquima almacenador de agua (Paniagua et
al.,2002); existen otros tipos de parénquima pero los mencionados anteriormente se
consideran los más relevantes.
5.3.5.3. Tejidos de sostén
Colénquima: tejido de sostén que se haya en órganos en vías de crecimiento, está constituido
por células vivas con cloroplastos. En las hojas se encuentra tanto protegiendo los haces
vasculares (venas) como a lo largo de los márgenes o con frecuencia en el peciolo Paniagua
et al., 2002).

17
Esclerénquima: está constituido por células que desarrollan una pared secundaria gruesa y
endurecida por el depósito de lignina mediante el proceso de lignificación, esto ofrece una
resistencia mayor que el colénquima. Este tejido de sostén es característico de órganos
adultos, que ya han dejado de crecer (Paniagua et al., 2002).
5.3.5.4. Tejidos vasculares o conductores:
Xilema: transporta agua y sustancias tales como sales minerales y compuestos nitrogenados,
se realiza desde la raíz y a través de toda la planta, este transporte es unidireccional hacia las
superficies de evaporación. Durante el crecimiento primario se forma el xilema primario
(primero el protoxilema y luego el metaxilema) y posteriormente el xilema secundario
(Paniagua et al., 2002).
Floema: transporta productos de la fotosíntesis, como azucares y materias orgánicas
elaboradas, en múltiples direcciones a través de la planta desde su sitio de producción
(fuentes) hasta los lugares donde son consumidos o almacenados (sumideros). Durante el
crecimiento primario se forma el floema primario (primero el protofloema y luego el
metafloema) y posteriormente el floema secundario (Paniagua et al., 2002).
5.3.5.5. Tejidos protectores:
Epidermis: es la capa celular más externa en hojas, verticilos florales, frutos, semillas, tallos
y raíces. Su función es la de intercambios entre el interior y el exterior de la planta, así
mismo, funciona como defensa ante los agentes externos físicos, ayuda en la regulación de la
transpiración y el intercambio de gases a través de los estomas, acumulación de sustancias
que posteriormente son segregadas, entre otras. No es completamente impermeable ya que
deja salir agua con algunos solutos, proceso que se denomina transpiración cuticular
(Paniagua et al., 2002).
Las células epidérmicas presentan diversas formas y tamaños lo cual está estrechamente
relacionado con su medio externo, pero por lo general estas células son aplanadas y se
encuentran unidas sin espacios intercelulares. El CO2 penetra a través de las células
epidérmicas, en algunas plantas se utilizan vías proporcionadas por los estomas y en algunas

18
cutículas existen diminutos canales que permiten la salida de secreciones (Paniagua et al.,
2002).
Cutícula: es una capa que se encuentra sobre la pared celular de las células epidérmicas la
cual está en contacto con el medio externo. Está constituida por cutina, una sustancia lipídica
que resulta de la polimerización de algunos ácidos grasos. Esta capa se forma en el estadio
más temprano de los órganos, los ácidos grasos insaturados que la conformaran se generaran
en el citosol y serán segregados por las células epidérmicas atravesando su pared externa a
través de los ectodermos o bien en forma de pequeñas gotas, estos ácidos grasos inician un
proceso de endurecimiento que comporta su oxidación y polimerización transformándose en
cutina, proceso que se conoce como cutinización (Paniagua et al. ,2002).

19
6. METODOLOGÍA
La presente investigación se dividió en tres fases, una fase de campo y una fase de
laboratorio, las cuales se plantearon teniendo en cuenta la metodología sugerida por
Sandoval (2005) y una última fase de medición de tejidos que se realizó con ayuda del
programa ConfocalUnioviImageJ.
6.1. Fase de Campo:
Se inició con el denominado muestreo, haciendo alusión a las formas en que se puede
muestrear sin que el material vivo tenga mayor variación y lo más importante sin realizar un
daño ecológico. La recolecta se efectuó directamente en el lugar donde se encontraban las
platas, identificando previamente la ubicación geográfica por medio de un GPS, luego para
cada ecosistema se tomaron 5 individuos de la especie al azar que midieran entre 80 a 120cm
de altura, posteriormente se tomaron 10 hojas de cada arbusto y se les corto transversalmente
el ápice y borde foliar y para que no existiera ningún daño en los tejidos las muestras
colectadas se preservaron en un fijador. (Tabla 1).
Tabla No. 1. Composición del Fijador.
FIJACIÓN CANTIDAD APROX
Ácido acético glacial 99% 50 ml
Formol 37-40% 100 ml
Agua destilada 350 ml
Etanol 96-100% 500 ml
Para transportar las muestras se colocó 5ml de fijador en frascos plásticos, se marcaron
teniendo en cuenta los siguientes aspectos: el número del individuo, el órgano, el ecosistema
y la hora. Con el fin de mantener una temperatura baja las muestras se colocaron en una
nevera de icopor. Por último el material biológico se dejó en el fijador durante 24 horas
(Sandoval, 2005).
20
6.2. Fase de Laboratorio:
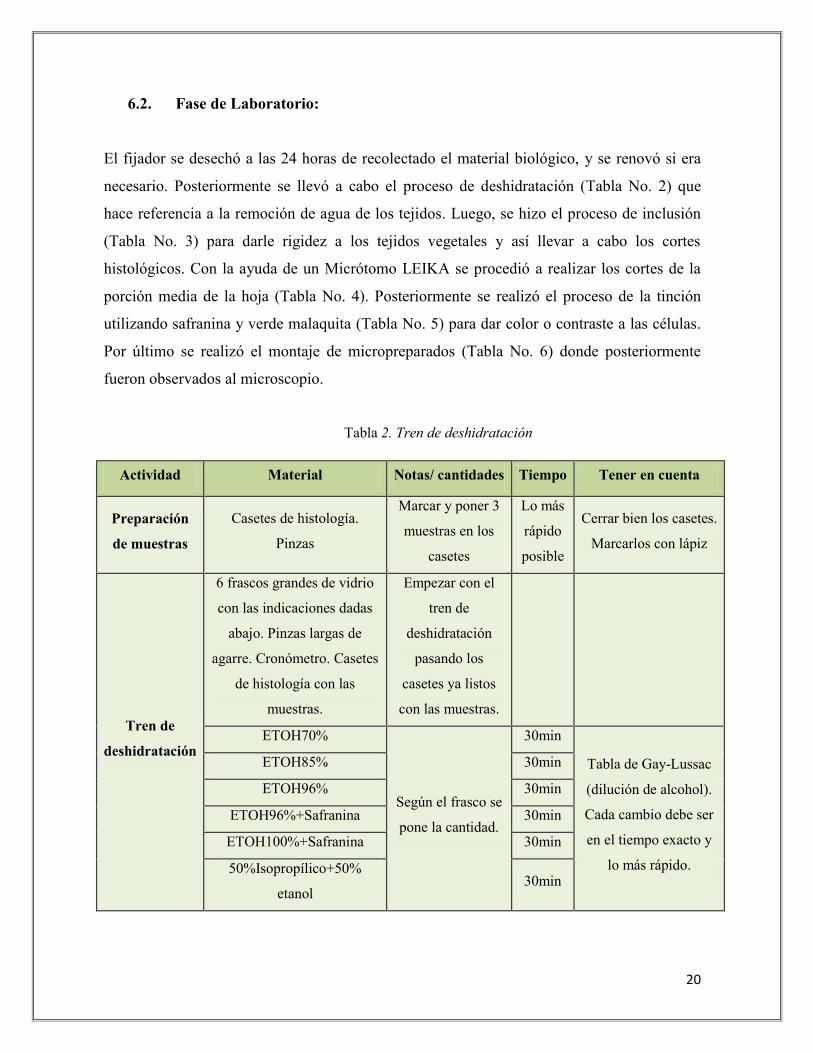
El fijador se desechó a las 24 horas de recolectado el material biológico, y se renovó si era
necesario. Posteriormente se llevó a cabo el proceso de deshidratación (Tabla No. 2) que
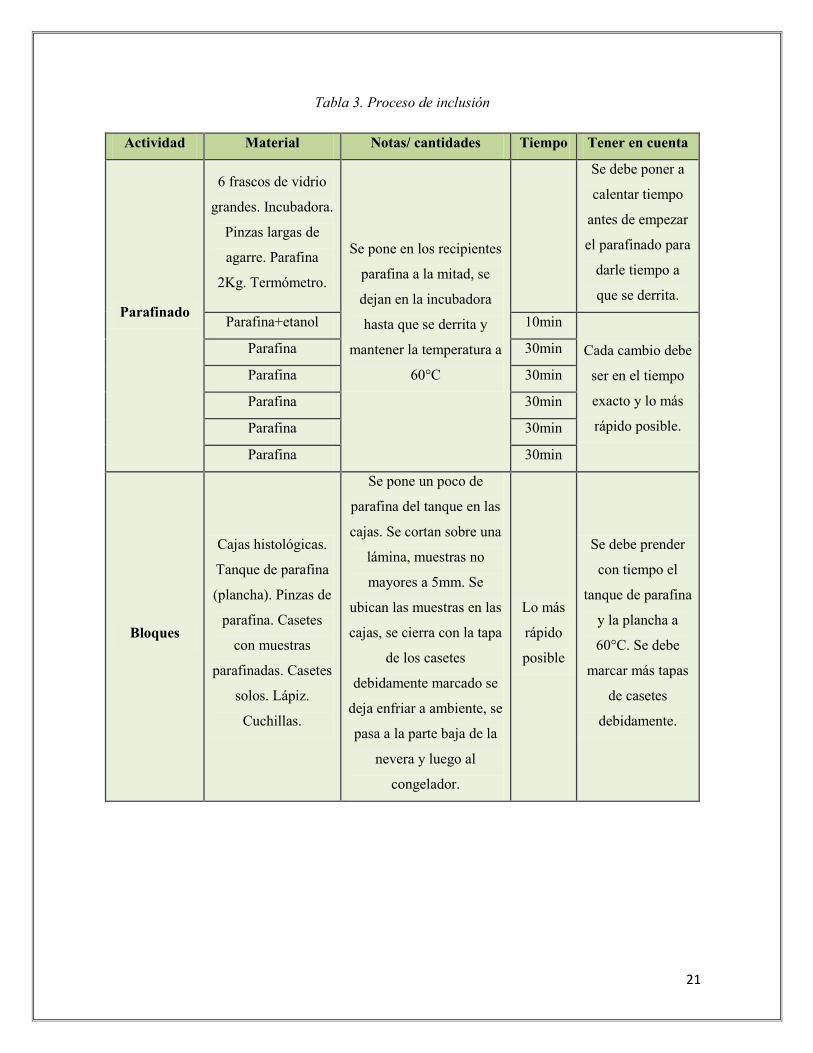
hace referencia a la remoción de agua de los tejidos. Luego, se hizo el proceso de inclusión
(Tabla No. 3) para darle rigidez a los tejidos vegetales y así llevar a cabo los cortes
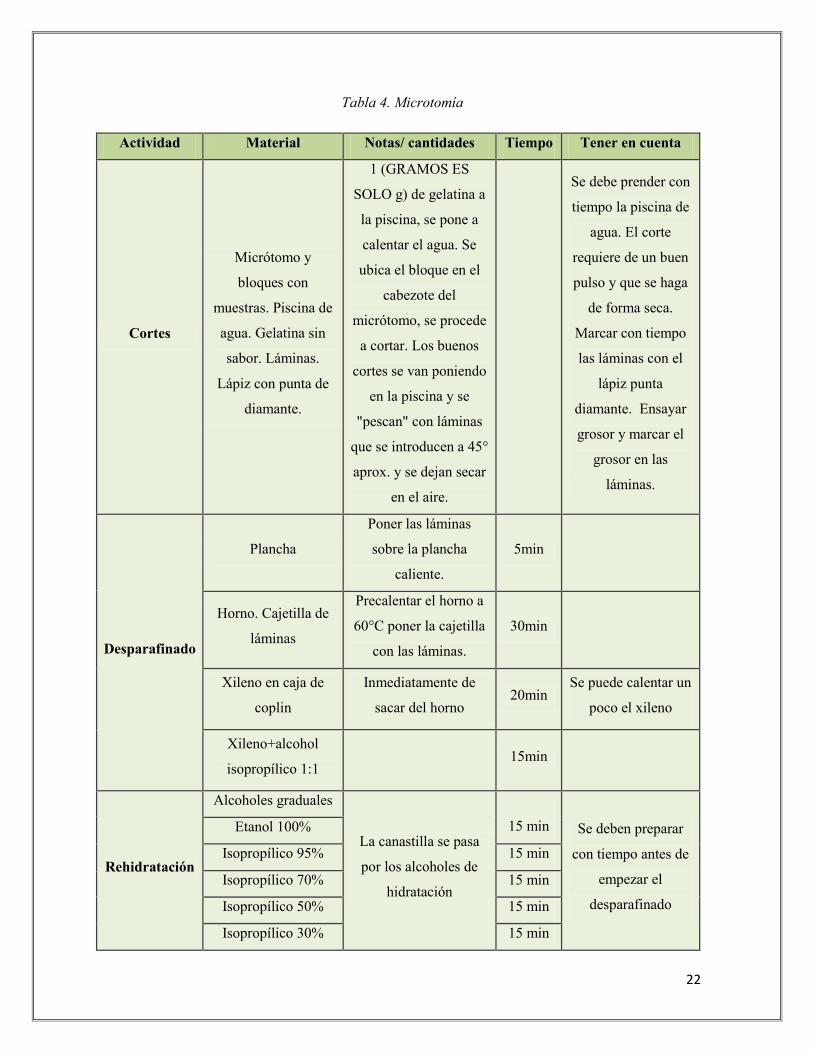
histológicos. Con la ayuda de un Micrótomo LEIKA se procedió a realizar los cortes de la
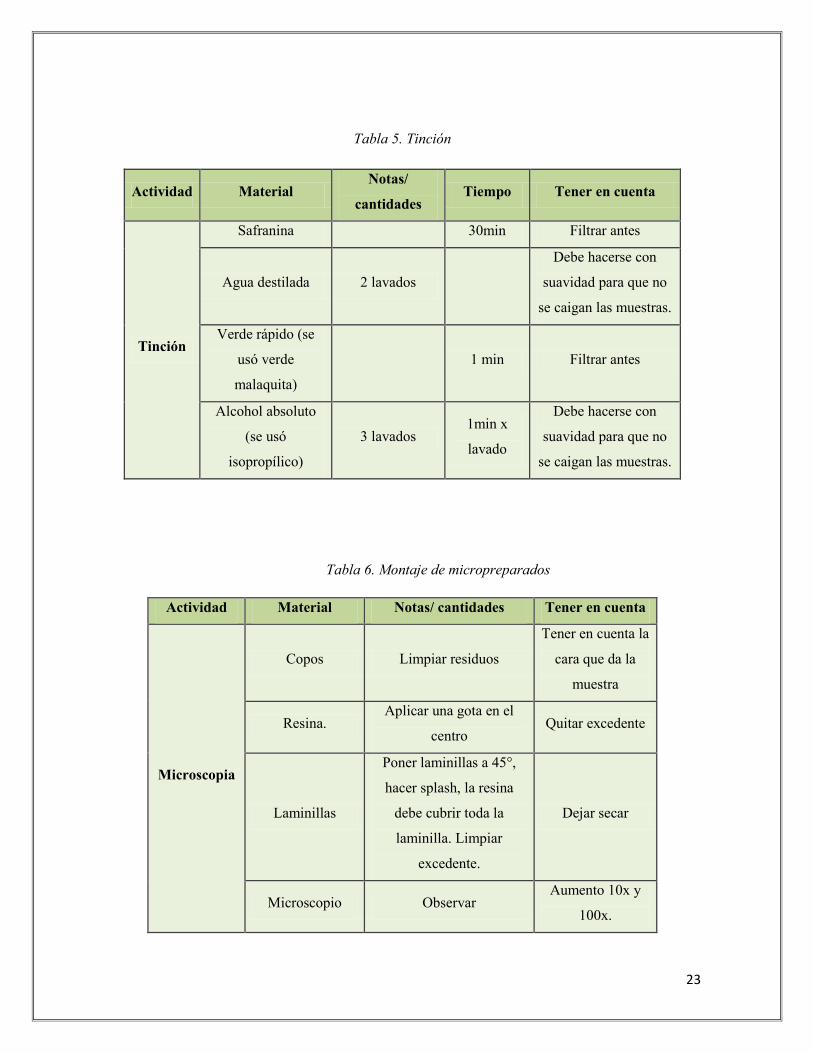
porción media de la hoja (Tabla No. 4). Posteriormente se realizó el proceso de la tinción
utilizando safranina y verde malaquita (Tabla No. 5) para dar color o contraste a las células.
Por último se realizó el montaje de micropreparados (Tabla No. 6) donde posteriormente
fueron observados al microscopio.
Tabla 2. Tren de deshidratación
Actividad Material Notas/ cantidades Tiempo Tener en cuenta
Preparación
de muestras
Casetes de histología.
Pinzas
Marcar y poner 3
muestras en los
casetes
Lo más
rápido
posible
Cerrar bien los casetes.
Marcarlos con lápiz
Tren de
deshidratación
6 frascos grandes de vidrio
con las indicaciones dadas
abajo. Pinzas largas de
agarre. Cronómetro. Casetes
de histología con las
muestras.
Empezar con el
tren de
deshidratación
pasando los
casetes ya listos
con las muestras.
ETOH70%
Según el frasco se
pone la cantidad.
30min
Tabla de Gay-Lussac
(dilución de alcohol).
Cada cambio debe ser
en el tiempo exacto y
lo más rápido.
ETOH85% 30min
ETOH96% 30min
ETOH96%+Safranina 30min
ETOH100%+Safranina 30min
50%Isopropílico+50%
etanol 30min
21
Tabla 3. Proceso de inclusión
Actividad Material Notas/ cantidades Tiempo Tener en cuenta
Parafinado
6 frascos de vidrio
grandes. Incubadora.
Pinzas largas de
agarre. Parafina
2Kg. Termómetro.
Se pone en los recipientes
parafina a la mitad, se
dejan en la incubadora
hasta que se derrita y
mantener la temperatura a
60°C
Se debe poner a
calentar tiempo
antes de empezar
el parafinado para
darle tiempo a
que se derrita.
Parafina+etanol 10min
Cada cambio debe
ser en el tiempo
exacto y lo más
rápido posible.
Parafina 30min
Parafina 30min
Parafina 30min
Parafina 30min
Parafina 30min
Bloques
Cajas histológicas.
Tanque de parafina
(plancha). Pinzas de
parafina. Casetes
con muestras
parafinadas. Casetes
solos. Lápiz.
Cuchillas.
Se pone un poco de
parafina del tanque en las
cajas. Se cortan sobre una
lámina, muestras no
mayores a 5mm. Se
ubican las muestras en las
cajas, se cierra con la tapa
de los casetes
debidamente marcado se
deja enfriar a ambiente, se
pasa a la parte baja de la
nevera y luego al
congelador.
Lo más
rápido
posible
Se debe prender
con tiempo el
tanque de parafina
y la plancha a
60°C. Se debe
marcar más tapas
de casetes
debidamente.
22
Tabla 4. Microtomía
Actividad Material Notas/ cantidades Tiempo Tener en cuenta
Cortes
Micrótomo y
bloques con
muestras. Piscina de
agua. Gelatina sin
sabor. Láminas.
Lápiz con punta de
diamante.
1 (GRAMOS ES
SOLO g) de gelatina a
la piscina, se pone a
calentar el agua. Se
ubica el bloque en el
cabezote del
micrótomo, se procede
a cortar. Los buenos
cortes se van poniendo
en la piscina y se
"pescan" con láminas
que se introducen a 45°
aprox. y se dejan secar
en el aire.
Se debe prender con
tiempo la piscina de
agua. El corte
requiere de un buen
pulso y que se haga
de forma seca.
Marcar con tiempo
las láminas con el
lápiz punta
diamante. Ensayar
grosor y marcar el
grosor en las
láminas.
Desparafinado
Plancha
Poner las láminas
sobre la plancha
caliente.
5min
Horno. Cajetilla de
láminas
Precalentar el horno a
60°C poner la cajetilla
con las láminas.
30min
Xileno en caja de
coplin
Inmediatamente de
sacar del horno 20min
Se puede calentar un
poco el xileno
Xileno+alcohol
isopropílico 1:1 15min
Rehidratación
Alcoholes graduales
La canastilla se pasa
por los alcoholes de
hidratación
15 min Se deben preparar
con tiempo antes de
empezar el
desparafinado
Etanol 100%
Isopropílico 95% 15 min
Isopropílico 70% 15 min
Isopropílico 50% 15 min
Isopropílico 30% 15 min
23
Tabla 5. Tinción
Actividad Material Notas/
cantidades Tiempo Tener en cuenta
Tinción
Safranina
30min Filtrar antes
Agua destilada 2 lavados
Debe hacerse con
suavidad para que no
se caigan las muestras.
Verde rápido (se
usó verde
malaquita)
1 min Filtrar antes
Alcohol absoluto
(se usó
isopropílico)
3 lavados 1min x
lavado
Debe hacerse con
suavidad para que no
se caigan las muestras.
Tabla 6. Montaje de micropreparados
Actividad Material Notas/ cantidades Tener en cuenta
Microscopia
Copos Limpiar residuos
Tener en cuenta la
cara que da la
muestra
Resina. Aplicar una gota en el
centro Quitar excedente
Laminillas
Poner laminillas a 45°,
hacer splash, la resina
debe cubrir toda la
laminilla. Limpiar
excedente.
Dejar secar
Microscopio Observar Aumento 10x y
100x.

24
6.3. Fase de medición:
Para finalizar, después de sellar los montajes con entellan en láminas de vidrio, se procedió a
observar y tomar fotografías de los cortes de hoja de Gaultheria anastomosans en un
microscopio en aumentos 10X, 40X y 100X, para realizar las correspondientes mediciones
de los tejidos de hoja con ayuda del programa ConfocalUnioviImageJ. Por último, se realizó
un análisis estadístico de epidermis con la prueba de Mann-Whitn U de dos variables, para
evidenciar diferencias significativas entre los ecosistemas.

25
7. RESULTADOS
7.1. Descripción de tejidos:
La hoja de Gaultheria anastomosans presentó tanto en bosque de niebla como en páramo
una distribución semejante, por lo cual no se evidencio una alteración o modificación de sus
tejidos en estos diferentes ecosistemas.
Imagen 2. Corte transversal foliar de Gaultheria anastomosans en páramo.
Aumento: 10X - Tinciones: Safranina y Verde Malaquita.
Cutícula adaxial (CAD); epidermis monoestratificada adaxial (EAD); parénquima clorofílico en empalizada
(PEM); parénquima lagunoso o esponjoso (PLA); epidermis abaxial (EAB); cutícula abaxial (CAB).
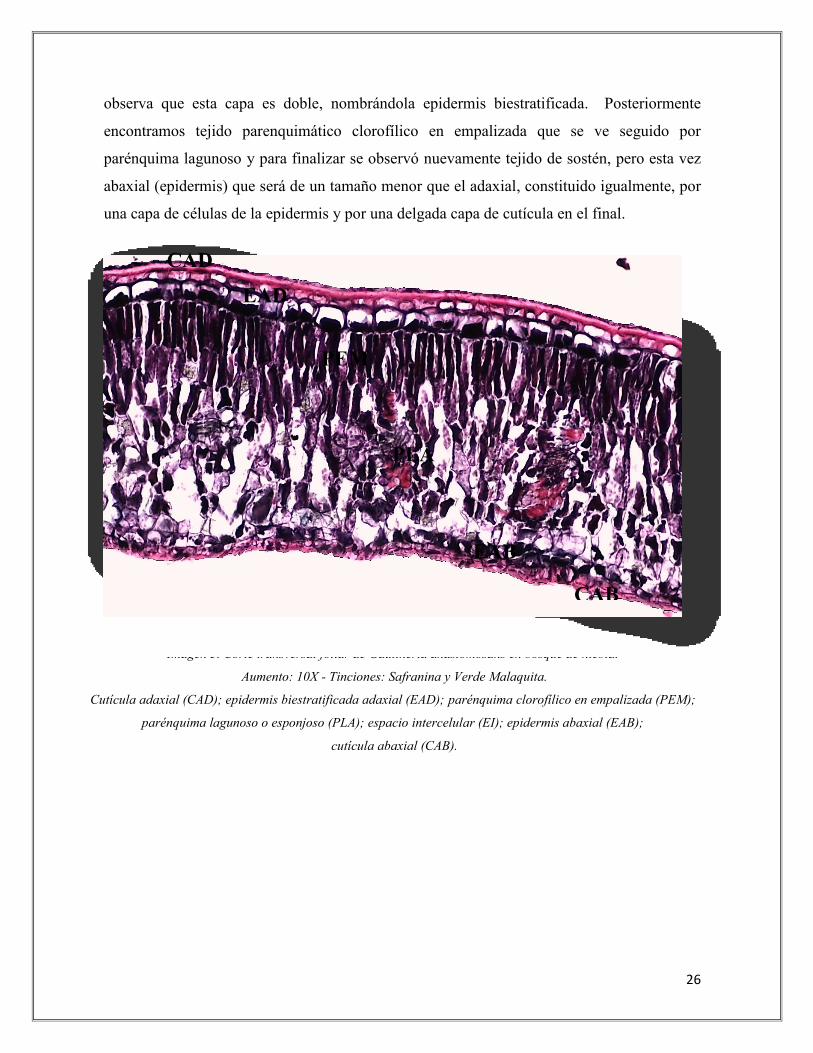
En la imagen 2, 3 y 4 se puede evidenciar totalmente un corte transversal de hoja de
Gaultheria anastomosans presente en páramo y bosque de niebla, en los cuales se observa la
distribución de los tejidos que los componen; este comienza con un tejido de sostén adaxial
(epidermis) compuesto por una capa de cutícula y seguido por una capa de células
epidérmicas, en la imagen 2 del corte de hoja esta capa es de una sola fila de células por lo
cual se denomina epidermis monoestratificada y en la imagen 3 y 4 del corte de hoja se
CADEAD
PEM
PLA
EAB CAB
26
observa que esta capa es doble, nombrándola epidermis biestratificada. Posteriormente
encontramos tejido parenquimático clorofílico en empalizada que se ve seguido por
parénquima lagunoso y para finalizar se observó nuevamente tejido de sostén, pero esta vez
abaxial (epidermis) que será de un tamaño menor que el adaxial, constituido igualmente, por
una capa de células de la epidermis y por una delgada capa de cutícula en el final.
Imagen 3. Corte transversal foliar de Gaultheria anastomosans en bosque de niebla.
Aumento: 10X - Tinciones: Safranina y Verde Malaquita.
Cutícula adaxial (CAD); epidermis biestratificada adaxial (EAD); parénquima clorofílico en empalizada (PEM);
parénquima lagunoso o esponjoso (PLA); espacio intercelular (EI); epidermis abaxial (EAB);
cutícula abaxial (CAB).
CAD
EAD
PLA
PEM
EAB
CAB
27
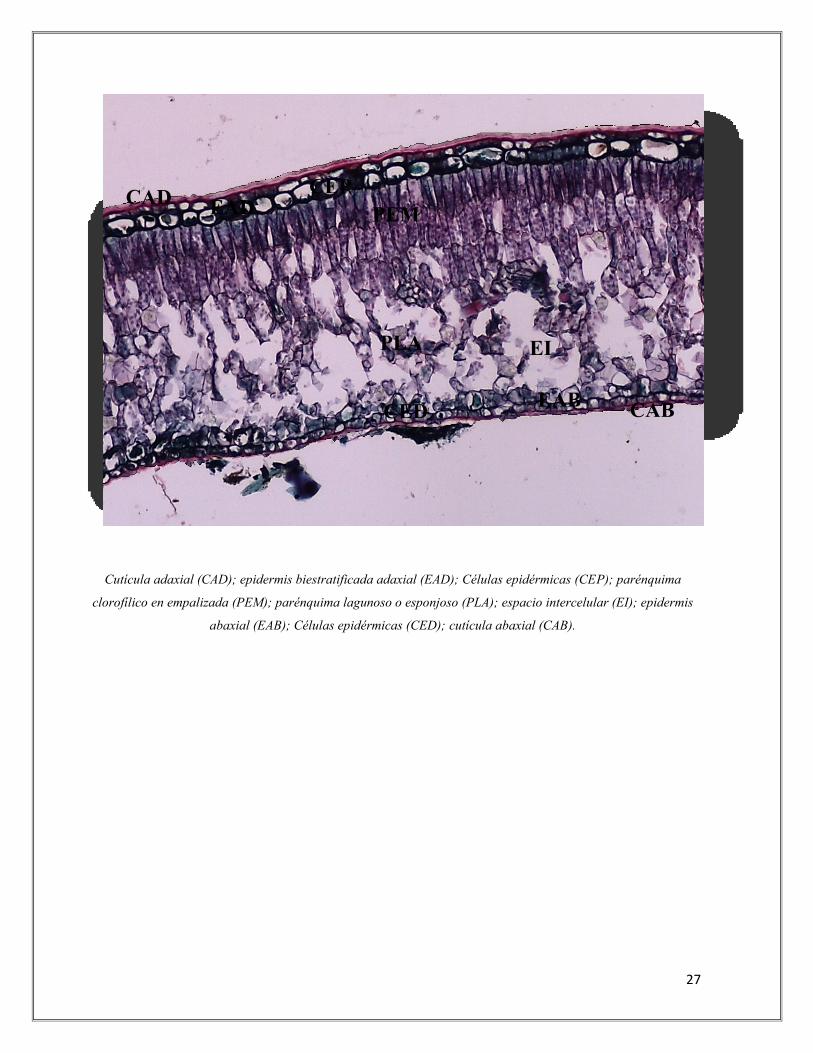
Imagen 4. Corte transversal foliar de Gaultheria anastomosans en páramo.
Aumento: 10X - Tinciones: Safranina y Verde Malaquita.
Cutícula adaxial (CAD); epidermis biestratificada adaxial (EAD); Células epidérmicas (CEP); parénquima
clorofílico en empalizada (PEM); parénquima lagunoso o esponjoso (PLA); espacio intercelular (EI); epidermis
abaxial (EAB); Células epidérmicas (CED); cutícula abaxial (CAB).
CADEAD
CEP
PEM
PLA EI
CED EAB
CAB

28
Imagen 4. Corte transversal foliar de Gaultheria anastomosans en bosque de niebla. Cara adaxial.
Aumento: 40X - Tinciones: Safranina y Verde Malaquita.
Cutícula adaxial (CAD); epidermis adaxial monoestratificada (EAD); Células epidérmicas (CEP);
parénquima clorofílico en empalizada (PEM).
En el corte anterior se pueden observar dos tipos de tejidos, uno de protección adaxial
(epidermis) constituido por una capa de cutícula y por una capa epidérmica
monoestratificada, que posee una sola fila de células, y continuamente un tejido
parenquimatico clorofílico en empalizada.
CAD
EAD
PEM
CEP
29
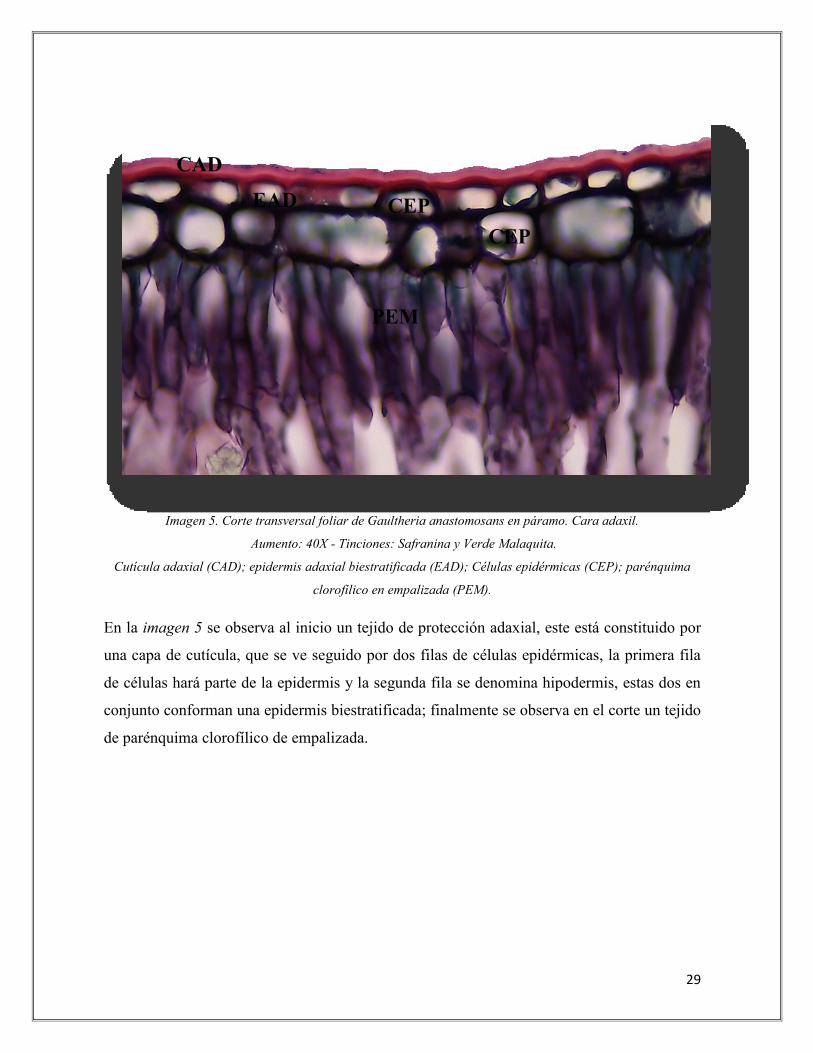
Imagen 5. Corte transversal foliar de Gaultheria anastomosans en páramo. Cara adaxil.
Aumento: 40X - Tinciones: Safranina y Verde Malaquita.
Cutícula adaxial (CAD); epidermis adaxial biestratificada (EAD); Células epidérmicas (CEP); parénquima
clorofílico en empalizada (PEM).
En la imagen 5 se observa al inicio un tejido de protección adaxial, este está constituido por
una capa de cutícula, que se ve seguido por dos filas de células epidérmicas, la primera fila
de células hará parte de la epidermis y la segunda fila se denomina hipodermis, estas dos en
conjunto conforman una epidermis biestratificada; finalmente se observa en el corte un tejido
de parénquima clorofílico de empalizada.
CAD
EAD CEP
CEP
PEM
30
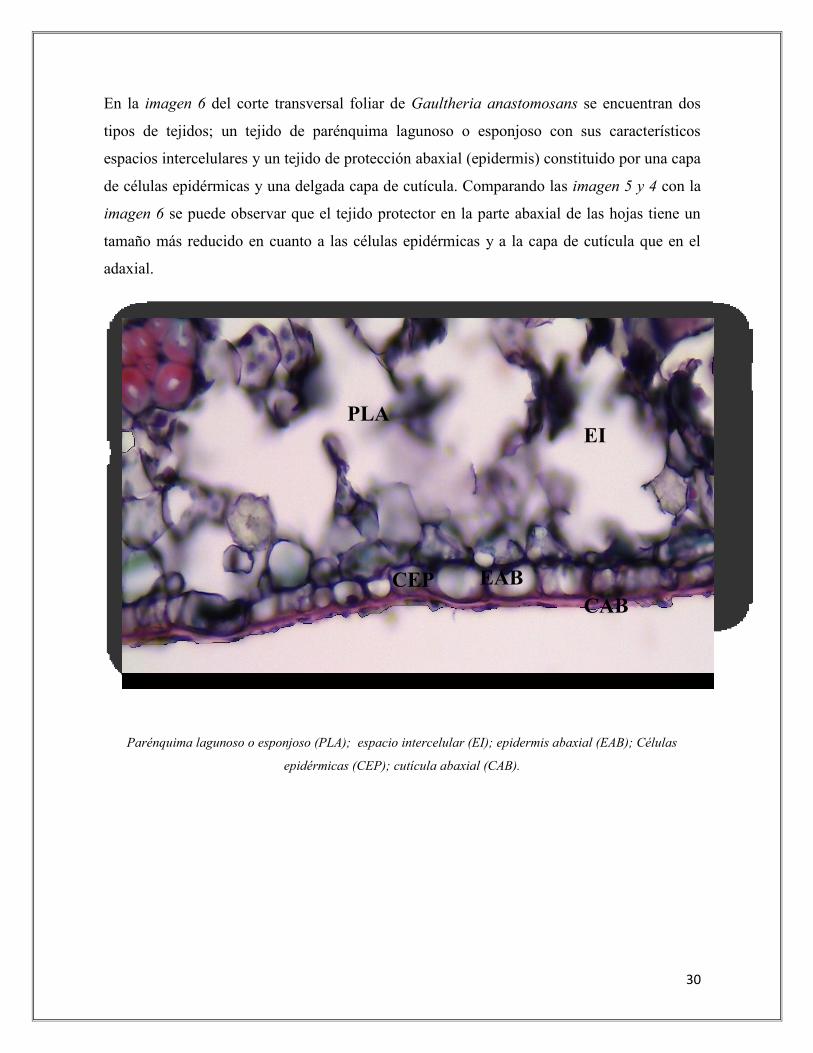
En la imagen 6 del corte transversal foliar de Gaultheria anastomosans se encuentran dos
tipos de tejidos; un tejido de parénquima lagunoso o esponjoso con sus característicos
espacios intercelulares y un tejido de protección abaxial (epidermis) constituido por una capa
de células epidérmicas y una delgada capa de cutícula. Comparando las imagen 5 y 4 con la
imagen 6 se puede observar que el tejido protector en la parte abaxial de las hojas tiene un
tamaño más reducido en cuanto a las células epidérmicas y a la capa de cutícula que en el
adaxial.
Imagen 6. Corte transversal foliar de Gaultheria anastomosans en páramo. Cara abaxial.
Aumento: 40X - Tinciones: Safranina y Verde Malaquita.
Parénquima lagunoso o esponjoso (PLA); espacio intercelular (EI); epidermis abaxial (EAB); Células
epidérmicas (CEP); cutícula abaxial (CAB).
PLA EI
CEP EAB
CAB
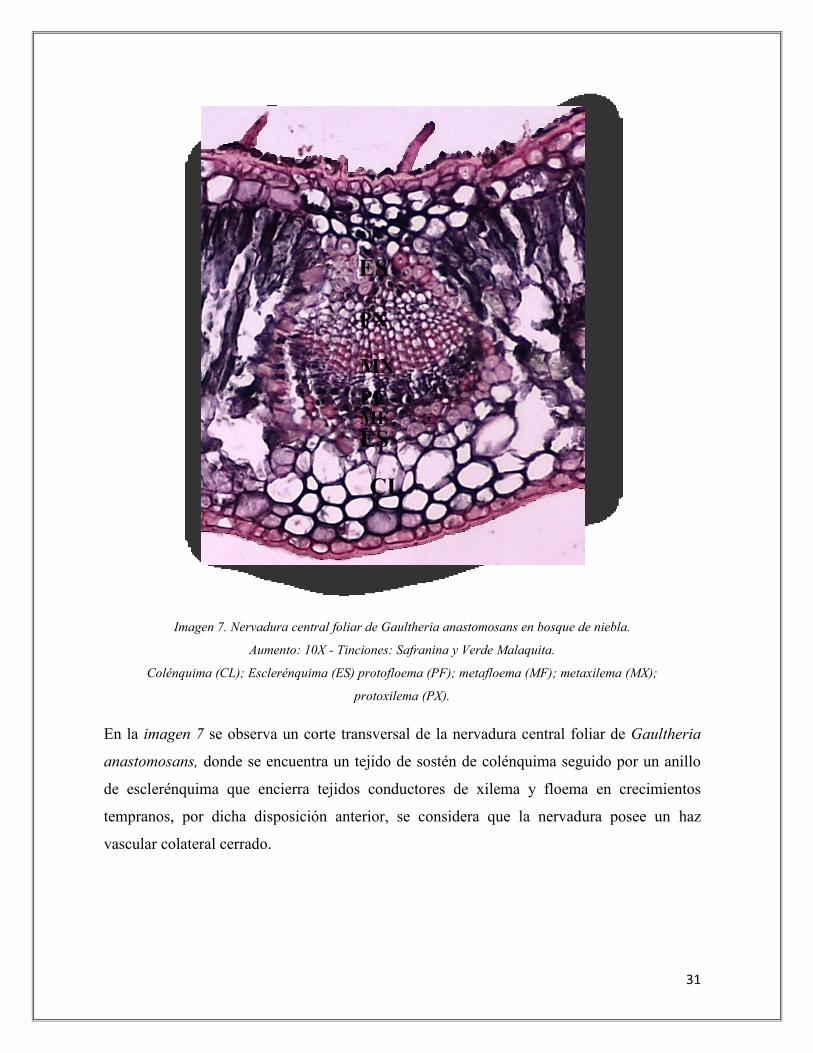
31
Imagen 7. Nervadura central foliar de Gaultheria anastomosans en bosque de niebla.
Aumento: 10X - Tinciones: Safranina y Verde Malaquita.
Colénquima (CL); Esclerénquima (ES) protofloema (PF); metafloema (MF); metaxilema (MX);
protoxilema (PX).
En la imagen 7 se observa un corte transversal de la nervadura central foliar de Gaultheria
anastomosans, donde se encuentra un tejido de sostén de colénquima seguido por un anillo
de esclerénquima que encierra tejidos conductores de xilema y floema en crecimientos
tempranos, por dicha disposición anterior, se considera que la nervadura posee un haz
vascular colateral cerrado.
CL
ES
MX
PX
ES
PF MF

32
7.2. Comparación estadística de epidermis:
La comparación estadística en primera instancia, consistió en realizar una matriz de datos
para cada ecosistema (Bosque de niebla y Páramo), se trabajó con un N (cantidad de
individuos) de 40, se les realizo a cada uno mediciones de: grosor de la cutícula tanto adaxial
como abaxial, grosor de la pared y área del lumen de 4 células adaxiales y 4 células
abaxiales. Posterior a tener la matriz totalmente diligenciada, se ingresaron los datos en el
programa PAST 3.17, para realizar el test de Mann-Whitn U, el cual se utiliza para datos no
paramétricos, con el fin de determinar si habían diferencias significativas entre las
mediciones de bosque de niebla y páramo, así pues, es importante tener en cuenta que para
que la comparación sea significativa el p del test de la prueba estadística debe tener un valor
inferior o igual a 0,05.
Teniendo en cuenta lo anterior, respecto a los resultado de los test de las imágenes 8, 9, 10 y
11 que se muestran a continuación se obtuvo un p en cada uno de estos que no fue inferior o
igual a 0,05 y las medias de cada ecosistema no mostraron una diferencia relevante la uno de
la otra, lo cual significo que las mediciones entre ecosistemas fueron similares haciendo que
no existiera una diferencia significativa para obtener una correcta comparación.
Imagen 8. Resultados de la comparación de cutícula adaxial
entre páramo y bosque de niebla. Páramo (A); Bosque de niebla (B); cantidad de individuos (N); Rango
promedio (Mean Rank); Probabilidad (p).

33
Imagen 9. Resultados de la comparación de cutícula abaxial
entre páramo y bosque de niebla. Páramo (D); Bosque de niebla (E); cantidad de individuos (N); Rango
promedio (Mean Rank); Probabilidad (p).
Imagen 10. Resultado de la comparación del lumen de las células epidérmicas abaxiales
entre páramo y bosque de niebla. Páramo (M); Bosque de niebla (N); cantidad de individuos (N); Rango
promedio (Mean Rank); Probabilidad (p).

34
Imagen 11. Resultados de la comparación de la pared de las células epidérmicas abaxiales
entre páramo y bosque de niebla. Páramo (P); Bosque de niebla (Q); cantidad de individuos (N); Rango
promedio (Mean Rank); Probabilidad (p).
En cuanto al resultado de la comparación del lumen de las células epidérmicas adaxiales de
páramo y bosque de niebla que se muestra en la imagen 12 se puede extraer que sí existe una
diferencia significativa ya que el p fue de 0,001 el cual es inferior de 0,05, esto también se
corroboro al observar que sus medias fueron considerablemente desiguales. Así pues,
analizando los datos, la diferencia del tamaño del lumen de las células epidérmicas adaxiales
entre páramo y bosque de niebla es relevante, encontrando que el lumen en páramo va a ser
más pequeño, con una media de 16,15 µm en comparación con el de bosque de niebla, que
va a tener una media de 24,35 µm.
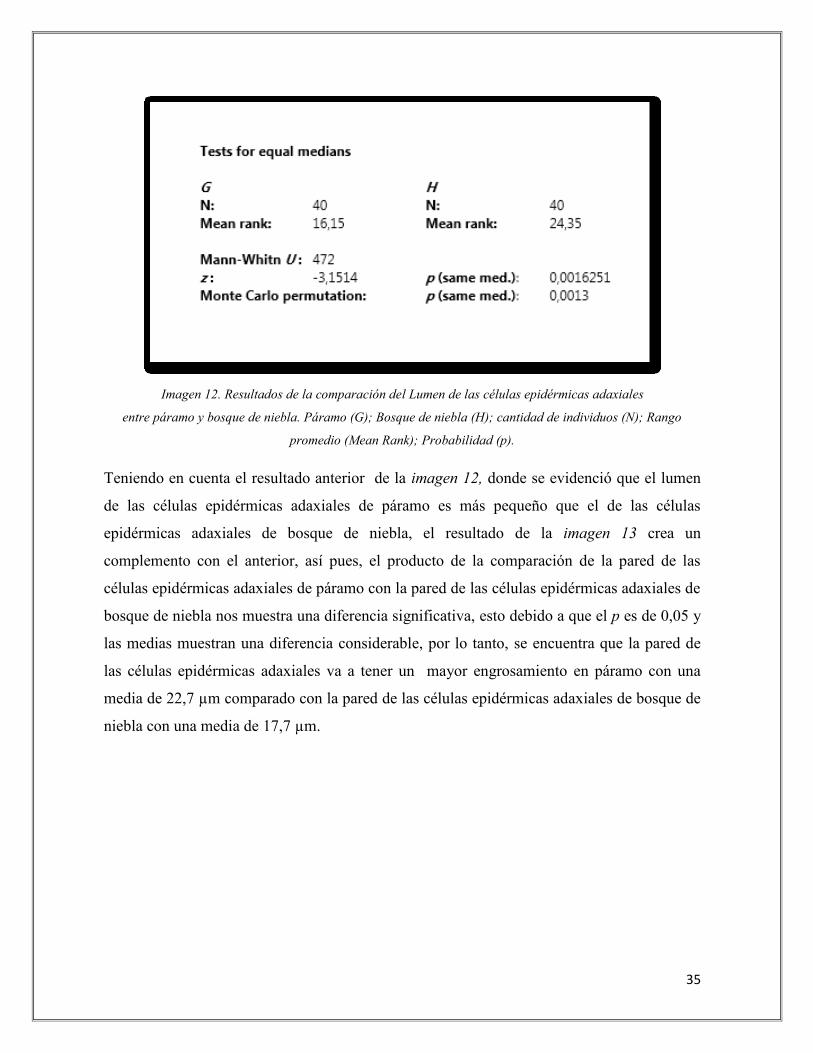
35
Imagen 12. Resultados de la comparación del Lumen de las células epidérmicas adaxiales
entre páramo y bosque de niebla. Páramo (G); Bosque de niebla (H); cantidad de individuos (N); Rango
promedio (Mean Rank); Probabilidad (p).
Teniendo en cuenta el resultado anterior de la imagen 12, donde se evidenció que el lumen
de las células epidérmicas adaxiales de páramo es más pequeño que el de las células
epidérmicas adaxiales de bosque de niebla, el resultado de la imagen 13 crea un
complemento con el anterior, así pues, el producto de la comparación de la pared de las
células epidérmicas adaxiales de páramo con la pared de las células epidérmicas adaxiales de
bosque de niebla nos muestra una diferencia significativa, esto debido a que el p es de 0,05 y
las medias muestran una diferencia considerable, por lo tanto, se encuentra que la pared de
las células epidérmicas adaxiales va a tener un mayor engrosamiento en páramo con una
media de 22,7 µm comparado con la pared de las células epidérmicas adaxiales de bosque de
niebla con una media de 17,7 µm.

36
Imagen 13. Resultados de la comparación de la pared de las células epidérmicas adaxiales
entre páramo y bosque de niebla. Páramo (J); Bosque de niebla (K); cantidad de individuos (N); Rango
promedio (Mean Rank); Probabilidad (p).
Por consiguiente, en los dos anteriores resultados al analizarse datos de células epidérmicas
adaxiales se considera una relación, la cual propone que dichas células epidérmicas presentes
en páramo van a tener un lumen menor (imagen 12) debido a que su pared celular va a
mostrar un engrosamiento significativo (imagen 13).
37
8. DISCUSIÓN
En la histología de los cortes transversales de hoja se encontró un tejido de protección que
está ubicado al inicio y final de cada corte, este según Megías et al., (2017) está constituido
por células epidérmicas las cuales sintetizan y secretan a la parte externa una sustancia
lipídica impermeable denominada cutina que se deposita en la superficie externa de la pared
celular para formar una capa llamada cutícula, se pudo observar claramente en las imágenes
de los cortes transversales de hoja de Gaultheria; está capa tiene un color rosado y se
encontró tanto en la cara adaxial como abaxial del limbo de la hoja. La cutícula se compone
por una cubierta superior de ceras epicuticulares, seguida por otra capa inferior formada por
cutina y ceras mezcladas con sustancias de la pared celular (pectinas, celulosa y otros
carbohidratos), cumpliendo la función de reducir la pérdida de agua actuando como una
eficaz barrera hidrofóbica protectora para minimizar dicha pérdida por evapotranspiración
(Tafolla et al., 2013).
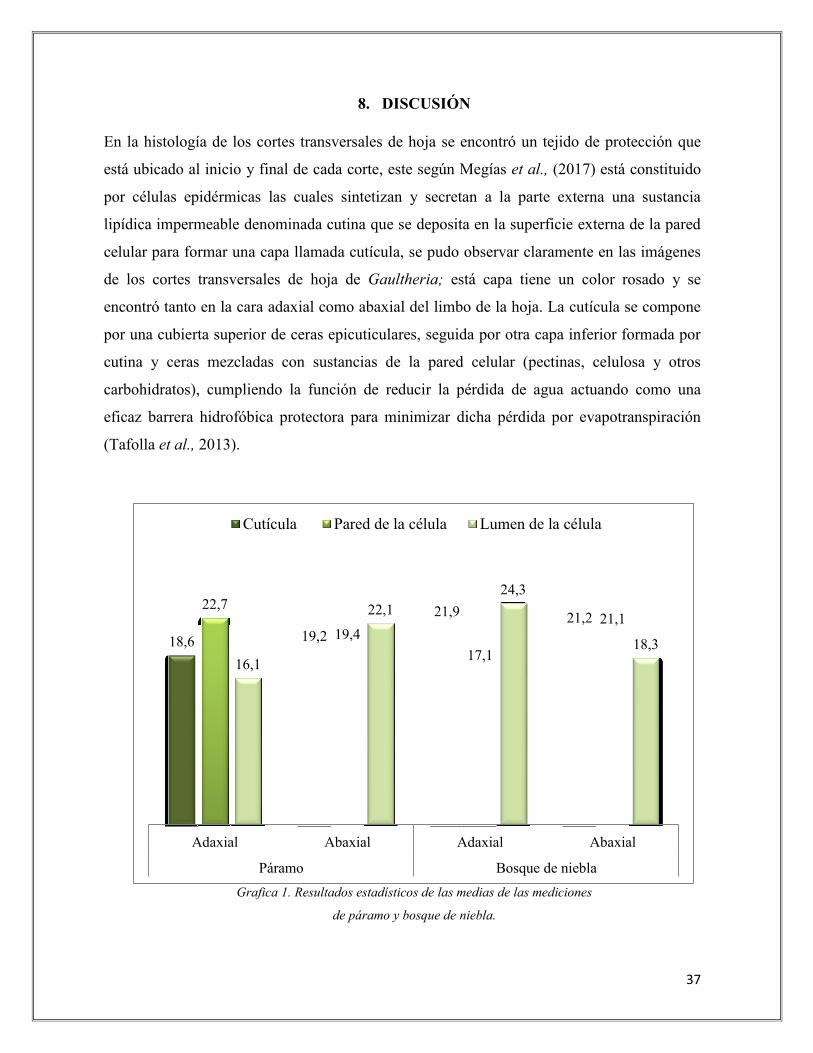
Grafica 1. Resultados estadísticos de las medias de las mediciones
de páramo y bosque de niebla.
18,6 19,2
21,9 21,2 22,7
19,4
17,1
21,1
16,1
22,1
24,3
18,3
Adaxial Abaxial Adaxial Abaxial
Páramo Bosque de niebla
Cutícula Pared de la célula Lumen de la célula

38
Analizando la gráfica 1 se pudo evidenciar que en cuanto a la cutícula, tanto de la cara
adaxial como abaxial, no existe una diferencia significativa entre los ecosistemas de páramo
y bosque de niebla, sin embargo, la capa de cutícula adaxial presentó un grosor mayor en
bosque de niebla con una media de 21.9 µm en comparación con páramo que fue de 18,6
µm; el grosor de la capa cuticular según Lallana et al., (2006) varía dependiendo las especies
vegetales, la zona de la planta y su edad o estado de desarrollo, tal cómo se evidencio en las
hojas de Gaultheria que crecen en bosque de niebla, cuyo grosor de la cutícula fue más
amplio, ya que esta zona no tiene tanta incidencia de luz como el ecosistema de páramo,
donde por factores como este provocarán un aumento de la temperatura dentro de las hojas,
haciendo que la transpiración sea mayor, por ende se presente pérdida de agua debido a la
reducción en esta capa cuticular.
Ahora bien, en cuanto a las células epidérmicas de las hojas de Gaultheria estas se disponen
unidas muy estrechamente, adquiriendo formas y tamaños muy variados que se suelen
adaptar a la forma de la estructura que recubren, como se evidencia en los cortes
transversales; teniendo en cuenta el análisis estadístico realizado se arrojaron resultados
donde existen diferencias significativas entre estas células epidérmicas de bosque de niebla
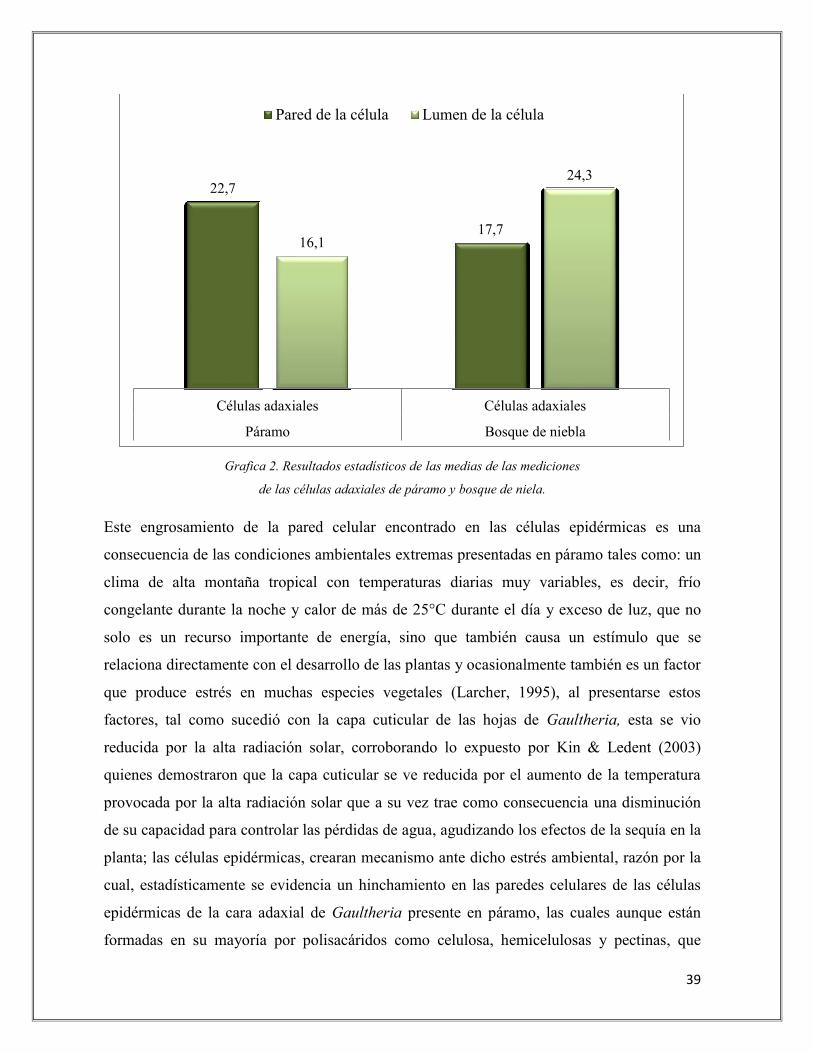
con las células epidérmicas de páramo, analizando la gráfica 2 se observó que la pared
celular de las células epidérmicas de la cara adaxial de las hojas presentes en páramo tienen
un promedio de 22,7 µm mientras que en bosque de niebla la pared celular de las células
epidérmicas de hoja tienen un promedio de 17,7 µm, indicando con ello que el grosor de las
paredes de las células epidérmicas es mayor en páramo. Así mismo, se obtuvo que el lumen
de las células epidérmicas de la cara adaxial de las hojas de Gaultheria presentes en páramo
tuvieron un promedio de 16,1 µm siendo menor frente al de las hojas de Gaultheria presentes
en bosque de niebla, el cual fue mayor con un promedio de 24,3 µm; por consiguiente al
presentarse un engrosamiento en la pared celular, el lumen se verá directamente afectado y
mostrará un tamaño más reducido tal como se pudo evidenciar en las hojas de Gaultheria
presentes en páramo.
39
Grafica 2. Resultados estadísticos de las medias de las mediciones
de las células adaxiales de páramo y bosque de niela.
Este engrosamiento de la pared celular encontrado en las células epidérmicas es una
consecuencia de las condiciones ambientales extremas presentadas en páramo tales como: un
clima de alta montaña tropical con temperaturas diarias muy variables, es decir, frío
congelante durante la noche y calor de más de 25°C durante el día y exceso de luz, que no
solo es un recurso importante de energía, sino que también causa un estímulo que se
relaciona directamente con el desarrollo de las plantas y ocasionalmente también es un factor
que produce estrés en muchas especies vegetales (Larcher, 1995), al presentarse estos
factores, tal como sucedió con la capa cuticular de las hojas de Gaultheria, esta se vio
reducida por la alta radiación solar, corroborando lo expuesto por Kin & Ledent (2003)
quienes demostraron que la capa cuticular se ve reducida por el aumento de la temperatura
provocada por la alta radiación solar que a su vez trae como consecuencia una disminución
de su capacidad para controlar las pérdidas de agua, agudizando los efectos de la sequía en la
planta; las células epidérmicas, crearan mecanismo ante dicho estrés ambiental, razón por la
cual, estadísticamente se evidencia un hinchamiento en las paredes celulares de las células
epidérmicas de la cara adaxial de Gaultheria presente en páramo, las cuales aunque están
formadas en su mayoría por polisacáridos como celulosa, hemicelulosas y pectinas, que
22,7
17,7 16,1
24,3
Células adaxiales Células adaxiales
Páramo Bosque de niebla
Pared de la célula Lumen de la célula

40
constituyen alrededor del 90% del peso seco de las paredes celulares, también acumularán
lignina y cutina que al ser de naturaleza hidrófoba desplazará el agua e impermeabilizara la
pared (Azcón & Talón 2000) ayudando de esta manera, para que la transpiración en esta
planta sea menor y no muera por deshidratación. De igual modo observando las imágenes 3 y
5 de los cortes transversales de Gaultheria anastomosans se evidencia dos capas de células
que hacen pate del tejido de protección, una capa externa que corresponde a la epidermis y
otra interna llamada hipodermis, esta última capa según Cosa & Dottori (2010) cumple la
función de aumentar la resistencia celular ante la deshidratación, acumular agua y proteger
contra el exceso de radiación solar.
La hipodermis también se presentó en los individuos presentes en bosque de niebla, ya que el
lugar de donde se colectaron los individuos no tiene una diferencia de altura relevante con
los individuos colectados en páramo, también en este bosque de niebla no se presenta una
cobertura vegetal abundante debido a que es un bosque con intervención antrópica haciendo
que estas plantas de Gaultheria queden expuestas a la radiación ultravioleta, lo cual es una
razón para justificar la presencia de dicha hipodermis en ambos ecosistemas.
Aunque Gaultheria anastomosans se considera como una planta mesófita que según Font
(2000) son organismos que se desarrollan en ambientes con temperaturas y humedad de tipo
medio, ni muy altas ni muy bajas, estas modificaciones en sus tejidos, como el
engrosamiento de la pared celular de las células epidérmicas y el posterior desarrollo de
hipodermis, se estiman como una adaptación xeromórfica en respuesta a su plasticidad de
adaptación a las condiciones adversas, presentándose, tanto en áreas con una alta intensidad
lumínica como en zonas con déficit hídrico o húmedas (Andersen et al., 2006).
Así pues, al ser la hoja el órgano de la planta más sensible a las condiciones ambientales, es
donde más se puede reflejar cambios como una forma de adaptación (Nughes et al., 2013)
esto se atribuye como un mecanismo de plasticidad, entendiendo la plasticidad según Reyes
y otros (2001) como un mecanismo que favorece a las especies ante condiciones
heterogéneas, creando la capacidad de adaptarse a diferentes condiciones ambientales,
ajustando su morfología y fisiología a través de la variación genética y la plasticidad en su
forma, pudiendo incluir respuestas a corto y largo plazo.

41
Siguiendo con la descripción de tejidos, después de las células epidérmicas se encontró el
mesófilo, constituido, como se observa en las imágenes 2, 3 y 4, por un parénquima
clorofílico en empalizada biestratificado y un parénquima lagunoso. El primer parénquima se
encuentra en la cara adaxial de la hoja y es el más expuesto al sol, tiene un mayor número de
cloroplastos, sus células estarán más empaquetadas y allí se lleva a cabo una mayor tasa de
fotosíntesis y el segundo parénquima se caracteriza por tener más espacios intercelulares
gracias a los cuales se dará un intercambio de gases y agua eficiente con la atmósfera
(Megías et al., 2017). El desarrollo del parénquima clorofílico en empalizada en la cara
adaxial de la hoja de Gaultheria anastomosans nos indicó que esta planta tiene una simetría
interna bifacial que externamente se corroboró al observar que la cara adaxial de dichas hojas
es de un color más oscuro frente al color de la cara abaxial que es más claro, por lo cual se
asume que la posición que tienen las hojas en las ramas es horizontal causando que estén
recibiendo los rayos del sol en mayor cantidad en la cara adaxial, esto se considera como otra
razón por las que se presenta un engrosamiento de la pared celular de las células
epidérmicas adaxiales de dicha planta y no en las células epidérmicas de la cara abaxial,
pues al crecer de manera horizontal esta cara no recibe directamente los rayos del sol.
En cuanto a la nervadura central de la hoja de Gaultheria anastomosans se encontró primero
un tejido de sostén compuesto por células de colénquima en la cara adaxial y abaxial,
caracterizado por ser vivo, tener paredes celulares desigualmente engrosadas y tener una
forma elongada en dirección del eje principal, dándole una gran resistencia a la tensión y a
otros tipos de estrés mecánico (Megías et al., 2017), y en segunda medida, como se puede
observar en la imagen 7, encontramos un anillo de esclerénquima de color rosado, compuesto
por sus células muertas, con las paredes lignificadas y engrosadas (Martín, 2012), que rodea
al tejido conductor de floema y xilema en estadios tempranos; con base en lo anterior, al no
evidenciar entre el xilema y floema primario procambium (Pérez et al., 2013) se considera
que la planta posee un haz vascular colateral cerrado, el cual según Gatusso & Gatusso
(2016) es típico de Angiospermas, plantas con flor como Gaultheria, dándole resistencia
mecánica a la nervadura principal y al limbo foliar de esta planta. La disposición de las
células del protoxilema frente a las de metaxilema nos indica que este xilema primario es
endarco, porque las células de protoxilema se encuentran hacia el interior y el metaxilema se

42
desarrolla de manera centrífuga, lo cual es típico de una Eustela presente en eudicotiledoneas
como Gaultheria anastomosans.

43
9. CONCLUSIONES
En cuanto a la histología de hoja de Gaultheria anastomosans presente en bosque de niebla y
páramo esta presentó una semejanza en la distribución de sus tejidos en ambos ecosistemas,
evidenciaron adecuadamente parénquima, tejidos de sostén, tejidos vasculares y tejidos de
protección.
Por otro lado, debido a la alta radiación solar que se da en los ecosistemas de páramo, la capa
cuticular se verá reducida, provocando que la planta cree mecanismos xeromórficos como
defensa, que se evidenciaron estadísticamente al encontrar diferencias significativas en las
células epidérmicas de los individuos presentes allí los cuales mostraron un engrosamiento
de la pared celular e histológicamente encontrando hipodermis, estas alteraciones se crean en
busca de la protección de los tejidos y como mecanismo para la retención de agua en la
planta, considerándose esto como una adaptación frente a los factores adversos en los que
crece Gaultheria anastomosans.
Así pues, teniendo en cuenta lo anterior, se estima que las modificaciones en los tejidos de
Gaultheria anastomosans son una adaptación de plasticidad de la planta para lograr una
supervivencia y permanencia en estas zonas con condiciones ambientales extremas.

44
10. RECOMENDACIONES
Metodológicamente se recomienda a la hora de realizar la colecta de los individuos, tener en
cuenta factores como pH del suelo, incidencia lumínica y el estrato del arbusto del cual se
tomó el órgano, ya que son variables que también están directamente relacionadas con las
diferentes modificaciones que realizan las plantas como mecanismo de defensa; por otro lado
en cuanto a fase de laboratorio, se recomienda hacer primero una prueba del protocolo y
según los resultados modificar tiempos o reactivos a la hora de trabajar con el material
biológico, esto debido a que con cada especie de planta y órgano de la misma puede variar
para un resultado óptimo.
También, debido a que la histología vegetal es un tema poco trabajado y aún más en nuestro
país, los referentes bibliográficos son escasos, por lo cual se sugiere una ampliación en la
investigación de esta área para extender el conocimiento de la misma.

45
11. BIBLIOGRAFÍA
Andersen, A., Lucchini, F., Moriconi, E. & Fernández, A. (2006). Variabilidad en la
morfo-anatomía foliar de Lippia turbinata (Verbenaceae) en la provincia de San Luis
(Argentina). Facultad de Química Bioquímica y Farmacia, Universidad Nacional de San
Luis.
Armenteras, D., Cadena, C. & Moreno, P. (2007). Evaluación del estado de los bosques
de niebla y de la meta 2010 en Colombia. Instituto de Investigación de Recursos
Biológicos Alexander von Humboldt.
Azcón, J. & Talón, M. (2000). Fundamentos de fisiología vegetal. Mc Graw-Hill.
Interamericana. Ediciones Universidad de Barcelona. Barcelona, España.
Cárdenas, L. (2015). Comparación histológica foliar de Gaultheria anastomosans (l. f.)
kunth de la familia Ericaceae presente en bosque de niebla y páramo del parque
ecológico Matarredonda vía Bogotá-Choachí. Facultad de ciencias y educación.
Universidad Distrital Francisco José de Caldas. Bogotá, Colombia.
CAR, Corporación Autónoma Regional de Cundinamarca. (2004). Estrategia corporativa
para la caracterización con fines de manejo y conservación de áreas de páramo en el
territorio CAR. Bogotá D.C.: CAR. Informe Final.
CORTOLIMA. (2007). Plan de Ordeneación y Manejo de la Cuenca Hidrográfica Mayor
del Río Totare. Obtenido de
www.cortolima.gov.co/default/files/images/stories/centro_documentos/pom_totare/diagn
ostico/kj_210111_flora.pdf.
Flórez, G., Núñez, O., Núñez, N., Ramírez, L., Ramírez, M. & Zusunaga, J. 100 plantas
útiles del páramo Rabanal. Guía para comunidades rurales. Instituto De Investigación De
Recursos Biológicos Alexander Von Humboldt. Corporación Autónoma Regional de
Cundinamarca. Corporación Autónoma Regional de Boyacá. Corporación Autónoma
Regional de Chivor. Boyacá, Colombia.
Fundación Humedales de Bogotá. (16 de Agosto de 2016). Matarredonda Parque
Ecológico. Obtenido de Caminata al Parque Ecológico de "Matarredonda" Parque

46
Ecológico: http://humedalesbogota.com/2016/08/16/caminata-al-parque-ecologico-
matarredonda/.
Garzón, O. (2017). Estudio histológico de tallo de Gaultheria anastomosans presente en
páramo y bosque de niebla. Facultad de ciencias y educación. Universidad Distrital
Francisco José de Caldas. Bogotá, Colombia.
Gatusso, M & Gatusso, S. (2016). Estructura del cuerpo de las espermatófitas.
Universidad Nacional de Rosario Facultad de Ciencias Bioquímicas y Farmacéuticas
Kin, A. & Ledent, J. (2003). Efecto del viento sobre las plantas. viento, suelo y planta.
Santa rosa.
Lallana, M., Billard, C., Elizalde, J., & Lallana, V. (2006). Breve revisión sobre
características de la cutícula vegetal y penetración de herbicidas. Cátedra Fisiología
Vegetal, Facultad de Ciencias Agropecuarias, Universidad Nacional de Entre Ríos –
UNER.
Larcher, W. (1995). Physiological plant ecology. 5th ed. Springer-Verlag. Berlin.
Laskowsk, E. (2000). Características anatómicas de la hoja y el tallo del Semeruco
(Malpighia emarginata dc) cultivado en dos localidades del estado de Lara. Dpto. de
Ciencias Biológicas. Universidad Centroccidental Lisandro Alvarado. Barquisimeto,
Venezuela.
Llambí, D., Soto, A., Celleri, R., Bievre, B., Ochoa, B. & Borja, P. (2012). Ecología,
hidrología y suelos de páramos. Proyecto Páramo Andino.
Lozano, G. & Schnetter, R. (1976). Estudios ecológicos en el Páramo de Cruz Verde,
Colombia II. Las comunidades vegetales. Departamento de Biología, Universidad
Nacional de Colombia. Bogotá, Colombia.
Madriñan, S. (2010). Flora ilustrada del Páramo de Chingaza. Guía de campo de plantas
comunes. Segunda edición. Facultad de ciencias. Departamento de ciencias biológicas.
Universidad de los Andes. Bogotá, Colombia.
Marín, S. & Parra, S. (2015). Bitácora de flora. Páramos vivos. Guía visual de plantas de
páramos en Colombia. Instituto de Investigación de Recursos Biológicos Alexander von
Humboldt.

47
Martín, S. (2012). Estudio de los tejidos para la caracterización de las plantas.
Departamento de Biología Vegetal II. Facultad de Farmacia. Universidad Complutense.
Madrid, España.
Megías, M., Molist, p. & Pombal, M. 2017. Atlas de histología vegetal y animal.
Departamento de biología funcional y ciencias de la salud. Facultad de biología.
Universidad de Vigo. España.
Morales, M., Otero, J., Van der Hammen, T., Torres, A., Cadena, C., Pedraza, C. &
Cárdenas, L. (2007). Atlas de Páramos de Colombia. Bogotá: Instituto de Investigación
de Recursos Biológicos Alexander von Humboldt.
Murillo, R., Piedra, G., & Leon, G. (2012). Absorción de nutrientes a través de la hoja.
UNICIENCIA Vol. 2. Costa Rica.
Nughes, L., Colares, M., Hernández, M. & Arambarri, N. (2013). Morfo-anatomía de las
hojas de celtis ehrenbergiana (celtidaceae) desarrolladas bajo condiciones naturales de
sol y sombra. Laboratorio de Morfología Comparada de Espermatofitas (LAMCE),
Facultad de Ciencias Agrarias y Forestales, Universidad Nacional de La Plata. La Plata,
Argentina.
Ortíz, L. & Reyes Bonilla, M. (2009). Páramos en Colombia: Un Ecosistema Vulnerable.
Medio Ambiente, 9.
Paniagua, R., Nistal, M., Sesma, P., Álvarez, M., Fraile, B., Anadón, R. & Sáez, F.
(2002). Citología e histología vegetal y animal. 3 edición. Editorial McGraw-Hill.
Pérez, S., Martín, D., Alemán, M., Etcheverry, A., Figueroa, T., Quiroga, M., López, D.
& Yañéz, C. (2013). Tejidos vegetales. Botánica agrícola- biología de las plantas –
botánica general.
Reyes, J., & Martínez, D. (2001). La Plasticidad de las Plantas. Elementos 41.
Rodriguez Erazo, N., Armenteras, D., Morales, M., & Romero, M. (2004). Ecosistemas
de los Andes Colombianos. Bogotá: Instituto de Investigaciones y Recursos Biológicos
Alexander Humboldt.

48
Sandoval, E. (2005). Técnicas aplicadas al estudio de la anatomía vegetal. México:
Cuadernos 38.
Sánchez, D. (1998). Inventario florístico del páramo de frontino (Urrao – Antioquia).
Departamento de Biología. Facultad de ciencias. Universidad Nacional de Colombia.
Medellín, Colombia.
Sánchez, L. & Gelviz, S. (2004). Aspectos florísticos y fitogeográficos de Pamplona.
Grupo de Recursos Naturales, Instituto de Ciencias Naturales y Biotecnología,
Universidad de Pamplona. Pamplona, Colombia.
Tafolla, J., González, A., Tiznado, M., García, L. & Báez, R. (2013). Composición,
fisiología y biosíntesis de la cutícula en plantas. Centro de Investigación en Alimentación
y Desarrollo, A. C. Sonora, México. Instituto de Agroquímica y Tecnología de
Alimentos, Consejo Superior de Investigaciones Científicas. Valencia, España.
Teillier, S. & Escobar, F. (2013). Revisión del género Gaultheria L. (Ericaceae) en Chile.
Escuela de Arquitectura del Paisaje. Universidad Central, Santa Isabel. Santiago de
Chile, Chile.
Valencia, D., Chongo, B. (2004). La pared celular. Influencia de su naturaleza en la
degradación microbiana ruminal de los forrajes. Instituto de Ciencia Animal. Revista
Cubana de Ciencia Agrícola, Tomo 38, No. 4. La Habana, Cuba.