efectos del estrÉs calorico en el piedemonte … · ii efectos del estrÉs calorico en el...
TRANSCRIPT

I
EFECTOS DEL ESTRÉS CALORICO EN EL PIEDEMONTE AMAZONICO COLOMBIANO SOBRE ALGUNOS
PARAMETROS FISIOLOGICOS Y ZOOTECNICOS EN DOS ESTIRPES DE POLLO DE ENGORDE
Por: Elvis Alexander Díaz López
Universidad Nacional de Colombia Maestría en estudios Amazónicos
Leticia, Colombia Año 2012

II
EFECTOS DEL ESTRÉS CALORICO EN EL PIEDEMONTE AMAZONICO COLOMBIANO SOBRE ALGUNOS
PARAMETROS FISIOLOGICOS Y ZOOTECNICOS EN DOS ESTIRPES DE POLLO DE ENGORDE
Elvis Alexander Díaz López
Trabajo de investigación presentado como requisito parcial para optar al título De:
MAGÍSTER EN ESTUDIOS AMAZÓNICOS
Director: Ph.D, M.Sc William Narvaez Solarte
Codirector:
Ph.D, M.Sc Germán Afanador Téllez
Línea de Investigación: Desarrollo regional
Universidad Nacional de Colombia Maestría en estudios Amazónicos
Leticia, Colombia Año 2012

III
Dedicatoria A mi familia, a mi padre y mi madre por sus consejos y ejemplo, a mi esposa Damaris y
mis hijos Erick, Dominick y Valery quienes son la luz de mi vida y la motivación diaria de
mí existir.

IV
Agradecimientos Al profesor Dr. William Narváez Solarte, por su orientación, enseñanza y ejemplo de
profesionalismo.
A la universidad de la Amazonia por su importante ayuda económica, al señor Pedro
Rojas (avicultor) y su familia por su valiosa colaboración económica y logística, al
profesor, Magister Luis Hernando Ortegón (Q.E.P.D) por su importante ayuda en el
laboratorio.
Al ICA y sus funcionarios del laboratorio diagnostico sede Florencia, a la Médica
Veterinaria y Zootecnista Paola Mantilla, a los alumnos de último semestre de Medicina
Veterinaria y Zootecnia de la Universidad de la Amazonia, Javier Gómez, Cristian
Vargas, Pedro Rojas y Johana Claros por su valiosa colaboración durante la etapa
experimental.

V
Contenido pág.
Resumen………………………………………………………………………………….………X
Abstract....................................................................................................................... XII
Introducción .................................................................................................................. 1
1. Objetivos ................................................................................................................... 2
1.1 Objetivo general ........................................................................................................ 2
1.2 Objetivos específicos ................................................................................................ 2
2. Variables evaluadas ................................................................................................. 3
2.1 Variables zootécnicas ................................................................................................ 3
2.2 Variables anatomofisiológicas .................................................................................. 3
3. Marco teórico ............................................................................................................ 4
3.1 Ambiente, temperatura ambiental y humedad relativa ambiental ............................... 4
3.2 Estrés ........................................................................................................................ 5
3.3 Estrés calórico ........................................................................................................... 6
3.4 Efectos del estrés calórico en el consumo de alimento y agua .................................. 7
3.5 La corticosterona ....................................................................................................... 7
3.6 Efectos del estrés calórico en los órganos ................................................................ 8
3.7 Efectos del estrés calórico en los órganos linfoides .................................................. 9
3.8 Cambios en parámetros sanguíneos y bioquímicos debidos al estrés calórico. ......... 9
3.9 Los electrolitos y ph durante el estrés calórico ...............................………………. 12
Bibliografia ........................................................................................................ .……..14
Capítulo 1………………………………………………………………………………………..22
ÍNDICES ZOOTÉCNICOS DE POLLO DE ENGORDE BAJO CONDICIONES DE
ESTRÉS CALÓRICO EN EL PIEDEMONTE AMAZÓNICO COLOMBIANO..….. …….22
1.1 Introducción ............................................................................................ …… ..22
1.2 Materiales y métodos ............................................................................................. .23
1.2.1 Localización .......................................................................................................... 23
1.2.2 Animales............................................................................................................... 23
1.2.3 Tratamientos ....................................................................................................... 24
1.2.4 Manejo del pollo de engorde................................................................................. 24

VI
1.2.5 Variables evaluadas ........................................................................................... 26
1.2.6 Análisis estadístico ............................................................................................... 26
1.2 Resultados y discusión ...................................................................................... 26
1.4 Conclusiones ........................................................................................................... 32
Capitulo 2. .................................................................................................................... 37
PERFIL BIOQUÍMICO SANGUÍNEO Y CUADRO HEMÁTICO DEL POLLO DE
ENGORDE BAJO CONDICIONES DE ESTRÉS CALORICO ...................................... 37
2.1 Introducción ............................................................................................................ 37
2.2 Materiales y métodos ............................................................................................. 38
2.2.1 Localización .......................................................................................................... 38
2.2.2 Animales............................................................................................................... 38
2.2.3 Tratamientos ....................................................................................................... 38
2.2.4 Manejo del pollo de Engorde ............................................................................... 38
2.2.5 Muestras.............................................................................................................. 40
2.3 Resultados y discusión ........................................................................................... 42
2.4 Conclusiones .......................................................................................................... 54
Capitulo 3. .................................................................................................................... 60
ANATOMOFISIOLOGIA DEL POLLO DE ENGORDE BAJO CONDICIONES DE
ESTRÉS CALORICO EN EL PIEDEMONTE AMAZÓNICO COLOMBIANO ............... 60
3.1 Introducción ............................................................................................................ 60
3.2 Materiales y métodos ............................................................................................. 61
3.3 Resultados y discusión ........................................................................................... 64
3.4 Conclusiones .......................................................................................................... 74

VII
Lista de figuras pág.
Figura 1: Relación ente la ganancia de Peso (GP) y temperatura ambiental del
Piedemonte Amazónico (TA) .......................................................................................... 67
Figura 2: Nivel de hiperventilación (NH) y la humedad relativa (HR) en función con la
temperatura ambiental del Piedemonte Amazónico ........................................................ 68
Figura 3: Relación entre la temperatura rectal (TR) y la temperatura ambiental del
Piedemonte Amazónico (TA) .......................................................................................... 69
Figura 4: Relación entre la temperatura ambiental (TA) y nivel de hiperventilación (NH)
bajo el Piedemonte Amazónico Colombiano .................................................................. 69
Figura 5: Relación entre la temperatura rectal (TR), humedad relativa (HR) y
temperatura ambiental del Piedemonte Amazónico (TA) ................................................ 70
Figura 6: Variabilidad de la temperatura ambiental bajo el Piedemonte Amazónico
durante diferentes horas del dia ..................................................................................... 71
Figura 7: Efecto de la hora del día sobre la temperatura rectal (TR) bajo el Piedemonte
Amazónico ..................................................................................................................... 72
Figura 8: Efecto de la hora del día sobre el nivel de hiperventilación (NH) bajo el
Piedemonte Amazónico .................................................................................................. 72

VIII
Lista de tablas pág. Tabla 1: Valores de referencia del perfil bioquímico sanguíneo en pollos de engorde de
32 a 47 días ................................................................................................................... 11
Tabla 2: Valores hematológicos para pollos de engorde ................................................ 12
Tabla 3: Composición de la dieta experimental para pollos de engorde de 22 a 42 días 25
Tabla 4: Ganancia de peso (g) en pollos de engorde sometidos a estrés calórico crónico
(30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano de los 22 a 42
días de edad .................................................................................................................. 26
Tabla 5: Valores medios de consumo de alimento acumulado (g/ave) en pollos de
engorde sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el
Piedemonte Amazónico Colombiano de los 22 a 42 días de edad ................................. 28
Tabla 6: Valores medios de conversión alimenticia acumulada (g/g) en pollos de engorde
de 22 a 42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura
ambiental en el Piedemonte Amazónico Colombiano ..................................................... 29
Tabla 7: Valores medios de mortalidad (%) acumulada en pollos de engorde de los 22 a
42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el
Piedemonte Amazónico Colombiano .............................................................................. 31
Tabla 8: Composición de la dieta experimental para pollos de engorde de 22 a 42 días.39
Tabla 9: Valores medios hematológicos (hematocrito, leucograma) de pollos de engorde
a los 42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental
en el Piedemonte Amazónico Colombiano ..................................................................... 42

IX
Tabla 10: Valores medios de hemoglobina (g/dL) en pollos de engorde a los 42 días de
edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el
Piedemonte Amazónico Colombiano .............................................................................. 43
Tabla 11: Valores medios plasmáticos de corticosterona (ng/mL) en pollos de engorde a
los 42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental
en el Piedemonte Amazónico Colombiano ..................................................................... 46
Tabla 12: Valores medios plasmáticos de glucosa (mmol/L) en pollos de engorde a los
42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el
Piedemonte Amazónico Colombiano .............................................................................. 48
Tabla 13: Valores medios plasmáticos de albumina, globulina y proteína total en pollos
de engorde a los 42 días de edad sometidos a estrés calórico crónico (30ºC) y
temperatura ambiental en el Piedemonte Amazónico Colombiano ................................. 49
Tabla 14: Valores medios plasmáticos de cloro, potasio, calcio y magnesio en pollos de
engorde a los 42 días de edad bajo estrés calórico crónico (30ºC) y temperatura
ambiental en el Piedemonte Amazónico Colombiano ..................................................... 50
Tabla 15: Valores medios plasmáticos de sodio y fosforo en pollos de engorde a los 42
días de edad sometidos a estrés calórico (30ºC) y temperatura ambiental en el
Piedemonte Amazónico Colombiano .............................................................................. 52
Tabla 16: Composición de la dieta experimental para pollos de engorde de 22 a 42 días
....................................................................................................................................... 62
Tabla 17: Valores medios de temperatura y humedad relativa experimental ................. 64
Tabla 18: Valores medios de temperatura rectal y nivel de hiperventilación en pollos de
engorde desde el día 22-42 de edad sometidos a estrés calórico crónico (30ºC) y
temperatura ambiental en el Piedemonte Amazónico Colombiano ................................. 65

X
Tabla 19: Valores medios del peso del bazo (PB) y de la bolsa de Fabricio (PF) en pollos
de engorde a los 42 días de edad bajo estrés calórico crónico (30ºC) y temperatura
ambiental en el Piedemonte Amazónico Colombiano ..................................................... 73

XI
Resumen Se realizo un experimento con tres objetivos específicos dirigidos a evaluar el efecto del estrés calórico en el Piedemonte Amazónico Colombiano sobre algunos parámetros fisiológicos y zootécnicos en dos estirpes de pollo de engorde durante 22 a 42 días de edad; se utilizo un diseño experimental irrestrictamente al azar con bajo cuatro tratamientos. T1 pollos macho Cobb-500 (n=125; de 22 días); T3 pollos macho Ross-308 (n=125; de 22 días), bajo temperatura estrés calórico crónico con temperatura media de
30C±1ºC; T2 pollos macho Cobb-500 (n=125; de 22 días) y T4 pollos macho Ross-308 (n=125; de 22 días) sometidos a temperatura ambiental del Piedemonte Amazónico Colombiano. El primer objetivo evaluó la ganancia de peso, consumo de alimento, conversión alimenticia y mortalidad. El segundo objetivo determino el comportamiento del perfil bioquímico sanguíneo, el cuadro hemático y el nivel de corticosterona plasmática. El tercer objetivo fue direccionado a evaluar los parámetros anatomofisiológicos temperatura rectal y nivel de hiperventilación durante los 22 a 42 días de edad; y el peso del bazo y de la bolsa de Fabricio al día 42 de edad. En general no se observaron diferencias (P>0,05) en los resultados de las variables entre ambas estirpes. Sin embargo, se hallaron diferencias (P<0,05) en los resultados de las variables entre la misma línea de pollo bajo las dos temperaturas. Las dos estirpes de aves no mostraron todo su potencial productivo. Los resultados bioquímicos y del leucograma no permitieron explicar claramente los posibles cambios séricos. Se observo una respuesta fisiológica análoga entre las aves Cobb y Ross. El bajo peso registrado en la bolsa de Fabricio y el bazo de las aves sometidas a estrés calórico crónico, sugieren un posible efecto inmunodepresor de esta forma de estrés. Palabras clave: Estrés, estirpes, zootécnicos, perfil bioquímico, hemático, anatomofisiológicos.

XII
Abstract
An experiment was conducted with three specific objectives aimed at evaluating the effect of heat stress in the Colombian Amazon Piedmont on some physiological and zootechnical parameters in two strains of broiler for 22 to 42 days of age, was used an experimental design randomly unconditionally with under four treatments. T1 male broiler Cobb-500 (n = 125; of 22 days), T3 male broiler Ross-308 (n = 125, 22 days), under
temperature of chronic stress with constant average temperature of 30C±1ºC, T2 male broiler Cobb-500 (n = 125; of 22 days), T4 male broiler Ross 308 (n = 125, 22 days) under ambient temperature of the Colombian Amazon Piedmont. The first objective sought to compare, body weight gain, feed intake, feed conversion and mortality. The second objective was aimed at assessing the blood chemistry profile, the complete blood count and plasma corticosterone level at 42 days of age. The third objective was oriented to assess the rectal temperature, level of hyperventilation for 22 to 42 days of age, and weight of spleen and bursa of Fabricius at day 42 of age. In general that there was not difference (P>0,05) in the results of all variables between both strains of broilers under any of the two experimental temperatures. However, significant differences were obtained (P<0,05) in the results of the variables between the same strain under both experimental temperatures. The two strains of broilers did not show their full productive potential. The biochemical and leukogram results did not allow to clearly explain any serum changes. Was observed similar physiological responses in Cobb and Ross broilers. The underweight of the bursa of Fabricius and spleen registered in broilers subjected to chronic heat stress, suggest a possible immunosupressor effect of this form of stress. Keywords: Stress, strains, animal, biochemical profile, blood count, anatomophysiological.

1
Introducción
La avicultura industrial colombiana ha crecido en la última década a un ritmo del 8,5% promedio anual, alcanzando las 1.075.136 toneladas de carne de pollo en el 2011 con un crecimiento del 3,8%, favorecida por el aumento en el consumo per cápita de la población, debido al precio competitivo de la carne de ave frente a la de bovino (FENAVI, 2011). En la región del Piedemonte Amazónico, la producción avícola está ampliando sus fronteras al convertirse en actividad económica alternativa a la producción bovina, como fuente de ingresos y de proteína para la dieta de la población; es así como en el Departamento del Caquetá, a pesar de las condiciones ambientales de altas temperaturas y humedad relativa propicias para la incidencia de estrés calórico en las aves, la población avícola anual de pollo de engorde asciende a un promedio de 1000.000 aves (FENAVI, 2011), criados en los sistemas de confinamiento ya sea de autoconsumo o tecnificado. Los avances tecnológicos, especialmente en genética y nutrición permiten que el pollo de engorde crezca rápidamente, lo que determina aumento en la demanda sanguínea, debido a la mayor tasa metabólica. Desafortunadamente, el sistema cardiorrespiratorio es ineficiente para oxigenar la abundante masa muscular depositada por el pollo moderno, ya que el mejoramiento genético de este sistema no ha acompañado a la evolución de la ganancia de peso (Macari et al., 2004). Paralelo a los avances alcanzados, la avicultura sufre hoy mayor incidencia de problemas sanitarios, mayor susceptibilidad al estrés con reducción en la calidad de la carne y aparición de defectos en el color y la textura (Akit et al., 2005), debido entre otras causas a que la capacidad termorreguladora de estos animales continúa deficiente para enfrentar las temperaturas y humedades relativas altas (Mitchel, 2001). Las estirpes de pollo moderno disponibles en el mercado han sido genéticamente mejoradas para las condiciones y exigencias de países templados. La producción a escala comercial de estos linajes en países tropicales y semitropicales ha generado la necesidad de reevaluar tanto las prácticas de manejo, así como los requerimientos nutricionales, con el fin de alcanzar su máximo rendimiento, bajo condiciones de temperatura y humedad relativa altas. En la región del Piedemonte Amazónico las altas temperaturas prevalecen en gran parte del año, para que las aves puedan expresar su potencial genético de rendimiento, se torna relevante el desarrollo de investigaciones encaminadas a generar conocimiento científico en la producción de las aves criadas bajo estas condiciones de estrés calórico.

2
1. Objetivos
1.1 Objetivo general Evaluar la respuesta fisiológica y de producción del pollo de engorde comercial moderno sometido a las condiciones de temperatura ambiental y de humedad relativa del Piedemonte Amazónico Colombiano
1.2 Objetivos específicos Evaluar los índices zootécnicos del pollo de engorde moderno criado bajo condiciones ambientales del Piedemonte Amazónico Colombiano Determinar el comportamiento del perfil bioquímico sanguíneo y el cuadro hemático del pollo de engorde moderno sometido a condiciones ambientales del Piedemonte Amazónico Colombiano Evaluar el efecto de las condiciones ambientales del Piedemonte Amazónico Colombiano sobre algunos parámetros anatomofisiológicos de estirpes comerciales de pollo de engorde moderno

3
2. Variables evaluadas
2.1 Variables zootécnicas
Ganancia de peso Consumo de alimento Conversión alimenticia Mortalidad
2.2 Variables anatomofisiológicas
Niveles de corticosterona plasmática Cuadro hemático (Hemoglobina, hematocrito, recuento leucocitario y leucograma diferencial) Perfil bioquímico sanguíneo (glucosa, proteína total, albúmina, globulina, sodio, cloro, potasio, calcio, magnesio y fósforo) Temperatura rectal Nivel de hiperventilación Peso del bazo y bolsa de Fabricio

4
3. Marco teórico
3.1 Ambiente, temperatura ambiental y humedad relativa ambiental El ambiente al que se someten las aves, definido como la sumatoria de impactos biológicos y físicos, es uno de los principales aspectos del que depende el éxito o el fracaso en la producción avícola. Entre los factores ambientales, las condiciones de temperatura, humedad relativa y movimiento del aire, son los factores que afectan directamente a las aves, pues estos comprometen el mantenimiento de la homotermia (Tinôco, 2001). A la temperatura como aspecto físico se le atribuye un papel importante ya que en la avicultura industrial las aves están confinadas, situación que puede interferir para realizar ajustes comportamentales necesarios para el mantenimiento de la homeostasis térmica (Macari et al., 2004). La respuesta a los cambios de temperatura ambiente está relacionada con la presentación de alteraciones metabólicas, aunque las variaciones de temperatura ambiente no superiores a 6°C no influyen en el rendimiento de los pollos, aquellas por encima de 10°C si interfieren (Macari y Gonzales, 1990). Cuando la temperatura ambiente se aproxima a los 42°C, o sea la temperatura corporal del pollo, la pérdida de calor latente se realiza mediante la respiración jadeante; sin embargo, para que este mecanismo de liberación de calor corporal sea eficiente, la humedad relativa del aire debe ser inferior al 70% (Lasiewski et al., 1966); condición difícil de lograr en las áreas tropicales, en las cuales siempre la humedad relativa ambiental supera ampliamente ese porcentaje. Los pollos criados bajo condiciones de temperaturas y humedad relativa altas no consiguen respirar lo suficientemente rápido para eliminar todo el calor que necesitan remover de su cuerpo; consecuentemente, el ave sufre un cambio térmico corporal, que puede generar postración y muerte, cuando la temperatura alcanza los 47°C, límite máximo fisiológico vital del animal (Mitchell, 1976; Nääs, 1994; Rutz, 1994). La situación anterior, es más preocupante en el pollo de engorde adulto, pues el área superficial necesaria para la disipación de calor disminuye, proporcionalmente con la edad y con el peso corporal. Del mismo modo, factores como cobertura de las plumas, el sexo, el grado de aclimatación de las aves pueden interactuar con la humedad relativa y determinar la respuesta de éstas a las altas temperaturas ambientales (Balnave, 2004). Según Yahav et al., (1995) el rendimiento de los pollos de engorde es mejor cuando están expuestos a humedades relativas entre 60 a 65%, aunque la temperatura ambiental alcance los 35°C. El confort térmico o zona termoneutral que varía en función de la edad y el peso del animal se lo define como la zona de temperatura ambiente donde el gasto metabólico es mínimo y la homotermia es mantenida con menos gasto energético, y por lo tanto, la cantidad de energía utilizada para la termogénesis es mínima y la energía utilizada para

5
la producción es máxima. Así, en pollitos de 1 a 7 días de edad la zona de confort está entre 31 y 33°C, disminuyendo hasta 21 a 23°C a la edad de 35 a 42 días, con una humedad relativa ambiental entre 65 y 70% (Furlan y Macari, 2002). Por lo anterior, en condiciones de confort térmico se presenta una buena regulación de la homeostasis, subsecuentemente se presenta un eficiente gasto de energía, generando mejor rendimiento en los parámetros ganancia de peso, conversión alimenticia y producción de huevos. A medida que la temperatura ambiente y la humedad relativa se elevan por encima de la zona termoneutral, disminuye la capacidad de disipación de calor de las aves. En consecuencia, la temperatura corporal del ave se incrementa y posteriormente aparecen los síntomas de estrés por calor, que se manifiestan inicialmente con la disminución del consumo de alimentos y subsecuentemente con la reducción de los substratos metabólicos o combustibles disponibles para el metabolismo, logrando de esta forma minimizar la producción de calor metabólico (Belay y Teeter, 1993).
3.2 Estrés El estado de estrés implica una necesidad de energía, por que caracteriza una gran actividad a nivel del sistema neuromuscular y demás tejidos. Un aumento basal del tono muscular esquelético surge luego del inicio, debido a que las catecolaminas, preparan al individuo para posibles actividades físicas de respuesta, lo que equivale a mantener un estado de alerta ante agentes que alteran el estado fisiológico normal del animal. Entre los agentes capaces de llevar los animales a un estado caracterizado como estrés se encuentran los de naturaleza mecánica (traumatismo, quirúrgico, lucha), física (calor, frio, ruido), química (medicamentos para tratamiento de enfermedades, promotores de crecimiento), biológica (estado nutricional y agentes patológicos), psicológica (cambio de ambiente y prácticas de manejo) y estresores de origen social, como jerarquía y dominancia entre los mismos animales (Baccari, 1998). El ave expuesta a un agente estresor activa su sistema nervioso simpático, lo cual conlleva a un incremento de las frecuencias respiratoria y cardiaca con redistribución de la irrigación sanguínea para los órganos centrales, preparando al organismo para la “fuga”. En pocos segundos, estos efectos son potencializados y prolongados por la liberación de epinefrina (adrenalina) y norepinefrina (noradrenalina) en la corriente sanguínea por la glándula adrenal. Posteriormente, esta misma glándula también libera corticosterona (hormona del estrés), la cual busca aumentar o suprimir la energía del organismo. Aunque estas respuestas son benéficas cuando son por un corto período de tiempo, permitiendo que el animal responda al agente estresor, la secreción de corticosterona asociada a un estrés persistente genera diversos efectos perjudiciales en el animal, entre los que se encuentran los disturbios en el sistema inmune, las ulceras gástricas y las alteraciones en la secreción de hormonas que regulan el crecimiento y la reproducción, (Elrom, 2000; Mench, 2002). Asimismo, el organismo demanda mayor cantidad de glucosa y otros nutrientes, pues el sistema nervioso es dependiente directo de la glucosa sanguínea, aminoácidos, sales y vitaminas, indispensables para sustentar el aumento de actividad (Cabral et al., 1997).

6
Dentro de la producción de pollo de engorde se encuentran factores estresantes como la alta densidad de animales por metro cuadrado a partir de los 22 días de edad (Costa, 2002); el manejo inadecuado previo al sacrificio de los animales (Bressan et al., 2002, citados por Bressan et al., 2003; Mitchell et al., 1992), y las situaciones de estrés térmico antes del sacrificio de los animales que afectan el metabolismo post mortem y las características de la carne que la predisponen a la incidencia de lesiones tipo Pálida, Blanda y Exudativa; y Oscura, Dura y Seca (Sayre et al., 1963; Marple y Cassens, 1973, citados por Bressan et al., 2003).
3.3 Estrés calórico Las aves son animales homeotermos que disponen de un centro termorregulador localizado en el hipotálamo capaz de controlar la temperatura corporal a través de mecanismos fisiológicos y respuestas del comportamiento que regulan la producción y liberación Las aves son animales homeotermos que disponen de un centro termorregulador localizado en el hipotálamo capaz de controlar la temperatura corporal a través de mecanismos fisiológicos de calor (Macari et al., 1994). Sin embargo ante el incremento de la humedad relativa y la temperatura ambiental por encima de la zona de confort, las aves reducen ostensiblemente su habilidad para disipar el calor (Teeter y Smith, 1985). La vasodilatación periférica, el aumento en la pérdida de calor no evaporativo con aumento de la producción de orina, el extender las alas rectando las plumas e intensificando la vasodilatación periférica, el aumento de la frecuencia respiratoria, resultando en pérdidas excesivas de dióxido de carbono (CO2), son algunas de las respuestas compensatorias de las aves frente al estrés calórico. Otra respuesta fisiológica es la disminución de la presión parcial de CO2 (pCO2), aumentando la concentración de acido carbónico (H2C-O3) e hidrógeno (H+), en donde como respuesta los riñones aumentan la excreción de HCO3 y reducen la excreción de (H+) intentando mantener el equilibrio acido-base del ave; para evitar la alteración del equilibrio acido-base denominada alcalosis respiratoria (Macari et al., 1994). Los avances tecnológicos, especialmente en genética y nutrición, han tenido efecto en la alta tasa de crecimiento del pollo de engorde moderno, lo que determina un aumento en la demanda sanguínea residual, debido al mayor requerimiento metabólico. No obstante esta evolución, la capacidad del sistema cardiorrespiratorio no ha tenido el mismo ritmo de mejoramiento, ocasionando deficiencias en los proceso de oxigenación de toda la masa muscular, que dan origen a trastornos en diversos órganos (Macari et al., 2004), situación acentuada en pollos adultos (Howlinder y Rose, 1987). Estas adaptaciones físicas y metabólicas que sufren los pollos para sobrevivir ante el estrés calórico, dan como consecuencia la disminución del consumo y peor eficiencia alimenticia, Deaton et al., (1968). El estrés por calor inicia cuando la temperatura ambiental sobrepasa los 25°C, si el ave está aclimatada a temperaturas más bajas. Sin embargo, aves adultas pueden responder eficientemente a temperaturas ambientales de hasta 32°C, cuando han alcanzado tolerancia al calor debido a la aclimatación (Arjona et al., 1988). Del mismo modo, las aves se adaptan mejor a una temperatura diaria alta cuando la temperatura en las

7
noches cae a 25°C o menos, ya que durante este período pueden recuperarse del estrés sufrido durante el día (Balnave, 2004). Los nutrientes de la dieta también han sido estudiados como atenuantes o agravantes de la respuesta de las aves al estrés calórico; se destacan la comparación entre diferentes fuentes de metionina, las proporciones entre Arg:Lis (Chen et al., 2003; Balnave et al., 1999), el porcentaje de grasa en la dieta (Dale y Fuller, 1980).
3.4 Efectos del estrés calórico en el consumo de alimento y agua Czarick y Tison (1990); Morgan (1990); Austic (1985); Howlider y Rose (1987); Bonnet et al., (1997) y Plavnik y Yahav (1998) observaron que la primera respuesta del ave al estrés por calor es la disminución en el consumo de alimento, seguida de la pérdida de agua en el organismo, la cual conlleva a un cuadro de deshidratación, y subsecuentemente disminución de la producción (Mardsen y Morris, 1975). Según Payne (1966) y Davis et al., (1973), las aves de postura disminuyen la producción de huevo entre 1,1% y 1,6% por cada grado centígrado que aumenta la temperatura del ambiente, situación que se agrava cuando la dieta ingerida da origen a una alta producción de calor metabólico; al respecto, Van kampen (1977), citado por Lesson (1986) observó que la temperatura corporal de aves criadas a una temperatura ambiente de 20°C aumenta en un 12% a las dos horas después de haber sido alimentadas; mientras que a 35°C la respuesta termogénica se manifesta a las cuatro horas pospandrial y con un incrementó de solo el 7%. Por lo anterior, Teeter et al., (1984) recomiendan no alimentar las aves durante periodos de alta temperatura ambiental en el que la producción de calor corporal no puede ser disipada y pueda llevar el animal a la muerte. La condición de estrés por calor en el ave también tiene influencia sobre el consumo de agua; es así como; el volumen de agua consumido puede duplicarse bajo estas circunstancias adversas (Fox, 1980). Según Bonnet et al., (1997) esto se debe a que el agua cumple un papel fundamental en los mecanismos de pérdida de calor, a través del proceso evaporativo respiratorio el cual también se altera bajo condiciones de estrés térmico (Macari, 1996), en donde las aves aumentan la tasa respiratoria de 25 a 250 movimientos respiratorios por minuto (Linsleey y Berger, 1964).
3.5 La corticosterona La hormona esteroidal corticosterona es liberada por la corteza suprarrenal con mediación del hipotálamo y la glándula pituitaria. A través de las vías neurales y endocrinas del sistema nervioso central es estimulada la producción del factor liberador de corticotropina (FLC) de las neuronas, en la eminencia media del hipotálamo, mediante el sistema vascular portal, para ser transportado a la glándula pituitaria, la cual estimula la producción de la hormona corticotropina (ACTH). Liberada la ACTH en el sistema circulatorio se transporta al tejido blanco principal y estimula a la corteza

8
suprarrenal a producir y liberar todas las hormonas secretadas en este órgano, destacándose entre ellas la corticosterona y la aldosterona (Eden,1978). El estrés por calor en las aves estimula la liberación de la corticosterona (Holmes y Phillips, 1976; Eden, 1978), con aumento en la concentración de corticosterona plasmática por períodos cortos, para hacer frente a la exposición aguda a las altas temperaturas (Edens y Siegel, 1975; Ben Nathan et al., 1976; El Halawani et al., 1976; Pilo et al., 1985). No obstante, la exposición continua de las aves a temperaturas altas, hace que la concentración plasmática de corticosterona disminuya después del pico inicial (Sammelwitz, 1967), acompañada por la disminución en los niveles plasmáticos de glucosa, fosforo, sodio y un pH plasmático elevado; con insuficiencia suprarrenal cortical, secreción masiva de catecolaminas, alteraciones cardiovasculares (Eden, 1978) y, a menos que otras alteraciones funcionales o las respuestas de comportamiento puedan ser implementadas para aliviar el estrés térmico, los animales pueden sufrir hipertermia y morir (Edens y Siegel, 1975; Pilo et al., 1985). La administración de corticosteroides y ACTH aumenta la concentración plasmática hormonal, disminuye el tamaño de los tejidos linfoides bazo, timo y bolsa de Fabricio (Garren y Shaffner, 1956; Glick,1967; Siegel y Beane, 1961) con disminución en el número de linfocitos circulantes y aumento en el número de neutrófilos o heterófilos (Dougherty y White, 1944; Gross y Siegel, 1983). Los corticosteroides se unen a receptores específicos en el citoplasma de las células linfáticas para reorientar la diferenciación y el metabolismo de este tipo de células (Thompson y Lippman, 1974; Sullivan y Wira,1979). La administración de ACTH y la exposición a altas temperaturas aumenta la unión de los corticosteroides a las células del sistema linfoide. El efecto de los corticosteroides en este tipo de células depende de la gravedad, persistencia del estrés y el rol de las células afectadas, tanto en la inmunidad celular así como en la inmunidad humoral (Gould y Siegel, 1981). Burger y Lorenz, 1960; Sammelwitz, 1967; Edens y Siegel, 1976 sostienen que la medicación con hidrocortisona disminuye la mortalidad en pollos de engorde expuestos a estrés calórico. Del mismo modo Gould y Siegel (1981) encontraron que el aumento en los niveles plasmáticos de aldosterona actúa en concierto con la hormona antidiurética para promover la absorción renal de agua, y así prevenir la deshidratación durante el enfriamiento evaporativo utilizado en la termorregulación.
3.6 Efectos del estrés calórico en los órganos Durante el crecimiento normal de los pollos, el tejido muscular presenta mayor desarrollo, seguido por la formación de tejido adiposo. Cuando la tasa de crecimiento es baja, ocurre un acondicionamiento de los tejidos, donde algunos responden más rápidamente que otros (viscerales > adiposo > muscular) como resultado de alteraciones endocrinas (Hornick et al., 2000). Esta característica hace que en condiciones de subnutrición post estrés calórico, los tejidos viscerales posean mayor capacidad de reducir su tamaño, y por ende, disminuir su actividad metabólica de manera más efectiva, que los tejidos de la carcasa (Lawrence y Fowler, 2002).

9
El estrés calórico en los pollos de engorde disminuye los pesos relativos y absolutos de los intestinos (Ribeiro et al., 2001a) y el peso relativo del corazón (Plavnik y Hurwitz, 1985). Teniendo en cuenta que durante los períodos de estrés calórico las aves restringen el consumo de alimento; Buhr et al., (1998); Zubair y Leeson (1994) concluyeron que en periodos largos de restricción alimentaria, el peso de las vísceras y del hígado disminuyen proporcionalmente con la pérdida de peso de los animales. Entretanto, Susbilla et al., (1994) no observaron efecto de la restricción de alimento sobre los pesos relativos del hígado, corazón, riñones y pulmones. Por el contrario Cattelan (1995) observó mayor peso del hígado en aves bajo restricción de alimento.
3.7 Efectos del estrés calórico en los órganos linfoides El peso proporcional de los órganos linfoides primarios y su histología son frecuentemente referenciados para evaluar la respuesta frente a los cuadros de estrés. Rosales et al., (1989) mencionan 0,18 y 0,20 como índices morfométricos bursales (Rbo) normales en pollos de engorde de cuatro a siete semanas de edad y de 0,44 en pollos de la misma edad libres de patógenos específicos, mantenidos en condiciones de laboratorio. Ya valores inferiores de 0,15% se consideran muy bajos e indican atrofia e inmunosupresión (Kuney et al., 1981, citados por Revidatti et al., 2002). La hipertrofia adrenal coexiste con una involución del sistema linfoide, inclusive con atrofia del páncreas, del bazo, de la bolsa y del timo e inmunosupresión prolongada, siendo más sensibles las aves de mayor tamaño y con mayor tasa de crecimiento, (Revidatti et al., 2002). Guimarães et al., (2003), afirman que el estrés producido por la temperatura ambiental por encima o por debajo de la zona termoneutral, tiene efecto directo sobre la bolsa de Fabricio con retraso en el desarrollo y maduración, disminución de la cantidad de parénquima y aumento del índice de apoptosis de los linfocitos bursales. La aplicación de corticosterona en pollos de cinco semanas de edad disminuye el peso relativo del timo entre el 65% y 71%, de la bolsa entre el 43% y 57% y del bazo entre el 27% y 35% (Donker y Beuving, 1989; Puvadolpirod y Thaxton, 2000)
3.8 Cambios en parámetros sanguíneos y bioquímicos debidos al estrés calórico
El sistema sanguíneo es particularmente sensible a los cambios de temperatura y se constituye en un importante indicador de las respuestas fisiológicas de las aves a los agentes estresores. Alteraciones cuantitativas y morfológicas en la células sanguíneas son asociadas al estrés por calor, manifiestas por variaciones en los valores de hematocrito, número de leucocitos circulantes, conteo de eritrocitos, y cantidad de hemoglobina en el eritrocito (Borges et al., 2003).

10
El número de leucocitos en la sangre de los pollos se encuentra entre 2000 a 3000 células /μL, número que varía en función del sexo, la edad, las condiciones de estrés y de enfermedad (Noriega, 2000a); es así como pollos bajo condiciones de estrés desencadenan la liberación de la hormona adrenocorticotropina (ACTH), determinando una disminución en la cantidad de linfocitos circulantes, aumento de la relación heterofilo: linfocito e incremento en la cantidad de heterofilos en la circulación (Macari y Luquetti, 2002) y aumento a la susceptibilidad a infecciones virales (Gross et al., 1980). El estrés puede ocasionar inmunosupresión o inmunoestimulación en el ave dependiendo de la intensidad y duración del estimulo estresor, del linaje y de la respuesta individual (Morgulis, 2002). El estrés afecta el conteo relativo y absoluto de células de la serie blanca (Gross y Siegel, 1983), aumenta el número de leucocitos totales (Coles, 1986; Ruckebush et al., 1994; Cunningham, 1999) con variación de la proporción de los componentes celulares (Bogin et al., 1981 y Maxwell et al., 1992). Leucocitosis baja a moderada, acompañada de heterofilia y linfopenia, puede ser el resultado de la presencia de glucocorticoides exógenos o endógenos producto del estrés (Campbell, 1994; Perea et al., 1997), con el consecuente aumento de heterófilos (H) y la reducción de linfocitos (L). Borges (2001) también destaca el aumento en la concentración de glucosa en respuesta directa a una mayor secreción de adrenalina y glucocorticoides, cuando las aves están bajo condiciones de estrés. Maxwell et al., (1992) sostienen que la heterofilia en los pollos de engorde es una respuesta para medir un estrés moderado, mientras que la basofilia el estrés severo. Revidatti et al., (2002) observaron disminución en la tasa de linfocitos en pollos machos expuestos durante cuatro horas a temperatura ambiental de 40°C, con humedad relativa de 60%, como respuesta al estrés agudo; mientras que Scope et al., (2002) observaron alteración en la relación heterófilos–linfocitos en aves estresadas durante el trasporte, pero no vislumbraron cambios sobre el número de basófilos, monocitos, eosinófilos, proteínas totales ni el hematocrito. Hasta el desarrollo de la técnica de dosificación de corticosterona plasmática, la relación H/L fue considerada por los investigadores una de las variables más importantes para evaluar el estrés en aves (Satterlee et al., 1980; Gross y Siegel, 1983). Sin embargo, el aumento de esta relación H/L solo ocurre en la fase inicial, pues bajo estrés crónico, esta relación no presenta alteraciones significativas (Gross y Siegel, 1983; McFarlane y Curtis, 1989). El volumen globular en el pollo de engorde está entre el 28% y 30% con 11,5 g/dL de hemoglobina (Furlan et al., 1999; Macari y Luquetti, 2002). Según Sturkie y Griminger (1986), los valores del hematocrito para pollo de engorde están entre 29 a 48% y 25,5 a 31% para machos y hembras, respectivamente, mientras que valores por encima de este intervalo pueden ser indicativos de deshidratación o policitemia (Campbell, 1994). Del mismo modo, índices bajos del hematocrito pueden ser indicativos de alteraciones agudas crónicas, septicemias, hemorragias y presencia de anemia (Campbell y Dein, 1984; Noriega, 2000a y Sturkie, 1976). Otros metabolitos a tener en cuenta en el plasma sanguíneo son las proteínas plasmáticas albuminas, globulinas y fibrinógeno. La concentración de proteína total se encuentra disminuida en insuficiencias hepáticas, trastornos intestinales y renales,

11
hemorragias o deficiencias de alimentación. La albumina representa el 50% del total de las proteínas, es sintetizada en el hígado y contribuye con el 80% de la osmolaridad del plasma sanguíneo, también cumple función importante en la regulación del pH sanguíneo, actuando como anión. La concentración de albúmina plasmática (hiperalbuminemia) aumenta en los cuadros de deshidratación y puede disminuir (hipoalbuminemia) en situaciones de daño hepático crónico, deficiencias de fuentes proteicas y parasitismo. Existe una correlación negativa entre la concentración de albúmina y globulinas; esta última mide el grado de adaptación al estrés, siendo que animales adaptados tienden a tener niveles normales, mientras que, los no adaptados niveles elevados (González y Silva, 2003). Otro metabolito vital para las necesidades energéticas del organismo es la concentración de glucosa sanguínea, la cual puede aumentar durante el estrés crónico (González y Silva, 2003). La concentración normal de glucosa sanguínea en las aves está entre 11,10 a 22,2 mmol/L. Cuando los valores de la glicemia están disminuidos, generalmente indican la presentación de enfermedades hepáticas, inanición o desnutrición (Noriega, 2000b) y fallas en la homeostasis, como ocurre en alteraciones metabólicas como la cetosis. Revidatti et al., (2002) al realizar análisis bioquímicos en sueros sanguíneos de pollos sometidos a estrés por manipulación física, no encontraron diferencia en las fracciones de proteínas plasmáticas, triglicéridos, colesterol y glicemia de las aves sometidas a los diferentes tratamientos; y concluyen que la hiperglicemia se relaciona con los cuadros agudos de estrés en los cuales el estresor actúa en forma súbita. Por el contrario, el comportamiento de la hipoglicemia es menos claro cuando se trata de estresores crónicos en los cuales influyen interacciones como el metabolismo lipídico (Puvadolpirod y Thaxton, 2000) o la disponibilidad de otros recursos energéticos. Tabla 1: Valores de referencia del perfil bioquímico sanguíneo en pollos de engorde de 32 a 47 días
Fuente: González et al., (2001) La exposición de pollo de engorde durante seis horas a estrés calórico con temperatura promedio de 35°C altera los índices sanguíneos aumentando el hematocrito de 26,1% a 28,1% y el de hemoglobina de 10,34 a 9,77 g/dL (Yahav et al., 1997). Según (Macari y Luquetti, 2002); Zhou et al., (1998), los cambios en la viscosidad sanguínea, dependientes de la luminosidad, la temperatura ambiente y la disponibilidad del agua, alteran la resistencia periférica y de esa manera la distribución de la sangre en los tejidos.
PARAMETRO (UNIDAD) EDAD (días)
32 47
Proteína total (g/dL) 29,7 35,9 Albumina (g/dL) 15,1 16,6 Globulina (g/dL) 15,6 21,6 Glucosa (g/dL) 14,4 13,5

12
Los valores hematológicos (Tabla: 2) de referencia en pollos de engorde cambian con la edad del pollo y el sitio de crianza, Latimer y Bienzle (2000). Tabla 2: Valores hematológicos para pollos de engorde
Hemograma Intervalo
Eritrocitos (millones/mm3) 2,5 a 3,5 Hemoglobina (g/dL) 7 a 13 Hematocrito (%) 22 a 35
Leucograma células/mm3
Leucocitos Totales 12000 a 30000 Heterófilos (Bastonados) Raros Heterófilos 3000 a 6000 Eosinófilos 0 a 1000 Basófilos raros Monocitos 150 a 2000 Linfocitos 7000 a 17500
Fuente: Latimer y Bienzle (2000)
3.9 Los electrolitos y ph durante el estrés calórico Los electrolitos son sustancias químicas, que se disocian en sus constituyentes iónicos. El sodio (Na+), el potasio (K+) y el cloro (Cl-) son los iones fundamentales en el mantenimiento de la presión osmótica y el equilibrio acido-base de los fluidos corporales (Mongin, 1981). El K+ es el principal catión de fluido intracelular, entre tanto el Na+ y Cl- son los principales iones del fluido extracelular. La osmoregulación se consigue por la homeostasia de estos iones intra y extracelulares. Consecuentemente cambios en la homeostasis del K+ pueden afectar las fluctuaciones celulares. Estudios con deshidratación térmica, seguidos de rehidratación demostraron que el grado de déficit de agua en el fluido intracelular se asocia a la pérdida de K+ intracelular y el déficit de fluido extracelular se relaciona con la pérdida de Na+ plasmático (Nose et al., 1988). La alcalosis respiratoria en mamíferos, provoca reducción de competencia entre el H+ y K+ para la excreción urinaria, por lo tanto, aumenta la pérdida de K+ en la orina. El exceso de iones de potasio es competencia de los aniones tampones del liquido tubular renal, impidiendo la remoción de H+, siendo este reabsorbido, pudiendo generar una acidosis (Bacila, 2003). Tal mecanismo, cuando se presenta en pollos, puede aumentar la necesidad de K+ durante el periodo de estrés calórico (Cunningham, 1999). El estrés calórico disminuye las concentraciones de Na+ y K+ en el plasma sanguíneo de las aves (Borges, 1997), mientras que la concentración de Cl- aumenta (Belay y Teeter, 1993). El aumento del Cl- deprime la excreción de H+ y la reabsorción de HCO3 por los riñones, lo cual contribuye con acidificación de la sangre, respuesta apropiada a la alcalosis fruto del estrés. Resultados opuestos fueron observados por Salvador et al.,

13
(1999) quienes encontraron reducción de los niveles séricos de Cl- al someter pollos de engorde de 42 días de edad a estrés crónico durante una semana. La excreción de K+ depende de factores hormonales, del equilibrio acido- base y el balance de cationes. La tasa de excreción de K+ a través de la orina es variable, está ligada a la concentración plasmática de Na+ y al estado de hidratación del ave. El aumento en la ingestión de K+ genera mayor pérdida urinaria, ya que el ave tiene poca capacidad de conservar el K+ corporal. El nivel sérico de K+ disminuye durante el estrés crónico debido al aumento en la excreción de este ion y al aumento del K+ intracelular comúnmente encontrado durante el estrés agudo (Borges, 1997; Salvador et al., 1999). Además del K, Na y Cl, otros cationes y aniones también participan en el equilibrio acido-base entre ellos el Mg, S, P, Ca, con mayor potencial eléctrico que el Fe, Mn, Zn, Cu, Se, Mo, Co y I. Estos elementos trazas tienen capacidad de funcionar como electrolitos estando presentes en pequeñas cantidades en las raciones y en bajas concentraciones en los tejidos de las aves. El impacto de la relación catión/anión sobre el equilibrio acido- base, pH sanguíneo y tasa de crecimiento del pollo de engorde, fue evaluado por Hurwitz et al., (1973), quienes observaron máxima tasa de crecimiento con pH sanguíneo de 7,28. Teeter et al., (1985) observaron que durante periodos de jadeo bajo estrés calórico, los valores de pH sanguíneo superiores a 7,25 deprimen la tasa de crecimiento y la eficiencia alimenticia. Los aumentos en el pH sanguíneo pueden ser el resultado de la disminución en la tasa respiratoria de las aves, que ocurre con temperaturas de aclimatación, mientras que durante el estrés agudo, la tasa respiratoria de las aves puede ser reducida y la eficiencia de la pérdida de calor puede ser mejorada por el mantenimiento de una alta ingestión de agua (Belay y Teeter, 1993). La exposición de pollos de engorde a una temperatura aérea elevada genera alcalosis respiratoria, llevando a un detrimento del rendimiento zootécnico (Borges, 1997). La alteraciones en el equilibrio acido–base como consecuencia del estrés calórico aunados a desequilibrios en la suplementación de Na+, K+, Cl-, causan inapetencia con reducción en la ganancia de peso, detrimento de la conversión alimenticia y aumentos de la mortalidad (Mongin, 1981).

14
Bibliografia
Akit, M.; Yalcin, S.; Ozkan, S.; Metin, K.; Ozdemin. D. Effects of temperature during rearing and crating on stress parameters and meat quality of broilers. Poultry Science. 85: p.1867-1874. 2005. Arjona, A. A.; Denbow, D.M. ; Weaver Jr, WD. Effect of heat stress early in life on mortality of broilers exposed to high environmental temperatures just prior to marketing. Ph. D. Department of Poultry Science, Virginia Polytechnic Institute and State University,1988. 54-58p. Austic, R. E. Feeding poultry in hot and cold climates. In: Stress Physiology in Livestock. Fla. USA: PressBoca Ratón.1985. Bacila, M. Bioquímica veterinária. 2.ed.São Paulo: Brasil: Varela, 2003. 211p. Balnave, D.; Hayat, J.; Brake, J. Dietary arginine: lysine ratio and methionine activity at elevated environmental temperatures. J. Appl. Poult. Res., Athens, v. 8: p.1–9, 1999. Balnave, D. Challenges of accurately defining the nutrient requeriments of heat-stressed poultry. Poult. Sci., Champaign, v. 83: p.5-14. 2004. Baccari, F. J. Manejo ambiental para produção de leite em climas quentes. In: Congresso Brasileiro de Biometeorologia. Ph. D. Campiña Grande Brasil: Goiânia SBBIOMET; 1998.p.32 Belay, T.; Teeter, R.G. Broiler water balance and thermobalance during thermoneutral and high ambient temperature exposure. Poult. Sci., Champaign. V. 72(2):p.116-124, 1993. Ben, N,D. ; Heller, E.D. ; Perek, M. 1976. The effect of short heat stress upon leucocyte count, plasma corticosterone level, plasma and leucocyte ascorbic acid content. British Poultry Science. 17: p. 481–485, 1976. Bonnet, S.; Geraert, P.A.; Lessire, M.; Carre, B.; Guillaumin, S. Effect of high ambient temperature on feed digestibility in broilers. Poult. Sci., Champaign, v. 76(6): p.857–863,1997. Bogin, E.; Weisman, Y.; Friedman, Y. The effect of heat stress on the levels of certain blood constituents in chickens. Refuah Vet Israel, v. 38(1): p.98-104, 1981. Borges, S.A. Suplementação de cloreto de potássio e bicarbonato de sódio para frangos de corte durante o verão. Jaboticabal. PH D. Unesp Dissertação Mestrado. Programa de Pós-Graduação em Zootecnia, Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal, Brasil,1997.

15
Borges, S.A. Balanço eletrolítico e sua interrelação com o equilibrio ácido-base em frangos de corte submetidos a estresse calórico. Jaboticabal. Unesp Tese Doutorado. Programa de Pós- Graduação em Zootecnia, Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Brasil, 2001. Borges, S.A.; Maiorka, A.; Silva, A.V.F. Fisiologia do estresse calórico e a utilização de eletrólitos em frangos de corte”. Ciênc. Rural, Santa Maria, v. 33(5): p.975-981, 2003. Bressan, C.; Ferrão, S.P.B.; Araújo, L.C.; Ferreira, M.W. Cómo diminuir o estresse causado pela apanha, transporte e abate visando o bemestar de frangos. In: Conferência Apinco de Ciência e Tecnologia Avícolas, Campinas. Anais. Campinas: FACTA. 1: p.255-268, 2003. Buhr, R.J.; Northcutt, J.K.; Lyon, C. E.; Rowland, G. N. Influence of time off feed on broiler viscera weight, diameter and shear. Poult. Sci., Champaign. v. 77(5): p.758-764,1998. Burger, R.E.; Lorenz, F.W. Pharmacologically induced resistance to heat shock. II. Modifications of activity of the central-nervous and endocrine systems. Poultry Science. 39: p.477–482, 1960. Cabral, A.P.T.; Luna, J.F.; Souza, K. N.; Macedo, L. M.; Mendes, M.G.A. Medeiros, P.A.; Gomes, R.M.; Souza, F.P. Estresse e as doenças psicossomáticas. Red de Revistas Científicas de América Latina, el Caribe, España y Portugal, Redalyc. 1997. Disponible en: < http://www.icb.ufmg.br/ipf/revista/index_revista.htm/> Accesado en: 6/06/2010. Campbell, T.W. 1995. Avian Hematology and Cytology. Ames: Iowa State Press. Campbell, T.W.; Dein, F.J. Avian Hematology: The Basics. Vet. Clin. North Am. Small Anim”. Pract, Orlando, v. 14(2): p. 223-248, 1984. Campbell, T.W. Hematology. In: Ritchie, B.W.; Harrison, G.J.; Harrison, L.R. Avian Medicine: Principles and application”. Fort Worth-FL: Wingers Publishing.1: p.177-198, 1994. Cattelan, E.V. Efeito da restrição alimentar no desempenho e na qualidade de carcaça em frangos de corte. Dissertação Mestrado. Programa de Pós-Graduação em Zootecnia, Faculdade de Agronomía, Universidade Federal do Rio Grande do Sul, Porto Alegre, Brasil, 1995. Chen, J.; Hayat, J.; Huang, B.; Balnave, D.; Brake, J. Responses of broilers at moderate or high temperatures to dietary arginine: lysine ratio and source of supplemental methionine activity”. Aust. J. Agric. Res., Melbourne, v. 54(1): p.177–181, 2003. Coles, E.H. Veterinary Clinical Patology. Philadelphia: W.B. Saunders, 1986. 143p. Costa, P.T.C. Ambiência na avicultura. In: Encontro Internacional Dos Negócios Da Pecuária 2002”. Cuiabá: ENIPEC, 2002. 1 CD-ROM.

16
Cunningham, J.G. Tratado de Fisiologia Veterinária. Rio de Janeiro, Brasil: Guanabara, 1999. 112p. Czarick, M.; Tison, B.L. Reflective roof coatings on commercial laying houses. Transac. ASAE, St Joseph, v.90, p.4512, 1990. Dale, N.M.; Fuller, H.L. Effect of diet composition on feed intake and growth of chicks under heat stress. II. Constant x cycling temperatures. Poult. Sci., Champaign, v. 59(9): p.1431-1441, 1980. Davis, R. H.; Hassan, O.E.M.; Sykes, A.H. Energy utilization in the laying hen in relation to ambient temperature.Journal Agric. Sci., Cambridge. v. 81(1): p.173-177, 1973. Deaton, J.W.; Reece, F.N.; Vardaman, 1968. The effect of temperature and density on broiler performance”. Poult. Sci., Champaign, v. 47(2): p.293- 300, 1968. Donker, R.A.; Beuving, G. Effect of corticosterone infusion on plasma corticosterone concentration, antibody production, circulating leukocytes and growth in chicken lines selected for humoral immune responsiveness. Br. Poult. Sci., Oxforshire, v. 30(3): p.361-369, 1989. Dougherty, T.F.; White, A. Influence of hormones on lymphoid tissue structure and function: the role of pituitary adrenotrophic hormone in the regulation of lymphocytes and other cellular elements of blood”. Endocrinology. 35: p.1–12, 1944. Edens, F.W. ; Siegel, H.S. Adrenal responses in high and low ACTH response lines of chickens during acute heat stress”. Gen. Comp. Endocrinol. 25: p.64-73, 1975. Edens, F.W.; Siegel, H.S. Modification of corticosterone and glucose responses by sympatholytic agents in young chickens during acute heat exposure. Poultry Science. 55: p.1704-1712, 1976. Edens, F.W. Adrenal cortical insufficiency in young chickens exposed to a high ambient temperature. Poultry Science. 57: p.1746–1750, 1978. El Halawani, M.E.; Waibel, P.E. Brain indole and catecholamines of turkeys during exposure to temperature stress”. American Journal of Physiology 230: p.110–114, 1976. Elrom, K. Handling and transportation of broilers: welfare, stress, fear and meat quality”. Israel J. Vet. Med., Raanana, v. 56(1): p.1–11, 2000. FENAVI, FONAV. La avicultura creció el 11,5%. Las cuentas del 2010. Federación Nacional de Avicultores de Colombia. Revista Avicultores edición especial. 180: p.34-36, 2010. Fox, T.W. Studies on heat tolerance in domestic fowl. Poult. Sci., Champaign, v.59, n.11, p.2391-2396, 1980.

17
Furlan, R.L.; Morais, V.M.; Malheiros, R.D.; Secato, E.R.; Macari, M. Alterações hematológicas e gasométricas em diferentes linhagens de frangos de corte submetidos ao estresse calórico agudo”. Rev. Br. Ciênc. Avíc., Campinas. v. 1(1): p.77-84, 1999. Furlan, R.L.; Macari, M. Termorregulação. In: Macari, M.; Furlan, R.L.; Gonzales, E. Fisiologia aviária aplicada a frangos de corte. Jaboticabal, Brasil: Funesp, 2002. 203p. Garren, H.W.; Shaffner, C.S. How the period of exposure to different stress stimuli affects the endocrine and lymphatic gland weights of young chicks”. Poultry Science. 35: p.266–272, 1956. Glick, B. Antibody and gland studies in cortisone and ACTH-injected birds. Journal of Immunology. 98: p.1076–1084, 1967. González, F.H.D.; Haida, K.S.; Mahl, D.; Giannesi, G.; Kronbauers, E. Incidência de doenças metabólicas em frangos de corte no sul do Brasil e uso do perfil bioquímico sanguíneo para o seu estudo. Brasil. Ciênc. Avic., Campinas. v. 3(2): p.141-147, 2001. González, F.H.D.; Silva, S.C. Introdução a Bioquímica Clínica Veterinária. PH D. Porto Alegre, Brasil: Universidade Federal do Rio Grande do Sul, 2003.147p. Gould, N.R.; Siegel, H.S. Viability of and corticosteroid binding in lymphoid cells of various tissues after corticotropin injection”. Poultry Science. 60: p.891–893, 1981. Gross, W.B.; Siegel, P.B.; Iegel, P.B.; Domermuth, C.H.; Dubose, R.T. Production and persistence of antibodies to sheep erytrocyte. 2. Resistence to infectious disease”. Poult. Sci., Champaign, v. 59(1): p.205- 210, 1980. Gross, W.B.; Siegel, H.S. Evaluation of the heterophil/ lymphocyte ratio as a measure of stress in chickens”. Avian Dis., Athens, v. 27(4): p.972-979, 1983. Guimaraes, E.B.; Vasconcelos, A.C.; Martins, N.R.S.; Oliveira, F.M.; Moro, L.; Nunes, J.E.S.; Santos, F.G.A. Porcentagem de parênquima e índice apoptótico da bolsa cloacal em frangos de corte em ambiente de conforto e estresse térmico”. Arq. Bras. Med. Vet. Zootec., Belo Horizonte, v. 55(2): p.178-186, 2003. Holmes, W.N.; Phillips, J.G. The adrenal cortex of birds. In: Chester Jones, I. and Henderson, I.W. (Eds) General, Comparative and Clinical Endocrinology of the Adrenal Cortex”. Academic Press, London. 1: p.293–420, 1976. Hornick, J.L.; Eenaeme, C.; Gérard, O.; Dufrasne, I.; Istasse, L. Mechanisms of reduced and compensatory growth. Dom. Anim. Endoc Auburn. v. 19(2): p.121-132, 2000. Howlider, M.A.R.; Rose, S.P. Temperature and the growth of broilers. World′s Poult. Sci. J., Ithaca. v. 43(2): p.228-237, 1987. Hurwitz, S.; Cohen, I.; Bar, A. Sodium and chloride requeriments of chick: relationship to acid-base balance. Poultry Science, v.52, p.903-909, 1973.

18
Lasiewski, R. C.; Acosta, A.; Berstein, M. H. Evaporative water loss in birds. I. Characteristics of the open flowmethod of determination and their relation to estimates ofthermoregulatory ability. Comp. Biochem Physiol., New York, v.19, n.3, p.455-457, 1966. Latimer, K.S.; Bienzle, D. The avian leukogram: Determination and interpretation. In: Latimer, K.S.; Bienzle, D. Schalm’s Veterinary Hematology. Baltimore: Lippincott Williams & Wilkins, 2000. 208-216p. Lawrence, T.L.J.; Fowler, V.R. Growth of farm animals. Aberdeen: CAB International, 2002. 254p. Leeson, S. Nutritional considerations of poultry during heat stress”. World's Poult. Sci. J., Ithaca, v. 42: p.69-81, 1986. Linsley, J.G.; Berger, R.R. Respiratory and cardiovascular responses in the hyperthermic domestic fowl. Poult. Sci.. Champaign, 43: 291-305, 1964. Macari, M.; Gonzales, E. Fisiopatogenia da síndrome da morte súbita em frangos de corte. In: Conferência Apinco de Ciência e Tecnologia Avícolas, Campinas. Anais... Campinas: FACTA; 1990. p. 65-73. Macari, M.; Furlan, R.L.; Gonzales, E. Fisiologia aviária aplicada a frangos de corte. Jaboticabal: FUNEP/ UNESP, 1994. 246p. Macari, M. Água na avicultura industrial. Jaboticabal: Funep/Unesp. 1996.1: p.128. Macari, M.; Luquetti, B.C. Fisiologia Cardiovascular. In: Macari, M.; Furlan, R.L.; Gonzales, E. Fisiologia aviária aplicada a frangos de corte. Jaboticabal: Funep/Unesp, 2002. 232-240p. Macari, M.; Furlan, R.L.; Maiorka, A. Aspectos fisiológicos e de manejo para manutenção da homeostase térmica e controle de síndromes metabólicas. In: Mendes, A. A.; Naas, I.A.; Macari, M. Produção de frangos de corte. Campinas: Facta; 2004. p.137-156. Mardsen, A.; Morris, T.R. Comparison between constant and cyclic environments on shell quality and other lay performance factors with leghorn pullets. Poult. Sci., Champaign. v. 54(1): p.36-46,1975. Maxwell, M.H.; Hocking, P.M.; Robertson, G.W. Differential leucocytes responses to various degrees of food restriction in broilers, turkeys and ducks”. Br. Poult. Sci., Oxforshire. v. 33(2): p.177-187, 1992. Mc. Farlane, J.M.; Curtis, S.E. Multiple concurrent stressors in chicks. 3. Effects on plasma corticosterone and the heterophil: lymphocyte ratio. Poult. Sci., Champaign. v. 68(4) p.522-527, 1989. Mench, J.A. Broiler breeders: feed restriction and welfare. World’s Poult”. Sci. J., Ithaca. v. 58(1): p.23-29, 2002.

19
Mitchell, J. W. Heat transfer from spheres and other animal forms. Biophys. J., Bethesda, v.16, n.6, p.561-569, 1976. Mitchell, M.A.; Carlisle, A.J. The effect of chronic exposure to elevated environmental temperature on intestinal morphology and nutrient absorption in the domestic fowl (Gallus domesticus)”. Comp. Biochem. Physiol., New York. v. 101(1): p.137-142, 1992. Mitchell, M.A. Anormalidades musculares: mecanismos fisiopatológicos. En: Richardson, R.I.; Mead, G.C. Ciência de la Carne de Ave. [S.l.:s.n], 2001. 497p. Mongin, P. “Recent advances in dietary cation-anion balance: applications. In: Poultry Proceedings Nutrition Society, 1981, Cambridge”. Proceedings... Cambridge. v. 40: p.285-294, 1981. Morgan, W.E. Heat reflective roof coatings. Transac. ASAE, St.Joseph. v. 90: p.4513, 1990. Morgulis, M.S. Inmunología aplicada. In: Macari, M.; Furlan, R.L.; Gonzales, E. Fisiologia aviária aplicada a frangos de corte. Jaboticabal: Funep/Unesp, 2002. 1: 231-243p. Naas, I.A. Aspectos físicos da construção no controle térmico do ambiente das instalações. In: Conferência Apinco de Ciencia e Tecnologia Avícolas, Santos. Anais... Campinas: FACTA; 1994. 1: p.111. Noriega, M.L.V.C. Apuntes de hematología aviar. México: Universidad Nacional Autónoma, 2000.121p. Noriega, M.L.V.C. Importância da hematologia no diagnóstico das aves. In: Encontro Técnico Sobre Avicultura de Corte da Região de Descalvado. Descalvado. Resumos. Descalvado, 2000. 4: p.1-11. Nose, H. Role of osmolality and plasma volume during dehydration in humans”. Journal Applied Physiology. v. 65: p.325- 331, 1988. Payne, G.C. Practical aspects of environmental temperature for laying hens”. World’s Poult. Sci. J., Ithaca, v. 22(1): p.126-139, 1966. Perea, A.T.; Isaías, G.I.; Maldonado, F.G. Técnicas de medición de estrés en aves. Vet. Mex., México. v. 28(4): p.345-351,1997. Pilo, B.; John, T.M.; George, J.C.; Etches, R.J. Liver Na+K+-ATPase activity and circulating levels of corticosterone and thyroid hormones following cold and heat exposure in the pigeon. Comparative Biochemistry and Physiology. 80: p.103–106, 1985. Plavnik, I.; Hurwitz, S. The performance of broiler chicks during and following a severe feed restriction at an early age”. Poult. Sci., Champaign. v. 64(2): p.348-355, 1985. Plavnik, I.; Yahav, S. Effect of environmental temperature on broiler chickens subjected to growth restriction at an early age. Poult. Sci., Champaign. v. 77(6): p.870-872,1998.

20
Puvadolpirod, S.; Thaxton, J.P. Model of Physiological Stress in Chickens 1. Response Parameters. Poult. Sci., Champaign. v. 79(4): p.363-369, 2000. Revidatti, F.A.; Fernandez, R.J.; Terraes, J.C.; Sandoval, G.L.; Luchi, P.E. Modificaciones del peso corporal y indicadores de estrés en pollos parrilleros sometidos a inmovilización y volteo. Rev. Vet. Arg., Corrientes. v. 12(1): p.1-8, 2002. Ribeiro, A.M.L.; Penz, A.M.; Teeter, R. Effects of 2-hydroxy-4-(methylthio) butanoic acid and DL-Methionine on broiler performance and compensatory growth after exposure to two different environmental temperatures. J. Appl. Poult. Res., Athens. v. 10(4): p.419-426, 2001. Rosales, A.G.; Villegas, P.; Lukert, P.D.; Mohamed, A.M.; Brown, J. Isolation. Identification and pathogenicity of two field strains of infectious Bursal Virus. Avian Dis., Athens. v. 33(1): p.35-41,1989. Ruckebush, L.P.; Phaneuf, L.P.; Dunlop, R. F. Fisiologia de pequeñas y grandes especies. 1. ed. México D.F, México: Manual Moderno, 1994. 243p. Rutz, F. Aspectos fisiológicos que regulam o conforto térmico das aves. In: Conferência Apinco de Ciencia e Tecnologia Avícolas, Santos. Anais... Campinas: FACTA. 1: p.99-110, 1994. Salvador, D. Suplementação de bicarbonato de sódio na ração e na água de bebida de frangos de corte submetidos ao estresse calórico. ARS Veterinária. v. 15; p.144-148, 1999. Sammelwitz, P.H. Adrenocortical hormone therapy of induced heat stress mortality in broilers”. Poultry Science. 46: p.1314, 1967. Satterlee, D.G.; Abdullah, R.B.; Gildersleeve, R.P. Plasma corticosterone radioimmuneassay levels in the neonate chick”. Poult. Sci., Champaign. v. 59, (6): p.900–905, 1980. Scope, A.; Filip, T; Glaber, C.; Resch, F. The influence of stress from transport and handling on hematologic and clinical chemistry blood parameters of racing pigeons (columba livia domestica). Avian Dis., Athens. v. 46(1): p.224-229, 2002. Siegel, H.S.; Beane, W.L. Time responses to single intramuscular doses of ACTH. Poultry Science. 40: p.216–219, 1961. Sturkie, P.D. Blood: physical characteristics, formed elements, hemoglobin, and coagulation. In: sturkie, P.D. Avian Physiology. New York: Springer-Verlag, 1976. p.245-250. Sturkie, P.D.; Griminger, P. Body fluids: blood. In: STURKIE, P.D. Avian Physiology. New York: Springer – Verlag, 1986. p.102-129.

21
Sullivan, D.A.; Wira, C.R. Sex hormone and glucocorticoid receptor in the bursa of Fabricius of immature chickens. Journal of Immunology. 122: p.2617–2623, 1979. Susbilla, J.P.; Frankel, T.L.; Parkinson, G.; Gow, C.B. Weight of internal organs and carcass yield of early food restricted broilers. Br. Poult. Sci., Oxforshire. v. 35(5): p.677-685, 1994. Teeter, R.G.; Smith, M.O.; Murray, E. Force feeding methodology and equipment for poultry”. Poult. Sci., Champaign. v. 63(4): p.573-575, 1984. Teeter, R.G.; Smith, M.O. Chromic ambient temperature stress effects on broiler acid-base balance and their response to supplemental ammonium chloride, potassium chloride and potassium carbonate. Poultry Science. v. 65: p.1777-1781, 1985. Thompson, E.B.; Lippman, M.E. Mechanism of action of glucocorticoids. Metabolism. 23: p.159–202, 1974. Tinôco, I.F.F. Avicultura Industrial: Novos conceitos de materiais, concepções e técnicas construtivas disponíveis para galpões avícolas Brasileiros. Rev. Bras. Ciênc. Avic., Campinas. v. 3(1): p.1-26, 2001. Yahav, S.; Goldfeld, S.; Plavnik, I.; Hurwitz, S. Physiological responses of chickens and turkeys to relative humidity during exposure to high ambient temperature. J. Thermal Biol., Southampton. v. 20(3): p.245-253, 1995. Yahav, S.; Straschnow, A.; Plavnik, I.; Hurwitz, S. Blood system response of chickens to changes in environmental temperature. Poult. Sci., Champaign. v. 76(4): p.627-633, 1997. Zhou, W.T.; Fujita, M.; Yamamoto, S.; Iwasaki, K.; Ikawa, R.; Oyama, H.; Horikawa, H. Effects of glucose in drinking water on the changes in whole blood viscosity and plasma osmolality of broiler chickens during high temperature exposure. Poult. Sci., Champaign. v. 77(4): p.644-647, 1998. Zubair, A.K.; Leeson, S. Effect of varying period of early nutrient restriction on growth compensation and carcass characteristics of male broilers. Poult. Sci., Champaign, v. 73(1): p.129-136, 1994.

22
1. Capitulo 1
ÍNDICES ZOOTÉCNICOS DE POLLO DE ENGORDE BAJO CONDICIONES DE ESTRÉS CALÓRICO EN EL PIEDEMONTE AMAZÓNICO COLOMBIANO
1.1 Introducción La industria avícola mundial tiene como objetivo producir la mayor cantidad de carne en el menor tiempo posible (Buxade, 1988; North, 1993) para lograrlo se han realizado avances generales en nutrición y genética con la generación de líneas modernas de alto potencial genético, las cuales manifiestan alta tasa de crecimiento y exigen aumento en la demanda sanguínea residual, debido al mayor requerimiento metabólico. No obstante a este mejoramiento, el desarrollo del sistema cardiorespiratorio de estos animales no ha acompañado esta evolución, lo cual ha conllevado a deficiencias en los proceso de oxigenación de toda la masa muscular, trastornos en diversos órganos y detrimento final del rendimiento productivo (Macari et al., 2004). En Colombia, como en muchas regiones del mundo la eficiencia de la productividad avícola y la ganancia económica (De Basilio et al., 2001) son afectadas en forma adversa por el estrés calórico, condición que altera el mantenimiento de la temperatura interna corporal en forma constante. El intervalo optimo de temperatura ambiente para las aves depende de su ciclo de vida, necesitando temperaturas entre 32°C a 30°C durante la primera semana, y después de los 21 días de edad y hasta finales del ciclo de engorde 22°C a 18°C (Gonzales et al., 2000). En ambientes cuyas condiciones son variables, estos ajustes requieren gasto de energía corporal, la que dentro de la zona de termoneutralidad debería ser empleada para la producción de carne, en lugar de ser utilizada para disipación de calor. Siendo así, los efectos del estrés calórico sobre el rendimiento zootécnico dependen del grado y duración del período que la temperatura exceda la zona de termoneutralidad; cuando la temperatura ambiente supera el límite de los 40°C el ave se encuentra en franco peligro de muerte (Macari et al., 2004). La temperatura y humedad ambiental son factores físicos de gran efecto en el rendimiento zootécnico de las aves de engorde, debido a su influencia directa sobre el consumo de alimento (Teeter et al., 1984). Czarick y Tison (1990); Morgan (1990), concluyen que la primera respuesta del ave al estrés por calor es la disminución en el consumo del alimento, con pérdida del apetito, estrategia que permite minimizar la cantidad de calor generado por la digestión y el metabolismo energético; aunque en forma simultánea se deja de recibir los nutrientes esenciales para la producción y bienestar en detrimento de la supervivencia, ganancia de peso y conversión alimenticia (Mc Geehin et al., 2001). Según Bonnet et al., (1997) y Hai et al., (2000), la situación se torna más crítica con el crecimiento de las aves, pues existe una correlación negativa entre la disipación de calor y el peso corporal.

23
En la zona de estudio bajo las condiciones climáticas del Piedemonte Amazónico Colombiano los resultados zootécnicos de la producción de pollo de engorde no son muy eficientes durante gran parte del año como efecto directo de las altas temperaturas, la que frecuentemente sobrepasan los límites de termorregulación de las aves. Debido a esta situación también se presenta influencia indirecta al crearse condiciones favorables en las casetas avícolas para la proliferación de microorganismos patógenos perjudiciales a la salud de las aves. Con el propósito de contribuir al conocimiento de la producción de pollo de engorde bajo condiciones climáticas de estrés calórico se planteo evaluar los índices zootécnicos del pollo de engorde moderno criado bajo condiciones ambientales del Piedemonte Amazónico Colombiano.
1.2 Materiales y métodos
1.2.1 Localización
El experimento se realizó en la granja “Pasarela” ubicada en la zona rural del municipio de Florencia Departamento del Caquetá, Colombia, ubicada a 1°34´35.58´´N y 75°32´39.68´´de longitud Occidental, 243 m.s.n.m. con temperatura media de 27°C, humedad relativa del 85% y precipitación anual de 3600 mm3.
1.2.2 Animales Fueron utilizados 250 pollos machos de la estirpe comercial Cobb-500 y 250 machos de la estirpe comercial Ross 308, previo al experimento las aves fueron clasificadas por peso y línea genética para conformar grupos homogéneos. Los animales con un peso promedio inicial de 773,56±8,28 g, se distribuyeron en corrales de 3,8 m2, para una densidad final de 6,9 aves/m2. Los tratamientos se organizaron en un diseño irrestrictamente al azar en un esquema bifactorial 2x2 (2 condiciones de temperatura y 2 estirpes comerciales de pollo de engorde) para un total de cuatro tratamientos, cinco repeticiones por tratamiento y 25 animales por unidad experimental.

24
1.2.3 Tratamientos
T1: Pollos de engorde Cobb 500 machos de 22 días de edad, mantenidos a una temperatura de 30,0±1ºC (estrés calórico crónico) durante 21 días. T2: Pollos de engorde Cobb 500 machos de 22 días de edad, sometidos a la temperatura ambiente del Piedemonte Amazónico Colombiano durante 21 días. T3: Pollos de engorde Ross 308 machos de 22 días de edad, mantenidos a una temperatura de 30,0±1ºC (estrés calórico crónico) durante 21 días. T4: Pollos de engorde Ross 308 machos de 22 días de edad, sometidos a la temperatura ambiente del Piedemonte Amazónico Colombiano durante 21 días.
1.2.4 Manejo del pollo de engorde Las aves recibieron las prácticas de manejo convencionales, en cuanto a planes sanitarios, nutricionales y alimenticios requeridos para cada etapa productiva, con disposición de alimento y agua a voluntad (May et al., 2001). Los pollos fueron criados durante las tres primeras semanas de vida, en un corral dentro del módulo experimental y sometidos a una temperatura inicial de 32ºC±2ºC (Sainsbury, 2000), la cual se redujo gradualmente hasta alcanzar los 29,87±7,20ºC a los 22 días, tiempo en el cual se inicio la etapa experimental. Las aves durante el período experimental recibieron alimento tipo migaja y agua a voluntad. La dieta experimental fue balanceada para contener 3100 Kcal de EM y 19,5% de proteína; a base de maíz, torta de soya y aceite vegetal, según las exigencias nutricionales, descritas por Rostagno (2005) y NRC (1994); para satisfacer los requerimientos nutricionales de las aves durante la fase de engorde (tabla 3). El monitoreo de la temperatura ambiente y la humedad relativa de cada ambiente fue realizado cada dos horas durante las 24 horas del día del periodo experimental, a través de un termo higrómetro digital de máximas y mínimas (Pacer industries Inc. Modelo HTA 4200, con sensibilidad de 0,1°C y 0,1%; Wisconsin, USA 1998) localizado a nivel de la altura de las aves.

25
Tabla 3: Composición de la dieta experimental para pollos de engorde de 22 a 42 días
Ingrediente % Maíz 64,1 Torta de soya 46% 29,5 Aceite vegetal 2,4 Carbonato de calcio 1,47 Fosfato bicálcico 1,63 Sal 0,46 Premezcla Vitamínica1 0,05 Premezcla mineral2 0,10 Metionina HA 0,26 Colina 0,03 Niveles nutricionales
EM (kcal/kg) 3100 PB (%) 19,5 Lisina (%) 1,14 Metionina+cistina (%) 0,83 Arginina (%) 1,27 Treonina (%) 0,75 Triptófano (%) 0,24 Relación Arg:Lys 1,11 Ca (%) 0,95 P disponible (%) 0,42 Para garantizar durante todo el periodo experimental (22-42 días) una temperatura ambiente superior a los 30ºC se utilizaron criadoras a gas. El programa de luz durante el periodo experimental fue continuo, 24 horas de luz natural/artificial (Fussell et al., 2003). Las aves se pesaron al inicio del experimento y semanalmente para monitorear el consumo de alimento y la ganancia de peso acumulada.
2 Premezcla vitamínica (Contenido por kg/ración): Vit.A. 10.000 UI; Vit D3 3.000 UI; Vit E 60 mg; Vit K3 3 mg;
Vit B1 3 mg; Vit. B2 8 mg; Vit B6 4 mg; Vit B12 0,014 mg; Ácido Pantoténico 20 mg; Niacina 50 mg; Ácido Fólico 2 mg; Biotina 0,15 mg 2 Premezcla mineral (Contenido por kg/ración): Fe 40 mg; Zn 80 mg; Mn 80 mg; Cu 10 mg; I 0,7 mg; Se 0,3
mg.

26
1.2.5 Variables evaluadas Ganancia de peso (g): Este índice se estableció determinando la diferencia entre el peso corporal acumulado al día 42 y el peso de las aves al día 22. Se determinó para cada repetición a través de una balanza digital de precisión Sartorius® (Pine Brook, New Yersey, USA 2003), para ello los animales fueron pesados cada semana en las horas de la mañana antes de recibir la primera ración de alimento. Consumo de alimento acumulado (g/ave): Se estableció para todas las repeticiones de cada tratamiento, calculando la diferencia entre el alimento ofrecido y el rechazado durante cada semana y el período experimental total. Conversión alimenticia (g/g): Se determinó de la relación entre el alimento total consumido y la ganancia de peso corporal de cada unidad experimental (Plavnik y Yahav, 1998). Mortalidad (%): Se obtuvo de la relación entre el número de pollos muertos de cada unidad experimental y tratamiento durante el período experimental dividido entre el número de pollos iniciales por unidad experimental y tratamiento multiplicado por 100.
1.2.6 Análisis estadístico Los resultados fueron sometidos a análisis de varianza, utilizando el procedimiento GLM (General Linear Models) de SAS-8.02 (2002). Las diferencias entre tratamientos fueron establecidas mediante la prueba de F y la prueba de tukey a un nivel de confianza de (P<0,05).
1.2 Resultados y discusión Tabla 4: Ganancia de peso (g) en pollos de engorde sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano de los 22 a 42 días de edad
ESTIRPE TEMPERATURA COBB ROSS PROMEDIO
Estrés calórico crónico
1073,93±277,9 Ab 991,17±172,8 Ab 1032,55±225,4 b
Piedemonte Amazónico
PROMEDIO
1470,92±188,3 Aa
1272,43±233,1 A
1385,50±296,3 Aa
1188,34±234,5 A
1428,21±242,3 a
C.V% 18,32 19,74
Medias de tratamientos dentro de cada fila seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.

27
Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
En la tabla 4, se observa el análisis de los datos el cual muestra que no hubo diferencia estadística significativa (P>0,05) entre los dos linajes de aves bajo ninguna de las dos variantes de temperatura. No obstante, al comparar los resultados de la ganancia de peso de cada uno de los dos linajes de aves sometidos a la temperatura ambiente del Piedemonte Amazónico, frente a los valores de referencia para este índice durante los 22 a 42 días de edad expuestos en las guías de manejo Cobb (1,954g) y Ross (1,947g ); se puede notar una diferencia inferior de 24,7% para los pollos Cobb y un 28,8% para la línea Ross respectivamente; este bajo rendimiento se explica debido a que las aves de rápido crecimiento requieren una temperatura confort inferior a los 21°C a partir de los 21 días de edad, ya que a esta edad las aves aumentan la producción de calor metabólico (Aviagen, 2008) y (Cobb-vantress, 2008). Los hallazgos experimentales obtenidos, permiten corroborar los reportes de Melitzer (1986) y Tinôco (1998) quienes determinan que el óptimo rendimiento zootécnico de las aves solo es obtenido cuando los pollos son engordados a una temperatura ambiente dentro de la zona de confort de acuerdo a la etapa de crecimiento. Cabe resaltar que cada uno de los híbridos sometidos a la temperatura ambiente del Piedemonte Amazónico mostró un rendimiento superior para la ganancia de peso, comparado con las aves bajo la temperatura estrés calórico crónico, representada en una diferencia de 26,9% para las aves Cobb y un 28,4% para los pollos Ross. El mejor rendimiento de los animales observado bajo la temperatura ambiental del Piedemonte Amazónico en cada uno de los linajes, fue favorecido por la variabilidad de la temperatura en donde probablemente los periodos de termoneutralidad (Deeb, 1999; Macari, 2001; Balnave, 2004) nocturnos registrados permitieron una recuperación parcial de las aves al maximizar el uso de la energía, mejorando el rendimiento de los animales evidenciado a través de la variable ganancia de peso; entre tanto la temperatura estrés calórico crónico no permitió una recuperación térmica de las aves. Otros factores como la reducción en el consumo de alimento (Emmans y Kyriazakiis, 2000), la selección genética de los pollos modernos, Cahaner et al., (1993); Emmans et al., (2000); Yahav et al., (2001); Bochi, (2005), y los niveles plasmáticos de corticosterona (Post et al., 2003), pueden aumentar la sensibilidad de los animales frente a condiciones ambientales no óptimas. Una digestión ineficiente del alimento, Har et al., (2000), la energía utilizada durante la contracción muscular asociada al jadeo durante las altas temperaturas Summer et al., (1990), acompañada de un deterioro en el metabolismo (Farrell y Swain, 1978) sumado al bajo consumo de algunos nutrientes en pobres cantidades, podrían ser agravantes del regular comportamiento de esta variable. La alta temperatura ambiental es el principal factor que afecta la ganancia de peso en la producción de pollo de engorde (Hacina et al., 1996; Abu-Dieyeh, 2006). En conclusión la variable ganancia de peso puede ser utilizada como indicador del comportamiento zootécnico de las aves bajo condiciones de estrés calórico.

28
Tabla 5: Valores medios de consumo de alimento acumulado (g/ave) en pollos de engorde sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano de los 22 a 42 días de edad
ESTIRPE TEMPERATURA COBB ROSS PROMEDIO
Estrés calórico crónico 2511,60±644,7 Ab 2438,60±518,8 Ab 2475,1 b
Piedemonte Amazónico
PROMEDIO
2855,60±477,6 Aa
2683,60±561,2 A
2837,2±555,2 Aa
2637,9±537,5 A
2846,4 a
C.V% 20,91 20,35
Medias de tratamientos dentro de cada fila seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
Al realizar el análisis de los datos para la variable consumo de alimento acumulado (tabla 5), se puede notar que no hubo diferencia significativa (P>0,05) entre las dos estirpes de pollo estudiadas bajo ninguna de las dos variantes de la temperatura experimental. Los mayores consumos de alimento fueron observados en las aves sometidas a las condiciones ambientales del Piedemonte Amazónico, cuyo resultado pudo ser favorecido por los diferentes rangos variables de temperatura diaria, permitiendo periodos de mayor consumo de alimento, asociado a un posible grado de aclimatación, (Teeter et al., 1992), en el cual probablemente las aves sometidas a estrés calórico ambiental cíclico aprendieron a comer menos durante las horas críticas de pico de calor, consumiendo más alimento en las horas más frescas del día. Sin embargo, al confrontar los resultados obtenidos en cada uno de los dos linajes de pollo bajo las condiciones ambientales del Piedemonte Amazónico, con el consumo de alimento acumulado de referencia desde el día 22 a 42 expuestos en las guías de manejo para el Ross (3,673 g/ave) y el Cobb (3,727 g/ave) respectivamente; se infiere que las aves Ross tuvieron un 22,7% menos de consumo de alimento respecto a su guía de manejo; en forma similar las aves Cobb mostraron también un inferior consumo de alimento representado por un 23,3%; los resultados hallados en cada uno los dos linajes se atribuye a las variables oscilaciones de temperatura ambiental del Piedemonte Amazónico Colombiano, las cuales no permitieron alcanzar una homeóstasis térmica constante para alcanzar los objetivos de consumo sugeridos por las dos líneas genéticas. Estos hallazgos corroboran los resultados obtenidos por Teeter et al., (1984), Bonnet et al., (1997) y Temim et al., (2000) quienes después de exponer pollos de engorde con

29
diferentes pesos corporales desde los 22 a 42 días de edad, con diferentes temperaturas generadoras de estrés calórico, evidenciaron una disminución del 22% al 24% del índice consumo de alimento acumulado; concluyendo que el peso corporal, el nivel, la duración y el tipo de estrés al que son sometidas las aves, son factores directos que afectan este parámetro. El bajo consumo de alimento es consecuencia directa de las elevadas temperaturas ambientales, explicado por la disminución del apetito como medios para reducir la carga calórica endógena dentro de los ajustes fisiológicos forzosos para el mantenimiento de la homeostasis térmica. La primera respuesta del ave al estrés por calor es la disminución del consumo de alimento (Veldkamp et., al 2005; Zeballos, 2001), lo que implica también una baja ingesta de nutrientes esenciales para la producción y mantenimiento. Durante el estrés calórico crónico, existe mayor disminución del consumo de alimento, debido a que las aves no pueden compensar el consumo de alimento en las horas más frescas del día como ocurre en el estrés agudo; este hecho puede explicar el mejor comportamiento de la variable evaluada bajo las condiciones del Piedemonte Amazónico. Tabla 6: Valores medios de conversión alimenticia acumulada (g/g) en pollos de engorde de 22 a 42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano
ESTIRPE TEMPERATURA COBB ROSS PROMEDIO
Estrés calórico crónico
2,340±2,02 Aa 2,460±0,03 Aa 2,40 a
Piedemonte Amazónico
PROMEDIO
1,941±0,02 Ab
2,141±1,02 A
2,048±1,01 Ab
2,254±0,52 A
1,99 b
C.V% 47,6 23,1
Medias de tratamientos dentro de cada fila seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
El análisis de los resultados para la variable conversión alimenticia (tabla 6), muestra que no hubo efecto (P>0,05) significativo entre las dos estirpes de aves bajo ninguna de las variantes de la temperatura experimental. Resultados diferentes a los hallados en este ensayo fueron obtenidos por Lovatto, (1989) y Flemming et al., (1999) quienes reportaron conversiones alimenticias significativas (P<0,05) en aves Cobb con referencia a otras líneas de engorde durante los últimos 21 días de crianza bajo condiciones de estrés calórico; entre tanto, Holsheimer y Veerkamp

30
(1992) y Souza et al., (1994) encontraron una mejor conversión alimenticia acumulada en pollos de engorde Ross después de establecer comparación con aves de engorde Cobb. No obstante, al analizar el comportamiento de esta variable en cada linaje y bajo cada una de las temperaturas generadoras de estrés calórico (tabla 6), se puede notar el mejor comportamiento (P<0,05) de la conversión alimenticia en ambas estirpes de pollos sometidas a la temperatura ambiental del Piedemonte Amazónico Colombiano; el mejor desempeño de las aves evidenciado a través de la variable evaluada, es consecuencia directa de un mejor aprovechamiento del alimento ingerido en relación con el peso acumulado; posiblemente este rendimiento productivo superior es efecto de una mejor adaptación o aclimatación, (Cahaner et al., 1993) y (Balnave, 2004), concedido por un mejor ajuste fisiológico y metabólico las aves bajo la temperatura ambiental del Piedemonte Amazónico Colombiano. En promedio los pollos de las dos estirpes bajo la temperatura del Piedemonte Amazónico (29,87±7,20ºC), mostraron la mejor conversión alimenticia representada por una diferencia del 17,08% con respecto a las aves sometidas a estrés calórico crónico. Sin embargo, al comparar los resultados experimentales de la variable conversión alimenticia en las aves sometidas a la temperatura ambiental del Piedemonte Amazónico, frente a los parámetros objetivos expuestos en las guías de manejo de los híbridos Cobb (1,90) y Ross (1,88) respectivamente desde los 22 a 42 días de edad, se puede concluir que los índices de conversión alimenticia hallados difieren en forma negativa con los rendimientos esperados para cada linaje; representados en una diferencia del 2,10% para el Cobb y un 7,84% para el Ross. El bajo rendimiento de los dos híbridos se explica como consecuencia de las altas temperaturas, condición que exige a las aves realizar cambios orgánicos para poner en marcha los mecanismos de termorregulación tendientes a disminuir el calor interno, Simmons et al., (2003) y Veldkamp et al., (2005), a través de los diferentes mecanismos de disipación de calor (Hillman et al., 1985) como por ejemplo, la evaporación respiratoria mediante el jadeo Butcher y Miles, (2003). Estos resultados corroboran los reportes de Oliveira, (2000); De Basilio et al., (2003), quienes afirman que el estrés por calor es responsable por grandes pérdidas en rendimiento de los lotes de pollos de engorde, generando perjuicio en la conversión alimenticia, especialmente cuando las condiciones de estrés calórico ocurren en la fase final del ciclo de engorde.

31
Tabla 7: Valores medios de mortalidad (%) acumulada en pollos de engorde de los 22 a 42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano
ESTIRPE TEMPERATURA COBB ROSS PROMEDIO
Estrés calórico crónico
13,10±3,34 Aa 12,80±3,06 Aa 12,9 a
Piedemonte Amazónico
PROMEDIO
3,20±1,57 Ab 8,15±2,46 A
3,60±1,78 Ab 8,20±2,42 A
3,4 b
C.V% 30,18 29,51
Medias de tratamientos dentro de cada fila seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
Al análisis estadístico de los resultados de la variable mortalidad, no se encontró diferencia estadística significativa (P>0,05) entre los promedios de ambas líneas genéticas como efecto de las dos variantes de la temperatura (tabla 7). Sin embargo, se puede observar una diferencia significativa (P<0,05) entre los valores promedio de la mortalidad para las dos líneas bajo la temperatura estrés calórico crónico, con respecto al promedio de las mismas bajo la temperatura ambiental del Piedemonte Amazónico, evidenciando una diferencia superior de 73,7% para las aves bajo estrés calórico crónico. Al comparar los resultados de la variable evaluada bajo la temperatura ambiental del Piedemonte Amazónico frente a los porcentajes de mortalidad esperados para los híbridos Cobb (2,5%) y Ross (2,2%) durante los 22 a 42 días de edad expuestos en las guías de manejo (Aviagen, 2008; Cobb-vantress, 2008); se puede inferir que los valores hallados se encuentran por encima de los valores de referencia de cada estirpe, representados en una mortalidad superior de 21,8% para el Cobb y 38,8% para el Ross; el alto porcentaje de mortalidad hallado para las dos estirpes de pollos probablemente fue ocasionado por el grado de confinamiento experimental, el cual pudo interferir con la elaboración de los ajustes comportamentales necesarios para el mantenimiento de la homeostasis térmica (Macari et al., 2004), con la cual las aves buscan mantenerse sanas y vivas. Los resultados encontrados en este trabajo ratifican los reportes de mortalidad bajo condiciones de estrés calórico en pollo de engorde durante los últimos 22 a 42 días de edad observados por Oliveira, (2000); De Basilio et al., (2003); Macari et al., (2004) y Mujahid et al., (2005, 2007a).

32
El ineficiente enfriamiento evaporativo de los pollos, a la postre pudo generar almacenamiento continuo de calor con incremento de la temperatura corporal, conduciendo a las aves a un nivel letal (Ito, 2002), mediante el agotamiento y postración. La muerte por agotamiento pudo ser debida a una insuficiencia adreno cortical o a una falla cardiovascular acompañada de un desbalance iónico sanguíneo (Harrison, 1995).
1.4 Conclusiones
Las dos estirpes de pollo de engorde evaluadas no expresaron su potencial genético de rendimiento, debido a las agrestes condiciones ambientales del Piedemonte Amazónico Colombiano.
La temperatura ambiental del Piedemonte Amazónico Colombiano, durante el
período experimental fue en promedio de 29,87±7,20ºC, la cual se convierte en generadora de estrés calórico agudo para las aves durante el día con cierto grado de atenuación en las noches.
No se evidencio diferencia en el rendimiento productivo entre los linajes Cobb y
Ross, bajo las dos temperaturas generadoras de estrés calórico.
La producción de pollo de engorde moderno en la región del Piedemonte
Amazónico Colombiano, no se ajusta a los patrones de producción de las regiones con temperaturas de climas templados, razón por la cual, además de adaptar todos los recursos tecnológicos para minimizar el efecto del estrés calórico en las aves, se hace necesario establecer las curvas de producción y rendimiento específicas para la región.

33
Bibliografía Abu-Dieyeh, Z.H.M. Effect of high ambient temperature per se on growth performance of broilers. Int. Poult. Sci., 5: p.19-21, 2006. Aviagen, Ross 308®. Objetivos de rendimiento y especificaciones nutricionales. Huntsville, Alabama.USA. p.3 -13, 2008. Balnave, D. Challenges of accurately defining the nutrient requeriments of heat-stressed poultry. Poult. Sci., Champaign, v. 83: p.5-14, 2004. Bochi, O.B. Efeito do cruzamento entre diferentes genótipos para o uso em sistemas alternativos de frangos de corte. Dissertação Mestrado. Programa de Pós-Graduação em Zootecnia, Área de Concentração em Produção Animal, Universidade Federal de Santa Maria centro de Ciências Rurais, R,S, Brasil, 2005. Bonnet, S.; Geraert, P.A.; Lessire, M.; Carre, B.; Guillaumin, S. Effect of high ambient temperature on feed digestibility in broilers. Poult. Sci., Champaign, v. 76(6): p.857–863,1997. Butcher, G.D.; Miles, R.D. Causes and prevention of wet litter in broiler houses. Cooperative Extension Service, Institute of Food and Agric. Sci. Univ. Florida. v. 80, p.67-69, 2003. Buxade, C. El pollo de carne. 2. ed. México D.F, México: Mundiprensa, 1988.105p. Cahaner, A.; Deeb, N.; Gutman, M. Effects of the plumage-reducing naked neck (Na) gene on the performance of fast-growing broilers at normal and high ambient temperatures. Poult. Sci., v. 72(5): p.767-775, 1993. Cobb-Vantress, Inc. Suplemento informativo de rendimiento y nutrición del pollo de engorde Cobb® 500. Siloam Springs, Arkansas, USA. p.6.2008. Czarick, M.; Tison, B.L. Reflective roof coatings on commercial laying houses. Transac. ASAE, St Joseph, v.90, p.4512, 1990. De Basilio, V.; Vilariño, M.; León, A.; Picard, M. Efecto de la aclimatación precoz sobre la termotolerancia en pollos de engorde sometidos a un estrés térmico tardío en condiciones de clima tropical”. Revista Científica, FCV-Luz. v. 11(1): p.60-68, 2001. De Basílio, V.; Requena, F.; León, A.; Vilariño, M.; Picard, M. Early-age thermal conditioning immediately reduces body temperature of broiler chicks under a tropical environment. Poultry Sci., 82(8): p.235-1242, 2003. Deeb, N.; Cahaner, A. The effects of naked neck genotypes, ambient temperature, and feeding status and their interactions on body temperature and performance of broilers”. Poult Sci., 78: p.1341-1346, 1999.

34
Emmans, G.; Kyriazakiis, I. Consequences of genetic change in farm animals on food intake and feeding behavior. Proc Nutr Soc 60: p.115-125, 2000. Farrell, D.J.; Swain, S. Effects of temperature treatments on the Energy and nitrogen metabolism of fed chickens. Br. Poult. Sci., 18: 735-748, 1978. Flemming, J.S.; Janzen S. A.; Endo, M.A. Parâmetros em linhagens comerciais de frangos de corte. Arch. Vet. Scienc., 4(1): 57-59, 1999. Fussell, L.; Rossi, A.Y.; Wilson, M. Lighting programs and cob 500 broiler performance. South África: Technical focus, 2003.107p. González, A.J.; Suárez, M.E.; Pró, M.C.; López, C. Restricción alimenticia y salbutamol en el control del síndrome ascítico en pollos de engorda: Comportamiento productivo y características de la canal. Montecillo, Edo. Méx. Agrociencia. 34: p.283-292, 2000. Hacina, A.B.; Geraert, P.A.; Padilha, J.C.; Solanage, G. Chronic heat exposure enhances fat deposition and modifies muscle and fat partition in broiler carcasses. Poult. Sci., 75: 505-513, 1996. Hai, L..; Rong, D.; Zhang, D.Z.Y. The effect of thermal environment on the digestion of broilers”. J. Anim. Physiol. A. Anim. Nutr., Verlag, v. 83: (1): p.57-64, 2000. Har, L.; Rong, D.; Zhang, Z.A. The effect of thermal environment on the digestion of broilers. Anim. Physiol., 83: p.75-61, 2000. Harrison, P.C. O estresse calórico nas aves-Fisiologia e conseqüências. In: Simpósio internacional de ambiência e instalação na avicultura industrial. Campinas. Anais… Campinas; 1995.p.25-32. Holsheimer, J.P.; Veerkamp, C.M. Effect of dietary energy, protein level and lysine content of on performance and yield of two strains of male broiler chicks. Poult Sci., v.66, n.5, p.872-879, 1992. Ito, N.M.K. Manejo e prevenção das doenças dos frangos de corte na época do verão. São Paulo: Elanco Saúde Animal, 2002. Manual técnico. 1:45 Jain, N.C. Essentials of veterinary hematology. Philadelphia: Lea & Febiger, 1993. 98p. Lovatto, A.A. Desempenho de seis linhagens comerciais de matrizes de frango de corte, criados com pesos corporais diferentes durante a fase de recria. Dissertação (Mestrado em Zootecnia) - Escola Veterinária da UFMG, Universidade Federal de Minas Gerais, Belo Horizonte, Brasil, 1889. Macari, M. Estresse de calor em aves. In: Reunião 38 da Sociedade Brasileira de Zootecnia, 2001, Piracicaba. Anais. Piracicaba ; 1996. p. 686-716. Macari, M.; Furlan, R.L.; Maiorka, A. Aspectos fisiológicos e de manejo para manutenção da homeostase térmica e controle de síndromes metabólicas. In: Mendes, A. A.; Naas, I.A.; Macari, M. Produção de frangos de corte. Campinas: Facta; 2004. p.137-156.

35
May, J.D.; Lott, B.D. Relating weight gain and feed: gain of male and female broilers to rearing temperature”. Poult Sci., 80: p.581-584, 2001. Mc. Geehin, M.A. Mirabelli, M. The potencial impacts of climate variability and change on temperature – related morbidity and mortality in the United State. Environ Health Perspect. 109 Suppl: p.185-189, 2001. Melitzer, A. Efficiency of effect of high ambient temperature on food utilization in male broilers. Brit. Poult. Sci., v. 27: p.349-351, 1986. Morgan, W.E. Heat reflective roof coatings. Transac. ASAE, St.Joseph. v. 90: p.4513, 1990. Mujahid, A.; Yoshiki, Y.; Akiba, Y.; Toyomizu, M. Superoxide radical production in chicken skeletal muscle induced by acute heat stress. Poult. Sci., 84, p.307–314, 2005. Mujahid, A.; Pumford, N.R.; Bottje, W.; Nakagawa, K.; Miyazawa, T.; Akiba, Y.; Toyomizu, M.. Mitochondrial oxidative damage in chicken skeletal muscle induced by acute heat stress. J. Poult. Sci., 44, p.439–445, 2007a. National Research Council. Nutrient Requirements of Poultry. Washington: National Academy of Sciences, 1994. North, M.O. Manual de Producción avícola. México D.F, México: El Manual Moderno S.A. Editores, 1993. 235p. Oliveira Neto, A. R.; Oliveira, R.F.M.; Donzele, J.L. Metabolizable energy level for broilers from 22 to 42 days of age maintained under thermoneutral environment. Rev. Bras. Zootec., Viçosa-MG. v. 29(4): p.1132-1140, 2000. Post, J.; Rebel, M.J.; Terhuurne, A.A. Physiological effect of elevated plasma corticosterone concentration in broiler chickens. An alternative means by which to assess the physiological effect of stress. Poult. Sci., 82: p.1313-1318, 2003. Plavnik, I.; Yahav, S. Effect of environmental temperature on broiler chickens subjected to growth restriction at an early age. Poult. Sci., Champaign. v. 77(6): p.870-872,1998. Rostagno, H.S.; editor. Tabelas brasileiras para aves e suínos: composição de alimentos e exigências nutricionais.2.ed, Viçosa, Brasil: UFV Departamento de Zootecnia, 2005. Sainsbury, D. Poultry health and management. 4. ed. Blackwell Science, 2000. 232p. Simmons, J.D.; Lott, B.D.; Miles, D.M. The effects of high air velocity on broiler performance. Poult Sci., 82: p.232-234, 2003. Souza, P.A.; Souza, H.B.A.; Campos, F.P. et al. Desempenho e características de carcaça de diferentes linhagens comerciais de frango de corte. Revista da Sociedade Brasileira de Zootecnia, v.23, n.5, p.782-791, 1994.

36
Summers, J.D.; Sprat, D.; Atkinson, J.L. Restricted feeding and Compensatory growth for broilers. Poult., Sci., 69: 1855-1861, 1990. Teeter, R.G.; Smith, M.O.; Murray, E. Force feeding methodology and equipment for poultry”. Poult. Sci., Champaign. v. 63(4): p.573-575, 1984. Teeter, R.G.; Smith, M.O.; Wiernusz, C.J. Effect of broiler acclimation to heat stress and feed intake on body temperature in birds exposed to thermoneutral and high ambient temperatures. Poult. Sci., 71: 1101-1104, 1992. Tinôco, I.F.F. Ambiência e instalações para a avicultura industrial. In: Encontro Nacional De Técnicos, Pesquisadores E Educadores De Construções Rurais, 3., 1998, Poços de Caldas, MG. Anais” Poços de Caldas: Sociedade Brasileira de Engenharia Agrícola; 1996. 3: p.1-86. Veldkamp, T.; Kwakkel, R.P.; Ferket, P.R; Verstegent, M.W. Growth responses to dietary energy and lysine at high and low ambient temperature in male turkeys. Poult Sci., 84: p.273-282, 2005. Yahav, S.; Straschnow, A.; Vax, E.; Raspakovski, V.; Shinder, D. Air velocity alters broiler performance under harsh environmental conditios. Poult Sci., 80: p.724-726, 2001. Zevallos, M.G. Condiciones ambientales. Disponible en: <http://www.portalveterinaria.com> Accesado en: 14/08/2010.

37
Capitulo 2. PERFIL BIOQUÍMICO SANGUÍNEO Y CUADRO HEMÁTICO DEL POLLO DE ENGORDE BAJO CONDICIONES DE ESTRÉS CALORICO
2.1 Introducción
La avicultura moderna ha venido acompañada del desarrollo de un pollo de engorde más precoz, con gran eficiencia para convertir alimento en proteína animal, a pesar de esto, una serie de problemas metabólicos y de manejo han surgido, destacándose entre estos el estrés calórico. La susceptibilidad de las aves al estrés calórico aumenta a medida que la temperatura ambiental y humedad relativa sobrepasan la zona de confort térmico. El efecto detrimental del estrés calórico en la producción avícola ha sido constantemente estudiado, yahav, (2000); Lin et al., (2006). Los conocimientos adquiridos sugieren diferencias en la tolerancia al estrés calórico entre diferentes estirpes de pollo (Yahav, 1998; Deeb y Cahaner, 1999; Cahaner 2008; Islam y Nishibori, 2009), indicando que los linajes de rápido crecimiento son más sensibles a las altas temperaturas ambientales (Yunis y Cahaner, 1999). El estrés se refiere a un efecto ambiental sobre un individuo que sobrepasa su sistema de control y reduce su adaptabilidad (Siegel, 1989). Este síndrome de desequilibrio orgánico, induce una serie de mecanismos defensivos ante situaciones adversas que podrían comprometer la vida del animal, en donde la respuesta orgánica inicial se halla integrada coordinadamente por el sistema nervioso, endocrino y cardiovascular; promoviendo profundas modificaciones metabólicas y bioquímicas (Carsia y Harvey, 2000). Existen muchas propuestas de evaluación del estrés en las aves durante la etapa productiva. Uno de los indicadores que ha sido frecuentemente utilizado por los investigadores desde hace algún tiempo, no sólo en las aves, es el cambio en las respuestas fisiológicas del animal; como los niveles de corticosterona plasmática la cual caracteriza la respuesta de estrés de las aves, las concentraciones de metabolitos sanguíneos, la interpretación de los recuentos de leucocitos y algunas enzimas que reflejan el estado interno del animal; son algunos de los análisis más utilizados para estimar el estrés. Sin embargo, el valor descriptivo y/o predictivo de estas variables dependen de las características del estresor, de la respuesta del agente estresado y del contexto en que el estrés ocurre, Elrom (2000). En ciertas circunstancias, como en el estrés crónico, estas variables no tienen un comportamiento claro. Con el objetivo de contribuir al conocimiento de la producción de pollo de engorde bajo temperaturas ambientales inductoras de estrés calórico, se planteo evaluar el perfil

38
bioquímico sanguíneo y cuadro hemático del pollo de engorde sometido a condiciones ambientales del Piedemonte Amazónico Colombiano.
2.2 Materiales y métodos 2.2.1 Localización El experimento se realizó en la granja “Pasarela” ubicada en la zona rural del municipio de Florencia, (Caquetá, Colombia) ubicada a 1°34´35.58´´N y 75°32´39.68´´de longitud Occidental, 243 m.s.n.m. con temperatura media de 27°C, humedad relativa del 85% y precipitación anual de 3600 mm3.
2.2.2 Animales Fueron utilizados 250 pollos machos de la estirpe comercial Cobb-500 y 250 machos de la estirpe comercial Ross 308, previo al experimento las aves fueron clasificadas por peso y línea genética para conformar grupos homogéneos. Los animales con un peso promedio inicial de 773,56±8,28 g, se distribuyeron en corrales de 3,8 m2, para una densidad final de 6,9 aves/m2. Los tratamientos se organizaron en un diseño irrestrictamente al azar en un esquema bifactorial 2x2 (2 condiciones de temperatura y 2 estirpes comerciales de pollo de engorde) para un total de cuatro tratamientos, cinco repeticiones por tratamiento y 25 animales por unidad experimental.
2.2.3 Tratamientos T1: Pollos de engorde Cobb 500 machos de 22 días de edad, mantenidos a una temperatura de 30,0±1ºC (estrés calórico crónico) durante 21 días. T2: Pollos de engorde Cobb 500 machos de 22 días de edad, sometidos a la temperatura ambiente del Piedemonte Amazónico Colombiano durante 21 días. T3: Pollos de engorde Ross 308 machos de 22 días de edad, mantenidos a una temperatura de 30,0±1ºC (estrés calórico crónico) durante 21 días. T4: Pollos de engorde Ross 308 machos de 22 días de edad, sometidos a la temperatura ambiente del Piedemonte Amazónico Colombiano durante 21 días.
2.2.4 Manejo del pollo de Engorde A los grupos de aves se les aplicaron prácticas de manejo convencionales, en cuanto a planes sanitarios, nutricionales y alimenticios requeridos para cada etapa productiva (el suministro de alimento y agua fue a voluntad) (May et al., 2001). Los pollos fueron criados durante las tres primeras semanas de vida, en un corral dentro del módulo experimental y sometidos a una temperatura inicial de 32ºC ± 2ºC (Sainsbury, 2000), la temperatura se redujo gradualmente hasta alcanzar una temperatura ambiente 29,87±7,20ºC a los 22 días, tiempo en el cual se inicio la etapa experimental.

39
Las aves durante el período experimental recibieron alimento tipo migaja y agua a voluntad. La dieta experimental fue balanceada para contener 3100 Kcal de EM y 19,5% de proteína; a base de maíz, torta de soya y aceite vegetal, según las exigencias nutricionales, descritas por Rostagno (2005) y NRC (1994); para satisfacer los requerimientos nutricionales de las aves durante la fase de engorde (tabla 8). El monitoreo de la temperatura ambiental y la humedad relativa de cada ambiente fue realizado cada dos horas durante las 24 horas del día del periodo experimental (21 días), a través de un termo higrómetro digital de máximas y mínimas (Pacer industries Inc.
Modelo HTA 4200, con sensibilidad de 0,1C y 0,1%; Wisconsin, USA 1998) localizado a nivel de la altura de las aves. Tabla 8: Composición de la dieta experimental para pollos de engorde de 22 a 42 días
Ingrediente % Maíz 64,1 Torta de soya 46% 29,5 Aceite vegetal 2,4 Carbonato de calcio 1,47 Fosfato bicálcico 1,63 Sal 0,46 Premezcla Vitamínica3 0,05 Premezcla mineral4 0,10 Metionina HA 0,26 Colina 0,03 Niveles nutricionales
EM (kcal/kg) 3100 PB (%) 19,5 Lisina (%) 1,14 Metionina+cistina (%) 0,83 Arginina (%) 1,27 Treonina (%) 0,75 Triptófano (%) 0,24 Relación Arg:Lys 1,11 Ca (%) 0,95 P disponible (%) 0,42
Para garantizar durante todo el periodo experimental (22-42 días) una temperatura ambiental superior a los 30ºC se utilizaron criadoras a gas (tratamiento1 y 3). El programa de luz durante el periodo experimental fue continuo, 24 horas de luz natural/artificial (Fussell et al., 2003).
3 Premezcla vitamínica (Contenido por kg/ración): Vit.A. 10.000 UI; Vit D3 3.000 UI; Vit E 60 mg; Vit K3 3 mg; Vit B1 3 mg;
Vit. B2 8 mg; Vit B6 4 mg; Vit B12 0,014 mg; Ácido Pantoténico 20 mg; Niacina 50 mg; Ácido Fólico 2 mg; Biotina 0,15 mg 4 Premezcla mineral (Contenido por kg/ración): Fe 40 mg; Zn 80 mg; Mn 80 mg; Cu 10 mg; I 0,7 mg; Se 0,3 mg

40
2.2.5 Muestras A los 42 días de edad de las aves, mediante punción de la vena braquial (Samour, 2000), fueron extraídas 100 muestras de sangre, 5 por unidad experimental y 25 por cada tratamiento. Para todas las pruebas sanguíneas, se obtuvieron 10 ml de sangre/ave, 5 ml en tubo colector estéril para la realización de los perfiles bioquímicos y 5ml en tubo colector estéril para las pruebas hematologicas, adicionando anticoagulante.
2.2.6 Variables evaluadas El hematocrito (%): se realizo a través de la técnica de micro centrifugación; se utilizo una microcentrífuga con capilar a 14000 RPM por 5 minutos, expresado en unidades de porcentaje (Fudge, 2000). Nivel de hemoglobina (g/dL): Se detecto usando el método de la cianometahemoglobina; las lecturas se realizaron con epectrofotometro a 540 nm. El recuento total de leucocitos (mm3): Se realizo a través del método de Natt y Herrick. Este método directo tiño ligeramente de azul todos los glóbulos blancos permitiendo su identificación. Leucograma diferencial (%): Consistió en el conteo del número de los distintos tipos de leucocitos (heterofilos, linfocitos, eosinofilos, basofilos y monocitos) de las dos líneas de aves estudiadas. El desarrollo de esta variable se realizo a través del análisis de frotis sanguíneo teñido con coloración de Wright (tinción de tipo Romanowsky), contando un total de 200 células cuyo resultado se reporto en porcentaje (Fudge, 2000); con determinación de la relación heterofilos/linfocitos (H/L). Perfil bioquímico sanguíneo: A partir de 100 muestras de plasma (5 por unidad experimental y 25 por cada tratamiento), fueron analizados, proteína total, albumina, globulina, glucosa, sodio, cloro, potasio, calcio, magnesio y fósforo, Los reactivos utilizados para determinar los niveles de electrolitos y de glucosa pertenecían a los laboratorios SPINREACT®. Todas las pruebas bioquímicas fueron realizadas a través del método colorimétrico (espectrofotometría), cuyas concentraciones se determinaron con un espectrofotómetro SPECTRONIC® 20D, con una longitud de onda amplia de 340 a 950 nanómetros. Niveles de albumina, globulina y proteínas plasmáticas totales (g/dL): Fue utilizando el método colorimétrico de Biuret, se determino la concentración de proteínas en 100 muestras de plasma mediante espectroscopia ultravioleta-visible a una longitud de onda de 540 nm (para detectar el ion Cu2+). Corticosterona plasmática (ng/mL): Fueron utilizadas 100 muestras de sangre, 5 por unidad experimental y 25 por cada tratamiento. A partir de sangre adicionada con anticoagulante se obtuvieron 2 mL de plasma por ave para obtener los niveles de corticosterona.

41
Los niveles de corticosterona plasmática fueron medidos por la técnica de inmunoensayo directo ("Enzyme-Linked Immunoasorbent Assay"), técnica fundamentada en el uso de anticuerpos marcados con una enzima, para detectar la concentración de la hormona en las muestras de estudio. La absorbancia se determino con un espectrofotómetro lector de microplaca (Multiskan plus-Fisher scientific®) a 540 nanómetros. Se realizo la curva estándar para calcular las concentraciones de corticosterona plasmática (ng/mL) sobre la base de estándares. El kit de Elisa utilizado para determinar los niveles de corticosterona pertenecía a la marca ACCUBIND®. Todos los análisis fueron realizados en los laboratorios de diagnostico veterinario de las universidades de la Amazonia, Tolima, e ICA (Instituto Colombiano Agropecuario) sede Florencia (Caquetá).
2.2.7 Análisis estadístico Los resultados fueron sometidos a análisis de varianza, utilizando el procedimiento GLM (General Linear Models) de SAS-8.02 (2002). Las diferencias entre tratamientos fueron establecidas mediante la prueba de comparación múltiple de Tukey o la prueba de F a un nivel de confianza de (P<0,05).
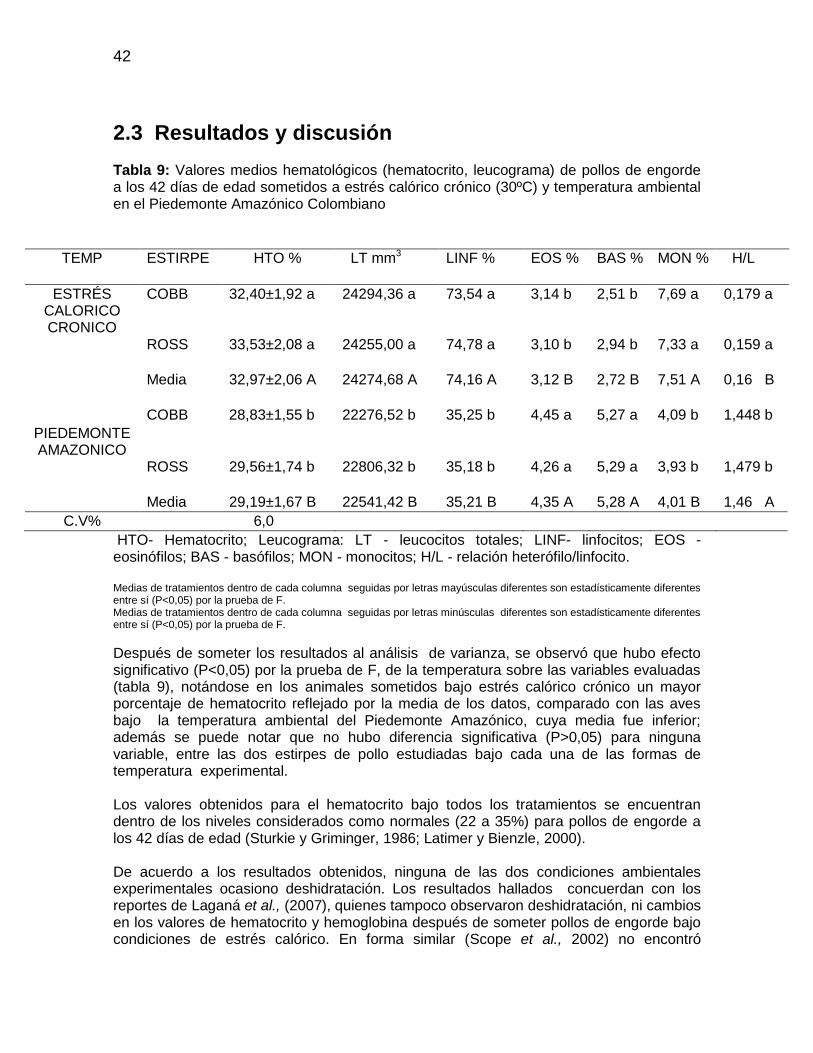
42
2.3 Resultados y discusión Tabla 9: Valores medios hematológicos (hematocrito, leucograma) de pollos de engorde a los 42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano
TEMP ESTIRPE HTO % LT mm3 LINF % EOS % BAS % MON % H/L
ESTRÉS CALORICO CRONICO
COBB 32,40±1,92 a 24294,36 a 73,54 a 3,14 b 2,51 b 7,69 a 0,179 a
ROSS 33,53±2,08 a 24255,00 a 74,78 a 3,10 b 2,94 b 7,33 a 0,159 a
Media 32,97±2,06 A 24274,68 A 74,16 A 3,12 B 2,72 B 7,51 A 0,16 B
PIEDEMONTE AMAZONICO
COBB 28,83±1,55 b 22276,52 b 35,25 b 4,45 a 5,27 a 4,09 b 1,448 b
ROSS 29,56±1,74 b 22806,32 b 35,18 b 4,26 a 5,29 a 3,93 b 1,479 b
Media 29,19±1,67 B 22541,42 B 35,21 B 4,35 A 5,28 A 4,01 B 1,46 A
C.V% 6,0 HTO- Hematocrito; Leucograma: LT - leucocitos totales; LINF- linfocitos; EOS - eosinófilos; BAS - basófilos; MON - monocitos; H/L - relación heterófilo/linfocito. Medias de tratamientos dentro de cada columna seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
Después de someter los resultados al análisis de varianza, se observó que hubo efecto significativo (P<0,05) por la prueba de F, de la temperatura sobre las variables evaluadas (tabla 9), notándose en los animales sometidos bajo estrés calórico crónico un mayor porcentaje de hematocrito reflejado por la media de los datos, comparado con las aves bajo la temperatura ambiental del Piedemonte Amazónico, cuya media fue inferior; además se puede notar que no hubo diferencia significativa (P>0,05) para ninguna variable, entre las dos estirpes de pollo estudiadas bajo cada una de las formas de temperatura experimental. Los valores obtenidos para el hematocrito bajo todos los tratamientos se encuentran dentro de los niveles considerados como normales (22 a 35%) para pollos de engorde a los 42 días de edad (Sturkie y Griminger, 1986; Latimer y Bienzle, 2000). De acuerdo a los resultados obtenidos, ninguna de las dos condiciones ambientales experimentales ocasiono deshidratación. Los resultados hallados concuerdan con los reportes de Laganá et al., (2007), quienes tampoco observaron deshidratación, ni cambios en los valores de hematocrito y hemoglobina después de someter pollos de engorde bajo condiciones de estrés calórico. En forma similar (Scope et al., 2002) no encontró

43
alteraciones en el valor del hematocrito y hemoglobina después de estresar aves de engorde; Puvadolpirod y Thaxton (2000), después de simular condiciones de estrés calórico, no evidenciaron diferencias en la cantidad de glóbulos rojos de aves inyectadas con la hormona ACTH. No obstante, en forma contraria Campbell y Dein, (1984); Latimer y Bienzle, (2000), determinan que bajo una alta temperatura y humedad relativa ambiental el porcentaje de hematocrito se encontraría por encima de los valores normales; igualmente Yahav et al., (1997) demostraron variación de los parámetros sanguíneos en pollos sometidos a estrés calórico agudo, en donde los valores del hematocrito aumentaron de 26,1% a 28,1%, con una disminución de los niveles de hemoglobina de 10,34 a 9,77g/dL. Los resultados hallados en este experimento bajo la temperatura estrés calórico crónico se explican como consecuencia de una adaptación a este tipo de estrés, en donde las aves no mostraron deshidratación aparente, representada en los valores del hemograma al día 42 de edad. Por otro lado los análisis hematológicos hallados en los animales bajo la temperatura ambiental del Piedemonte Amazónico, se pueden explicar por el efecto variable de la temperatura diurna y nocturna, en donde las aves tuvieron la opción de una recuperación homeostática en las horas más frescas de la noche, con retorno a los valores de hematocrito normales en un tiempo variable, según Puvadolpirod y Thaxton (2000), este fenómeno puede ocurrir dependiendo de la magnitud y duración del estimulo estresor inicial. Tabla 10: Valores medios de hemoglobina (g/dL) en pollos de engorde a los 42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano
ESTIRPE TEMPERATURA COBB ROSS PROMEDIO
Estrés calórico crónico
10,57±1,41 Aa 10,85±1,48 Aa 10,71 a
Piedemonte Amazónico
PROMEDIO
7,79±1,45 Ab 9,18±1,43 A
7,66±1,59 Ab 9,25±1,54 A
7,73 b
C.V% 15,58 16,65
Medias de tratamientos dentro de cada fila seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
En la tabla 10, se observa el análisis de los datos de la variable hemoglobina la cual muestra que no hubo diferencia estadística significativa (P>0,05) entre los dos linajes de aves bajo ninguna de las dos variantes de temperatura. Sin embargo, se puede notar una diferencia (P<0,05) entre los niveles de hemoglobina de las aves bajo la temperatura estrés calórico crónico respecto a los animales bajo la temperatura del Piedemonte Amazónico dentro de cada estirpe. Este resultado se puede explicar probablemente como

44
consecuencia de una mayor demanda de oxigeno de las aves al poner en marcha al máximo y en forma continua los mecanismos fisiológicos de termorregulación bajo esta forma de estrés crónico. Es importante precisar que los valores hallados para cada línea se encuentran dentro los rangos fisiológicos normales para la hemoglobina (7 -13 g/dL) de pollos de engorde a los 42 días de edad (Sturkie y Griminger,1986 ; Latimer y Bienzle, 2000). En la tabla 9, se exponen los resultados del leucograma, el cual evidencio diferencias estadísticas (P<0,05) por la prueba de F de la temperatura sobre el número de leucocitos totales, en donde las dos líneas de pollo bajo las dos temperaturas experimentales mostraron un recuento medio superior en el numero de leucocitos totales, resultado que se relaciona con una leucocitosis. Se puede notar en los dos linajes de aves bajo la temperatura estrés calórico crónico, un recuento medio superior en el número de leucocitos totales en relación con la temperatura ambiental del Piedemonte Amazónico, explicado posiblemente como el efecto de una mayor liberación de corticosterona durante el estrés crónico, resultados que coinciden con los reportes de Coles (1986); Ruckebush et al., (1994) y Cunningham (1999), quienes encontraron leucocitosis transitoria en aves bajo estrés calórico crónico comparado con aves bajo estrés calórico cíclico. Por otra parte no se observo una diferencia (P>0,05) entre los resultados del leucograma diferencial de las dos estirpes bajo cada una de las temperaturas experimentales. Después de analizar los valores obtenidos para los linfocitos bajo las dos temperaturas experimentales, se puede inferir la existencia de una linfocitosis, resultado que difiere a los hallados por Macari y Luquetti (2002), quienes afirman que bajo situaciones de estrés, la liberación de la hormona corticotropica (ACTH) genera una reducción en la cantidad de linfocitos circulantes, ayudando al incremento de la relación heterofilo/linfocito. Entre tanto, para las aves bajo la temperatura ambiental del Piedemonte Amazónico se encontró una moderada heteropenia, hallazgos que harían parte de la variabilidad de los componentes celulares, compatibles con las características del cuadro de estrés calórico, descrito dentro de la respuesta bifásica en las aves (Bogin et al., 1981; Maxwell et al., 1992), en donde en la fase secundaria se observa una linfocitosis y heteropenia (Perea et al., 1997). El valor calculado para los heterofilos de las aves Cobb y Ross bajo la temperatura del Piedemonte Amazónico, permite inferir heterofilia, comportamiento celular fisiológico que coincide con la respuesta bifásica de las aves en su fase primaria; pudiéndose explicar como parte del efecto estresor ocasionado por las condiciones ambientales variables del Piedemonte Amazónico Colombiano o como réplica de defensa frente a agentes agresivos bacterianos, ya que los heterofilos presentan enzimas lisosomales y actividad bactericida (Harmon, 1998; Campbell, 1995). Los hallazgos obtenidos en la presente investigación corroboran los reportes de Gross y Siegel (1983); Mitchell et al., (1992); Puvadolpirod y Thaxton (2000); Macari y Luquetti (2002) y Scope et al., (2002), quienes afirman que situaciones estresoras aumentan la cantidad de heterofilos en la circulación. No obstante, Laganá et al., (2007) hallo valores

45
bajos en el recuento de heterofilos (heteropenia) en pollos de engorde a los 42 días de edad, después de ser expuestos a condiciones ambientales de estrés calórico cíclico. En este trabajo se puede observar el efecto de la temperatura sobre las dos estirpes de aves, en donde bajo todos los tratamientos la relación heterofilo/linfocito (H/L) es diferente. La relación (H/L) hallada para las aves bajo estrés calórico crónico se encuentra más próxima a las condiciones de normalidad (tabla 9), comparado con los resultados de las aves bajo temperatura ambiental del Piedemonte Amazónico. Se puede determinar que la proporción normal de heterofilos/linfocitos (H/L) está alrededor de 0,5; cuando los pollos son sometidos a condiciones de estrés, esa relación aumenta (Gross y Siegel, 1983); (Macari y Luquetti, 2002). La menor relación (H/L) expresada por las aves Cobb y Ross bajo la temperatura estrés calórico crónico (tabla 9), se puede explicar posiblemente por la mejor adaptación de las aves a este tipo de estrés, en donde al día 42 de edad los animales no evidenciaron claras alteraciones en el aumento de la relación (H/L). El valor más alto de la relación heterofilos/linfocitos (H/L) se puede asociar con cuadros de estrés agudo, con estrésores que operan de manera intensa durante cortos periodos de tiempo (Davis et al., 2000). La relación (H/L) hallada (1,4) bajo la temperatura ambiental del Piedemonte Amazónico, puede ser asociada con factores generadores de estrés agudo propios de esta condición ambiental. Resultados similares han sido descritos por Altan et al., (2000), al reportar un aumento de la relación (H/L) de 1,6 en pollos de engorde después de someterlos bajo estrés calórico agudo durante dos horas a 39±1°C. Como efecto de la temperatura se encontraron diferencias entre los tratamientos respecto al número de eosinofilos y basofilos (tabla 9), observándose eosinofilia acompañada de basofilia, con un valor medio superior hallado en los animales bajo la temperatura ambiental del Piedemonte Amazónico Colombiano, el número elevado de eosinofilos posiblemente se explica por una respuesta inflamatoria aguda inusitada, favorecida por las condiciones experimentales. A pesar de aun no ser determinada totalmente la función exacta de los eosinofilos en las aves, se sabe que estas células participan en la fase inicial de la respuesta inflamatoria aguda en pollos de engorde (Campbell, 1994). Investigaciones realizadas por Laganá et al., (2007) no reportan diferencias en la cantidad de eosinofilos y basofilos después de someter pollos de engorde hasta el día 42 de edad, bajo condiciones de estrés calórico y temperatura termoneutral, de igual forma, Scope et al., (2002) no encontraron diferencias en la cantidad de eosinofilos y basofilos de palomas estresadas durante tres horas. Entre tanto, Gross y Siegel (1983); Maxwell y Robertson (1998), describen una basofilia significativa como respuesta de las aves al estrés calórico ambiental. Al observar el recuento de monocitos a través del leucograma, se evidencia en general un bajo número de monocitos (monocitopenia) bajo todos los tratamientos (tabla 9), resultados similares han sido reportados por Altan et al., (2000), quienes después de someter pollos de engorde bajo estrés calórico con temperaturas de 39±1°C al día 42 de edad, encontraron bajos recuentos en el número de monocitos.

46
Los resultados obtenidos pueden ser explicados como consecuencia directa del efecto quimiotactico de la hormona adreno corticotropina (ACTH) sobre los monocitos (Mc farlane et al., 1989), la cual es secretada bajo situaciones de estrés, como lo afirman Macari y Luquetti (2002) y Lin et al., (2006). Es posible notar un menor recuento de monocitos en los dos linajes de aves bajo la temperatura ambiental del Piedemonte Amazónico, comparado con los pollos sometidos a estrés calórico crónico, este valor inferior hace parte de los efectos del estrés calórico agudo, de acuerdo con Maxwell y Robertson (1998), quienes concluyen que un aumento súbito en la temperatura ambiental puede incrementar la temperatura corporal de las aves y generar cambios en los componentes de los leucocitos circulantes en los pollos. La monocitopenia hallada sugiere una baja respuesta fagocitica por parte de estas células, de acuerdo con (Morgulis, 2002), dentro de los estados de estrés agudo ocasionados por los factores ambientales del Piedemonte Amazónico Colombiano. El mayor valor hallado (7,69%) para los monocitos de las aves Cobb bajo la temperatura estrés calórico crónico, tentativamente represento una mejor capacidad de defensa celular de este linaje, denotado por una presunta mejor adaptación ante este tipo de estrés. Tabla 11: Valores medios plasmáticos de corticosterona (ng/mL) en pollos de engorde a los 42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano
ESTIRPE TEMPERATURA COBB ROSS PROMEDIO
Estrés calórico crónico
16,96±1,77 Aa 17,34 ±1,27 Aa 17,15 a
Piedemonte Amazónico
PROMEDIO
6,92±1,66 Ab
11,94±1,72 A
7,05±1,39 Ab
12,20±1,33 A
9,56 b
C.V% 14,40 10,90 Medias de tratamientos dentro de cada fila seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
Al analizar los índices plasmáticos de corticosterona se puede observar que no hubo diferencia estadística significativa (P>0,05) entre los dos linajes de aves bajo ninguna de las dos variantes de temperatura. No obstante, es posible apreciar una interacción significativa (P<0,05) entre la estirpe y la temperatura, evidenciada a través de un mayor nivel de corticosterona bajo la temperatura estrés calórico crónico, en cada una de las dos estirpes estudiadas. El resultado obtenido permite verificar un similar grado de susceptibilidad en los dos linajes frente a este tipo de estrés. Los mayores valores de corticosterona plasmática hallados, se pueden explicar por el efecto continuo de una alta temperatura, lo que a la postre genero una respuesta

47
hormonal de mayor frecuencia y duración, asociada con la adaptación y atenuación de la respuesta al estrés calórico. Una alta concentración de corticosterona plasmática, se puede considerar como el indicador más sensible y constante de la condición de estrés, Gross y Siegel, (1983). Los resultados obtenidos corroboran parcialmente los hallazgos de Virden et al., (2007), quienes después de someter pollos de engorde a estrés calórico crónico reportan valores plasmáticos altos de corticosterona (18,5 ng/mL) con una disminución en los niveles plasmáticos después del pico inicial. De manera análoga los resultados obtenidos se pueden relacionar con los reportes de varios investigadores, quienes determinan que niveles altos de corticosterona plasmática pueden indicar una forma de estrés crónico, (Edens y Siegel, 1975);(Eden, 1978); Puvadolpirod y Thaxton (2000); Post et al., (2003) y Shini et al., (2008). Entre tanto, los resultados de corticosterona plasmática hallados bajo la temperatura Piedemonte Amazónico, se relacionan con una exposición al estrés en forma aguda, lo cual pudo provocar un aumento rápido y de corta duración en la concentración plasmática de corticosterona, con una posible tendencia de retorno a los valores normales, una vez que el factor de estrés disminuyo durante las horas diurnas y nocturnas más frescas. Estos hechos confirman los hallazgos de Pitman (1990) y Yunianto et al., (1997), quienes
después de exponer pollos de engorde a temperaturas ambientales de 34C y 16C durante dos horas, reportan incrementos en los valores de corticosterona plasmática durante los siguientes 30 minutos, observando posteriormente disminución de los niveles de corticosterona respecto a los valores previos alcanzados a la exposición del estimulo estresor por más de dos horas, con retorno a los valores normales fisiológicos (2,49 - 6,40 ng/mL), según Edens y Siegel, (1975); Eden, (1978); Puvadolpirod y Thaxton (2000); Post et al., (2003) y Shini et al., (2008). La adaptación del eje hipotálamo-pituitaria-adrenal (HPA) al estrés se basa en la compleja interacción entre múltiples sistemas corporales (Fulford y Harbuz, 2000). El aumento en la concentración de corticosterona plasmática bajo la influencia del estrés calórico puede causar alteraciones prolongadas en los sistemas inmunológico, cardiovascular, digestivo y reproductivo, (Elrom, 2000). Es posible concluir, de acuerdo con Edens (1975); Siegel (1980); Beuving (1989); Puvadolpirod y Thaxton (2000); Shini et al., (2008), que los niveles de corticosterona plasmáticos pueden ser utilizados como un herramienta de medición de estrés calórico en los pollos de engorde.

48
Tabla 12: Valores medios plasmáticos de glucosa (mmol/L) en pollos de engorde a los 42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano
ESTIRPE TEMPERATURA COBB ROSS PROMEDIO
Estrés calórico crónico
13,07±1,06 Ab 12,37±1,17 Ab 12,72 b
Piedemonte Amazónico
PROMEDIO
15,84±0,89 Aa
14,45±1,70 A
16,51±1,53 Aa
14,44±2,48 A
16,18 a
C.V% 11,76 17,17
Medias de tratamientos dentro de cada fila seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
En la tabla 12, se puede observar una interacción significativa entre la estirpe y la temperatura, por lo tanto dentro de cada estirpe se comparan las aves sometidas a las dos temperaturas, (P<0,05) por la prueba de F. Las aves de las dos estirpes mostraron un comportamiento similar reflejado a través de la variable glucosa, frente a cada una de las dos formas temperatura experimental (tabla 12). El valor más alto para este índice en cada uno de los dos linajes se puede observar bajo la temperatura del Piedemonte Amazónico Colombiano. No obstante, todos los valores de glicemia hallados para ambas estirpes de pollo bajo las dos temperaturas se encuentran dentro los índices fisiológicos normales (11,10 a 22,2 mmol/L) según Noriega, (2000); la baja variabilidad en el nivel de glucosa se explica probablemente en función de los mecanismos homeostáticos eficientes del organismo, entre los que se involucra el control endocrino a través de la insulina y el glucagon sobre el glucógeno y los glucocorticoides en la gluconeogenesis. Resultados semejantes reporto Revidatti et al., (2002), al encontrar valores de glicemia normales en pollos de engorde a los 42 días de edad después de ser sometidos a estrés; contrario a estos resultados Puvadolpirod y Thaxton (2000), hallaron valores anormales de glucosa en pollos de engorde inyectados con ACTH. Entre tanto, González y Silva (2003), establecen que la concentración de glucosa puede aumentar durante el estrés crónico. Sin embargo, y aun sin observar una hiperglicemia, el valor significativo superior de la variable observado en los pollos Cobb y Ross bajo la temperatura del Piedemonte Amazónico, podría asociarse con cuadros de estrés agudo, en los cuales el estrés actúa en forma súbita, de acuerdo con Carsia et al., (2000) y Puvadolpirod y Thaxton (2000). El comportamiento glicemico de las aves bajo la temperatura estrés calórico crónico, no mostro cambios significativos entre las dos estirpes comparadas. La explicación de este resultado es confusa ya que los estresores crónicos, de acuerdo con Revidatti et al., (2002), generan interacciones que influyen con el metabolismo lipídico (Puvadolpirod y
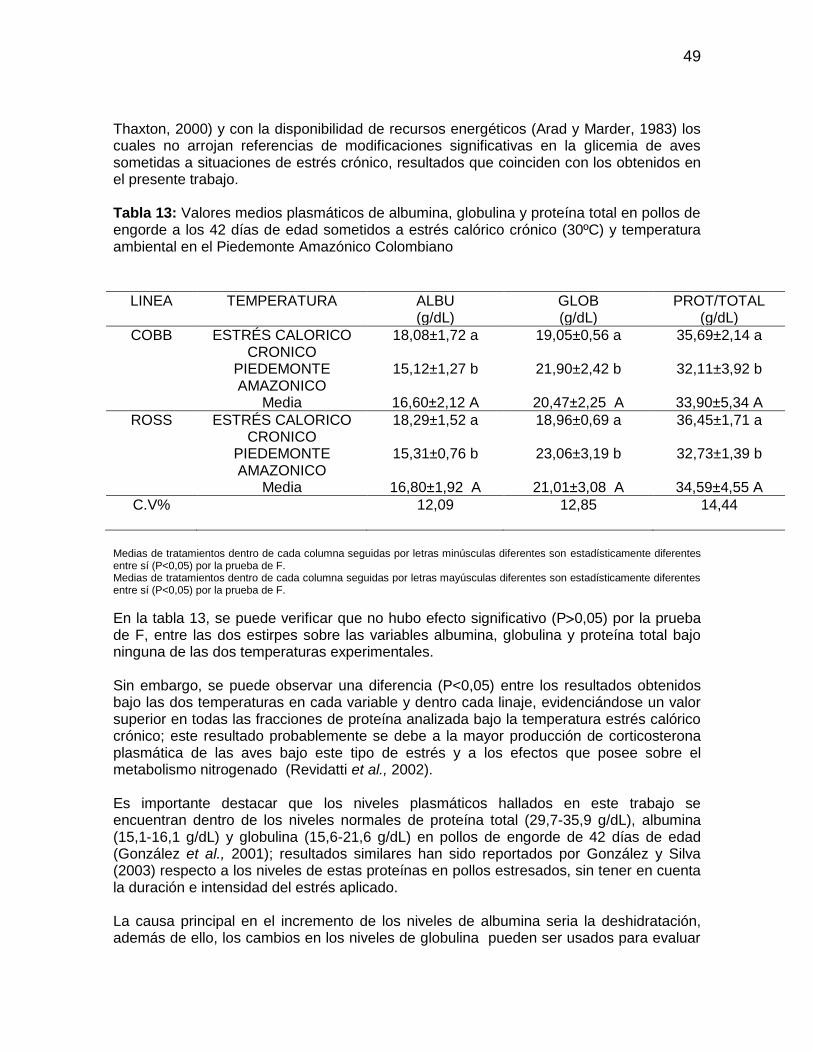
49
Thaxton, 2000) y con la disponibilidad de recursos energéticos (Arad y Marder, 1983) los cuales no arrojan referencias de modificaciones significativas en la glicemia de aves sometidas a situaciones de estrés crónico, resultados que coinciden con los obtenidos en el presente trabajo. Tabla 13: Valores medios plasmáticos de albumina, globulina y proteína total en pollos de engorde a los 42 días de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano
LINEA TEMPERATURA ALBU (g/dL)
GLOB (g/dL)
PROT/TOTAL (g/dL)
COBB ESTRÉS CALORICO CRONICO
18,08±1,72 a 19,05±0,56 a 35,69±2,14 a
PIEDEMONTE AMAZONICO
15,12±1,27 b 21,90±2,42 b 32,11±3,92 b
Media 16,60±2,12 A 20,47±2,25 A 33,90±5,34 A
ROSS ESTRÉS CALORICO CRONICO
18,29±1,52 a 18,96±0,69 a 36,45±1,71 a
PIEDEMONTE AMAZONICO
15,31±0,76 b 23,06±3,19 b 32,73±1,39 b
Media 16,80±1,92 A 21,01±3,08 A 34,59±4,55 A
C.V% 12,09 12,85 14,44
Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
En la tabla 13, se puede verificar que no hubo efecto significativo (P0,05) por la prueba de F, entre las dos estirpes sobre las variables albumina, globulina y proteína total bajo ninguna de las dos temperaturas experimentales. Sin embargo, se puede observar una diferencia (P<0,05) entre los resultados obtenidos bajo las dos temperaturas en cada variable y dentro cada linaje, evidenciándose un valor superior en todas las fracciones de proteína analizada bajo la temperatura estrés calórico crónico; este resultado probablemente se debe a la mayor producción de corticosterona plasmática de las aves bajo este tipo de estrés y a los efectos que posee sobre el metabolismo nitrogenado (Revidatti et al., 2002). Es importante destacar que los niveles plasmáticos hallados en este trabajo se encuentran dentro de los niveles normales de proteína total (29,7-35,9 g/dL), albumina (15,1-16,1 g/dL) y globulina (15,6-21,6 g/dL) en pollos de engorde de 42 días de edad (González et al., 2001); resultados similares han sido reportados por González y Silva (2003) respecto a los niveles de estas proteínas en pollos estresados, sin tener en cuenta la duración e intensidad del estrés aplicado. La causa principal en el incremento de los niveles de albumina seria la deshidratación, además de ello, los cambios en los niveles de globulina pueden ser usados para evaluar
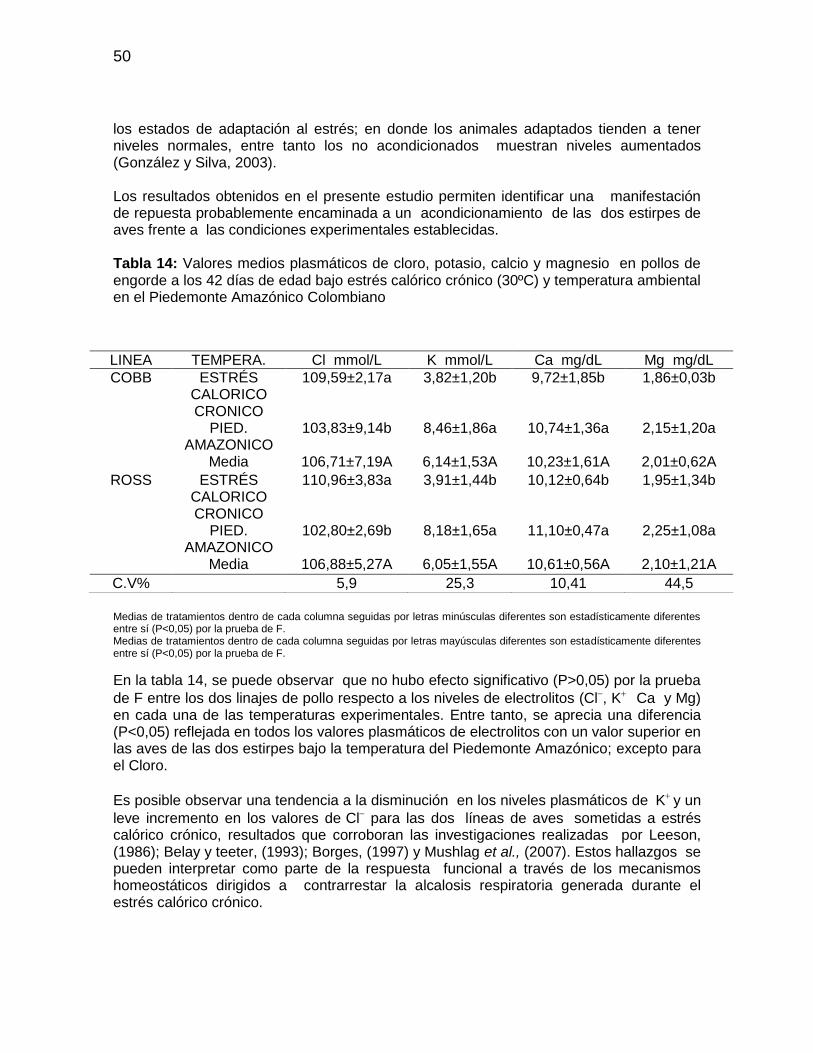
50
los estados de adaptación al estrés; en donde los animales adaptados tienden a tener niveles normales, entre tanto los no acondicionados muestran niveles aumentados (González y Silva, 2003). Los resultados obtenidos en el presente estudio permiten identificar una manifestación de repuesta probablemente encaminada a un acondicionamiento de las dos estirpes de aves frente a las condiciones experimentales establecidas. Tabla 14: Valores medios plasmáticos de cloro, potasio, calcio y magnesio en pollos de engorde a los 42 días de edad bajo estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano
Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
En la tabla 14, se puede observar que no hubo efecto significativo (P>0,05) por la prueba
de F entre los dos linajes de pollo respecto a los niveles de electrolitos (Cl, K Ca y Mg) en cada una de las temperaturas experimentales. Entre tanto, se aprecia una diferencia (P<0,05) reflejada en todos los valores plasmáticos de electrolitos con un valor superior en las aves de las dos estirpes bajo la temperatura del Piedemonte Amazónico; excepto para el Cloro.
Es posible observar una tendencia a la disminución en los niveles plasmáticos de K y un
leve incremento en los valores de Cl para las dos líneas de aves sometidas a estrés calórico crónico, resultados que corroboran las investigaciones realizadas por Leeson, (1986); Belay y teeter, (1993); Borges, (1997) y Mushlag et al., (2007). Estos hallazgos se pueden interpretar como parte de la respuesta funcional a través de los mecanismos homeostáticos dirigidos a contrarrestar la alcalosis respiratoria generada durante el estrés calórico crónico.
LINEA TEMPERA. Cl mmol/L K mmol/L Ca mg/dL Mg mg/dL COBB ESTRÉS
CALORICO CRONICO
109,59±2,17a 3,82±1,20b 9,72±1,85b 1,86±0,03b
PIED. AMAZONICO
103,83±9,14b 8,46±1,86a 10,74±1,36a 2,15±1,20a
Media 106,71±7,19A 6,14±1,53A 10,23±1,61A 2,01±0,62A ROSS ESTRÉS
CALORICO CRONICO
110,96±3,83a 3,91±1,44b 10,12±0,64b 1,95±1,34b
PIED. AMAZONICO
102,80±2,69b 8,18±1,65a 11,10±0,47a 2,25±1,08a
Media 106,88±5,27A 6,05±1,55A 10,61±0,56A 2,10±1,21A
C.V% 5,9 25,3 10,41 44,5

51
La disminución de los niveles plasmáticos de K+ se puede atribuir a un aumento en la excreción de estos iones durante el estrés calórico, consecuentemente la reducción en la retención de K+ (Smith y Teeter, 1987), afecta, asimismo, la concentración de K+ en el plasma (Borges, 1997; Salvador et al., 1999). El incremento en la excreción de K+ a través de la orina es variable, y está ligada a la concentración plasmática de Na+ y al estado de hidratación del ave, las perdidas pueden ser causadas por aumento en el consumo de agua, ya que el gradiente osmótico favorece el movimiento de agua del fluido celular para la formación de orina, pudiendo transportar el K+ (Belay y Teeter, 1993). Existen otros factores que también participan en la regulación de la excreción de K+, entre ellos están los hormonales (aldosterona, ADH-hormonaantidiurética y deoxicorticosterona), el equilibrio acido base y el balance de cationes (Borges, 1997).
El aumento en los niveles plasmáticos de Cl hallados en el presente experimento se justifica como parte del funcionamiento de un sistema tampón, en donde el cloro deprime la excreción de H+ y la reabsorción de HCO3 por los riñones, contribuyendo a la acidificación de la sangre como posible respuesta a la alcalosis Borges, (1997). Contrario a los anteriores hallazgos, Salvador et al., (1999), observaron una reducción de los
niveles séricos de Cl al someter pollos de engorde a estrés calórico crónico durante una semana desde los 42 a 49 días de edad. Las dos estirpes de pollo bajo la temperatura del Piedemonte Amazónico Colombiano mostraron una tendencia a incrementar los niveles de K+; el resultado obtenido probablemente se explica por una disminución en la excreción de este ion; este hallazgo confirma las investigaciones realizadas por Borges, (1997), las cuales reportan un aumento en los niveles de K+ en pollos de engorde bajo condiciones de estrés calórico agudo. Al observar los valores medios plasmáticos de calcio y magnesio (tabla 14), en las dos estirpes de pollo bajo la temperatura estrés calórico crónico, se evidencio una diferencia inferior en los índices plasmáticos de calcio (9,15%) y magnesio (13,4%) respectivamente, comparado con las aves bajo la temperatura del Piedemonte Amazónico; resultado que se explica como una consecuencia de la disminución en el consumo de alimento y por ende de electrolitos bajo la condición estrés calórico crónico (Klasing,1998). Sin embargo, los valores de calcio hallados como efecto de las dos temperaturas en cada uno de los dos linajes de aves, se encuentran dentro de los rangos fisiológicos normales de referencia (8.9-12.0 mg/dL) establecidos para pollos de engorde a los 42 días de edad (Kaneco,1989). Las concentraciones plasmáticas de calcio obtenidas, se pueden explicar probablemente mediante la relación directa con los valores normales de proteína plasmática hallados también en este experimento, debido a que un tercio de este calcio se encuentra unido a la albumina (Lumei, 1997). En forma similar los niveles de magnesio plasmático hallados bajo los cuatro tratamientos, también se encuentran dentro de los limites fisiológicos normales (1,82-2,78 mg/dL) para pollos de engorde con 42 días de edad (Kaneco,1989); resultado que se explica como parte de los ajustes homeostáticos realizados por las dos líneas genéticas bajo las

52
condiciones experimentales, en donde el nivel de estrés calórico experimental no afecto representativamente los valores de calcio y magnesio. Tabla 15: Valores medios plasmáticos de sodio y fosforo en pollos de engorde a los 42 días de edad sometidos a estrés calórico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano
Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
En la tabla 15, se puede observar una interacción significativa entre la estirpe y la temperatura, por lo tanto dentro de cada estirpe se comparan los resultados de las variables fosforo y sodio plasmático en las aves sometidas a las dos temperaturas, evidenciándose una diferencia (P<0,05) a favor de los animales bajo la temperatura del Piedemonte Amazónico. Por otra parte, se puede comprobar que no hubo efecto
significativo (P0,05) por la prueba de F, entre las dos líneas de pollo sobre los valores medios plasmáticos de sodio y fosforo bajo ninguna de las dos temperaturas experimentales. Al contrastar los resultados experimentales con los valores normales (7,2-7,81 mg/dL) plasmáticos para el fosforo en pollos de engorde a los 42 días de edad (Kaneco, 1989), se puede determinar una alta variación en los índices hallados para los dos linajes de aves sometidos bajo estrés calórico crónico, evidenciándose una diferencia inferior de 15,3% para el Cobb y 13,5% para el Ross. Los bajos niveles de fosforo encontrados pueden estar relacionados con un incremento en la excreción y disminución en la absorción a nivel intestinal y renal (Lumei, 1997 y Klasing, 1998). En cuadros de deshidratación asociados con poliuria y pasaje rápido del alimento bajo estrés calórico crónico, puede ocurrir un incremento en la excreción de algunos compuestos bioquímicos entre ellos el fosforo (Lumei, 1997). Entre tanto, los índices plasmáticos de fosforo obtenidos en los dos linajes bajo las condiciones ambientales del Piedemonte Amazónico, se encuentran dentro de los niveles normales de fosforo plasmático, probablemente en este experimento el estrés calórico cíclico ocurrido no genero alteraciones en su excreción y absorción a nivel renal e
LINEA TEMPERATURA P mg/dL Na mmol/L COBB ESTRÉS CALORICO
CRONICO 6,50±0,44 b 138,93±3,06 b
PIEDEMONTE AMAZONICO
7,76±0,68 a 145,15±23,05 a
Media 7,13±0,71 A 142,04±16,28 A
ROSS ESTRÉS CALORICO CRONICO
6,61±0,27 b 135,03± 5,24 b
PEDEMONTE AMAZONICO
7,63±0,44 a 148,89±5,68 a
Media 7,12±0,41 A 141,96±8,85 A
C.V% 7,86 8,85

53
intestinal; también existe la posibilidad de que este índice se pudo alterar solo en los momentos posteriores a la presentación del estrés agudo, regresando luego a su valor normal en un tiempo variable. Según Puvadolpirod y Thaxton, (2000) este fenómeno puede ocurrir dependiendo de la duración y magnitud del estimulo estresor inicial. Después de comparar los resultados de la variable sodio plasmático con los valores normales de referencia (140,50-149,50 mmol/L) para pollos de engorde a los 42 días de edad (Borges, 1997); se evidencia una disminución en el nivel de este ion para las dos líneas de aves expuestas bajo estrés calórico crónico observándose una diferencia inferior de 4,2% para el Cobb y 6,8% para el Ross (tabla 15); hallazgo que permite sugerir una alteración del equilibrio acido-básico con una probable variación en la presión osmótica, como resultado de una baja retención de sodio a nivel renal, favorecido ocasionalmente por un aumento en el consumo de agua en donde el gradiente osmótico pudo facilitar el movimiento del agua del fluido intracelular hacia la orina, transportando posiblemente más Na+ lo cual, incidiría en la presentación de una hiponatremia (Belay y Teeter, 1993); este fenómeno fisiológico es conocido como hemodilución (Borges et al., 2003), en donde además los tejidos empiezan a liberar K+ hacia el torrente sanguíneo debido a una alteración en la permeabilidad de las membranas, predisponiendo la presentación de signos neurológicos.

54
2.4 Conclusiones
Los resultados de las variables bioquímicas y hemáticas no fueron suficientemente contundentes, para explicar los posibles cambios séricos ocasionados por las dos condiciones de estrés calórico, sobre cada uno de los dos linajes de aves.
Las dos estirpes de aves mostraron igual grado de afectación bajo las dos
condiciones de estrés calórico experimental, representada por niveles similares de corticosterona plasmática.
Se pudo confirmar que la temperatura ambiental es uno de los factores abióticos
que pueden influir significativamente en el metabolismo y posterior rendimiento de los pollos de engorde.
Los niveles plasmáticos de corticosterona asociados correctamente a otros
parámetros fisiológicos, pueden ser utilizados como herramientas de medición de estrés calórico en aves de engorde.

55
Bibliografía Altan, O.; Altan, A.; Çabuk, M.; Bayraktar, H. Effects of heat stress on some blood parameters in broilers. Turk J. Vet. Anim. Sci., Tubitak, v. 24(2): p.45–148, 2000. Arad, Z.; Marder, W. Serum electrolyte and enzyme responses to heat stress and dehydration in the fowl (Gallus domesticus). Ph.D. Department of Zoology, The Hebrew University of Jerusalem.1983. 234p. Belay, T.; Teeter, R.G. Broiler water balance and thermobalance during thermoneutral and high ambient temperature exposure. Poult. Sci., Champaign. V. 72(2):p.116-124,1993. Beuving, G.; Jones R.B.; Blokhuis, H.J. Adrenocortical and heterophil/lymphocyte responses to challenge in hens showing short or long tonic immobility reactions. British Poultry Science. 30: p.175-184, 1989. Bogin, E.; Weisman, Y.; Friedman, Y. The effect of heat stress on the levels of certain blood constituents in chickens. Refuah Vet Israel, v. 38(1): p.98-104, 1981. Borges, S.A. Suplementação de cloreto de potássio e bicarbonato de sódio para frangos de corte durante o verão. Jaboticabal. PH D. Unesp Dissertação Mestrado. Programa de Pós-Graduação em Zootecnia, Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal, Brasil,1997. Borges, S.A.; Maiorka, A.; Silva, A.V.F. Fisiologia do estresse calórico e a utilização de eletrólitos em frangos de corte”. Ciênc. Rural, Santa Maria, v. 33(5): p.975-981, 2003. Cahaner, A.; Ajuh, J.A.; Siegmund-Schultze, M.; Azoulay, Y.; Druyan, S.; Valle Zarate, A. Effect of the genetically reduced feather coverage in naked neck and featherless broilers on their performance under hot conditions. Poult Sci. 87: p. 2517-2527, 2008. Campbell, T.W. Hematology. In: Ritchie, B.W.; Harrison, G.J.; Harrison, L.R. Avian Medicine: Principles and application”. Fort Worth-FL: Wingers Publishing.1: p.177-198, 1994. Campbell, T.W. 1995. Avian Hematology and Cytology. Ames: Iowa State Press. Carsia, R.V; Harvey, S. Adrenals. In: Sturkie’s Avian Physilogy, San Diego: Academic Press, 2000. 232p. Coles, E.H. Veterinary Clinical Patology. Philadelphia: W.B. Saunders, 1986. 143p. Cunningham, J.G. Tratado de Fisiologia Veterinária. Rio de Janeiro, Brasil: Guanabara, 1999. 112p. Davis, G.S.; Anderson, K.E.; Carrol, A.S. The effects of long-term caging and molt of single comb white leghorn hens on heterophil to lymphocyte ratios, corticosterone and thyroid hormones. Poult. Sci., Champaign, v. 79(4): p.514-518, 2000.

56
Deeb, N.; Cahaner, A. The effects of naked neck genotypes, ambient temperature, and feeding status and their interactions on body temperature and performance of broilers. Poult Sci. 78: p.1341-1346, 1999. Edens, F.W.; Siegel, H.S. Adrenal responses in high and low ACTH response lines of chickens during acute heat stress”. Gen. Comp. Endocrinol. 25: p.64-73, 1975. Edens, F.W. Adrenal cortical insufficiency in young chickens exposed to a high ambient temperature. Poultry Science. 57: p.1746–1750, 1978. Elrom, K. Handling and transportation of broilers: welfare, stress, fear and meat quality”. Israel J. Vet. Med., Raanana, v. 56(1): p.1–11, 2000. Fudge, A.M. Disorders of avian erythrocytes. In: Fudge AM. Avian Laboratory Medicine. Philadelphia PA: WB Saunders Company. 1: p.28-34, 2000. Fulford, A.J.; Harbuz, M.S. An introduction to the HPA axis. In: Steckler, T.; Kalin, N.H. Reul Handbook on stress, immunology and behavior”. JMHM, editors. Elsevier. 1: p.43–62, 2000. Fussell, L.; Rossi, A.Y.; Wilson, M. Lighting programs and coob 500 broiler performance. South África: Technical focus, 2003.107p. González, F.H.D.; Silva, S.C. Introdução a Bioquímica Clínica Veterinária. PH D. Porto Alegre, Brasil: Universidade Federal do Rio Grande do Sul, 2003.147p. Gross, W.B.; Siegel, H.S. Evaluation of the heterophil/ lymphocyte ratio as a measure of stress in chickens”. Avian Dis., Athens, v. 27(4): p.972-979, 1983. Harmon, B.G. Avian heterophils in inflammation and disease resistance”. Poult. Sci., Champaign. v. 77(6); p.972-977, 1998. Islam, M.A.; Nishibori, M. Indigenous naked neck chicken: a valuable genetic resource for Bangladesh. World Poult. Sci. J., p.65:125-138, 2009. Kaneko, J.J. Clinical Biochemistry of Domestic Animals. New York: Academic Press Inc, 1989.195p. Klasing, K. Comparative Avian Nutrition. Cab International. London. UK. 1998. Laganá, C. Níveis dietéticos de proteína e gordura e parâmetros bioquímicos, hematológicos e empenamento em frangos de corte estressados pelo calor. R. Bras. Zootec. v. 36(6): p.1783-1790, 2007. Latimer, K.S.; Bienzle, D. The avian leukogram: Determination and interpretation. In: Latimer, K.S.; Bienzle, D. Schalm’s Veterinary Hematology. Baltimore: Lippincott Williams & Wilkins, 2000. 208-216p.

57
Leeson, S. Nutritional considerations of poultry during heat stress”. World's Poult. Sci. J., Ithaca, v. 42: p.69-81, 1986. Lin, H. Acute heat stress induces oxidative stress in broiler chickens. Comparative Biochemistry and Physiology. Part A, Oxford, v. 144: p.11-17, 2006. Lumei, J.T. Avian clinical biochemistry. In: Clinical Biochemistry of Domestic Animals. Kaneko J.J.; Harvey J.W.; Bruss, M.L. (eds.). 5th ed. Academic Press. 1:1997 p.857-879. Macari, M.; Luquetti, B.C. Fisiologia Cardiovascular. In: Macari, M.; Furlan, R.L.; Gonzales, E. Fisiologia aviária aplicada a frangos de corte. Jaboticabal: Funep/Unesp, 2002. 232-240p. Mc farlane, J.M.; Stanley, C.E.; Shanks, R.D.; Carmer, S.G. Multiple concurrent stressors in chicks. 1. Effect on weight gain, feed intake, and behavior. Poultry Sci., 68: p.501-509, 1989. Maxwell, M.H.; Hocking, P.M.; Robertson, G.W. Differential leucocytes responses to various degrees of food restriction in broilers, turkeys and ducks”. Br. Poult. Sci., Oxforshire. v. 33(2): p.177-187, 1992. Maxwell, M.H.; Robertson, G.M. The avian heterophil leukocyte: a review. World’s Poultry Science Journal. 54: p.155-178, 1998. May, J.D.; Lott, B.D. Relating weight gain and feed: gain of male and female broilers to rearing temperature”. Poult Sci. 80: p.581-584, 2001. Mitchell, M.A.; Carlisle, A.J. The effect of chronic exposure to elevated environmental temperature on intestinal morphology and nutrient absorption in the domestic fowl (Gallus domesticus)”. Comp. Biochem. Physiol., New York. v. 101(1): p.137-142, 1992. Morgulis, M.S. Inmunología aplicada. In: Macari, M.; Furlan, R.L.; Gonzales, E. Fisiologia aviária aplicada a frangos de corte. Jaboticabal: Funep/Unesp, 2002. 1: 231-243p. Mushlag, J.; Mirza, M.A.; Athor, M.; Hooge, D.M.; Ahmad, T.; Ahmad, G.; Mushtag, M.M.; Noreen, U. Dietary sodium and chloride for 29 to 42 day old broilers at constant electrolyte balance under subtropical conditions. Jour. of Appl. Poultry Res. 16,p.161-170. 2007. National Research Council. Nutrient Requirements of Poultry. Washington: National Academy of Sciences, 1994. Noriega, M.L.V.C. Importância da hematologia no diagnóstico das aves. In: Encontro Técnico Sobre Avicultura de Corte da Região de Descalvado. Descalvado. Resumos... Descalvado, 2000. 4: p.1-11. Revidatti, F.A.; Fernandez, R.J.; Terraes, J.C.; Sandoval, G.L.; Luchi, P.E. Modificaciones del peso corporal y indicadores de estrés en pollos parrilleros sometidos a inmovilización y volteo. Rev. Vet. Arg., Corrientes. v. 12(1): p.1-8, 2002.

58
Rostagno, H.S.; editor. Tabelas brasileiras para aves e suínos: composição de alimentos e exigências nutricionais.2.ed, Viçosa, Brasil: UFV Departamento de Zootecnia, 2005. Perea, A.T.; Isaías, G.I.; Maldonado, F.G. Técnicas de medición de estrés en aves. Vet. Mex., México. v. 28(4): p.345-351,1997. Pitman, D.L.; Ottenweller, J.E.; Natelson, B.H. Effect of stressor intensity on habituation and sensitization of glucocorticoid responses in rats. Behav Neurosci. 104: p.28–36, 1990. Post, J.; Rebel, M.J.; Terhuurne, A.A. Physiological effect of elevated plasma corticosterone concentration in broiler chickens. An alternative means by which to assess the physiological effect of stress. Poult. Sci., 82: p.1313-1318, 2003. Puvadolpirod, S.; Thaxton, J.P. Model of Physiological Stress in Chickens 1. Response Parameters. Poult. Sci., Champaign. v. 79(4): p.363-369, 2000. Ruckebush, L.P.; Phaneuf, L.P.; Dunlop, R. F. Fisiologia de pequeñas y grandes especies. 1. ed. México D.F, México: Manual Moderno, 1994. 243p. Sainsbury, D. Poultry health and management. 4. ed. Blackwell Science, 2000. 232p. Salvador, D. Suplementação de bicarbonato de sódio na ração e na água de bebida de frangos de corte submetidos ao estresse calórico. ARS Veterinária. v. 15: p.144-148, 1999. Samour J. Clinical and diagnostic procedures. 3. ed. Livingston New York: Avian Medicine. Mosby, 2000.178p. Scope, A.; Filip, T; Glaber, C.; Resch, F. The influence of stress from transport and handling on hematologic and clinical chemistry blood parameters of racing pigeons (columba livia domestica). Avian Dis., Athens. v. 46(1): p.224-229, 2002. Shini, S.; Kaiser, P.; Shini, A.; Bryden, W.L. Biological response of chickens (Gallus gallus domesticus) induced by corticosterone and a bacterial endotoxin. Comp Biochem Physiol B; 149: p.324–33, 2008. Siegel, H.S. Physiological stress in birds. Bio Science. 301: p.529-533, 1980. Siegel, H.S. Stress, strains and resistance. Br. Poultry. Sci. 36: p.3-22, 1989. Sturkie, P.D.; Griminger, P. Body fluids: blood. In: STURKIE, P.D. Avian Physiology. New York: Springer – Verlag, 1986. p.102-129. Smith, M. O.; Teeter, R. G. Potassium balance of the 5 to 8-week-old broiler exposed to constant heat of cycling high temperature stress and the effects of supplemental potassium chloride on body weight gain and feed efficiency. Poult. Sci., Champaign. v. 66(3): p.487–492, 1987.

59
Virden, W.S.; Thaxton, J.P.; Corzo, A.; Dozier, IIIW.A.; Kidd, M.T. Evaluation of models using corticosterone and adrenocorticotropin to induce conditions mimicking physiological stress in commercial broilers. Poult Sci ;86: p.2485–91, 2007. Yahav, S.; Straschnow, A.; Plavnik, I.; Hurwitz, S. Blood system response of chickens to changes in environmental temperature. Poult. Sci., Champaign. v. 76(4): p.627-633, 1997. Yahav, S.; D.Lugar, A.; Cahaner, M.; Dotan, M.; Ruzanand.; Hurwitz, S. Thermoregulation in naked neck chickens subjected to different ambient temperatures. Br. Poult. Sci. 39: p.133-138, 1998. Yahav, S. Relative humidity at moderate ambient temperatures: its effect on male broiler chickens and turkeys”. Br. Poultry Sci., 41: p.94-100, 2000. Yuanianto, V.D.; K. Hayashi, S.; Kaneda, A.; Ohtsuka, Y.; Tomita, T. Effect of environmental temperature on muscle protein turnover and production in tube-fed broiler chickens. British Journal of Nutrition. 77: p.897-909, 1997. Yunis, R.; Cahaner, A. The effects of naked neck (Na) and frizzle (F) genes on growth and meat yield of broilers and their interactions with ambient temperatures and potential growth rate. Poult. Sci., 78: p.1347-1352, 1999.

60
Capitulo 3. ANATOMOFISIOLOGIA DEL POLLO DE ENGORDE BAJO CONDICIONES DE ESTRÉS CALORICO EN EL PIEDEMONTE AMAZÓNICO COLOMBIANO
3.1 Introducción La alta selección genética para obtener un rápido crecimiento en los pollos de engorde (Havenstein et al., 1994, 2003a) ha venido acompañada de un incremento del metabolismo (Janke et al., 2004), aumentando simultáneamente la producción de calor metabólico endógeno. La producción intensiva de estos linajes bajo una alta temperatura y humedad, hace a un más difícil la termorregulación de las aves, en parte debido a que el sistema cardio respiratorio no ha tenido un desarrollo análogo a los demás sistemas. Dentro de los mecanismos fisiológicos más eficientes para la disipación de calor utilizado por las aves, esta la evaporación respiratoria (Marder y Arad, 1989), la pérdida de calor evaporativo cutáneo (Webster y King, 1987; Ophir et al., 2002), la perdida de calor por radiación, la convección (Yahav et al., 2001), y la conducción (Wolfenson et al., 2001); No obstante estos procesos necesitan de un mayor esfuerzo muscular, generando a la postre un mayor aporte de energía con producción de más calor (Butcher y Miles, 2003), sobreviniendo la muerte rápidamente por agotamiento, principalmente en las aves más pesadas. El mayor problema en las zonas tropicales es el exceso de humedad relativa ambiental, la saturación del ambiente imposibilita que el ave elimine eficientemente calor a través de la respiración. La capacidad de las aves para soportar el calor es inversamente proporcional al porcentaje de humedad relativa ambiental, (Oliveira et al., 2000). La temperatura rectal normal de las aves esta alrededor de 41ºC (Chepete, 2008) con capacidad para conservarla dentro de un rango muy estrecho (Sturkie, 1986). El límite superior se ubica entre 41,5 - 42,0ºC y el inferior se encuentra entre 40,5-41,0ºC (Freeman, 1980). Bajo condiciones de estrés calórico, las aves reaccionan disminuyendo la producción de calor y aumentan la liberación del mismo (De Basilio et al., 2003). En consecuencia, la temperatura rectal aumenta y se sitúa en el rango de 42,0 - 43,5ºC (Yahav, 2000). De otra manera y en forma constante, la condición de estrés puede manifestarse a través de la una involución del sistema linforeticular acompañado de la atrofia y disminución del peso del timo, bolsa de Fabricio y bazo (Thaxton y Siegel, 1982; Cunningham, 1995; Aengwanich, 2008) e inmunodepresión más o menos prolongada (Espinet, 1987). El peso de los órganos linfoides se expresa como un porcentaje del peso corporal (Sandoval, 2002); con el cual usualmente se correlaciona en forma positiva (Ismail, 1990); al considerarlo de esta manera, es posible estimar con mayor precisión la incidencia de aquellos factores que actúan preponderantemente sobre el tejido linfoide (Rosales, 1989; Ismail, 1990). La pérdida de peso que acompaña a la atrofia y regresión

61
de los órganos linfoides constituye un indicador muy sensible del estrés en las aves (Dohms y Saif, 1984; Siegel, 1990; Cunningham, 1995). Con la finalidad de aportar al conocimiento de la producción de pollo de engorde bajo altas condiciones de temperatura y humedad relativa, el presente estudio tuvo como objetivo evaluar el efecto del estrés calórico en el Piedemonte Amazónico Colombiano sobre algunos parámetros anatomofisiológicos de dos estirpes de pollo de engorde.
3.2 Materiales y métodos
3.2.1 Localización El experimento se realizó en la granja “Pasarela” ubicada en la zona rural del municipio de Florencia, (Caquetá, Colombia) ubicada a 1°34´35. 58´´N y 75°32´39.68´´de longitud Occidental, 243 m.s.n.m. con temperatura media de 27°C, humedad relativa del 85% y precipitación anual de 3600 mm3.
3.2.2 Animales Fueron utilizados 250 pollos machos de la estirpe comercial Cobb-500 y 250 machos de la estirpe comercial Ross 308, previo al experimento las aves fueron clasificadas por peso y línea genética para conformar grupos homogéneos. Los animales con un peso promedio inicial de 773,56±8,28g, se distribuyeron en corrales de 3,8 m2, para una densidad final de 6,9 aves/m2. Los tratamientos se organizaron en un diseño irrestrictamente al azar en un esquema bifactorial 2x2 (2 condiciones de temperatura y 2 estirpes comerciales de pollo de engorde) para un total de cuatro tratamientos, cinco repeticiones por tratamiento y 25 animales por unidad experimental.
3.2.3 Tratamientos T1: Pollos de engorde Cobb 500 machos de 22 días de edad, mantenidos a una temperatura de 30,0±1ºC (estrés calórico crónico) durante 21 días. T2: Pollos de engorde Cobb 500 machos de 22 días de edad, sometidos a la temperatura ambiente del Piedemonte Amazónico Colombiano durante 21 días. T3: Pollos de engorde Ross 308 machos de 22 días de edad, mantenidos a una temperatura de 30,0±1ºC (estrés calórico crónico) durante 21 días. T4: Pollos de engorde Ross 308 machos de 22 días de edad, sometidos a la temperatura ambiente del Piedemonte Amazónico Colombiano durante 21 días.

62
3.2.4 Manejo del pollo de Engorde A los grupos de aves se les aplicaron prácticas de manejo convencionales, en cuanto a planes sanitarios, nutricionales y alimenticios requeridos para cada etapa productiva (el suministro de alimento y agua fue a voluntad), (May et al., 2001). Los pollos fueron criados durante las tres primeras semanas de vida, en un corral dentro del módulo experimental y sometidos a una temperatura inicial de 32ºC±2 ºC (Sainsbury, 2000), la temperatura se redujo gradualmente hasta alcanzar una temperatura ambiente 29,87±7,20ºC a los 22 días, tiempo en el cual se inicio la etapa experimental. Las aves durante el período experimental recibieron alimento tipo migaja y agua a voluntad. La dieta experimental fue balanceada para contener 3100 Kcal de EM y 19,5% de proteína; a base de maíz, torta de soya y aceite vegetal, según las exigencias nutricionales, descritas por Rostagno (2005) y NRC (1994); para satisfacer los requerimientos nutricionales de las aves durante la fase de engorde (Tabla 16). El monitoreo de la temperatura ambiente y la humedad relativa de cada ambiente fue realizado cada dos horas durante las 24 horas del día del periodo experimental (21 días), a través de un termo higrómetro digital de máximas y mínimas (Pacer industries Inc.
Modelo HTA 4200, con sensibilidad de 0,1C y 0,1%; Wisconsin, USA 1998) localizado a nivel de la altura de las aves. Tabla 16: Composición de la dieta experimental para pollos de engorde de 22 a 42 días
Ingrediente % Maíz 64,1 Torta de soya 46% 29,5 Aceite vegetal 2,4 Carbonato de calcio 1,47 Fosfato bicálcico 1,63 Sal 0,46 Premezcla Vitamínica5 0,05 Premezcla mineral6 0,10 Metionina HA 0,26 Colina 0,03 Niveles nutricionales
EM (kcal/kg) 3100 PB (%) 19,5 Lisina (%) 1,14 Metionina+cistina (%) 0,83 Arginina (%) 1,27 Treonina (%) 0,75
5 Premezcla vitamínica (Contenido por kg/ración): Vit.A. 10.000 UI; Vit D3 3.000 UI; Vit E 60 mg; Vit K3 3 mg; Vit B1 3 mg;
Vit. B2 8 mg; Vit B6 4 mg; Vit B12 0,014 mg; Ácido Pantoténico 20 mg; Niacina 50 mg; Ácido Fólico 2 mg; Biotina 0,15 mg6
Premezcla mineral (Contenido por kg/ración): Fe 40 mg; Zn 80 mg; Mn 80 mg; Cu 10 mg; I 0,7 mg; Se 0,3 mg

63
Tabla 16: (continuación) Triptófano (%)
0,24
Relación Arg:Lys 1,11 Ca (%) 0,95 P disponible (%) 0,42
Para garantizar durante todo el periodo experimental (22-42 días) una temperatura ambiente superior a los 30ºC se utilizaron criadoras a gas (tratamientos 1 y 3). El programa de luz durante el periodo experimental fue continuo, 24 horas de luz natural/artificial (Fussell et al., 2003).
3.2.5 Variables fisiológicas evaluadas Nivel de hiperventilación (NH): Se estableció en forma aleatoria en 5 aves por cada repetición y estirpe (Ross-Cobb) de cada tratamiento, para su medición, se empleó un cronómetro digital marca Casio® con precisión de 0,01 s. Se utilizó un cronómetro que se activaba al inicio del jadeo (el cual se reconoce por la apertura del pico, con aleteo en su base) con medición del tiempo que transcurre durante la realización de 10 inspiraciones consecutivas. A una distancia prudente se midió en forma individual a los pollos previamente identificados con colores por cada corral durante los 22 a 42 días de edad de la fase experimental, respectivamente, cada dos horas durante las 24 horas del día (mañana-tarde y noche). Para establecer el nivel de hiperventilación (NH) se multiplico el número de inspiraciones consecutivas medidas (10) por 60 segundos, el resultado obtenido fue dividido entre el tiempo transcurrido en el que ocurrieron las 10 inspiraciones consecutivas. El NH fue medido por dos observadores en todas las oportunidades, observando a cada ave marcada (Vilariño et al., 1999). Temperatura rectal (TR): Fue registrada aleatoriamente en 5 aves por cada repetición y estirpe (Ross-Cobb) de cada tratamiento, mediante un termómetro de alta precisión y rápida medida (Hansaplast® con sensibilidad de 0,4°C, Alemania 2002). Para ello, se introdujo el termómetro a través de la cloaca, localizado a nivel del recto terminal, hasta una profundidad de 3 cm (Yahav et a., 1998). La TR se midió de forma individual a los pollos previamente identificados por cada corral, durante todos los días a partir del día 22 hasta el día 42 de edad de la fase experimental, respectivamente, cada dos horas durante las 24 horas del día (mañana-tarde y noche).
3.5.2.3 Variables anatómicas evaluadas A los 42 días de edad y al momento del sacrificio (yugulación sin insensibilización) se registró el peso (g) del bazo (PB) y peso de la bolsa de Fabricio (PF), de 5 aves por repetición y estirpe (Ross-Cobb) de cada tratamiento, los órganos fueron retirados y secados con papel absorbente tipo toalla, posteriormente pesados mediante una balanza digital de precisión Sartorius® (Pine Brook, New Yersey, USA 2003), exactitud 0,1 g; con

64
una escala de medida mínima de un 0,1g y máxima de 7,500 g; para la determinación del peso relativo.
3.2.6 Análisis estadístico Los resultados fueron sometidos a análisis de varianza, utilizando el procedimiento GLM (General Linear Models) de SAS-8.02 (2002). Las diferencias entre tratamientos fueron establecidas mediante la prueba de comparación múltiple de Tukey o la prueba de F a un nivel de confianza de (P<0,05). Para observar el comportamiento e interacción de las variables estudiadas (temperatura rectal, nivel de hiperventilación) bajo las temperaturas experimentales, se realizaron pruebas de correlación, regresión lineal y cuadrática.
3.3 Resultados y discusión
3.3.1 Variables ambientales y fisiológicas La temperatura ambiental del Piedemonte Amazónico Colombiano durante todo el período experimental, mostro una diferencia inferior de 3,92% respecto a la temperatura registrada bajo la condición estrés calórico crónico (tabla 17). Las temperaturas registradas estuvieron dentro del rango establecido para las condiciones de estrés calórico (28-38ºC), con variaciones eventuales a temperaturas que configuraron un estrés agudo (Estrada y Márquez, 2005). La HR media registrada bajo la temperatura ambiental del Piedemonte Amazónico Colombiano, mostro un valor superior de 5,45% con respecto a la humedad relativa bajo la temperatura estrés calórico crónico. Tabla 17: Valores medios de temperatura y humedad relativa experimental
SEMANA/DIAS ESTRÉS CRÓNICO T°C
HR %
PIEDEMONTE AMAZONICO T°C
HR %
1 (28 días) 30,11±1,04 77,13±18,39 28,80±6,64 82,79±18,39 2 (35 días) 32,07±0,95 78,33±13,41 30,89±7,16 81,37±11,21 3 (42 días) 31,08±0,31 77,52±9,19 29,91±8,73 82,25±4,10
Media±DS 31,09±0,77 77,66±13,66 29,87±7,51 82,14±11,23
La humedad relativa estimada, revela que se supero el 65% citado como el óptimo para la cría de pollos de engorde en etapa de finalización (Yahav, 2000), alcanzando valores superiores al 80%, hecho que permite sugerir que la humedad relativa (HR) experimental pudo acentuar los efectos del estrés calórico en los pollos de engorde (Estrada y Márquez, 2005 y Farfán, 2008).
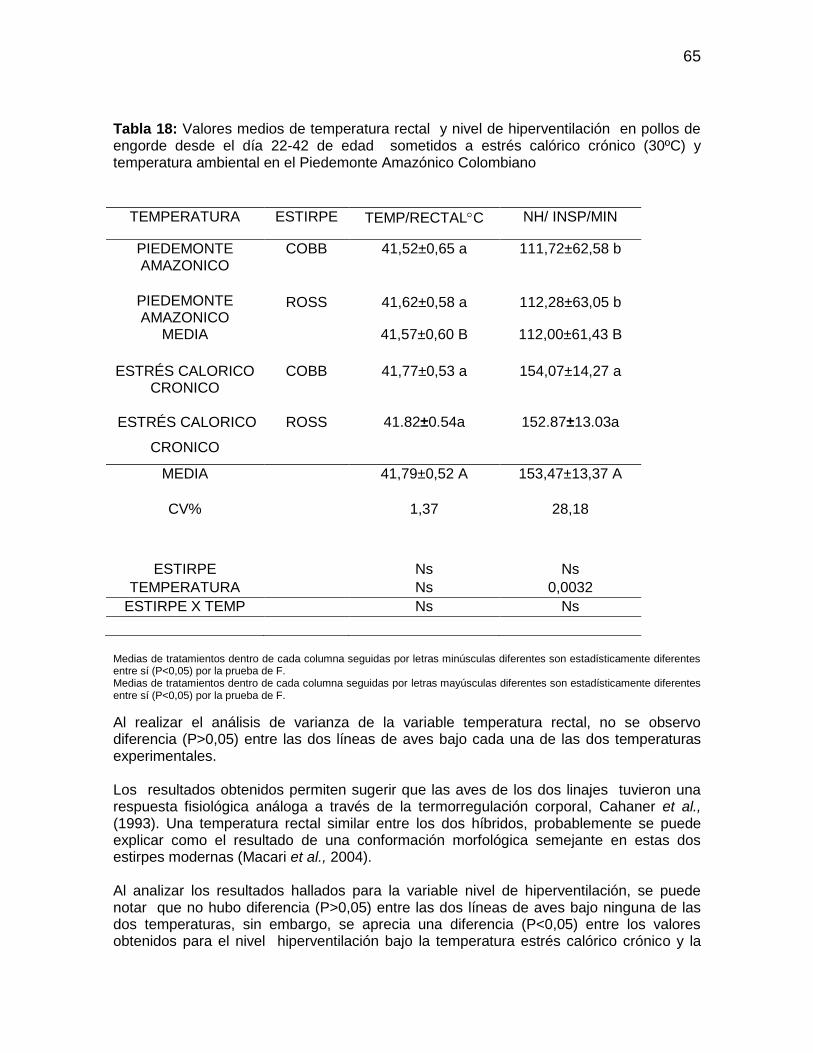
65
Tabla 18: Valores medios de temperatura rectal y nivel de hiperventilación en pollos de engorde desde el día 22-42 de edad sometidos a estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano
TEMPERATURA ESTIRPE TEMP/RECTALC NH/ INSP/MIN
PIEDEMONTE AMAZONICO
PIEDEMONTE AMAZONICO
COBB
ROSS
41,52±0,65 a
41,62±0,58 a
111,72±62,58 b
112,28±63,05 b
MEDIA 41,57±0,60 B 112,00±61,43 B
ESTRÉS CALORICO
CRONICO COBB 41,77±0,53 a 154,07±14,27 a
ESTRÉS CALORICO
CRONICO
ROSS 41.82±0.54a 152.87±13.03a
MEDIA 41,79±0,52 A 153,47±13,37 A
CV% 1,37 28,18
ESTIRPE Ns Ns
TEMPERATURA Ns 0,0032
ESTIRPE X TEMP Ns Ns
Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
Al realizar el análisis de varianza de la variable temperatura rectal, no se observo diferencia (P>0,05) entre las dos líneas de aves bajo cada una de las dos temperaturas experimentales. Los resultados obtenidos permiten sugerir que las aves de los dos linajes tuvieron una respuesta fisiológica análoga a través de la termorregulación corporal, Cahaner et al., (1993). Una temperatura rectal similar entre los dos híbridos, probablemente se puede explicar como el resultado de una conformación morfológica semejante en estas dos estirpes modernas (Macari et al., 2004). Al analizar los resultados hallados para la variable nivel de hiperventilación, se puede notar que no hubo diferencia (P>0,05) entre las dos líneas de aves bajo ninguna de las dos temperaturas, sin embargo, se aprecia una diferencia (P<0,05) entre los valores obtenidos para el nivel hiperventilación bajo la temperatura estrés calórico crónico y la

66
temperatura del Piedemonte Amazónico, notándose en las aves de las dos líneas bajo estrés calórico crónico una diferencia superior de 26,7% en el nivel de hiperventilación respecto a las mismas bajo la temperatura del Piedemonte Amazónico. Los resultados obtenidos en este trabajo corroboran los altos valores en el nivel de hiperventilación (140 – 200 insp/min) reportados por Mather et al., (1980) y Colina (2007) después de someter pollos de engorde a estrés calórico crónico; en forma similar El hadi
y Sykes, (1982) después de someter aves comerciales a 32C durante una hora, hallaron
un nivel de hiperventilación de 160 insp/min con una temperatura corporal de 41,8C; de manera semejante, Zhou y Yamamoto, (1997), al experimentar con pollos de engorde
bajo una temperatura ambiental artificial de 32C durante dos horas, registraron frecuencias inspiratorias superiores a 155 resp/min acompañada de una temperatura
rectal de 41,9C. El menor nivel de hiperventilación calculado en las aves bajo la temperatura del Piedemonte Amazónico, se explica como el efecto de los periodos de termoneutralidad registrados en algunas horas del dia y noche durante el periodo experimental, permitiendo a los animales almacenar menos calor. Estos hallazgos corroboran los reportes de Teeter et al., (1992), quienes afirman que bajo condiciones de estrés calórico cíclico las aves tienen la capacidad de disminuir su temperatura corporal y nivel de hiperventilación en las horas más frescas del día. La hiperventilación hallada, posiblemente se explica como parte de la respuesta fisiológica termorreguladora, en la cual las aves buscan elevar la evaporación del tracto respiratorio para mantener la homotermia durante el sobrecalentamiento corporal. La evaporación respiratoria a través de la hiperventilación, es el último recurso fisiológico eficiente disponible para la eliminación de calor después de un incremento en la temperatura corporal (Marder y Arad, 1989). No obstante, algunos investigadores cuestionan el posible efecto detonante del aumento de la temperatura corporal sobre el incremento en el nivel de hiperventilación de las aves. De Basilio et al., (2003), menciona que no se puede aseverar si realmente el nivel de hiperventilación es producto del aumento de la temperatura corporal, o si la temperatura corporal alta es consecuencia del nivel de hiperventilación. Al respecto Butcher y Miles, (2003) concluyen que durante la evaporación respiratoria a través de la hiperventilación, se incrementa el trabajo y contracción de los músculos participantes en el proceso respiratorio, con incremento del gasto energético y una consecuente mayor generación de calor e incremento de la temperatura corporal. Sin embargo, De Basilio et al., (2003), determina que bajo condiciones de estrés artificial si se puede establecer una relación clara entre el nivel de hiperventilación y la temperatura corporal. Por otra parte, al realizar el análisis de correlación entre las variables ganancia de peso y temperatura del Piedemonte Amazónico para los dos linajes de aves, se puede observar una correlación negativa significativa (P<0,05) con un coeficiente de (-O,32087), indicando que al incrementarse la temperatura ambiental disminuye la ganancia de peso (figura. 1); esto esencialmente se explica por una disminución en el consumo de alimento como tentativa para minimizar la cantidad de calor generado por la digestión y el metabolismo energético del alimento (Simmons et al., 1997; Simmons et al., 2003 y Veldkamp et al., 2005); presentándose consecuentemente una disminución en la ganancia de peso. Estos resultados son coherentes a los obtenidos por Melitzer (1986);
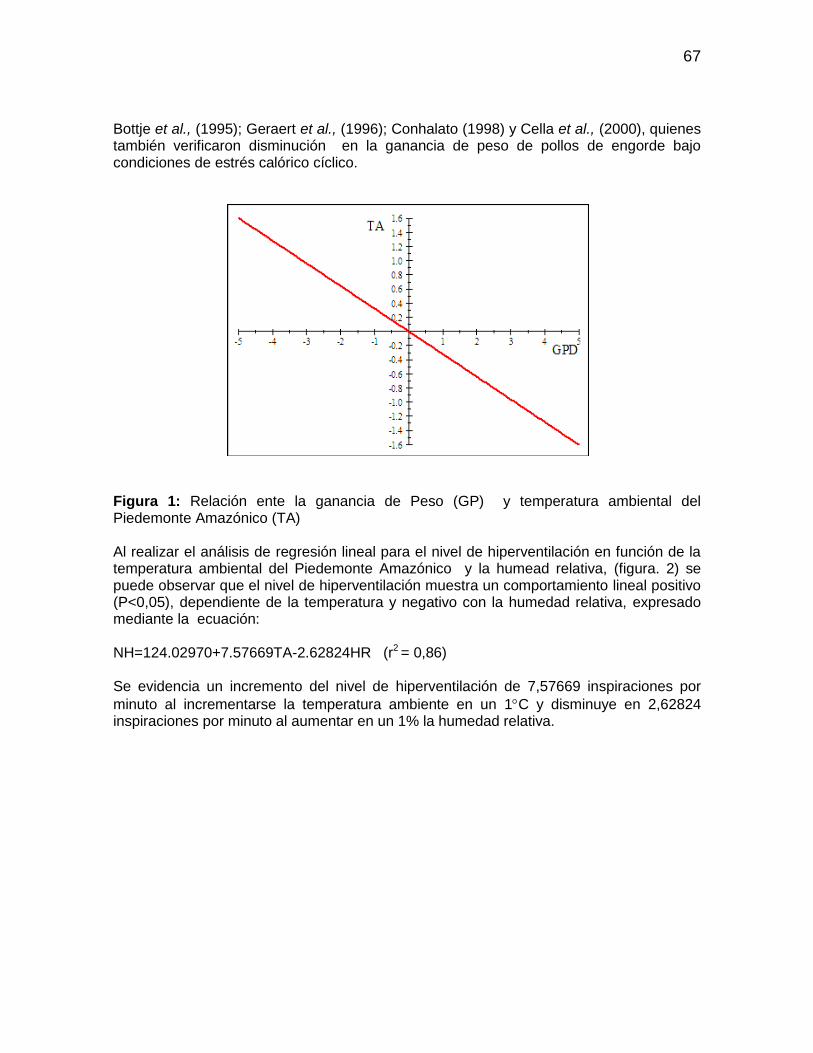
67
Bottje et al., (1995); Geraert et al., (1996); Conhalato (1998) y Cella et al., (2000), quienes también verificaron disminución en la ganancia de peso de pollos de engorde bajo condiciones de estrés calórico cíclico.
Figura 1: Relación ente la ganancia de Peso (GP) y temperatura ambiental del Piedemonte Amazónico (TA) Al realizar el análisis de regresión lineal para el nivel de hiperventilación en función de la temperatura ambiental del Piedemonte Amazónico y la humead relativa, (figura. 2) se puede observar que el nivel de hiperventilación muestra un comportamiento lineal positivo (P<0,05), dependiente de la temperatura y negativo con la humedad relativa, expresado mediante la ecuación: NH=124.02970+7.57669TA-2.62824HR (r2 = 0,86) Se evidencia un incremento del nivel de hiperventilación de 7,57669 inspiraciones por
minuto al incrementarse la temperatura ambiente en un 1C y disminuye en 2,62824 inspiraciones por minuto al aumentar en un 1% la humedad relativa.
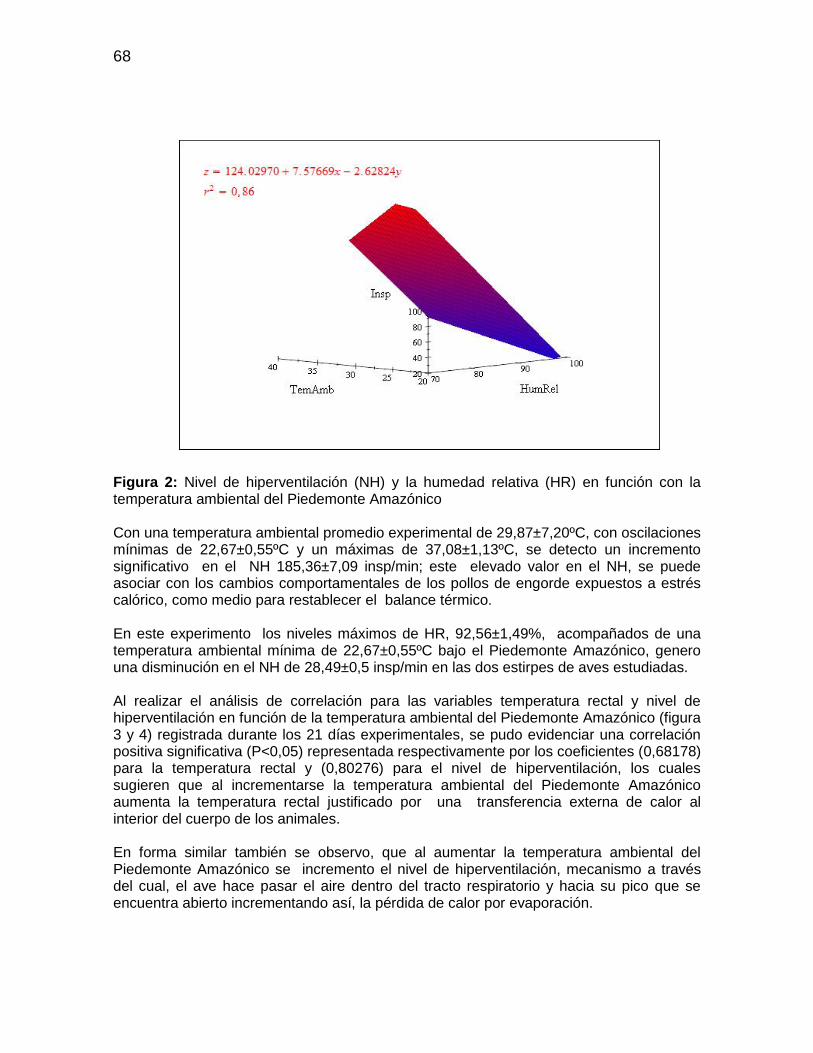
68
Figura 2: Nivel de hiperventilación (NH) y la humedad relativa (HR) en función con la temperatura ambiental del Piedemonte Amazónico Con una temperatura ambiental promedio experimental de 29,87±7,20ºC, con oscilaciones mínimas de 22,67±0,55ºC y un máximas de 37,08±1,13ºC, se detecto un incremento significativo en el NH 185,36±7,09 insp/min; este elevado valor en el NH, se puede asociar con los cambios comportamentales de los pollos de engorde expuestos a estrés calórico, como medio para restablecer el balance térmico. En este experimento los niveles máximos de HR, 92,56±1,49%, acompañados de una temperatura ambiental mínima de 22,67±0,55ºC bajo el Piedemonte Amazónico, genero una disminución en el NH de 28,49±0,5 insp/min en las dos estirpes de aves estudiadas. Al realizar el análisis de correlación para las variables temperatura rectal y nivel de hiperventilación en función de la temperatura ambiental del Piedemonte Amazónico (figura 3 y 4) registrada durante los 21 días experimentales, se pudo evidenciar una correlación positiva significativa (P<0,05) representada respectivamente por los coeficientes (0,68178) para la temperatura rectal y (0,80276) para el nivel de hiperventilación, los cuales sugieren que al incrementarse la temperatura ambiental del Piedemonte Amazónico aumenta la temperatura rectal justificado por una transferencia externa de calor al interior del cuerpo de los animales. En forma similar también se observo, que al aumentar la temperatura ambiental del Piedemonte Amazónico se incremento el nivel de hiperventilación, mecanismo a través del cual, el ave hace pasar el aire dentro del tracto respiratorio y hacia su pico que se encuentra abierto incrementando así, la pérdida de calor por evaporación.
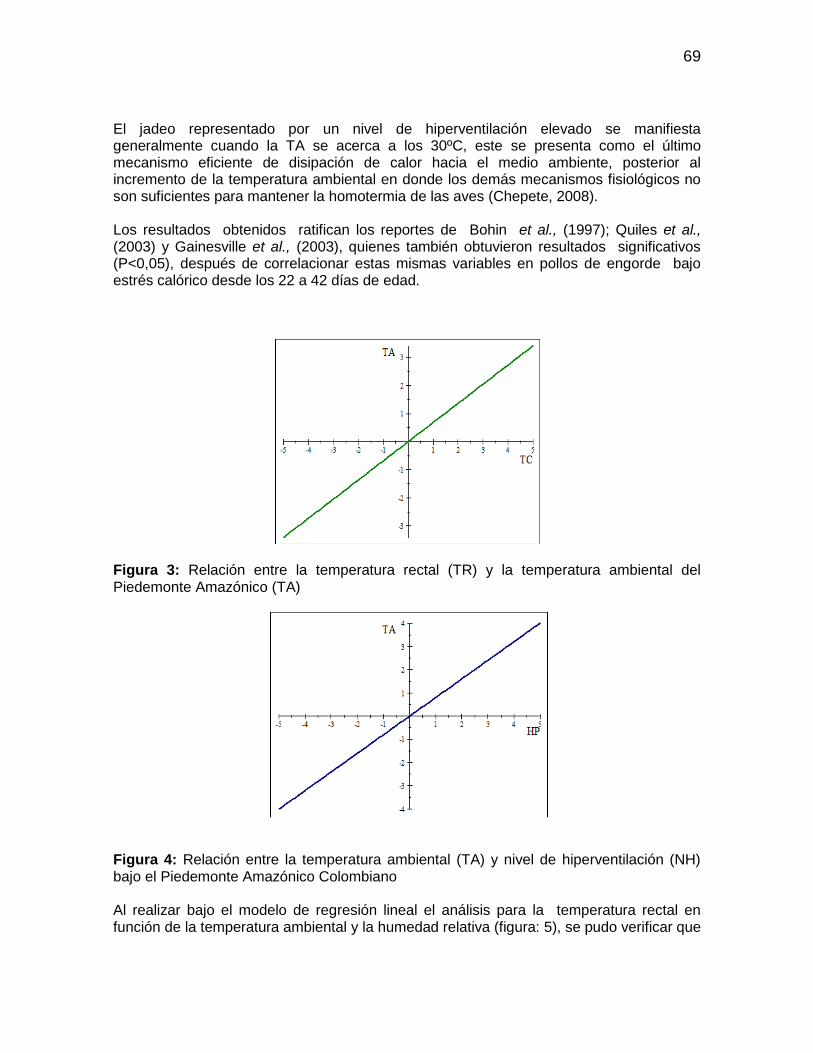
69
El jadeo representado por un nivel de hiperventilación elevado se manifiesta generalmente cuando la TA se acerca a los 30ºC, este se presenta como el último mecanismo eficiente de disipación de calor hacia el medio ambiente, posterior al incremento de la temperatura ambiental en donde los demás mecanismos fisiológicos no son suficientes para mantener la homotermia de las aves (Chepete, 2008). Los resultados obtenidos ratifican los reportes de Bohin et al., (1997); Quiles et al., (2003) y Gainesville et al., (2003), quienes también obtuvieron resultados significativos (P<0,05), después de correlacionar estas mismas variables en pollos de engorde bajo estrés calórico desde los 22 a 42 días de edad.
Figura 3: Relación entre la temperatura rectal (TR) y la temperatura ambiental del Piedemonte Amazónico (TA)
Figura 4: Relación entre la temperatura ambiental (TA) y nivel de hiperventilación (NH) bajo el Piedemonte Amazónico Colombiano Al realizar bajo el modelo de regresión lineal el análisis para la temperatura rectal en función de la temperatura ambiental y la humedad relativa (figura: 5), se pudo verificar que
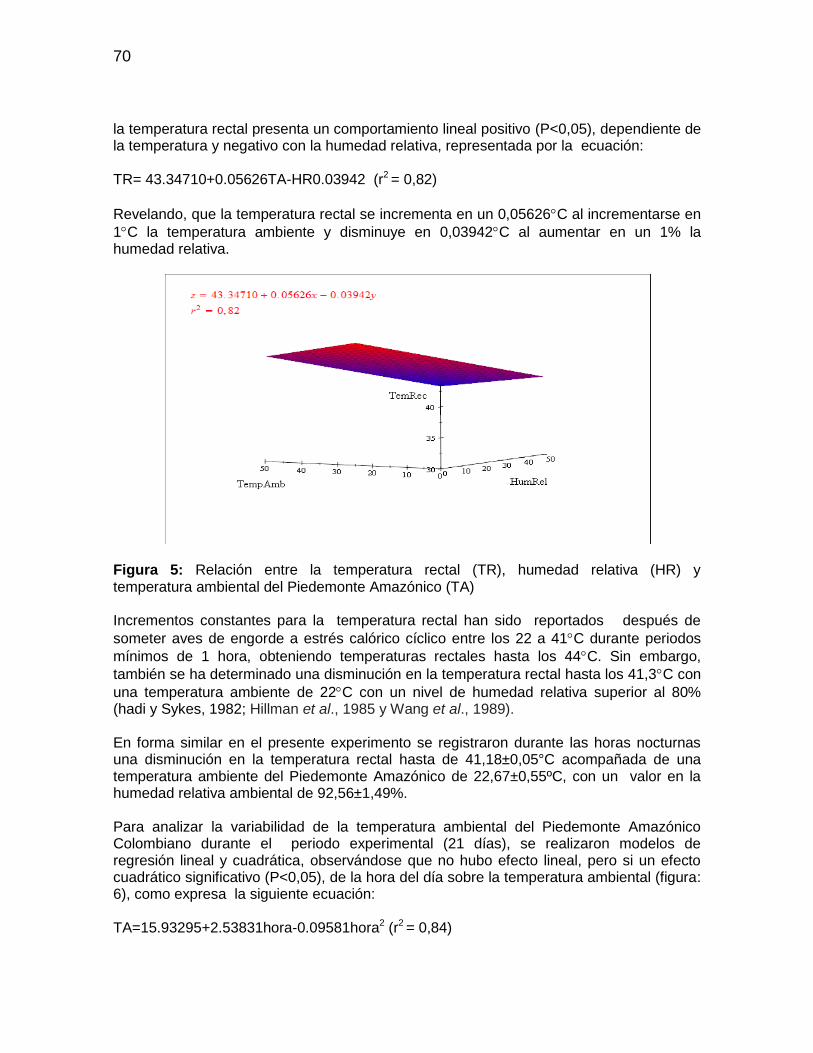
70
la temperatura rectal presenta un comportamiento lineal positivo (P<0,05), dependiente de la temperatura y negativo con la humedad relativa, representada por la ecuación: TR= 43.34710+0.05626TA-HR0.03942 (r2 = 0,82)
Revelando, que la temperatura rectal se incrementa en un 0,05626C al incrementarse en
1C la temperatura ambiente y disminuye en 0,03942C al aumentar en un 1% la humedad relativa.
Figura 5: Relación entre la temperatura rectal (TR), humedad relativa (HR) y temperatura ambiental del Piedemonte Amazónico (TA) Incrementos constantes para la temperatura rectal han sido reportados después de
someter aves de engorde a estrés calórico cíclico entre los 22 a 41C durante periodos
mínimos de 1 hora, obteniendo temperaturas rectales hasta los 44C. Sin embargo,
también se ha determinado una disminución en la temperatura rectal hasta los 41,3C con
una temperatura ambiente de 22C con un nivel de humedad relativa superior al 80% (hadi y Sykes, 1982; Hillman et al., 1985 y Wang et al., 1989). En forma similar en el presente experimento se registraron durante las horas nocturnas una disminución en la temperatura rectal hasta de 41,18±0,05°C acompañada de una temperatura ambiente del Piedemonte Amazónico de 22,67±0,55ºC, con un valor en la humedad relativa ambiental de 92,56±1,49%. Para analizar la variabilidad de la temperatura ambiental del Piedemonte Amazónico Colombiano durante el periodo experimental (21 días), se realizaron modelos de regresión lineal y cuadrática, observándose que no hubo efecto lineal, pero si un efecto cuadrático significativo (P<0,05), de la hora del día sobre la temperatura ambiental (figura: 6), como expresa la siguiente ecuación: TA=15.93295+2.53831hora-0.09581hora2 (r2 = 0,84)
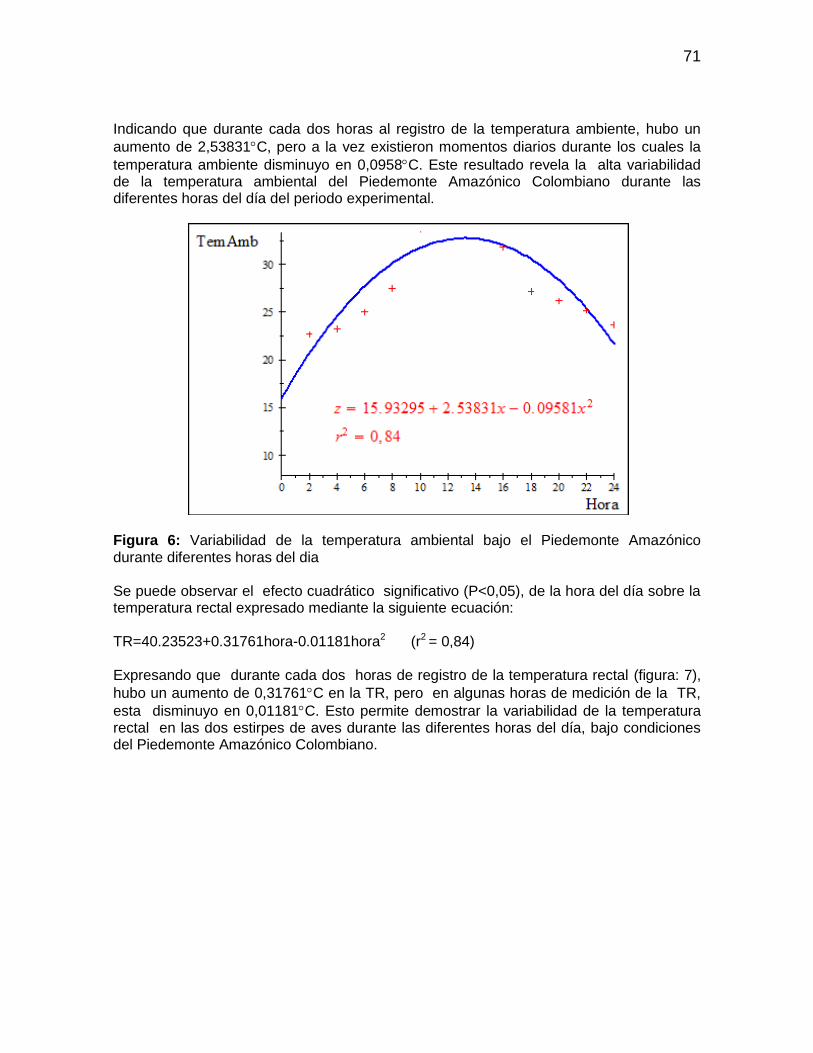
71
Indicando que durante cada dos horas al registro de la temperatura ambiente, hubo un
aumento de 2,53831C, pero a la vez existieron momentos diarios durante los cuales la
temperatura ambiente disminuyo en 0,0958C. Este resultado revela la alta variabilidad de la temperatura ambiental del Piedemonte Amazónico Colombiano durante las diferentes horas del día del periodo experimental.
Figura 6: Variabilidad de la temperatura ambiental bajo el Piedemonte Amazónico durante diferentes horas del dia Se puede observar el efecto cuadrático significativo (P<0,05), de la hora del día sobre la temperatura rectal expresado mediante la siguiente ecuación: TR=40.23523+0.31761hora-0.01181hora2 (r2 = 0,84) Expresando que durante cada dos horas de registro de la temperatura rectal (figura: 7),
hubo un aumento de 0,31761C en la TR, pero en algunas horas de medición de la TR,
esta disminuyo en 0,01181C. Esto permite demostrar la variabilidad de la temperatura rectal en las dos estirpes de aves durante las diferentes horas del día, bajo condiciones del Piedemonte Amazónico Colombiano.
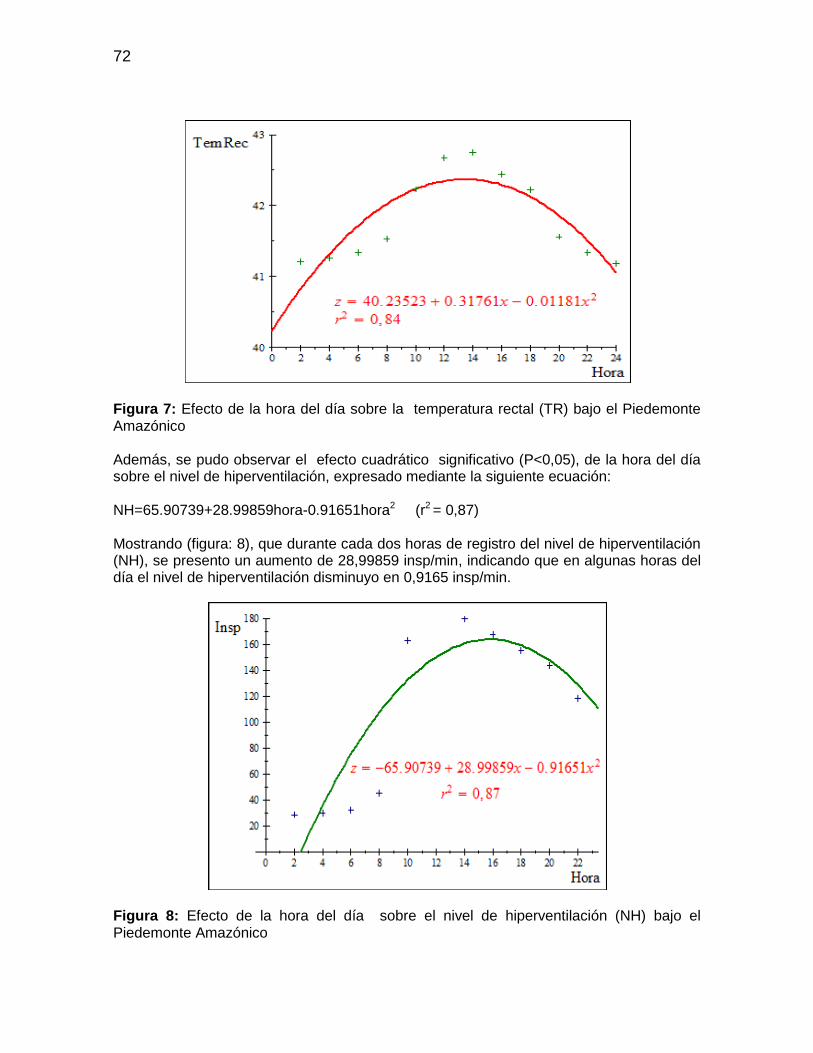
72
Figura 7: Efecto de la hora del día sobre la temperatura rectal (TR) bajo el Piedemonte Amazónico Además, se pudo observar el efecto cuadrático significativo (P<0,05), de la hora del día sobre el nivel de hiperventilación, expresado mediante la siguiente ecuación: NH=65.90739+28.99859hora-0.91651hora2 (r2 = 0,87) Mostrando (figura: 8), que durante cada dos horas de registro del nivel de hiperventilación (NH), se presento un aumento de 28,99859 insp/min, indicando que en algunas horas del día el nivel de hiperventilación disminuyo en 0,9165 insp/min.
Figura 8: Efecto de la hora del día sobre el nivel de hiperventilación (NH) bajo el Piedemonte Amazónico

73
Este resultado demuestra también la alta variabilidad del nivel de hiperventilación de las dos estirpes de aves durante las diferentes horas del día, bajo condiciones del Piedemonte Amazónico Colombiano. Los hallazgos anteriores permiten inferir que las variadas condiciones ambientales durante las diferentes horas del día, son los factores que más afectan al pollo de engorde, por comprometer el mantenimiento de la homeotermia con alteración en el equilibrio de las constantes fisiológicas. Tabla 19: Valores medios del peso del bazo (PB) y de la bolsa de Fabricio (PF) en pollos de engorde a los 42 días de edad bajo estrés calórico crónico (30ºC) y temperatura ambiental en el Piedemonte Amazónico Colombiano
TEMPERATURA ESTIRPE PB (g) PF(g) ESTRÉS
CALORICO CRONICO
COBB 3,64±0,21 b 3,69±0,19 b
ROSS 3,62±0,20 b 3,68±0,10 b
Media 3,63±0,21 B 3,68±0,15 B
PIEDEMONTE AMAZONICO
COBB
4,19±0,04 a 5,29±0,04 a
ROSS 4,20±0,04 a 5,31±0,02 a
C.V%
Media 4,20±0,04 A
3,19
5,30±0,03 A
2,0
Medias de tratamientos dentro de cada columna seguidas por letras minúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F. Medias de tratamientos dentro de cada columna seguidas por letras mayúsculas diferentes son estadísticamente diferentes entre sí (P<0,05) por la prueba de F.
Al observar los pesos del bazo (PB) y de la bolsa de Fabricio (PF), tabla 19, se puede evidenciar que no hubo diferencia significativa (P>0,05) entre las dos estirpes de aves bajo ninguna de las dos temperaturas experimentales; no obstante, el resultado hallado permite inferir un similar grado de afectación sobre el peso de estos dos órganos linfoides en los dos linajes de aves. Al comparar los pesos obtenidos bajo las dos temperaturas, con los pesos medios normales del bazo (4,19 g) y de la bolsa de Fabricio (5,38 g) en pollos de engorde a los 42 días de edad bajo termoneutralidad (Revidatti et al., 2002), se puede evidenciar una diferencia media inferior de 6,56% para el peso relativo del bazo y 16,5% para la bolsa de Fabricio. Este resultado se debe a que durante el estrés calórico los incrementos periódicos de corticosterona plasmática producen una atrofia temprana de la bolsa de Fabricio y el bazo con subsecuente efecto negativo sobre el peso de estos órganos (Kannan et al., 1996 y Post et al., 2003). La corticosterona actúa sobre el metabolismo in-termedio de proteínas, hidratos de carbono y lípidos, provocando su utilización como fuente de energía para la resistencia a la agresión, Bogin (1981); Arad (1983) y Angulo (1987).

74
En la tabla 19, se evidencia el efecto significativo (P<0,05) de la temperatura sobre la estirpe, notándose en las dos líneas de pollo sometidas a estrés calórico crónico el peso relativo más bajo de los dos órganos linfoides con una diferencia inferior de 13,6% para el bazo y 30,5% para la bolsa de Fabricio respecto a las aves bajo la temperatura del Piedemonte Amazónico Colombiano. Resultados semejantes han sido reportados por Rosales et al., (1989); Anwar et al., (2004) y Aengwanich, (2008), quienes encontraron disminución en el tamaño y peso de todos los órganos linfoides primarios (bazo, bolsa de Fabricio y timo) en pollos de engorde a los 42 días de edad después de ser sometidos a estrés calórico crónico. Los resultados obtenidos se relacionan con la mayor capacidad de reducción en el peso de los tejidos viscerales en condiciones de subnutrición post estrés calórico crónico, consecuentemente estos reducen su actividad metabólica más efectivamente, comparado con los tejidos de la carcasa (Lawrence y Fowler, 2002). El estrés calórico crónico afecta en mayor grado el desarrollo y maduración de la bolsa de Fabricio (Mashaly et al., 2004), disminuyendo la cantidad de parénquima, incrementando el índice de apoptosis (Guimarães et al., 2003; Anwar et al., 2004 y Aengwanich, 2008), de los linfocitos en la bolsa de Fabricio. Donker y Beuving, (1989), comprobaron que la infusión de corticosterona en pollos disminuyó el peso de la bolsa de Fabricio en un 57% y del bazo en un 35%; dichos cambios son consistentes con la idea de que los estresores y la corticosterona tienen efectos catabólicos sobre los órganos linfoides (Carsia y Harvey, 2000). Otras Investigaciones registran una disminución de hasta un 20% en el peso corporal y un 40% del peso de la bolsa de Fabricio en pollos inoculados con ACTH en forma continua durante 7 días (Puvadolpirod y Thaxton, 2000). El mayor detrimento en el peso de la bolsa (PF) y del bazo (PB) constituye en un buen indicador de los efectos deletéreos del estrés calórico crónico, más no del estrés calórico cíclico ocasionado por las temperaturas del Piedemonte Amazónico Colombiano; esta menor afectación sobre el peso de estos dos órganos linfoides, se puede explicar a través de las fluctuantes temperaturas durante el día y la noche, condición que mitigo parcialmente los efectos nocivos del estrés calórico, permitiendo periodos de mejor termorregulación en los pollos.
3.4 Conclusiones
El menor peso (g) hallado para el bazo y la bolsa de Fabricio en las aves sometidas a estrés calórico crónico, permite sugerir un marcado efecto inmuno depresor de este tipo de estrés.
El peso obtenido para el bazo y la bolsa de Fabricio bajo las temperaturas ambientales del Piedemonte Amazónico Colombiano, no constituye un buen indicador del estrés calórico cíclico en los pollos de engorde.
La alta variabilidad de las condiciones ambientales del Piedemonte Amazónico
Colombiano durante las diferentes horas diarias, son los factores que más afectaron las variaciones fisiológicas estudiadas en los dos linajes de aves, evidenciando alteraciones en la homotermia de las mismas.

75
Los pollos de engorde de las dos estirpes estudiadas mostraron una respuesta fisiológica análoga, demostrando baja eficiencia para eliminar calor a través del nivel de hiperventilación bajo las dos temperaturas experimentales.
Las condiciones ambientales de temperatura y humedad relativa nocturnas del Piedemonte Amazónico ofrecen periodos de termoneutralidad, los cuales pueden ser aprovechados para maniobras de alimentación y manejo.
El nivel de hiperventilación (NH) y la temperatura rectal (TR) se constituyen en indicadores fisiológicos fundamentales para medir los efectos del estrés calórico en los pollos de engorde, afianzándose como una herramienta para la investigación de esta problemática.

76
Bibliografía Aengwanich, W. Pathological changes and effect of ascorbic acid on lesion scores of bursa of fabricius in broilers under chronic heat stress. Res. J. Vet. Sci.1, p.62–66, 2008. Angulo-Chacon, I. Observaciones sobre nutrición en las aves en países de clima cálido. Sargento Cabral: Orient Avíc, v.12, p.23-24,1987. Anwar,B.;Khan, S.A.; Aslam, A.; Maqbool, A.; Khan, K.A. Effects of ascorbic acid and acetylsalicylic acid supplementation on the performance of broiler chicks exposed to heat stress. Pak. Vet. J. 24, p.109–111, 2004. Arad, Z.; Marder, W. Serum electrolyte and enzyme responses to heat stress and dehydration in the fowl (Gallus domesticus). Ph. D. Department of Zoology, The Hebrew University of Jerusalem.1983. 234p. Bogin, E.; Weisman, Y.; Friedman, Y. The effect of heat stress on the levels of certain blood constituents in chickens. Refuah Vet Israel, v. 38(1): p.98-104, 1981. Bohin, E.; Peh, H.C.; Avidar, Y.; Israelí, B. Sex and genotype dependence on effects of long-term high environmental temperatures on cellular enzyme activities from the chicken organs”. Avian Pathol 26: p.511-513, 1997. Bottje, W.G.; Harrison, P.C. Effects of carbonated water on growth performance of cockerels subjected to constant and cyclic heat stress temperature”. Poultry Science. 64: p.1285-1292, 1995. Butcher, G.D.; Miles, R.D. Causes and prevention of wet litter in broiler houses. Cooperative Extension Service, Institute of Food and Agric. Sci. Univ. Florida. v. 80, p.67-69, 2003. Cahaner, A.; Deeb, N.; Gutman, M. Effects of the plumage-reducing naked neck (Na) gene on the performance of fast-growing broilers at normal and high ambient temperatures. Poultry Science, v. 72(5): p.767-775, 1993. Carsia, R.V; Harvey, S. Adrenals. In: Sturkie’s Avian Physilogy, San Diego: Academic Press, 2000. 232p. Cella, P.S.; Donzele, J.L.; Oliveira, R.F.M. et al. Níveis de lisina mantendo a relação aminoacídica para pintos de corte, em diferentes ambientes térmicos. In: Reunião Anual da Sociedade Brasileira de Zootecnia: Viçosa, 2000.Anais 1: p.305. Chepete, H.J. Rectal temperatures changes in broilers kept under hot and dry conditions. Agr. Engin. Int.: The CIGR Ejournal 10: p.1-10, 2008. Colina, Y.C. Evaluación del proceso de hiperventilación como indicador del estrés térmico de pollos de engorde en condiciones de laboratorio y comerciales. Tesis de Maestría.

77
UCV. Postgrado en Producción Animal. Facultad de Agronomía. Maracay Edo. Aragua Venezuela, 2007. Conhalato, G.S. Exigência de lisina digestível para frangos de corte machos. Viçosa, MG: Universidade Federal de Viçosa. Dissertação Mestrado em Zootecnia. Universidade Federal de Viçosa, Brasil, 1998. Cunningham, J.G. Fisiología Veterinaria. México D.F, México: Interamericana, 1995. 202p. De Basilio,V.; F. Requena; A. León; M. Vilariño y M. Picard. Early age thermalconditioning immediately reduces body temperature of broiler chicks in a tropicalenvironment. Poult. Sci., 82: p.1235-1241, 2003. Dohms, J.; Saif, M. Criteria for evaluating immunosupression. Avian Dis. 28: p.305–310, 1984. Donker, R.A.; Beuving, G. Effect of corticosterone infusion on plasma corticosterone concentration, antibody production, circulating leukocytes and growth in chicken lines selected for humoral immune responsiveness. Br. Poult. Sci., Oxforshire, v. 30(3): p.361-369, 1989. El Hadi, H.; Sykes, A. Thermal panting and respiratory alkalosis in the laying hens. Br.Poult. Sci. 23: p.49-57, 1982. Espinet, R.G. Los “stress”– Su naturaleza – Sus reacciones. Vet. Arg. 4: p.882–888,1987. Estrada, M.; Márquez, S. Interacción de los factores ambientales con las respuestas del comportamiento productivo en pollos de engorde. Revista Colombiana de Ciencias Pecuarias.18: p.205-252, 2005. Farfán, C.H. Efecto de la adición de minerales en agua o en alimento sobre parámetros productivos y fisiológicos en pollos de engorde en finalización bajo estrés calórico. Maracay. Edo. Aragua. Venezuela, 2008.75p.Tesis (Facultad de Agronomía). Freeman, B.M. A stress hormone in the domestic fowl. Res. Vet. Sci. 28: p.389-390, 1980. Fussell, L.; Rossi, A.Y.; Wilson, M. Lighting programs and cob 500 broiler performance. South África: Technical focus, 2003.107p. Gainesville, G.; Gingerich E.N.; Donald, J. Influencia del calor en aves de carne y huevos. Control y manejo”. Revista avícola de Honduras, v.67, n7, 2003. Disponible en: <http://ww.visionveterinaria.com/prion/avescalor.html>. Accesado en: 23/04/2010. Geraert, P.A.; Gulillaumin, S.; Padilha J.C. Metabolic and endocrine changes induced by chronic heat exposure in broiler chickens: growth performance, body composition and energy retention”. Br J Nutr. 75: p.195-204, 1996.

78
Guimaraes, E.B.; Vasconcelos, A.C.; Martins, N.R.S.; Oliveira, F.M.; Moro, L.; Nunes, J.E.S.; Santos, F.G.A. Porcentagem de parênquima e índice apoptótico da bolsa cloacal em frangos de corte em ambiente de conforto e estresse térmico”. Arq. Bras. Med. Vet. Zootec., Belo Horizonte, v. 55(2): p.178-186, 2003. Havenstein, G.B.; Ferket, P.R.; Schiedeler, S.E.; Larson, B.T. Growth, livability and feed conversion of 1991 vs 1957 broilers when fed typical 1957 and 1991 broiler diets. Poultry Science 73: p.1785-1794, 1994. Havenstein, G.B., Ferket, P.R.; Qureshi, M.A. Growth, livability, and feed conversion of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets. Poultry Science 82: p.1500-1508, 2003a. Hillman, P.E. et al. Physiological responses and adaptations to hot and cold environments. In: YOUSEF, M.K. Stress physiology in livestock. Boca Raton: CRC. 3: p.1- 71, 1985. Ismail, N.M.; Saif, Y.M.; Wigle, G.B.; Havenstein, G.B.; Jackson, C. Infectious Bursal Disease virus variant from commercial Leghorn pullets. Avian Dis. 34: p.141-145, 1990. Janke, O.; Tzschentke, B.; Boerjan, B. Comparative investigations of heat production and body temperature in modern chicken breeds. Avian and Poultry Biological Reviews 15: p.191-196, 2004. Kannan, G.; Mench, J.A. Influence of different handling methods and crating periods on plasma corticosterone concentrations in broilers. British Poultry Science. 37(1): p.21-31, 1996. Lawrence, T.L.J.; Fowler, V.R. Growth of farm animals. Aberdeen: CAB International, 2002. 254p. Macari, M.; Furlan, R.L.; Maiorka, A. Aspectos fisiológicos e de manejo para manutenção da homeostase térmica e controle de síndromes metabólicas. In: Melitzer, A. Efficiency of effect of high ambient temperature on food utilization in male broilers. Brit. Poult. Sci. v. 27: p.349-351, 1986. Mashaly, M.M.; Hendricks, G.L.; Kalama, M.A.; Gehad, A.E.; Abbas, A.O.; Patterson, P.H. Effect of heat stress on production parameters and immune responses of commercial laying hens. Poult. Sci. 83, p.889–894, 2004. Mendes, A. A.; Naas, I.A.; Macari, M. Produção de frangos de corte. Campinas: Facta; 2004. p.137-156. May, J.D.; Lott, B.D. Relating weight gain and feed: gain of male and female broilers to rearing temperature”. Poult Sci. 80: p.581-584, 2001. Marder, J.; Arad, Z. Panting and acid base regulations in heat stressed birds. Comparative Biochemistry and Physiology, 1989. 395-400p.

79
Mather, F.; G. Barnas, Y.R. ; Burger, T. The Influencia of Alkalosis on Panting. Comp. Biochem Physiol. 67(A): p.265-268, 1980. National Research Council. Nutrient Requirements of Poultry. Washington: National Academy of Sciences, 1994. Oliveira Neto, A. R.; Oliveira, R.F.M.; Donzele, J.L. Metabolizable energy level for broilers from 22 to 42 days of age maintained under thermoneutral environment. Rev. Bras. Zootec., Viçosa-MG. v. 29(4): p.1132-1140, 2000. Ophir, E.; Arieli, Y.; Marder, J.; Horowitz, M. Cutaneous blood flow in the pigeon Columba livia: its possible relevance to cutaneous water evaporation. Journal of Experimental Biology 205: p.2627-2636, 2002. Post, J.; Rebel, M.J.; Terhuurne, A.A. Physiological effect of elevated plasma corticosterone concentration in broiler chickens. An alternative means by which to assess the physiological effect of stress. Poult. Sci., 82: p.1313-1318, 2003. Puvadolpirod, S.; Thaxton, J.P. Model of Physiological Stress in Chickens 1. Response Parameters. Poult. Sci., Champaign. v. 79(4): p.363-369, 2000. Quiles, A.; Hevia, M.L. Fisiologismo de la termorregulación en las gallinas. Departamento de producción animal. Facultad de Veterinaria. Universidad de Murcia, Argos On line Portal de Veterinaria. España. 2003. Disponible en: <http://www.portalveterinaria.com/modules.php?name=Articles&file=print&sid=182> Accesado en: 21/09/2010. Revidatti, F.A.; Fernandez, R.J.; Terraes, J.C.; Sandoval, G.L.; Luchi, P.E. Modificaciones del peso corporal y indicadores de estrés en pollos parrilleros sometidos a inmovilización y volteo. Rev. Vet. Arg., Corrientes. v. 12(1): p.1-8, 2002. Rosales, A.G.; Villegas, P.; Lukert, P.D.; Mohamed, A.M.; Brown, J. Isolation. Identification and pathogenicity of two field strains of infectious Bursal Virus. Avian Dis., Athens. v. 33(1): p.35-41,1989. Rostagno, H.S.; editor. Tabelas brasileiras para aves e suínos: composição de alimentos e exigências nutricionais.2.ed, Viçosa, Brasil: UFV Departamento de Zootecnia, 2005. Sainsbury, D. Poultry health and management. 4. ed. Blackwell Science, 2000. 232p. Sandoval, et al., Relación heterófilo/linfocito e índice morfométrico bursal como indicadores de estrés crónico en pollos parrilleros. Unne.. Argentina. 2002. Disponible en: <http://www1.unne.edu.ar/cyt/2002/04-Veterinarias/V-044.pdf> Accesado en: 20/09/2010. Siegel, B.P. Poultry stress, immunity interactions are analysed. Poultry Dig. 35: p.38–42, 1990. Simmons, J.D.; Lott, B.D.; May, J.D. Heat loss from broiler chickens subjected to various wind speeds and ambient temperatures. Appl Eng Agric. 13: p.665-669, 1987.

80
Simmons, J.D.; Lott, B.D.; Miles, D.M. The effects of high air velocity on broiler performance. Poult Sci. 82: p.232-234, 2003. Sturkie, P. Body Fluids: Blood. In. Sturkie, P ed. Avian Physiology. New York: Springer-Verlag, 1986. p.136-139. Teeter, R.G.; Smith, M.O.; Wiernusz, C.J. Effect of broiler acclimation to heat stress and feed intake on body temperature in birds exposed to thermoneutral and high ambient temperatures. Poult. Sci., 71: 1101-1104, 1992. Thaxton, P.; Siegel, H.S. Immunodepression in young chickens by high environmental temperature”. Poultry Sci. 49: p.202-205, 1982. Veldkamp, T.; Kwakkel, R.P.; Ferket, P.R; Verstegent, M.W. Growth responses to dietary energy and lysine at high and low ambient temperature in male turkeys. Poult Sci. 84: p.273-282, 2005. Vilariño, M.; León, A.; Picard, M. Comportamiento alimenticio de las aves. Conceptos básicos, metodologías y resultados”. V Encuentro sobre Nutrición y Producción de Animales Monogástricos. Production de Aves; 1999. 1: p.37-45. Wang, S.; Bottje, W.G.; Kinzler, S.; Neldon, H.L.; Koike, T.I. Effect of heat stress on plasma levels of arginine vasotocin and mesotocin in domestic fowl. (Gallus domesticus). Comparative Biochemistry and Physiology. 93A(4): p.721–724, 1989. Webster, M.D.; King, J.R. Temperature and humidity dynamics of cutaneous and respiratory evaporation in pigeons, Columba livia. Journal of Comparative Physiology 157: p.253-260, 1987. Wolfenson, D.; Bachrach, D.; Maman, M.; Graber, Y.; Rozenboim, I. Evaporative cooling of ventral regions of the skin in heat-stressed laying hens. Poultry Science 80: p.958-964, 2001. Yahav, S.; D.Lugar, A.; Cahaner, M.; Dotan, M.; Ruzanand.; Hurwitz, S. Thermoregulation in naked neck chickens subjected to different ambient temperatures. Br. Poult. Sci. 39: p.133-138, 1998. Yahav, S. Relative humidity at moderate ambient temperatures: its effect on male broiler chickens and turkeys”. Br. Poultry Sci., 41: p.94-100, 2000. Yahav, S.; Straschnow, A.; Vax, E.; Raspakovski, V.; Shinder, D. Air velocity alters broiler performance under harsh environmental conditios. Poult Sci. 80: p.724-726, 2001. Zhou, W.Y.; Yamamoto, S. Effects of environmental temperature and heat production due to food intake on abdominal temperature, shank skin temperature and respiration rate of broilers. British Poultry Science. 38: p.107-114, 1997.