estudio de la fracciÓn proteica de leche y fÓrmulas
TRANSCRIPT

UNIVERSIDAD AUTÓNOMA DE MADRID
Facultad de Ciencias
Departamento de Química-Física Aplicada
ESTUDIO DE LA FRACCIÓN PROTEICA DE
LECHE Y FÓRMULAS INFANTILES
SOMETIDAS A ALTAS PRESIONES
Fca Isabel Bravo Vázquez
Tesis doctoral
Junio 2012
CONSEJO SUPERIOR DE INVESTIGACIONES CIENTÍFICAS
UNIVERSIDAD AUTÓNOMA DE MADRID
INSTITUTO DE INVESTIGACIÓN EN CIENCIAS DE LA
ALIMENTACIÓN

UNIVERSIDAD AUTÓNOMA DE MADRID
Facultad de Ciencias
Departamento de Química-Física Aplicada
ESTUDIO DE LA FRACCIÓN PROTEICA DE LECHE Y FÓRMULAS INFANTILES SOMETIDAS A ALTAS PRESIONES
Memoria presentada por
FCA ISABEL BRAVO VÁZQUEZ
Para optar al grado de
Doctor en Ciencia y Tecnología de los Alimentos
Directoras:
Dras. Rosina López-Alonso Fandiño y Elena Molina Hernández
Consejo Superior de Investigaciones Científicas
Universidad Autónoma de Madrid
Instituto de Investigación en Ciencias de la Alimentación

CIAL Instituto de Investigación en Ciencias de la Alimentación
c/ Nicolás Cabrera, 9. Campus de Cantoblanco Universidad Autónoma de Madrid 28049 Madrid
ROSINA LÓPEZ‐ALONSO FANDIÑO, PROFESORA DE INVESTIGACIÓN y ELENA MOLINA
HERNÁNDEZ, CIENTÍFICA TITULAR, CON DESTINO EN EL INSTITUTO DE INVESTIGACIÓN EN
CIENCIAS DE LA ALIMENTACIÓN
INFORMAN:
Que el presente trabajo titulado “Estudio de la fracción proteica de leche y fórmulas infantiles
sometidas a altas presiones” y que constituye la Memoria que presenta Francisca Isabel Bravo
Vázquez para optar al grado de Doctora, se ha realizado bajo su dirección en el Departamento
de Bioactividad y Análisis de Alimentos del Instituto de Investigación en Ciencias de la
Alimentación (CIAL, CSIC‐UAM).
Y para que conste firmamos el presente informe a 8 de junio de 2012.
Fdo.: Rosina López‐Alonso Fandiño Fdo.: Elena Molina Hernández
VoBo Tutora:
Fdo.: Fco. Javier Señoráns

AGRADECIMIENTOS
Esta Tesis Doctoral ha sido desarrollada en el Departamento de Caracterización de Alimentos del Instituto de Fermentaciones Industriales (IFI, CSIC) y en el Departamento de Bioactividad y Análisis de Alimentos del Instituto de Investigación en Ciencias de la Alimentación (CIAL, CSIC-UAM). Durante la realización de la misma he disfrutado de un contrato de Técnico de Apoyo a la Investigación financiado por la Comunidad de Madrid.
Este trabajo ha sido realizado bajo la dirección de las Dras. Rosina López-
Alonso Fandiño y Elena Molina Hernández, a las cuales quiero agradecer la posibilidad que me han brindado de iniciarme en la investigación, trabajar en sus grupos, sus enseñanzas y orientaciones.
Mi agradecimiento y gratitud al Instituto de Fermentaciones Industriales y
al Instituto de Investigación en Ciencias de la Alimentación, a las directoras de los mismos, las Dras. Lourdes Amigo y Mª Victoria Moreno respectivamente, y a los equipos directivos por poner a mi disposición los medios técnicos y académicos necesarios para la realización de este trabajo. Mi agradecimiento al Dr. Fco Javier Señoráns por aceptar la tutoría de esta tesis y a las Dras. Mercedes Ramos, Mª Victoria Moreno y Nieves Corzo, jefas del Departamento de Caracterización de Alimentos del IFI y de Bioactividad y Análisis del CIAL durante este periodo, por su aceptación y apoyo para trabajar en su departamento.
El trabajo desarrollado ha contado con la ayuda y el suministro de muestras
de Laboratorio Ordesa S.A. También ha sido inestimable la colaboración de Xavier Felipe (y colaboradores) del IRTA.
Quiero agradecer a las Dras. Isidra Recio, Lourdes Amigo, Josefina Belloque,
Encarnación Pueyo y a los Dres. Adolfo Martínez e Iván López su apoyo y sus consejos. Y a Constanza por su constante amistad.
Agradecer de manera especial a mis compañeros de laboratorio su amistad,
sus consejos y por el buen ambiente del laboratorio. Este trabajo se lo dedico a mis padres, mi hermana y Juanma por haberme
ayudado a estar aquí, a seguir adelante y darme cariño aunque sea por teléfono.

ÍNDICE


ÍNDICE
RESUMEN
LISTA DE ABREVIATURAS
1. INTRODUCCIÓN
1.1. Las proteínas de la leche de vaca……………………………………………………….3
1.1.1. Las proteínas de suero………………………………………………………………3
1.1.2. Las caseínas………………………………………………………………………....6
1.1.3. La micela de caseína………………………………………………………………...8
1.1.3.1. Factores que afectan a la micela de caseína……………………………………...10
1.2. Nuevas técnicas de procesado de alimentos…………………………………………...11
1.2.1. Tratamientos con altas presiones…………………………………………………..12
1.2.1.1. Fundamentos de los tratamientos con alta presión…………………………….13
1.2.1.2. Equipos de alta presión……………………………………………………...…14
1.2.1.2.1. Tipos de equipos de alta presión…………………………………………...16
1.2.1.3. Efecto de la alta presión sobre las proteínas……………………………...……19
1.2.1.3.1. Efecto de la alta presión sobre las proteínas del suero……………………..19
1.2.1.3.2. Efecto de la alta presión sobre las micelas de caseína……………………..23
1.2.1.4. Aplicaciones tecnológicas de los tratamientos con alta presión…………….…28
2. OBJETIVO GENERAL Y PLAN DE TRABAJO……………………………………..35
3. MATERIALES Y MÉTODOS
3.1. Preparación de las muestras y tratamiento aplicado…………………………………...43
3.2. Obtención de fracciones proteicas……………………………………………………..43
3.3. Análisis de las muestras control y de las fracciones proteicas………………………....44
3.3.1. Determinación del contenido en nitrógeno………………………………………...44
3.3.2. PAGE-SDS………………………………………………………………………....44
3.3.3. Electroforesis capilar…………………………………………………………….…45
3.3.4. ELISA tipo sándwich………………………………………………………………46
3.3.5. Medida del tamaño de la micela de caseína………………………………………..47
3.3.6. Determinación del contenido en lactosa y lactulosa………………………….……48
3.3.7. Determinación del contenido en furosina………………………………………….49
3.4. Análisis estadístico……………………………………………………………………..50
4. RESULTADOS Y DISCUSIÓN
Capitulo I……………………………………………………………………………………..53
I.1. Introducción…………………………………………………………………………….55
I.2. Materiales y Métodos…………………………………………………………………..56

I.2.1. Preparación de las muestras y tratamientos aplicados……………………...…..…..56
I.3. Resultados……………………………………………………………………………....56
I.3.1. Efecto de los tratamientos UHT y con alta presión en las proteínas mayoritarias de la
leche………………………………………………………………………………………58
I.3.2. Efecto de los tratamientos UHT y con alta presión en las proteínas minoritarias del
suero…………………………………………………………………………………..…..63
I.3.3. Efecto del tratamiento UHT y de presión en el avance de la reacción de Maillard y la
isomerización de la lactosa……………………………………………………………….65
I.4. Discusión……………………………………………………………………………….66
I.5. Conclusiones……………………………………………………………………………68
Capítulo II…………………………………………………………………………………….71
II.1. Introducción…………………………………………………………………………....73
II.2. Materiales y Métodos………………………………………………………………….74
II.2.1. Preparación de las muestras……………………………………………………….74
II.2.2. Tratamientos de alta presión………………………………………………………75
II.3. Resultados……………………………………………………………………………..77
II.3.1. Caseínas no sedimentables y proteínas de suero presentes en la fracción soluble de
leche desnatada tratada con altas presiones………………………………………………77
II.3.2. Desnaturalización de las proteínas del suero en leche desnatada tratada por altas
presiones………………………………………………………………………………….82
II.3.3. Proteínas de suero desnaturalizadas y no sedimentables en suero de
ultracentrifugación tratado con altas presiones………………………………...…………86
II.4. Discusión………………………………………………………………………………88
II.4.1. Efecto del tipo de equipo de alta presión y de la velocidad de despresurización en la
liberación de caseínas no sedimentables a la fase soluble de la leche……………………88
II.4.2. Influencia de la desnaturalización de las proteínas de suero………………………91
II.5. Conclusiones…………………………………………………………………………..92
Capítulo III………………………………………………………………………………..…..93
III.1. Introducción…………………………………………………………………………..95
III.2. Materiales y Métodos………………………………………………………………....96
III.2.1. Preparación de las muestras………………………………………………………96
III.2.2. Tratamientos de alta presión……………………………………………………...97

ÍNDICE
III.3. Resultados………………………………………………………………………….....99
III.3.1. Solubilización de caseínas y desnaturalización de proteínas del suero en leche
desnatada tratada por altas presiones…………………………………………………..…99
III.3.2. Solubilización de caseínas y desnaturalización de proteínas del suero en leche
presurizada en presencia de agentes bloqueantes de grupos sulfhidrilo………………...108
III.4. Discusión…………………………………………………………………………….109
III.5. Conclusiones………………………………………………………………………...111
Capítulo IV…………………………………………………………………………………..113
IV.1. Introducción…………………………………………………………………………115
IV.2. Materiales y Métodos………………………………………………………………..116
IV.2.1. Preparación de las muestras……………………………………………………..117
IV.2.2. Tratamientos de presurización…………………………………………………..117
IV.2.3. Fraccionamiento y análisis………………………………………………………117
IV.3. Resultados………………………………………………………………………...…119
IV.3.1. Efecto de la presión en el tamaño de la micela de caseína……………………...119
IV.3.2. Efecto de la presión en la distribución proteica…………………………………121
IV.4. Discusión……………………………………………………………………………130
IV.5. Conclusiones………………………………………………………………………..134
5. CONCLUSIONES………………………………………………………………………135
6. BIBLIOGRAFÍA………………………………………………………………………..139

Resumen
En la actualidad, la industria alimentaria utiliza proteínas lácteas como
ingredientes en productos alimentarios, por su valor nutricional y, sobre todo, por sus
propiedades funcionales. Entre otros muchos productos, estas proteínas son utilizadas
para la elaboración de fórmulas infantiles. Aunque el mejor alimento para el recién
nacido es la leche materna, no siempre es posible la lactancia y es necesario recurrir a
fórmulas infantiles que sustituyan a la leche humana. Según el Codex Alimentarius, las
fórmulas infantiles deben promover el normal crecimiento y desarrollo del niño y
satisfacer, por sí solas, sus requerimientos nutricionales. Sin embargo, también se
persigue que, en la medida de lo posible, reproduzcan las propiedades beneficiosas que
caracterizan a la leche humana, muchas de las cuales, se deben a la fracción proteica.
Así, una ingesta adecuada de proteínas durante los primeros meses de vida, tanto en lo
que respecta a su contenido como a una elevada relación entre proteínas de suero y
caseínas, evita la ganancia excesivamente rápida de peso, facilita el proceso de digestión
y favorece una mayor colonización del intestino por bacterias probióticas.
En los últimos años el procesado con altas presiones hidrostáticas ha conseguido
un gran desarrollo comercial, ya que permite obtener productos con características
similares a los alimentos frescos, pero con una mayor vida útil. Además, las altas
presiones también modifican la estructura de las proteínas, conduciendo a propiedades
funcionales únicas o muy difíciles de conseguir mediante otros procesos. Estos cambios
dependen de factores tales como el nivel de presión, la duración del tratamiento, la
temperatura, el pH, etc. Actualmente, el efecto de la mayoría de estas variables se
conoce bien, pero rara vez se ha estudiado la influencia de parámetros como el tipo de
equipo de alta presión, la velocidad de despresurización o el empleo de presiones muy
elevadas, superiores a 600 MPa.
Esta tesis doctoral profundiza en la desnaturalización de las proteínas lácteas y
en los cambios en la distribución de proteínas que origina el tratamiento con altas
presiones hidrostáticas. El objetivo era explorar el potencial de las altas presiones para
mantener la calidad nutritiva de las proteínas de la leche y obtener fracciones lácteas de

diferente composición y concentración de proteínas, con aplicabilidad en la elaboración
de nuevas formulas infantiles.
Para desarrollar este estudio se obtuvieron diferentes fracciones lácteas a partir
de leche sometida a presiones comprendidas entre 250 y 900 MPa, en equipos a escalas
piloto e industrial, con diferentes tiempos de presurización y despresurización, y se
estudió su composición proteica mediante Kjeldahl, electroforesis capilar, SDS-PAGE
y ELISA, principalmente. Los resultados obtenidos han puesto de manifiesto que la
distribución de caseínas y seroproteínas en las fracciones estudiadas depende del equipo
utilizado, el nivel de presión y el tiempo de mantenimiento y/o liberación de la presión
aplicada.
Se ha comprobado que la aplicación de tratamientos con alta presión en distintos
equipos y a diferentes escalas, industrial y piloto, da lugar a diferentes distribuciones
proteicas, sobre todo en lo que se refiere a la desintegración micelar, puesto que los
diferentes equipos proporcionan similares grados de desnaturalización de las proteínas
de suero, a niveles de presión equivalentes. Además, la velocidad de despresurización
juega un papel fundamental en la distribución de proteínas entre las fases soluble y
coloidal de la leche presurizada, ya que el contenido en caseína soluble aumenta
significativamente durante la fase de despresurización respecto a la fase de
mantenimiento de la presión. Por su parte, la desnaturalización de la β-lactoglobulina
tiene lugar fundamentalmente durante la fase de mantenimiento de la presión,
produciéndose muy poca desnaturalización durante la fase de despresurización
En comparación con los tratamientos térmicos comúnmente empleados en la
industria, los tratamientos con alta presión no producen isomerización de la lactosa, ni
avance de la reacción de Maillard y conducen a una menor desnaturalización de las
proteínas mayoritarias y minoritarias del suero, por lo que podrían permitir la
estabilización de las fórmulas infantiles sin perjudicar su calidad nutritiva. También es
posible seleccionar las condiciones del tratamiento con alta presión que permitan
producir fracciones proteicas con una concentración de proteína entre el 1.05 y el 1.29
% y una relación de caseínas a proteínas de suero de, aproximadamente, 1.4-1.6, similar
a la de la leche humana, aunque contendrían una apreciable proporción de β-Lg
desnaturalizada soluble.

LISTA DE ABREVIATURAS
α-La: α-lactoalbúmina
β−Lg: β-lactoglobulina
4.6: Fracción soluble a pH 4.6
Arg: Arginina
BSA: Seroalbúmina bovina
Ca: Calcio
CCP: Fosfato cálcico coloidal
CG: Cromatografía de gases
CN: Caseína
Cys: Cisteína
DMF: N, N-Dimetilformamida
DTT: DL-ditiotreitol
EC: Electroforesis capilar
EDTA: Etilendiaminotetraacetato disódico
FITR: Transformada de Fourier
Fu: Furosina
g: Gramos
h: Hora
IgA: Inmunoglobulina A secretora
Igs: Inmunoglobulinas
IR: Tratamiento de presurización en un equipo industrial con despresurización rápida
kDa: Kilodaltons
L: Litro
La: Lactosa
Lf: Lactoferrina
LSD: Test de la menor diferencia significativa
Lu: Lactulosa
Lys: Lisina
Met: Metionina
MHEC: Metilhidroxietil celulosa
min: Minutos

MOPS: Ácido 3-morfolinopropanosulfónico
MPa: Megapascales
NEM: N-Etilmaleimida
NNP: Nitrógeno no proteico
OPD: Ortofenilendiamina
PAGE: Electroforesis en gel de poliacrilamida
PAGE-SDS: Electroforesis en gel de poliacrilamida en condiciones desnaturalizantes
PCS: Espectroscopía de correlación de fotones
Phe: Fenilalanina
pI: Punto isoeléctrico
PL: Tratamiento de presurización en un equipo piloto con despresurización lenta
PP: Polipropileno
PR: Tratamiento de presurización en un equipo piloto con despresurización rápida
Pro: Prolina
RP-HPLC: Cromatografía líquida de alta eficacia en fase inversa
SD: Desviación estándar
SDS: Dodecil sulfato sódico
Seg: Segundos
SU: Sobrenadante de ultracentrifugación
t: Tiempo
TCA: Ácido tricloroacético
Tris: Tris(hidroximetil)aminometano

1. INTRODUCCIÓN


INTRODUCCIÓN
- 3 -
- 3 -
Las proteínas son componentes fundamentales en la nutrición humana, no sólo por ser
fuente de nutrientes, sino porque su estructura y propiedades funcionales influyen en gran
medida en la calidad de los alimentos. La creciente demanda de productos con mejores
cualidades, tanto por su funcionalidad tecnológica y biológica como por sus características
organolépticas, ha hecho de las proteínas, sobre todo de las lácteas, una de las mejores
herramientas para desarrollar estos nuevos productos tan demandados.
En la actualidad, las proteínas lácteas son utilizadas como ingredientes multi-
funcionales ya que, además de ser solubles en un amplio rango de pH, de mejorar y de
estabilizar geles, emulsiones y espumas, su digestión da lugar a péptidos con actividades
fisiológicas, tales como antihipertensiva, antioxidante, etc (Barth y Schlimme, 1988; Bravo y
col., 2007, Martínez-Maqueda y col., 2012). Las propiedades derivadas de su estructura y de
las interacciones entre proteínas y otros elementos, así como la incidencia de diversos
procesos tecnológicos deben conocerse y estudiarse para aprovechar estas proteínas de la
manera más eficiente, en la mejora de los productos existentes o en el desarrollo de nuevos
productos.
1.1. Las proteínas de la leche de vaca
En la leche podemos identificar 2 clases de proteínas: las proteínas de suero y las
caseínas. Estas proteínas se separan mediante la acidificación de la leche a pH 4.6, el punto
isoeléctrico de las caseínas, que produce su precipitación. Sin embargo, existen otros métodos
para inducir su separación con mayor o menor precisión como la ultracentrifugación, la
centrifugación después de enriquecer con calcio, ultrafiltración y microfiltración a través de
diferentes tamaños de poro, precipitación por etanol o utilizando diferentes sales, etc.
1.1.1. Las proteínas de suero
Las proteínas de suero se encuentran presentes en la fracción soluble de la leche tras
su acidificación a pH 4.6 y representan el 20% de la proteína total. Dentro de esta
denominación se incluyen proteínas como la -Lactoglobulina (-Lg), -Lactoalbúmina (-
La), seroalbúmina bovina (BSA), lactoferrina (Lf) e inmunoglobulinas (Igs). La proteína
mayoritaria es la -Lg (50 % de la fracción proteica del lactosuero) con un masa molecular de
18.3 kDa (Figura 1). Es una proteína globular que posee 162 aminoácidos, de los cuales 5 son

- 4 -
cisteínas (Cys) implicadas en la formación de puentes disulfuro, uno localizado entre Cys66-
Cys160 y otro entre Cys106-Cys119 (Kilara, 2004). La conformación nativa de la proteína
consiste en 9 láminas- antiparalelas, de las cuales 8 forman una cavidad hidrofóbica (Zhai y
col., 2010). El grupo sulfidrilo libre de la Cys121 se encuentra en el interior de la molécula, por
lo que, durante la desnaturalización de la proteína, se expone al exterior, aumentando su
reactividad para formar puentes disulfuro y contribuyendo a la oligomerización de la proteína
(Considine, 2007).
Figura 1. Representación de la estructura tridimensional de la -Lg. Modelo obtenido mediante
el software RASMOL a partir de las coordenadas del Protein Data Bank (PD: 1BEB).
La -Lg puede aparecer en asociaciones diversas dependiendo de la temperatura y del
pH. A pH ácido (< 3.5) la proteína está en forma monomérica, mostrando una gran
estabilidad, aunque podrían formarse algunos dímeros (pH próximo a 3). Entre los pHs 3.5 y
5.2 se forman tetrámeros/octámeros reversibles que no conllevan cambios notables en la
estructura secundaria y aparece como un dímero en solución entre pH 5.2 y 7.5. El cambio de
monómero a dímero en el intervalo de pH entre 4.5 y 6 produce cambios en la compactación
de la proteína, sin embargo, a pH 7 se produce un cambio conformacional entre los residuos
85 y 90 de la proteína, producido posiblemente por una hendidura de los puentes de hidrógeno
entre las cadenas F y G. A este cambio conformacional se le denomina transición de Tanford.
A pHs superiores a 9 se produce una desnaturalización irreversible del monómero (Caessens y
col., 1997; Sakurai y Goto, 2007).
La temperatura es otro parámetro que puede incidir en la estructura y,
consecuentemente, en la funcionalidad de la proteína. La desnaturalización térmica de la -Lg
Lámina
-hélice
Lámina
-hélice

INTRODUCCIÓN
- 5 -
- 5 -
ocurre entre los 50-90ºC, pero es reversible siempre que la temperatura no supere los 65-70ºC
(Bertrand-Harb y col., 2002; Considine, 2007). Se ha propuesto que, a temperaturas entre 30 y
55ºC, la forma dimérica de la -Lg se disocia a monómeros. A temperaturas superiores, la
proteína se despliega incrementando la actividad y oxidación del grupo sulfidrilo (Hambling y
col., 1992) y produciéndose cambios estructurales irreversibles que pueden dar lugar a la
agregación de las moléculas cuando las temperaturas exceden los 70ºC (Bertrand-Harb y col.,
2002). La -Lg resiste en mayor medida la desnaturalización por calor a pH ácido, pero a pH
básico se desnaturaliza fácilmente (Caessens y col., 1997). La presión también afecta a su
estructura, produciéndose cambios reversibles a partir de 50 MPa e irreversibles a partir de
150 MPa (Tanaka y Kunugi, 1996).
La -lactoalbúmina representa el 25% de las proteínas del suero (Figura 2). Es una
molécula con 123 residuos aminoacídicos, cuatro puentes disulfuro y un masa molecular de
14.2 kDa (Considine y col., 2007). A diferencia de la -Lg, al no tener grupos sulfidrilo
libres, no forma homopolímeros (Monahan y col., 1993). Por su escasa estructura secundaria
(30% de -hélice y 9% de lámina-) (Alexandrescu y col, 1993) la molécula tiene una gran
flexibilidad. Sin embargo, la presencia de iones Ca2+ unidos a su estructura y los puentes
disulfuro la mantienen en una compacta estructura helicoidal con una zona interior hidrófoba
(Cayot y Lorient, 1997). La estabilidad en su conformación está relacionada con las
propiedades del puente de Ca2+. A pH <4, el calcio está libre y la -La se vuelve flexible y
puede proteolizarse fácilmente. Es altamente soluble incluso en su punto isoeléctrico (pI) y, ni
la fuerza iónica ni las variaciones de pH, influyen en la solubilidad de esta proteína
(Schlimme y Buchheim, 2002).
Figura 2. Representación de la estructura tridimensional de la -La. Modelo obtenido mediante
el software RASMOL a partir de las coordenadas del Protein Data Bank (PD: 1A4V).
Lámina
-hélice
Lámina
-hélice
Lámina
-hélice

- 6 -
Dentro del grupo de proteínas minoritarias de la leche se encuentran proteínas como
la BSA, Lf, transferrina, fibronectina, las inmunoglobulinas, enzimas, hormonas peptídicas,
etc. Cabe destacar la Lf y la IgA secretora (IgA), que juegan un papel muy importante en la
defensa del tracto gastrointestinal y de la mucosa de los recién nacidos. En concreto, la Lf
influye en la absorción del hierro, en la respuesta inmune y en la proliferación celular
(Lönnerdal, 2009) y la IgA en los mecanismos de defensa gastrointestinal (Bakker-
Zierikzee y col., 2006). La Lf es una glicoproteína fijadora de hierro con una masa
molecular de 82 kDa, que contiene dos lugares de unión para el hierro por cadena
peptídica. Su contenido en la leche es específico y diferente para cada especie y es máximo
en el calostro. La leche humana es, en comparación con la leche de vaca, rica en
lactoferrina. A mitad de la lactación, la leche humana y la de vaca contienen alrededor de
1.5 y 0.5 g/L, respectivamente (Schlimme y Buchheim, 2002). Las inmunoglobulinas
representan al menos el 2 % del total de las proteínas de la leche. Hay 4 tipos de Igs: IgG1,
IgG2, IgA y IgM.,con una estructura básica similar, compuesta por 2 cadenas ligeras con
masas moleculares entre 20-25 kDa, y 2 cadenas pesadas de 50-70 kDa (Kilara, 2004).
1.1.2. Las caseínas
Las caseínas son las proteínas de la leche insolubles a pH 4.6 y representan el 80% del
total de proteínas. Están constituidas por 4 proteínas, denominadas s1, s2, y κ-caseínas
(CN), que representan el 38, 10, 36 y 12% (3: 0.8: 3:1), respectivamente, del total de las
caseínas, con una masa molecular de 20-25 kDa (Schlimme y Buchheim, 2002). En general,
las caseínas tienen bajos niveles de estructura secundaria y terciaria, lo que les confiere gran
flexibilidad y una estructura fácilmente desnaturalizable por agentes como el calor o la urea y
susceptible a la acción de las enzimas. Los residuos hidrofóbicos, polares y cargados no se
distribuyen uniformemente en las moléculas sino que se concentran en zonas hidrofóbicas o
hidrofílicas, confiriendo a sus estructuras un fuerte carácter anfipático y una gran
hidrofobicidad superficial, responsable de sus propiedades espumantes y emulgentes. Estas
proteínas presentan gran heterogeneidad ya que, durante su formación en la glándula
mamaria, sufren cambios en sus estructuras primarias o modificaciones postraduccionales,
como fosforilación o glicosilación. En la leche, además, podemos encontrar fragmentos de las
anteriores producidos por la acción de diferentes enzimas endógenas en la leche, como la
plasmina, que es una serinproteinasa con gran afinidad por las uniones peptídicas de la lisina

INTRODUCCIÓN
- 7 -
- 7 -
(Lys) y, en menor medida, de la arginina (Arg). Todas las caseínas, excepto la -CN, son
sensibles a esta enzima (Fox y Kelly, 2004).
s1-CN
La s1-CN es la caseína mayoritaria de las micelas de caseína. Está compuesta por 199
aminoácidos, de los cuales ocho son residuos de serina fosforilados en las cadenas laterales, y
posee una masa molecular de 23.62 kDa. En cuanto a su estructura espacial, está formada por
un dominio hidrofóbico en el extremo carboxi terminal (100-199), compuesto
mayoritariamente por hoja beta, y por un dominio hidrofílico en el extremo amino terminal
(1-99). Además, precipita en presencia de iones calcio (Schlimme y Buchheim, 2002).
s2-CN
La s2-CN, al igual que la anterior, precipita en presencia deiones Ca2+. Contiene 207
aminoácidos y una masa molecular de 25.23 kDa. Además, tiene 2 moléculas de Cys en las
posiciones 36 y 40 (De Kruif, 2003).
-CN
La caseína está compuesta de 209 aminoácidos, de los que 35 son residuos de
prolina (Pro) distribuidos uniformemente en la molécula. Esta gran cantidad de prolina
previene la formación de -hélice, lamina y giros . También contiene 5 fosfoserinas en el
extremo N-terminal (la fosforilación es variable dependiendo de la variante genética) y
ninguna Cys. Es la caseína más anfipática de todas, con un extremo C-terminal fuertemente
hidrofóbico (Schlimme y Buchheim, 2002).
-CN
La -CN está compuesta por 169 aminoácidos y una masa molecular de 19kDa. En su
estructura tiene 2 Cys y entre 3 y 6 moléculas de galactosa, N-acetilglucosamina y ácido N-
acetilneuramínico unidas mediante enlaces O-glicosídicos a serina (Ser141) o treonina (Thr131, 133, 135 y 142). La región N-terminal es hidrofóbica y la C-terminal, que es hidrofílica, contiene
las variantes genéticas y las variaciones postraduccionales (los oligosacáridos y los grupos
fosfato) y posee una fuerte carga neta negativa (Fox y Kelly, 2004).

- 8 -
Las caseínas sensibles al Ca precipitan a concentraciones de Ca <6mM. Sin embargo,
en la leche se disponen en micelas lo que les permite ser estables a concentraciones 30 mM de
Ca. La principal responsable de la estabilización de la micela de caseína es la -CN que puede
unir de 8 a 10 veces su peso de caseínas sensibles al Ca (Fox y Kelly, 2004; Fox y Brodkord,
2008).
1.1.3. La micela de caseína
En la leche, las caseínas forman asociaciones coloidales llamadas micelas, con un
tamaño comprendido entre 50-300 nm (media 120 nm) y una masa molecular entre 106 y 3 x
109 Da (media 108 Da) (Fox y Kelly, 2004). Las micelas están altamente hidratadas (2-3 g de
H2O/g de proteína) y contienen, aproximadamente, un 6% de materiales inorgánicos
principalmente calcio y fosfato, que se conocen como fosfato cálcico coloidal (CCP) (Lucey y
col., 1996). Estas micelas confieren a la leche una serie de propiedadesmuy importantes
tecnológicamente, como su color, estabilidad al calor y al etanol, aptitud a la coagulación y
acidificación, etc.
A pesar de que las micelas de caseína se han estudiado durante muchos años, aún no
existe total consenso sobre la estructura de la micela. La mayoría de los modelos estructurales
propuestos coinciden en que la -CN está localizada mayoritariamente en la superficie,
jugando un papel esencial en la regulación del tamaño micelar y en el mantenimiento en
suspensión de las caseínas en la leche. La proporción de -CN varía en relación inversa con el
tamaño de la micela, mientras que la de -CN lo hace en forma directa (Riel, 1991; Ferrandini
y col., 2006).
El modelo más apoyado es aquel que define a la micela como una estructura formada
por la agrupación de submicelas más pequeñas, unidas mediante interacciones hidrofóbicas,
puentes de hidrógeno y/o CCP, recubiertas por una fina capa hidrofílica (5-10 nm). Esta capa,
que estaría formada por el extremo C-terminal de la κ-CN, con o sin otros extremos
hidrofílicos de otras caseínas (s1- y -CN) dependiendo del autor a seguir (Morr, 1967;
Waugh, 1971; Slattery, 1976; Schmidt, 1982; Walstra y Jenness, 1984), estabiliza a la micela
por medio del aumento del potencial zeta (-20mV). Además, el CCP también podría
localizarse dentro de la submicela (Figura 3) (Walstra y col., 1999).

INTRODUCCIÓN
- 9 -
- 9 -
Figura 3. Esquema del modelo de estructura de las micelas de caseína propuesto por Walstra
(1999).
Los últimos modelos propuestos retienen dos de las principales características del
modelo de la submicela, como son el papel que juega el CCP en la cementación de la micela y
el de la -CN estabilizando la micela por su localización en la capa superficial, pero difieren,
principalmente, en la composición de la estructura interna de la micela en forma de
submicelas (Visser, 1992; Holt, 1992; Horne 1998, 2002). En concreto, y entre otros, el
modelo propuesto por Visser (1992) presenta a la micela como conglomerados esféricos de
moléculas de caseína agregadas al azar y, el de Holt (1992, 1994), como una enredada red de
moléculas de caseína flexibles formando una estructura parecida a un gel (Figura 4).
Figura 4. Esquema del modelo de estructura de las micelas de caseína propuesto por Holt (1992).
Submicela
Fosfato cálcico
C-terminal de -CN
SubmicelaSubmicela
Fosfato cálcicoFosfato cálcico
C-terminal de -CNC-terminal de -CN
Caseínas
Fosfato cálcico
Caseínas
Fosfato cálcico

- 10 -
1.1.3.1. Factores que afectan a la estabilidad de la micela
Las micelas de caseína no se pueden considerar estructuras estables, sino que se
encuentran más bien en un equilibrio dinámico con el medio acuoso circundante, el suero
lácteo. La estructura y la estabilidad de las micelas de caseína se modifican, por ejemplo, a
consecuencia de las siguientes modificaciones (Fox y Kelly, 2004):
Las variaciones en la proporción entre el fosfato cálcico coloidal y el disuelto en el
suero modifican el tamaño de la micela. Si se acompleja el calcio libre, sin cambiar el
pH, o si se elimina mediante diálisis frente a una disolución libre de calcio, las micelas
se disgregan, y en casos extremos incluso dan lugar a fragmentos micelares.
El almacenamiento de leche cruda en frío durante un tiempo prolongado influye
especialmente en la -CN que, debido al debilitamiento de las interacciones
hidrofóbicas, se disocia de las micelas y pasa al suero lácteo. Sin embargo, este hecho
no conduce a una desestabilización profunda de las micelas y puede revertirse
parcialmente si se recalienta ligeramente la leche.
Al calentar la leche a temperaturas superiores a 70ºC se produce una agregación
creciente de las proteínas del suero, especialmente de la -Lg, con la κ-CN. Estos
complejos de caseína-proteína de suero confieren a las micelas de caseína propiedades
diferentes, por ejemplo en la coagulación por acidificación o por hidrólisis enzimática,
como se explica más adelante. A la particular interacción entre la -Lg y la κ-CN
micelar se le considera una propiedad tecnológica de extraordinaria importancia, ya
que permite esterilizar la leche sin que flocule a pesar de la casi completa
desnaturalización de las proteínas del suero.
La bajada del pH en la leche produce la solubilización del fosfato cálcico coloidal y,
por debajo de pH 5.2, las micelas de caseína empiezan a formar agregados que
precipitan en su pI (pH 4.6). Estos agregados se vuelven a disolver cuando el pH sube
a la neutralidad, aunque las micelas no vuelven a su estado natural. En la estructura de
la micela de caseína de la leche acidificada influye la temperatura, así como los

INTRODUCCIÓN
- 11 -
- 11 -
tratamientos térmicos previos a la acidificación, que producen la deposición de
proteínas séricas desnaturalizadas sobre la superficie de la micela.
Las micelas de caseína también pueden coagular por reacciones enzimáticas. La
quimosina escinde específicamente a la -CN a pH 6.7 en el enlace Phe105-Met106
dando lugar al caseinomacropéptido soluble (región C-terminal) y a la para--CN
insoluble. Cuando se ha escindido la mayor parte de la κ-CN, comienza la coagulación
de las micelas. El proceso de coagulación depende en gran medida de la temperatura y
de la concentración de iones calcio. Un precalentamiento intenso de la leche, que
conduce a la unión de proteínas del suero desnaturalizadas a la -CN que recubre a las
micelas, dificulta o impide el proceso de coagulación enzimática.
El tratamiento de las micelas de caseína a elevadas presiones hidrostáticas (por
ejemplo, 200 MPa, 5 min, 20ºC) produce su descomposición en subunidades o
pequeños agregados, como se explicará posteriormente.
1.2. Nuevas técnicas de procesado de alimentos
La utilización de altas temperaturas durante el procesado de los alimentos produce
daños en los componentes termolábiles como las proteínas, a las que causa desnaturalización
y agregación, además de pérdida de propiedades nutritivas y organolépticas. Sin embargo, el
procesamiento de los alimentos es necesario, ya que proporciona productos
microbiologicamente seguros y con una larga vida útil. Por ello, en los últimos 20 años, tanto
los científicos como la industria alimentaria, se han concentrado en estudiar y desarrollar
nuevas técnicas de procesado de alimentos que produzcan menores efectos en sus cualidades,
con el fin de obtener productos con características cada vez más similares a las de los
productos frescos, aunque con una mayor vida útil.
Estas tecnologías emergentes se pueden dividir en térmicas y no térmicas. El término
de procesado no térmico es utilizado normalmente para denominar aquellas tecnologías que
son efectivas a temperatura ambiente o subletal. Sin embargo, durante el procesado con estas
técnicas se pueden producir aumentos de temperatura, de modo que la eficacia del tratamiento

- 12 -
depende muchas veces de la combinación de ambos factores (Cullen y col., 2012). En la
Tabla 1 se muestran ejemplos de ambos tipos de tecnologías.
Tabla 1. Ejemplos de tecnologías emergentes de procesado y conservación de alimentos.
Tratamientos térmicos Tratamientos no térmicos
Altas presiones hidrostáticas
Pulsos eléctricos de alta intensidad
de campo
Irradiación
Microondas
Radio frecuencia
Calentamiento óhmico
Calentamiento inductivo Ultravioleta
Ultrasonido
Plasma frío
Ozono
Dióxido de carbono
Agua supercrítica
En la actualidad, muchas de estas técnicas ya son utilizadas en la industria alimentaria
como las altas presiones hidrostáticas, la irradiación, el ozono, etc. (Barbosa-Canovas y
Bermúdez-Aguirre, 2010).
1.2.1. Tratamientos con altas presiones
El tratamiento con altas presiones es una tecnología novedosa de procesado, de interés
comercial por su capacidad de aumentar la seguridad y la vida útil de los alimentos, sin
pérdidas indeseables de cualidades sensoriales y nutricionales.Un valor añadido de estos
tratamientos reside en su capacidad de modificar la funcionalidad y mejorar las propiedades
reológicas de éstos. En la actualidad, las altas presiones constituyen la tecnología más
desarrollada comercialmente, dentro de los métodos de conservación alternativos.
La tecnología de alta presión se utilizó originariamente en la producción de cerámicas,
aceros y superaleaciones. A finales del siglo XIX surge la aplicación de las altas presiones a

INTRODUCCIÓN
- 13 -
- 13 -
alimentos con Regnard (1884), Roger (1885) y Hite (1899) que aplicaron con éxito las altas
presiones a leche, carne y alimentos basados en frutas, para estudiar su efecto en la microbiota
de esos productos. No obstante, no fue hasta casi un siglo después cuando se comenzó a
aplicar esta técnica a alimentos a escala industrial, debido a que los problemas tecnológicos
que derivan de la aplicación de tan elevadas presiones supusieron un freno para el desarrollo
de esta técnica. Los primeros productos presurizados fueron mermeladas, salsas para
ensaladas, zumos y diferentes clases de yogures de frutas, que se comercializaron en 1992 en
Japón, probablemente por ser este país el pionero y líder en la fabricación de cámaras de alta
presión (Okpala y col., 2010).
Actualmente, la implantación de los equipos de alta presión en la industria alimentaria es
cada vez mayor (había 132 equipos de alta presión en el 2009 en todo el mundo, Barbosa-
Cánovas y Bermúdez-Aguirre, 2012), al igual que los productos tratados, utilizándose para
destruir microorganismos, inactivar enzimas o modificar proteínas. Además, en algunos
casos, las altas presiones permiten ejecutar y/o reducir el tiempo de algunas tareas, no
posibles o costosas de realizar por otros métodos de procesado, sin pérdidas de sus cualidades
nutricionales o sensoriales, por ejemplo, la abertura de las conchas de ostras u almejas (300
MPa, 1 min). La empresa japonesa Echigo Seika utiliza presiones entre 200-400 MPa para
modificar sin calor las proteínas y el almidón de algunos cereales, además de la velocidad de
penetración del agua en el grano. De esta manera, se obtienen cereales más dulces y
digestivos, que el consumidor prepara en 3 min al microondas, e incluso cereales
hipoalergénicos. Por otro lado, Fonterra comercializa calostro presurizado a 500 MPa
preservando compuestos bioactivos sensibles al calor (Lf, Igs y factores del crecimiento)
(Tonello, 2011).
1.2.1.1. Fundamentos de los tratamientos con alta presión
Se entiende por altas presiones la aplicación de presiones comprendidas entre 100 y
1000 MPa a un líquido en el que se sumergen los productos que van a ser sometidos al
tratamiento (Raventòs, 2005). Generalmente suele utilizarse agua debido a su baja
compresibilidad, de ahí que se le denomine también alta presión hidrostática. Esta técnica se
basa en dos principios (Cheftel 1991):

- 14 -
El primero indica que la presión se trasmite de manera uniforme e instantánea a
través de todo el material biológico, tratándose así de un proceso isostático. Esto
hace que el proceso sea homogéneo, evita la presencia de zonas sobretratadas y la
deformación del producto.
El segundo se refiere al principio de Le Chatelier, que indica que los fenómenos
acompañados de una disminución de volumen (reacciones químicas, modificación
de las conformaciones moleculares) son favorecidos por un aumento de presión y
viceversa. Según este principio la aplicación de alta presión desplaza el equilibrio
de las reacciones hacia el estado que ocupa menos volumen. En principio, en una
macromolécula proteica, la formación de enlaces de hidrógeno, la ruptura de
interacciones hidrofóbicas y de pares de iones se acompañan de una restricción de
volumen y se ven favorecidos por la presión (Cheftel, 1991). Sin embargo, otros
estudios suponen que los puentes de hidrógeno son prácticamente insensibles a la
presión (Mozhaev, 1994).
1.2.1.2. Equipos de alta presión
Los equipos de alta presión están formados por un recipiente donde se aplica la
presión (cámara de presión), un dispositivo de generación de presión, generalmente una
bomba hidráulica, y un sistema manual o automático para el control de las condiciones del
proceso (Zhou y col., 2010). El componente más importante es la cámara de presión y sus
cierres. El espesor de la pared de la cámara determina la presión máxima que puede alcanzar
el equipo, el diámetro del cilindro y el número de ciclos para los que se ha diseñado (Mertens
y Deplace, 1993). La cámara de presurización más simple es la monobloque, formada por un
cilindro de diámetro interno de menos de 15 cm. Generalmente el uso de estas cámaras está
limitado a presiones de trabajo inferiores a 400 MPa, porque su pared es delgada (Ting,
2011). Sin embargo, con el desarrollo de las paredes multicapas, de mayor espesor, se han
construido equipos de laboratorio capaces de alcanzar los 1400 MPa y con capacidad para 35
mL (como los fabricados por Stansted Fluid Power en el Reino Unido), además de equipos a
escala piloto e industriales. De forma general, ya que dependiendo del fabricante las
características de los equipos varían, el diámetro de las cámaras de los equipo piloto es de
unos 100 mm, con capacidad para unos pocos litros y pueden presurizar hasta 900 MPa en un
intervalo de temperatura ente 5 y 100ºC. Los equipos industriales, por otro lado, permiten

INTRODUCCIÓN
- 15 -
- 15 -
presurizar alimentos hasta 600-630 MPa, en el intervalo de temperatura de 5-30ºC. En este
caso, las cámaras tienen un diámetro entre 200 y 380 mm, albergando entre 55 y 420 L. En
los más pequeños (55 L) se ha podido aumentar la temperatura hasta 90ºC (NC Hyperbaric,
España).
Otro de los componentes a destacar es el sistema de cierre, que varía dependiendo de
la aplicación que se desee. Así, los cierres de rosca discontinua o de apertura y cierre rápidos
son idóneos para periodos cortos de presurización, ya que permiten minimizar el tiempo de
apertura y cierre y maximizar la productividad de la cámara (número de ciclos por unidad de
tiempo), lo cual es muy importante para la viabilidad económica del tratamiento de alta
presión. Sin embargo, cuando el tiempo de apertura y cierre de la cámara es despreciable en
comparación con la duración total del ciclo de procesado, se utilizan cierres de rosca continua,
más económicos (Raventòs, 2005).
Durante el tratamiento de presurización, la presión se transmite al alimento por medio
de un fluido transmisor, cuya elección varia dependiendo del material con el que esté
fabricada la cámara, ya que puede corroerla. Si es de acero inoxidable, permite el uso de agua
como medio transmisor. Sin embargo, pueden usarse otros líquidos como aceite de ricino,
aceite de silicona, mezclas de agua y glicol etc. La comprensión producida por la aplicación
de presión en el fluido transmisor y en el alimento produce el aumento de la temperatura
durante el tratamiento, es a lo que se le denomina calentamiento adiabático. El incremento en
la temperatura va a depender de la temperatura de partida y de la composición del alimento o
del fluido transmisor, variando dependiendo de sus características térmicas y físicas. A modo
de ejemplo, el calor de comprensión del agua es de 3ºC por cada 100 MPa, mientras que el del
aceite de silicona es de alrededor 20ºC por cada 100 MPa. La utilización de agua como fluido
transmisor es preferible ya que el comportamiento del calentamiento adiabático del agua es
similar al de la mayoría de los componentes de los alimentos, lo cual permitiría mimetizar el
gradiente térmico entre el fluido transmisor de la presión y el alimento. Además, el agua tiene
bajo coste, no es tóxico y se puede conseguir fácilmente (Nguyen y Balasubraniam, 2011).

- 16 -
1.2.1.2.1. Tipos de equipos de alta presión
Tras introducir el producto a presurizar dentro de la cámara, ésta se cierra y se llena
con el líquido transmisor de la presión. Llegados a este punto, unas bombas generan la
presión deseada y el producto se presuriza. Existen diferentes tipos de equipos de alta presión
dependiendo del producto a tratar: el tipo discontinuo, que se utiliza normalmente para
productos sólidos o líquidos previamente envasados, y el tipo semicontinuo para líquidos no
envasados.
Equipos de alta presión discontinuo
Son los más utilizados y se denominan así porque el producto a tratar, bien líquido o
sólido, está envasado cuando se introduce en la cámara de presión. La generación de la
presión puede hacerse de dos formas:
a) Por compresión directa, de modo que la presión se genera en el medio de trasmisión
por la parte final de un pistón. Este método permite una compresión muy rápida, pero
las limitaciones que impone el cierre dinámico entre el pistón y la superficie interna de
la cámara restringen su uso a diámetros pequeños, típicos de sistemas a escala de
laboratorio o a escala piloto (Figura 5).
b) Por compresión indirecta. Estos equipos utilizan un intensificador de alta presión para
bombear el medio trasmisor de la presión desde un tanque hasta la cámara, ya cerrada,
hasta conseguir la presión deseada. La mayoría de los sistemas industriales de presión
isostática utilizan este método de compresión (Figura 6) (Hogan y col., 2005;
Raventòs, 2005).
Los envases utilizados deben ser flexibles y se han de poder sellar herméticamente.
Algunos de los polímeros utilizados son el polietileno de baja densidad (LDPE), la goma
EVA (etilen vinil acetato), el polipropileno (PP) etc. (Balasubramaniam y col., 2004).
Además, debe evitarse la presencia de aire en el producto envasado, ya que la presencia de
éste puede incrementar el riesgo de ruptura del envase durante el tratamiento.

INTRODUCCIÓN
- 17 -
- 17 -
Figura 5. Esquema del diseño de un equipo de altas presiones con generación de presión por el
método de comprensión directa (adaptado de Raventòs, 2005).
Figura 6. Esquema del diseño de un equipo de altas presiones con generación de presión por el
método de comprensión indirecta (adaptado de Daoudi, 2004).
Tanque del mediode presión
Cierre inferior
Cierre superior
Cámara de presión
Medio de presión
Sistema de refrigeracióno calentamiento
Intensificador
Marco de la prensa
Tanque del mediode presión
Cierre inferior
Cierre superior
Cámara de presión
Medio de presión
Sistema de refrigeracióno calentamiento
Intensificador
Marco de la prensa
Bomba de baja presión
Sistema de refrigeracióno calentamiento
Cámara de presión
Medio de presión
Pistón
Marco de la prensa
Bomba de baja presión
Sistema de refrigeracióno calentamiento
Cámara de presión
Medio de presión
Pistón
Marco de la prensa

- 18 -
Equipos de tipo semicontinuo
Son utilizados con productos que pueden ser bombeados. En primer lugar, el alimento
se introduce en la cámara de presión, entrando en contacto directo con el acero de la cámara,
que debe ser adecuado para este fin y no experimentar corrosión durante su utilización .
Posteriormente, el alimento es comprimido por el líquido transmisor de la presión que está al
otro lado de un pistón. En estos equipos se utiliza una compresión directa para generar la
presión. Finalmente, el alimento presurizado es evacuado de la cámara a través de una válvula
aséptica de alta presión (Figura 7) (Balasubramaniam y col., 2008). Estos sistemas tienen
mejor rendimiento y mejor aprovechamiento de la energía utilizada, con el inconveniente de
un coste inicial mayor (Ting y Marshall, 2002; Hogan y col., 2005).
Figura 1.7. Esquema del diseño de un equipo de altas presiones de tipo semicontinuo (adaptado de
Daoudi, 2004).
De entre estos dos diseños, los más utilizados son los que presurizan alimentos
previamente envasados, ya que de esta manera se elimina el riesgo de contaminación del
producto con algún componente presente en el medio de presurización en de la maquinaria,
así como la posible contaminación de la cámara con el alimento y la necesidad de limpiarla
después de cada aplicación, disminuyendo, de esta manera, los costes de mantenimiento.
Tanque del mediode presión
Cierre inferior
Cierre superior
Cámara de presión
Medio de presión
Sistema de refrigeracióno calentamiento
Intensificador
Marco de la prensa
Producto a procesar
Producto procesado
Salida del mediode presión
Tanque del mediode presión
Cierre inferior
Cierre superior
Cámara de presión
Medio de presión
Sistema de refrigeracióno calentamiento
Intensificador
Marco de la prensa
Producto a procesar
Producto procesado
Salida del mediode presión

INTRODUCCIÓN
- 19 -
- 19 -
1.2.1.3. Efecto de la alta presión sobre las proteínas
Las altas presiones hidrostáticas producen la desnaturalización de las proteínas,
alterando el delicado equilibrio que existe entrelas interacciones que estabilizan la
conformación plegada de las proteínas nativas (Masson, 1992). Las dianas de los tratamientos
con alta presión son las interacciones hidrofóbicas y electrostáticas, cuya destrucción se
acompaña de una disminución de volumen. La formación de puentes de hidrógeno es casi
insensible a la presión, al igual que los enlaces covalentes que prácticamente no se modifican
hasta los 1000 MPa (Masson, 1992). La aplicación de alta presión induce cambios locales o
globales en la estructura de las proteínas, que pueden conducir a la desnaturalización
(Mozhaev y col., 1996). Presiones de 100-200 MPa son suficientes para provocar la
disociación de las proteínas poliméricas y de complejos multi-proteína (Silva y Weber, 1993),
mientras que las proteínas monoméricas suelen desnaturalizarse entre 400 y 800 MPa
(Heremans, 1982).
La exposición de residuos hidrofóbicos ocultos al agua y su resultante hidratación
durante el desplegamiento causa una disminución del volumen molar parcial de la proteína.
La hidratación de los grupos hidrofóbicos expuestos juega un papel decisivo en la estabilidad
termodinámica de la proteína y puede, además, ser considerada como un importante factor
para entender la modificación de su funcionalidad como resultado de los tratamientos de alta
presión. La disociación en subunidades y los desplegamientos localizados pueden exponer
grupos escondidos, que podrían unirse con otros grupos nuevamente expuestos y así producir
agregación (Masson, 1992).
1.2.1.3.1. Efecto de la alta presión sobre las proteínas de suero
Los estudios realizados sobre la desnaturalización por presión de las proteínas de suero
se centran en las proteínas mayoritarias, sobre todo en la -Lg y -La, ya que éstas son las
principales responsables de las propiedades de interés tecnológico. Los tratamientos con alta
presión producen la desnaturalización de las proteínas del suero. El efecto va a depender de
diferente factores, entre otros, la estructura de la proteína, la presión utilizada, el tiempo de
duración del tratamiento (López-Fandiño y col., 1996; Huppertz y col., 2004 a,b; Hinrichs y
Rademacher, 2004), la temperatura (Gaucheron y col., 1997; López-Fandiño y Olano, 1998;

- 20 -
García-Risco y col., 2000; Hinrichs y Rademacher, 2004; Huppertz y col., 2004b), el pH de la
leche (Arias y col., 2000; Huppertz y col., 2004a), etc.
De las proteínas mayoritarias del suero lácteo, la -Lg es la más sensible a los
tratamientos con alta presión (Patel y col., 2005). Estos tratamientos producen el
desplegamiento de la -Lg como se muestra en la Figura 8.
Figura 8. Cambios en la estructura de la -Lg presurizada (adaptada de Orlien y col., 2007).
A pH 4, la -Lg está en forma de dímero y al aplicarle presiones < 200 MPa se
produce la disociación de éste, lo que conlleva un aumento de la hidratación de la proteína. El
agua penetra en su interior y produce cambios reversibles en su conformación estructural. La
proteína está parcialmente plegada pero sin estructura terciaria específica (Orlien y col.,
2007). Panick y col. (1999) observaron mediante espectroscopía de infrarrojos por
transformada de Fourier (FITR) una disminución de la cantidad de lámina- y -hélice. A
presiones > 200 MPa, la desnaturalización de la proteína aumenta con el nivel de presión, ya
que se hidrata cada vez más, perdiendo su estructura terciaria a 500 MPa (se van rompiendo
los puentes de hidrógeno intramoleculares y se van formando interacciones intermoleculares
entre el agua y los nuevos residuos aminoacídicos expuestos) (Orlien y col., 2007), sin
embargo, la desnaturalización no es completa hasta los 1000 MPa (Panick y col., 1999).
Las altas presiones también pueden producir la agregación de la -Lg, ya que el
desplegamiento de su estructura ocasiona un incremento en la hidrofobicidad de la proteína,
aumentando de esta manera la interacción entre moléculas. Además, durante la presurización
se pueden dar oxidaciones de los grupos sulfidrilo o, más probablemente, reacciones de
intercambio sulfidrilo-disulfuro, en las que juega un papel importante el grupo sulfidrilo libre,
Presión (MPa)
-Lgnativa
-Lgdesnaturalizada
50 200 300 500Presión (MPa)
-Lgnativa
-Lgdesnaturalizada
50 200 300 500
-Lgnativa
-Lgdesnaturalizada
50 200 300 500

INTRODUCCIÓN
- 21 -
- 21 -
de esta proteína expuesto por efecto de la presurización, y que causan la agregación de la -
Lg (López-Fandiño y col., 1996; López-Fandiño, 2006a). Estos puentes disulfuro de nueva
creación son estables una vez que se elimina la presión.
La -La es menos sensible a la presión que la -Lg, debido a que su estructura
molecular es más rígida por la presencia de un mayor número de puentes disulfuro intra-
moleculares (4 frente a 2) y sitios de unión al Ca2+. Esta proteína empieza a desplegarse a
presiones de 200 MPa, aunque estos cambios son reversibles hasta presiones de 400 MPa. A
presiones comprendidas entre 200 y 400 MPa, la -La está en un estado intermedio de
desnaturalización, en la que la proteína mantiene su estructura secundaria, pero tiene una
estructura terciaria muy poco definida. A este estado de desnaturalización de la proteína se le
denomina en inglés molten globule (López-Fandiño, 2006b).
A diferencia de la -Lg, las altas presiones no producen la oligomerización de la -La
por si sola, incluso aunque se le apliquen largos periodos de presión. Sin embargo, en mezclas
de -La y -Lg, y debido al grupo sulfidrilo libre de esta última, la -Lg puede provocar la
oligomerización de la -La, llegando a producir la formación de una gran población de
oligómeros compuestos, presumiblemente, por varios cientos de moléculas de -La y -Lg
(Grinberg y Haertlé, 2000; López-Fandiño, 2006b).
En el caso de la BSA de leche bovina, López-Fandiño y col. (1996) indicaron que esta
proteína no se desnaturaliza a presiones entre 100 y 400 MPa. Esta gran baroestabilidad se
relaciona con el hecho de que la BSA tiene una estructura molecular extremadamente rígida
con 17 puentes disulfuro intramoleculares. Además, estudios realizados a 800 MPa mostraron
que la estructura secundaria de la proteína se ve sustancialmente afectada por esta presión,
aunque los cambios son mayoritariamente reversibles. Sin embargo, a muy altas presiones la
BSA puede también polimerizarse a través de uniones disulfuro mediadas por su grupo
sulfidrilo libre (López-Fandiño, 2006b).
En relación al efecto de las altas presiones sobre otras proteínas minoritarias, como Lf
e inmunoglobulinas, no existen estudios que indiquen el grado de desnaturalización o perdida
de solubilidad de éstas, por lo que queda patente la necesidad de determinar el daño que
pudieran sufrir por estos tratamientos.

- 22 -
Por otro lado, se ha observado que la desnaturalización de las seroproteínas producida
por la presión es mayor cuando se presuriza leche que suero de quesería (Huppertz y col.,
2004b). La presurización de la leche a ≥ 100 MPa, 25ºC produce una progresiva
desnaturalización de la -Lg, estimada mediante la pérdida de solubilidad a pH 4.6, mientras
que la -La y la BSA son resistentes hasta presiones de 400 MPa (López-Fandiño y col.,
1996). A presiones de 200 y 400 MPa, aplicadas a temperatura ambiente durante 15-30 min,
el porcentaje de desnaturalización de la -Lg es de 14-16 y 82-90%, respectivamente
(García-Risco y col., 2003; Huppertz y col., 2004b) mientras que a 600 MPa en las mismas
condiciones solamente se desnaturaliza un 15- 33 % de -La (Needs y col., 2000; Huppertz y
col., 2004b).
Además del nivel de presión, la temperatura es otro de los factores que influyen en el
grado de desnaturalización de las proteínas durante la presurización. López-Fandiño y Olano
(1998a) observaron que un incremento en la temperatura del tratamiento de alta presión hasta
60ºC no desnaturaliza la -Lg a 100 MPa pero, a presiones superiores, la desnaturalización se
incrementa a medida que aumenta la temperatura. También detectaron casi un 60 % de
desnaturalización de la -La tras tratamientos a 400 MPa y 60ºC.
El pH también es otro factor determinante. El pH alcalino favorece considerablemente
la desnaturalización de la -Lg como resultado del tratamiento de alta presión de la leche
debido a que aumenta la reactividad de los grupos sulfidrilo, mientras que el pH ácido reduce
la desnaturalización (Arias y col., 2000; Huppertz y col., 2004a).
Como ya se ha indicado, las proteínas de suero, -Lg y -La, son más susceptibles a la
desnaturalización inducida por la alta presión en la leche que en el suero, probablemente
debido a la ausencia de caseínas y a la menor concentración de calcio de este último (Hinrichs
y Rademacher, 2005; Huppertz y col., 2004b). Se ha sugerido la posibilidad de que la -Lg
desnaturalizada pueda unirse, mediante enlaces disulfuro, a la -CN (López-Fandiño y col.,
1997; Nabhan y col., 2004) y a la αS2-CN y así, formar agregados proteicos (Patel y col.,
2006). Además, el calcio podría facilitar el acercamiento de las proteínas de suero
desplegadas a otras proteínas a través de la neutralización de cargas. De hecho, se ha
observado que la desnaturalización de -Lg y -La inducida por presiones de hasta 800 MPa

INTRODUCCIÓN
- 23 -
- 23 -
se puede prevenir mediante la presencia de agentes bloqueantes de grupos sulfidrilo y reducir
mediante la eliminación de CCP (Huppertz y col., 2004b).
Así, en suero presurizado una importante proporción de las proteínas desnaturalizadas
permanece soluble, puesto que la concentración de caseínas es mínima (Huppertz y col.,
2004a). En la leche, la -Lg desnaturalizada por presión se encuentra asociada a la micela de
caseína, de manera análoga a lo que ocurre tras los tratamientos térmicos (Scollard y col.,
2000), aunque también puede permanecer en solución (García-Risco y col., 2003), ya sea en
forma de agregados formados exclusivamente por proteínas de suero o asociada a partículas
de caseína demasiado pequeñas para ser sedimentadas (López-Fandiño y col., 1998; Huppertz
y col., 2004a). Ye y col. (2004) también observaron algunas moléculas de -La y -Lg
asociadas a la membrana del glóbulo de grasa lácteo.
La asociación de la -Lg con las micelas de caseína inducida por las altas presiones
afecta a varias propiedades tecnológicas de la leche, por ejemplo, dificulta la coagulación
enzimática (Zobrist y col., 2005). Por otra parte, la interacción entre la -La y la -Lg puede
obstaculizar la aglutinación de los glóbulos de grasa inducida por frío y reducir la separación
de la grasa en la leche presurizada (Huppertz y col., 2006).
1.2.1.3.2. Efecto de las altas presiones sobre las micelas de caseína
Los tratamientos con alta presión pueden causar modificaciones sustanciales en las
micelas de caseína. De forma general, estos tratamientos producen la disgregación de la
micela de caseína, la cual se reorganiza en partículas más pequeñas e irregulares, dejando
libres pequeños fragmentos micelares o moléculas individuales de caseínas no
sedimentables,- junto con calcio y fósforo (López-Fandiño y col., 1998). Así, se modifica el
número, el tamaño y la composición de las micelas, lo que da lugar a cambios en las
propiedades ópticas de la leche, como disminución de la turbidez y cambios en la apariencia
visual (Needs y col., 2000a). Estas nuevas micelas generadas por la presión están más
hidratadas que sus predecesoras (Gaucheron y col., 1997; Huppertz y col., 2004c) ya que, por
un lado, la hidratación aumenta con la reducción del tamaño de la micela y es mayor para
partículas amorfas que para esféricas (Huppertz y col., 2006) y, por otro lado, la asociación a

- 24 -
éstas de la -Lg desnaturalizada aumenta la carga negativa y la solvatación micelar
(Huppertz y col., 2004c).
La disgregación micelar es consecuencia del debilitamiento de las interacciones
hidrofóbicas y electroestáticas que existen entre las caseínas (Johnston y col., 1992; Needs y
col., 2000a, b). Además, la ruptura de pares iónicos inducida por la presión altera la red de
CCP que estabiliza a la micela (Schrader y Buchheim, 1998; Huppertz y col 2004a, b). Así,
los tratamientos con altas presiones originan la solubilización del CCP, lo que conduce a un
aumento considerable de la concentración de minerales en la fase sérica (Huppertz y col.,
2006c). Entre 100 y 400 MPa se produce un incremento lineal en los niveles de Ca, P y Mg
(López-Fandiño y col., 1998; Hubbard y col., 2002), lo cual indica una progresiva
solubilización del CCP a medida que aumenta la presión, que conduce a la pérdida de la
integridad micelar. Parámetros como el nivel de presión, el tiempo de presurización, la
velocidad de despresurización, la temperatura y las condiciones del entorno (contenido en
calcio y caseína, pH) influyen en las propiedades de las micelas en la leche presurizada.
El nivel de presión es el parámetro que más influye en los cambios micelares. La
utilización de presiones < 200 MPa, a 20ºC , casi no modifica el tamaño de la micela de
caseína, aunque dependiendo del estudio se han indicado pequeños incrementos o reducciones
en su tamaño medio (Needs y col., 2000b; Huppertz y col., 2004a; Anema, 2008). Sin
embargo, sí se produce disgregación micelar, estimada a través del contenido en caseína no
sedimentable (Garcia-Risco y col., 2003), y aumento del volumen hidrodinámico de la micela.
Además, a partir de los 100 MPa comienza a disminuir la turbidez de la leche (Regnault y
col., 2004).
A partir de los 200 MPa, en tratamientos de 15 min, 20ºC, además de la disgregación
de la micela comienza a aparecer otro fenómeno, la agregación (Regnault y col., 2004). Ésta
se debe a que estas presiones intermedias (entre 250 y 300 MPa) favorecen las interacciones
hidrofóbicas respecto a la solvatación de los residuos hidrofóbicos expuestos. La asociación
se vería promovida, además, por la neutralización de las cargas negativas de las caseínas que
producen los iones calcio disueltos (Huppertz y De Kruif, 2006; Huppertz y col., 2006b), Por
esta razón, algunos autores han observado que el tamaño de la micela de caseína puede
aumentar a 250 MPa, aunque este hecho depende de varios factores, como la duración del
tratamiento, el pH y la temperatura, como se explica más adelante (Huppertz y col., 2004a).

INTRODUCCIÓN
- 25 -
- 25 -
Los tratamientos a presión >300 MPa reducen el tamaño de la micela de caseína, que
alcanza, a 400 MPa, aproximadamente la mitad del tamaño inicial (Gaucheron y col., 1997;
Needs y col., 2000b; Huppertz y col., 2004a, d, e), y no varía con el aumento de presión
(Anema y col., 2005a, Huppertz y col., 2004a), ni con posteriores presurizaciones (Huppertz y
col, 2004a). Estos cambios ocurren rápidamente y se acompañan de un importante aumento
en el nivel de caseínas de la fase sérica, que es máximo a presiones de 250-350 MPa (López-
Fandiño y col., 1998; Huppertz y col., 2004c). Las caseínas son liberadas durante la
disgregación micelar en el orden: -, -> S1-> S2-CN, el cual coincide con el número de
residuos por molécula capaces de unir CCP y, por tanto, de contribuir al mantenimiento de la
estructura micelar (López-Fandiño y col., 1998; Anema y col., 2005a). Medidas in situ han
demostrado que la máxima solubilización del CCP tiene lugar a 400 MPa, lo que coincide con
una disrupción micelar completa e irreversible (Huppertz y De Kruif, 2007a).Por otro lado, el
volumen de las micelas o subunidades resultantes es mayor, aproximadamente un 12%
superior que las micelas nativas (Regnault y col., 2004).
Dependiendo de la duración del tratamiento, los efectos de la alta presión en la micela
de caseína pueden variar. Los tratamientos a 250 MPa durante tiempos 5 min producen la
reducción del tamaño de la micela, aunque, posteriormente, éste va aumentando a medida que
se prolonga el tratamiento hasta los 40 min, momento en el que se alcanza el máximo
crecimiento (un 30% respecto al tamaño de la micela sin tratar), siendo a los 20 min cuando el
tamaño de la micela supera a la micela nativa. A presiones superiores (400 MPa) y tras sólo 1
seg de tratamiento, el tamaño de la micela se reduce (40% a 400 MPa) y tratamientos más
largos no reducen más el tamaño micelar (Huppertz y col., 2004a).
Existen muy pocos estudios que muestren el efecto de la velocidad de
despresurización en la micela de caseína, pero podría tratarse un factor importante, ya que
durante esta fase comenzarían a restablecerse interacciones electrostáticas e hidrofóbicas
destruidas durante el tratamiento con alta presión, originando la reasociación de las caseínas.
Se ha observado que la despresurización rápida de la leche (600 y 200 MPa/min) produce
micelas de caseína de menor tamaño y mayor volumen hidrodinámico que las obtenidas tras
una despresurización lenta (20 MPa/min) o sin tratar (Merel y col., 2007).
Al combinar presión y temperatura también se produce disgregación micelar, pero el
tamaño aumenta progresivamente a medida que aumenta la temperatura de presurización (20-

- 26 -
60ºC), presumiblemente porque ésta limite la ruptura de las interacciones hidrofóbicas o
promueva la agregación de las proteínas tras la descompresión (García-Risco y col., 2000;
Huppertz y col., 2004a). En concreto, en leche presurizada a 400 MPa durante 15 min el
tamaño medio, medido por microscopía electrónica de trasmisión, pasa de 38 a 135 nm al
cambiar la temperatura de presurización de 25 a 60ºC (García-Risco y col., 2000). Sin
embargo, a temperaturas inferiores (9ºC) y tras tratamientos a presiones ≤300 MPa no se
observa agregación, sino simplemente disgregación micelar, que aumenta a medida que sube
la presión, disminuyendo la turbidez de la leche (Regnault y col., 2004), y que conduce a un
tamaño micelar notablemente inferior al de la micela sin tratar (Huppertz y col., 2004). La
desintegración micelar inducida a bajas temperaturas, probablemente se debe a un
debilitamiento sinérgico de las interacciones hidrofóbicas (Regnault y col., 2004; Huppertz y
col., 2004a).
El pH también juega un papel importante en los cambios de tamaño de la micela de
caseína producidos por la alta presión. La aplicación de 250 MPa durante 30 min a leche
desnatada produce un aumento del tamaño de la micela (25 %) cuando el pH de la leche es de
6.7 y 7.0, disminuyendo posteriormente a medida que se incrementa el nivel de presión. Sin
embargo, a pHs de 6.5 no se produce este efecto, sino que el tamaño disminuye. Además, la
disminución del tamaño de la micela que se produce a presiones 300 MPa es mayor a pHs
más bajos (6.7 frente a 7.0) (Huppertz y col., 2004a). El pH bajo favorece la solubilización
del CCP y, por tanto, la destrucción de la estructura coloidal y el incremento en el contenido
de caseínas no sedimentables inducida por las altas presiones (Arias y col., 2000). Por el
contrario, a pH neutro o alcalino, la solubilización de CCP es limitada y ayuda a mantener la
estructura coloidal (Anema y col., 1997; Huppertz y col., 2004a), aunque la cantidad relativa
de todas las caseínas en la fracción del suero aumenta, probablemente a causa del aumento de
la carga negativa y la repulsión electrostática (Arias y col., 2000).
Todos los estudios presentados se han realizado en equipos de laboratorio o a escala
piloto, no existiendo actualmente evidencias del efecto que podría tener la presurización de
leche en equipos industriales. Dado que el diseño de estos últimos, incluyendo los sistemas de
liberación de la presión, termostatización etc., es diferente, no debe descartarse que pudiera
influir en el tamaño y la composición de la micela de caseína resultante o en la
desnaturalización de las proteínas séricas.

INTRODUCCIÓN
- 27 -
- 27 -
Algunos estudios sugieren que las proteínas de suero, desnaturalizadas por la presión,
podrían estar implicadas en los procesos de agregación proteica producidos por los
tratamientos con altas presiones. En concreto, la -Lg, al interaccionar con las micelas de
caseína mediante su unión con la -CN (Schrader y Buchheim, 1998; Huppertz y col., 2004a),
podría proteger a la micela de la disgregación (Kelly y col., 2002) o contribuir a su agregación
(Huppertz y col., 2004d). Sin embargo, estudios posteriores con sistemas de los cuales se
habían eliminado las proteínas de suero, mostraron que éstas influyen muy poco en los
cambios del tamaño de la micela inducidos por la presión, por lo que parece más probable que
el incremento de tamaño sea debido exclusivamente a la formación agregados caseínicos
(Anema y col., 2005a; Huppertz y col., 2007b). En concreto, Huppertz y col. (2007b)
observaron que tratamientos de presión de duración corta (≤10 min), aplicados a suspensiones
de caseínas micelares reconstituidas con adicción de proteínas de suero, producen micelas de
mayor tamaño que las obtenidas en el mismo sistema en ausencia de proteínas de suero. Sin
embargo, tras tratamientos de duración larga se elimina esta diferencia, lo cual apunta a una
influencia menor de las proteínas de suero en los cambios de la micela de caseína inducidos
por la alta presión.
La reversibilidad de los cambios micelares producidos por los tratamientos de alta
presión también ha sido objeto de estudio. Hasta 250 MPa, la mayoría de los cambios son
reversibles, sin embargo, a presiones superiores, su reversión depende del tiempo y la
temperatura de almacenamiento. Huppertz y col. (2004c) observaron que el aumento en el
tamaño micelar medio observado en leche tratada a 250 MPa, disminuye durante el
almacenamiento a 20ºC durante 24-48 h. En cambio, las partículas de pequeño tamaño que se
producen a mayores presiones son estables durante el mantenimiento posterior,
probablemente porque se necesite una estructura mínima que ejerza de núcleo para la
agregación, lo que se ve seriamente comprometido cuando la solubilización del CCP es muy
importante o completa (Huppertz y col., 2004c). En general, no se ha descrito reversibilidad
de los cambios producidos por las altas presiones durante almacenamientos a 5ºC
Gerhardt y col. (2006) explican en detalle como serían los cambios en la estructura
micelar producidos por la alta presión, a partir de sus estudios de espectroscopía de
correlación de fotones (PCS) in situ y microscopía de fuerza atómica (AFM) (Figura 9). A
presiones entre 100-250 MPa la micela se disgrega en mini-micelas, algunas de las cuales,
tienen un alto contenido en CCP, lo que les permite asociarse con otros fragmentos menos

- 28 -
ricos en CCP formando una nueva micela al cesar la presión. Esta nueva micela sería algo
más pequeña, más compacta y homogénea que la nativa, y en ella, las partículas ricas en CCP,
se encontrarían en el interior, mientras que las partículas hidrofóbicas, con menor contenido
en fosfato cálcico, se concentrarían en la superficie. Por lo tanto, el núcleo de esta micela
estaría estabilizado por uniones con el CCP, mientras que la superficie estaría dominada por
uniones hidrofóbicas entre las caseínas. Sin embargo, a presiones superiores a 280 MPa, las
micelas se disocian completamente en monómeros unitarios por la reducción de interacciones
electroestáticas y no-polares y aumenta la concentración de calcio libre y fosfato. Estos
monómeros se pueden reasociar, ya que el alto nivel de calcio libre induce agregación, pero
no se producirían micelas similares a la nativa, ni en tamaño ni en estructura, ya que la
disociación del CCP, necesario para formar la micela, a estas presiones es irreversible.
1.2.1.4. Aplicaciones tecnológicas de los tratamientos con alta presión
En los apartados anteriores se ha explicado que la alta presión reduce el tamaño de la
micela de caseína y produce disgregación micelar, desnaturaliza la estructura de las proteínas
de suero y altera el equilibrio mineral de la leche, en mayor o menor grado, dependiendo del
nivel de presión, el tiempo de tratamiento y el pH, entre otros factores. Estos cambios que se
producen en las proteínas mejoran sus propiedades funcionales y su aptitud tecnológica para
ciertos procesos, como la coagulación enzimática y ácida, por lo que pueden utilizarse en la
industria alimentaria para elaborar productos como quesos y yogures.
Coagulación enzimática y fabricación de quesos
La coagulación enzimática se basa en dos procesos interrelacionados, por un lado en la
hidrólisis específica de la κ-CN produciendo para-κ-CN y caseinmacropéptido y, por otro, en
la agregación de las micelas de caseína modificadas enzimaticamente (López-Fandiño,
2006a). Se ha observado que el tiempo de coagulación disminuye a presiones de hasta 200-
250 MPa, aumentando posteriormente, a medida que aumenta la presión aplicada, hasta
alcanzar tiempos similares a 400 MPa a los de la leche cruda, o superiores (15% más rápido a
600 MPa) (López-Fandiño y col., 1996; Needs y col., 2000b; Zobrist y col., 2005).

Presión normal(0.1 MPa)
Bajo presión(100-250 MPa)
Después de la despresurización(0.1 MPa)
100-200 nm 80-120 nm10-20 nmMicela nativa
100-200 nm 10-20 nm
Presión normal(0.1 MPa)
Bajo presión(280-400 MPa)
Después de la despresurización(0.1 MPa)
Figura 9. Representación esquemática de los cambios estructurales inducidos por la presión basado en el modelo de Walstra (1999) relacionado con los
modelos micelares descritos antes (adaptado de Gebhardt y col., 2006).

- 30 -
La alta presión acelera la coagulación enzimática de la leche debido a varios factores.
Por un lado, la disgregación de la micela libera caseínas a la fase soluble de la leche, entre
ellas la κ-CN, que es muy importante en la estabilidad de la micela. Se reduce así la carga
negativa y la repulsión electroestática en la micela. Además, la reducción de la capa
hidrofílica que recubre las micelas promueve las interacciones hidrofóbicas y la agregación
proteica (Johnston y col., 2002). Sin embargo, estos efectos se ven contrarrestados por la
desnaturalización de la β-Lg inducida por la presión. Esta proteína comienza a
desnaturalizarse a 200 MPa, su estructura comienza a desplegarse y el grupo sulfidrilo libre,
antes oculto en su estructura, se une a proteínas como la κ-CN (Patel y col., 2006), reduciendo
de esta manera la suceptibilidad de la κ-CN a hidrolizarse por la quimosina (López-Fandiño y
col., 1997). Además, la unión de ambas proteínas aumenta la repulsión electroestática, e
interfiere en la agregación micelar a presiones superiores a 250 MPa (López-Fandiño, 2006b).
Zobrist y col. (2005) observaron que el tiempo de coagulación de la leche no aumentó cuando
se presurizó a 400 y 600 MPa sin que hubiera proteínas de suero, o conteniendo KIO3, que
impide la unión de la -Lg a la micela de caseína.
Además de acortar el tiempo de coagulación, dependiendo del balance entre la
disgregación micelar y la desnaturalización de las proteínas de suero, los tratamientos con alta
presión dan lugar a cuajadas más firmes, hecho que se ha asociado a la disminución del
tamaño micelar (López-Fandiño y col., 1996). Las cuajadas con mayor firmeza se obtuvieron
en leches pasteurizadas a 65ºC durante 30 min con posterior tratamiento a 400 MPa, 15 min
(Molina y col., 2000).
En la fabricación de queso es también importante el rendimiento de la cuajada, en el
que influye la cantidad de proteínas desnaturalizadas asociadas a las caseínas y la cantidad de
agua retenida en la cuajada (Huppertz y col., 2004b). En la leche cruda, los tratamientos con
alta presión a 100-400 MPa aumentan el contenido en proteínas y la humedad de la cuajada y
mejoran su firmeza, mientras que se acelera o mantiene el tiempo de coagulación (López-
Fandiño y col., 1996). Ya se ha mencionado que la unión de la -Lg a las caseínas produce un
aumento de la carga neta negativa de la micela, y esto puede llevar asociado un aumento de la
solvatación (Imafidon and Farkye, 1996) y una reducción de la sinéresis (Smith and
McMahon, 1996). Además, la solvatación también aumenta a presiones entre 100 y 600 MPa
debido a la formación de submicelas o agregados de menor tamaño (Gaucheron y col., 1997;
Huppertz y col., 2004c). En cualquier caso, el aumento del rendimiento va estrechamente

RESULTADOS Y DISCUSIÓN: CAPÍTULO I
- 31 -
- 31 -
ligado a la desnaturalización de la -Lg: a presiones ≤ 250 MPa, el rendimiento de la cuajada
no varía (Huppertz y col., 2004f), mientras que tras tratamientos a presiones superiores, 300 y
400 MPa (30 min, 20-25ºC), se observaron aumentos del rendimiento del 14% y del 20%
respectivamente (López-Fandiño y col., 1996), que llegan a un 25% en leches presurizadas a
600-800 MPa (Huppertz y col., 2004f).
Algunos autores han estudiado la estructura de quesos elaborados a partir de leche
presurizada. Los quesos Cheddar elaborados a partir de leche presurizada a 586 MPa (3 ciclos
de 1 min) mostraron un contenido de humedad y un rendimiento mayor que los mismos
hechos con leche cruda o pasteurizada, aunque la textura fue pastosa y débil (Drake y col.,
1997). En quesos con bajo contenido en grasa fabricados a partir de leche pasteurizada (65ºC,
30 min) y posteriormente presurizada (400 MPa, 22ºC, 15 min) también se observó un
aumento del rendimiento frente a los elaborados con leche pasteurizada, al igual que una
proteolisis más rápida de las caseínas, lo que contribuyó a un desarrollo más rápido de la
textura, el aroma y el sabor (Molina y col., 2000). Además, el queso Fresco elaborado a partir
de leche presurizada (400 MPa, 20 min, 20ºC) fue más húmedo, menos firme, menos
quebradizo y más pegajoso (Sandra y col., 2004).
Coagulación ácida y fabricación de yogur
Los tratamientos con alta presión también favorecen la coagulación ácida. Factores
como la desintegración micelar, la pérdida de la estabilidad electroestática de la micela de
caseína cerca de pH 5.3 y la mayor frecuencia de colisión de las partículas influyen en la
calidad del coágulo formado por acidificación (Banon y Hardy, 1992; Schwertfeger y
Buchheim, 1998; Huppertz y col., 2002). En leches reconstituidas tratadas con presión, la
gelificación inducida por la acidificación se produce a pHs superiores que la producida en
leches no presurizadas, debido a los cambios inducidos por la presión en la capacidad
tamponadora (Desobry-Banon y col., 1994). Además, en leche bovina presurizada, la
acidificación producida por la inoculación con bacterias acido-lácticas se produce más
rápidamente, probablemente debido a que la rotura de la micela produce un aumento del
nitrógeno accesible a la bacteria (Huppertz y col., 2004g).
Algunos estudios demostraron que las propiedades de los geles formados por
acidificación mejoran al presurizar la leche. Por ejemplo, Johnston y col. (1993, 2002)

- 32 -
obtuvieron geles más rígidos, más fuertes y con mayor resistencia a la sinéresis, lo cual se
correlacionó con un aumento en la hidratación y en la densidad de la malla de proteína. La
agregación de pequeñas partículas micelares daría lugar a agregados compactos de menor
tamaño cuando se alcanza el punto isoeléctrico. Además, el hecho de que en su superficie se
encuentre -lg desnaturalizada contribuye a la firmeza del gel y a la resistencia a la sinéresis
(Harte y col., 2002).
Se ha observado que la firmeza de los yogures elaborados a partir de leche presurizada
aumenta a medida que aumenta la presión (Needs y col., 2000). Los yogures elaborados a
partir de leche presurizada a 55ºC, presentan una gran firmeza sin rotura micelar aparente, lo
cual pone de manifiesto la importancia de la desnaturalización de las proteínas de suero en
este proceso (Ferragut y col., 2000). Además, Harte y col. (2003) observaron que el uso de un
tratamiento térmico (85ºC, 30 min), antes o después de los tratamientos de alta presión, que
produce una amplia desnaturalización de las proteínas de suero, mejora las propiedades
reológicas y reduce la sinéresis. A este respecto debe indicarse que algunos estudios han
puesto de manifiesto que la firmeza optima se obtiene combinando la desnaturalización de las
proteínas de suero con una alta concentración de caseínas micelares disgregadas (Capellas y
Needs 2003).
Gelificación de la leche por la alta presión
Otro de los efectos que producen las altas presiones en la leche es la gelificación de
leche concentrada (66-114 g/ kg de proteína), en concreto, a presiones de 200 y 400 MPa. La
disminución del pH hacia 5.9 y el aumento del contenido proteico contribuyen a la formación
de geles de mayor firmeza (Famelart y col., 1998). Las propiedades relativas a la viscosidad,
viscoelasticidad y textura de los geles fabricados a partir de leche concentrada dependen del
nivel de presión, siendo la disociación micelar un requisito indispensable en la formación del
gel, mientras que la agregación de las proteínas del suero tiene un papel secundario (Keenan y
col., 2001). De hecho, la firmeza de los geles formados a partir de caseínas depende de la
velocidad de descompresión, a diferencia de los geles formados por proteínas de suero (Fertch
y col., 2003). La despresurización rápida produce geles más firmes, con una microestructura
más fina que los elaborados tras despresurizaciones lentas, lo que fue atribuido a que, una vez
que se produce la disociación de la micela de caseína durante la fase de mantenimiento de la
presión, una despresurización lenta favorecería el desarrollo de hiperestructuras caseínicas, al

RESULTADOS Y DISCUSIÓN: CAPÍTULO I
- 33 -
- 33 -
reformarse los puentes de calcio y las interacciones hidrofóbicas, lo que dificultaría una
gelificación homogénea (Fertsch y col., 2003). Sin embargo, Merel-Rausch y col. (2007), que
también encontraron geles de mayor firmeza a velocidades rápidas de liberación de la presión,
lo correlacionaron con la formación de grandes estructuras flexibles, construidas a partir de
micelas de caseína de menor tamaño.

2. OBJETIVO GENERAL Y
PLAN DE TRABAJO


OBJETIVO GENERAL Y PLAN DE TRABAJO
- 37 -
En la actualidad, la industria alimentaria utiliza frecuentemente proteínas lácteas como
ingredientes para productos alimentarios, por su valor nutricional y, sobre todo, por sus
propiedades funcionales. Entre otros muchos productos, estas proteínas son utilizadas para la
elaboración de fórmulas infantiles. Aunque el mejor alimento para el recién nacido es la leche
materna, no siempre es posible la lactancia y es necesario recurrir a fórmulas infantiles que
sustituyan a la leche humana.
Según el Codex Alimentarius, las fórmulas infantiles deben promover el normal
crecimiento y desarrollo del niño y satisfacer, por sí solas, sus requerimientos nutricionales.
El objetivo principal de los fabricantes no es obtener una fórmula que reproduzca fielmente la
composición de la leche humana, sino que, más bien, imite los efectos funcionales de la leche
materna. El conocimiento cada vez más exhaustivo de la importancia de algunos de sus
componentes, como citoquinas o factores de crecimiento, un adecuado balance de ácidos
grasos poliinsaturados (n-3/n-6) o la presencia de determinadas especies de bacterias
probióticas, a las que los oligosacáridos humanos (de los que se conocen más de 100
estructuras distintas) sirven como sustratos, ha conllevado a importantes avances en el
desarrollo de fórmulas infantiles mejoradas. También es sabido que muchas de las
propiedades beneficiosas de la leche humana se deben a la fracción de proteínas que, además
de proporcionar un aporte adecuado de nutrientes, contribuye a la defensa frente a infecciones
y facilita el óptimo desarrollo de funciones biológicas esenciales para el recién nacido. Sin
embargo, los requerimientos proteicos precisos son un tema controvertido (Lönnerdal, 2010).
Algunos autores señalan la importancia de una ingesta adecuada de proteínas durante
los primeros meses de vida, sobre todo, de la relación entre proteínas de suero/caseínas. La
presencia de caseínas en la leche humana es una de las menores comparada con la de otros
mamíferos. Así, mientras la leche bovina contiene unos 25 g/L, la leche humana contiene sólo
4.6 g/L (Ginger and Grigor, 1999). Un contenido bajo de proteínas y la predominancia de
proteínas de suero, que caracterizan a la leche humana, facilitaría el proceso de digestión e
incluso podría inducir una mayor colonización del intestino por bacterias probióticas. Por otra
parte, una ganancia rápida de peso durante la infancia se asocia con un mayor riesgo de
obesidad. De hecho, comparados con los niños alimentados con leche materna, los
alimentados con fórmulas ganan más peso durante la infancia. Esto se explica por una mayor
ingesta de sustratos metabolizables y, en particular, de proteínas: la ingesta de proteínas por

- 38 -
kg de peso corporal es del 55-80% mayor en los niños alimentados con fórmulas respecto a
los alimentados con leche materna (Koletko y col., 2009).
Durante la manufactura de las fórmulas infantiles, la aplicación de tratamientos
térmicos es imprescindible para garantizar su inocuidad. Sin embargo los procesos
convencionales afectan negativamente sus propiedades químicas y la biodisponibilidad de
nutrientes, incluyendo degradación de vitamina C, peroxidación de lípidos, formación de
agregados de lípidos y proteínas y formación de productos de la reacción de Maillard, que
disminuyen la disponibilidad de la lisina y tienen menor capacidad de unión a magnesio,
cobre, calcio y zinc, inhibiendo su absorción (Sarriá y col., 2001).
En los últimos 30 años se han desarrollado enormemente nuevas tecnologías de
procesado y conservación de alimentos, las llamadas tecnologías emergentes, si bien la alta
presión hidrostática es la única tecnología emergente que ha dado lugar a productos
comerciales, pudiéndose encontrar en los lineales de los supermercados una considerable
variedad de alimentos presurizados. La mayoría de las aplicaciones de la alta presión se basan
en su capacidad de destruir microorganismos sin afectar negativamente a las características
nutricionales de los alimentos. Pero, además, los tratamientos con alta presión pueden
modificar la estructura cuaternaria, terciaria y, en menor medida secundaria, de las proteínas,
conduciendo a propiedades funcionales únicas o muy difíciles de conseguir con otros
procesos. Así, como consecuencia de la desnaturalización de las proteínas de suero y de la
ruptura micelar, se forman nuevas estructuras proteicas, lo que, además de mejorar la aptitud
tecnológica de la leche para ciertos fines, podría permitir la obtención de fracciones con de
interés nutricional. En cualquier caso, la intensidad, la duración y la temperatura de los
tratamientos, así como, el pH y la concentración de proteína influyen en los cambios
originados por las altas presiones Actualmente, el efecto de la mayoría de estas variables se
conoce bien, pero rara vez se ha estudiado la influencia de parámetros como el tipo de equipo
de alta presión, la velocidad de despresurización o el empleo de presiones muy elevadas,
superiores a 600 MPa.
Esta tesis doctoral profundiza en el estudio de la desnaturalización de las proteínas de
suero de la leche, la desintegración micelar de las caseínas y en los cambios en la distribución
de estas proteínas que origina el tratamiento con altas presiones hidrostáticas. El objetivo era
explorar el potencial de las altas presiones para mantener la calidad nutritiva de las proteínas

OBJETIVO GENERAL Y PLAN DE TRABAJO
- 39 -
de la leche y obtener fracciones lácteas de diferente composición y concentración de proteínas
con aplicabilidad en la elaboración de nuevas formulas infantiles. Para desarrollar este
objetivo, nos centramos en los siguientes objetivos parciales:
o Estudio del efecto de las altas presiones hidrostáticas en las propiedades
nutricionales de fórmulas infantiles líquidas.
o Estudio del efecto del tipo de equipo de alta presión y del escalado en la distribución
proteica de diferentes fracciones lácteas.
o Estudio del efecto de la velocidad de despresurización en la distribución proteica de
diferentes fracciones lácteas.
o Estudio del efecto de presiones muy elevadas, entre 700, 800 y 900 MPa, aplicadas a
diferentes fracciones lácteas, sobre su distribución proteica.
Cada uno de los objetivos expuestos constituye un capítulo de la Tesis en el que se discute la
influencia de las distintas variables del tratamiento de alta presión en los cambios
encontrados.

- 40 -
Figura 10. Esquema del plan de trabajo desarrollado.
Mat
eria
pr
ima
Aná
lisis
Capítulo I Capítulo II Capítulo III Capítulo IV
Fórmula Infantil
(con o sin NEM o DMF)
UHT(142ºC, 4.7 seg)
Fact
ores
est
udia
dos
Tra
tam
ient
os a
plic
ados
Alta presión (HP)650 MPa, 15ºC,
5 min
HP0.1, 250, 350, 450, 550 y 650 MPa, 15ºC, 5min con
despresurización instantánea
Tipo de tratamiento: UHT v HP
Equipo utilizado:Industrial vs piloto
t mantenimiento presión:
5, 6, 10 y 15 min
t despresurización:0.07, 1, 5 y 10 min
HP0.1, 250, 450, 550,
700, 800 y 900 MPa, 15ºC, 5min
Nivel de presión:0.1- 650 MPa
t despresurización en equipo piloto:30 seg vs 5 min
Nivel de presión:0.1- 900 MPa
HP0.1, 250, 350, 450, 550 y 650 MPa, 15ºC, 5min con
despresurización de 5 min
HP0.1, 250 y 350 MPa, 25ºC, 5 - 15 min con
despresurización en 0.07 min
HP0.1, 250 y 350 MPa,
25ºC, 5 min con despresurización
0.07 -10 min
Nivel de presión:0.1- 350 MPa
Nitrógeno total
Furosina
Lactosa y lactulosa
Lactoferrina e IgA
Proteínas mayoritarias solubles en fracciones
Lactoferrina e IgA
Tamaño micela de caseína
Leche cruda desnatada
Mat
eria
pr
ima
Aná
lisis
Capítulo I Capítulo II Capítulo III Capítulo IV
Fórmula Infantil
(con o sin NEM o DMF)
UHT(142ºC, 4.7 seg)
Fact
ores
est
udia
dos
Tra
tam
ient
os a
plic
ados
Alta presión (HP)650 MPa, 15ºC,
5 min
HP0.1, 250, 350, 450, 550 y 650 MPa, 15ºC, 5min con
despresurización instantánea
Tipo de tratamiento: UHT v HP
Equipo utilizado:Industrial vs piloto
t mantenimiento presión:
5, 6, 10 y 15 min
t despresurización:0.07, 1, 5 y 10 min
HP0.1, 250, 450, 550,
700, 800 y 900 MPa, 15ºC, 5min
Nivel de presión:0.1- 650 MPa
t despresurización en equipo piloto:30 seg vs 5 min
Nivel de presión:0.1- 900 MPa
HP0.1, 250, 350, 450, 550 y 650 MPa, 15ºC, 5min con
despresurización de 5 min
HP0.1, 250 y 350 MPa, 25ºC, 5 - 15 min con
despresurización en 0.07 min
HP0.1, 250 y 350 MPa,
25ºC, 5 min con despresurización
0.07 -10 min
Nivel de presión:0.1- 350 MPa
Nitrógeno total
Furosina
Lactosa y lactulosa
Lactoferrina e IgA
Proteínas mayoritarias solubles en fracciones
Lactoferrina e IgA
Tamaño micela de caseína
Leche cruda desnatada

3. MATERIALES Y MÉTODOS


MATERIALES Y MÉTODOS
- 43 -
3.1. Preparación de las muestras y tratamiento aplicado
El procedimiento llevado a cabo para preparar las muestras (leches y fórmulas
infantiles), los tratamientos de presión o UHT aplicados y los equipos de alta presión
utilizados se explican con detalle en el apartado de Materiales y Métodos de cada capítulo.
3.2. Obtención de fracciones proteicas
Una vez tratadas las muestras, ya sean leche desnatada, leche desnatada con o sin NEM o
DMF, sobrenadantes de ultracentrifugación o fórmulas infantiles, se procedió al
fraccionamiento de las mismas como se indica a continuación:
• Fracción soluble de la ultracentrifugación a 100 000 x g, a 20 ºC durante 1 h. La
separación de las proteínas mediante esta técnica se basa en la diferente velocidad de
sedimentación de las moléculas en función de su peso molecular o del tamaño medio
de un conjunto de partículas. El sobrenadante obtenido se filtró a través de papel de
filtro Whatman nº 41.
• Fracción soluble a pH 4.6 se obtuvieron ajustando el pH de las muestras a 4.6 con HCl
2N, pH en el cual precipitan las caseínas. Las muestras se dejaron reposar durante 20
minutos, y se centrifugaron a 4 000 x g, 20 min, 5 ºC. La fracción soluble se filtró a
través de papel de filtro Whatman nº 40.
Fracción soluble de la precipitación con ácido tricloroacético al 4% (concentración
final). Las muestras se mezclaron con un volumen idéntico de ácido tricloroacético al
8% (v/v), se mantuvieron en reposo durante 1 h y se filtraron por un filtro Whatman nº
40.Eeste fraccionamiento sólo se realizó en las fórmulas infantiles (capitulo I).

MATERIALES Y MÉTODOS
- 44 -
3.3. Análisis de las muestras control y de las fracciones proteicas
3.3.1. Determinación del contenido en nitrógeno
Se empleó el método Kjeldahl para determinar el contenido en nitrógeno total
siguiendo el método prescrito en la Norma IDF (FIL-IDF, 1993). El contenido de proteínas se
calculó multiplicando dicho contenido por 6.38. Las muestras se digirieron en un digestor DK
20 (Velp Scientifica Srl., Usmate, Italy) y se destilaron en una unidad de destilación UDK 142
(Velp Scientifica Srl.).
3.3.2. PAGE-SDS
Las leches, las fórmulas infantiles y las fracciones obtenidas según los distintos
procedimientos de ambas se analizaron por electroforesis en gel de poliacrilamida (PAGE) en
presencia de agentes desnaturalizantes (SDS) y en condiciones reductoras (2-β-
mercaptoetanol). Las leches y fórmulas infantiles se diluyeron 1:9 y las fracciones solubles
1:3 en tampón de la muestra, consistente en 2.5% de SDS, 1 mM EDTA (ácido
dinitrotetraacético) y 5% de 2-β-mercaptoetanol en 10 mM de tampón Tris(hidroxi-
metil)amino metano pH 8. En los geles se incluyó una mezcla de proteínas estándar
compuesta por fosforilasa B (97 kDa), BSA (66 kDa), ovoalbúmina (45 kDa), anhidrasa
carbónica (30 kDa), inhibidor de tripsina (21.5 kDa) y α-La (14.4 kDa) (GE Healthcare,
Uppsala, Suecia). El desarrollo de la electroforesis se llevó a cabo en un equipo automatizado
(PhastSystem, Pharmacia), dotado de una unidad de separación y una de tinción. Se utilizaron
geles Homogeneus 20 y tiras de tampón (PhastGel SDS Strips) comerciales (GE Healthcare).
Se seguieron las condiciones electroforéticas indicadas en el manual del equipo (Pharmacia,
nota de aplicación nº 111, 1987), mostradas en la Tabla 2. Los geles se tiñeron con azul de
coomasie R-350 siguiendo las condiciones que se describen en el manual de instrucciones
(Pharmacia, notas de aplicación nº 200 y nº 210 de 1987, respectivamente).
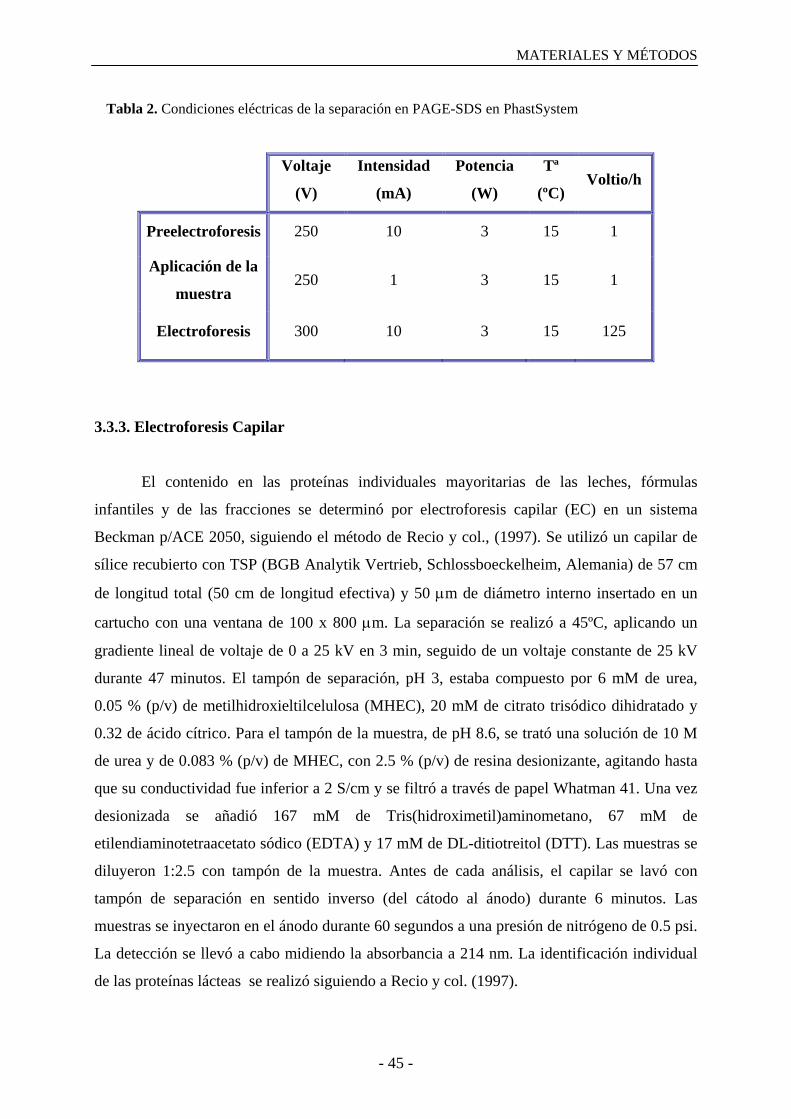
MATERIALES Y MÉTODOS
- 45 -
Tabla 2. Condiciones eléctricas de la separación en PAGE-SDS en PhastSystem
Voltaje
(V)
Intensidad
(mA)
Potencia
(W)
Tª
(ºC) Voltio/h
Preelectroforesis 250 10 3 15 1
Aplicación de la
muestra 250 1 3 15 1
Electroforesis 300 10 3 15 125
3.3.3. Electroforesis Capilar
El contenido en las proteínas individuales mayoritarias de las leches, fórmulas
infantiles y de las fracciones se determinó por electroforesis capilar (EC) en un sistema
Beckman p/ACE 2050, siguiendo el método de Recio y col., (1997). Se utilizó un capilar de
sílice recubierto con TSP (BGB Analytik Vertrieb, Schlossboeckelheim, Alemania) de 57 cm
de longitud total (50 cm de longitud efectiva) y 50 μm de diámetro interno insertado en un
cartucho con una ventana de 100 x 800 μm. La separación se realizó a 45ºC, aplicando un
gradiente lineal de voltaje de 0 a 25 kV en 3 min, seguido de un voltaje constante de 25 kV
durante 47 minutos. El tampón de separación, pH 3, estaba compuesto por 6 mM de urea,
0.05 % (p/v) de metilhidroxieltilcelulosa (MHEC), 20 mM de citrato trisódico dihidratado y
0.32 de ácido cítrico. Para el tampón de la muestra, de pH 8.6, se trató una solución de 10 M
de urea y de 0.083 % (p/v) de MHEC, con 2.5 % (p/v) de resina desionizante, agitando hasta
que su conductividad fue inferior a 2 S/cm y se filtró a través de papel Whatman 41. Una vez
desionizada se añadió 167 mM de Tris(hidroximetil)aminometano, 67 mM de
etilendiaminotetraacetato sódico (EDTA) y 17 mM de DL-ditiotreitol (DTT). Las muestras se
diluyeron 1:2.5 con tampón de la muestra. Antes de cada análisis, el capilar se lavó con
tampón de separación en sentido inverso (del cátodo al ánodo) durante 6 minutos. Las
muestras se inyectaron en el ánodo durante 60 segundos a una presión de nitrógeno de 0.5 psi.
La detección se llevó a cabo midiendo la absorbancia a 214 nm. La identificación individual
de las proteínas lácteas se realizó siguiendo a Recio y col. (1997).

MATERIALES Y MÉTODOS
- 46 -
3.3.4. ELISA tipo sandwich
La cuantificación de lactoferrina (Lf) e inmunoglobulina A secretora (IgA) se realizó
mediante esta técnica inmunológica, empleando kits comerciales de cuantificación de Lf e
IgA bovina de Bethyl Laboratories. Inc. (Montgomery, USA). En primer lugar se añadieron
100 μL del anticuerpo anti-lactoferrina bovina (anti-LF) o anti-IgA bovina (anti-IgA) a una
placa multipocillos (Nunc, Thermo Fisher Scientific, Waltham, USA ) y se incubó durante 60
min a temperatura ambiente. Posteriormente, la placa se lavó 3 veces con un tampón de
lavado (50 mM Tris, 0.14 M NaCl, 0.05 % Tween 20, pH 8) y se bloquearon los pocillos
durante 30 min con 200 μL de tampón de bloqueo (50 mM Tris, 0.14 M NaCl, 1% BSA, pH
8). Nuevamente, se volvió a lavar la placa 3 veces y, a continuación, se añadieron 100 μL de
las leches y fórmulas infantiles controles y presurizadas y de las fracciones de leche. Tanto las
leches como sus fracciones se diluyeron 1:2000 y 1:1000 dependiendo del tratamiento de
presión aplicado mientras que las fórmulas infantiles se testaron sin diluir. Además, también
se añadieron disoluciones de Lf en 50 mM Tris, 0.14 M NaCl, 1% BSA y 0.05% Tween 20 a
pH 8, en el intervalo de 500-7.8 ng/mL, o de IgA en el intervalo de 1000-15.6 ng/mL, con el
objetivo de elaborar una recta patrón. Tras 60 min de incubación a temperatura ambiente, los
pocillos se lavaron 5 veces con el tampón de lavado y se le adicionaron 100 μL del anticuerpo
anti-LF o anti-IgA unido a peroxidasa de rábano (HRP) (dilución 1:50000 en 50 mM Tris,
0.14 M NaCl, 0.05% Tween 20, pH 8), incubándose 1 hora. A continuación, se lavó
nuevamente 5 veces y se le adicionaron 100 μL del sustrato de la enzima, ortofenilendiamina
(OPD, Palex Medical SA, San Cugat del Vallés, España), y se dejó actuar durante 8 min en
oscuridad, parándose la reacción mediante la adición de 100 μL de H2SO4 2M. Se midió la
absorbancia a 492 nm con un espectrofotómetro Multiskan Ascent (Thermo Fisher Scientific).
La concentración de Lf en cada una de las muestras se determinó interpolando los valores de
absorbancia en la curva patrón. La ecuación de la curva se obtuvo mediante el programa
estadístico GraphPad Prism 4.0 y se ajustó a: y= ((A-D)/(1+(x/C)^B))+D, siendo: A=
0.01578; B= 1.403; C= 114.8; D= 2.304; R2= 0.9979 en el caso de la Lf (Figura 11) y A= -
0.0002165; B= 1.157; C= 755.2; D= 3.061; R2= 0.9986 para la IgA (Figura 12). Todas la
muestras se ensayaron por duplicado.

MATERIALES Y MÉTODOS
- 47 -
1 10 100 10000.0
0.5
1.0
1.5
2.0
2.5
Lactoferrina (ng/mL)
A 49
2 nm
Figura 11. Representación de la recta de calibrado de lactoferrina
10 100 10000.0
0.5
1.0
1.5
2.0
Ig A (ng/mL)
A 49
2 nm
Figura 12. Representación de la recta de calibrado de IgA
3.3.5. Medida del tamaño de la micela de caseína
Como medio de observación de la micela de caseína en la leche desnatada control y en
las presurizadas de 250 a 900 MPa (capítulo IV) se utilizó la microscopía electrónica de
transmisión siguiendo el método descrito por García-Risco y col. (2003). Para realizar el
procedimiento fue preciso solidificar la leche. Se mezclaron en una placa Petri 5 mL de leche
con 5 mL de agar estéril al 3 % (Scharlau, Barcelona, Spain) agitándose hasta quedar lo más
homogéneo posible. Una vez solidificada la leche, se cortaron rectángulos de 1 cm x 0.5 cm x
10 mm3 aproximadamente y se sumergieron en una solución de glutaraldehido al 2.5 %

MATERIALES Y MÉTODOS
- 48 -
(Sigma-Aldrich, St. Louis, MO, USA) en PBS (4.5 % NaCl y 3.9 % NaH2PO4) a pH 7,
durante 3 horas a 4ºC en oscuridad para la óptima fijación de las proteínas. Posteriormente,
los rectángulos se lavaron 3 veces con PBS, pH 7 dejándose en dicha solución durante toda la
noche a 4ºC. El siguiente paso consistió en sumergir los rectángulos en una solución de
tetróxido de osmio al 1 % (Ted Pella, Inc., Redding, CA, USA) durante 1 hora,
impregnándose de esta forma con metal de alto peso molecular, incrementando su densidad
electrónica. La eliminación del tetróxido de osmio se realizó mediante 2 lavados con agua
Milli-Q, seguidos de lavados consecutivos con proporciones crecientes de acetona en agua
(40, 60, 70, 90, 95 y 100%) para deshidratar la muestra. Finalmente, las muestras se fijaron en
resina Epoxi Spurr (Taab Laboratory and Microscopy, Aldermaston, United Kingdom)
mediante la adición de mezclas en proporciones crecientes de acetona-resina desde 2:1 hasta
100 % de resina. Tras cada una de estas adiciones se mantuvieron 1 hora a temperatura
ambiente. Los rectángulos se pusieron en un molde de silicona cubriéndose de 100 % de
resina, manteniéndose 72 h a 65ºC, con lo que la resina se solidificó y fue fácilmente
manipulable. Por último, se cortaron láminas de estas muestras con ultramicrotomo y se
tiñeron con acetato de uranilo al 2% y citrato de plomo Reynolds, ambos productos
proporcionados por Taab Laboratory and Microscopy (Aldermaston, United Kingdom). Una
vez teñidas se examinaron por el microscopio electrónico JEOL sem-1010 (JEOL Ltd., Tokio,
Japan) operando a 80 kV.
La medida del tamaño de las micelas de caseína se realizó mediante el programa
Measure IT (Olympus), midiéndose las caseínas incluidas en un rectángulo de 1.61x 2.16 μm
a partir de las fotografías realizadas con 40000 aumentos (2.89 x 3.6 μm). Se midieron
aproximadamente 100 micelas de caseínas para cada tratamiento a partir de varias fotografías.
3.3.6. Determinación del contenido en lactosa y lactulosa
El contenido en lactosa y lactulosa en las fórmulas infantiles (capítulo 1) se determinó
por cromatografía de gases siguiendo el método descrito por Montilla y col. (2005). Esto
requirió una extracción previa de los carbohidratos con metanol (1:9, v/v, fórmula
infantil/metanol), con objeto de precipitar las proteínas y la grasa que pudieran interferir en
las determinaciones. Tras 1 h de reposo, se recogieron 0.5 mL del sobrenadante, se mezclaron
con 0.5 mL de β-fenil-glucósido (patrón interno) y se procedió a su secado por rotavapor.

MATERIALES Y MÉTODOS
- 49 -
Posteriormente, se realizó la derivatización de los carbohidratos, añadiendo 0.1 mL de DMF,
con reposo de 1 h a 65ºC, y 0.1 mL de N-trimetilsililimidazol (TMSI, Sigma-Aldrich),
dejándolo reaccionar 30 min a 65ºC. Finalmente, la reacción se interrumpió por frío y adición
de 0.1 mL de hexano y 0.2 mL de agua y 1 μL de la fracción orgánica se utilizó para su
análisis por cromatografía de gases. Para ello, se utilizó un cromatógrafo HP 6890 equipado
con un detector de ionización de llama (FID) y con una columna de sílice fundida recubierta
con metilfenilsilicona (OV17) de 25 m, 0.32 mm de diámetro interno y 0.5 μm de espesor de
película. Se empleó N2 como gas portador con un flujo inicial de 1.8 ml/min y se realizó la
inyección con una relación de split de 1:40. El gradiente de temperaturas usado fue: 280ºC en
el inyector, 300 C en el detector, el horno se mantuvo a 235ºC durante 10 min y, se programó
un gradiente de 20ºC/ min hasta 270ºC que se mantuvo 5 min y de 20ºC/ min hasta 290ºC y
se mantuvo 2 min. Para la cuantificación se empleó el método de calibración interna,
utilizando β-fenil-glucósido como patrón.
3.3.7. Determinación del contenido en furosina
El análisis de la furosina en las fórmulas infantiles (capitulo I) se realizó del siguiente
modo: 2 mL de la fórmula se mezclaron con 6 mL de HCl 10.6 N en tubos pirex (10.2 x 2 cm)
de hidrólisis. A continuación, se pasó una corriente de helio durante 1 min para crear una
atmósfera inerte y, posteriormente, se realizó la hidrólisis a 110ºC durante 23 horas. Una vez
finalizada la hidrólisis, se filtró el hidrolizado a través de cartuchos C18 (Sep-Pak Plus, Waters
Corporation, Milford, Massachussets, USA), activados previamente con 5 ml de metanol y 10
ml de agua milli-Q. Por último, se pasaron 3 ml de HCl 3 N a través del cartucho C18 para
eluir la furosina retenida en el cartucho.
La determinación cuantitativa de la furosina se realizó por RP-HPLC empleando una
columna C8 específica para el análisis de furosina (Alltech, Lokeren, Bélgica) (250 mm x 4.6
mm d.i) (Resmini y col., 1990). El equipo utilizado estaba provisto de una bomba binaria
modelo 250 (PerkinElmer Life and Analytical Sciences, Inc, Waltham, Massachussets, USA),
un inyector manual Rheodyne (modelo 9125), un horno (Kariba Instruments, Cardiff, USA)
para termostatizar la columna, un detector UV (LDC Analytical, SM 4000, Riviera Beach,
FL, USA) y una interfase modelo 406 (Beckman). La adquisición y procesamiento de los
datos se realizó utilizando el programa System Gold versión 711 (Beckman Instruments). Las
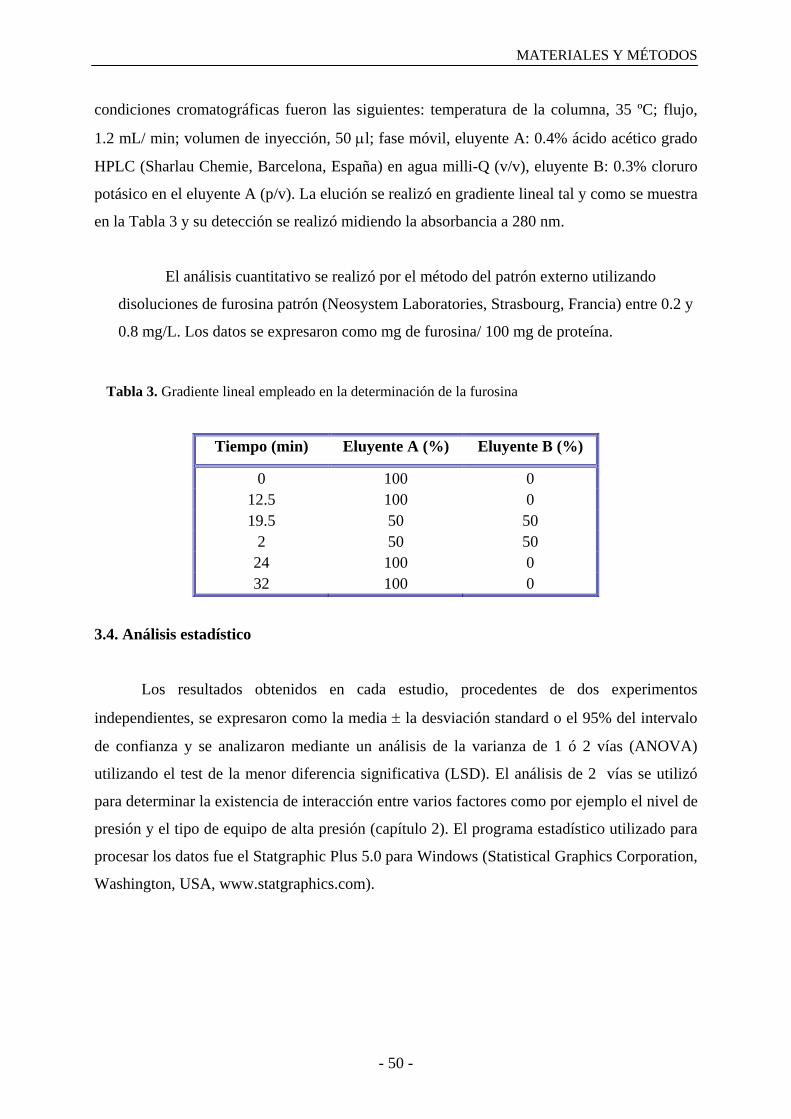
MATERIALES Y MÉTODOS
- 50 -
condiciones cromatográficas fueron las siguientes: temperatura de la columna, 35 ºC; flujo,
1.2 mL/ min; volumen de inyección, 50 μl; fase móvil, eluyente A: 0.4% ácido acético grado
HPLC (Sharlau Chemie, Barcelona, España) en agua milli-Q (v/v), eluyente B: 0.3% cloruro
potásico en el eluyente A (p/v). La elución se realizó en gradiente lineal tal y como se muestra
en la Tabla 3 y su detección se realizó midiendo la absorbancia a 280 nm.
El análisis cuantitativo se realizó por el método del patrón externo utilizando
disoluciones de furosina patrón (Neosystem Laboratories, Strasbourg, Francia) entre 0.2 y
0.8 mg/L. Los datos se expresaron como mg de furosina/ 100 mg de proteína.
Tabla 3. Gradiente lineal empleado en la determinación de la furosina
Tiempo (min) Eluyente A (%) Eluyente B (%)
0 100 0 12.5 100 0 19.5 50 50
2 50 50 24 100 0 32 100 0
3.4. Análisis estadístico
Los resultados obtenidos en cada estudio, procedentes de dos experimentos
independientes, se expresaron como la media ± la desviación standard o el 95% del intervalo
de confianza y se analizaron mediante un análisis de la varianza de 1 ó 2 vías (ANOVA)
utilizando el test de la menor diferencia significativa (LSD). El análisis de 2 vías se utilizó
para determinar la existencia de interacción entre varios factores como por ejemplo el nivel de
presión y el tipo de equipo de alta presión (capítulo 2). El programa estadístico utilizado para
procesar los datos fue el Statgraphic Plus 5.0 para Windows (Statistical Graphics Corporation,
Washington, USA, www.statgraphics.com).

4. RESULTADOS Y DISCUSIÓN


CAPÍTULO I
Estudio del efecto de las altas presiones sobre las propiedades
nutricionales de las fórmulas infantiles líquidas


RESULTADOS Y DISCUSIÓN: CAPÍTULO I
- 55 -
I.1. Introducción
La leche materna es el mejor alimento para el recién nacido, sin embargo, no siempre
es posible su administración. Las fórmulas infantiles, diseñadas para reemplazar a esta leche,
están elaboradas principalmente a base de proteínas lácteas bovinas, aunque se pueden
completar hasta con 60 ingredientes, como aceites vegetales, vitaminas o minerales (Naispour
y col., 2006; Thompkinson y Kharb, 2007), en cantidad necesaria para favorecer el
crecimiento y desarrollo normal del niño, además de prevenir enfermedades (Bosch y col.,
2008). La formulación de estas leches infantiles ha ido cambiando y mejorando a lo largo del
tiempo, si bien el objetivo principal de los fabricantes no es obtener una fórmula que imite la
composición exacta de la leche humana, ya que determinados ingredientes son muy difíciles
de obtener, sino conseguir los efectos funcionales que produce la leche materna, por lo que se
les adicionan, por ejemplo, probióticos o prebióticos. Además, algunos autores señalan la
importancia de una ingesta adecuada de proteínas, sobre todo, de la relación entre
caseínas/proteínas de suero, que es de 40/60 y 80/20 en la leche de humana y de vaca,
respectivamente. La ingestión de un mayor contenido de proteínas de suero facilita el proceso
de digestión (Riordan, 1998) e incluso podría favorecer la colonización del intestino por
bacterias probióticas (Balmer y col., 1989).
Por otro lado, tanto las fórmulas lácteas como los ingredientes que se les añaden se
someten a tratamientos térmicos con el fin de que el producto final sea seguro
microbiológicamente y tenga una vida útil mayor. Además, el producto final normalmente
pasa por torres de secado con el fin de obtener leches infantiles en polvo (Floris y col., 2007).
Los tratamientos térmicos aplicados pueden ser relativamente suaves, como la pasteurización,
que consiste en aplicar 71.7ºC durante 5 seg, mientras que el tratamiento UHT, más severo,
consiste en tratar el producto a 135ºC, aproximadamente, durante unos seg e, incluso, pueden
darse tratamientos de esterilización (110ºC, 10-30 min) (Pischetsrieder y Henle, 2012). Estos
tratamientos térmicos pueden causar importantes daños en la composición de las fórmulas,
como agregación de proteínas desnaturalizadas, degradación de vitamina C, peroxidación de
lípidos, etc. Además, tanto en estos procesos, como durante el posterior almacenamiento de
las fórmulas, se produce la reacción de Maillard, la unión de azúcares reductores como
glucosa, lactosa y maltosa al grupo ε-amino de la lisina (Ferrer y col., 2003), que genera
cambios de color, disminuye la calidad de las proteínas y su digestibilidad (Meade y col.,

- 56 -
2005; Dupont y col., 2010). Esta reacción es bastante frecuente en las fórmulas infantiles, ya
que tienen un alto contenido en proteínas y lactosa (Pischetsrider y Henle, 2012).
Por ello, se intentan buscar nuevas técnicas de procesado de alimentos que minimicen
los efectos secundarios producidos por los tratamientos térmicos. Dentro de las técnicas
alternativas, la alta presión hidrostática es la más utilizada industrialmente, ya que permite
obtener alimentos procesados con mejores cualidades sensoriales y nutricionales, sin
embargo, su aplicación industrial en productos lácteos es aún bastante escasa, puesto que
actualmente solo se comercializan calostros y yogures presurizados. Además, no existen
estudios sobre la aplicación de altas presiones a formulas infantiles.Así, el objetivo de este
estudio fue evaluar y comparar el efecto de un tratamiento UHT y de presión a 650 MPa
durante 5 min a 15ºC en una fórmula infantil. Los parámetros utilizados fueron el grado de
desnaturalización de las proteínas lácteas, y en particular de las proteínas minoritarias del
suero, como lactoferrina e inmunoglobulina A, y la formación de lactulosa y furosina.
I.2. Materiales y Métodos
La Figura 13 resume el procedimiento experimental seguido en este trabajo.
I.2.1. Preparación de las muestras y tratamientos aplicados
Para realizar este estudio se utilizó una fórmula infantil preparada por Ordesa S.A,
siguiendo el procedimiento habitual excepto que no se le aplicó el último tratamiento térmico,
ni se atomizó. Esta fórmula se preparó a partir de leche desnatada y pasteurizada a baja
temperatura, con adición de proteínas de suero. Tras su preparación, la fórmula se dividió en 3
alícuotas. Una se dejó en refrigeración, ya que se utilizó como control, otra se calentó a 142-
146ºC durante 4.7 seg (tratamiento UHT) y, finalmente, la última alícuota se sometió a un
tratamiento de presión a 650 MPa durante 5 min a 15ºC. En este último caso, se utilizó un
equipo de alta presión a escala industrial Wave 6000/120 (NC Hyperbaric, Burgos, España),
con capacidad para 120 L. Este equipo usa agua como fluido transmisor de la presión y tiene
3 bombas para generar la presión hidráulicamente, la cual sube a una velocidad de 3.04 MPa/
seg y se libera en 1.5 seg.
Las fórmulas se atemperaron a 37ºC durante 30 min, se centrifugaron a 3000 x g a 20
ºC durante 30 min y se filtraron por lana de vidrio con el fin de desnatarlas.
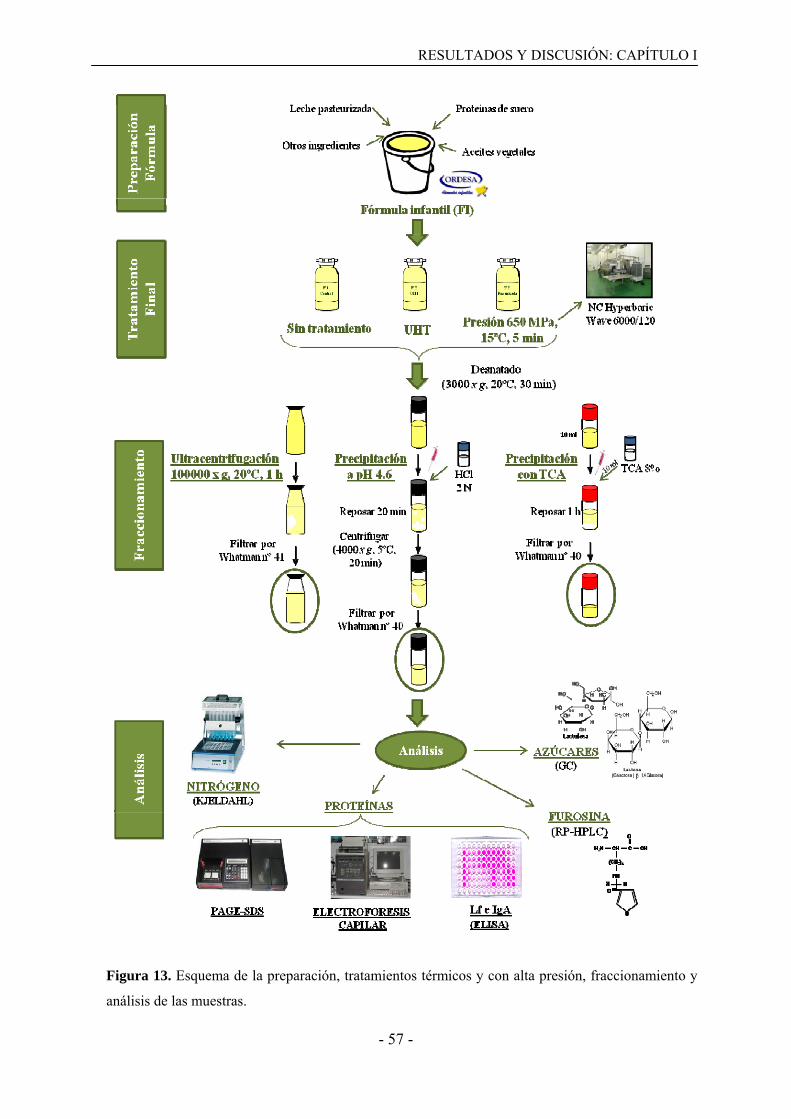
RESULTADOS Y DISCUSIÓN: CAPÍTULO I
- 57 -
Figura 13. Esquema de la preparación, tratamientos térmicos y con alta presión, fraccionamiento y
análisis de las muestras.

- 58 -
I.3. Resultados
I.3.1. Efecto de los tratamientos UHT y con alta presión en las proteínas mayoritarias de
la leche
La Figura 14A, muestra el electroforegrama (EC) de la fórmula infantil control y de
una leche humana cuyas proteínas se han identificado de acuerdo con Manso y col. (2007) (a
y b respectivamente). El perfil electroforético de la fórmula se asemejó más al de una leche de
vaca (Figura 14B), aunque con una mayor relación de proteína de suero a caseína, más
próxima a la de la leche de mujer. En la fórmula, destaca, en la zona de las proteínas de suero,
la aparición de varios picos adyacentes a cada pico principal. Éstos se atribuyeron a distintos
estados de glicosilación de las seroproteínas debido a la reacción de Maillard con la lactosa
presente en la leche. Los tiempos de migración mayores que presentan las formas lactosiladas
de las proteínas de suero se deben a que la unión covalente de la lactosa a la lisina reduce la
carga neta positiva a pH ácido y aumenta el peso molecular (Recio y col., 2001). El hecho de
que aparezcan picos correspondientes a varias formas glicosiladas de cada proteína
posiblemente se deba a la existencia de proteínas con diferentes grados de lactosilación.
La Tabla 4 muestra el contenido en proteína de las fórmulas infantiles, de los
sobrenadantes de ultracentrifugación (SU) y de las fracciones solubles a pH 4.6, así como el
nitrógeno no proteico (NNP, soluble en TCA al 4%). La concentración de proteína soluble en
los SU fue similar en todas las fórmulas estudiadas, pero el contenido en proteína soluble a
pH 4.6 disminuyó en el orden: fórmula control> fórmula presurizada> fórmula UHT. El
NNP disminuyó en, aproximadamente, un 15% en las fórmulas UHT y presurizada, respecto a
la fórmula control.
Resultados previos obtenidos en nuestro laboratorio (Manso y col, 2007) han puesto
de manifiesto que el contenido medio en proteína de la leche humana es de aproximadamente
un 0.94%, lo que supone menos de un 30% del contenido de la leche bovina y un 60% del de
la fórmula analizada en este estudio. Además, a lo largo de la lactación tiene lugar una
progresiva disminución, de una media en torno al 1.46% al final de la primera semana, hasta
un 0.66% a la semana 18. Por otra parte, la relación de caseínas a proteínas de suero
recomendada para la elaboración de fórmulas infantiles es 40:60, mientras que, en la fórmula,
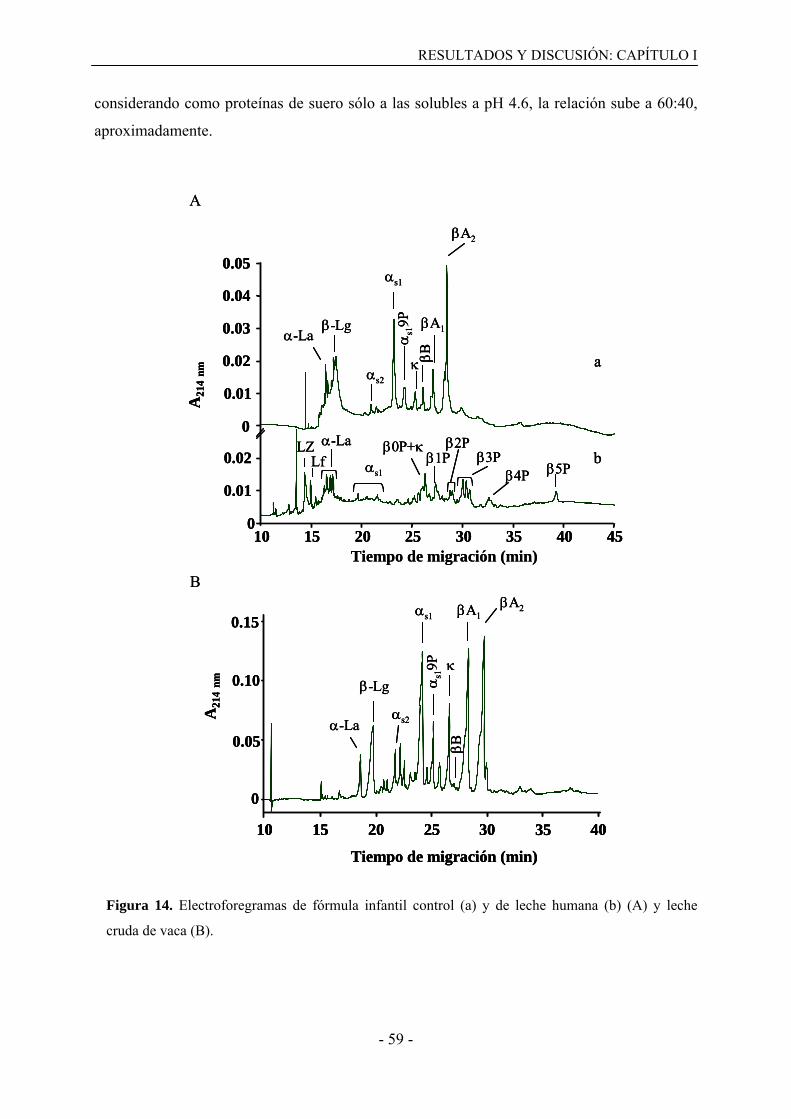
RESULTADOS Y DISCUSIÓN: CAPÍTULO I
- 59 -
considerando como proteínas de suero sólo a las solubles a pH 4.6, la relación sube a 60:40,
aproximadamente.
Figura 14. Electroforegramas de fórmula infantil control (a) y de leche humana (b) (A) y leche
cruda de vaca (B).
Tiempo de migración (min)
0
0.01
0.02
0.03
0.04
0.05
0
0.01
0.02
10 15 20 25 30 35 40 45
A21
4nm
a
b
β-Lg
αs2
αs1
αs1
9P
βA2
βA1
βBκ
α-La
LZLf
α-La
αs1
β0P+κβ1P
β2Pβ3P
β5Pβ4P
βA2
0.05
0.10
0.15
10 15 20 25 30 35 40
α-La
β-Lg
A21
4nm
Tiempo de migración (min)
αs2
αs1
αs1
9P
βA1
βB
κ
0
A
BTiempo de migración (min)
0
0.01
0.02
0.03
0.04
0.05
0
0.01
0.02
10 15 20 25 30 35 40 45
A21
4nm
a
b
β-Lg
αs2
αs1
αs1
9P
βA2
βA1
βBκ
α-La
LZLf
α-La
αs1
β0P+κβ1P
β2Pβ3P
β5Pβ4P
0
0.01
0.02
0.03
0.04
0.05
0
0.01
0.02
10 15 20 25 30 35 40 45
A21
4nm
a
b
β-Lg
αs2
αs1
αs1
9P
βA2
βA1
βBκ
α-La
LZLf
α-La
αs1
β0P+κβ1P
β2Pβ3P
β5Pβ4P
βA2
0.05
0.10
0.15
10 15 20 25 30 35 40
α-La
β-Lg
A21
4nm
Tiempo de migración (min)
αs2
αs1
αs1
9P
βA1
βB
κ
0
βA2
0.05
0.10
0.15
10 15 20 25 30 35 4010 15 20 25 30 35 40
α-La
β-Lg
A21
4nm
Tiempo de migración (min)
αs2
αs1
αs1
9P
βA1
βB
κ
0
A
B

- 60 -
Tabla 4. Contenido en proteína total, proteína soluble en los sobrenadantes de ultracentrifugación
(SU) y a pH 4.6 (4.6), y nitrógeno no proteico (NNP) de las fórmulas infantiles control, UHT y
presurizada a 650 MPa durante 5 min (%, p/v).
Proteína (%) Tratamiento
Total SU 4.6
NNP
(%)
Control 1.60±0.01 1.04a±0.01 0.66c±0.00 0.05b±0.00
UHT 0.91a±0.01 0.44a±0.00 0.04a±0.00
Presión (650 MPa) 0.99a±0.09 0.54b±0.00 0.04a±0.00 a-d Diferentes superíndices, en columnas, indican diferencias significativas (P< 0.05) producidas por el tratamiento.
Los análisis mediante PAGE-SDS y EC de estas fracciones (Figuras 15, 16 y 17)
complementaron los resultados anteriores. Como muestra la Figura 15, los sobrenadantes de
ultracentrifugación presentaron, en los tres casos, bandas correspondientes a las principales
caseínas (αS2-, αS1-, β- y κ-) y, tanto en la fórmula presurizada (calle 5) como en la control
(calle 3), bandas de proteínas minoritarias: Lf, BSA e inmunoglobulina, que, probablemente,
se desnaturalizaron parcialmente tras el tratamiento UHT (calle 4). En las fracciones solubles
a pH 4.6 (calles 6, 7, 8) se observó una disminución de la β-Lg y de la α-La solubles en las
fórmulas procesadas (UHT y alta presión) pero, sobre todo, en la fórmula UHT.
Figura 15. PAGE-SDS de las fórmulas infantiles. 1: Marcador de pesos moleculares; 2: fórmula de
partida (control); 3 a 5: sobrenadantes de ultracentrifugación (SU) de las fórmulas control, UHT y
presurizada; 6 a 8: fracciones solubles a pH 4.6 de las fórmulas control, UHT y presurizada.
1 2 3 4 5 6 7 8
β-CNα-Laβ-Lg
αS2-CN
κ-CN
αS1-CN
LfBSAIg
SU 4.6
UH
TPr
esió
n
Con
trol
UH
TPr
esió
n
Con
trol
1 2 3 4 5 6 7 8
β-CNα-Laβ-Lg
αS2-CN
κ-CN
αS1-CN
LfBSAIg
SU 4.6
UH
TPr
esió
n
Con
trol
UH
TPr
esió
n
Con
trol

RESULTADOS Y DISCUSIÓN: CAPÍTULO I
- 61 -
Se observó que el tratamiento a 650 MPa de la formula infantil no indujo la
solubilización de caseína micelar, siendo los niveles de caseínas no sedimentables similares
en la fórmula control y en la presurizada, mientras que el tratamiento UHT disminuyó los
niveles de caseína soluble (Figura 16).
Figura 16. Electroforegramas de la fórmula infantil control (a), de los sobrenadantes de
ultracentrifugación (SU) de la misma fórmula (b), y tratada mediante UHT (c) y alta presión (650
MPa, 5 min) (d). 1: α-La; 2: β-Lg; 3:αs2-CN; 4: αs1-CN; 5: αs1-CN 9P; 6: κ-CN; 7: βB-CN; 8:βA1-
CN; 9: βA2-CN.
En el caso de las fracciones solubles a pH 4.6, la modificación en la forma y área de
los picos debida a la glicosilación impidió la correcta cuantificación de las proteínas. Sin
embargo, los electroforegramas mostrados en la Figura 17 indican claramente que ambos
tratamientos, UHT y presión, produjeron una gran desnaturalización de las proteínas
mayoritarias del suero, especialmente en el caso de la fórmula UHT, como ya apuntaban los
0
0.01
0.02
0.03
0.04
0.05
0
0.01
0
0.01
10 15 20 25 30 35 40
0
0.01
Tiempo de migración (min)
A21
4 nm
a
b
c
d
2
3
4
5
9
8
76
1
0
0.01
0.02
0.03
0.04
0.05
0
0.01
0
0.01
10 15 20 25 30 35 40
0
0.01
Tiempo de migración (min)
A21
4 nm
a
b
c
d
a
b
c
d
2
3
4
5
9
8
76
12
3
4
5
9
8
76
1

- 62 -
análisis mediante PAGE-SDS (Figura 15). La comparación de las Figuras 16 y 17 también
reveló que, tanto en la formula UHT como en la presurizada, una importante proporción de
las proteínas desnaturalizadas permaneció soluble en los sobrenadantes de ultracentrifugación.
Figura 17. Electroforegramas la fórmula infantil control (a) y de la fracción soluble a pH 4.6 de la
misma fórmula (b) y tratada mediante UHT (c) y alta presión (650 MPa, 5 min) (d).
0
0.01
0.02
0.03
0.04
0.05
a
0
0.01
0.02 b
0
0.01
0.02 c
0
0.01
0.02
10 15 20 25 30 35 40
d
Tiempo de migración (min)
A21
4nm
0
0.01
0.02
0.03
0.04
0.05
a
0
0.01
0.02 b
0
0.01
0.02 c
0
0.01
0.02
10 15 20 25 30 35 40
d
Tiempo de migración (min)
A21
4nm

RESULTADOS Y DISCUSIÓN: CAPÍTULO I
- 63 -
I.3.2. Efecto de los tratamientos UHT y con alta presión en las proteínas minoritarias del
suero
El contenido en Lf de la fórmula infantil control detectado por ELISA fue de 0.75
mg/100 g de proteína, similar al de la fórmula tratada por presión, con 0.66 mg/100 g de
proteína. Por otro lado, no pudo detectarse Lf por ELISA en la fórmula UHT, a pesar de que
el análisis por PAGE-SDS de las fórmulas completas mostró la presencia de una banda
correspondiente a Lf en las 3 muestras estudiadas (control, presurizada y UHT), como se
muestra en la Figura 18.
1 2 3 4 5 6 7 8
β-CN
α−Laβ-Lgκ-CN
LFBSAIG
αs2-CNαs1-CN
Figura 18. PAGE-SDS de las fórmulas infantiles. 1: Marcador de pesos moleculares; 2: BSA; 3:
Lf; 4: Suero bovino enriquecido en Ig; 5: Fórmula de partida; 6: Fórmula presurizada a 650 MPa, 5
min; 7: Fórmula sometida a tratamiento UHT; 8: Leche cruda de vaca.
Es posible que el tratamiento UHT produjera una desnaturalización importante en la
Lf y, aun observándose la banda correspondiente mediante PAGE-SDS, a lo que podría
contribuir la presencia del detergente y de β-mercaptoetanol en el tampón de la muestra, el
anticuerpo no reconociera los sitios de unión. Los resultados obtenidos sugieren que el
tratamiento con alta presión no cambió su estructura, o por lo menos los sitios de unión con el
anticuerpo, siendo su unión igual de eficaz que en la fórmula sin tratar.
El contenido de IgA observado en las fórmulas infantiles fue de 22.32 y 116.98 ng/mL
en las fórmulas control y presurizada respectivamente. No se pudo detectar IgA en la fórmula
infantil UHT con el método utilizado (Figura 19), al igual que sucedió en el caso de la Lf, lo
que probablemente se debió a la desnaturalización de la proteína o a la alteración de los
puntos de unión con el anticuerpo.
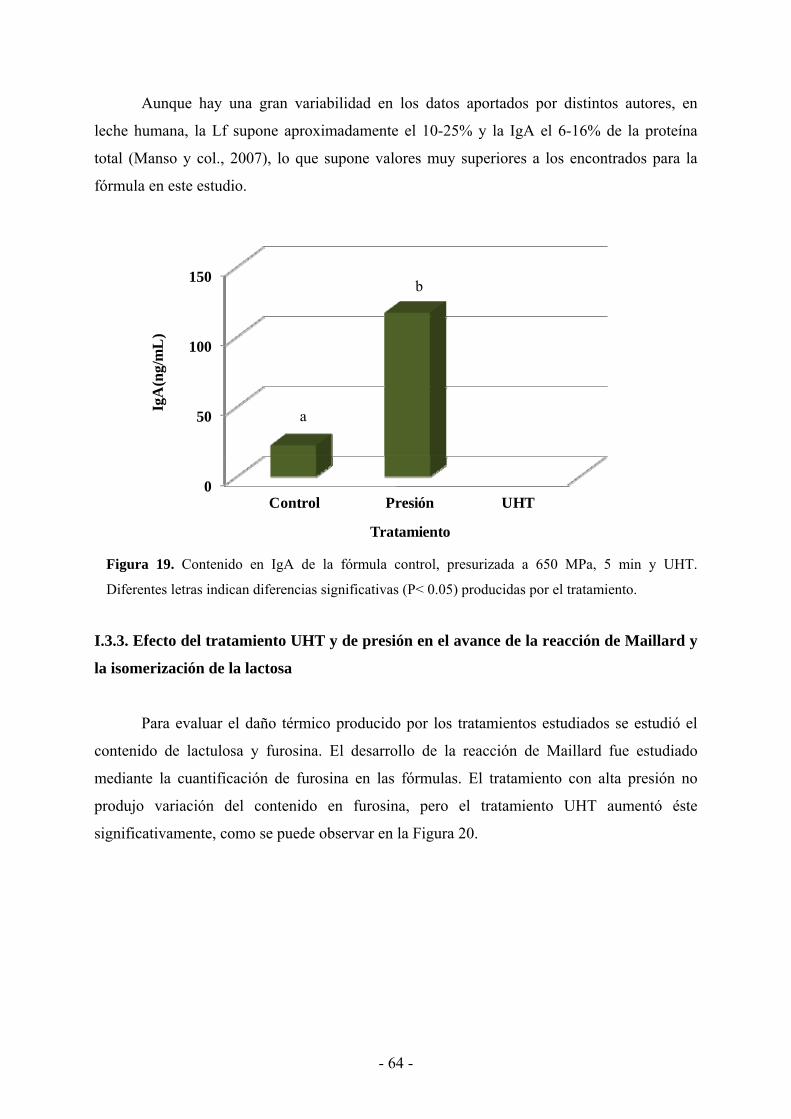
- 64 -
Aunque hay una gran variabilidad en los datos aportados por distintos autores, en
leche humana, la Lf supone aproximadamente el 10-25% y la IgA el 6-16% de la proteína
total (Manso y col., 2007), lo que supone valores muy superiores a los encontrados para la
fórmula en este estudio.
Presión UHTControl
Tratamiento
0
50
100
150
IgA
(ng/
mL
)
a
b
Figura 19. Contenido en IgA de la fórmula control, presurizada a 650 MPa, 5 min y UHT.
Diferentes letras indican diferencias significativas (P< 0.05) producidas por el tratamiento.
I.3.3. Efecto del tratamiento UHT y de presión en el avance de la reacción de Maillard y
la isomerización de la lactosa
Para evaluar el daño térmico producido por los tratamientos estudiados se estudió el
contenido de lactulosa y furosina. El desarrollo de la reacción de Maillard fue estudiado
mediante la cuantificación de furosina en las fórmulas. El tratamiento con alta presión no
produjo variación del contenido en furosina, pero el tratamiento UHT aumentó éste
significativamente, como se puede observar en la Figura 20.
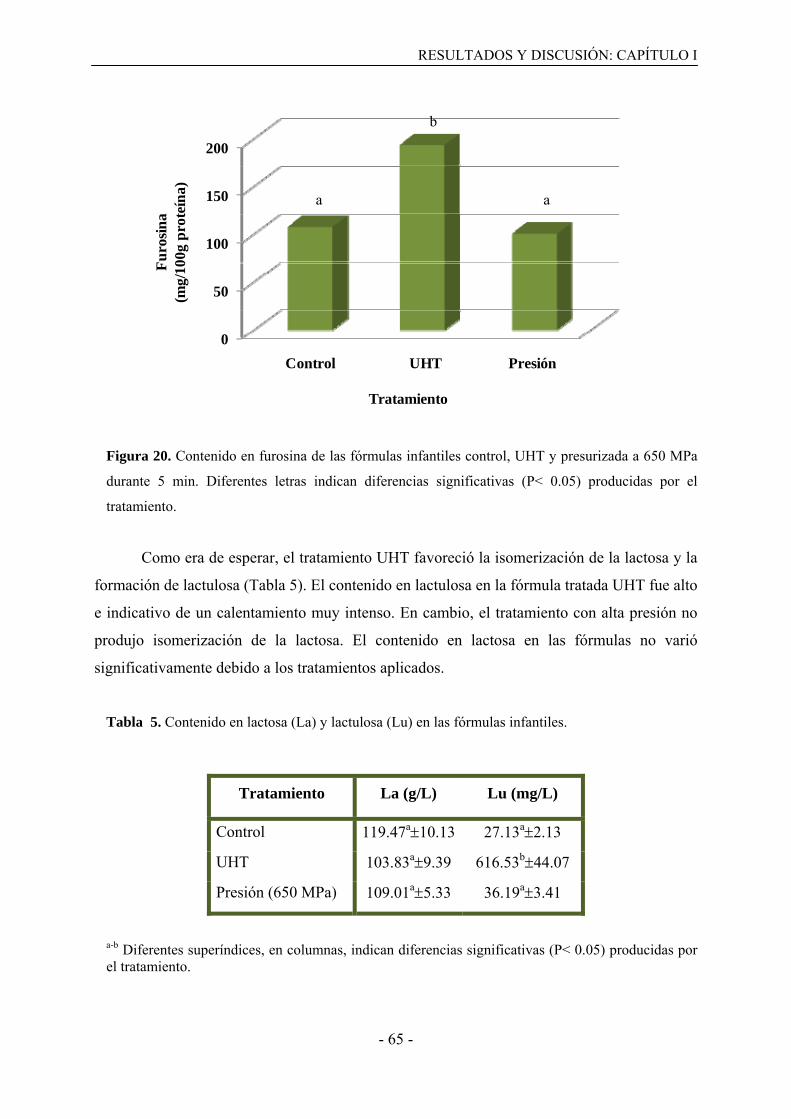
RESULTADOS Y DISCUSIÓN: CAPÍTULO I
- 65 -
0
50
100
150
200
Control UHT Presión
Tratamiento
Furo
sina
(mg/
100g
pro
teín
a)a a
b
Figura 20. Contenido en furosina de las fórmulas infantiles control, UHT y presurizada a 650 MPa
durante 5 min. Diferentes letras indican diferencias significativas (P< 0.05) producidas por el
tratamiento.
Como era de esperar, el tratamiento UHT favoreció la isomerización de la lactosa y la
formación de lactulosa (Tabla 5). El contenido en lactulosa en la fórmula tratada UHT fue alto
e indicativo de un calentamiento muy intenso. En cambio, el tratamiento con alta presión no
produjo isomerización de la lactosa. El contenido en lactosa en las fórmulas no varió
significativamente debido a los tratamientos aplicados.
Tabla 5. Contenido en lactosa (La) y lactulosa (Lu) en las fórmulas infantiles.
Tratamiento La (g/L) Lu (mg/L)
Control 119.47a±10.13 27.13a±2.13
UHT 103.83a±9.39 616.53b±44.07
Presión (650 MPa) 109.01a±5.33 36.19a±3.41
a-b Diferentes superíndices, en columnas, indican diferencias significativas (P< 0.05) producidas por el tratamiento.

- 66 -
I.4 Discusión
El uso de un tratamiento UHT para la estabilización final de una formula infantil
basada en distintos ingredientes lácteos produjo una modificación mucho más acusada en sus
componentes proteicos mayoritarios y minoritarios que un tratamiento con alta presión, lo que
puede afectar negativamente a sus propiedades nutricionales.
Se utilizaron los niveles de lactulosa y furosina como índices de daño térmico. La
furosina, indicador de las primeras etapas de la reacción de Maillard, es un aminoácido que se
forma durante la hidrólisis ácida de los compuestos de Amadori, fructosil-lisina, lactulosil-
lisina y maltulosil-lisina (Erbersdobler y Hupe, 1991). La reacción de Maillard se ve
promovida por la temperatura en mayor medida en condiciones de aw intermedia (entre 0.5 y
0.8), que concentra a los reactivos y disminuye su movilidad (Ames, 1990). Es de destacar
que el contenido en furosina de la fórmula de partida fue relativamente elevado (Figura 20), lo
que, de acuerdo con el contenido en proteínas de suero solubles a pH 4.6 (6.6 g/L -Tabla 4-
frente a unos 8 g/L, aproximadamente, de una leche cruda) y su modificación por
lactosilación, tal y como se observa en el análisis por electroforesis capilar (Figura 14),
indicaría que se habría empleado suero y/o leche en polvo en la fabricación de la fórmula
control, puesto que la deshidratación que se produce durante los procesos de atomización, así
como el almacenamiento posterior de los productos en polvo favorecen la reacción de
Maillard. El tratamiento UHT elevó significativamente el contenido en furosina hasta niveles
similares a los observados por otros autores (Guerra-Hernández y col., 2002; Ferrer y col.,
2003; Morales y col., 2004). El contenido en furosina varía dependiendo del tratamiento
térmico utilizado, la concentración azúcares reductores y el tiempo y las condiciones de
almacenamiento, lo que hace que exista un amplio rango en las medidas. Por su parte, el
tratamiento con alta presión no produjo un aumento del contenido en furosina, como se puede
observar en la Figura 20, por lo que no modificó la disponibilidad de la lisina en la fórmula.
La lactulosa es un disacárido producido por la isomerización de la lactosa en
disoluciones alcalinas y durante el tratamiento térmico de la leche, que se forma sobre todo
durante los tratamientos de esterilización y UHT de productos líquidos (Guerra-Hernández,
2002). Este disacárido no está presente en la leche cruda por lo que se le considera un buen
indicador para evaluar el daño térmico (López-Fandiño y Olano, 1999). El contenido en la
fórmula control (aproximadamente 27 mg/L, Tabla 5) se encontró dentro de los márgenes

RESULTADOS Y DISCUSIÓN: CAPÍTULO I
- 67 -
descritos para leche en polvo comercial (en torno a 23-78 mg/L de producto reconstituido,
Corzo y col., 1994), corroborando el uso de productos en polvo para su elaboración. Como
era de esperar, el tratamiento UHT favoreció la isomerización de la lactosa y la formación de
lactulosa (Tabla 5). El contenido en lactulosa en la fórmula UHT fue indicativo de un
calentamiento muy intenso, bastante superior a los encontrados en la bibliografía para
fórmulas comerciales reconstituidas elaboradas a base de leche o de leche con proteínas de
suero, (29-108 mg/L y 97-132 mg/L, respectivamente, Pereyra-Gonzáles y col., 2003) y más
típico de leche estéril (la Federación Internacional de Lechería propuso en 1992 el límite de
600 mg/L de lactulosa para distinguir la leche esterilizada de la leche UHT).
El tratamiento UHT condujo a una notable desnaturalización de las seroproteínas (el
nitrógeno soluble a pH 4.6 disminuyó en torno al 33% respecto a la fórmula control). En la
fórmula infantil tratada con presión la desnaturalización fue menor (con una disminución de,
aproximadamente, un 18%) (Tabla 4, Figura 5). Los tratamientos térmicos y los tratamientos
con alta presión alteran la estructura de las seroproteínas lácteas. Así, el desplegamiento
producido en la β-Lg tanto por temperatura como por presión produce la exposición del grupo
sulfhidrilo libre, favoreciendo que reaccione inter e intramolecularmente con otros grupos
sulfhidrilo o puentes disulfuro de proteínas como la propia β-Lg, la κ-CN y la αs2-CN,
formando de esta manera agregados, que pueden ser solubles o insolubles (Patel y col., 2006).
En tratamientos térmicos de larga duración (30 min) aplicados a leche reconstituida, también
se ha observado un aumento del contenido de la β-Lg y α-La desnaturalizadas a partir de los
70ºC, aunque la desnaturalización no es completa hasta los 100ºC, aproximadamente (Anema,
2008a). Por otra parte, debe tenerse en cuenta que la glicosilación aumenta la termoestabilidad
de las proteínas de suero y conduce a una menor oligomerización de éstas como consecuencia
de su desnaturalización (Mulsow y col., 2009). En el caso de los tratamientos con presión, en
leche reconstituida se ha observado que la α-La es más resistente a la desnaturalización por
presión que la β-Lg, comenzando a desnaturalizarse a presiones ≤500 MPa (30 min) mientras
que la β-Lg lo hace a presiones ≥200 MPa. A 600 MPa el contenido de β-Lg y α-La
desnaturalizadas es de en torno al 80 y 20%, respectivamente (Anema, 2008a).
El estudio realizado por EC mostró que la solubilización micelar en la fórmula
presurizada (Figura 16) fue diferente a la de una leche cruda presurizada. López-Fandiño y
col. (1998) observaron que la presión induce la liberación de todas las caseínas a la fracción

- 68 -
soluble, debido a la disgregación micelar que produce, siendo el orden de solubilización β-
CN> κ-CN> αs1-CN> αs0-CN> αs2-CN. Sin embargo, en la fórmula presurizada no se observó
solubilización de caseínas por el tratamiento aplicado. Este hecho podría deberse a que se
sometió al tratamiento con alta presión a una fórmula elaborada a partir de productos en polvo
reconstituidos, en la que, como se mencionó en el párrafo anterior ya había proteínas de suero
desnaturalizadas (Figura 17). Kelly y col. (2002) indicaron que la desnaturalización previa, ya
sea por calor o por presión de las proteínas de suero y su consecuente interacción con la κ-CN
podría proteger a la micela de disgregarse por la presión. En el caso de la leche cruda,
probablemente, las proteínas de suero se desnaturalizan bajo la alta presión, interaccionando
con fragmentos micelares, lo que podría limitar su reasociación al liberarse la presión. En el
caso de las fórmulas, las micelas podrían tener ya proteínas desnaturalizadas unidas a su
superficie debido a un tratamiento térmico previo, lo que facilitaría su reagregación (López-
Fandiño, 2006a).
En la actualidad hay una gran tendencia en suplementar a las fórmulas infantiles con
Lf, ya que esta proteína presenta actividad antibacterina, antiviral, y antifúngica y está
implicada en la respuesta inmune y en la regulación y consumo del hierro intestinal (Jenssen y
Hancock, 2009; Tomita y col., 2009). Además, la Lf está en una concentración más elevada
en la leche humana que en la leche de vaca (Fox y McSweeney, 1998). Junto a la IgA,
representa entre el 30 y 40% del contenido en proteínas de la leche humana (Chierici y Vigi,
1994). Indyk y Filonzi (2005) estimaron un contenido de 0.4 mg/100g y 14.8 mg/100g para
formulas no suplementadas y suplementadas con Lf usando el mismo sistema de detección,
aunque su contenido puede variar dependiendo del método de análisis utilizado (Indyk y
Filonzi, 2005) y de la composición de la fórmula. La Lf se desnaturaliza a temperaturas de
pasteurización (Paulsson y col., 1993). Scharcz y col. (2008) aportaron espectros de
discroismo circular a 70ºC que muestran que la estructura secundaria de la Lf cambia drástica
e irreversiblemente cuando la temperatura aumenta gradualmente. Sin embargo, en una
simulación de una pasteurización (72ºC durante 20 seg) no observaron ningún efecto en la
estructura secundaria de la proteína, lo que indica que, si la temperatura, se alcanza en un
breve periodo de tiempo, la Lf no pasa por el mismo estado de desplegamiento observado
cuando el aumento de la temperatura es gradual. Además, se ha observado que la forma apo
de esta proteína es más sensible al calor que la forma holo y, a pH 7, la apo LF se
desnaturaliza entre 60 y 65ºC mientras que la holo forma permanece nativa hasta los 80ºC

RESULTADOS Y DISCUSIÓN: CAPÍTULO I
- 69 -
(Sreedhara y col., 2010). Por otra parte, cuando la leche se calienta, la Lf establece
interacciones intermoleculares vía puentes disulfuro con las caseínas y otras proteínas séricas
(Brisson y col., 2007), lo que explicaría también la menor detección de Lf por anticuerpos en
leches tratadas por UHT a pesar de que la banda correspondiente a la Lf se resolubilice en el
tampón de la muestra usado en PAGE-SDS.
La IgA es otra proteína importante en la composición de las fórmulas, porque
interviene en los mecanismos de defensa gastrointestinal (Bakker-Zierikzee y col., 2006).
Ford y col. (1977) observaron una reducción del contenido de IgA en leche humana
pasteurizada (56ºC, 30 min) del 10%, pasando al 80% en tratamientos con temperaturas más
elevadas (80ºC, 15 min). Además, Viazis y col. (2007) observaron que la inmunoreactividad y
la concentración de IgA detectada en leche humana presurizada (400 MPa durante 30, 60, 90
y 120 min) fue significativamente mayor (P< 0.05) que en la misma leche pasteurizada. Sin
embargo, estos autores encontraron una menor concentración de IgA en la leche presurizada
que en la leche sin tratar, a diferencia de nuestro estudio, donde la fórmula presurizada
presentó un llamativo aumento del contenido de IgA. Esta diferencia podría deberse al hecho
de que el tiempo de presurización utilizado en nuestro caso fue menor, de 5 min frente a 30
min, lo que generaría menores cambios estructurales, o incluso podría producir un
desplegamiento que facilitase la unión al anticuerpo.
I.5. Conclusiones
Los resultados obtenidos sugieren que la aplicación de un tratamiento con alta presión
a 650 MPa durante 5 min a fórmulas infantiles, sustituyendo el tratamiento térmico final al
que se ven sometidas durante su fabricación, no produce isomerización de la lactosa, ni
avance de la reacción de Maillard y conduce a una menor desnaturalización de las proteínas
de suero, por lo que mejora la calidad nutritiva de las fórmulas. Además, la estructura de las
proteínas minoritarias del suero, lactoferrina e inmunoglobulina A secretora, se ve menos
afectada por el tratamiento con presión que por el tratamiento UHT, medido por su unión a
anticuerpos, por lo que se podrían obtener fórmulas infantiles con mejores actividades
biológicas para los recién nacidos. a pesar de producir menores cambios químicos, la
efectividad de los tratamientos con alta presión está condicionada a la comprobación de que
conducen a productos de aceptable calidad microbiológica.
.

CAPÍTULO II
Tratamientos con alta presión en equipos a escala industrial y piloto.
Efecto de las condiciones del tratamiento en la distribución proteica
en diferentes fracciones lácteas


RESULTADOS Y DISCUSIÓN: CAPÍTULO II
- 73 -
II.1. Introducción
La creciente demanda de los consumidores por productos que presenten nuevas o
mejores características sensoriales y nutricionales, junto con una mayor durabilidad, ha retado
a la industria alimentaria a desarrollar técnicas capaces de mantener los alimentos seguros por
más tiempo y de mejorar sus propiedades (Norton y Sun, 2008). En concreto, el procesado
con alta presión, que ha experimentado un enorme crecimiento en los últimos 20 años
llegando a ser una realidad industrial, puede facilitar la elaboración de productos similares a
los alimentos frescos con una mayor vida útil, además de proporcionar nuevos productos con
mejores cualidades, principalmente a través de cambios en la conformación de las proteínas
alimentarias que modifican sus propiedades funcionales (López-Fandiño, 2006b).
La leche fue el primer alimento en someterse a un tratamiento con alta presión (Hite,
1899), sin embargo, hasta el momento, hay solamente unos pocos productos lácteos
presurizados comerciales, como yogurt o calostro (Kelly y Zeece, 2009). Las aplicaciones
industriales de las altas presiones están orientadas principalmente a mejorar la seguridad y la
estabilidad, mientras que otras aplicaciones encaminadas a inducir cambios estructurales que
cambien la funcionalidad de las proteínas lácteas, se han explotado más escasamente. Los
tratamientos con alta presión producen importantes modificaciones en las proteínas y en el
balance mineral de la leche. Como resultado, se produce disgregación y reagregación de las
micelas de caseína, liberación de caseínas solubles y desnaturalización de las proteínas de
suero (Huppertz y col., 2006a; López-Fandiño, 2006a). Como consecuencia de la ruptura
micelar, se forman nuevas estructuras proteicas difíciles de lograr mediante el uso de métodos
convencionales de procesado, las cuales influyen en las propiedades de coagulación
enzimática, favorecen la coagulación ácida y pueden actuar como unidades funcionales para
la encapsulación y liberación de nutrientes en productos lácteos (Orlien y col., 2010). En
cualquier caso, la intensidad, la duración y la temperatura de los tratamientos, así como, del
pH y de la concentración de proteína influyen en los cambios originados por las altas
presiones (Law y col., 1998; López-Fandiño y col., 1998; Arias y col., 2000; Huppertz y col.,
2004b; Anema y col., 2005a; Huppertz y De Kruif, 2006; Anema, 2008b; Orlien y col., 2010).
Actualmente, el efecto de la mayoría de estas variables se conoce bien, pero rara vez se ha
estudiado la influencia del tipo de equipo de alta presión (Anema y col., 2005b).

- 74 -
Los equipos de alta presión están formados por una cámara, que es un cilindro lleno de
un fluido transmisor de la presión, generalmente agua potable o una mezcla de agua y aceite;
una o varias bombas generadoras de la presión; un dispositivo de control de la temperatura y
el sistema de control del equipo (Huppertz y col., 2002). Los equipos de procesado con alta
presión presentan diferentes características como el tamaño (existiendo a escala de
laboratorio, piloto e industrial), la orientación, vertical u horizontal, etc. Las especificaciones
del equipo indican la presión máxima de la cámara, su diámetro interno, la altura o longitud
interior de la cámara, la velocidad de subida y bajada de la presión, el fluido transmisor de la
presión y el rango de temperatura. En la industria alimentaria, se utilizan cámaras de
presurización con cientos de litros de volumen, que trabajan normalmente en el intervalo de
250 a 600 MPa, que se mantienen entre 1-8 minutos con tiempos rápidos de subida y bajada
de la presión, puesto que la duración del ciclo o la velocidad de producción son factores
importantes a controlar. También hay equipos a escala de laboratorio, capaces de alcanzar
presiones de hasta 1400 MPa, con diseños específicos o versátiles (Bermúdez-Aguirre y
Barbosa-Cánovas, 2011).
El objetivo de este trabajo ha sido estudiar la influencia del tipo de equipo de alta
presión y de la velocidad de despresurización en la distribución de las proteínas lácteas entre
las fases coloidal y soluble de la leche y en la desnaturalización de las proteínas de suero, en
tratamientos de presión entre 250 y 650 MPa. Además, también se presurizaron sobrenadantes
de ultracentrifugación obtenidos de leche desnatada con fines comparativos.
II.2. Materiales y Métodos
La Figura 21 resume el procedimiento experimental seguido en este trabajo.
II.2.1. Preparación de las muestras
Para llevar a cabo los experimentos de presurización, se utilizó tanto leche desnatada
de vaca como sobrenadantes de ultracentrifugación (SU) obtenidos a partir de ésta. Tras la
recogida de la leche cruda en una granja local, se procedió a su desnatado, calentándola a
37ºC durante 30 min y centrifugándola a 3000 x g, 20ºC durante 30 min. Finalmente, el
sobrenadante se filtró por lana de vidrio, para así retener los restos de grasa. Por otro lado, los
SU se obtuvieron ultracentrifugando alícuotas de leche desnatada a 100000 x g, 20ºC durante

RESULTADOS Y DISCUSIÓN: CAPÍTULO II
- 75 -
1 h en una ultracentrífuga Beckman L70 (Beckman Instruments Inc., San Ramón, CA) usando
el rotor 70 Ti (Figura 1). Estos sobrenadantes se guardaron en refrigeración a 4ºC por un
máximo de 18 h antes de los tratamientos con alta presión.
II.2.2. Tratamientos de alta presión
Las muestras (leche desnatada y SU), atemperadas a 15ºC, se presurizaron a 250, 350,
450, 550 y 650 MPa en un equipo industrial y en un equipo piloto (Figura 1). El equipo
industrial estaba formado por una cámara de presurización con capacidad de 120 L, con agua
como fluido transmisor de la presión y 3 bombas para generar la presión hidráulicamente
(Wave 6000/120, NC Hyperbaric, Burgos, España). El equipo piloto utilizado (modelo TE
9000, Thiot Ingenierie, NC Hyperbaric, Bretenoux, France-Burgos, España) tenía una cámara
de presurización de 2 L de capacidad, recubierta por una camisa de agua capaz de regular la
temperatura (5-90ºC), y una bomba que genera la presión mecánicamente. En este caso, el
fluido transmisor de presión fue aceite de silicona. A diferencia del industrial, el equipo piloto
permitió ajustar el tiempo de subida y bajada de la presión.
La presión aumentó a una velocidad de 3.04 y 6 MPa/seg en el industrial y en el piloto
respectivamente, se mantuvo durante 5 min y se liberó en 1.5 seg en el equipo industrial
(tratamientos IR – equipo industrial con despresurización rápida-) y en 30 seg o 5 min en el
equipo piloto (PR- equipo piloto con despresurización rápida y PL- equipo piloto con
despresurización lenta, respectivamente).
La temperatura del fluido transmisor de la presión se registró durante los tratamientos.
La temperatura inicial fue 14 ± 2ºC, la cual aumentó durante la subida de la presión. Así,
durante la fase de mantenimiento de la presión, la temperatura fue de 25.9 y 39.5ºC a 350
MPa o de 32.5 y 46.9ºC a 650 MPa, en los equipos industrial y piloto, respectivamente.
Se realizaron 2 experimentos independientes por cada tratamiento de alta presión en el
mismo día. Tras los tratamientos de alta presión, las muestras se almacenaron toda la noche a
4ºC antes de hacer los fraccionamientos y los análisis.

Figura 21. Esquema de la preparación, tratamientos con alta presión, fraccionamiento y análisis de las muestras
Leche cruda de vaca
Lechedesnatada
NC HyperbaricWave 6000/120
Subida de presión3.04 MPa/seg
Subida de presión6 MPa/seg
Thiot EngineerTE 9000
SobrenadanteUltracentrif.
Lechedesnatada
Lechedesnatada
Lechedesnatada
SobrenadanteUltracentrif.
SobrenadanteUltracentrif.
SobrenadanteUltracentrif.
Ultracentrifugación100000 x g, 20ºC, 1h
Lechedesnatada
Lechedesnatada
SobrenadanteUltracentrif.
Lechedesnatada
SobrenadanteUltracentrif.
LechepresurizadaI. 250 MPa
LechepresurizadaI. 350 MPa
LechepresurizadaI. 450 MPa
LechepresurizadaI. 550 MPa
LechepresurizadaI. 650 MPa
Sobr. UCpresurizadoI. 250 MPa
Sobr. UCpresurizadoI. 350 MPa
Sobr. UCpresurizadoI. 450 MPa
Sobr. UCpresurizadoI. 550 MPa
Sobr. UCpresurizadoI.650 MPa
Lechepresurizada
PR. 250 MPa
Lechepresurizada
PR. 350 MPa
Lechepresurizada
PR. 450 MPa
Lechepresurizada
PR. 550 MPa
Lechepresurizada
PR. 650 MPa
Sobr. UCpresurizado
PR. 250 MPa
Sobr. UCpresurizado
PR. 350 MPa
Sobr. UCpresurizado
PR. 450 MPa
Sobr. UCpresurizado
PR. 550 MPa
Sobr.UCpresurizadoPR.650 MPa
Lechepresurizada
PL. 250 MPa
Lechepresurizada
PL. 350 MPa
Lechepresurizada
PL. 450 MPa
Lechepresurizada
PL. 550 MPa
Lechepresurizada
PL. 650 MPa
Sobr. UCpresurizado
PL. 250 MPa
Sobr. UCpresurizado
PL. 350 MPa
Sobr. UCpresurizado
PL. 450 MPa
Sobr. UCpresurizado
PL. 550 MPa
Sobr. UCpresurizadoPL.650 MPa
Lechepresurizada
SobrenadanteUltracentrif.presurizado
SU
SU-P
Fracciones
Precipitacióna pH 4.6
Ultracentrifugación100000 x g, 20ºC, 1h
Análisis
Despresurización rápida(R) 30 seg
Despresurización lenta(L) 5 min
Equipo Industrial (I)
Equipo Piloto (P)
Despresurización Instantánea(R) 1.5 seg
4.6Precipitación
a pH 4.6
Ultracentrifugación100000 x g, 20ºC, 1h
ELECTROFORESISCAPILAR
NITRÓGENO(KJELDAHL)
PAGE-SDS
15ºC, 5 min
Desnatado y Preparación de las muestras
Tratamiento con alta presión Fraccionamiento Análisis
3000 x g, 20ºC, 30 min
PROTEÍNAS
Aná
lisis
4.6
Leche cruda de vaca
Lechedesnatada
Lechedesnatada
NC HyperbaricWave 6000/120NC HyperbaricWave 6000/120
Subida de presión3.04 MPa/seg
Subida de presión6 MPa/seg
Thiot EngineerTE 9000
Thiot EngineerTE 9000
SobrenadanteUltracentrif.SobrenadanteUltracentrif.
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
SobrenadanteUltracentrif.SobrenadanteUltracentrif.
SobrenadanteUltracentrif.SobrenadanteUltracentrif.
SobrenadanteUltracentrif.SobrenadanteUltracentrif.
Ultracentrifugación100000 x g, 20ºC, 1h
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
SobrenadanteUltracentrif.
SobrenadanteUltracentrif.
Lechedesnatada
Lechedesnatada
SobrenadanteUltracentrif.SobrenadanteUltracentrif.
LechepresurizadaI. 250 MPa
LechepresurizadaI. 350 MPa
LechepresurizadaI. 450 MPa
LechepresurizadaI. 550 MPa
LechepresurizadaI. 650 MPa
Sobr. UCpresurizadoI. 250 MPa
Sobr. UCpresurizadoI. 350 MPa
Sobr. UCpresurizadoI. 450 MPa
Sobr. UCpresurizadoI. 550 MPa
Sobr. UCpresurizadoI.650 MPa
LechepresurizadaI. 250 MPa
LechepresurizadaI. 350 MPa
LechepresurizadaI. 450 MPa
LechepresurizadaI. 550 MPa
LechepresurizadaI. 650 MPa
LechepresurizadaI. 250 MPa
LechepresurizadaI. 250 MPa
LechepresurizadaI. 350 MPa
LechepresurizadaI. 350 MPa
LechepresurizadaI. 450 MPa
LechepresurizadaI. 450 MPa
LechepresurizadaI. 550 MPa
LechepresurizadaI. 550 MPa
LechepresurizadaI. 650 MPa
LechepresurizadaI. 650 MPa
Sobr. UCpresurizadoI. 250 MPa
Sobr. UCpresurizadoI. 350 MPa
Sobr. UCpresurizadoI. 450 MPa
Sobr. UCpresurizadoI. 550 MPa
Sobr. UCpresurizadoI.650 MPa
Sobr. UCpresurizadoI. 250 MPa
Sobr. UCpresurizadoI. 250 MPa
Sobr. UCpresurizadoI. 350 MPa
Sobr. UCpresurizadoI. 350 MPa
Sobr. UCpresurizadoI. 450 MPa
Sobr. UCpresurizadoI. 450 MPa
Sobr. UCpresurizadoI. 550 MPa
Sobr. UCpresurizadoI. 550 MPa
Sobr. UCpresurizadoI.650 MPa
Sobr. UCpresurizadoI.650 MPa
Lechepresurizada
PR. 250 MPa
Lechepresurizada
PR. 350 MPa
Lechepresurizada
PR. 450 MPa
Lechepresurizada
PR. 550 MPa
Lechepresurizada
PR. 650 MPa
Sobr. UCpresurizado
PR. 250 MPa
Sobr. UCpresurizado
PR. 350 MPa
Sobr. UCpresurizado
PR. 450 MPa
Sobr. UCpresurizado
PR. 550 MPa
Sobr.UCpresurizadoPR.650 MPa
Lechepresurizada
PR. 250 MPa
Lechepresurizada
PR. 350 MPa
Lechepresurizada
PR. 450 MPa
Lechepresurizada
PR. 550 MPa
Lechepresurizada
PR. 650 MPa
Lechepresurizada
PR. 250 MPa
Lechepresurizada
PR. 250 MPa
Lechepresurizada
PR. 350 MPa
Lechepresurizada
PR. 350 MPa
Lechepresurizada
PR. 450 MPa
Lechepresurizada
PR. 450 MPa
Lechepresurizada
PR. 550 MPa
Lechepresurizada
PR. 550 MPa
Lechepresurizada
PR. 650 MPa
Lechepresurizada
PR. 650 MPa
Sobr. UCpresurizado
PR. 250 MPa
Sobr. UCpresurizado
PR. 350 MPa
Sobr. UCpresurizado
PR. 450 MPa
Sobr. UCpresurizado
PR. 550 MPa
Sobr.UCpresurizadoPR.650 MPa
Sobr. UCpresurizado
PR. 250 MPa
Sobr. UCpresurizado
PR. 250 MPa
Sobr. UCpresurizado
PR. 350 MPa
Sobr. UCpresurizado
PR. 350 MPa
Sobr. UCpresurizado
PR. 450 MPa
Sobr. UCpresurizado
PR. 450 MPa
Sobr. UCpresurizado
PR. 550 MPa
Sobr. UCpresurizado
PR. 550 MPa
Sobr.UCpresurizadoPR.650 MPa
Sobr.UCpresurizadoPR.650 MPa
Lechepresurizada
PL. 250 MPa
Lechepresurizada
PL. 350 MPa
Lechepresurizada
PL. 450 MPa
Lechepresurizada
PL. 550 MPa
Lechepresurizada
PL. 650 MPa
Sobr. UCpresurizado
PL. 250 MPa
Sobr. UCpresurizado
PL. 350 MPa
Sobr. UCpresurizado
PL. 450 MPa
Sobr. UCpresurizado
PL. 550 MPa
Sobr. UCpresurizadoPL.650 MPa
Lechepresurizada
PL. 250 MPa
Lechepresurizada
PL. 350 MPa
Lechepresurizada
PL. 450 MPa
Lechepresurizada
PL. 550 MPa
Lechepresurizada
PL. 650 MPa
Lechepresurizada
PL. 250 MPa
Lechepresurizada
PL. 250 MPa
Lechepresurizada
PL. 350 MPa
Lechepresurizada
PL. 350 MPa
Lechepresurizada
PL. 450 MPa
Lechepresurizada
PL. 450 MPa
Lechepresurizada
PL. 550 MPa
Lechepresurizada
PL. 550 MPa
Lechepresurizada
PL. 650 MPa
Lechepresurizada
PL. 650 MPa
Sobr. UCpresurizado
PL. 250 MPa
Sobr. UCpresurizado
PL. 350 MPa
Sobr. UCpresurizado
PL. 450 MPa
Sobr. UCpresurizado
PL. 550 MPa
Sobr. UCpresurizadoPL.650 MPa
Sobr. UCpresurizado
PL. 250 MPa
Sobr. UCpresurizado
PL. 250 MPa
Sobr. UCpresurizado
PL. 350 MPa
Sobr. UCpresurizado
PL. 350 MPa
Sobr. UCpresurizado
PL. 450 MPa
Sobr. UCpresurizado
PL. 450 MPa
Sobr. UCpresurizado
PL. 550 MPa
Sobr. UCpresurizado
PL. 550 MPa
Sobr. UCpresurizadoPL.650 MPa
Sobr. UCpresurizadoPL.650 MPa
Lechepresurizada
Lechepresurizada
SobrenadanteUltracentrif.presurizado
SobrenadanteUltracentrif.presurizado
SU
SU-P
Fracciones
Precipitacióna pH 4.6
Ultracentrifugación100000 x g, 20ºC, 1h
Análisis
Despresurización rápida(R) 30 seg
Despresurización lenta(L) 5 min
Equipo Industrial (I)
Equipo Piloto (P)
Despresurización Instantánea(R) 1.5 seg
4.6Precipitación
a pH 4.6
Ultracentrifugación100000 x g, 20ºC, 1h
ELECTROFORESISCAPILAR
NITRÓGENO(KJELDAHL)
PAGE-SDS
15ºC, 5 min
Desnatado y Preparación de las muestras
Tratamiento con alta presión Fraccionamiento Análisis
3000 x g, 20ºC, 30 min
PROTEÍNAS
Aná
lisis
4.6

RESULTADOS Y DISCUSIÓN: CAPÍTULO II
- 77 -
II.3. Resultados
II.3.1. Caseínas no sedimentables y proteínas de suero presentes en la fracción soluble de
leche desnatada tratada con altas presiones
En la Figura 22 se muestra el contenido proteico total de los SU de las muestras de
leche desnatada presurizada. El análisis estadístico reveló que el nivel de presión y el tipo de
equipo (IR, PR y PL) influyeron significativamente (P< 0.01) en la concentración de proteínas
solubles y que existió interacción entre ambos factores.
Los SU de las muestras de leche presurizadas en el equipo piloto (PR y PL) mostraron
mayor contenido en proteína soluble que las tratadas en el equipo industrial (IR) a todos los
niveles de presión estudiados (P< 0.01). Los mayores contenidos en proteínas no
sedimentables se encontraron a 550, 450 y 250 MPa tras los tratamientos IR, PR y PL,
respectivamente.
Figura 22. Contenido proteico (g/L) de los sobrenadantes de ultracentrifugación obtenidos a partir
de leche desnatada control (0.1 MPa) y de leche presurizada a 250, 350, 450, 550 y 650 MPa a 15ºC
durante 5 min, en un equipo industrial (IR) y en un equipo piloto con despresurización rápida (PR) y
lenta (PL). Las barras representan los intervalos de confianza del 95%.
IR PR PLIR PR PL
Presión (MPa)
Con
teni
do p
rote
ico
(g/L
)
10
12
14
16
18
20
0.1 250 350 450 550 650
Presión (MPa)
Con
teni
do p
rote
ico
(g/L
)
10
12
14
16
18
20
0.1 250 350 450 550 650

- 78 -
El estudio de los SU por PAGE-SDS permitió estimar cualitativamente la composición
en proteínas individuales solubles en estas fracciones (Figura 33). Las bandas
correspondientes a las caseínas en los sobrenadantes obtenidos a partir de leche presurizada
(calles 3-7) en ambos equipos presentaron una intensidad mayor de color que el obtenido a
partir de leche sin presurizar (calle 2), lo cual indica que la aplicación de presión hace que sea
mucho mayor la liberación de caseínas a esta fracción. Este hecho corrobora el aumento en el
contenido proteico total de estas muestras. Además, la intensidad de las bandas
correspondientes a las caseínas aumentó ligeramente debido al aumento de la presión en el
equipo industrial y piloto. Sin embargo, no se pudo comparar si este efecto fue similar en los
3 tratamientos IR, PR y PL debido a que la tinción de los geles se realizó en momentos
diferentes y la intensidad del color pudo cambiar.
Figura 23. PAGE-SDS de los sobrenadantes obtenidos por ultracentrifugación de leche presurizada
en un equipo industrial (IR) y en un equipo piloto con despresurización rápida (PR) y lenta (PL).
Calles: 1: Marcador de pesos moleculares; 2 a 7: tratamiento de 0.1, 250, 350, 450, 550 y
650 MPa.
El análisis de las distintas proteínas individuales se realizó por electroforesis capilar.
La Figura 24 muestra los electroforegramas obtenidos del análisis de los sobrenadantes de
ultracentrifugación por esta técnica. En ellos se puede observar que el tratamiento con el
equipo piloto a todas las presiones estudiadas produjo una mayor solubilización de caseínas
comparado con el equipo industrial (proteínas marcadas como 4-9 en la Figura 24). Además,
si se compara los dos tratamientos realizados en el equipo piloto, se produjo una mayor
solubilización de caseínas con la despresurización lenta (PL) al menos a presiones de 250 y
350 MPa (Figura 24 c y d).
IR PR PL
1 2 3 4 5 6 7
β-CN
α-Laβ-Lgκ-CN
LfBSAIg
αs2-CNαs1-CN
1 2 3 4 5 6 7 1 2 3 4 5 6 7
IR PR PL
1 2 3 4 5 6 7
β-CNβ-CN
α-Laβ-Lgκ-CN
LfBSAIg
αs2-CNαs1-CN
1 2 3 4 5 6 7 1 2 3 4 5 6 7
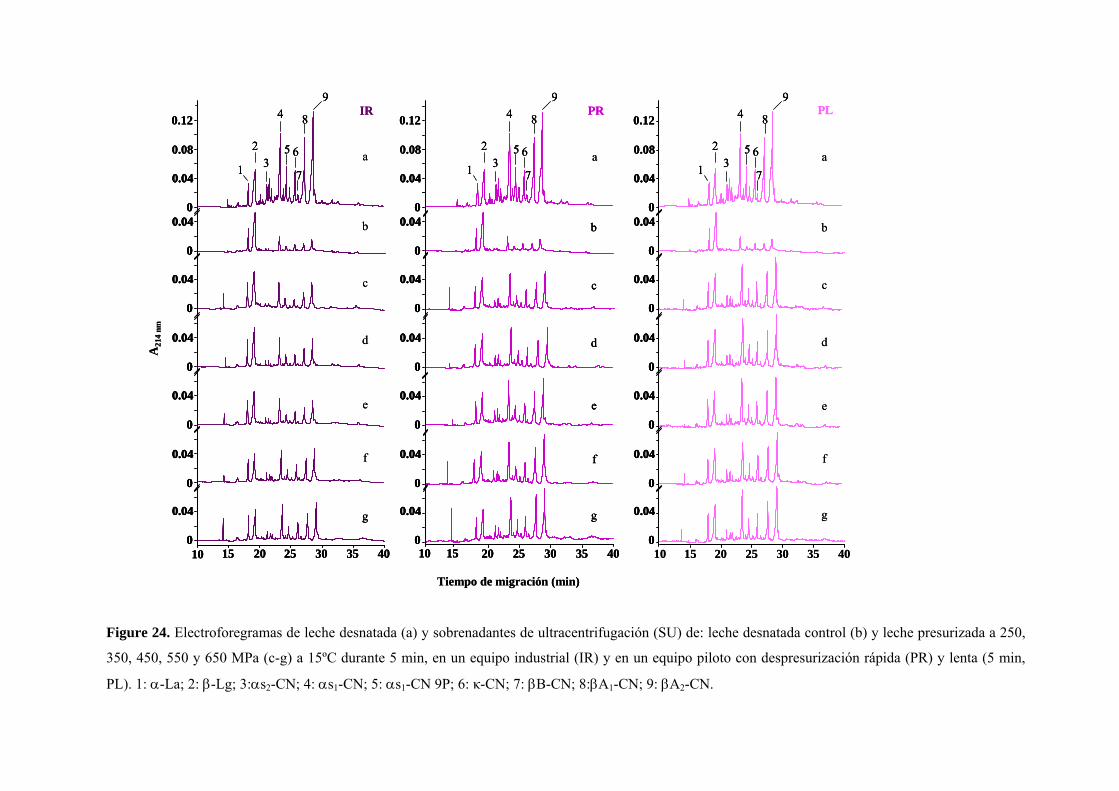
Figure 24. Electroforegramas de leche desnatada (a) y sobrenadantes de ultracentrifugación (SU) de: leche desnatada control (b) y leche presurizada a 250,
350, 450, 550 y 650 MPa (c-g) a 15ºC durante 5 min, en un equipo industrial (IR) y en un equipo piloto con despresurización rápida (PR) y lenta (5 min,
PL). 1: α-La; 2: β-Lg; 3:αs2-CN; 4: αs1-CN; 5: αs1-CN 9P; 6: κ-CN; 7: βB-CN; 8:βA1-CN; 9: βA2-CN.
c
d
e
f
b
10 15 20 25 30 35 40
g
Tiempo de migración (min)
c
d
e
f
10 15 20 25 30 35 40
g
b
a a
10 15 20 25 30 35 40
0
0.04
0.08
0.12
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0.08
0.12
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0.08
0.12
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
A21
4 nm
PRIR
g
f
e
d
c
b
a
PL
23
4
5
9
8
61 7
23
4
5
9
8
61 7
23
4
5
9
8
61 7
c
d
e
f
b
10 15 20 25 30 35 40
g
10 15 20 25 30 35 40
g
Tiempo de migración (min)
cc
dd
ee
ff
10 15 20 25 30 35 40
g
10 15 20 25 30 35 40
g
bb
a a
10 15 20 25 30 35 40
0
0.04
0.08
0.12
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0.08
0.12
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0.08
0.12
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0.08
0.12
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0.08
0.12
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0.08
0.12
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
0
0.04
A21
4 nm
PRIR
g
f
e
d
c
b
a
PL
23
4
5
9
8
61 7
23
4
5
9
8
61 7
23
4
5
9
8
61 7
23
4
5
9
8
61 7
23
4
5
9
8
61 7
23
4
5
9
8
61 7

- 80 -
La cuantificación de las proteínas individuales observadas por EC permitió estudiar
con más detalle el efecto de los tratamientos de presión aplicados en la liberación de caseínas
individuales a la fase soluble (Tabla 6). Como ya se había indicado anteriormente, el
tratamiento IR produjo una solubilización significativamente menor de todas las caseínas (P<
0.05) que el tratamiento PR. Los niveles de αS-caseínas solubles (αS1-9P-, αS1- o αS2-CN),
que en la leche control se correspondían con el 12.4-15% del contenido individual de cada
proteína, aumentaron hasta el 17.3-29.6% y 34.1-45.2% después de los tratamientos IR y PR a
250 MPa, respectivamente, mientras que las β-caseínas (βA1-, βA2- o β-CN B) aumentaron
del 10.5-18.4% (control) al 21.8-25.4 (IR) y 35.7-30.3% (PR). A esta presión (250 MPa), la κ-
CN soluble supuso el 28.8 y 50.8% de su contenido en leche después de los tratamientos IR y
PR, respectivamente. A mayores presiones, en términos generales, en el equipo industrial las
caseínas no sedimentables aumentaron ligeramente o se estabilizaron a 350 y 450 MPa,
observándose la máxima solubilización después de los tratamientos a 550 y 650 MPa. Esta
tendencia fue similar en el equipo piloto con despresurización rápida, aunque en este caso, el
contenido de caseínas no sedimentables alcanzó su máximo después de tratamientos de
presión a 450 y 550 MPa.
Las proteínas de suero no sedimentables presentes en los SU, principalmente la β-Lg,
disminuyeron gradualmente con el nivel de presión en ambos equipos (Figura 24 y Tabla 6).
Esto explicaría la reducción del contenido total en proteína soluble observado a las presiones
máximas (principalmente a 550 y 650 MPa después de los tratamientos IR y PR,
respectivamente, Figura 22). Aunque el nivel de β-Lg disminuyó algo más lentamente
después del tratamiento IR que después del PR, las diferencias no fueron significativas
excepto a 350 MPa.

RESULTADOS Y DISCUSIÓN: CAPÍTULO II
- 81 -
Tabla 6. Proteínas individuales presentes en los sobrenadantes de ultracentrifugación obtenidos a
partir de leche desnatada control (0.1 MPa) y leche presurizada a 250, 350, 450, 550 and 650 MPa, a
15ºC, durante 5 min, en un equipo industrial (IR) y un equipo piloto con despresurización rápida
(PR) (los valores se representan como porcentaje del contenido individual de cada caseína en la
leche cruda desnatada, estimado por EC).
Presión (MPa) Proteínas
Equipo alta
presión 0.1 250 350 450 550 650
SD total
IR A117.7b A116.9b A116.2b B112.8b A109.0ab A94.9a 4.0 α-La
PR A117.7c A107.4b A99.6ab A99.0ab A101.3ab A93.4a 2.9
IR A106.2d A100.8d B91.8c A76.4b A71.1ab A65.48a 2.4 β-Lg
PR A106.2d A93.3c A77.5b A70.3ab A71.3ab A63.6a 2.6
IR A14.4a A17.3a A17.2a A19.8ab A28.4b A29.3b 1.5 αS2-CN
PR A14.4a B34.1b B39.7c B46.2de B50.8e B43.5cd 1.4
IR A12.4a A24.1bc A21.1b A22.7b A27.5c A33.5d 0.8 αS1-CN
PR A12.4a B39.7b B40.5b B42.7b B44.3b B43.6b 1.5
IR A15.0a A29.6c A23.2b A26.2bc A30.3cd A35.0d 2.3 αS1-CN 9P PR A15.0a B45.2b B47.2b B49.1bc B57.4c B46.0bc 2.5
IR A19.0a A28.2bc A24.9b A28.5c A36.4d A39.2d 1.0
κ-CN PR A19.0a B50.8b B51.5b B55.1b B56.3b B55.8b 2.1
IR A18.4a A25.4b A23.6b A23.1b A30.5c A34.2c 1.0
β-CN B PR A18.4a B37.0b B42.1b B42.2b B46.0b A39.3b 2.2
IR A10.5a A21.8b A18.4b A19.6b A27.2c A31.2c 1.0
β A1-CN PR A10.5a B35.7b B37.7b B40.9b B45.2b B46.4b 2.2
IR A13.5a A25.2c A22.6b A22.9b A30.1c A34.9c 1.7
β A2-CN PR A13.5a B39.3b B41.0bc B44.4bc B46.6bc B47.7c 2.0
a-d Diferentes superíndices, a la derecha y en filas, indican diferencias significativas (P< 0.05) producidas por el tratamiento de presión. A-B Diferentes superíndices, a la izquierda y en columnas para cada proteína, indican diferencias significativas (P< 0.05) producidas por el equipo de alta presión. SD total: desviación estándar de los valores obtenidos para cada proteína a todas las presiones.

- 82 -
II.3.2. Desnaturalización de las proteínas del suero en leche desnatada tratada por altas
presiones
Los contenidos proteicos totales de las fracciones solubles a pH 4.6 de la leche
desnatada presurizada se muestran en la Figura 25. En todos los tratamientos, la cantidad de
proteína disminuyó significativamente con el nivel de presión (P< 0.01) debido a la
desnaturalización de las proteínas de suero. Se encontraron diferencias significativas entre
ambos equipos, IR y PR, y el equipo piloto con despresurización lenta, PL, a presiones ≥ 450
MPa. La desnaturalización de proteínas siguió el orden: PL> PR> IR.
Figura 25. Proteína soluble a pH 4.6 (g/L) de leche control (0.1 MPa) y leche presurizada a 250,
350, 450, 550 y 650 MPa, a 15ºC durante 5 min, en un equipo industrial (IR) y un equipo piloto con
despresurización rápida (PR) y lenta (PL). Las barras representan un intervalo de confianza del 95%.
Mediante el análisis de estas fracciones por PAGE-SDS se pudo determinar con mayor
precisión la causa de la disminución del contenido proteico por el aumento de la presión
(Figura 26). En estas fracciones, y como era de esperar, las proteínas predominantes fueron
las del suero. El aumento de la presión de 250 a 650 produjo una progresiva disminución del
contenido de β-Lg soluble (calles 3-7), observado por la disminución de la intensidad de color
Con
teni
do p
rote
ico
(g/L
)
5
7
Presión (MPa)
0.1 250 350 450 550 650
8
6
4
Con
teni
do p
rote
ico
(g/L
)
5
7
Presión (MPa)
0.1 250 350 450 550 650
8
6
4
IR PR PLIR PR PL

RESULTADOS Y DISCUSIÓN: CAPÍTULO II
- 83 -
de la banda correspondiente a esta proteína. Lo cual indicó que la proteína se desnaturalizó y
precipitó junto a las caseínas al descender el pH. En el caso de la α-La, la intensidad de la
banda no cambió de forma tan notable, observándose solamente una ligera disminución del
color. Esta proteína fue mucho más resistente a la presión que la β-Lg.
Figura 2.6. PAGE-SDS de las fracciones solubles a pH 4.6 de leche control (0.1 MPa) y leche
presurizada a 250, 350, 450, 550 y 650 MPa, a 15ºC durante 5 min, en un equipo industrial (IR) y un
equipo piloto con despresurización rápida (PR) y lenta (PL). Calles: 1:Marcador de pesos
moleculares; 2-7: tratamiento de 0.1, 250, 350, 450, 550 y 650 MPa.
El perfil electroforético de estas fracciones, obtenido por EC, corroboró el hecho de
que la β-Lg fue más sensible a la presión que la α-La, disminuyendo su contenido a medida
que aumentó la presión (Figura 27). Además, en los electroforegramas también fue
claramente visible que el tratamiento IR produjo una menor precipitación de la β-Lg que los
tratamientos PR y PL y la comparación de los dos tratamientos realizados en el equipo piloto
mostró que el contenido de β-Lg fue aún menor cuando se despresurizó de forma lenta (5
min).
1 2 3 4 5 6 7
α−La
β-Lg
IR PR PL
1 2 3 4 5 6 71 2 3 4 5 6 71 2 3 4 5 6 7
α−La
β-Lg
IR PR PL
1 2 3 4 5 6 71 2 3 4 5 6 7

84
Figura 27. Electroforegramas de las proteínas de suero (fracción soluble a pH 4.6) de leche desnatada y presurizada a 0, 250, 350, 450, 550 y 650 MPa (a-f)
en equipo industrial con despresurización rápida (IR) y en equipo piloto con despresurización rápida (PR) y lenta (PL).
A 21
4 nm
0.02
0.04
0.06
0.08
0.02
0
0
0.02
0
0
0.02
0
0.02
0
0.02
10 15 20 25
a
b
c
d
e
f
Tiempo de migración (min)
0
0.02
0.04
0.06
0.08
0.02
0.02
0
0.02
0
0.02
0
0.02
10 15 20 25
0
0
a
b
c
d
e
f
0
0.02
0.04
0.06
0.08
0
0.02
0
0.02
0
0.02
0
0.02
0
0.02
10 15 20 25
a
b
c
d
e
f
α-La
β-Lg
IR PR PL
α-La
β-Lg
α-La
β-Lg
A 21
4 nm
0.02
0.04
0.06
0.08
0.02
0
0
0.02
0
0
0.02
0
0.02
0
0.02
10 15 20 25
a
b
c
d
e
f
Tiempo de migración (min)
0
0.02
0.04
0.06
0.08
0.02
0.02
0
0.02
0
0.02
0
0.02
10 15 20 25
0
0
a
b
c
d
e
f
0
0.02
0.04
0.06
0.08
0
0.02
0
0.02
0
0.02
0
0.02
0
0.02
10 15 20 25
a
b
c
d
e
f
α-La
β-Lg
IR PR PL
α-La
β-Lg
α-La
β-Lg

RESULTADOS Y DISCUSIÓN: CAPÍTULO II
- 85 -
En la Figura 28 se muestra la cuantificación de la pérdida de solubilidad a pH 4.6 de
las proteínas de suero individuales, α-La y β-Lg medida por EC. Aproximadamente el 10% de
la α-La se desnaturalizó después de los tratamientos IR y PR a presiones ≥ 450 MPa,
aumentando al 25% a 650 MPa. La desnaturalización de la α-La fue significativamente mayor
después de los tratamientos PL a similares niveles de presión, alcanzando el 40% a 650 MPa.
La desnaturalización de la β-Lg aumentó progresivamente con el nivel de presión (P< 0.01),
alcanzando, aproximadamente, el 90% a 650 MPa. Se observaron diferencias significativas
entre los tres tipos de tratamientos a presiones ≥ 350 MPa (P< 0.01). La desnaturalización de
la β-Lg, estimada por la pérdida de solubilidad a pH 4.6, se produjo en el orden: PL> PR> IR.
Figura 28. Contenido de α-La y β-Lg solubles a pH 4.6, medido por CE, de leche presurizada a
250, 350, 450, 550 y 650 MPa, a 15ºC durante 5 min en un equipo industrial (IR) y un equipo piloto
con despresurización rápida (PR) y lenta (PL). Las barras representan la desviación estándar. Las
letras mayúsculas indican diferencias significativas (P< 0.05) debido al tipo de tratamiento (unidad
de alta presión o velocidad de despresurización, IR, PR y PL), mientras que las letras minúsculas
indican diferencias significativas (P< 0.05) debido al nivel de presión.
250 MPa 350 MPa 450 MPa 550 MPa 650 MPa250 MPa250 MPa 350 MPa350 MPa 450 MPa450 MPa 550 MPa550 MPa 650 MPa650 MPa
b)a)
0
20
40
60
80
100
IR PR PL
a a a
bb
b
cc
c
d d
d
e ee
β-L
gso
lubl
e (%
)
Equipo HP
0
20
40
60
80
100
IR PR PL
ab
ca
a
b
b
bbbccdccd
dd
α-L
a so
lubl
e (%
)
Equipo HP
A A B A B C
b)a)
0
20
40
60
80
100
IR PR PL
a a a
bb
b
cc
c
d d
d
e ee
β-L
gso
lubl
e (%
)
Equipo HP
0
20
40
60
80
100
IR PR PL
ab
ca
a
b
b
bbbccdccd
dd
α-L
a so
lubl
e (%
)
Equipo HP
A A B A B C
0
20
40
60
80
100
IR PR PL
a a a
bb
b
cc
c
d d
d
e ee
β-L
gso
lubl
e (%
)
Equipo HP
0
20
40
60
80
100
IR PR PL
a a a
bb
b
cc
c
d d
d
e ee
β-L
gso
lubl
e (%
)
Equipo HP
0
20
40
60
80
100
IR PR PL
ab
ca
a
b
b
bbbccdccd
dd
α-L
a so
lubl
e (%
)
Equipo HP
0
20
40
60
80
100
IR PR PL
ab
ca
a
b
b
bbbccdccd
dd
α-L
a so
lubl
e (%
)
Equipo HP
A A B A B C
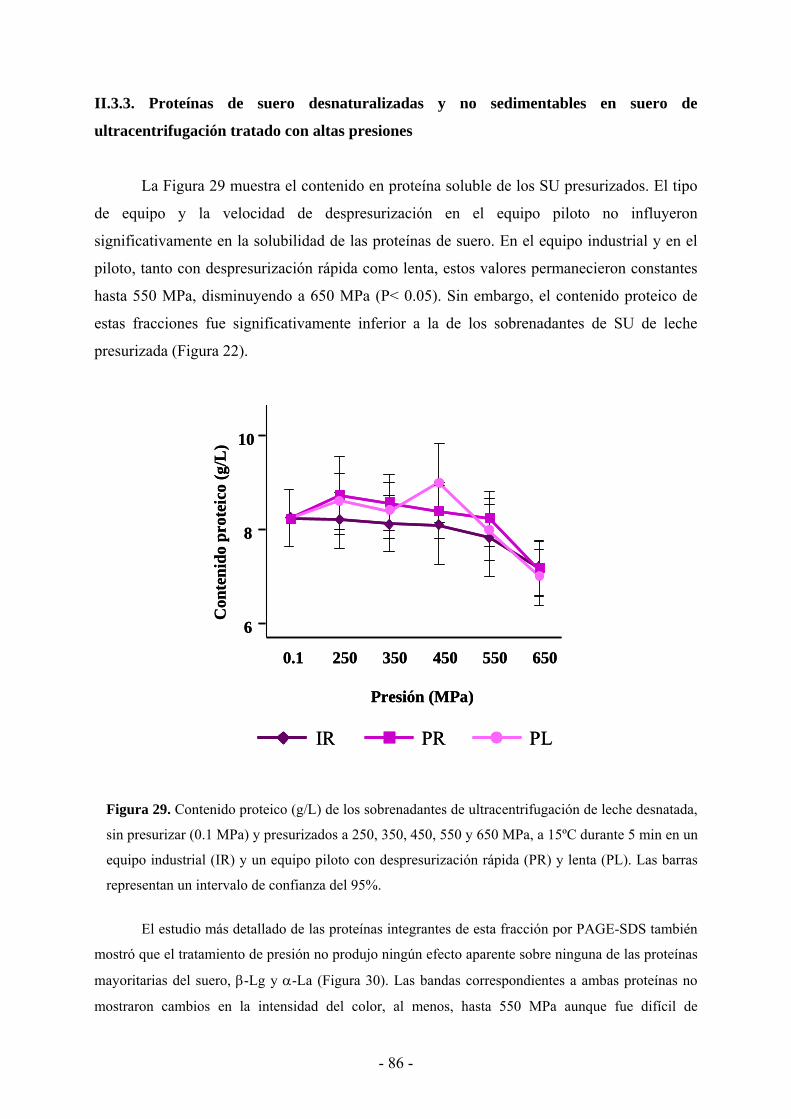
- 86 -
II.3.3. Proteínas de suero desnaturalizadas y no sedimentables en suero de
ultracentrifugación tratado con altas presiones
La Figura 29 muestra el contenido en proteína soluble de los SU presurizados. El tipo
de equipo y la velocidad de despresurización en el equipo piloto no influyeron
significativamente en la solubilidad de las proteínas de suero. En el equipo industrial y en el
piloto, tanto con despresurización rápida como lenta, estos valores permanecieron constantes
hasta 550 MPa, disminuyendo a 650 MPa (P< 0.05). Sin embargo, el contenido proteico de
estas fracciones fue significativamente inferior a la de los sobrenadantes de SU de leche
presurizada (Figura 22).
Figura 29. Contenido proteico (g/L) de los sobrenadantes de ultracentrifugación de leche desnatada,
sin presurizar (0.1 MPa) y presurizados a 250, 350, 450, 550 y 650 MPa, a 15ºC durante 5 min en un
equipo industrial (IR) y un equipo piloto con despresurización rápida (PR) y lenta (PL). Las barras
representan un intervalo de confianza del 95%.
El estudio más detallado de las proteínas integrantes de esta fracción por PAGE-SDS también
mostró que el tratamiento de presión no produjo ningún efecto aparente sobre ninguna de las proteínas
mayoritarias del suero, β-Lg y α-La (Figura 30). Las bandas correspondientes a ambas proteínas no
mostraron cambios en la intensidad del color, al menos, hasta 550 MPa aunque fue difícil de
6
8
10
Presión (MPa)
0.1 250 350 450 550 650
Con
teni
do p
rote
ico
(g/L
)
6
8
10
Presión (MPa)
0.1 250 350 450 550 650
Con
teni
do p
rote
ico
(g/L
)
IR PR PLIR PR PL
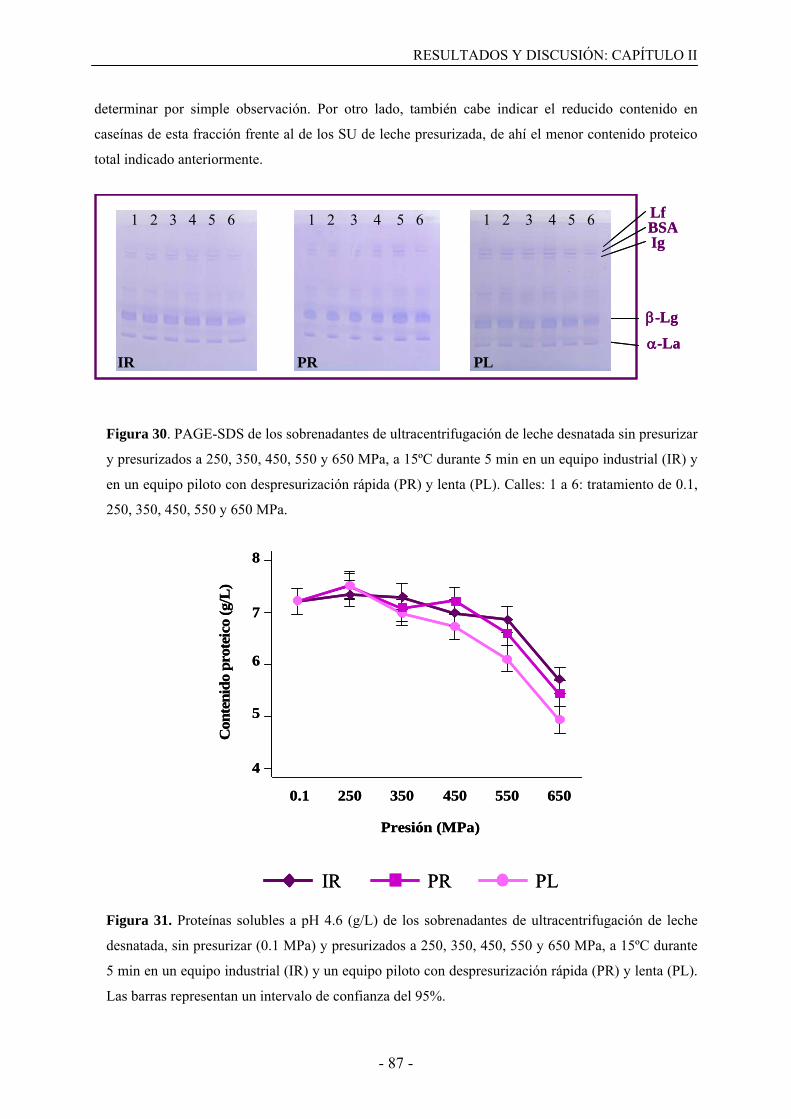
RESULTADOS Y DISCUSIÓN: CAPÍTULO II
- 87 -
determinar por simple observación. Por otro lado, también cabe indicar el reducido contenido en
caseínas de esta fracción frente al de los SU de leche presurizada, de ahí el menor contenido proteico
total indicado anteriormente.
Figura 30. PAGE-SDS de los sobrenadantes de ultracentrifugación de leche desnatada sin presurizar
y presurizados a 250, 350, 450, 550 y 650 MPa, a 15ºC durante 5 min en un equipo industrial (IR) y
en un equipo piloto con despresurización rápida (PR) y lenta (PL). Calles: 1 a 6: tratamiento de 0.1,
250, 350, 450, 550 y 650 MPa.
Figura 31. Proteínas solubles a pH 4.6 (g/L) de los sobrenadantes de ultracentrifugación de leche
desnatada, sin presurizar (0.1 MPa) y presurizados a 250, 350, 450, 550 y 650 MPa, a 15ºC durante
5 min en un equipo industrial (IR) y un equipo piloto con despresurización rápida (PR) y lenta (PL).
Las barras representan un intervalo de confianza del 95%.
4
6
8
Con
teni
do p
rote
ico
(g/L
)
Presión (MPa)
0.1 250 350 450 550 650
5
7
4
6
8
Con
teni
do p
rote
ico
(g/L
)
Presión (MPa)
0.1 250 350 450 550 650
5
7
IR PR PLIR PR PL
1 2 3 4 5 6
α-La
β-Lg
LfBSAIg
IR PR PL
1 2 3 4 5 61 2 3 4 5 61 2 3 4 5 6
α-La
β-Lg
LfBSAIg
IR PR PL
1 2 3 4 5 61 2 3 4 5 6

- 88 -
Como se muestra en la Figura 31, el contenido en proteínas solubles a pH 4.6 empezó
a disminuir a presiones ≥350 MPa. Se observaron diferencias significativas entre los dos
equipos (P< 0.05) y la velocidad de despresurización a presiones ≥ 550 MPa (P<0.01),
siguiendo el orden PL> PR> IR.
II.4. Discusión
II.4.1. Efecto del tipo de equipo de alta presión y de la velocidad de despresurización en
la liberación de caseínas no sedimentables a la fase soluble de la leche
Los resultados de este estudio mostraron que el tipo de equipo de alta presión y la
velocidad de despresurización influyeron en la composición de las fracciones coloidales y
solubles de la leche. El contenido en caseínas no sedimentables y la desnaturalización de las
proteínas de suero aumentaron significativamente con el nivel de presión bajo las tres
condiciones experimentales consideradas, aunque se encontraron diferencias entre el equipo
industrial y piloto y entre los dos tiempos de despresurización ensayados.
Durante los tratamientos de leche con alta presión, la micela de caseína se desintegra
debido a la ruptura de las interacciones electrostáticas intermoleculares y a la solubilización
del fosfato cálcico coloidal (CCP) (Huppertz y De Kruif, 2006, 2007a). Medidas in-situ de la
transmisión de la luz muestran que la ruptura de las micelas de caseína comienza a presiones
de 100 MPa, aunque, a 250 y 300 MPa, a la desintegración inicial de las micelas que ocurre
durante los primeros 5 min de tratamiento le sigue una progresiva agregación, debido a que, a
estas presiones intermedias, se potencian las interacciones hidrofóbicas sobre la solvatación
de los residuos hidrofóbicos (Huppertz y De Kruif, 2006; Huppertz y col., 2006b). Las
micelas de caseína resultantes están más hidratadas y, se produce un aumento de las proteínas
no sedimentables, posiblemente debido a la liberación fragmentos caseínicos o caseínas
individuales, a presiones ≥100 MPa, tal y como se observó en este estudio (López-Fandiño y
col., 1998; Huppertz y col., 2004b). Esto implica que el nivel de caseínas no sedimentables
está inversamente correlacionado con el tamaño de la micela de caseína, excepto para las
muestras agregadas a 250 y 300 MPa (Anema y col., 2005a).

RESULTADOS Y DISCUSIÓN: CAPÍTULO II
- 89 -
En términos generales, observamos niveles de caseína soluble (Figuras 22 y 24)
comparables a los publicados previamente para leche presurizada, con algunas diferencias que
se pueden atribuir al uso de diferentes velocidades de ultracentrifugación (Anema y col.,
2005a; Anema, 2008b). Normalmente, el máximo nivel de caseína soluble, que alcanza el 40
o 50% del total del contenido caseínico, se obtiene tras tratamientos a 300 MPa a 20ºC
durante 30 min (López-Fandiño y col., 1998; Huppertz y col., 2004b; Anema y col., 2005a;
Anema, 2008b). En nuestro estudio, y tras los tratamientos con despresurización rápida -IR y
PR-, los máximos niveles de proteínas no sedimentables se observaron a presiones superiores
a las ya publicadas. Esta diferencia se podría atribuir al corto tiempo de tratamiento utilizado
ya que, aunque la liberación de caseínas a la fase sérica es muy rápida durante los primeros 5
min de tratamiento, ésta continua aumentando gradualmente hasta los 60 min (Anema y col.,
2005a; Anema, 2008b). Por otra parte, cuanto mayor es la presión, más rápida es la ruptura de
la micela de caseína (Huppertz y col., 2006b).
En comparación con el tratamiento a escala piloto, el tratamiento a escala industrial
condujo a un contenido en proteínas no sedimentables significativamente menor y la máxima
liberación de caseínas a la fase soluble tuvo lugar a presiones más altas. Estas diferencias
podrían atribuirse, al menos parcialmente, a las diferentes temperaturas de presurización
alcanzadas en ambos sistemas. Durante la presurización, y asumiendo que la compresión es
uniforme y que no existen pérdidas de calor, se produce un aumento de la temperatura del
medio de presurización, que depende de diversos factores, como: el nivel de presión, el
tiempo de mantenimiento, la composición del líquido transmisor de la presión, los materiales
aislantes utilizados, etc. (Balasubramanian y Balasubramaniam, 2003; Buzrul y col., 2008;
Knoerzer y col., 2010a, b). A medida que sube la presión, la temperatura aumenta por
calentamiento adiabático, equilibrándose durante la fase de mantenimiento. En nuestras
condiciones experimentales, se eligió un tiempo de tratamiento de 5 min para simular
condiciones comerciales donde es muy importante el procesado de la mayor cantidad posible
de producto. Posiblemente, ese tiempo de mantenimiento sea demasiado corto para la
disipación de la temperatura alcanzada durante la etapa de subida de la presión, especialmente
en el equipo a escala industrial con un volumen 150 veces mayor que el equipo a escala piloto
(que a su vez estaba recubierto de una camisa de agua termostatizada a 15ºC). Si bien esto
habría dado lugar a una mayor temperatura efectiva durante los tratamientos a escala
industrial (IR), la temperatura real fue superior en los tratamientos PR y PL. Así, por ejemplo,

- 90 -
se alcanzaron temperaturas aproximadamente unos 14ºC superiores a 350 y 650 MPa,
durante los tratamientos con el equipo piloto. Esto se debió a la utilización de aceite de
silicona como fluido transmisor de la presión, que tiene mayor comprensibilidad y menor
calor específico y conductividad térmica que el agua (Balasubramanian y Balasubramaniam,
2003; Knoerzer y col., 2010a). La mayor temperatura alcanzada en los tratamientos a escala
piloto reduciría la solubilidad del CCP y promovería las uniones hidrofóbicas, estabilizando
las micelas de caseínas frente a la ruptura (Huppertz y col., 2006b). Sin embargo, los
resultados existentes sobre el efecto de la temperatura en la desintegración de la micela de
caseína indican que un aumento de la temperatura del tratamiento en el intervalo de 10 a 40ºC
causa una liberación más rápida de las caseínas al sobrenadante (Anema y col., 2005a). En
cualquier caso, debe tenerse en cuenta que Anema y col. (2005a) observaron diferencias
inferiores al 10%, tras tratamientos a 200-600 MPa durante 5 min, entre temperaturas de 30 y
40ºC. Dado que, en nuestro caso las diferencias son mayores no podría descartarse un efecto
adicional debido al tipo de equipo o al escalado (Tabla 6).
Respecto al efecto de la velocidad de despresurización, debe señalarse que los niveles
de proteínas no sedimentables obtenidos después de los dos tratamientos en el equipo piloto,
PR y PL fueron similares, aunque la liberación máxima de caseínas a la fase soluble se logró a
presiones más bajas en el tratamiento PL (Figure 22). Esto sugiere que una despresurización
lenta podría favorecer la desintegración de la micela. Previamente ya se había observado que,
a presiones superiores a las que causan la liberación máxima de caseínas, el nivel de caseínas
solubles tiende a disminuir, tal y como se ha observado en este estudio (Figura 22, 23 y 24,
Tabla 6) (López-Fandiño y col., 1998; Huppertz y col., 2004b; Anema y col., 2005a). Esto
podría deberse a una reagrupación parcial de las partículas micelares solubilizadas, puesto que
para que se dé una reagrupación total que origine grandes agregados de caseína (como ocurre
a presiones intermedias de 250-300 MPa) es necesario que la ruptura de la micela de caseína
inducida por la presión no sea ni demasiado limitada ni demasiado profunda (Huppertz y De
Kruif, 2006). Por otra parte, la disminución de la cantidad de sólidos solubles a 550 y 650
MPa (Figura 22) podría deberse , al menos en parte, a la pérdida de solubilidad de la β-Lg e
incluso de la α-La (Tabla 6 y Figura 24), explicando así, la reducción del contenido total de
proteína no sedimentable en los SU.

RESULTADOS Y DISCUSIÓN: CAPÍTULO II
- 91 -
II.4.2 Influencia de la desnaturalización de las proteínas de suero
Los resultados obtenidos en los SU presurizados, que contenían proteínas de suero
prácticamente en ausencia de micelas de caseína, indicaron que la presurización redujo tanto
los niveles de proteínas solubles como los de proteínas nativas a presiones ≥ 550 MPa. Sin
embargo, el examen de las fracciones solubles al pH normal de la leche y a pH 4.6 (Figuras
29, 30 y 31) indicó la existencia de proteínas de suero desnaturalizadas solubles.
Comparativamente, la pérdida de solubilidad de la α-La y de la β-Lg, tanto al pH normal de la
leche como a pH 4.6, fue mucho más importante en la leche desnatada presurizada (Figuras
25, 26, y 27). Estos resultados corroboraron el hecho de que las proteínas de suero son más
sensibles a la presión cuando están presentes en leche que en suero, puesto que, en el primer
caso, estas proteínas podrían asociarse con las caseínas y precipitar con ellas en su punto
isoeléctrico (Huppertz y col., 2004a, b). No obstante, en la leche presurizada, una importante
proporción de las proteínas de suero desnaturalizadas también permaneció soluble al pH
normal de la leche (Tabla 6), como ya habían descrito López-Fandiño y col. (1998) y Anema
(2008b). Esto sugiere que las proteínas de suero desnaturalizadas solubles interaccionan entre
sí o con las partículas de caseínas solubles liberadas durante la disgregación de la micela de
caseína inducida por la presión, y no influyen en los cambios micelares producidos durante
los tratamientos. A este respecto, aunque se ha sugerido que la asociación de las proteínas de
suero desnaturalizadas con las micelas de caseína podría compensar la disminución del
tamaño de la micela inducido por la presión (Huppertz y col., 2004a), experimentos de
presurización de micelas de caseína en ausencia y presencia de proteínas de suero indicaron
que la desnaturalización de las proteínas de suero no interfiere en la disociación o
reasociación micelar (Gaucheron y col., 1997; Anema y col., 2005a; Huppertz y De Kruif,
2007b).
El grado de desnaturalización de las proteínas de suero depende del nivel de presión,
el tiempo del tratamiento, el pH y la temperatura (López-Fandiño, 2006a). Sin embargo, este
estudio también muestra que otros factores, como el tipo de equipo o la velocidad de
despresurización, pueden influir en la desnaturalización de las seroproteínas, aunque sus
efectos sólo fueron significativos a presiones ≥ 350 MPa aplicadas a la leche (Figura 25), o ≥
550 MPa aplicadas a los SU (Figura 29). En el equipo industrial, a pesar de que se alcanzó
una temperatura mayor durante el tratamiento, se observaron las menores desnaturalizaciones.

- 92 -
En el equipo a escala piloto, el tiempo más largo de despresurización (PL) aumentó la
desnaturalización de las proteínas de suero frente al tiempo corto de despresurización (PR),
pero la mayor desnaturalización de las proteínas de suero se produjo durante la fase de
mantenimiento de la presión.
II.5. Conclusiones
Los resultados obtenidos sugieren que los tratamientos con alta presión en los equipos
estudiados, a escala industrial y piloto, producen diferentes distribuciones proteicas, pues
conducen a diferentes grados de desintegración micelar con similares niveles de
desnaturalización de las proteínas de suero. Si bien la presencia de caseína promovió la
desnaturalización de las proteínas del suero, el grado de desnaturalización de las proteínas de
suero no mostró ningún efecto en la ruptura micelar. Así, el equipo industrial condujo a una
menor desintegración micelar que el equipo piloto, pero la mayoría de las proteínas de suero
desnaturalizadas, unidas o no a las partículas de caseína no sedimentables, permanecieron
solubles en ambos sistemas. La influencia de la velocidad de despresurización, descuidada en
la mayoría de los estudios de alta presión, se tratará en más profundidad en el capítulo III.

CAPÍTULO III
Efecto de la velocidad de despresurización en la composición proteica de la
fracción soluble de la leche


RESULTADOS Y DISCUSIÓN: CAPÍTULO III
- 95 -
III.1. Introducción
Las micelas de caseína de la leche tratada con alta presión presentan propiedades
diferentes a las de la leche cruda o calentada, relacionadas con su tamaño, número,
composición e hidratación (Huppertz y col., 2006a; López-Fandiño, 2006a). El tratamiento
con alta presión produce la desintegración de las micelas de caseína que, posteriormente,
pueden volver a agregarse bajo ciertas condiciones. El grado de estos cambios depende de
diferentes factores, tales como el nivel de presión, la duración del tratamiento, la
temperatura, el contenido en calcio, el pH, etc (Arias y col., 2000; García-Risco y col., 2000;
Huppertz y col., 2004a). Además, la alta presión también desnaturaliza a las proteínas de
suero, que pueden asociarse con las micelas de caseína o con fragmentos micelares,
existiendo varios estudios que han investigado la interacción de ambos fenómenos (Huppertz
y col., 2004d; Anema y col., 2005a; Huppertz y De Kruif, 2007b).
Si bien se ha indicado que la velocidad de subida de la presión no tiene ningún efecto
en las modificaciones producidas por la presión sobre las caseínas (Merel-Rausch y col.,
2006), el efecto de la velocidad de liberación de la presión se ha ignorado en la mayoría de
los estudios. Así, los resultados mostrados en el capítulo anterior indican que la velocidad de
despresurización podría influir en la desintegración micelar y solubilización de las caseínas,
así como en la desnaturalización de las proteínas de suero. Según Fertsch y col. (2003), la
firmeza de los geles formados a partir de disoluciones de caseína micelar presurizadas a 600
MPa depende de la velocidad de descompresión, ya que las velocidades de despresurización
más rápidas proporcionan geles más firmes y microestructuras más finas. Estos autores
sugieren que, una vez que se produce la disociación de la micela de caseína durante la fase de
mantenimiento de la presión, una despresurización lenta favorece el desarrollo de
hiperestructuras caseínicas, debido a que los puentes de calcio y las interacciones
hidrofóbicas se vuelven a formar, interfiriendo en una gelificación homogénea (Fertsch y
col., 2003). Merel-Rausch y col. (2007) también encontraron geles de menor firmeza a
velocidades lentas de liberación de la presión. En este caso, la viscosidad de disoluciones de
caseína micelar tratadas a 400 y 600 MPa a 20 y 30ºC con velocidades de despresurización
rápidas, aumentó en un 5%, lo que se correlacionó con la formación de grandes estructuras
flexibles, construidas a partir de micelas de caseína de menor tamaño. Así, las
despresurizaciones rápidas (600 MPa/min) tenderían a diámetros hidrodinámicos grandes,

- 96 -
mientras que las lentas (20 MPa/min) disminuirían el diámetro, con respecto al de la leche
control (Merel-Rausch y col., 2006). A este respecto, cabe indicar, además, que todavía no
está clara la influencia de la desnaturalización de las proteínas de suero en la disociación y
reagregación de las caseínas inducida por la presión (Huppertz y col., 2006a; López-Fandiño,
2006a).
A la vista de la importancia que tienen las propiedades fisicoquímicas de las proteínas
de la leche en su aptitud tecnológica; el objetivo de este estudio fue el de profundizar en la
influencia de la velocidad de despresurización sobre la distribución proteica entre las
fracciones solubles y coloidales de la leche tratada con alta presión. Para llevar a cabo este
propósito, se presurizó leche desnatada, con y sin agentes bloqueantes de grupos sulfhidrilo, a
250 y 350 MPa de 5 a 15 min a 25ºC, aplicándole diferentes tiempos de despresurización
entre 0.07 y 10 min. Se evaluó el grado de liberación de caseínas a la fase soluble y la
influencia de la desnaturalización de las proteínas de suero en este proceso.
III.2. Materiales y Métodos
La Figura 32 resume el procedimiento experimental seguido en este trabajo.
III.2.1. Preparación de las muestras
La leche cruda de vaca se obtuvo de una granja local y se desnató calentándola a 37ºC
durante 30 min y centrifugándola a 3000 x g, 20ºC durante 30 min. Una parte de la leche
desnatada se trató con N-etilmaleimida (NEM; Sigma-Aldrich, St. Louis, MO, USA),
siguiendo el método descrito por Needs y col. (2000b), con el fin de bloquear los grupos
sufhidrilo de la proteína, implicados en la desnaturalización de las proteínas de suero. El
NEM es insoluble en agua por lo que se disolvió en dimetilformamida (DMF; Lab-Scan,
Gliwice, Poland) a una concentración de 20 mg/mL y se añadió, a la leche desnatada, 16.7
mL/L de esta disolución. Se usaron, como controles, muestras de leche con el mismo volumen
de DMF sin NEM. Tanto las muestras con DMF como con NEM se dejaron a temperatura
ambiente toda la noche antes de los tratamientos con alta presión.

RESULTADOS Y DISCUSIÓN: CAPÍTULO III
- 97 -
III.2.2. Tratamientos de alta presión
Se realizaron presurizaciones a 250 y 350 MPa, 25ºC durante 5 min de leche
desnatada, leche desnatada con NEM y leche desnatada con DMF. Para ello, las muestras se
dispensaron en botes de polietileno de 70 mL, llenándolos hasta arriba para no dejar espacio
libre en su interior y se introdujeron en un equipo de alta presión ACB 900 HP (Eurotherm
Automation, Lyon, France) el cual utiliza agua como fluido transmisor de la presión.
Para evaluar el efecto de la duración del tratamiento, la presión se elevó a una
velocidad de 2.5 MPa/seg, manteniéndose durante 5, 6, 10 o 15 min, y se liberó en 0.07 min
(4 seg), esto es, a una velocidad de despresurización de 62.5 y 87.5 MPa/seg, para los
tratamientos de 250 y 350 MPa, respectivamente.
Por otro lado, para evaluar el efecto de la despresurización, la presión se elevó a una
velocidad de 2.5 MPa/seg, manteniéndose durante 5 min, y se liberó en 0.07, 1, 5 o 10 min,
esto es, para los tratamientos de 250 MPa, las velocidades de despresurización fueron 62.5,
4.2, 0.8 o 0.4 MPa/seg y, para 350 MPa, 87.5, 5.8, 1.2 o 0.6 MPa/seg, respectivamente
(Figura 32). En adelante, y con el fin de simplificar la presentación de los resultados, se
utilizarán los tiempos de despresurización como indicadores de las diferentes velocidades de
despresurización. Todos los tratamientos con alta presión se realizaron al menos por
duplicado.

Esquema 1. Esquema de la preparación, tratamientos con alta presión, fraccionamiento y análisis de las muestras
250 y 350 MPa, 25ºC, 5 min
Leche cruda de vacaGec AlsthomACB 900 HP
Rampa de subida2.5 MPa/seg
Lechescon y sinpresurizar
SU
4.6
Fracciones
Precipitacióna pH 4.6
Ultracentrifugación100000 x g, 20ºC, 1h
Análisis
Despresurización 0.07 min
Despresurización 1 min
Despresurización 5 min
Despresurización 10 min
M. Presión6 min
M. Presión10 min
M. Presión15 min
Mantenimientode la presión 5 min, 25 ºC
Despresurización0.07 min
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatada
DMF NEMLeche
desnatada
Reposo 16 h Tª ambiente
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEM
Desnatado Tratamiento con alta presión Fraccionamiento Análisis
3000 x g, 20ºC, 30 min
Aná
lisis
Adición deDMF o NEM
ELECTROFORESISCAPILAR
NITRÓGENO(KJELDAHL)
PAGE-SDS
PROTEÍNAS
250 y 350 MPa, 25ºC, 5 min
Leche cruda de vacaGec AlsthomACB 900 HP
Rampa de subida2.5 MPa/seg
Lechescon y sinpresurizar
Lechescon y sinpresurizar
SU
4.6
Fracciones
Precipitacióna pH 4.6
Ultracentrifugación100000 x g, 20ºC, 1h
Análisis
Despresurización 0.07 min
Despresurización 1 min
Despresurización 5 min
Despresurización 10 min
M. Presión6 min
M. Presión10 min
M. Presión15 min
Mantenimientode la presión 5 min, 25 ºC
Despresurización0.07 min
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatadaLeche
desnatada
DMFLeche
desnatadaLeche
desnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatadaLeche
desnatada
DMFLeche
desnatadaLeche
desnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatadaLeche
desnatada
DMFLeche
desnatadaLeche
desnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatadaLeche
desnatada
DMFLeche
desnatadaLeche
desnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatadaLeche
desnatada
DMFLeche
desnatadaLeche
desnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatadaLeche
desnatada
DMFLeche
desnatadaLeche
desnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatadaLeche
desnatada
DMFLeche
desnatadaLeche
desnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatada
Lechedesnatada
Lechedesnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatadaLeche
desnatada
DMFLeche
desnatadaLeche
desnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatadaLeche
desnatadaLeche
desnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatadaLeche
desnatada
DMFLeche
desnatadaLeche
desnatada
DMF NEMLeche
desnatadaLeche
desnatadaLeche
desnatada
Reposo 16 h Tª ambiente
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEMLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEMLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEMLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEMLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEMLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEMLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizada
DMF NEM
Lechepresurizada
Lechepresurizada
Lechepresurizada
DMF NEMLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizadaLeche
presurizada
DMF NEM
Desnatado Tratamiento con alta presión Fraccionamiento Análisis
3000 x g, 20ºC, 30 min
Aná
lisis
Adición deDMF o NEM
ELECTROFORESISCAPILAR
NITRÓGENO(KJELDAHL)
PAGE-SDS
PROTEÍNAS

RESULTADOS Y DISCUSIÓN: CAPÍTULO III
- 99 -
III.3. Resultados
III.3.1. Solubilización de caseínas y desnaturalización de proteínas del suero en leche
desnatada tratada por altas presiones
Se determinó el contenido de proteínas no sedimentables y de proteínas solubles a pH
4.6 de la leche tratada con altas presiones a 250 y 350 MPa. La Tabla 7 muestra el efecto del
tiempo de mantenimiento de la presión con 0.07 min de despresurización y el efecto del
tiempo de despresurización con 5 min de mantenimiento de la presión.
A 250 MPa, los niveles de proteínas no sedimentables encontrados en los SU de leche
presurizada con tiempos de mantenimiento de la presión < 15 min fueron similares a los de la
leche control, mientras que, a 350 MPa, aumentaron significativamente con tiempos de
mantenimiento ≥ 5 min (Tabla 7). Los contenidos en proteínas de suero solubles a pH 4.6
disminuyeron significativamente después de 5 min a 250 y 350 MPa, y el mantenimiento de la
presión a 350 MPa hasta 15 min también los redujo significativamente (P< 0.05).
En comparación con la despresurización más rápida (0.07 min), el aumento del tiempo
de despresurización (de 1 a 10 min) aumentó significativamente los niveles de proteína no
sedimentable a ambos niveles de presión y, especialmente, a 350 MPa (Tabla 7). Los
tratamientos con una duración total de 15 min a 250 MPa y de 10 y 15 min a 350 MPa
produjeron contenidos de proteínas solubles significativamente mayores cuando el tiempo de
mantenimiento de la presión fue de 5 min, con 5 y 10 min de despresurización, que cuando
los tiempos de mantenimiento de la presión fueron de 10 y 15 min, respectivamente, con 0.07
min de despresurización (valores marcados con * en la Tabla 7). Sin embargo, los tiempos de
despresurización y de mantenimiento de la presión ejercieron una influencia similar en los
contenidos de proteínas en la fracción soluble a pH 4.6 después tratamientos a 250 y 350
MPa. Como era de esperar, la presurización durante 5 min, a ambas presiones, produjo una
disminución significativa del contenido proteico de esta fracción (P< 0.05) que fue
disminuyendo ligeramente con el aumento de la duración del tratamiento a ambas presiones
aunque estos cambios no fueron significativamente diferentes. No obstante, el contenido en
proteínas solubles fue menor en todos los tratamientos realizados a 350 MPa respecto a los
mismos a 250 MPa (Tabla 7).

- 100 -
Tabla 7. Contenidos en proteínas (g/L ± desviación estándar) de los sobrenadantes de
unltracentrifugación (SU) y de las fracciones soluble a pH 4.6 (4.6) de la leche desnatada
control (0 min de tiempo de mantenimiento de la presión y de despresurización) y leche
presurizada a 250 y 350 MPa y 25ºC a diferentes tiempos de mantenimiento y
despresurización.
a-d Diferentes superíndices en letras minúsculas, en columnas, indican diferencias significativas (P< 0.05) producidas por el tiempo de mantenimiento de la presión. A-E Diferentes superíndices en letras mayúsculas, en columnas, indican diferencias significativas (P< 0.05) producidas por el tiempo de despresurización. * Indica diferencias significativas (P< 0.05) entre valores correspondientes al mismo tiempo total de tratamiento con diferentes combinaciones de tiempo de mantenimiento y de despresurización a cada nivel de presión.
El análisis de los sobrenadantes de ultracentrifugación y de las fracciones solubles a
pH 4.6 por PAGE-SDS se muestran en la Figura 33, el cual reveló que el contenido en
caseínas no sedimentables en los sobrenadantes de ultracentrifugación aumentó con el
aumento del tiempo de despresurización (Figura 33, A y B, calles 5-8). Este hecho justifica el
incremento en el contenido proteico total observado anteriormente (Tabla 7). Además, la
intensidad de las bandas correspondientes a las caseínas en los sobrenadantes de
250 MPa 350 MPa Tiempo de mantenimiento
(min)
Tiempo de despresurización
(min) SU 4.6 SU 4.6
0 0 12.05bD±0.01 8.07aA±0.02 12.05dE±0.01 8.07aA±0.02
5 0.07 12.00bD±0.08 7.79bB±0.06 14.88cD±0.08 7.24bB±0.04
6 0.07 12.83ab±0.10 7.85ab±0.38 15.24b±0.10 7.22b±0.02
10 0.07 12.93ab±0.88 8.06a±0.01 *16.16a±0.01 6.93c±0.11
15 0.07 *13.30a±0.29 8.32a±0.15 *16.24a±0.05 6.86c±0.10
5 1 13.06C±0.20 7.71B±0.08 15.65C±0.10 7.36B±0.10
5 5 14.19 B±0.01 7.65B±0.10 *17.32B±0.31 7.26B±0.11
5 10 *14.62A±0.11 7.41C±0.06 *18.44A±0.30 7.17B±0.06
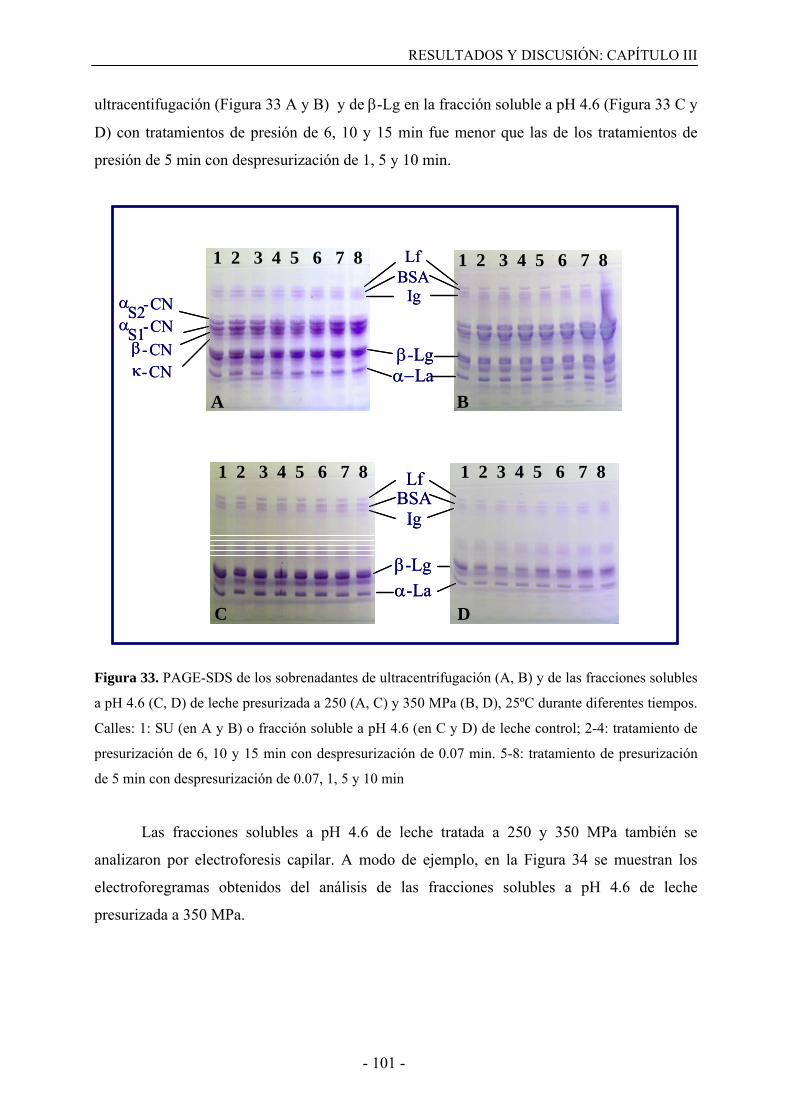
RESULTADOS Y DISCUSIÓN: CAPÍTULO III
- 101 -
ultracentifugación (Figura 33 A y B) y de β-Lg en la fracción soluble a pH 4.6 (Figura 33 C y
D) con tratamientos de presión de 6, 10 y 15 min fue menor que las de los tratamientos de
presión de 5 min con despresurización de 1, 5 y 10 min.
Figura 33. PAGE-SDS de los sobrenadantes de ultracentrifugación (A, B) y de las fracciones solubles
a pH 4.6 (C, D) de leche presurizada a 250 (A, C) y 350 MPa (B, D), 25ºC durante diferentes tiempos.
Calles: 1: SU (en A y B) o fracción soluble a pH 4.6 (en C y D) de leche control; 2-4: tratamiento de
presurización de 6, 10 y 15 min con despresurización de 0.07 min. 5-8: tratamiento de presurización
de 5 min con despresurización de 0.07, 1, 5 y 10 min
Las fracciones solubles a pH 4.6 de leche tratada a 250 y 350 MPa también se
analizaron por electroforesis capilar. A modo de ejemplo, en la Figura 34 se muestran los
electroforegramas obtenidos del análisis de las fracciones solubles a pH 4.6 de leche
presurizada a 350 MPa.
1 2 3 4 5 6 7 8
α-Laβ-Lg
LfBSA
Ig
1 2 3 4 5 6 7 8
1 2 3 4 5 6 7 8
B
C D
α−Laβ-Lg
Lf
β-CN
αS2-CN
κ-CN
αS1-CN
A
BSAIg
1 2 3 4 5 6 7 8
1 2 3 4 5 6 7 81 2 3 4 5 6 7 8
α-Laβ-Lg
LfBSA
Ig
1 2 3 4 5 6 7 81 2 3 4 5 6 7 8
1 2 3 4 5 6 7 81 2 3 4 5 6 7 8
B
C D
α−Laβ-Lg
Lf
β-CN
αS2-CN
κ-CN
αS1-CN
A
BSAIg
1 2 3 4 5 6 7 8

- 102 -
Figura 34. Electroforegramas de las fracciones solubles a pH 4.6 de leche desnatada control (a) y
presurizada a 350 MPa y 25ºC dutrante 5 con despresurización en 0.07 min (b). d-e muestran el
efecto del tiempo de mantenimiento de la presión (6, 10 y 15 min seguidos de 0.07 min de
despresurización, respectivamente) y f-i el efecto del tiempo de despresurización (5 minutos de
mantenimiento seguidos de 1, 5 y 10 min de despresurización, respectivamente)
Los contenidos individuales de α-La y β-Lg presentes en estas fracciones (4.6)
mostraron que, tanto a 250 como a 350 MPa, el tiempo de mantenimiento de la presión no
influyó en la desnaturalización de la α-La a ninguna de las presiones estudiadas (Figura 35 a).
Sin embargo, a 350 MPa, el aumento del tiempo de despresurización de 0.07 a 1 min produjo
una disminución del ---- en el contenido soluble de esta proteína aunque no disminuyó más
por efecto de la prolongación de la despresurización hasta los 10 min.
25
0
0.02
0.04
0.06
0.08
0
0.02
0.04
0
0.02
0
0.02
0.04
0
0.02
0.04
10 15 20
0.04 b
c
d
e
aα-La
β-Lg
10 15 20 25
α-La
β-Lg
b
g
h
i
a
A 21
4 nm
0
0.02
0.04
0.06
0.08
0
0.02
0.04
0
0.02
0
0.02
0.04
0
0.02
0.04
0.04
25
0
0.02
0.04
0.06
0.08
0
0.02
0.04
0
0.02
0
0.02
0.04
0
0.02
0.04
10 15 20
0.04 b
c
d
e
aα-La
β-Lg
10 15 20 25
α-La
β-Lg
b
g
h
i
a
A 21
4 nm
0
0.02
0.04
0.06
0.08
0
0.02
0.04
0
0.02
0
0.02
0.04
0
0.02
0.04
0.04
0
0.02
0.04
0.06
0.08
0
0.02
0.04
0
0.02
0
0.02
0.04
0
0.02
0.04
0.04
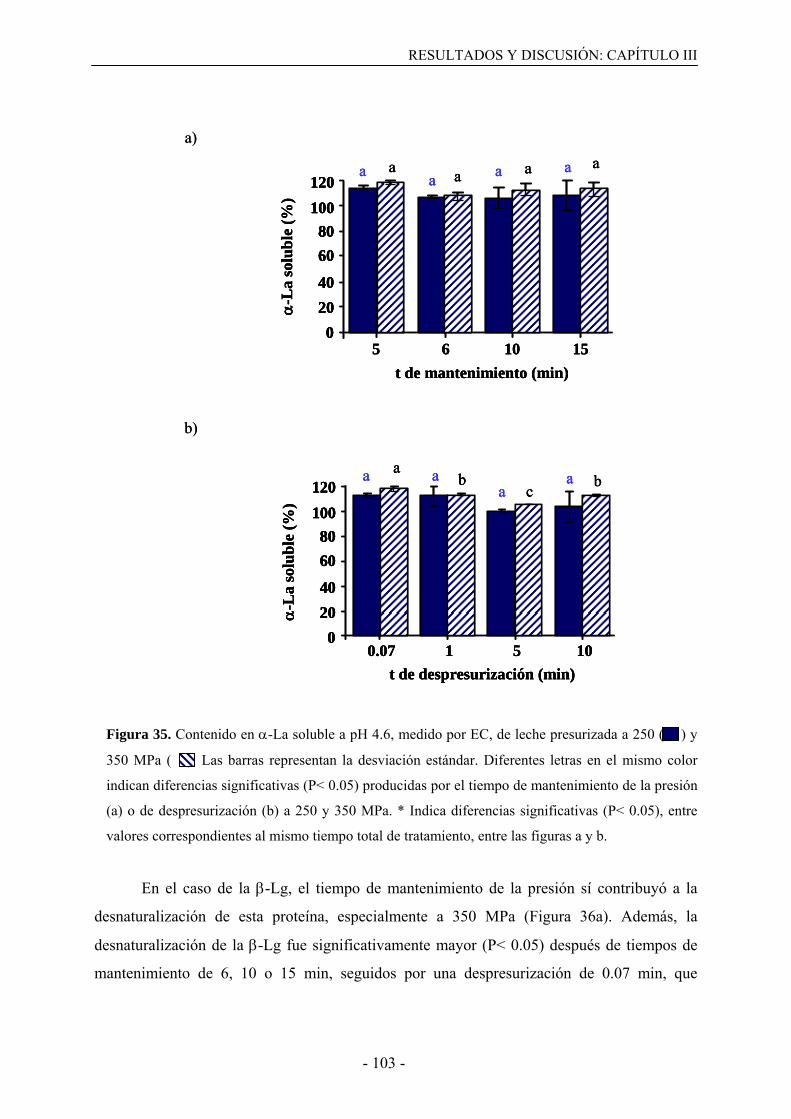
RESULTADOS Y DISCUSIÓN: CAPÍTULO III
- 103 -
Figura 35. Contenido en α-La soluble a pH 4.6, medido por EC, de leche presurizada a 250 ( ) y
350 MPa ( ). Las barras representan la desviación estándar. Diferentes letras en el mismo color
indican diferencias significativas (P< 0.05) producidas por el tiempo de mantenimiento de la presión
(a) o de despresurización (b) a 250 y 350 MPa. * Indica diferencias significativas (P< 0.05), entre
valores correspondientes al mismo tiempo total de tratamiento, entre las figuras a y b.
En el caso de la β-Lg, el tiempo de mantenimiento de la presión sí contribuyó a la
desnaturalización de esta proteína, especialmente a 350 MPa (Figura 36a). Además, la
desnaturalización de la β-Lg fue significativamente mayor (P< 0.05) después de tiempos de
mantenimiento de 6, 10 o 15 min, seguidos por una despresurización de 0.07 min, que
α-L
a so
lubl
e (%
)
t de mantenimiento (min)5 6 10 15
02040
6080
100120
a aaaa a
aa
α-L
a so
lubl
e (%
)
t de despresurización (min)
02040
6080
100120
0.07 1 5 10
a a aa
ab b
c
b)
a)
α-L
a so
lubl
e (%
)
t de mantenimiento (min)5 6 10 15
02040
6080
100120
a aaaa a
aa
α-L
a so
lubl
e (%
)
t de despresurización (min)
02040
6080
100120
0.07 1 5 10
a a aa
ab b
c
α-L
a so
lubl
e (%
)
t de mantenimiento (min)5 6 10 15
02040
6080
100120
02040
6080
100120
a aaaa a
aa
α-L
a so
lubl
e (%
)
t de despresurización (min)
02040
6080
100120
02040
6080
100120
0.07 1 5 10
a a aa
ab b
c
b)
a)

- 104 -
después de tiempos de mantenimiento de la presión de 5 min seguidos por despresurizaciones
de 1, 5 o 10 min (Figura 36 a y b).
Figura 36. Contenido en β-Lg soluble a pH 4.6, medido por EC, de leche presurizada a 250 ( ) y
350 MPa ( ). Las barras representan la desviación estándar. Diferentes letras en el mismo color
indican diferencias significativas (P< 0.05) producidas por el tiempo de mantenimiento de la presión
(a) o de despresurización (b) a 250 y 350 MPa. * Indica diferencias significativas (P< 0.05), entre
valores correspondientes al mismo tiempo total de tratamiento, entre las figuras a y b.
También se analizaron los sobrenadantes de ultracentrifugación por EC, mostrándose a
modo de ejemplo en la Figura 37 los electroforegramas de los SU de leche presurizada a 350
MPa. A tiempos de tratamiento totales equivalentes, los niveles individuales de todas la
b)
a)
020406080
100120
β-L
gso
lubl
e (%
)
5 6 10 15t de mantenimiento (min)
aab b b
ccba *
* *
020406080
100120
β-L
gso
lubl
e (%
)
0.07 1 5 10t de despresurización (min)
a a ab b
a aab
b* * *
b)
a)
020406080
100120
β-L
gso
lubl
e (%
)
5 6 10 15t de mantenimiento (min)
aab b b
ccba *
* *
020406080
100120
β-L
gso
lubl
e (%
)
0.07 1 5 10t de despresurización (min)
a a ab b
a aab
b* * *a a ab b
a aab
b* * *
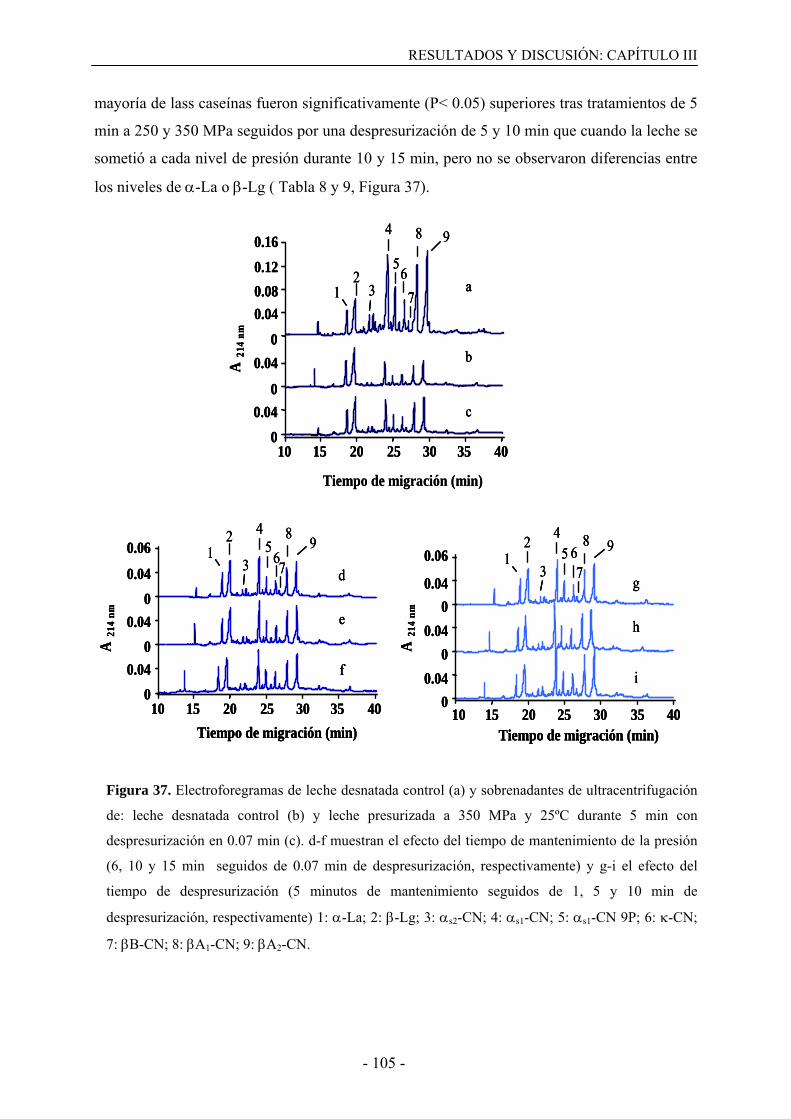
RESULTADOS Y DISCUSIÓN: CAPÍTULO III
- 105 -
mayoría de lass caseínas fueron significativamente (P< 0.05) superiores tras tratamientos de 5
min a 250 y 350 MPa seguidos por una despresurización de 5 y 10 min que cuando la leche se
sometió a cada nivel de presión durante 10 y 15 min, pero no se observaron diferencias entre
los niveles de α-La o β-Lg ( Tabla 8 y 9, Figura 37).
Figura 37. Electroforegramas de leche desnatada control (a) y sobrenadantes de ultracentrifugación
de: leche desnatada control (b) y leche presurizada a 350 MPa y 25ºC durante 5 min con
despresurización en 0.07 min (c). d-f muestran el efecto del tiempo de mantenimiento de la presión
(6, 10 y 15 min seguidos de 0.07 min de despresurización, respectivamente) y g-i el efecto del
tiempo de despresurización (5 minutos de mantenimiento seguidos de 1, 5 y 10 min de
despresurización, respectivamente) 1: α-La; 2: β-Lg; 3: αs2-CN; 4: αs1-CN; 5: αs1-CN 9P; 6: κ-CN;
7: βB-CN; 8: βA1-CN; 9: βA2-CN.
Tiempo de migración (min)
4
10 15 20 25 30 35 40
00.040.080.120.16
0
0.04
0.040
A 21
4 nm
b
c
a12
3
5
98
76
10 15 20 25 30 35 40
00.04
00.04
00.04
d
e
f
0.06
Tiempo de migración (min)
A 21
4 nm
A 21
4 nm
10 15 20 25 30 35 40
g
h
i
0.06
00.04
00.04
00.04
Tiempo de migración (min)
41
2
35
8
769
4
12
35
8
76 9
Tiempo de migración (min)
4
10 15 20 25 30 35 40
00.040.080.120.16
0
0.04
0.040
A 21
4 nm
b
c
a12
3
5
98
76
4
10 15 20 25 30 35 40
00.040.080.120.16
0
0.04
0.040
A 21
4 nm
b
c
a12
3
5
98
76
10 15 20 25 30 35 40
00.04
00.04
00.04
d
e
f
0.06
Tiempo de migración (min)
A 21
4 nm
A 21
4 nm
10 15 20 25 30 35 40
g
h
i
0.06
00.04
00.04
00.04
Tiempo de migración (min)
41
2
35
8
769
4
12
35
8
76 9
10 15 20 25 30 35 40
00.04
00.04
00.04
d
e
f
0.06
Tiempo de migración (min)
A 21
4 nm
A 21
4 nm
10 15 20 25 30 35 40
g
h
i
0.06
00.04
00.04
00.04
Tiempo de migración (min)
41
2
35
8
769
4
12
35
8
76 9
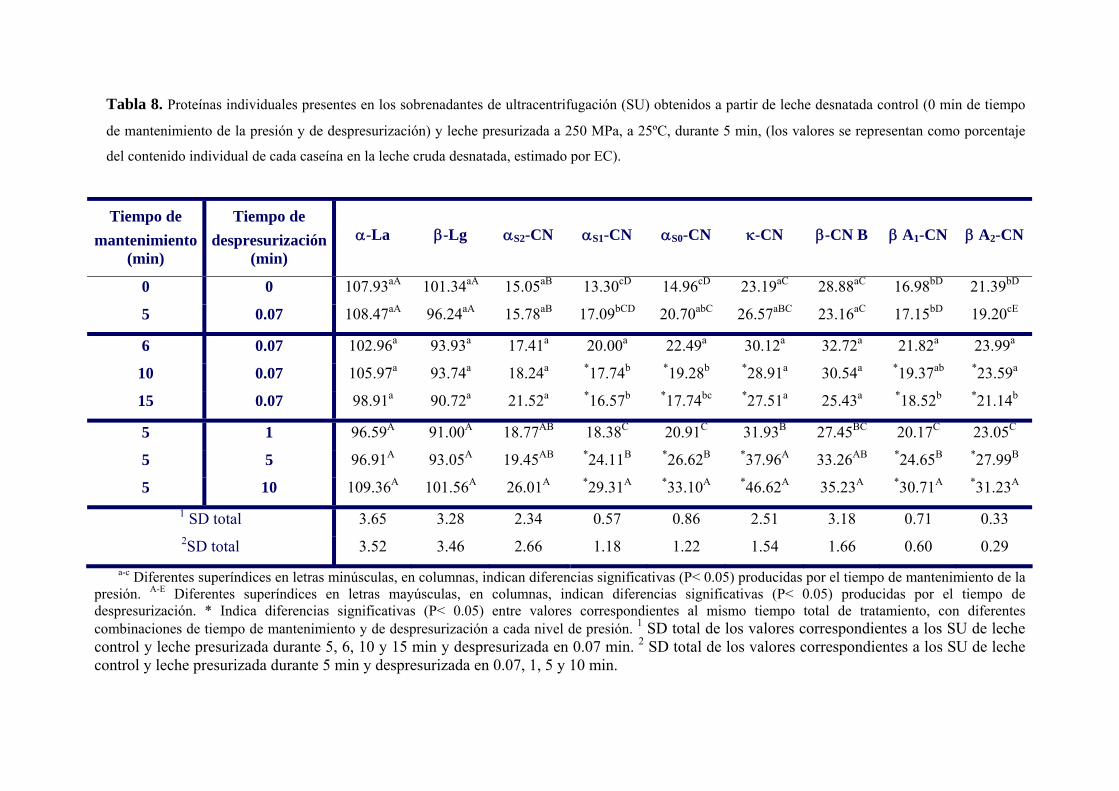
Tabla 8. Proteínas individuales presentes en los sobrenadantes de ultracentrifugación (SU) obtenidos a partir de leche desnatada control (0 min de tiempo
de mantenimiento de la presión y de despresurización) y leche presurizada a 250 MPa, a 25ºC, durante 5 min, (los valores se representan como porcentaje
del contenido individual de cada caseína en la leche cruda desnatada, estimado por EC).
a-c Diferentes superíndices en letras minúsculas, en columnas, indican diferencias significativas (P< 0.05) producidas por el tiempo de mantenimiento de la presión. A-E Diferentes superíndices en letras mayúsculas, en columnas, indican diferencias significativas (P< 0.05) producidas por el tiempo de despresurización. * Indica diferencias significativas (P< 0.05) entre valores correspondientes al mismo tiempo total de tratamiento, con diferentes combinaciones de tiempo de mantenimiento y de despresurización a cada nivel de presión. 1 SD total de los valores correspondientes a los SU de leche control y leche presurizada durante 5, 6, 10 y 15 min y despresurizada en 0.07 min. 2 SD total de los valores correspondientes a los SU de leche control y leche presurizada durante 5 min y despresurizada en 0.07, 1, 5 y 10 min.
Tiempo de mantenimiento
(min)
Tiempo de despresurización
(min) α-La β-Lg αS2-CN αS1-CN αS0-CN κ-CN β-CN B β A1-CN β A2-CN
0 0 107.93aA 101.34aA 15.05aB 13.30cD 14.96cD 23.19aC 28.88aC 16.98bD 21.39bD
5 0.07 108.47aA 96.24aA 15.78aB 17.09bCD 20.70abC 26.57aBC 23.16aC 17.15bD 19.20cE
6 0.07 102.96a 93.93a 17.41a 20.00a 22.49a 30.12a 32.72a 21.82a 23.99a
10 0.07 105.97a 93.74a 18.24a *17.74b *19.28b *28.91a 30.54a *19.37ab *23.59a
15 0.07 98.91a 90.72a 21.52a *16.57b *17.74bc *27.51a 25.43a *18.52b *21.14b
5 1 96.59A 91.00A 18.77AB 18.38C 20.91C 31.93B 27.45BC 20.17C 23.05C
5 5 96.91A 93.05A 19.45AB *24.11B *26.62B *37.96A 33.26AB *24.65B *27.99B
5 10 109.36A 101.56A 26.01A *29.31A *33.10A *46.62A 35.23A *30.71A *31.23A
1 SD total 3.65 3.28 2.34 0.57 0.86 2.51 3.18 0.71 0.33 2SD total 3.52 3.46 2.66 1.18 1.22 1.54 1.66 0.60 0.29

Tabla 9. Proteínas individuales presentes en los sobrenadantes de ultracentrifugación (SU) obtenidos a partir de leche desnatada control (0 min de tiempo
de mantenimiento de la presión y de despresurización) y leche presurizada a 350 MPa, a 25ºC, durante 5 min, (los valores se representan como porcentaje
del contenido individual de cada caseína en la leche cruda desnatada, estimado por EC).
a-c Diferentes superíndices en letras minúsculas, en columnas, indican diferencias significativas (P< 0.05) producidas por el tiempo de mantenimiento de la presión. A-D Diferentes superíndices en letras mayúsculas, en columnas, indican diferencias significativas (P< 0.05) producidas por el tiempo de despresurización. * Indica diferencias significativas (P< 0.05) entre valores correspondientes al mismo tiempo total de tratamiento, con diferentes combinaciones de tiempo de mantenimiento y de despresurización a cada nivel de presión. 1 SD total de los valores correspondientes a los SU de leche control y leche presurizada durante 5, 6, 10 y 15 min y despresurizada en 0.07 min. 2 SD total de los valores correspondientes a los SU de leche control y leche presurizada durante 5 min y despresurizada en 0.07, 1, 5 y 10 min.
Tiempo de mantenimiento
(min)
Tiempo de despresurización
(min) α-La β-Lg αS2-CN αS1-CN αS0-CN κ-CN β-CN B β A1-CN β A2-CN
0 0 107.93aA 101.34aB 15.05bB 13.30dD 14.96bD 23.19cD 24.88bC 16.98cD 21.39cC
5 0.07 102.61aA 90.57abA 29.18aA 25.34cC 27.19aC 33.89bCD 35.91aB 25.93bC 29.75bB
6 0.07 100.53a 88.61bc 27.66a 26.88bc 29.57a 37.94ab 37.76a 26.82b 29.58b
10 0.07 95.85a 78.33c 29.75a 29.64ab 26.86a 40.51a *34.66a *29.66a *32.56ab
15 0.07 91.26a 79.73bc 32.17a *30.29a *30.72a *40.97a *37.45a *31.19a *36.07a
5 1 97.93A 88.62A 30.73A 28.52BC 29.56BC 38.64 BC 37.48B 30.01C 35.07B
5 5 100.77A 88.41A 31.24A 32.51B 33.92AB 49.35 AB *45.97A *37.70B *43.06A
5 10 99.89A 88.00A 35.33A *37.91A *39.31A *53.49A *49.00A *44.18A *48.18A
1SD total 4.65 3.13 2.17 0.87 2.17 1.76 2.38 0.46 1.42 2SD total 3.79 1.82 2.30 1.23 1.31 3.14 2.22 1.20 1.90
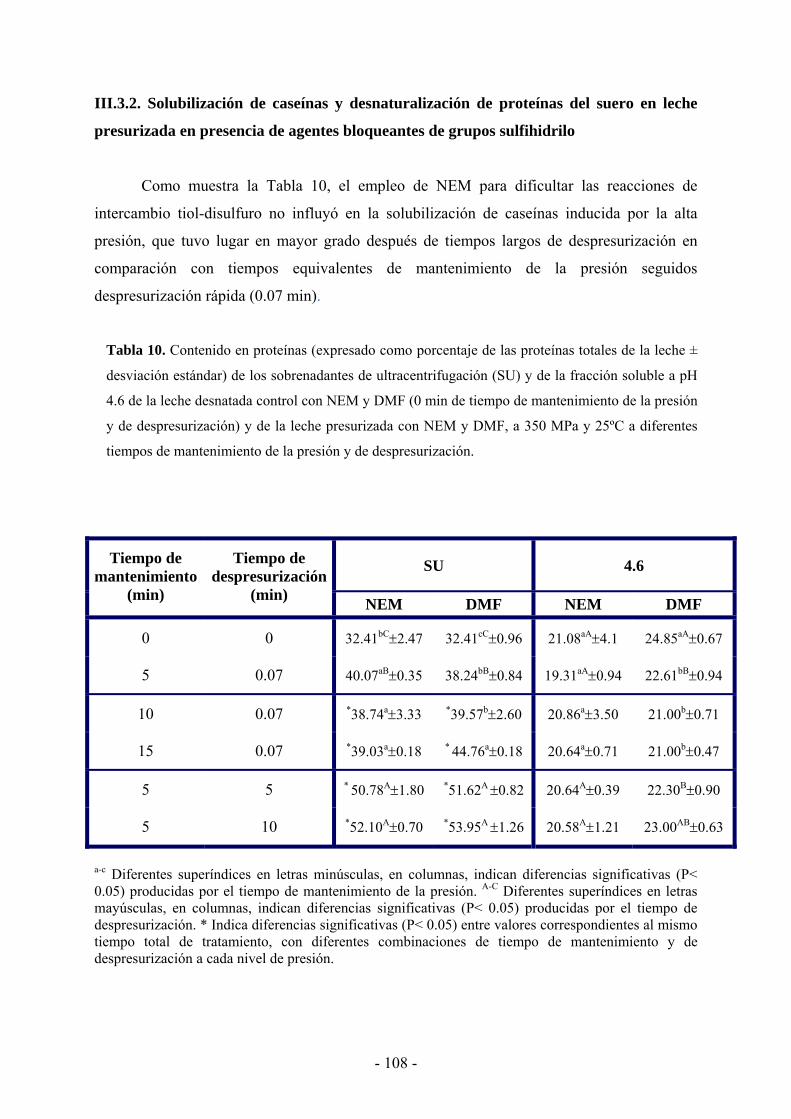
- 108 -
III.3.2. Solubilización de caseínas y desnaturalización de proteínas del suero en leche
presurizada en presencia de agentes bloqueantes de grupos sulfihidrilo
Como muestra la Tabla 10, el empleo de NEM para dificultar las reacciones de
intercambio tiol-disulfuro no influyó en la solubilización de caseínas inducida por la alta
presión, que tuvo lugar en mayor grado después de tiempos largos de despresurización en
comparación con tiempos equivalentes de mantenimiento de la presión seguidos
despresurización rápida (0.07 min).
Tabla 10. Contenido en proteínas (expresado como porcentaje de las proteínas totales de la leche ±
desviación estándar) de los sobrenadantes de ultracentrifugación (SU) y de la fracción soluble a pH
4.6 de la leche desnatada control con NEM y DMF (0 min de tiempo de mantenimiento de la presión
y de despresurización) y de la leche presurizada con NEM y DMF, a 350 MPa y 25ºC a diferentes
tiempos de mantenimiento de la presión y de despresurización.
a-c Diferentes superíndices en letras minúsculas, en columnas, indican diferencias significativas (P< 0.05) producidas por el tiempo de mantenimiento de la presión. A-C Diferentes superíndices en letras mayúsculas, en columnas, indican diferencias significativas (P< 0.05) producidas por el tiempo de despresurización. * Indica diferencias significativas (P< 0.05) entre valores correspondientes al mismo tiempo total de tratamiento, con diferentes combinaciones de tiempo de mantenimiento y de despresurización a cada nivel de presión.
SU 4.6 Tiempo de mantenimiento
(min)
Tiempo de despresurización
(min) NEM DMF NEM DMF
0 0 32.41bC±2.47 32.41cC±0.96 21.08aA±4.1 24.85aA±0.67
5 0.07 40.07aB±0.35 38.24bB±0.84 19.31aA±0.94 22.61bB±0.94
10 0.07 *38.74a±3.33 *39.57b±2.60 20.86a±3.50 21.00b±0.71
15 0.07 *39.03a±0.18 * 44.76a±0.18 20.64a±0.71 21.00b±0.47
5 5 * 50.78A±1.80 *51.62A ±0.82 20.64A±0.39 22.30B±0.90
5 10 *52.10A±0.70 *53.95A ±1.26 20.58A±1.21 23.00AB±0.63

RESULTADOS Y DISCUSIÓN: CAPÍTULO III
- 109 -
Por otro lado, como se esperaba, la adición de NEM a la leche previno la
desnaturalización de las proteínas de suero a 350 MPa. Los experimentos en los que se añadió
DMF, en la que se disolvió el NEM, a la leche también sugirieron una mayor incidencia del
tiempo de mantenimiento de la presión que del tiempo de despresurización en la
desnaturalización de las proteínas del suero, aunque no hubo diferencias significativas entre
los efectos de ambos.
III.4. Discusión
De acuerdo con los resultados obtenidos, a 250 MPa y, especialmente, a 350 MPa se
produjo la liberación de caseínas no sedimenables a la fase sérica, influyendo de manera más
importante el tiempo de despresurización que el de mantenimiento de la presión en este
proceso (Tablas 7-9, Figura 33 A y B, Figura 37). Se ha descrito que, a presiones superiores a
100 MPa, tiene lugar la solubilización de las caseínas como resultado de la ruptura micelar
(López-Fandiño y col., 1998; Huppertz y col., 2004c; Anema, 2008b). Sin embargo, a
presiones de 250 y 300 MPa, si bien se desintegra la micela de caseína durante los primeros 5-
10 min de tratamiento durante tratamientos más prolongados se produce agregación de las
proteínas debido a un aumento de las interacciones hidrofóbicas a esas presiones intermedias,
aunque tratamientos a 250 MPa de 5 a 15 min, como los usados en este estudio, son
demasiados cortos para inducir agregación micelar (Huppertz y col., 2006b; Huppertz y De
Kruif, 2006, 2007b). Por otra parte, otros autores también han observado agregación de
partículas micelares, debido al restablecimiento de los enlaces hidrofóbicos tras la liberación
de la presión, siempre y cuando estas micelas mantengan un cierto grado de integridad y parte
de la estructura del CCP intacto (Huppertz y col., 2006b; Orlien y col., 2006; Anema, 2008b).
A pesar de que la presión rompe los enlaces electrostáticos e hidrofóbicos éstos
podrían empezar a formarse de nuevo durante la despresurización. Sin embargo, nuestros
resultados mostraron claramente que durante la fase de despresurización se produce un
aumento significativamente mayor del contenido de caseínas no sedimentables que el que
tiene lugar durante la fase de mantenimiento de la presión. Según Merel-Rausch y col. (2006;
2007), a presiones superiores a 200 MPa, la velocidad de despresurización juega un papel
importante en la modificación de las caseínas, ya que, la despresurización lenta favorecería la
reasociación de las caseínas, mientras que la rápida perturbaría a los mecanismos de

- 110 -
asociación, formándose estructuras o agregados completamente nuevos. Teniendo en cuenta
las observaciones de Merl-Rausch y col. (2006; 2007), nuestros resultados indican que es
posible que tenga lugar una importante disociación micelar, que aumente las caseínas no
sedimentables, bajo condiciones que también promuevan la reagregación micelar. De hecho,
en muestras de leche sometidas a presiones de 250-300 MPa, en las que se ha producido
agregación micelar, se encuentran altos niveles de caseínas no sedimentables (Anema y col.,
2005a). Esto sugiere que la agregación micelar es un fenómeno independiente de la
disgregación micelar, aún cuando parece necesario un nivel mínimo de ruptura micelar para
que se produzca, posteriormente, agregación (Huppertz y De Kruif, 2006; Anema, 2008b).
Los resultados obtenidos apoyaron estudios previos que indican que la β-Lg se
desnaturaliza a presiones superiores de 100 MPa en función del nivel de presión y la duración
del tratamiento (López-Fandiño y col., 1996; Anema y col., 2005b). El desplegamiento
reversible de la β-Lg inducido por la presión (exposición de regiones hidrofóbicas y del grupo
sulfhidrilo libre) va seguido de una desnaturalización irreversible, por autoagregación o
asociación a otras proteínas de suero, o a la κ-CN (López-Fandiño y col., 1997; Needs y col.,
2000b; Patel y col. 2006). Al igual que nosotros, otros autores también han observado que la
mayoría de la β-Lg desnaturalizada permanece soluble en leche presurizada a 250 y 350 MPa,
al pH normal de la leche (Tablas 2 y 3, Figuras 36 y 33) (López-Fandiño y col., 1998; Anema,
1998), lo que sugiere que las moléculas de β-Lg interaccionan preferentemente entre ellas o
con partículas de caseína no sedimentable. Además, nuestros resultados (Tabla 7, Figuras 34 y
36) pusieron de manifiesto que la β-Lg se desnaturalizó principalmente durante la fase de
mantenimiento de la presión, bajo condiciones de alta solubilidad del fosfato cálcico e
inhibición de las interacciones hidrofóbicas, produciéndose muy poca desnaturalización
adicional durante la fase de despresurización. Esto apoya la hipótesis de que las reacciones de
intercambio -SH-S-S se dan principalmente durante el tratamiento de presión (Belloque y col.,
2000) y sugiere que las uniones hidrofóbicas que se restablecen durante la fase de liberación
de la presión no participan de manera importante en la desnaturalización de la β-Lg.
Nuestros resultados también sugirieron que la desnaturalización de las proteínas de
suero no ejerce una influencia apreciable en el aumento del contenido de caseína no
sedimentable producido durante la fase de despresurización, ya que dicho aumento no se
acompañó de cambios importantes en los niveles de proteínas de suero presentes en los SU

RESULTADOS Y DISCUSIÓN: CAPÍTULO III
- 111 -
(Tablas 2 y 3, Figuras 33 A,B y 37), ni de un aumento en la desnaturalización de las proteínas
de suero, determinado por la pérdida de solubilidad a pH 4.6 (Tabla 7, Figuras 33 C, D y 36).
Como era de esperar, la adición de NEM previno la formación de enlaces disulfuro, evitando
la en gran medida la desnaturalización de las proteínas de suero y su asociación con las
caseínas (Huppertz y col., 2004b), pero no ejerció ningún efecto en la liberación de caseínas a
la fase soluble de la leche que tuvo lugar durante la fase de despresurización (Tabla 10). De
modo similar, es probable que los cambios en el tamaño micelar inducidos por la presión sean
independientes de las proteínas de suero, ya que se encuentran tamaños micelares similares en
leche presurizada tanto con la adición de NEM como sin él (Huppertz y col., 2004d).
Igualmente la disgregación micelar y la reagregación que tienen lugar a 250 y 300 MPa son
similares en presencia y en ausencia de proteínas de suero (Anema y col., 2005a; Huppertz y
De Kruif, 2007b).
III.5. Conclusiones
Este trabajo muestra el efecto de la velocidad de despresurización en la distribución de
proteínas entre las fases soluble y coloidal de la leche presurizada. El contenido en caseína
soluble aumentó significativamente durante la fase de despresurización respecto a la fase de
mantenimiento de la presión, mientras que la desnaturalización de la β-Lg tuvo lugar,
mediante enlaces disulfuro, fundamentalmente durante la fase de mantenimiento. Estos
resultados corroboran los recogidos en el capítulo anterior, demostrando que la
desnaturalización de las proteínas de suero no influye en la ruptura micelar y en la
subsiguiente solubilización de caseínas que tiene lugar en la leche presurizada y evidencian la
importancia de la fase de despresurización en este proceso, de modo que cuanto más lenta sea
la velocidad de despresurización, mayor es el nivel de caseína soluble.

CAPITULO IV
Estudio de la distribución proteica en muestras tratadas
con altas presiones entre 700 y 900 MPa


RESULTADOS Y DISCUSIÓN: CAPÍTULO IV
- 115 -
IV.1. Introducción
En las últimas tres décadas, el conocimiento sobre el efecto de las altas presiones en
los alimentos se ha ampliado enormemente sobre todo, en la leche y sus proteínas,
encontrándonos varios trabajos en la bibliografía que resumen estos efectos (Huppertz y col.,
2002; López-Fandiño, 2006a; Huppertz y col., 2006a; Considine y col., 2007). Este gran
conocimiento ha permitido el desarrollo de esta técnica como medio para la conservación de
los alimentos, aumentado la implantación de equipos de alta presión, capaces de alcanzar los
650 MPa, en las industrias alimentarias. En concreto, en el 2009 los equipos industriales
instalados fueron 132 en todo el mundo, existiendo una gran variedad de productos
comerciales presurizados como calostros, zumos, guacamole, fiambre, arroz, etc (Barbosa-
Cánovas y Bermúdez-Aguirre, 2011; Tonello, 2011). Sin embargo, la tecnología continua
avanzando y actualmente podemos encontrar equipos a escala piloto que pueden llegar hasta
los 900 MPa e incluso hasta 1800 MPa en equipos de laboratorio (Bermúdez-Aguirre y
Barbosa-Cánovas, 2011). Lo que hace necesario continuar investigando en los efectos de las
muy altas presiones (> 650 MPa) en los alimentos.
En la leche, las altas presiones reducen sus propiedades de dispersión de la luz,
disminuyen su turbidez y cambian su apariencia visual, debido a la desintegración de la
micela de caseína en estructuras más pequeñas (Needs y col., 2000b; Considine y col., 2007).
Tanto las micelas de caseínas de la leche presurizada como las de leche sin presurizar se
mantienen primariamente por la interacciones hidrofóbicas e uniones con el CCP (Huppertz y
col., 2006a) aunque también juega un papel muy importante en su estabilidad la κ-CN (Fox y
Brodkorb, 2008). La desintegración de la micela inducida por la presión, se produce por la
solubilización del CCP que forma parte de las micelas (López-Fandiño y col., 1998; Huppertz
y De Kruif, 2007a) y por la ruptura de las interacciones hidrofóbicas, de hidrogeno y de van
der Waals (López-Fandiño, 2006a).
Como consecuencia de la desintegración de la micela inducida por la presión, el
tamaño medio de la micela cambia y el grado de su modificación variará dependiendo de
varios factores como la duración del tratamiento, la temperatura, la presencia de proteínas de
suero, el pH de la leche, la aplicación de varios ciclos de presión, etc (Huppertz y col., 2006a).
Por ejemplo, el tamaño de la micela en leche tratada a 250-300 MPa puede ser mayor o menor
que los de la leche sin tratar dependiendo de la duración y la temperatura del tratamiento

- 116 -
(Huppertz y col., 2004a; Regnault y col., 2004; Anema y col., 2005a). Sin embargo, los
tratamientos a presiones ≥400 MPa reducen considerablemente el tamaño micelar (Needs y
col., 2000b; Huppertz y col., 2004a; Anema y col., 2005a). Gebhardt y col. (2006) observaron
por microscopía de fuerza atómica ex-situ que las micelas nativas (170 nm) empiezan a
descomponerse a presiones bajas (>40 MPa) formando pequeñas partículas de entre 10 y 20
nm. Sin embargo, para una completa descomposición en monómeros es necesario presiones
superiores a 280 MPa.
Otro de los efectos de las altas presiones en la leche es que produce desnaturalización
de las proteínas del suero ya que produce el desplegamiento de estas proteínas. Sin embargo,
la estabilidad de las proteínas séricas a la presión varia dependiendo de su estructura, de
hecho la β-Lg es la más sensible y la α-La la más resistente a la presión (López-Fandiño,
2006a). Las altas presiones producen el desplegamiento de la estructura de la β-Lg
exponiendo el grupo sulfidrilo libre, que puede reaccionar con otras proteínas formando como
la κ-CN, αs2-CN, α-La o β-Lg (Huppertz y col., 2004b; Patel y col., 2006). La β-Lg empieza
a desnaturalizarse a partir de los 200 MPa, Anema (2008a) observó un ∼30% de
desnaturalización en tratamientos de 30 min que fue incrementando con el aumento de la
presión, llegando a una desnaturalización del 90% a 800 MPa. En cambio la desnaturalización
de la α-La no se produce hasta los 400-500 MPa (Anema y col., 2005b; Hinrichs y
Rademacher, 2005; Anema, 2008a) observándose desnaturalizaciones de en torno al 30-40%
a 800 MPa (Huppertz y col. 2004b; Anema, 2008a)
A pesar de que ya existen algunos estudios sobre el efecto de presiones hasta 800 MPa
en el tamaño micelar y la desnaturalización de las proteínas mayoritarias del suero aún se
necesita profundizar en el efecto de estas presiones en la disgregación micelar.
Así, el objetivo propuesto para este estudio fue investigar el efecto de la aplicación de
muy altas presiones (700, 800 y 900 MPa) a la leche, en la distribución proteica de diferentes
fracciones lácteas, profundizando en el estudio del tamaño de la micela de caseína y la
desnaturalización de las proteínas minoritarias del suero, entre otros aspectos.
IV.2. Materiales y métodos La Figura 38 resume el procedimiento experimental seguido en este trabajo.

RESULTADOS Y DISCUSIÓN: CAPÍTULO IV
- 117 -
IV.2.1. Preparación de las muestras
Para realizar este estudio se utilizó leche cruda de vaca procedente de una granja local. La leche se atemperó a 37ºC y se procedió a su desnatado.
IV.2.2. Tratamientos de presurización
Tras el desnatado de la leche, se llevaron a cabo los tratamientos con alta presión a
250, 350, 450, 550, 700, 800 y 900 MPa, a 15ºC durante 5 min. Para ello, la leche,
atemperada a 15ºC, se dispensó en bolsas selladas térmicamente que se introdujeron en un
equipo de alta presión TE 9000 (Thiot Ingenierie, NC Hyperbaric, Bretenoux, France-Burgos,
España). Este equipo es capaz de alcanzar 900 MPa a una velocidad de 6 MPa/seg, tiene una
cámara de presurización de 2 L de capacidad, y utiliza aceite de silicona como fluido
transmisor, como se ha explicado en detalle en el capítulo II, apartado II.2.2. Los tratamientos
se hicieron por duplicado.
IV.2.3. Fraccionamiento y análisis
Posteriormente, las muestras de leche presurizada se fraccionaron y se analizaron
como se indica en el apartadode Materiales y Métodos generales.(pag. 43-50).

- 118 -
Figura 38. Esquema de la preparación, tratamientos con alta presión, fraccionamiento y análisis de
las muestras
Lechedesnatada
Leche cruda de vaca
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
3000
x g
, 20º
C,
30 m
in15
ºC, 5
min
Thiot EngineerTE 9000
LechePresurizada250 MPa
Lechepresurizada
350 MPa
Lechepresurizada
450 MPa
Lechepresurizada550 MPa
Lechepresurizada700 MPa
Lechepresurizada800 MPa
Lechepresurizada900 MPa
Lechedesnatada0.1 MPa
Precipitacióna pH 4.6
Ultracentrifugación100000 x g, 20ºC, 1h
NITRÓGENO(Kjeldahl)
ELECTROFORESISCAPILAR
PAGE-SDS
PROTEÍNAS
MICELA DE CASEÍNA
(Microscopía electrónica)
Lf e IgA(ELISA)
Análisis
Des
nata
doTr
atam
ient
o co
n al
ta p
resi
ónFr
acci
onam
ient
oA
nális
is
Filtrar por Whatman nº 41
HCl2 N
Reposar 20 min,Centrifugar (4000 x g, 5ºC, 20 min)Filtrar
SU 4.6
Lechedesnatada
Lechedesnatada
Leche cruda de vacaLeche cruda de vaca
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
Lechedesnatada
3000
x g
, 20º
C,
30 m
in15
ºC, 5
min
Thiot EngineerTE 9000
Thiot EngineerTE 9000
LechePresurizada250 MPa
LechePresurizada250 MPa
Lechepresurizada
350 MPa
Lechepresurizada
350 MPa
Lechepresurizada
450 MPa
Lechepresurizada
450 MPa
Lechepresurizada550 MPa
Lechepresurizada550 MPa
Lechepresurizada700 MPa
Lechepresurizada700 MPa
Lechepresurizada800 MPa
Lechepresurizada800 MPa
Lechepresurizada900 MPa
Lechepresurizada900 MPa
Lechedesnatada0.1 MPa
Lechedesnatada0.1 MPa
Precipitacióna pH 4.6
Ultracentrifugación100000 x g, 20ºC, 1h
NITRÓGENO(Kjeldahl)
ELECTROFORESISCAPILAR
PAGE-SDS
PROTEÍNAS
MICELA DE CASEÍNA
(Microscopía electrónica)
Lf e IgA(ELISA)
Análisis
Des
nata
doTr
atam
ient
o co
n al
ta p
resi
ónFr
acci
onam
ient
oA
nális
is
Filtrar por Whatman nº 41
HCl2 N
Reposar 20 min,Centrifugar (4000 x g, 5ºC, 20 min)Filtrar
SU 4.6

RESULTADOS Y DISCUSIÓN: CAPÍTULO IV
- 119 -
IV.3. Resultados
IV.3.1. Efecto de la presión en el tamaño de la micela de caseína
Las fotografías obtenidas por microscopía electrónica de transmisión de la leche
sometida a altas presiones se presentan en la Figura 39. La medida del tamaño de las micelas
mostró que en la leche control presentaban una gran dispersión en el tamaño, desde 255 nm a
17 nm, que fue acortándose por efecto de la presión (Tabla 11, Figura 39). La aplicación de
presión provocó la ruptura de las micelas de caseína y disminuyó el número de micelas de
gran tamaño (Figura 39, Figura 40). A 250 MPa, todavía eran visibles algunas micelas
grandes (231 nm) que desaparecieron a presiones superiores. A partir de 450 MPa, las micelas
mayores medían entre 100 y115 nm, y estaban presentes en bajo número y a todas las
presiones (Tabla 11, Figura 40). Este hecho provocó el aumento del número de micelas de
tamaño comprendido entre 20 y 60 nm que predominantes a presiones ≤ 450 MPa (Figura 40).
Tabla 11. Tamaño de las micelas de caseína (nm) observadas por microscopía electrónica de
trasmisión de leche desnatada control (0.1 MPa) y leche sometida a diferentes presiones durante 5
min.
Tamaño de las micelas de caseína (nm)
Media SD Mínimo Máximo Nº micelas
medidas
0.1 83.82 44.45 16.86 254.89 157
250 56.49 38.39 21.94 234.38 187
450 45.97 15.32 18.84 105.30 179
550 48.10 17.40 14.05 114.63 149
700 52.69 15.34 22.47 103.60 182
800 51.86 15.77 19.86 112.44 164
900 53.65 16.89 19.86 106.90 170
SD indica desviación estándar.

- 120 -
Figura 39. Fotografías de microscopía electrónica de transmisión de leche desnatada control (a) y
leche presurizada a 250, 450, 700, 800 y 900 MPa, 15ºC durante 5 min.
a
b
c
d
e
f
a
b
c
d
e
f
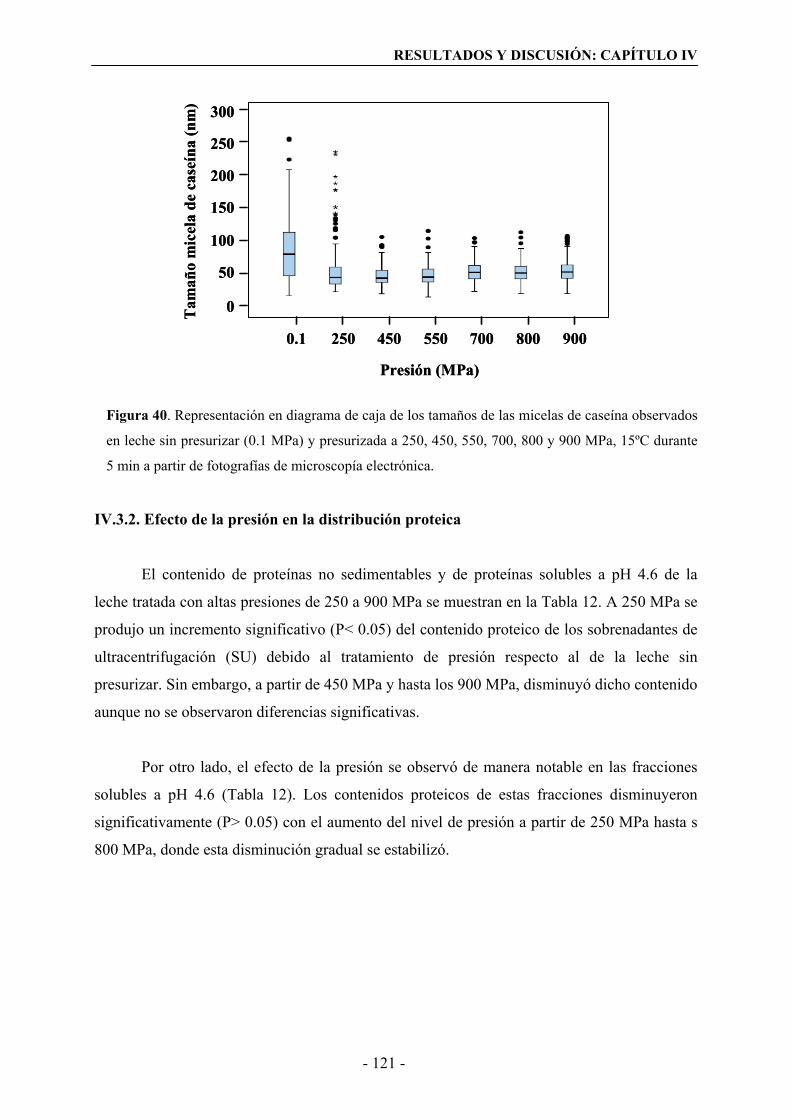
RESULTADOS Y DISCUSIÓN: CAPÍTULO IV
- 121 -
300
250
200
150
100
50
0Tam
año
mic
ela
de c
aseí
na (n
m)
0.1 250 450 550 700 800 900
Presión (MPa)
300
250
200
150
100
50
0Tam
año
mic
ela
de c
aseí
na (n
m)
0.1 250 450 550 700 800 900
Presión (MPa)
Figura 40. Representación en diagrama de caja de los tamaños de las micelas de caseína observados
en leche sin presurizar (0.1 MPa) y presurizada a 250, 450, 550, 700, 800 y 900 MPa, 15ºC durante
5 min a partir de fotografías de microscopía electrónica.
IV.3.2. Efecto de la presión en la distribución proteica
El contenido de proteínas no sedimentables y de proteínas solubles a pH 4.6 de la
leche tratada con altas presiones de 250 a 900 MPa se muestran en la Tabla 12. A 250 MPa se
produjo un incremento significativo (P< 0.05) del contenido proteico de los sobrenadantes de
ultracentrifugación (SU) debido al tratamiento de presión respecto al de la leche sin
presurizar. Sin embargo, a partir de 450 MPa y hasta los 900 MPa, disminuyó dicho contenido
aunque no se observaron diferencias significativas.
Por otro lado, el efecto de la presión se observó de manera notable en las fracciones
solubles a pH 4.6 (Tabla 12). Los contenidos proteicos de estas fracciones disminuyeron
significativamente (P> 0.05) con el aumento del nivel de presión a partir de 250 MPa hasta s
800 MPa, donde esta disminución gradual se estabilizó.
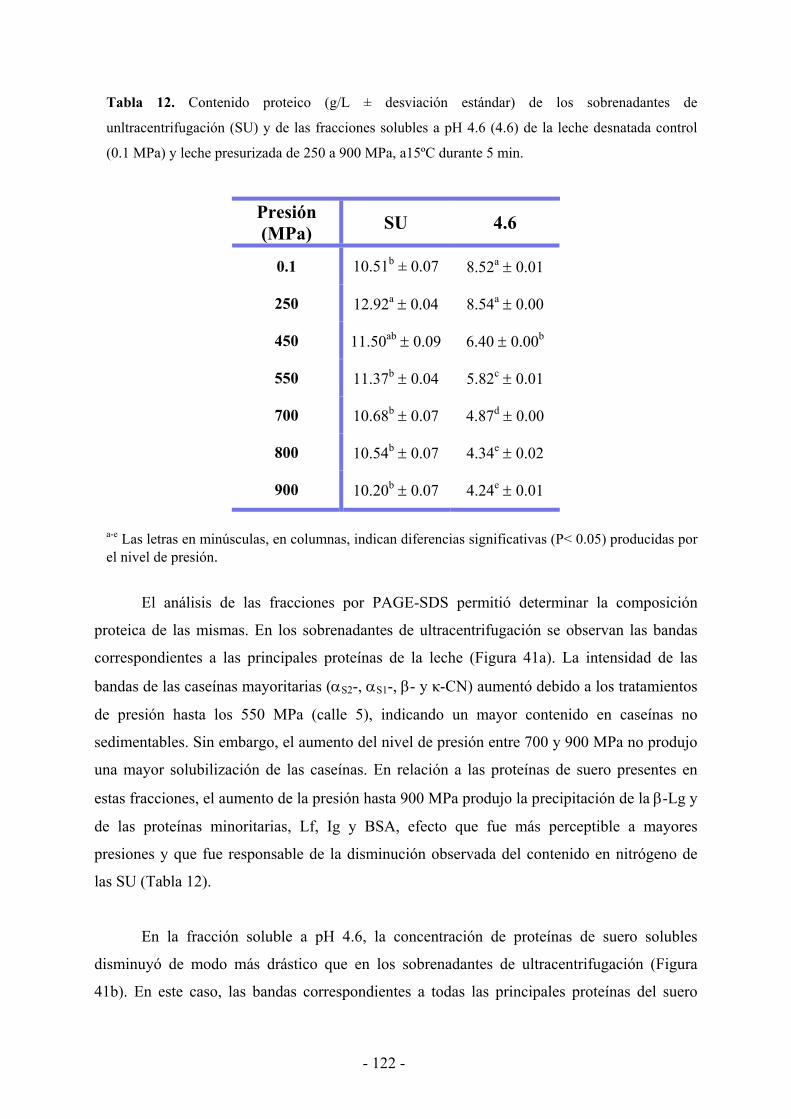
- 122 -
Tabla 12. Contenido proteico (g/L ± desviación estándar) de los sobrenadantes de
unltracentrifugación (SU) y de las fracciones solubles a pH 4.6 (4.6) de la leche desnatada control
(0.1 MPa) y leche presurizada de 250 a 900 MPa, a15ºC durante 5 min.
Presión (MPa) SU 4.6
0.1 10.51b ± 0.07 8.52a ± 0.01
250 12.92a ± 0.04 8.54a ± 0.00
450 11.50ab ± 0.09 6.40 ± 0.00b
550 11.37b ± 0.04 5.82c ± 0.01
700 10.68b ± 0.07 4.87d ± 0.00
800 10.54b ± 0.07 4.34e ± 0.02
900 10.20b ± 0.07 4.24e ± 0.01
a-e Las letras en minúsculas, en columnas, indican diferencias significativas (P< 0.05) producidas por el nivel de presión.
El análisis de las fracciones por PAGE-SDS permitió determinar la composición
proteica de las mismas. En los sobrenadantes de ultracentrifugación se observan las bandas
correspondientes a las principales proteínas de la leche (Figura 41a). La intensidad de las
bandas de las caseínas mayoritarias (αS2-, αS1-, β- y κ-CN) aumentó debido a los tratamientos
de presión hasta los 550 MPa (calle 5), indicando un mayor contenido en caseínas no
sedimentables. Sin embargo, el aumento del nivel de presión entre 700 y 900 MPa no produjo
una mayor solubilización de las caseínas. En relación a las proteínas de suero presentes en
estas fracciones, el aumento de la presión hasta 900 MPa produjo la precipitación de la β-Lg y
de las proteínas minoritarias, Lf, Ig y BSA, efecto que fue más perceptible a mayores
presiones y que fue responsable de la disminución observada del contenido en nitrógeno de
las SU (Tabla 12).
En la fracción soluble a pH 4.6, la concentración de proteínas de suero solubles
disminuyó de modo más drástico que en los sobrenadantes de ultracentrifugación (Figura
41b). En este caso, las bandas correspondientes a todas las principales proteínas del suero
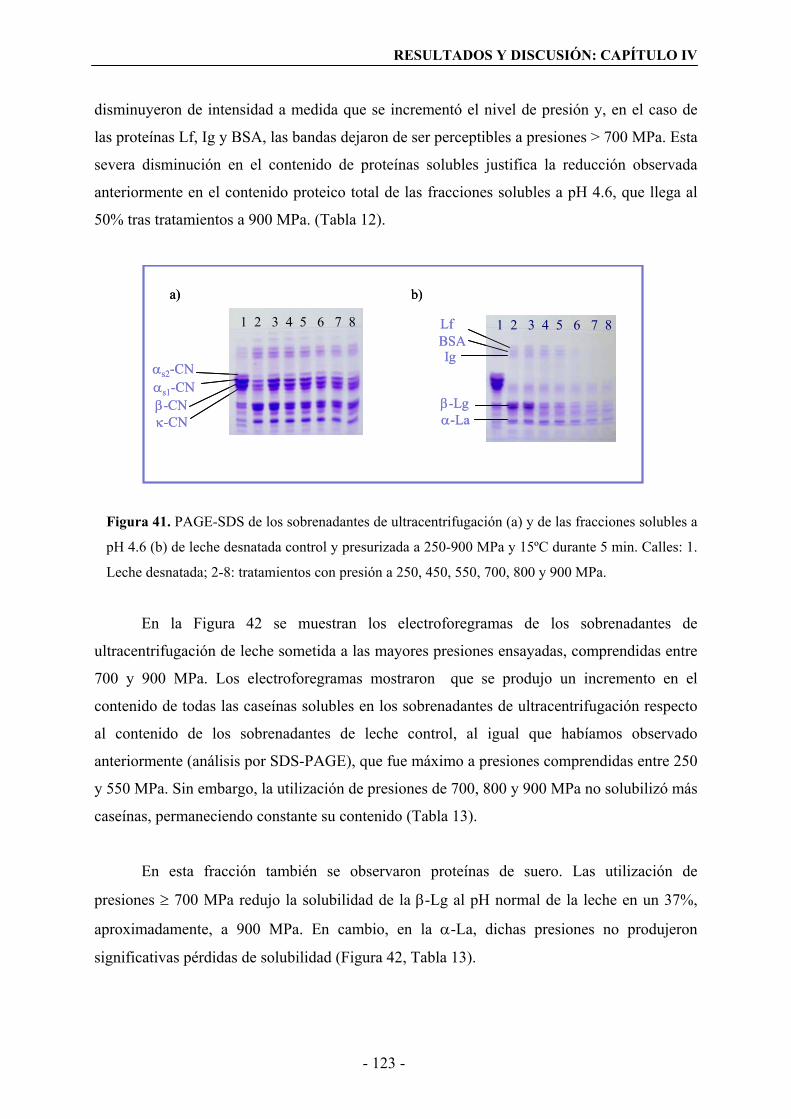
RESULTADOS Y DISCUSIÓN: CAPÍTULO IV
- 123 -
disminuyeron de intensidad a medida que se incrementó el nivel de presión y, en el caso de
las proteínas Lf, Ig y BSA, las bandas dejaron de ser perceptibles a presiones > 700 MPa. Esta
severa disminución en el contenido de proteínas solubles justifica la reducción observada
anteriormente en el contenido proteico total de las fracciones solubles a pH 4.6, que llega al
50% tras tratamientos a 900 MPa. (Tabla 12).
Figura 41. PAGE-SDS de los sobrenadantes de ultracentrifugación (a) y de las fracciones solubles a
pH 4.6 (b) de leche desnatada control y presurizada a 250-900 MPa y 15ºC durante 5 min. Calles: 1.
Leche desnatada; 2-8: tratamientos con presión a 250, 450, 550, 700, 800 y 900 MPa.
En la Figura 42 se muestran los electroforegramas de los sobrenadantes de
ultracentrifugación de leche sometida a las mayores presiones ensayadas, comprendidas entre
700 y 900 MPa. Los electroforegramas mostraron que se produjo un incremento en el
contenido de todas las caseínas solubles en los sobrenadantes de ultracentrifugación respecto
al contenido de los sobrenadantes de leche control, al igual que habíamos observado
anteriormente (análisis por SDS-PAGE), que fue máximo a presiones comprendidas entre 250
y 550 MPa. Sin embargo, la utilización de presiones de 700, 800 y 900 MPa no solubilizó más
caseínas, permaneciendo constante su contenido (Tabla 13).
En esta fracción también se observaron proteínas de suero. Las utilización de
presiones ≥ 700 MPa redujo la solubilidad de la β-Lg al pH normal de la leche en un 37%,
aproximadamente, a 900 MPa. En cambio, en la α-La, dichas presiones no produjeron
significativas pérdidas de solubilidad (Figura 42, Tabla 13).
α-Laβ-Lg
LfBSAIg
1 2 3 4 5 6 7 8
β-CN
αs2-CN
κ-CN
αs1-CN
1 2 3 4 5 6 7 8
a) b)
α-Laβ-Lg
LfBSAIg
1 2 3 4 5 6 7 8
β-CN
αs2-CN
κ-CN
αs1-CN
1 2 3 4 5 6 7 8
a) b)
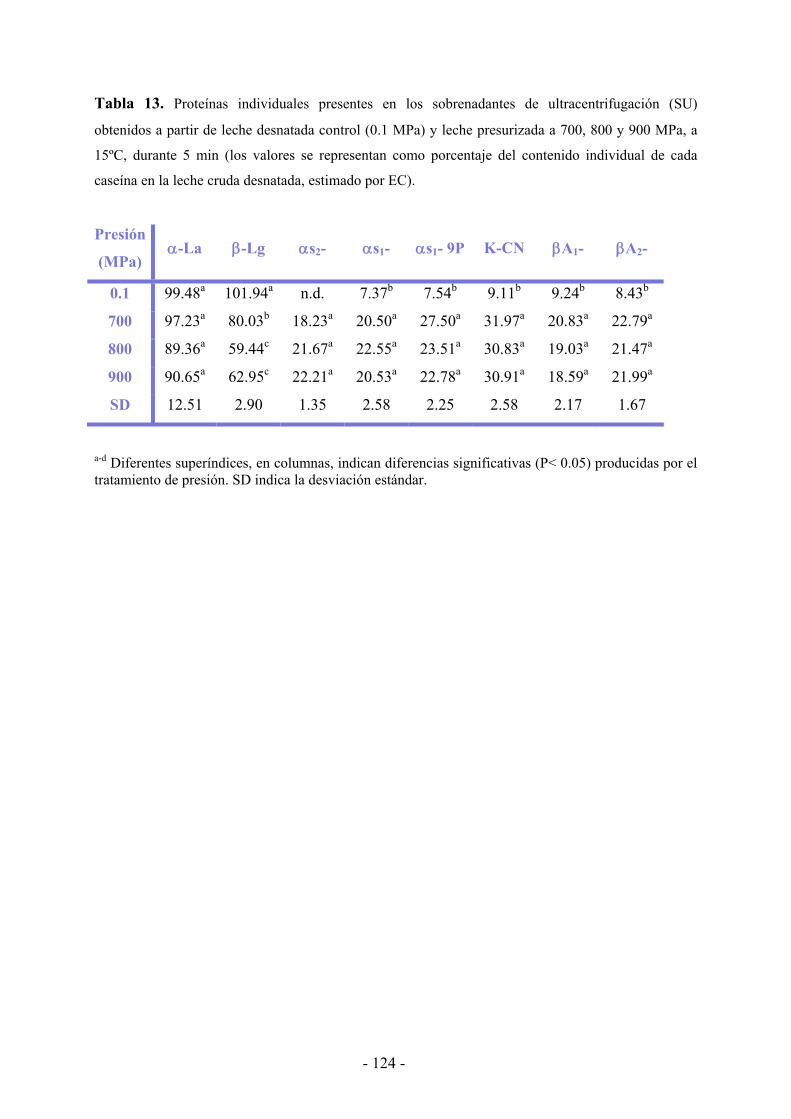
- 124 -
Tabla 13. Proteínas individuales presentes en los sobrenadantes de ultracentrifugación (SU)
obtenidos a partir de leche desnatada control (0.1 MPa) y leche presurizada a 700, 800 y 900 MPa, a
15ºC, durante 5 min (los valores se representan como porcentaje del contenido individual de cada
caseína en la leche cruda desnatada, estimado por EC).
Presión
(MPa) α-La β-Lg αs2- αs1- αs1- 9P Κ-CN βA1- βA2-
0.1 99.48a 101.94a n.d. 7.37b 7.54b 9.11b 9.24b 8.43b
700 97.23a 80.03b 18.23a 20.50a 27.50a 31.97a 20.83a 22.79a
800 89.36a 59.44c 21.67a 22.55a 23.51a 30.83a 19.03a 21.47a
900 90.65a 62.95c 22.21a 20.53a 22.78a 30.91a 18.59a 21.99a
SD 12.51 2.90 1.35 2.58 2.25 2.58 2.17 1.67
a-d Diferentes superíndices, en columnas, indican diferencias significativas (P< 0.05) producidas por el tratamiento de presión. SD indica la desviación estándar.

RESULTADOS Y DISCUSIÓN: CAPÍTULO IV
- 125 -
Figura 42. Electroforegramas de leche desnatada control (a) y sobrenadantes de ultracentrifugación
de: leche desnatada control (b) y leche presurizada a 700, 800 y 900 MPa y 25ºC (c-e). 1: α-La; 2:
β-Lg; 3: αs2-CN; 4: αs1-CN; 5: αs1-CN 9P; 6: κ-CN; 7: βB-CN; 8: βA1-CN; 9: βA2-CN.
a
4
1
23
5
98
7
6
Tiempo de migración (min)
A 21
4 nm
0
0.04
0
0.04
0
0.04
10 15 20 25 30 35 40
d
e
0.08
0.12
0.14
0
0.04
b
c
0
0.04
a
4
1
23
5
98
7
6
a
4
1
23
5
98
7
6
Tiempo de migración (min)
A 21
4 nm
0
0.04
0
0.04
0
0.04
10 15 20 25 30 35 40
d
e
0.08
0.12
0.14
0
0.04
b
c
0
0.04
Tiempo de migración (min)
A 21
4 nm
0
0.04
0
0.04
0
0.04
10 15 20 25 30 35 40
d
e
0.08
0.12
0.14
0
0.04
bbb
c
0
0.04
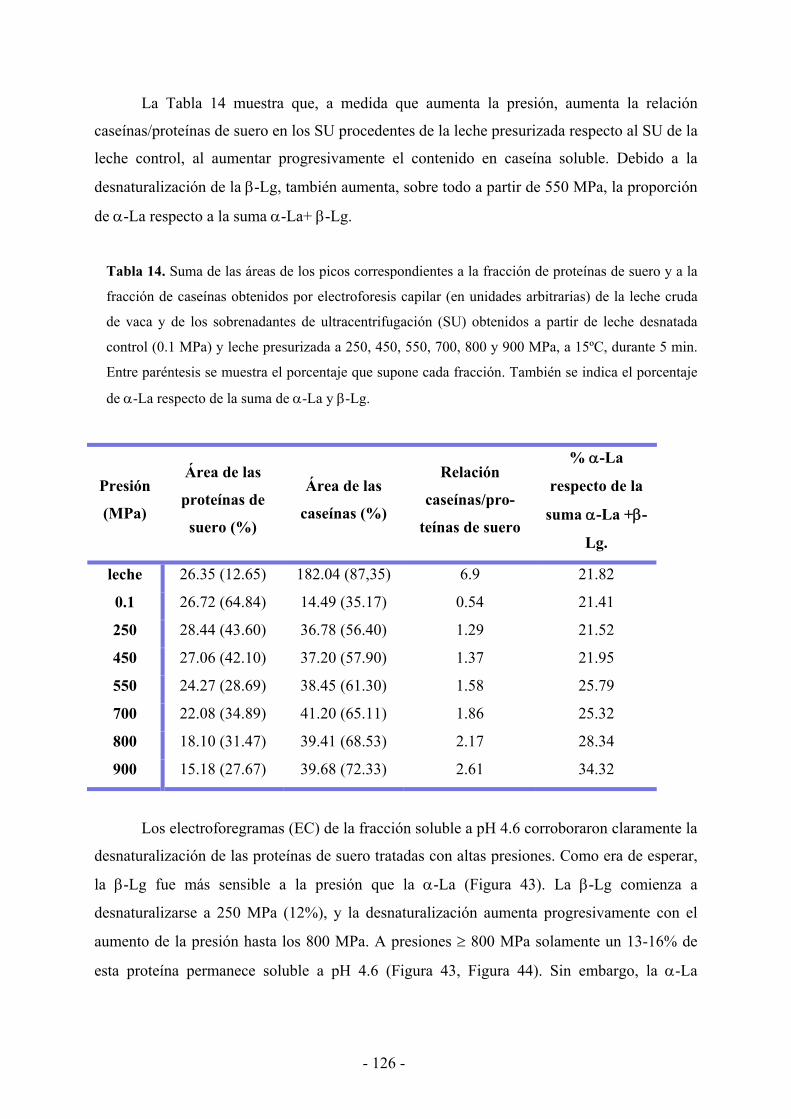
- 126 -
La Tabla 14 muestra que, a medida que aumenta la presión, aumenta la relación
caseínas/proteínas de suero en los SU procedentes de la leche presurizada respecto al SU de la
leche control, al aumentar progresivamente el contenido en caseína soluble. Debido a la
desnaturalización de la β-Lg, también aumenta, sobre todo a partir de 550 MPa, la proporción
de α-La respecto a la suma α-La+ β-Lg.
Tabla 14. Suma de las áreas de los picos correspondientes a la fracción de proteínas de suero y a la
fracción de caseínas obtenidos por electroforesis capilar (en unidades arbitrarias) de la leche cruda
de vaca y de los sobrenadantes de ultracentrifugación (SU) obtenidos a partir de leche desnatada
control (0.1 MPa) y leche presurizada a 250, 450, 550, 700, 800 y 900 MPa, a 15ºC, durante 5 min.
Entre paréntesis se muestra el porcentaje que supone cada fracción. También se indica el porcentaje
de α-La respecto de la suma de α-La y β-Lg.
Presión
(MPa)
Área de las
proteínas de
suero (%)
Área de las
caseínas (%)
Relación
caseínas/pro-
teínas de suero
% α-La
respecto de la
suma α-La +β-
Lg.
leche 26.35 (12.65) 182.04 (87,35) 6.9 21.82
0.1 26.72 (64.84) 14.49 (35.17) 0.54 21.41
250 28.44 (43.60) 36.78 (56.40) 1.29 21.52
450 27.06 (42.10) 37.20 (57.90) 1.37 21.95
550 24.27 (28.69) 38.45 (61.30) 1.58 25.79
700 22.08 (34.89) 41.20 (65.11) 1.86 25.32
800 18.10 (31.47) 39.41 (68.53) 2.17 28.34
900 15.18 (27.67) 39.68 (72.33) 2.61 34.32
Los electroforegramas (EC) de la fracción soluble a pH 4.6 corroboraron claramente la
desnaturalización de las proteínas de suero tratadas con altas presiones. Como era de esperar,
la β-Lg fue más sensible a la presión que la α-La (Figura 43). La β-Lg comienza a
desnaturalizarse a 250 MPa (12%), y la desnaturalización aumenta progresivamente con el
aumento de la presión hasta los 800 MPa. A presiones ≥ 800 MPa solamente un 13-16% de
esta proteína permanece soluble a pH 4.6 (Figura 43, Figura 44). Sin embargo, la α-La

RESULTADOS Y DISCUSIÓN: CAPÍTULO IV
- 127 -
permanece prácticamente invariable hasta los 550 MPa, alcanzando una desnaturalización del
42% a 900 MPa (Figura 43).
Figura 43. Contenido en α- La ( ) y β-Lg ( ) de la fracción soluble a pH 4.6, medido por EC,
de leche control (0.1 MPa) y presurizada a 700, 800 y 900 MPa. Las barras representan la desviación
estándar. Las letras en minúsculas con el mismo color indican diferencias significativas (P< 0.05)
producidas por el nivel de presión.
0
20
40
60
80
100
120
0.1 700 800 900
Presión (MPa)
Prot
eína
s de
suer
oso
lubl
es (%
)a
b bcc
a
b c c
0
20
40
60
80
100
120
0.1 700 800 900
Presión (MPa)
Prot
eína
s de
suer
oso
lubl
es (%
)a
b bcc
a
b c c

- 128 -
Figura 44. Electroforegramas de la fracción soluble a pH 4.6 de leche desnatada control (a) y de
leche presurizada a 250, 450, 550, 700, 800 y 900 MPa, 15ºC durante 5 min (b-g).
10 15 20 25
A 21
4 nm
Tiempo de migración (min)
b
c
d
e
f
aα-La
β-Lg
g
0
0.02
0.04
0.06
0.08
0
0.02
0.04
0.02
0.04
0
0.02
0.02
0.04
0.04
0
0.04
0
0.02
0
0.04
0.02
010 15 20 25
A 21
4 nm
Tiempo de migración (min)
b
c
d
e
f
aα-La
β-Lg
g
0
0.02
0.04
0.06
0.08
0
0.02
0.04
0.02
0.04
0
0.02
0.02
0.04
0.04
0
0.04
0
0.02
0
0.04
0.02
0
0
0.02
0.04
0.06
0.08
0
0.02
0.04
0.02
0.04
0
0.02
0.02
0.04
0.04
0
0.04
0
0.02
0
0.04
0.02
0

RESULTADOS Y DISCUSIÓN: CAPÍTULO IV
- 129 -
En este estudio también se evaluó el efecto de las altas presiones (250-900 MPa) sobre
las proteínas minoritarias del suero de interés nutricional, en concreto Lf e IgA. Los
contenidos en Lf de la leche presurizada hasta 900 MPa y de sus fracciones se muestran en la
Figura 45. La leche desnatada control presentó un contenido de 0.14 mg/mL que fue
disminuyendo por efecto de la presión hasta los 900 MPa, hasta alcanzar 0.0035 mg/mL. En
general, las fracciones solubles a pH 4.6 presentaron el mismo contenido en Lf que la leche de
la que procedían, mientras que, en los sobrenadantes de ultracentrifugación, el nivel de Lf
observado fue significativamente inferior a todas las presiones estudiadas. En cualquier caso,
todas las fracciones presentaron el mismo comportamiento frente a la presión: a partir de 550
hasta 800 MPa, el contenido de Lf se redujo significativamente (P< 0.05). A 900 MPa
también disminuyó, pero las diferencias no fueron significativas.
Figura 45. Contenido en lactoferrina de: leche desnatada control (0.1 MPa) y presurizada de 250 a
900 MPa ( ), los sobrenadantes de ultracentrifugación ( ) y las fracciones solubles a pH 4.6 ( ).
Las barras representan la desviación estándar. Las letras en minúscula indican diferencias
significativas (P< 0.05) producidas por el nivel de presión en cada fracción (indicada con el mismo
color). * indica diferencias significativas (P< 0.05) de los sobrenadantes de ultracentrifugación
respecto a las otras 2 muestras a la presión indicada y * indica diferencias significativas entre la
leche y la fracción soluble a pH 4.6 a la presión indicada.
El contenido de IgA en la leche control (0.24 mg/mL) fue similar en las tres muestras
estudiadas (Figura 46). Al aplicar alta presión, este contenido disminuyó progresivamente
0
0.05
0.1
0.15
0.2
0.1 250 450 550 700 800 900
Tratamiento de presión (MPa)
Lf(
mg/
mL
)
ba*
c
d
ef* g*
aabb
cd
e e
a*b* b*
c* d*e* e*
0
0.05
0.1
0.15
0.2
0.1 250 450 550 700 800 900
Tratamiento de presión (MPa)
Lf(
mg/
mL
)
ba*
c
d
ef* g*
aabb
cd
e e
a*b* b*
c* d*e* e*

- 130 -
hasta los 700 MPa, tanto en la leche como en los sobrenadantes de ultracentrifugación y las
fracciones solubles a pH 4.6, llegando a detectarse solamente 0.0079 mg/mL a 700 MPa. A
presiones superiores también se observaron reducciones de su contenido, pero las diferencias
no fueron significativas (se detectaron 0.0035 y 0.00011 mg/mL de IgA en leche presurizada
a 800 y 900 MPa, respectivamente).
Figura 46. Contenido en inmunoglobulina A de leche desnatada control (0.1 MPa) y presurizada de
250 a 900 MPa ( ), de los sobrenadantes de ultracentrifugación ( ) y de las fracciones solubles a pH
4.6 ( ). Las barras representan la desviación estándar. Las letras en el mismo color indican
diferencias significativas (P< 0.05) producidas por el nivel de presión en cada fracción. * indica
diferencias significativas (P< 0.05) de los sobrenadantes de ultracentrifugación respecto a las otras
dos muestras a la presión indicada y * indica diferencias significativas entre la leche y la fracción
soluble a pH 4.6 a la presión indicada.
IV.4. Discusión
Los resultados obtenidos indican que el tratamiento de leche a presiones muy
elevadas, en torno a 700-900 MPa, no disminuyó el tamaño micelar ni indujo mayor
solubilización de caseínas respecto a tratamientos a presiones inferiores de 450-550 MPa. Sin
embargo, estos tratamientos produjeron una severa desnaturalización de las proteínas del
suero, sobre todo de la β-Lg y de las proteínas minoritarias, Lf e IgA.
0
0.1
0.2
0.3
0.1 250 450 550 700 800 900
Tratamiento de presión (MPa)
IgA
(mg/
mL
)
ab*
c
d*
e* e* e
b
aa
c
d d d
a a
b
c*
d* d* d
0
0.1
0.2
0.3
0.1 250 450 550 700 800 900
Tratamiento de presión (MPa)
IgA
(mg/
mL
)
ab*
c
d*
e* e* e
b
aa
c
d d d
a a
b
c*
d* d* d

RESULTADOS Y DISCUSIÓN: CAPÍTULO IV
- 131 -
Hasta el momento no existían estudios sobre el efecto de presiones muy elevadas
(700-900 MPa) en las propiedades de las micelas de caseína o en la distribución proteica de la
leche. En general, y como ya se ha mencionado anteriormente, el tamaño micelar medio de la
leche tratada a 100-200 MPa a temperatura ambiente se considera comparable al de la leche
sin tratar; las micelas en la leche presurizada a 250 MPa durante periodos superiores a 15 min
presentan mayor tamaño; mientras que en la leche presurizada a más de 300 MPa se han
observado reducciones de tamaño de hasta el 50% (Huppertz y col., 2004a; Anema y col.,
2005a) y se mantienen así durante el posterior almacenamiento de la leche presurizada
(Huppertz y col., 2004a).
La observación de la micela de caseína por microscopía electrónica de transmisión nos
permitió evaluar el efecto de la presión en el tamaño micelar (Figura 39). Los tratamientos
con alta presión redujeron el diámetro medio de la micela de caseína hasta los 450 MPa,
presión a la cual se alcanzó un plateau. García-Risco y col. (2000) observaron, con esta
misma técnica, que a 300 y 400 MPa, (20ºC, 15 min) disminuía el diámetro de la micela de
caseína de 150-300 nm, en la leche sin presurizar, a 40 nm en la presurizada y que la
distribución de tamaño se hacía más homogénea. Por lo general, se considera que, en torno a
los 300 MPa en tratamientos de 15 min a temperatura ambiente, se produce la máxima
disgregación (Scollard y col., 2000; García-Risco y col., 2003; Anema, 2008a) no observando
modificaciones adicionales en el tamaño hasta los 800 MPa. En nuestro estudio el menor
tamaño se encontró a 450 MPa, aunque no se pudo comprobar si, efectivamente, era similar a
300 MPa como afirman otros autores (Figura 40, Tabla 11). De acuerdo con García-Risco y
col (2003), a 250 MPa se observaron micelas de gran tamaño, similares a las presentes en la
leche sin presurizar (en torno a 200 nm), sin embargo, a presiones ≥ 450 MPa todas las
micelas fueron inferiores a 115 nm, corroborando las observaciones hechas por Needs y col.
(2000b) a 400 y 600 MPa, y un 50 % de las micelas estuvieron comprendidas entre 20 y 60
nm.
La disgregación de las micelas de caseína se debe a la solubilización del CCP y a la
ruptura de las interacciones hidrofóbicas y electroestáticas intramicelares (Schrader y col.,
1997). De hecho, medidas in situ, han demostrado que la solubilización del CCP es
proporcional a la presión aplicada y es completa a 400 MPa, presión a la que la disrupción

- 132 -
micelar también es completa e irreversible (Huppertz y De Kruif, 2007a). Tal disgregación es
responsable de que, además de disminuir el tamaño micelar, aumente el número de micelas de
caseína y el contenido en proteína no sedimentable por ultracentrifugación, como ya se
explicó en los capítulos II y III. En este estudio, los cambios observados en el tamaño micelar
fueron concordantes con los niveles de caseína soluble en los sobrenadantes de
ultracentrifugación a presiones de 700-900, puesto que no se detectó un incremento
significativo en el contenido de caseínas no sedimentables respecto a los observados a
menores presiones (250-550 MPa) (Tablas 12, 13 y Figura 42). Así, Anema y col. (2005a)
indicaron que el nivel de caseínas no sedimentables está inversamente correlacionado con el
tamaño de la micela de caseína, excepto para las muestras agregadas a 250 y 300 MPa.
Los tratamientos a presiones muy elevadas produjeron cambios en las proteínas de
suero solubles a pH 4.6 (Figura 41, 43, 44). Se observó una severa desnaturalización de la β-
Lg a presiones ≥ 700 MPa, en torno al 77%. Garcia-Risco y col. (2000) ya habían observado
este grado de desnaturalización a presiones inferiores (400 MPa, 30 min) probablemente
debido a la mayor duración del tratamiento. Esta proteína comienza a desnaturalizarse a 100
MPa, aunque la desnaturalización depende del nivel de presión, de la temperatura (López-
Fandiño y col., 1996; Huppertz y col., 2002; Anema y col., 2005b) y, como se muestra en el
capítulo anterior, de la duración del tratamiento A partir de 200 MPa durante 30 min, Anema
(2008a) observó, aproximadamente, un 30% de desnaturalización de la β-Lg, que se fue
incrementando con el aumento de la presión, llegando al 90% a 800 MPa.
La α-La se desnaturaliza por alta presión notablemente menos que la β-Lg (Huppertz
y col., 2004b); Patel y col., 2006), debido al mayor número de puentes disulfuro que presenta
la molécula (4 frente a 2 de la β-Lg) y a la ausencia de grupos sulfidrilo libres que pudieran
interaccionar con otras proteínas (Hinrichs y col., 1996). Esta proteína permanece estable
hasta los 400-500 MPa (Anema y col., 2005b; Hinrichs y Rademacher, 2005), sin embargo, a
mayores presiones se hace patente la desnaturalización de la α-La (Figura 41, 43, 44), como
ya se indicó en el Capítulo II. Tras tratamientos a 800MPa, a 20ºC durante 30 min, se han
observado desnaturalizaciones de, aproximadamente el 35% (Huppertz y col., 2004b, Anema
y col. (2008a), similares a las obtenidas en nuestro estudio.

RESULTADOS Y DISCUSIÓN: CAPÍTULO IV
- 133 -
La solubilización de caseínas producida por la alta presión, junto con el hecho de que
la desnaturalización de la β-Lg se produzca en mayor medida que la de la α-La, hace posible
seleccionar las condiciones que proporcionan sobrenadantes de ultracentrifugación de leche
presurizada de vaca que simulen la composición de la leche de mujer. Como se muestra en la
Tabla 14, presiones en torno a 450 o 550 MPa proporcionan, en los sobrenadantes de
ultracentrifugación, relaciones de caseínas a proteínas de suero de, aproximadamente, 1.4-1.6,
que se asemejan a la relación recomendada en las fórmulas infantiles de 1.5 (60:40
caseínas:seroproteínas). La relación en la leche de partida fue de 6.9, superior a la descrita
normalmente para una leche de vaca (4), posiblemente porque, con nuestro método, sólo
cuantificamos β-Lg y α-La, con lo que no se tiene en cuenta la contribución de otras proteínas
minoritarias del suero como la BSA, etc. Cabría esperar, entonces, que las relaciones fueran
ligeramente superiores a las que se muestran en la Tabla 14. Por otra parte, el contenido en
proteína soluble de los sobrenadantes de ultracentrifugación de las muestras empleadas en
este estudio (comprendido entre el 1.05 y el 1.29 %. Tabla 12) también resultó similar al de la
leche humana (aproximadamente 0.94% de media, Manso et al. 2007). Finalmente, debe
tenerse en cuenta que la leche humana no contiene β-Lg, por lo que la α-La supone
aproximadamente el 30% del total de proteínas. Presiones ≥ 700 MPa desnaturalizan más β-
Lg y aumentan la proporción de α-La dentro del total de las proteínas de suero (Tabla x.3),
pero también incrementan el contenido en caseína soluble sin reducir considerablemente el de
β-Lg. Además, presiones en torno a 450 o 550 MPa mantendrían en mayor medida los niveles
de proteínas minoritarias, Lf e IgA, nativas, respecto a presiones más elevadas.
Los efectos de la alta presión en estas proteínas minoritarias han sido bastante
ignorados. El contenido de Lf en la leche bovina puede variar entre 0.02 y 0.2 mg/mL según
el periodo de lactación (Steijns y Van Hooijdonk, 2000). El contenido en Lf determinado en
la leche control antes del fraccionamiento fue de 0.14 mg/mL y disminuye al aumentar la
presión hasta llegar a 0,00011 mg/mL a 900 MPa (Figura 45), debido probablemente a la
desnaturalización de la Lf. Los niveles de Lf encontrados en la fracción soluble a pH 4.6
fueron similares a los de la leche antes de fraccionar (0.14 a 0.0026 mg/mL), pues la
recuperación de Lf en suero de leche cruda sin calentar es prácticamente completa y sólo se
reduce a medida que se incrementa la desnaturalización (Brisson y col., 2007). Sin embargo,
el contenido fue considerablemente menor en el sobrenadante obtenido por
ultracentrifugación (0.09 a 0.0015), probablemente debido a que una parte de la Lf

- 134 -
desnaturalizada pueda estar unida a fragmentos caseínicos no sedimentables (Figura 45). La
Lf establece interacciones intermoleculares vía puentes disulfuro con las caseínas y otras
proteínas séricas durante el calentamiento de la leche (Brisson y col., 2007) por lo que durante
los tratamientos de alta presión también podrían formarse estas uniones.
Nuestros resultados indican que la IgA se desnaturaliza a presiones comprendidas
entre 250-450 MPa (Figura 46) y su contenido va disminuyendo a medida que se incrementa
el nivel de presión, hasta niveles muy bajos a presiones ≥ 700 MPa. En leche humana,
también se ha observado la desnaturalización de la IgA a presiones ≥ 400 MPa, aumentando
con la aplicación de presiones crecientes durante 5 min. Sin embargo, Viazis y col. (2007)
observaron que la inmunoreactividad y la concentración de IgA en leche humana presurizada
(400 MPa durante 30, 60, 90 y 120 min) fue significativamente mayor (P< 0.05) que en la
misma leche pasteurizada.
IV.5. Conclusiones
El tratamiento de leche a presiones muy elevadas, en torno a 700-900 MPa, no
disminuyó el tamaño micelar ni indujo mayor solubilización de caseínas respecto a
tratamientos a presiones inferiores de 450-550 MPa. Sin embargo, estos tratamientos
produjeron una severa desnaturalización de las proteínas del suero, sobre todo de la β-Lg y de
las proteínas minoritarias, Lf e IgA, aunque también condujeron a una apreciable
desnaturalización de la α-La. Por ello, el mejor compromiso para producir fracciones
proteicas con una concentración de proteína y una relación de caseínas a proteínas de suero
similar a la de la leche humana, seria utilizar presiones de 450-550 MPa aplicadas durante 5
min. Estos tratamientos proporcionan, sobrenadantes de ultracentrifugación, con contenidos
en proteína entre el 1.05 y el 1.29 % y relaciones de caseínas a proteínas de suero de,
aproximadamente, 1.4-1.6, aunque contienen una apreciable proporción de β-Lg
desnaturalizada soluble.

5. CONCLUSIONES


CONCLUSIONES
- 137 -
1. Los tratamientos con alta presión a 650 MPa durante 5 min, aplicados a fórmulas infantiles,
no producen isomerización de la lactosa, ni avance de la reacción de Maillard y, comparados
con tratamientos UHT, conducen a una menor desnaturalización de las proteínas mayoritarias
y minoritarias (lactoferrina e inmunoglobulina A secretora) del suero, por lo que mejoran la
calidad nutritiva de las fórmulas.
2. La aplicación de tratamientos con alta presión en distintos equipos y a diferentes escalas,
industrial y piloto, da lugar a diferentes distribuciones proteicas, sobre todo en lo que se
refiere a la desintegración micelar. Los diferentes equipos proporcionan grados de
desnaturalización de las proteínas de suero más homogéneos, a niveles de presión
equivalentes.
3. La velocidad de despresurización juega un papel fundamental en la distribución de
proteínas entre las fases soluble y coloidal de la leche presurizada, puesto que el contenido en
caseína soluble aumenta significativamente durante la fase de despresurización respecto a la
fase de mantenimiento de la presión. Cuanto más lenta es la velocidad de despresurización,
mayor es el nivel de caseína soluble.
4. La desnaturalización de la β-Lg tiene lugar, mediante enlaces disulfuro, fundamentalmente
durante la fase de mantenimiento de la presión, produciéndose muy poca desnaturalización
durante la fase de despresurización. La mayoría de las proteínas de suero desnaturalizadas,
unidas o no a las partículas de caseína no sedimentables, permanecen solubles al pH normal
de la leche.
5. La presencia de caseína promueve la desnaturalización de las proteínas del suero, pero el
grado de desnaturalización de las proteínas de suero no influye en la ruptura micelar y en la
subsiguiente solubilización de caseínas que tiene lugar en la leche presurizada.
6. El tratamiento de leche a presiones muy elevadas, en torno a 700-900 MPa, no disminuye
el tamaño micelar ni induce mayor solubilización de caseínas respecto a tratamientos a
presiones inferiores de 450-550 MPa. Sin embargo, estos tratamientos producen una severa
desnaturalización de las proteínas del suero, sobre todo de la β-Lg y de las proteínas
minoritarias, Lf e IgA.

- 138 -
7. Presiones de 450-550 MPa aplicadas durante 5 min permiten producir fracciones proteicas
con una concentración de proteína entre el 1.05 y el 1.29 % y una relación de caseínas a
proteínas de suero de, aproximadamente, 1.4-1.6, similar a la de la leche humana, aunque
contienen una apreciable proporción de β-Lg desnaturalizada soluble.

6. BIBLIOGRAFÍA


BIBLIOGRAFÍA
- 141 -
Alexandrescu, A.T., Evans, P.A., Pitkeathly, M., Baum, J. y Dobson, C.M. (1993). Structure
and dynamics of the acid-denatured molten globule state of α-lactalbumin: A two-
dimensional NMR study. Biochemistry (Easton) 32, 1707-1718.
Ames, J. M. (1990). Control of the Maillard reaction in food systems. Trends in Food Science
and Technology 1, 150-154.
Anema, S.G. (2008a). Heat and/or high-pressure treatment of skim milk: changes to the
casein micelle size, whey proteins and the acid gelation properties of the milk. International
Journal of Dairy Technology 61, 245-252.
Anema, S.G. (2008b). Effect of milk solids concentration on whey protein denaturation,
particle size changes and solubilization of casein in high-pressure treated skim milk.
International Dairy Journal 18, 228-235.
Anema, S.G., Lee, S.K., Schrader, K. y Buchheim, W. (1997). Effect of pH on the turbity of
pressure-treated calcium caseinate suspensions and skim milk. Milchwissenschaft 52, 141-
146.
Anema, S.G., Lowe, E.K. y Stockman, G. (2005a). Particle size changes and casein
solubilisation in high-pressure-treated skim milk. Food Hydrocolloids 19, 257-267.
Anema, S.G., Stockmann, R. y Lowe, E. K. (2005b). Denaturation of β-Lactoglobulin in
pressure-treated skim milk. Journal of Agricultural and Food Chemistry 53, 7783-7791.
Arias, M., López-Fandiño, R. y Olano, A. (2000). Influence of pH on the effects of high
pressure on milk proteins. Milchwissenschaft 55, 191-194.
Bakker-Zierikzee, A.M., Van Tol, E.A.F, Kroes, H., Alles, M.S., Kok, F.J. y Bindels, J.G.
(2006). Faecal sIgA secretion in infants fed on pre- or probiotic infant formula. Pediatric
Allergy and Immunology 17, 134-140.
Bakker-Zierikzee, A.M., van Tol, E.A.F., Kroes, H., Alles, M.S., Kok, F.J. y Bindels, J.G.
(2006). Faecal sIgA secretion in infants fed on pre- or probioic infant formula. Pediatric
Allergy and Immunology 17, 134-140.
Balasubramaniam, V.M., Farkas, D. y Turek, E. (2008). Preserving foods through high-
pressure processing. Food Technology 62, 32-38.

- 142 -
Balasubramaniam, V.M., Ting, E.Y., Steward, C.M. y Robbins, J.A. (2004). Recommended
laboratory practices for conducting high pressure microbial inactivation experiments.
Innovative Food Science and Emerging Technologies 5, 299-306.
Balasubramanian, S. y Balasubramaniam, V.M. (2003). Compression heating influence of
pressure transmitting fluid son bacteria inactivation during HP processing. Food Research
International 36, 661-668.
Balmer, S.E. y Wharton, B.A. (1989). Diet and faecal flora in the newborn: breast milk and
infant formula. Archives of Disease in Childhood 64, 1672-1677.
Banon, S. y Hardy, J. (1992). A colloidal approach of milk acidification by glucono-delta-
lactone. Journal of Dairy Science 75, 935-94.
Barbosa-Cánovas, G. y Bermúdez-Aguirre, D. (2011). Introduction. En: Nonthermal
processing technologies for food, Zhang, H. Q., Barbosa-Cánovas, G. V., Balasubramaniam,
V. M., Dunne, C. P., Farkas, D. F. y Yuan, T. C. (Eds.). Wiley-Blackwell. Iowa, USA, pp.
XX-XXX.
Barbosa-Cánovas, G.V. y Bermúdez-Aguirre, D. (2010). Novel food processing
technologies and regulatory hurdles. En: Ensuring global Food Safety: Exploring Global
Harmonization. Boisrobert, C., Stjepanovic, A., Oh, S. y Lelieveld, H. (Eds.). Elservier Inc.
London, UK, pp. 281-288.
Barth, C.A. y Schlimme, E. (1988). Milk proteins: nutritional, clinical, functional and
technological aspects. Springer-Verlag, Nueva York.
Belloque, J., López-Fandiño, R. y Smith, G.M. (2000). A 1H-NMR study on the effect of
high pressures on beta-lactoglobulin. Journal of Agricultural and Food Chemistry 48, 3906-
3912.
Bermúdez-Aguirre, D. y Barbosa-Cánovas, G. V. (2011). An update on high hydrostatic
pressure, from the laboratory to industrial applications. Food Engineering Reviews 3, 44-61.
Bertrand-Harb, C., Baday, A., Dalgalarrondo, M., Chobert, J.M. y Haertlé, T. (2002).
Thermal modifications of structure and co-denaturation of α-lactalbumin and β-lactoglobulin
induce changes of solubility and susceptibility to proteases. Nahrung-Food 46, 283-289.

BIBLIOGRAFÍA
- 143 -
Bosch, L., Alegría, A., Farré, R. y Clemante, G. (2000). Effect of storage conditions on
furosine formation in milk-cereal based baby foods. Food Chemistry 107, 1681-1686.
Bravo, F.I., Villamiel, M. y Molina, E. (2007). Emulsifying properties of α-lactalbumin after
high-pressure treatment and subsequent lactosylation. High Pressure Research 27, 115-119.
Brisson, G., Britten, M. y Pouliot, Y. (2007). Effect of iron saturation on the recovery of
lactoferrin in rennet whey coming from heat-treated skim milk. Journal of Dairy Science 90,
2655-2664.
Buzrul, S., Alpas, H., Largeteau, A., Bozoglu, F. y Demazeau, G. (2008). Compression
heating of selected pressure transmitting fluids and liquids foods during high hydrostatic
pressure treatment. Journal of Food Engineering 85, 466-472.
Caessens, P.W.J.R., Visser, S. y Gruppen, H. (1997). Method for the isolation of bovine β-
lactoglobulin from a cheese whey protein fraction and physicochemical characterization of the
purified product. International Dairy Journal 7, 229-235.
Capellas, M. y Needs, E. (2003). Physical properties of yogurt prepared from pressure-
treated concentrated or fortified milks. Milchwissenschaft 58, 46–48.
Cayot, P. y Lorient, D. (1997). Structure-function relationships of whey proteins. En: Food
Proteins and their Applications. Damodaran, S. y Paraf, A. (Eds.). Marcel Dekker, Inc., pp.
225-256.
Cheftel, J.C. (1991). Application des hautes pressions en technologie alimentaire. Actualités
des Industries Alimentaires et Agro-Alimentaire 108, 141-153.
Chierici, R. y Vigi, V. (1994). Lactoferrin in infant formula. Acta Pediatrica 83, 83-88.
Considine, T., Patel, H.A., Anema, S.G., Singh, H. y Creamer, L.K. (2007). Interactions of
milk proteins during heat and high hydrostatic pressure treatments- a review. Innovative Food
Science and Emerging Technologies 8, 1-23.
Corzo, N., Delgado, T., Troyano, E. y Olano A. (1994). Ratio of lactulose to furosine as
indicador of quality of commercial milks. Journal of Food Protection 57, 737-739.

- 144 -
Cullen, P.J., Tiwari, B.K. y Valdramidis, V.P. Status and trends of novel termal and non-
thremal technologies for fluids foods. En: Novel Thermal and Non-Thermal Technologies for
Fluid Foods. Cullen, P. J., Tiwari, B. K. y Valdramidis, V. P. (Eds.). Elservier, Inc. London,
UK, pp. 1-6
Daoudi, L. (2004). Efecto de las altas presiones hidrostáticas sobre el gazpacho y zumo de
uva. Universitat Autònoma de Barcelona. Tesis doctoral.
De Kruif, C.G. (2003). Caseins. En: Progress in Biotechnology, volume 23. W.Y.
Aalbersberg, W. Y., Hamer, R. J., Jasperse, P., De Jongh, H. H. J., De Kruif, C. G., Walstra,
P. y De Wolf, F. A. (Eds). Elservier Science B.V, London, UK, pp. 219-269.
Desobry-Banon, S., Richard, F. y Hardy, J. (1994). Study of acid and rennet coagulation of
high pressurized milk. Journal of Dairy Science 77, 3267–3274.
Drake, M.A., Harrison, S.L., Asplund, M., Barbosa-Cánovas, G. y Swanson, B.G. (1997).
High pressure treatment of milk and effects on microbiological and sensory quality of
Cheddar cheese. Journal of Food Science 62, 843–860.
Dupont, D., Boutrou, R., Menard, O., Jardin, J., Tanguy, G., Schuck, P., Haab, B.B. y Leonil,
J. (2010). Heat treatment of milk during powder manufacture increases casein resistance to
simulated infant digestion. Food Digestion 1, 28-39.
Erbersdobler, H.F. y Hupe, A. (1991). Determination of lysine damage and calculation of
lysine bioavailability in several processed foods. Zeitschrift fur Ernahrungswissenschaft 30,
46-49.
Famelart, M.H., Chapron, L., Piot, M., Brulé , G. y Durier, C. (1998). High pressure-induced
gel formation of milk and whey concentrates. Journal of Food Engineering 36, 149–164.
Ferragut, V., Martínez, V.M., Trujillo, A.J. y Guamis, B. (2000). Properties of yogurts made
from whole ewe’s milk treated by high hydrostatic pressure. Milchwissenschaft 55, 267–269.
Ferrandini, E., Castillo, M., López, M.B. y Laencina, J. (2006). Modelos estructurales de la
micela de caseína. Anual. Veterinaria 22, 5-18.

BIBLIOGRAFÍA
- 145 -
Ferrer, E., Alegría, A., Farré, R., Abellán, P., Romero, F. y Clemente, G. (2003). Evolution
of available lysine and furosine contents in milk-based infant formulas throughout the shelf-
life storage period. Journal of the Science of Food and Agriculture 83, 465-472.
Fertsch, B., Müller, M. y Hinrichs, J. (2003). Firmeness of pressure-induced casein and whey
protein gels modulated by holding time and rate of pressure release. Innovative Food Science
and Emerging Technologies 4, 143-150.
Floris, R., Lambers, T., Alting, A. y Kiers, J. (2007). Trends in infant formulas: a dairy
perspective. En: Improving the safety and quality of milk. Volume 2: improving quality in
milk products. Griffiths, M.W. (ed). Woodhead, Cambridge, pp 454-474.
Ford, J.E., Law, B.A., Marshall, V.M.E. y Reiter, B. (1977). Influence of heat treatment of
human milk on some of its protective constituents. Journal of Pediatrics 90, 29-35.
Fox, P.F. y Brodkorb, A. (2008). The casein micelle: Historical aspects, current concepts
and significance. International Dairy Journal 18, 677-684.
Fox, P.F. y Kelly, A.L. (2004). The caseins. En: Proteins in Food Processing. Yada, R.Y.
(Eds.). Woodhead Publishing Limited. Cambridge, pp. 29-71.
García-Risco, M.R., Olano, A., Ramos, M. y López-Fandiño, R. (2000). Micellar changes
induced by high pressure. Influence in the proteolytic activity and organoleptic properties of
milk. Journal of Dairy Science 83, 2184-2189.
García-Risco, M.R., Recio, I., Molina, E. y López-Fandiño, R. (2003). Plasmin activity in
pressurized milk. Journal of Dairy Science 86, 728-734.
Gaucheron, F., Famelart, M.H., Mariette, F., Raulot, K., Michel, F. y Le Graet, Y. (1997).
Combined effects of temperature and high-pressure treatments on physicochemical
characteristics of skim milk. Food Chemistry 59, 439-447.
Geghardt, R., Doster, W., Friedrich, J. y Kulozik, U. (2006). Size distribution of pressure-
decomposed casein micelles studied by dynamic light scattering and AFM. European
Biophysics Journal 35, 503-509.
Ginger, M.R. y Grigor, M.R. (1999). Comparative aspects of milk caseins. Comparative
Biochemistry Physiology B-Biochemistry & Molecular 124, 133-145.

- 146 -
Grinberg, V.Y. y Heartlé, T. (2000). Reducer driven baric denaturation and oligomerisation
of whey proteins. The Journal of Biochemistry 79, 205-209.
Guerra-Hernández, E., León, C., Corzo, N., García-Villanova, B. y Romera, J.M. (2002).
Chemical changes in powdered infant formulas during storage. International Journal of Dairy
Technology 55, 171-176.
Hambling, S.G., Mc Alpine, A.S. y Sawyer, L. (1992). β-Lactoglobulin. En: Advanced Dairy
Chemistry, volume 1: Proteins. Fox, P.F. (Eds.), Elsevier Inc. London, pp. 141-190.
Harte, F., Amonte, M., Luedecke, L., Swanson, B.G. y Barbosa-Cánovas, G.V. (2002).
Yield-stress and microstructure of set yogurt made from high hydrostatic pressure-treated full
fat milk. Journal of Food Science 67, 2245–2250.
Heremans, K. (1984). High-pressure effects on proteins and other biomolecules. Annual
Review of Biophysics and Bioengineering 11, 1-21.
Hinrichs, J. y Rademacher, B. (2005). Kinetics of combined thermal and pressure-induced
whey protein denaturation in bovine skim milk. International Dairy Journal 14, 315-323.
Hinrichs, J. y Rademacher. B. (2004). High pressure thermal denaturation kinetics of whey
proteins. Journal of Dairy Research 71, 480-488.
Hite, B.H. (1899). The effects of pressure in the preservation of milk. West Virginia
Agricultural Experiment Station Bulletin 58, 15-35.
Hogan, E., Kelly, A.L. y Sun, D.W. (2005). High pressure processing of foods: an overview.
En: Emerging Technologies for Food Processing. Sun, D.W. (Eds.). Elservier Academic
Press, San Diego, pp. 3-32.
Holt, C. (1992). Structure and properties of bovine casein micelles. Advances in Protein
Chemistry 43, 63-151.
Holt, C. (1994). The biological function of casein. En: Yearbook 1994. The Hannah Institute.
Scottland, pp 60-68.
Horne, D.S. (1998). Casein interactions: Casting light on the black boxes, the structure in
dairy products. International Dairy Journal 8, 171-177.

BIBLIOGRAFÍA
- 147 -
Hubbard, C.D., Caswell, D., Ludemann, H.D. y Arnold, M. (2002). Characterisation of
pressure-treated skimmed milk powder dispersion: application of NMR spectroscopy. Journal
of the Science of Food and Agriculture 82, 1107-1114.
Huppertz, T. y De Kruif, C.G. (2006). Disruption and reassociation of casein micelles under
high pressure: influence of milk serum composition and casein micelle concentration. Journal
of Agricultural and Food Chemistry 54, 5903-5909.
Huppertz, T. y De Kruif, C.G. (2007a). High pressure-induced solubilisation of micellar
calcium phosphate from cross-linked casein micelles. Colloids and Surfaces A:
Physicochemical and Engineering Aspects 295, 264-268.
Huppertz, T. y De Kruif, C.G. (2007b). Disruption and reassociation of casein micelles
during high pressure treatment: influence of whey proteins. Journal of Dairy Research 74,
194-197.
Huppertz, T., Fox, P.F. y Kelly, A.L. (2004a). High pressure treatment of bovine milk:
effects on casein micelles and whey proteins. Journal of Dairy Research 71, 97-106.
Huppertz, T., Fox, P.F. y Kelly, A.L. (2004b). High pressure-induced denaturation of α-
lactalbumin and β-lactoglobulin in bovine milk and whey: a possible mechanism. Journal of
Dairy Research 71, 489-495.
Huppertz, T., Fox, P.F. y Kelly, A.L. (2004c). Dissociation of caseins in high pressure-
treated bovine milk. International Dairy Journal 14, 675-680.
Huppertz, T., Fox, P.F. y Kelly, A.L. (2004d). Properties of casein micelles in high pressure-
treated bovine milk. Food Chemistry 87, 103-110.
Huppertz, T., Fox, P.F. y Kelly, A.L. (2004f). Effects of high pressure treatment on the yield
of cheese curd from bovine milk. Innovative Food Science and Emerging Technologies 5, 1-8.
Huppertz, T., Fox, P.F. y Kelly, A.L. (2004g). Influence of high pressure treatment on the
acidification of bovine milk by lactic acid bacteria. Milchwissenschaft-Milk Science
International 59, 246-249.
Huppertz, T., Fox, P.F., De Kruif, K.G. y Kelly, A.L. (2006a). High pressure-induced
changes in bovine milk proteins: a review. Biochimica et Biophysica Acta 1764, 593-598.

- 148 -
Huppertz, T., Grosman, S., Fox, P.F. y Kelly, A.L. (2004e). Heat and ethanol stabilities of
high pressure-treated bovine milk. International Dairy Journal 14, 125-133.
Huppertz, T., Kelly, A.L. y De Kruif, C.G. (2006b). Disruption and reassociation of casein
micelles under high pressure. Journal of Dairy Research 73, 294-298.
Huppertz, T., Kelly, A.L. y Fox, P.F. (2002). Effects of high pressure on constituents and
properties of milk. International Dairy Journal 12, 561-572.
Huppertz, T., Smiddy, M.A., Upadhyay, V.K. y Kelly, A.L. (2006c). High-pressure-induced
changes in bovine milk: a review. International Journal of Dairy Technology 59, 58-66.
Imafidon, G.I. y Farkye, N.Y. (1996). Solvation of rennet curd from high-heat-treated milk
influenced by acid type and pH. Journal of Agricultural and Food Chemistry 44, 2560-2562.
Indyk, H.E. y Filonzi, E.L. (2005). Determination of lactoferrin in bovine milk, colostrum
and infant formulas by optical biosensor analysis. International Dairy Journal 15, 429-438.
International Dairy Federation (1992). Influence of technology on the quality of heat
treated milk and fluid milk products. B-Doc. 222, IDF, Bruselas, Bélgica.
Jenssen, H. y Hancock, R.E.W. (2009). Antimicrobial properties of lactoferrin. Biochimie
91, 19-29.
Johnston, D.E., Austin, B.A. y Murphy, R.J. (1992). Effects of high hydrostatic pressure on
milk. Milchwissenschaft 47, 760-763.
Johnston, D.E., Austin, B.A. y Murphy, R.J. (1993). Properties of acidset gels prepared from
high pressure treated milk. Milchwissenschaft 48, 206–209.
Johnston, D.E., Murphy, R.J., Rutherford, J.A. y McCreedy, R.W. (2002). Acidification of
high pressure treated milk: The role of whey protein denaturation. Milchwissenschaft 57, 605–
608.
Keenan, R.D., Young, D.J., Tier, C.M., Jones, A. D. y Underdown, J. (2001). Mechanism of
pressure-induced gelation of milk. Journal of Agricultural and Food Chemistry 49, 3394–
3402.

BIBLIOGRAFÍA
- 149 -
Kelly, A.L. y Zeece, M. (2009). Applications of novel technologies in processing of
functional foods. Australian Journal of Dairy Technology 64, 12-15.
Kelly, A.L., Huppertz, T. y Fox, P.F. (2002). Structural properties of casein micelles in high-
pressure-treated milk. Technical Program Abstract 197 IFT Annual Meeting.
Kilara, A. (2004). Whey proteins. En: Proteins in food processing. Yada, R.Y. (Eds.).
Woodhead Publishing Limited. Cambridge, pp. 72-99.
Knoerzer, K., Buckow, R. y Versteeg, C. (2010b). Adiabatic compression heating
coefficients for high-pressure processing- A study of some insulating polymer materials.
Journal of Food Engineering 98, 110-119.
Knoerzer, K., Buckow, R., Sanguansri, P. y Versteeg, C. (2010a). Adiabatic compression
heating coefficients for high-pressure processing of water, propylene-glycol and mixtures- A
combined experimental and numerical approach. Journal of Food Engineering 96, 229-238.
Koletko B., von Kries, R., Closa, R., Escribano, J., Scagliolini et al. (2009). Can infant
feeding choices modulate later obesity risk? American Journal of Clinical Nutrition 89,
1502S-158S.
Law, A. J. R., Leaver, J., Felipe, X., Ferragut, V., Pla, R. y Guamis, B. (1998). Comparison
of the effects of HP and thermal treatments on the casein micelles in goat´s milk. Journal of
Agricultural and Food Chemistry 46, 2523-2530.
Lönnerdal, B. (2009). Nutritional roles of lactoferrin. Current Opinion in Clinical Nutrition
and Metabolic Care 12, 293-297.
Lönnerdal, B. (2010). Novel insights into human lactation as a driver of infant formula
development. Nestle Nutrition Workshop Series. Pediatric Program 66, 19-29.
López-Fandiño, R. (2006a). High pressure-induced changes in milk proteins and possible
applications in dairy technology. International Dairy Journal 16, 1119-1131.
López-Fandiño, R. (2006b). Functional improvement of milk whey proteins induced by high
hydrostatic pressure. Critical Reviews in Food Science and Nutrition 46, 351-363.

- 150 -
López-Fandiño, R. y Olano, A. (1998). Effects of high pressure combined with moderate
temperatures on the rennet coagulation properties of milk. International of Dairy Journal 8,
623-627.
López-Fandiño, R. y Olano, A. (1999). Selected indicators of the quality of thermal
processed milk. Food Science And Technology International 5, 121-137.
López-Fandiño, R., Carrascosa, A.V. y Olano, A. (1996). The effects of high pressure on
whey protein denaturation and cheese-making properties of raw milk. International Dairy
Journal 79, 929-936.
López-Fandiño, R., De la Fuente, M.A., Ramos, M. y Olano, A. (1998). Distribution of
minerals and proteins between the soluble and colloidal phases of pressurized milks from
different species. Journal of Dairy Research 65, 69-78.
López-Fandiño, R., Ramos, M. y Olano, A. (1997). Rennet coagulation of milk subjected to
high pressures. Journal of Agricultural and Food Chemistry 45, 3233-3237.
Lucey, J.A., Gorry, C., OKennedy, B., Kalab, M., Tankinita, R. y Fox, P.F. (1996). Effect of
acidification and neutralization of milk on some physico-chemical properties of casein
micelles. International Dairy Journal 6, 257-272.
Maede, S.J., Reid, E. A. y Gerrard, J.A. (2005). The impact of processing on the nutritional
quality of food proteins. Journal of AOAC International 88, 904-922.
Manso, M., Miguel, M. y López-Fandiño, R. (2007). Application of capillary zone
electrophoresis to the characterisation of the human milk protein profile and its evolution
throughout lactation. Journal of Chromatograhy A 1146, 110-117.
Martínez-Maqueda, D., Miralles, B., Recio, I. y Hernández-Ledesma, B. (2012).
Antihypertensive peptides from food proteins: a review. Food & Function 3, 350-361.
Masson, P. (1992). Pressure denaturation of proteins. En: Pressure and biotechnology. Balny,
B., Hayashi, R. y Heremans, K. (Eds.). Libby Eurotext Ltd. Montrouge, France, pp. 89-99
Merel-Rausch, E., Duma, I. P. y Hinrichs, J. (2006). Pressure-induced modification of casein
micelles- Influence of pressure built-up rate, pressure level, release rate and temperature on
viscosity and particle size. Milchwissenschaft 61, 255-259.

BIBLIOGRAFÍA
- 151 -
Merel-Rausch, E., Kulozik, U. y Hinrichs, J. (2007). Influence of pressure release rate and
protein concentration on the formation of pressure-induced casein structures. Journal of Dairy
Research 74, 283-289.
Mertens, B. y Deplace, G. (1993). Engineering aspects of high-pressure technology in the
food industry. Food Technology 47, 164-169.
Molina, E., Álvarez, M.D., Ramos, M., Olano, A. y López-Fandiño, R. (2000). Use of high-
pressure treated milk for the production of reduced-fat cheese. International Dairy Journal
10, 467–475.
Monahan, F.J., McClemants, D.J. y Kinsella, J.E. (1993). Polymerization of whey proteins
in whey protein-stabilized emulsions. Journal of Agricultural and Food Chemistry 41, 1826-
1829.
Montilla, T., Moreno, F. J. y Olano, A. (2005). A reliable gas capillary chromatographic
determination of lactulose in dairy samples. Chromatographia 62, 311-314.
Morales, V., Olano, A. y Corzo, N. (2004). Ratio maltose to maltulose and furosine as quality
parameters for infant formula. Journal of Agricultural and Food Chemistry 52, 6732-6736.
Morr, C.V. (1967). Effect of oxalate and urea on the ultra-centrifugation properties of raw
and heated skim milk casein micelles. Journal of Dairy Science 50, 1744-1751.
Mozhaev, V.V., Heremans, K., Frank, J., Masson, P. y Balny, C. (1996). High pressure
effects on protein structure and function. Proteins-Structure Function and Genetics 24, 81-91.
Mozhaev, V.V., Heremans, K., Frank, J., Masson, P. y Balny, C. (1994). Exploiting the
effects of high hydrostatic-pressure in biotechnological applications. Trends in Biotechnology
12, 493-501.
Mulsow, B. B., Jacob, M. y Henle, T. (2009). Studies on the impact of glycation on the
denaturation of whey proteins. European Food Research and Technology 228, 643-649.
Nabhan, M.A., Girardet, J.M., Campagna, S., Gaillard, J.L. y Le Roux, Y. (2004). Isolation
and characterization of copolymers of β-lactoglobulin, α-lactalbumin, κ-casein, and αs1-
casein generated by pressurization and thermal treatment of raw milk. Journal of Dairy
Science 87, 3614-3622.

- 152 -
Nasirpour, A., Scher, J. y Desobry, S. (2006). Baby foods: formulations and interactions (a
review). Critical Reviews in Food Science and Nutrition 46, 665-681.
Needs, E.C., Capellas, M., Bland, A.P., Manoj, P., MacDougal, D. y Paul, G. (2000a).
Comparison of heat and pressure treatments of skimmed milk, fortified with whey
concentrate, for set yogurt preparation: effects on milk proteins and gel structure. Journal of
Dairy Research 67, 329-348.
Needs, E.C., Stenning, R.A., Gill, A.L., Ferragut, V. y Rich, G.T. (2000b). High-pressure
treatment of milk: effects on casein micelle structure and on enzymatic coagulation. Journal
of Dairy Research 67, 31-42.
Nguyen, L. T. y Balasubramaniam, V. M. (2011). Fundamentals of food processing using
high pressure. En: Nonthermal processing technologies for food. Zhang, H. Q., Barbosa-
Cánovas, G. V., Balasubramaniam, V. M., Dunne, C. P., Farkas, D. F. y Yuan, T. C. (Eds).
Wiley-Blackwell. Iowa, USA, pp. 3-19.
Norton, T. y Sun, D. W. (2008). Recent advantages in the use of HP as an effective
processing technique in the food industry. Food and Bioprocess Technology 1, 2-34.
Okpala, C.O.R, Piggott, J.R. y Schaschke, C.J. (2010). Influence of high-pressure processing
(HPP) on physico-chemical properties of fresh cheese. Innovative Food Science and
Emerging Technologies 11, 61-67.
Orlien, V., Boserup, L. y Olsen, K. (2010). Casein micelle dissociation in skim milk during
high-pressure treatment: effects of pressure, pH and temperature. Journal of Dairy Science
93, 12-18.
Orlien, V., Knudsen, J.C., Colon, M. y Skibsted, L.H. (2006). Dynamics of casein micelles in
skim milk during and after high pressure treatment. Food Chemistry 98, 513-521.
Orlien, V., Olsen, K. y Skibsted, L.H. (2007). In situ measurements of ph changes in β-
lactoglobulin solutions under high hydrostatic pressure. Journal of Agricultural and Food
Chemistry 55, 4422-4428.

BIBLIOGRAFÍA
- 153 -
Panick, G., Melessa, R. y Winter, R. (1999). Differences between the pressure- and
temperature-induced denaturation and aggregation of β-lactoglobulin A, B and AB monitored
by FTIR spectroscopy and small-angle X-ray scattering. Biochemistry 38, 6512-6519.
Patel, H. A., Singh, H., Anema, S. G. y Creamer, L. K. (2006). Effect of heat and high
pressure hydrostatic pressure treatments on disulfide bonding interchanges among the proteins
in skim milk. Journal of Agricultural and Food Chemistry 54, 3409-3420.
Patel, H.A., Sigh, H., Havea, P., Considine, T. y Creamer, L.K. (2005). Pressure-induced
unfolding and aggregation of the proteins in whey protein concentrate solutions. Journal of
Agricultural and Food Chemistry 53, 9590-9601.
Paulsson, M. A., Svensson, U., Kishore, A. R. y Naidu, A. S. (1993). Thermal-behaviour of
bovine lactoferrin in water and its relation to bacterial interaction and antibacterial activity.
Journal of Dairy Science 76, 3711-3720.
Pereyra-Gonzáles, A. S., Naranjo, G. B., Malec, L. S. y Vigo, M. S. (2003). Available
lysine, protein digestibility and lactulose in commercial infant formulas. International Dairy
Journal 13, 95-99.
Pischetsrieder, M. y Henle, T. (2012). Glycation products in infant formulas: chemical,
analytical and physiological aspects. Amino Acids 42, 1111-1118.
Raventós, M. (2005). Tratamientos por altas presiones en la conservación y transformación
de alimentos. En: Industria alimentaria. Tecnologías emergentes. Edicions de la Universidat
Politècnica de Catalunya. Barcelona, Spain, pp. 25-76.
Recio, I., Amigo, L., Ramos, M. y López-Fandiño, R. (1997). Application of capillary
electrophoresis to the study of proteolysis of caseins. Journal of Dairy Research 64, 221-230.
Recio, I., Ramos, M. y López-Fandiño, R. (2001). Capillary electrophoresis for the analysis
of food proteins of animal origin. Electrophoresis 22, 1489-1502.
Regnard, P. (1884). Effet des hautes pressions sur les animaux marins. Comptes Rendus des
Seances de la Societe de Biologie et de ses Filiales 36, 394–395.

- 154 -
Regnault, S., Thiebaud, M., Dumay, E. y Cheftel, J.C. (2004). Pressurization of raw skim
milk and of a dispersion of phosphocaseinate at 9 ºC or 20 ºC: effects on casein micelle size
distribution. International Dairy Journal 14, 55-68.
Resmini, P., Pellegrino, L. y Batelli, G. (1990). Accurate quantification of furosine in milk
and dairy products by a direct HPLC method. Italian Journal of Food Science 2, 173–183.
Riel, R. (1991). Composición y estructura físico-química de la leche. En: Ciencia y
tecnología de la leche. Amiot, J. (Eds.) Acribia. Zaragoza, Spain, pp. 1-53.
Riordan, J. (1998). The biologic specificity of breastmilk. En: Breastfeeding and human
lactation. 2ªed. Riordan, J. y Auerbach, K. G. (Eds). Boston, Jones and Barlett Publisher. pp.
121-161.
Sakurai, K. y Goto, Y. (2007). Principal component analysis of the pH-dependent
conformational transitions of bovine β-lactoglobulin monitored by heteronuclear NMR.
Proceedings of the National Academy of Sciences of the United States of America 104, 15346-
15351.
Sandra, S., Stanford, M.A. y Meunier Goddik, L. (2004). The use of high pressure processing
in the production of Queso Fresco cheese. Journal of Food Science 69, FEP153–FEP158.
Sarriá, B., López-Fandiño, R. y Vaquero, M.P. (2001). Does processing of a powder or in-
bottle-sterilized liquid infant formula affect calcium bioavailability? Nutrition 17, 326-331.
Scheedhara, A., Flengsrud, R., Langsrud, T., Kaul, P., Prakash, V. y Vegarud, G. E. (2010).
Structural characteristic, pH and thermal stabilities of apo and holo forms of caprine and
bovine lactoferrins. Biometals 23, 1159-1170.
Schlimme, E. y Buchheim, W. (2002). Proteínas lácteas: caseínas, proteínas del suero y
proteínas minoritarias. En: La leche y sus componentes. Propiedades químicas y físicas.
Acribia. Zaragoza, Spain, pp. 33-75.
Schmidt, D.G. (1982). Association of caseins and casein micelle structure. En: Developments
in Dairy Chemistry, volume 1: Proteins. Fox, P.F. (Eds). Applied Science Publishers. London,
UK, pp. 63-110.

BIBLIOGRAFÍA
- 155 -
Schrader, K. y Buchheim, W. (1998). High pressure effects on the colloidal calcium
phosphate and the structural integrity of micellar casein in milk II. Kinetics of the casein
micelle disintegration and protein interactions in milk. Kieler Milchwirtschaft- lithe
Forschungsber 50, 79-88.
Schrader, K., Buchheim, W. y Morr, C.V. (1997). High pressure effects on the colloidal
calcium phosphate and the structural integrity of micellar calcium. Part 1. High pressure
dissolution of colloidal calcium phosphate in heated milk systems. Nahrung 41, S133-S138.
Schwarcz, W., Carnelocce, L., Silva, J., Oliveira, A. C. y Goncalves, R. B. (2008).
Conformational changes in bovine lactoferrin induced by slow or fast temperature increases.
Biological Chemistry 389, 1137-1142.
Schwertfeger, M. y Buchheim, W. (1999). Coagulation of skim milk under high hydrostatic
pressure with acidification by glucono-dlactone. International Dairy Journal 9, 487–492.
Scollard, P.G., Beresford, T.P., Murphy, P.M. y Kelly, A.L. (2000). Plasmin activity, β-
lactoglobulin denaturation and proteolysis in high pressure treated milk. International Dairy
Journal 10, 835-841.
Silva, J.L. y Weber, G. (1993). Pressure stability of proteins. Annual Review of Physical
Chemistry 44, 89-113.
Slattery, C.W. (1976). Casein micelle structure; an examination of models. Journal of Dairy
Science 59, 1547-1556.
Smith, M. y McMahon, D.J. (1996). Aseptic rennet coagulation of ultrahigh temperature
processed milk concentrates. Journal of Food Science 79, 1513-1520.
Steijns, J.M. y van Hooijdonk, A.C.M. (2000). Occurrence, structure, biochemical
properties and technological characteristics of lactoferrin. British Journal of Nutrition 84,
S11-S17.
Tanaka, N. y Kunugi, S. (1996). Effect of pressure on the denaturation exchange reaction of
α-lactalbumin and β-lactoglobulin. International Journal of Biological Macromolecules 18,
33-39.

- 156 -
Thompkinson, D. K. y Kharb, S. (2007). Aspects of infant food formulation.
Comprehensive Reviews in Food Science and Food Safety 6, 79-102.
Ting, E. (2011). High-pressure processing equipment fundamentals. En: Nonthermal
processing technologies for food. Zhang, H. Q., Barbosa-Cánovas, G. V., Balasubramaniam,
V. M., Dunne, C. P., Farkas, D. F. y Yuan, T. C. (Eds.). Wiley-Blackwell. Iowa, USA, pp. 20-
27
Ting, E.Y. y Marshall, R.G. (2002). Production issues related to UHP food. En: Engineering
and food for the 21st century. Welti-Chanes, J., Barbosa-Canovas, G.V. y Aguilera, J.M.
(Eds.). CRC Press, Boca Raton, pp 727-738.
Tomita, M., Wakabayashi, H., Shin, K., Yamauchi, K., Yaeshima, T. y Iwatsuki, K. (2009).
Twenty-five years of research on bovine lactoferrin applications. Biochimie 91, 52-57.
Tonello, C. (2011). Case studies on high-pressure processing of foods. En: Nonthermal
processing technologies for food, Zhang, H. Q., Barbosa-Cánovas, G. V., Balasubramaniam,
V. M., Dunne, C. P., Farkas, D. F. y Yuan, T. C. (Eds.). Wiley-Blackwell. Iowa, USA, pp. 36-
50.
Viazis, S., Farkas, B.E. y Allen, J.C. (2007). Effects of high-pressure processing on
immunoglobulin A and lysozyme activity in human milk. Journal of Human Lactation 23,
253-261.
Visser, H. (1992). A new casein micelle model and its consequences for pH and temperature
effects on the properties of milk. En: Protein interactions. Visser, H. (Eds.). VCH, Weinheim,
Germany, pp 135-165.
Walstra, P. (1999). Casein sub-micelles: do they exist?. International Dairy Journal 9, 189-
192.
Walstra, P. y Jenness, R. (1984). Dairy Chemistry and Physics. John Wiley and Sons, New
York, USA.
Walstra, P., Geurts, T.J., Noomen, A., Jellma, A. y van Boekel, M. (1999). Dairy
technology: Principles of milk properties and processes. Marcel Dekker Inc. New York,
USA.

BIBLIOGRAFÍA
- 157 -
Waugh, D.F. (1971). Formation and structure of casein micelles. En: Milk proteins:
Chemistry and molecular biology. Volume 2. McKenzie, H.A. (Eds). Academic Press, New
York, pp 3-85.
Ye, A., Anema, S.G. y Singh, H. (2004). High-pressure-induced interactions between milk fat
globule membrane proteins and skim milk proteins in whole milk. Journal of Dairy Science
87, 4013-4022.
Zhai, J., Miles, A.J., Pattenden, L.K., Lee, T.H., Augustin, M.A., Wallace, B.A., Marie-
Isabel Aguilar, M.I. y Wooster, T.J. (2010). Changes in β-lactoglobulin conformation at the
oil/water interface of emulsions studied by synchrotron radiation circular dichroism
spectroscopy. Biomacromolecules 11, 2136-2142.
Zhou, G.H., Xu, X.L. y Liu, Y. (2010). Preservation technologies for fresh meat- A review.
Meat Science 86, 119-128.
Zobrist, M.R., Huppertz, T, Uniacke, T., Fox, P.F y Kelly, A.L. (2005). High pressure –
induced changes in rennet-coagulation properties of bovine milk. International Dairy Journal
15, 655-662.