
I
UNIVERSIDAD DE GUAYAQUIL UNIDAD DE POSGRADO INVESTIGACIÓN Y DESARROLLO
MAESTRÍA EN BIOTECNOLOGÍA MOLECULAR
TESIS PRESENTADA PARA OPTAR EL GRADO ACADÉMICO DE MAGÍSTER EN BIOTECNOLOGÍA MOLECULAR
“CLONACIÓN DE LA MELITINA ORIGINARIO DE LA ABEJA
(Apis mellifera) EN UN VECTOR DE EXPRESIÓN”
AUTOR (A): DIEGO JAVIER JÁUREGUI SIERRA
TUTOR: SANTIAGO XAVIER MAFLA ANDRADE CO-TUTOR: PhD MIQUEL BLASCO CARLOS
GUAYAQUIL – ECUADOR JUNIO-2016

II
CERTIFICADO DEL TUTOR
REPOSITORIO NACIONAL EN CIENCIA Y TECNOLOGÍA
FICHA DE REGISTRO DE TESIS
TÍTULO Y SUBTÍTULO: Clonación de la melitina originaria de la abeja (Apis mellifera) en un vector de expresión.
AUTOR: Diego Javier Jáuregui Sierra
TUTOR: MSc. Santiago Xavier Mafla Andrade CO-TUTOR: PhD Miquel Blasco Carlos REVISORES:
INSTITUCIÓN: UNIVERSIDAD DE GUAYAQUIL
FACULTAD: Unidad de Posgrado, Investigación y Desarrollo
CARRERA: Maestría en Biotecnología Molecular FECHA DE PUBLICACIÓN:
No. DE PÁGS: 85
TÍTULO OBTENIDO: Magíster en Biotecnología Molecular
ÁREAS TEMÁTICAS: Ingeniería Genética
PALABRAS CLAVE: Apis mellifera; melitina, vector de expresión. RESUMEN: El veneno de abeja está compuesto por varios péptidos uno de ellos es la melitina, que actúa como un bactericida potente y, además, ayuda a mejorar el sistema inmune. El objetivo de este estudio es clonar la melitina originaria de la abeja (Apis mellifera) en un vector de expresión. Para conseguirlo se extrajo el ARNm total de la abeja, después se realizó un RT-PCR para tener un ADNc y posteriormente una PCR con primers específicos con el fin de obtener un ADNc de melitina originario de la abeja e incrustarlo en el vector de expresión pGEM-T easy vector, con el que se transformó una bacteria competente de E. coli JM109 de fácil transformación, dando lugar a colonias blancas que indican la transformación de las células en un medio con IPTG y X-gal. Con este ensayo se pretende conseguir en un futuro un organismo eucariota transgénico que a base de melitina con el objetivo que exprese este gen y sirva para mejorar el sistema inmune de animales y personas. No. DE REGISTRO (en base de datos): No. DE CLASIFICACIÓN:
DIRECCIÓN URL (tesis en la web): ADJUNTO PDF: x SI NO CONTACTO CON AUTOR/ES Teléfono: 0996506235
(06) 2950592 (02) 2533510
E-mail: [email protected] [email protected]
CONTACTO EN LA INSTITUCIÓN: Nombre: Unidad de Posgrado Investigación y Desarrollo
Teléfono: 2325530-38 Ext. 114 E-mail: [email protected]
x

III
En mi calidad de tutor del Programa de Maestría en Biotecnología Molecular,
nombrado por el Director General de la Unidad de Posgrado, Investigación y
Desarrollo, CERTIFICO: que he analizado la Tesis presentada, como requisito
para optar el grado académico de Magíster en Biotecnología Molecular
,titulada: “Clonación de la melitina originaria de la abeja (Apis mellifera) en un
vector de expresión.” la cual cumple con los requisitos académicos, científicos
y formales que demanda el reglamento de posgrado.
Firma -------------------------------------- Santiago Xavier Mafla Andrade C.I. 100265839-9 Guayaquil, 26 de agosto de 2016

IV
CERTIFICACIÓN DE REDACCIÓN Y ESTILO (Nombre del gramatólogo) Título profesional, con registro del SENESCYT xxxxxxxxxxxx, por medio del presente tengo a bien CERTIFICAR: Que he revisado la redacción, estilo y ortografía de la tesis de grado, elaborado (a) por el Señor (a) Diego Javier Jáuregui Sierra, con C.I. 172028207-6, previo a la obtención del título de MAGISTER EN BIOTECNOLOGÍA MOLECULAR. Tema de tesis: “Clonación de la melitina originaria de la abeja (Apis mellifera)
en un vector de expresión.”
Trabajo de investigación que ha sido escrito de acuerdo a las normas ortográficas y de sintaxis vigentes. ------------------------------------------------------ Nombre N° Cédula Registro SENESCYT
Número de teléfono Correo

V
DECLARACIÓN JURADA DEL AUTOR Yo, Diego Javier Jáuregui Sierra, declaro bajo juramento ante la Dirección de Posgrado de la Universidad de Guayaquil, que el trabajo aquí descrito, así como sus resultados, conclusiones y recomendaciones presentadas es de mi autoría y exclusiva responsabilidad, que es inédita y no ha sido previamente presentado para ningún grado o calificación profesional. La reproducción total o parcial de esta tesis en forma idéntica o modificada, no autorizada por los editores transgrede los derechos de autoría. Cualquier utilización debe ser previamente solicitada a la Universidad de Guayaquil, a través de la Dirección de Posgrado o al autor. Firma. ------------------------------------------------ DIEGO JÁUREGUI C.I. 172028207-6

VI
ÍNDICE
INTRODUCCIÓN .................................................................................................... 13
HIPÓTESIS: ........................................................................................................... 13
OBJETIVOS: .......................................................................................................... 13
OBJETIVO GENERAL: ........................................................................................... 13
OBJETIVOS ESPECÍFICOS: .................................................................................. 14
DESCRIPCIÓN DEL PROYECTO .......................................................................... 15
CAPÍTULO I ............................................................................................................ 17
1. EL PROBLEMA ............................................................................................ 17
CAPÍTULO II ........................................................................................................... 22
2. JUSTIFICACIÓN .......................................................................................... 22
CAPÍTULO III .......................................................................................................... 26
3. MARCO TÉORICO ...................................................................................... 26
3.1. LA ABEJA ............................................................................................. 26
3.2. VENENO DE ABEJA ................................................................................ 27
3.3. LA MELITINA ........................................................................................ 30
3.3.1. ESTRUCTURA DE LA MELITINA...................................................... 31
3.3.2. MODO DE ACCIÓN .......................................................................... 33
3.3.3. ACTIVIDAD BIOLÓGICA ................................................................... 35
3.3.4. ACTIVIDAD DE TRANSFECCIÓN..................................................... 37
3.3.5. ACTIVIDAD ANTI VIRAL ................................................................... 39
3.3.6. ACTIVIDAD MICROBIOLÓGICA ....................................................... 40
3.4. PÉPTIDOS ANTIMICROBIANOS (AMP) .................................................. 41
3.4.1. CLASIFICACIÓN DE LOS PÉPTIDOS ANTIMICROBIANOS ............ 42
3.4.2. AMP: MECANISMOS DE ACCIÓN .................................................... 43
CAPÍTULO IV ......................................................................................................... 46
4. METODOLOGÍA: ............................................................................................. 46
4.1. Tipo de Investigación: ............................................................................... 46

VII
4.2. Lugar del Ensayo: ..................................................................................... 46
4.3. Materiales y Reactivos: ............................................................................. 46
4.4. Elaboración de primers ............................................................................. 48
4.5. Extracción de ARN ................................................................................... 49
4.5.1. Preparación de la muestra ................................................................. 49
4.5.2. Preparación de lisados de muestras de tejido de abejas. .................. 50
4.5.3. Limpieza del lisado ............................................................................ 50
4.5.4. Purificación del ARN .......................................................................... 51
4.6. Retrotranscripción .................................................................................... 53
4.7. Amplificación por PCR. ............................................................................. 56
4.8. Electroforesis ............................................................................................ 58
4.9. Purificación del ADN ................................................................................. 59
4.10. Inserción de la secuencia del Gen en el pGEM®-T Easy Vector ........... 61
4.10.1. Ligadura del gen de Interés en el Vector de Expresión ...................... 61
4.10.2. Preparación de Medios para el Proceso de Transformación .............. 63
4.10.3. Preparación del medio Caldo Super Optimo con Represión Catabólica (S.O.C) (100 ml) .............................................................................................. 63
4.10.4. IPTG solución madre (0,1 M) ............................................................. 64
4.10.5. Preparación de placas de Caldo de Lysogeny (LB) con ampicilina .... 64
4.10.6. Preparación de placas de LB con ampicilina / IPTG / X-Gal .............. 65
4.11. Transformación de la célula competente ............................................... 65
CAPÍTULO V .......................................................................................................... 68
5. RESULTADOS Y DISCUCIÓN ........................................................................ 68
CONCLUSIONES Y RECOMENDACIONES .......................................................... 75
CONCLUSIONES ............................................................................................... 75
RECOMENDACIONES ....................................................................................... 77
BIBLIOGRAFÍA ....................................................................................................... 78
ANEXOS ................................................................................................................ 83

VIII
ÍNDICE DE TABLAS
Tabla 1. Materiales y reactivos utilizados en el ensayo .......................... 47
Tabla 2. Primers Foward y Reverse para la secuencia del gen de la
Melitina ........................................................................................................ 49
Tabla 3. Componentes de para la formación de la primera cadena de
ADNc. ........................................................................................................... 54
Tabla 4. Componentes sistema de reacción transcripción inversa. ...... 55
Tabla 5 PCR Mix .......................................................................................... 57
Tabla 6 Ligación del gen de Interés al Vector de Expresión. .................. 62
Tabla 7. Medio S.O.C .................................................................................. 63

IX
ÍNDICE DE FIGURAS
Figura 1. Aparato del aguijón y glándulas de la abeja ............................. 27
Figura 2. Diagrama explicativo de usos terapéuticos del veneno de
abeja. ............................................................................................................ 29
Figura 3. Melitina Componente principal del Veneno de Abeja. ............ 30
Figura 4. Estructura helicoidal de la melitina ........................................... 32
Figura 5 Estructura tetrámera de la melitina ............................................ 33
Figura 6. Estructuras de los Péptidos Antimicrobianos (AMP). ............ 43
Figura 7. Mecanismos de Acción de los AMP en la célula. ..................... 45
Figura 8 Cantidad de ARN de Melitina ...................................................... 68
Figura 9 Gen de interés en electroforesis a distintas temperaturas de
hibridación (Tm). ......................................................................................... 69
Figura 10. Cantidad de ARN de Melitina ..........¡Error! Marcador no definido.
Figura 11 Electroforesis en gel de agarosa con dos tipos de muestras:
1) Control positivo, 2) ADNc de melitina originario de la abeja
(Apis mellifera) purificado. ......................................................................... 71
Figura 12. Representación Esquemática del vector de Expresión pGEM
Easy Vector con el gen de interés. ............................................................ 72
Figura 13. Representación Esquemática de la transformación del vector
de Expresión pGEM Easy Vector con el gen de interés en la célula
competente de E coli JM109. ..................................................................... 73

X
RESUMEN
La abeja occidental (Apis mellifera) es un insecto 'eusociale', que pertenece a
la familia de las Hymenópteras. Las poblaciones se diferencian en reinas que
producen crías y trabajadores no reproductivos, los cuales recogen y procesan
los alimentos y producen la apitoxina. El veneno de abeja está compuesto por
varios péptidos, uno de los cuales es la melitina, siendo este una proteína que
posee un efecto bactericida y ayuda a mejorar el sistema inmunológico.
El objetivo de este estudio ha sido clonar la melitina originaria de la abeja en
un vector de expresión con el fin de transformar una célula competente. Para
esto se extrajo el ARNm total de la abeja, obteniendo una concentración de
47.4ng/µl, con el que se realizó un RT-PCR para obtener un ADNc, dando una
concentración de 78ng/ µl y posteriormente una PCR con primers específicos
y a temperaturas de hibridación distintas (Tm), con el fin de obtener un ADNc
de melitina originario de la abeja e incrustarlo en el vector de expresión pGEM-
T® easy vector. Posteriormente, con el vector se transformaron bacterias
competentes de E. coli JM109® de fácil transformación, con lo que se
consiguieron colonias blancas que indicaban el éxito de la transformación en
un medio con IPTG y X-gal.
Con este ensayo se pretende conseguir en un futuro un organismo eucariota
transgénico con el gen de la melitina, con el objetivo que exprese este gen y
sirva para mejorar el sistema inmune de animales y personas.
Palabras claves: Apis mellifera; melitina, vector de expresión, transformación.

XI
ABSTRACT
Western honey bee (Apis mellifera) is an insect 'eusociale', which belongs to
the family of Hymenópteras. Populations differ in queens produce offspring and
non-reproductive workers who collect and process food and produce apitoxina.
Bee venom is composed of several peptides like melittin, which is a protein
having a bactericidal effect and improving the immune system. The objective
of this study was to clone the native melittin bee in an expression vector, to this
total mRNA bee, with whom further RT-PCR was performed to obtain a cDNA.
The PCR was carried out with specific primers and various hybridization
temperatures (Tm): Tm1 at 68 ° C; Tm2 to 68.2 ° C; Tm3 to 68.4 ° C; Tm4 to
68.6 ° C; TM5 to TM6 68.8 ° C and 69 ° C. Tm3 produced the highest
concentration of amplified DNA. This cDNA was embed into the expression
vector pGEM-T® easy vector, to transform, after that, bacteria E. coli JM109®.
The delivery of white colonies indicated the transformation of cells in medium
containing IPTG and X-gal. This test is intended to achieve in the future a
transgenic eukaryotic organism with melittin gene in order to enhance the
immune system of animals and people.
Keywords: Apis mellifera; melittin, expression vector, transformation.

13
INTRODUCCIÓN
HIPÓTESIS:
El gen que codifica para la melitina se expresa de manera óptima en un
vector de expresión.
OBJETIVOS:
OBJETIVO GENERAL:
Clonar el gen de la melitina originaria de la abeja (Apis mellifera)
mediante un vector de expresión para introducirla en una célula competente.

14
OBJETIVOS ESPECÍFICOS:
1. Aislar el gen que codifica a la melitina de abejas (Apis mellifera) mediante
técnicas de PCR para insertarlo en una célula competente.
2. Obtener un constructo con el gen de la melitina mediante técnicas
moleculares con el fin de que una célula competente sea transformada.
3. Transformar una célula competente mediante una técnica molecular para
la comprobación del funcionamiento del constructo.

15
DESCRIPCIÓN DEL PROYECTO
La “Clonación de la melitina originaria de la abeja (Apis mellifera) en un
vector de expresión.” es una investigación de tesis para la obtención previa del
título de cuarto nivel de Magíster en Biotecnología Molecular, la misma que
consta de cinco capítulos que incluyen: Problema, Justificación, Marco
Teórico, Metodología, Resultados y Discusión, a más de Introducción,
Conclusiones, Recomendaciones y Bibliografía.
En la Introducción se detalla la hipótesis, los objetivos de la tesis, que
fueron planteados para el desarrollo de la misma y finalmente concluir con una
breve descripción del contenido de los capítulos.
Los objetivos que se plantearon en el estudio fueron los siguientes:
Como Objetivo General: Clonar la melitina originaria de la abeja (Apis
mellifera) en un vector de expresión para introducirla en una célula
competente.
Mientras que los objetivos específicos fueron tres, de los cuales el
primero hace referencia a identificar del gen que codifica a la melitina de abejas
mediante técnicas de PCR para insertarlo en una célula competente, mientras
que el segundo pretende obtener un constructo con el gen de la melitina
mediante técnicas moleculares con el fin de que una célula competente sea
transformada. Y finalmente el tercero tiene como meta transformar una célula

16
competente mediante una técnica molecular para la comprobación del
funcionamiento del constructo.
El primer capítulo describe el problema, donde se analiza mediante
bibliografía a los antibióticos y su resistencia, además de las investigaciones
que se han realizado sobre los péptidos microbianos de este tipo, lo cual
conlleva a enfrentar muchas enfermedades.
El segundo capítulo se justifica la investigación mediante un análisis
bibliográfico exhaustivo, donde se explica las potencialidades y bondades de
la melitina, que realiza en un organismo.
El tercer capítulo concierne al marco teórico, el cual es una recopilación
bibliográfica, que abarca una breve descripción de la abeja (Apis mellifera), así
como del veneno de abeja y generalidades sobre investigaciones de la
melitina, además de sus diferentes tipos de acciones, potencialidades.
El cuarto capítulo indica la metodología empleada para el desarrollo de
este experimento, que va desde la extracción y purificación del ARN total y
pasarlo a ADNc, mediante la técnica de la transcripción reversa, con la ayuda
de primers, para después insertar la secuencia encontrada en un vector de
expresión, y así poder transformar una célula competente con el fin de que
exprese la secuencia encontrada.
El quinto capítulo trata sobre los resultados obtenidos en la
investigación que se detalla en el cuarto capítulo.
Las conclusiones y recomendaciones en base a los resultados de la
investigación por cada objetivo planteado.

17
CAPÍTULO I
1. EL PROBLEMA
Los seres vivos se encuentran constantemente rodeados por agentes
patógenos (radiaciones, toxinas, bacterias, virus, hongos) que pueden debilitar
el sistema inmune y afectar a su salud.
Los agentes que provocan las enfermedades infecciosas y las toxinas,
presentes en las poblaciones animales, constituyen un riesgo para la sanidad
animal, la inocuidad de los alimentos y la salud pública (OIE, 2014). La mayoría
de los brotes de enfermedades y las contaminaciones alimentarias surgen
naturalmente y son también un riesgo real de que se introduzca en poblaciones
humanas o animales susceptibles como consecuencia de la liberación
accidental de un agente infeccioso o de una toxina. Esto acarrea grandes
riesgos ya que estos patógenos, al haber sido manipulados, pueden resultar
aún más peligrosos que los naturales (Park y col, 2014).
Debido a la limitación de vacunas y péptidos antimicrobianos, persisten
algunas enfermedades de gran impacto en la salud mundial. Dentro de estas
enfermedades, unas pocas o bien no cuentan con tratamiento, o bien cuentan
con tratamientos paliativos. Debido a esto, la investigación biomédica es
todavía esencial para mejorar la salud, no solo humana, sino también animal
(Park y col, 2014).

18
Los primeros agentes antimicrobianos eficaces, se introdujeron en 1937 y,
después de 2 años, se informó de la aparición resistencia. Estos mismos
mecanismos de resistencia a antibióticos clínicos se encuentran presentes en
la actualidad (Watkins & Bonomo, 2016).
La administración general como medida profiláctica de los antibióticos ha
aumentado en más del 30% en la década 2001-2011. Esto ha desarrollado la
resistencia de los mismos que sean más frecuentes, dando lugar a la aparición
de enfermedades ya controladas anterior mente (Becattini y col., 2016).
Los antibióticos son unos de los descubrimientos más importantes del siglo
XX y han sido ampliamente utilizados para el tratamiento de enfermedades en
los animales en el siglo XXI. Sin embargo, la resistencia a los antibióticos entre
patógenos bacterianos es una preocupación generalizada con respecto a su
uso en animales (Wang y col, 2016).
La adquisición de resistencia a los antibióticos modifica las interacciones
ecológicas dentro de una comunidad bacteriana homogénea, contribuyendo a
su propio bienestar. En ambientes bien mezclados, donde coexisten diferentes
taxones microbianos en las proximidades, cepas distintas generalmente tienen
la capacidad de producir antibióticos para inactivar conjuntos específicos de
moléculas antibacterianas (Becattini y col., 2016).
Una manera útil de entender los mecanismos básicos de la resistencia a
antibióticos es a través del concepto de bala y de destino, por el que los sitios
de actividad de la droga (el blanco) pueden ser cambiados por modificación
enzimática, transformados por mutaciones genómicas o pasando por alto

19
metabólicamente. Así pues, el antibiótico (la bala) puede someterse a la
inactivación enzimática y a la degradación, la reducción del acceso a la célula
(por ejemplo, pérdida de porinas) y el aumento de su eliminación por parte de
la célula (por ejemplo, bombas de expulsión) (Watkins & Bonomo, 2016).
Las Mutaciones Novo forman parte de la generación a la Resistencia a los
antibióticos, donde los antibióticos ejercen una presión selectiva que impulsa
el rápido desarrollo de cepas resistentes. Este proceso generalmente requiere
de múltiples mutaciones en el ADN (Becattini y col., 2016).
El mal uso de los antibióticos plantea muchas preocupaciones ya que
existe una trayectoria evolutiva en la adquisición de mutaciones que confieren
resistencia a los antibióticos (Becattini y col., 2016).
Uno de los factores que causan resistencia a los antibióticos son los
Biofilms, que son comunidades de células bacterianas agregadas, embebidas
en una matriz polimérica extracelular de producción propia. Estas biopelículas
son recalcitrantes contra el tratamiento de antibioticos y las defensas inmunes
que están implicadas en infecciones bacterianas y fúngicas (Hathroubi y col.,
2016).
El suministro de antibióticos podría llevar a una alteración de la
composición y funciones de la microbiota, que pueden producir efectos nocivos
para el huésped (Becattini y col., 2016).

20
La rápida y progresiva resistencia a los antibióticos representa una seria
amenaza para la salud pública mundial. El uso indiscriminado de antibióticos
en la agricultura y la medicina humana, junto con sociedades cada vez más
interconectadas han alimentado la aparición de bacterias resistentes a los
antibióticos. Estos factores juntos han dado lugar a un número de infecciones
causadas por bacterias resistentes a múltiples fármacos y, junto a este
aumento de la resistencia, se está dando también un aumento de la morbilidad
y la mortalidad (Watkins & Bonomo, 2016).
Los antibióticos se utilizan ampliamente en la ganadería en dosis bajas con
el objetivo principal de mejorar el crecimiento animal. Esto da como resultado
la selección de bacterias resistentes que pueden infectar a los humanos
mediante la resistencia a los patógenos por transferencia horizontal (Becattini
y col., 2016).
La resistencia a antibióticos en la agricultura podría poner en peligro la
salud humana en tres casos posibles:
(1) Un ser humano es infectado por un patógeno resistente a través de
zoonosis o por la ingestión de bacterias en alimentos o agua contaminados.
(2) Un ser humano es colonizado por bacterias resistentes a través de uno
de estos medios y luego se extiende a otra persona que posteriormente se
enferma.

21
(3) Los genes de resistencia que surgen en la agricultura se transmite a los
humanos a través de la transferencia horizontal de genes y las cepas
resistentes resultantes son seleccionadas por el uso de antibióticos en las
personas (Watkins & Bonomo, 2016).
En este proyecto se plantea una aportación a este campo de investigación
mediante la creación de un constructo que contiene el gen codificante de la
melitina (Linguo y col., 2014; Kim y col., 2015).
Uno de los genes diana de este campo de la investigación es el gen
codificante de la melitina, que es un oligopéptido de 26 aminoácidos y que
forma el principal componente de la apitoxina de la abeja, el cual, como se
describe más adelante, tiene múltiples efectos terapéuticos, incluyendo el de
antibiótico natural (Fenell y col., 1967; Fenell y col., 1968; Choi y col., 2015;
Leandro y col., 2015) e incluso el de agente antitumoral (Linguo y col., 2014;
Kim y col., 2015).
En este proyecto se plantea la creación de un constructo con la secuencia
parcial del gen que codifica a la melitina, lo cual abre muchas posibilidades
para su aplicación terapéutica, como la posibilidad de obtener bacterias
superproductoras de este oligopéptido, o incluso del mejoramiento vegetal
mediante la inserción de éste constructo en callo embriogénico de plantas
alimentarias que podrían dar lugar a vegetales productores de melitina.

22
CAPÍTULO II
2. JUSTIFICACIÓN
Los animales cumplen una función importante como sensores biológicos
de las liberaciones, accidentales o deliberadas de toxinas y agentes
infecciosos y de enfermedades emergentes (OIE, 2014).
En el caso de enfermedades zoonóticas, es esencial contar con una
respuesta coordinada entre los responsables de la sanidad animal y la salud
pública, puesto que los procedimientos de control se suelen orientar hacia la
eliminación del agente patógeno en la fuente animal (OIE, 2014). Con el fin de
establecer la causa del brote de enfermedad, es necesario que tanto las
autoridades sanitarias como los laboratorios de investigación realicen las
pertinentes investigaciones y se desarrollen, además, nuevos mecanismos de
control de estas enfermedades.
Muchos péptidos antimicrobianos ya se han estudiado tanto in vivo como
in vitro. Ventajas obvias de péptidos antimicrobianos que más de antibióticos,
en el tratamiento de enfermedades por su amplio espectro de actividad, su
eficacia en concentraciones micromolares, la práctica imposibilidad de
desarrollo de resistencia de los microbios y la oportunidad de sintetizar
análogos de péptidos naturales con propiedades (Lazarev y col., 2004).

23
El uso de antibióticos para el ganado ha sido criticado y la FDA ha
recomendado evitar los innecesarios, pero esto sigue siendo una política
voluntaria. Es de destacar que se ha calculado que la prohibición de este
procedimiento no aumentaría significativamente los costos para los
productores o consumidores (Becattini y col., 2016).
Dado el vínculo entre la resistencia a antibióticos y su uso en animales de
granja, una posible solución es prohibirlos, con el fin de controlar a organismos
menos resistentes en el ambiente, sin embargo un mejor argumento puede ser
la reducción de la prevalencia de patógenos resistentes y, por lo tanto,
menores son las posibilidades para la transferencia horizontal de genes de
resistencia (Watkins & Bonomo, 2016).
Diversos materiales tales como enzimas, prebióticos, probióticos,
minerales, péptidos antimicrobianos, acidificantes, plantas y extractos de
plantas, han sido probados como posibles antibióticos alternativos contra la
alteración que causan los antibióticos sobre la microbiota intestinal y la función
inmune. Por este motivo, nuevas investigaciones se están enfocado sobre la
aplicación de estos suplementos en la alimentación (Wang y col., 2016).
Así pues, actualmente se está investigando sobre las alternativas a los
antibióticos en la cría de animales, que se espera que limiten los efectos
adversos y garanticen la seguridad de los alimentos de origen animal y el
ambiente (Wang y col., 2016).
La creciente resistencia a los antibióticos se ha convertido en un importante
problema de salud y ha alentado a muchos investigadores a encontrar clases

24
de antibióticos alternativos. En este contexto, los péptidos antimicrobianos
(AMP) han aparecido como nuevos agentes prometedores para combatir a los
patógenos (Mularski y col., 2015; Reinhardt & Neundorf, 2016).
Por este motivo, los AMP han acaparado cada vez más el foco de las
investigaciones, aunque con el primer uso clínico de la AMP, la posibilidad de
desarrollo de cepas resistentes a AMP es inevitable y está siendo
intensamente investigado (Reinhardt & Neundorf, 2016).
Durante los últimos años, muchos esfuerzos se han hecho en la
identificación y desarrollo de AMP sintéticos. Estos han sido dirigidos hacia el
acortamiento del tamaño y, por lo tanto, la optimización de la estabilidad
metabólica, biodisponibilidad y las cuestiones relativas a la seguridad y la
inmunogenicidad. Además, las secuencias más cortas disminuyen
drásticamente los costos de producción (Reinhardt & Neundorf, 2016).
Otro factor interesante de los AMPs, además de su actividad
antimicrobiana, es su poca toxicidad hacia las células de los animales (Peters
y col., 2010). También la evolución de la resistencia contra los péptidos
antimicrobianos ha sido considerada poco probable debido a sus diversos
mecanismos de acción (Johnston y col, 2016).
Otra de las posibles aplicaciones de los AMPs es la combinación con otras
sustancias antimicrobianas con el fin de encontrar efectos sinérgicos; así los

25
AMPs alterarían las membranas del patógeno permitiendo que otras
moléculas, como los antibióticos clásicos, penetren más fácilmente y
ejercieran su acción bactericida (Johnston y col., 2016).
Por todos estos motivos, la ciencia Biomédica está centrándose
actualmente en la obtención de nuevas formas de tratar enfermedades
infecciosas mediante el uso de estos péptidos. Muchas familias de péptidos
antimicrobianos han sido clasificados (Lazarev y col., 2004), incluyendo a la
melitina a partir de veneno de abeja. Mediante estudios de biotecnología se
han determinado algunos péptidos antimicrobianos derivados de insectos
(AMPS) que podrían ayudar a combatir infecciones, inflamaciones e incluso,
en algunos casos, el cáncer (Park y col., 2014).
Otra posibilidad sobre la que se está trabajando hoy en día es diseñar
AMPs sintéticos con actividad microbiológica mejorada para células
microbianas, especialmente aquellos con resistencia a los antibióticos, así
como efectos sinérgicos con agentes antibióticos convencionales que carecen
de actividad citotóxica o hemolítica (Park & Hahm, 2005).

26
CAPÍTULO III
3. MARCO TÉORICO
3.1. LA ABEJA
Weinstock y col. (2006) describen la abeja occidental (Apis mellifera), como
un insecto 'eusociale', que pertenece al orden de las Hymenópteras y a la
familia de la Apidae (Biló y col., 2005). Existe una distinción de roles dentro de
esta especie, donde las reinas son las encargas de producir crías, las
trabajadoras no reproductivas recogen y procesan los alimentos y los insectos
jóvenes hacen sus nidos y defienden las colonias.
Su aguijón que tiene múltiples barbillas en la parte posterior del cuerpo y
está en comunicación con glándulas del veneno (Figura 1), las cuales se
atascan y ocasionan que parte del intestino con el sistema glandular y el saco
del veneno se desprenda del insecto al intentar sacar el mismo del tegumento
de su víctima provocando posteriormente la muerte de la abeja (Peña y col.,
2006). La glándula de veneno se compone de uno o más filamentos secretores
que se abren en un depósito en el que se almacena veneno.
Las abejas, generalmente obreras, producen un veneno llamado apitoxina,
que consta de muchos péptidos y proteínas tóxicas que funciona como un
arma eficaz para la defensa y su organización morfológica (Peiren y col.,
2008).

27
En la abeja occidental, la glándula de veneno está presente no solo en las
obreras sino también en las castas de matrimonio e incluso en las reinas, las
cuales tienen glándulas significativamente más grandes que las trabajadoras
(Peiren y col., 2008).
FIGURA 1. APARATO DEL AGUIJÓN Y GLÁNDULAS DE LA ABEJA
Fuente: (Apicola el Nebrillo, 2011)
3.2. VENENO DE ABEJA
El veneno de abeja es un arma única en el reino animal primordial en la
defensa de la colonia de abejas. Dicho veneno está formado por una mezcla
compleja de proteínas eficientes destinadas a proteger a las abejas contra una
amplia diversidad de depredadores (Han & Lee, 2012; Oršolić, 2012).

28
El veneno de abeja se encuentra en la cavidad abdominal (dentro de una
glándula) y contiene contiene al menos 18 componentes activos (Anexo 3),
incluyendo enzimas, péptidos, y aminas biogénicas, que tienen una amplia
variedad de propiedades farmacéuticas, incluyendo la melitina, apamina,
adolapina, el péptido de mastocitos degranulación (MCD), enzimas (la
fosfolipasa [PLA] ), aminas biológicamente activas (histamina y epinefrina) y
componentes no peptídicos que tienen una variedad de propiedades
farmacéuticas (Han & Lee, 2012; Oršolić, 2012).
Posee diversas actividades biológicas y farmacológicas (Matysiak y
col., 2001) entre las cuales destaca la capacidad de modificar las funciones
del sistema inmune en el cuerpo (Son y col., 2007).
Su eficacia farmacológica se ha demostrado en muchas enfermedades,
siendo usada en inmunoterapia (Han & Lee, 2012) e incluso como un
cosmético para las funciones antienvejecimiento, antiinflamatorios y
antibacterianos.
El veneno de abeja contiene varios compuestos tóxicos que causan la
muerte o inflingen dolor, lo que les permite defender la colmena contra los
depredadores y las amenazas externas.
En los mamíferos, la exposición al veneno de abeja provoca la
producción de IgG1, IgG2 y, en menor medida, las respuestas de anticuerpos
IgG4 de la respuesta humoral. Mientras que la alergia a la picadura de una
abeja está mediada por anticuerpos IgE (Van Vaerenbergh y col., 2013).

29
El veneno de abeja se está usando como un fármaco anti-inflamatorio
no esteroideo usado para el alivio del dolor y el tratamiento de enfermedades
inflamatorias crónicas, como la artritis reumatoide y la esclerosis múltiple, así
como también en tratamiento de tumores. El papel beneficioso que juega el
veneno de abeja en la salud del ser humano es: anti-mutagénica,
antiinflamatorios, antinociceptivos y actividades radioprotectoras contra el
cáncer. Además, actúa sobre la inducción de la apoptosis y la necrosis y
efectos sobre la proliferación, citotoxicidad y la inhibición del crecimiento de
diferentes tipos de células del cáncer (Oršolić, 2012) (Figura 2).
FIGURA 2. DIAGRAMA EXPLICATIVO DE USOS TERAPÉUTICOS DEL VENENO DE
ABEJA.
Fuente: (Oršolić, 2012).
Efecto de los componentes de veneno de abeja sobre el complejo calmodulina (Ca2 + / CaM)
y NF-kappa B (NF-kB) y su interection sobre la inmunidad y el crecimiento tumoral y su
supervivencia.
30
3.3. LA MELITINA
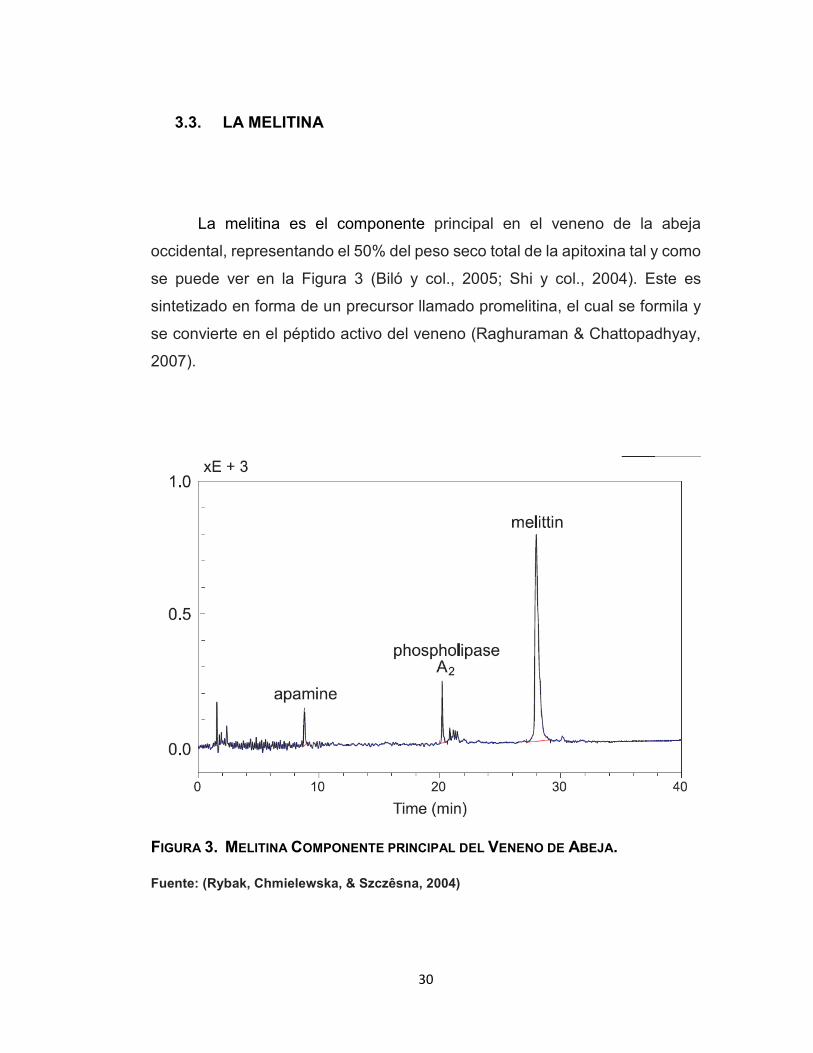
La melitina es el componente principal en el veneno de la abeja
occidental, representando el 50% del peso seco total de la apitoxina tal y como
se puede ver en la Figura 3 (Biló y col., 2005; Shi y col., 2004). Este es
sintetizado en forma de un precursor llamado promelitina, el cual se formila y
se convierte en el péptido activo del veneno (Raghuraman & Chattopadhyay,
2007).
FIGURA 3. MELITINA COMPONENTE PRINCIPAL DEL VENENO DE ABEJA.
Fuente: (Rybak, Chmielewska, & Szczêsna, 2004)

31
Los principales componentes que contiene el veneno de abeja es la melitina con el 65 %, la
apamina con el 3.13% y la fosfolipasa A2 con un 12.98%.
La separación, identificación y determinación cuantitativa de los
principales constituyentes del veneno de las abejas por cromatografía líquida
de alto rendimiento (HPLC) muestra distintos compuestos principales de las
proteínas: la melitina, fosfolipasa A2 y apamina, donde el contenido melitina
tiene un promedio de 64,40% con relación a las demás proteínas (Rybak y col.,
2004).
3.3.1. ESTRUCTURA DE LA MELITINA
Es una molécula peptídica líneal
(GIGAVLKVLTTGLPALISWIKRKRQQ-NH2), compuesta de 26 residuos de
aminoácidos, que tiene como características que es anfipática, catiónica y
presenta función hemolítica (Figura 4). La región amino-terminal de esta
cadena polipeptídica es predominantemente hidrófoba, mientras que la región
carboxi-terminal es hidrófila, debido a la presencia de un tramo de aminoácidos
cargados positivamente (Raghuraman & Chattopadhyay, 2007) y, debido a
esta anfipatía, es capaz de interactuar con las membranas biológicas (Huang,
2013).

32
FIGURA 4. ESTRUCTURA HELICOIDAL DE LA MELITINA
Fuente: (Raghuraman & Chattopadhyay, 2007)
La estructura tridimensional de la melitina forma un tetrámero donde los
cuatro monómeros de melitina son casi idénticos en la conformación (amarillo,
azul, verde, turquesa). Cada cadena de melitina se compone de dos
segmentos α-helicoidales y la forma general es la de una "varilla doblada” en
un ángulo aproximado de 120° (Figura 5). La gran curva hélice permite el
embalaje óptimo de cadenas laterales hidrofóbicas dentro del tetrámero
melitina. Debido a la distribución asimétrica de las cadenas laterales polares y
apolares, la cadena de melitina tiene una región terminal hidrófoba amino, una
región central con las caras hidrófobas e hidrófilicas y una región totalmente
hidrófila C-terminal (puntos azules) (Raghuraman & Chattopadhyay, 2007).

33
FIGURA 5 ESTRUCTURA TETRÁMERA DE LA MELITINA
Fuente: (Raghuraman & Chattopadhyay, 2007)
3.3.2. MODO DE ACCIÓN
3.3.2.1. Acciones de melitina en las membranas
Actividad hemolítica
La característica de la melitina es su actividad hemolítica, que tiene
como objetivo la unión de la melitina con la membrana de los eritrocitos. Esta
se une rápidamente a la membrana de los eritrocitos e induce la liberación de
hemoglobina en el medio extracelular, siendo el sitio primario de la interacción

34
la de lípidos de membrana en lugar de receptores específicos. Se ha
demostrado que la melitina induce a la hemólisis a temperatura ambiente,
mediante un mecanismo osmótico coloidal. Esta conclusión se basa en la
observación de que la permeabilidad de los iones es rápida durante la
exposición a la melitina y la liberación de hemoglobina es secundaria, dando
formación a poros (Raghuraman & Chattopadhyay, 2007).
La hemolisis consta de fases rápidas y lentas que oscilan entre los 4° y
37° C, respectivamente. Curiosamente, los tipos de las fases cinéticas rápidas
y lentas son dependientes de la concentración de la melitina. La fase rápida
se interpreta como el resultado de la perturbación de la estructura y
organización de la membrana debido a la rápida acumulación de melitina en
el exterior de la membrana de los eritrocitos, y su descomposición en una fase
lenta como resultado de la reorganización de los péptidos y lípidos de
membrana para recuperar una geometría de embalaje favorable
(Raghuraman & Chattopadhyay, 2007).
3.3.2.2. Formación de canales dependientes de voltaje.
La melitina interrumpe la función de la barrera de las membranas
celulares al formar canales, induciendo al aumento de la permeabilidad de los
iones en las membranas lipídicas. Los poros formados muestran selectividad
para aniones más de cationes, debido a la acumulación de cargas positivas en
la región C-terminal de la melitina. Curiosamente la melitina muestra que sus
cargas positivas en la región amino terminal desempeñan un papel importante
pero no exclusiva en el gating de voltaje de los canales de melitina en bicapas
(Raghuraman & Chattopadhyay, 2007).

35
3.3.2.3. Formación de micelas y la fusión de las bicapas.
La permeabilización inducida por la melitina en membranas causa la
ruptura de las membranas en micelas a alta concentración. Curiosamente, la
formación de micelas de melitina se produce específicamente en las
membranas compuestas de fosfatidilcolinas saturadas, teniendo una
dependencia de la temperatura de transición de fase de lípidos. Sin embargo,
la presencia de cantidades intermedias de melitina induce al liposoma a
fragmentarse en pequeñas partículas (micelas). Así pues, la melitina induce a
una reorganización de los conjuntos de lípidos que incluyen vesicularization
de multibiliares, la fusión de pequeñas vesículas de lípidos y la fragmentación
en discos y micelas.
3.3.3. ACTIVIDAD BIOLÓGICA
La melitina tiene capacidad para afectar a la dinámica de las proteínas
de membrana, provocando la agregación de estas e inmovilizándolas en el
plano de la bicapa lipídica. Además, es un péptido formador de poros que
induce la permeabilización de la membrana y puede incluso llegar a romperla
(Husseneder y col., 2016). También actúa como potente inhibidor de
Ca2+ATPasa; H+K+ATPasa; Na+K+ATPasa y de la proteína quinasa
(Raghuraman & Chattopadhyay, 2007).
La melitina es uno de los péptidos más utilizados para comprender las
interacciones lípido-proteína a nivel molecular, esto se debe a su tamaño
pequeño y su fácil disponibilidad en la naturaleza (Raghuraman &
Chattopadhyay, 2007).

36
El péptido citolítico melitina es un candidato potencial contra el cáncer
que puede ser capaz de superar la resistencia a fármacos de ciertos tumores
debido a sus propiedades líticas (Huang, 2013). De hecho, provoca la
inducción de la muerte de células cancerosas por apoptosis mediante la
activación de la caspasa y metaloproteinasas de la matriz (Oršolić, 2012).
El polimorfismo genético que presentan estos péptidos tiene una
influencia directa e indirecta sobre sus efectos. De hecho, se han identificado
nueve polimorfismos en la región codificante de la melitina de las poblaciones
de abejas; uno de ellos da lugar a la sustitución de serina (Ser) por asparagina
(Asp), afectando potencialmente a las actividades biológicas del péptido. Así
pues, con serina, la melitina mostró un mayor efecto citotóxico que con
asparagina contra E. coli (Park y col., 2014). Además, ambas variedades
presentaban diferentes efectos inhibidores sobre la producción de factores
inflamatorios tales como la interleucina IL-6 y la citocina TNF-α en células BV-
2 de la microglia (Park y col, 2014).
La melitina y la fosfolipasa A2 (PLA2) tienen la capacidad de inducir la
necrosis en células del músculo (Fletcher y col., 1990), ya que les
inducecambios tempranos relacionados con la ruptura de la membrana
plasmática y la hipercontracción de las miofibrillas. Estos resultados indican
que tanto la melitina y la PLA2 del veneno de abeja son capaces de inducir a
actividades catalíticas y miotóxicas. Además, pueden actuar sinérgicamente
para inducir un efecto miotóxico más fuerte (Ownby y col., 1997).

37
En un estudio se demuestra la capacidad de la melitina de hiperactivar
la fosfolipasa A2 (PLA2), en células transformadas con el oncogen ras por la
mediación de una mayor afluencia de iones de calcio (Fletcher y col., 1990).
Tanto la melitina como la hiperactivación mediada por PLA2 requieren de
Ca+2. Sin embargo, la acción de la melitina es fuertemente dependiente del
Ca+2 extracelular. Otros estudios revelan una estrecha correlación entre el
grado de hiperactivación de PLA2 y la movilización de Ca+2, lo que sugiere una
relación causal (Sharma, 1993).
3.3.4. ACTIVIDAD DE TRANSFECCIÓN
Los péptidos antimicrobianos forman canales en la membrana celular
de los microbios, seguido por su fragmentación, para lo cual no hay receptores
específicos de membrana que unan estos péptidos que se han encontrado en
esta estructura celularla membrana celular (Lazarev y col., 2004).
La entrada de ADN exógeno en el citoplasma y el transporte posterior
hasta el núcleo son las principales barreras celulares para los vectores de
suministro de genes no virales (Lazarev y col., 2004).
Se ha demostrado que la melitina permite una liberación eficaz de las
partículas de transferencia de genes no virales en el citoplasma, debido a su
actividad lítica de membrana, por lo que ha sido utilizada con éxito para
mejorar la transfección de complejos de ADN en una amplia gama de líneas
celulares (Oršolić, 2012).

38
Algunos estudios sugieren que la melitina de origen natural posee una
doble funcionalidad endosomolítico y nuclear-homing, que puede formar la
base de un agente de transfección potente y hace que sea un candidato
interesante para el desarrollo de la administración génica sistémica in vivo
(Oršolić, 2012). Además, los policationes de la melitina poseen función de
endosomolisis, lo cual permite una eficiente transfección de ARNm no viral de
las células quiescentes y post-mitóticas (Raghuraman & Chattopadhyay,
2007).
Se han empleado técnicas de ingeniería genética para crear
construcciones con este gen tan interesante desde el punto de vista
farmacológico. Así, se ha sintetizado un hibrido del gen cecropina A
(codificante del alcaloide cecoprina) con el de la melitina, e insertado en el
plásmido pGEMEX-1. Se ha infectado E. coli con esta construcción y se ha
determinado positivamente su expresión génica (Zhu y col., 2002).
Además, con el objetivo de obtener un nuevo péptido antibacteriano, se
ha sintetizado un gen codificante para un péptido híbrido LfcinB - melitina,
compuesto de 1 a 15 residuos de aminoácidos de lactoferricina bovina y
residuos de aminoácidos de melitina. Este gen ha sido insertado en el
plásmido pET -32a para su expresión en E. coli (Bi y col., 2009).
En un estudio realizado por Vutto y col. (2010), se han obtenido vectores
binarios construidos para expresar los genes con péptidos antimicrobianos
(APs) del tipo cectropin melitina, bajo el control de un promotor del virus del
mosaico de la coliflor 35S ARN, para la transformación mediada por

39
Agrobacterium de plantas con el objetivo de que estas tuvieran propiedades
antimicrobianas.
Además, se ha desarrollado una levadura genéticamente modificada,
con capacidad de síntesis de melitina, y se ha demostrado su capacidad de
aniquilación de los protozoos del intestino de las termitas mediante su ingesta.
Como efecto colateral, la melitina causó debilitamiento en el intestino, pero no
se observó un efecto sinérgico con la acción bactericida en los protozoos por
la necrosis intestinal (Husseneder y col., 2016).
Estudios han demostrado la capacidad de la melitina para para ingresar
con éxito a las células competentes y así transformalas. (Sharma, 1993).
3.3.5. ACTIVIDAD ANTI VIRAL
Se ha demostrado que la melitina reduce la producción de VIH-1 de una
manera dependiente de la dosis. Además, se ha indicado que la melitina tiene
un efecto supresor directo sobre la actividad del VIH (Raghuraman &
Chattopadhyay, 2007) y que los péptidos de melitina citolíticos podrían actuar
como virucidas si son transportados mediante un constructo inserto en un
vector, inhibiendo la infectividad del VIH-1 (Hood y col., 2013).

40
En otro estudio se ha demostrado que la melitina inhibe la infección por
el virus de la planta de tabaco, y se han diseñado análogos sintéticos de
melitina que tienen secuencia y similitudes estructurales a un dominio esencial
de la cubierta proteica del virus del mosaico del tabaco, con actividad antiviral
altamente específica (Raghuraman & Chattopadhyay, 2007).
Sin embargo, las aplicaciones in vivo de la melitina están limitadas
debido a su principal efecto secundario, la hemólisis, que está especialmente
pronunciada después de la administración intravenosa (Huang, 2013).
3.3.6. ACTIVIDAD MICROBIOLÓGICA
Se realizaron mediciones biofísicas in situ mediante una Microscopia de
Fuerza Atómica para entender cómo el péptido melitina modula varios
comportamientos biofísicos en bacterias individuales de Klebsiella
pneumoniae. El péptido tuvo un efecto significativo en la presión de turgencia
y el módulo de la pared celular (Mularski y col., 2015).
Se ha visto que la adición de melitina aumenta la presión de turgencia de
la célula, seguida posteriormente de una disminución en la presión de esta, lo
que sugiere que se produce lisis celular y debido a la destrucción de la

41
envoltura celular de la bacteria. De hecho, la cápsula es afectada por la
exposición al péptido, así como su grosor (Mularski y col., 2015).
3.4. PÉPTIDOS ANTIMICROBIANOS (AMP)
Los péptidos antimicrobianos (AMP) son polipéptidos esenciales en la
defensa del huésped y juegan un papel importante en el sistema inmune
innato. Ellos actúan contra un amplio espectro de organismos, tales como
bacterias Gram-positivas y Gram-negativas, así como hongos, parásitos y
virus. Presentan un modo de acción único que está principalmente relacionado
con sus propiedades anfipáticas catiónicas, haciéndolos capaces de
permeabilizar las membranas microbianas (Reinhardt & Neundorf, 2016).
Los péptidos antimicrobianos (AMP) se han aislado y caracterizado a partir
de tejidos y organismos que representan prácticamente todos los reinos y
phylums. Su composición de aminoácidos, anfipaticidad, carga catiónica, y las
dimensiones les permiten adherirse e insertarse en bicapas de membrana para
formar poros de 'barril duela', 'alfombra' o 'poro toroidal' (Park & Hahm, 2005).
Los péptidos antimicrobianos (AMP) derivados de las abejas, y en especial
la melitina, tienen propiedades antimicrobianas y actividades farmacológicas
tales como anti-inflamación y propiedades contra el cáncer de origen natural
(Park y col., 2014).

42
Los AMPs inducen la formación de poros en la membrana que sería como
una posible vía alternativa para combatir cepas bacterianas resistentes a los
antibióticos (Park & Hahm, 2005).
3.4.1. CLASIFICACIÓN DE LOS PÉPTIDOS ANTIMICROBIANOS
La clasificación de los péptidos antimicrobianos es diversa por la
complejidad en sus moléculas. Se pueden dividir en cuatro grupos (Figura 6)
basándose en características estructurales tales como: α-helicoidal, β-hoja,
extendida y péptidos de bucle (Reinhardt & Neundorf, 2016). La mayoría de
los péptidos α-helicoidales comparten un carácter anfipático con una carga
neta positiva y un contenido alto (50%) de residuos hidrófobos (Reinhardt &
Neundorf, 2016).
La estructura de AMP-ß sheet- o similares a hebra beta, propia de las
familias de la defensina y protegrina, se caracteriza por la presencia de dos o
más puentes disulfuro que estabilizan su conformación (Reinhardt & Neundorf,
2016).
AMP a menudo contienen un alto porcentaje de residuos de prolina,
triptófano, arginina e histidina en su secuencia primaria. En la mayoría de los
casos, estos péptidos forman sólo las estructuras secundarias irregulares
(Reinhardt & Neundorf, 2016).

43
El grupo más pequeño de los AMPs son los péptidos altamente estables
que exhiben una estructura de bucle en horquilla (loop) la cual está
interconectada por al menos un puente disulfuro (Reinhardt & Neundorf, 2016).
Figura 6. Estructuras de los Péptidos Antimicrobianos (AMP).
Fuente (Peters y col., 2010)
3.4.2. AMP: MECANISMOS DE ACCIÓN
Muchos AMPs son activos de membrana, pero su modo de acción
consiste en matar bacterias o inhibir su crecimiento (Mularski y col., 2015) y

44
emplean mecanismos y dinámicas de acción para llevar a cabo sus posibles
papeles en la defensa del huésped contra el microbio (Park & Hahm, 2005).
Su actividad antimicrobiana puede neutralizar a una amplia gama de
microbios, incluidos virus, bacterias, protozoos y hongos (Peters y col., 2010).
La formación de poros transmembranales no es el único mecanismo de
destrucción microbiana por parte de los AMPs. De hecho, varias
observaciones sugieren que la traslocación de los AMPs puede alterar la
formación de la membrana citoplasmática, reducir la pared celular, el ácido
nucleico, la síntesis de proteínas, e inhibir la actividad enzimática (Park &
Hahm K, 2005).
Los AMP por ser de naturaleza catiónica se sienten atraídos por las
cargas negativas de las membranas externas microbianas, soportando una
interacción altamente selectiva, lo que permite al AMP atacar selectivamente
las membranas microbianas. Sin embargo, la toxicidad de los AMP contra las
células se debería a la modulación de parámetros físico-químicos como la
hidrofobicidad, la carga neta y la helicidad (Reinhardt & Neundorf, 2016).
Por lo general, los AMP adoptan estructuras secundarias bien definidas
cuando entran en contacto con las membranas de los patógenos. Después de
este paso inicial de la unión por fuerzas electrostáticas, se lleva a cabo la
permeabilización de la membrana, en donde las membranas bacterianas
finalmente son interrumpidas. Esto induce varios procesos, por ejemplo, la
ruptura de la membrana y la filtración de componentes intracelulares, lo que
lleva a la muerte celular (Reinhardt & Neundorf, 2016) (Figura 7).

45
FIGURA 7. MECANISMOS DE ACCIÓN DE LOS AMP EN LA CÉLULA.
Fuente (Peters, Shirtliff, & Jabra-Rizk, 2010)
A Membrana celular (1) los AMPs se insertan al azar en la membrana, (2) e interaccionan entre
sí a través de sus secuencias hidrofóbicas lo que causa (3) que una parte de la membrana
sea removida y se forme un poro. B Inhibición de la síntesis de DNA. C Bloqueo de la síntesis
de RNA. D Inhibición de las enzimas necesarias para la síntesis la estructura de la pared
celular. E Inhibición de los ribosomas. F Bloqueo de las chaperonas impidiendo el plegamiento
de las proteínas. G Inactivación de la mitocondria. (1) Inhibición de la cadena respiratoria, (2)
ruptura de la membrana mitocondrial.

46
CAPÍTULO IV
4. METODOLOGÍA:
4.1. Tipo de Investigación:
Es de tipo aplicada, experimental y descriptiva.
4.2. Lugar del Ensayo:
Este trabajo se desarrolló en el Laboratorio de Biotecnología de la
Pontificia Universidad Católica del Ecuador - Sede Ibarra (PUCESI)
4.3. Materiales y Reactivos:
Materiales y reactivos que se utilizaron se detallan a continuación en la Tabla
1

47
TABLA 1. MATERIALES Y REACTIVOS UTILIZADOS EN EL ENSAYO
Equipos:
Microcentrifuga de alta velocidad PRISM™ C2500
Cámara de flujo ESCORT©
Termociclador Life Technologies™ A24811
Electroforesis C.B.S SCIENTIFIC© EPS-300X
Termobloque ACCUBLOCK™ D1100
Incubadora agitadora N-BIOTEK NB205L
Incubadora Thermo Scientific© 3621
Autoclave Centurion Scientific ltd. C2006
Materiales:
Micropipetas de 0.5 hasta 1000µl marca Eppendorf®
Puntas de 10 hasta 1000 µl Eppendorf®
Microtubos de 100 hasta 1500 µl Eppendorf®
Cajas petri
Reactivos:
Nitrógeno Líquido
Kit de extracción de ARN (PureYield™ RNA Midiprep System)
Kit de Retrotranscripción (PureYield™ RNA Midiprep
System)

48
Kit de transformación (pGEM®-T and pGEM®-T
Easy Vector Systems)
Kit de purificación del ADN (Wizard® DNA Clean-Up
System)
Medio S.O.C
x-gal de Promega®
IPTG de Promega®
Primers Revers y Foward
Agarosa
Fuente: Diego Jáuregui
4.4. Elaboración de primers
La elaboración de los primers de la melitina, tanto forward como
reverse, se realizó mediante la búsqueda de su secuencia en Gen Bank
(NCBI), con la accesión NC_007073.3 en donde se obtuvo la secuencia que
se puede ver (Anexo 1 y 2).
Posteriormente por medio del software analítico Primer 3 se elaboraron
y validaron los primers, en base a su estabilidad por su contenido de guanina
y citosina, además de la ausencia de harpins y solapamientos de las pares de
bases entre los cebadores y se obtuvo como resultado lo siguiente (Tabla 2):

49
TABLA 2. PRIMERS FOWARD Y REVERSE PARA LA SECUENCIA DEL GEN DE LA
MELITINA
start len tm gc% any 3' seq PRIMER FORWARD 16 21 57.92 42.86 4.00 0.00 TTAACAGGAAGGAAGGAAGGA PRIMER REVERSE 1416 20 60.02 45.00 6.00 2.00 AGATCGATAAATCGGCATCG
Fuente: Diego Jáuregui
A los primers estudiados por bioinformática se agregaron las
secuencias de sitios de restricción para ApalI y SacII, que amplifica un
fragmento de 500 pares de bases, con el fin que empalmen correctamente
dentro del vector, ya que este contiene estos sitios de restricción en su sitio de
clonaje. Así, la secuencia final fue la siguiente:
Primer Forward 5’TTTTGGGCCCTTAACAGGAAGGAAGGAAGGAA3’
Primer Reverse 5’AAAACCGCGGAGATCGATAAATCGGCATCG3’
4.5. Extracción de ARN
4.5.1. Preparación de la muestra
Con el fin de obtener muestras de ARN total se recolectaron alrededor
de 50 abejas, en frascos de vidrio debidamente esterilizados de 250 mL, del

50
apiaro de la PUCESI y se depositaron en un congelador a una temperatura de
-30°C durante 30 minutos con el fin de conservar el material genético.
4.5.2. Preparación de lisados de muestras de tejido de abejas.
Se depositaron las 20 abejas congeladas en un mortero y se
sumergieron en nitrógeno líquido para su trituración manual.
Posteriormente se aplicaron 2 ml de Solución de Lisis de ARN
PureYield™ por cada 150 mg de abejas y se continuó con su trituración. Para
completar la lisis se incubaron los lisados en hielo durante 10 minutos.
4.5.3. Limpieza del lisado
Se transfirieron 2 ml de lisado a un tubo Falcon® de 15 ml y se
añadieron 4 ml de tampón de dilución de ARN PureYield™ a 2 ml de lisado.
La mezcla se cerró y se agitó por inversión (3-4 veces) y luego se vorteó.
Seguido se añadió 1 ml diluido del Agente de Compensación
PureYield™ al lisado, el mismo que elimina selectivamente los desechos de
ADN, dejando libre al ARN.
Una vez sellado el tubo se mezcló por inversión de 2-3 veces, para
posteriormente agitar hasta obtener homogeneidad en el lisado, momento en

51
el cual se colocaron los tubos en baño María a 70 °C durante 5 minutos para
la desnaturalización de las proteínas.
Se retiraron los tubos del baño María y se enfriaron las muestras a
temperatura ambiente durante al menos 5 minutos.
La muestra se mezcló por agitación hasta su homogeneización, para
después colocar en una columna PureYield™ Clearing, para que su
membrana retuviera las impurezas y permitiera el paso del ARN a un tubo de
extracción de 50 ml.
Se centrifugó el conjunto de la columna/tubo a 2.000 x g a 25 ° C durante
10 minutos para eliminar el lisado y permitir que un líquido azul-verde aclarado
se acumulara en el tubo de extracción de 50 ml.
Se desechó la columna PureYield™ Clearing y se guardó el lisado
aclarado en el tubo de recolección.
4.5.4. Purificación del ARN
Se añadieron 4 ml de isopropanol libre de nucleasas al lisado aclarado
y posteriormente se mezclaron por agitación los tubos.

52
Después se colocó una columna PureYield™ Binding en un tubo de
extracción de 50 ml para cada muestra, para promover la interrupción de
moléculas de agua por las sales caotrópicas y el alcohol, lo que favorece la
adsorción de ARN en la membrana de sílice.
Inmediatamente se vertió la mezcla en un conjunto conformado por la
columna y el tubo de recolección, que se centrifugó a 2000 × g durante 10
minutos con el fin de capturar el ARN en el filtro de la columna.
Posteriormente se retiró la columna PureYield™ Binding y se desechó
el caudal del tubo para después volver a unir la columna al tubo.
Se añadieron 20 ml de ARN solución de lavado a la columna y se
centrifugó a 2000 x g durante 5 minutos.
Se vació el contenido del tubo y se realizó un lavado adicional aplicando
10 ml de Solución de Lavado de ARN PureYield™ a la columna y realizando
una centrifugación por 10 minutos.
Entonces se pasó la columna PureYield™ Binding a un tubo de
extracción de 50 ml fresco y se colocó 1 ml de agua libre de nucleasa a la
columna PureYield™ Binding.
Posteriormente se incubó a temperatura ambiente durante 2 minutos
para liberar el ARN total en la solución y después se centrifugó a 2000 x g
durante 3 minutos para recoger el ARN.

53
Finalmente se retiraron las columnas PureYield™ Binding del tubo y se
desecharon, con el fin de almacenar en alícuotas el ARN purificado a -30 ° C
dentro de tubos estériles.
4.5.5. Cuantificación del ARN extraído
Se empezó extrayendo ARN de las abejas con el fin de cuantificarlo,
para lo cual se utilizó el equipo de fluorómetro ™ Quantus. Este sistema de
detección de ARN QuantiFluor® contiene un colorante fluorescente que
permite la cuantificación sensible de pequeñas cantidades de ARN en
solución. Cabe resaltar que el ARN es un material susceptible a la degradación
debido a la acción de las ribonucleasas, para lo cual se requiere de una
adecuada toma de muestra, condiciones de frío durante la manipulación y
conservación, con el objetivo de inhibir la degradación enzimática. En general
se obtiene muy poca cantidad de este material por la concentración, en
ocasiones puede llegar a no ser detectable (Díaz-Alonso y col, 2013).
4.6. Retrotranscripción
Con el objetivo de realizar la retrotranscripción a ADNc, el ARN
purificado se mezcló con los reactivos necesarios según Tabla 3, tal y como
se detalla a continuación:

54
TABLA 3. COMPONENTES DE PARA LA FORMACIÓN DE LA PRIMERA CADENA DE
ADNC.
Componente Volumen (μl) Concetración
Final
ARN Experimental 0.5 47.4 ng/μl
Primer [Oligo(dT)] 0.5 500 μg/ ml
Agua Libre de
Nucleasas 4.0 -
Final volumen 5.0 -
Fuente: Diego Jáuregui
Se colocaron los tubos en un termobloc a 70 ° C durante 5 minutos.
Inmediatamente se enfriaron en agua con hielo durante 5 minutos.
Se centrifugó a 2000 x g durante 10 segundos en una microcentrífuga
para recoger el condensado y mantener el volumen original.
Se preparó la mezcla de reacción de transcripción inversa mediante la
combinación de los siguientes componentes del sistema (Tabla 4) en un tubo
de microcentrífuga estéril depositado sobre hielo:

55
TABLA 4. COMPONENTES SISTEMA DE REACCIÓN TRANSCRIPCIÓN INVERSA.
Componente Volumen
(μl)
Concentración
Final
Agua Libre de Nucleasas 3.8 -
Buffer de Reacción 5X
GoScript™ 4.0 5x
MgCl2 5.0 1 mM
Mezcla de Nucleóticos de PCR
(Promega) 1.0 0.5mM
Inhibidor de Ribonucleasa
(Recombinant RNasin®) 1.0 40u/ μl
Transcriptasa Reversa
GoScript™ 1.0 -
Volumen final 15.0 -
Fuente: Diego Jáuregui
A cada tubo de microcentrífuga con los 15μl de la mezcla de reacción,
se le añadieron 5μl de la muestra de extracción de ARN a cada reacción para
obtener un volumen final de 20μl por tubo.
Para la alineación del ADNc, se colocaron los tubos en un termobloc a
25 ° C de temperatura y se incubaron durante 5 minutos.

56
Posteriormente se incubaron a 42°C durante una hora para la extensión
de la cadena. Con el fin de amplificar el ADN mediante PCR.
Se inactivó térmicamente la transcriptasa inversa incubando los tubos a
70 ° C durante 15 minutos.
4.7. Amplificación por PCR.
Se preparó la mezcla de PCR según los componentes y las cantidades
de la Tabla 5, en un tubo estéril de microcentrífuga (1.5 ml de volumen) situado
sobre hielo, que se vortexó suavemente para el mezclado y se mantuvo en
hielo antes de la distribución a los tubos de reacción.
Se dispensó el volumen adecuado de mezcla PCR, según Tabla 5, a
cada tubo de reacción que se sentó sobre hielo.

57
TABLA 5. PCR MIX
Componentes Volumen (μl) Concentración
Final
Agua Libre de
Nucleasas 13.25 -
5X GoTaq® Flexi
Reaction Buffer 5.00 5x
MgCl2, 2.00 25mM
Mezcla de
Nucleótidos de
PCR
0.50 10mM
upstream primer 1.50 133.1pmol
downstream primer 1.50 147.9 pmol
Polimerasa
GoTaq® Flexi DNA 0.25 5u/μl
cDNA volumen 1.10 78ng/ μl
Volumen final 25.00 -
Fuente: Diego Jáuregui
Se colocaron las reacciones en un termociclador precalentado a 94 °C
y una vez terminado el ciclo térmico de 2 horas, cuyas rampas térmicas fueron
para la desnaturalización de 94°C, para la anidación (Tm) de 68, 68.2, 68.4,
68.6, 68.8, 69, y para la elongación a 72°C, posteriormente se analizaron los
productos en electroforesis de gel de agarosa al 1.5%.

58
4.8. Electroforesis
Con el fin de preparar el gel de agarosa se disolvió 1.5g de agarosa en
100ml de agua destilada caliente con el fin de que la agarosa se disuelva.
Posteriormente se colocó el gel en la bandeja de molde y se dejó que se
gelifique a temperatura ambiente.
Se preparó la cámara de electroforesis y se colocó la placa que se formó
en el paso anterior.
El ADN a evaluar se puso en los pocillos del gel de agarosa situados dentro
de la cámara de electroforesis y se dejaron correr las muestras por un lapso
de 45 min a 90 voltios.
Finalmente, el gel de agarosa, ya corrido, se transladó al Transiluminador
de UV con el fin de que observe las bandas que se presentan en cada una de
las muestras.

59
4.9. Purificación del ADN
Con el fin de purificar el ADN se usó una jeringa desechable de 3 ml por
cada limpieza. El volumen de la muestra de un ADN de 85 µg/µl fue 50 μl.
Se usó una mini columna Wizard® para cada muestra.
Después se retiró y dejó de lado el émbolo de una jeringa desechable
de 3 ml y se conectó el barril de la jeringa a la extensión Luer-Lok Wizard® de
cada mini columna.
Previamente se mezcló la resina de limpieza del ADN Wizard®, antes
de extraer una alícuota, disolviendo por calentamiento a 37°C durante 10
minutos con el fin de eliminar los cristales presentes, ya que la resina en sí es
insoluble.
Para lo cual se añadió 1 ml de la resina de limpieza del ADN Wizard®
a un tubo de microcentrífuga de 1,5 ml.
Posteriormente se añadió la muestra (50μl) a la resina de limpieza y se
mezcló por inversión suavemente varias veces.

60
Se pipeteó la resina de limpieza que contenía el ADN y se introdujo al
barril de la jeringa, para después insertar lentamente el émbolo de la jeringa y
empujarla suavemente a la suspensión en la minicolumna con el émbolo de la
jeringa.
Se separó la mini columna y el émbolo del barril de la jeringa con el fin
de separar la jeringa de la mini columna y se retiró el émbolo de la jeringa. Y
se colocó una jeringa nueva a la mini columna.
Para lavar la columna se colocó con la pipeta con 2 ml de isopropanol
al 80% en la jeringa y, con la ayuda del émbolo, se empujó suavemente la
solución a través de la mini columna.
Se transfirió la mini columna a un tubo de microcentrífuga de 1,5 ml.
Para posteriormente se centrifugó durante 2 minutos a una velocidad de
10.000 x g con el fin de eliminar la resina.
Se colocó la mini columna en un nuevo tubo de microcentrífuga y se
aplicaron 50μl de agua ultrapura precalentada a 70 ° C a la minicolumna y se
esperó 1 minuto.
Para elucionar el fragmento de ADN unido, se centrifugó la mini
columna durante 20 segundos a velocidad máxima 10.000 x g.

61
Finalmente se retiró y desechó la mini columna. Y el ADN purificado se
almacenó en un tubo de microcentrífuga a -20 ° C.
4.10. Inserción de la secuencia del Gen en el pGEM®-T Easy Vector
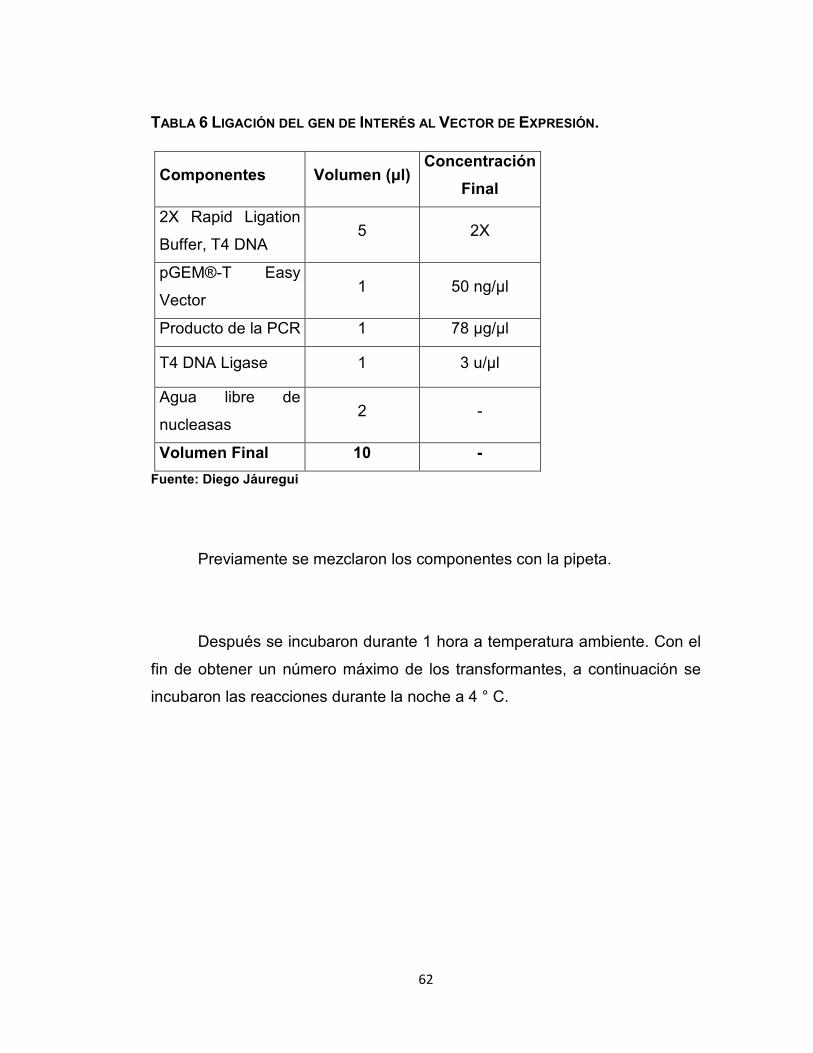
4.10.1. Ligadura del gen de Interés en el Vector de Expresión
El producto de PCR se purificó utilizando el PCR Clean-Up System™,
recomendado para la limpieza de las reacciones antes de la ligadura del ADNc
con el vector para eliminar dímeros de cebadores u otros productos de
reacción no deseados con el fin de mejorar la eficiencia de ligación.
Se centrifugaron rápidamente los tubos que contenían al vector de
expresión de fácil transformación (pGEM®-T Easy Vector) y el ADNc de la
melitina inserto en el vector de expresión con el fin recoger el contenido en la
parte inferior de los tubos. Se establecieron las reacciones de ligación según
la Tabla 6.
62
TABLA 6 LIGACIÓN DEL GEN DE INTERÉS AL VECTOR DE EXPRESIÓN.
Componentes Volumen (μl) Concentración
Final
2X Rapid Ligation
Buffer, T4 DNA 5 2X
pGEM®-T Easy
Vector 1 50 ng/µl
Producto de la PCR 1 78 µg/µl
T4 DNA Ligase 1 3 u/µl
Agua libre de
nucleasas 2 -
Volumen Final 10 -
Fuente: Diego Jáuregui
Previamente se mezclaron los componentes con la pipeta.
Después se incubaron durante 1 hora a temperatura ambiente. Con el
fin de obtener un número máximo de los transformantes, a continuación se
incubaron las reacciones durante la noche a 4 ° C.

63
4.10.2. Preparación de Medios para el Proceso de Transformación
4.10.3. Preparación del medio Caldo Super Optimo con Represión
Catabólica (S.O.C) (100 ml)
Para la preparación del medio Caldo Super Optimo con Represión
Catabólica (S.O.C) se añadió agua de triptona, extracto de levadura-Bacto®,
NaCl y KCl a 97ml de agua destilada y se procedió a agitar para disolver la
solución.
TABLA 7. MEDIO S.O.C
Componente Cantidad
Agua de Triptona 2.0 g
Extracto de levadura-Bacto®, 0,5 g
1M NaCl 1 ml
1M KCl 0.25ml
2M Mg2 + esterilizada por filtración 1 ml
2M glucosa esterilizada por filtración 1 ml
Fuente: Diego Jáuregui
Con el fin de esterilizar la solución se sometió a la autoclave y se lo
enfrió a temperatura ambiente.
Posteriormente se añadió Mg+2 2M y glucosa 2 M, cada uno a una
concentración final de 20 mM.

64
Después se llevó esta solución a 100 ml con agua destilada estéril.
Además, se midió el pH final a 7.0.
4.10.4. IPTG solución madre (0,1 M)
Con el fin de obtener una solución madre del IPTG a 0.1 M se pesó 1.20
g de este y se añadió agua filtrada y esterilizada hasta 50 ml de volumen final.
4.10.5. Preparación de placas de Caldo de Lysogeny (LB) con
ampicilina
Se mezclaron 15 g de agar en 1 litro de medio LB. Autoclavado.
Después al medio se lo enfrió a 50 ° C antes de añadir a la ampicilina a una
concentración final de 100 ppm
Se vertieron 30ml de medio preparado en placas Petri de 85 mm. Y se
dejó cuajar el agar.

65
4.10.6. Preparación de placas de LB con ampicilina / IPTG / X-Gal
Las placas de LB con ampicilina ya preparadas se complementaron con
0,5 mM IPTG y 80μg / ml de X-Gal y se vertieron en las placas.
4.11. Transformación de la célula competente
Se prepararon dos placas con medio sólido de LB / ampicilina / IPTG /
X-Gal. Para lo cual se añadieron 15 g de agar en 1 litro de medio BD Difco™
LB Agar, Miller y se dejó que el medio se enfríe a 50 °C antes de la adición de
ampicilina a una concentración final de 100μg / ml.
Se vertieron 30 ml de medio en cada placa Petri de 85 mm y, una vez
solidificado el medio, este se complementó con IPTG 0.5 mM y 80 μg/ml de X-
Gal.
Esto se hizo para cada reacción de ligación y se equilibraron las placas
a temperatura ambiente.
Se centrifugaron a 2000 x g los tubos que contenían las reacciones de
ligación con el fin de recoger el contenido en la parte inferior.

66
Además, se añadieron 2 μl de cada reacción de ligación a un tubo de
microcentrífuga de 1.5 ml en hielo.
Se retiraron los tubos con células competentes de E. coli JM109 de alta
eficiencia congeladas.
Se colocaron en un baño de hielo hasta que esté descongelado.
Después se procedió a mezclar el contenido del tubo con un movimiento
rápido suavemente el tubo.
Se transfirieron 50 μl de este contenido con células en cada tubo
preparado.
Se golpearon suavemente los tubos para mezclar y colocarlos en hielo
durante 20 minutos y se expusieron a un choque térmico las células mediante
su inmersión en un baño seco a 42 ° C durante 45-50 segundos.
Inmediatamente se devolvieron los tubos a hielo durante 2 minutos y se
añadieron 950 μl de medio S.O.C., siendo este un medio rico en nutrientes que
contiene péptidos, aminoácidos, vitaminas solubles en agua y glucosa en una
formulación baja en sal que ayuda al crecimiento microbiano.

67
La mezcla se incubó durante 1.5 horas a 37 ° C con agitación a 150
rpm.
Se colocaron entonces 100 μl de cada cultivo en las placas de
transformación. Y finalmente se incubaron las placas durante 24 horas a 37°C.

68
CAPÍTULO V
5. RESULTADOS Y DISCUCIÓN
Con el fin de identificar la secuencia del gen que codifica a la melitinia de
abejas (Apis mellifera) mediante técnicas de PCR, para insertarlo en un vector
de expresión se obtuvo los siguientes resultados.
Mediante un sistema para la detección de ARN QuantiFluor®, se midió la
cantidad de 47 ng/μl de ARN (figura 8). Cabe destacar que en ensayos
espectrofotométricos tradicionales no pueden determinar claramente las
concentraciones de ARN inferiores a 2 μg/ml; sin embargo, muchas
concentraciones de ARN aisladas están muy por debajo de ese nivel
(Corporation, Promega, 2014). y cuando se obtiene muy poca cantidad de este
material por la concentración en ocasiones puede llegar a no ser detectable
(Carmen y col., 2013).
Figura 8 Cantidad de ARN de Melitina
Fuente: Diego Jáuregui (Laboratorio PUCESI)

69
Tras las pruebas de RT-PCR a diferentes temperaturas (Tm1 a 68.0°C;
Tm2 a 68. 2°C; Tm3 a 68.4°C; Tm4 a 68.6°C; Tm5 a 68.8°C y Tm6 a 69°C;
Figura 9), la temperatura de hibridación a 68.4°C fue la que dio como resultado
una mejor visualización a una concentración del 1.5% del gel de agarosa y,
por tanto, mayor cantidad de ADNc. Cabe resaltar que todas las muestras
de ADNc hibridaron con los primers a temperaturas de hibridación distintas.
Figura 9 Gen de interés en electroforesis a distintas
temperaturas de hibridación (Tm).
Fuente: Diego Jáuregui (Laboratorio PUCESI)

70
Con el fin de cuantificar el ADNc purificado de la secuencia de la melitina
se utilizó el equipo de fluorómetro ™ Quantus. Que en general se obtuvo una
cantidad significativa de este material de 78 ng/μl, tal como se aprecia en la
Figura 10, en ocasiones puede llegar a no ser detectable por su baja
concentración (Carmen y col., 2013).
Figura 10. Cantidad de ARN de Melitina
Fuente: Diego Jáuregui (Laboratorio PUCESI)

71
El fragmento de ADNc que codifica para la melitina a partir del ARN total
de las abejas obreras que se obtuvo mediante RT-PCR y posteriormente con
una PCR convencional a una Tm de 68.4 °C, pasó por un proceso de
purificación tal y como se observa en la Figura 11 donde 1 es el control positivo
y el 2 es el ADNc purificado de melitina.
Figura 11 Electroforesis en gel de agarosa con dos tipos de
muestras: 1) Control positivo, 2) ADNc de melitina
originario de la abeja (Apis mellifera) purificado.
Fuente: Diego Jáuregui (Laboratorio PUCESI)

72
El fragmento de la secuencia de ADNc melitina que fue amplificado por
PCR a partir de un ADN recombinante de melitina previamente secuenciado y
fue insertado en el vector pGEM-T easy, tal como se encuentra representado
en tal como se muestra en la Figura 12.
Figura 12. Representación Esquemática del vector de
Expresión pGEM Easy Vector con el gen de interés.
Fuente: modificado por Diego Jáuregui de
https://worldwide.promega.com/products/pcr/pcr-
cloning/pgem_t-easy-vector-systems/
La transformación de la célula competente de E coli JM109 con el
inserto en el vector pGEM-T easy, se encuentra representada
esquemáticamente en la Figura 13.

73
Figura 13. Representación Esquemática de la
transformación del vector de Expresión pGEM Easy Vector
con el gen de interés en la célula competente de E coli
JM109.
Fuente: http://www.promegaconnections.com/tag/cloning/
Se detectaron bacterias transformadas, con el método de prueba azul/blanco,
donde se mutó la región lacZ Δ, y se eliminó una región resultado un péptido
no funcional llamado fragmento ω, y no codificó para la enzima β-
galactosidasa funcional dando una coloración blanca (Figura 14). Porque el
plásmido suministró un prorción, llamada fragmento α. Debido a que en este
experimento se obtuvieron bacterias con esta coloración, se puede afirmar que
se han obtenido bacterias transformadas con el gen de la melitina (Figura 15).

74
Figura 1 Colonias de bacterias transformandas con el ADNc
de la melitina.
Fuente: Diego Jáuregui

75
CONCLUSIONES Y RECOMENDACIONES
CONCLUSIONES
Una vez acabada la investigación se pudo clonar el gen de la melitina
originaria de la abeja (Apis mellifera) en un vector de expresión lo cual esto
abre las puertas de investigación con finalidades terapéuticas.
Para la identificación del gen que codifica a la melitina de abejas se
obtuvo un ARNm total de la abeja a una concentración de 47 ng/μl, lo que
indica que su concentración es baja sin embargo se pudo trabajar con este
ARNm para que mediante la técnica de la Retro Transcriptasa Inversa RT-
PCR, lograr retrotranscribir el ARNm total a un ADNc total de la abeja y esto
ayudó a encontrar el gen de la melitina.
Se elaboraron primers específicos parara el gen de la melitina, con la
ayuda de las bases de datos del banco de genes del NCBI y un software libre
específico, a los que se les añadieron adaptadores moleculares con las
secuencias de sitios de restricción para ApalI y SacII, con el fin de añadirse al
sitio de clonaje en el vector de expresión. Estos primers fueron sometidos
conjuntamente con el ADNc total de la abeja a la técnica de la PCR
convencional con lo que se consiguió el gen específico de la melitina.

76
Para la obtención del constructo con el gen de la melitina se utilizó un
vector de expresión, el cual fue introducido en una célula competente E. coli
JM109 de fácil transformación.
Con el fin de evidenciar la correcta inserción, transformación y
expresión del gen en el plásmido y la célula transformada, se utilizó el método
de clonación azul / blanco, con el cual se logró diferenciar los plásmidos con
inserto de los sin inserto por el color de la colonia, donde las colonias de color
blanco tuvieron vectores que con insertos mientras que las colonias que
presentaron color azul sus vectores no tuvieron insertos.

77
RECOMENDACIONES
Para obtener mejores concentraciones de ARNm se debe mantener el
lugar de trabajo sumamente limpio, libre de nucleasas ya que este ácido
nucleico es muy lábil además se debe buscar otros protocolos de extracción
de ARNm.
La elaboración de otros primers ayudaría a evaluar de mejor manera el
segmento del gen de la melitina para su inserción y su posterior expresión.
Así también elaborar constructos a base de melitina con otros
promotores con el fin de evaluar su nivel de expresión mediante la técnica de
la Real Time PCR.
Probar la transformación con nuevas técnicas como la electroporación
para determinar la eficiencia de transformación, así como otras opciones de
células competentes.
Purificar los medios de cultivo que contienen las células transformadas
con el gen de melitina para obtener melitina pura y poder realizar aplicaciones
terapeúticas.

78
BIBLIOGRAFÍA
Apicola el Nebrillo. (2011). El aguijón de las abejas y su veneno . Obtenido
de Pepe miel: http://www.pepemiel.es/blog/el-aguijon-de-las-abejas-y-
su-veneno
Becattini, S., Taur, Y., & Pamer, E. (2016). Antibiotic-Induced Changes in the
Intestinal Microbiota and Disease. Trends Mol Med, (6):458-78.
Bi y col. (2009). Cloning and expression of a gene encoding shortened LfcinB
(1-15)-Melittin(5-12) hybrid peptide in Escherichia coli BL21(DE3).
Sheng Wu Gong Cheng Xue Bao 2009 , 7:975-981.
Biló y col. (2005). Diagnosis of Hymenoptera venom allergy. EAACI.
Carmen, Garrote, H., Amor, A., Suárez, Y., & González, R. (2013).
Cuantificación de ácido ribonucleico para la realización para de la
técnica de RT- PCR. Revista Cubana Hematología, Inmunología y
Hemoterapia, 7: 298-303.
Corporación, Promega. (17 de abril de 2015). Cloning. Obtenido de
Successful Ligation and Cloning of Your Insert:
http://www.promegaconnections.com/tag/cloning/
Corporation, Promega. (2014). Measuring RNA Concentration Using the
Quantus™ Fluorometer with the QuantiFluor® RNA System.
MADISON: MADISON, WI 53711-5399 USA.
Fletcher, J., Michaux, K., & Jiang, M. (1990). Contribution of bee venom
phospholipase A2 contamination in melittin fractions to presumed
activation of tissue phospholipase A2. Toxicon, 28(6):647-656.

79
Hahn, A., Jain, R., & Spach, D. (2016). New Approaches to Antibiotic Use
and Review of Recently Approved Antimicrobial Agents. Med Clin
North Am. 2016 Jul;100(4):911-26., : 911- 926.
Han, S., & Lee, G. a. (2012). Skin Sensitization Study of Bee Venom (Apis
mellifera L.) in Guinea Pigs. Toxico, 28:1-4.
Hathroubi, S., Mekni, M., Domenico, P., Nguyen, D., & Jacques, M. (2016).
Biofilms: Microbial Shelters Against Antibiotics. Microb Drug Resist.
Hood y col. (2013). Cytolytic nanoparticles attenuate HIV-1 infectivity. Toxicol,
28:1-4.
Huang. (2013). Hybrid Melittin Cytolytic Peptide-Driven Ultrasmall Lipid
NanoparticlesBlock Melanoma Growth in Vivo. American Chemical
Society.
Husseneder y col. (2016). Genetically Engineered Yeast Expressing a Lytic
Peptide from Bee Venom (Melittin) Kills Symbiotic Protozoa in the Gut
of Formosan Subterranean Termites. PLOS ONE, 3:1-9.
Johnston, P., Dobson, A., & Rolff, J. (2016). Genomic Signatures of
Experimental Adaptation to Antimicrobial Peptides in Staphylococcus
aureus. G3 (Bethesda).
Lazarev, V., Stipkovits, L., Biro, J., Miklodi, D., Shkarupeta, M., Titova, G., . . .
Govorun, V. (2004). Induced expression of the antimicrobial peptide
melittin inhibits experimental infection by Mycoplasma gallisepticum in
chickens. Microbes and Infection, 6: 536–541.
Matysiak y col. (2001). Characterization of honeybee venom by MALDI-TOF
and nanoESI-QqTOF mass spectrometry. J Pharm Biomed Anal.,
54:273-278.

80
Mularski, A., Wilksch, J., Wang, H., Hossain, M., Wade, J., Separovic, F., . . .
M.L., G. (2015). Atomic Force Microscopy Reveals the
Mechanobiology of Lytic Peptide Action on Bacteria. Langmuir,
(22):6164-6171.
OIE. (22 de 09 de 2014). Organización Mundial de Sanidad Animal,
Estrategia de la OIE para la reducción de las amenazas biológicas.
Obtenido de http://www.oie.int/es/nuestra-experiencia-
cientifica/reduccion-de-las-amenazas-biologicas/
Oršolić. (2012). Bee venom in cancer therapy. Springer Science+Business
Media, 31:173–194.
Ownby, C., Powell, J., Jiang, & Fletcher, J. (1997). Melittin and phospholipase
A2 from bee (Apis mellifera) venom cause necrosis of murine skeletal
muscle in vivo. Toxicon., 35(1):67-80.
Park y col. (2014). Functional characterization of naturally occurring melittin
peptide isoforms in two honey bee species, Apis mellifera and Apis
cerana. Peptides., 53:185-193.
Park, Y., & Hahm K, S. (2005). Antimicrobial peptides (AMPs): peptide
structure and mode of action. Biochem Mol Biol., (5):507-516.
Peiren y col. (2008). Proteomic analysis of the honey bee worker venom
gland focusing on the mechanisms of protection against tissue
damage. Toxicon, 52:72-83.
Peña, L., Pineda, M., Hernández, m., & Rodríguez, A. (2006). Toxinas
Naturales: abejas y sus venenos. Archivos Venezolanos de
Farmacología y Terapéutica, , 25(1): 6-10.
Peters, B., Shirtliff, M., & Jabra-Rizk, M. (2010). Antimicrobial Peptides:
Primeval Molecules or Future Drugs? ONEPLUS.

81
Raghuraman, H., & Chattopadhyay, A. (2007). Melittin: a Membrane-active
Peptide with Diverse Functions. Springer Science+Business Media,
27:189–223.
Reinhardt, A., & Neundorf, I. (2016). Design and Application of Antimicrobial
Peptide Conjugates. Int J Mol Sci., 5: 11-17.
Rybak, Chmielewska, & Szczêsna, T. (2004). HPLC STUDY OF CHEMICAL
COMPOSITION OF HONEYBEE (Apis mellifera L.) VENOM.
Apicultural Science, 48(2):103-109.
Sharma, S. (1993). Melittin-induced hyperactivation of phospholipase A2
activity and calcium influx in ras-transformed cells. Oncogen, 8(4):939-
947.
Shi, W., Xu, H., Cheng, J., & y Zhang, C. (2004). Expression of the melittin
gene of Apis cerana cerana in Escherichia coli. Protein Expression and
Purification, 37:213–219.
Son y col. (2007). Therapeutic application of anti-arthritis, pain-releasing, and
anti-cancer effects of bee venom and its constituent compounds.
Pharmacol., 115:246-270.
Van Vaerenbergh, M., Cardoen, D., & E.M., F. (2013). Extending the honey
bee venome with the antimicrobial peptide apidaecin and a
proteinresembling wasp antigen 5. Royal Entomological Society, 1-12.
Vutto, N., T.A, G., Pundik, A., Tretiakova, T., & Volotovskiĭ, I. (2010).
Transgenic Belarussian-bred potato plants expressing genes for
antimicrobial peptides of the cecropin-melittin type. AGRIS, 17:208-
218.
Wang, J., Han, M, Zhang, G., Qiao, S., & Li, D. y. (2016). The Signal Pathway
of Antibiotic Alternatives on Intestinal Microbiota and Immune Function.
Curr Protein Pept Sci.

82
Watkins, R., & Bonomo, A. (2016). Overview: Global and Local Impact of
Antibiotic Resistence. Infect Dis Clin N Am, 30: 313-322.
Weinstock, G., Robinson, G., Gibbs, R., Weinstock, G., Robinson, G., Worley,
K., . . . M., B. (2006). Insights into social insects from the genome of
the honeybee Apis mellifera. NIH Public Access, 443:931–949.
Zhu y col. (2002). Cloning and expression of a gene encoding shortened
cecropin A-melittin hybrid in E.coli. Yi Chuan, 24:31-34.
83
ANEXOS
Anexo 1

84
Anexo 2.
Secuencia de la Melitina
Fuente: (NCBI, 2016)

85
Anexo 3.
COMPONENTES DE LA APITOXINA
Fuente: (Oršolić, 2012).

86
Anexo 4.
ALINEACIÓN DE LA SECUENCIA
Fuente: (NCBI-BLAST, 2016)







