efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos de … · 2014-03-21 ·...
TRANSCRIPT

María Angeles del Castillo Alonso
Encarnación Núñez Olivera y Javier Martínez Abaigar
Facultad de Ciencias, Estudios Agroalimentarios e Informática
Programa de doctorado Ecosistemas agrícolas sostenibles (formación)
2012-2013
Título
Director/es
Facultad
Titulación
Departamento
TRABAJO FIN DE ESTUDIOS
Curso Académico
Efectos de la exclusión de radiación ultravioleta sobrehojas y hollejos de Vitis vinifera cv. Graciano
Autor/es

© El autor© Universidad de La Rioja, Servicio de Publicaciones, 2014
publicaciones.unirioja.esE-mail: [email protected]
Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos de Vitisvinifera cv. Graciano, trabajo fin de estudios
de María Angeles del Castillo Alonso, dirigido por Encarnación Núñez Olivera y Javier Martínez Abaigar (publicado por la Universidad de La Rioja), se difunde bajo una Licencia
Creative Commons Reconocimiento-NoComercial-SinObraDerivada 3.0 Unported. Permisos que vayan más allá de lo cubierto por esta licencia pueden solicitarse a los
titulares del copyright.


FACULTAD DE CIENCIAS, ESTUDIOS AGROALIMENTARIOS E INFORMÁTICA
Efectos de la exclusión de la radiación ultravioleta sobre hojas y hollejos de Vitis vinifera cv. Graciano
Trabajo presentado por:
MARIA ÁNGELES DEL CASTILLO ALONSO
para conseguir los 30 créditos correspondientes a la elaboración del Trabajo de Investigación (Programa de Doctorado Ecosistemas Agrícolas Sostenibles).
Logroño, a 7 de Junio de 2013
Fdo. Maria Ángeles del Castillo Alonso

FACULTAD DE CIENCIAS, ESTUDIOS AGROALIMENTARIOS E INFORMÁTICA
Los Doctores Encarnación Núñez Olivera y Javier Martínez Abaigar, Catedráticos de Fisiología Vegetal y Botánica del Departamento de Agricultura y Alimentación de la Universidad de La Rioja.
Certifican:
Que el Trabajo de Investigación titulado “Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos de Vitis vinifera cv. Graciano” ha sido realizado por la alumna Mª Ángeles del Castillo Alonso bajo su inmediata dirección y reúne las condiciones exigidas para conseguir los 30 créditos correspondientes al mismo.
Logroño, a 7 de Junio de 2013
Fdo. Encarnación Núñez Olivera Fdo. Javier Martínez Abaigar

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Índice
Índice Resumen..............................................................................................................................................1 Introducción ........................................................................................................................................2
La radiación UV ................................................................................................................................2 Efectos de la RUV sobre los organismos fotosintéticos ................................................................3 Vitis vinifera L. cv. Graciano ............................................................................................................4 Efectos de la RUV sobre la vid ........................................................................................................5
Objetivos .......................................................................................................................................... 10 Materiales y métodos ..................................................................................................................... 11
Material vegetal y sitio de estudio ................................................................................................ 11 Diseño experimental ..................................................................................................................... 11 Variables fisiológicas medidas ..................................................................................................... 14
Fluorescencia de clorofilas ....................................................................................................... 14 Índice de Esclerofilia (IE) .......................................................................................................... 15 Pigmentos fotosintéticos ........................................................................................................... 15 Compuestos absorbentes de radiación ultravioleta (CARUV)................................................ 16
Análisis estadístico ....................................................................................................................... 18 Resultados ....................................................................................................................................... 19
Comparaciones metodológicas .................................................................................................... 19 Variaciones en función del tiempo y del tratamiento (régimen de radiación) ............................ 20
Discusión.......................................................................................................................................... 32 Comparaciones metodológicas .................................................................................................... 32 Composición fenólica de la variedad Graciano ........................................................................... 33 Variaciones en función del tiempo ............................................................................................... 34 Variaciones en función del tratamiento (régimen de radiación) ................................................. 35
Conclusiones ................................................................................................................................... 40 Agradecimientos ............................................................................................................................. 41 Referencias bibliográficas ............................................................................................................. 42

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resumen
1
Resumen Se ha realizado, en condiciones de campo, el primer estudio acerca de los efectos de la exclusión de radiación ultravioleta (RUV) en Vitis vinifera cv. Graciano, una de las variedades autorizadas para el cultivo por la D.O.Ca. Rioja. Para ello se establecieron tres regímenes de radiación: Ambiente (plantas expuestas a niveles naturales de radiación solar), Filtro UV+ (plantas cubiertas por un filtro que dejaba pasar radiación fotosintéticamente activa (PAR), UV-A y UV-B), y Filtro UV- (plantas cubiertas por un filtro que únicamente dejaba pasar PAR). Los filtros se colocaron en el envero y se mantuvieron hasta la vendimia. Los efectos de la exclusión de RUV se analizaron en hojas y hollejos. En las hojas se midieron la fluorescencia de clorofilas, el Índice de Esclerofilia (IE), la composición pigmentaria fotosintética y los compuestos absorbentes de RUV (CARUV), mientras que en los hollejos se analizaron los CARUV. Tanto en hojas como en hollejos, los CARUV se midieron de manera global e individual, y en ambos casos se distinguió, por primera vez en la vid, su localización en las vacuolas (fracción soluble) y en las paredes celulares (fracción insoluble). Se ha comprobado que algunas variables de gran importancia fisiológica, como el rendimiento cuántico máximo del fotosistema II (Fv / Fm) y las concentraciones de clorofila y CARUV, se pueden medir de manera fiable, rápida y repetitiva mediante técnicas no destructivas, lo que constituye una sustancial simplificación metodológica aplicable a futuros experimentos. Los cambios temporales que mostraron las plantas desde envero a vendimia fueron más intensos que los debidos a la exclusión de RUV. En las hojas, estos cambios se relacionaban con la senescencia (descensos de Fv / Fm y pigmentos fotosintéticos, y aumentos de IE), aunque también con la RUV y la PAR (aumentos de CARUV, tanto solubles como insolubles, medidos globalmente e individualmente). Las respuestas de las hojas a la exclusión de la RUV solar fueron ligeras, destacando los descensos de un ácido hidroxicinámico y un flavonol presentes en la fracción soluble. En los hollejos, al eliminar la RUV solar descendieron significativamente los CARUV solubles medidos globalmente y las concentraciones de 13 de los 25 CARUV individuales analizados, todos ellos presentes en la fracción soluble. Por tanto, la fracción soluble es más reactiva a la RUV que la insoluble. Los CARUV más reactivos fueron los flavonoles (principalmente la miricetina-3-O-glucósido y las quercetinas glucosiladas), mientras que los flavanoles no respondieron al tratamiento. En conclusión, la débil respuesta fisiológica de las hojas de la variedad Graciano a la exclusión de RUV solar sugiere que esta variedad se encuentra bien adaptada a los niveles actuales de RUV. En esta adaptación podrían ser importantes los CARUV que respondieron a la exclusión de RUV, presentes en la fracción vacuolar de la célula. Además, la presencia de RUV es un factor crucial para la acumulación de diversos compuestos del hollejo importantes en la elaboración del vino, como los flavonoles. Todo esto realza la importancia potencial de la manipulación de la RUV en la vitivinicultura.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Introducción
2
Introducción
La radiación UV La radiación ultravioleta (RUV) es un componente minoritario (aproximadamente un 6%) de la radiación solar, en comparación con las fracciones dominantes: la visible/fotosintética y la infrarroja. Sin embargo, la RUV es un factor ambiental que ha provocado diversos cambios adaptativos en los organismos en el proceso de desarrollo de la vida sobre la Tierra (Cockell y Knowland, 1999). La RUV induce ciertos procesos en todos los seres vivos incluida la especie humana, y muchos de ellos son lesivos, aunque pueden ser contrarrestados hasta cierto punto por mecanismos de reparación y aclimatación. Entre las tres bandas en las que se divide la RUV según el criterio de la CIE (Commission Internationale d’Eclairage), la más dañina es la UV-C (<280 nm), pero no resulta importante en la época presente porque es completamente absorbida por el oxígeno y el ozono de la estratosfera. Por su parte, las bandas UV-B (280-315 nm) y UV-A (315-400 nm) penetran en la biosfera y provocan efectos biológicos significativos. Estos efectos dependen en gran medida de la longitud de onda asociada, y por ello se han desarrollado diferentes funciones de ponderación para calcular la RUV efectiva biológicamente (RUVBE). La RUVBE agrupa tanto la UV-A como la UV-B, pero está dominada por esta última porque la efectividad biológica aumenta logarítmicamente al disminuir la longitud de onda. Por tanto, la mayoría de los estudios sobre los efectos de la RUV han tenido en cuenta la UV-B. Este hecho se ha intensificado desde el descubrimiento de la degradación antropogénica del ozono estratosférico, porque sólo la UV-B (y no la UV-A) es absorbida por este gas, y por consiguiente la degradación de ozono causa un aumento en la UV-B que llega a la superficie terrestre. No obstante, la tendencia actual es considerar también la UV-A en el diseño de funciones de ponderación biológica (Flint et al., 2003). La medida de la RUV como variable meteorológica resulta delicada, y por ejemplo hay discrepancia entre las medidas al nivel del suelo y las estimaciones realizadas por satélite (Seckmeyer et al., 2008). La irradiancia UV al nivel del suelo depende de numerosos factores, como la latitud, la estación del año, la hora del día, la altitud, la presencia de nubes o aerosoles, el albedo y los niveles de ozono (McKenzie et al., 2007). La pérdida de ozono como consecuencia de las emisiones antropogénicas de halocarburos y otros gases destructores ha sido más intensa en la Antártida, aunque también se han observado reducciones en latitudes árticas y medias. En latitudes medias, se estima que la radiación UV-B solar ha aumentado entre un 6% y un 12% desde 1980 (McKenzie et al., 2003), aunque este aumento puede resultar enmascarado por grandes cambios estacionales y por las diferencias geográficas (Häder et al., 2007). Se espera que los niveles altos de RUV continúen varias décadas por una insuficiente recuperación del ozono estratosférico, a pesar de los beneficios derivados del Protocolo de Montreal (1987). Por lo tanto, los estudios acerca de los efectos de la RUV (tanto de sus niveles actuales como de su aumento) sobre los organismos y los ecosistemas, están revestidos de una gran importancia, teniendo también en cuenta que pueden existir interacciones entre la degradación de ozono y otros factores de cambio climático, como las variaciones en la nubosidad, la concentración de aerosoles y el albedo (McKenzie et al., 2007).

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Introducción
3
Efectos de la RUV sobre los organismos fotosintéticos En los organismos fotosintéticos, el aumento de RUV puede provocar daños en el aparato fotosintético: degradación de pigmentos, fotoinhibición, y descensos en el rendimiento cuántico, la tasa de fotosíntesis y la actividad de enzimas del ciclo de Calvin (Jansen et al., 1998). Así mismo, pueden ocurrir alteraciones en el ADN, daños oxidativos y cambios en la absorción mineral (Day y Neale, 2002; Prado et al., 2012). Todo ello puede conducir a alteraciones generales del crecimiento y el desarrollo. Sin embargo, persiste todavía cierta controversia sobre la relevancia ecológica de estos efectos, ya que muchos de los estudios iniciales acerca de los efectos de la RUV se realizaron en condiciones de laboratorio utilizando dosis excesivas y poco realistas de RUV, así como proporciones UV/PAR (PAR son las siglas inglesas de Photosynthetically Active Radiation o radiación activa para la fotosíntesis, que se extiende entre 400 y 700 nm) alejadas de las naturales. Por lo tanto, resulta inapropiada la extrapolación de estos resultados a las condiciones de campo (Searles et al., 2001; Day y Neale, 2002). En el nivel ecosistémico, la RUV puede afectar a la descomposición de los restos vegetales, los ciclos biogeoquímicos, las interacciones tróficas y el equilibrio competitivo entre las especies (Caldwell et al., 2007). Los organismos fotosintéticos desarrollan varios mecanismos de protección, reparación y aclimatación frente a los efectos adversos de la RUV (Jansen et al., 1998): acumulación de compuestos absorbentes de RUV (CARUV) (flavonoides, fenil-propanoides, aminoácidos micosporínicos, etc.), procesos antioxidantes y fotoprotectores, y reparación o recambio de las biomoléculas dañadas, como el ADN o las proteínas. A nivel metabólico, la RUV afecta a la cantidad de clorofilas y por tanto a la capacidad fotosintética de la planta, ya que dicha radiación puede ocasionar daños en los pigmentos fotosintéticos y en la estructura de los cloroplastos (Teramura y Sullivan, 1994; Kakani et al., 2003), así como la inhibición del fotosistema II (PSII). Adicionalmente, la fotosíntesis puede verse afectada de forma indirecta por la reducción de la conductancia estomática (Day y Vogelmann, 1995), que limita la capacidad de intercambio gaseoso de la planta. El aumento de RUV-B en la Biosfera, provocado por la degradación antropogénica de ozono estratosférico, ha motivado que la investigación sobre los efectos de la RUV se haya enfocado en las últimas décadas especialmente al estudio de sus efectos dañinos, tanto en el ser humano como en las plantas. Sin embargo, en los últimos años, los estudios realizados han mostrado que en condiciones naturales, las plantas están adaptadas a los niveles actuales de RUV (Núñez Olivera et al., 2006; Prado et al., 2012) y no les supone estrés, a no ser que estén sometidas simultáneamente a otras condiciones ambientales adversas. Un descubrimiento relativamente nuevo es que las plantas son capaces de detectar RUV, gracias a la presencia de un fotorreceptor (UVR8), y responder a ella (Rizzini et al., 2011). Este fotorreceptor controla la expresión de numerosos genes implicados en la aclimatación y protección de las plantas a la radiación UV-B. Junto a la regulación de genes que codifican para las fotoliasas (enzimas necesarias para la reparación de los daños en ADN) y otros relacionados con los mecanismos de activación de sistemas antioxidantes, UVR8 regula la expresión de los genes implicados en la biosíntesis de los derivados fenólicos. Estos descubrimientos han modificado la percepción que tiene la comunidad científica sobre la RUV.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Introducción
4
Vitis vinifera L. cv. Graciano Graciano es una variedad cuyo origen se sitúa en La Rioja. La superficie empleada en su cultivo en España ronda las 1800 Ha y se encuentra presente en 8 Comunidades Autónomas, entre las que destaca La Rioja con casi un 40% del total de la superficie dedicada a este cultivo. Se encuentra autorizada por 7 Denominaciones de Origen (D.O.), entre ellas Rioja, lo que le otorga cierta relevancia dentro de las variedades minoritarias en dicha D.O. (Cabello Saénz de Santamaría et al., 2012). Graciano es una variedad de ciclo largo y brotación tardía, muy interesante en climas cálidos, en los que mantiene altos niveles de acidez. Es bastante sensible a quemaduras foliares, y requiere muy buenas condiciones climáticas para completar su maduración. Tanto su fertilidad como su producción son medias-bajas. Esta variedad es poco sensible a plagas y enfermedades en general, aunque manifiesta una sensibilidad muy alta al mildiu y media a botritis. La planta es de porte erecto, sin apenas nietos, lo que facilita su manejo. Sus hojas son trilobuladas, de forma pentagonal, con los senos laterales inferiores ausentes y los superiores muy marcados (Figura 1). El haz es verde oscuro y el envés algodonoso. Tiene racimos cilíndricos o cónicos cortos, compactos y de tamaño mediano, formados por bayas azul-negras, esféricas y medianas, con hollejo grueso y bastante contenido en pruina. La uva tiene abundantes pectinas, elevado contenido en azúcares, bajo contenido en taninos y contenido medio en compuestos fenólicos no siendo muy característica su concentración de antocianos (Suberviola Ripa, 1998). La pulpa no tiene pigmentación, es de baja consistencia y muy jugosa. Posee unas buenas aptitudes enológicas para mejorar a otras variedades como Tempranillo, gracias a su acidez elevada, color rojo vivo y aromas a regaliz. Se emplea tanto para la elaboración de vinos de crianza como reserva y gran reserva (Cabello Saénz de Santamaría et al., 2012).
Figura 1. Hoja y racimo de Vitis vinifera L. cv. Graciano.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Introducción
5
Efectos de la RUV sobre la vid En los organismos fotosintéticos, los efectos de la RUV se han estudiado principalmente en plantas terrestres, especialmente Arabidopsis thaliana (la planta modelo) y especies cultivadas. Sin embargo, son relativamente escasos los estudios realizados en vid (Tabla 1), y sobre todo en variedades cultivadas en La Rioja a nivel local como Graciano, objeto de este estudio (Guerrero et al., 2010). La respuesta de la vid a la RUV depende fundamentalmente de la variedad, de las condiciones ambientales (temperatura del aire y humedad relativa), y de la dosis de RUV incidente y la relación entre ésta y la PAR (Berli et al., 2013), ya que las ratios UV-B/PAR más altas de las que se pueden medir en condiciones naturales, provocan efectos exagerados sobre las plantas. Por tanto, dicha respuesta es variable y es función del órgano estudiado y del estado de desarrollo de éste (Berli et al., 2013): se puede producir una reducción del crecimiento de los tallos, el área y peso de las hojas, y el volumen y peso de las bayas. Las variedades más estudiadas han sido Cabernet Sauvignon, Malbec, Chardonnay y Silvaner, aunque existen estudios de diversas variedades de varios lugares del mundo, incluida La Rioja. Los experimentos se han realizado tanto en condiciones de campo, más realistas, como en invernadero y laboratorio. La manipulación de la RUV se ha realizado mediante suplementos con lámparas y exclusión con filtros, y también se han hecho algunos experimentos con niveles naturales de RUV. La duración de los experimentos ha sido muy variable, desde unos días hasta todo el periodo de desarrollo de la planta, e incluso hay estudios plurianuales (Tabla 1). Diversos estudios se han ocupado de la influencia de la RUV sobre la fotosíntesis y la fluorescencia de clorofilas. Una fuerte disminución tanto en la captación de CO2 como en la conductancia estomática en las hojas de Chardonnay, no provoca una disminución apreciable en Fv / Fm (Majer y Hideg, 2012). Este factor puede verse influenciado por la variedad (Pollastrini et al., 2011), pero de forma general la vid está adaptada a las dosis naturales de RUV, gracias a la protección proporcionada por el ciclo de las xantofilas, los mecanismos antioxidantes y los CARUV (Núñez-Olivera et al., 2006; Doupis et al., 2011). En contraposición, cuando la vid se somete a suplemento de RUV, Fv / Fm sí se reduce drásticamente por la inhibición del PSII (Pfündel, 2003). La protección epidérmica que poseen las plantas del cultivar Silvaner es suficiente para que una exposición corta a RUV no provoque una reducción significativa de Fv / Fm con respecto a las plantas que no han sufrido estrés de ningún tipo (Kolb et al., 2001; Kolb y Pfündel, 2005). Los efectos de la RUV-B, en condiciones naturales de radiación, son tan pequeños y transitorios que indican que la inhibición de la fotosíntesis no está relacionada con la inhibición del PSII provocado directamente por la RUV (Berli et al., 2013). Se ha demostrado que la RUV produce cambios en la morfología de las hojas, aumentando su grosor por una mayor acumulación de células epidérmicas, lo que incrementa la distancia que tiene que recorrer la RUV hasta los tejidos internos más sensibles a esta radiación (Berli et al., 2013). Aquellas plantas sometidas a una mayor RUV serán más esclerófilas (Pollastrini et al.,

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Introducción
6
2011). Las diferencias en el Índice de Esclerofilia estarán íntimamente relacionadas con la duración de la exposición a la radiación. Si ésta no es lo suficientemente prolongada, este mecanismo de defensa y/o adaptación no se manifiesta de forma apreciable (Núñez-Olivera et al., 2006). Por otro lado hay resultados contradictorios con respecto a la evolución de los pigmentos fotosintéticos. El contenido de clorofilas está muy influenciado por la variedad y por el estado de desarrollo de la vid (Berli et al., 2013; Majer y Hideg, 2012). Así la RUV no afecta al contenido total de clorofilas en Cabernet Sauvignon, Malbec y Chardonnay (Keller et al., 2003; Berli et al., 2010; Majer y Hideg, 2012), pero con la exposición a RUV sí disminuye el contenido total del clorofilas en Tempranillo (Núñez-Olivera et al., 2006) o el contenido de clorofila b en Malbec (Berli et al., 2013). La RUV induce un aumento en la producción de pigmentos de protección, ya que juegan un papel decisivo en la disipación de la energía excedente (fotoprotección). En frutos con un bajo contenido de carotenoides, como las uvas, esta variable disminuye durante el desarrollo y la maduración. La concentración de carotenoides al inicio de la maduración es mayor en las bayas expuestas directamente a la radiación, mientras que en fechas cercanas a la vendimia se observa una reducción drástica por la degradación directa de estos pigmentos por la RUV (Schultz et al., 1998). Cerca de la vendimia, la reducción de radiación provoca una disminución en el contenido de carotenoides totales en hojas de Cabernet Sauvignon (Steel et al., 2000). Este efecto puede ser muy variable entre variedades de vid, pudiendo ser más sensibles unas que otras en cuanto a la degradación de carotenoides por la RUV, (Núñez-Olivera et al., 2006). Entre los efectos de la RUV sobre la vid, son especialmente importantes desde un punto de vista aplicado, los relacionados con los derivados fenólicos. Estos compuestos son metabolitos secundarios que tienen varias funciones biológicas en las plantas, además son de vital importancia en las características organolépticas y nutricionales de los vinos. Estos compuestos incluyen los ácidos fenólicos (ácidos hidroxibenzoicos y ácidos hidroxicinámicos), estilbenos como el resveratrol, y flavonoides (antocianos, flavanoles y flavonoles) (Berli et al., 2011). Los ácidos fenólicos están presentes fundamentalmente en las paredes celulares, y el resto en las vacuolas de la baya (hollejo, pulpa y semillas). Los flavonoles son los compuestos fenólicos más abundantes en las bayas de vid. Son pigmentos amarillos dentro de los cuales se incluyen la quercetina, el kaempferol y la miricetina, además de sus formas glucosiladas. Contribuyen a potenciar el color en los vinos tintos a través de la copigmentación con antocianos. Éstos últimos son pigmentos de color rojo sintetizados en el hollejo de las bayas al comienzo del envero. Los flavanoles, entre ellos la catequina, son los responsables de la astringencia y la estructura de los vinos. Dentro de los estilbenos, el más importante en la vid es el resveratrol, que juega un papel muy importante en los mecanismos de defensa contra el ataque de patógenos, y también está relacionado con la RUV. Es el compuesto más conocido debido a sus cualidades cardioprotectoras para el ser humano. La acumulación de compuestos absorbentes de RUV (CARUV), en su gran mayoría derivados fenólicos, es probablemente el principal mecanismo de adaptación de la vid a la RUV. La

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Introducción
7
biosíntesis de estos metabolitos secundarios es, sin duda, el tema más abordado en los trabajos realizados con RUV y vid (Tabla 1). Incluso, algunos estudios tratan de los efectos de la RUV-C, por lo tanto en condiciones no naturales (Bonomelli et al., 2004; González-Barrio et al., 2009; Guerrero et al., 2010; Pan et al., 2009; Petit et al., 2009; Wang et al., 2010). La biosíntesis de derivados fenólicos, además de contribuir a la formación del color, puede disminuir la penetración de la RUV a través de la epidermis (Kolb et al., 2001, 2003). En relación con los derivados fenólicos y la RUV, se ha demostrado que los polifenoles, especialmente antocianinas y flavonoles, se acumulan en hojas y bayas en respuesta a un aumento de RUV (Pontin et al., 2010; Berli et al., 2011). En cuanto a los compuestos individuales, en la última década se ha prestado especial interés a la síntesis de estilbenos en baya (Pan et al., 2009; Zhang et al., 2012; 2013), especialmente del resveratrol por aportar múltiples beneficios a los vinos. También existen trabajos sobre la respuesta de otros compuestos a la RUV, tanto en hoja (Berli et al., 2010, 2013) como en baya (Berli et al., 2008, 2011). Por ejemplo, las concentraciones de algunos flavonoides mayoritarios en vid, como catequinas, quercetinas, kaempferoles y miricetinas, están íntimamente relacionadas con la RUV, ya que disminuyen cuando se reduce esta fracción de radiación (Berli et al., 2008, 2011). Por otra parte, la estimulación de la síntesis de compuestos fenólicos se produce en función de la longitud de onda que incida sobre la planta. Así, la radiación visible (PAR) estimula la síntesis de ácidos hidroxicinámicos mientras que la RUV induce la de los flavonoides (Kolb et al., 2001; Koyama et al., 2012). La altitud del viñedo influye de forma muy notoria en la síntesis de CARUV. Las plantaciones situadas por encima de los 1000 m sobre el nivel del mar, donde los niveles de RUV-B son muy considerables, muestran concentraciones muy elevadas de compuestos fenólicos totales en baya (Berli et al., 2008, 2011, 2013).

Tabla 1. Artículos publicados sobre los efectos de la RUV en vid. VARIEDAD: NE, no especificada. ÓRGANO UTILIZADO: B, baya; F, flor; H, hoja. TIPO DE EXPERIMENTO: C, campo; I, invernadero; L, laboratorio. MANIPULACIÓN DE LA RUV (distinguiendo las distintas fracciones de UV: UV-A, UV-B, UV-C): E, exclusión; N, niveles naturales; S, suplemento. DURACIÓN DEL EXPERIMENTO: C, corta (< 15 días); M, media (16-60 días); L, larga (> 60 días). VARIABLES UTILIZADAS: C, crecimiento; E, esclerofilia; F, fotosíntesis; Fl, fluorescencia de clorofilas; Gn, respuestas genéticas; H, relaciones hídricas; Hr, hormonas; M, morfología; Mt1, metabolismo primario (glúcidos, proteínas, lípidos…); Mt2, metabolismo secundario, incluyendo compuestos absorbentes de RUV; Ox, variables de estrés oxidativo; P, pigmentos fotosintéticos; R, respiración; U, ultrastructura.
REFERENCIA VARIEDAD ÓRGANO UTILIZADO
TIPO DE EXPERIMENTO
MANIPULACIÓN DE LA RUV
DURACIÓN DEL
EXPERIMENTO
VARIABLES UTILIZADAS
Berli et al. (2008) Malbec B C E (UV-B) L Mt2
Berli et al. (2011) Malbec B C E (UV-B) L C, Mt1, Mt2 Berli et al. (2010) Malbec H C E (UV-B), S (UV-B) L Mt2, Ox, P Berli et al. (2013) Malbec H C E (UV-B) L C, F, Fl, Mt2,
Ox, P
Bonomelli et al. (2004) Chardonnay; Vitis rupestris cv. Rupestris du Lot H L S (UV-C) C Gn, Mt1, Mt2
Doupis et al. (2011) Soultanina H C S (UV-B) M C, H, Hr, Mt2, Ox,
González-Barrio et al. (2009) NE B L S (UV-C) M Mt2
Guerrero et al. (2010) Syrah, Merlot, Graciano, Tempranillo, Palomino fino, Palomino negro, Tintilla roja, Orion, Regent, V9, V15, V16
B L S (UV-C) C Mt2
Keller et al. (2003) Cabernet Sauvignon, Chardonnay H C E (UV) M F, Mt2, P, R, Kolb et al. (2001) Silvaner H C, I E (UV) C F, Fl, Mt2, U Kolb y Pfündel (2005) Silvaner H I E (UV) C Fl, Mt2
Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Introducción
8
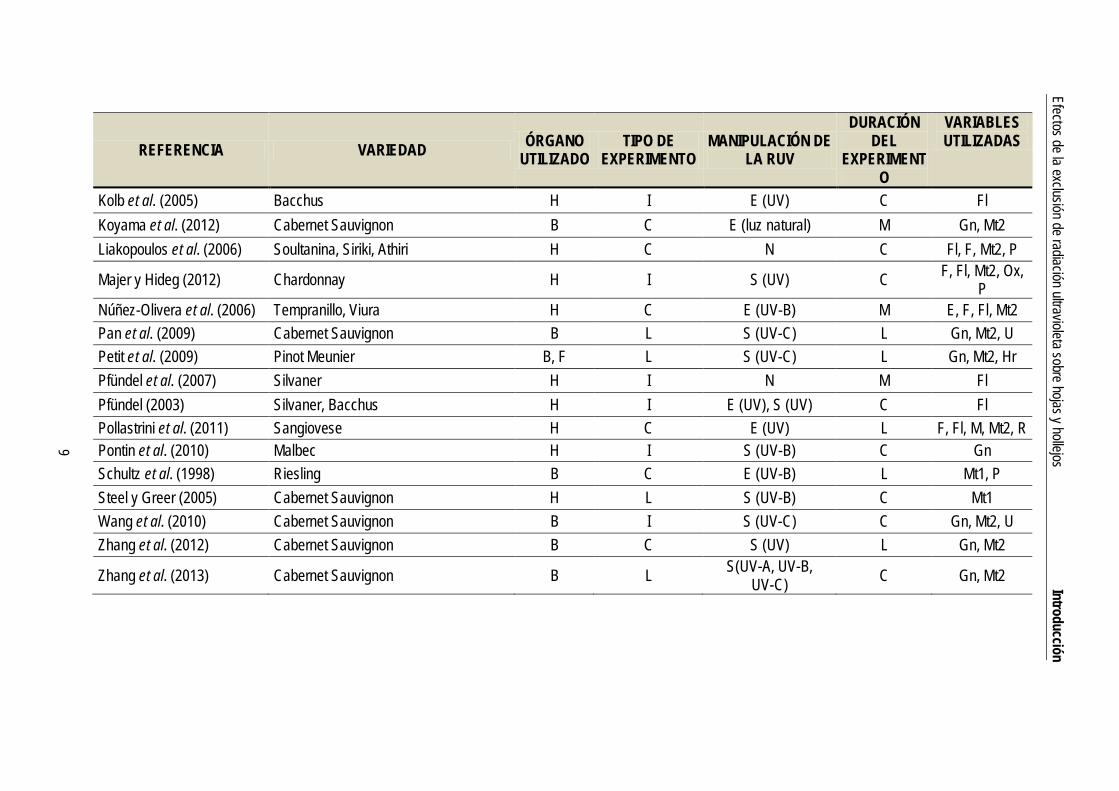
REFERENCIA VARIEDAD ÓRGANO UTILIZADO
TIPO DE EXPERIMENTO
MANIPULACIÓN DE LA RUV
DURACIÓN DEL
EXPERIMENTO
VARIABLES UTILIZADAS
Kolb et al. (2005) Bacchus H I E (UV) C Fl Koyama et al. (2012) Cabernet Sauvignon B C E (luz natural) M Gn, Mt2 Liakopoulos et al. (2006) Soultanina, Siriki, Athiri H C N C Fl, F, Mt2, P
Majer y Hideg (2012) Chardonnay H I S (UV) C F, Fl, Mt2, Ox, P
Núñez-Olivera et al. (2006) Tempranillo, Viura H C E (UV-B) M E, F, Fl, Mt2 Pan et al. (2009) Cabernet Sauvignon B L S (UV-C) L Gn, Mt2, U Petit et al. (2009) Pinot Meunier B, F L S (UV-C) L Gn, Mt2, Hr Pfündel et al. (2007) Silvaner H I N M Fl Pfündel (2003) Silvaner, Bacchus H I E (UV), S (UV) C Fl Pollastrini et al. (2011) Sangiovese H C E (UV) L F, Fl, M, Mt2, R Pontin et al. (2010) Malbec H I S (UV-B) C Gn Schultz et al. (1998) Riesling B C E (UV-B) L Mt1, P Steel y Greer (2005) Cabernet Sauvignon H L S (UV-B) C Mt1 Wang et al. (2010) Cabernet Sauvignon B I S (UV-C) C Gn, Mt2, U Zhang et al. (2012) Cabernet Sauvignon B C S (UV) L Gn, Mt2
Zhang et al. (2013) Cabernet Sauvignon B L S(UV-A, UV-B, UV-C) C Gn, Mt2
Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Introducción
9

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Objetivos
10
Objetivos Debido a la todavía escasa investigación sobre los efectos de la RUV en variedades de vid cultivadas en La Rioja, el objetivo de este trabajo era estudiar, en condiciones de campo, el efecto de la exclusión de RUV sobre la fisiología de las hojas y la composición fenólica de los hollejos en la variedad Graciano, que no se había estudiado anteriormente en este contexto. Este objetivo global se puede dividir en los siguientes objetivos parciales:
- Estudiar los efectos citados en un período considerable de tiempo (73 días desde el envero hasta la vendimia), superior al utilizado en los estudios previos realizados con variedades riojanas.
- Utilizar diferentes equipos para la medición no destructiva de ciertas variables fisiológicas, para comparar sus resultados con los obtenidos mediante técnicas destructivas.
- Analizar las respuestas de las hojas mediante una gran diversidad de variables
fisiológicas: fluorescencia de clorofilas, esclerofilia, composición pigmentaria fotosintética, niveles globales de compuestos absorbentes de RUV (CARUV) y concentraciones de compuestos individuales (ácidos fenólicos, estilbenos y flavonoides).
- Analizar las respuestas de los hollejos en cuanto a sus derivados fenólicos, por su
importante papel en la calidad de los vinos.
- Evaluar la acumulación de CARUV distinguiendo, por primera vez en vid, las fracciones vacuolar y ligada a las paredes celulares, tanto en hojas como en hollejos.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Materiales y métodos
11
Materiales y métodos
Material vegetal y sitio de estudio El estudio se realizó en 2011 en una parcela de viñedo de Vitis vinifera L. cv. Graciano, plantada en 1995 sobre suelo franco arcilloso, situada en la localidad de Ollauri (La Rioja) (lat. 42°31’N; long. 2°49’W; alt. 527 m). El marco de plantación del viñedo es de 2.70 m x 1.15 m y la orientación de las filas es de Este a Oeste. La viña se encuentra formada en espaldera VSP a doble cordón con 12 yemas por cepa. Este sistema cuenta con un alambre de formación situado a 0.70 m, uno de protección contra el viento a 1.00 m y un par de alambres móviles a 1.45 m. Todas las medidas están referidas sobre el suelo. Las plantas se deshojaron el 03/06/2011, en plena floración. Esta práctica consistió en la eliminación manual de 8 hojas adultas de cada pámpano, sin tocar los nietos. La parcela no dispone de riego, por lo que el régimen hídrico al que se sometieron las plantas correspondió a las precipitaciones naturales, registrándose un valor acumulado de 21.8 mm durante el periodo de estudio (02/08/2011 a 14/10/2011). En cuanto a la temperatura se obtuvieron valores de Tmed, Tmáx y Tmín de 17.8 ± 0.2 ºC, 36.2 ± 0.2 ºC y 3.3 ± 0.2 ºC, respectivamente, en el mismo intervalo de tiempo (estación agroclimática El Naval, alt. 510 m, Santos Salinas, Casalarreina, La Rioja, España).
Diseño experimental Se eligieron tres filas representativas y consecutivas dentro de la parcela. En cada fila, se establecieron los tres tratamientos utilizados, y cada uno de ellos se aplicó en tres cepas (Figura 2). Los tres tratamientos fueron los siguientes:
- AMBIENTE: tratamiento en el que la radiación recibida es la proveniente directamente del Sol.
- FILTRO UV+: tratamiento en el que la radiación recibida es filtrada por una plancha de 24 mm de espesor, de polimetacrilato de metilo de extrusión PMMA XT Vitroflex 295 Incoloro (Polimertecnic, Girona, España), que elimina la radiación de longitud de onda menor de 295 nm. Permite por tanto el paso tanto de la radiación fotosintéticamente activa (PAR) como de la radiación UV-A y de la UV-B.
- FILTRO UV-: tratamiento en el que la radiación recibida es filtrada por una plancha de 24 mm de espesor, de PMMA XT Vitroflex 395 Solarium (Polimertecnic, Girona, España), que elimina la radiación de longitud de onda menor de 395 nm, es decir, bloquea el paso de la radiación UV-A y UV-B, y por tanto las plantas reciben solamente radiación PAR.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Materiales y métodos
12
Los filtros se instalaron el día 02/08/2011 coincidiendo con el envero de la viña y se quitaron el 14/10/2011 coincidiendo con la vendimia.
Figura 2. Esquema de la distribución de los tratamientos en cada fila.
Los filtros se dispusieron sobre un bastidor doble fabricado con listones de pino de 3 cm de lado. Éstos estaban fijados sobre postes del mismo material, de 10 cm de lado y 250 cm de altura (Figura 3). Tanto los listones como los postes se trataron con fungicida (sulfato de cobre) para favorecer su resistencia en el exterior. Toda la estructura se clavó en el suelo con una profundidad de unos 20 cm, de tal manera que el marco inferior del filtro quedaba a una distancia perpendicular al suelo de 50 cm.
Figura 3. Estructuras con los filtros de RUV en la viña de Graciano.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Materiales y métodos
13
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
280 330 380 430 480 530 580 630 680
Irrad
ianc
ia e
spec
tral
(W m
-2nm
-1)
Longitud de onda (nm)
0.0
0.1
0.2
0.3
0.4
0.5
290 315 340 365 390
Las irradiancias espectrales en los tres tratamientos se midieron con un espectrorradiómetro (Macam SR9910, Macam Fotometrics Ltd., Livingstone, Escocia), al comienzo del tratamiento y periódicamente a lo largo de todo el periodo de estudio. Dado que los filtros no sufrieron cambios en sus características espectrales, en la Figura 4 se muestra una de las medidas de irradiancia espectral realizada en los tres tratamientos.
Figura 4. Irradiancias espectrales a las que se expusieron las plantas en cada uno de los tres tratamientos: AMBIENTE (azul); FILTRO UV+ (amarillo) y FILTRO UV- (rosa). Las dosis totales de radiación PAR (fotosintética), UV-A y UV-B recibidas por las plantas en la viña durante el periodo de estudio, se midieron en continuo con radiómetros de banda ancha (sensores Skye Quantum SKP 215, SKU 420 y SKU 430, respectivamente) situados en la Universidad de La Rioja y conectados a una unidad automática remota de adquisición y transmisión de datos (Meteodata 3000, Geonica, Madrid). La radiación UV biológicamente efectiva (UVBE) se calculó a partir de las irradiancias espectrales ponderadas con el espectro de acción de Flint y Caldwell (2003) (Tabla 2).
TRATAMIENTO PAR (MJ m-2) UV-A (MJ m-2) UV-B (MJ m-2) UVBE (MJ m-2) AMBIENTE 654 65.17 2.620 0.869
FILTRO UV+ 616 45.62 2.130 0.686 FILTRO UV- 639 5.58 0.290 0.085
Tabla 2. Dosis de PAR, UV-A, UV-B y UVBE recibida por las plantas de cada tratamiento (régimen de radiación) en el período de estudio del 02/08/2011 al 14/10/2011 (nº días = 73).

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Materiales y métodos
14
Variables fisiológicas medidas Todas las medidas fisiológicas se tomaron en hojas y bayas situadas en la cara sur de la espaldera y en las fechas de envero (02/08/2011) y vendimia (14/10/2011), salvo las excepciones que se indican en las variables correspondientes.
Fluorescencia de clorofilas
La fluorescencia de clorofilas in vivo del fotosistema II (PSII) se midió con un fluorímetro portátil (MINI-PAM, Walz, Effeltrich, Alemania) mediante la aplicación de la técnica de pulsos saturantes (Schreiber et al., 1995) (Figura 5). La fluorescencia máxima y mínima (Fm y F0) en hojas intactas, previamente adaptadas a la oscuridad, se determinó utilizando un haz modulado de 600 Hz y 0.03 µmol m-2 s-1 de densidad de flujo fotónico fotosintético (PPFD) y un pulso saturante de 8000 µmol m-2 s-1 PPFD y una duración de 0.8 s. El rendimiento cuántico máximo del PSII se expresa como el cociente Fv / Fm, donde Fv = Fm – Fo (Schreiber et al., 1995).
Figura 5. Inducción típica de señal de fluorescencia obtenida por el método de pulsos saturantes. ML, luz de medida. SP, pulso saturante. AL, luz actínica.
La fluorescencia se midió también con un equipo AquaPen AP-P 100 (Photon Systems Instruments, Brno, República Checa). Las curvas OJIP se realizaron a partir de los datos de fluorescencia de clorofilas inducida mediante la saturación con 3000 µmol m-2 s-1 PPFD. Dicha fluorescencia se registró a diferentes frecuencias entre 10 µs y 2 s. Los datos de los ejes de la curva OJIP y ciertos parámetros derivados se calcularon con el software Fluorpen 2.0 basado en Strasser et al. (2000).

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Materiales y métodos
15
Índice de Esclerofilia (IE)
Para el cálculo del Índice de Esclerofilia (IE) se tomaron con un sacabocados discos de hojas de una superficie total S = 3.98 cm2. Previamente se pesó la masa fresca (PF) y posteriormente las muestras se secaron a 60 ºC durante 24 h en estufa para obtener la masa seca (PS). El IE se calculó como el cociente entre PS / S (mg PS cm-2).
Pigmentos fotosintéticos
El contenido en clorofilas se midió periódicamente en las hojas, de forma no destructiva, con el medidor SPAD (SPAD-502, Minolta) y con el DUALEX® 4 SCIENTIFIC (FORCE-A, Dynamax, Houston, USA). También se analizaron clorofilas y carotenoides en hojas mediante métodos destructivos. Para ello se recogieron en campo 2 discos de hojas de S = 3.98 cm2 de cada una de las plantas de vid en cada tratamiento y repetición. En hollejos, se analizó únicamente el contenido en clorofilas y sólo en envero. Para ello se recogieron en campo 6 bayas de cada cepa seleccionándolas por su flotabilidad en soluciones con diferentes densidades de NaCl, hasta recoger como válidas aquéllas que se encontraban en el intervalo de estratificación (150-170 g l-1) correspondiente al estado de maduración de cosecha (Lijavetzky et al., 2012), que corresponde a un contenido en azúcar de 24 ºBrix aproximadamente. Para ello se prepararon 3 disoluciones de cloruro sódico (NaCl) con una concentración de 190, 170 y 150 g l-1, respectivamente. Las bayas de cada tratamiento (régimen de radiación), se fueron introduciendo en cada solución, de la más concentrada a la menos concentrada, recogiendo en las dos primeras soluciones, únicamente aquellas bayas que flotaban. En la tercera solución, se seleccionaron las que se hundían en el fondo del recipiente, bayas con una densidad de entre 150-170 g l-1.
Tanto los discos de hojas como las bayas se congelaron in situ en N2 líquido y se trasladaron al laboratorio en un equipo de criopreservación (modelo 8040, Thermo Fisher Scientific Forma®). En el caso de los hollejos, se separaron de la pulpa siempre congelados en N2 líquido. Para la extracción de los pigmentos fotosintéticos, los discos de hojas y los hollejos se molieron con un Tyssue-Lyser (Qiagen, Hilden, Germany) realizando dos ciclos de 45 s a 30 Hz. De la masa de hollejos obtenida se recogieron aproximadamente 100 mg por muestra. Posteriormente se añadió acetona 100% (con 0.5 g l-1 de CaCO3). Tras 24 h a 4 ºC, las muestras se filtraron con filtro de diámetro 0.45 µm (SimplePure MCE) y se enrasaron a un volumen de 3 ml.
La identificación y cuantificación de pigmentos fotosintéticos en hoja se realizó mediante HPLC (HP1100 HPLC, Agilent Technologies, Palo Alto, CA, USA) utilizando el método descrito por García-Plazaola y Becerril (2001) con modificaciones de Otero et al. (2006). Los diferentes pigmentos se separaron en una columna en fase reversa Hypersil ODS-C18 (5 µm, 250 x 4 mm) protegida con un guardacolumna Hypersil ODS (5 µm, 4 x 4 mm i.d.). La fase móvil estaba compuesta por dos solventes, A (acetonitrilo: metanol: tris 0.1 M pH = 8 84:2:14) y B (metanol: acetato de etilo 68:32). Se usó un gradiente lineal de 100% del solvente A a 100% de solvente B para los primeros 12 min, seguido de una elución isocrática con 100% de B para los siguientes 6 min; después de esto, 1 min de gradiente lineal desde 100% de B a 100% de A, y durante otros 5

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Materiales y métodos
16
min otra elución isocrática con 100% de A que permite el reequilibio de la columna. La tasa de flujo de solvente fue de 1.2 ml min-1, con un volumen de inyección de 15 µl. Durante todo el proceso de separación, la columna se mantuvo a una T de 20 ºC. Durante el análisis en HPLC las muestras se mantuvieron a 4ºC en un compartimento refrigerado del automuestreador Agilent 1200. El espectro de absorción de cada pigmento se registró desde 250 a 700 nm. La integración de los cromatogramas se hizo a 445 nm. Para la cuantificación de los pigmentos se emplearon rectas de calibrado calculadas a partir de estándares comerciales de clorofila a y b (Fluka) y carotenoides: luteína, zeaxantina y β-caroteno (CaroteNature). Se determinó el contenido total de clorofilas (Clo a + b), el cociente entre clorofilas (Clo a / b), los carotenoides totales y las xantofilas (luteína, neoxantina, violaxantina, anteraxantina y zeaxantina). El índice de xantofilas se calculó como (Anteraxantina + Zeaxantina) / (Anteraxantina + Zeaxantina + Violaxantina), (A+Z) / (A+Z+V). En hollejo, se midieron las absorbancias para longitudes de onda de 661.6 y 644.8 nm con un espectrofotómetro Perkin-Elmer λ35 (Perkin-Elmer, Wilton, CT, USA). A partir de dichos datos de absorbancia y en base a las ecuaciones acordes con la solución de extracción (Lichtenthaler et al., 1987), se calcularon las concentraciones de clorofila a y b así como Clo a + b y Clo a / b. Tanto el proceso de extracción como de medición se realizó en condiciones de baja iluminación y a T = 4 ºC.
Compuestos absorbentes de radiación ultravioleta (CARUV)
Los compuestos que absorben RUV se cuantificaron periódicamente en hojas, de forma no destructiva, con un medidor DUALEX® 4 SCIENTIFIC (FORCE-A, Dynamax, Houston, USA). Además, al inicio (envero) y al final (vendimia) del estudio se analizaron y cuantificaron los CARUV por métodos destructivos tanto en hojas como en hollejos. En ambos casos se analizaron tanto los solubles (CARUV-S), presumiblemente localizados en las vacuolas, como los insolubles (CARUV-I), unidos a la pared celular. El análisis se realizó siguiendo los métodos descritos por Schmitz-Hoerner y Weissenbock, 2003; Arróniz-Crespo et al., 2006 y Fabón et al. 2010. La recogida, transporte y molienda de muestras de hojas y hollejos se hizo tal y como se ha descrito en el apartado de pigmentos fotosintéticos.
Una vez molida la muestra, para la extracción de los CARUV se añadió metanol: agua: 7M HCl (70:29:1 v/v/v), 5 ml en el caso de las hojas y 2 ml para los hollejos. A continuación las muestras se mantuvieron 24 h a 4 ºC, en oscuridad. El extracto se centrifugó a 6000 g durante 15 min. Se separaron el sobrenadante y el pellet para la obtención de CARUV-S y CARUV-I, respectivamente (Clarke y Robinson, 2008).
El contenido global de CARUV-S se calculó, en unidades arbitrarias, como el área bajo la curva de absorbancia en el intervalo 280 – 315 nm (AUC280-315) y 280–400 nm (AUC280-400), por unidad de peso seco y por unidad de superficie en hoja, y por unidad de peso fresco en hollejo, usando un espectrofotómetro Perkin-Elmer λ35 (Perkin-Elmer, Wilton, CT, USA). El primer intervalo corresponde a la absorción de UV-B, y el segundo a UV-A y UV-B. Ambas variables estaban estrechamente correlacionadas (P<0.001), y por tanto solamente se han reflejado los resultados del intervalo 280-400 nm.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Materiales y métodos
17
El pellet obtenido de la extracción con metanol se hidrolizó añadiendo 1 ml de NaOH. A continuación las muestras se metieron en un baño durante 3 h con agua a 80 ºC. Tras acidificación con 1 ml de HCl (5.6 N), se lavaron tres veces con acetato de etilo. Tras la evaporación del sobrenadante recogido en los lavados, utilizando un rotavapor (Büchi R-200) a 40ºC (Büchi Heating Bath B-490), el material se resuspendió en metanol 100%, obteniendo un volumen final de 2 ml (muestras de hoja) y 1 ml (muestras de hollejo). El contenido global de CARUV-I se midió también por espectrofotometría en las mismas unidades que se han descrito para los niveles de CARUV-S. La identificación y cuantificación de los CARUV individuales de las hojas se realizó mediante HPLC (Agilent HP1100 HPLC System, Agilent Technologies, Palo Alto, CA, USA), siguiendo el método descrito por Arróniz-Crespo et al., 2006. Los diferentes compuestos se separaron con una columna de fase reversa Hypersil ODS-C18 (5 µm, 250 x 4 mm i.d.) protegida por un guardacolumna Hypersil ODS (5 µm, 4 x 4 mm i.d.). La fase móvil estaba compuesta por dos solventes: solvente A (ácido acético pH 2.4) y solvente B (acetonitrilo: metanol: agua (v/v/v)). Para el análisis se bombeó primero un 90 % de A y un 10 % de B durante 10 min, seguido de un gradiente lineal de 100 % de B durante los siguientes 15 min y una elución isocrática con 100 % de B durante un periodo de 5 min. Se restauraron las condiciones iniciales con el equilibrado de la columna durante los últimos 8 min. La tasa de flujo de solvente fue de 1.2 ml min-1, con un volumen de inyección de 15 µl, estando la columna a una temperatura de 20 ºC. Tanto la identificación como la cuantificación de los compuestos se hizo con un detector Agilent (photodiode array) (Agilent 1100 DAD) a 324 nm (ref. 700 nm) y el espectro de absorción se registró en un rango de longitudes de onda entre 200 y 500 nm. La identificación y cuantificación de los CARUV individuales de los hollejos (Sáenz-Navajas et al., 2010, con algunas modificaciones) se realizó con un cromatógrafo Waters Acquity UPLC TM (Milford, MA, USA) equipado con una columna ACQUITY UPLC® BEH-C18 (1.7 µm x 100 x 2.1 mm) mantenida a 40ºC y una precolumna Waters VanGuard del mismo material. La fase móvil estaba compuesta de dos solventes: solvente A (acetonitrilo con 0.1 % (v/v) de ácido fórmico) y solvente B (solución acuosa con 0.1% (v/v) de ácido fórmico). El volumen de inyección fue de 5 µl. La composición de la fase móvil varió de acuerdo al siguiente gradiente lineal: 0 a 7 min, 99.5-80% de B; 7 a 9 min, 80-50% de B; 9-11.7 min, 50-0% de B; 11.7 a 15 min, 0-99.5% de B. El sistema UPLC estaba acoplado a un espectrómetro de masas (Microtof-Q, Q-TOF, Bruker Daltonik GMBH, Germany). Se empleó un módulo detector de UV de longitud de onda 324 nm (Acquity UV Detector). Los datos obtenidos fueron procesados mediante el software Bruker Compass DataAnalysis 4.0.234.0 (Bruker, Bremen, Alemania). La fuente de electrospray operó siempre en negativo. El potencial capilar se fijó a 4 kV. La temperatura del gas de secado fue de 200 ºC y su flujo de 9 l min-1. El gas nebulizador se estableció a una presión de 3.5 bar y 25 ºC. Los espectros se adquirieron entre 120-1505 m / z en modo negativo. Para la cuantificación de los diferentes compuestos se emplearon rectas de calibrado calculadas a partir de estándares comerciales: ácido protocatéquico (03930590), kaempferol-3-O-glucósido (68437), quercetina-3-O-glucurónido (90733) y procianidina B1 (1952) de FLUKA; ácido trans p-cumárico (C9008), ácido trans ferúlico (46278), ácido sirínguico (S6881), resveratrol (R5010), kaempferol (60010), epicatequina (E4018), catequina (C1251), quercetina (Q0125), epigallocatequina (E3768), ácido

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Materiales y métodos
18
cafeoil-tartárico (15029), isorhamnetina (17794), miricetina (70050), quercetina-3-O-glucopiranósido (91802), quercetina-3-O-galactósido (83388) y quercetina-3-O-rutinósido (R5143) de SIGMA. Para aquellos compuestos para los que no se disponía estándar comercial, se empleó la recta de calibrado del estándar con cromóforo similar al del compuesto a analizar.
Análisis estadístico Las relaciones entre variables se examinaron calculando los coeficientes de correlación de Pearson (r). Para el estudio estadístico de los efectos del tratamiento (régimen de radiación) y del tiempo sobre las variables fisiológicas, se realizó un análisis de varianza (ANOVA) de dos vías, con medidas repetidas para el tiempo, si los datos eran normales (test de Shapiro–Wilks) y homocedásticos (test de Levene). En caso contrario, se realizaron pruebas no paramétricas (test de Kruskal-Wallis). En el caso de encontrar diferencias significativas, la comparación de medias se realizó mediante un test de Tukey (en variables que eran normales y homocedásticas), o un test de Mann-Whitney en variables que no lo eran. Para comparar las diferencias entre algunas variables medidas en envero y vendimia, se utilizó la t de Student. Todos los procedimientos estadísticos se desarrollaron con el paquete SPSS 19.0 para Windows (SPSS Inc., Chicago, Illinois, USA).

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
19
0
50
100
150
200
250
300
350
0.9 1.1 1.3 1.5 1.7 1.9
CAR
UV-
S (A
UC
280-
400 cm
-2)
(a)
p < 0.001
0
20
40
60
80
100
120
0.6 0.8 1.0 1.2 1.4 1.6 1.8
CAR
UV-
I (AU
C 28
0-40
0 cm
-2)
(b)
NS
0
10
20
30
40
50
60
0.6 0.8 1.0 1.2 1.4 1.6 1.8
CAR
UV-
I (AU
C 28
0-40
0 m
g-1 )
Flavonoides (ud DUALEX)
(d)
NS
0
20
40
60
80
100
120
140
160
0.9 1.1 1.3 1.5 1.7 1.9
CAR
UV-
S (A
UC
280-
400 m
g-1 )
Flavonoides (ud DUALEX)
(c)
NS
Resultados
Comparaciones metodológicas Las variables que se midieron en hojas utilizando diferentes métodos se correlacionaban significativamente (P<0.001): clorofilas DUALEX vs. clorofilas SPAD, Fv / Fm MINIPAM vs. AQUAPEN, contenido de clorofilas medido con el DUALEX y el medido en laboratorio mediante extracción con acetona expresado por unidad de superficie, y la variable Flav con el DUALEX y el contenido de CARUV-S expresado por unidad de superficie (Figura 6.a) pero no por peso seco (Figura 6.c). No se obtuvo tampoco una buena correlación entre las medidas Flav del DUALEX y el contenido en CARUV-I (Figuras 6.b y 6.d).
Figura 6. Correlaciones entre el contenido en flavonoides medido con DUALEX y CARUV (azul: solubles; rojo: insolubles), expresados por superficie (a y b) y por peso seco (c y d). Se muestran Medias, línea de tendencia y significación (N = 54).

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
20
Variaciones en función del tiempo y del tratamiento (régimen de radiación) Se observaron diferencias significativas con respecto al tiempo en todas las variables medidas en hoja, salvo en el cociente Clo a / Clo b (Tabla 3). La zeaxantina, índice de xantofilas, contenido de carotenoides totales, ácido férulico, ácido p-cumaroil-tartárico, ácido feruloil-tartárico y cis kaempferol-3-O-glucósido, no se pudieron analizar con respecto a dicho factor, por no ser posible cuantificarlos en envero. Únicamente se obtuvieron diferencias significativas con respecto al tratamiento (régimen de radiación) en cinco variables medidas en las hojas: Fv / Fm, ácido p-cumárico, ácido p-cafeoil-tartárico, quercetina-3-O-glucurónido y cis kaempferol-3-O-glucósido (Tabla 4). Entre estos resultados, se puede destacar que el carotenoide más abundante en hoja es la luteína. Además, los CARUV están en mucha mayor concentración (aproximadamente el triple) en la fracción soluble que en la insoluble. Dentro de los compuestos individuales, destaca el ácido p-cafeoil-tartárico como el mayoritario en hoja, seguido de las quercetinas, siendo el resto mucho menos abundante. En el hollejo, se observaron diferencias significativas con respecto al tratamiento (régimen de radiación) en las siguientes variables: Clo a + b, Clo a / Clo b, CARUV-S, ácido protocatéquico, ácido p-cumárico, catequina, miricetinas glucosiladas, kaempferoles glucosilados, quercetinas glucosiladas, isorhamnetina-3-O-glucósido y siringetina-3-O-glucósido (Tabla 5). Dentro de los compuestos individuales en hollejo, destaca como mayoritaria la miricetina-3-O-glucósido, seguida de la quercetina-3-O-glucopiranósido y la epigallocatequina, siendo el resto de compuestos bastante menos abundantes (Tabla 5).

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
21
VARIABLE TIEMPO
Fv / Fm *** Índice de Esclerofilia (IE) * Clorofila (DUALEX) ** Clo a *** Clo b *** Clo a + b *** Clo a / Clo b NS β-Caroteno *** Luteína *** Anteraxantina *** Violaxantina ** Neoxantina *** Flav (DUALEX) ** CARUV-S *** CARUV-I *** Ácido p-cumárico *** Ácido p-cafeoil-tartárico *** Quercetina- 3-O-glucopiranósido *** Quercetina-3-O- glucurónido *** Trans kaempferol-3-O-glucósido ***
Tabla 3. Efectos del tiempo en las variables fisiológicas medidas en hoja, analizados mediante ANOVA, t de Student o test de Kruskal-Wallis (*** P < 0.001, ** P < 0.01, * P < 0.05, NS no significativo).

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
22
VARIABLE AMBIENTE FILTRO UV+ FILTRO UV- SIG.
Fv / Fm 0.538 ± 0.034 0.585 ± 0.025 0.445 ± 0.045 *Índice de esclerofilia IE (mg cm-2) 7.04 ± 0.35 6.94 ± 0.27 6.95 ± 0.51 NSClorofila (DUALEX) 28.3 ± 1.0 29.8 ± 0.8 29.4 ± 1.0 NSClo a (µg mg-1) 3.10 ± 0.36 3.88 ± 0.37 3.55 ± 0.41 NSClo b (µg mg-1) 1.17 ± 0.14 1.49 ± 0.15 1.37 ± 0.17 NSClo a + b (µg mg-1) 6.54 ± 0.87 7.54 ± 0.51 8.20 ± 0.85 NSClo a / Clo b 2.68 ± 0.07 2.59 ± 0.05 2.52 ± 0.05 NSβ-Caroteno (µg mg-1) 0.839 ± 0.094 1.011 ± 0.091 0.956 ± 0.099 NSLuteína (µg mg-1) 2.07 ± 0.21 2.64 ± 0.15 2.54 ± 0.40 NSZeaxantina (µg mg-1) 0.612 ± 0.066 0.664 ± 0.035 0.676 ± 0.096 NSAnteraxantina (µg mg-1) 0.490 ± 0.067 0.461 ± 0.041 0.563 ± 0.081 NSViolaxantina (µg mg-1) 0.653 ± 0.053 0.694 ± 0.098 0.886 ± 0.153 NSNeoxantina (µg mg-1) 0.443 ± 0.053 0.547 ± 0.038 0.518 ± 0.084 NSÍndice xantofilas (A + Z) /( A + Z + V) 0.629 ± 0.018 0.640 ± 0.037 0.597 ± 0.051 NSCarotenoides totales (µg mg-1) 5.92 ± 0.66 6.48 ± 0.35 7.41 ± 0.70 NSFlav (DUALEX) 1.42 ± 0.02 1.35 ± 0.03 1.37 ± 0.03 NSCARUV- S (AUC280-400 cm-2) 235 ± 11 209 ± 18 239 ± 20 NSCARUV-I (AUC280-400 cm-2) 68.6 ± 4.9 69.4 ± 5.5 72.8 ± 2.9 NSÁcido p -cumárico (µg cm-2) 0.676 ± 0.037 1.040 ± 0.070 0.974 ± 0.038 ***Ácido ferúlico (µg cm-2) 0.418 ± 0.073 0.302 ± 0.016 0.382 ± 0.037 NSÁcido p -cumaroil-tartárico (µg cm-2) 1.28 ± 0.11 1.65 ± 0.15 1.40 ± 0.18 NSÁcido p -cafeoil-tartárico (µg cm-2) 77.0 ± 2.6 78.9 ± 4.2 62.7 ± 3.3 **Ácido feruloil-tartárico (µg cm-2) 1.41 ± 0.10 1.80 ± 0.23 1.48 ± 0.13 NSQuercetina- 3-O -glucopiranósido (µg cm-2) 22.2 ± 2.8 26.5 ± 5.8 17.5 ± 3.5 NSQuercetina-3-O - glucurónido (µg cm-2) 37.3 ± 2.4 31.9 ± 4.6 33.3 ± 3.0 *Cis kaempferol-3-O -glucósido (µg cm-2) 3.30 ± 0.42 4.15 ± 0.59 2.35 ± 0.41 *Trans kaempferol-3-O -glucósido (µg cm-2) 3.90 ± 0.49 3.68 ± 0.24 3.03 ± 0.27 NS
Tabla 4. Valores de las variables medidas en hoja en vendimia, en función del tratamiento (régimen de radiación). Se muestra MEDIA ± ET. Los efectos del tratamiento se analizaron mediante ANOVA de un factor o test de Kruskal-Wallis (*** P < 0.001, ** P < 0.01, * P < 0.05, NS no significativo).

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
23
VARIABLE AMBIENTE FILTRO UV+ FILTRO UV- SIG.
Clo a (µg mg-1) 0.026 ± 0.002 0.035 ± 0.003 0.028 ± 0.002 *Clo b (µg mg-1) 0.040 ± 0.005 0.054 ± 0.008 0.033 ± 0.003 *Clo a + b (µg mg-1) 0.065 ± 0.007 0.090 ± 0.010 0.062 ± 0.005 **Clo a / Clo b 0.679 ± 0.043 0.688 ± 0.054 0.855 ± 0.020 *CARUV-S (AUC280-400 mg-1) 15.5 ± 0.9 12.6 ± 0.4 12.3 ± 0.5 **CARUV-I (AUC280-400 mg-1) 2.75 ± 0.25 2.52 ± 0.26 2.20 ± 0.19 NSÁcido protocatéquico (µg g-1) 6.35 ± 0.52 4.12 ± 0.82 3.32 ± 0.74 *Ácido siríngico (µg g-1) 178 ± 23 175 ± 56 175 ± 33 NSResveratrol (µg g-1) 4.59 ± 0.55 5.47 ± 0.54 5.30 ± 0.52 NSResveratrol-3-O - glucósido (µg g-1) 107 ± 10 123 ± 11 91.9 ± 7.7 NSÁcido p -cumárico (µg g-1) 211 ± 22 354 ± 34 183 ± 31 **Ácido p -cumaroil-tartárico (µg g-1) 99.2 ± 17.1 154 ± 27 181 ± 32 NSÁcido p-cafeoil-tartárico (µg g-1) 86.1 ± 16.6 55.0 ± 23.8 94.3 ± 25.6 NSCatequina (µg g-1) 32.3 ± 1.5 45.6 ± 3.7 43.6 ± 1.7 **Epicatequina (µg g-1) 7.13 ± 0.54 9.49 ± 0.93 8.25 ± 0.89 NSEpigallocatequina (µg g-1) 238 ± 21 305 ± 45 218 ± 13 NSAstilbina (µg g-1) 10.67 ± 0.94 8.24 ± 1.90 7.87 ± 1.17 NSProcianidina B1 (µg g-1) 32.4 ± 1.9 38.1 ± 2.4 37.2 ± 2.8 NSMiricetina (µg g-1) 77.3 ± 19.6 72.6 ± 9.6 43.7 ± 9.5 NSMiricetina-3-O -glucósido (µg g-1) 837 ± 76 710 ± 69 474 ± 25 **Miricetina-3-O -glucurónido (µg g-1) 34.6 ± 4.3 27.9 ± 2.3 21.7 ± 1.5 *Kaempferol-3-O -galactósido (µg g-1) 21.8 ± 2.6 29.6 ± 3.3 16.1 ± 1.8 **Kaempferol-3-O -glucósido (µg g-1) 113.1 ± 2.3 61.7 ± 8.3 46.4 ± 7.2 ***Quercetina-3-O -glucósido (µg g-1) 38.5 ± 8.8 43.8 ± 6.7 27.1 ± 3.9 NSQuercetina-3-O -galactósido (µg g-1) 68.8 ± 3.9 39.4 ± 6.6 38.7 ± 3.2 ***Quercetina-3-O -glucopiranósido (µg g-1) 436 ± 19 301 ± 25 254 ± 18 ***Quercetina-3-O -glucurónido (µg g-1) 576 ± 13 429 ± 24 468 ± 57 *Quercetina-3-O -rutinósido (µg g-1) 10.67 ± 0.40 6.00 ± 0.69 11.32 ± 2.02 *Isorhamnetina-3-O -glucósido (µg g-1) 184 ± 16 135 ± 15 106 ± 9 **Isorhamnetina-3-O -glucurónido (µg g-1) 13.8 ± 1.6 11.4 ± 1.4 12.4 ± 1.7 NSSiringetina-3-O -glucósido (µg g-1) 104.1 ± 3.7 78.0 ± 3.3 71.7 ± 4.5 ***
Tabla 5. Valores de las variables medidas en hollejo en vendimia, en función del tratamiento (régimen de radiación). Se muestra MEDIA ± ET. Los efectos del tratamiento se analizaron mediante ANOVA de un factor o test de Kruskal-
Wallis (*** P < 0.001, ** P < 0.01, * P < 0.05, NS no significativo).

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
24
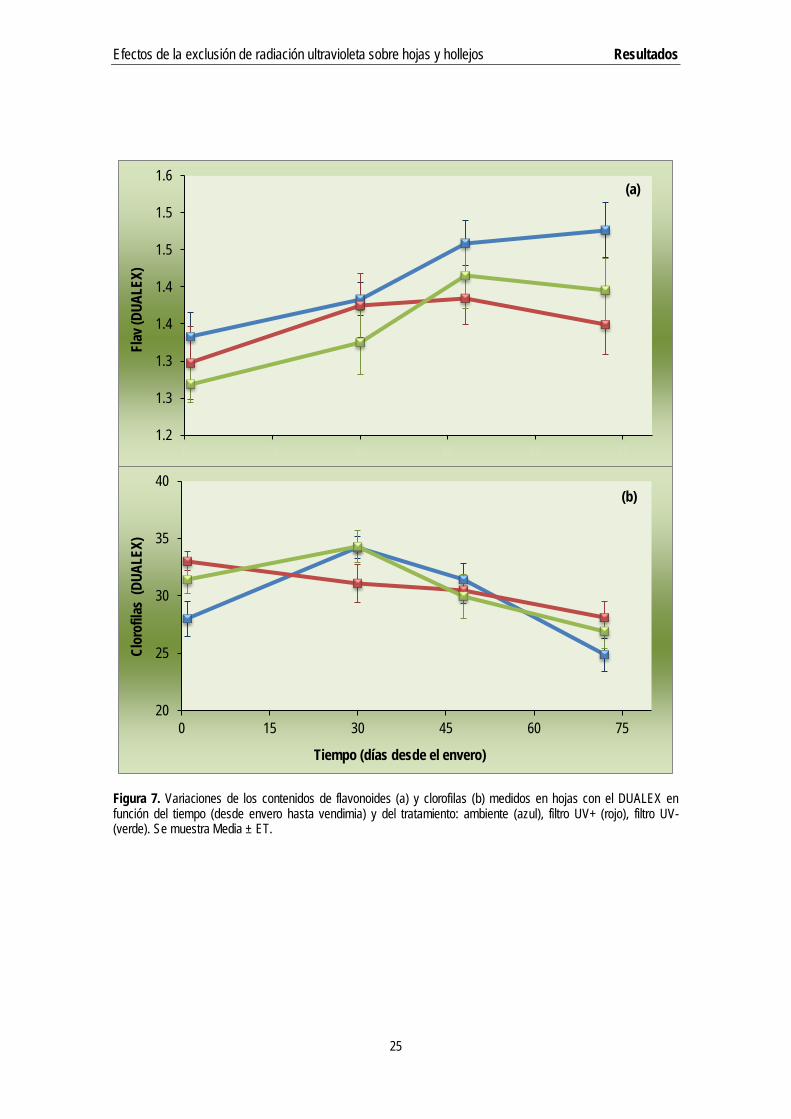
Entre las variables medidas periódicamente en hojas, el contenido de flavonoides con respecto al tiempo aumentó significativamente desde el envero hasta septiembre, estabilizándose prácticamente desde entonces hasta la vendimia (Figura 7.a). Esto ocurría en todos los regímenes de radiación, aunque menos acusadamente en el tratamiento filtro UV+. El contenido en clorofilas, en cambio, solía alcanzar su nivel máximo a primeros de septiembre para luego seguir una tendencia descendente hasta la vendimia (Figura 7.b). No se apreciaron diferencias significativas entre tratamientos (regímenes de radiación) en ninguna de las dos variables (ANOVA de dos vías).
En las variables medidas en hojas sólo en dos momentos, los cambios registrados desde envero a vendimia fueron los siguientes (todos ellos significativos): aumentos en Índice de Esclerofilia, anteraxantina, CARUV-S, CARUV-I, ácido p-cumárico, ácido p-cafeoil-tartárico, quercetina-3-O-glucopiranósido, quercetina-3-O-glucurónido y trans kaempferol-3-O-glucósido, y descensos en Fv / Fm, clorofilas (medidas mediante extracción), β-Caroteno, luteína, violaxantina y neoxantina. Fueron especialmente destacables los aumentos en las quercetinas y el ácido p-cumárico, que se multiplicaron 3-4 veces. Dentro de las variables medidas en hojas en vendimia, hubo diferencias significativas entre tratamientos en Fv / Fm, siendo menor en ausencia de RUV (Figura 8.a). En el Índice de Esclerofilia no se encontraron diferencias significativas entre regímenes de radiación (Figura 8.b). Lo mismo sucedió en las variables relacionadas con el contenido en pigmentos fotosintéticos, aunque en éstas se observó una tendencia creciente en la cantidad de clorofila y carotenoides totales, y decreciente en el cociente Clo a / Clo b, en el tratamiento filtro UV- (Figura 8.c, 8.d, 8.e). El ciclo de las xantofilas muestra una tendencia a una mayor síntesis de violaxantina en tratamientos con filtro UV+ y UV- (Figura 8.f). Tanto los CARUV-S como CARUV-I no mostraron diferencias significativas entre tratamientos (régimen de radiación) (Figura 9.a, 9.b). En cuanto a los compuestos individuales identificados en hoja, sólo se encontraron diferencias significativas en cuatro (ácido p-cumárico, ácido cafeoil-tartárico, quercetina-3-O-glucurónido y cis kaempferol-3-O-glucósido). En el primer caso se observó que se produce una mayor acumulación de dicho compuesto en aquellos tratamientos que poseen filtro (UV+ y UV-), independientemente de que éste deje pasar o no la RUV (Figura 9.c). En las otras tres variables la tendencia general es a acumular menos cantidad de dichos compuestos cuando la planta no se ve sometida a RUV (Figura 9.d, 9.e, 9.f).
Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
25
Figura 7. Variaciones de los contenidos de flavonoides (a) y clorofilas (b) medidos en hojas con el DUALEX en función del tiempo (desde envero hasta vendimia) y del tratamiento: ambiente (azul), filtro UV+ (rojo), filtro UV- (verde). Se muestra Media ± ET.
1.2
1.3
1.3
1.4
1.4
1.5
1.5
1.6
0 15 30 45 60 75
Flav
(DUA
LEX)
(a)
20
25
30
35
40
0 15 30 45 60 75
Clor
ofila
s (D
UALE
X)
Tiempo (días desde el envero)
(b)

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
26
Figura 8. Efecto del tratamiento (ambiente, azul; filtro UV+, rojo; filtro UV-, verde) sobre diversas variables analizadas en muestras de hoja recogidas en vendimia (N = 9): Fv / Fm (a), Índice de Esclerofilia (b), contenido de Clo a + b (c), cociente Clo a / Clo b (d), carotenoides totales (e) e Índice de Xantofilas (f). Se muestra Media ± ET. Se indican mediante letras distintas las diferencias significativas entre los regímenes de radiación.
0.00
0.10
0.20
0.30
0.40
0.50
0.60
0.70
AMBIENTE FILTRO UV FILTRO NO UV
F v / F
m
a
b
a (a)
3.5
4.0
4.5
5.0
5.5
6.0
6.5
7.0
7.5
8.0
AMBIENTE FILTRO UV FILTRO NO UV
Índi
ce d
e Esc
lerof
ilia (m
g cm
-2) a a
a
(b)
0.0
2.0
4.0
6.0
8.0
10.0
AMBIENTE FILTRO UV FILTRO NO UV
Clo
a+ b
(µg
mg-1
)
a
aa
(c)
0.5
1.0
1.5
2.0
2.5
3.0
AMBIENTE FILTRO UV FILTRO NO UV
Clo
a/ b
a aa(d)
0.5
1.5
2.5
3.5
4.5
5.5
6.5
7.5
8.5
AMBIENTE FILTRO UV+ FILTRO UV-
Caro
teno
ides
tota
les (µ
g m
g-1)
a
a
a
(e)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
AMBIENTE FILTRO UV+ FILTRO UV-
Índi
ce X
anto
filas
a aa(f)

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
27
Figura 9. Efecto del tratamiento (ambiente, azul; filtro UV+, rojo; filtro UV-, verde) sobre diversas variables analizadas en muestras de hoja recogidas en vendimia (N = 9): CARUV-S (a), CARUV-I (b), ácido p-cumárico (c), ácido p-cafeoil-tartárico (d), quercetina-3-O-glucurónido (e) y cis kaempferol-3-O-glucósido (f). Se muestra Media ± ET. Se indican mediante letras distintas las diferencias significativas entre los regímenes de radiación.
0
50
100
150
200
250
300
1 2 3
CARU
V-S
(AUC
280-
400
cm-2
)
a aa
(a)
0
10
20
30
40
50
60
70
80
1 2 3
CARU
V-I (
AUC 2
80-4
00cm
-2)
a aa (b)
0
20
40
60
80
100
AMBIENTE FILTRO UV FILTRO NO UV
Ácid
o p-
cafe
oil-t
artá
rico
(µg
cm-2
)
a
b
a(d)
0
5
10
15
20
25
30
35
40
45
AMBIENTE FILTRO UV+ FILTRO UV-
Quer
cetin
a-3-
O-gl
ucur
ónid
o (µ
g cm
-2)
abb
(e)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1 2 3
Ácid
o p-
cum
árico
(µg
cm-2
)
a
bb (c)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
AMBIENTE FILTRO UV+ FILTRO UV-
Cis k
aem
pfer
ol-3
-O-g
lucó
sido
(µg
cm-2
)
ab
b
a (f)

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
28
El análisis de los pigmentos fotosintéticos en hollejo indica que se produjo una disminución del contenido de clorofila y un aumento del cociente Clo a / Clo b en el tratamiento filtro UV- (Figura 10).
Figura 10. Efecto del tratamiento (ambiente, azul; filtro UV+, rojo; filtro UV-, verde) sobre diversas variables analizadas en muestras de hollejo recogidas en vendimia (N = 9): Clo a + b (a), y cociente Clo a / b (b). Se muestra Media ± ET. Se indican mediante letras distintas las diferencias significativas entre los regímenes de radiación. Los CARUV-S mostraron diferencias significativas con respecto al régimen de radiación, siendo su acumulación menor en los tratamientos con filtro (UV+ y UV-) (Figura 11.a). Los CARUV-I, en cambio, no mostraron diferencias significativas entre tratamientos (régimen de radiación) (Figura 11.b).
Figura 11. Efecto del tratamiento (ambiente, azul; filtro UV+, rojo; filtro UV-, verde) sobre diversas variables analizadas en muestras de hollejo recogidas en vendimia (N = 9): CARUV-S (a) y CARUV-I (b). Se muestra Media ± ET. Se indican mediante letras distintas las diferencias significativas entre los regímenes de radiación.
0.00
0.02
0.04
0.06
0.08
0.10
0.12
AMBIENTE FILTRO UV+ FILTRO UV-
Clo
a + b
( µg
mg-1
) aa
b(a)
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
AMBIENTE FILTRO UV+ FILTRO UV-
Clo
a/ b
b
a a
(b)
0
2
4
6
8
10
12
14
16
18
AMBIENTE FILTRO UV+ FILTRO UV-
CARU
V-S
(AUC
280-
400
mg-1
) b
a
b
(a)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
AMBIENTE FILTRO UV+ FILTRO UV-
CARU
V-I (
AUC
280-
400
mg-1
) a
aa
(b)
Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
29
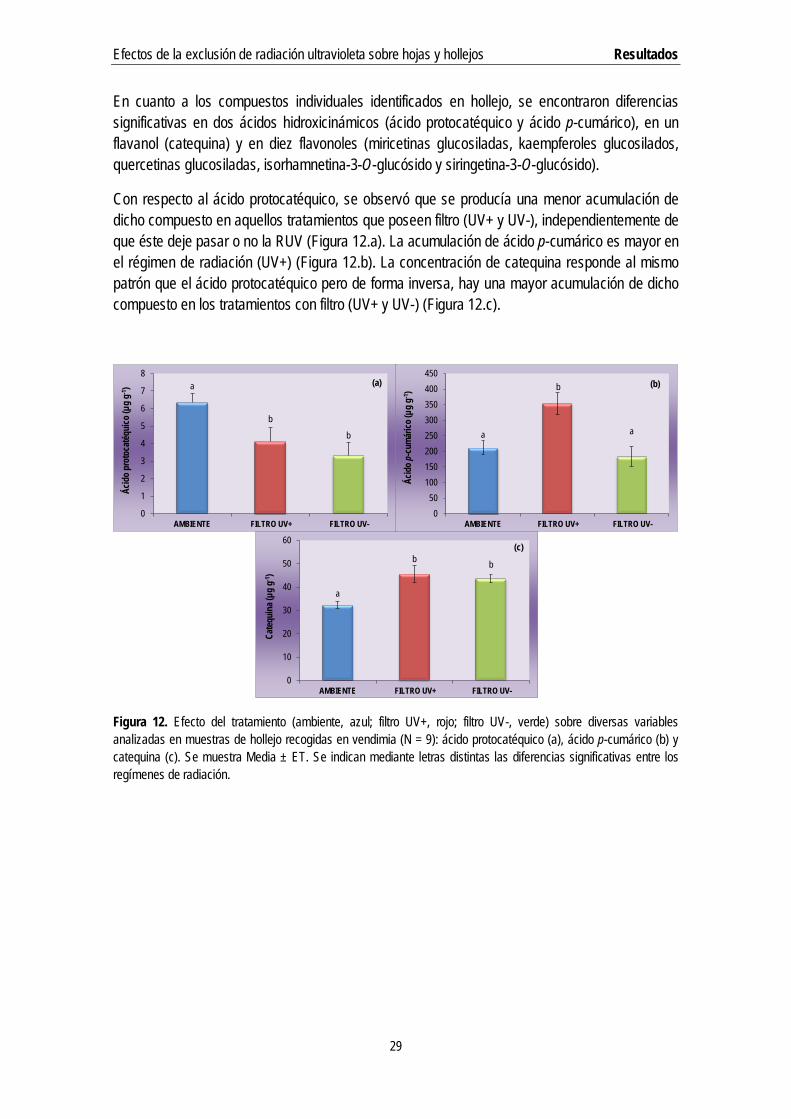
En cuanto a los compuestos individuales identificados en hollejo, se encontraron diferencias significativas en dos ácidos hidroxicinámicos (ácido protocatéquico y ácido p-cumárico), en un flavanol (catequina) y en diez flavonoles (miricetinas glucosiladas, kaempferoles glucosilados, quercetinas glucosiladas, isorhamnetina-3-O-glucósido y siringetina-3-O-glucósido).
Con respecto al ácido protocatéquico, se observó que se producía una menor acumulación de dicho compuesto en aquellos tratamientos que poseen filtro (UV+ y UV-), independientemente de que éste deje pasar o no la RUV (Figura 12.a). La acumulación de ácido p-cumárico es mayor en el régimen de radiación (UV+) (Figura 12.b). La concentración de catequina responde al mismo patrón que el ácido protocatéquico pero de forma inversa, hay una mayor acumulación de dicho compuesto en los tratamientos con filtro (UV+ y UV-) (Figura 12.c).
Figura 12. Efecto del tratamiento (ambiente, azul; filtro UV+, rojo; filtro UV-, verde) sobre diversas variables analizadas en muestras de hollejo recogidas en vendimia (N = 9): ácido protocatéquico (a), ácido p-cumárico (b) y catequina (c). Se muestra Media ± ET. Se indican mediante letras distintas las diferencias significativas entre los regímenes de radiación.
a
b
b
0
1
2
3
4
5
6
7
8
AMBIENTE FILTRO UV+ FILTRO UV-
Ácid
o pr
otoc
atéq
uico
(µg
g-1) (a)
a
b
a
050
100150200250300350400450
AMBIENTE FILTRO UV+ FILTRO UV-
Ácid
o p-
cum
árico
(µg
g-1)
(b)
a
b b
0
10
20
30
40
50
60
AMBIENTE FILTRO UV+ FILTRO UV-
Cate
quin
a (µg
g-1
)
(c)

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
30
a
a
b
0100200300400500600700800900
1,000
AMBIENTE FILTRO UV FILTRO NO UV
Miric
etin
a-3-
O-gl
ucós
ido
(µg
g-1)
(a) a
ab
b
05
1015202530354045
AMBIENTE FILTRO UV FILTRO NO UVMi
ricet
ina-
3-O-
gluc
urón
ido
(µg
g-1)
(b)
ab
a
b
0
5
10
15
20
25
30
35
AMBIENTE FILTRO UV+ FILTRO UV-
Kaem
pfer
ol-3
-O-g
alact
ósid
o(µ
g g-1
)
(c) a
b
b
0
20
40
60
80
100
120
140
AMBIENTE FILTRO UV+ FILTRO UV-
Kaem
pfer
ol-3
-O-g
lucó
sido
(µg
g-1)
(d)
En cuanto a los flavonoles (Figuras 13 y 14), la tendencia general es que la concentración tiende a ser mayor en el tratamiento ambiente, salvo en kaempferol-3-O-galactósido (en el que el tratamiento filtro UV+ es el que tiene mayor concentración) y en la quercetina-3-O-rutinósido (en la que los mayores valores se registran en el filtro UV-). En general, las concentraciones en el régimen de radiación filtro UV- eran menores que en el filtro UV+.
Figura 13. Efecto del tratamiento (ambiente, azul; filtro UV+, rojo; filtro UV-, verde) sobre diversas variables analizadas en muestras de hollejo recogidas en vendimia (N = 9): miricetina-3-O-glucósido (a), en miricetina-3-O-glucurónido (b), en kaempferol-3-O-galactósido (c) y kaempferol-3-O-glucósido (d). Se muestra Media ± ET. Se indican mediante letras distintas las diferencias significativas entre los regímenes de radiación.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Resultados
31
a
b b
0
10
20
30
40
50
60
70
80
AMBIENTE FILTRO UV FILTRO NO UV
Quer
cetin
a-3-
O-ga
lactó
sido
(µg
g-1)
(a)a
b
b
050
100150200250300350400450500
AMBIENTE FILTRO UV FILTRO NO UV
Quer
cetin
a-3-
O-gl
ucop
iranó
sido
(µg
g-1)
(b)
a
bb
0
100
200
300
400
500
600
700
AMBIENTE FILTRO UV FILTRO NO UV
Quer
cetin
a-3-
O-gl
ucur
ónid
o(µ
g g-1
)
(c)
a
b
b
0
50
100
150
200
250
AMBIENTE FILTRO UV+ FILTRO UV-
Isorh
amne
tina-
3-O-
gluc
ósid
o(µ
g g-1
)
(e)
a
b
a
0
2
4
6
8
10
12
14
16
AMBIENTE FILTRO UV FILTRO NO UV
Quer
cetin
a-3-
O-ru
tinós
ido
(µg
g-1)
(d)
a
bb
0
20
40
60
80
100
120
AMBIENTE FILTRO UV+ FILTRO UV-
Sirin
getin
a-3-
O-gl
ucós
ido
(µg
g-1)
(f)
Figura 14. Efecto del tratamiento (ambiente, azul; filtro UV+, rojo; filtro UV-, verde) sobre diversas variables analizadas en muestras de hollejo recogidas en vendimia (N = 9): quercetina-3-O-galactósido (a), quercetina-3-O-glucopiranósido (b), quercetina-3-O-glucurónido (c), quercetina-3-O-rutinósido (d), isorhamnetina-3-O-glucósido (e) y siringetina-3-O-glucósido (f). Se muestra Media ± ET. Se indican mediante letras distintas las diferencias significativas entre los regímenes de radiación.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Discusión
32
Discusión
Comparaciones metodológicas
Las medidas de Fv / Fm realizadas con MINIPAM y AQUAPEN mostraron una correlación muy significativa (P<0.001), por lo que ambos aparatos son aceptables para medir la citada variable en hojas de Graciano. Esto resulta lógico a la vista de los numerosos estudios que utilizan estos aparatos para este fin en diversos tipos de plantas (ver por ejemplo Fabón (2012) y Fernández Marín (2012)). Dado que el AQUAPEN es un instrumento más sencillo y fácil de transportar que el MINIPAM, los resultados obtenidos en este trabajo indican que se podrían simplificar las medidas de Fv / Fm en vid en condiciones de campo. También mostraron correlaciones altamente significativas (P<0.001) las concentraciones de clorofilas obtenidas en hojas de Graciano con los tres métodos utilizados: DUALEX, SPAD (ambos métodos no destructivos y aplicados en campo) y extracción con acetona (método destructivo aplicado en laboratorio), aunque en este último caso solamente cuando las clorofilas se expresaban por unidad de superficie. Al igual que ocurría con Fv / Fm, esta concordancia metodológica es interesante porque podría simplificar la medida de clorofila en vid en condiciones naturales, ya que evitaría las medidas destructivas, mucho más laboriosas y consumidoras de tiempo que las no destructivas. Cerovic et al. (2012) obtuvieron resultados similares al comparar los valores que dieron DUALEX y SPAD en la variedad Merlot. En su trabajo incluso se establece una ecuación consensuada, válida para una gran diversidad de especies, con la que es posible determinar de una forma fácil, rápida y no destructiva, el contenido de Clo a + b (µg cm-2) a partir de los datos reportados por el SPAD-502. Al utilizar esta ecuación para los datos SPAD de nuestro estudio y comparar los valores calculados con los obtenidos mediante la extracción en laboratorio, se observó que ambos grupos de datos estaban correlacionados signficativamente (P<0.01). Por lo tanto, la ecuación consensuada de Cerovic et al. (2012) es también válida para la variedad Graciano de vid. La variable Flav (flavonoides epidérmicos) medida por el DUALEX y los CARUV-S se correlacionaban significativamente por superficie. Nuevamente, esto supone una medida más fácil, rápida y repetitiva (y además no destructiva) de la capacidad de absorción UV de la fracción soluble de las hojas de Graciano en campo, que las medidas espectrofotométricas, más laboriosas y además destructivas. Sin embargo, Flav no se correlacionaba con CARUV-S por peso. Esto puede deberse a que el aparato basa su funcionamiento en la emisión de un haz de luz sobre una superficie foliar concreta, sin tener en cuenta el peso de esa fracción de superficie sobre la que se realiza la medición, y por tanto las medidas DUALEX estarían más relacionadas con la superficie foliar que con el peso (Cerovic et al., 2002). No obstante, en algunos estudios se ha encontrado correlación entre las medidas del DUALEX y la concentración de flavonoides totales por peso medidos mediante HPLC (Morales et al., 2010). La falta de correlación en nuestras medidas se puede deber a que la medida de CARUV-S incluye no sólo flavonoides, sino también otros derivados fenólicos como los ácidos hidroxicinámicos. Dado que el DUALEX es un instrumento de diseño relativamente reciente, habría que hacer más estudios para

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Discusión
33
comprobar la relación que existe entre sus medidas de flavonoides y las obtenidas mediante otras técnicas. Por otra parte, se están diseñando otros instrumentos de este tipo que son capaces de evaluar también otros derivados fenólicos además de flavonoides. El hecho de que Flav. (DUALEX) no se correlacionase con CARUV-I puede ser debido a que los compuestos absorbentes de RUV localizados en la pared celular son principalmente ácidos hidroxicinámicos (ácido p-cumárico y ácido ferúlico) y tienen un máximo de absorción en torno a 320 nm, mientras que la medida de flavonoides con el DUALEX se basa en la fluorescencia emitida por la clorofila a 375 nm (Cerovic et al., 2002). Pfündel et al. (2007) ofrecen una explicación similar en su trabajo con la variedad Silvaner, ya que sus valores del DUALEX no se correlacionaban con los de otro fluorímetro utilizado (UV-A-PAM) debido a la presencia de antocianos, que tienen un máximo de absorción en el azul en torno a 500 nm.
Composición fenólica de la variedad Graciano La concentración y tipo de compuesto fenólico varían mucho en función del tejido u órgano vegetal de que se trate (Petit et al., 2009). Además, su síntesis parece ser selectiva en función de la radiación que incida sobre la planta. En hojas de Graciano se identificaron 2 ácidos hidroxicinámicos, 3 ácidos hidroxibenzoicos y 4 flavonoles, mientras que en hollejo fueron 3 ácidos hidroxicinámicos, 4 ácidos benzoicos, 5 flavanoles y 13 flavonoles (Tablas 4 y 5). Por tanto, el grupo de compuestos fenólicos más diverso en ambos órganos de esta variedad de vid es el de los flavonoles. Al separar las fracciones vacuolar y ligada a la pared celular, se encontró que en ésta había únicamente ácidos hidroxicinámicos: cumárico y siríngico en hollejos, y cumárico y ferúlico en hojas. En la fracción vacuolar se identificaron el resto de compuestos, incluidos ácidos hidroxicinámicos unidos a ácidos orgánicos para facilitar su solubilidad, como ocurre en otras plantas (Hause et al., 2002; Meissner et al., 2008; Fabón et al., 2010). Por lo tanto, la fracción vacuolar actúa como principal reservorio de compuestos fenólicos. Como compuestos mayoritarios destacaron en hoja el ácido cafeoil-tartárico y las quercetinas glucosiladas, y en hollejo la miricetina-3-O-glucósido, las mismas quercetinas que en hoja, el ácido cumárico y la epigallocatequina. La quercetina es más eficiente a la hora de eliminar los radicales libres que el kaempferol, debido a un mayor número de grupos hidroxilo, lo que dotaría de una mayor capacidad antioxidante tanto a las hojas como a los hollejos. Estos resultados concuerdan con los obtenidos por Berli et al. (2010, 2011), que llegan a cuantificar hasta 20 veces más quercetinas que kaempferoles en hojas y bayas de la variedad Malbec. En Graciano esta proporción se encuentra en torno a 10, pero estas diferencias pueden estar relacionadas con la variedad de vid. Nuestros resultados también se relacionan con el aumento en la expresión de genes implicados en la biosíntesis de flavonoles y fenilpropanoides en vid, que son activados por la RUV (Pontin et al., 2010). La composición fenólica depende en gran medida de la variedad de vid utilizada. Gómez et al. (2007) también observaron que los compuestos mayoritarios en hollejos de la variedad Cencibel

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Discusión
34
eran los flavonoles, aunque ellos encontraron como mayoritarios kaempferoles e isorhamnetina glucosilados. Por su parte, Koyama et al. (2012) encuentran concentraciones mucho más pequeñas de flavanoles en bayas de Cabernet-Sauvignon que las obtenidas en Graciano. Dentro de la misma variedad, otros factores que afectan a la composición fenólica son las condiciones meteorológicas de las diferentes campañas (Ferrer-Gallego et al., 2012). Estas condiciones podrían ser las responsables de las diferencias encontradas en las concentraciones de flavonoles en hollejo entre este estudio y el nuestro.
Variaciones en función del tiempo Se observaron diferencias significativas con respecto al tiempo (desde el envero hasta la vendimia) en todas las variables medidas en hoja, salvo en el cociente Clo a / Clo b. Por ejemplo, una variable de gran importancia fisiológica como Fv / Fm disminuyó de envero a vendimia, un proceso lógico ya que está relacionado con la senescencia, tanto en la vid en particular (Berli et al., 2012) como en las plantas en general (Maxwell y Johnson, 2000). Por su parte, el Índice de Esclerofilia (IE) fue significativamente mayor en vendimia que envero, concordando con los resultados obtenidos por Berli et al. (2012) en hojas de Malbec. Esto se debe a que las hojas se vuelven más esclerófilas con el tiempo, como un proceso natural relacionado con el desarrollo de la hoja y, en nuestro caso, también con la senescencia. Las variables relacionadas con el contenido de clorofilas (Clo a, Clo b, Clo a + b) tenían un valor menor en vendimia que en envero. Como en el caso de las variables anteriores, este descenso se relaciona con la senescencia en diversos tipos de plantas (Martínez-Abaigar y Núñez-Olivera, 1998). Berli et al. (2012) obtienen los mismos resultados en la variedad Malbec. En este sentido, resulta extraño que el cociente Clo a / Clo b no haya variado con el tiempo, ya que suele ser una variable indicativa de vitalidad fisiológica (Martínez-Abaigar y Núñez-Olivera, 1998) y hubiera sido esperable un descenso hacia la vendimia. Los carotenoides (tanto los totales como la mayor parte de los individuales) también mostraban la mínima concentración en vendimia, tal y como observan Berli et al. (2012) en hojas de Malbec y Schultz et al. (1998) en bayas de la variedad Riesling. Este descenso es lógico, ya que hacia la senescencia foliar se degradan los complejos pigmento-proteína de los fotosistemas, lo que incluye la degradación no sólo de las clorofilas sino también de los carotenoides, aunque éstos en menor medida (Martínez-Abaigar y Núñez-Olivera, 1998). Al comparar los valores obtenidos en envero y vendimia, se observa que tanto Flav. (DUALEX) como el contenido de CARUV (solubles e insolubles) y la concentración de los compuestos individuales aumentaron significativamente con el tiempo (P<0.001). Además, Flav (DUALEX), que se midió en dos momentos más entre envero y vendimia, aumentó desde envero hasta septiembre pero después se estabilizó hasta la vendimia. Se ha demostrado que el aumento de compuestos fenólicos en hojas de vid, como ocurre en las plantas en general, se produce porque la RUV activa la expresión de genes que codifican algunas enzimas de las vías de síntesis de fenilpropanoides y flavonoides, como PAL y CHS (Berli et al., 2012). Por lo tanto, los resultados descritos eran esperables y se asocian positivamente con la RUV (y posiblemente también con la PAR) recibida por las hojas, que se mantiene alta desde el envero hasta el equinoccio de

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Discusión
35
septiembre. Pollastrini et al. (2011) obtuvieron resultados similares en la variedad Sangiovese. Por otra parte, no se puede descartar la influencia de la fenología de la planta sobre los cambios temporales encontrados, ya que, como las hojas jóvenes tienen mayor actividad fotosintética, es más necesaria su protección que la de las hojas maduras próximas a la senescencia (Majer y Hideg, 2012), éstas, al ser más esclerófilas, tienen por tanto más protecciones estructurales.
Variaciones en función del tratamiento (régimen de radiación) El efecto que el tratamiento (régimen de radiación) provocó en las hojas fue relativamente ligero, ya que solamente se vieron afectadas significativamente unas pocas variables. Una de ellas fue Fv / Fm, pero los valores más bajos, que corresponden a una menor vitalidad fisiológica (Maxwell y Johnson, 2000), se encontraron en ausencia de RUV. Esto indicaría que la presencia de niveles naturales de RUV es necesaria para una adecuada fotoprotección y por lo tanto para evitar la caída de Fv / Fm. Estos resultados concuerdan con lo observado por Kolb et al. (2001), que encontraron que los cambios en Fv / Fm en hojas de la variedad Bacchus fueron pequeños y transitorios, en comparación con los producidos en la asimilación de CO2. Se sugiere así que, en condiciones de luz natural, la inhibición de la fotosíntesis por la RUV no se puede medir a través de Fv / Fm. Se ha demostrado que la luz natural estimula el efecto reparador del PSII y los compuestos pantalla (CARUV), que evitan el paso de la RUV hacia tejidos internos más sensibles. Ambos mecanismos evitan el daño provocado por la RUV en el PSII (Jansen et al., 1998), y por tanto ese daño no se refleja en Fv / Fm. En la misma línea apunta el trabajo de Pollastrini et al. (2011), al no encontrar tampoco diferencias significativas en Fv / Fm. Berli et al. (2013) dan un paso más allá sugiriendo que los complejos antena están protegidos contra el daño por RUV a través de un mecanismo eficaz para la disipación de energía. Por lo tanto, los niveles naturales de RUV-B afectarían a la fotosíntesis en cuanto al intercambio gaseoso y la conductancia estomática, pero no provocarían una alteración importante en el aparato fotoquímico. Esto encaja con lo observado por Núñez-Olivera et al. (2006), que sugieren que la vid se encuentra bien adaptada a las dosis naturales de RUV que recibe el cultivo actualmente. Parece entonces que Fv / Fm no es una variable muy adecuada para medir el efecto de la RUV sobre la vid en condiciones naturales. Pfündel et al. (2003) sugieren así que puede ser más ilustrativo el estudio de las variables F0 y Fm por separado, ya que sí se observan diferencias significativas entre regímenes de radiación (con RUV, Fm disminuye significativamente y Fo aumenta en menor medida). Sin embargo, si sometemos a la planta a un suplemento de RUV-B o incluso a dosis de RUV-C, los mecanismos de aclimatación y protección no son suficientes, y se provocan daños irreparables en el PSII, ocasionando así una disminución drástica de Fv / Fm (Pfündel et al., 2003; Majer y Hideg, 2012). El IE en hoja no mostró diferencias significativas entre tratamientos. La variación del espesor de la hoja se debe fundamentalmente a un aumento del grosor del mesófilo y la RUV refuerza este efecto morfogenético, como se ha encontrado en algunas variedades de vid como Sangiovese y Malbec (Pollastrini et al., 2011; Berli et al., 2012). Sin embargo, en Viura y Tempranillo no se encontraron diferencias en IE entre hojas expuestas a niveles naturales de RUV y hojas que no recibían RUV, aunque esto sucedía utilizando tiempos experimentales relativamente cortos (Núñez-Olivera et al., 2006). En nuestro caso, la ausencia de efectos de la RUV solar sobre el IE

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Discusión
36
se podría deber a que los filtros se colocaron en envero, cuando las hojas ya estaban bien desarrolladas. Ninguna variable relacionada con las clorofilas mostró diferencias significativas en hoja debidas al tratamiento. Estos resultados concuerdan con los obtenidos por Keller et al. (2003) en Cabernet Sauvignon y Chardonnay, Núñez-Olivera et al. (2006) en Viura, y Berli et al. (2010) en Malbec. Sin embargo, Núñez-Olivera et al. (2006) sí encontraron una disminución significativa en la concentración de clorofila (medida tanto con SPAD como mediante extracción en laboratorio) en la variedad Tempranillo. Una disminución significativa del contenido de clorofilas reduce la sobreexcitación de los complejos antena, lo que contribuiría a la protección del aparato fotosintético. De acuerdo con los resultados descritos, los cambios en la concentración de clorofilas pueden estar muy influenciados por la variedad de vid utilizada. Así, las variedades Cabernet Sauvignon, Chardonnay, Viura y Malbec, junto con Graciano, estarían mejor adaptadas en este aspecto a la RUV que Tempranillo, ya que sus clorofilas no se ven afectadas en condiciones normales de radiación. El hecho de que Berli et al. (2013), estudiando la misma variedad que Berli et al. (2010) (Malbec), sí que encontraran una disminución significativa en Clo a + b en hojas expuestas a RUV natural, puede ser debido a que el cultivo en el primer caso se encontraba a 1450 m de altitud y el segundo a sólo 900 m, y por tanto con menor RUV incidente. En nuestro trabajo tampoco se apreciaron diferencias significativas entre tratamientos en ninguna variable relacionada con los carotenoides (Tabla 4). Berli et al. (2010) y Núñez-Olivera et al. (2006) llegan a los mismos resultados en experimentos de exclusión de RUV realizados en otras variedades de vid (Malbec y Viura). Sin embargo, en Tempranillo sí se encontró un aumento significativo en el Índice de Xantofilas en presencia de RUV-B, lo que indica una mayor actividad del ciclo de las xantofilas y por tanto una mayor fotoprotección a través de la disipación del exceso de energía (Núñez-Olivera et al., 2006). Por lo tanto, al igual que ocurría con las clorofilas, existen diferencias intervarietales en las respuestas de los carotenoides a la exclusión de RUV. En experimentos con suplementos de RUV, es más habitual encontrar respuestas de los carotenoides (Majer y Hideg, 2012). Los derivados fenólicos confieren una mejor protección contra la RUV, ya que por una parte absorben esta radiación (más energética y por tanto más lesiva) y además protegen del estrés oxidativo, ya que poseen una estructura química capaz de eliminar los radicales libres, siendo eficaces antioxidantes. En nuestro trabajo, los derivados fenólicos en hojas de Graciano se midieron de una manera global, mediante la absorción del extracto metanólico en la región UV, y también de manera individual mediante HPLC. En ambos casos se analizaron las fracciones soluble e insoluble, lo cual constituye una novedad en vid, ya que hasta el momento solamente se había analizado la fracción soluble (ver por ejemplo Keller et al. (2003), Núñez-Olivera et al. (2006), Berli et al. (2010, 2013)). Esto es importante, ya que la distinta localización celular de ambas fracciones (la vacuolar para la fracción soluble y las paredes celulares para la insoluble) puede conferir distintos mecanismos de protección frente a la RUV. En las medidas globales de CARUV, no se observaron diferencias significativas en CARUV-S y CARUV-I entre los distintos regímenes de radiación en hojas de Graciano. Como se ha

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Discusión
37
mencionado anteriormente, en la bibliografía solamente existen datos comparativos para los CARUV-S. El efecto de la exclusión de RUV sobre CARUV-S depende de la variedad, ya que se ha encontrado un efecto significativo en Cabernet Sauvignon (Keller et al., 2003), Tempranillo y Viura (Núñez-Olivera et al., 2006), pero no en Chardonnay (Keller et al., 2003). Además, la falta de respuesta en Graciano se pudo deber a que las muestras se tomaron en la época de vendimia, en un estado de desarrollo muy avanzado, ya que la mayor síntesis de CARUV se produce después del envero, mientras que en la vendimia la mayoría de estos compuestos ya han sido sintetizados y la hoja tiene menos capacidad de respuesta a la RUV. En este sentido, Majer y Hideg (2012) sugieren que la capacidad de respuesta a la RUV es mayor en hojas jóvenes, que son las fotosintéticamente activas y necesitan proteger su aparato fotosintético, que en hojas próximas a la senescencia, que no muestran un aumento significativo del contenido en CARUV. En otros estudios de exclusión sí se detectaron aumentos significativos de CARUV-S en presencia de RUV (Núñez-Olivera et al., 2006; Kolb y Pfündel, 2005; Pollastrini et al., 2011; Koyama et al., 2012), y esta discrepancia con nuestro trabajo pudo deberse también al momento de recogida de las muestras. Por otra parte, la falta de respuesta de CARUV-S y CARUV-I en Graciano pudo deberse a que se encontraron diferentes respuestas en los distintos compuestos individuales analizados, que se pudieron compensar entre sí, con lo cual no se hicieron patentes las diferencias en la absorbancia global. En estudios de suplemento de RUV-B sí se aprecia más fácilmente un aumento significativo de la concentración de CARUV-S (Doupis et al., 2011). Esto también ocurre en los estudios con suplemento de UV-C (Bonomelli et al., 2004; González-Barrio et al., 2009; Pan et al., 2009; Petit et al., 2009), ya que la RUV-C estimula la expresión de genes que codifican algunas enzimas de la ruta de biosíntesis de los fenilpropanoides y la vía de los flavonoides, fundamentalmente estilbenos (Bonomelli et al., 2004). En el análisis de los compuestos individuales de hojas con respecto al régimen de radiación, sólo algunos de ellos mostraron diferencias significativas en sus concentraciones: el ácido p-cumárico, ácido p-cafeoil-tartárico, quercetina-3-O-glucurónido y cis kaempferol-3-O-glucósido. De ellos, los que muestran una respuesta más clara a la exclusión de RUV son el ácido p-cafeoil-tartárico y el cis kaempferol-3-O-glucósido (Figura 9). Por lo tanto, podría concluirse que estos dos compuestos juegan algún papel en la protección frente a la RUV. Los otros dos compuestos estarían influidos por algún factor adicional a la RUV. Nuestros resultados concuerdan con los de Kolb et al. (2001) y Berli et al. (2010), que señalan que los principales compuestos fenólicos relacionados con la protección epidérmica frente a la RUV son los ácidos hidroxicinámicos y los flavonoles. No obstante, dentro de estos dos grandes grupos de compuestos, cada compuesto individual puede mostrar una respuesta diferente debido a la influencia de otros factores, como la variedad, la etapa de desarrollo y el tipo de tejido. Se necesita un mayor número de estudios para extraer conclusiones más significativas acerca de la influencia de la exclusión de RUV sobre los compuestos fenólicos individuales en hojas de vid. En el caso del hollejo, los CARUV-S eran significativamente mayores en el tratamiento Ambiente que en los dos filtros (tanto UV+ como UV-), mientras que no había diferencias significativas entre tratamientos en CARUV-I. La mayor capacidad de respuesta en los CARUV-S parece lógica, ya que los CARUV-I estarían más inmovilizados que los CARUV-S por su localización en las paredes celulares, donde muchos compuestos se encuentran unidos covalentemente a los

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Discusión
38
polisacáridos de la pared. Además, los principales compuestos fenólicos presentes en las paredes estarían relacionados con funciones más estructurales que fotoprotectoras. Por otra parte, el mayor efecto de la exclusión de RUV sobre bayas que sobre hojas (donde no había respuesta ni de CARUV-S ni de CARUV-I) se deba a que los racimos estaban completamente cubiertos por los filtros, mientras que algunas hojas no lo estaban, y esto pudo enmascarar las diferencias. La mayor concentración de CARUV-S en el tratamiento Ambiente que en los otros dos tratamientos se podría deber a la influencia tanto de la RUV (que, aunque no muy marcadamente, era mayor en el tratamiento Ambiente) como a la temperatura, ya que bajo los filtros suelen alcanzarse temperaturas más altas y eso puede contribuir a la degradación de algunos compuestos (Berli et al., 2008). La falta de diferencia entre los tratamientos UV+ y UV- en nuestro estudio concuerda con lo encontrado por Berli et al. (2008) en la variedad Malbec, por lo que harían falta dosis más altas de UV solar para provocar un incremento significativo en compuestos globales. Los compuestos individuales del hollejo que mostraron diferencias significativas con respecto al tratamiento fueron aproximadamente la mitad (13 de 25): los ácidos protocatéquico y p-cumárico, un flavanol (catequina) y diez flavonoles (miricetinas glucosiladas, kaempferoles glucosilados, quercetinas glucosiladas, isorhamnetina-3-O-glucósido y siringetina-3-O-glucósido) (Tabla 5, Figuras 12-14). Claramente, los compuestos que respondían más fielmente a la RUV eran los flavonoles (10 compuestos de un total de 13). En general, las concentraciones de estos compuestos eran mayores en el tratamiento Ambiente que en los otros dos, y mayores en UV+ que en UV-, por lo que su aumento estaría ligado a la presencia de RUV. Esto concuerda con lo encontrado por Kolb et al. (2001) y Koyama et al. (2012) en las variedades Silvaner y Cabernet Sauvignon, respectivamente. Estos autores señalan que la síntesis de flavonoles está relacionada directamente con la RUV. La falta de respuesta de los flavanoles en nuestro estudio también resulta congruente con los hallazgos de Kolb et al. (2001) y Koyama et al. (2012). En este sentido, los flavanoles responderían más a la radiación visible que a la RUV. Con respecto a otros compuestos individuales del hollejo que respondían a la RUV, en el caso de ácido p-cumárico su comportamiento fue diferente en hollejo y en hoja, lo que sugiere que su síntesis no sólo está regulada por la RUV sino por algún factor adicional. Esto podría ser aplicable también al ácido protocatéquico, que mostraba mayores concentraciones en el tratamiento Ambiente que en los otros dos. Algunos resultados irregulares hallados en nuestro estudio, como el comportamiento del kaempferol-3-O-galactósido (concentración máxima en UV+) o la quercetina-3-O-rutinósido (concentración mínima en UV+), pudieron deberse a las pequeñas concentraciones en las que se encontraban. En definitiva, los flavonoles se asocian específicamente con la RUV (Kolb et al., 2001; Koyama et al., 2012; nuestro estudio), mientras que los ácidos hidroxicinámicos y los flavanoles se relacionan más con la radiación visible y probablemente otros factores. Resulta llamativo que el resveratrol era uno de los compuestos individuales del hollejo que no respondieron a la exclusión de RUV. Esto se contrapone a los datos de varios autores, que encontraron aumentos de este compuesto en bayas en presencia de RUV (Bonomelli et al., 2004; Berli et al., 2008; González-Barrio et al., 2009; Pan et al., 2009; Petit et al., 2009). Posiblemente, el resveratrol responde más a RUV-C (en estudios de laboratorio) que a niveles

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Discusión
39
naturales de otras bandas de RUV, ya que se trata de una fitoalexina que responde a numerosos factores ambientales adversos, así como a ciertas enfermedades (Bonomelli et al., 2004). Una consecuencia importante de nuestro estudio es que, dado que los flavonoles son muy importantes en la elaboración del vino ya que contribuyen a la estabilización del color, hemos puesto de relieve que la manipulación de la RUV es un factor crucial para su acumulación en bayas de Graciano en la época de vendimia.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Conclusiones
40
Conclusiones 1. Se han obtenido correlaciones muy significativas entre las técnicas destructivas y no destructivas utilizadas, en condiciones de campo, para medir algunas variables de gran importancia fisiológica, como el rendimiento cuántico máximo del fotosistema II (Fv / Fm), las concentraciones de clorofila y los niveles globales de compuestos absorbentes de radiación UV (CARUV). Por lo tanto, las técnicas no destructivas suponen una alternativa rápida, fácil y repetitiva con respecto a los métodos destructivos.
2. Por primera vez en vid, los CARUV se midieron, tanto en hojas como en hollejos, distinguiendo su localización en las vacuolas y en las paredes celulares. Dado que la primera fue más reactiva a la RUV que la segunda, sus funciones serían diferentes (respectivamente, protección frente a la RUV y funciones estructurales).
3. Los cambios temporales registrados en las hojas desde envero a vendimia fueron muy acusados y estaban determinados principalmente por la senescencia foliar, ya que se produjeron descensos de Fv / Fm y pigmentos fotosintéticos, y aumentos del Índice de Esclerofilia (IE). No obstante, algunos cambios temporales, como el aumento en CARUV, también estaban ligados a factores ambientales como la radiación UV y fotosintética. 4. La exclusión de la RUV solar influyó sólo débilmente sobre la fisiología de las hojas, y no se produjo ninguna alteración de las variables indicativas de vitalidad, como Fv / Fm y la concentración de clorofila. Por lo tanto, la variedad Graciano se encuentra bien adaptada a los niveles de RUV a los que está expuesta actualmente de forma natural. 5. La composición fenólica de los hollejos respondió muy significativamente a la exclusión de RUV, con descensos de 13 de los 25 CARUV individuales analizados, todos ellos presentes en la fracción soluble. En particular, los compuestos más reactivos fueron los flavonoles, mientras que los flavanoles no respondieron al tratamiento. 6. Con este estudio se ha puesto de relieve que la RUV, incluso en los niveles actuales recibidos en la Biosfera, influye decisivamente sobre la composición fenólica de las bayas de vid y por lo tanto sobre las características del vino resultante. Incluso, se podrían potenciar las respuestas de la planta si se excluyera la RUV desde estados fenológicos anteriores. Todo ello aconseja continuar estudiando las potencialidades que tiene la manipulación de la RUV en vitivinicultura.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Agradecimientos
41
Agradecimientos Al Ministerio de Ciencia e Innovación y FEDER (Proyecto CGL2011-26977 del Plan Nacional de I+D+i) por la financiación de este estudio. En especial a mis directores, Encarna y Javier, sin su esfuerzo, sin sus consejos, sin su apoyo, este trabajo nunca hubiera visto la luz. A Rafa, por su ayuda técnica y consejos prácticos. A mis compañeros de grupo de investigación: Gabi, Laura y Gonzalo, sin ellos todo hubiera sido mucho más difícil. A Javier Tardáguila y Mª Paz Diago, por su estrecha colaboración en la realización de este estudio. A Fernando Martínez de Toda por su aportación de conocimientos sobre Graciano. A Ernesto y Nines por su paciencia y buen hacer. Por compartir su pasión por la espectrometría de masas. Por último, a las personas que desinteresadamente y con toda la predisposición posible, han ayudado a que la costosa labor experimental, tanto de campo como de laboratorio, fuera mucho más llevadera. Gracias a Juanan, María e Inma.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Referencias bibliográficas
42
Referencias bibliográficas Arróniz-Crespo M, Núñez-Olivera E, Martínez-Abaigar J, Becker H, Scher J, Zapp J, Tomás
R, Beaucourt N. 2006. Physiological changes and UV protection in the aquatic liverwort Jungermannia exsertifolia subsp. cordifolia along an altitudinal gradient of UV-B radiation. Functional Plant Biology 33: 1025-1036.
Berli F, D'Angelo J, Cavagnaro B, Bottini R, Wuilloud R, Silva MF. 2008. Phenolic composition in grape (Vitis vinifera L. cv. Malbec) ripened with different solar UV-B radiation levels by capillary zone electrophoresis. Journal of Agricultural and Food Chemistry 56: 2892-2898.
Berli FJ, Alonso R, Bressan-Smith R, Bottini R. 2013. UV-B impairs growth and gas exchange in grapevines grown in high altitude. Physiologia Plantarum (en prensa; doi: 10.1111/ppl.12012).
Berli FJ, Fanzone M, Piccoli P, Bottini R. 2011. Solar UV-B and ABA Are Involved in Phenol Metabolism of Vitis vinifera L. Increasing Biosynthesis of Berry Skin Polyphenols. Journal of Agricultural and Food Chemistry 59: 4874-4884.
Berli FJ, Moreno D, Piccoli P, Hespanhol-Viana L, Silva MF, Bressan-Smith R, Cavagnaro JB, Bottini R. 2010. Abscisic acid is involved in the response of grape (Vitis vinifera L.) cv. Malbec leaf tissues to ultraviolet-B radiation by enhancing ultraviolet-absorbing compounds, antioxidant enzymes and membrane sterols. Plant, Cell and Environment 33: 1-10.
Bonomelli A, Mercier L, Franchel K, Baillieul F, Benizri E, Mauro MC. 2004. Response of grapevine defenses to UV-C exposure. American Journal of Enology and Viticulture 55: 51-59.
Cabello Sáenz de Santamaría J, Ortiz Marcide JM, Muñoz Organero G, Rodríguez Torres I, Benito Barba A, Rubio de Miguel C, García Muñoz S, Sáiz Sáiz R. 2012. Variedades de vid en España. Madrid: Editorial Agrícola Española.
Caldwell MM, Bornman JF, Ballaré CL, Flint SD, Kulandaivelu G. 2007. Terrestrial ecosystems, increased solar ultraviolet radiation, and interactions with other climate change factors. Photochemical and Photobiological Sciences 6: 252-266.
Cerovic ZG, Masdoumier G, Ghozlen NB, Latouche G. 2012. A new optical leaf-clip meter for simultaneous non-destructive assessment of leaf chlorophyll and epidermal flavonoids. Physiologia Plantarum 146: 251-260.
Cerovic ZG, Ounis A, Cartelat A, Latouche G, Goulas Y, Meyer S, Moya I. 2002. The use of chlorophyll fluorescence excitation spectra for the non-destructive in situ assessment of UV-absorbing compounds in leaves. Plant, Cell and Environment 25: 1663-1676.
Clarke LJ, Robinson SA. 2008. Cell wall-bound ultraviolet-screening compounds explain the high ultraviolet tolerance of the Antarctic moss, Ceratodon purpureus. New Phytologist 179: 776-783.
Cockell CS, Knowland J. 1999. Ultraviolet radiation screening compounds. Biological Review 74: 311-345.
Day TA, Neale PJ. 2002. Effects of UV-B radiation on terrestrial and aquatic primary producers. Annual Review of Ecology and Systematics 33: 371-396.
Day TA, Vogelmann TC. 1995. Alterations in photosynthesis and pigment distributions in pea

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Referencias bibliográficas
43
leaves following UV-B exposure. Physiologia Plantarum 94: 433-440. Doupis G, Chartzoulakis K, Beis A, Patakas A. 2011. Allometric and biochemical responses of
grapevines subjected to drought and enhanced ultraviolet-B radiation. Australian Journal of Grape and Wine Research 17: 36-42.
Fabón G. 2012. Ultraviolet radiation and aquatic bryophytes: physiological damage and protection mechanisms under laboratory and field conditions. Tesis Doctoral, Universidad de La Rioja.
Fabón G, Martínez-Abaigar J, Tomás R, Núñez-Olivera E. 2010. Effects of enhanced UV-B radiation on hydroxycinnamic acid derivatives extracted from different cell compartments in the aquatic liverwort Jungermannia exsertifolia subsp. cordifolia. Physiologia Plantarum 140: 269-279.
Fernández Marín B. 2012. The dark side of violaxanthin cycle. Tesis Doctoral, Universidad del País Vasco.
Ferrer-Gallego R, Hernández-Hierro JM, Rivas-Gonzalo JC, Escribano-Bailón MT. 2012. Influence of climatic conditions on the phenolic composition of Vitis vinifera L. cv. Graciano. Analytica Chimica Acta 732: 73-77.
Flint SD, Caldwell MM. 2003. A biological spectral weighting function for ozone depletion research with higher plants. Physiologia Plantarum 117: 137-144.
García-Plazaola JI, Becerril JM. 2001. Seasonal changes in photosynthetic pigments and antioxidants in beech (Fagus sylvatica) in a Mediterranean climate: implications for tree decline diagnosis. Australian Journal of Plant Physiology 28: 225-232.
Gómez I, Orostegui M, Huovinen P. 2007. Morpho-functional patterns of photosynthesis in the south pacific kelp Lessonia nigrescens: Effects of UV radiation on C-14 fixation and primary photochemical reactions. Journal of Phycology 43: 55-64.
González-Barrio R, Vidal-Guevara ML, Tomás-Barberán FA, Espín JC. 2009. Preparation of a resveratrol-enriched grape juice based on ultraviolet C-treated berries. Innovative Food Science and Emerging Technologies 10: 374-382.
Guerrero RF, Puertas B, Fernández MI, Palma M, Cantos-Villar E. 2010. Induction of stilbenes in grapes by UV-C: Comparison of different subspecies of Vitis. Innovative Food Science and Emerging Technologies 11: 231-238.
Häder DP, Lebert M, Schuster M, Del Campo L, Helbling EW, McKenzie R. 2007. ELDONET - A decade of monitoring solar radiation on five continents. Photochemistry and Photobiology 83: 1348-1357.
Hause B, Meyer K, Viitanen PV, Chapple C, Strack D. 2002. Immunolocalization of 1-O -sinapoylglucose:malate sinapoyltransferase in Arabidopsis thaliana. Planta 215: 26-32.
Jansen MAK, Gaba V, Greenberg BM. 1998. Higher plants and UV-B radiation: balancing damage, repair and acclimation. Trends in Plant Science 3: 131-135.
Kakani VG, Reddy KR, Zhao D, Sailaja K. 2003. Field crop responses to ultraviolet-B radiation: a review. Agricultural and Forest Meteorology 120: 191-218.
Keller M, Rogiers SY, Schultz HR. 2003. Nitrogen and ultraviolet radiation modify grapevines' susceptibility to powdery mildew. Vitis 42: 87-94.
Kolb CA, Käser MA, Kopecky J, Zotz G, Riederer M, Pfündel EE. 2001. Effects of natural intensities of visible and ultraviolet radiation on epidermal ultraviolet screening and photosynthesis in grape leaves. Plant Physiology 127: 863-875.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Referencias bibliográficas
44
Kolb CA, Kopecky J, Riederer M, Pfündel EE. 2003. UV screening by phenolics in berries of grapevine (Vitis vinifera). Functional Plant Biology 30: 1177-1186.
Kolb CA, Pfündel EE. 2005. Origins of non-linear and dissimilar relationships between epidermal UV absorbance and UV absorbance of extracted phenolics in leaves of grapevine and barley. Plant, Cell and Environment 28: 580-590.
Kolb CA, Schreiber U, Gademann R, Pfündel EE. 2005. UV-A screening in plants determined using a new portable fluorimeter. Photosynthetica 43: 371-377.
Koyama K, Ikeda H, Poudel PR, Goto-Yamamoto N. 2012. Light quality affects flavonoid biosynthesis in young berries of Cabernet Sauvignon grape. Phytochemistry 78: 54-64.
Liakopoulos G, Nikolopoulos D, Klouvatou A, Vekkos KA, Manetas Y, Karabourniotis G. 2006. The photoprotective role of epidermal anthocyanins and surface pubescence in young leaves of grapevine (Vitis vinifera). Annals of Botany 98: 257-265.
Lichtenthaler HK. 1987. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods in Enzymology 148: 350-382.
Lijavetzky D, Carbonell-Bejerano P, Grimplet J, Bravo G, Flores P, Fenoll J, Hellín P, Oliveros JC, Martínez-Zapater JM. 2012. Berry Flesh and Skin Ripening Features in Vitis vinifera as Assessed by Transcriptional Profiling. PLoS ONE 7: e39547.
Majer P, Hideg E. 2012. Developmental stage is an important factor that determines the antioxidant responses of young and old grapevine leaves under UV irradiation in a green-house. Plant Physiology and Biochemistry 50: 15-23.
Martínez-Abaigar J, Núñez-Olivera E. 1998. Ecophysiology of photosynthetic pigments in aquatic bryophytes. En: Bates JW, Ashton NW, Duckett JG (eds.), Bryology for the Twenty-first Century. Leeds: Maney Publishing and the British Bryological Society, 277-292.
Maxwell K, Johnson GN. 2000. Chlorophyll fluorescence - a practical guide. Journal of Experimental Botany 51: 659-668.
McKenzie RL, Aucamp PJ, Bais AF, Björn LO, Hyas M. 2007. Changes in biologically-active ultraviolet radiation reaching the Earth's surface. Photochemical and Photobiological Sciences 6: 218-231.
McKenzie RL, Björn LO, Bais A, Ilyasd M. 2003. Changes in biologically active ultraviolet radiation reaching the Earth's surface. Photochemical and Photobiological Sciences 2: 5-15.
Meissner D, Albert A, Bottcher C, Strack D, Milkowski C. 2008. The role of UDP-glucose : hydroxycinnamate glucosyltransferases in phenylpropanoid metabolism and the response to UV-B radiation in Arabidopsis thaliana. Planta 228: 663-674.
Morales LO, Tegelberg R, Brosche M, Keinanen M, Lindfors A, Aphalo PJ. 2010. Effects of solar UV-A and UV-B radiation on gene expression and phenolic accumulation in Betula pendula leaves. Tree Physiology 30: 923-934.
Núñez-Olivera E, Martínez-Abaigar J, Tomás R, Otero S, Arróniz-Crespo M. 2006. Physiological effects of solar ultraviolet-B exclusion on two cultivars of Vitis vinifera L. from La Rioja, Spain. American Journal of Enology and Viticulture 57: 441-448.
Otero S, Núñez-Olivera E, Martínez-Abaigar J, Tomás R, Arróniz-Crespo M, Beaucourt N. 2006. Effects of cadmium and enhanced UV radiation on the physiology and the concentration of UV-absorbing compounds of the aquatic liverwort Jungermannia

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Referencias bibliográficas
45
exsertifolia subsp. cordifolia. Photochemical and Photobiological Sciences 5: 760-769. Pan QH, Wang L, Li JM. 2009. Amounts and subcellular localization of stilbene synthase in
response of grape berries to UV irradiation. Plant Science 176: 360-366. Petit AN, Baillieul F, Vaillant-Gaveau N, Jacquens L, Conreux A, Jeandet P, Clement C,
Fontaine F. 2009. Low responsiveness of grapevine flowers and berries at fruit set to UV-C irradiation. Journal of Experimental Botany 60: 1155-1162.
Pfündel EE. 2003. Action of UV and visible radiation on chlorophyll fluorescence from dark-adapted grape leaves (Vitis vinifera L.). Photosynthesis Research 75: 29-39.
Pfündel EE, Ben Ghozlen N, Meyer S, Cerovic ZG. 2007. Investigating UV screening in leaves by two different types of portable UV fluorimeters reveals in vivo screening by anthocyanins and carotenoids. Photosynthesis Research 93: 205-221.
Pollastrini M, Di Stefano V, Ferretti M, Agati G, Grifoni D, Zipoli G, Orlandini S, Bussotti F. 2011. Influence of different light intensity regimes on leaf features of Vitis vinifera L. in ultraviolet radiation filtered condition. Environmental and Experimental Botany 73: 108-115.
Pontin MA, Piccoli PN, Francisco R, Bottini R, Martinez-Zapater JM, Lijavetzky D. 2010. Transcriptome changes in grapevine (Vitis vinifera L.) cv. Malbec leaves induced by ultraviolet-B radiation. BMC Plant Biology 10: Art. No. 224.
Prado FE, Rosa M, Prado C, Podazza G, Interdonato R, González JA, Hilal M. 2012. UV-B Radiation Its effects and Defense Mechanism in Terrestrial Plants. En: Ahmad P, Prasad MNV (eds.), Environmental Adaptations and Stress Tolerance of Plants in the Era of Climate Change. New York: Springer, 57-83.
Rizzini L, Favory JJ, Cloix C, Faggionato D, O'Hara A, Kaiserli E, Baumeister R, Schafer E, Nagy F, Jenkins GI, Ulm R. 2011. Perception of UV-B by the Arabidopsis UVR8 Protein. Science 332: 103-106.
Sáenz-Navajas MP, Ferreira V, Dizy M, Fernández-Zurbano P. 2010. Characterization of taste-active fractions in red wine combining HPLC fractionation, sensory analysis and ultra performance liquid chromatography coupled with mass spectrometry detection. Analytica Chimica Acta 673: 151-159.
Schmitz-Hoerner R, Weissenböck G. 2003. Contribution of phenolic compounds to the UV-B screening capacity of developing barley primary leaves in relation to DNA damage and repair under elevated UV-B levels. Phytochemistry 64: 243-255.
Schreiber U, Bilger W, Neubauer C. 1995. Chlorophyll fluorescence as a nonintrusive indicator for rapid assessment of in vivo photosynthesis. En: Schulze ED, Caldwell MM (eds.), Ecophysiology of photosynthesis. Berlin: Springer, 49-70.
Schultz HR, Löhnertz O, Bettner W, Bálo B, Linsenmeier A, Jähnisch A, Müller M, Gaubatz B, Váradi G. 1998. Is grape composition affected by current levels of UV-B radiation? Vitis 37: 191-192.
Searles PS, Flint SD, Caldwell MM. 2001. A meta-analysis of plant field studies simulating stratospheric ozone depletion. Oecologia 127: 1-10.
Seckmeyer G, Pissulla D, Glandorf M, Henriques D, Johnsen B, Webb A, Siani AM, Bais A, Kjeldstad B, Brogniez C, Lenoble J, Gardiner B, Kirsch P, Koskela T, Kaurola J, Uhlmann B, Slaper H, Den Outer P, Janouch M, Werle P, Grobner J, Mayer B, De la Casiniere A, Simic S, Carvalho F. 2008. Variability of UV irradiance in Europe. Photochemistry and Photobiology 84: 172-179.

Efectos de la exclusión de radiación ultravioleta sobre hojas y hollejos Referencias bibliográficas
46
Steel CC, Greer LA. 2005. Catalase activity and susceptibility of grapevine callus culture (cv. Cabernet Sauvignon) to Botrytis cinerea infection: Effects of UV-B exposure. Vitis 44: 149-150.
Steel CC, Keller M. 2000. Influence of UV-B irradiation on the carotenoid content of Vitis vinifera tissues. Biochemical Society Transactions 28: 883-885.
Strasser RJ, Srivastava A, Tsimilli-Michael M. 2000. The fluorescence transient as a tool to characterize and screen photosynthetic samples. En: Yunus M, Pathre U, Mohanty P (eds.), Probing photosynthesis: mechanisms, regulation and adaptation. London: Taylor & Francis, 445-483.
Suberviola Ripa J. 1998. Aspectos enológicos de la variedad Graciano en Navarra. En: Gobierno de La Rioja (ed.), Viticultura y enología de la variedad Graciano. Logroño: Gobierno de La Rioja, 95-123.
Teramura AH, Sullivan JH. 1994. Effects of UV-B radiation on photosynthesis and growth of terrestrial plants. Photosynthesis Research 39: 463-473.
Wang W, Tang K, Yang HR, Wen PF, Zhang P, Wang HL, Huang WD. 2010. Distribution of resveratrol and stilbene synthase in young grape plants (Vitis vinifera L. cv. Cabernet Sauvignon) and the effect of UV-C on its accumulation. Plant Physiology and Biochemistry 48: 142-152.
Zhang ZZ, Che XN, Pan QH, Li XX, Duan CQ. 2013. Transcriptional activation of flavan-3-ols biosynthesis in grape berries by UV irradiation depending on developmental stage. Plant Science 208: 64- 74.
Zhang ZZ, Li XX, Chu YN, Zhang MX, Wen YQ, Duan CQ, Pan QH. 2012. Three types of ultraviolet irradiation differentially promote expression of shikimate pathway genes and production of anthocyanins in grape berries. Plant Physiology and Biochemistry 57: 74-83.