
Manual QIR
VOLUMEN III: BIOQUÍMICA GENERAL Y
CLÍNICA
Autoras: María del Carmen Enjo Mallou
Hélade Sotomayor Pérez
Dra. Idalmys Perdomo López
Editora: Dra. Iliana Perdomo López

Manual QIR. VOLUMEN III: BIOQUÍMICA GENERAL
Y CLÍNICA
Autoras: María del Carmen Enjo Mallou Hélade Sotomayor Pérez Dra. Idalmys Perdomo López
© Iliana Perdomo López (editora). Depósito legal: C 610-2012
Reservado todos los derechos. Está prohibido, bajo las sanciones penales y el resarcimiento civil previsto en las leyes, reproducir, registrar o transmitir esta publicación, íntegra o parcialmente por cualquier sistema de recuperación y por cualquier medio, sea mecánico, electrónico, magnético, por fotocopia o por cualquier otro, sin la autorización previa por escrito de la editora.

ÍNDICE BIOQUÍMICA GENERAL Y CLÍNICA
UNIDAD I: BIOQUÍMICA GENERAL 1
1.- INTRODUCCIÓN. COMPOSICIÓN DE LOS SERES VIVOS 1
2.- GLÚCIDOS 7
2.1- OSAS O MONOSACÁRIDOS 8
2.1.1.- ACTIVIDAD ÓPTICA 8
2.1.2.- ESTRUCTURA CÍCLICA DE LOS MONOSACÁRIDOS 11
2. 1.3.- ISÓMEROS CONFORMACIONALES 14
2. 1.4.- PROPIEDADES GENERALES DE LAS OSAS 14
2. 1.5.- DERIVADOS DE MONOSACÁRIDOS 16
2.2- OLIGOSACÁRIDOS: DISACÁRIDOS 18
2.2.1.-CARACTERÍSTICAS DEL ENLACE GLUCOSÍDICO 19
2.2.2.-TIPOS DE ENLACE GLUCOSÍDICO 19
2.2.3.-OLIGOSACÁRIDOS MÁS IMPORTANTES 19
2.3- POLISACARIDOS 20
2.3.1.- HOMOPOLISACÁRIDOS 20
2.3.2.- HETEROPOLISACÁRIDOS 22
2.4- HETERÓSIDOS 26
2.4.1.- PROTEOGLUCANOS 26
2.4.2.- GLUCOPROTEÍNAS 28
2.4.3.- OTROS HETERÓSIDOS 29
3.- LÍPIDOS 31
3.1- LÍPIDOS SAPONIFICABLES 32
3.1.1.- LÍPIDOS SAPONOFICABLES SIMPLES 32
3.1.2.- LÍPIDOS SAPONOFICABLES COMPLEJOS 40
3.2- LÍPIDOS INSAPONIFICABLES 46
3.2.1.- TERPENOS 47
3.2.2.-ESTEROIDES 47
4.- AMINOÁCIDOS, PÉPTIDOS Y PROTEINAS 49
4.1- AMINOÁCIDOS 49
4.1.1.- CARÁCTERISTICAS ESTRUCTURALES 49
4.1.2.- ESTEREOQUIMICA DE LOS AMINOACIDOS 49
4.1.3.- CLASIFICACIÓN 50
4.1.4.- PROPIEDADES FISICAS 52
4.1.5.- IONIZACIÓN 53
4.1.6.- REACCIONES DE IDENTIFICACION DE AMINOACIDOS 54

4.2- PÉPTIDOS 55
4.2.1.- EL ENLACE PEPTIDICO 55
4.2.2.- CARACTERISTICAS DEL ENLACE PEPTÍDICO 55
4.2.3.- CARACTERIZACIÓN DE PÉPTIDOS Y PROTEINAS 56
4.2.4.- PÉPTIDOS DE IMPORTANCIA BIOLÓGICA 57
4.3- PROTEÍNAS 57
4.3.1.- CLASIFICACIÓN DE LAS PROTEÍNAS 58
4.3.1.1. ATENDIENDO A SU COMPOSICIÓN 58
4.3.1.2. ATENDIENDO A SU FORMA 59
4.3.1.3. ATENDIENDO A SU SOLUBILIDAD 59
4.3.1.4. ATENDIENDO A SU FUNCION BIOLÓGICA 59
4.3.2.- ESTRUCTURA DE LAS PROTEÍNAS 60
4.3.2.1. ESTRUCTURA PRIMARIA 61
4.3.2.2. ESTRUCTURA SECUNDARIA 61
4.3.2.3. ESTRUCTURA TERCIARIA 65
4.3.2.4. ESTRUCTURA CUATERNARIA 66
4.4- ENZIMAS 77
5.- COMPOSICIÓN, ESTRUCTURA Y FUNCIONES DE LAS BIOMEMBRANAS 91
5.1- LÍPIDOS DE MEMBRANAS 92
5.2- PROTEÍNAS DE MEMBRANAS 97
5.3- TRANSPORTES A TRAVÉS DE MEMBRANAS 98
6.- INTRODUCCIÓN AL METABOLISMO. 105
7.- GLUCOLISIS 113
8.- FERMENTACIONES 125
9.- DESCARBOXILACIÓN OXIDATIVA DEL PIRUVATO 129
10.- CICLO DE KREBS 133
11.- CADENA DE TRANSPORTE ELECTRÓNICO Y FOSFORILACIÓN OXIDATIVA 139
12.- LANZADERAS 153
13.- GLUCONEOGÉNESIS 155
14.- METABOLISMO DEL GLUCÓGENO 167

15.- OTRAS RUTAS DE OXIDACIÓN DE LA GLUCOSA 181
16.- OXIDACIÓN DE ÁCIDOS GRASOS 191
17.- BIOSÍNTESIS DE LÍPIDOS 205
18.- METABOLISMO DE CUERPOS CETONICOS 221
19.- BIOSÍNTESIS DE COLESTEROL 225
20.- METABOLISMO DE LAS LIPOPROTEÍNAS 231
21.- METABOLISMO DE AMINOÁCIDOS 243
22.- METABOLISMO DE NUCLEÓTIDOS 261
23.- INTEGRACIÓN DEL METABOLISMO 275
UNIDAD II: BIOQUÍMICA CLÍNICA 285
24.- BIOQUÍMICA CLÍNICA 285
GENERALIDADES DE ANÁLISIS CLÍNICOS 285
SANGRE 289
HECES 299
ORINA 299
LÍQUIDOS ESPECIALES 301
HORMONAS 304
INFARTO AGUDO DEL MIOCARDIO 306
PRINCIPALES REACTANTES DE FASE AGUDA 307
MARCADORES TUMORALES 308
DIAGNÓSTICO DE LABORATORIO DE ENFERMEDADES AUTOINMUNES 309
ANEXOS 311
BIBLIOGRAFÍA 315

UNIDAD I
BIOQUÍMICA GENERAL

Bioquímica InspiracleQIR/2015
1
TEMA 1. COMPOSICIÓN DE LOS SERES VIVOS. [p. 206 (2012)]
Todos los seres vivos están constituidos, cualitativa y cuantitativamente por los mismos
elementos químicos, aproximadamente 30, lo que confirma la idea de que la vida se ha
desarrollado sobre unos elementos concretos que poseen unas propiedades físico-químicas
acordes con los procesos químicos que se desarrollan como parte de ésta.
Se denominan bioelementos o elementos biogénicos aquellos elementos químicos que
forman parte de los seres vivos. Atendiendo a su abundancia (no importancia) se pueden
agrupar en tres categorías: mayoritarios primarios, mayoritarios secundarios y
oligoelementos.
Bioelementos mayoritarios primarios o principales: C, H, N, O.
Son los elementos más abundantes de la materia viva (constituyen más del 95 % de la masa
total). Éstos son idóneos por sus propiedades físico–químicas, ya que:
1. Pueden compartir electrones entre ellos para completar sus capas electrónicas
externas y formar enlaces. El hidrógeno necesita solamente 1 e-, el oxígeno 2 e-, el
nitrógeno 3 e- y el carbono 4 e-.
2. Son los elementos más ligeros con capacidad de formar enlaces estables:
particularmente significativa es la capacidad del carbono para formar enlaces
covalentes C-C de gran estabilidad (348 KJ/mol).
3. Carbono, nitrógeno y oxígeno pueden compartir más de un par de electrones,
formando incluso enlaces dobles y triples, lo cual les dota de una gran versatilidad
para la formación de otras estructuras químicas (cadenas más o menos largas,
lineales, ramificadas, anillos…).
4. A causa de la configuración tetraédrica de los enlaces del carbono, los diferentes
tipos de moléculas orgánicas adoptan estructuras tridimensionales en muchos
casos biológicamente activas.
5. Las combinaciones del carbono con otros elementos químicos permite la aparición
de diversos grupos funcionales que dan lugar a las diferentes biomoléculas que
conforman a los seres vivos.
Bioquímica InspiracleQIR/2015
7
TEMA 2. GLÚCIDOS. [p. 157, 228, 229 (2010); 139, 140, 170, 183, 239 (2011); 112, 140,
175, 189, 205 (2012); 148 (2013)].
Los glúcidos son moléculas orgánicas caracterizadas por la presencia de cadenas
carbonadas portadoras de grupos hidroxilos y funciones aldehídicas, cetónicas, ácidas o
aminadas.
DISTRIBUCIÓN EN LA NATURALEZA
Son compuestos naturales ampliamente distribuidos:
a) Como elementos estructurales; confiriendo resistencia
celulosa; pared celular de los vegetales
quitina: exoesqueleto de invertebrados
polisacáridos de la pared bacteriana
b) Como reserva energética
almidón, reserva vegetal
glucógeno, reserva animal
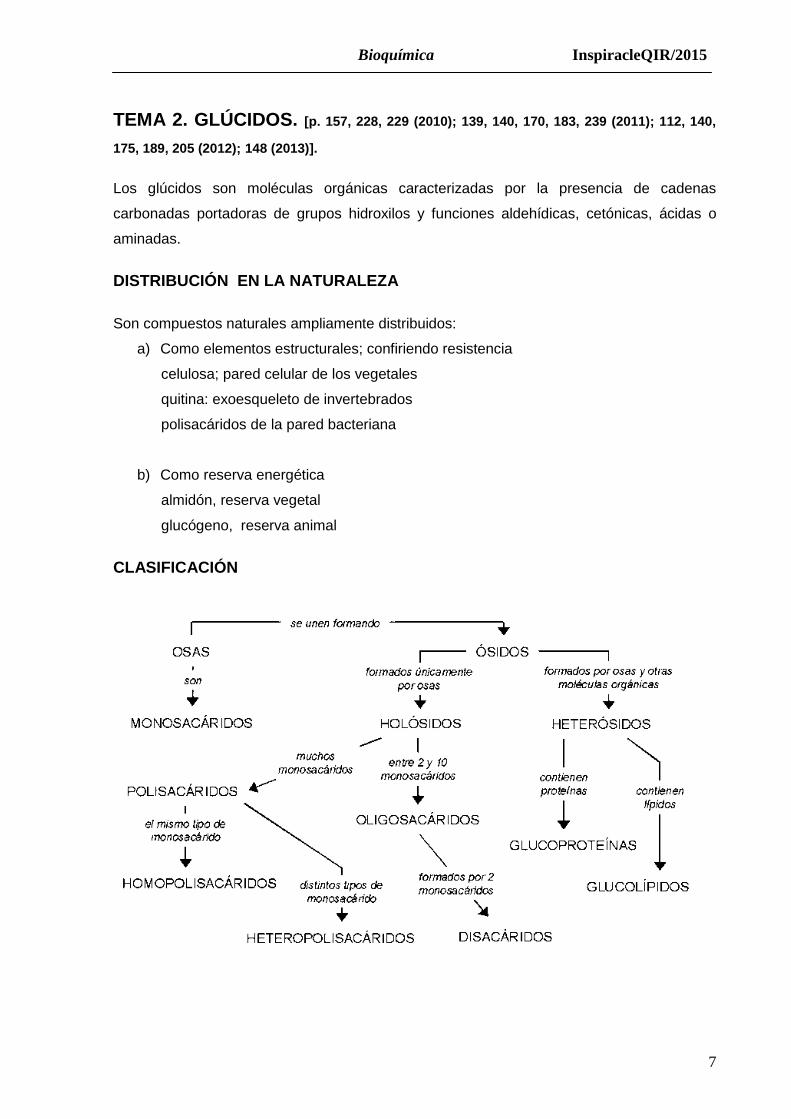
CLASIFICACIÓN

InspiracleQIR/2015 Bioquímica
10
En el caso de las cetosas, para un nº igual de átomos de carbono, comparándolas con sus
aldosas correspondientes, tienen un átomo de carbono menos sustituido asimétricamente (el
carbono que porta la función cetónica no es un carbono asimétrico). Así, tomando como
ejemplo una cetotetrosa, observamos que tiene dos isómeros (21), uno de la serie D y otro
de la serie L, frente a los 4 que apreciábamos en el caso de las aldotetrosas, pues tiene sólo
un carbono asimétrico.
Fuente: Fig 7-3 (b) (pag 241) del libro “Lehninger. Principios de Bioquímica. Ediciones Omega, Barcelona, 4ª ed., 2006”.
La mayor parte de las osas naturales pertenecen a la serie D
*TIPOS DE ESTEREOISÓMEROS [p. 259 (2004); 131 (2008)]
ENANTIÓMEROS: se definen como imágenes especulares no superponibles, son las parejas
D y L; así, D-glucosa y L-glucosa son enantiómeros. La mezcla equimolecular de
compuestos enantiómeros se denomina mezcla racémica
DIASTEREOISÓMEROS: la configuración de los grupos hidroxilo es diferente en un carbono o
más, aquí se incluyen:
Epímeros, si sólo se diferencian en la configuración del hidroxilo de un C asimétrico, por ejemplo, la
D-Glucosa y la D-Manosa son 2-epímeros, la D-Glucosa y la D-Galactosa son 4-epímeros.
Anómeros: difieren en la configuración entorno al carbono anomérico

InspiracleQIR/2015 Bioquímica
24
negativamente. Los glucosaminoglucanos están presentes en la matriz extracelular de los
tejidos de animales multicelulares, están interconectados con proteínas fibrosas (elastina, la
fibronectina y la minina).[p. 9 (2007); p. 224 (2009)]
MUCOPOLISACÁRIDO UNIDAD REPETITIVA DISACÁRIDA LOCALIZACIÓN
FUNCIÓN ESTRUCTURAL
Elevada tendencia a la hidratación, actuando como lubricantes
Confieren viscosidad y elasticidad (conforman el cemento intercelular y son
amortiguadores)
Suelen unirse de forma covalente a proteínas (excepto el ácido hialurónico),
formando los proteoglucanos2
Compuestos sensibles al ataque por hialuronidasa (presente en
microorganismos y espermatozoides)
ÁC HIALURÓNICO
No está sulfatado
Líquido sinovial
Cordón umbilical
Humor vítreo
CONDROITÍN
SULFATO
Es el más
abundante
Cartílago
Ligamentos
Tendón
DERMATÁN
SULFATO
Piel
Vasos
sanguíneos
Válvulas
2 Nombre reservado a las moléculas híbridas de polisacáridos y proteínas.

Bioquímica InspiracleQIR/2015
31
TEMA 3. LÍPIDOS [p. 181, 188, 199, 241, 251 (2010); 134, 152, 154, 199, 221, 223, 228, 231,
243 (2011); 113, 119, 121, 124, 151, 199, 209, 217, 219, 222 (2012); 114, 187, 203 (2013); 135, 139,
145, 201, 206 (2014)]
Los lípidos son compuestos orgánicos que se definen por su insolubilidad en agua y
solubilidad en disolventes polares (éter, cloroformo, benceno..), no obstante se contemplan
excepciones.
Entre las características más sobresalientes:
- compuestos hidrofóbicos
- no son moléculas poliméricas
- exhiben una mayor variedad estructural
Funciones biológicas
- energética
- estructural
- aislante
- funciones especiales: ciertos lípidos desempeñan funciones especiales en el
organismo. Por ejemplo, los esteroides, los eicosanoides y algunos metabolitos de
los fosfolípidos funcionan como señales. Actúan como hormonas, mediadores y
segundos mensajeros. Algunos son cofactores de reacciones enzimáticas (vit. K..).
otros se utilizan como anclas para fijar las proteínas a las membranas.
Clasificación
3.1 Lípidos saponificables
- Contienen ácidos grasos
-Tras la hidrólisis alcalina se
obtienen jabones
Simples
(C, H, O)
Ácidos grasos
Acilglicéridos
Céridos
Estéridos
Otros: etólidos y eteroglicéridos
Complejos
Además de (C, H, O)
pueden contener N,
P, S
Fosfoacilglicéridos
Esfingolípidos
Lipoproteínas
3.2.Lípidos insaponificables
- No contienen en su
estructura ácidos grasos
- Derivan del isopreno
Terpenoides Vit. A, E, K Ubiquinonas
Esteroides
Vit D Colesterol3 Ácidos biliares Hormonas esteroideas4
3 Las células procariotas carecen de colesterol, pero éste se encuentra en distintas cantidades en prácticamente
todas las membranas de animales, fundamentalmente mamíferos. [Preg. 157 (2008)]

Bioquímica InspiracleQIR/2015
47
3.2.1. TERPENOS
Se forman a partir de cadenas isoprenoides que permanecen lineales. Entre los más
importantes:
- vitaminas A, E y K (vitaminas liposolubles). [p. 97 (2007)]
- ubiquinona o coenzima Q (su síntesis requiere selenio), junto con el citocromo c
participa como elemento móvil en la cadena de transporte electrónico
3.2.2. ESTEROIDES [p. 209 (2004); 147; 217 (2005); 207 (2008)]
Derivan fisiológicamente del colesterol, aunque estructuralmente procedan del
ciclopentanoperhidrofrenanteno. Mayoritariamente son de origen eucariota.
Dentro de este grupo se clasifican las hormonas esteroideas, los ácidos biliares, la vitamina
D (también es un derivado isoprenoide al igual que el resto de esteroides) y el colesterol.
Colesterol (27C)
Fuente: Fig 10-16 (pag 355) del libro “Lehninger. Principios de Bioquímica. Ediciones Omega, Barcelona, 4ª ed., 2006”.
El colesterol es el principal esteroide en los tejidos animales, muy abundante en mamíferos,
es un compuesto anfipático, con:
- un grupo de cabeza polar, un hidroxilo en C3 (polo hidrofílico y posición de
esterificación con ácidos grasos),
- un cuerpo hidrocarbonato apolar formado por el núcleo esteroide y la cadena lateral
hidrocarbonada ramificada en C20 (cola apolar)
- una insaturación en C5
- grupos metilo en C18 y C19
El colesterol ejerce un papel fundamental regulando la fluidez de las membranas de las
células eucariotas, puesto que ello depende del cociente fosfolípidos/colesterol, de este
modo, a mayor contenido en colesterol, menor será la fluidez de la membrana.

Bioquímica InspiracleQIR/2015
49
TEMA 4. AMINOÁCIDOS, PÉPTIDOS Y PROTEÍNAS [p. 131, 137, 138, 139,
147, 152, 156, 161, 163, 166, 172, 174, 176, 178, 185, 187, 190, 193, 202, 203, 204, 206, 212, 219,
221, 224, 225, 237, 240, 243, 244, 249, 256, 260 (2010); 131, 136, 141, 143, 144, 145, 151, 153, 157,
163, 166, 178, 179, 180, 181, 187, 195, 198, 200, 208, 214, 215, 216, 225, 232, 235,243, 244, 246,
256, 260 (2011); 116, 117, 122, 125, 126, 131, 135, 142, 144, 145, 154, 158, 161, 169, 190, 191, 192,
193, 200. 213, 225, 235 (2012); 118, 119, 121, 122, 127, 134, 139, 144, 150, 153, 154, 155, 158, 163,
180, 184, 185, 189, 192, 194, 205, 208, 227, 229, 235 (2013); 117, 130, 131, 134, 138, 158, 162, 171,
173, 175, 176, 183, 203, 208, 216, 218, 235 (2014)].
4.1 AMINOÁCIDOS.
Los α-aminoácidos son las unidades estructurales básicas que se obtienen tras la hidrólisis
ácida de las proteínas. En las proteínas encontramos 20 aminoácidos que denominamos
estándar.
4.1.1 Características estructurales
- Todos los aminoácidos contienen un grupo carboxilo y un grupo amino (la prolina posee su
grupo nitrogenado en forma de amina secundaria) unidos al mismo átomo de carbono, que
denominamos carbono α.
- Difieren unos de otros en sus cadenas laterales, o grupos R,, que varían en estructura,
tamaño y carga eléctrica, y que influyen en su solubilidad en agua.
4.1.2 Estereoquímica de los aminoácidos [p. 40; 77 (2007); 220 (2008)]
- En todos los aminoácidos, con excepción de la glicina, el Cα está unido a 4 sustituyentes
diferentes, es un centro quiral, y confiere a los aminoácidos actividad óptica.
- De la misma manera que para las osas, se definen dos series de aminoácidos, en este
caso, en función de la orientación del grupo NH2 que porta el Cα: la serie D (el grupo amino
se dispone igual que en el D-gliceraldehido) y la serie L (como en el L-gliceraldehido).
- Los aminoácidos naturales son de la serie L. los aminoácidos de la serie D se encuentran
en las paredes de las bacterias y en la estructura de ciertos antibióticos peptídicos.
- Algunos aminoácidos poseen un segundo átomo de carbono asimétrico, y por tanto
posibilitan la existencia de 4 estereoisómeros: treonina e isoleucina.
Bioquímica InspiracleQIR/2015
53
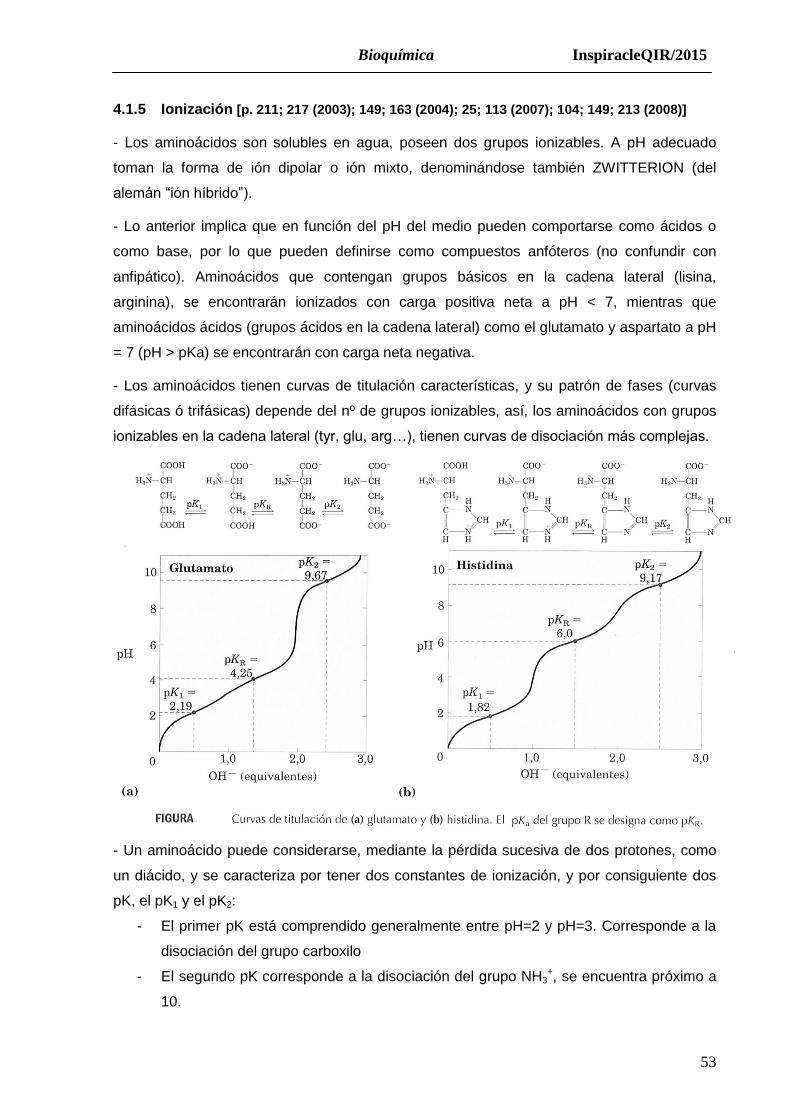
4.1.5 Ionización [p. 211; 217 (2003); 149; 163 (2004); 25; 113 (2007); 104; 149; 213 (2008)]
- Los aminoácidos son solubles en agua, poseen dos grupos ionizables. A pH adecuado
toman la forma de ión dipolar o ión mixto, denominándose también ZWITTERION (del
alemán “ión híbrido”).
- Lo anterior implica que en función del pH del medio pueden comportarse como ácidos o
como base, por lo que pueden definirse como compuestos anfóteros (no confundir con
anfipático). Aminoácidos que contengan grupos básicos en la cadena lateral (lisina,
arginina), se encontrarán ionizados con carga positiva neta a pH < 7, mientras que
aminoácidos ácidos (grupos ácidos en la cadena lateral) como el glutamato y aspartato a pH
= 7 (pH > pKa) se encontrarán con carga neta negativa.
- Los aminoácidos tienen curvas de titulación características, y su patrón de fases (curvas
difásicas ó trifásicas) depende del nº de grupos ionizables, así, los aminoácidos con grupos
ionizables en la cadena lateral (tyr, glu, arg…), tienen curvas de disociación más complejas.
- Un aminoácido puede considerarse, mediante la pérdida sucesiva de dos protones, como
un diácido, y se caracteriza por tener dos constantes de ionización, y por consiguiente dos
pK, el pK1 y el pK2:
- El primer pK está comprendido generalmente entre pH=2 y pH=3. Corresponde a la
disociación del grupo carboxilo
- El segundo pK corresponde a la disociación del grupo NH3+, se encuentra próximo a
10.

InspiracleQIR/2015 Bioquímica
62
Fuerzas que estabilizan la hélice
La hélice está estabilizada por enlaces de hidrógeno que se establecen entre el átomo
de hidrógeno unido al átomo de nitrógeno electronegativo de un enlace peptídico y el
átomo de oxígeno carbonílico electronegativo del cuarto aminoácido del lado amino
terminal de ese polipétido. Se dice que en este tipo de estructura, el aminoácido
espacialmente más próximo a un aminoácido (n) es el (n+3) ó (n+4).
Cada uno de los enlaces peptídicos, salvo los próximos a cada extremo de la hélice
participa en el establecimiento de enlaces de hidrógeno, de manera que cada vuelta
sucesiva de la hélice se mantiene unida a las vueltas adyacentes mediante 3 ó 4 enlaces
de hidrógeno.
Factores que afectan a la estabilidad de la hélice
- Repulsión (o atracción) electrostática entre residuos sucesivos con grupos R
cargados (ej: glutamatos o lisinas adyacentes, a pH fisiológico).
- Interacciones entre cadenas laterales de aminoácidos separados tres (o 4) residuos
- Volumen de los grupos R adyacentes (también la forma). Aminoácidos
desestabilizantes son Asn, Ser, Thr y Cys

Bioquímica InspiracleQIR/2015
65
Giros beta [p. 226 (2005)]
Son elementos de conexión que unen tramos sucesivos de hélices alfa o conformaciones
beta. Son muy frecuentes los giros beta que conectan los extremos adyacentes de dos
segmentos de hojas beta antiparalelas.
Estas estructuras forman giros cerrados de 180º mediante enlaces de H establecidos entre
los grupos peptídicos del primer y cuarto aminoácidos de los 4 implicados en el giro.
A menudo se encuentran Gly y Pro.
Los giros B suelen ubicarse en la superficie de las proteínas globulares
En el plegamiento de una proteína existe un estado intermedio denominado glóbulo fundido, en el que
se han formado casi todos los elementos de estructura secundaria.
4.3.2.3. Estructura terciaria. [p. 142; 146; 162 (2003); 84 (2007); 155; 214 (2009)]
Hace referencia al plegamiento espacial completo y compacto de cada cadena.
Incluye el conjunto de interacciones, covalentes o de otro tipo, como puentes disulfuro,
puentes de hidrógeno, interacciones hidrofóbicas, fuerzas de Van der Waals e interacciones
iónicas.
El efecto hidrofóbico favorece el plegamiento de las proteínas. En la estructura de las
proteínas hay aminoácidos apolares, los cuales poseen alta tendencia a asociarse entre sí,
en el interior de la proteína plegada. La interacción entre estos fragmentos polares, provoca
un incremento de la entropía del agua, lo cual compensa la propia pérdida de entropía del
inherente al proceso de plegamiento. El incremento de la entropía de algua viene producido
por la liberación de moléculas de agua en el proceso de interacción (unión) entre los
fragmentos apolares, debido a la propia tendencia del agua a minimizar el contacto con
moléculas hidrofóbicas. Preg 197 (2012).
En esta estructura, los fragmentos con estructuras secundarias variadas pueden combinarse
con zonas sin estructura secundaria definida, o zonas de giro donde las cadenas se pliegan
con un patrón determinado.
La disposición de los aminoácidos difiere según la proteína sea fibrosa o globular, así, en las
proteínas globulares (que suelen ser solubles):
- los aminoácidos no polares se sitúan preferentemente hacia el interior de la proteína,
para evitar el contacto con el disolvente acuoso
- los residuos con carga suelen ubicarse hacia el exterior , interaccionando con el
medio acuoso
- los grupos polares de los aminoácidos sin carga se distribuyen a lo largo de la
cadena polipeptídica, pero con cierta preferencia, aparecen también hacia la zona
externa.

Bioquímica InspiracleQIR/2015
67
PROTEÍNAS FIBROSAS PROTEÍNAS GLOBULARES
Constan mayoritariamente de un único
tipo de estructura secundaria
Suelen contener varios tipos de
estructura secundaria
Proteínas de los tejidos estructurales Incluyen enzimas, proteínas de
transporte, proteínas motoras, proteínas
reguladoras, inmunoglobulinas…
α-queratinas, β- queratinas, colágeno Mioglobina, hemoglobina
PROTEÍNAS FIBROSAS
Colágeno [p. 207 (2003); 151 (2004); 194 (2006); 50 (2007); 137; 165; 225 (2008); 241 (2009)]
Es la proteína más abundante del cuerpo humano.
Se caracteriza por formar fibras muy insolubles y muy resistentes a la tracción, pero, a la
vez, flexibles.
El colágeno es una glucoproteína, pues se unen covalentemente monómeros de galactosa o
disacáridos galactosa –glucosa a los hidroxilos de la lisina.
Desde el punto de vista estructural presenta una estructura secundaria única, está
compuesto por secuencias de unidades de tropocolágeno, que se repiten de forma regular.
Cada unidad de tropocolágeno consta de tres cadenas polipeptídicas llamadas cadenas α.
Cada una de estas cadenas posee un arrollamiento levógiro, y las tres cadenas se
entrelazan entre sí mediante enlaces cruzados, formando una triple hélice dextrógira
(tropocolágeno).
Existen muchos tipos de colágeno en vertebrados. Normalmente contienen alrededor de un
35% de Gly, un 11 % de Ala y un 21 % de Pro y de 4-OH-Pro (formada por hidroxilación
catalizada por la *prolil hidroxilasa y requiere ascorbato (Vitamina C) para su síntesis).
La secuencia de aminoácidos del colágeno suele corresponder a la repetición de un
tripéptido del tipo Gly-X-Y, donde X suele ser Pro e Y con frecuencia es 4OH-Pro.
Las uniones entre las cadenas alfa son tan estrechas, que prácticamente es la Gly el único
aminoácido que puede acomodarse.
*La prolil hidroxilasa y la lisil hidroxilasa requieren para catalizar la reacción de hidroxilación, vit C (el
colágeno es menos estable cuando hay ausencia de esta vitamina) y Fe+2
[p. 218 (2014)].
EL fallo en la hidroxilación de la prolina y lisina tiene importantes consecuencias patológicas.

InspiracleQIR/2015 Bioquímica
68
Las fuerzas que estabilizan las cadenas de colágeno:
- Entrecruzamientos covalentes cruzados (ó “puentes de piridinolina”), por acción
de una lisil oxidasa (requiere Cu2+ y vit B6), que transforma la lis y la *OH-Lis en
derivados aldehidicos (lisinal ó al-lisina)
- Puentes de hidrógeno intracatenarios
* El colágeno y la elastina son las dos únicas proteínas que contienen hidroxilisina y se glucosilan en
los hidroxilos.
Las moléculas de colágeno contienen en sus extremos unas cortas secuencias de
aminoácidos, que son esenciales para el establecimiento de los enlaces cruzados y el
correcto alineamiento de las mismas, se denominan telopéptidos.
En total, se han postulado más de 30 especies de colágeno, de las cuales se han
caracterizado 9.
Tipos, composición y ubicación principal del colágeno
Tipo Característica Tropocolágeno Ubicación Contenido
I Forma fibras
(90% total) [1(I)]2[2(II)] En hueso, tendón,
ligamentos, piel,
córnea y pulpa dental
HOLys bajo
Glicosilación bajo
Fibras finas
II Forma fibras [1(II)]3 Cartílago, humor
vítreo
HOLys alto
Glicosilación alto
Fibra ancha
III Forma fibras [1(III)]3 Piel joven, vasos
sanguíneos, pulpa
dental
HOPro alto
HOLys bajo
Glicosilación bajo
Puentes disulfuro
extras en C-
terminal
IV Forma mallas [1(IV)]2[2(IV)] Lámina basal HOLys alto
Glicosilación alto
Pobre en Ala
V Forma fibras [1(V)]2[2(V)] Idem al tipo I, menor
abundancia
HOLys alto
Glicosilación alto
Pobre en Ala
VII Forma mallas [1(VII)]3 Lámina del epitelio
estratificado
---
IX Asociado a fibras [1(IX)][2(IX)]
[3(IX)]
Cartílago ---
XI Forma fibras [1(XI)][2(XI)]
[3(XI)]
Idem al tipo II, menor
abundancia
---
XII Asociado a fibras [1(XII)]3 Tendón, ligamentos ---

InspiracleQIR/2015 Bioquímica
70
PROTEÍNAS GLOBULARES [p. 144; 185; 222; 235 (2003); 172 (2004); 236 (2005); 192 (2006);
175; 203 (2008); 206; 248 (2009)]
MIOGLOBINA HEMOGLOBINA9
FUNCIÓN Almacenamiento de oxígeno Transporte de oxígeno
ESTRUCTURA Monomérica Oligomérica (4 sub)
AFINIDAD POR O2 Alta Baja y es menor cuando
desciende el pH, se regula por
fosfatos orgánicos.
CINÉTICA Hiperbólica.
Cinética de M-M
(no cooperatividad)
Sigmoidal
Alostérica (se da el fenómeno
de cooperatividad) Un efector
alostérico es el 2,3-
bisfosfoglicerato, que se une a
la desoxihemoglobina, y
disminuye la afinidad del
oxígeno por la hemoglobina A.
Metal Fe (II) Fe (II)
La oxidación del Fe (II) de la mioglobina, así como la hemoglobina conduce a
metamioglobina y metahemoglobina, perdiéndose la capacidad de unión al oxígeno.
En relación con la estructura de la mioglobina y la hemoglobina es importante que el
grupo hemo se encuentre en un ambiente apolar con el fin de prevenir la oxidación del
ión ferroso.
El CO2 se une a la a la hemoglobina en los grupos alfa-amino. Con respecto al CO, su
toxicidad para organismos aeróbicos viene dada porque se une al grupo hemo con
mayor afinidad que el O2 [(Preg. 131 (2014)].
Tipos de Hemoglobina según su estructura:
TIPO Hb FÓRMULA OBSERVACIONES
A Adulto α2 β2 97 % de la Hb del adulto. A la glicosilada se le llama A1.
A2
Adulto
2 alfa + 2 delta
< 3 % de la Hb del adulto. ↑ en beta talasemia.
F
Fetal
2 alfa + 2 gamma
Aparece en el 2º y 3º trimestre del embarazo. 97 % de la Hb fetal y 1% en el adulto.
9 El metabolismo de la hemoglobina separa la porción glogina del grupo hemo. Este último es
metabolizado por la enzima hemo-oxigenasa originando biliverdina y posteriormente el metabolimo de
la misma por la biliverdina reductasa origina Bilirrubina. [Preg. 235 (2009); 138 (2014)]

Bioquímica InspiracleQIR/2015
71
E
Embrionaria Hb-Gower I
Hb-Gower II * Hb-Portland
2 zeta + 2 epsilon 2 alfa + 2 epsilon 2 zeta + 2 gamma
Aparecen durante el 1º trimestre del embarazo (eritropoyesis en saco vitelino). La más importante: 60 % de Hb E.
Una técnica de gran utilidad en el diagnóstico de hemoglobinopatías es la electroforesis de
hemoglobina. En la electroforesis de hemoglobina en acetato de celulosa se obtienen
tres fracciones diferenciadas: Hb A, Hb A2 y anhidrasa carbónica. Las condiciones
experimentales para la electroforesis pueden variarse, y con ellas los resultados obtenidos,
por ejemplo: en la electroforesis capilar se elimina la banda correspondiente a la anhidrasa.
Técnicas para la determinación de estructuras de macromoléculas:
[p. 189 (2004)]
1. Estructura primaria: espectrometría de masas (también es la técnica que se utiliza
para determinar con mayor exactitud el peso molecular de una proteína.
2. Estructura secundaria (análisis conformacional): métodos ópticos (dicroismo
circular fundamentalmente y dispersión rotatoria óptica) y espectroscopia (de IR, UV
y fluorescencia).
3. Estructura terciaria (análisis tridimensional): resonancia magnética nuclear (RMN) y
difracción de rayos X.
AGENTES DESNATURALIZANTES O CAOTRÓPICOS: [p. 228 (2006); 17
(2007)]
La conformación nativa de una proteína es la estructura tridimensional biológicamente
activa, por lo que la pérdida de la misma implica la pérdida de su actividad. Dicha
conformación es sensible a varios agentes desnaturalizantes. Estos compuestos pueden
facilitar la acción de proteasas, provocan la pérdida de estructura secundaria y terciaria,
pero no afectan la estructura primaria de la proteína. Por lo general implica un aumento de la
viscosidad. La desnaturalización en algunos casos puede ser reversible.
Agentes desnaturalizantes:
pH extremo o temperaturas elevadas
Cloruro de guanidinio: rompe enlaces no covalentes.
β-mercaptoetanol, ditiotreitol y ácido perfórmico: rompen los puentes disulfuro.
Urea: rompe puentes de H.
Detergentes (ej: dodecilsulfato de sodio = SDS): desnaturaliza dotando de una carga
superficial negativa a la proteína. Muy utilizado en SDS-PAGE (electroforesis en gel

InspiracleQIR/2015 Bioquímica
88
REGULACIÓN ENZIMÁTICA. [p. 179; 187 (2003); 219 (2004); 142; 230 (2005); 73; 133;
160 (2008); 133; 222; 233 (2009)] La actividad enzimática puede regularese a nivel de la síntesis de la propia proteína con al
función enzimática, o a nivel de la proteína ya sintetizada.
Existen diferentes niveles de regulación de la actividad enzimática:
- Control de la cantidad / concentración de enzima: regulación LENTA. Se lleva a
cabo a nivel de la expresión de los genes que codifican las enzimas, implicando un
control de la síntesis y la degradación de las mismas. (ADN → ARN → proteína).
- Compartimentación: de forma general las enzimas que participan en una misma ruta
suelen ubicarse en el mismo compartimiento subcelular. La compartimentación permite
la separación de dos rutas opuestas que podrían interferirse mutuamente (ej: biosíntesis
de ácidos grasos transcurre en el citosol, mientras su degradación transcurre en la matriz
mitocondrial), y permite regular la velocidad de funcionamiento de ambas rutas
controlando la disponibilidad de sustrato. Algunas rutas poseen enzimas en
compartimentos diferentes (ej: gluconeogénesis, ruta para sintetizar glucosa a partir de
precursores no glucídicos, posee etapas citosólicas, la mayor parte, y una mitocondrial).
- Existencia de Isoenzimas: Formas múltiples de una enzima con diferente localización
en el organismo, que catalizan la misma reacción pero exhibiendo parámetros
cinéticos diferentes (afinidad diferente por un sustrato). Presentan diferente pHi, PM
(movilidad electroforética), sensibilidad a moduladores de la actividad enzimática, etc.
(Ej: LDH, CPK, FAlc).
- Activación por proteólisis o existencia de zimógenos (proenzimas): Muchas
enzimas son sintetizadas de una forma inactiva como zimógenos o proenzimas y se
activan por proteólisis. Este tipo de activación es irreversible y transcurre por la acción
de proteasas. (Ej: proteasas digestivas: quimotripsinógeno / quimotripsina, pepsinógeno /
pepsina; proteínas de la cascada de la coagulación: fibrinógeno/ fibrina; proteínas del
complemento, proinsulina / insulina por pérdida del péptido C).
- Control de la actividad enzimática: regulación RÁPIDA, mediante enzimas
reguladoras, implica el control de la velocidad de la ruta. Puede ser: regulación
covalente reversible (Ej: por fosforilación, adenilación, metilación, etc.) y/o
regulación alostérica reversible por sustrato o moduladores.
Regulación covalente reversible: Generalmente por fosforilación reversible de una
enzima que alterna entre dos formas: una más activa y otra menos activa. Suele estar
mediada por la acción hormonal. Mecanismo más habitual: H → H – R → activación de
proteína G → activación de Adenilato Ciclasa → ↑ AMPc → activación de proteína kinasa A →
fosforilación de la enzima. La enzima fosforilada puede ser más o menos activa. La
regulación por modificación covalente requiere de la acción de otras enzimas. En el caso

InspiracleQIR/2015 Bioquímica
90
Se distinguen dos tipos de interacciones como resultado de la naturaleza oligomérica de
las enzimas alostéricas:
Homotrópicas (COOPERATIVIDAD): Efecto de un ligando sobre otro ligando
idéntico con sitios de unión equivalentes (Ej: la unión de O2 a una molécula de Hb
estimula la unión de otros O2 a la Hb). Los procesos de cooperatividad suelen ser
positivos.
Heterotrópicas (ALOSTERISMO): Efecto de un ligando sobre otro ligando diferente
con sitios de unión no equivalentes (Ej: la unión del 2,3-diP- glicerato a la Hb afecta la
unión de O2 a la propia Hb). Pueden ser positivas o negativas.
Modelos para explicar la CINÉTICA ALOSTÉRICA:
Las enzimas alostéricas no siguen una cinética de Michaelis-Menten, por lo que no tienen un
comportamiento hiperbólico típico de ésta, sino que en una representación de actividad
enzimática vs [sustrato] la curva es de tipo sigmoidal (de manera que pequeños cambios en
la [S] provocan grandes cambios en la actividad enzimática).
Modelos explicativos:
1) Modelo concertado o simétrico de MONOD-WYMAN-CHANGEAUX: Propone que
el enzima existe solo en dos estados: tenso (T) y relajado (R) (no existiendo estados
híbridos), que se encuentran en equilibrio. Los sustratos y activadores, aunque
puedan unirse al estado T poseen mayor afinidad por el estado R, de ahí que lo
favorecen, mientras que los inhibidores o la ausencia de sustrato favorecen el estado
T.
2) Modelo secuencial o asimétrico de KOSHLAND: Propone que el efecto de la
fijación del ligando se transmite secuencialmente, mediante una sucesión de estados
híbridos, a través del oligómero, dando lugar a una afinidad creciente o decreciente
hacia el ligando por los protómeros contiguos. Explica tanto alosterismo como
cooperatividad (tanto positiva como negativa, infrecuente).
Nota: la representación de Scatchard se basa en el estudio de la unión de ligando a receptor y suele
utilizarse para determinar el número de sitios de unión de un ligando a la enzima. Permite calcular
constantes de afinidad ligando – proteína.
InspiracleQIR/2015 Bioquímica
96
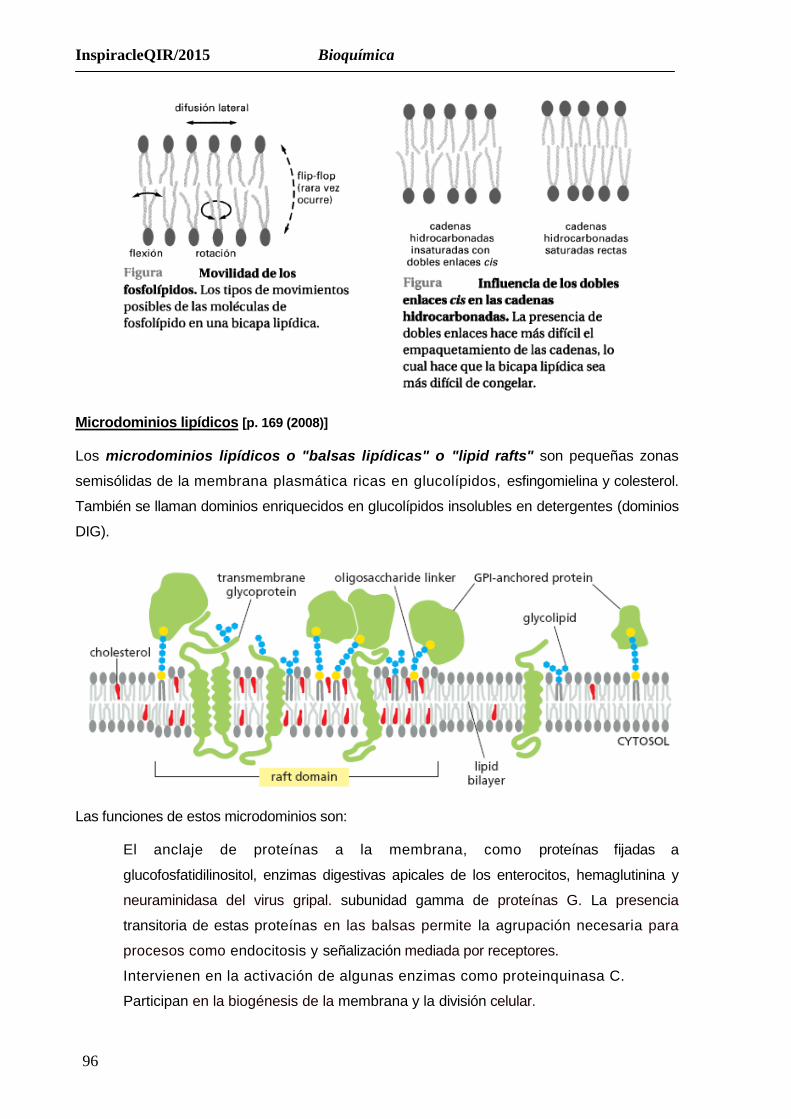
Microdominios lipídicos [p. 169 (2008)]
Los microdominios lipídicos o "balsas lipídicas" o "lipid rafts" son pequeñas zonas
semisólidas de la membrana plasmática ricas en glucolípidos, esfingomielina y colesterol.
También se llaman dominios enriquecidos en glucolípidos insolubles en detergentes (dominios
DIG).
Las funciones de estos microdominios son:
El anclaje de proteínas a la membrana, como proteínas fijadas a
glucofosfatidilinositol, enzimas digestivas apicales de los enterocitos, hemaglutinina y
neuraminidasa del virus gripal. subunidad gamma de proteínas G. La presencia
transitoria de estas proteínas en las balsas permite la agrupación necesaria para
procesos como endocitosis y señalización mediada por receptores.
Intervienen en la activación de algunas enzimas como proteinquinasa C.
Participan en la biogénesis de la membrana y la división celular.

Bioquímica InspiracleQIR/2015
137
FUNCIONES DEL CICLO DE KREBS [p. 220 (2004)]
- Ciclo anfibólico: tiene tanto carácter catabólico (degradando las moléculas de AcCoA)
como anabólico, pues varios de los intermediarios del ciclo pueden utilizarse para
biosintetizar distintos compuestos.
Fuente: Fig 16-15 (pag 617) del libro “Lehninger. Principios de Bioquímica. Ediciones Omega, Barcelona, 4ª ed., 2006”.
- Reacciones anapleróticas: se definen como reacciones que abastecen de intermediarios a
un ciclo. A medida que los intermediarios van siendo retirados del ciclo de krebs para servir
como precursores biosintéticos, son repuestos mediante reacciones denominadas
anapleróticas. Las más importantes se recogen en la tabla.
Fuente: Tabla 16-12 (pag 618) del libro “Lehninger. Principios de Bioquímica. Ediciones Omega, Barcelona, 4ª
ed., 2006”.

InspiracleQIR/2015 Bioquímica
140
los dos al mismo tiempo. Estos centros ferrosulfurados van desde estructuras
sencillas con un solo átomo de Fe coordinado con 4 residuos cys mediante el azufre
de sus cadenas laterales, hasta centros Fe-S más complejos con dos o cuatro
átomos de Fe. Las proteínas ferrosulfuradas de Rieske, son una variante de las
anteriores, en las que un átomo de Fe está coordinado con dos residuos de His en
lugar de con dos de cys.
Los transportadores implicados en la cadena de transporte electrónico funcionan en orden
de potenciales de reducción crecientes, ya que los electrones tienden a fluir
espontáneamente desde los transportadores con un E’º (potencial de reducción estándar)
más bajo hacia los transportadores con un E’º más elevado.
- Según lo comentado, el orden deducido de los transportadores es como sigue:
NADH Q (ubiquinona) cit b cit c1 cit c cit a cit a3 O2 .
FLUJO DEL TRANSPORTE DE LOS ELECTRONES
- Los transportadores de electrones de la cadena respiratoria están organizados en
complejos supramoleculares incrustados en la membrana mitocondrial interna:
COMPLEJO I
También llamado NADH:ubiquinona oxidorreductasa o NADH deshidrogenasa; canaliza
el flujo de electrones desde el NADH a la ubiquinona. Es una flavoproteína compuesta por
varias cadenas polipeptídicas que contiene un grupo prostético FMN y varios centros
ferrosulfurados; cataliza dos tipos de procesos acoplados:

Bioquímica InspiracleQIR/2015
231
TEMA 20. METABOLISMO DE LAS LIPOPROTEÍNAS [p. 145 (2003); 232
(2004); 216, 222 (2010); 142, 176, 240 (2011); 118 (2012); 128 (2013); 202, 214 (2014)]
Los lípidos provenientes de la dieta, sintetizados por el hígado o liberados por el tejido
adiposo, deben ser trasladados hasta los tejidos que necesiten emplearlos.
• Como los lípidos son insolubles en agua, el problema de cómo transportarlos se resuelve
asociando los lípidos apolares (TAG y CE) con otros lípidos anfipáticos (FL y CL) y con
proteínas (apolipoproteínas) para constituir lipoproteínas.
CONSTITUYENTES DE LAS LIPOPROTEÍNAS
- Capa anfipática periférica, constituida por
fosfolípidos y colesterol libre en la interfase,
disponiendo sus grupos polares en contacto
con el medio acuoso.
- Interior hidrofóbico, formado por
triglicéridos y colesterol esterificado
- Contiene además una fracción protéica
denominada apoproteína
InspiracleQIR/2015 Bioquímica
232
APOPROTEÍNAS MÁS IMPORTANTES
Apolipoproteínas de las lipoproteínas plasmáticas humanas
Apolipoptroteína Masa molecular Lipoproteína asociada Función (si se conoce)
ApoA-I 28.331 HDL Activa la LCAT; interacciona con
el transportador ABC
ApoA-II 17.380 HDL
ApoA-IV 44.000 Quilomicrones. HDL
ApoB-48 240.000 Quilomicrones
ApoB-100 513.000 VLDL, LDL Se une al receptor de LDL
ApoC-I 7000 VLDL, HDL
ApoC-II 8837 Quilomicrones, VLDL,
HDL
Activa la lipoptoteína lipasa
ApoC-III 8751 Quilomicrones, VLDL,
HDL
Inhibe la lipoproteína lipasa
ApoD 32.500 HDL
ApoE 34.145 Quilomicrones, VLDL,
HDL
Desencadena la eliminación de
VLDL y quilomicrones residuales
*ApoE: presenta varias isoformas, la mayoritaria es la ApoEIII. Las isoformas EI y EII interaccionan
ineficazmente con los receptores hepáticos, por lo que son lentamente catabolizadas (pasan a LDL),
mientras las isoformas EIII y EIV son reconocidas y rápidamente catabolizadas por el hígado.
Además, se han relacionado las isoformas EII conla hiperlipoproteinemia tipo III (ver más adelante) y
el fenotipo E IV (el minoritario?) parece conferir a sus portadores un mayor riesgo de padecer la
enfermedad enfermedad cardiovascular (por elevación del colesterol LDL) y la enfermedad de
Alzheimer.
PRINCIPALES LIPOPROTEÍNAS EN PLASMA
Composición (%)
Lipoproteína Densidad
(g/ml)
Proteína Fosfolípido Colesterol
libre
Ésteres de
colesterol
Triacilgliceroles
Quilomicrones <0,95 2 9 1 3 85
VLDL <1,006 10 18 7 12 50
IDL 1,006-1,019 18 12 8 30 31
LDL 1,019-1,063 23 20 8 37 10
HDL 1,063-1,210 55 24 2 15 4

Bioquímica InspiracleQIR/2015
243
TEMA 21. METABOLISMO DE AMINOÁCIDOS [p. 157 (2004), 145, 169, 180,
197, 242 (2010); 132, 133, 137, 150, 190, 219 (2011); 127, 150, 163, 166, 185, 203, 224 (2012; 132,
146, 170, 180, 185, 188, 191, 192, 195, 200, 233 (2014)]
La fracción de energía metabólica generada a partir de los aminoácidos, ya sean
procedentes de las proteínas de la dieta o de proteínas de los tejidos, varía mucho según el
tipo de organismos y las situaciones metabólicas.
En los animales, los aminoácidos sufren degradación oxidativa en tres situaciones:
- Durante la síntesis y degradación normales de proteínas celulares (recambio
protéico), algunos de los aá liberados durante la degradación de las proteínas se
degradan oxidativamente si no se necesitan para la síntesis de nuevas proteínas.
- Cuando una dieta es rica en proteínas y los aás ingeridos exceden las necesidades
corporales para la síntesis de proteínas, el excedente se cataboliza; los aás no se
pueden almacenar.
- Durante la inanición o la diabetes mellitus, en las que no hay glúcidos disponibles, o
estos no son utilizados adecuadamente, se recurre a las proteínas celulares como
combustible.
Los aá obtenidos a partir de las proteínas de la dieta son la fuente de la mayor parte de
grupos amino. La digestión de proteínas ocurre en estómago e intestino por proteasas de los
jugos gástrico y pancreático: el jugo gástrico presenta PEPSINA, proteasa que genera
péptidos y algunos aa libres. El jugo pancreático contiene varias proteasas (tripsina,
quimotripsina, carboxipeptidasa A y B, aminopeptidasa). Las proteasas y peptidasas se
sintetizan como zimógenos y se activan por proteólisis. De los zimógenos, la activación del
tripsinógeno es iniciada por la enteropeptidasa. La absorción de aa se produce en
intestino delgado y de ahí los aminoácidos pasan a la circulación.
La mayoría de aás se metabolizan en el hígado. Parte del amoníaco generado en este
proceso se recicla se utiliza en diversas rutas biosintéticas; el exceso se excreta
directamente o se convierte en urea o ácido úrico para su excreción, según el organismo.
El exceso de amoníaco generado en otros tejidos (extrahepáticos) se transporta al hígado
(en forma de grupos amino) para su conversión en la forma en que se excreta.
El glutamato y la glutamina desempeñan papeles especialmente críticos en el metabolismo
del nitrógeno, actuando como una especie de puntos de recogida de grupos amino. En el
citosol se los hepatocitos, los grupos amino de la mayoría de los aminoácidos se
transfieren al alfacetoglutarato formando glutamato. A continuación se transporta el
glutamato a la mitocondria, donde se elimina el grupo amino en forma de NH4+.

InspiracleQIR/2015 Bioquímica
244
El exceso de amoníaco generado en la mayor parte de los tejidos se convierte en el
nitrógeno amídico de la glutamina, que pasa al hígado y seguidamente a las mitocondrias
hepáticas. En la mayoría de los tejidos, la glutamina o el glutamato se encuentran en
concentraciones más elevadas que el resto de aminoácidos.
En el músculo esquelético, los grupos amino en exceso se transfieren al piruvato formando
alanina, otra molécula importante en el transporte de grupos amino al hígado.
CATABOLISMO DE LOS AMINOÁCIDOS EN EL HÍGADO
TRANSAMINACIÓN [p. 250 (2003); 166 (2004); 1 (2007); 164 (2009)]
El primer paso es la eliminación de los grupos amino, catalizado por enzimas
transaminasas, que catalizan la transferencia de un grupo amino al átomo de carbono del
alfacetoglutarato, dejando el correspondiente alfacetoácido análogo del aminoácido.
El cometido de las reacciones de transaminación es recoger los grupos amino de muchos
aminoácidos diferentes en forma de glutamato. El glutamato funciona a continuación como
grupo dador de grupos amino para rutas biosintéticas o para rutas de excreción que
conducen a la eliminación de productos nitrogenados de deshecho (ciclo de la urea).
Las células contienen varios tipos de aminotransferasas, todas contienen como grupo
prostético el piridoxal fosfato (PLP) forma coenzimática de la vitamina B6.
Las distintas transaminasas se diferencian en su especificidad por el aminoácido; entre las
más abundantes en los tejidos destacan:
- GOT: glutamato /oxalacetato transaminasa; su par aá/alfacetoácido sería
alfacetoglutarato/aspartato; también se denomina aspartato aminotransferasa.
- GPT: glutamato/piruvato transaminasa, cuyo par sería alfacetoglutarato/alanina
El nivel elevado de estas transaminasas es un indicio de enfermedad hepática o cardíaca.
Por acción de las transaminasas el nitrógeno de los aás se recoge en forma de glutamato.
DESAMINACIÓN OXIDATIVA [p. 104 (2005)]
Los grupos amino de muchos aminoácidos se recogen en el hígado en forma del grupo
amino del glutamato. A continuación estos grupos amino son eliminados del glutamato para
prepararlos para la excreción. En los hepatocitos el glutamato se transporta desde el citosol
a la mitocondria, en donde experimenta desaminación oxidativa catalizada por la glutamato
deshidrogenasa.
En mamíferos este enzima se encuentra en la matriz mitocondrial; es el único enzima que
puede utilizar tanto NAD+ como NADP+ como aceptor de los equivalentes de reducción.
La acción combinada de una transaminación y una desaminación oxidativa se denomina
TRANSDESAMINACIÓN. Algunos aminoácidos se saltan esta ruta de desaminación y se
desaminan directamente.

Bioquímica InspiracleQIR/2015
245
La reacción inversa de la desaminación oxidativa es la aminación reductora y se lleva a cabo
mediante fosfato del piridoxal y fosfato de piridoxamina.
TRANSPORTE DE AMONÍACO AL HÍGADO POR LA GLUTAMINA
El amoníaco es muy tóxico para los tejidos animales, por lo que sus niveles en sangre están
regulados. En algunos tejidos, incluído cerebro, se genera amoníco libre en algunos
procesos tales como la degradación de nucleótidos. En la mayoría de animales el exceso de
amoníaco se convierte en un compuesto no tóxico, antes de ser exportado de los tejidos
extrahepáticos a la sangre y transportado al hígado o los riñones.
El glutamato, que es tan importante para el metabolismo intracelular de grupos amino es
sustituido por la glutamina para esta función de transporte. El amoníaco libre producido en
los tejidos se combina con glutamato dando glutamina (glutamina sintasa).
La glutamina constituye una forma de transporte no tóxica del amoníaco; normalmente está
presente en la sangre en concentraciones mucho mayores que otros aminoácidos, además
la glutamina sirve de fuente de grupos amino en muchas reacciones de biosíntesis.
En la mayoría de los animales terrestres, la glutamina en exceso respecto a la necesaria
para la biosíntesis se transporta por la sangre al intestino, hígado y riñones para su
tratamiento.
En estos tejidos el nitrógeno amídico se libera en forma de un ión amonio dentro de las
mitocondrias, en donde el enzima glutaminasa convierte la glutamina en glutamato y NH4+.
TRANSPORTE DE AMONÍACO DESDE LOS MÚSCULOS AL HÍGADO EN FORMA DE
ALANINA
La alanina también juega un papel importante en el transporte de grupos amino al hígado en
una forma no tóxica, mediante el ciclo de la glucosa-alanina (esquema en tema de
gluconeogénesis). En el músculo y algún otro tejido que degrada aás como combustible, los
grupos amino se recogen en forma de glutamato por transaminación. El glutamato puede
entonces convertirse en glutamina para su transporte al hígado o puede o puede transferir
su grupo amino al piruvato por acción de la alanina aminotransferasa, la alanina así formada
pasa a la sangre y es transportada hasta el hígado. En el citosol de los hepatocitos, la
alanina aminotransferasa, transfiere el grupo amino de la alanina al alfacetoglutarato,
formando piruvato y glutamato. El glutamato puede entrar en las mitocondrias, donde la
reacción de la glutamato DHasa libera NH4+.

InspiracleQIR/2015 Bioquímica
246
CICLO DE LA UREA [p. 165; 197 (2003); 228 (2005); 178 (2006); 4; 15; 57; 74 (2007); 137
(2009); p. 242 (2010)]
Si no se reutilizan para la síntesis de nuevos aminoácidos u otros productos nitrogenados,
los grupos amino se canalizan a un único producto final de excreción. En función de la
naturaleza del compuesto de excreción, podemos clasificar a los organismos en:
amonotélidosexcretan el nitrógeno amínico en forma de amoníaco (peces)
uricotélidos el nitrógeno es excretado en forma de ácido úrico (aves y mamíferos)
ureotélidos la excreción de nitrógeno se lleva a cabo mediante la producción de urea (la
mayoría de animales terrestres)
En los animales ureotélidos el amoníaco depositado en las mitocondrias de los hepatocitos,
se convierten en urea mediante el ciclo de la urea. La producción de urea tiene lugar casi
exclusivamente en el hígado y representa el destino de la mayor parte del amoníaco allí
canalizado. La urea pasa al torrente sanguíneo y de ahí a los riñones y se excreta en la
orina.
La urea se sintetiza gracias a una ruta de 5 etapas enzimáticas, y cuyas dos primeras
reacciones transcurren en la matriz mitocondrial; las restantes tienen lugar en el citosol de
los hepatocitos.
El primer grupo amino que entra en el ciclo de la urea proviene del amoníaco de la matriz
mitocondrial (NH4+). El NH4
+ generado en las mitocondrias hepáticas se utiliza
inmediatamente, junto con el CO2 (en la forma HCO3-) producido por la respiración
mitocondrial, para dar carbamil-P en la matriz, en una reacción catalizada por la carbamilP
sintetasa I, un enzima regulador. El carbamil-P, que funciona como un dador activado del
grupo carbamilo, entra ahora en el ciclo de la urea que consta de 4 etapas enzimáticas (ver
esquema). El ciclo de la urea en eucariotas requiere transportadores de membrana
específicos para determinados metabolitos porque la membrana interna mitocondrial es
impermeable.

Bioquímica InspiracleQIR/2015
247
El déficit de cualquiera de las enzimas del ciclo de la urea da lugar al acúmulo de NH4
(hiperamonemias) lo que se trata con una dieta pobre en proteínas, rica en glúcidos y alfa-
cetoácidos y con benzoato (sódico). Si la deficiencia es total, puede llegar a ser incompatible
con la vida.
Tipos Enzima deficitaria
Hiperamonemia tipo I y II Carbamil fosfato sintasa I y Ornitina transcarbamilasa, respectivamente.
Citrulinemia Argininosuccinato sintetasa
Aciduria arginino-succínica Argininosuccinato liasa
Argininemia Arginasa
Otros N-acetil glutamato sintetasa
Otras situaciones que también provocan hiperamonemia son: ayuno (por el aumento del
catabolismo proteico, deshidratación e insuficiencia hepática (cirrosis, síndrome de Reye y
encefalopatía hepática).
REGULACIÓN
El flujo de nitrógeno a través del ciclo de la urea varía con la dieta de un individuo a otro.
Cuando la dieta es mayoritariamente protéica, la utilización de los esqueletos carbonados de
los aminoácidos como combustible da lugar a la producción de mucha urea a partir del
exceso de grupos amino.

InspiracleQIR/2015 Bioquímica
248
Durante la inanición prolongada, en la que la degradación de la proteína muscular empieza
a suministrar gran parte de la energía metabólica del organismo, también aumenta de forma
sustancial la producción de urea.
Estos cambios en la demanda de actividad del ciclo se consiguen a largo plazo mediante la
regulación de las velocidades de síntesis de los cuatro enzimas del ciclo de la urea y de la
carbamil-P-sintetasa I.
Los cinco enzimas se sintetizan a velocidades más elevadas durante la inanición o en los
animales con dietas muy ricas en proteínas. Los animales con dietas más pobres en
proteínas producen niveles más bajos de los enzimas.
Una regulación a más corto plazo es la ejercida por un metabolito, el N-acetil glutamato,
que es sintetizado por la N-acetil-glutamato sintasa a partir de glutamato y acetilCoA. Este
metabolito activa alostéricamente a la carbamil-P-sintetasa I; sus niveles vienen
determinados por las concentraciones de acetilCoA y glutamato, así como de la arginina.
DEGRADACIÓN DEL ESQUELETO CARBONADO DE LOS AMINOÁCIDOS [p. 195 (2003);
212 (2004); 220 (2005); 240 (2009)]
Para la degradación oxidativa de los 20 aminoácidos protéicos exiten 20 rutas diferente. En
el hombre estas rutas aportan entre el 10 y el 15% de la producción energética. Esto implica
que las rutas degradativas de los 20 aás individuales son mucho menos energéticas.
Estas 20 rutas distintas convergen en 5 productos finales que forman parte del ciclo de
krebs. Desde el ciclo de krebs, los carbonos de esos compuestos se pueden desviar a la
gluconeogénesis o a la cetogénesis o bien pueden oxidarse completamente a CO2 y H2O
(en este caso tienen que entrar como acetilCoA) a través del ciclo de Krebs y la cadena de
transporte electrónico.
De los 20 aminoácidos sólo 10 degradan sus esqueletos carbonados total o parcialmente,
generando en última instancia acetilCoA.
Sólo hay dos aminoácidos que son exclusivamente cetogénicos (rinden acetilCoA o
acetoacetato): lisina y leucina.
*Los aminoácidos ramificados leucina, isoleucina, y valina son oxidados como combustible en el
músculo, tejido adiposo, riñón y cerebro, ya que contienen una aminotransferasa específica que actúa
sobre esos aminoácidos, generando los alfacetoácidos correspondientes.

Bioquímica InspiracleQIR/2015
315
BIBLIOGRAFÍA
- NELSON, D.L. y COX, M.M.: Lehninger. Principios de Bioquímica. Ediciones
Omega, Barcelona, 4ª ed., 2006.
- LOZANO, J. A., et al: Bioquímica y Biología Molecular. Editorial McGraw-Hill
Interamericana. Madrid, 3ª ed., 2005.
- LOUISOT, P.: Bioquímica Estructural. Editorial AC. España, 1977.
- DEBLIN, T. M.: Bioquímica, libro de texto con aplicaciones clínicas, Editorial
Reverté, S.A. España. 4ª ed., 2004.
- KOOLMAN, J.A., ROHM, K-H.: Bioquímica, texto y atlas. Editorial Médica
Panamericana, 3ª Ed., 2004.
![Cancionero Confirma 2009[1]](https://cdn.vdocuments.co/doc/165x107/55cf8efa550346703b97ac33/cancionero-confirma-20091.jpg)






