
UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTOS DE CIENCIAS DEL MAR
DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
TESIS:
EVALUACIÓN DEL POTENCIAL DE Sargassum lapazeanum
Setchell & N.L. Gardner (Ochrophyta: Fucales, Phaeophyceae)
COMO FUENTE DE COMPUESTOS ANTIBACTERIANOS,
ANTIOXIDANTES Y ANTICOAGULANTES.
QUE PARA OBTENER EL TITULO DE:
BIÓLOGA MARINA
PRESENTA:
VALERIA ALEXANDRA VILLEGAS SILVA
DIRECTOR:
DR. MAURICIO MUÑOZ OCHOA
LA PAZ, B.C.S., MÉXICO. JUNIO DE 2014



Agradecimientos
Este proyecto se realizó en las instalaciones del Centro Interdisciplinario de Ciencias
Marinas del Instituto Politécnico Nacional, bajo la dirección del Dr. Mauricio Muñoz
Ochoa, financiado por el proyecto "Sargassum lapazeanum como fuente de
compuestos bioactivos” IPN-SIP20131037”.
Al Dr. Mauricio Muñoz Ochoa por su incondicional apoyo y paciencia, así como por
su orientación y guía en laboratorio durante las pruebas realizadas incluso en
momentos de desesperación académica. Gracias por enseñarme las bases de la
química de productos naturales, motivándome siempre a seguir adelante.
A la Dra. Gabriela Andrade Sorcia, por la agradable lección de botánica y la
identificación taxonómica del alga. A mi comité de tesis Dr. Rafael Riosmena
Rodríguez, Dr. Juan Manuel López Vivas, Dra. Gabriela Andrade Sorcia y Dr. Jesús
Iván Murillo Álvarez por sus acertados comentarios y observaciones, así como el
tiempo y dedicación a la hora de aclarar dudas.
A las M. en C. Yoloxochitl Elizabeth Rodríguez Montesinos y Dora Luz Arvizu
Higuera por el apoyo, la confianza y las facilidades que me permitieron disponer del
laboratorio hasta altas horas de la noche. Asimismo agradezco a mis compañeros de
laboratorio de química de algas por hacer del tiempo trabajando algo más ameno.
Finalmente a aquellas personas que me apoyaron y confiaron en mí durante la
realización de este proyecto, principalmente mis hermanos y amigos por escucharme
cuando más lo necesite y acompañarme a lo largo de este proceso, muchas gracias.

“... pero sus estridentes ladridos sólo son señal de que cabalgamos.”
(Goethe, 1808).

RESUMEN
La península de Baja California cuenta con una gran variedad de hábitats y
condiciones climáticas, por lo que alberga una enorme riqueza de algas marinas destacando
la presencia de las algas pertenecientes al género Sargassum por su alta dominancia en
biomasa. (Suárez Castillo et al., 2013). Se seleccionó la especie Sargassum lapazeanum,
para el estudio de su potencial actividad biológica como fuente de compuestos con actividad
antibacteriana, antioxidante y anticoagulante. Los resultados obtenidos indican que el
extracto etanólico de S. lapazeanum, así como sus diferentes fracciones presentan
una actividad selectiva contra bacterias del género Vibrio como V. harveyi y V.
parahaemolyticus, además de actividad secuestrante de radicales libres en la
mayoría de los extractos analizados, obteniendo la menor EC50=39.96 µg mL-1 a
partir de F6. Por su parte el fucoidan crudo y sus fracciones mostraron valores
comparables a los observados por la heparina a concentraciones de 10 µgmL -1.
Finalmente el análisis estructural del fucoidan demostró que se trata de un
heterofucano con sustituciones de grupos sulfato en posición C4 mayoritariamente.
Palabras clave: actividad biológica, algas pardas, caracterización, proximal,
fucoidan.

ÍNDICE
Páginas
LISTA DE FIGURAS I
LISTA DE TABLAS III
LISTA DE ABREVIACIONES IV
GLOSARIO V
I. INTRODUCCIÓN 1
II. ANTECEDENTES 2
1. Análisis del potencial antibacteriano 3
1.1. Antibacterianos y su clasificación 4
1.2. El género Sargassum como fuente de compuestos con
actividad antibacteriana. 5
2. Análisis del potencial antioxidante 6
2.1. Radical libre DPPH 6
2.2. Algas del género Sargassum como fuente de compuestos
antioxidantes 7
3. Análisis del potencial anticoagulante 9
3.1. Polisacáridos sulfatados: Fucoidan y actividad anticoagulante
en phaeophytas 9
3.2. Mecanismos de coagulación sanguínea 12
3.3. Evaluación de la actividad anticoagulante 16
III. JUSTIFICACIÓN 18
IV. OBJETIVOS 21
V. MATERIAL Y MÉTODOS 22
1. Recolección e identificación taxonómica 22
2. Análisis proximal del alga S. lapazeanum 23
3. Obtención del extracto 23
3.1. Obtención del extracto etanólico 23
3.2. Obtención del extracto acuoso: Fucoidan 23

4.1. Fraccionamiento por gradiente de polaridad 26
4.2. Fraccionamiento de F2 en columna cromatográfica 26
4.3 Purificación parcial del fucoidan 26
5. Pruebas de actividad biológica 26
5.1. Evaluación de la actividad secuestrante de radicales libres
(DPPH): análisis del potencial antioxidante 26
5.2. Evaluación de la actividad antibacteriana por medio del método
de difusión en agar con discos 28
5.3. Análisis de la actividad anticoagulante 28
6. Caracterización parcial del fucoidan 29
VI. RESULTADOS 30
1. Análisis proximal 30
2. Análisis de la actividad biológica 30
2.1. Actividad antibacteriana 32
2.2. Actividad secuestrante de radicales libres 34
2.3 Actividad anticoagulante 35
VII. DISCUSIÓN
1. Análisis proximal 36
2. Análisis de la actividad antibacteriana 37
3. Análisis de la actividad antioxidante 41
4. Análisis de la actividad anticoagulante 44
4.1 Caracterización parcial del fucoidan 44
4.2. Ensayo de actividad anticoagulante. 46
VIII. CONCLUSIONES 50
IX. BIBLIOGRAFÍA 51
X. ANEXOS 64

I
LISTA DE FIGURAS
Páginas
Figura 1. Estructura química del dioctil ftalato 5
Figura 2. Difenilpicrilhidracilo (radical libre). 7
Figura 3. Difenilpicrilhidracina. 7
Figura 4. Estructura de algunos compuestos químicos aislados a partir de
especies del género Sargassum: A) sargothunbergol-A, B y C)
Tetrapreniltoluquinoles, D Y E) monogalactosil-diacilgliceroles.
8
Figura 5. Estructura química promedio del fucoidan. 10
Figura 6. Diagrama que muestra la cascada de coagulación. Imagen
tomada de Martínez-Murillo (2003). 13
Figura 7. Vía afectada por la prueba de tiempo de protrombina. Imagen
tomada de Kitchen et al. (2010). 17
Figura 8. Mapa que muestra la zona de recolecta del alga S. lapazeanum
en San Juan de la Costa, Bahía de La Paz B.C.S, México.
Modificado a partir de NOAA: http://www.ngdc.noaa.gov/mgg/
shorelines/shorelines.html.
22
Figura 9. Obtención del extracto crudo (EC) y fraccionamiento del alga S.
lapazeanum. 24
Figura 10. Esquema que muestra el proceso de obtención y
fraccionamiento del fucoidan. 25
Figura 11. Cromatografía en placa fina que muestra la actividad
antioxidante de S. lapazeanum por el método bioautográfico,
revelada con una solución de DPPH al 0.4%. E.E.C= Extracto
crudo.
32
Figura 12.
.
Gráfica comparativa de los valores obtenidos para las EC50 en
las fracciones obtenidas por gradiente de polaridad a partir de
S. lapazeanum. 1= Menor polaridad, 7= Mayor polaridad E.EC.=
Extracto Crudo.
32

II
Figura 13. Curva dosis efecto la fracción F6 de S. lapazeanum, donde se
observa el porcentaje de reducción del DPPH (%) en relación a
la concentración del extracto (µg mL-1). En negro: línea de
tendencia y desviación estándar. En rojo: concentración efectiva
media (EC50).
33
Figura 14. Curva dosis efecto del ácido ascórbico (AA), utilizada como
blanco de comparación, donde se observa el porcentaje de
reducción del DPPH (%) en relación a la concentración del AA
(µg mL-1). En negro: línea de tendencia y desviación estándar.
EC50, obteniendo por extrapolación= 1.5 µg mL-1
33
Figura 15. Gráfico que muestra la relación entre la concentración de los
extractos obtenidos a partir de la precipitación fraccionada del
Fucoidan (FC) de S. lapazeanum y su efecto sobre el tiempo
normal de coagulación según el ensayo de TTPA36
35
Figura 16. Espectros de FTIR-ATR de las fracciones obtenidos por
precipitación fraccionada del extracto acuoso (Fucoidan) de S.
lapazeanum: FC1, FC2 y FC3 en la región de los 500-1800cm-1.
44

III
LISTA DE TABLAS
Páginas
Tabla I. Estudios realizados con el alga Sargassum lapazeanum 4
Tabla II. Listado de algunas especies del género Sargassum que han
presentado actividad anticoagulante. 12
Tabla III. Nombre y función de los diferentes factores y cofactores que
actúan en las diferentes etapas de la cascada de coagulación15 14
Tabla IV. Resultados de análisis proximal del alga S. lapazeanum. 30
Tabla V. Resultados de la prueba de difusión en agar con discos [2
mg/disco] del extracto crudo y las fracciones obtenidas a partir de
S. lapazeanum. Se reporta el promedio del halo de inhibición en
milímetros ± sd, n = 2.
31
Tabla VI. Resultados de la prueba de difusión en agar con discos [2
mg/disco] del fraccionamiento del extracto F2 obtenido a partir de
S. lapazeanum. Se reporta el promedio del halo de inhibición en
milímetros ± sd, n = 2.
31
Tabla VII. Actividad anticoagulante de los extractos obtenidos por
precipitación fraccionada a partir de S. lapazeanum 34
Tabla VIII. Comparación de actividad de los extractos acuosos obtenidos a
partir de Sargassum lapazeanum en el ensayo de tiempo de
protrombina (TP)47
46
Tabla IX. Listado de algunos valores de TP y TPPA reportados para algas
del género Sargassum 47

IV
LISTA DE ABREVIACIONES
CH2Cl2 Diclorometano
BHA Butil-hiroxi-anisol
BHT Butil-hidroxi-tolueno
DPPH 2,2-difenil-1-pricilhidracilo
ECV Enfermedades Cardio Vasculares
ELN Extracto libre de Nitrógeno
ERO Especies Reactivas de Oxígeno
EtOH Etanol
IR Espectroscopia de infrarrojo
H2O Agua
MEtOH Metanol
TIH Trombocitopenia inducida por heparina
TLC Cromatografía en placa fina, del inglés: Thin Layer Cromatography.
TTPA Tiempo de Tromboplastina Parcial Activada
TP Tiempo de Protrombina
PS Polisacáridos sulfatados

V
GLOSARIO
Absorbancia Medida de la fracción de luz que es absorbida por una
muestra.
Aerocistos Estructuras vegetativas que contienen gas nitrógeno y
contribuyen a la flotación del alga.
Antibacteriano Químicos que ayudan a eliminar bacterias patógenas para los
organismo.
Anticoagulante Sustancia que prolonga el tiempo de coagulación de la sangre.
Antioxidante Cualquier sustancia, ya sea sintética o natural, que al
encontrarse concentraciones menores en relación a las del
sustrato oxidable retrasará o inhibirá de manera significativa la
oxidación de éste.
Cininógeno Proteína plasmática que existe normalmente en el plasma.
Estipe Eje principal cilíndrico y ramificado que le proporciona
flexibilidad y rigidez al talo.
Espectroscopia Técnica analítica experimental que se basa en detectar la
absorción de la radiación electromagnética, y relacionarla con
los niveles de energía implicados en la transición cuántica
Extrínseca Vía dependiente del factor tisular (FT).
Filoides Estructuras fotosintéticas con apariencia de hojas.
Hemodiálisis Paso de la sangre a través de membranas semipermeables
para liberarla de productos nocivos de bajo peso molecular,
como la urea.

VI
Hemostasia Fenómeno fisiológico que detiene el sangrado.
Hirudina Polipéptido de 65 residuos aislado de las glándulas salivales
de las sanguijuelas. Es un inhibidor específico de la trombina.
Intrínseca La vía intrínseca recibió este nombre debido a que todos los
componentes necesarios se encontraban presentes en la
sangre.
Precalicreína Proteína plasmática precursora de la calicreína, enzima que a
su vez activa el factor FXII, su ausencia en plasma produce
anormalidades en la formación de tromboplastina y en la
generación de cinina.
Radicales libres Átomos o moléculas con un electrón impar que es
extremadamente reactivo y capaz de participar en cadenas de
reacción rápida que desestabilizan otras moléculas y generan
la formación de muchos otros radicales libres.
Sujetador Parte del talo que sirve para el anclaje del mismo al sustrato.
Zimógenos Precursores enzimáticos. Durante las diferentes etapas de la
coagulación se convierten de pro-enzimas a enzimas activas.
Trombocitopenia
inducida por
heparina (TIH) Trombosis inducidas por el uso de heparina.

1
I. INTRODUCCIÓN
Sargassum C. Agardh es un género de algas café (Phaeophyceae) que cuenta
con al menos 335 especies aceptadas taxonómicamente (Guiry y Guiry, 2013), lo
que lo convierte en el más abundante dentro del orden de los Fucales (Rindi et al.,
2012). Éste género se extiende a lo largo de casi todos los océanos del mundo,
por lo que incluye especies tropicales, subtropicales y templadas, las cuales
suelen formar densos bosques submarinos, constituyendo así un hábitat clave en
el suministro y refugio de muchas especies (Suárez Castillo et al., 2013). En el
Golfo de California, son consideradas de gran importancia por su alta dominancia
en biomasa, así como por su capacidad para la síntesis de metabolitos
secundarios, entre los que destacan los terpenoides, florotaninos, polifenoles,
hidrocarburos volátiles, polisacáridos sulfatados y productos de origen biogenético
mixto. Lo anterior, les proporciona una composición química única y las convierte
en un recurso altamente cultivable de potencial uso como fertilizante, forraje,
suplemento alimenticio o materia prima para la obtención de ficocoloides y
compuestos de interés farmacéutico (Mascheck y Baker, 2008; Echeverría et
al.,2009).
Existen 5 especies de Sargassum conformando los bosques submarinos del Golfo
de California: S. johnstonii, S. herporhizum, S. sinicola, S. horridum y S.
lapazeanum (Suárez Castillo et al., 2013), siendo la gran mayoría de éstas
estudiadas para el análisis de su composición química. No obstante, aún no se
cuenta con ningún registro de actividad biológica para S. lapazeanum, un alga
endémica del Golfo que podría representar un recurso de potencial explotación
para la zona (Andrade-sorcia, 2008). Por lo anterior, se seleccionó el alga
Sargassum lapazeanum (Anexo I) para el análisis de su actividad biológica como
fuente de compuestos antibacterianos, antioxidantes y anticoagulantes.

2
II. ANTECEDENTES
Fue a principios de 1960 que los investigadores comenzaron a concentrarse en los
océanos como un novedoso e inexplorado recurso con gran potencial para la
obtención de compuestos bioactivos. Como resultado de esto, más de 10,000
metabolitos marinos se han aislado y caracterizado a lo largo de las últimas cinco
décadas, demostrando tener diferentes tipos de actividad biológica (Attaway y
Zaborsky 1993). Actualmente son muchos los organismos marinos a partir de los
cuales se han aislado e identificado compuestos con actividad biológica, sin
embargo de entre todos destacan las macroalgas por su capacidad para la
síntesis de metabolitos secundarios (Mascheck y Baker, 2008).
La biotecnología con macroalgas comenzó a desarrollarse a mitad del siglo
pasado, sin embargo en México no ha avanzado lo suficiente, pues representa el
estudio de una comunidad compleja, de alta diversidad biológica en la que aún
hace falta una mayor dedicación de esfuerzos e inversión económica (Zertuche-
González, 1993). Es probable que lo anterior sea la razón por la cual la mayoría
de los trabajos realizados con macroalgas en el País se enfocan principalmente en
su ecología, diversidad, variaciones espacio-temporales, su aprovechamiento
como bioindicadores, así como su caracterización y descripción taxonómica (ej:
Calva et al., 2008; Mateo-Cid et al., 2013; Saad y Riosmena, 2005), siendo aún
más escasos los trabajos para la obtención de compuestos bioactivos a partir de
éstas. Esto también pudiera deberse a la necesidad de conocer y clasificar
adecuadamente los organismos con los que trabajamos, ya que nos permite
reconocer con mayor exactitud la procedencia del compuesto aislado, así como
comprender parte del ciclo de vida y la ecología química de los organismos
estudiados.
Por su parte, las algas café (Phaeophyceae) han sido altamente estudiadas
debido a que producen una amplia variedad de metabolitos secundarios.
Actualmente se han reportado al menos 1,140 compuestos químicos obtenidos a
partir esta clase de algas, los cuales ofrecen un amplio espectro de actividades

3
farmacológicas tales como: antimicrobiana, antioxidante, o anticoagulante
(Mascheck y Baker, 2008; Echeverría et al.,2009). No obstante, al menos un tercio
de lo que se conoce de la química de éstas algas proviene tan sólo del estudio del
género Dictyota, mientras el resto corresponde a estudios en su mayoría con el
género Cystoseira, Padina y Sargassum (Mascheck y Baker, 2008).
En ese sentido, los estudios realizados con Sargassum lapazeanum son muy
escasos (Tabla I), estos comienzan con su descubrimiento en 1924 (Setchell, W.A.
& Gardner, 1924). y han mejorado conforme el avance de la tecnología con
estudios de biogeografía (Phillips, 1995), dinámica poblacional (Rivera y Scrosati,
2006), taxonomía (Norris, 2010; Andrade-Sorcia et al. 2008) y capacidad de
bioadsorción de cadmio (Patrón-Prado et al., 2011), hasta un replanteamiento
actual de su sistemática (Andrade-Sorcia et al., 2013). Cabe destacar que a pesar
de los avances tecnológicos, no se ha reportado aún ningún estudio sobre la
actividad biológica de ésta alga, no obstante, existen referencias bibliográficas que
demuestran la actividad biológica de algas pertenecientes al género Sargassum, lo
que hacen del presente estudio un trabajo novedoso que contribuirá al desarrollo
del conocimiento sobre la química en las algas pertenecientes a éste género.
1. Potencial de las algas como agentes antibacterianos.
La mayoría de los antibióticos que se utilizan en la actualidad son de origen
terrestre, por lo que el análisis se ha enfocado mayoritariamente a este ambiente,
lo que sugiere que la probabilidad de encontrar nuevos compuestos antibióticos a
partir de ambientes distintos como el mar es mucho mayor (Song et al., 2011).
Algunos estudios indican que el potencial antibacteriano de las algas se debe a su
capacidad para la síntesis de diterpenos en plantas chlorophytas, terpenos
halogenados en rodophytas y metabolitos mixtos de origen terpeno-aromático en
las algas phaeophytas (Magallanes et al. 2003).

4
Tabla I. Estudios realizados con el alga Sargassum lapazeanum.
Año Autor Tipo de estudio
1924 Setchell, W.A. & Gardner Taxonomía
1955 Phillips, N. Biogeografía
2006 Rivera, M. & Scrosati Dinámica Poblacional
2008 Andrade-sorcia, G., Riosmena-Rodríguez, R., & Paúl-Chávez, L.
Taxonomía
2008 Rivera, M. & Scrosati, R.. Journal of Phycology 44(1):
45-49.
Dinámica Poblacional
2010 Norris, J.N. Smithsonian Contributions to Botany 94: i-x, 1-276.
Taxonomía
2011 Patrón-Prado M., Casas-Valdez M., Serviere-Zaragoza E., Zenteno-Savín T., Lluch-Cota D. B. y L. Méndes-Rodríguez. Water Air Soil Pollution. 221:137 144.
Biotecnología
2013 Andrade-sorcia, G., Riosmena-Rodríguez, R., Lee K.M, Boo G.H., Muniz-Salazar R., Lopez Vivas J. M., G. González-Barba & S.M. Boo.
Biología Molecular
1.1. Antibacterianos y su clasificación.
Los agentes antibacterianos son químicos que ayudan a eliminar bacterias
patógenas para el paciente que los consume (Sussman y Bates, 2007). De
acuerdo a Goering et al. (2012), existen al menos tres modos de clasificar a los
agentes antibacteriales, uno es por el efecto que tienen sobre las bacterias
(bactericidas y bacteriostáticos). Si el agente antibacteriano elimina las bacterias,
entonces se denomina bactericida y es un proceso irreversible. En cambio, los
agentes bacteriostáticos producen un efecto reversible, sólo previenen el
crecimiento de la población bacteriana. Otro modo de clasificación consiste en
identificar su sitio de acción, es decir en qué parte de la célula actúa, ya sea la
síntesis de la pared celular, de proteínas, de ácidos nucleicos, en rutas
metabólicas o sobre las funciones de la membrana celular. Finalmente, también
pueden clasificarse en base a su estructura química, sin embargo no suele ser una
clasificación práctica debido a la diversidad de estructuras observadas en éste tipo

5
de compuestos. No obstante el presente estudio se limita a realizar el análisis de
la actividad antibacteriana de los extractos analizados y con ello elucidar el posible
potencial de estos contra ciertas bacterias marinas como terrestres.
1.2. El género Sargassum como fuente de compuestos con actividad
antibacteriana
Son muchos los estudios que demuestran el potencial antibacteriano de algas
phaeophytas, entre los que destacan estudios con especies pertenecientes al
género Sargassum, como lo son S. tenerrimum (Kumar et al., 2013), S. boveanum
(Khan y Qari, 2012), S. oligocystum (Tajbakhsh et al. 2012), S. latifolium
(Dashtiannasab et al. 2012) y S. wightii (Vijayabaskar y Shiyamala, 2011). No
obstante, son pocos los compuestos antibacterianos que se han logrado aislar y
caracterizar a partir de este género de algas, destacando el dioctil ftalato (Figura
1) (Sastry y Rao, 1995) y el sulfoglicerolípido 1-0 palmitoil-3-0 (6’-sulfo-α-
quinovopiranosil)-glicerol (Arunkumar et al., 2005), ambos obtenidos a partir del
alga Sargassum wightii. Cabe destacar que esta alga presenta un gran potencial
en este campo, ya que trabajos como los de Vijayabaskar y Shiyamala (2011) han
demostrado que los polifenoles aislados de S. wightii son activos contra al menos
9 patógenos diferentes, pudiendo inhibir cepas de Escherichia coli y Aeromonas
hydrophila, unas de las bacterias más patógenas para el hombre.
Figura 1. Estructura química del dioctil ftalato.
En México se ha estudiado el potencial antibacteriano de algas del género
Sargassum tales como S. cymosum, S. filipendula y S. vulgare en el Golfo de
México y el Caribe Mexicano (Lara-Issasi et al., 1999), S. liebmani en las costas

6
de Oaxaca (Lara-Issasi et al., 1995) y S. filipéndula en las costas de Yucatán
(Freile-Pelegrín y Morales, 2004). No obstante la mayoría de los extractos
obtenidos a partir de estas algas mostraron un bajo potencial antibacteriano contra
cepas de Staphylococcus aeureus, Shygella sonnei, Streptococcus pyogenes,
Streptococcus faecalis y Micrococcus luteus, ya que sólo dos algas mostraron
actividad significativa: Sargassum liebmani contra cepas de S. aeureus (Lara-
Issasi et al., 1999) y S. pyogenes (Lara-Issasi et al., 1995) y Sargassum
filipéndula contra cepas de S. aureus (Lara-Issasi et al., 1999), y Bacillus subtilis
(Freile-Pelegrín y Morales, 2004). Finalmente, cabe destacar que los trabajos
realizados con algas del género Sargassum en la zona del Golfo de California en
México son escasos y sólo estudian las especies S. horridum y S. sinicola, la
primera de ellas activa contra Mycobacterium tuberculosis (Pardo-Fuentes, 2012),
S. aureus y S. pyogenes (Muñoz-Ochoa, 2010), mientras que la segunda contra
Pseudomona aeruginosa, S. aureus y B. subtilis (Castro-Reyes, 1997).
2. Análisis del potencial antioxidante
2.1. Radical Libre DPPH.
La molécula 2,2 Difenil-1-Picrilhidracilo, también conocida como DPPH (Figura 2),
es un radical libre estable que se caracteriza por la deslocalización de un electrón
libre. Dicha deslocalización provee a la molécula de cierta estabilidad, lo que
provoca que esta no se dimerice como es el caso de la mayoría de los otros
radicales libres. Esta característica también da lugar al color violeta oscuro del
DPPH, sin embargo, cuando una solución de este se mezcla con una sustancia
capaz de donar un átomo de hidrógeno, el DPPH se reduce (Figura 3) y suele
presentar una coloración amarillo pálido debido al grupo picril que aún se
encuentra presente (Molyneux, 2004). De esta manera, los ensayos realizados
con DPPH, se basan en el cambio de coloración de la molécula, monitorizando la
disminución de la absorbancia en función de la capacidad antioxidante de los
extractos a diferentes concentraciones y a un tiempo fijo (Mendiola-León, 2008)

7
Figura 2. Difenilpicrilhidracilo (radical libre) Figura 3.Difenilpicrilhidracina
2.2. Algas del género Sargassum como fuente de compuestos antioxidantes
Las algas marinas, al igual que todos los organismos fotosintéticos, se encuentran
expuestas a diferentes combinaciones de luz y altas concentraciones de oxígeno
que conllevan a la formación de radicales libres y otros agentes oxidantes. Los
ácidos grasos poliinsaturados son componentes estructurales de la membrana
tilacoide, esto hace que los elementos de los aparatos fotosintéticos se encuentren
vulnerables al daño fotodinámico. No obstante, la ausencia de este tipo de daño
en las algas, indica que las células de sus aparatos fotosintéticos cuentan con
diferentes mecanismos y compuestos antioxidantes protectores, entre los que
destacan compuestos terpenoides, especialmente de tipo sesquiterpeno y
diterpeno, de los cuales muchos contienen grupos bromofenol (Echeverría et al.,
2009; Vadlapudi, 2012).
Las algas pardas son reconocidas por la producción de una amplia variedad de
metabolitos secundarios, todos ellos con una amplia gama de actividades
farmacológicas, de entre las que destaca su actividad antioxidante (Echeverría et
al., 2009). Un ejemplo de éstos compuestos activos es el sargothunbergol A
(Figura 4A) un nuevo cromeno obtenido por Seo et al. en el 2007 a partir del alga
Sargassum thunbergii. El sargothunbergol A, los tetrapreniltoluquinoles (Figura 4B
y 4C), y los monogalactosil-diacilgliceroles (Figura 4D y Figura 4E) aislados de
ésta misma alga (Seo et al., 2006; Kim et al., 2007), funcionan como eliminador de
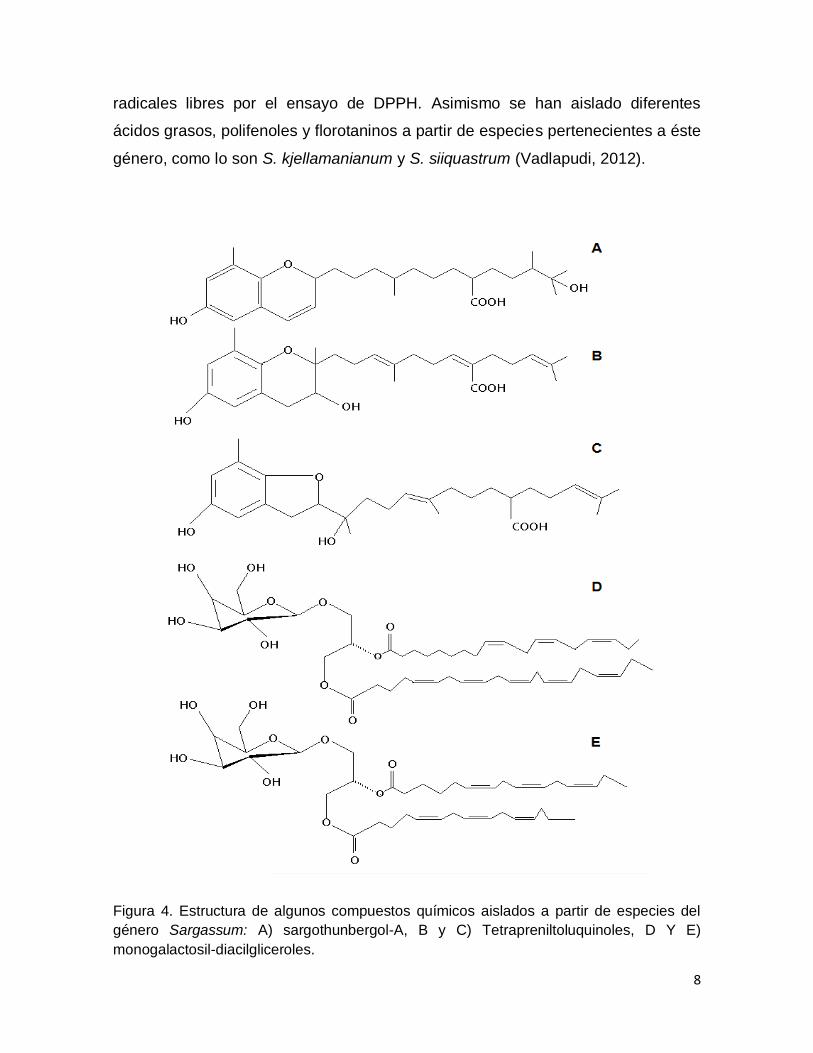
8
radicales libres por el ensayo de DPPH. Asimismo se han aislado diferentes
ácidos grasos, polifenoles y florotaninos a partir de especies pertenecientes a éste
género, como lo son S. kjellamanianum y S. siiquastrum (Vadlapudi, 2012).
Figura 4. Estructura de algunos compuestos químicos aislados a partir de especies del
género Sargassum: A) sargothunbergol-A, B y C) Tetrapreniltoluquinoles, D Y E)
monogalactosil-diacilgliceroles.

9
Aunque son pocos los compuestos antioxidantes ya aislados, identificados y
caracterizados obtenidos a partir de algas marinas, existen muchos estudios
dedicados al análisis de ésta actividad, destacando en México los trabajos
realizados por Zubia et al. en el 2007, quienes estudiaron la actividad antioxidante
en macroalgas marinas tropicales de la península de Yucatán, México, analizando
con ello dos especies de Sargassum, S. pteropleuron y S. ramifolium.
3. Análisis del potencial anticoagulante en algas.
3.1. Polisacáridos sulfatados: Fucoidan y actividad anticoagulante de
Phaeophytas.
Los polisacáridos sulfatados (PS) constituyen una clase de macromoléculas
biológicas con estructuras diversas en la que algunas unidades de monosacáridos
cuentan con grupos semiéster sulfatado (Stewart, 1974). Estos se encuentran en
la pared celular y algunas regiones intercelulares de los organismos, por lo que es
posible aislarlos a partir de fuentes naturales como los tejidos de animales,
plantas superiores, invertebrados marinos, cianobacterias y algas marinas
(Senthilkumar et al., 2013). Entre las propiedades más estudiadas de los
polisacáridos sulfatados están la actividad anticoagulante y antitrombótica. Autores
como Matsubara (2004) han reportado que algunas algas presentan PS con
actividad moduladora de la coagulación sanguínea, por lo que su estudio
representa una alternativa para el desarrollo de fármacos anticoagulantes más
seguros.
Cada división de algas sintetiza su propio tipo característico de polisacáridos, no
obstante, existen variaciones importantes en la estructura, composición y
secuencia monomérica, peso molecular, configuración anomérica, posición del
enlace glucosídico y densidad de cargas, entre cada grupo. Estas diferencias
estructurales dependen en gran medida del entorno y la presión ambiental a la que
estos organismos se encuentran sometidos, ya que le confieren a los polisacáridos
sulfatados distintas propiedades fisicoquímicas responsables de los diferentes
tipos de actividad biológica que son la base de sus aplicaciones en medicina y

10
farmacología. De esta manera las plantas marinas del filo Rodophyceae suelen
distinguirse por la producción de agar, carragenanos, galactanos, xilanos y
porphyran, mientras que las Chlorophytas por presentar estructuras complejas
como los arabinanos sulfatados, galactoarabinoxilanos, galactanos sulfatados y el
ulvan (Stewart, 1974; Muñoz, 2006).
Por su parte las Ochrophytas del orden Phaeophyceae (también conocidas como
algas pardas), se caracterizan por la producción de ácido algínico, laminaran (β-
1,3 glucano), sargassan y principalmente fucanos o fucoidan. Son muchos los
estudios que demuestran la presencia de polisacáridos sulfatados con actividad
anticoagulante extraídos a partir de éste orden de algas (Athukorala et al., 2007;
Gupta y Abu-Ghannam, 2011; Jiao et al., 2011; García-Ríos et al., 2012),
destacando el análisis del fucoidan. Éste último (Figura 5) es un heteropolisacárido
de L-fucosa sulfatada que se encuentra en la pared celular o matriz extracelular de
las phaeophytas pudiendo alcanzar hasta el 40% de su peso seco (Hold y Kraan,
2011). No obstante también se han aislado fucoidanos similares a partir de
invertebrados marinos como medusas, huevos de erizos marinos y pepinos de
mar (Cumashi et al., 2007).
Figura 5. Estructura química promedio del fucoidan
El término fucoidan, se utiliza para definir a una familia de polisacáridos
sulfatados de peso molecular altamente variable. Están compuestos
principalmente de L-fucosa, sulfato y ácidos urónicos, con menor contenido de
otros azúcares como xilosa, galactosa, arabinosa, manosa, glucosa e incluso en

11
ocasiones puede llegar a contener proteínas. No obstante, a pesar de los estudios
sobre su composición y estructura química, aún no ha sido posible definirle
completamente. Esto se debe a que tanto su heterogeneidad y complejidad
estructural se encuentran condicionadas no sólo a diferencias entre las especies,
sino también entre partes de un mismo organismo, por las variaciones
estacionales, las condiciones climáticas locales e incluso por el método de
extracción empleado (García-Ríos et al., 2012). Por lo anterior, cada que se
describe un nuevo fucoidan, se le considera como un compuesto único en cuanto
a sus características estructurales y composición química, siendo posible también
observar variaciones en la actividad biológica (Gómez-Ordóñez, 2012).
Ya que existe una correlación entre el número y la posición de los grupos sulfato
con la actividad biológica de los fucoidanos, es importante el análisis de su
estructura. Este análisis se realiza en el laboratorio por medio de espectroscopía
de infrarrojo, lo que permite determinar la posición de los grupos sulfato en los
polisacáridos, así como la diferenciación de bandas características de sulfatos en
posición axial o ecuatorial dentro de la molécula (Li et al., 2008; García-Ríos et al.,
2012).
De acuerdo a Shanmugam y Mody, para el año 2000 se habían registrado una
mayor cantidad de estudios de actividad anticoagulante en las algas phaeophytas
con más de 60 especies reportadas, en comparación con las rodophytas y
clorofhytas de las que sólo se tenía registro de 40 y 10 especies respectivamente.
En México los estudios de la actividad anticoagulante son escasos en
comparación a los realizados en lugares como Europa, Asia y Estados Unidos. A
pesar de ello, se encontraron más referencias de estudios sobre actividad
anticoagulante realizados con algas pertenecientes al género Sargassum en
México que en otras partes del mundo (Tabla II). No obstante cabe señalar que
todos los trabajos consistían únicamente en el registro de actividad.

12
Tabla II. Listado de algunas especies del género Sargassum que han
presentado actividad anticoagulante.
Especie Área de estudio Referencia bibliográfica
Sargassum cinctum India Shanmugam y Mody, 2000 Sargassum fulvellum Isla Jeju, Korea De Zoysa et al., 2008
Sargassum horneri Isla Jeju, Korea Athukorala et al., 2007
Sargassum coreanum Isla Jeju, Korea Athukorala et al., 2007
Sargassum siliquastrum Isla Jeju, Korea Athukorala et al., 2007
Sargassum thunbergii Isla Jeju, Korea Athukorala et al., 2007
Sargassum stenophyllum Suramérica Duarte et al., 2001
Sargassum hystrix Golfo de México Lara-Issasi y Álvarez-Hernández, 1994
Sargassum cymosum Golfo de México Lara-Issasi y Álvarez-Hernández, 1999
Sargassum filipendula Golfo de México Lara-Issasi y Álvarez-Hernández, 1999
Sargassum pteropleuron Golfo de México Lara-Issasi y Álvarez-Hernández, 1999
Sargassum vulgare Golfo de México Lara-Issasi y Álvarez-Hernández, 1999
Sargassum horridum Golfo de California, México
Muñoz-Ochoa, 2006
3.2. Mecanismo de coagulación sanguínea.
En 1904 Morawitz descubrió que los tejidos vasculares liberan una tromboplastina
tisular (Factor III, o Factor Tisular) necesaria para que se inicie el proceso de
coagulación y propuso los cuatro componentes principales para la coagulación en
plasma: Protrombina, fibrinógeno, calcio y tromboplastina, ideas que prevalecen
en la actualidad (Martínez-Murillo, 2003).
Los estudios realizados para la determinación de la actividad anticoagulante se
han desarrollado conforme los avances en la investigación hemostática, de
manera que actualmente se cuenta con un mejor entendimiento de los diferentes
procesos fisiológicos y bioquímicos que dan lugar a la coagulación, descrita en la
década de los 60 como un proceso enzimático en cascada, mejor conocido
actualmente como “cascada de coagulación” (Figura 6), la cual consta de una

13
serie de cambios bioquímicos y enzimáticos modulados por diferentes factores y
zimógenos que participan en las diferentes etapas del mecanismo de coagulación
para la formación de trombina. La trombina obtenida, actúa sobre el fibrinógeno
liberando fibrina que es el material cementante que genera una red o coágulo
que refuerza el tapón hemostático plaquetario, dando lugar a la coagulación
(Rosenberg y Rosenberg, 1984; Martínez-Murillo, 2003).
Los zimógenos que participan en la coagulación reciben el nombre de cofactores
y son los encargados activar las proteínas plasmáticas denominadas factores de
coagulación. Estos últimos se identifican con números romanos de acuerdo al
orden en que fueron descubiertos, además se les añade el sufijo “a” después del
número romano para indicar la forma activa del factor (Martínez-Murillo, 2003). En
la Tabla III se puede consultar una lista de los diferentes cofactores y factores de
coagulación, así como sus respectivas funciones.
Figura 6. Diagrama que muestra la cascada de coagulación (Martínez-Murillo, 2003).

14
Tabla III. Nombre y función de los diferentes factores y cofactores que actúan en las diferentes etapas de la cascada de coagulación
Factores y cofactores
Sinónimo
Función
FI Fibrinógeno Precursor de fibrina FII Protrombina Zimógeno de proteasa
FT (FIII) Factor tisular o Tromboplastina Iniciador, cofactor de FVIIa en activación de FIX y FX.
FIV Ca2+ Cofactor pro-coagulante
FV Proacelerina o factor lábil Cofactor de Xa en activación de FII
FVI -- --
FVII Autoprotrombina Zimógeno de proteasa
FVIII Factor antihemofílico Cofactor de los factores Ixa en activación de X.
FIX Globulina antihemofílica Zimógeno de proteasa
FX Factor de Stuart-Prower Zimógeno de proteasa
FXI Antecedente tromboplástico del plasma Zimógeno de proteasa
FXII Factor de Hageman Zimógeno de proteasa
FXIII protransglutamidasa, fibrinasa o fibrinoligasa
Factor estabilizante de la fibrina
Precalicreína Factor de Fletcher Cofactor
Cininógeno de alto peso molecular
Factor de Fitzgerald-Williams-Flaujeauc
Cofactor
La coagulación suele ser descrita por dos vías diferentes: intrínseca y extrínseca.
El modelo tradicional de cascada de coagulación describe ambas vías como vías
independientes para la generación del factor FXaA. La vía intrínseca inicia la
coagulación cuando existe un daño en el sistema vascular y las superficies
cargadas negativamente interactúan con los zimógenos FXII y precalicreina (PK) y
el cofactor denominado cininógeno de alto peso molecular (CAMP) (Martínez-
Murillo, 2003).
Por lo tanto la auto-activación del FXII y la PK tienen lugar en el momento en que
la sangre entra en contacto con una superficie artificial como lo son los tubos de
hemodiálisis. Esta es la razón por la cual la sangre formará coágulos en una
jeringa que no contenga ningún agente anticoagulante. Cabe destacar que

15
mientras más negativa sea la carga de la superficie, más rápida será la formación
de coágulos. Se habla de una “auto-activación” del FXII y PK ya que éstos
zimógenos no son dependientes de calcio por lo que seguirán activándose a pesar
de que exista citratro en el anticoagulante (McMichael, 2012).
Por su parte, la vía extrínseca se activa cuando se daña algún tejido diferente al
vascular, provocando la liberación del factor tisular (FT). El FT inmediatamente se
une al FVIIa en plasma, formándose el complejo FT/FVII, mientras que la
autoconversión catalítica del FVII a FVIIa amplifica la respuesta hemostática,
generando así más complejos FT/FVIIa. El complejo FVIIa/FT actúa como
iniciador de la coagulación activando a los factores FIX y FX en una etapa inicial
de activación. Como resultado de esto se obtienen los factores FIXa y FXa. El
primero es necesario para que exista suficiente producción de trombina, mientras
que el segundo es requerido en la activación plaquetaria. El FXa actúa activando a
la protrombina en presencia del factor FVa, fosfolípidos y calcio (complejo
protrombinasa). El resultado de esta reacción es la formulación de la trombina, la
cual al estar presente en la superficie celular, activa otros procesos enzimáticos
como lo son los factores FV, FVIII, FXI y las plaquetas, que a su vez dan como
resultado la formación de fibrina. Cuando el complejo FT/FVIIa da lugar al factor
FX, se activa un inhibidor de la coagulación mediante la vía de inhibición del FT
(IVFT), por lo que se requiere de una vía alterna para producir cantidades
suficientes de trombina (Martínez-Murillo, 2003). Para explicar esto, actualmente
se cuenta con otros modelos de coagulación como es el modelo celular, el cual
describe la coagulación por etapas. Ya que durante la etapa de iniciación la
trombina producida es escasa por la presencia del factor de inhibición (IVFT),
mientras que en las fases de amplificación y propagación se produce una mayor
cantidad debido a la participación de los factores FXI, FVIII y FIX (Díaz-
Concepción y Almagro-Vázquez, 2001)
En la actualidad, se sabe que los sistemas intrínseco y extrínseco de la
coagulación no pueden funcionar de manera independiente uno del otro, ya que
todos los factores de coagulación se interrelacionan entre sí. Además se ha

16
descubierto que algunos pro-coagulantes son realmente cofactores y no poseen
actividad enzimática, como es el caso de los cofactores FV y FVIII (Martínez-
Murillo, 2003; McMichael, 2012). Por lo anterior, los modelos originales en
cascada han sido subsecuentemente modificados para incluir nuevos conceptos y
observaciones, sin embargo ninguno logra explicar el modelo real de la
coagulación in vivo. No obstante, el conocimiento actual del mecanismo de
coagulación sanguínea nos permite comprender de mejor manera el inicio de la
coagulación sin la necesidad de dividir el sistema en vías separadas, sino en una
sola vía que inicia con diferentes factores de coagulación los cuales actúan entre
sí para realizar de manera adecuada el sistema de coagulación sanguínea
(McMichael, 2012).
3.3. Evaluación de la actividad anticoagulante
A pesar de que el modelo de cascada de coagulación no explique completamente
el sistema de coagulación sanguínea, el esquema sigue siendo útil para explicar
las pruebas de laboratorio empleadas para el monitoreo de la hemostasia, o su
prevención. Siendo ésta última de importancia particular para el presente trabajo.
Esto se debe a la posibilidad de simular la cascada de coagulación en condiciones
de laboratorio por medio de dos ensayos: Tiempo de protrombina (TP) para la vía
extrínseca y tiempo de tromboplastina parcial activado (TTPA) para la intrínseca
(Quintana-González, 2002).
El ensayo de TP es sensible a los factores FV, FVII y FX y en menor medida al FII
(Protrombina). El reactivo utilizado en éste ensayo, a menudo denominado
tromboplastina, contiene factor tisular y fosfolípidos. La figura 7 muestra la vía que
mide el tiempo de protrombina, ésta se basa en las reacciones para generar el
activador del FX, el activador de protrombina, la generación de trombina y el paso
de fibrógeno a fibrina donde intervienen los factores antes mencionados. El tiempo
normal en plasma humano para este ensayo suele variar dependiendo del reactivo
utilizado y las condiciones en que se realiza el ensayo, no obstante el tiempo para
este ensayo suele encontrase entre los 11 y 14 segundos. En las condiciones de
la mayoría de las pruebas de TP, la activación del factor X es tan potente, que el

17
ensayo no resulta sensible a la deficiencia de los factores IX u VIII (Kitchen et al.,
2010).
Por su parte la prueba de TTPA consiste en un análisis no específico de la vía
intrínseca. Aunque al utilizarse con otras pruebas resulta eficiente para la
detección en la deficiencia de los factores FVIII, FIX, FXI y FXII. Por si sola, ésta
prueba puede detectar la presencia de anticoagulantes si el tiempo normal para
plasma humano, que suele oscilar entre los 35 y 45 segundos, se ve prolongado
(Kitchen et al., 2010).
Cabe destacar que ambas pruebas se verán alteradas y los tiempos de
coagulación prolongados dependiendo no sólo de los aumentos o deficiencias en
factores de coagulación, sino también de la presencia de inhibidores de la
coagulación como lo son los anticoagulantes (Kitchen et al., 2010).
Figura 7. Vía afectada por la prueba de tiempo de protrombina. Imagen tomada de
Kitchen et al. 2010.

18
III. JUSTIFICACIÓN
Muchos de los tratamientos actuales para los padecimientos trombóticos,
infecciosos y crónico degenerativos son muy efectivos, no obstante con el uso
prolongado suelen generar efectos secundarios nocivos para la salud. Un ejemplo
de esto es la heparina, utilizada para el tratamiento de enfermedades trombóticas,
que si bien es efectiva, su uso por largos periodos de tiempo puede llegar a causar
problemas de infección, así como trombocitopenia, osteoporosis y hemorragias
(Muñoz, 2010). Otro ejemplo son los antioxidantes sintéticos como el Butil-hiroxi-
anisol (BHA), Butil-hidroxi-tolueno (BHT), α-tocoferol y galato de propilo, utilizados
para la reducción de los daños oxidativos en el cuerpo humano, los cuales pueden
provocar diversos efectos secundarios tales como daño hepático y carcinogénesis
(Lee et al. 2010). Además, el uso indiscriminado, así como la prescripción
incorrecta de antibióticos han contribuido al problema de la resistencia bacteriana,
que si bien corresponde a un proceso natural, estos factores lo han acelerado,
conduciendo así al uso de tratamientos que son cada vez más agresivos, de
mayor costo y además pueden generar peligrosos efectos secundarios como lo
son arritmias, hepatotoxicidad o insuficiencia renal aguda (Granowitz y Brown,
2008; Muñoz, 2010).
El daño celular producido por las ERO puede generar una serie de
enfermedades crónico degenerativas, entre las que destacan enfermedades
coronarias, ateroesclerosis, cáncer, diabetes mellitus, envejecimiento prematuro y
enfermedades neurodegenerativas como Alzheimer (Dorado-Martínez et al. 2003;
Li et al. 2007). Este tipo de enfermedades son responsables de la muerte de más
de 36 millones de personas cada año (OMS a, 2013), siendo causa de mortalidad
en todas las edades y constituyendo más de la mitad de la carga mundial de
morbilidad, provocando muchas veces invalidez y deterioro de la vida de las
personas (Castillo-Guzmán et al. 2008). Además la secretaría de Salud (INEGI
2011) declaró que estas enfermedades representan uno de los más importantes
problemas de salud en México, causando la mayor cantidad de muertes al año
principalmente con padecimientos como enfermedades cardiovasculares (ECV) y

19
diabetes mellitus, siendo las ECV consideradas la principal causa de muerte en
todo el mundo. Se calcula que en 2008 murieron por esta causa 17,3 millones de
personas, lo que representa un 30% del total de muertes registradas en el mundo,
de las cuales 7,3 millones de se debieron a la cardiopatía coronaria, y 6,2 millones
a los accidentes vascular cerebral (AVC) los cuales pueden ser ocasionados por
hemorragias de los vasos cerebrales o coágulos de sangre (OMS b, 2013).
Lo anterior ha despertado el interés de los investigadores por la búsqueda de
nuevos compuestos bioactivos de origen natural como una alternativa para el
tratamiento de éstos padecimientos. Las algas, al tratarse de organismos
fotoautróficos expuestos a una mayor cantidad de estrés, han desarrollado
sistemas de defensa eficientes, como lo son la síntesis de metabolitos secundarios
protectores (Mascheck y Baker, 2008), lo que las ha convertido en una de las
principales fuentes de compuestos bioactivos con un amplio espectro de
actividades biológicas de potencial uso en el tratamiento de enfermedades
trombóticas, infecciosas y crónico degenerativas. Es por ello que en la actualidad
los estudios dirigidos al aislamiento de nuevos compuestos biológicamente activos
a partir de macroalgas han incrementado en diferentes partes del mundo (Frikha
et al., 2011). Utilizando estrategia principal la búsqueda de actividad y el
fraccionamiento guiado por bioensayo (Murillo-Álvarez et al., 2001).
México presenta una gran riqueza y diversidad de macroalgas, a pesar de ello los
estudios de su uso para la obtención de compuestos activos con actividad
biológica aún son muy escasos, desaprovechando así la diversidad de este
recurso en el País (Zubia, 2007). La península de Baja California cuenta con una
gran variedad de hábitats y condiciones climáticas, por lo que alberga una enorme
riqueza de algas marinas (Suárez Castillo et al., 2013), destacando la presencia
de los algas pertenecientes al género Sargassum debido a la alta dominancia en
biomasa. No obstante, de las 5 especies de Sargassum que se encuentran
conformando los bosques submarinos del Golfo de California, sólo S. horridum y
S. sinicola cuentan con registros de estudios para el análisis de su actividad
biológica. Por lo anterior, impulsados por la falta de documentación sobre las

20
propiedades químicas de S. lapazeanum, se seleccionó la especie para el estudio
de su actividad biológica como fuente de compuestos antibacterianos,
antioxidantes y anticoagulantes. Éstos estudios no sólo son esenciales para el
entendimiento de la forma de vida del organismo y su interacción con el medio
(Amsler, 2008), sino que también contribuyen al conocimiento de las algas
pertenecientes al género Sargassum y el futuro desarrollo de nuevos fármacos o
productos funcionales como cosméticos y suplementos alimenticios a partir de las
mismas (Querellou et al., 2010).

21
IV. OBJETIVOS
1. Objetivo general
Evaluar el potencial de Sargassum lapazeanum como fuente de compuestos con
actividad antioxidante, antibacteriana y anticoagulante.
2. Objetivos particulares
1) Obtener y fraccionar el extracto por medio de distintas técnicas
cromatográficas.
2) Evaluar la actividad antibacteriana por medio del método de difusión en
agar con discos.
3) Evaluar la actividad antioxidante, utilizando el radical libre estable 2,2-
difenil-1-picrilhidracilo (DPPH).
4) Obtener el fucoidan y evaluar su la actividad anticoagulante por medio de
los ensayos de tiempo de protrombina (TP) y tiempo de tromboplastina
parcial activada (TTPA)

22
V. MATERIAL Y MÉTODOS
1. Recolección e identificación taxonómica.
El alga Sargassum lapazeanum se recolectó manualmente a una profundidad
máxima de un metro, en la zona intermareal de San Juan de la Costa (La Paz,
B.C.S) (Figura 8). Posteriormente fue lavada con agua corriente para eliminar los
epífitos y restos de sedimento presentes, y se dejó secar al sol para facilitar su
maceración y tamizado.
La identificación taxonómica se realizó en el laboratorio de botánica marina de la
Universidad Autónoma de Baja California Sur, con el apoyo de la Dra. Gabriela
Andrade Sorcia.
Figura 8. Mapa que muestra la zona de recolecta del alga S. lapazeanum en San Juan de
la Costa, Bahía de La Paz B.C.S, México. Modificado a partir de NOAA.
http://www.ngdc.noaa.gov/mgg/shorelines/shorelines.html.

23
2. Análisis proximal del alga S. lapazeanum.
El análisis proximal del alga se realizó por medio de servicio externo, determinando el
porcentaje de humedad por diferencia de peso, calentando a 105°C durante 4 hrs y el de
cenizas por diferencia de peso y calcinación a 600°C durante 5 hrs. Para la determinación
de proteínas se utilizó el método de Dumas (equipo LECO FP528), mientras que el
cálculo del extracto etéreo se realizó por el método Soxtec-Avanti 2050 (TECATOR) y el
contenido de fibra cruda por medio de hidrólisis sucesiva (ácido/base). Finalmente el
extracto libre de Nitrógeno (ELN) y la energía (cal/g) se calcularon utilizando un
calorímetro.
3. Obtención del extracto
3.1. Obtención del extracto etanólico
Se maceraron trescientos gramos del alga previamente seca y molida con EtOH
100 %, realizando recambios cada tercer día durante dos semanas. La mezcla se
separó por filtración y la solución obtenida fue concentrada a sequedad a 40 °C y
presión reducida en un rotavapor Yamato. Finalmente el extracto crudo (12-009-
EC) resultante, fue sometido a un fraccionamiento sólido-líquido (Figura 9).
3.2 Obtención del extracto acuoso: Fucoidan
Para la extracción del fucoidan se pesaron 100 g del alga previamente seca y
molida, a los cuales se les agregó 600mL de agua destilada y se dejó en baño
María con agitación continua durante 2 hrs a una temperatura constante de 55ºC,
Posteriormente se removió el tejido algal por medio de filtración simple,
obteniendo 500 mL de solución la cual fue centrifugada durante media hora. El
centrifugado fue recuperado por decantación y precipitado con 40 mL de una
solución de Cl2Ca al 10% previamente preparada, con el fin de remover el
alginato disuelto, el cual fue removido por centrifugación a 1770 x g durante 30
minutos. La solución clarificada (400 mL) se precipitó con tres volúmenes de EtOH
(1200 mL). El precipitado final se recuperó por centrifugación, se dejó secar en
una estufa eléctrica a 50ºC durante un día y se etiquetó como 12-009-FC (Figura
10).

24
4. Fraccionamiento
4.1 Fraccionamiento por gradiente de polaridad.
Se fraccionaron quince gramos del extracto crudo en una columna cromatográfica
empacada con sílica gel fase normal, eluyendo la columna con un gradiente de
polaridad compuesto por CH2Cl2 100, CH2Cl2 - EtOH 90:10, CH2Cl2 - EtOH 25:75,
CH2Cl2 - EtOH 50:50, EtOH 100, EtOH:H2O 50:50 y H2O 100%. Los eluatos se
compararon analizando la similitud por medio de cromatografía en capa fina
(Anexo III) y se concentraron en siete fracciones de la 1-7 respectivamente (Figura
9).
Figura 9. Obtención del extracto crudo (EC) y fraccionamiento del alga S. lapazeanum.

25
Figura 10. Esquema que muestra el proceso de obtención y fraccionamiento del fucoidan.

26
4.2. Fraccionamiento de F2 en columna cromatográfica.
La fracción más activa en los ensayos de actividad se seleccionó para un segundo
fraccionamiento en columna cromatográfica con sílica gel en diclorometano
(Figura 9). Para ello se utilizó una relación 1:60 de muestra-sílica, es decir 600 mg
de la fracción más activa se disolvieron en diclorometano para ser adsorbido en
36g de sílica gel, utilizando como eluyente una fase móvil en gradiente de
polaridad creciente con los siguientes sistemas: CH2Cl2 100%, CH2Cl2:MEtOH
95:5, CH2Cl2:MEtOH 90:10, CH2Cl2:MEtOH 80:20, CH2Cl2:,EtOH 70:30,
CH2Cl2:MEtOH 50:50, EtOH 100% y H2O 100%.
4.3. Purificación parcial del fucoidan.
La purificación parcial del fucoidan (FC) obtenido se realizó por medio de
precipitación fraccionada con 3 volúmenes de etanol, recuperando el precipitado
entre adición y adición por medio de centrifugación. Los sólidos obtenidos en cada
precipitación se secaron en estufa eléctrica a 55ºC y se almacenaron para el
posterior análisis de su actividad biológica (Figura 10).
5. Pruebas de Actividad Biológica.
Tanto el extracto etanólico crudo como sus fracciones fueron sometidas a los
ensayos de actividad antibacteriana por el método de difusión con discos y al
ensayo de actividad secuestrante de radicales libres utilizando el método del
radical libre estable 2,2-difenil-1-pricilhidracilo (DPPH).
5.1. Evaluación de la actividad secuestrante de radicales libres (DPPH):
Ensayo de actividad antioxidante.
Se evaluó la actividad secuestrante de radicales libres del extracto crudo, así
como de las fracciones, utilizando el radical libre estable 2,2-difenil-1-picrilhidracilo
(DPPH) por medio de dos métodos:1) Bioautográfico y 2) Colorimétrico

27
Método bioautográfico
Para este ensayo se siguió la metodología descrita por Choi et al. (1994), que
consiste en un análisis cromatográfico de placa fina (TLC), depositando 10 µl de
una solución de cada uno de las fracciones a evaluar sobre una placa de sílica gel
fase normal, la cual se desarrolló en una cámara de saturación utilizando una
solución de diclorometano-metanol a una relación 9:1 como eluyente.
Posteriormente se dejó secar la placa y se roció de manera uniforme con una
solución metanolica de DPPH al 0.4 %. La placa cromatográfica se dejó reposar
en oscuridad durante 30 min. y después de este tiempo se registraron las zonas
decoloradas sobre la placa como indicativos de la actividad secuestrante.
Método colorimétrico
Para los ensayos de colorimetría se utilizó una solución de DPPH al 0.02 % en
metanol. Se prepararon las fracciones por duplicado en una solución con etanol a
concentraciones de 400, 200, 100, 50 y 25 µg mL-1, agregando 4 mL de la solución
de DPPH. Además se preparó un blanco con el que se obtuvo el factor de
corrección del color. Una vez hecho esto se dejaron en oscuridad durante media
hora y posteriormente se realizaron las lecturas en un espectrofotómetro Milton
Roy Company SPECTRONIC 20D a una longitud de onda de 517nm. A partir de
estos valores se obtuvieron curvas dosis-efecto graficando el porcentaje de
reducción del DPPH en relación a la concentración de extracto utilizado. Este
porcentaje se obtuvo con la siguiente fórmula:
AB: Absorbancia del Blanco
AM: Absorbancia de la muestra corregida (AM= A-FC)
A= Absorbancia
FC= Factor de corrección de color.

28
Además se calculó la concentración efectiva media (EC50) mediante la ecuación
de la recta obtenida en las curvas dosis-efecto realizadas para cada uno de los
extractos, utilizando ácido ascórbico como estándar de referencia.
5.2. Evaluación de la actividad antibacteriana por medio del método de
difusión en agar con discos.
La actividad antibacteriana fue evaluada por el método de difusión en agar con
discos Kirby-bauer (Jorgensen y Turnidge, 2007) contra 3 bacterias terrestres y 2
marinas. En base a lo anterior, se prepararon pequeños discos de papel filtro
Whatman No.4 de 6.4 mm impregnados con 2mg de las fracciones a probar. Una
vez impregnados los discos, estos se colocaron sobre la superficie de placas de
agar Mueller-Hinton (Bioxon) preparadas con [NaCl 25%] y sin sal, las cuales
fueron previamente inoculadas con una suspensión de los microorganismos de
prueba: Staphylococcus aureus, Escherichia coli, Bacillus subtilis, Vibrio harveyi y
Vibrio parahaemoliticus, a una concentración de 1x10-8 células mL-1.
Para la inoculación se utilizó el método de siembra por estrías, realizando dos
replicas para cada extracto. Las placas se incubaron a una temperatura de 37ºC
durante 24 h y al término de la incubación se midió el halo de inhibición y se
registró el resultado en una tabla (Tabla VI). En base a los resultados obtenidos
de este ensayo, se repitió el mismo proceso para las fracciones obtenidas a partir
de la fracción F2, las cuales sólo se probaron contra las cepas de bacterias del
género Vibrio previamente analizadas (Tabla VII).
5.3. Análisis de la actividad Anticoagulante
La actividad anticoagulante del fucoidan, se evaluó por medio de los ensayos de
tiempo de protrombina (TP) y tiempo de tromboplastina parcial activada (TTPA),
utilizando plasma sanguíneo obtenido de humanos sanos, siguiendo las
instrucciones del fabricante. Todos los extractos fueron ensayados por duplicado.
La sangre utilizada en ambas pruebas se obtuvo por venopunción y se mezcló con
una solución de citrato de sodio al 3.5% en una proporción de 9 a 1 (v/v). El

29
paquete celular se removió por medio de centrifugación, mientras que el plasma
recuperado se almacenó en refrigeración hasta el momento de su utilización.
El ensayo de tiempo de protrombina (TP) consistió en mezclar 100 μL de plasma
humano citrado con 10 μL de una solución del extracto de prueba, dejando incubar
la mecla por 1 min. a 37°C. Una vez concluido el tiempo de incubación, se
agregaron 200 μL del reactivo de TP (Biopool® by Trinity Biotech, plc)
preincubado a la misma temperatura (37°C) por 10 min., y se midió el tiempo de
formación del coágulo a partir de ese momento.
Por su parte para el ensayo de tiempo de tromboplastina parcial activada (TTPA),
se mezclaron 100 μL de plasma humano citrado con 10 μL de una solución del
extracto crudo de prueba e igualmente se dejó incubar la mezcla por un periodo de
1 min a 37°C. Después del tiempo de incubación, se agregaron 100 μL del
reactivo de TTPA (Biopool® by Trinity Biotech, plc), la nueva mezcla se dejó
incubar por 3 min a 37°C y se agregaron 100 μL de CaCl2 al 0.25%, midiendo el
tiempo de formación del coagulo a partir de ese momento. Finalmente la
inspección de la formación del coagulo en ambos ensayos, se realizó de manera
visual, registrando los tiempos en una tabla (Tabla VIII)
6. Caracterización parcial del fucoidan.
Las fracciones obtenidas a partir de la purificación del fucoidan, se caracterizaron
estructuralmente por medio de espectroscopia de infrarrojo (IR). Además se
calculó el contenido de sulfatos por el método de línea base a través de las
relaciones del área bajo la curva de las señales a los 1250 y 1040 cm-1 (Lijour et
al., 1994).

30
VI. RESULTADOS
1. Análisis proximal
El análisis proximal mostró que Sargassum lapazeanum se encuentra conformado
principalmente por un alto contenido de material inorgánico y carbohidratos y un
bajo contenido de extracto etéreo. Esto se observa en la Tabla IV, donde se
presenta que S. lapazeanum cuenta con los valores más elevados para ELN
(extracto libre de nitrógeno) y cenizas.
Tabla IV. Resultados de análisis proximal del alga.
Humedad (%)
Proteína (%)
Extracto Etéreo
(%)
Fibra Cruda
(%)
Cenizas (%)
ELN (%)
Energía
Cal/g
8.51 8.54 0.28 6.06 30.52 85.12 2522.34
0.03 0.07 0.03 0.15 0.1 - 4.86
2. Análisis de la actividad biológica
2.1. Actividad antibacteriana
Los resultados del ensayo antibacteriano, indican una actividad selectiva sobre
bacterias del género Vibrio (Tabla V), donde el extracto crudo (EC) y la fracción 2
(F2) mostraron la mayor actividad con un halo de inhibición de 8.5 ± 0.70 y 8 ± 0
para V. parahaemolyticus, y de 10 ± 0 y 8.5 ± 0.70 para V. harveyi
respectivamente. Además las fracciones F6 y F7 presentaron una actividad
comparable contra V. harveyi, con halos de inhibición de 8.75 ± 0.35 y 9 ± 0
respectivamente. Las fracciones F3 y 4 F4 también mostraron halos de inhibición
aunque menos representativos cuando fueron probados contra V. harveyi,
mientras que las fracciones F6 y F7 tuvieron un efecto similar contra V.
parahaemolyticus.
En base a lo anterior, considerando la relación entre actividad y rendimiento de la
muestra, se seleccionó F2 para su fraccionamiento por cromatografía en columna.
Las fracciones obtenidas a partir de F2 mostraron tener una mayor actividad sobre

31
V. harveyi (Tabla VI), siendo F2F9 la fracción más activa con un halo de inhibición
de 11.75 ± 0.35. Mientras que la fracción F2F8 resultó ser más activa sobre V.
parahaemolyticus con un halo de inhibición de 8.5 ± 0.71. Cabe destacar que sólo
se probaron aquellas fracciones que contaban con muestra suficiente para la
realización de las pruebas antibacterianas.
Tabla V. Resultados de la prueba de difusión en agar con discos [2 mg/disco] del extracto crudo y las fracciones obtenidas a partir de S. lapazeanum. Se reporta el promedio del halo de inhibición en milímetros ± sd, n = 2 .
Halos de inhibición (mm)
Gram + Gram -
Fracciones S. aureus B. subtilis E. coli V. parahaemolitycus V. harveyi
F1 - - - - - F2 - - - 8 ± 0 8.5 ± 0.70 F3 - - - - * F4 - - - - * F5 - - - - - F6 - - - * 8.75 ± 0.35 F7 - - - * 9 ± 0 EC - - - 8.5 ± 0.70 10 ± 0
- No Activo, * Ligera actividad
Tabla VI. Resultados de la prueba de difusión en agar con discos [2 mg/disco] del fraccionamiento del extracto F2 obtenido a partir de S. lapazeanum. Se reporta el promedio del halo de inhibición en milímetros ± sd, n = 2 .
Halos de inhibición (mm)
Fracciones V. harveyi V. parahaemolitycus
F2F1 N.P N.P F2F2 N.P N.P F2F3 N.P N.P F2F4 N.P N.P F2F5 - - F2F6 10.75 ± 1.06 7.75 ± 0.35 F2F7 N.P N.P F2F8 11.75 ± 1.06 8.5 ± 0.71 F2F9 11.75 ± 0.35 8 ± 0.71 F2F10 11.5 ± 0.71 7.75 ± 0.35 F2F11 - - F2F12 10 ± 0 * F2F13 N.P N.P
F2F14 N.P N.P
N.P No se probó, - No Activo, * Ligera actividad.

32
2.2. Actividad antioxidante
El método bioautográfico reveló una franja decolorada a lo largo de la placa
cromatográfica, indicativo de actividad antiradicalaria en la mayoría de las
fracciones (Figura 11). Lo anterior, fue corroborado por medio del método
colorimétrico, el cual demostró el potencial de las fracciones, destacando la
fracción 6 con la menor EC50 de 39.96 µg mL-1 (Figura 12 y 13), mientras que la
más elevada fue la fracción F1 con un EC50 de 394.21 µg mL-1.
Figura 11. Cromatografía en placa fina que muestra la actividad antioxidante de S.
lapazeanum por el método bioautográfico, revelada con una solución de DPPH al 0.4%.
EC= Extracto crudo.
Figura 12. Gráfica comparativa de los valores obtenidos para las EC50 en las fracciones
obtenidas por gradiente de polaridad a partir de S. lapazeanum. 1= Menor polaridad, 7=
Mayor polaridad E.C.= Extracto Crudo
0
100
200
300
400
1 2 3 4 5 6 7 E.C.
EC50
[µ
g m
L-1]
Fracciones de S. lapazeanum
EC F1 F2 F3 F4 F5 F6 F7

33
Figura 13.- Curva dosis efecto la fracción F6 de S. lapazeanum, donde se observa el
porcentaje de reducción del DPPH (%) en relación a la concentración del extracto (µg mL -
1). En negro: línea de tendencia y desviación estándar. En rojo: concentración efectiva
media (EC50).
Figura 14.- Curva dosis efecto del ácido ascórbico (AA), utilizada como blanco de
comparación, donde se observa el porcentaje de reducción del DPPH (%) en relación a la
concentración del AA (µg mL-1). En negro: línea de tendencia y desviación estándar. EC50,
obteniendo por extrapolación= 1.5 µg mL-1
Finalmente, la curva control realizada con ácido ascórbico presentó una EC50 de
1.5 µg mL-1 (Figura 14); mientras que la fracción más activa de S. lapazeanum
(Figura 13) demostró una actividad 26 veces menor a éste valor, con una EC50 de
39.96 µg mL-1.
39.96
y = 0.5709x + 27.186 R² = 0.9726
0
20
40
60
80
100
0 20 40 60 80 100 120
Po
rce
nta
je d
e r
ed
ucc
ión
de
l D
PP
H (
%)
Concentración del extracto [µg mL-1]
DPPH
y = 23.325x - 14.965 R² = 0.9823
45.00
50.00
55.00
60.00
65.00
70.00
75.00
80.00
85.00
3 3.2 3.4 3.6 3.8 4
Po
rcen
taje
de
red
ucc
ión
d
el D
PP
H (
%)
Concentración del extracto [µg mL-1]
DPPH
Lineal (DPPH)

34
2.3. Actividad Anticoagulante
A partir del proceso de extracción y precipitación fraccionada del extracto acuoso
de Sargassum lapazeanum se obtuvieron un total de 3 extractos (FC1, FC2 y
FC3). Como puede observarse en la Tabla VII los extractos obtenidos, así como el
extracto acuoso de fucoidan (FC) presentaron una alta actividad en ambas
pruebas. Todos los extractos analizados sobrepasaron el tiempo límite de
coagulación establecido (300 segundos) para la prueba de TTPA. Mientras que
para la prueba de TP, el extracto FC2 fue el que mostró una mayor actividad con
un tiempo de coagulación de 41.93 ± 2.49 s.
Tabla VII. Actividad anticoagulante de los
extractos obtenidos por precipitación
fraccionada a partir de S. lapazeanum.
Extracto TTPA TP
Control 33.54 14
FC >300 30.25 ± 0.79
FC1 >300 24.50 ± 1.30
FC2 >300 41.93 ± 2.49
FC3 >300 23.079 ± 1.90
En base a lo anterior, se realizaron curvas dosis-efecto (Figura 15) de los
extractos (FC, FC1, FC2 y FC3) las cuales confirmaron el alto potencial
anticoagulante de S. lapazeanum, pues la mayoría de los extractos duplicó el
tiempo de coagulación establecido por el control (32.84s) incluso a sus
concentraciones mínimas.

35
Figura 15. Gráfico que muestra la relación entre la concentración de los extractos
obtenidos a partir de la precipitación fraccionada del Fucoidan (FC) de S. lapazeanum y
su efecto sobre el tiempo normal de coagulación según el ensayo de TTPA.
2.3.1. Caracterización parcial del fucoidan
El análisis del fucoidan por medio de IR indicó la presencia de diferentes bandas
de absorción que identifican el fucoidan obtenido como un heterofucano con un
contenido de 27.85% de sulfato para el fucoian crudo FC y de 8.10, 47.48 y 12.98
% para las fracciones FC1, FC2, y FC3 obtenidas por medio de precipitación
fraccionada.
0
50
100
150
200
250
0 5 10 15 20 25 30
TTP
A (
S)
Concentración del extracto [µg mLˉ¹]
FC1 FC2 FC3 FC Control

36
VII. DISCUSIÓN
1. Análisis proximal del alga.
Como se puede observar en la Tabla IV, los constituyentes principales de S.
lapazeanum fueron el ELN y las cenizas. El ELN obtenido fue de 85.12%, esta
porción suele estar conformada principalmente por carbohidratos solubles como
almidón y azúcares pero también incluye una considerable cantidad de agar, ácido
algínico y polisacáridos sulfatados (Carrillo-Domínguez, 2002;). Estos son de gran
importancia en la industria alimenticia ya que carbohidratos como el almidón y
laminaran son fácilmente utilizados por los humanos y animales debido a los
enlaces que presentan de tipo α-glicosídico 1,4 y 1,6, así como 1,3. Otros
polisacáridos como el ácido algínico y el agar sólo pueden ser utilizados por
animales que presenten las enzimas específicas para ello. No obstante estos
polisacáridos son altamente utilizados en la industria alimentaria y farmacéutica
por sus diferentes propiedades gelificantes (Carrillo-Domínguez, 2002). Además el
fucoidan obtenido a partir de algas pardas se caracteriza por presentar una gran
gama de actividades biológicas de interés en la industria farmacéutica, cosmética
y alimentaria (Li et al., 2008), un ejemplo de ello es la comercialización del
fucoidan líquido como aditivo inmuno-estimulatorio para camarones (Casas, 2006).
S. lapazeanum presentó además un alto contenido de materia inorgánica, con un
porcentaje de cenizas de 30.52 % esto es comparable con lo obtenido por otros
estudios en plantas marinas y ocrophytas (Carrillo-Domínguez, 2002; Casas-
Valdez, 2006), los cuales indican que los valores elevados en el contenido de
materia orgánica se deben a la capacidad de estos organismos para almacenar
elementos minerales, los cuales abundan en el ambiente marino.
Los resultados obtenidos mostraron un bajo valor de proteína de 8,54%, sin
embargo éste valor sobrepasa lo reportado por Marín (1999), Casas et al. (2002) y
Casas et al. (2006) de 7,0, 7,7, y 6.09% respectivamente para algas del género
Sargassum. Estas diferencias observadas podrían deberse a factores como la
especie y la época de recolecta del alga, ya que estos organismos presentan

37
variaciones estacionales que afectan la concentración de sus constituyentes. A
pesar de que el contenido de proteína es reportado en la literatura como “bajo”
para algas phaeophytas, cabe destacar que estudios realizados con algas del
género Sargassum han demostrado que la calidad de estas proteínas es buena,
pues presenta una gran cantidad de aminoácidos esenciales y su digestibilidad es
mayor al 70% (Casas, 2006 ).
La cantidad de fibra cruda observada fue del 6.06%,, a pesar de ser un valor bajo,
debe considerarse que el método subestima el contenido de fibra dietética, ya que
se disuelve gran parte de la hemicelulosa y lignina, así como cantidades variables
de celulosa y toda la fibra soluble (Marín, 1999), por lo que se sugiere analizar el
contenido de fibra dietética por medio de otras metodologías. Algunos alimentos
de consumo humano como cereales y algunos vegetales contienen valores de
fibra similares, tal es el caso de la avena 7% y algunas leguminosas como el frijol
y el garbanzo con un 5% de fibra cruda (Carrillo-Domínguez, 2002).
La calidad y digestibilidad de las proteínas, así como el bajo contenido energético
y de extracto etéreo de S. lapazeanum, le proporcionan al alga un alto valor
nutricional en la industria alimentaria. Lo anterior, aunado a su potencial actividad
biológica hacen de esta alga un recurso explotable para la elaboración de
diferentes tipos de suplementos.
2. Análisis de la actividad antibacteriana
La mayoría de los trabajos realizados con extractos crudos de algas marinas,
mencionan una gran actividad contra bacterias gram positivas, considerando S.
aureus como una de las especies más susceptibles a los extractos algales (Ríos et
al., 2009). Esto probablemente debido a la estructura más compleja de la pared
celular en bacterias gram negativas (Demirel et al., 2009). No obstante, el extracto
crudo y los extractos obtenidos por medio del fraccionamiento sólido-líquido de S.
lapazeanum no fueron activos contra las bacterias gram positivas utilizadas en el
ensayo.

38
Aunque son más los estudios que demuestran la actividad de las especies
pertenecientes al género Sargassum contra bacterias gram positivas,
particularmente S. aureus (Lara-Issasi et al., 1995; Castro-Reyes, 1997; Muñoz-
Ochoa, 2010; Vijayabaskar y Shiyamala, 2011; Kumar et al., 2013), también hay
estudios que reportan una baja o nula actividad contra éste patógeno. En este
sentido, nuestros resultados concuerdan con lo observado por investigadores
como Lara-Issasi et al., (1999) quienes reportaron que algunas especies de
Sargassum del Caribe y Golfo de México (S. cymosum y S. vulgare) no
presentaron actividad contra S. aureus, explicando que estas diferencias de
actividad podrían no sólo deberse a la especie, sino también a un
enmascaramiento en los extractos de los metabolitos secundarios responsables
de esta actividad. Por lo anterior, aunque los extractos presentaron una actividad
selectiva contra las bacterias del género Vibrio, es probable que al purificar las
otras fracciones, éstas incrementen la actividad actual o presenten actividad
contra bacterias que no presentaron antes.
Además, especies del género Sargassum han demostrado presentar un alto
potencial contra bacterias del género Vibrio, entre ellas destacan Sargassum
oligocystum (Nuestro-Baleta et al., 2011) y Sargassum latifolium (Dashtiannasab
et al., 2012). En este último estudio se demostró que Sargassum latifolium
presenta halos de inhibición de 9±0.7 y 9.2±1.1 contra V. parahaemolyticus y V.
harveyi respectivamente a concentraciones de 200 mgl-1. Esto es 10 veces mayor
a lo obtenido en el presente estudio. No obstante, estas diferencias de actividad
pueden explicarse por diversos factores, tales como la especie, la variación
estacional, la variación geográfica e incluso el estado reproductivo del alga (Lara-
Issasi et al., 1999).
Algunos estudios demuestran que Vibrio harveyi suele ser más sensible a los
ensayos de actividad antibacteriana, presentando en la mayoría de los casos
halos de inhibición más definidos que Vibrio parahaemolyticus (Nuestro-Baleta et
al. 2011; Dashtiannasab et al., 2012). Lo anterior es similar a lo obtenido en el
presente estudio, donde la mayor actividad observada se registró contra Vibrio

39
harveyi (Tabla V y VI) tanto en el extracto crudo (EC) como en la fracción F2
obtenida con diclorometano-etanol (9:1). Esto concuerda con estudios previos, los
cuales indican que el uso de solventes orgánicos provee una mayor eficiencia en
la extracción de compuestos con actividad antibacteriana comparado a aquellos
que utilizan agua (Tüney et al., 2006). Además estudios recientes elaborados por
Al-Saif et al. (2014) reportan que las extracciones etanólicas permiten la
obtención de compuestos con una mayor actividad antibacteriana, no obstante
otros autores como Tüney et al. (2006) sugieren el cloroformo como mejor
solvente para la extracción.
La actividad antibacteriana de S. lapazeanum se presentó tanto en fracciones
polares como no polares (Tabla V). No obstante, se seleccionó una de las
fracciones apolares (F2) para su posterior fraccionamiento debido a que mostró la
mayor actividad con un halo de inhibición de 8 ± 0 para V. parahaemolyticus, y 8.5
± 0.70 para V. harveyi. Por su parte, las fracciones obtenidas por cromatografía en
columna a partir de F2, presentaron un incremento en la actividad antibacteriana
con halos de inhibición más definidos, siendo F2F9 la fracción más activa contra
V. harveyi (11.75 ± 0.35) y F2F8 contra V. parahaemolyticus (8.5 ± 0.71) (Tabla
VI). Esto demuestra que el proceso de fraccionamiento fue efectivo, de manera
que purificó el extracto incrementando así su actividad.
Los principales compuestos bioactivos producidos por las algas marinas se
conforman por una gran variedad de metabolitos secundarios, los cuales según
Magallanes et al. (2003), presentan una función específica dentro de su medio,
entre las que se encuentran la defensa química contra herbívoros marinos, así
como la disminución de epífitos mediante la inhibición de los organismos
competidores y patógenos microbianos. Se ha reportado que la actividad
antibacteriana en diferentes tipos de plantas y algas marinas se debe a la
presencia de metabolitos de tipo terpenoides (sesterpenoides), fenoles,
polifenoles, o incluso hidroquinonas. Estudios realizados con especies del orden
phaeophyta, demuestran la presencia de compuestos antibacterianos
halogenados como los bromofenoles y compuestos lipofílicos conformados de

40
ácidos grasos saturados e insaturados con la predominancia de los ácidos
palmítico, mirístico, oleico y eicosapentaenico. Además, este tipo de ácidos grasos
ha demostrado ser efectivo contra un amplio espectro de bacterias patogénicas
gram positivas y negativas (Al-Saif et al., 2014). Por lo que la actividad
antibacteriana observada en los extractos más activos, podría atribuirse a alguno
de los compuestos mencionados anteriormente.
No obstante, los resultados obtenidos no permiten identificar qué compuestos o
mezcla de compuestos se encuentran dando lugar a esta actividad, por lo cual se
sugiere realizar un segundo fraccionamiento con las muestras F2F8 y F2F9. Esto
no sólo podría incrementar la actividad registrada, sino que también permitirá
determinar si la actividad de S. lapazeanum sobre bacterias del género Vibrio se
debe a alguno de los compuestos ya reportados para algas del género, o si se
trata efectivamente de un compuesto novedoso.
Los compuestos antibacterianos aislados a partir de algas del género Sargassum
son muy escasos, ya que sólo se encontró registro de la obtención del ácido
mirístico (Muñoz-Ochoa, 2010), el dioctil ftalato extraído con cloroformo-metanol
(2:1) (Sastry y Rao, 1995) y el sulfoglicerolípido 1-0 palmitoil-3-0 (6’-sulfo-α-
quinovopiranosil) -glicerol aislado con metanol (Arunkumar et al., 2005), siendo los
últimos dos obtenidos a partir del alga Sargassum wightii. Esto sugiere que aún
queda mucho por hacer en la investigación de compuestos antibacterianos en
algas de este género. Además, se debe considerar que las sustancias contenidas
en las algas que presentan propiedades antimicrobianas suelen fluctuar en
cantidad a lo largo del año, siendo muy probablemente de naturaleza distinta, ya
que se extraen con solventes de diferente polaridad. Por lo que se recomienda
elaborar un estudio de la variación estacional de éstos compuestos que
complemente los resultados obtenidos.
Finalmente, la actividad selectiva contra bacterias del género Vibrio observada en
los ensayos, permite considerar a S. lapazeanum como una alternativa en
acuacultura para la obtención de compuestos antibacterianos naturales con
aplicaciones en el tratamiento contra la vibriosis, una de las principales causas de

41
enfermedades y pérdidas económicas en granjas acuícolas, que suele afectar
principalmente los cultivos de camarón. Esta enfermedad genera lesiones en la
cutícula, apéndices y branquias de los camarones, infecciones localizadas en las
heridas, perdidas de miembros, así como musculatura blanda, que al no ser
tratada adecuadamente puede causar pérdidas de producción aproximados al
100% (Venkateswara, 2008; Limonta y Coffigny, 2012).
3. Análisis de la actividad antioxidante
Los ensayos de actividad secuestrante de radicales libres, demostraron que S.
lapazeanum presenta un alto potencial como fuente de compuestos con actividad
antioxidante. Esto coincide con diversos estudios que indican el alto potencial de
las algas pertenecientes al género Sargassum como fuente de este tipo de
compuestos (Anggadiredja y Andyani, 1997; Budhiyanti, 2012; Zhang et al, 2012;
Agardh, 2013). Los resultados obtenidos en el presente estudio son similares a
aquellos reportados por Zubia y colaboradores (2007) quienes obtuvieron EC50 de
6.64 y 7.14 µg mL−1 para S. ramifolium y S. pteropleuron respectivamente, siendo
estos resultados 6.01 y 5.59 veces más activos que el extracto de mayor actividad
obtenido en el presente estudio, el cual presentó una EC50 de 39.96 µg mL-1.
Estas variaciones de actividad podrían deberse a la composición química de las
algas analizadas, así como a los solventes utilizados durante el método de
extracción, ya que estudios recientes demuestran que la actividad secuestrante de
radicales libres a partir de extractos de Sargassum se ve afectada por diferentes
factores, tales como la especie, la zona y la temporada de colecta, así como el
método de extracción utilizado (Budhiyanti, 2012).
Además puede observarse que la actividad antiradicalaria fue mayor en los
extractos más polares (Figura 14) lo cual, como sugiere Zubia y colaboradores
(2007) podría deberse a la presencia de compuestos polifenólicos ya que éstos
han demostrado una correlación significativamente alta entre el contenido fenólico
y la actividad antioxidante en extractos de algas marinas, tal es el caso de lo
descrito por Wei et al., (2003) quienes demostraron la actividad antioxidante de
compuestos fenólicos puros obtenidos a partir de Sargassum kjellmanianum. Es

42
importante destacar que estudios previos realizados con algas del género
Sargassum analizan el contenido de polifenoles en relación a las variaciones
ambientales, obteniendo variabilidades muy altas en especies como Sargassum
furcatum y Sargassum mangarevense; mientras que en otras especies como
Sargassum hystrix las variaciones eran menores (González-Batista, 2009;
Budhiyanti, 2012).
Todas las fracciones analizadas presentaron una menor EC50 en relación a lo
obtenido en la curva control de ácido ascórbico EC50: 1.5 µg mL-1 (Figura 5). No
obstante, la fracción más activa de S. lapazeanum resultó 26 veces menor a éste
valor, con una EC50 de 36.96 µg mL-1. En este sentido, es importante considerar
que los extractos utilizados son extractos que no han sido purificados, por lo que
los compuestos responsables de la actividad observada pudieran incrementar su
efectividad una vez fraccionados. Por lo anterior, podría incluso considerarse el
ácido ascórbico, como uno de los compuestos relacionados a esta actividad, ya
que se ha reportado se encuentra presente en algunas macroalgas (Gonzáles-
Batista et al., 2009). Cabe destacar que en la composición química de las algas
existen diferentes metabolitos relacionados en mayor o menor medida a su
capacidad antioxidante, es por ello que realizar los ensayos de actividad se debe
considerar que esta puede atribuirse no sólo a una molécula, sino también al
efecto sinérgico entre varias (Gonzáles-Batista et al., 2009).
Generalmente los organismos fotoautróficos se encuentran expuestos a un gran
estrés por oxígeno, y se han adaptado a este medio desarrollado diferentes
sistemas de protección eficientes contra las especies reactivas de este (ERO).
Uno de estos sistemas, constituye en la producción de potentes antioxidantes
como lo son polifenoles, carbohidratos y compuestos nitrogenados, fitosteroles,
carotenoides, derivados de la clorofila y otros pigmentos fotoprotectores (Gouveia
et al., 2008). Además, las algas contienen metales como Se, Zn, Mn y Cu los
cuales son componentes fundamentales de enzimas antioxidantes, por lo que
podrían contribuir en esta propiedad al ser consumidos por el hombre (González-
Batista, 2009).

43
La actividad antioxidante observada, se puede atribuir a tres diferentes tipos de
compuestos que se han reportado como responsables de esta actividad en las
algas: Liposolubles, hidrosolubles y polisacáridos. Entre los liposolubles se
encuentran los carotenoides y la clorofila A. No obstante, no siempre es posible
ubicar los metabolitos de acuerdo a su polaridad, ya que Iwashima et al (2005)
aislaron 3 plastoquinonas a partir de Sargassum micracathum que presentaban
actividad antioxidante, la cual podía atribuirse a la porción polifenólica, el
segmento similar al alfa-tocoferol, o ambos, ya que no pudo ser determinado
(González-Batista, 2009).
Por su parte, entre los compuestos hidrosolubles destacan los polifenólicos como
los flavonoides y los ácidos fenólicos. Estos constituyen uno de los grupos de
metabolitos secundarios más numerosos y representativos en las plantas; son de
suma importancia ya que forman parte de diferentes eventos fisiológicos y
metabólicos. Actualmente se sabe que los compuestos fenólicos son los más
efectivos en la mayoría de las algas pardas estudiadas, conformando entre el 20 y
el 30% de su peso seco, siendo los florotaninos los polifenoles de mayor
presencia, los cuáles sólo se han aislado a partir de diferentes especies de algas
pardas, entre las que destacan Sargassum kjellamanianum, Sargassum
siliquastrum, y el bifuhalol polimerizado obtenido a partir de Sargassum
ringgoldianum (Nakai et al., 2006; González-Batista, 2009; Budhiyanti, 2012).
Finalmente, los polisacáridos sulfatados de algunas algas marinas también han
demostrado presentar un alto potencial antioxidante en diversos estudios que
incluyen el análisis hepatoprotector en modelos de ratón, sin embargo aún es
escaso el conocimiento que se tiene de estos compuestos y su relación con la
actividad antioxidante (González-Batista, 2009).

44
4. Análisis de la actividad anticoagulante.
4.1 Caracterización parcial del Fucoidan
El espectro de IR de las fracciones presentó un patrón de absorción similar del
infrarrojo, sugiriendo que todas las fracciones contienen los mismos grupos
funcionales, no obstante presentan variaciones en su composición, las cuales
pueden apreciarse por medio de las diferencias de intensidad de los picos de
absorción (Figura 16). Las bandas que se encuentran entre 960-970
corresponden a los residuos el grupo metilo de la fucosa (CH3), confirmando la
presencia de ésta. Por su parte las bandas entre el 1417-1616 cm-1 corresponden
al enlace amida N-H y demuestran la presencia de amino-azúcares o uronatos
(Gómez-Ordóñez, 2012), mientras que las bandas ubicadas entre 1210 y 1250
cm−1 corresponden a las vibraciones de estrechamiento del S-O. La banda a 830
cm−1 fue asignada a la vibración del grupo C-O-S. Estas vibraciones indican la
presencia del grupo sulfato en posición axial del C4 con sustituciones menores en
los carbonos C2 y C3 de la fucosa, dadas por un pico de absorción menos intenso
a los 805 cm-1 (Li et al., 2008).
Además, la banda de absorción a los 570 cm-1 confirmó la presencia de un
número significativo de grupos sulfato (Muñoz-Ochoa, 2006). No obstante, a parte
de las vibraciones C-O-S, en la región de 820-850 cm -1 existen vibraciones dadas
por las uniones C-H de la terminación de azúcares reductores, lo cual podría
afectar el criterio de la posición del grupo sulfato. En base a lo anterior Li et al.,
(2008) sugieren comparar y complementar los resultados obtenidos por IR con la
estabilidad de los esteres sulfatados por medio de análisis de álcali y de
metilación para determinar la posición del grupo sulfato. Es por ello que se
recomienda continuar con la investigación y realizar estudios posteriores, no sólo
con el fin de investigar la conformación de la estructura química del fucoidan
obtenido a partir de S. lapazeanum, sino también para esclarecer los mecanismos
de acción de este compuesto.

45
Fig
ura
16
. P
rin
cip
ale
s
pic
os
de
a
bso
rció
n
de
lo
s
esp
ectr
os
de
F
TIR
-AT
R
de
l e
xtr
acto
a
cuoso
(F
uco
ida
n:
FC
) de
S
.
lap
aze
anu
m y
la
s f
raccio
ne
s o
bte
nid
os p
or
pre
cip
ita
ció
n f
raccio
nad
a a
pa
rtir d
e é
ste
: F
C1,
FC
2 y
FC
3 e
n la
re
gió
n d
e lo
s 4
00
-
200
0cm
-1.

46
En base a lo anterior se describe el fucoidan obtenido a partir de S. lapazeanum
como un heterofucano debido a la presencia de ácidos urónicos y azúcares
neutros detectados en las banas de IR (Gómez-Ordóñez, 2012). Por lo que se
sugiere hacer un análisis de su composición monomérica para la identificación de
azúcares como manosa, xilosa o galactosa.
4.2 Ensayo de actividad anticoagulante
El proceso de coagulación se lleva a cabo a través de ciertos factores que
permiten detener el flujo sanguíneo en el lugar donde se ha producido daño tisular.
En la medida que los anticoagulantes (endógenos o exógenos), interfieren con
estos factores de coagulación inactivándolos o limitándolos, la coagulación de la
sangre se prolonga o se detiene (Gómez-Ordóñez, 2012). La actividad in vitro del
fucoidan y los precipitados obtenidos a partir de este, fue evaluada por medio del
tiempo de protrombina (TP) y el tiempo de tromboplastina parcial activada (TTPA).
El ensayo de TP sugiere que los heterofucanos obtenidos a partir de S.
lapazeanum actúan en la vía extrínseca de la cascada de coagulación ya que
todos los extractos probados prolongaron el tiempo de formación del coágulo,
siendo este hasta 28 s mayor que el control en FC2 (Tabla VIII).
En base a los resultados observados en los primeros ensayos de TTPA, se
realizaron curvas dosis-efecto con cada uno de los extractos para definir las
concentraciones a las que se presenta esta actividad. Las curvas obtenidas
demostraron el alto potencial anticoagulante de S. lapazeanum, ya que todos los
extractos prolongaron el tiempo de formación del coágulo, duplicando el tiempo de
coagulación establecido por el control (32.84 s) incluso a concentraciones
mínimas de 3.06 µg mL-1 (Figura 15), siendo FC2 el más activo. Las fracciones
de fucoidan obtenidas en el presente estudio mostraron una actividad similar a la
de la heparina. Ronghua et al., (2003) reportaron que el tiempo de coagulación
para la heparina fue de 125 s a concentraciones de 10 µgmL-1, en base a ello es
posible decir que FC2 la fracción más activa y de mayor potencial ya que duplicó
este tiempo a la misma concentración (Figura 15). Esto concuerda con el
potencial observado en otras especies del género, ya que Dore-Guerra et al.,

47
(2013) reportaron que el fucoidan obtenido a partir de Sargassum vulgare resultó
ser hasta 10 veces más activo que la heparina.
Tabla VIII. Comparación de actividad de los extractos
acuosos obtenidos a partir de Sargassum lapazeanum en
el ensayo de tiempo de protrombina (TP).
Extracto TP Tiempo prolongado (s) comparación al control
Control 14
FC 30.25 ± 0.79 16.25
FC1 24.50 ± 1.30 10.5
FC2 41.93 ± 2.49 27.93
FC3 23.079 ± 1.90 9.079
Por su parte, los ensayos de TTPA mostraron tiempos de coagulación mayores a
los 300 s establecidos para el ensayo, siendo este resultado casi 9 veces mayor
que lo mostrado por el control (33.54 s) (Tabla VII). Esto coincide con lo observado
por diferentes autores (Tabla IX), tales como Athukorala et al. (2007) quienes
registraron valores similares a partir de extractos acuosos obtenidos a partir de S.
horneri (>300 s) y S. siliquastrum (200 s). Asimismo, Lara-Issasi y Álvarez-
Hernández (1999), registraron una menor actividad a partir de algas como S.
cymosum, S. filipéndula y S. pteropleuron, así como tiempos igual de elevados
(>300 s) a partir de extractos de Sargassum vulgare. Estas diferencias podrían
deberse al tipo de especie estudiada y su composición química que, como lo
indica García-Ríos et al. (2012), se ve afectada por diversos factores como el
clima y la zona geográfica. Sus estudios señalan que existe una correlación entre
el contenido de fucoidan y la profundidad de las algas, obteniendo un mayor
contenido de este compuesto en aquellas que crecen cerca de la superficie. Lo
anterior podría explicar el alto potencial de S. lapazeanum, un alga que habita en
la zona intermareal.

48
Tabla IX. Listado de algunos valores de TP y TPPA reportados para algas del género Sargassum.
Especie Referencia TP TTPA (s)
Sargassum horneri Athukorala et al., 2007. > 300
Sargassum siliquastrum Athukorala et al., 2007. 200
Sargasum cymosum Lara-Isassi y Álvarez-Hernández, 1999. 90 90
Sargassum filipendula Lara-Isassi y Álvarez-Hernández, 1999. 16 82
Sargassum
pteropleuron
Lara-Isassi y Álvarez-Hernández, 1999. 60 100
Sargassum vulgare Lara-Isassi y Álvarez-Hernández, 1999. 25 > 300
La actividad observada en los ensayos de TTPA sugiere que los heterofucanos
obtenidos también podrían estar actuando en la vía intrínseca o la vía común del
FX, factor que es esencial para la formación de la trombina y la subsecuente
fibrina. Se ha reportado que los fucoidanos obtenidos a partir de algas pardas
pueden, al igual que la heparina, inhibir la actividad de la trombina actuando de
manera directa en la enzima o mediante la activación de los inhibidores de
trombina, incluyendo la antitrombina III y el cofactor II de la heparina. No obstante
algunos activan sólo la antitrombina. La heparina es una biomolécula que
contiene glucosaminoglicano altamente sulfatado que junto con las heparinas de
bajo peso molecular, representan los anticoagulantes de mayor uso en el
mercado (Gómez-Ordóñez, 2012). Sin embargo, se ha registrado que el uso
prolongado de estos compuestos puede tener efectos adversos como lo son
trombocitopenia, osteoporosis y hemorragias.
Con respecto al grado de sulfatación, el contenido de sulfatos en FC2 (47.48%)
fue mucho mayor en comparación a lo observado en las otras fracciones, siendo
igualmente la fracción que presentó más actividad (Tabla VII y Figura 15). Esto
coincide con estudios previos, los cuales indican el incremento en el contenido de
sulfatos como uno de los factores más importantes que definen la actividad
anticoagulante, la cual incrementa conforme el grado de sulfatación de los fucanos
(Muñoz-Ochoa, 2006). Además se ha reportado que pequeñas diferencias en la
proporción y distribución de los residuos de sulfatos en las cadenas del fucoidan,

49
podrían resultar críticas para la interacción de las proteasas, inhibidores y
activadores del sistema de coagulación, alterando así el valor observado en los
ensayos de actividad anticoagulante (Gurgel-Rodrigues et al., 2011)
Debido al alto potencial anticoagulante del fucoidan obtenido a partir de S.
lapazeanum, se sugiere continuar con futuras investigaciones enfocadas en el
aislamiento y caracterización de este compuesto que permitan complementar el
estudio con métodos como resonancia magnética nuclear (RMN).

50
VIII. CONCLUSIONES
Los ensayos elaborados en el presente estudio demostraron que Sargassum
lapazeanum es un alga con un alto potencial como fuente de compuestos con
actividad antioxidante, antibacteriana y anticoagulante.
El análisis de la actividad antibacteriana indicó la selectividad de S. Lapazeanum
por bacterias del género Vibrio, como V. harveyi y V. parahaemolyticus y V.
parahaemoliticus
Los ensayos de actividad secuestrante de radicales libres demostraron que S.
lapazeanum es un alga de gran potencial antioxidante con una EC50 = 39.96 µg
mL-1 comparable a lo obtenido en otras especies de algas de este género.
Los heterofucanos FC1, FC2 y FC3 obtenidos a partir de la precipitación
fraccionada de FC, mostraron una actividad anticoagulante comparable a la de la
heparina, siendo FC2 el más activo. Finalmente el análisis estructural del fucoidan
demostró que se trata de un heterofucano compuesto por fucosa con sustituciones
de grupos sulfato en posición C4 principalmente.

51
X. BIBLIOGRAFÍA
Amsler, C. D. (2008). Algal Sensory Chemical Ecology 297 - 305. En: Amsler, C.D.
Andrade-sorcia, G., Riosmena-Rodríguez, R., & Paúl-Chávez, L. (2008).
Variabilidad morfológica y morfométrica de Sargassum lapazeanum
(Sargassaceae: Phaeophyta) en el Suroeste del Golfo de California. INSULA,
(37), 67–80.
Anggadiredja J., & Andyani R. (1997). Antioxidant activity of Sargassum
polycystum (Phaeophyta ) and Laurencia obtusa ( Rhodophyta ) from Seribu
Islands. Journal of Applied Phicology. 9(5): 477-479.
Al-Saif S.S., Abdel-Raouf N., El-Wazanani H. & I. A. Aref. (2014). Antibacterial
substances from marine algae isolated from Jeddah coast of Red sea. Saudi
Journal of Biological Sciences. 21(1): 57–64.
Andrade-sorcia, G., Riosmena-Rodríguez, R., Lee K.M, Boo G.H., Muniz-Salazar
R., Lopez Vivas J. M., G. González-Barba & S.M. Boo. (2013). Systematic
reassessment of Sargassum (Fucales, Phaephyceae) species from the Gulf of
California, México. Phycologia. 52(4): 4-4.
Arunkumar K., Selvapalam N. & R. Rengasamy. (2005). The antibacterial
compound sulphoglycerolipid 1-0 palmitoyl-3-0(6´sulpho-α-quinovopyranosyl)-
glycerol from Sargassum wightii Greville (Phaephyceae). Botánica Marina.
48(5): 441-445.
Athukorala Y., Lee K-W., Kim S-K. & Y-J Jeon. (2007). Anticoagulant activity of
marine green and brown algae collected from Jeju Island in Korea.
Bioresource Technology. 98(9): 1711-1716.
Attaway H.D & O.R Zaborsky. 1993. Marine Biotechnology: Pharmaceutical and
bioactive natural products. Springer, New York. 524 p.

52
Budhiyanti, S. A., Raharjo, S., Marseno, D. W., & Lelana, I. Y. B. (2012).
Antioxidant activity of brown algae Sargassum species extract from the
coastline of java island. American Journal of Agricultural and Biological
Sciences. 7(3), 337–346.
Carrillo-Domínguez S., Casas-Valdez M., Ramos-Ramos F., Pérez-Gil F. & I.
Sánchez-Rodríguez. (2002). Algas marinas de Baja California Sur, México:
Valor nutrimental. ALAN. 52(4): 400-405.
Casas Valdez M., C. Hernández, R. Aguila, B. González, A. Marín, S. Rodríguez,
S. Carrillo, F. Pérez Gil, E. Cruz, M. Rique & M. Tapia. (2002). Sargassum
spp. como fuente potencial de alimento para camarón. Informe Técnico Final.
CGPI. Instituto Politécnico Nacional, 34 pp.
Casas-Valdez, M., Portillo-Clark G., Aguila-Ramírez N., Rodríguez-Astudillo S.,
Sánchez-Rodríguez I. & S. Carrillo-Domínguez. (2006). Efecto del alga marina
Sargassum spp. sobre las variables productivas y la concentración de
colesterol en el camarón café, Farfantepenaeus californiensis (Holmes,
1900). Revista de biología marina y oceanografía.41(1), 97-105.
Castillo-Guzmán A., Arocha-Meriño C., Armas-Rojas N.B., Castillo-Arocha I.,
Cueto-Cañabate M.E & M. L. Herrera- Giró. 2008. Calidad de vida relacionada
con la salud en personas con enfermedades crónicas degenerativas. Rev
Cubana Investigaciones Biomédicas. 27:3-4.
Calva B., G.L., & R. Torres-Alvarado. (2008). Macroalgas y pasto marino, útiles
bioindicadores de contaminación por hidrocarburos fósiles en sistemas
acúaticos. Contactos. 68, 38-46.
Castro-Reyes, M.C. (1997). Actividad antibacteriana de Sargassum sinicola
(Sargassaceae, Phaeophyta) & Laurencia johnstonii (Rhodomelaceae,
Rodophyta) de Bahía de La Paz, B.C.S., México. Tesis Maestría. CICIMAR-
IPN, B.C.S., México. 64p.
Choi J.S., Lee H.J. & S.S. Kang, Arch. (1994). Pharmacal. Res. 17, 462.

53
Cumashi A., Ushakova N. A, Preobrazhenskaya M. E., D’Incecco A., Piccoli A.,
Totani L., Tinari, N., Morozevich G. E, Berman A. E., Bilan M. I., Usov A. I.,
Ustyuzhanina N. E., Grachev A. A, Sanderson C. J., Kelly M., Rabinovich G.
A., Iacobelli S. & N.E. Nifantiev. (2007). A comparative study of the anti-
inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine
different fucoidans from brown seaweeds. Glycobiology, 17(5), 541–52.
Dashtiannasab A., Kakoolaki S., Sharif Rohani M. & V. Yeganeh. (2011). In vitro
effects of Sargassum latifolium (Agardeh, 1948) against selected bacterial
pathogens of shrimp. Iranian Journal of Fisheries Sciences. 11(4):765-775.
Demirel, Z., F. Y. Ferda, N. K. Ulku, G. Ozdemir & A. Sukatar. (2009). Antimicrobial
and antioxidant activity of brown algae from the Aegean. J. Serb. Chem. Soc.
74 (6) 619–628
De Zoysa., Nikapitiya C., Jeon Y.J, Jee Y. & J. Lee. (2008). Anticoagulant activity
of sulfated polysaccharide isolated from fermented brown
seaweed Sargassum fulvellum. Journal of Applied Phycology. 20(1): 67-74 p.
Díaz-Concepción, A. & D. Almagro-Vázquez. (2001). Estado del mecanismo de la
coagulación sanguínea. Revista Cubana de Hematología, Inmunología y
Hemoterapia, 17: 77-89.
Dorado-Martínez C., Rugerio-Vargas C. & S. Rivas-Arancibia. (2003). Estrés
oxidativo & neurodegeneración. Rev. Facultad de Medicina UNAM. 46 (6):
229-235.
Dore-Guerra C. M., Faustino-Alves M. G. D., Pofírio Will E. L. S., Costa G T.,
Sabry D. A., De Souza-Rêgo L. A. R., Accardo M. C., Rocha O. H. A.,
Guimarães Filgueira G. A. & E.. Lisboa Leite. (2013). A sulfated
polysaccharide, fucans, isolated from brown algae Sargassum vulgare with
anticoagulant, antithrombotic, antioxidant and anti-inflammatory effects.
Carbohydrate Polymers. 91(1): 467-475.

54
Duarte E. R. M., Cardoso M. A., Noseda M. D. y A. S. Cerezo. (2001). Structural
studies on fucoidans from the Brown seaweed Sargassum stenophyllum.
Carbohydrate Research. 333(4): 281-293 p.
Echeverría Z. B., Franco S. A. & M. A. Martínez. (2009). Evaluación de la actividad
antioxidante y determinación del contenido de compuestos fenólicos en
extractos de macroalgas del Caribe Colombiano. Redalyc, 16(1), 126–131.
Flora, S. J. S. (2009). Structural, chemical and biological aspects of antioxidants
for strategies against metal and metalloid exposure. Oxidative medicine and
cellular longevity, 2(4), 191–206.
Freile-Pelegrín & J. L. Morales. (2004). Antibacterial activity in marine algae from
the coast of Yucatán, México. Botánica Marina. 47: 140-146.
Frikha, F, Kammoun, M, Hammami, N, Mchirgui, RA, Belbahri, L, Gargouri, Y,
Miled, N, & Ben-Rebah, F. (2011). Composición química y algunas actividades
biológicas de algas marinas recolectadas en Túnez. Ciencias marinas, 37(2),
113-124.
García-Ríos, V., Ríos-Leal, E., Robledo, D., & Y. Freile-Pelegrin, (2012).
Polysaccharides composition from tropical brown seaweeds. Phycological
Research, 60(4), 305–315.
Goering R., Dockrell H., Zuckerman M., Riott I. & P. L. (2012). Chiodini. Mim’s
Medical Microbiology. Elsevier Health Sciences. 624 pp.
Gómez-Ordóñez E., (2012). Evaluación nutricional y propiedades biológicas de
algas marinas comestibles. Estudios in vitro e in vivo. Tesis Doctoral.
Universidad Computense de Madrid CSIC-ICTAN, Madrid, España. 218 pp.
González-Batista, A. E., Charles B., M., Mancini-filho, J., & Vidal Novoa, A. (2009).
Las algas marinas como fuentes de fitofármacos antioxidantes. Revista
Cubana de Plantas Medicinales. 14(2): 1–18.

55
Gouveia L., Batista A.P., Sousa I., Raymundo A. & N.M Bandarra. (2008).
Microalgae in Novel Food Products. Food Chemestry Research
Developments. 37 pp.
Granowitz, E. V, & Brown, R. B. (2008). Antibiotic adverse reactions and drug
interactions. Critical care clinics, 24(2): 421–442.
Gupta S. & N. Abu-Ghannam. (2011). Bioactive potential and possible health
effects of edible brown seaweeds. Trends in Food Science & Technology.
1(1): 12.
Gurgel Rodrigues, A. J., Oliveira Vanderlei, E. D. S., Bessa Façanha, É.,
Magalhães Araújo, F. De, Monteiro De Paula, C. R., Lima, V., & Barros
Benevides, M. N. (2011). Anticoagulant Activity of a Sulfated Polysaccharide
Isolated from the Green Seaweed Caulerpa cupressoides. Brazilian Archives
of Biology and Technology., 54(4), 691–700.
Haas S. (2008). New oral Xa and IIa inhibitors: updates on clinical trial
results. Journal of Thromb Thrombolysis. 25(1):52-60.
Holdt, S. L. & S. Kraan. (2011). Bioactive compounds in seaweed: Functional food
applications and legislation. Journal of Applied Phycology. 23(3), 543-597.
INEGI. 20011. Perspectiva estadística de México: Diciembre 2011. INEGI. 99 pp.
Jiao, G., Yu, G., Zhang, J., & Stephen, H. E. (2011). Chemical structures and
bioactivities of sulfated polysaccharides from marine algae. Marine Drugs,
9(2): 196–223.
Jorgensen, J. H. & Turnidge J. D. (2007). Susceptibility test methods: dilution and
disk diffusion methods, p. 1152–1172. In P. R. Murray, E. J. Baron, J. H.
Jorgensen, M. L. Landry, & M. A. Pfaller (ed.), Manual of clinical microbiology,
9th ed. ASM Press, Washington, D.C.

56
Khan A. R. & R. Qari. 2012. Antibacterial activities of brown seaweed Sargassum
boveanum (J. AG.) against diarrhea along the coast of Karachi, Pakistan.
Journal of Enviromental Research And Development. 6(3A): 753-757.
Kim Y.H., Kim E.H., C. Lee, M.H. Kim & J.R. Rho. (2007). Two new
monogalactosyl diacylglycerols from brown alga Sargassum thunbergii. Lipids,
42: 395–399.
Kitchen, S., McCraw, A., & Echenagucia, M. (2010). Diagnóstico de la hemofilia y
otros trastornos de la coagulación. Federación Mundial de Hemofilia. 144 pp.
Kumar P., Senthamilselvi S. & M. Govindaraju. 2013. GC-MS profiling and
antibacterial activity of Sargassum tenerrimum. Journal of pharmacy research.
6: 88-92.
Lara-Isassi, G. & Álvarez-Hernández, S. (1994). Actividad biológica de las
macroalgas marinas de México. Revista de la Sociedad Mexicana de Historia
Natural. 45: 51-60.
Lara-Issasi G., Álvarez-Hernández S. & C. Lozano-Ramírez. (1996). Actividad
antibacteriana de algas marinas de Oaxaca, Pacífico Tropical Mexicano. Biol.
Trop. 44(2): 885-898.
Lara-Issasi G., Álvarez-Hernández S., Lozano-Ramírez C. & N. Hernádez-Soto.
(1999). Nuevas adiciones al conocimiento de la actividad antibiótica de
macroalgas marinas mexicanas. Hidrobiología. 9(2): 159-168.
Lara-Isassi, G. & Álvarez-Hernández, S. (1999). Evaluación de la actividad
anticoagulante de algas marinas presentes en las costas del Golfo de México
y Caribe Mexicano. Revista de la Sociedad Mexicana de Historia Natural. 49:
75-82.
Li B., Lu, F., Wei, X., & R. Zhao (2008). Fucoidan: Structure and bioactivity.
Molecules, 13(8), 1671-1695.

57
Li H., Cheng K., Wong C., Fan K., Chen F. & Y. Jiang. (2007). Evaluation of
antioxidant capacity and total phenolic content of different fractions of selected
microalgae. Food Chemistry, 102(3), 771-776.
Lijour, Y., E. Gentric, E. Deslandes & J. Guezennec. 1994. Estimation of the
sulfate content of hydrothermal vent bacterial polysaccharides by Fourier
transform infrared spectroscopy. Analytical Biochemistry, 220: 244-248
Limonta, M. R. & R. S. Coffigny. 2012. Enfermedades infecciosas en Panulirus.
Situación actual camarones Penaeus y langosta. REDVET, 13(7):1-17.
Magallanes C., Córdova C. & R. Orozco. 2003. Actividad antibacteriana de
extractos etanólicos de macroalgas marinas de la costa central del Perú. Rev.
Perú biol. 10 (2): 125-132.
Marín Álvarez A. 1999. Utilización del alga Sargassum spp. como complemento
alimenticio de mantenimiento en dietas de ganado ovino. Tesis de Maestría
en Manejo de Recursos Marinos. Centro Interdisciplinario de Ciencias
Marinas-IPN. La Paz, B.C.S., México, 86 pp.
Martínez-Murillo, C. (2003). Bases de la hemostasia y trombosis. Gaceta Médica
de México. 139(S2): 28-30
Mascheck, J. A. & B. J. Baker. (2008). The Chemistry of Secondary Metabolism.
En: Amsler, C.D. (Ed). Algal Chemical Ecology. Springer-Verlag, 313pp.
Mateo-Cid, L. E., Mendoza-Gonzáles, A. C., Ávila-Ortiz, A. G., & Díaz-Martínez, S.
2013. Algas marinas bentónicas del litoral de campeche, méxico l. Acta
Botánica Mexicana, 104, 53–92.
Matsubara, K. (2004). Recent advances in marine algal anticoagulants. Curr. Med.
Chem. Cardiovasc. Hematol. Agents. 2: 13-19.
McMichael M. (2012). New models of hemostasis. Top Companion Anim Med.
27(2): 40-45.

58
Mendiola-León, J. A. (2008). Extracción de compuestos bioactivos de microalgas
mediante fluidos supercríticos. Universidad Autónoma de Madrid. 151p.
Michael Guiry in Guiry, M.D. & Guiry, G.M. (2014). AlgaeBase. World-wide
electronic publication, National University of Ireland, Galway.
http://www.algaebase.org; searched on 20 February 2014.
Molyneux, P. (2004). The use of the stable free radical diphenylpicryl- hydrazyl
(DPPH) for estimating antioxidant activity. Songklanakarin J. Sci. Technol.
26(2): 211-219.
Muñoz-Ochoa (2006).Extracción y caracterización de polisacáridos con actividad
anticoagulante a partir de algas colectadas en Baja California Sur. Tesis
Maestría. CICIMAR-IPN, B.C.S., México. 64 p.
Muñoz-Ochoa, M., Murillo-Álvarez, J. I., Zermeño-Cervantes, L. a, Martínez-Diaz,
S., & Rodríguez-Riosmena, R. (2010). Screening of extracts of algae from
Baja California sur, Mexico as reversers of the antibiotic resistance of some
pathogenic bacteria. European Review for Medical and Pharmacological
Sciences, 14(9), 739–47.
Murillo-Álvarez, J. I., D. R. Encarnación & S. G. Franzblau. (2001). Antimicrobial
and cytotoxic activity of some medicinal plants from Baja California Sur
(México). Pharmaceutical Biology, (39): 445 – 449.
Nakai M, Kageyama N, Nakahara K & Miki W. (2006). Phlorotannins as Radical
Scavengers from the extract of Sargassum ringgoldianum. Marine
Biotechnology. 8:409-14.
Norris, J.N. (2010). Marine algae of the Northern Gulf of California: Chlorophyta
and Phaeophyceae.Smithsonian Contributions to Botany 94(9): 1-276.
Nuestro-Baleta F., Laureta V.L., Jane M., Apines-Amar S., Ian P., Padilla P. & G.
F. Quinitio. (2011). Biological Activity of Extracts of Sargassum oligocystum

59
(Magnaye) against Aquaculture Pathogenic Bacteria. The Israeli Journal of
Aquaculture. Bamidgeh. 2(63): 667-5.
Organización Mundial de las Salud [OMS a]. (2013). Enfermedades no
transmisibles. 18/03/2014. En línea:
http://www.who.int/mediacentre/factsheets/fs355/es/
Organización Mundial de las Salud [OMS b]. (2013). Enfermedades
cardiovasculares. 18/03/2014. En línea:
http://www.who.int/mediacentre/factsheets/fs317/es/
Pardo-Fuentes P.E. (2012). Estudios del alga Sargassum horridum como fuente
de compuestos activos contra Mycobacterium tuberculosis. Tesis Maestría.
CICIMAR-IPN, B.C.S., México. 73 p.
Patrón-Prado M., Casas-Valdez M., Serviere-Zaragoza E., Zenteno-Savín T.,
Lluch-Cota D. B. & L. Méndes-Rodríguez. (2011). Biosorption capacity for
cadmium of brown seaweed Sargassum sinicola and S. lapazeanum in the
Gulf of California. Water Air Soil Pollution. 221:137 144.
Pedroche, P.F., Silva, P.C., Aguilar Rosas, L.E., Dreckmann, K.M. & Aguilar
Rosas, R. (2008). Catálogo de las algas benthónicas del Pacífico de México II.
Phaeophycota. pp. [i-viii], i-vi, 15-146. Mexicali & Berkeley: Universidad
Autónoma Metropolitana and University of California Berkeley.
Phillips, N. (1995). Biogeography of Sargassum (Phaeophyta) in the Pacific basin.
In: Taxonomy of Economic Seaweeds. (Abbott, I.A. Eds) Vol.5, pp. 107-145.
La Jolla, California: California Sea Grant College System.
Querellou J, Børresen T, Boyen C, Dobson A, Höfle M, Ianora A, Jaspars M, Kijjoa
A, Olafsen J, Rigos & G, Wijffels R. (2010). Marine Biotechnology: A New
Vision and Strategy for Europe: Marine Board-ESF Postition Paper 15.
Beernem, Belgium: European Science Foundation. 91 p. (Marine Board-ESF
Position Paper; No. 15).

60
Quintana-González, S. (2002). Nuevos conceptos en la fisiología de la
coagulación. Gaceta Médica de México, 138 (S1): 47-51.
Rindi F., Soler-Vila A. & D. Guiry. (2012). Taxonomy of Marine Macroalgae Used
as Sources of Bioactive Compounds. En: Maria Hayes (Ed.) Marine Bioactive
Compounds: Sources, Characterization and Applications. 229p.
Ríos N., Medina G., Jiménez J., Yáñez C. García Y. M., Di Bernardo M. L. & M.
Gualtieri. (2009). Actividad antibacteriana y antifúngica de extractos de algas
marinas venezolanas. Revista Peruana de Biología. 16(1): 97-100.
Rivera, M. & Scrosati, R. (2006). Population dynamics of Sargassum
lapazeanum (Fucales, Phaeophyta) from the Gulf of California,
Mexico. Phycologia 45: 178-189.
Rivera, M. & Scrosati, R. (2008). Self-thinning and size inequality dynamics in a
clonal seaweed (Sargassum lapazeanum, Phaeophyceae) (Note). Journal of
Phycology 44(1): 45-49.
Ronghua, H, D. Yumin & Y. Jianhong. (2003). Preparation and in vitro
anticoagulant activities of alginate sulfate and its quaterized derivatives.
Carbohydrates polymers, 52: 19-24.
Saad-Navarro, G. & Riosmena-rodríguez, R. (2005). Variación espacial y temporal
de la riqueza florística de macroalgas en la zona rocosa de Bahía de Muertos,
B.C.S. México. Ciencia y Mar, 9(26), 19–32.
Sastry V.M.V.S. & G.R.K. Rao (1995). Dioctyl phthalate, and antibacterial
compound from the marine brown alga Sargassum wightii. Journal of Applied
Phycology. 7(2): 185-186.
Senthilkumar, K., Manivasagan, P., Venkatesan, J., & S.K. Kim. (2013). Brown
seaweed fucoidan: biological activity and apoptosis, growth signaling
mechanism in cancer. International Journal of Biological Macromolecules,
60:366–74.

61
Seo Y., Park K.E., Kim Y.A., Lee H.I., Yoo I.S., Ahn I.W. & Lee B.J. (2006).
Isolation of tetraprenyltoluquinols from the brown alga Sargassum
thunbergii. Chem. Pharm. Bull. 54: 1730-1733.
Seo Y., Park K.E. & T.J Nam Bull (2007). Isolation of a new chromene from the
brown alga Sargassum thunbergii. Korean Chem. Soc. 28: 1831-1833.
Setchell, W.A. & Gardner, N.L. (1924). New marine algae from the Gulf of
California.Proceeding of the California Academy of Science, Series 4 12: 695-
949, 77 plates.
Shanab Sanaa MM. (2007). Antioxidant and antibiotic Activities of some Seaweed
(Egyptian Isolates). International J Agric & Biol. 2: 220-5.
Shanmugam, M. & K. H. Mody. (2000). Heparinoid-active sulphated
polysaccharides from marine algae as potential blood anticoagulant agents.
Current Science, 79: 1672-1683.
Song Qin, W. E., Müller G. & E. L. Cooper. (2011). “Marine Biotechnology,”
Evidence-Based Complementary and Alternative Medicine. 2pp.
Spyropoulos A.C. (2007). Investigational treatments of venous thromboembolism.
Expert Opin Investig Drugs. 16(4): 431-440.
Stewart W. D. P. (1974). Algal Physiology and Biochemistry. University of
California Press. 989 p.
Suárez-Castillo A. N., R. Riosmena-Rodríguez, M. Rojo-Amaya, J. Torre-Cosio, R
Rioja Nieto, A. Hudson-Weaver, T. Pfister, G Hernández-Carmona, G. Hiojosa
Arango, O. Aburto Oropeza & A. L. Figueroa Cárdenas. (2013). Bosques de
algas pardas en el golfo de California: Sargassum, un hábitat esencial.
CONABIO. Biodiversitas, 108: 12-16.
Stella R. L. & D. C. Marin. (2009). Metodologías para evaluar in vitro la actividad
antibacteriana de compuestos de origen vegetal. Scientia et Technica. XV,
42: 263-268.

62
Sussman C., B. M. & Bates-Jensen. (2007). Wound Care: A collaborative Practice
Manual for Health Professionals. Lippincott Williams & Wilkins 720 pp.
Tajbakhsh S., Pouyan M., Zandi K., Bahramian P., Sarvati K., Fouladvand M.,
Asayesh G. & A. Barazesh. (2011). In vitro study of antibacterial activity of the
alga Sargassum oligocystum from the Persian Gulf. Eur Rev. Med. Pharmacol.
Sci. 15(3): 293-8.
Tüney Ü. Çadirci D. & A. Sukatar. (2006). Antimicrobial Activities of the Extracts of
Marine Algae from the Coast of Urla (Üzmir, Turkey). Turk Journal of Biology.
30: 171-175.
Vadlapudi, V. (2012). Antioxidant activities of marine algae : A review. 661 (2).
Medicinal Plants as Antioxidant Agents: Understanding Their Mechanism of
Action and Therapeutic. Research signpost. Anna capasso.
Venkateswara, R. 2008. Vibriosis in Shrimp Aquaculture. Neospark Drugs and
Chemicals. Private limited Corporate.
Vijayabaskar, P., & V. Shiyamala, 2011. Antibacterial Activities of Brown Marine
Algae (Sargassum wightii and Turbinaria ornata ) from the Gulf of Mannar
Biosphere Reserve. Advances in Biological Research, 5(2), 99–102.
Wei Y, Li Z, Hu Y & Xu Z.(2003).Inhibition of mouse liver lipid peroxidation by high
molecular weight phlorotannins from Sargassum kjellmanianum. Journal of
Applied Phycology.15:507–511
Zertuche-González, J.A. (1993). Situación actual de la industria de macroalgas
productoras de ficocoloides en América Latina y el Caribe. FAO. 66 pp.
Zhang C. Y., Wu W. H., Wang J. & M. B. Lan. (2012). Antioxidant Properties of
Polysaccharide from the Brown Seaweed Sargassum graminifolium and Its
Effects on Calcium Oxalate Crystallization. Marine Drugs. 10(1): 119-30.

63
Zubia, M., Robledo, D., & Freile-Pelegrin, Y. (2007). Antioxidant activities in
tropical marine macroalgae from the Yucatan Peninsula, Mexico. Journal of
Applied Phycology, 19(5): 449–458.

64
ANEXO I.
Descripción de la especie: Sargassum lapazeanum.
Las especies de Sargassum se caracterizan morfológicamente por presentar un
talo (estructura de forma arbustiva), que se divide en sujetador, estipe, filoides y
aerocistos. No obstante, las estructuras en éste género suelen verse modificadas
debido a variaciones espacio-temporales de distintos factores ambientales, y es
precisamente ésta plasticidad morfológica lo que por mucho tiempo complicó la
clasificación taxonómica de Sargassum lapazeanum (Andrade-Sorcia, 2005).
Tabla X. Ubicación taxonómica del alga
Sargassum lapazeanum.
Taxa Descripción
Imperio: Eukaryota Chatton.
Reino: Chromista T. Cavalier-Smith.
Phylum: Ochrophyta T. Cavalier-Smith.
Clase: Phaeophyceae F.R. Kjellman.
Orden: Fucales Bory de Saint-
Vincent.
Familia: Sargassaceae Kützing.
Género: Sargassum C. Agardh.
Morfología
Ls talos de S. lapazeanum llegan a medir entre ~10 a ~1.5 m de largo, estos
surgen de un sujetador discoidal parenquimatoso y sólido que inicia como un disco
hasta formar un cono o hapteros. El estipe es cilíndrico y liso, presenta además
numerosas ramas lisas y cilíndricas. Las ramas primarias son delgadas y pueden
llegar a presentar ramas secundarias. Los filoides son asimétricos, más amplios
en la zona apical del filoide, con pedicelos cortos y miden entre 0.5 – 2.5 cm de
largo. Algunos filoides presentan una corona sobre uno de los lados del filoide (no

65
son completamente planos), otros se encuentran torcidos de la mitad basal del
margen superior, que es liso y cóncavo. El remanente del filoide es agudo,
dentado y con una nervadura de poco evidente a evidente. Los criptostomatas
están presentes y evidentes, mientras que los aerocistos son escasos entre los
receptáculos que son elípticos. Los aerocistos en su mayoría son similares a la
forma de los filoides, tienen el margen modificado con remanencias de filoides, lo
que forma una cresta. Los receptáculos se observan en trinchete con algunas
espinas, entremezclados receptáculos de anteridios y oogonias, creciendo en
ramilletes (Andrade-Sorcia et al., 2013)
Figura 17. Fotografías de Sargassum lapazeanum. © Valeria Villegas
Hábitat
Los organismos de esta especie crecen sobre rocas a lo largo de la zona
intermareal a submareal. Además se ha observado que pueden formar mantos
con S. horridum, S. sinicola, y S. johnstonii.

66
Ciclo de vida
No se cuenta con un registro del ciclo de vida de esta especie en particular, no
obstante se trata de un alga del género Sargassum, por lo que su reproducción es
sexual y asexual. La reproducción sexual se da por medio de la producción de
gametos, su fertilización y la fijación de zigotos al sustrato; mientras que la asexual
consiste en la regeneración de un nuevo talo a partir de un estolón (talo de años
anteriores, menor a 17 cm y con pocos o sin filoides y aerocistos).
Estudios de la dinámica poblacional de S. lapazeanum (Rivera y Scrosati, 2008)
indican que presenta sus máximos de crecimiento en primavera (Marzo-Abril tasa
de crecimiento máxima) y los mínimos durante el periodo de otoño, con escasez
de plantas reproductivas durante Junio-Agosto y la mayor mortalidad en los meses
de Agosto y Septiembre.
Distribución
S. lapazeanum fue descrita en 1924 por Setchell y Gardner, con ejemplares de La
Paz B.C.S., ya que sólo se tenía registro del alga en esta zona la especie se
consideró como endémica de la parte Sur del Golfo por mucho tiempo. Estudios
moleculares recientes (Andrade-Sorcia et al., 2013) demuestran que su
distribución es aún más amplia, pudiéndose observar en las costas de Sonora,
Baja California y Baja California Sur (Figura 18).

67
Figura 18. Mapa que muestra la distribución actual de S. lapazeanum. Modificado a partir
de NOAA. http://www.ngdc.noaa.gov/mgg/shorelines/shorelines.html.
68
ANEXO II.
Tabla XI. Extracto crudo (g) y rendimiento (%) obtenido del alga S.lapazeanum.
Especie
Peso del alga (g)
Extracto crudo (g)
Rendimiento (%)
S. lapazeanum 300 34.73 11.57
Tabla XII. Peso (g) y rendimiento (%) de las fracciones obtenidas por fraccionamiento
sólido líquido a partir de S. lapazeanum.
Especie Fracciones obtenidas por extracción sólido-líquido
12-009 F1 F2 F3 F4 F5 F6 F7
Sargassum lapazeanum
CH2Cl
2
100%
CH2Cl2-EtOH 90-10%
CH2Cl2-EtOH 75-25%
CH2Cl2-EtOH 50-50%
EtOH 100%
EtOH- H2O 50-50%
H2O 100%
Peso (g) 1.52 0.79 1.54 2.04 0.84 0.46 0.078 Rendimiento(%) 10.13 5.26 10.26 13.6 5.6 3.06 0.52

69
ANEXO III.
Figura 19. TLC de los eluatos obtenidos por medio del fraccionamiento sólido-lìquido del
extracto crudo E.E.C.
Figura 20. Cromatografìas en placa fina (TLC) de los eluatos obtenidos a partir de la
columna cromatográfica realizada con F2.

70
ANEXO IV.
Figura 21. Halos de inhibición de los extractos obtenidos a partir de S. lapazeanum en
cajas de petri inoculadas con Vibrio harveyi.
Figura 21. Halos de inhibición de los extractos obtenidos a partir de S. lapazeanum en
cajas de petri inoculadas con Vibrio parahaemolyticus.

71
Figura 23. Halos de inhibición de los extractos obtenidos a partir de F2 en cajas de petri
inoculadas con Vibrio harveyi.
Figura 24. Halos de inhibición de los extractos obtenidos a partir de F2 en cajas de petri
inoculadas con Vibrio parahaemolyticus.







