glucolisis
TRANSCRIPT

METABOLISMO INTERMEDIARIO
Glucólisis

Importancia de la glucólisis
• Ruta casi universal• Primer ruta que fue conocida con detalle• Es una ruta central del metabolismo:
– En el sentido catabólico las hexosas son degradadas hasta piruvato con lo que se gana energía química y poder reductor
– En el sentido anabólico, el piruvato y otros metabolitos aportan esqueletos carbonados para biosíntesis.

Antecedentes: La fermentación de los azúcares
• Transformación por distintos microorganismos para obtener diferentes productos: vino, cerveza (etanol), pan, lácteos fermentados (ácido láctico)
• Buchner - 1897 – Fermentación sin células• Harden/Young - 1905 El fosfato estimula la
fermentación de la glucosa• Embden/Meyerhof/Warburg - 1930s –
descubren las reacciones de la glucólisis

Panorama general
• Es una ruta lineal• Consta de 10 reacciones• El balance global es:
1 glucosa → 2 piruvato 2 ADP → 2 ATP 2 NAD+ → 2 NADH
• Presenta dos fases:– Fase preparativa o de
inversión de energía– Fase de beneficios o de
generación de energía

Fase preparativa o de inversión de energía
• Se requiere ATP para activar (o cebar) la molécula de glucosa.
• Se usan dos mol de ATP por mol de glucosa.
• El resultado son dos triosas fosfato

Reacción 1
• Fosforilación de la glucosa por la hexocinasa (HK) usando ATP con una ∆Gº’ muy negativa.
• La hexocinasa puede fosforilar otras hexosas: manosa, galactosa, fructosa
• La reacción es inhibida por el producto• En eucariontes la hexocinasa está unida a la
membrana mitocondrial externa

Reacción 2
• Catalizada por la fosfohexosa isomerasa (PGI o PHI)
• Isomerización aldosa – cetosa a través de un intermediario enediol.

Reacción 3
• Segunda fosforilación• Catalizada por la fosfofructocinasa (PFK1)• En este punto se han completado todas las entradas de
energía de la glucólisis.• Es una reacción clave de la ruta• La enzima es regulada alostéricamente por distintos
factores

Reacción 4
• Lisis no hidrolítica de la fosfohexosa en dos triosas fosfato• Es la reacción que da nombre a la ruta• La enzima es la Fructosa-1,6-bisfosfato aldolasa (ALD)
El sustrato es estabilizado formando una base de Schiff con la enzima

Reacción 5
• Isomerización de una triosa fosfato en otra (dihidroxiacetona fosfato en gliceraldehído-3-fosfato)
• Catalizada por la triosa fosfato isomerasa (TPI)• El posterior uso del G3P desplaza el equilibrio
hacia su formación

Fase de beneficios o ganancias
• Conversión del gliceraldehído -3-fosfato en piruvato y formación acoplada de ATP

Reacción 6• Oxidación y fosforilación catalizada por la
gliceraldehído-3-fosfato deshidrogenasa (G3PDH)
• El producto es una molécula de energía superelevada
La fosforilación utiliza fosfato inorgánico
• Los electrones son donados para reducir al NAD+
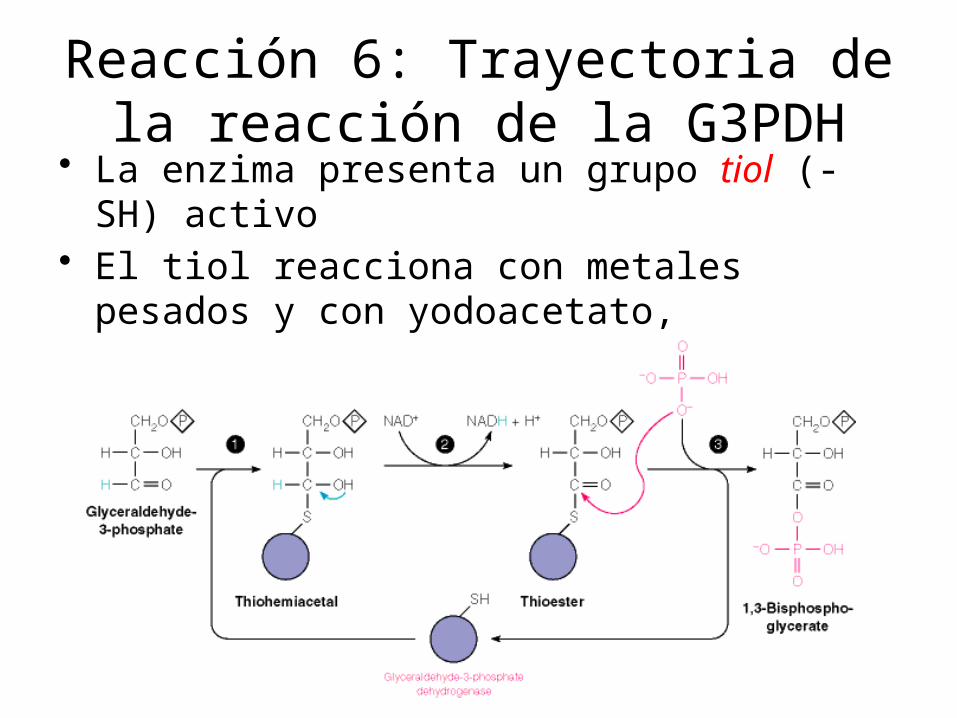
Reacción 6: Trayectoria de la reacción de la G3PDH
• La enzima presenta un grupo tiol (-SH) activo• El tiol reacciona con metales pesados y con
yodoacetato, inhibiendo a la enzima

Reacción 7
• Primera fosforilación a nivel de sustrato catalizada por la fosfoglicerato cinasa (PGK)
• El metabolito de energía súper-elevada dona el fosfato al ADP
• Hasta este paso, la ganancia neta de ATP es cero
• Como se trata de dos triosas, son dos ATP

Reacción 8
• Isomerización que convierte al fosfoglicerato en un buen donador de grupo fosfato
• La enzima es la fosfoglicerato mutasa (PGM)

Reacción 9
• Reducción por deshidratación del 2-fosfoglicerato que produce otra molécula de energía súper-elevada: el fosfoenolpiruvato (PEP)
• El cambio en el potencial de energía libre de hidrólisis del fosfato es de -15.6 kJ/mol en el 2PG a -61.9 kJ/mol en el PEP
• La enzima es la enolasa (ENO)

Reacción 10
• El PEP dona su fosfato al ADP en una reacción virtualmente irreversible
• Es la segunda fosforilación a nivel de sustrato• El ATP formado representa una ganancia de 2
mol• La enzima piruvato cinasa (PK) es regulada
alostéricamente

Balance global de la glucólisis• La variación
total de energía libre en condiciones fisiológicas hace más favorable la glucólisis en células con alta demanda de ATP

Perfil energético de la glucólisisLas reacciones de la HK, la PFK1 y la PK son muy exergónicas y limitan la velocidad de flujo de la ruta (o sea, una vez entrando a esos pasos, la dirección de la ruta está comprometida)
El resto de las reacciones son prácticamente reversibles

Principales tipos de reacción
• Transferencias de fosfato: participan un donador y un aceptor
• Transferencias de electrones (H:-) por óxido-reducción
• Rompimiento de enlace C-C

Aclaración 1: Fosforilación a nivel de sustrato
• Es una de las 3 formas de producir ATP
• Consiste en que una molécula sirve como donadora de fosfato para el ADP debido a que presenta una energía libre de hidrólisis de fosfato más negativa que el mismo ATP

Aclaración 2: La glucosa no es el único sustrato de la glucólisis
• Distintas moléculas pueden ser utilizados, alimentando la glucólisis en pasos diferentes

Aclaración 3: La glucólisis no es la única vía de oxidación de la
glucosa• Vías alternativas son:
– Vía de las pentosas fosfato o del fosfogluconato: se obtiene NADPH y ribosa-P que puede ser reciclada a G6P (6 pentosas-P→5 hexosas-P)
– Conversión en ácido urónico: importante en la desintoxicación y excreción de compuestos ajenos al cuerpo
– Conversión en ácido ascórbico: presente sólo en algunos organismos pero no en vertebrados

Aclaración 4: El destino del piruvato depende del blance redox• En condiciones anaerobias, es necesario
reoxidar al NADH en NAD+ mediante la transferencia de electrones a un aceptor
• Algunos microorganismos usan como aceptor iones inorgánicos (sulfato, nitrato) y otros sustratos orgánicos
• La mayoría de los organismos obtienen lactato al reducir al piruvato usando la lactato deshidrogenasa en la fermentación láctica (LDH)
• Otra fermentación importante es la alcohólica

Destino del piruvato

Glucólisis aerobia y anaerobia
• La oxidación de la glucosa y otras fuentes de carbono puede ocurrir en presencia o ausencia de oxígeno.
• El oxígeno actúa como aceptor final externo de electrones, y en este caso hablamos de respiración aerobia.
• En ausencia de oxígeno pero otros aceptores finales externos de electrones muchos organismos llevan a cabo una respiración anaerobia
• En ausencia total de aceptores finales ocurre la fermentación.

Fermentación
• Ruta metabólica productora de energía en la cual no hay un cambio neto en el estado de oxidación de los productos en comparación con los sustratos
• Literalmente significa “ebullición a temperatura ambiente”
• Algunos productos de importancia son: – Etanol– Ácidos orgánicos (acético, láctico, propiónico,
butírico, fórmico)– Amoniaco: a partir de aminoácidos (de la
fermentación putrefactiva y alcalina)

Glucólisis aerobia
• Es parte de la respiración aerobia• El piruvato es descarboxilado
oxidativamente por el complejo piruvato deshidrogenasa
• El NADH es oxidado en la cadena respiratoria
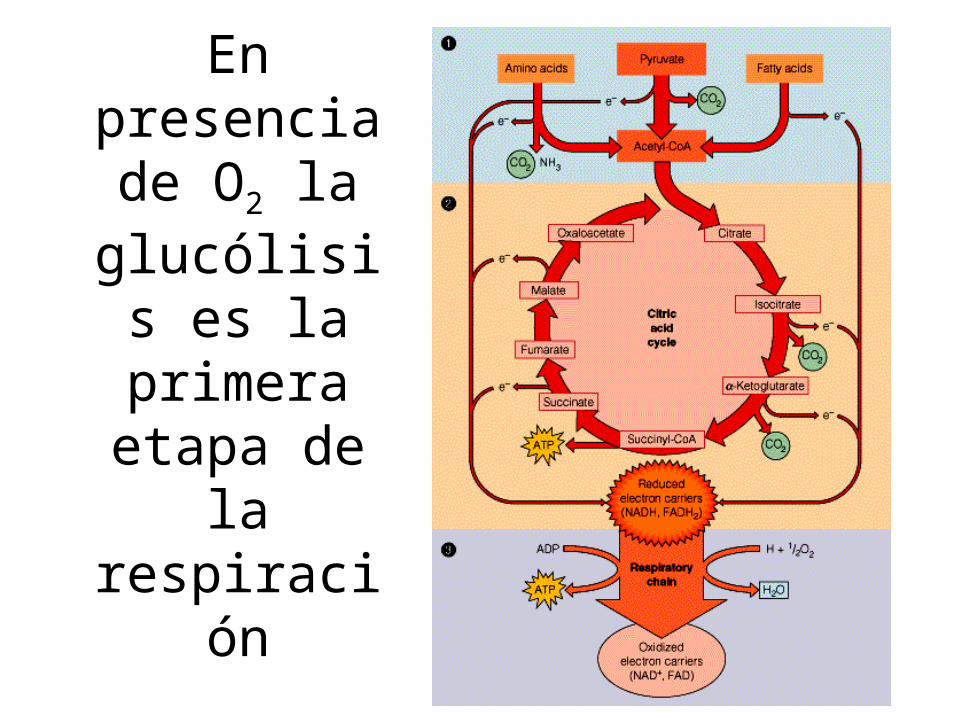
En presencia de O2 la
glucólisis es la primera etapa de la respiración

Regulación de la glucólisis
• Evita que ocurran simultáneamente la glucólisis y la gluconeogénesis (ruta “inversa”)
• Hay mecanismos de:– Control enzimático (alosterismo e inhibición por
producto)– Retroinhibición (feed-back) y proactivación (o
activación anterógrada, feed-forward)– Control hormonal por insulina, glucagón, etc.– Compartimentación en el citoplasma

Regulación de la glucólisis
• La hexocinasa es inhibida por el producto (G6P)
• La fosfofructocinasa 1 (PKF1) está regulada por la carga energética (niveles de ATP/ADP) y por los moduladores alostéricos fructosa-2,6-bisfosfato, y citrato, entre otros
• La piruvato cinasa es estimulada por proactivación e inhibida por retroinhibición

Las fluctuaciones en NADH demuestran que la PFK1 está regulada por los niveles de O2
• Al añadir una cantidad limitada de sustrato fermentable (glucosa), las oscilaciones en niveles de ATP están desfasadas de las de NADH

La PFK1 es regulada alostéricamente
La fructosa-2,6-bisfosfato aumenta la afinidad de la PFK1: modulador alostérico positivo
El ATP modula negativamente: a mayor concentración, menor afinidad, es decir, la PFK1 está regulada por la carga energética

Compartimentación
• Las reacciones se llevan a cabo en regiones localizadas del
citoplasma en muchos organismos

Compartimentación
• En eucariontes la HK está unida a la cara externa de las mitocondrias

Compartimentación
• En tripanosomas existe el glucosoma: orgánulo que contiene las enzimas de las primeras 7 reacciones

Compartimentación
• Hay canalización de sustratos: éstos pasan directamente de un sitio activo al siguiente, sin ser hidratados en el paso (deducido a partir de las velocidades de reacción comparadas con la velocidad de difusión)
• Las reacciones ocurren en complejos multienzimáticos

Regulación del metabolismo de la
glucosa

Carácter anabólico de la
glucólisis