fecundación y desarrollo temprano de animales · - descripción de los estadíos de desarrollo del...
TRANSCRIPT

Enero 2016
Fecundación y desarrollo temprano de animales
Taller del Proyecto de Laboratorios Portátiles para la enseñanza de la Biología
en la Educación Media
11 al 15 de enero de 2016
Facultad de Ciencias, Universidad de Chile
www.uchile.cl www.genomacrg.cl

Enero 2016
Geraldine Aedo Consuelo Anguita Myra Chávez Sandra Edwards Rodrigo Gómez Javiera de La Paz Daniela Gutiérrez Victor Guzmán
Fernanda Lourido Claudia Molina Carlos Muñoz Cristina Muñoz Salomé Muñoz Rodrigo A. Morales Luisa Pereiro Diego A. Rojas
Prof. Juan Fernández Facultad de Ciencias, Universidad de Chile
Prof. Alvaro Glavic Centro de Regulación del Genoma Facultad de Ciencias, Universidad de Chile
Prof Miguel L Allende Centro de Regulación del Genoma Facultad de Ciencias, Universidad de Chile
www.uchile.cl www.genomacrg.cl
Instructores
Laboratorios

Enero 2016
Introducción.
La biología del desarrollo es una disciplina que surge de la fusión de numerosas áreas de la biología moderna, incluyendo la embriología, la genética, la biología celular, la evolución y la genómica. Numerosos descubrimientos de vital importancia científica y de relevantes aplicaciones biomédicas han surgido de la biología del desarrollo. Se hace necesario que la sociedad conozca, en lo posible, algunos conceptos elementales para entender las consecuencias de estos avances y para tener opinión respecto a políticas públicas que se basen en este conocimiento. Las células madre, la clonación, la edición de genomas, los organismos transgénicos son todos desarrollos tecnológicos que involucran aspectos de la biología del desarrollo y que aparecen cotidianamente.
El objetivo de este taller es brindar un conocimiento básico del desarrollo embrionario y post embrionario de las especies animales. Por razones de tiempo y espacio, hemos dividido el curso en sesiones teóricas, para proporcionar el conocimiento necesario para la comprensión de los fenómenos, y experiencias prácticas, para visualizar in situ muchos de los conceptos y descripciones hechas de manera teórica. La biología del desarrollo es una disciplina eminentemente visual; es necesario experimentar y observar directamente los fenómenos para comprenderlos en su justa dimensión. Además, los embriones tienden a ser estéticamente atractivos, siendo muchas veces hermosas estructuras vivientes que impresionan y motivan. Es decir, se prestan especialmente bien a la difusión de la ciencia entre los niños y jóvenes ya que se maravillan fácilmente al tener un organismo en desarrollo frente a sus ojos. La gran pregunta que nos hacemos los que cultivamos esta disciplina es: “cómo se forma un organismo adulto a partir de una sola célula?” Si bien, en las últimas décadas hemos avanzado enormemente en el campo, debemos reconocer que no estamos cerca aún de responder satisfactoriamente esta pregunta. Este misterio también genera gran curiosidad y por ello, muchos jóvenes se sienten atraidos por el estudio del desarrollo embrionario.
El taller se centrará principalmente en dos modelos animales, la mosca del vinagre Drosophila melanogaster y el pez cebra, Danio rerio. Estos modelos se han usado por décadas para estudiar el desarrollo gracias a la facilidad con que se pueden mantener en el laboratorio y a las poderosas herramientas genéticas y celulares disponibles con ellos.
Recomendamos, si se quiere profundizar en el tema, el libro Developmental Biology de W. Gilbert, que tiene algunas ediciones traducidas al español.
www.uchile.cl www.genomacrg.cl

Enero 2016
Conceptos fundamentales del taller.
Esta guía describe las actividades a realizar en el taller de “Fecundación y desarrollo embrionario en animales” para profesores de enseñanza media. El taller tiene por objeto familiarizar a los profesores con el material y las actividades que se podrán llevar a cabo con el laboratorio portátil del mismo nombre. Se procura proveer de un marco teórico para las actividades que exponga los conceptos que interesa transmitir, resumidos a continuación:
1. La meiosis es clave para mantener la dotación cromosómica constante (reduciendo a la mitad la dotación cromosómica en cada gameto) y permite además la variabilidad genética de las especies. En la fecundación, el espermio contribuye con su material genético nuclear y otros componentes limitados. El oocito en cambio, aporta nutrientes, proteínas, ribosomas, mitocondrias, y el grueso de los elementos que participan en la construcción del embrión temprano.
2. La fecundación inicia el desarrollo embrionario. Se gatillan procesos gobernados por componentes maternos, primero, y luego cigóticos. El clivaje aumenta el número de células y prepara el embrión para la morfogénesis y la diferenciación celular.
3. Cada especie tiene un programa determinado genéticamente, que define la secuencia de eventos que formarán el embrión, incluyendo los clivajes y plan corporal que dará origen a los diferentes tejidos. Mutaciones en los genes involucrados en el desarrollo, generan defectos o malformaciones que nos revelan cuál es la función de esos genes.
4. A medida que la embriogénesis progresa, el número de células totipotenciales (células madre) va disminuyendo y la mayoría se diferencia en células de tejidos específicos.
5. El desarrollo se organiza gracias a un programa genético intrínseco pero puede modificarse externamente por factores ambientales. Existen etapas del desarrollo embrionario muy sensibles a estas perturbaciones (físicas o químicas) y que pueden ser muy perjudiciales para la sobrevivencia del individuo.
www.uchile.cl www.genomacrg.cl

Enero 2016
Consideraciones bioéticas.
En este trabajo práctico o laboratorio portátil, se trabajará con seres vivos. Los animales de experimentación o de enseñanza deben ser tratados con el máximo respeto y cuidado. En los laboratorios científicos, los animales están protegidos por rigurosas normativas bioéticas que garantizan su bienestar y el uso restringido y limitado de ellos. Lo mismo debemos intentar cuando llevamos estas experiencias fuera del laboratorio. Las normas para el uso de animales de experimentación respetadas internacionalmente sugieren regirse por el concepto de las tres “R”: replacement, reduction, refinement (reemplazo, reducción, refinamiento). La idea es usar el menor número de animales posible, cuando sea posible, evitarlo, y minimizar o impedir su sufrimiento. En la legislación de muchos países se hace distinción entre animales vertebrados (como el pez cebra) e invertebrados (como Drosophila). También se hace una diferencia según el estado de desarrollo ya que los embriones muchas veces no son considerados “animales” en el sentido de un ente con algún grado de consciencia (sentience). En este laboratorio portátil procuraremos evitar la muerte o el daño innecesario a embriones y adultos de ambas especies en los estadíos donde ya exista un sistema nervioso funcional. Es importante seguir las instrucciones de los profesores y ayudantes en el manejo de los animales y observar de cerca cómo se realizan las manipulaciones. Cuando sea necesario, se anestesiarán los animales para evitarles sufrimiento. La eutanasia, será realizada sólo por personas entrenadas en ello. No se debe desechar ningún animal vivo o muerto en basureros o desagues; usaremos lugares especialmente designados y dedicados a ello. Por último, el trabajo de laboratorio con animales vivos debe realizarse de manera calmada y silenciosa para no perturbarlos.
www.uchile.cl www.genomacrg.cl
Clairvoyance (autorretrato). Rene Magritte, 1936.

Enero 2016
PROGRAMA DEL TALLER DE FECUNDACION Y DESARROLLO EMBRIONARIO
UNIDAD 1: GAMETOGÉNESIS Y FECUNDACIÓN
Objetivo Específico 1:
Analizar el origen y características de los gametos
Contenidos:
Teórico: - Diferenciación celular durante la gametogénesis - Proceso de reducción y recombinación del material genético (meiosis) - Transcripción, organelogénesis y vitelogénesis
Objetivo Específico 2:
Analizar los tipos de reproducción
Contenidos:
Teórico: - Reproducción interna vs externa - Variabilidad genética
Objetivo Específico 3:
Conocer los eventos moleculares y celulares que ocurren durante la fecundación
Contenidos:
www.uchile.cl www.genomacrg.cl

Enero 2016
Teórico: - Requerimientos del proceso de Fecundación - Activación del huevo - Reconstitución de la dosis genética
UNIDAD 2: DESARROLLO TEMPRANO DEL PEZ CEBRA Danio rerio
Objetivo Específico 1:
Conocer el proceso de ovogénesis
Contenidos:
Teórico: - Estructura del ovario - Fases de la ovogénesis - Características morfológicas y particularidades - Síntesis y distribución de los determinantes maternos - Estructura del ovocito al momento de la fecundación
Práctico: - Disección del ovario - Observación de la diferentes etapas de la ovogénesis
Objetivo Específico 2:
Distinguir los cambios que ocurren en la activación y fecundación del huevo
Contenidos:
Teórico: - Penetración del espermio y prevención de la poliespermia
www.uchile.cl www.genomacrg.cl

Enero 2016
- Reacción cortical y formación de la envoltura vitelina (corión) - Movimientos de los organelos internos del huevo fecundado - Flujo citoplásmico y formación del disco embrionario
Práctico: - Observación del desarrollo de huevos activados y fecundados y no
fecundados - Observar los cambios de forma que acompañan el proceso de
fecundación: formación del blastodisco - Observar la formación de canales de flujo citoplasmático rápido
(”streamers”).
Objetivo Específico 3:
Analizar las características del proceso de clivaje y la blastulación, Reconocer el momento de transición de la blástula intermedia.
Contenidos:
Teórico: - Analizar los procesos que llevan a la primera división mitótica en el
embrión - Tipo de clivaje y sus características durante los ciclos celulares 1-5 y 6 en
adelante - Características y clasificación de la blástula - Modificaciones del desarrollo durante la transición de la blástula
intermedia (TBM) y sus consecuencias - Fin de la blastulación e inicio de la gastrulación. Desplazamiento
epibólico del blastodermo
Práctico: - zigoto de D.rerio con énfasis en el crecimiento del disco embrionario y
su clivaje regular durante lasa primeras divisiones celulares - Distinguir tipos celulares los diferentes tipos de blástula hasta el 50% de
epibolía
www.uchile.cl www.genomacrg.cl

Enero 2016
- ¿Cómo establecer la TMB? - Observación de mutantes de efecto materno vs cigótico
UNIDAD 3: DESARROLLO DEL INSECTO Drosophila melanogaster
Objetivo Específico 1:
Conocer el ciclo de vida y características principales
Contenidos:
Teórico: - Ciclo de vida - Embriogénesis. Gastrulación y establecimiento del plan corporal
larvario. Proceso de segmentación. - Organogénesis. Regulación de la expresión génica y la selección de la
identidad celular - Desarrollo larvario y metamorfosis. Control hormonal del desarrollo
Práctico: - Observación de cultivos de Drosophila melanogaster - Reconocer las diferentes etapas del desarrollo y sus principales
características - Observación de las estructuras larvarias internas
Objetivo Específico 2:
Reconocer cómo el genoma es determinante del desarrollo
Contenidos:
Teórico:
www.uchile.cl www.genomacrg.cl

Enero 2016
- Regulación génica, jerarquías de regulación, patrones de expresión y diferenciación en el tiempo y espacio.
- Genes de efecto materno - Genes controladores y selección de tipos celulares - Mutantes y transgénicos
Práctico: - Observación las etapas tempranas del desarrollo - Observar los cambios de forma que acompañan el desarrollo - Observación de mutantes y transgénicos.
Objetivo Específico 3:
Conocer los mecanismo de control hormonal del desarrollo
Contenidos:
Teórico: - Cromosomas politénicos y las diferencias con cromosomas mitóticos e
interfásicos - Señales hormonales e integración con el medio - Metamorfosis y transiciones del desarrollo - Reestructuración del plan corporal - Gametogénesis y terminación el ciclo generacional.
Práctico: - Observación las glándulas salivales - Realizar tinción de cromosomas politénicos - Observar estadios pupales - Disección de línea germinal.
www.uchile.cl www.genomacrg.cl

Enero 2016
UNIDAD 4: DESARROLLO TARDIO DEL PEZ CEBRA Danio rerio
Objetivo Específico 1:
- Reconocer los estadíos de la embriogénesis tardía, desde la gástrula hasta la etapa de somitogénesis y organogénesis
Contenidos:
Teórico: - Descripción de los estadíos de desarrollo del embrión de pez cebra - Principales órganos presentes en los embriones tardíos y larvas
Práctico: - Observación de embriones de pez cebra. Desde gástrula a larva
eclosionada. Estructuras y órganos.
Objetivo Específico 2:
- El pez cebra como modelo para visualizar la expresión génica in vivo
Contenidos:
Teórico: - Regulación génica, jerarquías de regulación, patrones de expresión y
diferenciación en el tiempo y espacio. - Mecanismos celulares y moleculares del desarrollo tardío - La transgénesis y el uso de las proteínas fluorescentes
Práctico: - Observación de embriones tardíos y larvas que expresan GFP en
distintos tejidos u órganos
www.uchile.cl www.genomacrg.cl

Enero 2016
UNIDAD 5: EFECTOS DEL AMBIENTE SOBRE EL DESARROLLO ANIMAL
Objetivo Específico 1:
Los efectos de compuestos presentes en el ambiente externo al organismo como perturbadores del desarrollo embrionario.
Contenidos:
Teórico: - Características de los compuestos químicos - Distinguir la diferencia entre teratógenos y mutágenos - Efectos somáticos vs en la línea germinal - Dominancia y recesividad - Etapas y procesos del desarrollo sensibles a agentes externos - Efectos de la nutrición sobre el desarrollo animal - Uso de modelos animales como biosensores
Práctico: - Ensayar efectos deletéreos de compuestos químicos - Ensayar efectos de la nutrición sobre el desarrollo - Utilizar transgénicos como biosensores ambientales
www.uchile.cl www.genomacrg.cl

Calendario del TallerHora Lunes Martes Miércoles Jueves Viernes
9:00 Unidad 1. Gametogénesis, meiosis (SB)
TP Unidad 2. Fecundación y desarrollo temprano en el pez cebra (JF)
TP Unidad 3. Desarrollo de Drosophila (AG)
Unidad 5. Efectos del ambiente sobre el desarrollo (AG, MA)
TP Unidad 5. Observación de efectos ambientales y de otros experimentos (AG, MA)
10:00
11:00 Unidad 1. Ovogénesis y fecundación (JF)
TP Unidad 4. Desarrollo tardío del pez cebra (MA)
12:00
13:00 Almuerzo Almuerzo Almuerzo Almuerzo Almuerzo
14:00
15:00 Unidad 1 y 2. Segregación ovoplásmica, clivaje, blastulación (JF)
Unidad 3. Desarrollo de Drosophila (AG)
Unidad 4. Desarrollo tardío en pez cebra (MA)
TP Unidad 5. Efectos ambientales en el desarrollo de pez cebra y Drosophila (AG, MA)
Discusión final, encuesta y evaluación
16:00
17:00 TP Unidad 1. Disección de ovario
TP Unidad 4. Gastrulación en pez cebra. (MA)
Convivencia
18:00
19:00
Clase teórica (Auditorio Niemeyer)
Trabajo práctico (TP)(laboratorios de docencia)
SB: Soledad Berrios AG: Alvaro Glavic
JF: Juan Fernández MA: Miguel Allende
�1

TRABAJOPRACTICOUNIDADES1Y2

DESARROLLOTEMPRANODELPEZCEBRAElPezcebracomomodeloparaelestudiodeldesarrollo.Elembrióndelpezcebracombinaunacantidadimportantederasgosbiológicosque lohan convertidoenunmodelomuyadecuadoparaelestudiodelosmecanismosqueregulaneldesarrolloatravés de enfoques estructurales, fisiológicos, genéticos y moleculares. GeorgeStreisinger, de la Universidad de Oregon, fue el primer investigador en utilizar elembrión del pez cebra como modelo para el estudio del desarrollo utilizando unenfoquegenético.Suprimertrabajosobrelamateria(1981)fuepioneroenlograrelestablecimiento de líneas homozigotas y la descripción del primer mutante. Elembrióndelpezcebraes transparentey sudesarrolloembrionariomuyrápido. EsfácildemanteneryreproducirylosadultospuedenserexitosamentemutagenizadosconN-etil-N-nitrosourea.Lafecundaciónesexternaycadaposturaincluyeembrionescuyodesarrollo es razonablemente sincrónico.Cadahembrapuedeponeralrededorde 100 huevos relativamente grandes (0,7 µm). Estos y los embriones pueden serobjeto de variadas manipulaciones experimentales tales como microinyección ytransplantes. A las 24 horas post fecundación (pf) ya es posible distinguir losprincipalesrasgoscorporalescaracterísticosdelosvertebradoscomoeltuboneural,losojosysomitos.Lalarvaempiezaaalimentarsealrededordelos6-7díaspfyalos3mesespfelpezhaalcanzadolamadurezsexualycomienzasureproducción.Mantención de adultos. Los investigadores que utilizan el pez cebra cuentan conbioteriosdondesepuedenmantenerdecenas,cientosomilesdeestanquesconaguapurificadayaireadaencirculaciónpermanente,Cadaacuariodebeconteneralrededorde2pecesporlitrodeagua.Paramanteneraguaenlacondicionesseñaladassedebecontarconseriesde filtrospararemoversedimentos,sales, bacterias,yenespecialhipoclorito, que es muy nocivo para los peces. Además hay que remover del aguacirculante fecas, amoníaco, restosde alimentoymicroorganismosnocivosmediantelimpiezadiariayexposicióndelaguaaradiaciónultravioleta.ElpHdebemantenersealrededor de 7 y la conductividad en aproximadamente 500 µS. Los peces quemueren por la edad u otras razones deben removerse en forma inmediata. El aguadebemantenerseaunatemperaturadealrededorde28°C(rango24-32°C.SchironeandGross,1968).Porúltimo,sedebemantenerunritmodiariode14hdeluz/10hde oscuridad. Los animales son alimentados con una doble dieta: por la mañanaalimentoseco,comolashojuelasdelpopularTetraAmin,yporlatardealimentovivo,como es el caso de quistes del crustaceo Artemia salina cultivados por 24 h ensolución salinaparaArtemia.Esteúltimoalimentovivomejora la efectividadde lasposturas.Hayqueevitarquelospecesengordenendemasíaporqueestoperjudicasureproducción.Elpezcebraesunamascotaquepuedetambiénmantenerseenformaartesanalconlossistemassuministradosporelcomercioespecializadodeacuariosyaccesorios.Losmismoscuidadosaplicadosenlosbioteriosdebenimplementarseenlossistemasartesanales,queusualmentenotienenaguaencirculación.Portanto,eseste caso es recomendable cuidar de la densidad de peces por volumen de agua yreemplazaréstaalmenossemanalmente.Lospecespuedensobrevivirhasta10días

sin alimentación y cambio de agua. Por supuesto que bajo estas condiciones sureproducciónesfuertementeafectada.Utilizartanquesde20-50litrosycuandoelpHestábajosubirloconbicarbonatodesodio.Paralailuminaciónsepuedeutilizarunalámpara de bajowataje y un timer casero y paramantener la temperaturamás omenosconstanteuncalentadorcontermostato.Sielacuarioestáenunlugarqueseusatambiénparaotrospropósitos,estepuedecubrirseconunacajadecartónopañonegro.Estopermitirámantenerelritmoluz/oscuridad.Cruce y postura. El día antes de colectar loshuevos, lospecesmachosyhembrasdebenmantenerseencontenedoresseparados.Losmachossereconocenporsermásdelgadosyamarillentos,mientraslashembrasgrávidastienenelvientreabultadoysonplateadas.Luegodequesehayaencendidolaluzlamañanasiguiente,colocardoshembras y unmacho en una cámara de apareamiento diseñada para evitar que losprogenitoressecomanloshuevos.Simultáneamentesepuedenprepararotroscrucespara obtener más huevos. La cámara de apareamiento (cámara nupcial) se puedeprepararcolocandounacapadebolitasdevidrioenelfondodeunacajadeplásticotransparente que contiene los peces o sobreponiendo dos cajas de plásticotransparentededistinto tamaño, lamáspequeñade las cuales contiene los peces ytiene como base unamallametálica con orificios de aproximadamente 2mm. En elprimer caso los huevos fecundados se extraen de entre las bolitas con un sifón,mientras que en el segundo caso los huevos atravesarán la malla metálica y serecogeránseparandolosdoscompartimientosdelacajadeapareamiento.Loshuevos,colectados con un colador de te, se lavan con una pizeta que contiene aguaacondicionadafiltradadelsistema(aguadelosacuarios)ysetransfierenaplacasdePetri de vidrio o plástico conteniendo agua acondicionada filtrada con miliporo omedio E3. Se puede realizar fecundación in vitro colectando espermatozoides yovocitosdeejemplaresanestesiados.Paraestosemasajeasuavementeconundedoelvientre de machos o hembras en la dirección anterior-posterior, con el pez “bocaabajo”. Colectar separadamente los gametos y después reunirlos en aguaacondicionada o medio E3. La fecundación in vitro asegura el desarrollo mássincrónicodelosembriones.Anestesia.Paraobtenergametos,extraerlasgonadasoejecutaroperacionescomolaamputacióndelaaletacaudal(queregenera),losadultossonanestesiadosporunosminutos en Tricaína (etil 3 -aminobenzoatometanosulfonato), hasta que cesen susmovimientos.Preparación y observación del material embrionario. Se requiere de unmicroscopiodedisecciónolupaquepermitaaumentosde hastaalmenos40X,conunaplatinaquecombineelcolorblancodeunadesussuperficiesconelnegrodelaotra. Las observaciones pueden ser in vivo o postfijación. Para esto último losembriones se fijan en Formaldehído/ácido acético por un tiempo corto. Elmaterialfijado puede lavarse con agua destilada o PBS 1X , descorionarse y teñirse con uncolorantebásico.Losembrionessedescorionanconpinzasderelojerooenpronasa(0.5 mg/ml de agua acondicionada). Para orientar la posición de los embrionesdescorionadosestossepuedenmontarenunmedioviscosocomolametilcelulosa.

.Ovogénesis.Eselprocesomedianteelcualsegeneran célulascompetentesparaserfecundadas.EnelpezcebracorrespondenaovocitosVquecompletanlameiosiscomoresultadodelapenetracióndelespermatozoide.Enesemomentosegeneraelzigoto.Durante la ovogénesis ocurren tres procesos fundamentales: (1) reduccióncromosómica,(2)recombinacióngenéticaentrecromosomashomólogosy(3)intensatranscripción. A estos procesos se agrega el ensamblaje de glóbulos de vitelo ygránulos corticales, la proliferación de organelos y la elaboración del corion y lamicropila,estructuraalolargodelacualpenetranlosespermatozoides.Laactividadtranscriptiva es de lamayor importancia para el desarrollo porque los transcriptosmaternosacumuladosenelovocito regulannosoloeldesarrollodelpropioovocitosino también la activación del huevo y la embriogénesis temprana (Pelegri, 2003;Lindeman and Pelegri, 2010; Dosch, 2014). La hembra tiene un par de ovariossuspendidosdelacavidaddelcuerpoporunvascularizadomesovarioysecomunicanalexteriorporcortosgonoductos.Elovariodeanimalesmadurosocupaunvolumenimportantede la cavidadpleuroperitoneal, 10-20% delpesode lahembra cuandoestágrávida.Selmanetal.(1993)reconocen5estadioseneldesarrollodelovocito.(a)Ovocitos del estado I (estadodecrecimientoprimario)son transparentes,tienenunvoluminosonúcleocentral(vesículagerminal)ymidende7-140micronesde diámetro. Los ovocitos más pequeños o prefoliculares (estado IA) están en elleptotenedelaprimeradivisiónmeióticamientraslosmásvoluminosos,ofoliculares(estado 1B), han entrado al diplotene de la primera división meiótica y estánclaramente rodeados por una capa epitelial plana de células foliculares. Algunos onumerososnucleólosestánpresentesenlosovocitos,muchoscercanosalaenvolturanuclear.(b)OvocitosdelestadoII(estadoalveolarcortical)sontransparentesymidende140-340micronesdediámetro.Sunúcleoescentralyestádetenidoenlaprofasede la meiosis I. El citoplasma de aspecto espumoso se debe al comienzo de lasformacióndelosgránuloscorticales.(c) Ovocitos del estado III (estado de vitelogénesis) empiezan a opacarsecomo resultado del depósito de glóbulos de vitelo que desplazan los nacientesgránuloscorticaleshacialaperiferiacelular.Midende340-690micronesysunúcleoempiezaamoversehacialaperiferiamanteniéndoseenlaprimeradivisiónmeiótica.(d) Ovocitos del estado IV (estado de maduración) son muy opacos comoresultado de la acumulación de gran cantidad de vitelo. La ruptura de la vesículagerminal marca la conclusión de la primera división meiótica y el inicio de lassegunda,ElovocitosedetieneenlametafaseII.

(e)OvocitosdelestadoV(estadodehuevo)sontransparentesymiden0,73-0,75 micrones de diámetro y la meiosis continua detenida en la metafase II. Sonaplanadosyflotanenlacavidaddelovario.ElovocitoVyahainiciadoelprocesodesegregaciónovoplásmicamedianteel cual lagunasdecitoplasmaricasenorganelos,citoesqueleto y determinantes maternos que han empezado a acumularse en elcasquete animal del ovocito para formar el naciente blastodisco o preblastodisco(Fernándezetal.,2006).Estedaráfinalmenteorigenalembrión.Características generales del desarrollo. En verdad el desarrollo se inicia en elovarioycontinúaenunaseriedeprocesosquesehanordenadoenestados.LatablaI,elaborada por Kimmel et al. (1995), resume las características de tales estadosbasadoenrasgosmorfológicosdeembrionesvivosobtenidosdelamismamadrequese han fecundado in vitro, situación que mejora la sincronía de su desarrollo(consultar también el “Zebrafish Book” en la dirección http:/zfish.uoregon.edu o ellibro Zebrafish editado por Nüsslein Volhard andDahm, 2002). La descripción deldesarrollobasadaenestadosesmássignificativaquelabasadaenedad.Lavelocidaddedesarrolloaumentaodisminuyesegúnlatemperatura.Sinembargo,eldesarrollonormal ocurre dentro de un rango de temperatura ya indicado. Con frecuencia sehabladedesarrollotempranoytardío.Elprimeroseextiendehastaelcomienzodelagastrulaciónmientraselsegundohastaquesehayacompletadolaorganogénesisylaespecificacióndelosejescorporales.Enelpezcebrayotrosorganismosseagregaunlargoperíodo larvarioqueculminacon la formacióndeun juvenil cuyamaduraciónsexualsealcanzaalos3mesespf.UnacompletadescripcióndelaanatomíadelalarvaendesarrolloseencuentraenSalgadoyal.(2012).Fecundación y activación. La fecundación puede ocurrir en forma natural oejecutarseinvitro.Comienzacuandounespermatozoidepenetraporlamicropilaysumembrana se fusiona con la del ovocito, fenómeno que ocurre 20-60 segundosdespués que los gametos son descargados o combinados. En contacto con el aguaambosgametossonactivados.Poreso,alobtenerlosgametospormasajeabdominalhay que prevenir que tomen contacto con el agua. Esto se logra removiendo losespermatozoidesconunamicropipeta.Losovocitosquefueronactivadosporelaguanopodránfecundarsedespuésporquehanformadolaenvolturavitelina,ocorion,quepreviene la penetración de espermatozoides. Los espermatozoides permanecenactivos por aproximadamente 1 minuto. La activación del ovocito tiene variasmanifestaciones comocambios en el potencial demembrana, explosivo aumentodecalciolibredentrodelhuevo,terminacióndelameiosisconexpulsióndelpolocitoII,exocitosis de los gránulos corticales y levantamiento de la envoltura vitelina queformala“membranadefecundación”ocorión.CambiosenelpHdelzigoto,aumentodelconsumodeoxígeno,síntesisdeproteínasyDNAymovimientoscitoplasmáticos,también toman lugar (ver Gilbert, 2013). Muchos de estos fenómenos tambiénocurrenenelovocitonofecundadoactivadoencontactoconelagua.Desarrollo del zigoto y movimientos citoplasmáticos (segregaciónovoplásmica).Unavezqueelhuevocompletalameiosis,aproximadamentealos10min pf, pasa a llamarse zigoto y consiste de un citoplasma rico en vitelo, o

viteloplasma, y un citoplasma rico en organelos, citoesqueleto y RNAm de origenmaterno, llamado ovoplasma. El ovoplasma se distribuye en tres dominioscitoplasmáticos:blastodisco,endoplasmayectoplasma.Elendoplasmaformaunaredde numerosas lagunas de bordes irregulares que se conectan con el blastodiscomediante cortos canales (“short axial streamers”), mientras el ectoplasma rodea elzigotoy también se conecta conelblastodisco.Amedidaque transcurre laprimerainterfase el ovoplasma fluye lentamente desde las lagunas de endoplasma y delectoplasmahaciaelpoloanimal,conlocualelblastodiscocrecediscretamenteenlaregiónanimaldelzigoto.Mientrastanto,lospronúcleosseacercanyformanelnucleodelzigoto.Esteinicialamitosisaproximadamentealos30minpf,cuandoseproducelacontraccióndeunanillodeactinaensambladoenel rebordedelblastodisco.Estefenómenoseacompañadeunflujorápidodeovoplasmahaciaelpoloanimal,procesoque semanifiesta por la formación de numerosos y voluminosos canales llamados“streamers”,queporsutrayectoriayrelacionesconelblastodiscopuedenseraxialesomeridionales. El blastodisco crece rápidamente y cuando el anillo de contracciónempieza a relajarse se inicia la formación del primer surco de citoquinesis que daorigenalosdosprimerosblastómeros.Porlotanto,partedelaprimeramitosisocurredurante la etapa de flujo rápido de ovoplasma. La formación de canales axiales ymeridionalesmenosrobustos se repiteen las siguientesdivisionesdeclivaje, juntocon la formación de anillos de contracción más débiles. Ambos procesos songeneralmente indetectables a partir de los 8-16 blastómeros . A estas alturas lamayoríadelaslagunasdeendoplasmahandesaparecidoyensulugarlosglóbulosdeviteloseacercanentresiyformanlaboladevitelo,queenlalarvaconstituiráelsacovitelino.Lasegregacióndecitoplasmasepuedeseguirenembrionesvivosobservadosen la lupa o elmicroscopio, instrumentos que generalmente están conectados conunacámaradevideoyuncomputadorpremunidodeunprogramacapazdeinvertirelcontraste entre ovoplasma y viteloplasma. Esto también se logra observandoembriones,colectadosadistintotiempo,quehansidosometidosaunafijaciónácida(Fernándezetal,2006;FuentesandFernández,2010).Divisiones de clivaje.Las5primerasdivisionesdeclivajesonmeridionales, tienenuna orientación regular y ocurren desde el centro al borde del blastodisco. Cadadivisiónesortogonalalaprecedentedemaneraqueelblastodiscoessucesivamenteparceladoenhilerasdecélulas.Primerounahileradedoscélulas,segundodoshilerasdedoscélulas,tercero2hilerasde4células,cuarto4hilerasde4célulasyquinto4hilerasde8célulasAlfinaldelestadode16célulashay4centralesy12marginales,que permanecen conectadas con la bola de vitelo. La sexta división de clivaje esecuatorialygeneraunacapasuperficialyotraprofunda.Deaquíhastalatransicióndelablástulaintermedialamayoríadelascélulasnomarginalesclivancompletamente,persistiendodivisionesincompletasenlascélulasmarginales.A28,5ºClasdivisionesdeclivajeocurrencada15-20mindeacuerdoalasiguientesecuenciatemporal:2células:45-50minpf4células:1:00-1:05hpf8células:1:15-1:20hpf16células:1:30-1:35hpf

32células:1:45-1:50hpf64células:2:00-2.05hpfBlastulación (2:15-5:15h) y transición de la blástula intermedia.Lassiguientesdivisionescelularessonrelativamentesincrónicas(metasincrónicas)eirregularesenelplanoenqueseensamblanloshusosdeclivajeyportantoenladirecciónenqueocurre la citoquinesis. A las 2:30-2:35 h pf se alcanza el ciclo celular 8 con 256células,alas2:45-2:50helciclocelular9con512célulasy15a20minmástardeelciclocelular10conmilcélulas (“1kgdeçélulas”) .El ciclocelular9correspondealmomento en que ocurre la transición de la blástula intermedia (Kimmel and Law,1985b).Esteesunestadodegrandescambioseneldesarrolloembrionarioporqueladivisiones celulares se hacen asincrónicas (con las células animales dividiéndoseantesdelasmarginales),loscicloscelularessealarganycomienzalatranscripcióndelgenoma zigótico. Es decir el control materno del desarrollo es reemplazado por elcontrol zigótico. Esto no debe entenderse como un cambio brusco de la acción deproductosgénicosmaternosazigóticos.Porelcontrarioyporuntiempo,laaccióndelos dos productos génicos se sobreponen. La progresiva acumulación de célulasdespuésdelciclocelular 11 transformaelblastodermoenuncúmulodecélulasdedistinta forma, situación que permite reconocer distintos tipos de blástulas: alta,oblonga, esfera y cúpula. Son estereoblástulas porque carecen de una cavidad. Lascélulasmarginalessufrenuncolapso,liberanelcitoplasmaylosnúcleossereúnenenelcitoplasmaquerodeaelviteloparaformarlacapasincicialdelvitelo.Aestasalturasno hay puentes citoplasmáticos entre los blastómeros. La blastulación termina y lagastrulación se inicia cuando el desplazamiento o epibolía, en la dirección animalvegetal del blastodermo alcanza el ecuador del embrión (50% de epibolía). Lacronologíadelablastulaciónaunatemperaturade28,5°Ceslasiguiente:128células2:15-2:20hpf256células2:30-2:35hpf512células2:45-2:50hpf1000células3:00-3:05hpfBlástulaalta3:15-3:20hpfBlástulaoblonga:3:40-3:45hpfBlástulaesfera4:00-4:05hpfBlástulacúpula4:15-4:20hpf50%deEpibolía5:15-5:20hpf.

PROTOCOLO1
DESARROLLOTEMPRANODELPEZCEBRA
Para la ejecución de este y otros protocolos del pez cebra se le suministrarátablas del desarrollo, diagramas y fotografías que facilitarán las observacionesprogramadas.
OVOGENESIS
1.-Elijahembrasgrávidas,anestésielasycolóquelassobreunportaobjetos.Hagaunaescisiónventraly remueva losovariosqueaparecenrepletosdeovocitos.TransfieralosovariosaunacápsuladePetriconsoluciónsalinaoaguaacondicionada.Esta última no es una solución fisiológica adecuada peromantiene razonablementebienlaestructuradelosovocitosparasuobservacióninmediatayfijación.2.-Observandoabajoaumentodescubriráquecadaovarioestaformadodeacúmulosoquistesdeovocitoscadaunodeloscualesesposiblementeelclondeunacélulamadre.Cadaquiste tieneovocitosdedistinto tamañoqueestánendiferentesestadosdelaovogénesis.Amayoraumentopodrádistinguirovocitosdelosestados1-4deldesarrollo.3.-Fijeuntrozodeovarioconformaldehído/ácidoacéticoporalmenos30min,laveporelmismotiempoenvarioscambiosdePBS1Xoaguadestiladaytiñaporunosminutos con azul de toluidina (colorante básico) en borato de sodio. Lave denuevoconPBSpararemoverelexcesodecolorante.MonteeltrozodeovarioconPBSentre un porta y cubre objetos, este último con topes de plasticina en sus cuatroángulos.Visualizelosiguiente:(a) OvocitoI.Sucitoplasmaapareceintensamenteteñidodecolorazul(basofiliadebidaalaacumulaciónderibonucleoproteínas).Elnúcleocentraldeestosovocitospresentaunoomásacúmulosdematerialbasófiloenlaperiferiadelnúcleoquecorrespondenanúcleolos.(b) Ovocito II. La basofilia del citoplasma es reemplazada por laaparicióndeuncitoplasmadeaspectoesponjosodebidoalaacumulacióndealveólosprecursores de los gránulos corticales. La silueta del núcleo de posición central estodavíadetectable

(c) Ovocito III y IV. El citoplasma empieza a oscurecerse debido aldepósitogradualdeglóbulosdevitelo.Ladiferenciaentreunoyotroovocitosepuededeterminarporeltamañodelacélula.(d)OvocitoV.Sepuedeobtenerdelliquidoqueseacumulaenelcentrodelovariodeunahembragrávidaprontodespuésquelaluzsehaencendido.Esunacélulaaplanadayfrágilquesehinchaenpresenciadeaguayseactiva.Sepuedefijaren formaldehído/ácido acético y así visualizar la distribución temprana delovoplasma.
FECUNDACION
Tan pronto detecte que el apareamiento de machos y hembras fue exitoso,remuevay lave loshuevospara luego transferirlos aunacápsuladePetriconaguaacondicionada. Observe la formación del corion y de la cámara perivitelina. Consuertepodrávisualizar lamicropila,queune lasuperficiedelhuevoconelcorión,yuna pequeña célula adherida al zigoto o flotando en el líquido perivitelino y quecorrespondealpolocito2.Tantohuevosfecundadoscomonofecundados,encontactoconelagua, formanelcorionyhabráqueesperar ladivisióndesublastodiscoparadeterminar si fue o no fecundado La estructura de los gametos no activados podráestudiarse enmontajes de ovocitos V extraídos directamente del abdomen de unahembra grávida y de espermatozoides extraídos directamente del abdomen demachos. La observación deberá hacerse a aumentos altos de la lupa omejor en unmicroscopio.
SEGREGACIONOVOPLASMICAYPRIMERCICLOCELULARDesdeelmomentode la fecundaciónhasta laterminacióndelprimerciclocelularUd. podrá combinar la observacionde zigotos vivos conaquellos fijados cada5-10minmedianteformaldehido/ácidoacético.Laposicióndelzigotovivopuedeajustarsemontándolos enmetil celulosa y orientando su posición con unmondadientes. Laobservacionesdebendirigirsealavisualizacióndelossiguientesprocesos:(1) Etapa de crecimiento gradual del blastodisco en la región animal delzigoto durante la primera interfase. Asegúrese que la temperatura se mantienealrededordelos28,5°C.(2)Gradualdesaparecimientodelaslagunasdeendoplasma(3)Contraccióndelanillodeactinaenelrebordedelblastodisco(4)Formacióndecanaleso“streamers”axialesymeridionales.Estosúltimossevisualizanmejorenhuevosfijadosymontadosenformaoblicua.(5)Relajacióndelanillodeactina,desaparicióndeloscanales,yaparicióndelprimersurcodedivisióndelblastodisco.Conestosecompletaelprimerciclocelularalos30-40minpf.
CLIVAJE

Este proceso será estudiado tanto en huevos vivos como fijados, estos últimosprocesadoshaciaelfinaldecadaciclocelular.Conunmondadientespodráorientarlaposicióndelosembrionesquemejormuestreelprogresodelprocesodeclivaje.(1) Visualize la dirección en la cual avanza el surco de clivaje de cadadivisióndelblastodisco.(2)Corroborelaregularidaddelasdivisionescelulares1-5ylosplanosenqueocurreestefenómeno.(3)Verifique como lamayorparte del endoplasmaha sido trasladado alblastodisco(4)Identifiqueembrionesde64-1.000células,quenopodrácontar,perosicomparar con la imágenes de la tabla de desarrollo que se le suministrará. En losembrionesmásavanzadospodráidentificarlascélulassincisialesdelviteloporfueradelrebordedelblastodermo
BLASTULACION
Esteprocesoenverdadseiniciamástempranoeneldesarrolloperoesapartirde1kgde células cuando aparecen cambios sistemáticos en la formadel blastodermoquedanorigenalosdistintostiposdeblástula.Reconozca:(1) La blástula alta que presenta un promontorio de blastómeros que seagudizahaciaelpoloanimaldelembrión.(2) La blástula oblonga que presenta un aplanamiento del blastodermosobre laboladevitelo.Lacapasincisialdelviteloaparece formadaporvariascapasconcéntricas.denúcleoscelulares.(3)Lablástulaesferatiene,comodicesunombre,formaesféricayellímiteentre el blastodermo y la bola de vitelo es una horizontal. Los núcleos de la capaenvolventedelvitelosufrenlasúltimasdivisiones.(4)Lablástulacúpulasufreundesplazamientodelaboladevitelohaciaelpolo animal mientras el blastodermo se aplana como resultado del inicio de sudesplazamiento,oepibolía,haciaelpolovegetal.Losnúcleosdelacapaenvolventedelvitelopuedenserfácilmentevisualizados.Laepibolíadelblastodermoa lo largode labolavitelosehaceaexpensasdeunprogresivo adelgazamiento del blastodermo que cuando alcanza el ecuador delembrión(50%deepibolía)dainicioalprocesodegastrulación.
MUTANTESQUEAFECTANELDESARROLLOEMBRIONARIOTEMPRANO
La observación demutantes de efectomaterno brinda la oportunidad de visualizarfenotipos en los que se han alterado procesos tempranos del desarrollo como lagametogénesis, fecundación y activación, segregación ovoplásmica, clivaje yblastulación. Muchos de losmutantesmueren en las primeras horas de desarrollomientrasotrosformangástrulasylarvasanormalesquenocompletaneldesarrollo.Elmutante emulsion/cax1 tiene alterada la estructura de una proteína que funcionacomo un intercambiador de Ca+2/H+. (Fuentes et al., 2014 ). Las alteraciones másconspicuas de este mutante tienen que ver con el retardo de los movimientos

citoplasmáticos, alteracionesen la sincroníaypatrónde lasdivisiones celularesdeclivaje,disminucióndelaadhesividadcelularyventralizacióndelalarva(Doschetal.,2004;Wagneretal.,2004).
MATERIALESREQUERIDOSMedio E3: 15mMNaCl, 0,5mMKCl, 1mMCaCl2, 1mMMgSO4, 0,15mMKH2PO4,0,050mMNa2HPO4,0,70mMNaHCO3(sepuedeomitir)pH7-7.5.Tricaína:Disolver40mgdetricaínaen9,79mldeaguadestiladay0,21mldeTris1MpH7.AlicuotarentubosEppendorfde630µlycongelara-20°C.DescongelarenelmomentodeanestesiarytransferiraunaplacadePetridondesecolocaelpez.Pronasa:0,5mgdePronasaenaguaacondicionada.Incubarporunosminutosa28°CPipetasPasteurdevidriooplásticoCápsulasdePetrideplásticoovidriode50x12o90x15mmdediámetroyaltura.Pizetasde100o250ml.PBS1X:8gdeNaCl,0,2gdeKCL,1,44gdeNa2HPO4,0,24gdeKH2PO4enunlitrodeaguadestiladapH7.4.Mantenerenelrefrigeradora4°Cyentibiarantesdeusar.Metilcelulosa:2-3%enaguaacondicionadafiltradaomedioE3.Alicuotarymantenerenelrefrigerador(siesposiblea-20°C).Entibiarantesdeusar.Sepuedeutilizarparaorientarlaposicióndeembrionesvivosofijados.PortaycubreobjetosPlasticina.Filtrosdemiliporode0,2micrómetros.Aguaartificialdeacuario:60mgdelamezcladesales“InstantOcean”enunlitrodeaguadesmineralizada.SoluciónsalinaparaArtemia:300gdeNaClen1ltdeaguadestilada.MantenerquistesdeArtemiaconaireaciónconstanteenembudodedecantación.SoluciónsalinaRinger:116mMNaCl,2,9mMKCl,1,8mMCaCl2,5mMHepespH7,2.SoluciónsalinaHank:137mMNaCl,5,4mMCaCl2,1,3mMCaCl2,1mMMgSO4,0,44mMKH2PO4,0,25mMNa2HPO4,4,2mMNaHCO3.

METODOSFijación ácida: Preparar formaldehído al 5% fresco a partir de la solución stock al37%,quedebeconsiderarsecomosi fueseal100%.Pipetearen lasolución fijadora10-15 embriones y acto seguido agregar 5 gotas de acético acético glacial. Tapar lacápsulayagitarrotatoriamentehastaquelosembrionessetornanblanquizcos.Fijarpor30-120min..Elformaldehídoyácidoacéticosonmuytóxicosyportantocuandose utilizan para la fijación de material biológico debe contarse con una buenaventilación.Tinción con azul de Toluidina. Preparar una solución al 0,001% de Azul deToluidinaenPBS1X.Partirpreparandounasoluciónstockdeazuldetoluidinaal1%enboratodesodioal1%.Tomar15µldelasoluciónstockydiluiren10mldePBS1X.Teñirlosovariosoembrionesenunagotadepositadasobreunportaobjetos.DespuésdeunosminutosdetinciónremoverelexcesodelcolorantemediantelavadosconPBS1X. Montar un trozo pequeño del ovario o algunos embriones entre porta y cubreobjetoscontopesdeplasticina.
REFERENCIAS
Dosch,R.2014.Nextgenerationmothers:maternalcontrolofgermlinedevelopmentinzebrafish.CritRevBiochemMolBiol.M.M.Coxeditor.Dosch, R;Wagner, D. S; Mintzer, K. A; Runke, G;Wiemelt, A.P; Mullins, M.M. 2004.Maternal control of vertebrate development before the midblastula transition:Mutantsfromthezebrafish1.DevCell6:771-780.Fernández,J;Valladares,M;Fuentes,R;Ubilla,(2006).Reorganizationofcytoplasminthezebrafishoocyteandeggduringearlystepsofooplasmicsegregation.DevDyn235:656-671.Fuentes,R; Fernández, J. (2010).Ooplasmic segregation in the zebrafish zygote andearly embryo: Pattern of ooplasmic movements and transport pathways. Dev Dyn242:503-517.Fuentes, R; Escobar, M; Kugath, A; Montecinos-Franjola, F; López, P; Fernández, J;Mullins, M (2014) Cation/proton exchanger 1 protein (Cax1), a maternal-effectregulatorofzebrafishcytoplasmicsegregationandmRNAlocalization.CongressoftheAmericanSocietyofCellBiology.Philadelphia.Penn.

Gilbert, S. Developmental Biology (2013). Tenth Edition. Sinauer Associates inc.Publisher. SunderlandMassachusetts.USA.Este libro es tambiénútil para consultarsobreotrosaspectosdeldesarrollodepeces.Kimmel,C.B;Law,R,D(1985).Celllineageofzebrafishblastomeres.II.Formationoftheyolksyncitiallayer.DevBiol108:86-93.Kimmel.C.B;Ballard,W.W;Kimmel,S.R;Ullmann,B;Schilling,T.F.(1995).Stagesofembryonicdevelopmentofthezebrafish.DevDy.203:253-310.Lindeman, R. E; Pelegri, F. 2010. Vertebrate Matternal-effect genes: Insights intofertilization, early cleavage divisions, and germ cell determinant localization fromstudiesinthezebrafish.MolReprodDev77:299-313.Nüsslein-Volhard,Cr;DahmR.2002.Zebrafish.Practicalapproach.OxfordUniversityPress.Pelegri,F.Maternalfactorsinzebrafishdevelopment.(2003).DevDyn228:535-554.Salgado, D; Marcelle, C; Currie, P. D; Bryson-Richardson. (2012). The zebrafishanatomyportal:Anovelintegratedresourcetofacilitatezebrafishresearch.DevBiol372:1-4.Schirone, R. C; Gross, L. (1968). Effect of temperature on early embryologicaldevelopmentofzebrafishBrachydaniorerio.JExpZool169:43-52.Selman,K;Wallace,R.A;Sarka,A;Xiaoping,Q.(1993).StagesofoocytedevelopmentinthezebrafishBrachydaniorerio.JMorph218:203-224.Streisinger,G;Walker,C;Dower,N;Knauber,D;Singer,F.(1981).Productionofclonesofhomozygousdiploidzebrafish(Brachydaniorerio).Nature291:293-296.TheZebrafishBook.Aguide for the laboratoryuseofzebrafishDanio(Brachydanio)rerio(1993).UniversityofOregonPress.Eugene.Wagner, D. S; Dosch, R;Mintzer, K. A;Wiemelt, A. P;Mullins,M.M. 2004.MaternalcontrolofDevelopmentatthemidblastiulatransitionandbeyond:MutantsfromthezebrafishII.DevCell6:781-790.


Kimmelatal.,1995

Kimmeletal.,1995
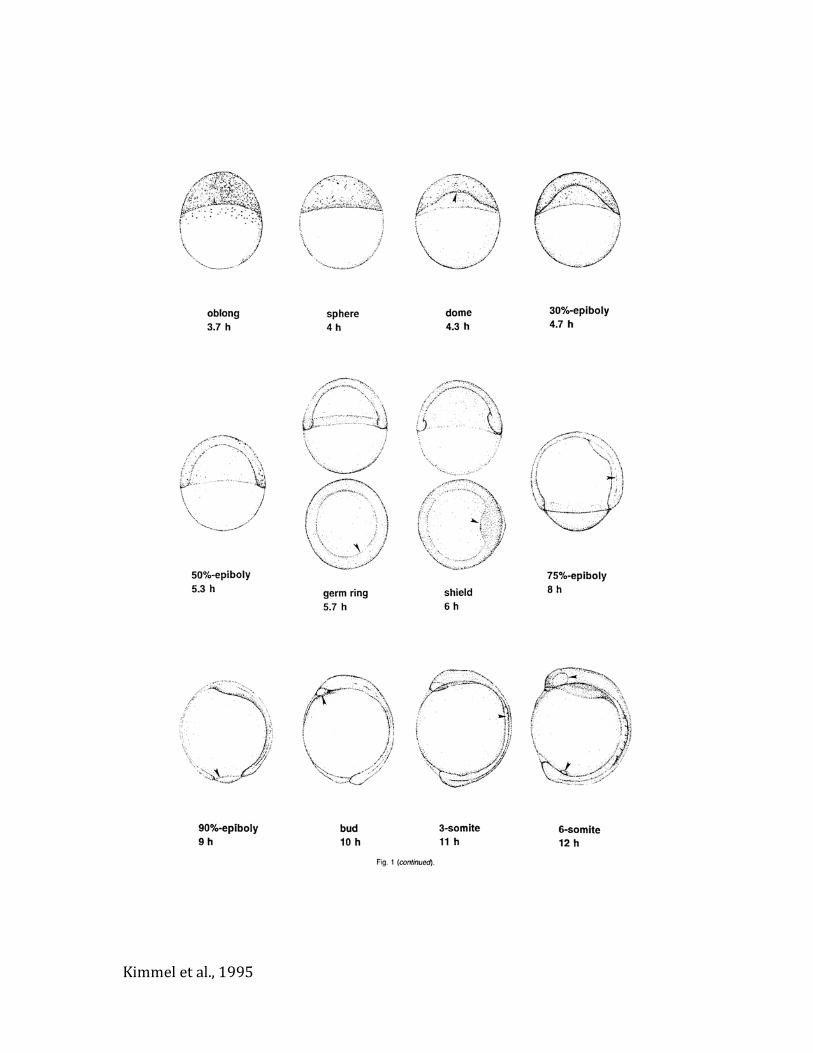
Kimmeletal.,1995

Formaciónde“streamers”enembrionessometidosafijaciónácidaFuentesandFernández,2010


Kimmeletal.,1995
A:2BlastómerosB.4BlastómerosC:8BlastóterosD.16BlastómerosE:32BlastómerosF:64Blastómeros

A. 256BlastómerosB. BlástulaaltaC. Blástulaalta/oblongaD. Blástulaoblonga/esferaE. BlástulacúpulaF. E.30%deepibolía
Kimmeletal.,1995

TRABAJO PRACTICO UNIDAD 3

APOYO TEÓRICO PARA UNIDAD 3:
DESARROLLO DEL INSECTO DROSOPHILA MELANOGASTER Objetivo Específico 1: Conocer el ciclo de vida y característica principales SECCIÓN TEÓRICA INTRODUCCIÓN La especie Drosophila melanogaster ha sido utilizada ampliamente para experimentación en genética puesto que presenta una serie de ventajas para su manipulación y estudio, entre las cuales se encuentra el ciclo de vida corto, la generación de un alto número de descendientes, fácil mantención de un alto número de individuos en un laboratorio por un bajo costo y un número de cromosomas relativamente reducido (4 pares), a lo que se suma la existencia de un gran número de herramientas para su manipulación genética. Fue introducido como animal de experimentación genética por Thomas Morgan a principios del siglo XX gracias a las ventajas mencionadas, junto con características heredables que podían ser reconocidas a simple vista. Su genoma fue completamente secuenciado y publicado en marzo del año 2000, y hoy en día es utilizado como modelo de estudio en un sinnúmero de laboratorios alrededor del mundo para la comprensión de diversos procesos biológicos, como lo es el desarrollo, conducta, patologías, entre otros. Cabe destacar que una vasta gama de dichos procesos son homologables a otras especies, incluyendo los mamíferos. Aún más, aproximadamente el 60% de los genes asociados a enfermedades en humanos que se han descrito a la fecha presentan un homólogo identificable en el genoma de D. melanogaster. Drosophila melanogaster: características principales El insecto Drosophila melanogaster, es una especie de díptero braquícero perteneciente a la familia Drosophilidae. Este animal se conoce también como “mosca del vinagre” o “mosca de la fruta”, dado que se alimenta de frutas en proceso de fermentación tales como manzanas y uvas, entre otras. D. melanogaster es una especie de insecto holometábolo, es decir que lleva cabo un proceso de metamorfosis completa desde el estadío larvario hasta el adulto con una fase intermedia pupal. Las distintas fases del ciclo de vida de estos animales se muestran en la Figura 1, que incluyen huevo o embrión, larva I, II y III, pupa y adultos, que presentan dimorfismo sexual. El ciclo de vida de estos animales demora alrededor de 10 días a 25°C, pudiendo ser modulado en función de la temperatura, siendo más lento a menores temperaturas y más rápido a mayores temperaturas, por ejemplo, el ciclo demora aproximadamente 19 días a 18°C y 7 a 29°C. A su vez, existen determinadas condiciones genéticas que podrían variar la temporalidad de desarrollo de estos animales.
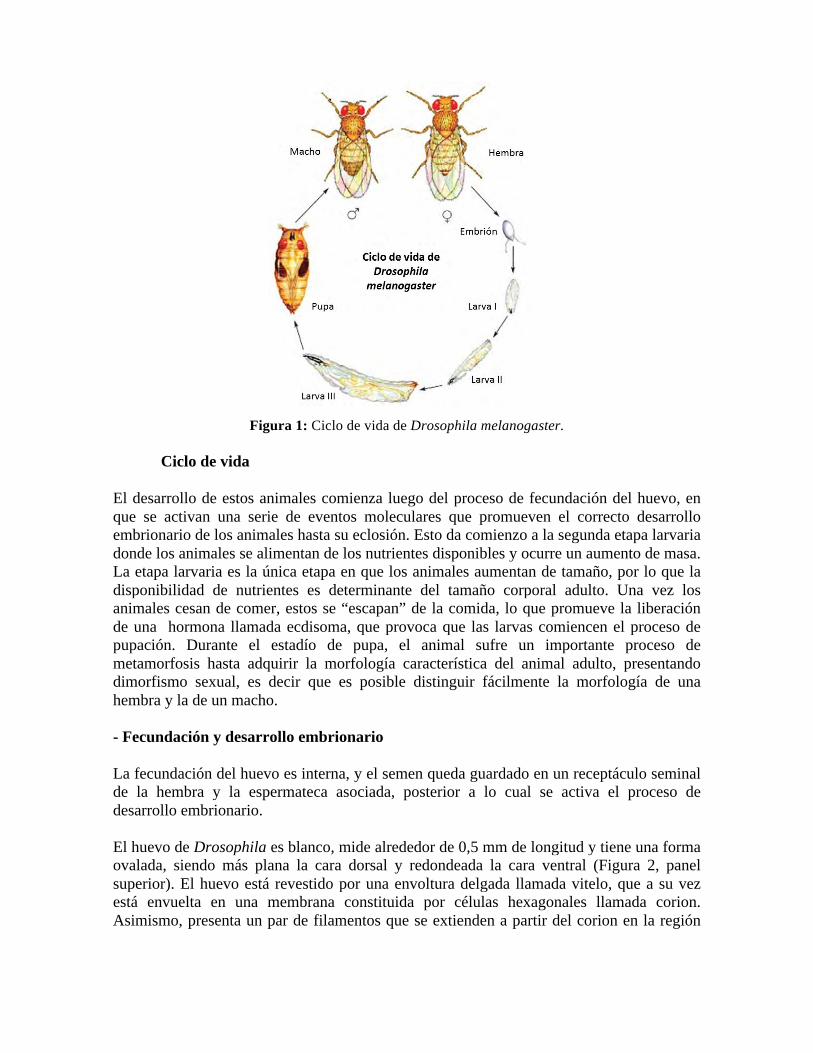
Figura 1: Ciclo de vida de Drosophila melanogaster.
Ciclo de vida
El desarrollo de estos animales comienza luego del proceso de fecundación del huevo, en que se activan una serie de eventos moleculares que promueven el correcto desarrollo embrionario de los animales hasta su eclosión. Esto da comienzo a la segunda etapa larvaria donde los animales se alimentan de los nutrientes disponibles y ocurre un aumento de masa. La etapa larvaria es la única etapa en que los animales aumentan de tamaño, por lo que la disponibilidad de nutrientes es determinante del tamaño corporal adulto. Una vez los animales cesan de comer, estos se “escapan” de la comida, lo que promueve la liberación de una hormona llamada ecdisoma, que provoca que las larvas comiencen el proceso de pupación. Durante el estadío de pupa, el animal sufre un importante proceso de metamorfosis hasta adquirir la morfología característica del animal adulto, presentando dimorfismo sexual, es decir que es posible distinguir fácilmente la morfología de una hembra y la de un macho. - Fecundación y desarrollo embrionario
La fecundación del huevo es interna, y el semen queda guardado en un receptáculo seminal de la hembra y la espermateca asociada, posterior a lo cual se activa el proceso de desarrollo embrionario. El huevo de Drosophila es blanco, mide alrededor de 0,5 mm de longitud y tiene una forma ovalada, siendo más plana la cara dorsal y redondeada la cara ventral (Figura 2, panel superior). El huevo está revestido por una envoltura delgada llamada vitelo, que a su vez está envuelta en una membrana constituida por células hexagonales llamada corion. Asimismo, presenta un par de filamentos que se extienden a partir del corion en la región

anterior de la cara dorsal de los embriones (Figura 2, panel inferior), que corresponden a filamentos respiratorios que como su nombre indica, permiten el intercambio gaseoso. Los embriones eclosionan luego de 22-24 horas a 25°C, luego de lo cual comienza el periodo larvario.
Figura 2: Vista lateral (panel superior) y dorsal (panel inferior) de un embrión de Drosophila melanogaster. Se observa la geometría hexagonal de las células del corion que lo recubre y los
filamentos respiratorios en la región anterior. - Desarrollo larvario
La larva de Drosophila tiene un cuerpo suave y flexible, que está cubierto por una cutícula no celular e internamente por una epidermis celular. La cutícula muestra en la región anterior de cada segmento numerosos anillos con garfios quitinosos. En la región ventral del segmento de la cabeza se puede distinguir la compleja armadura mandibular. La larva es bastante transparente y es posible observar, bajo luz transmitida, los cuerpos grasos e intestino (aparato digestivo), tráqueas y espiráculos (aparato respiratorio), tubos de Malpighi (aparato excretor) y gónadas (aparato reproductor). La observación de los testículos en el tercio posterior de la larva es utilizado para la selección de larvas por su sexo. Además de estas estructuras puramente larvarias, la larva contiene grupos celulares que originarán las estructuras del adulto, llamadas discos imaginales (imaginal, porque son importantes para la formación del imago). El estadío larvario se caracteriza por un aumento de masa a partir de nutrientes externos y está constituido por tres etapas, comenzando el primero luego de la eclosión de los embriones. La pequeña larva I se alimenta de la comida en la que los huevos fueron depositados, y luego de ± 25 horas, ésta muda su cubierta, desechando completamente su cutícula, aparato mandibular y espiráculos, convirtiéndose en una larva más grande, comenzando así el segundo estadío larvario, en que debe reconstruir los elementos desechados, lo cual permite la generación de estructuras nuevas. La larva II se alimenta durante ± 24 horas más, luego de lo cual muda y se convierte en una larva aún más grande, la larva III. Ésta última también se alimenta y crece, pudiendo alcanzar dimensiones de hasta 4,5 mm de largo, pero a su vez, comienza a “arrancar” de la comida hacia áreas más limpias y secas para comenzar el proceso de pupación. Esta etapa dura aproximadamente

30 horas luego de lo cual se libera un pulso de ecdisoma, hormona que provoca que la larva se convierta en pupa. Es posible acortar esta última etapa si se retiran los animales antes de la comida, siempre y cuando hayan alcanzado lo que se conoce como masa crítica, concepto sobre el que se detallará en la unidad 5. - Estadío pupal
La pupa es estacionaria (no se mueve), y es durante esta etapa que ocurre el proceso de metamorfosis desde larva hasta la anatomía del imago. Durante dicho proceso, ocurre una lisis masiva de las estructuras larvarias, aunque algunos de los órganos larvarios se conservan y reestructuran, como por ejemplo, el sistema nervioso que pasa por un proceso de reestructuración importante durante el proceso de morfogénesis. Sin embargo, la mayoría de las estructuras presentes en los adultos se forman a partir de tejidos indiferenciados que se replican durante el desarrollo larvario, que se llaman discos imaginales y los histoblastos. En las etapas tempranas, las pupas presentan una coloración entre blanca y amarilla, la cual se va oscureciendo a medida que el animal se desarrolla (Figura 3).
Figura 3: Durante la etapa P1, la pupa tiene una coloración blancuzca, que se va volviendo amarilla
en la transición P1-P4. De P5-P12, la cutícula se oscurece, por lo que las fotos corresponden a pupas a las que se les retiró la cutícula para una mejor ilustración. En P5 se comienza a observar la
morfología del animal en desarrollo, mientras que en P8 los ojos en formación comienzan a desarrollar pigmento. En P12, el animal está completamente desarrollado y a punto de eclosionar.
- Estadío adulto o imago
Una vez culminado el proceso de metamorfosis durante el estadío pupal, el adulto o imago eclosiona por el extremo anterior de la pupa. La mosca recién nacida es alargada y con alas retraídas, que luego de unos minutos se expanden y el animal se va redondeando gradualmente hasta adquirir la forma característica del adulto. A su vez, las moscas jóvenes tienen un color claro, y se translucen los desechos remanentes del proceso de pupación, que se observa como una mancha oscura en el abdomen, llamado meconio, pero al cabo de unas pocas horas, los animales se oscurecen, por lo que es fácil identificar a simple vista dentro de un cultivo, cuáles son las moscas jóvenes (Figura 4). Como ya se mencionó, estos animales presentan dimorfismo sexual (Figura 4), y los

machos son sexualmente activos alrededor de 8 horas luego de su eclosión, mientras que las hembras no poseen huevos maduros hasta 2 días posteriores a su eclosión. Por lo tanto, el reconocimiento de moscas jóvenes es una herramienta útil para el reconocimiento de hembras vírgenes (que no se hayan cruzado previamente con machos dentro del cultivo), que es importante a la hora de realizar cruces experimentales pues se tendrá noción del genotipo de la descendencia sin lugar a dudas. Una vez se realice un cruce, los machos fecundarán a las hembras, y el ciclo se retomará nuevamente.
Figura 4: Fotografía de animales adultos revelando dimorfismo sexual de la placa anal. Izq:
machos presentan una placa anal oscurecida y un tamaño corporal menor que el de las hembras (medio y derecha). Der: hembra virgen, presenta abdomen blancuzco y se trasluce claramente el
meconio.
Embriogénesis, Gastrulación, Establecimiento del plan corporal larvario y Proceso de segmentación.
La embriogénesis se refiere al conjunto de procesos biológicos que conllevan a la transformación controlada de una célula única a un individuo maduro. Posterior al evento de fecundación, en que los gametos masculino y femenino, ambos haploides, fusionan su material genético, generando un cigoto diploide, comienza el proceso de embriogénesis. - Embriogénesis temprana
El cigoto formado contiene toda la información genética del nuevo individuo a desarrollarse, el cual comienza a sufrir una serie de divisiones sincrónicas rápidas sin la ocurrencia de citoquinesis (sin división de citoplasma), con lo que se genera un gran sincicio (gran célula con múltiples núcleos) y luego se establece una monocapa de núcleos en la periferia que albergan al vitelo central, pero carecen de membranas plasmáticas que definan a las células individualmente; esta estructura se denomina blastodermo sincicial (típica del desarrollo de insectos). Cabe destacar que el ciclo celular 10 (Figura 5, estadío 4) marca el inicio de la transcripción cigótica, mientras que el ciclo celular 14 (Figura 5, estadío 5) marca el inicio

del estadío de blastodermo celular (3 horas post fecundación), y a partir de este momento, las divisiones celulares dejan de ser sincrónicas.
Figura 5: Esquema de un embrión de Drosophila durante distintas etapas de desarrollo embrionario
temprano. Estadío 1, una célula; estadío 2, sincicio; estadío 3, núcleos se desplazan hacia la periferia; estadío 4, blastodermo sincicial; estadío 5, blastodermo celular.
Durante el desarrollo embrionario temprano, ocurre un proceso de diferenciación de diversos tipos celulares en un orden espacial determinado, lo cual ocurre en parte gracias a la acción combinada de proteínas llamadas morfógenos. Estas proteínas se sintetizan o liberan de regiones determinadas y difunden hacia el resto del embrión, generándose gradientes de concentración. Por lo tanto, células que se encuentren más cerca de la fuente estarán expuestas a mayores concentraciones del morfógeno, mientras que lo opuesto ocurre para las que se encuentren más lejos. Esto es sumamente importante, puesto que los niveles de morfógeno a las que se encuentre sometida la célula blanco, determinarán su destino celular de manera dosis-dependiente, y una de las formas de describir este fenómeno es mediante el modelo de la bandera francesa, propuesto por Lewis Wolpert en 1969, cuya idea consiste básicamente en que una célula indiferenciada adquiere un destino celular u otro si es que la concentración de morfógeno a la que está expuesta excede un determinado umbral o no (Figura 6). Esto se explica dado que los morfògenos activan la expresión diferencial de genes por sobre determinados umbrales de concentración, pudiendo un mismo morfógeno activar la expresión de varios genes, cada uno con sus propios umbrales de activación. Por consiguiente, las células lejanas a la fuente del morfógeno solo expresarán genes con un

bajo umbral de activación, mientras que los que se encuentren cerca, expresarán tanto los que tengan un bajo como alto umbral de activación.
Figura 6: Si una célula tiene una concentración de morfógeno por sobre el umbral 1, tiene un
destino celular “azul”. Por su parte, si el morfógeno tiene una concentración menor al umbral 2, la célula tiene un destino “rojo”. Por último, si la concentración de morfógeno es intermedia, el
destino celular será “blanco”. Es evidente que el modelo de la bandera francesa es una simplificación de lo que ocurre realmente durante el desarrollo del animal, pues durante dicho proceso participa un conjunto de morfógenos, cada uno con su propia fuente de emisión y gradiente, por lo que cada una de las células estaría expuesta a una combinación de concentraciones de morfógeno, existiendo un verdadero código posicional para la identidad de cada una de ellas dentro del animal en desarrollo. - Gastrulación
La gastrulación comienza inmediatamente luego del establecimiento del blastodermo celular, y corresponde al proceso de reorganización de las células del embrión en desarrollo para formar las 3 capas germinales primarias: ectodermo, mesodermo y endodermo. La primera señal del proceso de gastrulación es la invaginación de la banda de células al centro de la cara ventral del embrión, formando el surco ventral. Las células invaginando van a formar el mesodermo presuntivo, mientras que las células a los bordes de la invaginación formarán los neuroblastos que formarán parte del sistema nervioso central de los animales. Durante la formación del mesodermo se genera un surco transversal llamado surco cefálico que se localiza en el borde anterior al surco ventral (Figura 7). Esta es una zona de ingreso de células que posteriormente formarán el intestino anterior, llamada la invaginación del intestino medio anterior. En la región posterior de los animales se genera una invaginación similar, llamada la invaginación del intestino medio posterior. Eventualmente, estas dos invaginaciones se encuentran en el medio, para formar el intestino endodermal.

Figura 7: A) Diagramas de vista lateral de embriones durante la gastrulación, indicando regiones precursoras de ectodermo (celeste), mesodermo (amarillo) y endodermo (rosa). Arriba hacia abajo: A1, blastodermo celular, el primordio está en la superficie del embrión; A2, el mesodermo presuntivo ha formado el surco ventral; A3, la
parte posterior del endodermo ha invaginado y la banda germinal ha comenzado a extender hacia el lado dorsal del embrión; A4, mesodermo completamente internalizado y ha comenzado a dispersarse para formar una capa
de célula única. B) Diagramas de visión lateral de los embriones en los mismos estadíos que en (A). Inmediatamente posterior al proceso de gastrulación, comienza el proceso de extensión de la banda germinal, donde aparecen los primeros signos visibles de segmentación. Nueve horas post fecundación, la banda germinal se retrae y el cuerpo adopta un ordenamiento antero-posterior convencional. - Segmentación
Una de las características comunes entre muchos animales es la formación de unidades morfológicas repetitivas llamadas segmentos o metámeros. Estos segmentos pueden ser esencialmente indistinguibles como lo es el caso de un ciempiés o bien cumplir funciones más especializadas, como por ejemplo, los segmentos torácicos que se asocian a funciones locomotoras, donde se desarrollan estructuras como alas y patas. En el caso de Drosophila melanogaster, la segmentación se cataloga de acuerdo a dos registros: los parasegmentos embrionarios y los segmentos larvarios/adultos, los cuales se

definen durante etapas tempranas de desarrollo mediante mecanismos moleculares sumamente regulados. La simetría bilateral que presentan estos animales está asociada a la existencia de dos ejes principales, ortogonales entre sí: antero-posterior (A/P) y dorso-ventral (D/V). A lo largo del eje A/P, la larva presenta una división con 12 segmentos: cabeza en la región anterior, seguida por una división regular de 3 segmentos torácicos y 8 segmentos abdominales, cada uno de los cuales posee una identidad definida, tanto en su cara externa como interna (Figura 8). En cuanto al eje D/V, éste define cuatro regiones en el cuerpo larval.
Figura 8: Segmentación, de izquierda a derecha: Cabeza, segmentos torácicos T1-T3 y segmentos
abdominales A1-A8. Por su parte, los animales adultos presentan una segmentación análoga a las larvas: la cabeza, 3 segmentos torácicos y 8 segmentos abdominales.
• La cabeza tiene a su vez tres segmentos, más una región no segmentada, y lleva la boca dotada de un complejo aparato mandibular. Hay un par de antenas y de voluminosos ojos facetados de color rojo.
• El tórax consiste de tres segmentos (protórax, mesotórax y metatórax), cada uno de los cuales lleva un par de patas provistas de numerosas articulaciones. Presenta a su vez un par de alas insertas en la superficie dorsal del mesotórax, y otro par de “alas” reducidas, o halterios, que se encuentra en la región dorsal del metatórax.
• El abdomen está compuesto por 8 segmentos que superficialmente adoptan la forma de anillos.
La determinación de segmentos y adquisición de características únicas para cada uno de ellos ocurre gracias a la acción concertada de diversos genes. Por su parte, existen genes cuya función no radica en la formación del patrón corporal, sino en la asignación de identidad a cada una de los parasegmentos una vez ya establecidos. Estos son los genes homeóticos (también conocidos como genes Hox), que corresponden a un grupo de genes relacionados entre sí que controlan la identidad de los distintos segmentos (antena, patas, etc). Los genes Hox codifican factores de transcripción capaces de encender y apagar genes, mediante un dominio de interacción con el DNA denominado “homedominio” (codificado en la secuencia nucleotídica por un dominio llamado homeobox). Asimismo, se ha descrito que en la mayoría de los animales, los genes Hox se encuentran ordenados en el

cromosoma de la misma manera en que su expresión se distribuye a lo largo del eje A/P del animal (Figura 9).
Figura 9: Esquema ilustrando los segmentos determinados por los genes homeóticos lab (labial),
dfd (deformed), scr (sex combs reduced), Antp (antennapedia), Ubx (ultrabithorax), abdA (abdominal-A) y AbdB (abdominal-B), cuyo código de colores representa los segmentos que
determina durante el estadio adulto (arriba) y larvario (abajo). Organogénesis. Regulación de la expresión génica y la selección de la identidad celular
La organogénesis corresponde al conjunto de procesos que permiten que células provenientes de las tres capas germinales, ectodermo, mesodermo y endodermo se diferencien hacia tejidos específicos para constituir los distintos órganos que forman parte del animal completo. En el caso de Drosophila, estos son:
• Ectodermo: Es la capa de tejido más externo de las 3 capas germinativas. Da origen a la epidermis del embrión, larva y adulto, que a su vez son células responsables de la secreción de cutícula en el adulto. Asociada fuertemente a la epidermis se encuentran los discos imaginales de las larvas, que posteriormente se convertirán en apéndices en el adulto. Esta capa germinal tambien da origen a las glándulas salivales de la larva.
• Mesodermo: Corresponde a la capa germinal intermedia. Da origen a células musculares y cardioblastos.
• Endodermo: Es la capa más interna de tejido, y dará origen al intestino. La división del blastodermo en los primordios de dichas tres capas germinales ocurre gracias a la acción conjunta de diversos genes. En la ausencia de dichos genes, solo se especifican destinos ectodermales, por lo que el estado ectodermal pareciera ser el estado por defecto. Existen vías de señalización que guían la fina regulación temporal y espacial durante este proceso, la cual es particular para cada uno de los tejidos. Cabe destacar que para el

establecimiento de identidad celular de los órganos internos, también participan algunos de los genes homeóticos. Asimismo, en etapas tempranas de desarrollo, las células se “comprometen” hacia determinados linajes, esto quiere decir que las posibilidades de los tejidos a los que potencialmente se podría transformar, son limitadas y a medida que la célula se compromete, este pool de posibilidades se va acotando aún más. Una vez la célula ha adquirido una identidad determinada, ésta expresa de manera regulada los genes requeridos para la mantención de las características del tejido al que pertenece. Los discos imaginales se forman a partir de células que se escinden del epitelio embrionario y heredan características genéticas particulares (genes del complejo Hox) que los definen en su destino adulto. Así encontramos discos de ala, halterios, pata (3 pares), ojo-antena, genitales y labiales. Desarrollo larvario y metamorfosis. Control hormonal del desarrollo - Desarrollo larvario
Las larvas de Drosophila tienen principalmente dos tipos de poblaciones celulares: las células larvarias, presentes en los tejidos utilizados para las funciones durante este estadío; y las células presentes en el tejido imaginal, que se encuentran dentro de las larvas en un estado indiferenciado, pero que corresponden a precursores para los tejidos presentes en el adulto o imago, y de ahí su nombre. Durante el desarrollo larvario de estos animales existen distintos mecanismos para el crecimiento de los tejidos. Por ejemplo, los tejidos larvarios como el intestino, las glándulas salivales, los cuerpos grasos y los músculos crecen principalmente por aumento del tamaño celular mientras que el número de células permanece constante. Dichas células aumentan su masa como consecuencia de un aumento en la carga genética de los núcleos celulares, esto quiere decir que las células replican su DNA pero no atraviesan la fase de mitosis, lo que lleva a la generación de núcleos de gran tamaño. Durante el proceso de metamorfosis estos tejidos son degradados, salvo los túbulos de Malpighi (sistema excretor). Otras estructuras necesarias en el adulto, como las glándulas salivales, se regeneran a partir de anillos imaginales. Por su parte, los discos imaginales y anillos imaginales crecen mediante proliferación celular, y a diferencia de los tejidos larvarios, no se degradan durante el proceso de metamorfosis sino que sufren un proceso importante de reestructuración. En Drosophila existen 10 pares de discos imaginales, que se convertirán luego en órganos adultos, y un disco genital (singular) que formará las estructuras reproductivas. A su vez, la epidermis abdominal forma un pequeño grupo de células imaginales llamadas histoblastos que yacen en la región del intestino de la larva. Los discos imaginales, al comienzo de su desarrollo corresponden a grupos reducidos de aproximadamente 30 a 40 células cada uno, las cuales proliferan rápidamente a tiempos

específicos y característicos, hasta alcanzar un número total de células al final del estadío larvario III cuando detienen el ritmo proliferativo. El disco más grande es el de ala, que prolifera hasta alcanzar aproximadamente 60.000 células, mientras que los de pata y halterio alcanzan alrededor de 10.000 cada uno. Asimismo, mientras las células proliferan, forman un epitelio tubular que se pliega en una especie de espiral compacto. - Metamorfosis
En insectos holometábolos, el proceso de metamorfosis (transformación de larva a adulto) ocurre adentro de la cutícula de la pupa. La mayor parte de los órganos larvarios son sistemáticamente destruidos, mientras que nuevos órganos adultos se desarrollan a partir de tejido precursor indiferenciado, el tejido imaginal. Durante el proceso de metamorfosis, los discos evierten (como “dar vuelta un calcetín”) y adquieren la forma y características particulares del apéndice en el individuo adulto.
Figura 10: Esquema del interior de una larva mostrando los discos con código de colores,
señalizando la estructura correspondiente en el animal adulto, conservando el mismo código.
Tanto los procesos de muda larvarios como el de metamorfosis de estos insectos están regulados por hormonas efectoras, que son controladas por neurohormonas en el cerebro. Estas son 20-hidroxiecdisona y la hormona juvenil lipídica. La ecdisona inicia y coordina cada muda y regula los cambios en la expresión génica que ocurren durante el proceso de metamorfosis. La hormona juvenil previene, durante las etapas juveniles, que la ecdisona induzca cambios en la expresión génica asociados a la metamorfosis, y como consecuencia, su presencia durante cada muda asegura que el resultado de dicha muda sea una larva más grande y no una pupa o un adulto.

SECCIÓN PRÁCTICA
1. Observación de cultivos de Drosophila melanogaster Se entregarán a los alumnos viales con individuos de Drosophila durante las distintas etapas de desarrollo: embriones, larvas y adultos. Los alumnos observarán el comportamiento de los animales, en cuanto a sus hábitos de alimentación (larvas) y vuelo (adultos), así como la distinta coloración que adquieren las pupas de acuerdo al estadío de desarrollo.
2. Reconocer las diferentes etapas del desarrollo y sus principales características
Los alumnos observarán bajo la lupa, animales en distintos estadíos de desarrollo para observar y dibujar las características morfológicas principales.
2.1.Embriones
Los alumnos observarán muestras de embriones que serán entregadas por el personal del laboratorio para observar características como el corion y filamentos respiratorios.
Para ello, se deberán aislar los embriones a partir de unas placas conteniendo levadura (que promueve la puesta de huevos por parte de los animales, pues es afrodisíaco para ellas) mediante lavado con agua de la llave. Luego se pasarán los embriones por un colador fino para lavarlos completamente y posterior a ello, se sacarán los embriones del colador con la ayuda de un pincel y se pondrán sobre una superficie de vidrio llamada portaobjeto.
Posteriormente se procederá a remover el corion de los embriones para poder observar las estructuras presentes bajo dicha capa de células. Para ello se deben repetir los pasos 1 y 2 del punto anterior y proseguir como se detalla a continuación.
Sumergir los embriones (dentro del colador) en una solución de cloro al 10% (preparada previamente con una parte de cloro doméstico y una parte de agua), procurando que los embriones no salgan del colador. Dejar dentro del cloro durante un minuto agitando el colador hacia los lados suavemente (como un pequeño “tiritón” de manos). Lavar con abundante agua para retirar todo exceso de cloro y sacar los embriones suavemente con un pincel y apoyar sobre un portaobjetos para mirar bajo la lupa. Reconocer estructuras como el surco cefálico.

2.2.Larvas
2.2.1. Estructuras externas Los alumnos deberán observar larvas bajo la luz de la lupa y notar detalles como los segmentos y los órganos que se puedan traslucir bajo luz blanca.
2.2.2. Estructuras internas Se proveerá a los alumnos de muestras preparadas de distintos órganos aislados de las larvas para que puedan observar con mayor detalle.
2.3.Pupas
Los alumnos deberán observar pupas de distintos estadíos bajo la lupa y notar la rigidez de la cutícula. A su vez los alumnos deberán distinguir estructuras como los espiráculos y el animal en desarrollo que se transluce a través de la cutícula. A su vez se entregará a los alumnos unas muestras de pupas fijadas a las que se les retiró la cutícula para que observen el animal que se encuentra dentro, durante distintas etapas del proceso de metamorfosis.
• Notar definición de estructuras adultas y aparición de pigmentación en los ojos de los animales.
2.4.Adultos
Se entregará a los alumnos viales con moscas adultas eutanasiadas para que reconozcan bajo la lupa los distintos segmentos de los animales, así como las diferencias entre sexos e individuos adultos y jóvenes. Para ello deberán:
Depositar entre 5 y 10 moscas en una placa de plástico y observar bajo la lupa. Si los alumnos desean mover los animales para observar desde distintos ángulos, podrán hacerlo suavemente con un pincel. Observar detenidamente para discriminar las características especificadas a continuación y dibujar las características más importantes.
2.4.1. Distinción entre moscas jóvenes y viejas
Existen características fáciles de identificar a simple vista para distinguir si un animal es joven o viejo.
Las moscas viejas son más oscuras y rígidas que las moscas jóvenes. Las moscas muy jóvenes (pocas horas posteriores a su eclosión de la pupa) serán aún más claras y será posible distinguir una mancha oscura en el abdomen, llamada “meconio”, que corresponde a desechos residuales del proceso de metamorfosis. Si la mosca es aún más joven podría incluso visualizar que sus alas no se han desplegado del todo, se observará una estructura plegada y poco rígida.

2.4.2. Reconocimiento de segmentos y apéndices La mosca presenta una serie de segmentos fácilmente distinguibles, que se le solicitará al alumno que observe y posteriormente dibuje. Primero, el alumno deberá observar que el cuerpo de la mosca se divide en 3 segmentos principales: la cabeza, el tórax y el abdomen, dentro de los cuales podrán observar características con mayor detalle: - Cabeza: ojos, antenas, aparato chupador. - Tórax: alas, halterios, patas (1 par por segmento). - Abdomen: bandas oscuras, placa anal.
2.4.3. Identificación del sexo Existen varias diferencias morfológicas permiten distinguir los machos de las hembras de Drosophila.
El extremo del abdomen es alargado en la hembra y algo redondeado en el macho. La distribución de bandas oscuras en los segmentos abdominales permite distinguir a ambos sexos sin necesidad de usar microscopio: El abdomen de la hembra tiene 7 segmentos y el macho 5. Los machos poseen el denominado peine sexual, un cepillo de más o menos 10 gruesas cerdas negras ubicadas en la superficie distal de la articulación tarsal del primer par de patas. El disco anal de los machos es más oscuro que el de las hembras, y probablemente ésta la característica más fácil de identificar para la distinción entre machos y hembras.

Objetivo específico 2: Reconocer cómo el genoma es determinante del desarrollo
SECCIÓN TEÓRICA
INTRODUCCIÓN
Una de las principales preguntas de la biología es cómo a partir de una célula se origina un individuo completo y en este mismo sentido, cómo a partir de una célula se originan los distintos tipos celulares que poseen la misma secuencia de ADN, preguntas que los biólogos del desarrollo tratan de responder, principalmente mediante estudios genéticos (utilizando mutantes). El control preciso de la expresión genética tejido-específica es central para la diferenciación celular y el desarrollo. Para controlar estos procesos, los programas de desarrollo utilizan redes genéticas regulatorias que consisten de múltiples componentes que ejecutan los distintos procesos involucrados en la diferenciación, donde se requiere una regulación génica altamente dinámica, con el fin de encender y apagar genes cuando sea necesario durante el desarrollo y para responder a señales ambientales específicas. Todo esto se ha estudiado en diversos modelos animales, donde el más utilizado es el uso de Drosophila melanogaster debido a las invaluables herramientas genéticas que este organismo proporciona. Los primeros genes que participan en el desarrollo son aquellos heredados por la madre en forma de ARN mensajeros (ARNm), por lo que comenzaremos estudiando su producción y posterior participación durante el desarrollo. Desarrollo en Drosophila melanogaster. Como se mencionó, la formación de patrones del embrión de Drosophila empieza con señales proveídas maternalmente, las que se transforman posteriormente en gradientes de factores de transcripción que controlan la expresión de genes blanco río abajo a lo largo de los ejes antero-posterior y dorso-ventral. Los blancos de estas vías incluyen proteínas regulatorias y estructurales que colaboran para definir la posición y la identidad de los segmentos larvales y para controlar la diferenciación de las capas germinales. Los precursores de los gametos se dividen mediante meiosis para generar células haploides (con un solo conjuntos de cromosomas, a diferencia de las diploides que poseen dos). En este punto cabe recordar que las células eucariontes pueden dividir sus núcleos tanto por mitosis como por meiosis, la primera ocurre en células somáticas, precede a la división celular y consiste en repartir de manera equitativa del material hereditario (ADN) y finalmente produce dos células hijas idénticas, es un proceso esencial en el crecimiento, reparación de tejidos y en la reproducción asexual. Mientras que la meiosis permite la división de los gametos y produce células genéticamente distintas y es el fundamento de la reproducción sexual y de la variabilidad genética. Durante la meiosis, se producen dos divisiones sucesivas y así se generan cuatro células haploides. Ambas divisiones comprenden las etapas profase, metafase, anafase y telofase, pero no existe replicación de ADN entre ambas.

Oogénesis. El sistema reproductor femenino contiene dos ovarios, cada uno con aproximadamente una docena de ovaríolos que contienen una línea de ensamblaje de cámaras de huevo en maduración. Cada cámara de huevo contiene un oocito y 15 células nodrizas, las que derivan de la línea germinal y están rodeadas por una monocapa de células foliculares somáticas (esto se analizará con más detalle en el Objetivo 3). Las células nodrizas endoreplicadas pasan por un período de transcripción masiva y son responsables de la síntesis de la mayoría de los productos maternos, los que son depositados en el oocito en desarrollo a través de puentes citoplasmáticos. Durante su residencia de tres días en la cámara de huevos, el oocito es detenido en la Profase I de la meiosis. Tres horas antes de ser liberado, el oocito madura y progresa hacia Metafase I y ciertos ARNm maternos latentes, se activan transcripcionalmente. Activación del huevo. Este fenómeno sirve para gatillar eventos celulares y moleculares adicionales que preparan al oocito maduro para la embriogénesis temprana. En Drosophila, la activación del huevo gatilla la completación de la Meiosis, la depolimerización de microtúbulos y cambios postranscripcionales que incluyen poliadenilación citoplasmática, activación traduccional y desestabilización de ARNm. Además, el huevo se hincha debido a la activación y la membrana vitelina, que es el componente más interno de la cáscara del huevo, se hace impermeable. La activación precisa del huevo en Drosophila ha sido difícil de estudiar debido a que ocurre muy rápidamente y dentro de la mosca adulta. Fertilización. En el reino animal, los individuos se originan mediante la fusión de dos células haploides altamente especializadas (gametos), el espermio y el oocito. Luego de la fertilización, se forman los pronúcleos femenino y masculino y las proteínas y ARNm maternos se depositan en el oocito fertilizado donde ejercen su función. Durante esta primera etapa el genoma cigótico es transcripcionalmente inactivo. Embriogénesis temprana. La particularidad de Drosophila en este proceso es que ocurren 13 divisiones nucleares rápidas y sincrónicas, sin citoquinesis, produciendo 5000 núcleos dentro de un mismo citoplasma. Los eventos de la embriogénesis temprana ocurren en ausencia de la transcripción cigótica y por lo tanto son dependientes de las moléculas proveídas maternalmente. Las primeras siete divisiones nucleares ocurren en el interior del embrión, pero durante los ciclos octavo y noveno, la mayoría de los núcleos migran hacia la periferia. Los primeros núcleos en llegar al polo posterior formarán las células del polo, que darán lugar a la futura línea germinal. Para los núcleos restantes (totipotenciales), los ciclos 10 a 13 ocurren en la superficie del embrión y corresponden al blastodermo sincicial. Durante la interfase del ciclo 14, aproximadamente 2,5 horas después de la fertilización, la membrana plasmática se invagina entre los núcleos para formar el blastodermo celular (Figura 5).

Con la excepción de unos pocos genes cigóticos tempranos, es durante la interfase del ciclo 14 donde ocurre la mayoría de la transcripción cigótica (en este punto ya hay aproximadamente 2000 núcleos), representando la Transición materna a cigótica (MZT) en la expresión genética, proceso que incluye la degradación de los transcritos maternos que son sustituidos por los transcritos cigóticos. Esta programación transcripcional resulta en cambios morfológicos drásticos. La activación del genoma cigótico ocurre en dos oleadas distintas: una menor que activa sólo una decena de genes seguida por una mayor que activa varios cientos de genes. Unos 40 minutos después de esta transición ocurren tres rondas más de divisiones nucleares y luego comienzan a migrar hacia la periferia, formando un blastodermo sincicial con unos 6000 núcleos dispuestos en monocapa en la superficie. Después de una larga interfase, durante la cual las membranas celulares invaginan para formar el blastodermo celular, se establece el plan corporal básico y comienza la gastrulación. La celularización marca la transición de la blástula media como el primer evento del desarrollo que depende de productos cigóticos. Durante las siguientes horas del desarrollo, los patrones de expresión genética especifican la identidad de segmento, las tres capas germinales y los tipos celulares dentro de estos tejidos. Factores maternos. La formación de patrones en el embrión de Drosophila ocurre por la información posicional establecida por los factores maternos, también denominados genes de polaridad del huevo, ya que son los genes que determinan la constitución de los ejes principales de desarrollo de Drosophila: el eje antero-posterior (A/P) y el eje dorso-ventral (D/V). La transición inicial de las células indiferenciadas está mediada por gradientes de dos factores de transcripción: Dorsal establece el eje dorso-ventral del embrión mientras Bicoide determina el eje antero-posterior. bicoide es un gen materno acumulado específicamente en la región cefálica (anterior) del oocito y es traducido después de la deposición del huevo, resultando en una alta concentración de la proteína Bicoide en el polo anterior con niveles progresivamente más bajos hacia el polo posterior. Esta proteína funciona como un factor transcripcional y los altos niveles de Bicoide median la formación de las estructuras más anteriores, los niveles intermedios de este mismo median la formación de las estructuras de la cabeza y niveles bajos las estructuras abdominales anteriores y torácicas. Además, Bicoide induce la expresión del gen hunchback, uniéndose a la zona promotora de éste y así ambos se acumulan en la zona anterior del embrión (Figura 11). Cambiando los niveles de la proteína Bicoide resulta en un cambio en la identidad posicional, lo que permite sostener que Bicoide actúa como un morfógeno que instruye el destino celular en una manera dependiente de su concentración. De la misma forma, Dorsal también activa genes de manera dependiente de concentración. Otro gen materno que se distribuye de manera opuesta a bicoide es nanos, ya que este último se acumula especialmente en la región posterior del embrión y su concentración va disminuyendo hacia el segmento anterior (Figura 11). Esta distribución es regulada por otros genes como oskar, que mantiene la ubicación del ARNm de nanos antes de la fecundación. La actividad de Nanos es inhibir la traducción del ARNm del gen hunchback. Este último se distribuye por todo el oocito antes de la fertilización, pero luego Bicoide promueve su expresión cigótica exclusivamente en la zona anterior, mientras nanos inhibe su traducción en el extremo posterior. Otro de los genes de origen materno que participa en
el establecimiento del eje antero-posterior es caudal, que originalmente está distribuido en todo el oocito, pero luego Bicoide inhibe su traducción en la región anterior. El establecimiento del eje dorso-ventral es uno de los eventos más tempranos de la formación de patrones en el embrión de Drosophila en desarrollo. Eventos durante la oogénesis, incluyendo el proceso de transporte de morfógenos, resulta en una activación localizada del receptor transmembrana Toll. Luego, diversas etapas de señalización llevan a la localización nuclear en un gradiente dorso-ventral del factor de transcripción Dorsal, que actuará como referencia para la formación de patrones posteriores, subdividiendo al embrión en regiones como aquellas con altos niveles de Dorsal, que darán origen al mesodermo y aquellas regiones con niveles intermedios darán lugar al neuroectodermo. Respecto de Dorsal, se ha observado que al inicio de la embriogénesis esta molécula se encuentra dispersa por todo el embrión y tras la migración de los núcleos a la periferia, esta proteína penetra en los núcleos celulares de las regiones ventrales y laterales, pero permanece en el citoplasma en la región dorsal. Este mecanismo de transporte dorsal está regulado por una proteína de señalización denominada Spätzle, que presenta un gradiente ventral a dorsal y es un ligando de los receptores Toll y como se activa dependiendo de la abundancia de su ligando, está en plena actividad en la zona ventral del embrión y decrece progresivamente en las regiones más laterales. Cuando Toll inicia su cascada de señalización ocasiona la degradación de un inhibidor citoplasmático denominado Cactus, lo que provoca su activación y así se genera el gradiente de Dorsal, especialmente en la zona ventral y más diluido en las regiones laterales. Dorsal afecta la actividad de otras proteínas dependiendo del umbral de sensibilidad de éstas ya que para algunas es activo como estimulante a bajas concentraciones y para otras requiere altas concentraciones. El gradiente generado de Dorsal repercute en la regulación de tres genes diana activados por altas, intermedias y bajas concentraciones de Dorsal: twist, rhomboid y sog, respectivamente.
Figura 11: Distribución diferencial de los ARNm (izquierda) y proteínas (derecha) de bicoid, nanos, hunchback y caudal en el embrión temprano de Drosophila.

Segmentación. Después de la acción de los genes maternos, se activan los genes cigóticos o genes de segmentación que se expresan tras la fertilización. La segmentación a lo largo del eje antero-posterior comienza con el establecimiento de cuatro regiones anchas por acción de los genes gap. Después de la gastrulación se forma una serie de surcos denominados parasegmentos y definen las áreas corporales (Figura 12). Bicoide y Hunchback activan la expresión del gen even-skipped. La identidad de cada parasegmento está dada por la acción de los genes pair-rule (eve y fushi tarazu) y de los genes de polaridad de segmentos, ya que cada uno se expresa en series de siete elementos transversales al eje antero-posterior de forma alternada (pares e impares). Finalmente, participan los genes de polaridad de segmento, los que determinan la conversión de los parasegmentos en segmentos definitivos (Figura 12).
Figura 12. Esquema que muestra los dominios de expresión de los genes Gap, Pair-rule y de polaridad de segmentos en el embrión en desarrollo para el establecimiento de parasegmentos.
Luego, el grupo de genes que controlan el establecimiento de la identidad de segmento se denominan complejos de genes homeóticos o Hox y son los últimos genes cigóticos que actúan a lo largo del desarrollo y se activan tras el efecto de los genes de la polaridad de los segmentos, la expresión de estos genes determina las estructuras del adulto que se formarán en cada segmento corporal. En Drosophila, se sabe que estos genes se agrupan en dos complejos. Uno es el Complejo Antennapedia que controla la identidad de los segmentos

de la cabeza y de los dos primeros segmentos torácicos y está compuesto de 5 genes homeobox: labial, proboscidea, deformed, sex combs reduced y antennapedia. El segundo es el Complejo Bithorax que controla la identidad del tercer segmento torácico y de los segmentos abdominales y se compone de tres genes con homeobox: ultrabithorax, abdominal-A y abdominal-B. Morfogénesis. Durante esta etapa, las células adquieren información posicional en un proceso que involucra una clase particular de moléculas de señalización: los morfógenos. Los morfógenos son ligandos que son secretados a partir de un grupo restringido de células. Luego de su secreción, los morfógenos se mueven a partir de la fuente para formar gradientes de concentración. La información posicional está codificada en la distribución graduada de los morfógenos: las células son capaces de “leer” su distancia a la fuente de sereción midiendo las concentraciones de morfógeno que las rodean. Los morfógenos se mueven rápido desde su fuente de producción y establecen un gradiente estable que dura varios días. Un ejemplo es Dpp, un morfógeno que provee información a lo largo del eje antero-posterior durante el desarrollo del ala. Dpp activa directamente la expresión de genes blanco en una manera dependiente de concentración. Los sitios de unión de los factores de transcripción son cruciales para controlar la expresión de sus genes blanco, pero no se sabe cómo integran la información para producir patrones de expresión gen-específico. Durante la morfogénesis es primordial que se genere un cambio en la adhesión celular (dado por la presencia diferencial de moléculas de adhesión, como cadherinas, por ejemplo), lo que implica que un grupo de células presenta una adhesión selectiva respecto de otro grupo. Esto favorece la formación de las tres capas germinales: ectodermo, mesodermo y endodermo. Estudio en mutantes. Para conocer todos los procesos mencionados, los biólogos del desarrollo han utilizado principalmente organismos (como Drosophila melanogaster) mutantes en diferentes estadíos del desarrollo y así se ha determinado la importancia genética de las distintas etapas del desarrollo, donde cada etapa está controlada por un grupo de genes diferente. Debido a que los genes reguladores del patrón corporal están en una posición alta de jerarquía, mutaciones en algunos de los componentes de estos complejos pueden provocar grandes cambios en el fenotipo de las larvas, fenómeno denominado homeosis y corresponde a la conversión de una parte del cuerpo en otra. El ejemplo más notorio de este efecto son mutaciones de ganancia de función del complejo Antennapedia que convertirá las antenas en patas.

APOYO TEÓRICO PARA PROTOCOLO Protocolo 1: Observación de embriones en distintos estadíos Para esta etapa, previamente se habrá realizado una “puesta” de moscas wild type (o silvestres) para así obtener al día siguiente embriones en distintos estadíos del desarrollo. Esta “puesta” implica dejar las moscas cruzándose toda la noche en una placa especial, que contiene levadura para promover la obtención de un alto número de huevos. A continuación se muestran distintas imágenes de la morfología que presentarán los embriones en los distintos estadíos, que corresponde a lo que deberán observar los alumnos.
Blastodermo sincicial
Aparición células del polo (flecha)
Blastodermo celular
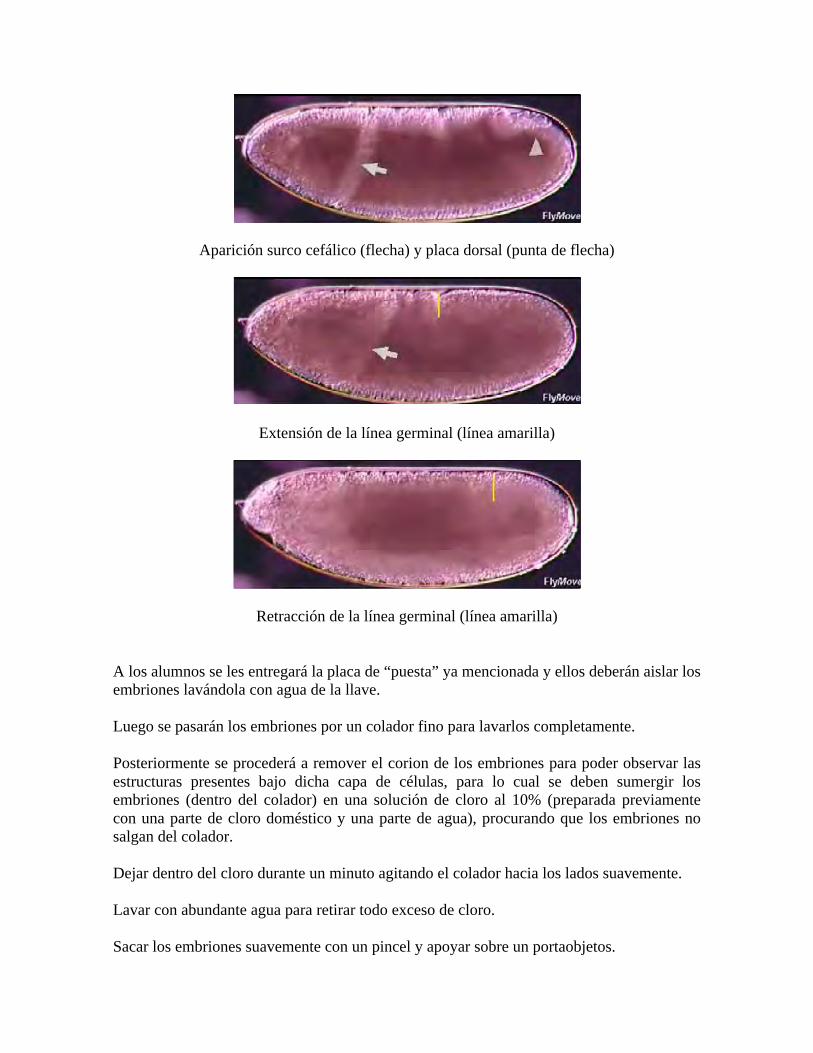
Aparición surco cefálico (flecha) y placa dorsal (punta de flecha)
Extensión de la línea germinal (línea amarilla)
Retracción de la línea germinal (línea amarilla) A los alumnos se les entregará la placa de “puesta” ya mencionada y ellos deberán aislar los embriones lavándola con agua de la llave. Luego se pasarán los embriones por un colador fino para lavarlos completamente. Posteriormente se procederá a remover el corion de los embriones para poder observar las estructuras presentes bajo dicha capa de células, para lo cual se deben sumergir los embriones (dentro del colador) en una solución de cloro al 10% (preparada previamente con una parte de cloro doméstico y una parte de agua), procurando que los embriones no salgan del colador. Dejar dentro del cloro durante un minuto agitando el colador hacia los lados suavemente. Lavar con abundante agua para retirar todo exceso de cloro. Sacar los embriones suavemente con un pincel y apoyar sobre un portaobjetos.

Mirar bajo la lupa. Indicar a los niños que observen los embriones obtenidos e identifiquen en ellos los siguientes estadíos: - Observar sincicio - Observar formación células del polo - Observar celularización - Movimientos gastrulación - Observar extensión y retracción de la banda germinal Nota: Discutir con los alumnos los cambios de forma que ocurren durante el desarrollo de Drosophila melanogaster. Propuesta: Tinción con hematoxilina para ver proliferación en embrión (mitosis) Protocolo 2: Observación de mutantes antennapedia Como se mencionó al principio de esta unidad, el homeobox corresponde a un sector del DNA genómico qua incluye una secuencia de 180 pares de nucleótidos que codifican para un dominio proteico de 60 aminoácidos. Los genes homeóticos (como antennapedia), al igual que los genes involucrados en el control de la segmentación de la mosca, tienen un homeohox. Si uno genera alteraciones en la expresión de estos genes se observarán aberraciones en el desarrollo como errores en la identidad de órganos. El ejemplo más claro de esto es la mutación en el gen antennapedia que genera la aparición de patas en el lugar de las antenas (Figura 13).
Figura 13: A la izquierda se muestra una mosca silvestre y a la derecha una mosca mutante
para antennapedia.

A los estudiantes se les entregará un tubo con diversos individuos en etanol, incluyendo moscas silvestres y mutantes para el gen antennapedia y ellos deberán distinguirlos. Protocolo 3: observación individuos transgénicos Sistema Gal4/UAS El sistema de expresión GAL4-UAS es una poderosa herramienta genética desarrollada por Andrea Brand y Norbert Perrimon en 1933 y es ampliamente utilizada en Drosophila melanogaster para estudiar la expresión de genes, ya que permite la expresión tejido-específica de distintos genes de interés. Este sistema consiste de dos componentes: el activador transcripcional de levadura Gal4, expresado en un patrón específico (dado por un promotor particular, de interés) y el enhancer UAS (Upstream Activation Sequence) al cual se une específicamente Gal4 para activar la transcripción génica. Por lo tanto, uno puede utilizar este sistema para evaluar la expresión de los distintos genes necesarios durante el desarrollo, teniendo el ADN que codifica para GFP junto a una secuencia UAS y el activador Gal4 con el promotor del gen que nos interesa evaluar, y así veremos que las células se verán verde exclusivamente donde se exprese nuestro gen en cuestión.
Figura 14: Esquema del funcionamiento del sistema Gal4/UAS. En esta sección utilizaremos este sistema para observar los patrones de expresión de los siguientes genes en Drosophila melanogaster: engrailed, nanos y ultrabithorax. Estos genes tienen en común que actúan como factores de transcripción involucrados en el desarrollo de este insecto, ya que actúan confiriendo una identidad específica a regiones definidas del cuerpo y también coordinan la expresión de genes río abajo.

Se entregará a los alumnos los animales ya mencionados en estadío larvario y se observará los patrones de expresión de los genes ya mencionados ya que expresarán una proteína fluorescente verde (GFP), que se podrá observar al iluminar las larvas con un láser verde, pues las larvas que expresen GFP emitirán más luz que las que no lo expresen.
1. Tomar los viales conteniendo larvas. 2. Sacar las larvas de los tubos con pincel y ponerlas sobre una placa de plástico con
agua para lavarlas. 3. Agarrar suavemente las larvas con una pinza y traspasarlas a un pocillo conteniendo
agua limpia (este paso es importante pues la presencia de comida en el agua puede dificultar la observación de las larvas y dañar el experimento).
4. Observar las larvas a simple viste y comparar con animales silvestres. 5. Apagar las luces y apuntar las larvas con un láser verde y observar la tonalidad de
las larvas. 6. Registrar observaciones.

Objetivo específico 3: Control hormonal del desarrollo de Drosophila melanogaster. SECCIÓN TEÓRICA INTRODUCCIÓN. La mosca en un excelente sistema de estudio de los mecanismos moleculares que controlan las transiciones en el desarrollo de organismos multicelulares, principalmente, por la presencia de etapas claramente diferenciables, como la muda de la piel y la metamorfosis. Estos eventos de transición están controlados estrictamente por una señalización endocrina, la cual es mediada por moléculas denominadas hormonas, que actúan en células dianas distantes de sus órganos de síntesis. El desarrollo de la mosca está controlado por diversas hormonas, pero el regulador maestro es una hormona esteroidea llamada Ecdisona (E). La ecdisona es sintetizada en las glándulas protorácicas y luego secretada a la hemolinfa, donde es convertida a 20-hidroxi-ecdisona (20E). La 20E es recepcionada por sus tejidos diana, donde gatilla cambios fisiológicos, morfológicos y de comportamiento, asociados a la muda de la piel y la metamorfosis. Durante el ciclo de vida de la mosca, distintos peaks de ecdisona marcan importantes transiciones del desarrollo postembrionico. La respuesta más dramática a ecdisona se produce en el término del estadio de larva III, donde una alta concentración de esta hormona gatilla la formación del pupario. 10 hrs más tarde, un nuevo pulso de ecdisona induce la transición de prepupa a pupa.
Figura 15: Cambios en los niveles de Ecdisona en relación al ciclo de vida de la mosca. Las etapas
de la larva están separadas por mudas, que son controlados por pulsos de Ecdisona. Luego se produce nuevamente un pulso de Ecdisona para marcar la transición de larva a pupa y gatillar la
metamorfosis.

Los primeros antecedentes del mecanismo de cómo la ecdisona inicia la transformación proviene de los cromosomas politénicos presentes en las glándulas salivales de la larva. Los cromosomas politénicos son cromosomas gigantes en interfase, en que el ADN se replica por endomitosis, las cromátidas hermanas no se separan y se encuentran alineadas. Los órganos que poseen este tipo de cromosomas no crecen por proliferación celular, sino por el aumento de tamaño de las células, es decir, estas células no se dividen, produciéndose ciclos S-G1, manteniendo por tanto un estado permanente de interfase (sólo replican su ADN, sin división celular). La característica más distintiva de los cromosomas politénicos es su patrón de bandas. Este patrón puede ser descrito citológicamente como una secuencia repetitiva y aperiódica de bandas de altas intensidades de coloración y concentración de masa, denominadas bandas o cromómeros, conectadas por secciones con muy poca concentración de masa o coloración, llamadas interbandas o intercromómeros. Los cromosomas politénicos permiten el estudio de la expresión génica y de la diferenciación celular debido a que se producen cambios estructurales de la región cromosómica, en que ocurre una expansión denotando zonas de alta transcripción génica. Estas zonas son denominadas “puffs”. Así, cada cromosoma politénico posee un conjunto definidos de “puffs” en cada estadio de larval.
Figura 16: Esquema de cromosomas politénicos. De esta forma, se pudo analizar el efecto de la ecdison sobre la expresión génica para iniciar las transiciones de una etapa a otra en el ciclo de vida de la mosca, dada la aparición de “puffs” característicos en regiones definidas de los cromosomas en los periodos de aumento de esta hormona.

Como se mencionó anteriormente, la ecdisona es importante para la formación de la pupa. Durante el estadío pupal ocurre la metamorfosis, que es la reorganización del plan corporal de la mosca. Esta reorganización dura alrededor de 4 días y consiste en la destrucción por histólisis de la mayoría los órganos larvales y que son reemplazados por tejidos adultos que se desarrollan a partir de los discos imaginales. Los discos imaginales son estructuras epiteliales que se forman a partir de un pequeño número celular, que prolifera y diferencia para formar las estructuras adultas. Otro aspecto importante en el desarrollo de la mosca es el estado nutricional y como este es censado para gatillar el avance en las distintas etapas del ciclo de vida. Durante el estadío larval III temprano la larva crece hasta alcanzar un punto importante denominado, masa crítica, en que la larva ha adquirido suficientes nutrientes para completar el estadío larval sin la necesidad de alimentarse nuevamente. La restricción de nutrientes antes de alcanzar la masa crítica retrasa el desarrollo de la larva, sin embargo, cuando esta restricción ocurre después de este punto, la larva es capaz de pupar, sin un retraso en el desarrollo. Cuando la larva ha crecido en condiciones normales de nutrientes, el cuerpo graso, un tejido encargado del almacenamiento de energía, señaliza al cerebro para la liberación de insulina, esta última actúa sobre la glándula protorácica induciendo la liberación de ecdisona, la que finalmente inicia la pupación. En la última década se ha determinado que la ecdisona no solo actúa en el desarrollo de la larva o en la metamorfosis, sino que también juega un importante rol en adultos, como en la formación del ovario y la oogénesis. La ecdisona participa a la mitad de la oogénesis, como punto de control para asegurar una correcta producción del huevo. El ovario de la Drosophila está compuesto por 15-20 ovariolos que se encuentran en un desarrollo continuo. Cada ovariolo se puede dividir en 14 etapas basándose en un criterio morfológico. La etapa 14 corresponde al huevo maduro y la primera etapa es la cámara del huevo naciente en la región anterior de los ovariolos, llamado el germario. La etapa 8 es un importante punto de control durante la formación del huevo, debido a que se determina si la cámara del huevo sigue desarrollándose o muere. En esta etapa los niveles de ecdisona son relevantes, debido a que bajos niveles de esta hormona son necesarios para un correcto desarrollo del huevo y altos niveles van a inducir la muerte. En mayor detalle, la cámara del huevo está compuesta por un oocito, las células nodrizas y el folículo somático. El germario se encuentra en la punta del ovaríolo y es en esta región donde residen las células madres que se dividen de forma asimétrica, generando una nueva célula madre y una célula denominada cistoblasto. El cistoblasto experimenta 4 divisiones mitóticas generando 16 células que se encuentran unidas por puentes citoplasmáticos. Este grupo de células se conoce como quiste de la línea germinativa. Una de estas células se convertirá en el oocito y las otras 15 en las células nodrizas, que producen gran cantidad de proteínas y ARN mensajero que será transportado al oocito. Por otra parte, las células somáticas formaran el folículo.
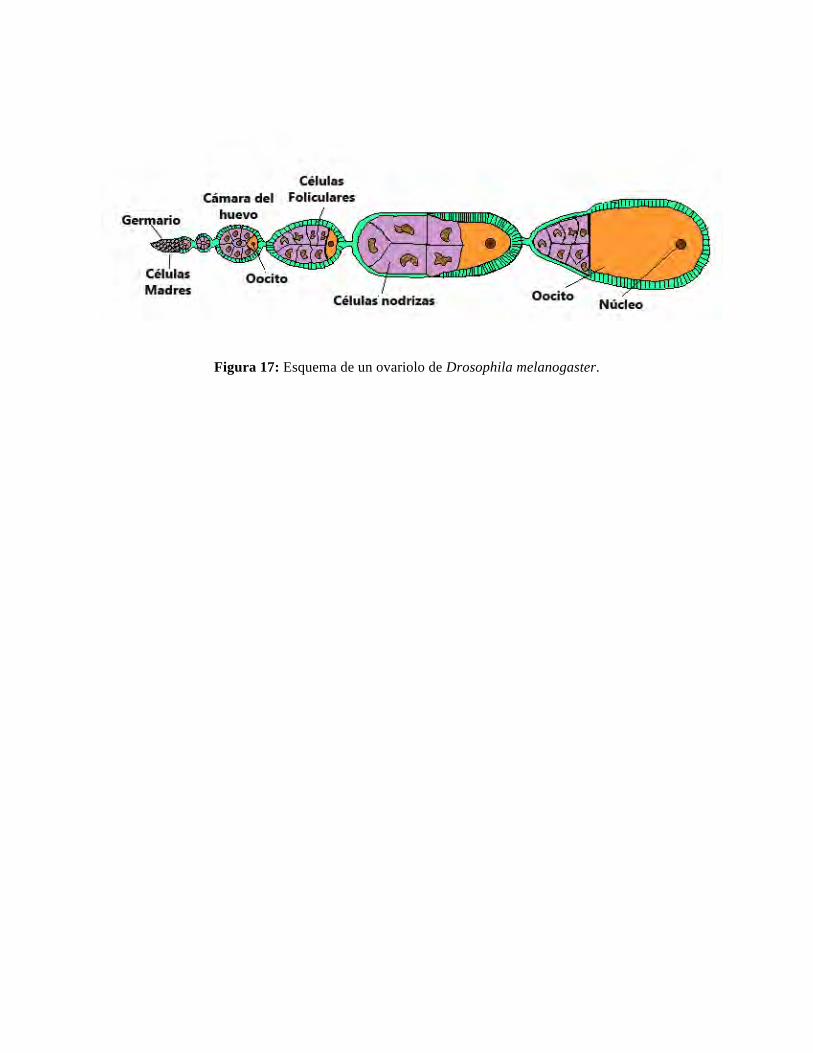
Figura 17: Esquema de un ovariolo de Drosophila melanogaster.

APOYO TEÓRICO PARA PROTOCOLO Disección de glándulas salivales
Coloque la larva en un portaobjeto con solución de PBS. Con ayuda de sus pinzas divida la larva en dos. Inserte unas de las pinzas en la mandíbula y de vuelta la larva como un calcetín. Asociadas a la mandíbula encontrará las glándulas salivales que son dos tejidos delgados y alargados que se encuentran unidos a un lóbulo de cuerpo graso. Separe las glándulas salivales de la mandíbula y del cuerpo graso. Dado que las células de las glándulas salivales poseen cromosomas politénicos, son células de gran tamaño. Observe las células.
Observación de cromosomas politénicos.
Observe en el microscopio y reconozca en las preparaciones el cromocentro, bandas e interbandas y las regiones de activa expresión o “puffs”.
Observación de Pupas.
Reconozca los distintos estadíos pupales y observe la aparición de estructuras adultas.
Disección de ovaríolos.
El instructor le ayudará a disecar el abdomen de una hembra y a colocar su contenido en una gota de solución salina. Reconozca los ovaríolos que aparecen como un par de conjuntos de estructuras filamentosas y blanquecinas que se insertan en la región más posterior del abdomen. Con la ayuda de un par de alfileres entomológicos separe los ovaríolos, retire la solución salina con papel filtro y adhiera el tejido al porta objetos por desecación. Después de algunos segundos fije la preparación con formaldehido al 5%.

Después de 5-10 min lave con varios cambios de agua destilada y tiña con una solución diluida de azul de metileno por algunos minutos. Lave nuevamente con agua destilada y monte la preparación con un cubre objeto premunido de topes de plasticina. Algunas preparaciones podrán observarse sin tinción.
NOTA: Cada ovaríolo consiste de una sucesión de cámaras, cada una llamada germario o quiste germinal, cuyo tamaño aumenta desde el extremo distal al proximal del ovaríolo. Las más distales son las menos desarrolladas. Cada germario está revestido por el epitelio folicular y los menos desarrollados contienen los primeros productos de división de una ovogonia.
Examine los quistes más desarrollados y podrá distinguir las células del clon. Detecte aquellos quistes en los cuales es posible distinguir el ovocito y las células nodrizas. El crecimiento del ovocito se acompaña de un aumento progresivo de su densidad óptica y de una disminución del tamaño de las células nodrizas. Elabore los esquemas correspondientes.

BIBLIOGRAFÍA Gilbert, Biología del Desarrollo, Séptima edición. Wunderlich, DePace, 2011. Modeling transcriptional networks in Drosophila development at multiple scales. Current Opinion in Genetics snd Development, 21: 711-718. MacNamara, 2014. Multiscale modeling of dorsoventral patterning in Drosophila. Seminars in Cell and Developmental Biology, 35: 82-89. Semotok, Lipshitz, 2007. Regulation and function of maternal mRNA destabilization during early Drosophila development. Differentiation, 75: 482-506. González-Gaitán, 2003. Endocytic trafficking during Drosophila development. Mechanisms of Development, 120: 1265-1282. Boija, Mannervik, 2015. A time of change: dynamics of chromatin and transcriptional regulation during nuclear programming in early Drosophila development. Molecular Reproduction and development, 82: 735-746.

TRABAJOPRACTICOUNIDAD4

TALLERDEFECUNDACIÓNYBIOLOGÍADELDESARROLLO
Elpezcebracomomodelodeldesarrollodevertebrados

ParteI.Dimorfismosexual,fecundaciónydesarrollotempranodelpezcebra
Identificacióndemachosyhembras
Identificar hembras y machos es una habilidad fundamental que debe ser dominada por uninvestigadorquepretendeutilizaralpezcebracomosumodelodeestudio.Machosyhembrassevenmuyparecidosalojopocoentrenado.Sólolaprácticaayudaadominarestahabilidad.EnlaFigura1sepresentaunalistadecaracterísticasqueayudanasuidentificación.
Figura 1: Comparación entre peces cebra de distintossexos.
Hembras:· Coloraciónmásclaraquelosmachos· Engeneralsonmásgrandesquelosmachos· Son más redondas que los machos en la región
ventral· Presentan una coloración rojiza cerca de las
aletasMachos:
· Presentan una coloración más oscura que lashembras
· Sucoloresamarillo/café,conrayascontrastantesSonmásplanosensuregiónventral(abdomen)Tienen una coloración amarilla cerca de las
aletas
Observacióndelcomportamientodeapareamiento
1. Separarmachosyhembrasutilizandocomoreferencia lafigura1y las instruccionesdadasporlosprofesores.
2. JuntarlospecesseparadosenuntanquededoblefondocomoseesquematizaenlaFigura23. Observarelcomportamientodelospeces.¿Cómosecomportaelmacho?¿Enquémomentola
hembraliberaloshuevos,quequedanseparadosdelospadres.4. Anotarlasobservacionesmásrelevantes.

Figura2:Usodetanquededoblefondoparaapareamientodepeces
ParteII.Desarrolloembrionario
Observacióndeldesarrollotemprano:lasprimerasdivisionesdelosblastómeros
1. Luegodelapareamientodelospeces,sedeberecuperarloshuevosutilizandouncoladordeltamañoadecuado.Seguirlasinstruccionesdelosprofesores.
2. Unavezobtenidoloshuevos,estosdebenserpreparadosparasuobservaciónmicroscópicaenlupas.Seguirlasinstruccionesdelosprofesores.
3. Observarloshuevosenlalupaydibujarlascaracterísticasmásconspicuasdeestos.¿Haymovimientosenelovoplasma?¿Cuántascélulaspuedesdistinguir?
4. Losprofesoresprepararánunadelaslupasdemaneradesacarfotografíasaintervalosdeterminadosparaseguirloscambiosmorfológicosdeloshuevoseneltiempo.
5. Alfinaldeesteperíodoobservaremosdichoscambiosregistradosenlasucesióndeimágenes,sedibujaráloscambiosmásllamativos.¿Cuántasdivisionesselogróobservar?¿Cuántascélulaslograsdistinguir?
6. Discusióndelosresultadosobtenidos
Observacióndeldesarrollotemprano:gastrulación
La gastrulación corresponde a un proceso durante el cual se mueven los blastómeros,cambiando sus interaccionesy resultandoenel establecimientode las3 capasgerminalesqueconelpaso del tiempo darán origen a los distintos órganos del cuerpo. Estos movimientos dramáticosempiezancuandolavelocidaddedivisióndelosblastómerosdisminuye.
Figura3:Esquemadelagastrulaciónenembrionesdepezcebra
1. La clave para observar este dramático proceso, es obtener embriones en el estadio correcto.Estosseránprovistosporlosprofesores.

2. Conlosembrionesyaidentificados,seprocederáaobservarlosbajolalupa,talcomosehizoconlosembrionesmástempranos.Esnecesariodibujarlascaracterísticasmásllamativas.
3. Luego, se procederá a registrar la gastrulación tomando fotografías bajo la lupa de losembrionesporuntiempodeterminado.Losprofesoresrealizaránestaparte.
4. Transcurridoeltiempocorrespondiente,pasaremosaobservarlaseriedefotografíasobtenidas.¿Cómo semueven las células? ¿Pueden describir la forma en que ocurren losmovimientos?Dibujeesquemas
5. Discusióndelosresultadosobtenidos

ParteIII.Identificacióndeestructurasanatómicas:usodeanimalestransgénicos
Observacióndeestructurasanatómicas.
Unadelascaracterísticasdelpezcebraquelohatransformadoenunpotenteanimalmodelodeestudio es su transparencia, la que permite una sencilla observación de estructuras anatómicas yprocesosbiológicossinelusodesofisticadossistemasópticos.
Las larvas que observaremos tienen entre 1 y 5 días de desarrollo. En especial estas últimasnadan activamente, por lo cual necesitamos inmovilizarlas. Esto se logra agregando tricaina, unanestésicoalmediodecultivo.
Figura4:Anatomíadeunalarvadepezcebrade5díaspost-fertilización(5dpf)
1. Agregartricainaalmediodecultivo.Esperarhastaquelaslarvasdejendemoverse.2. Llevar las larvas a la lupa, basándose en la figura 4 y las instrucciones entregadas por los
profesoresidentificarlamayorcantidaddeestructurasanatómicasposibles.Dibujar.

Analizaranimalestransgénicosquemarcandiversasestructurasanatómicas
Utilizando lupas de fluorescencia (ubicadas en el Laboratorio de Biología del Desarrollo) sevisualizarálaexpresióndeproteínasfluorescentes(GFP,mCherry),quereportanlaexpresióndegenesespecíficos en distintas estructuras anatómicas de larvas de 3 dpf. Se cuentan con distintas líneastransgénicasquemarcandistintoscomponentescelularesenlalarva,comosemuestraenlafigura5.Elobjetivoesqueelestudianteseacapazdediscriminartiposdemarcasyaquéestructuracorresponde.
Figura5:ExpresióndeGFPenelsistemacirculatorioyneuronasenlarvasdepezcebra
Enestasecciónanalizaremosdostiposdetransgénicos:
1.NeuroD: factor de transcripción basic hélix loop hélix, gen pan neuronal de diferenciacióntempranadeneuronasdelsistemanerviosocentralyperiférico,ademásdeladiferenciacióndelpáncreas, expresándose en las células beta pancreáticas y enteroendocrinas. Se expresa eninterneuronasyneuronasmotorasdelamédulaespinal,elnerviodelalínealateralposterioryproyecciones anterior y dorsal, nervio óptico y cerebro anterior-medio-posterior, cerebro,ganglioscraneales,etc.
2.Fli1:(proto-oncogenFli1)permiteobservarlaangiogénesisdelpezdesdeestadiostempranos.Seobservaelmesodermolateraly lavasculaturacompleta, laaortaanteriorydorsal,elcorazón,lascélulasendoteliales,elendocardio,losvasossanguíneos,lavasculaturacranial,entreotros.
Yprocedemosconlaactividad:
1. Agregartricainaalmediodecultivo.Esperarhastaquelaslarvasdejendemoverse2. Llevar las larvas a la lupa de fluorescencia, y con la ayuda de los profesores observará
estructurasmarcadasconfluorescenciaenlaslarvas.Dibuje.3. Según loobservadoen la lupadefluorescencia,¿aquécreequecorresponden lasestructuras
queseencuentranmarcadasconfluorescenciaenlalarva?.Discutasobrecómolaobservacióndecomponentescelularesinvivopuedeserútilenelestudiodelabiologíadeldesarrollo.

ParteIV.Elpezcebraencomomodeloparaelestudiodelosefectostóxicosdelcigarrilloyelalcohol
Muchasdelascualidadesembrionariasdelpezcebra,talescomolafecundaciónyeldesarrolloexternos,lagrancantidaddeembrionesqueseobtienenporcadacruceylatransparenciadelostejidosdelpezdurantelasetapastempranas,asícomosufácilmantenciónydebajocosto,hanpromovidosuimplementacióncomomodeloenensayosdetoxicidadambientalydeevaluaciónderiesgosdesalud.Estosensayospermiten,porejemplo,analizarelaguaysuelodeunlugarenespecíficoenestudiosdeecotoxicidad,evaluar latoxicidadasociadaconproductosderivadosdelpetróleoyestudiar losefectosdealgunospesticidas.
El humo del cigarrillo contienemás de 4.000 compuestos conocidos, incluyendomás de 300agentescancerígenos.Elconsumoregulardelcigarro,estádirectamenteasociadoconeldesarrollodeenfermedades cardiorrespiratorias, tales como enfermedades coronarias, enfermedad pulmonarobstructiva crónica y cáncer de pulmón, así como riesgos de infertilidad y complicaciones durante elembarazo. Mientras que el abuso del consumo de alcohol se relaciona con problemas de hígado,páncreasyestómago,aligualqueretrasoyanomalíaseneldesarrollofetal.
En esta sección, embriones de pez cebra serán expuestas a distintas concentraciones deextractodehumodecigarro(EHC)oetanolylosefectosqueestoscompuestostengaseneldesarrolloseránevaluados.
Actividad
1. Preparacióndedilucionesdeextractodehumodecigarro
1.1. Instalar labombadevacíoy labotellaespecialcondossalidas (Figura6)siguiendo lasinstruccionesdelosprofesores.
1.2. Obtenerelextractodehumodeuntotaldetrescigarrospormediodelburbujeocreadoporlasuccióndelabomba.
1.3. Preparardilucionesdelextractoobtenidoenmediodecultivo,deacuerdoalasiguientetabla.
Dilución 0% 0.25% 0.5% 0.75% 1.0% 1.5% 2.0% 2.5%
EHC 0 12.5�l 25.0�l 37.5�l 50�l 75�l 100�l 125�l
Medio 5ml 5ml 5ml 5ml 5ml 5ml 5ml 5ml

Figura6:Equiponecesarioparalapreparacióndeextractoadehumodecigarro.Enlafigurase observa la botella especial con salidas, una con cigarrillo y la otra unida a la bomba devacío.

ObjetivoIdentificar alteraciones del desarrollo del pez cebra al ser incubado con alcohol.MaterialesPipetas plásticasMicroplacas de 6 pocillosReactivosSolución E3Soluciónes de Etanol al 0,5% v/v y 1,5% v/v en E3Inicio del ensayo: día 0
Temperar cada solución de exposición a 26°C±1°C antes de comenzar el ensayo.
Para comenzar la exposición tan pronto como sea posible, transferir inmediatamente cerca de 60 huevos a la solución control temperada. Posteriormente, transferir a las microplacas de 6 pocillos solo los huevos fecundados y sin ningún tipo de anomalía, de manera que cada pocillo contenga 10 huevos en 5 ml de solución, y existan 2 réplicas para cada condición, como lo muestra la siguiente imagen:
Figura 1
Control Negativo: Huevos en medio E3. Tratamiento 1: Huevos en Etanol 0,5% v/v. Tratamiento 2: Huevos en Etanol 1,5% v/v.
Cubrir las microplacas con su respectiva tapa e incubar a 26 °C ± 1°C durante 48 a 96 horas.

Registre la hora de inicio del ensayo.
Desarrollo del ensayo: días 1 a 4
Temperar un volumen de muestra adecuado al número de grupos de exposición utilizado (10 ml de E3 y 10 ml para cada condición).
Cada 24 horas luego del inicio de la exposición examinar cada uno de las sub-poblaciones y registrar las observaciones diarias según corresponda a cada día de ensayo.
Remover de cada pocillo a todos los individuos muertos y restos de corion.
Después de finalizar la observación y cuantificación de los parámetros correspondientes, diariamente realice una renovación de la solución de exposición de cada pocillo en todas las microplacas (tanto de control como las condiciones). Para ello debe ser reemplazada al menos 3⁄4 partes del volumen inicial con solución de exposición fresca y previamente temperada a 26°C±1°C.
Parámetros básicos de observación Número de individuos muertosAl día 1 un embrión se puede considerar muerto si se observan huevos coagulados, estos pueden ser claramente identificados, son opacos y parecen ser oscuros cuando se observan a través de un microscopio. A partir del día 2 sólo la total ausencia de latidos cardiacos es indicador inequívoco de la muerte de los embriones (ver figura 2).
Figura 2
Número de individuos eclosionados En los días 2, 3 y 4 del ensayo es necesario cuantificar el número de individuos que ha
eclosionado en cada grupo. Para ello, es vital que en estos días ésta sea la primera respuesta a evaluar. Cuidadosamente, abrir cada placa multipocillo y sin tocar los pocillos contar el número de individuos que se encuentran fuera del corión. Si luego de terminar el registro diario de esta respuesta, algún embrión eclosiona al ser manipulado para el registro de las respuestas siguientes, este no debe ser contado como eclosionado dicho día, sino al día siguiente. Se considera que un individuo ha eclosionado cuando se encuentra total o parcialmente fuera de su corion.
1 dpt 2 dpt 4 dpt
Condición Muertos Muertos Eclosionados Muertos Eclosionados
Control
Etanol 0,5%
Etanol 1,5%
Número de individuos con malformaciones generalesEn los días 2 y 4 cuantificar la cantidad de individuos que presenta cualquiera de las malformaciones según Tabla 1, sin tomar en cuenta la severidad de la respuesta, sólo su presencia o ausencia.
Malformación Día 2 Día 4
Microftalmia X X
Microcefalia X X
Curvatura corporal X
Aletas pectorales anormales X
Vesícula ótica anormal X X
Saco vitelino anormal X XTabla 1

Figura7:Estadosembrionariosylarvariosdepezcebra

TRABAJO PRACTICO UNIDAD 5

UNIDAD 5: EFECTOS DEL AMBIENTE SOBRE EL DESARROLLO ANIMAL
Objetivo 1. Efecto de la nutrición sobre el desarrollo del animal
SECCIÓN TEÓRICA
En el desarrollo de los seres vivos las condiciones ambientales y nutricionales son de gran importancia. Como se mencionó en la unidad 3, las larvas en estadío III, se alimentan hasta alcanzar la masa crítica. Si se produce una privación de nutrientes antes de que la larva alcanza este punto de control, el animal pausa su desarrollo, extendiéndose el periodo de desarrollo larval hasta reestablecerse la alimentación. En contraste, cuando la larva sufre de privación de nutrientes luego de haber alcanzado la masa crítica, el animal continúa su desarrollo normal. Por otra parte, existe una interesante relación entre el tiempo de privación de nutrientes durante el periodo larval y el tamaño final del adulto. El tamaño corporal aumenta drásticamente durante el periodo larval, contribuyendo con la mayor parte del tamaño de la mosca adulta, por tanto, la duración del periodo larval es un parámetro importante para la determinación del tamaño del adulto. La privación de nutrientes antes de adquirir la masa crítica extiende la duración de la etapa larval, sin embargo, no afecta el tamaño del adulto. En cambio, si la falta de nutrientes se produce después del punto de masa crítica sí se reduce el tamaño del adulto, debido a que no se extiende la duración del periodo larval. Esto sugiere que el estímulo de privación de nutrientes disminuye la tasa de crecimiento (incremento de la masa corporal en el tiempo) en el periodo larval, pero en el caso que la falta de nutrientes precede la masa crítica, la extensión del periodo larval puede compensar la disminución en la tasa de crecimiento para la obtención de un adulto de tamaño normal. Cabe destacar, que si la larva no ha alcanzado la masa crítica no continuará el desarrollo para la formación de la pupa. PARTE PRÁCTICA
• Observe las diferencias de tamaño de larvas en estadio III que han sido o no sometidas a condiciones de privación de nutrientes.
• Compare el tamaño de las pupas y adultos provenientes de larvas alimentadas normalmente y de larvas sometidas a privación de nutrientes luego de alcanzar la masa crítica.

Objetivo 2. Teratogénesis y Mutagénesis
SECCIÓN TEÓRICA.
La teratogénesis (del griego teratos, que significa monstruo), se refiere a malformaciones anatómicas, tanto macroscópicas como sutiles, incluyendo malformaciones físicas y deficiencias mentales o conductuales, así como también a retrasos en el desarrollo intrauterino, que ocurren como consecuencia de la exposición a un agente teratogénico durante el periodo gestacional. Estos agentes pueden ser de variada naturaleza, como lo son:
• Sustancias químicas como el alcohol, la nicotina, determinados tipos de hormonas, ciertos antibióticos (tetraciclinas), talidomida, entre otros.
• Agentes físicos como la radiación solar o la exposicón a rayos X. • Agentes infecciosos como virus o bacterias • Enfermedades maternas • Carencia nutricional de la madre
Cabe destacar que el efecto post-natal que pueda producir el agente teratogénico dependerá de varios factores, como el tipo de agente y exposición (etapa del desarrollo embrionario y tiempo de exposición al agente), siendo el periodo de 3-8 semanas de gestación el periodo más susceptible, dado que en el caso de los humanos es cuando está ocurriendo el proceso de organogénesis. Asimismo, son importantes características de la madre como hábitos y susceptibilidad a enfermedades. Dada la naturaleza variada de estos agentes, los mecanismos de acción también lo son, pero para ejemplificar, pensemos en la talidomida, que es un fármaco que se utilizó en el periodo comprendido entre los años 1958 y 1963 para el control de las náuseas que ocurren durante el primer periodo del embarazo, pero que presentaba efectos adversos terribles: tras su uso nacieron miles de niños con malformaciones en las extremidades, siendo éstas excesivamente cortas e incluso inexistentes.
Por su parte, la mutagénesis, se refiere al proceso en que la información genética de una célula es alterada de manera estable. Este proceso puede ocurrir espontáneamente en la naturaleza o bien, ser consecuencia de la exposición a agentes mutágenos. Las consecuencias de un proceso de mutagénesis pueden ser de variada naturaleza, desde ser neutras (sin efectos aparentes), a causar enfermedades como cáncer u otro tipo de enfermedades heredables, pero a su vez también representa una de las causas importantes que ha impulsado la evolución de las especies. Por su parte, el mecanismo por el cual aparece mutagénesis dependerá fuertemente del agente causal.
Una de las causas más comunes de mutagénesis endógena corresponde a fallas en la maquinaria de replicación del ADN, o por daños físicos en dicha molécula a causa de especies reactivas de oxígeno (generadas normalmente mediante procesos asociadas al metabolismo celular).
En cuanto a los agentes exógenos, éstos pueden ser de diversa naturaleza:

• Mutágenos químicos: compuestos capaces de alterar estructura del ADN como el ácido nitroso (agente desaminizante), brominas y derivados.
• Mutágenos físicos: radiaciones que pueden alterar la secuencia y estructura del ADN, como por ejemplo la radiación ultravioleta que pude generar dímeros de pirimidina (tipo de nucleótidos, bloques esenciales de la molécula de ADN), y la radiación gamma, que es ionizante. Por su parte se ha descrito que el ultrasonido, con una frecuencia de 400.000 vibraciones/segundo, es capaz de inducir mutaciones en Drosophila.
• Mutágenos biológicos: aquellos organismos o entidades que contienen ADN propio, capaces de alterar las secuencias del material genético de su hospedador, como por ejemplo bacterias, virus y transposones.
Cabe destacar que el evento de mutagénesis que ocurre en las células de un individuo puede ser transmitido a la descendencia o no, de acuerdo a las células que estén afectadas. Si las células que mutan son somáticas (cualquier parte del cuerpo exceptuando la línea germinal), entonces la mutación generada por el agente mutágeno no será heredable. Pero, si la mutación ocurre en células de la línea germinal, entonces ésta formará parte de la descendencia.
Las mutaciones somáticas podrían o no generar un efecto macroscópico evidente, que a su vez podría ser nocivo o neutro. En el caso desafortunado de que la mutación ocurra en genes llamados supresores de tumores, existe la posibilidad de que dicha célula adquiera características que la vuelvan más susceptible a que ante un nuevo evento de mutagénesis, se convierta en tumoral.
Las mutaciones que ocurran en la línea germinal en cambio, por lo general no presentan sintomatología en el individuo que sufre la mutación, sino que esta, en el caso de no ser neutra, generará un efecto en la descendencia, siendo el individuo completo afectado por dicha mutación. Ahora bien, existen mutaciones en determinados genes que pueden generar patologías graves, las cuales se encuentran en la base de datos de OMIM, del inglés Online Mendelian Inheritance in Man (http://www.omim.org/). Estas son enfermedades heredables de forma mendeliana, es decir que se puede predecir fácilmente su herencia pues se encuentran ligadas a un único gen (denominadas también como enfermedades monogénicas). Asimismo, esta mutación puede tener un carácter recesivo, dominante o ligado al cromosoma X, que se basa en el hecho de que todos los humanos tienen dos copias de cada gen, llamado alelo, uno proveniente de la madre y el otro del padre.
• Recesividad: Los dos alelos del mismo gen deben estar mutados para que ocurra la patología, por lo que tanto la madre como el padre deben ser portadores del gen mutado. Ejemplo de este tipo de enfermedad es la fibrosis quística que genera problemas graves en los pulmones.
• Dominancia: Basta con que uno de los alelos se encuentre mutado para que ocurra la patología, que puede provenir tanto de la madre como del padre. Un ejemplo de enfermedad autosómica dominante es el Huntington, que es una enfermedad

neurodegenerativa progresiva, que culmina afectando habilidades tan básicas como la capacidad de caminar, el habla y el razonamiento.
• Ligado al X: Dado que la determinación del sexo en humanos está determinado de la forma XX para la mujer y XY para el hombre. Si la mutación es dominante, cualquier individuo, sea hombre o mujer será afectado por la enfermedad. Si la mutación es recesiva, el hombre siempre se verá afectado (pues los hombres tienen un solo cromosoma X), mientras que la mujer se verá afectada en el caso de que sus dos cromosomas X estén afectados por la mutación. Ejemplo de enfermedad recesiva ligada al cromosoma X es la Hemofilia, caracterizada por problemas en factores de coagulación y por tanto generan procesos de sangrado excesivo y sangrados internos espontáneos.
Dado que las patologías monogénicas ocurren producto de la mutación en un único gen, es predecible que dicho gen cumple una función importante en los individuos, y por ende, generalmente se encuentran evolutivamente conservados. Es así que aproximadamente el 60% de los genes asociados a enfermedades monogénicas en humanos están codificados en el genoma de Drosophila, por lo que este modelo representa una plataforma útil para un estudio de dichas enfermedades que permita mejor comprensión lo que a su vez posibilita la generación de terapias específicas en el futuro.
Es evidente entonces que los agentes teratogénicos y mutagénicos pueden generar graves consecuencias durante el desarrollo de los individuos, por lo que es menester que existan herramientas de fácil uso que permitan determinar la presencia de dichos agentes en un determinado medio. Es así que se han generado especies de plantas y animales como biosensores ambientales, por ejemplo mediante el uso de promotores inducibles por la presencia de un determinado agente (Figura 1), de manera tal que de una manera dosis dependiente, el animal exprese una proteína reportera bajo el control de dicho promotor, como por ejemplo del uso de proteínas fluorescentes, que en la naturaleza no son expresadas por los animales, y la expresión ectópica puede ser fácilmente determinada mediante el uso de microscopios de fluorescencia.
Figura 1. Panel superior: En ausencia del factor ambiental de interés, ni el promotor inducible ni el gen reportero se expresan. Panel inferior: En presencia del factor ambiental, el promotor inducible

se enciende y activa la expresión del gen reportero, de manera tal que los individuos que porten dicho constructo podrán ser bioindicadores de la calidad del ambiente al que se expongan.
SECCIÓN PRÁCTICA
1. Ensayar efectos deletéreos de compuestos químicos sobre la viabilidad y morfología de las moscas.
1.1.Viabilidad
Existen compuestos químicos con una toxicidad tal que pueden afectar la sobrevida de los animales, tanto teratogénicos como mutagénicos. En este caso se evaluará el efecto del cobre, que en concentraciones altas genera un alto nivel de estrés oxidativo, que eventualmente daña las células hasta generar su muerte.
Por lo tanto, se entregará a los alumnos viales con animales que fueron previamente sometidos a concentraciones distintas de sulfato de cobre (CuSO4), y los alumnos deberán contar el número de animales que nace, y calcular entonces el porcentaje de sobrevida ante las distintas condiciones mencionadas.
1.2.Morfología
Se ha descrito que es posible fenocopiar (imitar el fenotipo, características visibles de un individuo producto de lo codificado en el genoma) los efectos de mutaciones en genes homeóticos de manera artificial con agentes químicos. Un ejemplo de ello es la exposición de moscas sin mutaciones a altas concentraciones de éter, lo que provoca alrededor de un 30% de animales con fenotipo de animales con mutaciones en bithorax (Figura 2)
Figura 2: Izquierda: Drosophila adulta normal tiene los segmentos apropiados, con un solo par de alas, y un par de halterios. . Derecha: Drosophila adulta mutante para el gel bithorax presenta un
cambio de identidad de segmento, lo que se traduce en el cambio del segmento T3 por T2, provocando que los animales reemplacen el par de halterios por un segundo par de alas.

Protocolo (la parte inicial 1-4 será realizada por los miembros del laboratorio)
1. Colectar embriones que tengan entre 2 y 3 horas de desarrollo. 2. Someter a un ambiente con éter saturado durante 10 minutos. 3. Dejar en ambiente normal y permitir que los animales se desarrollen normalmente. 4. Esperar que las moscas eclosionen. 5. Evaluar las moscas adultas y cuantificar el porcentaje de animales con fenotipo bithorax.
2. Utilizar animales transgénicos como biosensores ambientales.
Se entregará a los alumnos animales en estadío larvario que contienen una construcción reportera de altas temperaturas, de manera tal que si los animales son expuestos al calor, expresarán una proteína fluorescente verde (GFP), que se podrá observar al iluminar las larvas con un láser verde, pues las larvas que expresen GFP emitirán más luz que las que no lo expresen.
1. Tomar los viales conteniendo larvas y sumergir en agua a 37°C durante una hora. 2. Esperar 2-3 horas 3. Sacar las larvas de los tubos con espátula y ponerlas sobre una placa de plástico con
agua para lavarlas. 4. Agarrar suavemente las larvas con una pinza y traspasarlas a un pocillo conteniendo
agua limpia (este paso es importante pues la presencia de comida en el agua puede dificultar la observación de las larvas y dañar el experimento).
5. Observar las larvas a simple viste y comparar con animales que no fueron expuestos a las temperaturas altas como control.
6. Apagar las luces y apuntar las larvas con un láser verde y observar la tonalidad de las larvas.
7. Registrar observaciones.