facultad de medicina centro universitario de ...digeset.ucol.mx/tesis_posgrado/pdf/jorge ramon calvo...
TRANSCRIPT

FACULTAD DE MEDICINA Centro Universitario de Investigaciones Biomédicas
CORRIENTES DE CALCIO EN VESICULAS FORMADAS A PARTIR DE LA MEMBRANA PLASMATICA DEL
MUSCULO ESQUELETICO DEL ANFIBIO
Que para obtener el grado de:
MAESTRO EN CIENCIAS MEDICAS
P r e s e n t a :
Biol. Jorge Ramón Calvo Martínez
Asesores: Dr. Jorge Alberto Sánchez Rodríguez
Dr. Miguel Huerta Viera Dra. Maria del Carmen García García
Colima, Colima., Marzo de1 2003

INDICE
RESUMEN…………………………………………..……..…………………………………...…………….1
ABSTRACT…………….……………………….……………………………………………………………2
INTRODUCCIÓN.................................................................................................……........……...….3
Los canales iónicos de calcio ........................................................………………….................................…5
La subunidad αl de1 canal de calcio ...............................................................…………………....................7
El papel fisiológico de la corriente iónica de calcio ........................................................…………………..8
Estudios electrofisiológicos ..................................................................................................…………………9
Modelos para estudiar directamente a 10s canales iónicos ...............................................………………….11
Vesículas derivadas de1 músculo esquelético ......................................................................……………12
ANTECEDENTES DIRECTOS.................................................................................…………….…14
La corriente iónica de calcio .......................................................................................………………..........14
Características electrofisiológicas de las corrientes de bario ...............................................………………14
Corrientes de cola .................................................................................................…………………………..17
Inactivación de1 canal de calcio .............................................................……………………………………18
El receptor a la dihidropiridinas .............................................................................…………………………18
Movimiento de carga ............................................................................................…………………………..20
PLANTEAMIENTO DEL PROBLEMA ........................................................................……………25
Problema ......………………………………………………………………………………………………...25
objetivos de1 presente trabajo ...................................................................................………………………25
METODOLOGÍA .............................................................................................................…………27
Obtención de vesículas...............................................................................................……………………27
Soluciones .....………………………………………………………………………………………………..27
Registro electrofisiológico ..............................................................................................…………………...28
Protocolo de pulsos y captura de datos .............................................................................…………………..30
Diseño experimental ............................................................................................................………………...30
Registro de las corrientes de bario ................................................….................................…………………30
Corrientes de cola.. ..………………………………………………………………………………………..31
Inactivación .............…...…………………………………………………………………………………….31
Movimiento de carga ………………………………………………………………………………………31
RESULTADOS ..............................................................................................................……………33
Controles …………………………………………………………………………………………………………33
Corrientes de bario........................………………….............................................................................................33
Corrientes de cola .......................……………………………………………............................................................... 39
Inactivación....................................……………………………………………………………………………..41
El receptor a las dihidropiridinas ..............................................………………………………………………44
Movimiento de carga en las vesículas ....................................……………………………………….....................48

Carga 2..........…………………………………………………………………………………………….53
CONCLUSIONES .............................................................................................................55
Corrientes de bario ......................................................................................................................………..55
Corrientes de cola .......................................................................................................................………...58
Inactivación ...……………………………………………………………………………………………58
El receptor a las dihidropiridinas ..............................................................................................…………58
Movimiento de carga 1................................................................................................................………...58
Movimiento de carga 2................................................................................................................………60
BIBLIOGRAFIA ................................................................................................................61
TABLA DE FIGURAS …………..........................................................................................34
Figura 1.........……………………………………………………………………………………………34
Figura 2.........……………………………………………………………………………………………37
Figura 3.........……………………………………………………………………………………………40
Figura 4 ..………………………………………………………………………………………………..42
Figura 5.........……………………………………………………………………………………………45
Figura 6.........……………………………………………………………………………………………47
Figura 7 49
TABLA DE GRAFICAS ……………………………………………………………………35
Grafica 1………………………………………………………………………………………………...35
Grafica 2....................................................................................................……………………………...38
Grafica 3......……………………………………………………………………………………………..43
Grafica 4.......……………………………………………………………………………………………46
Grafica 5…………………………………………………………………………………………………51
Grafica 6…………………………………………………………………………………………………52
Grafica 7…………………………………………………………………………………………………54

1
ABSTRACT
We here describe L-type Ca channel currents recorded in
vesicles derived from skeletal muscle plasma membranes with theWhole-cell voltage clamp technique, The time course of activationwas voltage and time dependent. The inward current was blockedby Cobalt and it was sensitive to Bay K. The main differences ofthese current with those present in cells are the follows: Activationis ten times faster and voltage dependent inactivation is morepronunced. Main differences in charge movements were: Highvalue of Qmax and no indication of QY component. Likewise, therewas no evidence of non linear charge movement in depolarizedvesicles. No major facilitation of tail currents was observed bypulses of increasing duration.
This work shows recording of channels present in thetransverse tubular membrane system in a preparation where it ispossible to control the composition of the external and internalmedia. These features make the vesicles a good preparation to testthe role of channels subunits and other components like ryanodinereceptors under controlled conditions.

2
RESUMEN
Se registraron corrientes iónicas a través de canales de calciotipo L en vesículas obtenidas de la membrana plasmática delmúsculo esquelético, utilizando la técnica de Patch Clamp. Lacorriente mostró una cinética dependiente del tiempo y del voltaje,fue bloqueada por el cobalto y potenciada por el Bay K. Lasprincipales diferencias con las fibras musculares son: unaactivation diez veces mas rápida y un proceso de inactivationdependiente del voltaje mas pronunciado. Las diferencias en elmovimiento de carga fueron: un valor mayor de Qmax y la ausenciadel componente Qy. Las vesículas despolarizadas no mostraroncarga 2 y mostraron solamente una pequeña facilitation mantenidapor el componente lento de la corriente de cola.
Este trabajo muestra el registro directo de canales presentesen el sistema de túbulos transversos, donde fue posible controlar lacomposition iónica extra e intravesicular. Estas característicashacen de las vesículas un modelo adecuado para evaluar laparticipación de las subunidades del DHPR o bien su interaccióncon otros componentes.

3
Abbreviations: + + Convincing electrical and pharmacological evidence + Some evidence K, IR Delayed rectifier and inward rectifier K channels (a) The Na channel of tunicate eggs binds Leirus scorpion toxin, which modifies its inactivation gating, but the channel is insensitive to 15 µM TTX. (b) Cnidarian axons and ctenophore smooth muscles have brief Na-dependent action potentials that are insensitive to 10 µM TTX References: 1Ccrschenfeld (1973), 2Hagiwara and Kidokoro (l978), .3Ohmori (1978), Ohmori and Yoshii (IY77), 4Hagiwara and Takahashi (1974a). Hagiwara (11)X3), 5Schwartz and Stühmer (1984), 6Byerly and Masuda (1979), 7Koopowitz (1989), 8Dubas et al. (IY8H). 9Anderson and Schwab (1982), Anderson (1989b), 10Eckert and Brehm (1979). Naitoh (1982), Deitmer (198Y), Wood (1989), 11Gustin et al. (19%), Caldwell, Van Brunt and Harold (1986), 12Tester (199U), 13Hedrich and Schroeder (1989).
De entre estos dos canales se ha llegado a pensar que los canales de calcio tienen un carácter más general (Glossmann, Lubbecke, Mewes y Hofmann, 1982, Tsien y Tsien, 1990) ya que los iones de calcio pueden regular procesos celulares tan diversos coma: La contracción, la secreci6n de neurotransmisores, la fecundación, la proliferaci6n celular, la intercomunicaci6n en neuronas y muchas funciones más (Berridge, 1997). El principal mecanismo por el cual opera este ion es por incrementar brevemente su concentraci6n Citoplasmática en regiones muy especificas, alcanzando valores del orden micromolar (Berridge, 1997). Esta alta concentración contrasta con la condici6n en reposo, la cual se mantiene dentro de un estrecho rango, entre 20 y 100 X 10 -9 M (Trump y Berezesky, 1995).

4
CORRIENTES DE CALCIO EN VESICULAS FORMADAS A PARTIR
DE LA MEMBRANA PLASMATICA DEL MUSCULO ESQUELETICO
DEL ANFIBIO.
INTRODUCCIÓN
Los canales iónicos representan a una variedad de proteínas presentes en
las membranas biológicas, los cuales dependiendo de la especialización celular
desempeñan muy diversas funciones. En las células excitables son responsables
de los cambios en el potencial eléctrico transmembranal en respuesta a un
estimulo.
La activación de un canal iónico abre un poro selectivo en la membrana de
las células excitables permitiendo la difusión de cationes o aniones de acuerdo a
su gradiente electroquímico. En la mayoría de los cases la activation de un canal
cationico resulta en la formación de una corriente entrante de iones de sodio o de
calcio, causando la disminución del potencial de reposo. Mientras que la
activation de los canales de potasio permite la salida de estos iones con la
consecuente repolarización o hiperpolarización de la célula.
Los canales de sodio y de calcio comparten ciertas semejanzas
electrofisiológicas es decir tienen compuertas dependientes del voltaje, muestran
alguna forma de inactivation con la despolarización sostenida, tienen una
selectividad iónica moderada, un poro con un radio pequeño y son bloqueados
por compuestos cuaternarios hidrofóbicos (Hille, 1992), además sus secuencias
de aminoácidos son similares resaltando así su estrecha cercanía evolutiva
(Goldin, 1995 y Stea, Soong y Snutch, 1995). Otra característica de estos canales
es su amplia distribución en la naturaleza ya que se encuentran presentes en un
gran numero de células desde protozoarios hasta células de mamíferos (ver el
esquema a continuación; Tsien, Ellinor y Horne, 1991 y Hille, 1992).

5
Uno de los principales mecanismos por el cual se logra incrementar la
concentración del calcio intracelular es a través de los canales iónicos de calcio,
de tal forma que la corriente iónica desempeña un importante papel en la
fisiología de las células, sin embargo en algunos tipos celulares, coma es el
músculo esquelético las corrientes iónicas de los canales de calcio no se han
relacionado a una función especifica.
En la presente tesis se caracterizo la corriente iónica de los canales de
calcio mediante la técnica de célula intacta utilizando vesículas del músculo
esquelético coma un modelo nuevo que hizo posible el registro directo.
En la exposición de este trabajo primero se hablara sobre las
características de los canales iónicos y sus corrientes, posteriormente se
abordaran los procedimientos empleados en el estudio de estos sistemas y los
problemas que se han observado en preparaciones coma el músculo esquelético.
Finalmente se hará una breve descripción de las vesículas.
En los antecedentes directos se revisa específicamente los parámetros
electrofisiológicos que delimitan al presente trabajo en el estudio de los canales
de calcio en su papel tanto de canales iónicos como en su participación de sensor
al voltaje en el proceso de la contracción; Cubriendo aspectos coma son: la curva
corriente voltaje, la cinética de activación de la corriente iónica y su inactivation
dependiente del voltaje también se abordan otros aspectos relacionados con los
canales de calcio como son: El proceso de la potenciación observado en las
corrientes de cola, así como la sensibilidad a agonistas y antagonistas de las
dihidropiridinas y una evaluación del movimiento de carga y su inmovilización por
el voltaje ( carga 1 y carga 2 respectivamente).
Los canales iónicos de calcio
Los canales iónicos por los cuales el calcio puede cruzar la membrana
celular, se agrupan en tres grandes categorías; los dependientes del voltaje, los
controlados por un receptor químico y los presentes en fuentes de
almacenamiento (Burnashev, 1998; Randall, 1998).
Los canales de calcio dependientes del voltaje se clasifican a su vez en los
tipo T, L, N, P y Q de acuerdo a sus características electrofisiológicas y
farmacológicas (Zhang, et al. 1993).
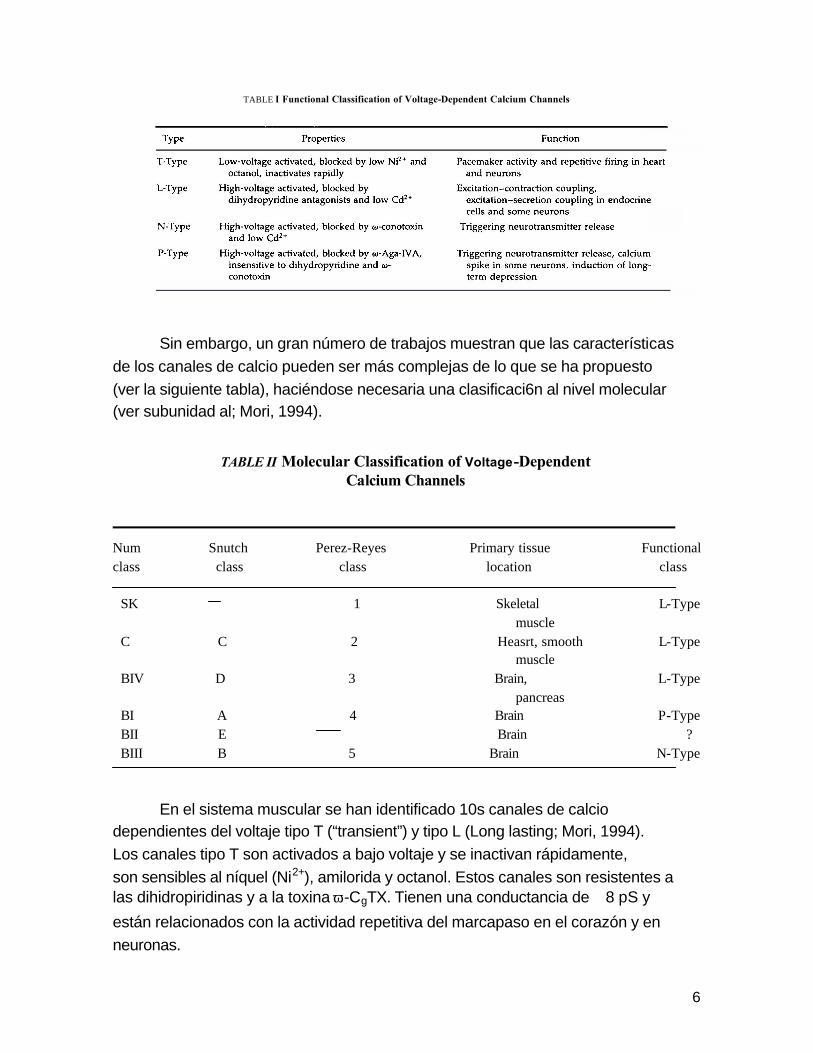
6
TABLE I Functional Classification of Voltage-Dependent Calcium Channels
Sin embargo, un gran número de trabajos muestran que las características
de los canales de calcio pueden ser más complejas de lo que se ha propuesto
(ver la siguiente tabla), haciéndose necesaria una clasificaci6n al nivel molecular (ver subunidad al; Mori, 1994).
TABLE II Molecular Classification of Voltage-Dependent Calcium Channels
Num Snutch Perez-Reyes Primary tissue Functional class class class location class SK 1 Skeletal L-Type muscle C C 2 Heasrt, smooth L-Type muscle BIV D 3 Brain, L-Type pancreas BI A 4 Brain P-Type BII E Brain ? BIII B 5 Brain N-Type
En el sistema muscular se han identificado 10s canales de calcio dependientes del voltaje tipo T (“transient”) y tipo L (Long lasting; Mori, 1994).
Los canales tipo T son activados a bajo voltaje y se inactivan rápidamente,
son sensibles al níquel (Ni2+), amilorida y octanol. Estos canales son resistentes a las dihidropiridinas y a la toxina ω-CgTX. Tienen una conductancia de �8 pS y
están relacionados con la actividad repetitiva del marcapaso en el corazón y en
neuronas.

7
Los canales tipo L son activados a altos voltajes, son sensibles a las
dihidropiridinas y muestran una conductancia unitaria de 22-27 pS. Estos canales
se encuentran en prácticamente todos los tejidos excitables y en muchas células
no excitables, participan en el proceso del acople entre la excitación y la
contracción del músculo esquelético, del músculo cardíaco y del músculo liso, así
coma en la liberación del neurotransmisor de células endocrinas y de neuronas.
Los canales de calcio activados por el voltaje e incluso los de sodio y de
potasio son proteínas oligoméricas formadas por varias subunidades. De entre
estas se distingue una subunidad coma la principal ya que por si sola puede
formar un canal funcional; contiene al poro acuoso del canal y al sensor del
voltaje. En los canales de sodio y de potasio esta subunidad se representa por “α”
y en los canales de calcio por “α,” (Cambell, Leung y Sharp, 1988; Catterall,
Seagar y Takahashi, 1988 y Glossmann, 1990). Las subunidades restantes se
consideran auxiliares y tienen la capacidad de modular al canal (Isom, De Jong y
Catterall, 1994 y Adelman, 1995)
La subunidad α1 del canal de calcio
La diversidad molecular de la subunidad α1 ayuda a explicar la gran
variedad de canales de calcio. Así, las subunidades α1C y α1D dan lugar a canales
tipo L, la subunidad α1Bforma a los canales tipo N, los canales P, Q y R cuentan
con subunidades α1A (Hofmann, Lacinová y Klugbauer, 1999)
Con base a la semejanza en la secuencia de aminoácidos que tienen las
diferentes subunidades al en relación con la α1s, del músculo esquelético, se
reconocen dos grandes grupos. El primero es un grupo sensible a las
dihidropiridinas e incluye a las subunidades α1s, α1c y α1D. El Segundo grupo tiene
características electrofisiológicas y farmacológicas mas diversas e incluye las
subunidades α1A, α1B y α1E (Snutch, Leonard, Gilbert, Lester y Davidson, 1990;
Soong, Stea, Hodson, Dubel, Vincent y Snutch, 1993).
La subunidad al contiene a los receptores a las dihidropiridinas (en
adelante DHPR) y se le ha atribuido una doble función, la de comportarse coma
un canal de calcio y la de ser el sensor al voltaje, el cual interviene en el proceso
del acople entre la excitación y la contracción (Beam y Knudson, 1988)..

8
La subunidad α1s del canal de calcio del músculo esquelético fue la primera
subunidad a la que se le determine su estructura (Tanabe et al. 1987; Hosey et al.
1987 y Cambell et al. 1988). La expresión del cDNA de la subunidad al, del
músculo cardiaco en Ovocitos de Xenopus ha resultado en la formación de
canales funcionales, sin embargo la subunidad α1s, del músculo esquelético no ha
dado el mismo resultado y solo se ha expresado funcionalmente en miotubos de
músculos disgenicos de ratón (Tanabe, Powell y Numa, 1988).
Los DHPR del músculo esquelético y del músculo cardiaco permiten la
formación de una corriente entrante de calcio, sin embargo el papel fisiológico de
estas dos corrientes es muy diferente (Adams, Tanabe, Mikami, Numa y Beam,
1990).
El papel fisiológico de la corriente iónica de calcio.
El papel fisiológico que desempeñan tanto los DHPR coma la corriente
iónica que a través de ellos se genera, depende fundamentalmente del tipo de
célula en cuestión, así por ejemplo en el músculo cardiaco la entrada de calcio
por los DHPR es necesaria en la transmisión de la señal para la liberación de
calcio del retícula sarcoplásmico (Tanabe, Numa y Beam 1990), esta propiedad
ha permitido utilizar diversos fármacos que tienen la capacidad de alterar las
características de esta corriente de calcio, coma agentes terapéuticos para el
tratamiento de ciertas enfermedades cardiovasculares coma la hipertensión, la
taquicardia o la angina de pecho.
El músculo esquelético también posee este tipo de canal iónico, pero su
corriente de calcio no lleva a cabo la misma función que en el músculo cardiaco,
de hecho esta corriente no tiene una función plenamente identificada (Tanabe,
Adams, Niidome y Numa, 1990, Lamb, 1992, Lu y Meissner, 1994). La
incertidumbre sobre el papel que desempeña la corriente iónica de calcio en el
músculo esquelético es paradójica, puesto que esta preparation se considera
clásica para el estudio del papel del calcio en la contracción.
Parte de la dificultad para aclarar la función de la corriente de calcio es la
falta de un modelo biológico donde se tengan a 10s canales de calcio en la
superficie de tal forma que permita hacer estudios directos sobre el canal.

9
Estudios electrofisiológicos
Tradicionalmente los canales iónicos han sido estudiados a través de sus
corrientes iónicas, utilizando para ello diversos métodos electrofisiológicos, en los
cuales ha sido de suma importancia seleccionar adecuadamente tanto al modelo
biológico coma al sistema de registro y análisis de estas señales. Sin embargo
esta selección no siempre se ha completado satisfactoriamente, tal es el case de
los estudios realizados en el músculo esquelético, donde a la fecha no se cuenta
con un modelo que permita estudiar adecuadamente a las corrientes de calcio.
Las propiedades biofísicas de los canales pueden ser determinadas
rápidamente si los canales ocurren en alta densidad sobre una gran superficie
donde pueda ser fijado el potencial de membrana, sin embargo los canales de
calcio del músculo esquelético se localizan en lugares de difícil acceso coma el
sistema de túbulos transversos (Peachy, 1965).
Las técnicas de fijación de voltaje que se han usado tradicionalmente no
son capaces de lograr una buena fijación de voltaje en el sistema de túbulos
transversos (Jack, Noble y Tsien, 1983), Heiny y Vergara utilizando colorantes

10
sensibles al voltaje fueron capaces de discriminar entre el potencial al cual se
encontraba el sarcolema y el presente en el sistema de túbulos transversos
después de la aplicación de un pulse cuadrado en la superficie de la fibra.
Observaron que el potencial en la superficie era muy similar al pulse cuadrado
mientras que en el sistema de túbulos transversos el voltaje cambiaba muy
lentamente, esto se debe principalmente a la gran cantidad de membrana
contenida en este sistema, aproximadamente el 85 % del total de membrana
plasmática de la fibra muscular (Heiny y Vergara, 1984 y Vergara y Bezanilla,
1881).
Un Segundo problema que enfrentan las corrientes de calcio es su pequeña
magnitud por lo que son fácilmente enmascaradas por la participación de otros
canales, especialmente por los canales de potasio. Este inconveniente es
superado finalmente con métodos farmacológicos que permiten la separación de
las corrientes ya sea usando bloqueadores de canales i6nicos (ver la tabla a
continuación) o mediante la sustitución de iones; teniendo presente que la
sustitución extracelular de los iones de calcio por algún otro catión puede alterar
las características cinéticas de estos canales. (McDonald, Pelzer, Trautwein y
Pelzer, 1994).
TABLE 1. REVERSIBLE BLOCKING AGENTS FOR DIFFERENT CHANNELS

11
Pese a estas dificultades en la década de los setenta y ochenta se
realizaron un gran numero de trabajos de fijación de voltaje, obteniendo una Clara
imagen del canal de calcio.
Las corrientes de calcio fueron registradas a partir de pequeños ases de
fibras de tejido cardíaco o de músculo liso usando las técnicas de uno o dos sellos
de sacarosa (Attwell y Cohen, 1977; Beeler y Reuter, 1970 y Bolton, 1979).
También la disección de fibras únicas de músculos de anfibio o de mamífero
permitió usar satisfactoriamente las técnicas de dos o tres microelectrodos
(Adrian, Chandler y Hodgkin, 1970; Gamboa-Aldeco, Huerta y Stefani 1988 y
Stanfield, 1977) y la técnica del sello de vaselina (Almers, Fink y Palade, 1981 y
Mejia-Alvarez, Fill y Stefani, 1991), pero estas técnicas aún se ven limitadas para
fijar el voltaje (García, McKinley, Appel y Stefani, 1992), además los registros se
vieron afectadas por los procesos de diálisis o de lavado del interior celular, causa
del conocido “rundown” (Francini, Pizza y Traina, 1992).
Modelos para estudiar directamente a los canales iónicos
La utilización de métodos muy precisos coma es la técnica de registro de
canales unitarios o “Patch-Clamp” o bien la incorporación de canales en
membranas artificiales permitió acercarse al estudio directo de los canales, sin
embargo estas preparaciones también mostraron ciertas restricciones.
El disponer de células únicas disociadas de tejido muscular de fetos o de
roedores neonatos permitió usar la técnica de “Whole cell” (Beam y Knudson,
1988 y Cognard, Lazdunski y Romey, 1986) y aunque el tratamiento enzimático
parece no afectar a los canales de calcio (Burnashev, Edwards y Verkhratsky,
1990 y lsenberg y Klockner, 1980) las células en cultivo pueden sufrir
reordenación celular y con ello alterar la actividad de los canales de calcio
(Bechem y Pott, L. 1985 y Yuan, Goldman, Tod, Rubin y Blaustein, 1993).
La incorporación de canales de la membrana del sistema de túbulos
transversos del músculo esquelético en bicapas ha sido uno de los métodos que
mejor aceptación ha tenido (Flockerzi, Oeken, Hofmann, Pelzer, Cavalié y
Trautwein, 1986; Pelzer, et al. 1989; Smith, et al. 1987) debido a que permite
controlar adecuadamente el voltaje y es posible determinar la composition iónica
tanto de las soluciones que representan al medio intracelular coma el medio

12
extracelular (Wonderlin, Finkel y French, 1990). En estas condiciones registros de
“Patch clamp” muestran una conductancia de 20 pS en soluciones simétricas de
90 mM de Ba2+ y los canales conservan su sensibilidad al D-600. Las corrientes
se incrementan por el BAY K 8644 (ver mas adelante, el receptor a la
dihidropiridinas) y el PKA pero esto solo ocurre ocasionalmente (Pelzer, et al.
1989; Affolter y Coronado, 1985 y Ma, Mundiña-Weilenmann, Hosey y Ríos,
1991). También es común observar diferentes valores de conductancia, entre 7 y
15 pS (Hymel, Striessinig, Glossmann y Schindler, 1988; Talvenheimo, Worley y
Nelson, 1987 y Vaghy, et al. 1987).
Por otra parte los DHPR incorporados en bicapas lipídicas no muestran
inactivation (Affolter y Coronado, 1985 y Valdivia y Coronado, 1990). Por lo
anterior se ha sugerido que el fraccionamiento de membranas puede alterar la
estructura de los canales, resultando en la remoción del mecanismo de
inactivation (Mejia-Alvarez, et al. 1991).
Vesículas derivadas del músculo esquelético.
Un modelo que se vislumbra coma una alternativa para el estudio de los
DHPR es el descrito por Standen y colaboradores (1984) donde el tratamiento del
músculo esquelético con colagenasa y la incubación en una solución de alto
potasio resulta en la formación de vesículas.
Estudios de nuestro grupo muestran que las vesículas conservan su
orientación; el interior de la vesícula corresponde al interior celular y pese a que
no se ha determinado el origen de la membrana que forma a la vesícula, se ha
sugerido una naturaleza tubular (Camacho, Vázquez, Arguello y Sánchez, 1996).
Dada su característica vesicular es posible formular anticipadamente la
composition extravesicular (baño) e intravesicular (micropipeta), también es
posible la fijación de voltaje mediante la técnica de “Whole-cell” lo cual ha hecho
posible registrar corrientes iónicas microscópicas coma es la corriente de potasio
tipo A (Camacho, et al. 1996).
Las ventajas operativas del uso de las vesículas, pueden hacer de estas,
un modelo adecuado para la caracterización directa de las corrientes de los
DHPR, de acuerdo a los protocolos tradicionales coma son: Empleo de
herramientas farmacológicas y técnicas de registro electrofisiológico.

13
El análisis detallado de las corrientes iónicas de membrana puede
evidenciar la participación de mas de un canal iónico por lo tanto para estudiar la
cinética de un tipo especifico de canal es necesario hacer ciertas manipulaciones
experimentales coma sustituir los demás iones permeables por sus equivalentes
impermeables de tal manera que se cancele su participación en el registro. Otra
alternativa es el empleo de agentes farmacológicos que bloqueen al canal, una
practica frecuente desde los años 60, donde se utilizan drogas o toxinas que al
actuar a concentraciones muy bajas no presentan efectos tóxicos y su efecto solo
se explica por una acción directa sobre sitios específicos.

14
ANTECEDENTES DIRECTOS
La corriente iónica de calcio.
En las fibras musculares esqueléticas de los vertebrados se observan
potenciales de acción similares a los descritos en el axón gigante del calamar.
Donde una breve despolarización activa una corriente entrante de sodio rápida y
posteriormente una corriente saliente de potasio (Hille, 1992), además en el
músculo esquelético cuando la despolarización se prolonga varios cientos de
milisegundos se activa una corriente lenta de calcio. Estudios bajo fijación de
voltaje muestran que estas corrientes son más pequeñas que las corrientes de
sodio (0.1 mA/cm2 y 2.5 mA/cm2 respectivamente; Arreola, Calvo, García y Sánchez,
1987). Cuando la despolarización se mantiene durante algunos segundos estas
corrientes son suprimidas. Los bloqueadores de canales de calcio del músculo
cardiaco coma son la nifedipina o el D-600 también la bloquean (McDonald et al.,
1994).
Características electrofisiológicas de las corrientes de bario.
En términos biofísicos los canales iónicos suelen ser representados por
diagramas de circuitos eléctricos simples, donde la membrana se representa
coma un conductor en serie a la fuerza electromotriz (E) causada por la diferencia
en la concentración de un ion permeable (Eeq=RT/zFIn C1/P2 Donde R es la
constante universal de los gases, T la temperatura absoluta, z la valencia, F la
constante de Faraday y C1 y C2 la concentración del ion. De acuerdo a la ley de
Ohm se establece una relación entre la corriente y el voltaje denominada curva
corriente voltaje (I-E) la cual es descrita por la ecuación I=g(E-Eeq) donde la
pendiente (g) indica la magnitud de la conductancia, en términos generales es una
estimación del numero de canales abiertos. Cuando la probabilidad de apertura
de un canal depende del voltaje entonces la conductancia cambia con el voltaje
resultando así en una relación no lineal.

15
Las propiedades de las corrientes iónicas son frecuentemente resumidas
en términos de esta relación corriente voltaje, por ejemplo en una neurona de
caracol las despolarizaciones a -20 ó -10 mV abren una pequeña fracción de
canales y cuando la despolarización es a +20 ó, +30 mV se abren la mayoría de
los canales iónicos resultando en una corriente iónica máxima (ver la figura a
continuación). En despolarizaciones mayores pese a que la mayoría de los
canales están abiertos la corriente es menor o incluso puede no haber corriente,
esto es debido a que la fuerza impulsora es menor (E=Eeq). El potencial en el cual
la corriente es cero se denomina el potencial de inversión, ya que posterior a
este punto la corriente puede fluir en la dirección contraria a su gradiente químico.
De esta forma la manipulación del voltaje permite analizar las propiedades
cinéticas de los canales que dependen del voltaje y que participan en la corriente
de membrana. El curso temporal de dicho cambio es característico a cada tipo de
canal.
Las fibras musculares esqueléticas del anfibio presentan corrientes de
calcio que se activan cuando se aplica un pulso de 500 mseg a un potencial de

16
membrana de 10 mV. La activation de estos canales es 500 veces mas lenta que
los canales de sodio, Aunque esta tasa puede variar entre las diferentes fibras
musculares. Las fibras musculares de los vertebrados poseen diferentes tasas de
activation debido a las diversas clases de canales de calcio. La activación de las
corrientes tipo L es descrita por la distribución Boltzmann. El grafico de esta
función describe una curva sigmoidea en el rango de 0 a 1 para los voltajes de
-40 a +10 mV, haciéndose más rápida con voltajes mas positivos (Cota y Stefani,
1986 y Sánchez y Stefani, 1983).
El tiempo que se mantiene la despolarización también es importante, ya
que la conductancia de la membrana no es constante. Una vez que son activados
los canales iónicos estos comenzaran a cerrarse siguiendo un proceso
característico a cada canal (ver inactivation del canal de calcio). De esta forma
para un mismo potencial la corriente de calcio inicialmente aumentara pero casi
inmediatamente también ira disminuyendo. Por lo tanto la corriente de calcio
alcanzara una amplitud máxima denominada corriente al pica.
Los canales de calcio muestran diferente permeabilidad a los cationes
divalentes, por ejemplo los iones mas permeables son el Ba++ y el Sr++ siendo los
iones de Ba++ los que dan las corrientes iónicas mas grandes, razón por la cual
este ion es frecuentemente usado en estudios biofísicos.
TABLE 4. IONIC PERMEABILITY RATIOS, Px PCa FOR L-TYPE Ca CHANNELS
Calculated from reversal potentials in cardiac ventricular cells of adult guinea pigs using Equation 13-11 and 10 mM extracellular divalent ion concentrations (Hess et al., 1986; Tsien et al., 1987).

17
Corrientes de cola
Los canales de calcio tipo L se activan con la despolarización y forman una
corriente iónica cuya magnitud esta determinada por la diferencia de potencial
entre el pulse de prueba y el gradiente electroquímico del calcio (E-Eeq). En el
case de los canales de sodio del axón gigante del calamar, el mayor numero de
canales abiertos durante el pulso de despolarización se alcanza en
aproximadamente 800 microsegundos, pero si el axón se repolariza a los 700
microsegundos, cuando la mayoría de los canales están-r abiertos, aparece una
gran pero breve corriente de sodio denominada corriente de cola, la cual
disminuye rápidamente conforme se cierran los canales.
En algunos sistemas el curso temporal de las corrientes de cola ha sido
ajustado a una exponencial simple, y de acuerdo al modelo clásico de Hodgkin y
Huxley (1952) predice que la caída de la corriente de cola representa el proceso
de cierre de los canales que fueron activados. Cuando la caída de la corriente de
cola presenta mas de una constante de tiempo, indica la participación de otros
procesos, coma la inactivation del canal o bien revela la participación de mas
canales iónicos. De esta forma el estudio de las corrientes de cola constituye un
procedimiento adecuado para estudiar diverso aspectos del comportamiento de
los canales iónicos.
Fleig y Penner (1995) observaron que la corriente de cola de miotubos del
músculo esquelético de mamífero puede ser modulada por el voltaje, un efecto
potenciador similar al descrito por Fenwick y colaboradores (1982) en las células
cromafin de bovino. Sin embargo en el músculo esquelético esta potenciación
únicamente se observa en las corrientes de cola y no en las corrientes lentas de
calcio tipo L, coma si se tratase de dos poblaciones distintas de canales de calcio
(Fleig y Penner, 1996) o bien que el efecto potenciador resulta de algún tipo de
interacción dependiente del voltaje con otros elementos, coma puede ser el
receptor a las rianodinas (Fleig y Penner, 1996).
Se ha sugerido que los DHPR pueden funcionar coma canales de calcio
activados por el voltaje y además pueden actuar coma el sensor al voltaje en la
liberación de calcio del retículo sarcoplasmico (Ríos y Brum, 1987). A partir de la
discrepancia observada en la proporción de canales de calcio y el numero de

18
sitios a los que se unen las dihidropiridinas (Lamb y Walsh, 1987), parece sugerir
que solo una pequeña proporción de DHPR pueda actuar coma canales iónicos y
la mayoría actuarían entonces como el sensor al voltaje.
Sin embargo en el músculo esquelético del adulto la falta de un modelo
adecuado hace imposible establecer una conclusión Clara sobre si los de DHPR
actúan coma un canal de calcio silente el cual es activado por la despolarización o
bien si la potenciación observada en miotubos es el resultado de la interacción
con otros elementos.
lnactivación del canal de calcio
Existe una gran controversia acerca de la existencia de un proceso de
inactivación dependiente del voltaje en los canales de calcio. Almers y
colaboradores concluyen que la supresión de estas corrientes es causada por un
agotamiento de iones de calcio en los túbulos T (Almers, et al. 1981), sin embargo
el grupo de Stefani lo atribuye a una inactivación dependiente del voltaje (Cota,
Siri y Stefani, 1984 y Sánchez y Stefani, 1983). Otros trabajos también apoyan al
proceso de inactivación (Beam y Knudson, 1988 y Francini, et al. 1992). Sin
embargo los estudios de reconstitución de canales tipo L en bicapas no muestran
inactivación con el voltaje (Affolter, H., y Coronado R. 1985 y Mejia-Alvarez, et al.
1991 y Rosenberg, R. L., et a/. 1986).
La imposibilidad para aclarar la naturaleza de la inactivación resalta la falta
de un modelo que permita hacer estudios directos sobre el canal.
El receptor a la dihidropiridinas
Fármacos que bloquean al canal actúan coma antagonistas de calcio.
Clínicamente estos inhibidores se han agrupado en tres generaciones, los
representantes de la ultima generación tales como la amlodipina y lacidipina o
bien tratamientos combinados con otras drogas son los tratamientos terapéuticos
que actualmente se recomiendan en padecimientos cardiovasculares (Herman ZS
1999 y Muino Miguez A 1999).
Estructuralmente los antagonistas de calcio se clasifican en tres principales
grupos:
a) 1, 4-dihidropiridinas (DHP) que incluyen a la nitrendipina, nifedipina y PN
200-l 10.

19
b) Fenilalquilaminas coma el verapamil, D600 y bepridil
c) Benzodiazepinas (diltiazem).
Recientemente se ha desarrollado una cuarta categoría encabezada por el
Mibefradil el cual puede bloquear a los canales de calcio tipo T (Pitt B 1997).
El verapamil fue el primer fármaco utilizado en los Estados Unidos de
Norteamérica (McGoon, Vlietstra, Holmes y Osborn 1982), este farmaco actúa en
la boca interna del canal de calcio causando alteraciones en su cinética o gating
(Glossmann et al. 1982). Este mecanismo de acción contrasta con el de las 1,4-
dihidropiridinas (coma la nifedipina), las cuales bloquean la corriente lenta
entrante de calcio en el rango nanomolar sin afectar la cinética, son estéreo
especificas y también tejido especifico recordando el modo de acción de la
tetrodotoxina en el canal de sodio. Estas propiedades sitúan a la nifedipina coma
uno de los principales agentes bloqueadores usado en estudios
electrofisiológicos.
Las dihidropiridinas son fármacos sintéticos liposolubles derivados de la
papaverina (ver figura a continuación). Pequeñas modificaciones en la molécula
de la nifedipina pueden resultar en el efecto opuesto esto es incrementar la
corriente de calcio. De esta forma los diferentes enantiomeros ópticos de la
nifedipina afectan significativamente a la corriente de calcio (Kokubun, 1987 y
Bechem et al., 1988). En el músculo cardiaco estos derivados causan un efecto
inotrópico positivo y en el músculo liso inducen la contracción, por esta razón
estos compuestos se denominan agonistas de la DHP. El más potente de estos
compuestos es el Bay K 8644. Registros unitarios de células cardiacas en cultivo
muestran que se incrementa la amplitud de las corrientes en forma proporcional a
la concentración de la droga usada; coma resultado de un incremento en el
tiempo de apertura de los canales de calcio (Hille B., 1992).

20
16 ORGANIC Ca CHANNEL ANTAGONISTS AND AGONISTS
Three chemical classes of Ca channel antagonist block L-type Ca chan- nels reversibly, usually acting more potently on vertebrate cells than on invertebrate cells. Verapamil is a phenalkylamine, diltiazem a ben- zothiazepine, and nifedipine a dihydropyridine. Their relatives include D-600, which is verapamil plus another methoxy group on the leftmost ring, and nitrendipine, which is nifedipine with the nitro group moved to the 3-position and with one methyl ester converted to an ethyl ester. Such blockers have clinical usefulness in the treatment of supra- ventricular cardiac arrhythmias, angina pectoris, and hypertension. BAY K 8644, a dihydropyridine Ca-channel agonist, increases the open- ing probability of L-type Ca channels. All of these compounds are modeled after papaverine, a smooth muscle relaxant found in opium.
Gracias a la alta afinidad que tienen las dihidropiridinas para bloquear al
canal de calcio o bien para potenciar sus corrientes, hacen de estas una
excelente herramienta farmacológica para estudiar a los DHPR.
Movimiento de carga
El proceso del acople entre la excitación y la contracción en el músculo
esquelético de los vertebrados se inicia cuando la membrana citoplasmática se
despolariza, dando lugar a una serie de eventos que culminan con la elevación en
los niveles de calcio intracelular y la consecuente activación de la maquinaria
contractil. Este proceso esta constituido por tres etapas, la primera es un proceso
sensible al voltaje, caracterizado por un movimiento de carga intramembranal,
después ocurre la liberación del calcio, este proceso es independiente de la
corriente de calcio a través de los DHPR constituyendo así un mecanismo
diferente al observado en el músculo cardiaco donde la corriente de calcio es el

21
elemento clave del acople entre la excitación y la contracción (Cleemann, L., y Morad, M. 1991). El tercer proceso, aun no esclarecido, es el enlace entre la percepción al voltaje y la compuerta del canal,
Los primeros indicios de que el canal de calcio podría actuar coma el censor al voltaje en este proceso proviene de los estudios de Schneider y Chandler en 1973, quienes observaron el movimiento de una carga no lineal dependiente del voltaje. Este movimiento de carga se ve disminuido considerablemente por la nifedipina, por lo que Ríos y Brum (1987) concluyeron que el movimiento de carga es debido a la activación del sensor al voltaje del DHPR localizado en la membrana del sistema de túbulos. Así la despolarización induce un cambio en el DHPR, dando coma resultado un movimiento de carga (Melzer, W., et al. 1986 y Ríos, E., y Pizarro, G. 1991),

22
17 MODELS OF T-SR COUPLING (A) Schneider and Chandler (1973) envisioned a charged sensor in the T-tubular membrane pulling on a plunger to open the release channel in the SR membrane. [From Chandler et al., 197613.1 (B) Two membrane proteins, the dihydropyridine receptor with voltage sensors and the ryanodine receptor with a foot and pore, are now known to be compo-nents of the release machinery. [After Takeshima et al., 1989.1
18 SLOW ICa OF SKELETAL MUSCLE The unusual, dihydropyridine-sensitive ICa., of skeletal muscle T tu-bules. Frog muscle fiber filled with isotonic (TEA), EGTA and bathed in 100 mM Ca methansulfonate. Voltage-clamp step from - 70 to - 10 mV. Note the slow time scale. T= 22°C. [From Almers et al, 1981.1
Hodgkin A. L. y Huxley A. F. en 1952 postularon que la activación de un
canal i6nico dependiente del voltaje puede ser debida a una alteración
conformacional dando coma resultado la apertura del canal, esto puede estar
causado por un movimiento de cargas o compuertas en respuesta al cambio del
voltaje. El primer movimiento de compuertas registrado fue en el canal de sodio
del axón gigante del calamar (Armstrong, C. M., y Bezanilla, F. 1973). Para pulsos

23
despolarizantes cortos donde se ha eliminado la participación de corrientes
iónicas, es posible registrar la apertura y cierre de los canales iónicos. Durante el
inicio del pulso el movimiento de carga se denomina Qon y representa la apertura
de los canales de sodio, debe ser igual al movimiento de carga al final del pulso
Qon, lo cual indica el cierre del mismo numero de canales abiertos. En el case de
los canales de sodio, los procesos de inactivación resultan en una disminución de
la carga Qon, en este case un pulso de 10 milisegundos es suficiente para
inmovilizar el 70 % de la carga. La inmovilización de la carga y la inactivación de
los canales de sodio son procesos estrechamente relacionados (Lamb, G. D.
1987). Sin embargo en el case del músculo esquelético las corrientes de
compuerta y las corrientes del sensor al voltaje no necesariamente son las
mismas, puesto que pueden participar otros elementos, coma pueden ser los
mecanismos liberadores de calcio denominados Qy. (Ríos, E., y Pizarro, G. 1991)
Por otra parte en el músculo cardiaco están claramente identificadas las
corrientes de compuerta del canal de calcio (Cleemann, L., y Morad, M. 1991). El
músculo esquelético del anfibio también presenta un movimiento de carga, el cual
se muestra coma una breve corriente exponencial, cuya magnitud cambia con el
voltaje de una forma descrita por la función Boltzmann. En el músculo rápido del
anfibio esta función sigmoidea entre la carga y el voltaje muestra un punto de
inflexión entre los potenciales de -35 a -45 mV y desplaza una carga máxima de
24-30 ηC/ µF (Adrian y Almers, 1976; Chandler, et al. 1976).
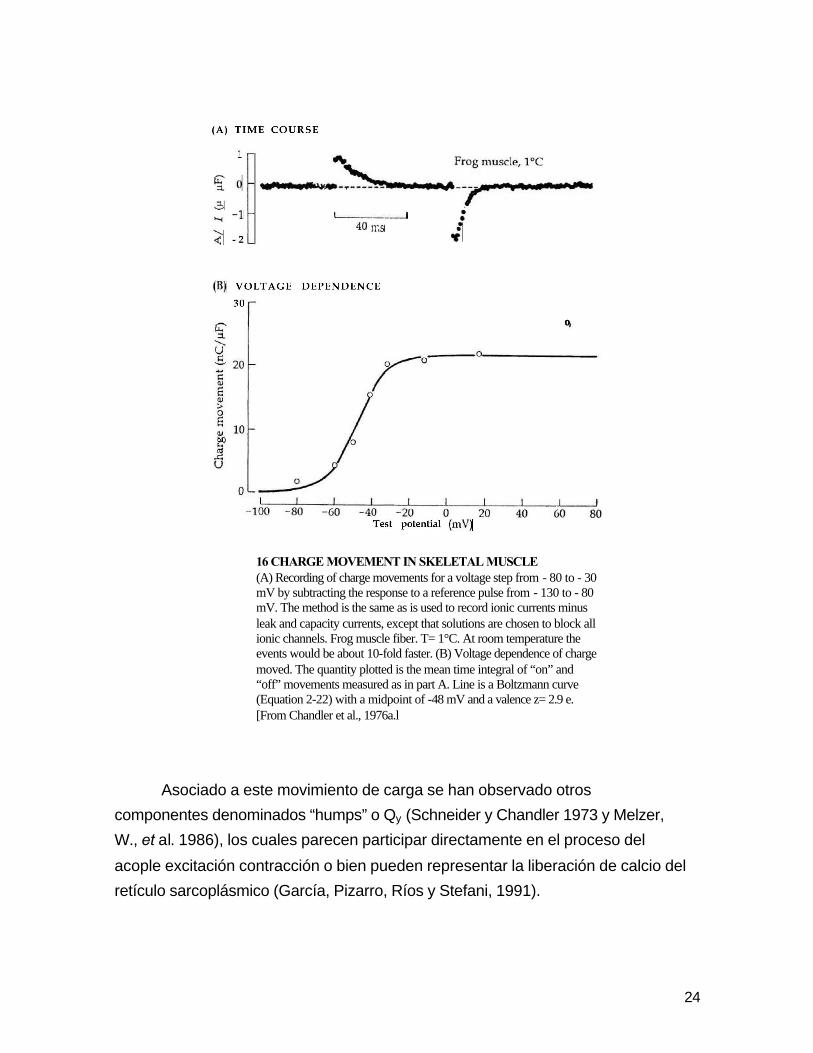
24
16 CHARGE MOVEMENT IN SKELETAL MUSCLE (A) Recording of charge movements for a voltage step from - 80 to - 30 mV by subtracting the response to a reference pulse from - 130 to - 80 mV. The method is the same as is used to record ionic currents minus leak and capacity currents, except that solutions are chosen to block all ionic channels. Frog muscle fiber. T= 1°C. At room temperature the events would be about 10-fold faster. (B) Voltage dependence of charge moved. The quantity plotted is the mean time integral of “on” and “off” movements measured as in part A. Line is a Boltzmann curve (Equation 2-22) with a midpoint of -48 mV and a valence z= 2.9 e. [From Chandler et al., 1976a.l
Asociado a este movimiento de carga se han observado otros
componentes denominados “humps” o Qy (Schneider y Chandler 1973 y Melzer,
W., et al. 1986), los cuales parecen participar directamente en el proceso del
acople excitación contracción o bien pueden representar la liberación de calcio del
retículo sarcoplásmico (García, Pizarro, Ríos y Stefani, 1991).

25
PLANTEAMIENTO DEL PROBLEMA
Problema
La localización de los canales de calcio en el sistema de túbulos
transversos ha dificultado la realización de los experimentos bajo condiciones controladas. Esto es debido principalmente al intrincado patrón que forma este
sistema y al espacio tan restringido que existe en ellos, así es que la búsqueda de
nuevos métodos o modelos que permitan estudiar directamente tanto a los
canales iónicos presentes en el sistema de túbulos y a sus corrientes ha sido un
gran reto en los estudios electrofisiológicos.
Hipótesis
Se ha pensado que las vesículas, formadas por el tratamiento de alto
potasio y colagenasa, provienen del sistema de túbulos transversos razón por la
cual debe ser posible registrar en ellas corrientes de calcio. Las ventajas
operativas que ofrece las vesículas pueden situarla coma un modelo adecuado
para la caracterización fisiológica de las corrientes de los canales sensibles a las
dihidropiridinas. objetivos de/ presente trabajo
1) Situar a las vesículas derivadas del músculo esquelético como un modelo
adecuado para el estudio directo de los canales de calcio tipo L.
2) Caracterizar electrofisiológicamente la corriente ionica de los canales de calcio
en las vesículas bajo fijación de voltaje con la técnica de Patch clamp en la
modalidad de vesícula completa
a) Elaborar la curva corriente voltaje de la corriente ionica registrada en una
solución donde el único ion permeable-es el bario.
b) Determinar la cinética de activación de la corriente ionica.
c) Determinar si la corriente ionica de bario presenta inactivación dependiente
del voltaje.

26
d) Comprobar si el músculo esquelético del anfibio adulto presenta procesos
de potenciación en las corrientes de cola.
3) Comprobar la sensibilidad de estas corrientes a las dihidropiridinas coma la
nifedipina y el Bay K 8644.
4) Verificar la presencia de movimientos de carga 1 y 2.

27
METODOLOGÍA
Obtención de vesículas
La obtención de vesículas esféricas a partir de la membrana del músculo
esquelético del anfibio se hizo siguiendo el protocolo descrito por Standen y
colaboradores en 1984 adaptado para el registro electrofisiológico en la
modalidad de “whole-vesicles” (Camacho, et al.1 996).
Brevemente el protocolo estuvo formado por los siguientes pasos:
1. Se usaron ranas Rana montezumae.
2. Las ranas fueron sacrificadas por decapitación.
3. Disección del músculo semitendinoso.
4, Limpieza de tejido conectivo en solución Ringer para rana (ver tabla 1).
5 .Incubación del músculo semitendinoso en una solución de 120 mM KCI con
colagenasa (tipo IA, 50 unidades/ml, de Sigma).
6. Incubación por 90 minutos a temperatura ambiente.
7 . Lavado del músculo con una solución de 120 mM KCI.
8. Las vesículas se formaron espontáneamente después de un period0 de
aproximadamente 60 minutos a 20-22 “C.
Soluciones
Para el registro de las corrientes de bario (Ba2+) se empleo la solución
extravesicular (Ver tabla 1) donde el bario fue el único ion permeable y la
conductancia al potasio se bloqueo con TEA. La composición de la solución
utilizada para el registro del movimiento de carga también se muestra en la tabla I.
Todos los químicos fueron obtenidos de Sigma Chemical Co. 6 de Aldrich
Chemical Co. (St. Louis, M. 0.).
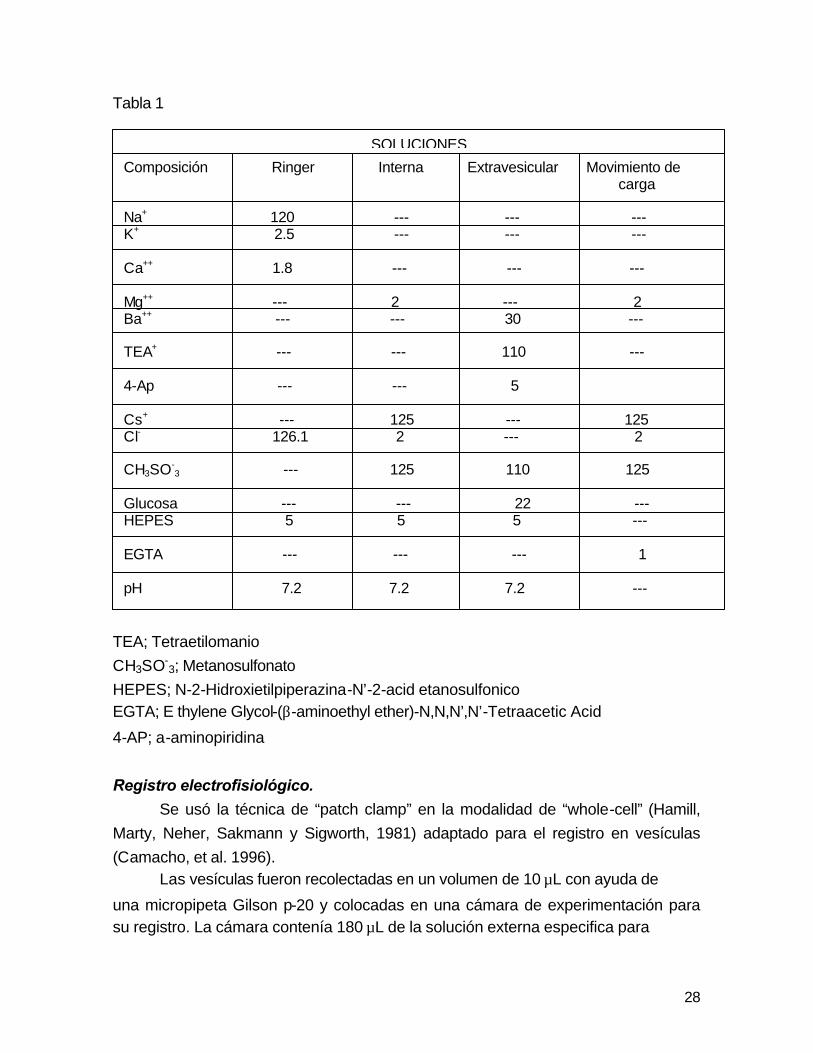
28
Tabla 1
TEA; Tetraetilomanio
CH3SO-3; Metanosulfonato
HEPES; N-2-Hidroxietilpiperazina-N’-2-acid etanosulfonico EGTA; E thylene Glycol-(β-aminoethyl ether)-N,N,N’,N’-Tetraacetic Acid
4-AP; a-aminopiridina
Registro electrofisiológico.
Se usó la técnica de “patch clamp” en la modalidad de “whole-cell” (Hamill,
Marty, Neher, Sakmann y Sigworth, 1981) adaptado para el registro en vesículas
(Camacho, et al. 1996). Las vesículas fueron recolectadas en un volumen de 10 µL con ayuda de
una micropipeta Gilson p-20 y colocadas en una cámara de experimentación para su registro. La cámara contenía 180 µL de la solución externa especifica para
SOLUCIONES
Composición Ringer Interna Extravesicular Movimiento de carga Na+ 120 --- --- --- K+ 2.5 --- --- --- Ca++ 1.8 --- --- --- Mg++ --- 2 --- 2 Ba++ --- --- 30 --- TEA+ --- --- 110 --- 4-Ap --- --- 5 Cs+ --- 125 --- 125 Cl- 126.1 2 --- 2 CH3SO-
3 --- 125 110 125 Glucosa --- --- 22 --- HEPES 5 5 5 --- EGTA --- --- --- 1 pH 7.2 7.2 7.2 ---

29
cada tipo de registro (ver tabla 1), una vez que se le adicionaban las vesículas se
trasladaba a la platina de un microscopio óptico invertido. La cámara se conecta a
tierra mediante un puente de agar (ver diagrama a continuación)
Los electrodos consistieron en micropipetas de vidrio duro (KIMAX-51,
Kimble Glass, Toledo, OH), estas micropipetas están libres de artefactos que
puedan afectar el registro de las corrientes (Camacho, et al. 1996).
Las micropipetas fueron estiradas siguiendo un protocolo de dos pasos,
usando un estirador vertical David Kopf 700 D (Tujunga, CA, USA) y fueron
llenadas con una solución interna, cuya composición se define en la tabla 1. La
resistencia eléctrica de las micropipetas estuvo dentro del rango de 6 y 8 M B
El electrodo fue implantado en la vesícula con ayuda de un
micromanipulador hidráulico. Se mantuvo una presión positiva en el interior de la
micropipeta mientras se acercaba el electrodo a la vesícula, lo cual permito la
formación de buenos gigasellos entre el vidrio y la membrana de la vesícula.
Después de la formación del sello se aplico una débil presión negativa para
romper el segmento de la membrana contenido en la micropipeta, logrando así la
configuración de vesícula completa.

30
Protocolo de pulses y captura de datos
Los registros de la corriente de bario fueron realizados a una temperatura
de15a17%.
El sistema de registro utilizado consistió en:
1. Las corrientes de membrana (Im) fueron capturadas con un amplificador
Axopatch (modelo 200 A, Axon Instruments) usando un microprocesador
Pentium compatible con IBM-PC/AT.
2. Las señales analógicas fueron digitalizadas a una resolución de 12 bits a
través de una interface LabMaster (TL-1DMA interface, Axon Instruments)
que también genera los pulsos comando.
3. La Im, fue amplificada y filtrada con un filtro pasa bajos Bessel de cuatro
polos, a una frecuencia “corner” de no mas de la mitad de la frecuencia de
muestreo.
4. Cada protocolo de pulsos fue flanqueado por 5 pulsos hiperpolartizantes
consecutivos a -20 mV del potencial fijado coma reposo (Et, = -80 mV). Las
corrientes generadas durante los pulsos hiperpolarizantes fueron usadas
para calcular la capacitancia lineal de membrana y para restar las
corrientes “leakage” de los pulsos de prueba. En algunos cases, se usaron
corrientes generadas por pulsos de +20 mV a partir de un Et,= -100 mV.
5. Los datos fueron analizados usando programas tanto de pCLAMP (versión
6.0, de Axon Instruments) coma programas desarrollados en el laboratorio.
Diseño experimental
Registro de /as corrientes de bario
Para medir la activación de las corrientes de bario (IBa), se aplicaron pulsos
comando de 500 ms de duración y amplitud variable partiendo del Eh hasta +70
mV en pasos de 20 mV. El intervalo entre pulsos fue de al menos 1 Segundo para
evitar los cambios en la cinética del canal causados por la despolarización
(Feldmeyer et al. 1990).

31
Corrientes de cola
A diferencia de las corrientes de bario evocadas durante los pulses
despolarizantes, las corrientes que se observan al final del pulso de prueba,
durante la repolarización, se denominan corrientes de cola [en mamíferos tienen
una amplitud promedio de 6.6kO.9 ηA y una constante de activación de 50 ms].
Las corrientes de cola requieren de un prepulso de 600 ms para que alcancen su
valor máximo ya que con prepulsos menores las corrientes son pequeñas pese a
que se activa el numero máximo de los canales de calcio
No se realizó compensation de la resistencia en serie ya que esto podría
subestimar la amplitud de las corrientes de cola.
Inactivación
Las propiedades de inactivación de las corrientes fueron examinadas
siguiendo el protocolo de dos pulsos descrito inicialmente para los canales de
sodio (Hodgkin y Huxley, 1952). La inactivación en el estado estacionario fue
investigada por aplicar prepulsos de 5000 ms a diferentes potenciales seguidos
por un pulso de prueba de 500 ms a +30 mV. La relación de la corriente obtenida
durante el pulso de prueba y el potencial de membrana del prepulso se ajusta a
una función Boltzmann del tipo:
I=ImaJ( 1 +exp( (E-E,)/k))
Donde Imax es el valor máximo de la corriente, E, es el potencial de
membrana, E es el punto en el cual el potencial alcanza la mitad de su valor
máximo y K es una medida de la pendiente de la curva.
Movimiento de carga
El movimiento de carga 1 fue medido en vesículas polarizadas mantenidas
a -70 mV a las cuales se les aplico un protocolo de pulsos de 100 ms a diferentes
potenciales despolarizantes. Para restar los componentes lineales se integro la
corriente obtenida durante un pulso hiperpolarizantes al Eh (-100 mV), el valor
promedio de cuatro de estos pulsos fue escalado al potencial usado durante el
pulso de prueba. Este procedimiento produce menores niveles de ruido que los
protocolos en los cuales se usa solamente una de las corrientes hiperpolarizantes.
Para mantener la estabilidad de los registros en los experimentos de
movimiento de carga la solución extracelular contenía Ca2+, sin embargo coma

32
esto favorece la formación de corrientes de cola de Ca2+ durante las espigas “off”
solo se determine el movimiento de carga durante las espigas “on”.
Para determinar la presencia de la carga 2 en las vesículas, se aplicaron
pulsos de +60 a -170 mV en pasos de 10 mV a partir de un potencial fijado en Et,
=0 mV.
La dependencia del voltaje de la activación del movimiento de carga fue
ajustada a una función Boltzmann de la siguiente forma:
Q=Qmax/(1 +exp((E-Em)/k))
Donde Qmax es el valor máximo de la carga, E, es el potencial de
membrana, E es el potencial donde Q=0.5 Qmax, y K es una medida de la
pendiente de la curva.
El ajuste de los datos experimentales a la formula se hizo mediante un
algoritmo de mínimos cuadrados no lineal. Los valores de los parámetros están
expresados coma media Terror estándar. Se uso la prueba t de student con un
valor de significancia a un nivel de p<0.05.

33
RESULTADOS
Con troles
Después de cancelar electrónicamente los componentes capacitivos
asociados a la pipeta durante la formación de un gigasello se procedió a romper la
membrana contenida en la pipeta, lo que resulta en grandes espigas capacitivas
correspondientes a la capacitancia de la membrana. La pendiente obtenida al
graficar la magnitud de la corriente capacitiva en función del potencial de
mantenimiento determina el valor de la capacitancia de la vesícula. Las vesículas
utilizadas tenían un diámetro de aproximadamente 60 µm y una capacitancia de
membrana de aproximadamente 79 ρF. Para el registro de las corrientes iónicas,
en algunos experimentos se compenso electrónicamente hasta un 60% la
capacitancia de la vesícula y la resistencia en serie, en los otros cases se
aplicaron cinco pulsos hiperpolarizantes consecutivos de -20 mV a partir del Eh
de -80 mV, para restar estos elementos lineales.
Corrientes de bario.
Las vesículas mostraron sellos muy estables en el lapso en el que se
realizaron los registros (aproximadamente 20 min.), durante este tiempo fue
posible registrar corrientes iónicas sin que mostraran algún tipo de alteración.
Se registro una corriente iónica entrante (ver figura 1) en las soluciones que
contienen bario coma único ion permeable, los demás iones fueron reemplazados
por sus equivalentes impermeables (ver tabla 1 en la metodología). La activación
de las corrientes de bario ocurre a un potencial cercano a 0 mV. La corriente se
incrementa a potenciales despolarizantes y a potenciales superiores a 20 mV la
corriente comienza a disminuir.

34
Figura 1.
Figura 1. Corrientes de bario Se muestran las corrientes macroscopicas de Bario registradas en la membrana de las vesículas, obtenidas durante los pulsos de voltaje al potencial indicado en cada trazo. Se restaron los componentes lineales a cada registro.

35
Gráfica 1. Grafica 1 Se muestra la relación entre corriente al pica y potencial de membrana durante el pulso comando de un experimento típico (Eh=-80 mV).

36
En la curva corriente voltaje (ver grafica 1) se muestra que la corriente al
pica alcanzo su máximo [-3.0 + 0.7 µA µF-1 (n=14)] a un potencial de membrana
de +20 mV, por encima de este potencial la magnitud de la corriente comienza a
declinar, sin embargo a +70 mV aun es una corriente entrante, lo cual indica un
potencial de inversión muy positivo.
El curso temporal de la corriente entrante de bario se hace mas rapid0
conforme la despolarización aumenta (ver figura 2 y grafica 2)
Estas corrientes no se mantuvieron en el tiempo y su magnitud disminuyo
casi en su totalidad con largos pulsos despolarizantes (ver inactivación).

37
Figura 2. Figura2. Tiempo al pica Serie de corrientes de Bario registradas en la membrana de las vesículas, al potencial indicado en cada trazo. Los componentes lineales fueron restados.

38
Gráfica 2 Gráfica 2. Tiempo al Pica Se muestra la relación entre el tiempo al pica y el potencial de membrana en condiciones control.

39
Corrientes ;de cola
De acuerdo al protocolo descrito en la metodología en las vesículas se
observaron corrientes de cola al final de un pulso despolarizante (ver figura 3).
En miotubos de mamíferos las despolarizaciones prolongadas incrementan
la corriente de cola en más de un 400 %, (Fleig y Penner, 1996). Sin embargo en
las vesículas se encontró solamente un pequeño incremento en la amplitud de las
corrientes de cola coma resultado de incrementar la duración del pulse (ver figura

40
Figura 3. Figura 3. Corrientes de cola La influencia de la duración del pulse sobre las corrientes de cola de Bario, Los registros muestran corrientes sin restar generadas con el protocolo de pulsos mostrado en el inciso C. A.- Corrientes durante un pulso de 50 ms a +60 mV de Eh=-80 mV. B.- Corrientes durante un pulso de 600 ms al mismo potencial. D.- La resta de las corrientes de cola, “b-a”. La longitud del pulso induce una corriente de cola que es ligeramente mayor que la producida por un pulso menor. En “D” la escala de tiempo y amplitud es diferente a las usadas en “A” y B”. En todos los cases se muestra el mismo experimento.

41
lnactivación
Las corrientes tipo L alcanzan un máximo y luego comienzan a decaer
durante el pulse despolarizante. Las propiedades de inactivación de las corrientes
fueron examinadas siguiendo el protocolo de dos pulsos descrito en la
metodología.
Un experimento representativo se ilustra en la figura 4 donde se muestran
las corrientes de membrana después de restarle los componentes lineales. Las
corrientes de bario se inactivan durante el pulso de prueba.
Se encontró que conforme se incrementa la magnitud del prepulso, la
amplitud de la IBa disminuye paulatinamente hasta desaparecer coma se espera
de un proceso de inactivación dependiente del voltaje. En tres experimentos las
corrientes se inactivaron en promedio a -40.2 f 4.7 mV.
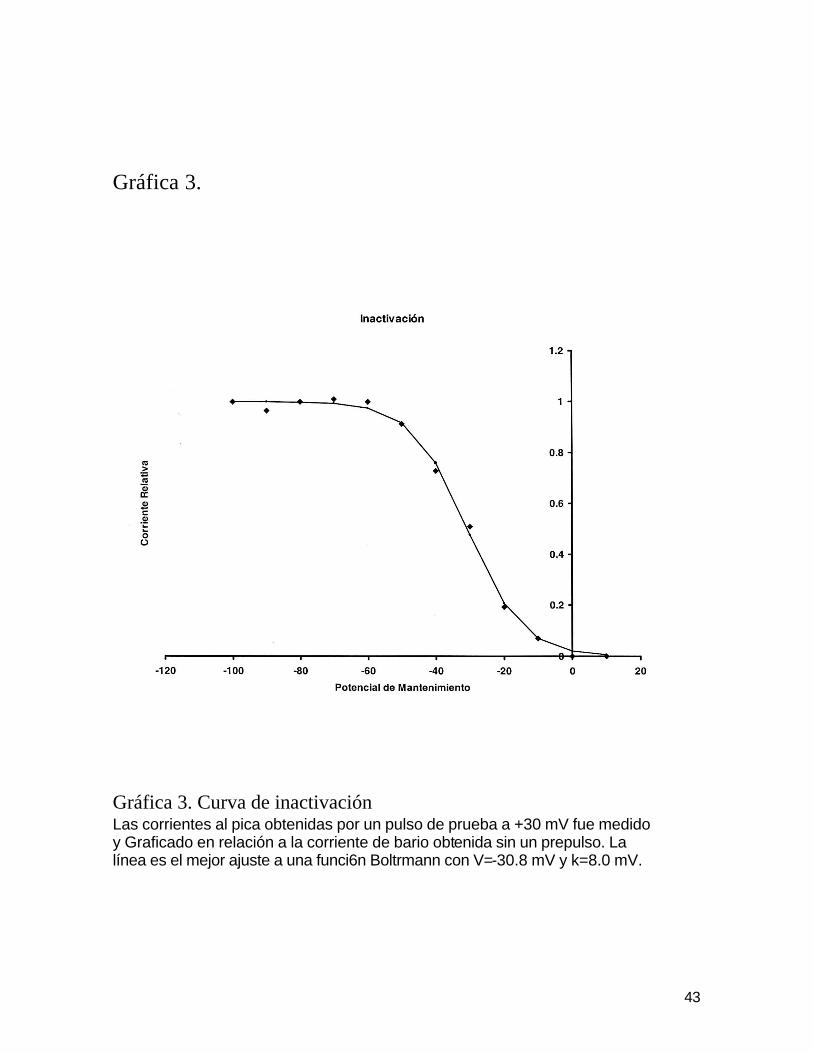
En la grafica 3, los símbolos representan valores relativos de corrientes al
pica y la línea continua es el mejor ajuste de una función de inactivación de
acuerdo a la función de Boltzmann con V=-30.8 mV y k=8.0 mV.

42
Figura 4
Figura 4. lnactivacion La lnactivación de las corrientes de bario en el estado estacionario. Las vesículas fueron fijadas a un pulso comando de amplitud variable por 5 segundos y luego llevadas por un pulso de prueba de 30 mV. Eh= -80 mV.
43
Gráfica 3. Gráfica 3. Curva de inactivación Las corrientes al pica obtenidas por un pulso de prueba a +30 mV fue medido y Graficado en relación a la corriente de bario obtenida sin un prepulso. La línea es el mejor ajuste a una funci6n Boltrmann con V=-30.8 mV y k=8.0 mV.

44
El receptor a las dihidropiridinas Se investigo la posibilidad de que la corriente de bario fluya a través del
receptor a las dihidropiridinas. Al comparar los registros de las corrientes de bario de las vesículas
incubadas en presencia del BAY K 8644, un agonista a las DHP, con registros obtenidos en condiciones controles se observa que ambas corrientes se activan a potenciales similares y tienen un curso temporal semejante, sin embargo, la amplitud de la corriente fue significativamente mayor en todos los potenciales probados (ver figura 5). En 14 experimentos la corriente al pica en soluciones control fue -3.0 ± 0.7 µA µF-1 y en presencia de BAY K el promedio fue de -6.8 ± 1.5 µA µF-1 (ver grafica 4)
El empleo de cobalto a una concentración de 2 mM, un bloqueador de canales de calcio tipo L, suprime totalmente las corrientes de bario registradas en las vesículas, como se muestra en la figura 6.
Este trabajo muestra la presencia de canales de calcio sensibles a las DHP en las vesículas, coma se indica por las corrientes iónicas de Ba2+ potenciadas por el Bay K y suprimidas por el cobalto.
La presencia de canales de calcio tipo L sugiere que para la formación de las vesículas hay una importante contribución de la membrana del sistema tubular.

45
Figura 5 Figura 5 Efecto de1 BAY K A.Corrientes de Bario control B.Corrientes de Bario en presencia de Bay K

46
Gráfica 4. Gráfica 4. Corriente al pica en presencia del BAY K Se muestra la relación entre corriente al pica de las corrientes de bario registradas en la membrana de la vesícula, obtenidas a diferentes potenciales del pulso de prueba y en presencia del BAY K (1 PM) después de restar los componentes lineales. Eh=-80 mV.

47
Figura 6 Figura 6. Efecto de1 Cobalto
El cobalto a la concentración de 2 mM inhibe totalmente las corrientes de Bario.
A. Corrientes de Bario
B. Corrientes de Bario en presencia de 2 mM de cobalto

48
Movimiento de carga en /as versículas
La aplicación de un protocolo de estimulación a las vesículas mantenidas
en soluciones carentes de iones permeables y después de restar los
componentes lineales, mostraron corrientes capacitivas no lineales de magnitud
muy pequeña. Fueron distinguibles dos corrientes no lineales denominadas
corrientes “on” al inicio del pulso y corriente “off” al final del pulso (ver figura 7).
Ambas corrientes disminuyeron de magnitud con el tiempo y no mostraron
la participación de otros elementos o “humps”. Se observo que el valor de carga
on y off no era el mismo, observándose una Clara separación en el valor de la
carga a despolarizaciones de -30 mV, lo cual sugiere la presencia de corrientes
de cola de calcio, sin embargo la activación de IBa se hace evidente a 0 mV, Esta
diferencia puede deberse a la composición iónica de las soluciones. La presencia
de altas concentraciones de cationes divalentes puede desviar la dependencia del
voltaje de la activación de los canales hacia potenciales mas positivos debido a
los efectos de carga de superficie (Hille, 1992).
La participación de una corriente de cola de Bario fue confirmada por
bloquear las corrientes de calcio tipo L con nifedipina (10µM). Bajo estas
condiciones la proporción entre la carga “on” y “off a +50 mV fue de 0.96kO.05
(n=5).

49
Figura 7. Movimiento de carga en las vesículas. Corrientes macroscopicas de membrana en la ausencia de iones permeables. Los registros muestran corrientes no lineales durante los potenciales de prueba (potenciales en mV).
50
Las corrientes off están formadas por dos componentes uno rápido y otro
lento relativamente mas pequeño. La amplitud del componente rápido se
incrementa con el voltaje. El componente lento no cambia significativamente en el
mismo rango de voltaje. Esto representa la participación de dos distintos
elementos; Una corriente de cola entrante de Ba2+ y una corriente capacitiva no
lineal, por este motivo el análisis del movimiento de carga se centro solo en la
fase “on”.
Los valores para el movimiento de carga en las vesículas polarizadas
fueron obtenidos por restarle a la carga total “on” de cada potencial de membrana
la carga lineal extrapolada a dicho valor. La grafica 5 muestra la relación entre la
carga movilizada durante los pulsos despolarizantes coma una función del
potencial de membrana. Tal coma se espera de una corriente de carga hay una
saturación a grandes despolarizaciones. Los valores de la carga durante los
pulsos hiperpolarizantes son muy cercanos a cero.
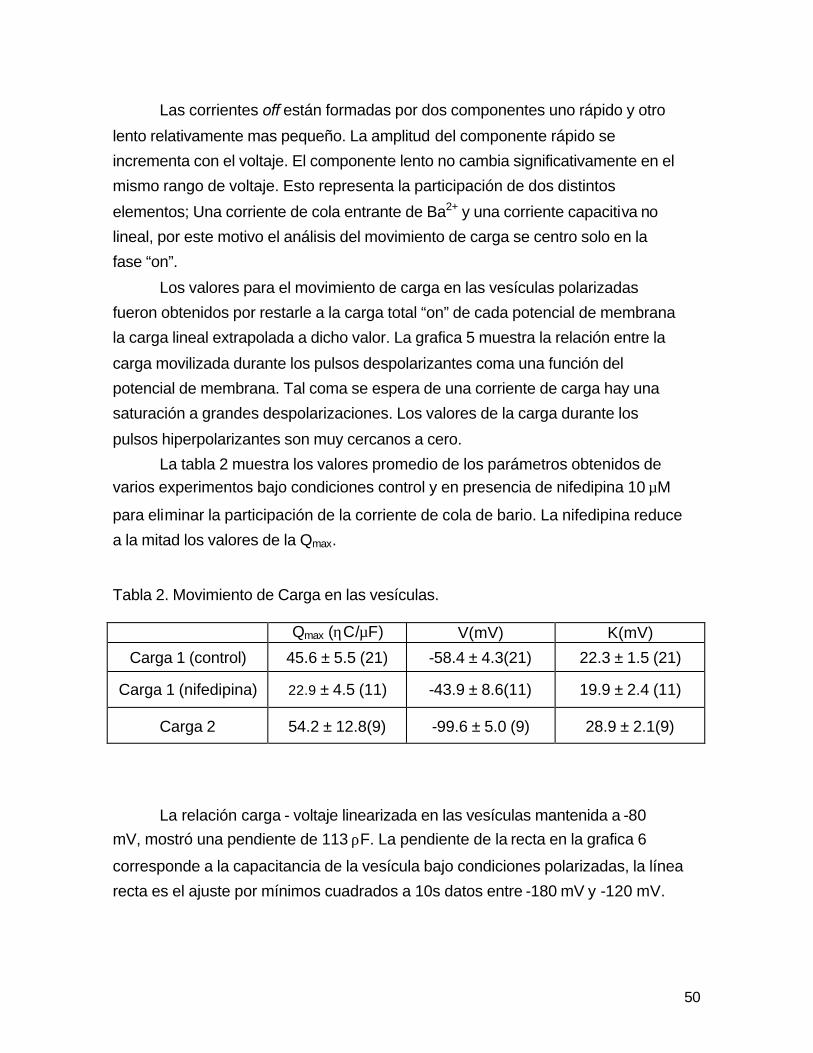
La tabla 2 muestra los valores promedio de los parámetros obtenidos de
varios experimentos bajo condiciones control y en presencia de nifedipina 10 µM
para eliminar la participación de la corriente de cola de bario. La nifedipina reduce
a la mitad los valores de la Qmax. Tabla 2. Movimiento de Carga en las vesículas.
Qmax (ηC/µF) V(mV) K(mV)
Carga 1 (control) 45.6 ± 5.5 (21) -58.4 ± 4.3(21) 22.3 ± 1.5 (21)
Carga 1 (nifedipina) 22.9 ± 4.5 (11) -43.9 ± 8.6(11) 19.9 ± 2.4 (11)
Carga 2 54.2 ± 12.8(9) -99.6 ± 5.0 (9) 28.9 ± 2.1(9)
La relación carga - voltaje linearizada en las vesículas mantenida a -80
mV, mostró una pendiente de 113 ρF. La pendiente de la recta en la grafica 6
corresponde a la capacitancia de la vesícula bajo condiciones polarizadas, la línea
recta es el ajuste por mínimos cuadrados a 10s datos entre -180 mV y -120 mV.

51
Gráfica 5.
Gráfica 5. Movimiento de carga Movimiento de carga no lineal coma una función del voltaje. La línea es el mejor ajuste a una función Boltzmann con Qmax,=37.2 ηC/µF, V=65.2 mV y k-1 6.4 mV.

52
Gráfica 6. Gráfica 6. Movimiento de carga 1 Relación carga voltaje en las vesículas. Los símbolos representan la integración de las espigas “on” de una vesícula mantenida a -80 mV. La línea recta es el ajuste por mínimos cuadrados a los datos entre -180 mV y -120 mV, con una pendiente de 113 ρF.

53
Carga 2
En las vesículas despolarizadas se registraron corrientes no lineales a
potenciales hiperpolarizantes (Eh, = 0 mv) y a potenciales positivos siguieron un
comportamiento lineal. Se observe un incremento en la capacitancia de la
membrana con relación a las vesículas polarizadas. La proporción entre las
pendientes de las vesículas despolarizadas y repolarizadas tuvo un valor
promedio de 1.23 ± 0.03 (8).
El movimiento de carga en los potenciales hiperpolarizantes no fue
bloqueado por la nifedipina.
La carga off a los mismos potenciales presenta un comportamiento lineal
en todo el rango del voltaje probado. Resultados similares fueron obtenidos en
otros 8 experimentos (ver gráfica 7).

54
Gráfica 7. Gráfica 7. Movimiento de carga 2 Relación carga voltaje en las vesículas. Los símbolos representan la Integración de las espigas “on” de una vesícula mantenida a 0 mV. La línea recta es el ajuste por mínimos cuadrados a los datos entre 0 mV y +60 mV, con una pendiente de 146 ρF.

55
CONCLUSIONES
El presente estudio describe las propiedades de la corriente iónica y el
movimiento de carga de los DHPR en vesículas obtenidas del músculo
esquelético de la rana.
Aunque no se ha reportado aun, la contribución relativa de la membrana
plasmática en comparación a la tubular, la presencia de canales de calcio tipo L
en las vesículas sugiere que el sistema de túbulos transversos contribuye en la
formación de las vesículas, dado que solo en el sistema T del músculo
esquelético del adulto se encuentran los DHPR (Nicola Siri et al.,1 980; Fosset et
al., 1983; Jorgensen et al., 1989 y Flucher et al., 1990). No obstante Jorgensen y
colaboradores sugieren que también pueden encontrarse en regiones del
subsarcolema coma las caveolas (Jorgensen et al., 1989)
Como en las vesículas no se observan partículas u organelos
citoplasmáticos ni tampoco se han reportado invaginaciones (Camacho, et al.
1996) son un modelo ideal para estudiar la actividad de los canales de calcio
sensibles a las dihidropiridinas bajo condiciones de fijación de voltaje, evitando así
las complicaciones que confiere el sistema de túbulos transversos. Corrientes de bario
La activación de las IBa es dependiente del voltaje y del tiempo. La
activación se hace más rápida a despolarizaciones mayores y largos prepulsos
despolarizantes las inactivan.
Hay algunas diferencias entre las corrientes tipo L de las vesículas con
respecto a las registradas en las fibras musculares. Primero, la amplitud de las
corrientes de bario en las vesículas es menor que en las fibras. Los valores
promedios al pica registrados fueron de 3 µA µF-1, comparados con los valores de
10 a 20 µA µF-1 de las corrientes de Ca2+ y Ba2+ de las fibras musculares de
anfibio, asumiendo una capacitancia media de 6 µFcm-2 (Sánchez y Stefani, 1983;
Almers, et al. 1981).
La menor amplitud de la corriente en las vesículas puede deberse a que
solo una pequeña fracción de la membrana del sistema T participe en la
formación de la vesícula y que la mayor parte provenga de la membrana

56
superficial, o bien puede ser que la densidad de DHPR en las vesículas sea
menor que en las fibras.
Otra posibilidad para explicar que la densidad de la corriente de calcio en
las vesículas sea menor es el proceso conocido coma “rundown” o lavado de las
fibras musculares cuando son cortadas (Arreola, et al. 1987). Se ha sugerido que
este proceso puede estar relacionado con la perdida de los mecanismos
reguladores del calcio y del ATP los cuales son importantes para la fosforilación
de los canales de calcio o bien puede haber una perdida de elementos que
confieren resistencia a la proteolisis enzimatica (Belles et a/. 1988 y Francini y
Stefani, 1989)
Finalmente otra posibilidad involucra la perdida de una serial de
retroalimentación proporcionada por el receptor a las rianodinas. Recientemente
se ha descrito que los miotubos obtenidos de ratones que no expresan el receptor
a las rianodinas (miotubos dispédicos), presentan corrientes de calcio
considerablemente menores (Nakai et al., 1996).
Las propiedades cinéticas de los canales de calcio de las vesículas son
claramente diferentes a los presentes en las fibras musculares del anfibio. La
activación de los canales tipo L en las vesículas es significativamente mas rápida,
la relación obtenida entre el tiempo al pica y el potencial de membrana muestra
valores de aproximadamente 5 veces mas rápidos que los reportados en el
músculo esquelético (Ver tabla 3; Cota, Nicola Siri y Stefani, 1983 y Francini, F.,
Bencini, C. y Squecco, R. 1996).
Tabla 3. Tiempo al Pica vs Voltaje
Potencial de Membrana (Eh)
Datos reportados
Datos Experimentales
-10 2140 433.5
0 1365 354.56
10 1100 212.61
20 1000 148.38
30 600 78.21
40 400 43.38

57
Es improbable que las diferencias cinéticas puedan ser explicadas por el
retardo de la carga asociado con el sistema tubular de las fibras musculares. El
potencial de membrana tubular del músculo de la rana ha sido medido con
marcadores potenciométricos y se ha observado que cuando se aplica un pulso
cuadrados de voltaje en la superficie, hay un retardo de 13 ms para que el
sistema T alcance el 95 % de este pulso (Kim y Vergara, 1998). Claramente este
retard0 tendría un efecto mínimo sobre estas corrientes que duran segundos.
No se descarta la posibilidad de que las altas concentraciones de bario
tengan efectos sobre la cinética del canal. Dirkensen y Beam (1995) reportaron
que la tasa de activación de los canales tipo L en miotubos de mamífero es
acelerada cuando el calcio en el medio extracelular es reemplazado por una alta
concentración de bario. Sin embargo este efecto aun es muy pequeño coma para
explicar nuestros resultados, por ejemplo usando los parámetros cinéticos
reportados por Stefani y colaboradores a +50 mV y corrigiendo las diferencias en
temperatura (Cota, Nicola Siri y Stefani, 1983) se calculo un tiempo al pica de 110
ms, aproximadamente 5 veces mayor que los registrados en las vesículas.
Por otra parte es posible que en las vesículas se pierda algún tipo de factor
modulador sobre la cinética del canal el cual este presente en las fibras. A este
respecto es interesante notar que en los experimentos de Nakai y colaboradores
(1996) y Fleig y Penner (1996) la activación de las corrientes de calcio (por
ejemplo a +10 mV en miotubos dispédicos) muestran un curso temporal
comparativamente mas rápido en células que carecen del receptor a las
rianodinas con respecto a los miotubos controles.
Una última posibilidad para explicar la diferencia en valores cinéticos de las
corrientes registradas en las vesículas es que estas Sean generadas por un nuevo
canal que se activa con un curso temporal menor. En el pasado se han usado la
diferencia en cinéticas de los componentes de la corriente de cola para diferenciar
a dos distintos tipos de canales de calcio en las células excitables (Matteson y
Armstrong, 1986) pero no tenemos evidencia experimental que sugiera la
presencia de mas de un canal de calcio en las vesículas.

58
Corrientes de cola
La ausencia de potenciación en las vesículas puede ser explicada por los
pronunciados efectos en la inactivación de los DHPR causado por la
despolarización.
Otra posibilidad es que las vesículas no posean ciertos componentes
celulares coma el Receptor a las Rianodinas y su ausencia puede resultar en un
pronunciado efecto sobre el proceso de la potenciación, a este respecto Fleig y
Penner (1996) observaron que en los miotubos dispédicos no se presenta el
incremento causado por la despolarización
lnactivación
Las corrientes Bario disminuyen con la despolarización prolongada al igual
que ocurre en las fibras musculares de la rana, un proceso que puede ser
explicado por una inactivación dependiente del voltaje. El valor promedio de la
inactivación fue similar al valor obtenido en las fibras intactas (Sánchez y Stefani,
1983). En las fibras musculares intactas ha sido difícil determinar si ocurre un
proceso de inactivación (Sánchez y Stefani, 1983) o bien a un agotamiento de
cationes divalentes en la luz del túbulo (Almers, et al. 1981).
Dado que las vesículas carecen de membranas internas, estos resultados
apoyan a un proceso de inactivación dependiente del voltaje para explicar la
disminución observada en las corrientes.
El receptor a las dihidropiridinas
Nuestros resultados muestran que las vesículas contienen canales de
calcio tipo L. Esta conclusión de basa en el registro de corrientes entrantes de
Bario (ΙBa ) activadas por la despolarización y potenciadas por BAY K, un agonista
de las dihidropiridinas. Por otra parte estas corrientes son suprimidas por su
antagonista coma la nifedipina y bloqueadas por el Cobalto.
Movimiento de carga 1
Se encontró que las vesículas mantenidas en soluciones que carecen de
iones permeables presentan corrientes capacitivas no lineales activadas por
pulses despolarizantes.

59
Estas corrientes mostraron características similares al movimiento de carga
1 registrado en las fibras musculares (Chandler, Rakowski y Schneider, 1976), al
igual que en las fibras, las corrientes de carga se saturan cuando se aplican
grandes voltajes (para una revisión ver Ríos y Pizarro, 1991). La nifedipina
bloquea una cantidad importante del movimiento de carga de las vesículas,
apoyando la relación entre la carga y los DHPR (Ríos y Brum 1987).
El movimiento de las compuertas de otros canales sensibles al voltaje
puede tener poca participación en los registros obtenidos en las vesículas. La
apertura de canales de potasio en el músculo del anfibio, presenta alguna
similitud con el movimiento de carga. Sin embargo, coma fue revisado por Huang
(1993), la densidad de canales de K en el músculo es considerablemente menor
coma para explicar todo el movimiento de carga y dado que en las vesículas la
conductancia de K es considerablemente menor que en la fibra muscular
(Camacho, et al. 1996), la contribución de estos canales de K al movimiento de
carga registrado debe ser mínima. Por otra parte la apertura de canales de sodio
tiene un curso temporal mas rápido, por lo que es poco probable su participación
en el movimiento de carga registrado.
Se observaron algunas diferencias por ejemplo: el valor de Qmax fue mayor,
ya que se registraron valores entre 22 µC µF-1 y 38 ηC µF-1 en presencia de calcio
extracelular esto puede deberse a una mejora en la capacidad de registro por
parte del modelo experimental. Kim y Vergara (1998) recientemente mostraron un
incremento del 37% en el Qmax por la participación de componentes lentos que no
habían sido registrados anteriormente.
La diferencia mas importante en el movimiento de carga registrado en las
vesículas es la ausencia de “humps” identificadas coma Qmax las cuales fueron
descritas en las fibras musculares. Como se menciono en los antecedentes se ha
propuesto que este componente de la carga participa directamente en el proceso
del acople entre la excitación y la contracción o bien que representa la liberación
de calcio del retícula sarcoplásmico (García, et al. 1991). El presente trabajo no
provee evidencia directa que apoye a alguna de estas dos posibilidades ya que
las vesículas carecen de retícula sarcoplásmico por lo que no hay liberación de
calcio.
La disminución en la amplitud del componente rápido de las corrientes “off”
cuando la duración del pulso se incrementa puede deberse a los procesos de

60
inactivación de los canales de calcio. En realidad es de esperar que parte de la
corriente “off” represente el cierre o desactivación de canales de calcio. Conforme
la inactivación se presenta, menos canales están disponibles cuando las
vesículas son repolarizadas y la magnitud de los saltos en la corriente es más
pequeña, Sin embargo es probable que no todo los componentes rápidos
representen una corriente de Ba2+ ya que aun cuando los canales están
principalmente inactivados, la amplitud del componente rápidos es aun
significativamente mayor. Es posible que parte de los componentes rápidos de la
corriente “off” sea una corriente capacitiva no lineal.
Por otra parte, no se identifico el origen de los componentes lentos
observados en el movimiento de carga. En el músculo esquelético no se han
observado estos componentes posiblemente por que las lentas despolarizaciones
del sistema T limitan el registro de los componentes lentos.
Movimiento de carga 2
En las fibras musculares esqueléticas despolarizadas pueden registrarse
corrientes no lineales con pulsos hiperpolarizantes (Adrian y Almers, 1976) , el
cual corresponde a un movimiento de carga no lineal denominado Carga 2. Se ha
propuesto que esta carga representa al sensor del voltaje en el estado inactivado
(Brum y Ríos, 1987). La activación por el voltaje de la carga 2 ocurre a valores de
-74 a -108 mV (Brum y Ríos, 1987).
No se encontró evidencia de movimiento de carga 2 en las vesículas
despolarizadas. En realidad se observo que la relación entre la carga de las
espigas “off” y el voltaje fue lineal en todo el rango del voltaje probado. Esto
podría indicar la ausencia de la carga 2 o bien una dependencia extremadamente
baja al voltaje.
Se registraron corrientes no lineales en las vesículas despolarizadas pero
solo durante las espigas “on”. Estas corrientes son similares a aquellas descritas
por Brum y Ríos (1987) quienes propusieron que son llevadas por calcio, ya que
son eliminadas al quitar el calcio de la solución, sin embargo no se confirmó esta
dependencia ya que los registros en las vesículas son inestables en ausencia de
calcio. La naturaleza de esta conductancia aun no se ha identificado, sin
embargo los DHPR pueden no estar involucrados debido a que la nifedipina no la
afecta.

61
BIBLIOGRAFÍA
Adams, B. A., Tanabe, T., Mikami, A., Numa, S., y Beam, K. G. (1990).
lntramembrane charge movement restored in dysgenic skeletal muscle by
injection of dihydropiridine receptor cDNAs. Nature, Lond., 346, 569-572
Adelman, J. P. (1995). Proteins that interact with the pore-forming subunits of
voltage-gated ion channels. Curr. Coin. Neurobiol., 5,286-295.
Adrian, R. H. y Almers, W. (1976). Charge movement in the membrane of striated
muscle. J. Phvsiol. Lond., 254,339-360.
Adrian, R. H., Chandler, W. K., y Hodgkin, A. L. (1970). Voltage clamp
experiments in striated muscle fibres. J. Phvsiol. Lond., 208,607-644.
Affolter, H., y Coronado R. (1985). Agonists Bay-K 8644 and CGP-28392 open
calcium channels reconstituted from skeletal muscle transverse tubules.
Biophvs. J.. 48,341-347.
Almers, W., Fink, R., y Palade, P. T. (1981). Calcium depletion in frog muscle
tubules, the decline of calcium current under maintained depolarization. J
Phvsiol. Lond, 312,177-207.
Armstrong, C. M., y Bezanilla, F. (1973). Currents related to movement of the
gating particles of the sodium channels. Nature Lond., 242,459-461.
Arreola, JCalvo, J.Garcia, M.C. I. Sanchez, J.A. (1987). Modulation of calcium
channels of twitch skeletal muscle fibres of the frog by adrenaline and cyclic
adenosine monophosphate. J. Physiol., 393, 307-330.
Attwell, D., y Cohen, I. (1977). The voltage clamp of multicellular preparations.
Proq. Biophvs. Mol. Biol., 31,201-245.
Beam, K. G., y Knudson, C. M. (1988). Calcium currents in embrionic and
neonatal mammalian skeletal muscle. J. Gen. Phvsiol., 91,781-798.
Bechem, M., Hebisch, S. y Schramm, M. (1988). Ca++ agonists, new, sensitive
probes for channels. Trends Pharmacoloq. Sci., 9,257-261.
Bechem, M., y Pott, L. (1985). Removal of Ca current inactivation in dialysed
guinea-pig atrial cardioballs by Ca chelators. Pflueqers Arch., 404,10-20.
Beeler, G. W., Jr., y Reuter, H. (1970). Membrane calcium current in ventricular
myocardial fibres. J. Phvsiol. Lond., 207,191-209.

62
Belles, B., Malécot, C. O. Hescheler, J. y Trautwein, W. (1988). Run-down of the
Ca current during long whole-cell recordings in guinea pig heart cells, role
of phosphorylation and intracelular calcium. Pfluesers Arch. 411,353-360.
Berridge, M. J. (1997). The AM and FM of calcium signalling. Nature Lond., 386,
759-760.
Bolton, T. B. (1979). Mechanisms of action of trasmitters and other substances on
smooth mjuscle. Physiol. Rev., 59,606-718.
Brum G, y Rios E. (1987). lntramembrane charge movement in frog skeletal
muscle fibres. Properties of charge 2. J. Phvsiol., 387,489-517.
Burnashev, N. (1998). Calcium permeability of ligand-gated channels. Cell
Calcium, 24(5-6), 325-332.
Burnashev, N. A., Edwards, F. A., y Verkhratsky, A. N. (1990). Patch-clamp
recordings on rat cardiac muscle slices. Pluegers Arch., 417,123-125.
Camacho, J, D. M., Vazquez, M, Arguello, C, Sanchez, J. A. (1996). Transient
outward K+ channels in vesicles derived from frog skeletal muscle plasma
membranes. Biophys J., 71 (I), 171-181,
Campbell, K. P., Leung, A. T. y Sharp, A. H. (1988). The biochemistry and
molecular biology of the dihydropyridine-sensitive calcium channel. Trends
Neurosci., 11, 425-430.
Catterall, W. A., Seagar, M. J. y Takahashi, M. (1988). Molecular properties of the
dihydropyridine-sensitive calcium channel. J. Biol. Chem., 263, 3535-3538.
Cleemann, L., y Morad, M. (1991). Role of Ca2+ channel in cardiac excitation-
contraction coupling in the rat, evidence from Ca2+ transients and
contraction. J. Phvsiol. Lond., 432,283-312.
Cognard, C., Lazdunski, M. y Romey, G. (1986). Different types of Ca2+ channels
in mammalian skeletal muscle cells in culture. Proc. Natl. Acad. Sci. USA,
83,517-521.
Cota G, Nicola Siri L, Stefani E. (1983). Calcium-channel gating in frog skeletal
muscle membrane, effect of temperature. J. Phvsiol., 338,395-412.
Cota, G., Siri, L. N., y Stefani, E. (1984). Calcium channel inactivation in frog
(Rana pipiens and Rana moctezuma) skeletal muscle fibres. J.Physiol.
Lond., 354,99-l 08.

63
Cota, G., y Stefani, E., (1986). A fast-activated inward calcium current in twitch
muscle fibres of the frog (Rana montezuma). J. Physiol. Lond., 370,151-
163.
Chandler, W. K., Rakowski, R. F., y Schneider, M. F. (1976). A non-linear voltage
dependent charge movement in frog skeletal muscle. J. Physiol.. 254,243-
283.
Dirksen RT, Beam KG. (1995). Single calcium channel behavior in native skeletal
muscle. J. Gen. Phvsiol, 105(2),227-47
Feldmeyer, D., Melzer, W., Pohl, B. y Zollner, P. (1990). Fast gating kinetics of the
slow Ca2+ current in cut skeletal muscle fibres of the frog. J. Phvsiol. Lond.,
425, 347-367.
Fenwick, E. M., Marty, A. y Neher, E. (1982). Sodium and calcium channels in
bovine chromaffin cells. J. Physiol., 331,599-635.
Fleig A. y Penner R. (1995). Excessive repolarization-dependent calcium currents
induced by strong depolarizations in rat skeletal myoballs. J. Phvsiol.,
489,41-53.
Fleig A. y Penner R. (1996). Absence of Ca2+ current facilitation in skeletal
muscle of transgenic mice lacking the type 1 ryanodine receptor. J. Phvsiol.,
496,339-346.
Fleig A. y Penner R. (1996). Silent calcium channels generate excessive tail
currents and facilitation of calcium currents in rat skeletal myoballs. J.
Physiol., 494,141-153.
Flockerzi, V., Oeken, H. J., Hofmann, F., Pelzer, D., Cavalie, A., y Trautwein, W,
(1986). Purified dihydropyridine-binding site from skeletal muscle t-tubules
is a functional calcium channel. Nature Lond., 323,66-68.
Flucher BE, Morton ME, Froehner SC, Daniels MP. (1990). Localization of the
alpha 1 and alpha 2 subunits of the dihydropyridine receptor and ankyrin in
skeletal muscle triads. Neuron., 5(3),339-51.
Fosset M, Jaimovich E, Delpont E, Lazdunski M. (1983). [3H]nitrendipine
receptors in skeletal muscle. J. Biol. Chem., 258,6086-6092.
Francini, F. y Stefani, E. (1989). Decay of slow calcium current in twitch muscle
fibres of the frog is influenced by intracellular EGTA. J. Gen. Physiol.,
94,953-969.

64
Francini, F., Bencini, C. Y Squecco, R. (1996). Activation of L type calcium
channel in twitch skeletal muscle fibres of the frog. J. Physiol. Land,
494,121-140.
Francini, F., Pizza, L., y Traina, G. (1992). Inactivation of the slow calcium current
in twitch skeletal muscle fibres of the frog. J. Phvsiol. Lord, 448,633-653.
Gamboa-Aldeco, R., Huerta M., y Stefani, E. (1988). Effect of Ca2+channel
blockers on K+ contractures in twitch fibres of the frog (Rana pipiens). J-
Phvsiol. Lond, 397,389-399.
Garcia, J., McKinley,K., Appel, S. H., y Stefani , E. (1992). Ca2’currentand charge
movement in adult single human skeletal muscle fibres. J. Physiol. Lond.,
454,183-196.
Garcia, J., Pizarro, G., Rios, E. y Stefani , E. (1991). Effect of the calcium buffer
EGTA on the hump component of charge movement in skeletal muscle. J-
Gen. Physiol., 97(5),885-896.
Glossmann H, F. D., Lubbecke F, Mewes R. y Hofmann F. (1982).
CalciumChannel, direct identification with radioligand binding studies.
Trends Pharmacol Sci, 3,431-437.
Glossmann H, S. J. (1990). Molecular properties of calcium channels. Rev Physiol
Biochem Pharmacol., 114,1-l 05.
Goldin A. L. (1995). Voltage-gated sodium channels. En R. A. North (Ed.).
Handbook of receptors and channels, Ligand- and voltage-gated ion
channels (pp. 73-111). Boca Raton, Florida: CRC Press.
Hamill, 0. P., Marty, A., Neher, E., Sakmann, B. y Sigworth, F. J. (1981). Improved
patch-clamp techniques for high-resolution current recording from cells and
cell-free membrane patches. Pfluqers Archiv 391, 85-l 00.
Hamilton, S. L., Yatani, A., Hawkes, M. J., Redding, K., y Brown, A. M. (1985).
Atrotoxin, a specific agonist or calcium currents in heart. Science Wash. D.
C., 229,-l 82-184,
Heiny JA, Vergara J. (1984). Dichroic behavior of the absorbance signals from
dyes NK2367 and WW375 in skeletal muscle fibers. J. Gen. Phvsiol.,
84(5),805-37.
Herman ZS(1999). Application of calcium channel blockers in the treatment of
cardiovascular diseases. Pol. J. Pharmacol., 51(2), 201-206.

65
Hille, B. (1992). Ionic channels of excitable membranes. Massachusetts: Sinauer
Assoc. Inc. Sunderland.
Hodgkin, A. L., Huxley, A. F. y Katz, B. (1949). Ionic currents underlaying activity
in the giant axon of the squid. Arch. Sci. Physiol., 3,129-150.
Hodgkin, A. L., y Huxley, A. F. (1952). A quantitative description of membrane
current and its application to conduction and excitation in nerve. J. Physiol.
Lond., 117,500-544.
Hofmann, F., Lacinova, L. y Klugbauer, N. (1999). Voltage-dependent calcium-
channels, from structure to function. Rev. Phvsiol. Biochem. Pharmacol.,
139,33-85.
Hosey MM, Barhanin J, Schmid A, Vandaele S, Ptasienski J, O’callahan C,
Cooper C, y Lazdunski M. (1987). Photoaffinity labelling and
phosphorylation of a 165 kilodalton peptide associated with dihydropyridine
and phenylalkylamine sensitive calcium channels. Biochem. Biophvs, Res.
Commun., 147, 1137-l 145.
Huang CL. (1993). Charge inactivation in the membrane of intact frog striated
muscle fibers. J. Physiol., 468,107-24.
Hymel, L., Striessnig, J., Glossmann, H., y Schindler H. (1988). Purified skeletal
muscle 1, 4-dihydropyridine receptor forms phosphorylation-dependent
oligomeric calcium channels in planar bilayers. Proc. Natl. Acad. Sci. USA,
85,4290-4294.
Isenberg, G., y Klockner, U. (1980). Glycocalyx is not required for slow inward
calcium current in isolated rat heart myocytes. Nature Lond., 284,358-360.
Isom, L. L., De Jongh, K. S., y Catterall, W. A. (1994). Auxiliary subunits of
voltage-gated ion channels. Neuron, 12,1183-l 194.
Jack, J. J. B., Noble, D. y Tsien, R. W. (1983). Electric current flow in excitable
cells. Oxford: Oxford University Press.
Jorgensen AO, Shen AC, Arnold W, Leung AT, Campbell KP. (1989). Subcellular
distribution of the 1,4-dihydropyridine receptor in rabbit skeletal muscle in
situ, an immunofluorescence and immunocolloidal gold-labeling study. J-
Cell. Biol., 109(1),135-47.
Kim AM, Vergara JL. (1998). Fast voltage gating of Ca2+ release in frog skeletal
muscle revealed by supercharging pulses. J. Physiol., 511 ( Pt 2),509-18

66
Kokubun, S., Prod’hom, B,, Becker, C., Porzig, H., y Reuter, H. (1987). Studies on
Ca channels in intact cardiac cells, Voltage-dependent effects and
cooperative interactions of dihydropyridine enantiomers. Mol. Pharmacol.,
30,571-584.
Lamb GD. (1987). Asymmetric charge movement in polarized and depolarized
muscle fibres of de rabbit. J. Physiol. Lond., 383,349-367.
Lamb, G. D. (1992). DHP receptors and excitation-contraction coupling. J. Muscle
Res. Cell Motil., 13(4), 394-405.
Lamb, G. D. y Walsh, T. (1992). Calcium currents, charge movement and
dihydropyridine binding in fast- and slow- twitch muscle of rat and rabbit. J-
Phvsiol., 393, 595-617.
Lu X, X. L., Meissner G. (1994). Activation of the skeletal muscle calcium release
channel by a cytoplasmic loop of the dihydropyridine receptor. J. Biol.
Chem., 269(g), 651 I-651 6.
Ma, J., Mundiiia-Weilenmann, C., Hosey, M. M., y Rios E. (1991).
Dihydropyridine-sensitive skeletal muscle Ca channels in polarized planar
bilayers. 1. Kinetics and voltage dependence of gating. Biophvs. J., 60,890-
901.
Marty, A., y Neher, E. (1995). Tight-seal whole-cell recording. En, B. Sakmann y
E. Neher (Eds.). Single channel recording (pp 31-52). New York: Plenum
Press.
Matteson, D. R. y Armstrong, C. M. (1986). Properties of two types of calcium
channels in clonal pituitary cells. J. Gen. Phvsiol., 87,161-l 82.
McDonald, T. F., Pelzer, S., Trautwein W. y Pelzer D. J. (1994). Regulation and
modulation of calcium channels in cardiac, skeletal, and smooth muscle
cells. Physiol. Rev., 74(2),365-507.
McGoon M.D., Vlietstra, R. E., Holmes, D. R. Jr, &born, J. E. (1982). The clinical
use of verapamil. Mayo Clin. Proc., 57(8), 495-510.
Mejia-Alvarez, R., Fill, M., y Stefani, E. (1991). Voltage dependent inactivation of
T-tubular skeletal calcium channels in planar lipid bilayers. J. Gen. Physiol.,
97,393-412.
Melzer, W., Schneider, M. F., Simon, B. J., y Szucs, G. (1986). lntramembrane
charge movement and calcium release in frog skeletal muscle. J. Physiol.
Lond., 373,481-511.

67
Mori, Y. (1994). Molecular biology of voltage-dependent calcium channels. En
C.Peracchia (Ed.). Handbook of membrane channels (pp 163-176). San
Diego: Academic Press.
Muino Miguez A, L. G.-C. C., Villalba Garcia MV, Segado Soriano A, Granda
Martin MJ, Gil Gomez J. (1999). Fixed-dosage combinations in the
treatment of hypertension. An. Med. Interna, 16(2), 92-96.
Nakai, J., Adams, 6. A., Imoto, K., y Beam, K.G. (1994). Critical roles of the S3
segment and S3S4 linker of repeat I in activation of L-type channels. Proc.
Natl. Acad. Sci. U. S. A., 91, 1014-1018.
Nicola Siri, L., Sanchez, J. A. y Stefani, E. (1980). Effect of glycerol treatment on
the calcium current of frog skeletal muscle. J. Physiol. Lond., 305,87-96.
Peachey, L. D. (1965). The sarcoplasmic reticulum and transverse tubules of the
frog’s sartorius. J. Cell Biol., 25,209-231.
Pelzer, D., Grant, A. 0, Cavalie A., Pelzer, D., Sieber, M., Hofmann, F., y
Trautwein, W. (1989). Calcium channels reconstituted from the skeletal
muscle dihydropyridine receptor protein complex and its a’ peptide subunit
in lipid bilayers. Ann. NY Acad. Sci., 560,138-154.
Pitt B. (1997). Diversity of calcium antagonists. Clin Ther, lS(Suppl A), 3-17.
Randall A. D. (1998). The molecular basis of voltage-gated Ca2+ channel
diversity, is it time for T? J. Membr. Biol., 161(3), 207-213.
Rios, E. y Brum, G. (1987). Involvement of dihydropyridine receptors in excitation
contraction coupling in skeletal muscle. Nature Lond., 325,717-720.
Rios, E., y Pizarro, G. (1991). Voltage sensor of excitation-contraction coupling in
skeletal muscle. Physiol. Rev., 71,849-908.
Rosenberg, R. L., Hess, P., Reeves, J. P., Smilowitz, H., y Tsien R. W. (1986).
Calcium channels in planar lipid bilayers, insights into mechanisms of ion
permeation and gating. Science Wash. D.C., 231 ,I 564-1566.
Sanchez, J. A., y Stefani, E. (1983). Kinetic properties of calcium channels of
twitch muscle fibres of the frog. J.Phvsiol. Lond., 337,1-17.
Schneider, M. F. y Chandler, W. K. (1973). Voltage-dependent charge in skeletal
muscle, a possible step in excitation-contraction coupling. Nature Lond.,
342, 244-6.

68
Smith, J. S., McKenna, E. J., Ma, J., Vilven, J., Vaghy, P. L., Schwartz, A., y
Coronado R. (1987). Calcium channel activity in a purified dihydropiridine-
receptor preparation of a skeletal muscle. Biochemestry, 26,7182-7188.
Snutch, T. P., Leonard, J. P., Gilbert, M. M., Lester, H. A. y Davidson, N. (1990).
Rat brain expresses a heterogeneous family of calcium channels. Proc.
Natl. Acad. Sci. U. S. A, 87,3391-3395
Soong TW, Stea A, Hodson CD, Dubel SJ, Vincent SR, Snutch TP.(1993).
Structure and functional expression of a member of the low voltage-
activated calcium channel family. Science., 260(511 1),1133-6.
Standen NB, Stanfield PR, Ward TA, Wilson SW. (1984). A new preparation for
recording single-channel currents from skeletal muscle. Proc. R. Sot. Lond.
B Biol. Sci., 221(1225),455-64.
Stanfield, P. R. (1977). A calcium dependent inward current in frog skeletal muscle
fibres. Pfluqers Arch., 368;267-270.
Stea, A., Soong T. W. y Snutch T. P. (1995). Voltage-gated calcium channels. En
R. A. North (Ed.). Handbook of receptors and channels, Ligand- and
voltage-gated ion channels (pp 113-l 51). Boca Raton, Florida: CRC Press.
Talvenheimo, J. A., Worley III J, F., y Nelson, M. T. (1987). Heterogeneity of
calcium channels from a purified dihydropyridine receptor preparation.
Biophvs. J., 52,891-899.
Tanabe, T., Adams, B. A. Niidome, T. Numa, S. (1990a). Regions of the skeletal
muscle dihydropyridine receptor critical for excitation-contraction coupling.
Nature Lond., 346(6284), 567-569.
Tanabe, T., Numa, S., Beam, K. G. (1990b). Cardiac-type excitation-contraction
coupling in dysgenic skeletal muscle injected with cardiac dihydropiridine
receptor cDNA. Nature Lond., 344(6265), 451-453.
Tanabe, T., Takeshima, H., Mikami, A., Flockerzi, V., Takahashi, H., Kangawa, K.,
Kojima, M., Matsuo, H., Hirose, T. y Numa, S. (1987). Primary structure of
the receptor for calcium channel blockers from skeletal muscle. Nature
Lond., 328, 313-318.
Tanabe. T. B. K., Powell, J. A. Numa, S. (1988). Restoration of excitation
Contraction coupling and slow calcium current in dysgenic muscle by
dihydropyridine receptor complementary DNA. Nature Lond., 336, 134-9.

69
Trump, 9. F. y Berezesky, I. K. (1995). Calcium, cell death, and tumor promotion.
Proq. Clin. Biol. Res, 391, 121-l 31.
Tsien, R. W. y Tsien R. Y. (1990). Calcium channels, stores and oscillations”
Annu. Rev. Cell biol., 6,715760.
Tsien, R. W., Ellinor, P. T., y Horne W. A. (1991). Molecular diversity of voltaje-
dependent Ca2+ channels. Trends Pharmacol. Sci., 12,349-354.
Vaghy, P. L., Striessnig, J., Miwa, K., Knaus, H. -G., ltagaki, K., McKenna, E.,
Glossmann, H., y Schwartz, A. (1987). Identification of a novel 1, 4-
dihydropyridine- and phenylalkylamine-binding polypeptide in calcium
channel preparations. J. Biol. Chem., 262,14337-14342.
Valdivia, H. H., y Coronado, R., (1990). Internal and external effects of
dihydropiridines in the calcium channel of skeletal muscle. J. Gen. Phvsiol.,
95,1-27.
Vergara, J. Y Bezanilla, F. (1981). Optical studies of E-C coupling with
potentiometric dyes. En A. D. Grinnell y M. A. 9. Brazier (Eds.). Regulation
of muscle contraction . Excitation contraction coupling (pp 66-77). Nueva
York: Academic Press.
Wonderlin, W. F., Finkel, A., y French, R. J. (1990). Optimizing planar lipid bilayer
single-channel recordings for high resolution with rapid voltage steps.
Biophys. J., 58,289-297.
Yuan, X. -J., Goldman, W. F., Tod, M. L., Rubin, L. J., y Blaustein, M. P. (1993).
Ionic currents in rat pulmonary and mesenteric arterial myocytes in priamry
culture and subculture. Am. J. Physiol., 264 (2 pt l),L107-L115.
Zhang, J. -F., Randall, A. D., Ellinor, P. T., Horne, W. A., Sather, W. A., Tanabe,
T., Schwartz, T. L. y Tsien, R. W (1993). Distinctive pharmacology and
kinetics of cloned neuronal Ca channels and their possible counterparts in
mammalian CNS neurons. Neuropharmacology, 32,1075-l 088