estructura del adn
TRANSCRIPT

276 CAPITULO 11
Surco menor
[•'¡gura 11.9. (arriba) Modelo de
espacios atómicos de la doble hélice
de
DNA. (abajo) Representación de un
segmento corto desenrollado de
pares
de nucleótidos, en el que se aprecia
cómo los emparejamientos A-T y
G-C producen las proporciones de
Chargaff. Este modelo corresponde a
una de las diversas formas del DNA,
llamada forma B (véase fig. 11.10).
(Modela de espacios atómicos
tomado
de C. Yanofsky, «Gene structure and
Protein Structure». Copyright © 1967
de Scientific American, Inc.
Reservados
todos los derechos. Estructura
desenrollada basada en A. Konberg,
«The Synthesis of DNA». Copyright
© 1968 de Scientific American Inc.
Reservados todos los derechos.)
otros) en un libro fascinante, titulado «La doble hélice»,
que revela el intrincado juego entre personalidades cien-
tíficas, las inteligentes apreciaciones, el trabajo duro y el
puro azar, que rodearon tan importante avance cientí-
fico.
Estructuras alternas
Además de las formas A y B del DNA, se ha encontrado
un nuevo tipo de estructura en cristales de un DNA
sintético que contiene purinas y pirimidinas, alternadas
sobre la misma cadena. Esta forma de DNA, la forma Z,
tiene un esqueleto en forma de zigzag y forma una hélice
«a izquierdas». Los dibujos en perspectiva de las figuras
11.11, 11.12 y 11.13 nos permiten comparar estas tres
formas sobre la base de unos datos precisos obtenidos
con estructuras cristalinas de secuencias sintéticas cortas
de DNA, llamadas oligonucleótidos. Que la forma Z ocu-
rra o no en la naturaleza es una cuestión que todavía se
sigue debatiendo.
propiedad esencial de la molécula genética había
sido un misterio.
2. La estructura hace pensar que quizás la secuencia
de pares de nucleótidos en el DNA es la que deter-
mina la secuencia de aminoácidos de la proteína
dictada por un gen. En otras palabras, algún tipo
de código genético podría escribir información en
el DNA como una secuencia de pares de nucleóti-
. dos y luego traducirla en un lenguaje diferente de
secuencias de aminoácidos en la proteína.
Estos datos básicos sobre el DNA son ahora familiares
a casi cualquiera que haya leído un texto de biología en
una escuela primaria o secundaria, o incluso en periódi-
cos y revistas. Puede parecer trivial y obvio, pero trate de
traladarse al escenario de 1953 e ¡imagine la excitación
de entonces! Hasta ese momento, la evidencia de que el
poco interesante DNA era el material genético resultaba
descorazonadora y decepcionante. Pero la estructura de
Watson y Crick abría de repente la posibilidad de expli-
car dos de los grandes «secretos» de la vida. James Wat-
son ha contado-la historia de este descubrimiento (desde
su peculiar punto de vista, fuertemente cuestionado por
LA ESTRUCTURA DEL DNA
\
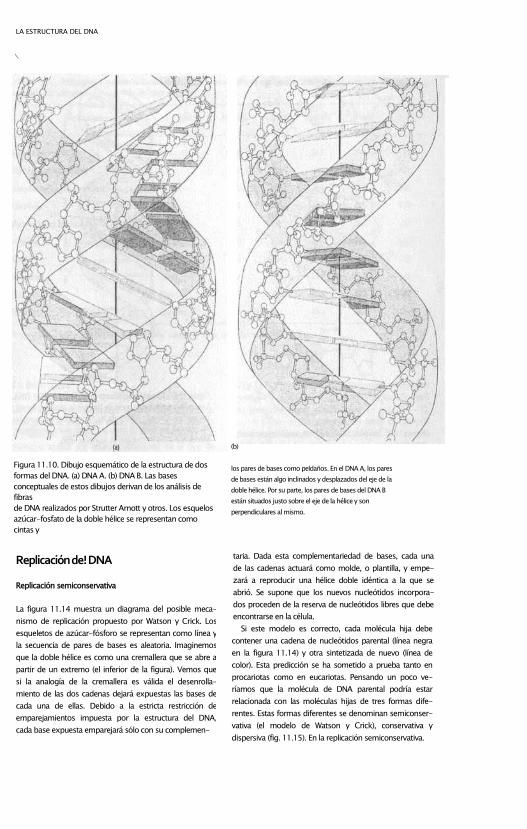
Figura 11.10. Dibujo esquemático de la estructura de dos
formas del DNA. (a) DNA A. (b) DNA B. Las bases
conceptuales de estos dibujos derivan de los análisis de
fibras
de DNA realizados por Strutter Arnott y otros. Los esquelos
azúcar-fosfato de la doble hélice se representan como
cintas y
Replicación de! DNA
Replicación semiconservativa
La figura 11.14 muestra un diagrama del posible meca-
nismo de replicación propuesto por Watson y Crick. Los
esqueletos de azúcar-fósforo se representan como línea y
la secuencia de pares de bases es aleatoria. Imaginemos
que la doble hélice es como una cremallera que se abre a
partir de un extremo (el inferior de la figura). Vemos que
si la analogía de la cremallera es válida el desenrolla-
miento de las dos cadenas dejará expuestas las bases de
cada una de ellas. Debido a la estricta restricción de
emparejamientos impuesta por la estructura del DNA,
cada base expuesta emparejará sólo con su complemen-
(b)
los pares de bases como peldaños. En el DNA A, los pares
de bases están algo inclinados y desplazados del eje de la
doble hélice. Por su parte, los pares de bases del DNA B
están situados justo sobre el eje de la hélice y son
perpendiculares al mismo.
taria. Dada esta complementariedad de bases, cada una
de las cadenas actuará como molde, o plantilla, y empe-
zará a reproducir una hélice doble idéntica a la que se
abrió. Se supone que los nuevos nucleótidos incorpora-
dos proceden de la reserva de nucleótidos libres que debe
encontrarse en la célula.
Si este modelo es correcto, cada molécula hija debe
contener una cadena de nucleótidos parental (línea negra
en la figura 11.14) y otra sintetizada de nuevo (línea de
color). Esta predicción se ha sometido a prueba tanto en
procariotas como en eucariotas. Pensando un poco ve-
ríamos que la molécula de DNA parental podría estar
relacionada con las moléculas hijas de tres formas dife-
rentes. Estas formas diferentes se denominan semiconser-
vativa (el modelo de Watson y Crick), conservativa y
dispersiva (fig. 11.15). En la replicación semiconservativa.
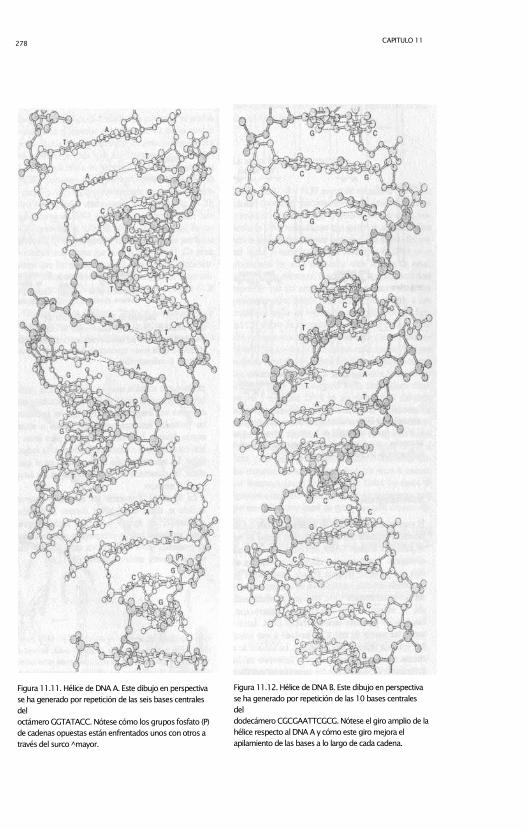
278 CAPITULO 11
Figura 11.11. Hélice de DNA A. Este dibujo en perspectiva
se ha generado por repetición de las seis bases centrales
del
octámero GGTATACC. Nótese cómo los grupos fosfato (P)
de cadenas opuestas están enfrentados unos con otros a
través del surco ̂ mayor.
Figura 11.12. Hélice de DNA B. Este dibujo en perspectiva
se ha generado por repetición de las 10 bases centrales
del
dodecámero CGCGAATTCGCG. Nótese el giro amplio de la
hélice respecto al DNA A y cómo este giro mejora el
apilamiento de las bases a lo largo de cada cadena.
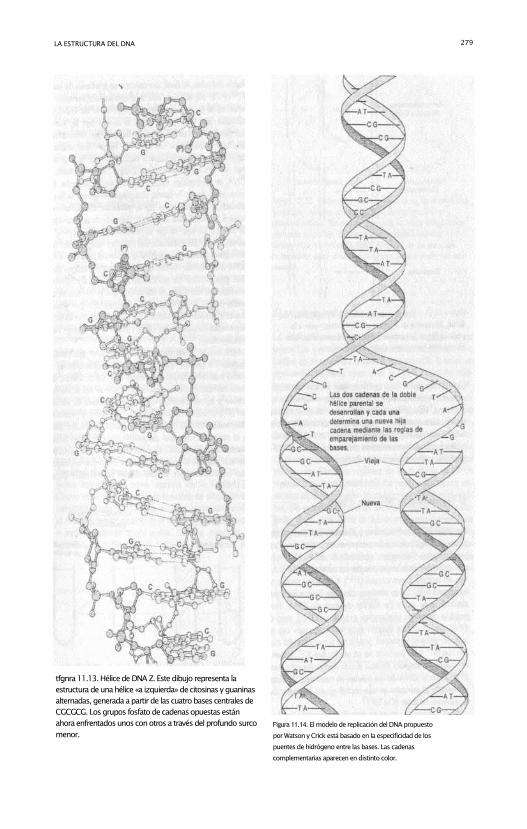
LA ESTRUCTURA DEL DNA 279
tfgnra 11.13. Hélice de DNA Z. Este dibujo representa la
estructura de una hélice «a izquierda» de citosinas y guaninas
alternadas, generada a partir de las cuatro bases centrales de
CGCGCG. Los grupos fosfato de cadenas opuestas están
ahora enfrentados unos con otros a través del profundo surco
menor.
Figura 11.14. El modelo de replicación del DNA propuesto
por Watson y Crick está basado en la especificidad de los
puentes de hidrógeno entre las bases. Las cadenas
complementarias aparecen en distinto color.

280 CAPITULO 11
Semiconservativa
(Watson y Crick)
Conservativa
Dispersiva
Figura 11.15. Tres formas alternativas de replicación del
DNA. El modelo de Watson y Crick corresponde a la
primera forma (semiconservativa). Las líneas coloreadas
representan las cadenas sintetizadas de nuevo.
cada hélice doble hija contiene una cadena parental y
otra recién sintetizada. En la replicación conservativa, sin
embargo, una hélice doble hija tiene las dos cadenas
sintetizadas de nuevo y se conserva la hélice doble paren-
tal. La replicación dispersiva da lugar a dos moléculas
hijas cuyas cadenas contienen ciertos segmentos de DNA
parental y otros sintetizados de nuevo.
El experimento de Meselson y Stahl
En 1958, Matthew Meselson y Franklin Stahl se propu-
sieron distinguir de manera experimental entre esas tres
posibilidades. Cultivaron células de E. coli en un medio
que contenía un isótopo pesado del nitrógeno (N15), en
vez del isótopo ligero normal (N14). El isótopo pesado se
incorporó a las bases nitrogenadas, que fueron incorpo-
rándose a su vez a las nuevas cadenas de DNA. Tras
muchas divisiones celulares en presencia de N15, todo el
DNA de las células debería estar bien marcado con el
isótopo pesado. Entonces, separaron las células del me-
dio con N15 y las pusieron en un medio con Nu, toman-
do muestras después de una y dos divisiones celulares.
Extrajeron el DNA de las células de cada una de estas
muestras y lo pusieron en una centrífuga, en una solución
de cloruro de cesto (CsCl).
Si el cloruro de cesio se centrifuga durante muchas
horas a velocidades enormes (50000 r.p.m.), los iones del
cloruro son empujados por la fuerza centrifuga hacia el
fondo del tubo. Al final, se establece un gradiente de iones
Cs! y CI" en el tubo, con mayores concentraciones de
ellos en el fondo. También las moléculas de DNA en
solución son empujadas hacia el fondo por la fuerza
centrifuga. Pero en su trayecto hacia abajo, encuentran
una concentración de sal cada vez mayor que tiende a
empujarlas de nuevo hacia arriba, debido a la flotabili-
dad del DNA (su tendencia a flotar). De esa manera, el
DNA se «deposita» finalmente en cierta posición del
tubo, justo donde la fuerza centrífuga compensa la flota-
bilidad de las moléculas en el gradiente de cloruro de
cesio. La flotabilidad del DNA depende de su densidad
(la cual, a su vez, refleja la relación de pares G-C a
pares A-T). La presencia del isótopo más pesado del
nitrógeno cambia la densidad de flotación del DNA. El
DNA extraído de células que han crecido durante mu-
chas generaciones en medio con N15 puede distinguirse
con facilidad del DNA de las células que lo han hecho en
medio con N14, por la posición de equilibrio que alcan-
zan en el gradiente de cloruro de cesio. Tales muestras se
denominan familiarmente DNA «pesado» y «ligero», res-
pectivamente.
Meselson y Stahl comprobaron que una generación
después de que las células «pesadas» se trasladaron a
medio con N14, el DNA formaba una única banda, de
densidad intermedia entre las densidades de los controles
pesado y ligero. Después de dos generaciones en medio
con N14, el DNA formaba dos bandas: una en la posición
intermedia y la otra en la posición ligera (fig. 11.16). Este
es el resultado esperado del modo de replicación semi-
conservativa; de hecho, el resultado sólo es compatible
con este modelo, si el experimento se inicia con cromoso-
mas constituidos por hélices dobles individuales (figu-
ra 11.17).
Incubación de células
«pesadas» en N11
Centrifugación de DNA en un gradiente de
cloruro de cesio (CsCl). Los cultivos crecidos durante muchas
generaciones en medio con N15 o N14 permiten establecer los
controles de posición de las bandas de DNA «pesado» y
«ligero», respectivamente. Cuando células cultivadas en N,s
se transfieren a medio con N14, la primera generación
produce una banda de DNA intermedia, y la segunda
generación produce dos bandas: una intermedia y otra ligera.

LA ESTRUCTURA DEL DNA 281
¡■¡gura 11.17. Sólo el modelo semiconservativo de
replicación del DNA predice resultados como los de la figura
11.16; una banda intermedia en la primera generación y una
intermedia y otra ligera en la segunda generación. (Véase la
explicación de los símbolos en la figura 11.15.)
Autorradiografía
El experimento con E. coli de Meselson y Stahl fue prác-
ticamente repetido con cromosomas de células de raíces
de judías por Herbert Taylor, en 1958, utilizando técnicas
citológicas. Taylor puso las células de raíz en una solu-
ción que contenía timidina tritiada (tim¡dina-[H3]), el
nucleótido timidina marcado con un isótopo radioactivo
del hidrógeno llamado tritio. Dejó que las células realiza-
ran la mitosis en esta solución, de manera que la
timidína-[H3] pudiera incorporarse al DNA. Lavó en-
tonces las puntas de raíz y las trasladó a una solución
que contenía timidina no radioactiva. La adición de col-
chicina a esta preparación inhibe la formación del huso
acromático, de manera que los cromosomas metafásicos
no se separan y las cromátidas hermanas permanecen
unidas entre sí por el centròmero.
La localización celular del H3 puede hacerse mediante
autorradiografía. Al descomponerse, el H3 emite partícu-
las beta (electrón con energía). Al colocar una lámina de
emulsión fotográfica sobre una célula que contiene H3, se
Aspecto cuando p¡n ¡a Aspecto cuando
se añade primera sfi añade
colchicina en la división colchicina en la
primera división segunda división
¡■'¡¡jura 11.13. Esquema de la autorradiografía de
cromosomas de células cultivadas durante un ciclo de
división en presencia del isótopo radioactivo del hidrógeno
H! (tritio). Cada punto representa la huella dejada por una
partícula de radioactividad.
produce una reacción química allí donde una partícula
beta choque con la emulsión. Esta puede revelarse enton-
ces como una fotografía, de forma que las emisiones de
las partículas beta aparecen como granos o manchas
negras. Además, la célula puede teñirse para hacer visi-
bles sus estructuras e identificar la fuente de la radioacti-
vidad. La autorradiografía es un método en el que, en
efecto, las estructuras celulares radioactivas «se fotogra-
fían a sí mismas».
La figura 11.18 muestra los resultados observados
cuando se añade la colchicina durante la división en
timidina-[H3] o durante la división mitótica siguiente.
Estos resultados pueden interpretarse representando ca-
da cromátida como una molécula única de DNA que se
replica de forma semiconservativa (fig. 11.19).
Hi'iiva 1 U‘.' Posible interpretación de la figura 11.18 a
nivel del DNA. Las lineas coloreadas representan cadenas
radioactivas.

282 CAPITULO 11
Cromosomas en arlequín
Usando una técnica de tinción más moderna, es posible
visualizar la replicación semiconservativa de los cromo-
somas en mitosis, sin necesidad de autorradiografia. En
este método, se deja que los cromosomas sufran dos
ciclos de replicación en presencia de bromodeoxiuridína
(BUdR). Entonces, se tiñen los cromosomas con Giemsa
y con un colorante fluorescente; este procedimiento pro-
duce los llamados cromosomas en arlequín (fig. 11.20).
Las cadenas de DÑA sintetizadas en presencia de bro-
modeoxiuridina se tiñen de manera diferente que las ca-
denas del DNA «original». La razón de este patrón es
exactamente la misma que la de la figura 11.19. (De paso,
nótese que. los cromosomas en arlequín son particular-
mente adecuados para detectar intercambios entre cro-
mátidas hermanas durante la mitosis; en la figura 11.20
se ven dos ejemplos.)
Usando técnicas similares, Taylor demostró que la re-
plicación cromosómica durante la necrosis es también
semiconservativa. Este resultado da otro empujoncito a
la tumba al modelo de elección de copia del entrecruza-
miento (capítulo 5), que propone una replicación cromo-
sómica durante la meiosis.
La estructura del cromosoma
Las figuras 11.18 y 11.19 ponen de manifiesto una de las
grandes cuestiones genéticas que quedan por resolver: un
cromosoma eucaríótico ¿es esencialmente una única mo-
lécula de DNA rodeada por una matriz de proteína? Dos
hechos apoyan fuertemente que así es, en efecto. En pri-
mer lugar, si hubieran muchas moléculas de DNA en el
cromosoma (ya sean dispuestas unas al lado de otras,
extremos con extremos, u orientadas al azar), sería casi
imposible que los cromosomas se replicaran de forma
semiconservativa (yendo todo el material marcado a una
cromátida, según los resultados de Taylor). Mire la figu-
ra 11.21 y trate de imaginar cómo esto sería posible. Estu-
dios recientes con cromosomas aislados y largas molécu-
las de DNA apoyan la idea de que cada cromátida es una
molécula única de DNA. Esto significa una molécula muy
larga. Por ejemplo, sólo en un cromosoma humano hay
suficiente DNA para alargarlo hasta tres o cinco centí-
metros, y suficiente DNA en un núcleo para alargarlo
hasta un metro. (Ello plantea otro problema interesante:
¿en qué forma está empaquetada esta larguísima molécu-
la que permite una fácil replicación?) El segundo hecho
que apoya la hipótesis de una única molécula es que el
DNA y los genes se comportan como si estuvieran uni-
dos extremo con extremo en una única ristra o cadena,
que llamamos grupo de ligamiento. Todos los datos de
ligamiento (capítulo 5) nos dicen que no necesitamos más
que una única serie lineal de genes por cada cromosoma
para explicar los fenómenos genéticos.
Como acabamos de mencionar, hay demasiado DNA
en un cromosoma como para que pueda extenderse li-
Figura 11.20. Cromosomas en arlequíwde células de ovario
de hámster chino (CHO). El método consiste en dejar que los
cromosomas pasen dos ciclos de replicación en presencia de
bromodesoxiuridina (BUdR), que sustituye a la timidina en el
DNA sintetizado de nuevo. Los cromosomas al teñirse con
Giemsa y un colorante fluorescente, toman la apariencia de
la figura. Las cadenas de DNA replicadas de nuevo en BUdR
se tiñen de forma diferente a las cadenas de DNA «original».
Arriba (véanse flechas), un cromosoma ha sufrido dos
intercambios entre cromátidas hermanas. (Fotografía cortesía
de Sheldon Wolf y Judy Bodycote.)
¿Lado a lado?
¿Extremo con extremo? ■—
¿Apiladas?
¿Aleatoriamente? (
Figura 11.21. Algunas alternativas teóricas de
empaquetamiento del DNA en un cromosoma eucaríótico.
Ninguno de estos modelos puede reconciliarse fácilmente con
los datos que apoyan el modelo de replicación
semiconservativa del DNA. El tema del empaquetamiento de
una molécula larga de DNA se trata en el capítulo 14.

LA ESTRUCTURA DEL DNA 283
nealmente a lo largo del mismo. Debe estar empaquetado
muy eflcientemente. Las ideas actuales (apoyadas por
buenas pruebas microscópicas) apuntan a un proceso de
enrollamiento y superenrollamiento del DNA. Gire una
cinta de goma con sus dedos y compruebe cómo se enro-
lla; así puede ocurrir con los cromosomas. Volveremos a
este asunto en el capitulo 14.
La horquilla de replicación
Una predicción del modelo de replicación del DNA de
Watson y Crick es que debe encontrarse una horquilla en
la molécula de DNA durante su replicación. John Cairns,
en 1963, puso a prueba esta predicción dejando que el
DNA en replicación de células bacterianas incorporara
timidina tritiada. Teóricamente, cada molécula hija sinte-
tizada de nuevo debe contener entonces una cadena ra-
dioactiva («caliente») y otra no radioactiva («fría»). Tras
variar el período y el número de ciclos de replicación en
medio «caliente», Cairns extrajo el DNA de las células, lo
puso en un portaobjetos y lo autorradiografió, para ob-
servarlo luego al microscopio electrónico. Tras un ciclo
de replicación en timidina-[H3], aparecieron anillos de
puntos en la autorradiografia. Cairns interpretó los ani-
llos como se muestra en la figura 11.21 En ella resulta
aparente que el cromosoma bacteriano es circular, hecho
que se deducía también de los datos genéticos descritos
anteriormente (capítulo 10).
Durante el segundo ciclo de replicación, se vieron real-
mente las horquillas predichas por el modelo. Además, la
densidad de granos en los tres segmentos era tal, que
podía interpretarse como aparece en la figura 11.23.
Cairns observó toda clase de tamaños en estos patrones
autorradiográficos en forma de luna, correspondiéndose
al movimiento progresivo de la cremallera de replicación,
u orquilla, alrededor del anillo. Formas como las que
aparecen en la figura 11.23 se llaman formas theta (0).
Autorradiografia
Figura 11.23. (izquierda) Autorradiografia de un cromosoma
bacteriano durante el segundo ciclo de replicación en
timidina tritiada. En esta estructura 0, la hélice doble recién
replicada que cruza el círculo tendría ambas cadenas
radioactivas. (derecha) La doble densidad de las manchas
radioactivas sobre el autorradiograma parece confirmar este
modelo.
£1 origen de replicación
La replicación se inicia en las bacterias a partir de un
origen fijo, y progresa luego bidireccionalmente, con hor-
quillas en movimiento a ambos extremos de la zona en
replicación, tal como aparece en la figura 11.24. En célu-
las superiores la replicación progresa a partir de múlti-
ples puntos u orígenes. Supongamos que exponemos una
célula eucariótica brevemente a timidina-[H3], paso que
denominamos pulso, y luego añadimos un exceso de timi-
dina «fría», paso que denominamos caza; se extrae en-
tonces el DNA y se hace una autorradiografia. La figura
11.25 muestra el resultado de tal experimento, con lo que
parecen ser diferentes regiones de la molécula de DNA
replicándose simultáneamente. La replicación parece co-
menzar en distintos sitios de estos cromosomas eucarióti-
cos. Un experimento similar de pulso, caza y autorradio-
grafia de la replicación de los cromosomas politécnicos
(gigantes) de Drosophila, revela muchas regiones en repli-
cación dentro de un mismo brazo cromosómico (figu-
ra 11.26). Así y todo, no hay una prueba contundente de
que estas regiones correspondan en realidad a diferentes
Figura 11.22. (izquierda) Autorradiografia de un cromosoma
bacteriano después de un ciclo de replicación de timidina
tritiada. Según el modelo de replicación semiconservativa,
una de las dos cadenas debe de ser radioactiva, [derecha)
Interpretación de la autorradiografia.
Figura 11.24. Diagrama de la replicación del DNA
avanzado en las dos direcciones a partir del origen. (Tomado
de A. Kornberg, DMA synthesis. Copyright © 1974 de W. H.
Freeman and Company.)

284 CAPITOLO 11
Figura ¡1.25. Una forma de replicación del DNA, revelada
por autorradiografia. Una célula es expuesta brevemente a
timidlna (H3) (pulso) y transferida luego a timidina no
radioactiva (fría) en exceso (caza). Se extiende el DNA sobre
un portaobjeto y se autorradiografia. Según la interpretación
que se muestra, existirían varios puntos de replicación en una
misma hélice doble de DNA.
puntos de iniciación de una única molécula de DNA;
pueden interpretarse también como prueba de que los
cromosomas están hechos de varias moléculas de DNA
separadas. La estructura del cromosoma eucariótico es
uno de los problemas genéticos más apasionantes no
resueltos todavía (véase capitulo 14).
Enzimologia de la replicación
A finales de los años 50, Arthur Kornberg consiguió
identificar y purificar una enzima, llamada polimerasa de
DNA, que cataliza la siguiente reacción de replicación:
DNA (parental) cebados +
Esta reacción funciona sólo con la forma trifosfato de los
nucleótidos (como el trifosfato de desoxiadenosina o
dATP). Al término de la reacción, la cantidad total de
DNA puede ser hasta 20 veces la cantidad del DNA de
entrada, de manera que el DNA presente al final debe ser
fundamentalmente DNA descendiente. Por tanto, el
aná-
lisis de esta mezcla final de DNA puede considerarse
como indicativa en gran manera de la naturaleza del
DNA descendiente. La figura 11.27 ilustra la reacción de
elongación de la cadena, o reacción de polimerización,
catalizada por las polimerasas de DNA.
Aunque en la figura 11.27 la replicación parece un
proceso sencillo, se dan ciertas complicaciones
(conforme
vamos describiendo estas complicaciones, mire la figu-
ra 11.28, que resume muchos de los pasos intermedios
de
la síntesis de DNA):
1. La doble hélice debe rotar durante el proceso de
replicación, ya que las dos cadenas están
entrelaza-
das. Esta rotación se lleva a cabo con la ayuda de
catalizadores biológicos, enzimas llamadas topo-
isomerasas del DNA, que transforman anillos de
DNA de una forma topològica a otra. Por ejem-
plo, una toposiomerasa, la girasa de DNA, puede
inducir retorcimientos del DNA, llamados
superen-
roliamientos (fig. 11.29). La forma superenrollada
puede facilitar el desenrollamiento de la hélice (fi-
gura 11.30). La proteina «rep», una helicasa,
proba-
blemente está implicada en el desenrollamiento
de
la hélice. Las zonas de cadena sencilla están ex-
puestas a ser degradadas, pero las protege otra
proteina, llamada «proteína de unión a DNA de
cadena simple» (SSB) (fig. 11.28).
2. La idea simple de una molécula abriéndose como
una cremallera no es realmente correcta. Todas
las
polimerasas de DNA conocidas sintetizan nuevas
fioiirs ¡U6. Forma de replicación de un cromosoma de
Drosoplüla revelada por autorradiografia. En un mismo
cromosoma se ven varios puntos de replicación señalados
con flechas.
gura ¡ 1.27. Reacción de elongación de la cadena
catalizada por las polimerasas de DNA. (Figs. 11.27 y 11.31,
tomadas de L. Stryer, Biochemistry, 3.3 ed. Copyright © 198?
de W. H. Freeman and Company.)

LA ESTRUCTURA DEL DNA 285
cadenas sólo en la dirección 5'-»3’. (La primera
enzima estudiada por Kornberg, llamada ahora
polimerasa I, no es la enzima principal para la
replicación del DNA, mientras que si lo es, proba-
blemente, la polimerasa III.) Sabemos que mien-
tras una cadena es sintetizada de forma continua
por la polimerasa III, la otra se fabrica de manera
discontinua. En este último caso, la polimerasa III
inicia su acción sobre muchos puntos distintos del
molde, dejando huecos que son rellenados por la
polimerasa I y luego unidos por la acción de la
enzima ligasa del DNA.
ira i 12!/. Superenrollamiento catalizado por la
girasa de DNA. La replicación del DNA genera un
superenrollamiento «positivo» (parte inferior del
dibujo) que resulta de la rotación rápida del DNA
en la horquilla de replicación. La girasa puede abrir
y cerrar enlaces fosfodiésteres, relajando así el
superenrollamiento (DNA relajado). La girasa
también puede generar superenrollamientos en la
otra dirección, llamado superenrollamiento
«negativo»; este último facilita la disociación de la
doble hélice (véase la íig. 11.28). (Modificado de L.
Strycr, Biochemistry, 3.a ed., Copyright © 1988 de
W. H. Freeman and Company.)

CAPITULO 11
(a) (b) (c)
Figura 11.30. Para que una célula se divida, es necesario
replicar la información constituida por la secuencia de pares
de bases. La repücación de una molécula circular de DNA de
cadena doble puede depender de la capacidad de una
topoisomerasa de permitir que el DNA se desenrolle. Uno de
los mecanismos más frecuentes para duplicar una doble
cadena circular es la fabricación de una cadena
complementaria de cada cadena original. Para construir las
nuevas cadenas, deben separarse las cadenas originales. En
las bacterias, la girasa superenrollada negativamente el anillo
de DNA (a). En esta forma superenrollada negativamente,
resulta más fácil desenrollar la doble hélice. El
(d)
desenrollamiento es necesario probablemente para que
comience la repücación en varios organismos (b). La
fabricación de las nuevas cadenas comienza entonces,
empleándose las cadenas originales como moldes (c). Para
que continúe la replicación, la doble hélice debe ser
desarrollada progresivamente. Ello debe de ocurrir con la
ayuda de una o más moléculas de topoisomerasa (d), después
de lo cual los dos anillos de doble cadena se separarán; cada
anillo está formado por una cadena original y otra nueva.
(Tomado de J. G. Wang, «DNA topoisomerases». Copyright
© 1982 de Scientific American, Inc. Reservados todos los
derechos.)
Tanto la polimerasa de DNA I como la polimera-
sa III poseen además una actividad exonucleasa,
en dirección 3'->5', que cumple una función de
«corrección de pruebas», eliminando aquellas ba-
ses mal apareadas, inscritas erróneamente durante
la polimerización. La figura 11.31 muestra la acti-
vidad exonucleasa 3'-*5', que requiere un extremo
libre 3'—OH no apareado. La escisión regenera
un grupo 3'—OH en el nucleótido anterior, permi-
tiendo que continúe la síntesis, por incorporación
de un trifosfato de nucleótido libre. En la adición
de cada base se requiere la hidrólisis de un enlace
fosfato de alta energía. Así pues, una polimerasa
con una función de síntesis 5'-*3' y otra de correc-
ción por exonucleasa 3'->5 mantiene esta condi-
ción necesaria de la síntesis de DNA. Si las polime-
rasas actuaran en dirección 3'->5', entonces el tri-
fosfato estaría sobre la cadena en crecimiento (véa-
se la fig. 11.27) y el grupo —OH estaría en el
nucleótido libre que se va a añadir. La actividad
exonucleasa de corrección sería en dirección 5'-*3'
y liberaría la base con el enlace fosfato de alta
energía, ¡bloqueando la continuación de la síntesis! Figura 11.31.
de DNA I.
Acción exonucleásica 3'-» 5' de la polimerasa

LA ESTRUCTURA DEL DNA 287
Por esta razón no han aparecido en la evolución
poiimerasas 3'-*5'. La consecuencia, a su vez, de
que todas las poiimerasas actúen en dirección
5' -» 3' es que una de las cadenas deba fabricarse
de forma discontinua.
4. La polimerasa de DNA no puede empezar una
nueva cadena sobre un molde de cadena sencilla,
sin que haya al menos una pequeña región de
doble cadena que actúa como cebador. En las bac-
terias, la enzima primasa, junto con una segunda
protema cifrada por dnuB, sintetiza un cebador de
RNA para la polimerasa III.
La síntesis simultánea de las dos nuevas cadenas de
DNA se muestra en la figura 11.28. En la parte izquierda
del esquema, conforme la hélice se va desenrollando con
la ayuda de la proteína rep, la nueva cadena se fabrica de
manera continua, en dirección 5'-» 3'. Las regiones de
cadena sencilla son estabilizadas por la proteina SSB. En
la parte derecha del esquema, la nueva cadena debe fabri-
carse de forma discontinua, ya que todas las poiimerasas
de DNA actúan en dirección 5'-* 3'. Así, al hacerse dispo-
nibles nuevos segmentos de la cadena antigua, puede
comenzar la síntesis de fragmentos de la cadena nueva.
Esta síntesis requiere una primasa para sintetizar prime-
ro un fragmento corto de RNA que sirva de «cebador»,
sobre el que la polimerasa de DNA III va añadiendo
desoxinucleótidos hasta separarse de la cadena. Como
puede verse en la figura, este proceso deja unos huecos
que son rellenados por la polimerasa de DNA I, la cual
elimina también el pequeño cebador situado al comienzo
del fragmento de doble cadena al que ha llegado ahora.
Entonces, la enzima ligasa de DNA sella el último enlace.
El DNA y el gen \
Hemos visto ya que el DNA es el material genético y que
consiste en una secuencia lineal de pares de nucleótidos.
La conclusión inmediata es que el mapa de alelos consti-
tuye el equivalente genético de las secuencias de pares de
nucleótidos en el DNA. Confirmaríamos esta suposición
si pudiéramos demostrar que los mapas genéticos son
coherentes con los mapas del DNA. Este paso fue realiza-
do por primera vez mediante algunas manipulaciones
bioquímicas y genéticas elegantes.
El DNA del fago Á resultó ser un trozo del DNA lineal
que puede circularizarse, ya que el extremo 5' de cada
cadena tiene una prolongación terminal de 12 bases,
complementaria del otro extremo 5' (figura 11.32). Por
ello, ambos extremos pueden emparejarse y convertir el
DNA en un círculo; se conocen como extremos «cohesi-
vos» o «pegajosos».
Cuando un objeto lineal como la molécula de DNA se
somete a tensiones mecánicas (mediante pipeteo o agita-
ción, por ejemplo) pueden producirse roturas, sobre todo
en la parte central de la molécula. Al romperse el DNA
Figura 11.32. Circulación del DNA.
Dentro del fago, el DNA de A es
lineal, pero una vez en el interior de la
célula hospedadora se circulariza,
como paso previo a su replicación o
inserción. La circularización se
produce al unirse los extremos
complementarios («cohesivos») de
cadena sencilla. (Tomado de A.
Kornberg, DNA Synthesis. Copyright
© 1974 de W. H. Freeman and
Company.)