fÓsiles virtuales
TRANSCRIPT

FÓSILES VIRTUALES
Nuevos avances tecnológicos en Paleontología
Introducción
Una de las principales obsesiones de los paleontólogos es el poder observar y comprender detalladamente el interior de los fósiles sin alterarlos o romperlos. Durante muchos años se utilizó técnicas de preparación, con un grado destructivo, para acceder y estudiar al misterio que ocultaban las rocas o restos fósiles. Estas técnicas desfavorables permitían acceder al interior, pero por consecuencia, se alteraba las muestras.
En la actualidad, mediante el avance tecnológico, existe un método favorable para acceder al interior de los fósiles sin dañarlos y así estudiar sus características internas. Esta técnica, muy avanzada, es la tomografía de rayos X y originalmente se creó para la medicina, con el fin de visualizar tejidos y huesos, pero ha sido de una gran ayuda en la disciplina paleontológica.
Este informe describe la técnica empleada en un pequeño fósil marino desarrollado en el Cámbrico de España.
La primera diversificación importante de la vida animal tuvo lugar durante la "explosión" del Cámbrico. En aquella época, la vida animal se hallaba confinada en el océano. Durante gran parte del período Cámbrico (hace entre 542 y 490 millones de años), la mayoría de los animales carecían de la capacidad de excavar profundamente en el sedimento. Por lo que, las corrientes de fango submarinas sepultaron a los animales que vivían en el fondo, sus cadáveres quedaban al abrigo de carroñeros excavadores, con lo que dejaron un registro fósil muy rico.
Descripción del fósil
Nombre: Protocinctus mansillaensis Rahman y Zamora 2009 Ubicación: España, Zaragoza Distribución: Cámbrico medio Descripción: pocos centímetros de longitud y de color blanco brillante

Figura 1. Ejemplar perfectamente conservado del carpoideo cincta Protocinctus mansillaensis Rahman y Zamora 2009 procedente del Cámbrico medio, España. Museo Paleontológico de la
Universidad de Zaragoza-Gobierno de Aragón.
Con apenas un par de centímetros de longitud y de color blanco brillante, este fósil resalta claramente sobre la matriz rojiza que lo rodea (Figura 1). Este animal pertenece a los equinodermos, grupo en el que hoy en día se incluyen también, animales tan familiares para todos nosotros, como el erizo o la estrella de mar.
Los equinodermos presentan un registro fósil bastante completo, si lo comparamos con el que muestran otros filos animales, debido a la facilidad de fosilización de sus componentes calcíticos. Esto les permite ser unos buenos candidatos para estableces los cambios evolutivos sufridos desde sus primero representantes.
Paleoecología de los primeros equinodermos
Existen numerosas estrategias que los primeros equinodermos utilizaron para alimentarse y para interaccionar con el sustrato. Las principales modificaciones que aparecen en sus planes corporales pueden explicarse, en parte, por el desarrollo de un modo de vida u otro. Existen formas libres como los carpoideos que vivieron apoyados sobre el sustrato. Los cincta presentan una forma de raqueta muy apta para la vida en fondos blandos. A lo largo de la evolución del grupo, los cincta desarrollaron túmulos ventrales que les permitieron elevar la teca sobre el sustrato. Su apéndice posterior lo utilizaban para estabilizar al organismo frente a las corrientes (Rahman y Zamora, 2009) (Fig. 11). La alimentación se produciría a partir de la inyección de flujos de agua a través de la boca hacia el interior de la teca. Allí, una faringe interna captaría las partículas nutritivas. El sobrante de agua se expulsaba a través del opérculo (Friedrich, 1993; Smith, 2005; Zamora et al., 2007b) Los tenocistoideos presentan una morfología muy parecida, y algunos

autores han sugerido que su movimiento se realizaría por propulsión a chorro (Robison y Sprinkle, 1969), aunque otros descartan esta idea y los consideran principalmente sésiles (Rahman y Clausen, 2009). Se alimentaban captando partículas nutritivas en la interfase agua-sedimento mediante el tenoide (Fig. 11). Los estilóforos también son formas libres que se alimentaban filtrando el agua y captando partículas alimenticias cerca de la interfase agua-sedimento. Según la interpretación funcional de sus partes, el alimento lo capturarían mediante el paso del agua por una faringe interna, de forma análoga a los cincta (Smith, 2005). En este caso la boca se situaría en la parte opuesta al apéndice. La otra interpretación explica que la alimentación se producía utilizando el apéndice a modo de brazo, en cuyo caso la boca se situaría en la zona más proximal de la región distal del apéndice (Ubaghs, 1968; Lefebvre, 2003).
Figura. Clase Soluta. A. El soluto Scalenocystites strimplei, del Devónico inferior de Minnesota, Norte América (Universidad de Zaragoza). B. Reconstrucción del soluto Castericystis vali, del
Cámbrico medio de Norte América (tomado de Ubaghs y Robison, 1985).
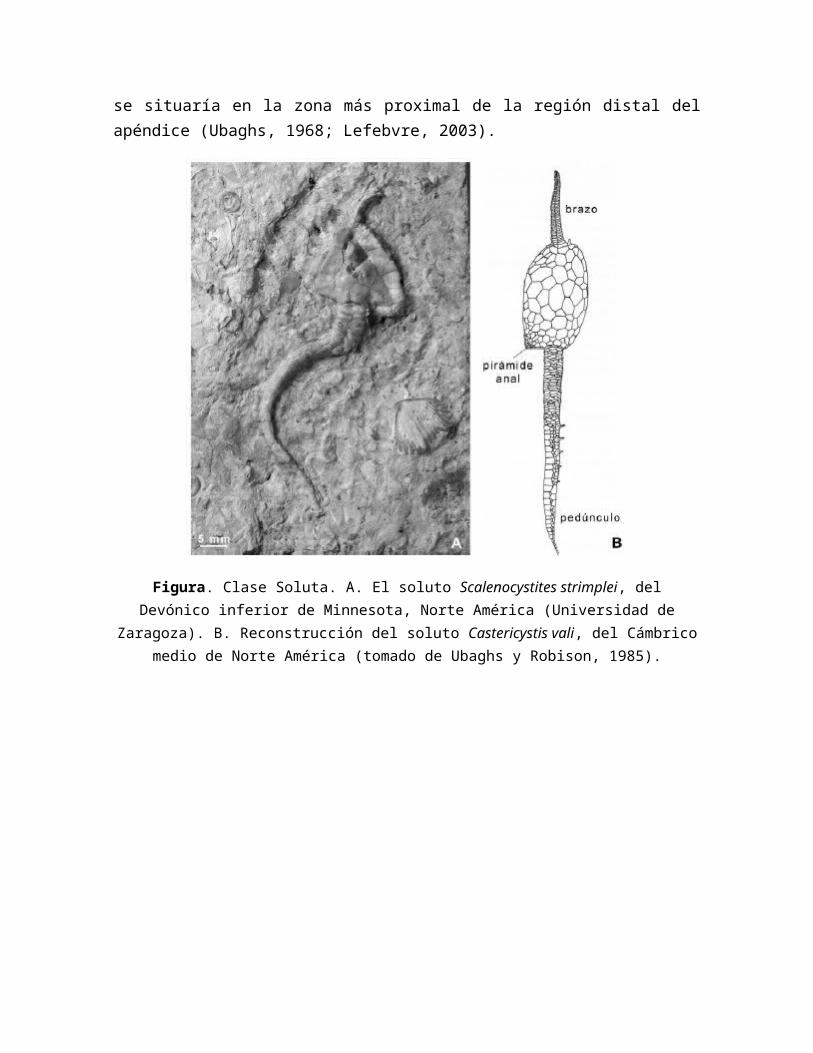
Figura. Clase Helicoplacoidea. A. Ejemplar del helicoplacoideo, Helicoplacus gilberti del Cámbrico inferior de Norte América. B. Tipos de ambulacros descritos en helicoplacoideos
(según Wilbur, 2006), el ambulacro de Helicoplacus gilberti (B1-2) es más simple y presenta menos series de placas coberteras que el de Waucobella nelsoni (C1-C2). Abreviaciones: series
de placas coberteras (1º-5º cp), placas basales del ambulacro (fp), placas romboidales (rp).
Lo que parece estar claro es que el apéndice estaría relacionado en parte con la locomoción de estos organismos y así lo han demostrado unas pistas fósiles procedentes del Devónico de Alemania (Rahman et al. 2009) (Fig. 10). Los solutos tienen formas de vida más variadas, géneros como Coleicarpus, y los estadios juveniles de Castericystis, se fijaban a fragmentos de conchas por lo que eran sésiles.
Los adultos de Castericystis (Fig. 11), y todos los taxones post-cámbricos tienen un modo de vida libre (vágil) con cierta capacidad de movimiento (Daley, 1995; 1996).
El brazo situado en posición anterior lo utilizaban para captar el alimento, seguramente partículas orgánicas del sedimento en descomposición.
Los edrioasteroideos, eocrinoideos y helicoplacoideos son formas sésiles. Los edrioasteroideos vivían en el Cámbrico fijados directamente al sustrato (Fig. 11). La revolución de los sustratos cámbricos (Dornbos, 2006) obligó a estos taxones a buscar otras formas de fijación, como los restos de conchas o los “hardgrounds”. Estos últimos empezaron a ser abundantes a partir del Ordovícico inferior proporcionando un nuevo nicho para ser colonizado.
Hasta ahora se pensaba que los edrioasteroideos se cementaban a restos de conchas a partir del Ordovícico inferior, pero el reciente descubrimiento de los primeros isorófidos fijados a mudas de trilobites en las rocas del Cámbrico medio español, sugiere que estas estrategias comenzaron mucho antes de lo que se pensaba (Zamora y Smith, en revisión).
Los eocrinoideos cámbricos son formas que vivieron fijas a restos de conchas (Fig. 11) o directamente sobre el sustrato (Sprinkle, 1973; Lin et al., 2008; Zamora et al., 2009). Las braquiolas se utilizaban para captar el alimento mediante filtración. Existe un grupo, los lichenoididos, que aparentemente vivían apoyados directamente sobre el sustrato (Parsley y Prokop, 2004) (Fig. 11). No es descartable que algunos taxones introdujeran parte de la teca en el sedimento para fijarse (Dornbos, 2006).
Los helicoplacoideos vivían insertando uno de sus polos en el sustrato (Sprinkle y Wilbur, 2005) (Fig. 11), y captaban alimentos abriendo las placas coberteras de los ambulacros y enfrentándolos a las corrientes.
En resumen se puede hablar de cuatro estrategias utilizadas por los equinodermos cámbricos en relación al sustrato: formas libres como los carpoideos, que tienen una estrategia tipo raqueta de nieve, formas que viven permanentemente fijadas al sustrato mediante succión (algunos edrioasteroideos y eocrinoideos), formas que se fijan a restos de conchas (eocrinoideos y edrioasteroideos isorófidos) y formas que introducen parte de la teca en el sustrato (helicoplacoideos y algunos eocrinoideos).
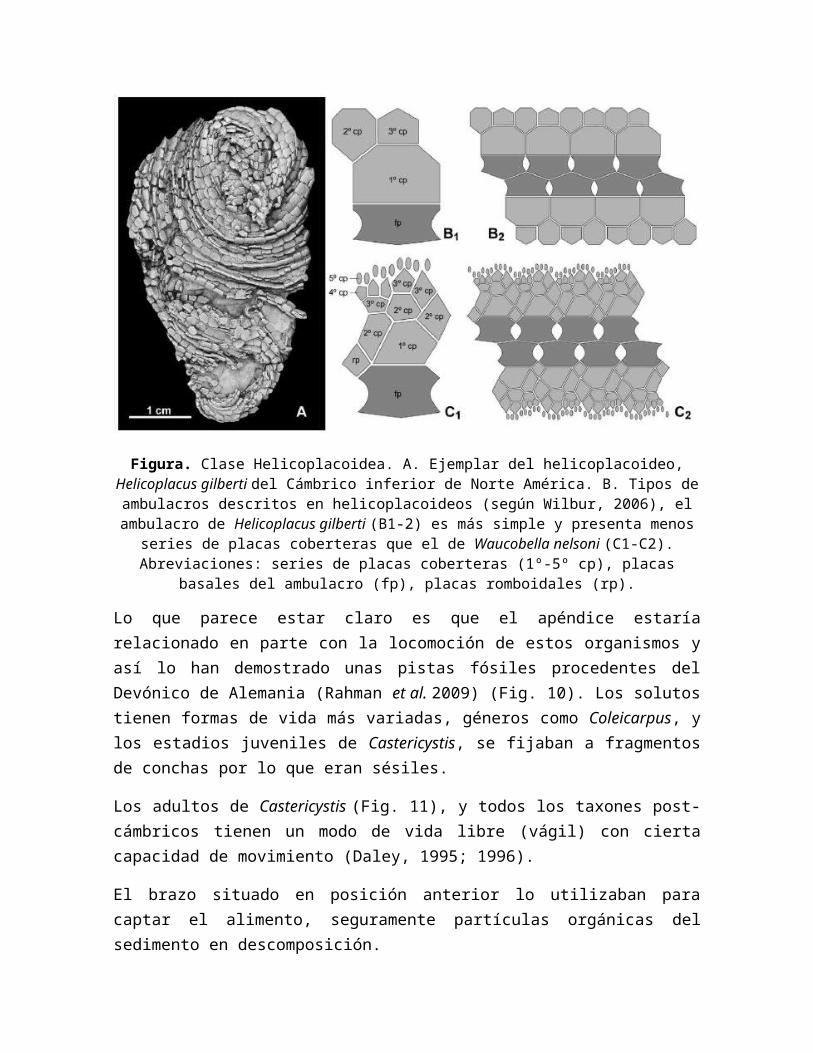
Figura. A. Ejemplar del estilóforo mitrado Rhenocystis que quedó atrapado tras dar sus últimos coletazos (Devónico inferior, Alemania). Estos ejemplares sirvieron para demostrar
que, al menos algunos estilóforos, usaban el apéndice para propulsarse. B. Reconstrucción de dicho movimiento (Tomado de Sutcliffe et al. 2000).

Respecto al tipo de alimentación, los equinodermos cámbricos fueron filtradores o depositívoros. La principal diferencia entre los distintos equinodermos filtradores radica en la altura de la columna de agua que explotaban. Algunos carpoideos explotaban la interface agua-sedimento captando partículas re-suspendidas, mientras que formas como eocrinoideos capturaban las partículas que caían verticalmente en suspensión. Los edrioasteroideos son formas filtradoras que captan alimento de la columna de agua a diferentes alturas. Los solutos, algunos estilóforos y quizás otros carpoideos capturaban partículas nutritivas sobre el sustrato (depositívoros o deposit feeders). Sobre los helicoplacoideos se sabe muy poco en este aspecto.
Los primeros equinodermos
Los equinodermos presentan un registro fósil bastante completo, si lo comparamos con el que muestran otros filos animales, debido a la facilidad de fosilización de sus componentes calcíticos. Esto les permite ser unos buenos candidatos para establecer los cambios evolutivos sufridos desde sus primeros representantes.
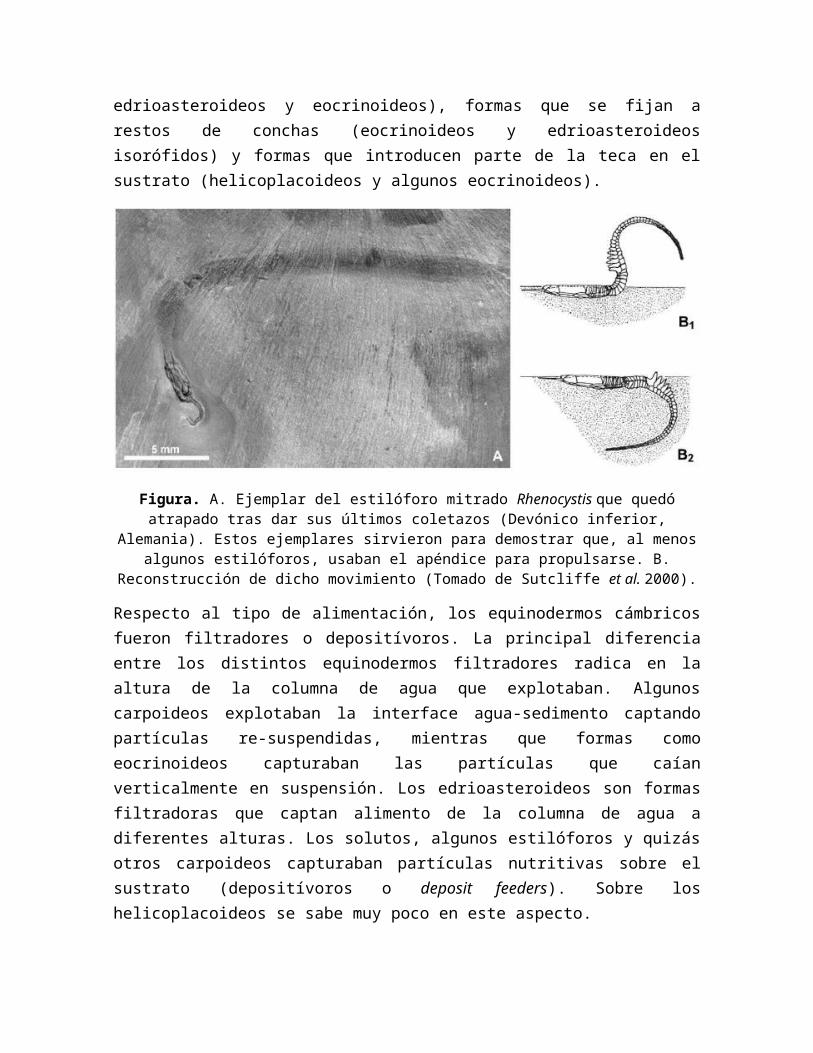
Figura. Reconstrucción de una comunidad de equinodermos del Cambrico hipotética donde aparecen taxones de diferentes edades y distintas regiones geográficas. 1. Helicoplacoideo
(Helicoplacus), 2. Estilóforo (Ceratocystis), 3. Eocrinoideo (Gogia), 4. Soluto (Castericystis), 5. Edrioasteroideo (Cambraster), 6. Eocrinoideo (Lichenoides), 7. Tenocistoideo (Ctenocystis), 8.
Cincta (Gyrocystis).
El registro fósil de equinodermos comienza en el Cámbrico inferior con la primera aparición en Norte América de los Helicoplacoideos y edrioasteroideos (Sprinke, 1992). A estos les siguen los eocrinoideos (Durham, 1978), que también aparecen a la vez en otras regiones de Gondwana (Ubaghs y Vizcaino, 1990; Zhao et al. 2007; Hu et al., 2007). La presencia en el Cámbrico medio de numerosos clados de equinodermos que están ampliamente distribuidos geográficamente (Sprinkle, 1992; Zamora, en prensa), sugiere fuertemente que muchos de estos clados debieron originarse durante el Cámbrico inferior, pero todavía no han sido encontrados. Esto nos advierte de lo incompleto que es el registro fósil de equinodermos en el Cámbrico inferior y de lo mucho que queda por descubrir.
El Cámbrico y el origen de los equinodermos
Durante el Cámbrico, hace más de 500 millones de años el mundo era muy diferente al de hoy. Las masas continentales se dividían en cuatro grandes continentes (Laurentia, Siberia, Báltica, Gondwana), no existían casquetes polares y los mares estaban poblados de criaturas extrañas. La gran explosión de la vida, que se dio durante este periodo, produjo la aparición de casi todos los filos de animales que hoy viven en nuestro planeta, como por ejemplo los equinodermos.
Los equinodermos son organismos exclusivamente marinos y cuya principal característica es la presencia de un esqueleto mesodérmico de calcita con una microestructura tridimensional denominada esteroma (Figura 2). La construcción de este esqueleto tan particular y exclusivo de los equinodermos fue una novedad importante que se dio en el Cámbrico inferior cuando los principales representantes de este grupo aparecieron en los mares.
Cualquier curioso de la naturaleza se ha fijado alguna vez en la perfecta simetría que presenta una estrella de mar con cinco brazos dispuestos en ángulos de 72° cada uno (fifura 3). Este plan corporal se ajusta a lo que se conoce como simetría radial de tipo pentámero y es otra de las principales características que presentan los equinodermos actuales (figura 3).
El sistema vascular acuífero es una especie de sistema hidráulico, y es otra de las características importantes de los equinodermos. Se compone de una serie de conductos y reservorios de líquido cuya función principal es controlar la presión hidráulica en el

interior del caparazón. Este sistema se abre al exterior por una serie de aberturas en la teca (caparazón) situadas en las ambulacros y que permiten proyectar al exterior los pies ambulacrales. Estos pueden verse, por ejemplo, en los erizos o las estrellas de mar actuales y controlan funciones básicas como el movimiento, respiración o la alimentación. (fig 3).
De todas estas características solo el esqueleto de calcita está presente en todos los equinodermos cámbricos. Algunos (fig 4), como los carpoideos cincta o los tenocistoideos (fig 5 6) no presenta simetría radial y son asimétricos o casi bilaterales. Otros carecen de ambulacros, como los estilófiros, pero presenta internamente una faringe para alimentarse. Algunos desarrollan una simetría muy particular, por ejemplo los helicoplacoideos que tienen tres ambulacros enroscadas formando un helicoide. Todas estas formas de equinodermos mantienen en jaque a los paleontólogos, ya que son muy raras y poseen estructuras nunca antes vistas. Actualmente, a partir de nuevos fósiles, como el que se muestra en este informe, gracias al avance de la biología molecular, se está tratando de esclarecer y comprender el origen de este grupo de animales.
Rocas del Cámbrico en el Parque Natural de Moncayo
Estas rocas son el resultado de la sedimentación en una plataforma marina poco profunda y con temperaturas cálidas.
Figura. Sistema vascular acuífero de un equinodermo actual con simetría radial.
Esta forma de raqueta es una característica de un grupo primitivo y extinto de equinodermos, los cincta. Se desarrollaron sólo durante el Cámbrico medio, en mares someros de Godnwana y Siberia, es decir de lo que en la actualidad es Europa, Africa y Asia.

En un equinodermo encontrado en prácticas de exploración, se encontraron fósiles con una excelente conservación, aunque es imposible obtener de forma directa información de ellas. Los datos más importantes se encuentran en las partes dorsal y frontal que en este caso están ocultas en la matriz rojiza. Puede perderse información muy valiosa si se intenta preparar el fósil con técnicas tradicionales.
Para aplicar este tipo de técnica es preferible que el fósil se encuentre en un buen estado de conservación. Esto permite realizar estudios muy novedosos acerca de cuestiones nunca antes abordadas, como el modo de vida o la reconstrucción de la anatomía interna, pero primero es necesario acceder a estar partes.
Etapas del desarrollo de la técnica
El complejo estudio que se desarrolla es por medio de una microtomografía computerizada de rayos X. Se debe primero, preparar las muestras mecánicamente, para después escanearlas y ver si la técnica es posible de aplicar. Si se puede aplicar esta técnica, es necesario tratar de una forma digital los resultados para después completar una reconstrucción del fósil. La siguiente etapa es el estudio e interpretación de los datos obtenidos.
Este tipo de trabajo abarca varios años de investigación para luego ser publicado y dar a conocerlo al mundo.
Escáner del fósil
Se utiliza sierras con un disco de diamante para reducir la laja donde se encuentra el fósil al menor tamaño posible y así poder introducir la muestra en el tomógrafo y obtener una mejor resolución ante los rayos X. Este procedimiento se debe de realizar con mucho cuidado, ya que algún movimiento en falso puede causar que se parta la roca y con ella el fósil.
El resultado del procedimiento se obtiene una pequeña pastilla de 25x15x5 milímetros lista para ser escaneada.
A través de la roca: Más allá de lo que el ojo humano puede ver
Los análisis de escaneo se efectuaron en el laboratorio de microtomografía de rayos X del Imperial College de Londres.
Luego de 3 horas se obtiene el escáner del fósil. Se obtiene un total aproximado de 500 rodajas en 2 dimensiones del fósil. Cada rodaja se realiza cada 20 a 30 micras aproximadamente.

Cada rodaja se analiza independientemente en una computadora. El objetivo es doble, por un parte se debe de separar el fósil de la roca encajonante. Este proceso es relativamente sencillo, ya que los cristales calcíticos tienen una composición muy diferente de la roca arcillosa. Por ello los rayos X atraviesan cada composicoon de manera difenrente lo cual se obtiene diferentes respuestas en el receptor. Estos resultados se muestran con un alto contraste en las imágenes que se obtienen, gris-blanco (calcita) y negro (roca arcillosa).
La segunda parte del análisis es la más difícil, consiste en distinguir y separar por colores todas las partes que constituyen el esqueleto del equinodermo (fig 7). Cada imagen se trata de manera cuidadosa, finalmente se obtiene la anatomía original del fósil, lo cual estará al servicio de los paleontólogos para su estudio detallado y clasificación.
Figura. Diferentes etapas de la reconstrucción del cincta Protocinctus mansillaensis Rahman y Zamora, 2009. A. Sección directamente extraída del microtomógrafo,; las diferentes placas del
esqueleto aparecen en tonos grises. B. Sección donde se individualiza el fósil (blanco) de la matriz circundante. C. Sección con la interpretación de las placas que constituyen el esqueleto
en distintos colores. D. Reconstrucción frontal del ejemplar a partir de la combinación de todas las secciones.

Figura. Reconstrucción en vista frontal del tenecisteoideo Ctenocystis del Cámbrico medio de Utah (EE. UU.). En el centro de las placas con forma de gancho (ctenidiales) y cubiertas por
ellas se encontraría la boca.
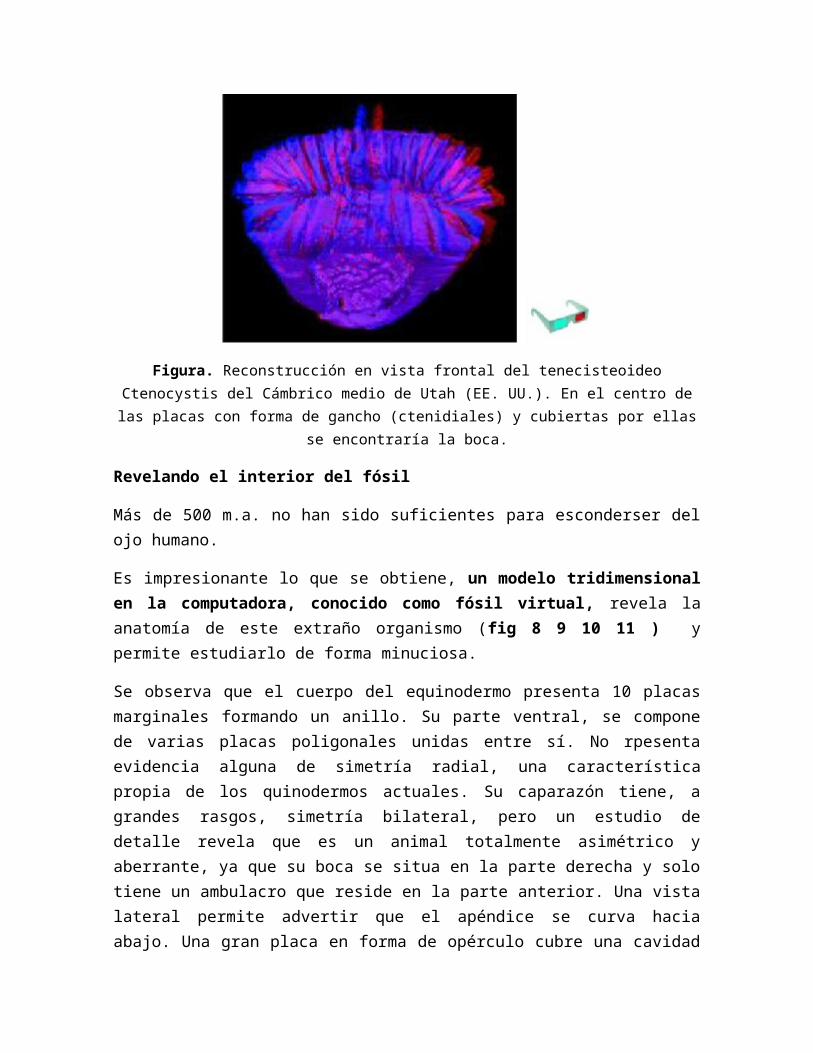
Revelando el interior del fósil
Más de 500 m.a. no han sido suficientes para esconderser del ojo humano.
Es impresionante lo que se obtiene, un modelo tridimensional en la computadora, conocido como fósil virtual, revela la anatomía de este extraño organismo (fig 8 9 10 11 ) y permite estudiarlo de forma minuciosa.
Se observa que el cuerpo del equinodermo presenta 10 placas marginales formando un anillo. Su parte ventral, se compone de varias placas poligonales unidas entre sí. No rpesenta evidencia alguna de simetría radial, una característica propia de los quinodermos actuales. Su caparazón tiene, a grandes rasgos, simetría bilateral, pero un estudio de detalle revela que es un animal totalmente asimétrico y aberrante, ya que su boca se situa en la parte derecha y solo tiene un ambulacro que reside en la parte anterior. Una vista lateral permite advertir que el apéndice se curva hacia abajo. Una gran placa en forma de opérculo cubre una cavidad anterior (fis 11). En su interior aparece una gran depresión, lugar que en vida ocuparon los órganos principales.
Presenta muchas características únicas, por lo que se bautizó como con nuevo nombre, Protocinctus mansillaensis.

Figura. Reconstrucción final de Protocinctus mansillaensis Rahman y Zamora, 2009 en vista oblicua. En ella se observan las diferentes partes anatómicas y el interior del esqueleto.
Figura. Reconstrucción de Protocinctus mansillaensis Rahman y Zamora, 2009 en vista oblicua. Observar las placas supracentrales que están desarticuladas y se disponen flotando
sobre la teca.

Comparar a Protocinctus con sus parientes actuales
Cuando se estudian fosiles que se tiene representantes entre los animales actuales, es relativamente fácil hacer comparaciones directas y obtener conclusiones. Es los que en Geología se llama Actualismo, y que consiste en estudiar los procesos actuales para comprender como ocurrieron en el pasado. A medida que se retrocede en el tiempo existen un mayor número de variables que dificultan estas comparaciones. Protoncinctus es un casi extremo, ya que pertenece a un periodo muy antiguo y su morfología no se parece en nada a la que presentan sus parientes actuales.
Si se tuviera que incluir a Protoncinctus dentro de algún grupo de animales se situaría junto a los equinodermos, ya que presenta el esqueleto formado por placas de calcita. Un estudio reciente de la secuencia genética del erizo de mar, ha revelado que los genes que construyen el esqueleto de calcita típico de los equinodermos son exclusivamente de este filo animal; por lo tanto, cualquier animal del pasado con estas placas fue seguramente un equinodermo. Sin embargo Protoncinctus presenta unas características que lo hacen muy diferente de otros equinodermos actuales. Presenta cola, no tiene simetría radial, una gran placa opercular cubre su parte anterior y sólo tiene un ambulacro.
La biología molecular actual reconoce que los equinodermos están muy emparentados con otro grupo de animales marinos, los hemicordados o gusanos bellota (fig 12), cuyo pariente fósil más familiar para los paleontólogos son los graptolitos. Ambos, hemicordados y equinodermos compartieron un ancestro común. Estos dos grupos junto al de los cordados (al que nosotros pertenecemos) forman los deuteróstomos (fig 12). Esto significa que, a grandes rasgos, la estrella de mar, los humanos y los graptolitos compartimos un ancestro común.
Los genes de hemicordados y equinodermos son muy parecidos. Pero, existen unas características morfológicas que nos llaman tremendamente la atención cuando comparamos a un hemicordados con un equinodermo. Mientras que los primeros son bilaterales, los segundos tienen simetría radial, además los hemicordados no tienen caparazón de calcita y presenta hendiduras branquiales. Las larvas de hemicordados y equinodermos son muy parecidas entre sí. Ambas presentan simetría bilateral. En el caso de los hemicordados la larva se desarrolla progresivamente hasta constituir un adulto muy parecido a la larva, pero más grande. Es un crecimiento más o menos continuo.
Sin embargo, el desarrollo de la larva de los equinodermos revela cambios muy drásticos en su morfología. En un momento dado, la larva bilateral sufre una metamorfosis en la que algunas partes se atrofian y nunca son desarrolladas en el adulto. Tras una etapa de

metamorfosis, en la que tenemos una larva aberrante asimétrica, el adulto emerge ya con simetría radial.
Regresemos a Protoncinctus, muchas de sus características recuerdan a las que se observan en los hemicordados y en las larvas de los equinodermos. Su esqueleto asimétrico con cola, su boca en posición derecha, la presencia de una sólo ambulacro o la faringe interna perforada por hendiduras branquiales, son características que seguramente fueron primitivas dentro del grupo de los equinodermos. A medida que los equinodermos evolucionaron introdujeron novedades en su morfología, alejándose de su condición ancestral (primitiva). No es posible comparara directamente a Protoncinctus con los equinodermos actuales porque, a pesar de pertenecer al mismo grupo, es muy primitivo. Más de 500 millones de años de evolución lo alejan de sus representantes actuales, de ahí que su morfología sea tan diferente.
La importancia de Protocinctus
Durante la gran “explosión de la vida” que se dio durante el Cambrico, los equinoermos experimetaron su origen y diversificación. Algunos grupos como los concta surgieron durante un breve espacio de tiempo, con una anatomía muy extraña y terminaron por extinguirse. fueron un experimento más de la vida, que nucan llegó a tener el suficiente éxito para llegar a nuestros días. Us estudio arroja luz importante a como eran los primero equinoermos un grupo de animales que sigue con éxito en todos los mares actuales.
Conclusiones
Las formas de equinodermos cámbricos presentados muestran un amplio rango de planes corporales. O existe un consenso sobre su paleobiología o posición filogenética. Esto se debe principalmente a la ausencia de formas actuales parecidas y que permitan estableces comparaciones.
Los fósiles son la única herramienta que tenemos para reconstruir los primeros pasos en la evolución de los equinodermos.
Los Protocinctus presentaban una raqueta que era ideal apra apoyarse sobre el sustrato fangoso donde vivía. La estela (cola) se curvaba para penetrar en el sedimento y poder anclarse, de esta manera el Protocinctus evitaba ser desplazado por las correintes.
Con la ayuda de nuevas tecnologías se está logrando alcanzar y obtener nuevas fronteras en el conocimiento paleontológico.
Figura. El cladograma representado permite situar a Protocinctus en la base del grupo de los equinodermos. Sin embargo, muchas de sus características son comunes a los hemicordados más primitivos y a otros enigmáticos equinodermos, como los tenocystoideos. Estos grupos
no presentan simetría radial, característica típica de los equinodermos actuales.
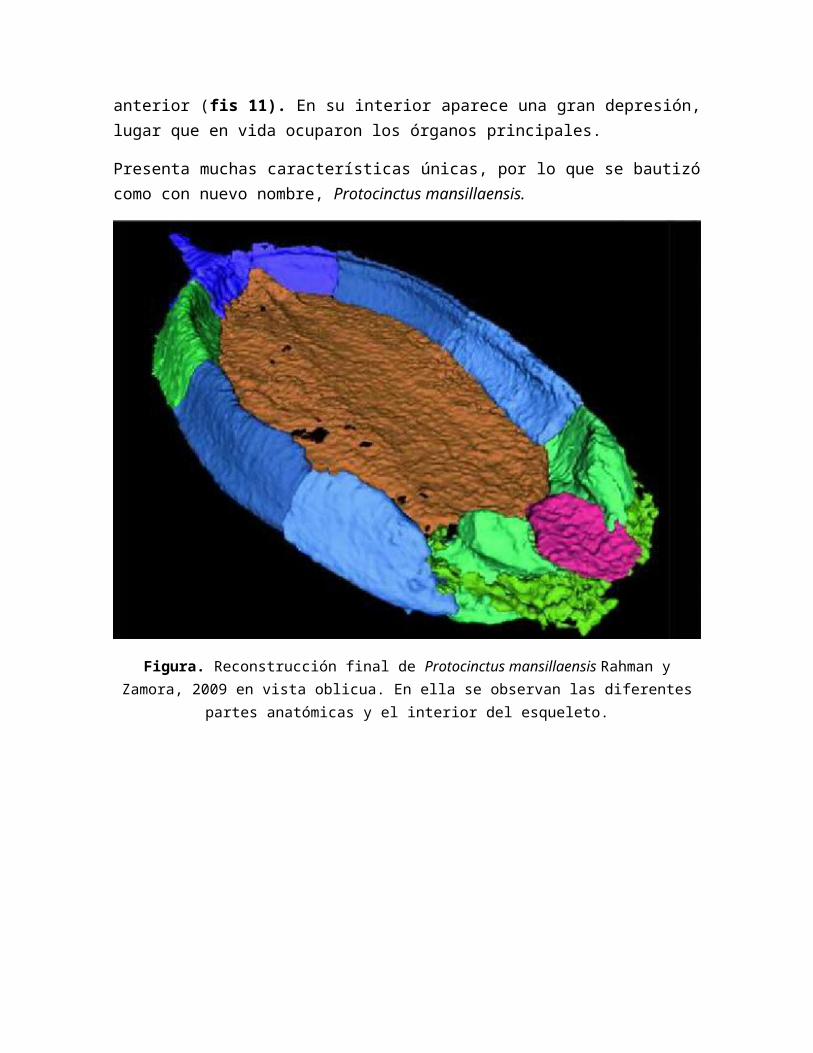
Figura. Otro de los ejemplares de Protocinctus escaneados. Se observa como una gran opérculo cubre la cavidad del cuerpo.
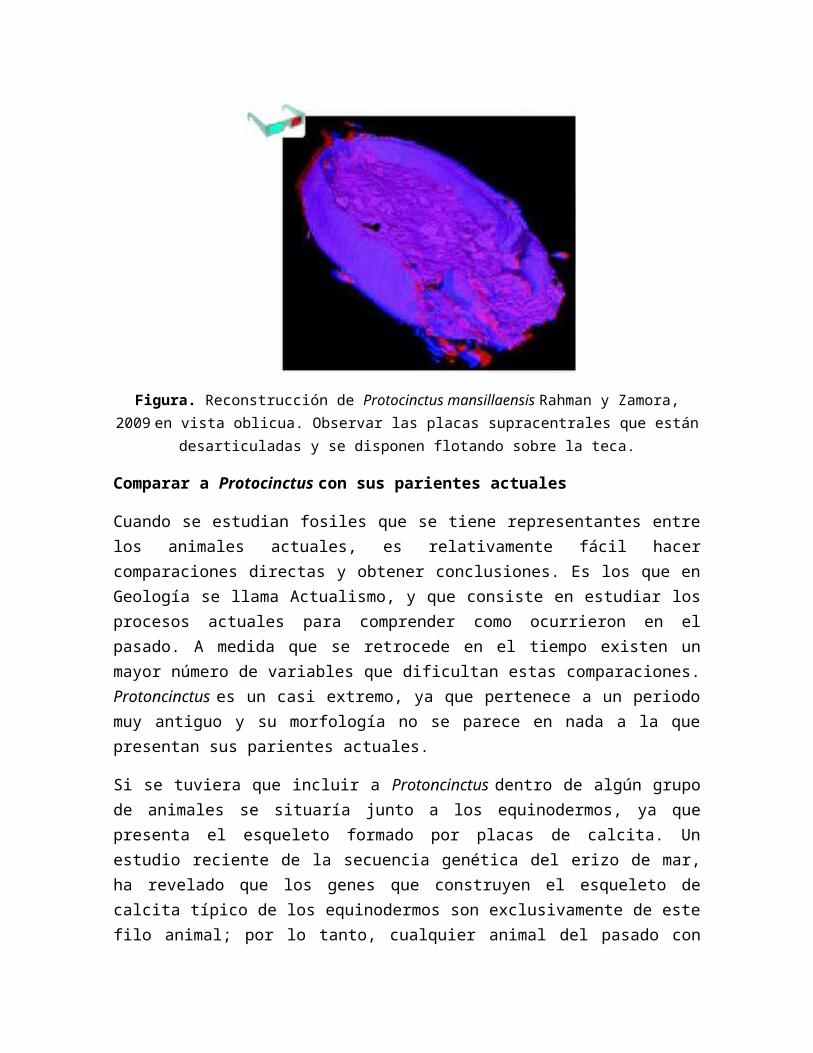
Figura. Protocinctus revela su anatomía. Vista frontal, lateral y ventral.

Bibliografía
Zamora, S. & Rahman, I. A. 2009. Fósiles virtuales cámbricos del Parque Natural del Moncayo. Naturaleza Aragonesa, 23: 12-20.
Zamora, S. 2010. Equinodermos del Cámbrico: Planes corporales, Paleoecología y registro fósil en España.
Revista de Investigación y Ciencia, Boletín N° 389. Febrero, 2009. Fósiles virtuales en cenizas del Silúrico
http://www.academia.edu/2544055/Zamora_S._and_Rahman_I.A._2009._Fosiles_virtuales_cambricos_del_Parque_Natural_del_Moncayo._Naturaleza_Aragonesa_23_12-20
Anexos
Una nueva técnica, la Tomografía Computarizada de Rayos X en 3D, que permite el Sincrotón europeo, está revolucionando el mundo de la Paleontología, ya que permite escanear los fósiles con un detalle que hasta ahora resultaba impensable, sobre todo cuando están incrustados en rocas.
La utilización de esta avanzada tecnología, que ya tiene una amplia utilización en la medicina y la biología, ha permitido dar a conocer algunos detalles asombrosos de los hallazgos recientes más espectaculares, como ha sido el caso del primate bautizado como “Ida”, de hace 37 millones de años.
En un artículo en “New Scientist”, los autores hacen un repaso de las muchas posibilidades que presenta esta técnica, que permite ver los objetos en 3D, a la vez que se les gira para examinarlos desde cualquier ángulo o se les pone del revés para ver que ocultan dentro. En definitiva, una revolución de la forma de ver los fósiles.
Las primeras experiencias en el ámbito de la Paleontología se presentaron en congresos especializados hace unos 3 años desde entonces, como recuerda el investigador Paul Barret, del Museo de Historia Natural de Londres, han aumentado mucho el tipo de muestras que pueden ser analizadas.
Entre los ejemplos más claros está el de los fósiles que son tan delicados que resulta imposible separarlos de las rocas o materiales en lo que están incrustados. Volviendo a “Ida”, el famoso primate, la tomografía computarizada permitió estudiar sus dientes sin necesidad de destruir el cráneo en el que estaba.

Alta precisión
En otras ocasiones, permite saber si hay fósiles. Utilizando el Sincrotón europeo, instalado en Grenoble, el paleontólogo Paul Tafforeau descubrió 356 fósiles ocultos en ámbar en tan solo cuatro días. Entre ellos, un centenar de caracoles, cucarachas, avispas y otros insectos con un millón de años de antigüedad, que pudo ver en tres dimensiones.
“Por primera vez podemos observar lo que hay dentro de los ejemplares sin necesidad de romperlos”, explica Jorn Hurum, descubridor de “Ida”. De hecho, entre los más espectaculares estudios está el de un embrión de gusano de hace 1.000 millones de años o los lóbulos ópticos en el cerebro de un pescado fosilizado o la distancia de las vértebras en un dinosaurio de cuatro toneladas.
Tal es su precisión que la TC que permite comparar el sistema auditivo de los pájaros actuales con el de criaturas prehistóricas como el “Archaeopteryx”, de donde concluyen que su oído interno era parecido al del emú actual. También permite el análisis de los diente, en los que están escritos los acontecimientos más importantes de la vida del individuo (como creció que comía, que enfermedades tuvo, etc.).
El problema es que hay pocos aparatos con esta tecnología en el mundo y, además, es muy cara. Los investigadores españoles recurren en la actualidad al ya mencionado Sincrotón europeo, en el que España tiene una participación del 4% en su presupuesto.
Además, continúa construyendo el Sincrotón Alba, en Cerdanyola (Barcelona), que podría estar en marcha el año próximo. De hecho, ya existe una asociación de posibles usuarios de esta tecnología.
Publicado: 9 de Junio del 2009 en el Blog “Retazos de la Historia” por el director Jesús Manuel Castillo Ramos.