
INSTITUTO POLITECNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
RESPUESTA FISIOLÓGICA DE Pteria sterna
(GOULD, 1851) EN RELACIÓN A LA
CONCENTRACIÓN DE ALIMENTO Y
TEMPERATURA
TESIS
QUE PARA OBTENER EL GRADO DE
MAESTRÍA EN CIENCIAS EN MANEJO DE RECURSOS MARINOS
PRESENTA
ANA KAREN MEZA BUENDIA
LA PAZ, B. C. S., DICIEMBRE DE 2016



CONTENIDO
GLOSARIO ................................................................................................................ VI
LISTA DE TABLAS ................................................................................................. VIII
LISTA DE FIGURAS .................................................................................................. IX
RESUMEN .................................................................................................................. X
ABSTRACT ................................................................................................................ XI
1. INTRODUCCIÓN .................................................................................................... 1
2. ANTECEDENTES ................................................................................................... 4
2.1. Generalidades de P. sterna .............................................................................. 4
2.2. Hábitat y distribución ........................................................................................ 4
2.3. Ecología y biología ........................................................................................... 5
2.4. Balance energético y potencial de crecimiento ................................................ 7
2.5. Influencia de la temperatura en el metabolismo y balance energético de los
moluscos bivalvos ................................................................................................... 8
2.6. Influencia de la disponibilidad de alimento en el metabolismo y balance
energético de los moluscos bivalvos ..................................................................... 11
3. JUSTIFICACIÓN ................................................................................................... 13
4. HIPOTESIS ........................................................................................................... 13
5. OBJETIVOS .......................................................................................................... 14
5.1. General .......................................................................................................... 14
5.2. Específicos, bajo condiciones de laboratorio para P. sterna .......................... 14
6. METODOLOGÍA ................................................................................................... 14
6.1. Obtención de organismos .............................................................................. 14
6.2. Traslado y mantenimiento de los organismos ................................................ 16
6.3. Pre-aclimatación de los organismos .............................................................. 16
6.4. Bioensayo 1: Determinación de la concentración óptima de alimento en
relación a la temperatura del agua ........................................................................ 17
6.4.1. Aclimatación de los organismos .............................................................. 17
6.4.2. Diseño experimental ............................................................................... 19
6.4.3. Estimación de la concentración óptima de alimento mediante la tasa de
ingestión ........................................................................................................... 21

6.5. Bioensayo 2: Efecto de la temperatura en la ecofisiología de P. sterna ........ 22
6.5.1. Tasa de ingestión .................................................................................... 22
6.5.2. Tasa respiratoria ..................................................................................... 23
6.5.3. Tasa de excreción ................................................................................... 24
6.5.4. Eficiencia de absorción ........................................................................... 25
6.5.5. Tasa de absorción .................................................................................. 26
6.5.6. Determinación del peso del tejido seco del organismo ........................... 26
6.5.7. Determinación del Potencial de Crecimiento .......................................... 26
6.6. Tratamiento estadístico de los datos .............................................................. 27
7. RESULTADOS ...................................................................................................... 28
7.1. Bioensayo 1: efecto de la concentración de alimento sobre la tasa de ingestión
a diferentes temperaturas ..................................................................................... 28
7.2. Bioensayo 2: efecto de la temperatura sobre la ecofisiología de P. sterna .... 31
7.2.1. Tasa de ingestión .................................................................................... 31
7.2.2. Tasa respiratoria ..................................................................................... 32
7.2.3. Tasa de excreción de amonio ................................................................. 33
7.2.4. Eficiencia de absorción ........................................................................... 34
7.2.5. Tasa de absorción .................................................................................. 35
7.2.6. Potencial de crecimiento ......................................................................... 36
8. DISCUSIÓN .......................................................................................................... 38
8.1.Bioensayo 1: efecto de la concentración de alimento sobre la tasa de ingestión
a diferentes temperaturas……………………………………………………………….38
8.2. Efecto de la temperatura sobre las tasas fisiológicas de P. sterna ................ 40
9. CONCLUSIONES ................................................................................................. 46
10. BIBLIOGRAFÍA .................................................................................................. 47

VI
GLOSARIO
Aclimatación: Proceso por el cual un organismo se adapta fisiológicamente a los
cambios en su ambiente.
Balance energético: Ecuación que permite estimar el potencial de crecimiento y que
contempla las entradas y salidas de energía en un organismo en relación con el
ambiente.
Bioenergética: Estudio de los intercambios de energía dentro de la célula.
Consumo de oxígeno: Cantidad de oxígeno que un organismo toma del medio para
sus procesos metabólicos.
Ecofisiología: Rama de la fisiología que estudia las adaptaciones y respuestas del
organismo en relación a variaciones en los factores ambientales.
Ectotermos: Son organismos que toman el calor procedente del medio externo.
Eficiencia de absorción: Porcentaje de materia orgánica del alimento que atraviesa
la pared del tubo digestivo. La eficiencia con la que se absorbe la concentración de
alimento ingerida.
Fisiología energética: Se ocupa del estudio de las ganancias y pérdidas de energía,
y la eficiencia de transformación de energía, desde el punto de vista de todo el
organismo.
Pseudo-heces: Partículas rechazadas de la filtración de un organismo y embebidas
en una matriz de mucopolisacáridos.
Potencial de crecimiento: Energía libre que tiene un organismo y que puede
destinarse a actividades como el crecimiento, almacenamiento de reservas

VII
energéticas, movimiento y reproducción. Ésta puede estimarse por medio de la
ecuación de balance energético.
Poiquilotermos: Son aquellos organismos que no mantienen su temperatura
corporal constante, ésta varía en función de la del medio.
Tasa de absorción: Materia orgánica del alimento (microalgas) que atraviesan la
pared del tubo digestivo de un animal por unidad de tiempo.
Tasa de crecimiento: Incremento de peso corporal de un animal por unidad de
tiempo.
Tasa de excreción de heces: Materia que es ingerida por un organismo y que no
fue absorbida por el tubo digestivo por unidad de tiempo.
Tasa de ingestión: Material que pasa a través de la boca hacia el tubo digestivo por
unidad de tiempo.
Tasa de excreción de desechos nitrogenados: Energía que fue absorbida por un
organismo y que no fue aprovechada y es desechada en forma de desechos
nitrogenados por unidad de tiempo.
Tasa respiratoria: Medida indirecta de la tasa metabólica de los organismos,
estimada por el consumo de oxígeno por unidad de tiempo.

VIII
LISTA DE TABLAS
Tabla Página
I Temperaturas a las cuales fueron expuestos organismos de P.
sterna de 6.0 ± 0.05 cm de altura de la concha y concentraciones de
I. galbana utilizadas para observar la concentración óptima a cada
temperatura.
19
II
Prueba de comparación múltiple de medias de Tukey para definir
diferencias en la tasa de ingestión de P. sterna bajo diferentes
concentraciones de alimento y temperaturas experimentales (n=10).
Las letras distintas en la columna representan diferencias
significativas entre los tratamientos a P<0.05.
30
III
Prueba de comparación múltiple de medias de Tukey para definir
diferencias en la tasa de ingestión de P. sterna bajo diferentes
temperaturas experimentales (n=10). Las letras distintas en la
columna representan diferencias significativas entre los
tratamientos a P<0.05
32
IV
Prueba de comparación múltiple de medias de Tukey para definir
diferencias en la tasa respiratoria de P. sterna bajo diferentes
temperaturas experimentales (n=10). Las letras distintas en la
columna representan diferencias significativas entre los
tratamientos a P<0.05
33
V
Prueba de comparación múltiple de medias de Tukey para definir
diferencias en la tasa de excreción de amonio de P. sterna bajo
diferentes temperaturas experimentales (n=10). Las letras distintas
en la columna representan diferencias significativas entre los
tratamientos a P<0.05.
34
VI
Prueba de comparación múltiple de medias de Tukey para definir
diferencias en la tasa de absorción de P. sterna bajo diferentes
temperaturas experimentales (n=10). Las letras distintas en la
columna representan diferencias significativas entre los
tratamientos a P<0.05.
36
VII
Prueba de comparación múltiple de medias de Tukey para definir
diferencias en el potencial de crecimiento de P. sterna bajo
diferentes temperaturas experimentales (n=10). Las letras distintas
en la columna representan diferencias significativas entre los
tratamientos a P<0.05.
37

IX
LISTA DE FIGURAS
Figura Página
1 Medidas morfométricas de la concha nácar (P. sterna). Tomado de
Saucedo (1995). 15
2 Sistema de cultivo tipo long-line para P. sterna. 16
3 Sistema de alimentación de flujo continuo utilizado para aclimatar y
alimentar a los organismos de P. sterna, a diferentes temperaturas. 18
4 Sistema empleado para la determinación del potencial de
crecimiento de la concha nacara P. sterna a diferentes
temperaturas.
20
5 Sensor de oxígeno de fibra óptica del Oximetro TX, colocado a la
salida del distribuidor, para determinar la concentración de oxígeno
disuelto dentro de las cámaras de incubación.
23
6 Tasa de ingestión de P. sterna bajo diferentes concentraciones de
alimento y temperaturas experimentales. Las barras presentan la
media ± el error estándar (n=10).
29
7 Tasa de ingestión de P. sterna bajo diferentes temperaturas
experimentales. Las barras presentan la media ± el error estándar
(n=10).
31
8 Tasa respiratoria de P. sterna bajo diferentes temperaturas
experimentales. Las barras presentan la media ± el error estándar
(n=10).
32
9 Tasa de excreción de amonio de P. sterna bajo diferentes
temperaturas experimentales. Las barras presentan la media ± el
error estándar (n=10).
34
10 Eficiencia de absorción de P. sterna bajo diferentes temperaturas
experimentales. Las barras presentan la media ± el error estándar
(n=10).
35
11
Tasa de absorción de P. sterna bajo diferentes temperaturas
experimentales. Las barras presentan la media ± el error estándar
(n=10).
36
12
Potencial de crecimiento de P. sterna bajo diferentes temperaturas
experimentales. Las barras presentan la media ± el error estándar
(n=10).
37

X
RESUMEN
Se estudió el efecto de la concentración de alimento y temperatura sobre las
respuestas fisiológicas en la concha nácar Pteria sterna, como una estrategia para
definir las condiciones óptimas para futuros experimentos sobre acondicionamiento
de reproductores y cultivo de larvas de esta especie. En un primer bioensayo, se
utilizaron 60 adultos (6.0 ± 0.05 cm de altura de la concha) para determinar, la
concentración óptima de alimento con respecto a la temperatura mediante la tasa de
ingestión (TI). Para ello se empleó la microalga Isochrysis galbana a cinco
concentraciones (3, 8, 15, 20 y 25×104 células/mL) a cinco temperaturas (16, 19, 22,
25 y 28 °C). Las variaciones en la tasa de ingestión fueron significativas entre
temperaturas; se registró el valor más alto (122.32×103 células/g/h) a 25°C a una
concentración de 25×104 células/mL, mientras que el valor más bajo (19.33×103
células/g/h) a 28 °C a 25×104 células/mL. La concentración óptima de alimento se
observó a 15×104 células/mL. En un segundo bioensayo, los mismos organismos
fueron expuestos a cinco temperaturas (16, 19, 22, 25 y 28 °C) y se alimentaron a la
concentración óptima de 15×104 células/mL de I. galbana, con el fin de determinar el
balance energético y potencial de crecimiento de la especie. La TI más alta se
observó entre 22°C y 25°C (24.74×103 y 24.43×103 células/g/h), sin embargo no fue
significativo, a diferencia de las temperaturas de 16° y 28 que arrojaron los valores
significativamente más bajos (14.14×103 y 16.28×103 células/g/h, respectivamente).
Valores más altos de la tasa respiratoria (1.72 mLO2/g/h) se obtuvieron a 28°C, y de
la eficiencia de absorción y tasa de absorción a 25 °C (82.34% y 19.94×103
células/g/h). Para la tasa de excreción, los mayores valores se presentaron a 28°C
(6.54 ± 0.94 µg NH4/g/h). Se obtuvo un potencial de crecimiento significativamente
alto de 0.09 J/g/h, lo cual indicó un gasto energético mayor que sugiere que el
intervalo óptimo fisiológico de la especie se encuentra entre 19 y 22 °C, con valores
promedio de 24.74 ± 2.2 y 22.97 ± 1.2 J/g/h, respectivamente.
Palabras clave: Pteria sterna, temperatura, potencial de crecimiento, balance
energético, tasa de ingestión.

XI
ABSTRACT
Physiological response of the winged pearl oyster Pteria sterna in relation to
food concentration and temperature
The effect of food concentration and water temperature on physiological responses in
the nacre shell Pteria sterna was studied as a strategy to define the optimal
conditions for future experiments on broodstock conditioning and larval rearing of this
species. In a first bioassay, 60 adults (6.0 ± 0.05 cm of shell height) were used to
determine the optimum food concentration with respect to temperature by ingestion
rate (IR). For this purpose the microalgae Isochrysis galbana was employed at five
concentrations (3, 8, 15, 20 and 25×104 cells / mL) at five temperatures (16, 19, 22,
25 and 28 ° C). Variations in the ingestion rate were significant between
temperatures; the highest value (122.32×103 cells /g/h) at 25 ° C at a concentration of
25×104 cells/mL was recorded, whereas the lowest value (19.33×103 cells /g/h) 28 °C
to 28×104 cells/mL. The optimum concentration of food was observed at 15×104
cells/mL. In a second bioassay, the same organisms were exposed to five
temperatures (16, 19, 22, 25 and 28 °C) and fed to the optimal concentration 15×104
cells/ mL of I. galbana, in order to determine the energy balance and scope for growth
of the species. The IR highest was observed between 22 °C and 25 °C (24.74×103
and 24.43×103 cells/g/h), however it was not significant, unlike the temperatures of 16
and 28 °C threw values significantly lower (14.14×103 and 16.28×103 cells/g/h,
respectively). Higher values of respiratory rate (1.72 mLO2 /g/h) were obtained at 28 °
C, and the absorption efficiency and absorption rate at 25 ° C (82.34% and 19.94×103
cells/g/h). For excretion rate, the highest values were presented to 28 °C (6.54 ± 0.94
µg NH4 /g/h). It was obtained a scope for growth significantly high of 0.09 J/g/h, which
indicated a greater energy expenditure suggesting that the physiological optimum
range of the species is between 19 and 22 °C, with average values of 24.74 ± 2.2 and
22.97 ± 1.2 J/g/h, respectively.
Keywords: Pteria sterna, temperature, scope for growth, energy balance, rate of
ingestion.

1
1. INTRODUCCIÓN
Dentro del grupo de los moluscos bivalvos, las ostras perleras del género Pinctada
(P. fucata, P. margaritifera, P. maxima) son especies de importancia comercial
debido a su aprovechamiento en el cultivo de perlas (perlicultura). Desde los años
70s, esta actividad se ha manejado a través de Programas de Integración Familiar-
Comunitarios que han permitido promover un activo desarrollo social y económico de
muchas regiones en países como Japón, Polinesia Francesa, Australia, Indonesia,
Islas Cook, Islas Marshall, Tonga, Kiribati y otros (Monteforte et al., 2005; Monteforte
2013; Saucedo et al., 2015).
En México existen dos especies nativas de ostras perleras, la madreperla P.
mazatlanica (Hanley 1856) y la concha nácar Pteria sterna (Gould 1851). Esta última
ha sido utilizada en algunos sitios de las costas del Golfo de California para la
producción de perlas de gran calidad y tonos multicolores que se consideran exóticos
y muy atractivos (McLaurin et al., 1999; Kiefert et al., 2004; Ruíz-Rubio et al., 2006).
Además, la especie se aprovecha para la elaboración de productos de valor
agregado como jabones y crema de concha nácar con aplicación en la industria
cosmética y farmacéutica (Rangel y Chávez, 1994; Lamghari et al., 1999; Pereira-
Mourèis et al., 2002; Saucedo et al., 2015). Estas características hacen que la
especie sea de gran interés en la acuicultura, lo cual ha estimulado la investigación
básica para optimizar los protocolos de cultivo de la especie.
Aunque la tecnología para el cultivo extensivo de ostras perleras en México se
encuentra científicamente adelantada y tecnológicamente validada para ser
transferida (Monteforte, 2013; Monteforte y Cariño, 2013), aún existe un vacío de
información sobre los requerimientos fisiológicos y nutricionales de la especie en
condiciones de laboratorio (Saucedo, 2016). A la fecha, sólo se han analizado de
forma preliminar las condiciones para el acondicionamiento gonádico de los
reproductores (Mazón-Suástegui y Avilés-Quevedo, 1988), la tasa de ingestión y
digestión de larvas (Martínez-Fernández et al., 2004) y las condiciones básicas para
el cultivo de larvas, en donde la fijación de semilla se ha logrado alrededor del día 26
(Serrano-Guzmán y Salinas-Ordáz, 1993), del día 33 (McAnally-Salas y Valenzuela-

2
Espinoza, 1990), del día 39 (Araya-Nuñez et al., 1995), o nunca se logró (Araya-
Nuñez et al., 1991). Por ello, la producción de semilla no ha logrado ser suficiente y
consistente para apoyar las actividades de producción de perlas.
Al igual que otros organismos, P. sterna se encuentra expuesta a cambios en el
ambiente. Los efectos de variables como la temperatura del agua, disponibilidad de
alimento, salinidad, oxígeno, entre otros, sobre la fisiología de los organismos son
importantes y pueden ser estudiados a través de la ecofisiología (Bayne, 1976;
Riisgard y Larsen, 2000; Peck, 2002; Soria y Merino, 2007). Esta disciplina permite
estudiar las adaptaciones fisiológicas del organismo al ambiente con el fin de
interpretar las interrelaciones del funcionamiento físico del organismo en su medio,
prediciendo el estado de salud, desarrollo, condiciones óptimas para su crecimiento,
e indicar situaciones de estrés y de deterioro ambiental (Widdows et al., 1997, 2002;
Spicer, 2014).
Entre diversos factores ambientales, la temperatura del agua juega un papel muy
importante en la fisiología y metabolismo de los organismos, ya que regula procesos
vitales como el crecimiento y la reproducción de cualquier especie acuática
(Hochachka y Somero, 2002; Brown et al., 2004; Helmuth et al., 2010). En
condiciones de cultivo, el conocimiento sobre el efecto y control de la temperatura del
agua en los organismos es esencial, ya que asegura un crecimiento más rápido, la
reducción del tiempo de producción y una mejor eficacia alimenticia (Sarà et al.,
2008; Helmuth, 2009). A su vez, la temperatura actúa como un factor controlador que
determina los requerimientos metabólicos de los organismos, rigiendo los procesos
relacionados con la transformación del alimento (Yukihira et al., 2000; Velasco, 2007;
Ezgeta-Balic et al., 2011). La mayoría de las especies presentan un rápido
crecimiento con el aumento de la temperatura hasta un cierto punto (temperatura
óptima), donde generalmente el crecimiento desciende gradual o drásticamente, por
lo que las altas temperaturas resultan adversas en la fisiología de los organismos
(Somero, 2002; Helm et al., 2004; Anestis et al., 2008).
De forma similar, las variaciones estacionales en la disponibilidad de alimento
regulan diversos procesos fisiológicos en los moluscos bivalvos (Wong y Cheung,
2003; Rueda y Smaal, 2004) y están relacionadas con factores como la talla y edad

3
de los organismos, así como a procesos exógenos como los tiempos de
aclimatación, cantidad y calidad de partículas suministradas, temperatura ofrecida y
flujo predominante (Yukihira et al., 1998a, 2000; Velasco y Navarro, 2002). La
cantidad de alimento que ingiere un organismo en un tiempo determinado es una
variable de gran interés ecológico, debido a que indica la forma en la que éste
adquiere energía del medio para llevar a cabo muchas de sus funciones biológicas,
como el crecimiento y la reproducción (Newell, 2004). Por ello, el conocimiento y
control de las variaciones en la cantidad y calidad de alimento permite determinar y
prever el posible impacto de la especie sobre los recursos alimenticios de un
determinado lugar, así como seleccionar sitios de cultivo, optimizar la producción y
gasto de alimento, entre otros (Ibarrola et al., 1998; Yukihira et al., 1998a, b;
Kesarcodi-Watson et al., 2001; Velasco, 2007).
Dentro de las variables ecofisiológicas, el potencial de crecimiento es utilizado
para determinar directamente la cantidad de energía destinada al crecimiento
(Warren y Davis, 1967). Permite visualizar el modo en que los factores ambientales
afectan el metabolismo del crecimiento, simulando las condiciones ambientales de
interés que permitan conseguir una buena concordancia entre el crecimiento
estimado y el medido (Dame, 2011). Además describe, explica y predice la condición
fisiológica del organismo mediante el cálculo de la energía consumida, gastada y/o
asimilada dentro por el organismo, por medio de la ecuación de balance energético.
Éste, es un concepto útil que permite comprender los fenómenos ecológicos e
interpretar aquellos relacionados con la distribución de los organismos y su
adecuación al medio (Pouvreau et al., 2006; Sokolova et al., 2012).
El presente trabajo estuvo dirigido a estudiar las respuestas fisiológicas de P.
sterna ante cambios combinados en la temperatura del agua y la concentración de
alimento. El estudio de dicha interacción de factores es importante, pues podría
predecir el estado de bienestar y/o estrés de esta especie (Widdows et al., 2000) y
determinar los procesos fisiológicos relacionados con la adquisición (ingestión,
absorción) y utilización (respiración y excreción) de energía en organismos
mantenidos en condiciones de laboratorio. Esto es fundamental para optimizar los
protocolos de manejo y cultivo de la especie.

4
2. ANTECEDENTES
2.1. Generalidades de P. sterna
La ostra perlera P. sterna, comúnmente conocida como concha nácar, pertenece a la
familia Pteriidae (Keen, 1971). La morfología de sus valvas es variable, relativamente
delgada, a oblicuamente ovalada con una coloración marrón oscura en el exterior y
nacarado intenso en el interior (Saucedo, 1991; Poutiers, 1995; Pérez-Estrada,
2000). La especie posee una extensión en la charnela, la cual le da origen al nombre
(Pterion en griego significa ala). El ala es más larga en la parte anterior que la
posterior (Saucedo, 1991).
P. sterna es una especie de talla mediana a grande considerando los rangos de
talla que presentan los miembros de la familia (Monteforte, 2013); alcanza una
longitud máxima de 12 cm de altura de la concha (Keen, 1971; Poutiers, 1995),
aunque se han reportado organismos de tallas mayores de 18 cm (Sevilla, 1969;
Shirai y Sano, 1972; Singh et al., 1982).
2.2. Hábitat y distribución
P. sterna es un organismo bentónico, con una forma de alimentación por filtración no
selectiva (Arizmendi-Castillo, 1996; Martínez-Fernández et al., 2004); habita desde la
zona intermareal hasta los 35-40 m de profundidad (Poutiers, 1995) y aunque se
adhiere a sustratos rocosos, tiene preferencia por aquellos de tipo lodoso-fangoso
menos firmes (Bervera-León, 1994). A diferencia de otras especies solitarias, P.
sterna se fija formando agrupaciones o conglomerados masivos que localmente se
conocen como “macoyos”. Se distribuye a lo largo de un amplio margen de la línea
costera tropical, subtropical y templada en el Pacífico oriental, que comprende desde
la parte alta del Golfo de California, México, hasta Perú (Keen, 1971; Alamo y
Valdivieso, 1997).
El rango de temperatura en el que se desarrolla P. sterna es de 18°C a 32 °C,
muy similar al reportado para P. mazatlanica (Gervis y Sims, 1992), aunque la

5
concha nácar presenta una marcada preferencia por las temperaturas bajas, ya que
su temporada de reproducción se extiende de los meses de diciembre a mayo del
siguiente año, cuando las temperaturas oscilan entre 21 y 24 °C (Saucedo y
Monteforte, 1997a; Vite-García y Saucedo, 2008; Cáceres-Puig et al., 2009).
2.3. Ecología y biología
A la fecha se han estudiado diversos aspectos de la biología y ecología de P. sterna
que afectan el cultivo en campo. Estos incluyen la recolecta de semilla mediante
colectores artificiales de diferentes materiales y colores (Cáceres-Martínez et al.,
1992; Monteforte y Aldana, 1994; Monteforte y García-Gasca, 1994; Monteforte y
Wright, 1994; Monteforte et al., 1995; Torres-Zepeda et al., 2002) y la pre-engorda y
engorda de juveniles y adultos hasta talla comercial, empleando diferentes métodos
de armado de líneas y estructuras en campo (Gaytán-Mondragón et al., 1992;
Monteforte, 2013). También se ha analizado el crecimiento de juveniles y adultos
bajo condiciones de cultivo extensivo (Bückle-Ramírez et al., 1992; Saucedo y
Monteforte, 1997b; Vite-García y Saucedo, 2008; Cáceres-Puig et al., 2009),
estimando algunos parámetros de crecimiento importantes (Cáceres-Martínez et al.,
1991, 1992; Saucedo, 1991, 1995; Bückle-Ramírez et al., 1992; Del Río-Portilla et al.,
1992; Monteforte y García-Gasca, 1994; Rangel-Dávalos y Cáceres, 1994; Wright y
Monteforte, 1995; Monteforte et al., 1995; Wright-López, 1997; Saucedo y
Monteforte, 1997b; MacLaurin et al., 1999; Young, 1999; Monteforte et al., 2005;
Wright-López et al., 2009).
Por su parte, estudios sobre el ciclo reproductivo y la temporalidad de los
principales eventos relacionados con la maduración y el desove de P. sterna en
diversos sitios del Pacífico y Golfo de California han aportado información importante.
Se sabe que en Bahía de Los Ángeles, B.C., P. sterna desova de manera continua a
lo largo del año presentando tres picos reproductivos; en diciembre, febrero y agosto
(Hernández-Díaz y Bückle-Ramírez, 1996); en Bahía de La Paz, la especie presenta
dos picos de desove a lo largo del año, uno en febrero y otro en mayo a una
temperatura de entre 22.2 y 23.4 °C (Saucedo y Monteforte, 1997a). En Guaymas,

6
Sonora, se ha reportado un solo periodo de desove, cuando la temperatura del agua
varía de 19 a 22 °C (Arizmendi-Castillo, 1996); mientras que en Laguna Ojo de
Liebre, B.C.S., se reporta la ocurrencia del desove a temperaturas de entre 23 y 25
°C (Hernández-Olalde et al., 2007).
A su vez, se han analizado las estrategias de almacenamiento y movilización de
reservas energéticas entre los tejidos somáticos y la gónada durante la
gametogénesis, señalando que la cantidad de energía de los tejidos somáticos
(músculo, manto y glándula digestiva) decrece cuando la temperatura disminuye.
Esto presumiblemente es debido a que las temperaturas bajas (∽21 °C) estimulan el
movimiento de energía hacia la gónada, mientras que las temperaturas elevadas
(∽28.8°C) muestran un evento de acumulación en tejidos somáticos, así como los
mínimos de energía en la gónada (Vite-García y Saucedo, 2008, Cáceres-Puig et al.,
2009).
El que existiera la limitante de obtener gametos maduros durante la temporada
reproductiva canalizó los esfuerzos de investigación hacia los principales factores
reguladores de la gametogénesis en reproductores acondicionados en condiciones
de laboratorio. En este sentido, se estudió el efecto de la dieta microalgal
complementada con productos ricos en carbohidratos (fécula de maíz) y en lípidos
(microcápsulas de ácidos grasos) en el desarrollo y composición de la gónada y
tejidos somáticos (Hernández-López, 2012) observándose que su utilización favorece
el desarrollo y propicia una mejor condición general en los organismos. Por otra
parte, se estudió el efecto de la temperatura en el desarrollo y composición gónada,
en el cual P. sterna es expuesta cinco regímenes térmicos: 22° C, ascendente (19 a
27°C), descendente (27 a 19° C) y dos temperaturas oscilantes (21 y 22 °C) durante
34 días (Granados-Amores, 2012), teniendo como resultado un transporte de
carbohidratos desde el músculo aductor hacia la gónada en los machos, reflejado en
la presencia de estadios de desarrollo gonádico avanzado (régimen térmico
descendente), mientras que en las hembras hubo acumulación de lípidos en la
gónada por parte del régimen constante y ascendente.
Por su parte, Cáceres-Puig (2012) estudió los factores bióticos y abióticos que
influyen sobre la dinámica de reclutamiento de semillas de P. sterna en Bahía de La

7
Paz. Los autores señalan que tanto la temperatura (19 y 24°C) como la energía
contenida en el alimento (seston ~1.3×10-2 mg/L) provocan un mayor impacto sobre
el reclutamiento de semillas y una mejor condición fisiológica de los organismos
reproductores.
Asimismo, se ha determinado la relación que existe entre la condición de los
reproductores de P. sterna y el desempeño larvario a lo largo de toda la temporada
reproductiva en la Bahía de La Paz (Gómez-Robles et al., 2013). De forma particular
se reporta que a una temperatura de 21.8°C con una concentración de clorofila a de
1.3 mg/m3 se presenta la mejor condición reproductiva con base en indicadores de la
calidad gonádica.
2.4. Balance energético y potencial de crecimiento
Una de las herramientas de la ecofisiología es el balance energético, que permite
medir el estado energético desde un ecosistema a hasta una célula y puede predecir
la cantidad de energía disponible para el crecimiento, movimiento y la reproducción
(Bayne y Newell, 1983; Newell, 2004).
El balance energético contempla las principales ganancias y pérdidas de energía
en el organismo, reflejadas en las tasas fisiológicas, como ingestión, respiración,
absorción y excreción (Bayne, 1976). Esta energía neta es el resultado del balance
entre la energía: 1) adquirida del alimento (asimilación), 2) el costo energético de la
respiración, 3) el metabolismo de proteínas y 4) la energía no asimilada y excretada
por medio de las heces (Bühringer y Danischewski, 2001).
La energía adquirida proviene de la tasa de ingestión (TI), mientras que los
gastos de energía están representados por la tasa respiratoria (TR), la tasa de
excreción de compuestos nitrogenados (TU) y la tasa de excreción de heces (TH).
Warren y Davis (1967) expresaron esta compleja red de interacciones ecofisiológicas
a través de la siguiente fórmula:
Balance energético = TI-(TR+TU+TH)

8
De acuerdo con la clasificación de Widdows et al. (1995a), los valores del
potencial de crecimiento indican condiciones que van desde de menor a mayor
estrés. Como este equilibrio puede ser alterado por el estrés inducido por la
contaminación, el potencial de crecimiento ha sido también recomendado como
medida sensible de los efectos contaminantes en animales marinos (Widdows et al.,
1995a). De forma particular, el potencial de crecimiento ha sido estimado en algunas
especies de moluscos bivalvos, como Ctenopharyngodon idella (Espina et al., 1986),
Mytilus edulis (Widdows et al., 1988, 1995a, b, 2002; Widdows y Johnson, 1988;
Widdows y Page, 1993; Bühringer y Danischewski, 2001), M. galloprovincialis
(Widdows et al., 1997), Arca zebra (Widdows et al., 1990) y Meretrix meretrix (Scarlet
et al., 2015).
2.5. Influencia de la temperatura en el metabolismo y balance energético de los
moluscos bivalvos
La mayoría de los organismos marinos, incluyendo los bivalvos, son ectotermos y
muchos aspectos de su balance energético, como la cantidad total de la energía
invertida y/o la proporción de energía consumida asignada al crecimiento son
especialmente sensibles a los cambios de temperatura externos (Kooijman, 2010).
Por lo tanto, la temperatura es un factor principal que tiene influencia en las
respuestas fisiológicas de los organismos (Gillooly et al., 2001; Sarà et al., 2011;
Kearney et al., 2010) como la alimentación, respiración y reproducción (Newell y
Bayne, 1973). Esto debido a que un aumento en la temperatura tiende a aumentar el
metabolismo, y como consecuencia, las necesidades energéticas del organismo
(Bernabé, 1991). Por ello, se considera que la temperatura actúa como un factor
ambiental clave en la determinación del metabolismo (Bayne et al., 1976) y el nicho
fundamental de una especie (Schoener, 1986; Chase y Leibold, 2003).
Los efectos de la temperatura sobre la fisiología y la bioenergética de los
bivalvos que se alimentan de materia particulada en suspensión han sido bien
documentados, tanto en especies intermareales como Geukensia demissa (Wilbur y
Hilbish, 1989) y Brachidontes pharaonis (Sara et al., 2008), así como en otros de

9
hábitos submareales como Modiolus barbatus (Ezgeta-Balic et al., 2011), M.
modiolus (Navarro y Thompson, 1996), M. galloprovinciallis (Pérez-Camacho et al.,
2000; Sarà y Pusceddu, 2008), M. edulis (Widdows, 1976; Sukhotin et al., 2003),
Perna viridis (Wong y Cheung, 2003), Argopecten purpuratus (Navarro et al., 2000),
P. margaritifera y P. maxima (Yukihira et al., 2000).
Por lo general, las tasas de filtración de los bivalvos que se alimentan de materia
particulada en suspensión aumentan con el incremento de temperatura hasta una
cierta temperatura, por encima de la cual disminuyen (Winter, 1978; Shumway, 1982;
Griffiths y Griffiths, 1987). Sin embargo, Bayne (1976) argumentó que el aumento de
la temperatura disminuye ligeramente la eficiencia de absorción del mejillón Mytilus
sp. (0.7% de disminución por cada 1 °C), pero Elvin y Gonor (1979) reportaron una
mayor eficiencia de absorción en M. californianus a 15 °C que a 9 °C. Por su parte,
M. barbatus presentó una tasa de aclaramiento más alta al incrementar la
temperatura de 20°C a 28°C, mientras que la tasa respiratoria fue máxima a 9 °C y
mínima a 26°C, con valores promedio más altos de energía absorbida a los 20 y 26
°C. El potencial de crecimiento de la especie arrojó valores negativos para 9, 15 y 28
°C y valores positivos a 20 y 26°C (Ezgeta-Belic et al., 2011).
Navarro et al. (2000) reportan que el ostión del norte A. purpuratus no muestra
diferencias significativas en la tasa de aclaramiento entre 16 y 20 °C cuando se
alimenta con microalgas puras y microalgas adicionadas con lípidos. Ello muestra la
capacidad de la especie para modular sus tasas fisiológicas dentro de un cierto
rango de temperatura, así como la independencia de la eficiencia de absorción de la
temperatura. Para el caso de M. edulis, parece que se adapta a temperaturas cíclicas
(6 a 20°C) al reducir la amplitud de respuesta de consumo de oxígeno y la tasa de
filtración, y con ello aumenta su independencia de la temperatura dentro del intervalo
del régimen fluctuante (Widdows, 1976).
La mayoría de los estudios sobre los efectos de la temperatura en los procesos
fisiológicos en las ostras perleras han sido reportadas para P. fucata martensii
(Miyauchi, 1962; Seki, 1972; Numaguchi y Tanaka, 1986; Numaguchi, 1994). Para el
caso particular de la semilla mantenida en laboratorio, se registra una alta mortalidad
a <12.5 °C y 35 °C, así como un incremento significativo en la tasa de crecimiento

10
entre 15 y 32 °C. El rango de temperatura óptimo para la semilla fue de 17.5 a 29 °
(Numaguchi y Tanaka, 1986). Esta especie tiene una tasa metabólica muy baja en
invierno, cuando la temperatura del agua está por debajo de 13 °C (Kobayashi y
Tobata, 1949).
Miyauchi (1962) y Numaguchi (1994) encontraron que los más altos niveles de
aclaramiento de P. fucata martensii ocurrieron entre 24 y 27 °C y 25 y 28 °C,
respectivamente. Las tasas de aclaramiento disminuyeron a temperaturas >28 °C
(verano) y <13 °C (invierno) (Numaguchi, 1994).
Por su parte, Yukihira et al. (2000) expusieron a P. margaritifera y P. maxima a
temperaturas de 19, 23, 28 y 32°C y determinaron diferentes parámetros
ecofisiológicos. Los autores reportaron que la tasa de aclaramiento, en P.
margaritifera fue significativamente mayor que para P. maxima a 19 ° C, mientras
que P. maxima tenía una eficiencia de absorción mayor que P. margaritifera a los 28
y 32 °C. A su vez, la temperatura afecta significativamente la energía respirada de P.
margaritifera sobre un rango de temperatura más amplio (19-32 °C) que P. maxima
(19-23 °C). Sin embargo, el potencial de crecimiento es mayor a 32 ° C y 19°C
respectivamente. Estos resultados están de acuerdo con las observaciones de la
presencia de P. margaritifera en latitudes más altas y los hábitats de temperatura
más bajos, y que la diferencia de las tasas fisiológicas en las dos especies es debido
probablemente a las diferencias en distribución y ocupación de hábitats.
La capacidad de modular las tasas fisiológicas en función de los cambios en la
temperatura ambiental ha sido considerada como una adaptación para los
organismos que habitan en entornos inestables térmicamente (Wilbur y Thompson,
1989). Por tanto, la aclimatación de las tasas metabólicas en respuesta a la
temperatura variables se ha considerado como un importante mecanismo de
compensación por los animales (Segal, 1961; Precht et al., 1973; Prosser, 1973;
Newell, 1979).
El régimen térmico del medio en el que habitan los organismos condiciona las
actividades alimenticias (Navarro, 2001), lo que se ve reflejado en un historial térmico
y el tiempo de aclimatación. Organismos que habitan en un ambiente con pocas
fluctuaciones de temperatura, la tasa de aclaramiento es termodependiente en un

11
amplio intervalo de temperaturas de exposición, caso contrario para los organismos
sometidos a marcadas fluctuaciones quienes presenten una zona de
termodependecia comprendida en el rango habitual de temperaturas (Widdows,
1976).
2.6. Influencia de la disponibilidad de alimento en el metabolismo y balance
energético de los moluscos bivalvos
Además de la calidad del alimento que afecta el crecimiento y maduración de los
bivalvos (Bayne y Newell, 1983; Heasman et al., 1996), se sabe que la presencia o
ausencia de ciertas especies de microalgas en una dieta es más importante que su
contenido total de proteínas, lípidos, carbohidratos, aminoácidos y hasta ácidos
grasos (Epifanio, 1979; Velasco y Navarro, 2002, 2003; Maeda-Martínez et al., 2016).
Esto debido a que el ciclo gametogénico de los bivalvos marinos se acopla con la
dinámica de síntesis de los lípidos durante la vitelogénesis, lo cual ocurre a expensas
del glucógeno almacenado o tomado de la dieta (Gabbot, 1975; Navarro et al., 2000;
Velasco, 2007; Gómez-Robles et al., 2013).
Varios estudios de las respuestas fisiológicas respecto a los diferentes tipos de
microalgas utilizadas con bivalvos filtradores han concluido que las tasas fisiológicas
varían de acuerdo a la especie de microalga, suplemento alimenticio y especie de
bivalvo (Velasco, 2007). En C. virginica y M. edulis bajo diferente calidad de
partículas se reportó el tiempo de residencia, tiempo de manipulación de las
partículas en los palpos labiales probando Rhodomonas lens, Spartina sp. y 50/50 de
ambas, siendo C. virginica a la cual le afecta el cambio de dieta, disminuyendo el
tiempo de manipulación de la partícula en los palpos labiales con el incremento de la
calidad de la dieta (Ward et al., 2003).
Otros estudios se han enfocado en estudiar las variaciones en las tasas de
ingestión de los bivalvos, así como su crecimiento y reproducción por efecto de la
disponibilidad de alimento. Ejemplos de esto se han reportado en especies como
Perna viridis (Hawkins et al., 1997), M. edulis (Schulte, 1975; Bayne et al., 1987;
Hawkins et al., 1997), C. edule (Navarro y Widdows, 1997), Crassostrea gigas

12
(Bougrier et al., 1997) y C. virginica (Schulte, 1975). En condiciones ambientales
naturales, los bivalvos que habitan estuarios y los hábitats costeros poco profundos
están expuestos con frecuencia a niveles de seston por encima del límite en el que
algún material filtrado de la suspensión es rechazado como pseudoheces, de forma
que la proporción de material filtrado rechazado de esta manera varía tanto con la
cantidad y el contenido orgánico del seston. En el proceso de rechazo, un grado de
clasificación se afecta por el cual el material relativamente enriquecido en contenido
orgánico se selecciona para la ingestión (Riisgard, 2001; Ward et al., 2003). Por ello,
muchos autores utilizan suspensiones muy concentradas de algas (Allen, 1962;
Davids, 1964; Ali, 1970), que no tienen relación con las concentraciones de
partículas de alimentos que se producen normalmente en el medio natural.
Inoportunamente, esta estrategia genera valores bajos o incorrectos de filtración de
alimento, ya que el mecanismo se encuentra muy perturbado por la sobrecarga de
partículas en suspensión (Schulte, 1975) y suele traducirse en formación de
pseudoheces. Para miembros de la familia Pteriidae, se ha reportado que no existe
alimentación selectiva de partículas orgánicas (Yukihira et al. 1999, 2000), y que
especies como P. margaritifera y P. maxima presentan una alta capacidad de adquirir
la energía en condiciones de poco fitoplancton, lo que las coloca en una categoría
excepcional en términos de flujos de energía, con tasas de aclaramiento de 50 a 100
L h-1. Ambas especies reportan entre los valores más altos de tasa de aclaramiento,
tasa respiratoria, tasa de excreción de amonio y potencial de crecimiento registrados
para muchos bivalvos, incluidos los alimentados a altas concentraciones (Yukhira et
al. 1998a).
Yukihira et al. (1998a) también demostraron que en condiciones oligotróficas
simuladas (0.5 mg L-1 de Isochrysis galbana variedad Thaitiana), P. margaritifera y P.
maxima presentan altas tasas de aclaramiento y altos valores de potencial de
crecimiento en comparación con muchas otras especies de bivalvos, incluyendo los
que se alimentan de las concentraciones más altas de fitoplancton. De hecho, P.
maxima parece estar mejor adapta a una amplio rango de concentraciones de
alimento que P. margaritifera. Se han reportado máximos valores positivos de
potencial de crecimiento de 7 mg L-1 de Dunaliella primolecta) y hasta de 9 mg L-1 de

13
I. galbana (variedad Tahiti) para P. maxima, sin embargo para P. margaritifera estos
valores son de 7 mg L-1 y 5 mg L-1 (Yukhira et al., 1998b). Parece que ambas
especies son capaces de crecer y reproducirse en condiciones de bajo suministro de
fitoplancton.
3. JUSTIFICACIÓN
P. sterna es considerada una especie de interés comercial en el noroeste de México,
debido a que presenta la capacidad de aprovechamiento integral en productos de
alto valor agregado, particularmente perlas (medias perlas y perlas esféricas). La
concha se puede utilizar para fabricar artesanía, así como jabón para la industria
cosmética y polvo de nácar para la industria farmacéutica y biomédica. Sin embargo,
falta información sobre sus requerimientos fisiológicos y nutricionales de esta especie
que coadyuven al establecimiento del cultivo en laboratorio. Debido a la influencia
que tiene la temperatura del agua y la disponibilidad de alimento sobre los procesos
fisiológicos, el crecimiento y la reproducción, es particularmente importante definir
condiciones adecuadas de manejo de ambos factores en el laboratorio, con la
finalidad de definir sus necesidades energéticas y optimizar una tecnología para la
maduración de reproductores y la producción de semilla con aplicación para el cultivo
de perlas. En P. sterna, ninguno de estos aspectos han sido estudiados y son
fundamentales para construir una base de conocimientos para el establecimiento de
un cultivo sustentable.
4. HIPOTESIS
Dada la importancia de la temperatura del agua y la disponibilidad de alimento en la
fisiología y metabolismo de los moluscos bivalvos filtradores, se espera que al
someter a individuos de P. sterna a temperaturas y concentraciones de alimento
opuestas en su límite de distribución natural ocurran ajustes significativos en sus
tasas de ingestión, eficiencia y tasa de absorción y tasas de respiración y excreción.

14
Igualmente, se espera que estas variaciones afecten de forma significativa la
ecuación general de balance energético y el potencial de crecimiento de la especie.
5. OBJETIVOS
5.1. General
Estudiar el efecto de la concentración de alimento y temperatura del agua sobre las
respuestas fisiológicas implicadas en el potencial de crecimiento de P. sterna, con el
fin de determinar las condiciones óptimas para el manejo y crianza de la especie bajo
condiciones controladas de laboratorio.
5.2. Específicos, bajo condiciones de laboratorio para P. sterna:
Definir el intervalo óptimo de la concentración de alimento en función de la
temperatura.
Determinar el efecto de la temperatura sobre la eficiencia de absorción de
energía.
Determinar el efecto de la temperatura sobre la tasa de respiración y
excreción.
Determinar el potencial de crecimiento óptimo en relación a la temperatura.
6. METODOLOGÍA
6.1. Obtención de organismos
Se utilizaron 60 ejemplares de P. sterna con una talla promedio de 6.0 ± 0.05 cm de
altura de la concha (eje dorso-ventral) y 7.0 ±0.04 cm de longitud de la concha (eje
antero-posterior), según se muestra en la Figura 1. Los organismos provinieron de la
granja Perlas del Cortez S. de R. L. M. I., ubicada en la Bahía de La Paz, B. C. S.,
México (24°46’ y 24°07’ N y 110° 18’ y 110° 38’ O).

15
Figura 1. Medidas morfométricas de la concha nácar (P. sterna). Tomado de
Saucedo (1995).
La recolecta de los ejemplares se realizó en el mes de junio de 2012, cuando los
organismos se encuentran fuera de la temporada reproductiva (Saucedo y
Monteforte, 1997a). Con esto se evitó que una parte del presupuesto energético se
destinara al proceso reproductivo. La temperatura promedio del agua en estas fechas
se encontraba en 23 °C y la salinidad en 37 ups. Cabe señalar que el cultivo utilizado
es de tipo extensivo por medio de jaulas de plástico suspendidas en la columna de
agua, manteniéndose cerca de la superficie mediante boyas (sistema “long line”),
como se muestra en la Figura 2.

16
Figura 2. Sistema de cultivo tipo long-line para P. sterna.
6.2. Traslado y mantenimiento de los organismos
Los organismos fueron trasladados al Laboratorio de Ecofisiología de Organismos
Acuáticos del Centro de Investigaciones Biológicas del Noroeste (CIBNOR) para la
realización de dos bioensayos. El traslado se llevó a cabo en medio húmedo dentro
de una hielera plástica, envolviendo a los organismos con papel mojado antes de
colocarlos en la hielera para evitar la desecación de los tejidos blandos. Para
mantener bajo el metabolismo de los organismos, la temperatura se mantuvo a 20 °C
con ayuda de un hidrogel congelado en el fondo de la hielera. El traslado duró
aproximadamente 45 min y una vez que los organismos llegaron al laboratorio, se
limpiaron de fauna acompañante e incrustante.
6.3. Pre-aclimatación de los organismos
Previó a la realización de los bioensayos 1 y 2 (ver abajo), los organismos
permanecieron en el laboratorio en etapa de pre-aclimatación durante 15 días, con la

17
finalidad de permitir su recuperación al posible estrés producido por el trasporte y la
manipulación (Ocaño-Higuera 2003). Para esto, los organismos se colocaron dentro
de un sistema de mantenimiento de flujo continuo de agua, consistente de cinco
tanques de plástico de 80 L con 12 organismos cada uno.
El sistema estaba provisto de un mecanismo de recirculación de agua de mar
con alimento, el cual era bombeado desde un tanque de 150 L mediante una bomba
sumergible Wizard modelo 5-MSP de 1/6 HP. La concentración de alimento se
mantuvo a 15×104 células/mL/día de una mezcla de las microalgas I. galbana y
Chaetoceros calcitrans en proporción 1:1 (en cuanto a conteo celular).
El sistema también estaba provisto de agua de mar filtrada a 1 μm, manteniendo
condiciones constantes de temperatura (20 ± 0.5 °C), salinidad (37 ups), fotoperiodo
(12h luz: 12 h oscuridad) y aireación. El control térmico se logró manteniendo el
sistema dentro de una cámara a temperatura controlada a 18°C y a cada tanque se
le colocó un termocalentador sumergible (RENA-Cal® de 300 W) para mantenerla
estable a 20°C. Para cada tanque con organismos se mantuvo un tanque adicional
con agua de recambio a las mismas condiciones. Diariamente las heces fecales
fueron removidas por medio de un sifón y se efectuaron recambios del 50% del agua
en cada tanque.
6.4. Bioensayo 1: Determinación de la concentración óptima de alimento en
relación a la temperatura del agua
6.4.1. Aclimatación de los organismos
Una vez finalizada la etapa de pre-aclimatación, los 60 organismos se sometieron a
dos diferentes bioensayos; este primero dirigido a determinar la concentración óptima
de alimento bajo diferentes temperaturas experimentales (16, 19, 22, 25 y 28°C) y el
segundo a determinar el balance energético y potencial de crecimiento de la especie
(ver subsección 6.5). Estas temperaturas se mantuvieron con termocalentadores
RENA-Cal® 300W que permitieron disminuir o aumentar 1°C por día la temperatura

18
del agua hasta alcanzar el valor experimental deseado, partiendo de la temperatura
de pre-aclimatación (20°C).
Asimismo, a la cámara en donde se llevaron a cabo los dos bioensayos se le
disminuyó la temperatura 1°C por día hasta alcanzar 13°C, con la finalidad de
mantener la temperatura de 16°C, siendo esta la más baja en ambos bioensayos
(Figura 3). Cada temperatura fue verificada mediante un termómetro (Ever Ready
Thermometer Co) y adicionalmente se colocó un termógrafo digital (Onset HOBO-
Pendant Temperature/Light Data Logger) programado para registrar cada 10 min la
temperatura de los tanques y observar las variaciones en cada temperatura
expuesta. El mantenimiento y aclimatación de los organismos se realizó por una
semana, bajo las mismas condiciones que durante el periodo de pre-aclimatación
(subsección 6.3).
Figura 3. Sistema de alimentación de flujo continuo utilizado para aclimatar y
alimentar a los organismos de P. sterna, a diferentes temperaturas.

19
6.4.2. Diseño experimental
Para determinar la concentración óptima de alimento a cada temperatura se estimó
la tasa de Ingestión (TI) de una sola especie de microalga (I. galbana), la cual se
suministró a cinco diferentes concentraciones (Tabla 1). Este diseño experimental bi-
factorial permitió analizar el efecto combinado de la temperatura y concentración de
alimento de forma individual para cada organismo (ver subsección 6.4.3.). Esto se
realizó colocando a los organismos dentro de un sistema de cámaras de incubación
con flujo abierto descrito por Sicard (2006).
Tabla I. Temperaturas a las cuales fueron expuestos
organismos de P. sterna de 6.0 ± 0.05 cm de altura
de la concha y concentraciones de Isochrysis
galbana utilizadas para observar la concentración
óptima a cada temperatura.
El sistema de cámaras de incubación consiste de frascos de vidrio de 1.5 L con
tapa hermética de polietileno rígido adaptada con un empaque de silicón. El agua
con alimento es bombeada hacia el interior de cada cámara, insertando una
manguera a través de un orificio hecho en la tapa de cada cámara y llevándola hasta
el fondo de la misma. La exclusión de las burbujas y el exceso de agua se consiguen
por la forma interior cónica de la tapa y la perforación del ápice del cono, donde fue
colocada una manguera. El flujo de entrada al interior de cada cámara se mantiene
Temperatura
experimental (°C)
Concentración de alimento
(células/mL×104)
16 3, 8, 15, 20 y 25
19 3, 8, 15, 20 y 25
22 3, 8, 15, 20 y 25
25 3, 8, 15, 20 y 25
28 3, 8, 15, 20 y 25
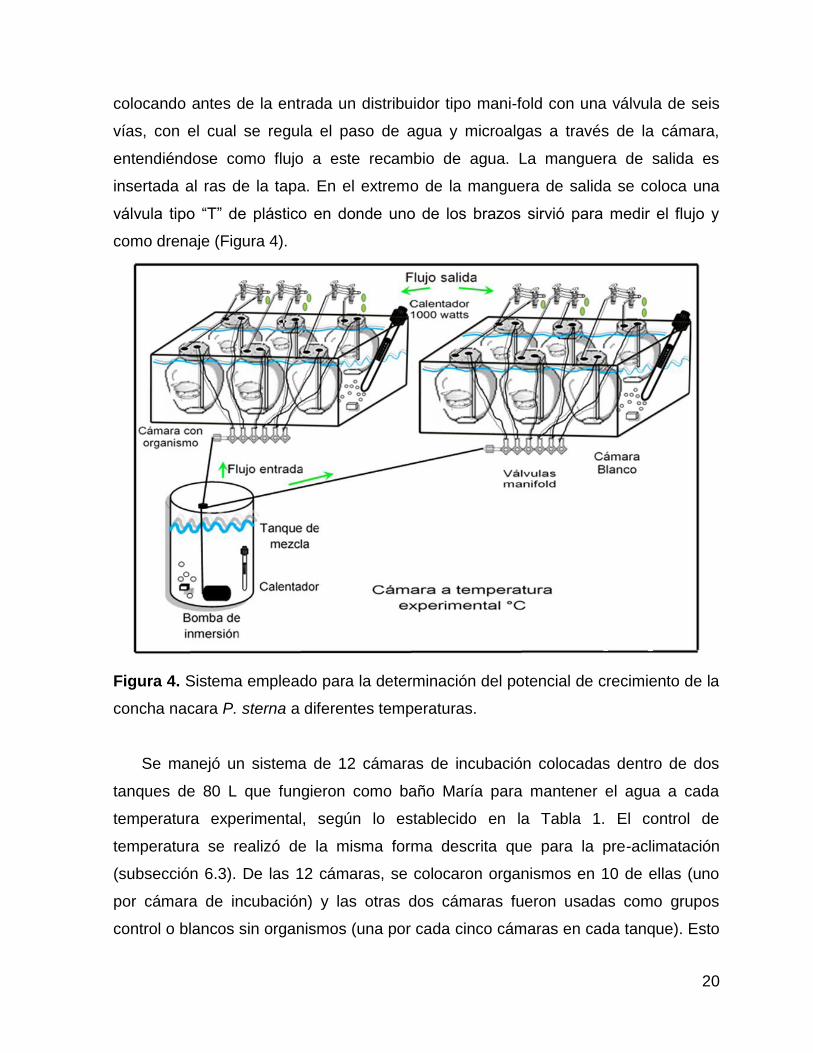
20
colocando antes de la entrada un distribuidor tipo mani-fold con una válvula de seis
vías, con el cual se regula el paso de agua y microalgas a través de la cámara,
entendiéndose como flujo a este recambio de agua. La manguera de salida es
insertada al ras de la tapa. En el extremo de la manguera de salida se coloca una
válvula tipo “T” de plástico en donde uno de los brazos sirvió para medir el flujo y
como drenaje (Figura 4).
Figura 4. Sistema empleado para la determinación del potencial de crecimiento de la
concha nacara P. sterna a diferentes temperaturas.
Se manejó un sistema de 12 cámaras de incubación colocadas dentro de dos
tanques de 80 L que fungieron como baño María para mantener el agua a cada
temperatura experimental, según lo establecido en la Tabla 1. El control de
temperatura se realizó de la misma forma descrita que para la pre-aclimatación
(subsección 6.3). De las 12 cámaras, se colocaron organismos en 10 de ellas (uno
por cámara de incubación) y las otras dos cámaras fueron usadas como grupos
control o blancos sin organismos (una por cada cinco cámaras en cada tanque). Esto

21
permitió evaluar, por duplicado, el efecto de la temperatura y concentración de
alimento en la TI (Figura 4). El flujo de agua con alimento hacia las cámaras se llevó
a cabo mediante una bomba sumergible colocada dentro de un tanque de 200 L con
agua marina filtrada a 1 µm y la microalga a la concentración y temperatura deseada
(Tabla 1).
El tanque con alimento se mantuvo con aireación continua por medio de un
difusor para homogenizar la concentración de células y mantener la concentración de
oxígeno a saturación. Se colocó una tapa en el tanque para inhibir la producción de
las microalgas. Previo al bioensayo, los organismos se mantuvieron en inanición
durante 12 h y posteriormente se colocaron en las cámaras experimentales descritas,
bombeando previamente agua marina filtrada a 1 µm a cada temperatura
experimental, por un tiempo de 25 min. El oxígeno se mantuvo a saturación y la
salinidad a 37 ups. Los bioensayos se realizaron manteniendo constante un flujo de
agua con alimento a la concentración de interés a 50 mL/min.
6.4.3. Estimación de la concentración óptima de alimento mediante la tasa de
ingestión
La tasa de ingestión (TI) se calculó de manera individual, midiendo el consumo de
alimento de cada organismo por unidad de tiempo, el cual fue estandarizando al peso
seco de sus tejidos. Los organismos se alimentaron mediante el sistema descrito
anteriormente (subsección 6.4.2) permitiendo que se estabilizaran durante 2 h.
Posterior a este tiempo, el consumo se midió recolectando muestras de 20 mL, por
triplicado, del agua con alimento a las salidas de cada cámara de incubación, así
como la cámara blanco. Las muestras se fijaron con 0.2 mL de una solución de lugol
y formaldehído al 4% neutralizado con borato de sodio. La cuantificación de células
se realizó por medio de un contador de partículas Coulter Multisizer 3, empleando la
fórmula:

22
El consumo de alimento se estimó por diferencia de la concentración celular de la
cámara blanco menos la concentración celular de cada una de las cámaras de
incubación con alimento. Por su parte, el peso seco se determinó de una muestra de
10 organismos de la misma talla (subsección 6.5.7.).Los resultados de la TI se
expresaron como número de células/g/h y luego fueron convertidos a unidades
energéticas utilizando el equivalente energético de materia particulada (POM), que
es de 23500 J/g (Widdows et al. 1979).
6.5. Bioensayo 2: Efecto de la temperatura en la ecofisiología de P. sterna
Una vez determinada la concentración óptima de alimento para los organismos, se
realizó un segundo bioensayo dirigido a estimar su potencial de crecimiento (PC) en
función de las mismas temperaturas experimentales (Tabla 1). Para ello, se
determinaron una serie de variables o tasas fisiológicas que componen la ecuación
general de balance energético propuesta por Winberg (1960). Estas variables
incluyen la tasa de ingestión (TI), tasa de respiración (TR), tasa de excreción (TE),
tasa de absorción (TA) y eficiencia de absorción (EA). El diseño experimental y
manejo de los organismos en las cámaras de incubación fue el mismo que se
describió para el Bioensayo 1 (ver subsección 6.4.3.).
6.5.1. Tasa de ingestión
Para cada una de las cinco temperaturas experimentales, la tasa de ingestión (TI) se
calculó de forma individual, midiendo el consumo de alimento por unidad de tiempo y
estandarizado al peso de tejido seco del organismo. Lo anterior se realizó siguiendo
la metodología y los cálculos descritos en el apartado 6.4.3. del bioensayo 1, a una
concentración de 15 x 104 células/mL de I. galbana, que resultó la óptima (ver
sección 7 de Resultados). Los organismos se alimentaron durante 1 h y
posteriormente se tomaron las muestras para la evaluación de la TI. El consumo de
alimento se estimó a partir de la diferencia en la concentración de células de la

23
cámara blanco menos la concentración de células de cada una de las cámaras
experimentales.
6.5.2. Tasa respiratoria
La tasa respiratoria (TR) se determinó mediante la concentración de oxígeno con un
oxímetro Microx TX equipado con un sensor de fibra óptica de 50 µm de diámetro, el
cual fue instalado a una celda de flujo continuo y colocado en un distribuidor donde
se conectaron las mangueras de la salida de las cámaras de incubación (Figura 5).
Figura 5. Sensor de oxígeno de fibra óptica del
Oximetro TX, colocado a la salida del distribuidor,
para determinar la concentración de oxígeno
disuelto dentro de las cámaras de incubación.
El oxímetro fue conectado a una computadora equipada con el software Oxyview
(vers. 4.16), que permitió realizar las lecturas de oxígeno disuelto y expresarlas en
unidades de mgO2/L. Los resultados se expresaron en mLO2/mg/h después de
transformar los mg de oxígeno a mL dividiendo entre 1.423 (Widdows et al. 1979). De
este modo, para el cálculo de la TR se utilizó la siguiente fórmula:

24
Donde:
O2b= Concentración de oxígeno disuelto en la cámara blanco (mL/mL)
O2c= Concentración de oxígeno disuelto en la cámara con organismo (mL/mL)
f= Flujo de agua a través de la cámara (mL/h)
Pseco= Peso del tejido seco del organismo dentro de la cámara de incubación (g)
Los resultados de la TR expresados en unidades de mL de O2/g/h fueron
finalmente convertidos a unidades energéticas, considerando que un mL de O2
consumido es igual a 20.2 J (Elliot y Davison 1975).
6.5.3. Tasa de excreción
Se estimó la tasa de excreción (TE) mediante la producción de amonio por unidad de
tiempo. Para ello, a las 2 h de haber retirado el alimento no ingerido se tomaron
muestras de 45 mL en tubos falcon, los cuales se colocaron a la salida de cada
cámara de incubación. Asimismo, después de retirar las heces, los organismos se
dejaron dentro de las cámaras de incubación con un volumen de 250 mL durante 1 h
con aireación, de la misma manera para la cámara de incubación blanco.
Posteriormente se recolectaron con tubos eppendorf muestras de agua de 2 mL, por
triplicado, del flujo de salida de las cámaras de incubación y blanco, las cuales se
preservaron a –80°C hasta su posterior análisis. Las muestras se analizaron
mediante el método Solórzano (1969); para el caso de las muestras recolectadas en
los tubos falcon, el procesamiento fue por medio de un analizador automatizado Auto
Sampler y los tubos de eppendorf se utilizó la técnica de microplaca para
cuantificación de nutrientes propuesta por Hernández-López y Vargas-Albores
(2003). El cálculo de la TE se realizó mediante la siguiente fórmula:

25
La producción de amonio se estimó por diferencia de la concentración de amonio
de la cámara de incubación de cada organismo, menos la concentración de amonio
de la cámara blanco. Los resultados de la TE fueron expresados en unidades de μg
de NH4/g/h, y posteriormente convertidas en unidades energéticas utilizando el
equivalente energético de 7.37×10-3 J/μg de NH4 (Logan y Epifanio 1978).
6.5.4. Eficiencia de absorción
La eficiencia de absorción (EA) se estimó por el método de Conover (1966), el cual
relaciona el contenido de materia orgánica (MOP) e inorgánica (MIP) particulada en
muestras de alimento y heces recolectadas de las cámaras de incubación y blanco.
Con estos datos se estimó la EA mediante la siguiente ecuación:
Donde:
F=Contenido de material orgánico del alimento entre el peso total de alimento.
E= Contenido de material orgánico de las heces entre el peso total de las heces.
El contenido de material orgánico del alimento se obtuvo de los datos de la TI
(subsección 6.5.1), mientras que el contenido de heces se obtuvo a partir de la
extracción de heces desde el fondo de cada una de las cámaras de incubación
mediante sifoneo después de un periodo de alimentación de 2 h. Posteriormente, se
filtraron las muestras mediante una bomba de vacío, se retuvieron en filtros de fibra
de vidrio de 1 μm (Whatman GF/C), se lavaron con agua destilada, se secaron
durante 24 h a 60°C y finalmente se incineraron en una mufla a 450°C para eliminar
la materia orgánica (12 h) hasta obtener un peso constante y el peso del filtro. Los
filtros con heces se lavaron con una solución de formato de amonio al 3% para

26
eliminar las sales de sodio, se secaron a 65°C para obtener un peso constante, y
también se incineraron a 450°C por 12 h para conseguir el peso de las cenizas.
6.5.5. Tasa de absorción
La tasa de absorción (TA) de los organismos se calculó mediante la siguiente
ecuación, expresada en unidades de células/g/h:
TA= Tasa de ingestión (TI) * Eficiencia de absorción (EA)
6.5.6. Determinación del peso del tejido seco del organismo
Al término del Bioensayo 2, se extrajo de cada organismo la masa visceral y se
colocó en tubos falcon de 15 mL, previamente rotulados y pesados. Cada tubo se
peso de nuevo y se almacenó a –80°C en un ultracongelador. La masa visceral de
cada organismo se introdujo en un sistema liofilizador Telstar® (Cryodos-50) por 24
h, destapando cada tubo y tapándolo con parafilm y realizando perforaciones por
medio de una aguja de disección. El peso del tejido seco (Pseco) de cada organismo
se estimó mediante la siguiente fórmula:
6.5.7. Determinación del potencial de crecimiento
Los datos de la concentración óptima de alimento a las diferentes temperaturas (16,
19, 22, 25 y 28 °C) y los valores de las diferentes variables fisiológicas (TI, TR, TE,
EA) se utilizaron para determinar el potencial de crecimiento de los organismos. Esto
implicó a su vez la integración de estas variables a la ecuación general de balance
energético propuesta por Warren y Davis (1967):
PC= TA- (TR+TE)

27
Donde:
TA = Tasa de Absorción (energía absorbida)
TR = Tasa Respiratoria (energía perdida por respiración)
TE = Tasa de Excreción (energía perdida por desechos nitrogenados)
6.6. Tratamiento estadístico de los datos
La normalidad de los datos fue determinada inicialmente con la prueba de Levene
(Sokal y Rohlf, 1981). En función de los resultados, se utilizó un análisis de varianza
de una vía para detectar diferencias significativas en las diversas variables
fisiológicas en función de la temperatura del agua (factor T con cuatro niveles). En
caso de existir diferencias significativas a una P<0.05, la homogeneidad de las
diferencias de medias se evaluó mediante comparaciones post hoc con la prueba de
rangos múltiples de Tukey, asumiendo un igual número de observaciones a cada
temperatura experimental (n=10).

28
7. RESULTADOS
7.1. Bioensayo 1: efecto de la concentración de alimento sobre la tasa de
ingestión a diferentes temperaturas
La tasa de ingestión (TI) de P. sterna varió en función de la concentración de
alimento suministrado y de las temperaturas del agua. El aumento de la
concentración de alimento de 3×104 a 25×104 células/mL en todas las temperaturas
(16, 19, 22, 25 y 28°C) favoreció también un incremento en los valores de la TI, hasta
alcanzar un valor máximo a partir del cual disminuyen. Para este caso la
concentración que provocó el punto de inflexión fue de 15×104 células/mL (Figura 7).
En todas las temperaturas del agua evaluadas en este bioensayo (Figura 6), la TI
fue menor en la concentración más baja de alimento (3×104 células/mL) cuyos
valores oscilaron de 19.33 ± 1.1×103 a 23.79 ± 1.2×103 células/g/h. Sin embargo, el
valor significativamente (P<0.05) más alto de la TI (25.94 ± 4.1×103 células/g/h) a
esta concentración (3×104 células/mL) se observó a 22°C, mientras que la
temperatura del agua de 28°C provocó el valor más bajo en la TI (Tabla II). Por su
parte, los valores de la TI registrados para la concentración de 8×104 células/mL
fueron de 48.55 ± 6.3×103 a 92.46 ± 2.1×103 células/g/h (16 y 28 °C,
respectivamente).
Para las concentraciones de alimento de 15×104, 20×104 y 25×104 células/mL,
los valores más bajos de la TI se presentaron a la temperatura de 16 °C, los cuales
corresponden a 58.48 ± 1.2×103, 47.25 ± 6.6×103 y 43.14 ± 3.2×103 células/g/h,
respectivamente. Mientras que los valores más altos de TI se observaron a 22°C
(134.82 ± 5.1×103, 119.47 ± 4.1×103 y 110.26 ± 1.2×103 células/g/h, respectivamente
para 15×104, 20×104 y 25×104 células/mL). Estas diferencias fueron significativas
(P<0.05). Los resultados del análisis de Tukey se presentan en la Tabla II y
confirman que la concentración de 15×104 células/mL arrojó valores de TI
significativamente mayores con respecto a las otras concentraciones, independiente
de la temperatura.
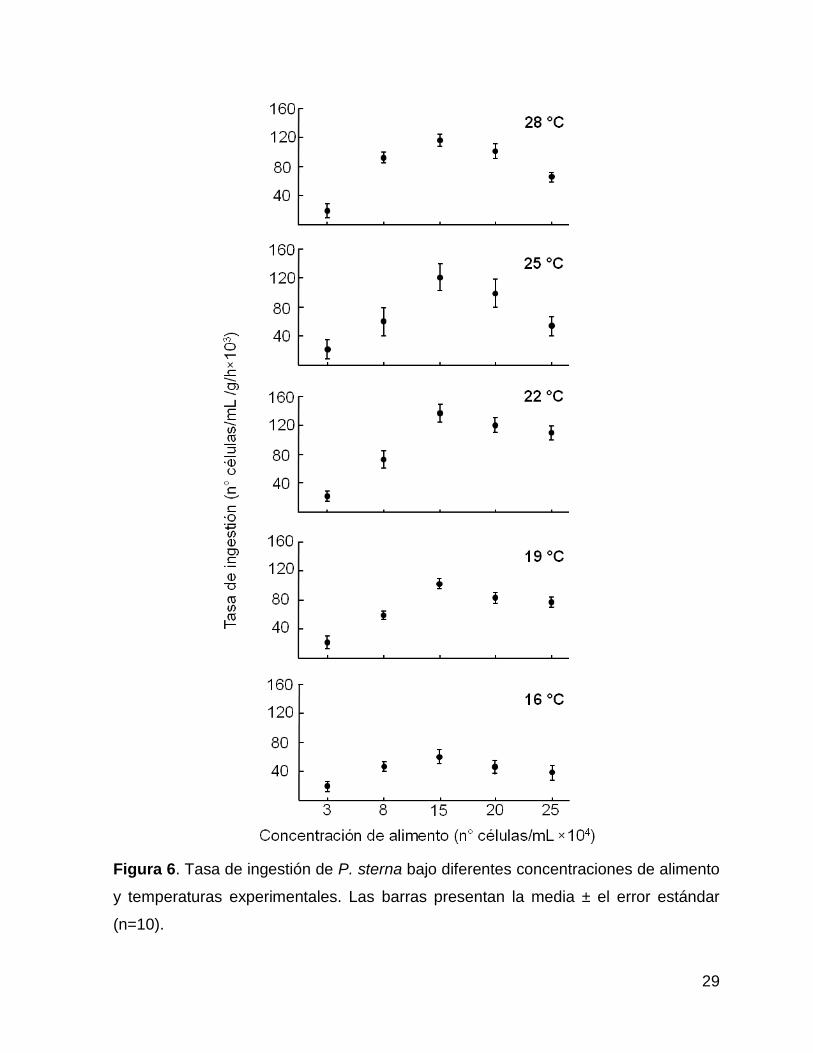
29
Figura 6. Tasa de ingestión de P. sterna bajo diferentes concentraciones de alimento
y temperaturas experimentales. Las barras presentan la media ± el error estándar
(n=10).
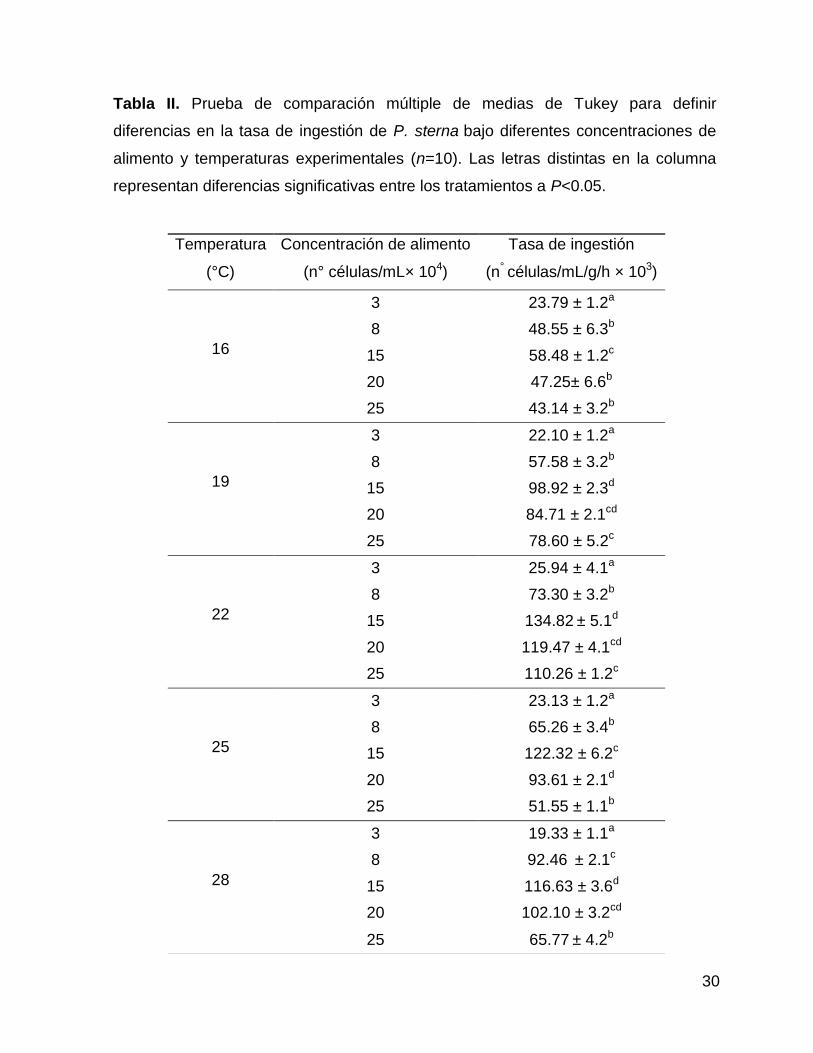
30
Tabla II. Prueba de comparación múltiple de medias de Tukey para definir
diferencias en la tasa de ingestión de P. sterna bajo diferentes concentraciones de
alimento y temperaturas experimentales (n=10). Las letras distintas en la columna
representan diferencias significativas entre los tratamientos a P<0.05.
Temperatura
(°C)
Concentración de alimento
(n° células/mL× 104)
Tasa de ingestión
(n° células/mL/g/h × 103)
16
3 23.79 ± 1.2a
8 48.55 ± 6.3b
15 58.48 ± 1.2c
20 47.25± 6.6b
25 43.14 ± 3.2b
19
3 22.10 ± 1.2a
8 57.58 ± 3.2b
15 98.92 ± 2.3d
20 84.71 ± 2.1cd
25 78.60 ± 5.2c
22
3 25.94 ± 4.1a
8 73.30 ± 3.2b
15 134.82 ± 5.1d
20 119.47 ± 4.1cd
25 110.26 ± 1.2c
25
3 23.13 ± 1.2a
8 65.26 ± 3.4b
15 122.32 ± 6.2c
20 93.61 ± 2.1d
25 51.55 ± 1.1b
28
3 19.33 ± 1.1a
8 92.46 ± 2.1c
15 116.63 ± 3.6d
20 102.10 ± 3.2cd
25 65.77 ± 4.2b
31
7.2. Bioensayo 2: efecto de la temperatura sobre las tasas fisiológicas de P.
sterna
7.2.1. Tasa de ingestión
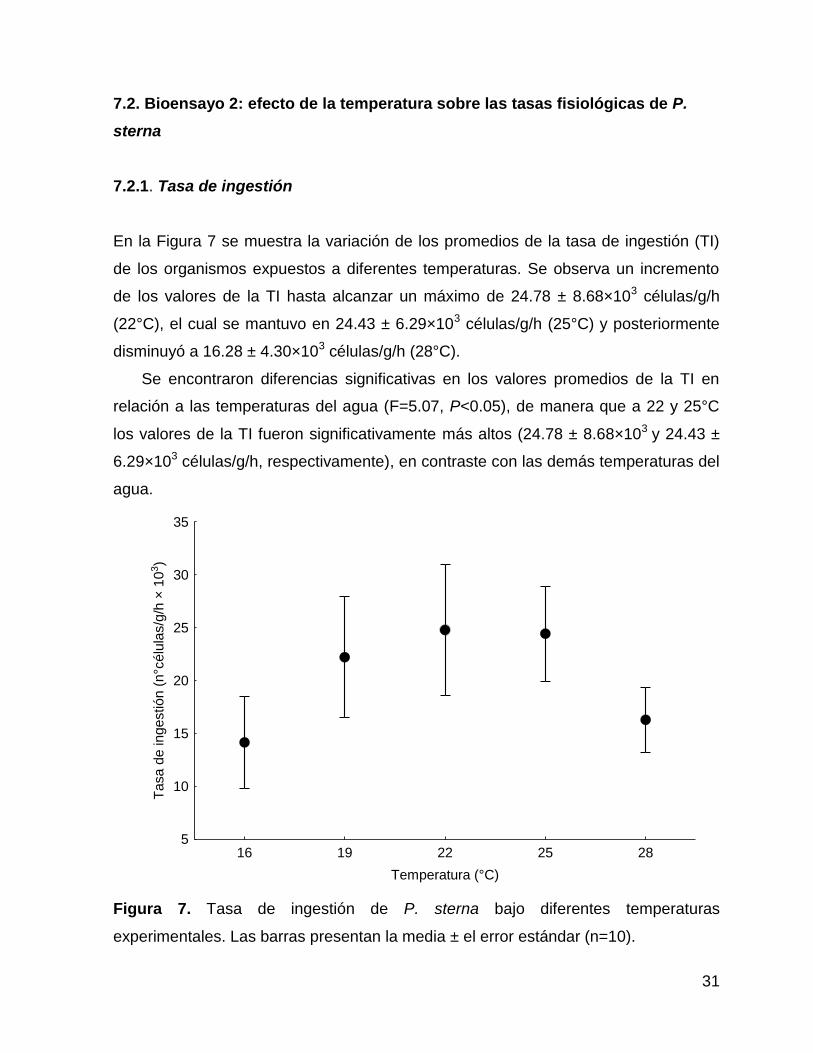
En la Figura 7 se muestra la variación de los promedios de la tasa de ingestión (TI)
de los organismos expuestos a diferentes temperaturas. Se observa un incremento
de los valores de la TI hasta alcanzar un máximo de 24.78 ± 8.68×103 células/g/h
(22°C), el cual se mantuvo en 24.43 ± 6.29×103 células/g/h (25°C) y posteriormente
disminuyó a 16.28 ± 4.30×103 células/g/h (28°C).
Se encontraron diferencias significativas en los valores promedios de la TI en
relación a las temperaturas del agua (F=5.07, P<0.05), de manera que a 22 y 25°C
los valores de la TI fueron significativamente más altos (24.78 ± 8.68×103 y 24.43 ±
6.29×103 células/g/h, respectivamente), en contraste con las demás temperaturas del
agua.
16 19 22 25 28
Temperatura (°C)
5
10
15
20
25
30
35
Ta
sa
de
inge
stió
n (
n°c
élu
las/g
/h ×
10
3)
Figura 7. Tasa de ingestión de P. sterna bajo diferentes temperaturas
experimentales. Las barras presentan la media ± el error estándar (n=10).
32
Tabla III. Prueba de comparación múltiple de medias de Tukey para definir
diferencias en la tasa de ingestión de P. sterna bajo diferentes temperaturas
experimentales (n=10). Las letras distintas en la columna representan diferencias
significativas entre los tratamientos a P<0.05.
Temperatura (°C) Tasa de ingestión
(n° células /g/h×103)
Grupo homogéneos
(P< 0.05)
16 14.14 ± 6.07 **** 28 16.28 ± 4.30 **** ****
19 22.21 ± 7.95 **** ****
25 24.43 ± 6.29 ****
22 24.78 ± 8.68 ****
7.2.2. Tasa respiratoria
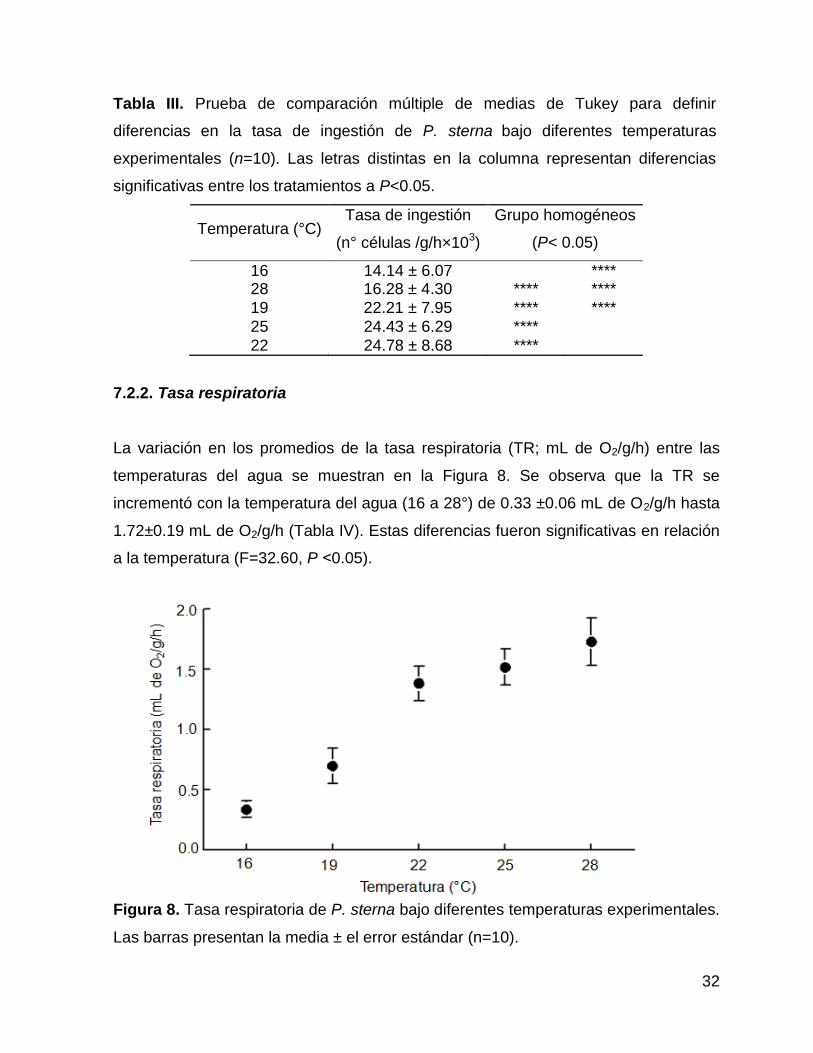
La variación en los promedios de la tasa respiratoria (TR; mL de O2/g/h) entre las
temperaturas del agua se muestran en la Figura 8. Se observa que la TR se
incrementó con la temperatura del agua (16 a 28°) de 0.33 ±0.06 mL de O2/g/h hasta
1.72±0.19 mL de O2/g/h (Tabla IV). Estas diferencias fueron significativas en relación
a la temperatura (F=32.60, P ˂0.05).
Figura 8. Tasa respiratoria de P. sterna bajo diferentes temperaturas experimentales.
Las barras presentan la media ± el error estándar (n=10).

33
Tabla IV. Prueba de comparación múltiple de medias de Tukey para definir
diferencias en la tasa respiratoria de P. sterna bajo diferentes temperaturas
experimentales (n=10). Las letras distintas en la columna representan diferencias
significativas entre los tratamientos a P<0.05.
Temperatura (°C) Tasa respiratoria
(mLO2/g/h)
Grupo homogéneos
(P< 0.05)
16 0.33 ± 0.06 **** 19 0.69 ± 1.44 ****
22 1.38± 1.44 ****
25 1.51 ± 0.15 ****
28 1.72 ± 0.19 ****
7.2.3. Tasa de excreción de amonio
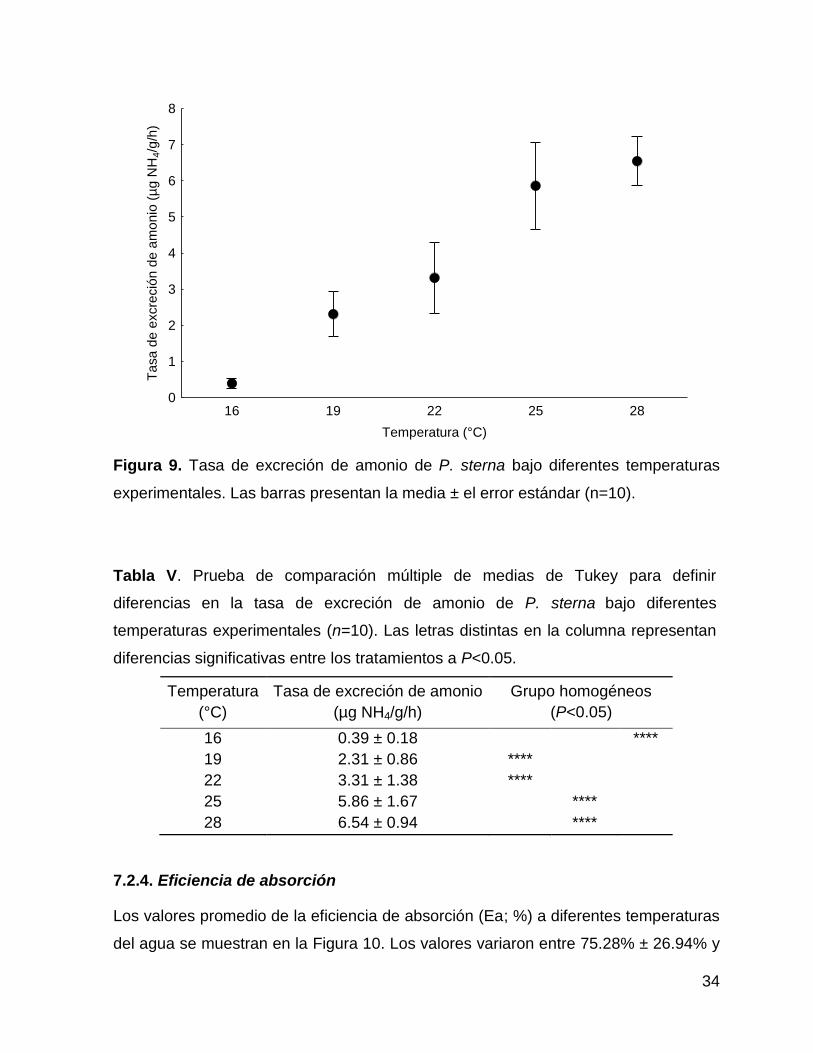
Los valores promedio de la tasa de excreción de amonio (TE; µg NH4/g/h) en relación
a las diferentes temperatura del agua (16 a 28°C) se muestran en la Figura 9. Se
observó que la TE aumentó en relación a la temperatura, registrándose el valor más
alto (6.54 ± 0.94 µg NH4/g/h) a 28°C, agrupado con la temperatura de 25 °C (5.86 ±
1.67 µg NH4/g/h). Por el contrario, el valor significativamente más bajo de la TE se
observó a los 16°C (0.39 ± 0.18 µg NH4/g/h). Existen diferencias significativas
(F=50.31, P˂0.05) en la TE a diferentes temperaturas (Tabla V).
34
16 19 22 25 28
Temperatura (°C)
0
1
2
3
4
5
6
7
8
Ta
sa
de
excre
ció
n d
e a
mon
io (
µg
NH
4/g
/h)
Figura 9. Tasa de excreción de amonio de P. sterna bajo diferentes temperaturas
experimentales. Las barras presentan la media ± el error estándar (n=10).
Tabla V. Prueba de comparación múltiple de medias de Tukey para definir
diferencias en la tasa de excreción de amonio de P. sterna bajo diferentes
temperaturas experimentales (n=10). Las letras distintas en la columna representan
diferencias significativas entre los tratamientos a P<0.05.
Temperatura
(°C)
Tasa de excreción de amonio
(µg NH4/g/h)
Grupo homogéneos
(P<0.05)
16 0.39 ± 0.18 ****
19 2.31 ± 0.86 ****
22 3.31 ± 1.38 ****
25 5.86 ± 1.67 ****
28 6.54 ± 0.94 ****
7.2.4. Eficiencia de absorción
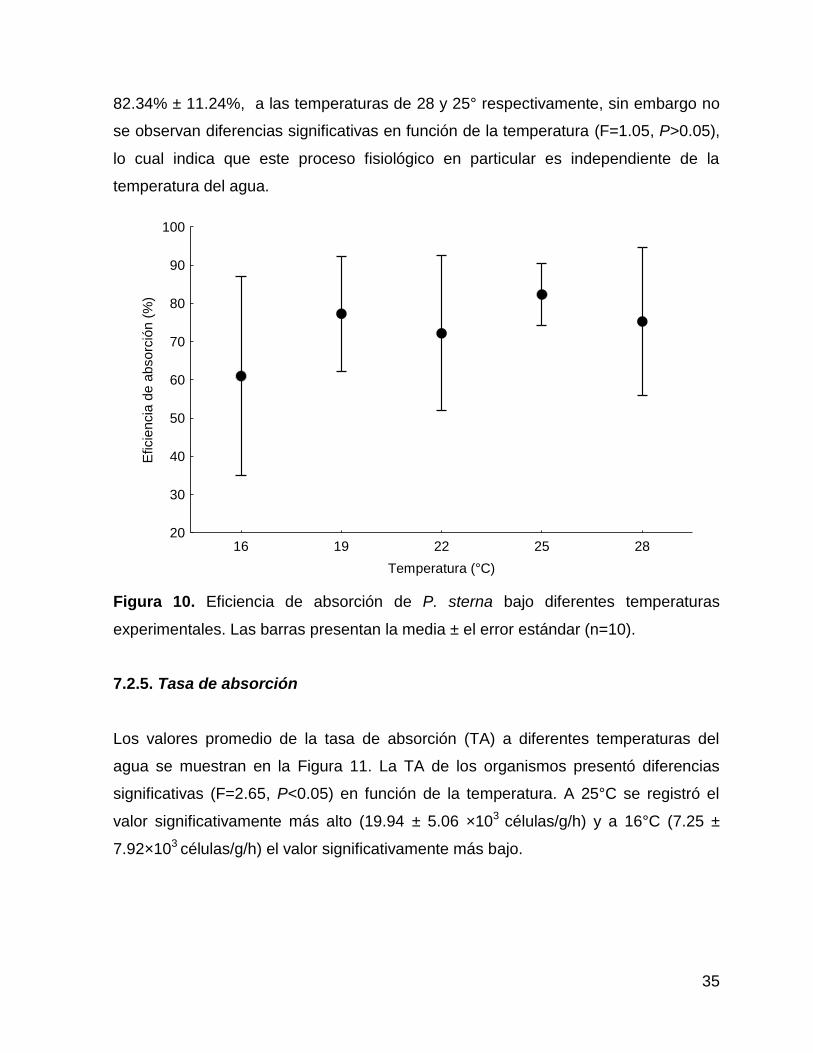
Los valores promedio de la eficiencia de absorción (Ea; %) a diferentes temperaturas
del agua se muestran en la Figura 10. Los valores variaron entre 75.28% ± 26.94% y
35
82.34% ± 11.24%, a las temperaturas de 28 y 25° respectivamente, sin embargo no
se observan diferencias significativas en función de la temperatura (F=1.05, P>0.05),
lo cual indica que este proceso fisiológico en particular es independiente de la
temperatura del agua.
16 19 22 25 28
Temperatura (°C)
20
30
40
50
60
70
80
90
100
Eficie
ncia
de
ab
so
rció
n (
%)
Figura 10. Eficiencia de absorción de P. sterna bajo diferentes temperaturas
experimentales. Las barras presentan la media ± el error estándar (n=10).
7.2.5. Tasa de absorción
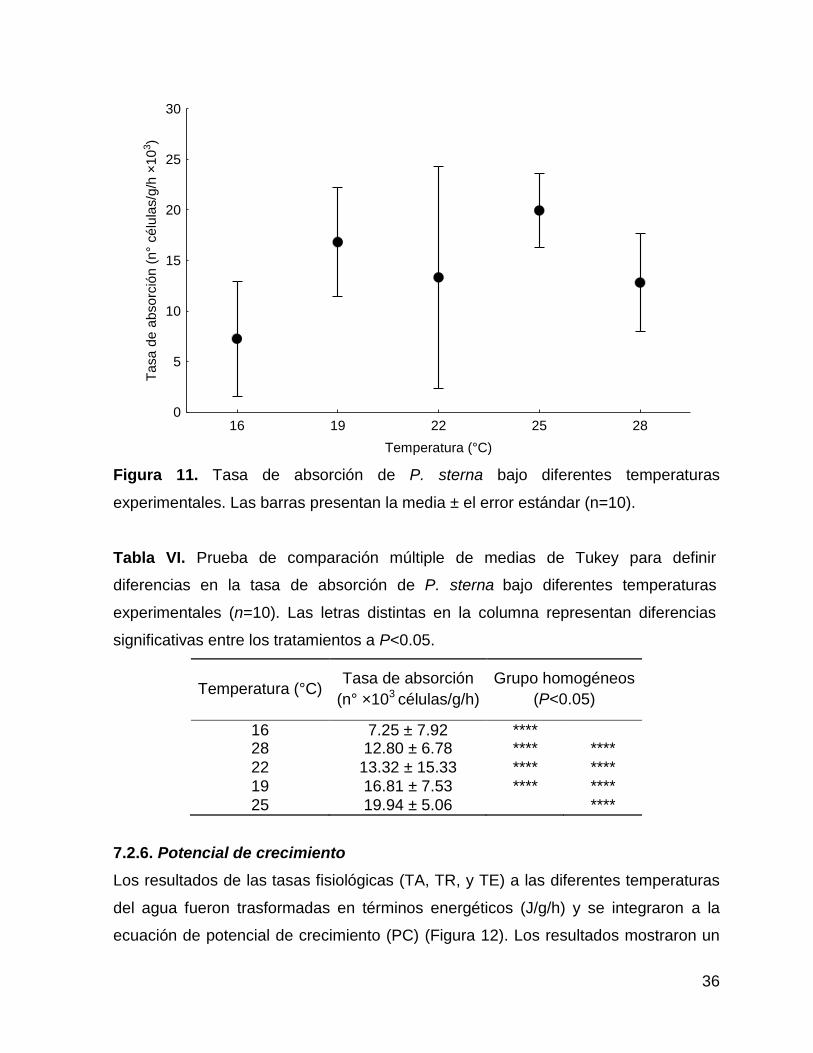
Los valores promedio de la tasa de absorción (TA) a diferentes temperaturas del
agua se muestran en la Figura 11. La TA de los organismos presentó diferencias
significativas (F=2.65, P<0.05) en función de la temperatura. A 25°C se registró el
valor significativamente más alto (19.94 ± 5.06 ×103 células/g/h) y a 16°C (7.25 ±
7.92×103 células/g/h) el valor significativamente más bajo.
36
16 19 22 25 28
Temperatura (°C)
0
5
10
15
20
25
30
Tasa
de
ab
so
rció
n (
n°
cé
lula
s/g
/h ×
10
3)
Figura 11. Tasa de absorción de P. sterna bajo diferentes temperaturas
experimentales. Las barras presentan la media ± el error estándar (n=10).
Tabla VI. Prueba de comparación múltiple de medias de Tukey para definir
diferencias en la tasa de absorción de P. sterna bajo diferentes temperaturas
experimentales (n=10). Las letras distintas en la columna representan diferencias
significativas entre los tratamientos a P<0.05.
Temperatura (°C) Tasa de absorción
(n° ×103 células/g/h)
Grupo homogéneos
(P<0.05)
16 7.25 ± 7.92 **** 28 12.80 ± 6.78 **** ****
22 13.32 ± 15.33 **** ****
19 16.81 ± 7.53 **** ****
25 19.94 ± 5.06 ****
7.2.6. Potencial de crecimiento
Los resultados de las tasas fisiológicas (TA, TR, y TE) a las diferentes temperaturas
del agua fueron trasformadas en términos energéticos (J/g/h) y se integraron a la
ecuación de potencial de crecimiento (PC) (Figura 12). Los resultados mostraron un
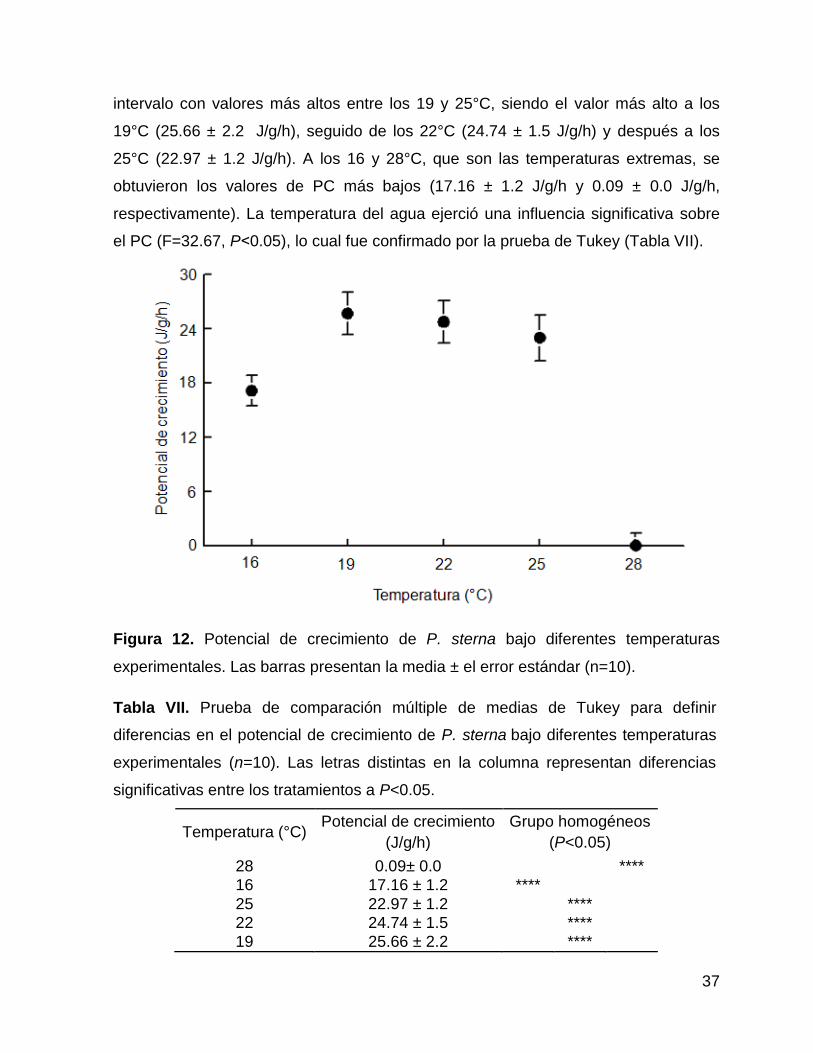
37
intervalo con valores más altos entre los 19 y 25°C, siendo el valor más alto a los
19°C (25.66 ± 2.2 J/g/h), seguido de los 22°C (24.74 ± 1.5 J/g/h) y después a los
25°C (22.97 ± 1.2 J/g/h). A los 16 y 28°C, que son las temperaturas extremas, se
obtuvieron los valores de PC más bajos (17.16 ± 1.2 J/g/h y 0.09 ± 0.0 J/g/h,
respectivamente). La temperatura del agua ejerció una influencia significativa sobre
el PC (F=32.67, P˂0.05), lo cual fue confirmado por la prueba de Tukey (Tabla VII).
Figura 12. Potencial de crecimiento de P. sterna bajo diferentes temperaturas
experimentales. Las barras presentan la media ± el error estándar (n=10).
Tabla VII. Prueba de comparación múltiple de medias de Tukey para definir
diferencias en el potencial de crecimiento de P. sterna bajo diferentes temperaturas
experimentales (n=10). Las letras distintas en la columna representan diferencias
significativas entre los tratamientos a P<0.05.
Temperatura (°C) Potencial de crecimiento
(J/g/h)
Grupo homogéneos
(P<0.05)
28 0.09± 0.0 ****
16 17.16 ± 1.2 ****
25 22.97 ± 1.2 ****
22 24.74 ± 1.5 ****
19 25.66 ± 2.2 ****

38
8. DISCUSIÓN
8.1. Bioensayo 1: efecto de la concentración de alimento sobre la tasa de
ingestión a diferentes temperaturas
Es importante mencionar que las temperaturas del agua seleccionadas en este
estudio (16 a 28°C) se encuentran dentro del intervalo de distribución natural de la
especie en la Bahía de La Paz, Baja California Sur (Mazón-Suástegui y Avilés-
Quevedo, 1988; Araya-Nuñez et al., 1991, 1995; Saucedo y Monteforte, 1997a). Esta
selección de temperatura se estableció de esta forma para identificar las
temperaturas óptimas para la fisiología de P. sterna, particularmente para controlar la
reproducción y la producción de semilla en el laboratorio (Gómez-Robles et al.,
2013).
La concentración óptima de ingestión del flagelado I. galbana en este bioensayo
fue de 15×104 células/mL, independientemente de la temperatura del agua. El patrón
observado para la TI fue de una curva de Gauss. Lo anterior es consistente con lo
propuesto por Shumway (1982) y Bayne y Newell (1983), quienes mencionan que las
tasas metabólicas (tasa de ingestión, de excreción, etc.,) de los moluscos bivalvos
normalmente se incrementan con la temperatura, hasta un máximo o límite óptimo,
más allá del cual disminuyen rápidamente. Lo anterior está relacionado con el límite
fisiológico de la especie que cuando sobrepasan un cierto tiempo, se rompe la
homeostásis y no es posible compensar. Fuera de estos límites se localizan los
niveles críticos e incluso letales de una especie (Bayne, 1976; Yukihira et al., 2000;
Saucedo et al., 2004; MacDonald et al., 2006).
Diversos estudios previos han demostrado que las tasas fisiológicas varían de
acuerdo al tipo de microalgas y/o suplemento alimenticio suministrado (Velasco,
2007). En este caso, la utilización de I. galbana como dieta monoalgal permitió una
observación precisa de las respuestas fisiológicas de P. sterna al cambio en las
concentraciones de alimento. Además, esta microalga es comúnmente utilizada en la
acuicultura para la alimentación de moluscos bivalvos durante sus tres estadios de
vida (larvas, juveniles y adultos) y con base a numerosos reportes se cataloga como

39
una de las mejores especies microalgales para cubrir los requerimientos nutricionales
de bivalvos marinos de distribución tropical y subtropical (Navarro et al., 2000;
Martínez-Fernández et al., 2004; Velasco, 2007). Esto es debido a su tamaño
adecuado (5 µm de diámetro), fácil ingestión y fácil digestión debido a la ausencia de
paredes celulares (Laing y Utting, 1980).
Se sabe que la disponibilidad de alimento es un factor importante para el
desarrollo de los bivalvos, y que afecta diversos aspectos de la fisiología,
particularmente las reservas de energía que se utilizarán para la gametogénesis. En
el presente estudio la alimentación de los organismos fue evaluada a través de la TI
(# células consumidas estandarizadas a gramos de peso de tejido seco por hora) y
se observó que fue dependiente de la concentración de alimento. La concentración a
la cual se observó mayor consumo fue cuando los organismos disponían de 15×104
células/mL, por arriba o debajo de esta concentración la ingestión se reduce. Este
patrón es consistente con el comportamiento reportado para P. margaritifera, donde
la TI aumenta con la densidad del alimento a una tasa de desaceleración hasta que
se alcanza un máximo por encima del cual la TI se mantiene constante (Le Moullac
et al., 2013). Se sabe que los bivalvos filtradores compensan activamente las
variaciones naturales en la abundancia y composición de seston cuando estas son
considerables, y para ello utilizan mecanismos fisiológicos pre-ingestivos (e. g.
producción de pseudoheces) y post ingestivos (tasa de excreción) (Ward y
Shumway, 2004).
La capacidad de ingestión de alimento reportada para diferentes especies de
bivalvos filtradores (Crassostrea madrasensis, Perna viridis y Paphia malabárica)
llega a ser notablemente diferente, lo cual suele atribuirse a variaciones en la
distribución natural de las especies y al tipo de hábitat ocupado (Rajesh et al., 2001;
Somero, 2002). Esto es particularmente importante para el caso del hábitat, pues los
cambios en los tiempos de exposición al aire o de permanencia bajo el agua (e.g.
zona intermareal, mareal, o submareal) suelen reflejar las adaptaciones de las
especies y los mecanismos que utilizan para compensar fisiológicamente. Por
ejemplo, bivalvos filtradores de hábitats bentónicos intermareales como Mulina edulis
(infauna) y Mytilus chilensis (epifauna) están sujetos a grandes fluctuaciones en la

40
cantidad y calidad del seston, principalmente debido a la exposición a la intemperie
y/o la re-suspensión de los sedimentos por acción de los vientos y mareas. A pesar
de las variaciones en la disponibilidad de alimento en suspensión, estas especies
muestran tasas de crecimiento rápido durante la mayor parte del año (Velasco y
Navarro, 2002). Para el caso de P. sterna, esta se encuentra en la zona submareal (a
profundidades mayores a 30 m), donde no existe exposición al aire o intercambio de
sedimentos por el continente, por lo que la especie presente un rango de adaptación
estrecho.
En la mayor concentración de alimento evaluada (25×104 células/mL) se observó
la producción de pseudoheces, aunque la TI fue similar a la observada a 20×104
células/mL. Esto nos permite sugerir que una concentración de 20×104 células/mL es
el máximo punto de ingestión para P. sterna antes de que ocurra el cierre de valvas y
rechazo de alimento en forma de pseudoheces, independientemente de la
temperatura del agua. Esto parece operar como una estrategia de la especie para
controlar el excedente de alimento y maximizar la ingesta.
Si bien la concentración de alimento es altamente variable, tanto en abundancia
y composición en ambientes costeros (Erga et al., 2005), para comprender la
respuesta de la TI funcional de bivalvos a la gama de condiciones de la dieta en la
naturaleza, es necesario exponer los bivalvos a una amplia gama de condiciones de
alimentación e incluir los extremos inferiores de las concentraciones de origen
natural.
8.2. Efecto de la temperatura sobre las tasas fisiológicas de P. sterna
La integración de las tasas fisiológicas en la ecuación del PC indicó que la
temperatura fisiológicamente óptima para P. sterna se encuentra entre los 19 y 22
°C, en función de la mayor disponibilidad de energía libre ofrecida a los organismos
(24.74 ± 2.2 J/g/h y 22.97 ± 1.2 J/g/h). La mayor TI (59.75 ± 6.61 J/g/h y 53.57 ± 6.06
J/g/h) y moderadas perdidas de energía en la TR (14.07 ± 0.22 J/g/h) y de excreción
(27.91 ± 1.2 J/g/h) se encontraron en este intervalo térmico. La temperatura de 28° C
ofreció poca disponibilidad de energía (0.09± 0.0 J/g/h) y promovió pérdidas de

41
energía más altas (34.87 ±1.2 J/g/h), por lo que no se consideró una temperatura
óptima para el crecimiento de P. sterna. Esto confirma que las temperaturas más
frías promueven que el metabolismo sea más eficiente y por lo tanto la energía libre
es mayor. Contrariamente, la otra especie de ostra perlera que habita en el Golfo de
California y Bahía de La Paz (P. mazatlanica), alcanza su óptimo funcionamiento
fisiológico (respiración y excreción) a una temperatura mucho más cálida, cerca de
los 28 °C (Saucedo et al., 2004). A diferencia de P. sterna, P. mazatlanica habita en
la zona intermareal, y aunque no suele estar expuesta al aire, está adaptada a
condiciones de agua oligotróficas. Esto es un patrón común a ciertas especies del
género Pinctada de otras partes del mundo (Yukihira et al., 1998a, b, 2000; Pouvreau
et al., 2000) y contrasta con los hábitos de P. sterna, que habita más en fondos
lodoso-arenosos donde las concentraciones de materia orgánica disuelta y en
suspensión son más altas (Wright-López, 1997; Saucedo y Southgate, 2008; Wright-
López et al., 2009).
Por su parte, la TI en P. margaritifera es independiente de la temperatura del
agua en un rango entre 24 y 27 °C (Chávez-Villalba et al., 2013). Este
comportamiento de alimentación se reportó también para P. margaritifera y P.
maxima entre 23°C y 32°C (Yukihira et al., 1998). Para el caso de P. sterna a una
temperatura de 16°C, se observó una TI baja, lo cual coincide con lo reportado para
una ostra perlera de distribución más templada, como P. fucata, en donde la
demanda energética, en términos de la TI, decrece a bajas temperaturas. Esto
permite que una fracción mayor de la ración disponible de alimento se convierta en
energía útil para el crecimiento, no obstante la apetencia disminuye debido a este
menor requerimiento (Mondal, 2006). Por el contrario, el efecto de las temperaturas
del agua elevadas (28 °C) sobre la TI de P. sterna fue más marcado, mostrando un
descenso que sugiere una afectación en las tasas fisiológicas. Los resultados del
presente estudio coincidieron con los reportes de Yukihira et al. (2000), quienes
observaron que el incremento de la temperatura del agua en un rango de 19 a 23°C
ocasionó también un aumento en la TI de las ostras perleras P. margaritifera y P.
maxima.

42
No se encontraron variaciones significativas de la EA en relación a la
temperatura en el rango probado, los valores encontrados fluctuaron entre 75.28 %
en la temperatura de 28°C y 82.34% en la temperatura de 25°C, sin tener un
comportamiento consistente. El efecto de la temperatura sobre este indicador ha
recibido poca atención en bivalvos marinos, aunque por lo general es relativamente
independiente de los cambios térmicos, lo que sugiere mecanismos de
compensación adecuados en la mayoría de las especies (Laing et al., 1987; Wilbur y
Hilbish, 1989). De hecho, Buxton et al. (1981) y Hutchinson y Hawkins (1992)
encontraron que un rango de temperatura entre 5 y 25 °C ejerce poco efecto sobre la
eficiencia de absorción en especies de aguas templadas como Ostrea edulis, e
incluso no cambia como es el caso de M. edulis (Widdows y Bayne, 1971). Por el
contrario, la temperatura tiene un efecto positivo en la eficiencia de absorción de
otras especies como Artica islandica y Modiolus modiolus al incrementarse en un
16% y 15% en un intervalo de 4 a 20°C, respectivamente (Winter, 1969).
Nuevamente, los mecanismos de compensación de cada especie varían en función
de su distribución geográfica en una latitud templada, subtropical o templada, donde
las variaciones en la temperatura del agua y la disponibilidad de alimento son
diferentes (MacDonald y Thompson, 2006; Freites et al., 2010). Sin embargo, en P.
margaritifera, este indicador fisiológico tampoco está influenciado por la temperatura
del agua, mientras que en P. maxima el incremento de 19 a 32 °C provocó un cambio
significativo en el mismo (Yukihira et al. 2000).
Complementando lo anterior, la TA de P. sterna fue mínima en las temperaturas
del agua extremas (16 y 28°C), presentándose el valor fisiológicamente óptimo a 22
°C (13.32 células/g/h). Los resultados obtenidos en este estudio difieren de los
encontrados en A. purpuratus y A. irradians, donde se observó que la TA se
mantiene constante a pesar de las variaciones en la temperatura del agua (Kirby-
Smith, 1970; Navarro et al., 2000). Es posible que las variaciones inter-específicas en
la eficiencia de absorción, se deban a diferencias fisiológicas compensatorias por
cambios en la temperatura del agua u otros factores ambientales.
Por su parte, la TR estuvo fuertemente influenciada por la temperatura,
obteniendo los máximos a las temperaturas más altas de 25 °C (1.51 mLO2/g/h) y 28

43
°C (1.72 mLO2/g/h). Lo anterior es un reflejo de la demanda metabólica que imponen
estas temperaturas sobre los organismos y es a su vez un indicador de estrés De
forma opuesta al comportamiento que sucede en P. mazatlanica (Saucedo et al.,
2004), la cual tolera temperaturas más altas. Así pues, es claro que cuando P. sterna
es mantenida a temperaturas bajas (16 y 19°C) reduce considerablemente su
metabolismo en comparación con las temperaturas más altas (22 a 28°C), donde la
demanda metabólica es muy alta. Estos resultados coinciden con lo reportado por
Araya-Nuñez et al. (1991, 1995) y Saucedo (2016) quienes reportan que las larvas
de P. sterna se desarrollan mejor a una temperatura entre 22 y 24 °C. Igualmente,
Mazón-Suástegui y Avilés-Quevedo (1988) resaltaron que la maduración gonádica
de esta especie promueve un desarrollo óptimo de los gametos en temperaturas
inferiores a los 25 °C.
Uemoto (1968) encontró que la TR de P. fucata martensiii se incrementa
consistentemente de 2.7 a 4.4 mLO2/g/h cuando se aumenta la temperatura del agua
de 13 a 28°C. Asimismo, Zhang et al. (2004) observaron que la TR de Chlamys
farreri se incrementaba conforme aumentaba la temperatura por encima de los 26 °C,
de la misma manera a temperaturas ascendentes de 10 hasta 31°C (Hong-Sheng et
al., 2004). Por su parte, Sugiyama y Tomori (1988) reportaron una disminución de la
TR de la misma especie a los 33°C, cuando esta fue sometida a un rango de
temperatura crecientes de 15 a 35°C, reportándose el límite superior de la especie a
35 ° C. A su vez, el aumentar la temperatura en un intervalo de 10 a 25°C incrementa
la TR de la almeja Venerupis pullastra de 0.5 a 2 mLO2/g/h (Albentosa et al., 1994).
Lo mismo ocurrió para Tapes semidecussata, en la que la TR aumentó de 0.92 a 2.0
mLO2/g/h cuando la temperatura del agua se elevó de 15 a 25 °C (Laing et al., 1987).
En A. ventricosus, ocurrió lo mismo de 1.05 a 3.05 mLO2/g/h en un intervalo de
temperatura de 12 a 28 °C (Sicard-González, 1999). Con base en lo anterior, parece
claro que los valores de TR varían mucho para cada especie de bivalvos mantenidos
a diferentes regímenes térmicos, lo que indica que cada una posee su propio umbral
de respuesta (Velasco, 2007; Pouvreau et al., 2006). Por tanto, los organismos
expuestos a temperaturas altas podrían haber experimentado una situación de estrés
que los obligó a canalizar más energía a la supervivencia, más no a otros procesos

44
fisiológicos (Sokolova et al., 2012). Dicho de otra forma, los organismos mantenidos
a 16 y 28°C operaron mayormente en metabolismo basal, no en metabolismo activo
(Bayne, 1976; Newell y Branch, 1980).
En cuanto a la TE, también se detectó un incremento proporcional con la
temperatura del agua, desde 0.39 a 6.54 µgNH4/g/h entre 16 y 28°C. En particular, la
temperatura más alta de 28 °C sugiere una situación de estrés por efecto de la
actividad metabólica del incremento en la TE, lo cual provoca pérdida de energía
asociada con la excreción de amonio. Las pérdidas de energía representadas por las
tasas respiratorias y de excreción variaron de acuerdo a las temperaturas
experimentales, por lo que las pérdidas más importantes se deben al proceso de
respiración. En el caso de P. sterna hay una tendencia hacia un mayor costo
energético entre los 25 y 28 °C. De acuerdo a Sadok et al. (1999), la reducción de la
tasa de flujo de salida de amoniaco durante ciertas condiciones ambientales es una
estrategia para la conservación de la energía como en M. edulis.
Además un alto valor de la TE puede representar la metabolización de
aminoácidos importantes, y como consecuencia se obtiene una disminución en la TA,
posiblemente debida al estrés por la poca disponibilidad de energía libre y reducción
en el metabolismo a esta temperatura (Anestis et al., 2008). Por el contrario, en P.
mazatlanica la TE fue profundamente afectada por las temperaturas (18, 23, 28 y 33
° C), pero en este caso, se observaron diferencias significativas solamente en la
temperatura más alta de (33 °C), de modo que la producción alta de amonio
excretada en 33 °C (0.09 µgNH4/g/h) fue el doble que en otras temperaturas,
revelando que en este punto, las ostras se sometieron a condiciones de estrés, pero
que no fueron capaces de compensarlo por mecanismos fisiológicos. Por el contrario,
las pequeñas cantidades de amoníaco se excretan a 23 ° C (0.02 µgNH4/g/h)
sugieren que esta temperatura óptima para la regulación de flujo de salida
metabólico (Sauedo et al., 2004).
Con base en los resultados obtenidos de este estudio, podemos concluir que P.
sterna es una especie que se beneficia más para sus procesos fisiológicos de las
temperaturas bajas del agua, encontrando un óptimo entre los 22 y 25 °C, situación
que se observó claramente reflejada en el PC.

45
Adicionalmente, es importante realizar otros estudios que impliquen el análisis de
otros parámetros fisiológicos. Por otra parte, es importante probar otros factores
importantes como oxígeno disuelto y establecer niveles críticos, salinidad para
establecer la capacidad de sobrevivir a rangos extremos de este factor para ser
cultivada en bahías hipo o hipersalinas, etc..

46
9. CONCLUSIONES
La TI fue mayor a 150,000 céls/mL e independientemente de la temperatura.
A concentraciones mayores de 200,000 céls/mL P. sterna forma pseudoheces
indicando estrés.
La TI fue significativamente menor a las temperaturas extremas de 16 y 28 °C,
mientras que la mayor TI ocurrió a los 22°C.
La TR fue mayor a 28 ° obteniendo un valor 1.72 ± 0.19 mLO2/g/h, indicando una
alta demanda metabólica a esta temperatura.
La TE aumentó conforme incremento la temperatura del agua, presentándose el
valor más alto de 23.57 ± 3.39 mgNH4/g/h a los 28°C, indicando menor eficiencia
metabólica.
La EA en P. sterna no se modifica, en relación a la temperatura dentro del rango
de 16 a 28°C.
La TA no presentó una tendencia clara debida la dispersión de los datos.
P. sterna es una especie que tiene mayor energía potencial (PC) para crecimiento
y otros procesos en el intervalo entre 19 y 25°C, fuera de este los organismos
presuntamente crecerán menos.

47
10. BIBLIOGRAFÍA
Alamo V., Valdivieso V. 1997. Lista sistemática de Moluscos Marinos del Perú.
(Segunda edición, revisada y actualizada). Publicación especial. Instituto del
Mar del Perú, Callao, Perú, 183 p.
Albentosa M. U., Labarta A., Pérez-Camacho M. J., Fernández-Reiriz, Beiras R.
1994. Fatty acid composition of Venerupis pullastra spat fed on different
microalgae diets. Comparative Biochemistry and Physiology Part A., 108:639-
648.
Ali R. M. 1970. The influence of suspension density and temperature on the filtration
rate of Hiatella arctica. Marine Biology, 6:291-302.
Allen J. A. 1962. Preliminary experiments on the feeding and excretion of bivalves
using Phaeodactylum labelled with 32P. Journal of the Marine Biological
Association of the United Kingdom, 42:609-623.
Anestis A., Pörtner H. O., Lazou A., Michaelidis B. 2008. Metabolic and molecular
stress responses of sublittoral bearded horse mussel Modiolus barbatus to
warming sea water: implications for vertical zonation. Journal of Experimental
Biology, 211:2889-2898.
Araya-Nuñez O., Ganning B., Bückle-Ramírez F. 1991. Gonad maturity, induction of
spawning, larval breeding, and growth in the American pearl-oyster (Pteria
sterna, Gould). California Fish and Game, 77:181-193.
Araya-Nuñez O., Ganning B., Bückle-Ramirez F. 1995. Embryonic development,
larval culture, and settling of American pearl-oyster (Pteria sterna, Gould)
spat. California Fish and Game, 81:10-21.
Arizmendi-Castillo E. 1996. Ciclo reproductivo de las ostras perleras, Pinctada
mazatlanica (Hanley, 1856) y Pteria sterna (Gould, 1851) (PTERIIDAE), en el
área de Guaymas, Sonora, México. Tesis de Maestría. Instituto Tecnológico
de Estudios superiores de Monterrey. Monterrey, México 140 p.
Bayne B. L. 1976. Marine Mussels: Their physiology and ecology. Cambridge
University Press, 506 p.

48
Bayne B. L., Widdows J., Thompson R. J. 1976. Physiological integrations. En: Bayne
B. L. (Ed.) Marine mussels. Cambridge University Press, Cambridge, p. 261-
299.
Bayne B. L., Newell C. 1983. Physiological energetics of marine molluscs. En: Wilbur
K. M., Saleuddin A. S. M., (Eds.) The Mollusca. Volumen 4. Physiology, Part
1. Academic Press, New York, p. 407-515.
Bayne B. L., Hawkins A. J. S., Navarro E. 1987. Feeding and digestion by the
common mussel Mytilus edulis L. (Bivalvia: Mollusca) in mixtures of salt and
algal cells at low concentrations. Journal of Experimental Marine Biology and
Ecology, 111:1-22.
Bernabé G. 1991. Acuicultura, Volumen I. Omega S.A. Barcelona España, 1100 p.
Bervera-León H. 1994. Evaluación de la captación de semilla de Pinctada
mazatlanica (Hanley 1856) en diferentes células colectoras durante el
periodo 1991-1992 y tratamiento de juveniles en la pre-engorda a partir de
1992 en la Bahía de la Paz, B. C. S., México. Tesis de licenciatura.
Universidad Autónoma de Baja California Sur. La Paz, Baja California Sur,
México 80 p.
Bougrier S., Hawkins A. J. S. Héral M. 1997. Preingestive selection of different
microalgal mixtures in Cassostrea gigas and Mytilus edulis, analysed by flow
cytometry. Aquaculture 150:123-134.
Brown J. H., Gillooly J. F., Allen A. P., Savage V. M., West G. B. 2004. Toward a
metabolic theory of ecology. Ecology, 85:1771-1789.
Buxton C. D., Newell R. C., Field J. G. 1981. Response-surface analysis of the
combined effects of exposure and acclimation temperatures on filtration,
oxygen consumption and scope for growth in the oyster Ostrea edulis. Marine
Ecology Progress Series, 6:73-82.
Bückle-Ramírez L. F., Voltolina-Lobina D., Morales-Guerrero E., Valenzuela-Buriel F.
1992. Spat settlement and growth of Pteria sterna (Gould) (Mollusca,
Bivalvia) in Bahía de Los Ángeles, Baja California, Mexico. Journal of
Tropical Ecology, 33:137-147.

49
Bühringer H., Danischewski D. 2001. Laboratory studies on the scope for growth in
blue mussels, Mytilus edulis L. Archive of Fishery and Marine Research,
49:61-68.
Cáceres-Martínez C., Cortés-Salazar J., Chávez-Villalba J. 1991. Reclutamiento de
jóvenes de moluscos bivalvos en Baja California Sur, México. En: Guzmán
del Proo S. (Ed.) Taller México-Australia sobre Reclutamiento de Recursos
Bentónicos de Baja California, 25-29 de Noviembre 1991, La Paz, B.C.S.,
México. SEPESCA-IPN, p 151-165.
Cáceres-Martínez C., Ruiz-Verdugo C. A., Ramírez-Filippini D. H. 1992. Experimental
collection of pearl oyster, Pinctada mazatlanica and Pteria sterna, spat on a
filament substrate. Journal of the World Aquaculture Society, 23:232-240.
Cáceres-Puig J. I., Cáceres-Martínez C., Saucedo P. E. 2009. Annual reproductive
effort of pacific winged pearl oyster Pteria sterna and its relation with the
timing for planning pearl seeding operations. Journal of Shellfish Research,
28:471-476.
Cáceres-Puig J. I. 2012. Dinámica del reclutamiento de semilla de la concha nácar
Pteria sterna en la Bahía de La Paz Baja California Sur, México. Tesis de
Doctorado, Centro de Investigaciones Biológicas de Noroeste, México 131 p.
Chase J. M., Leibold M. A. 2003. Ecological niches: Linking classical and
contemporary approaches: Interspecific interactions. University of Chicago
Press. Chicago, Illinois, 221 p.
Chávez-Villalba J., Soyez C., Aurentz H., Le Mollac G. 2013. Physiological responses
of female and male black-lip pearl oysters (Pinctada margaritifera) to different
temperatures and concentrations of food. Aquatic Living Resource, 26:263-
271.
Dame R. F. 2011. Ecology of marine bivalves: An ecosystem approach. 2 ed. CRC
Press, 283 p.
Davids C. 1964. The influence of suspensions of microorganisms of different
concentrations on the pumping and retention of food by the mussel (Mytilus
edulis L.). Netherlands Journal of Sea Research, 2:233-249.

50
Del Río-Portilla M. A., Re-Araujo A. D., Voltolina D. 1992. Growth of the pearl oyster
Pteria sterna under different thermic and feeding conditions. Marine Ecology
Progress Series, 89:221-227.
Elvin D. W., Gonor J. J. 1979. The thermal regime of an intertidal Mytilus californianus
Conrad population on the central Oregon coast. Journal of Experimental
Marine Biology and Ecology, 39:265-279.
Epifanio C. E. 1979. Comparison of yeast as foods for juvenile bivalve molluscs.
Aquaculture, 16:187-192.
Erga S. R., Aursland K., Frette O., Hamre B., Lotsberg J. K., Stamnes J. J., Aure J.,
Rey F., Stamnes K. 2005. UV transmission in Norwegian marine waters:
controlling factors and possible effects on primary production and vertical
distribution of phytoplankton. Marine Ecology Progress Series, 305:79-100.
Espina S., Díaz F., Rosas C., Rosas I. 1986. Influencia del detergente sobre el
balance energético de Ctenopharyngodon idella a través de un bioensayo
crónico. Contaminación Ambiental, 2:25-37.
Ezgeta-Balic D., Rinaldi A., Peharda M., Prusina I., Montalto V., Niceta N., Sarà G.
2011. An energy budget for subtidal bivalve Modiolus barbatus (Mollusca) at
different temperatures. Marine Environmental Research, 71:79-85.
Freites L., Montero L., Arrieche D., Babarro J. M. F., Saucedo P. E., Córdova C.,
García N. 2010. Influence of environmental factors on the reproductive cycle
of the eared ark Anadara notabilis (Röding, 1798) in Northeastern Venezuela.
Journal of Shellfish Research, 29: 69-75.
Gabbot P. A. 1975. Storage cycles in marine bivalve molluscs: a hypothesis
concerning the relationship between glycogen metabolism and
gametogenesis. En: Barnes H. (Ed.) Proc. 9th European Marine Biology
Symposium, Aberdeen University Press, Aberdeen, UK, p. 191-211.
García-Esquivel Z., Parés-Sierra G., García-Pámanes L. 2000. Efecto de la velocidad
de flujo y concentración de alimento sobre el crecimiento de juveniles de la
escalopa Nodipecten subnodosus. Ciencias Marinas, 26:621-641.
Gaytán-Mondragón I., Cáceres-Martínez C., Tobias-Sánchez M. 1992. Growth of the
pearl oysters Pinctada mazatlanica and Pteria sterna in different culture

51
structures at La Paz Bay, Baja California Sur, México. Journal of the World
Aquaculture Society, 24:541-546.
Gervis M. H., Sims N. A. 1992. The biology and culture of pearl oysters (Bivalvia:
Pteriidae). International Center for Living Aquatic resources, Manila,
Philippines, 49 p.
Gillooly J. F., Charnov E. L., West G. B., Savage V. M., Brown J. H. 2001. Effects of
size and temperature on developmental time. Nature, 417:70-73.
Gómez-Robles E., Mazón-Suástegui J. M., Acosta-Salmón H., Hawkyns-Martínez M.,
Saucedo P. E. 2013. Internal nutrient management associated with gonad
quality and successful reproduction in the winged pearl oyster Pteria sterna.
Aquaculture, 412-413:45-51.
Granados-Amores A. 2012. Efecto de la temperatura en el desarrollo y composición
gonadal de la concha nácar Pteria sterna (Gould 1951) en condiciones
controladas de laboratorio. Tesis de Maestría. Centro Interdisciplinario de
Ciencias Marinas, México, 99 p.
Griffiths C. L., Griffiths R. J. 1987. Bivalvia. En: Pandian T. J., Vernberg F. J. (Eds.)
Animal energetics. Volume 2. Academic Press, New York, p. 1-88.
Hawkins A. J. S., Smith R. F. M., Bougrier S., Bayne B. L., Héral M. 1997.
Manipulation of dietary conditions for maximal growth in mussels, Mytilus
edulis L., from the Marennes-Oléron Bay, France. Aquatic Living Resources,
10:13-22.
Heasman M. P., O’ Connor W. A., Allen W. F. 1996. Temperature and nutrition as
factors in conditioning broodstock of the commercial scallop Pecten fumatus
Reeve. Aquaculture, 143:75-90.
Helm M. M., Bourne N., Lovatelli A. 2004. Hatchery culture of bivalves. A practical
manual. Food and Agriculture Organization (FAO) of the United Nations,
Rome, 177 p.
Helmuth B. 2009. From cells to coastlines: how can we use physiology to forecast the
impacts of climate change. Journal of Experimental Biology, 212:753-760.
Helmuth B., Broitman B. R., Yamane L., Gilman S. E., Mach K., Mislan K. A. S.,
Denny M. W. 2010. Organismal climatology: analyzing environmental

52
variability at scales relevant to physiological stress. Journal of Experimental
Biology, 213:995-1003.
Hernández-Díaz A., Bückle-Ramírez. 1996 Ciclo gonádico de Pteria sterna (Gould,
1852) (Mollusca:Bivalvia) en Baja California, México. Ciencias Marinas,
22:495-509.
Hernández-López C. G. 2012. Efecto de la dieta en el desarrollo y composición de la
gónada y tejidos somáticos relacionados con la reproducción de la concha
nácar Pteria sterna (Gould, 1851). Tesis de Maestría, Centro de
Investigaciones Biológicas del Noroeste, México 93 p.
Hernández-Olalde L., García-Domínguez F., Arellano-Martínez M., B. P. Ceballos-
Vázquez. 2007. Reproductive cycle of the Pearl oyster Pteria sterna
(Pteriidae) in the Ojo de Liebre Lagoon, B.C. S., Mexico. Journal Shellfish
Research, 26:543-548.
Hochachka P. W., Somero G. N. 2002. Biochemical Adaptation: Mechanism and
process in physiological evolution. Oxford University Press, 480 p.
Hutchinson S., Hawkins L. E. 1992. Quantification of the physiological responses of
the European flat oyster Ostrea edulis L. to temperature and salinity. Journal
of Molluscan Studies, 58:215-226.
Ibarrola I., Navarro E., Iglesias J. I. P. 1998. Short-term adaptation of digestive
processes in the cockle Cerastoderma edule exposed to different food
quantity and quality. Journal of Comparative Physiology B, 168:32-40.
Kearney M., Stephen J. S., Raubenheimer D., Helmuth B. 2010. Modelling the
ecological niche from functional traits. Philosophical Transactions of the
Royal Society B, 365:3469-3483.
Keen M. 1971. Sea shells of tropical west America. Marine mollusks from Baja
California to Perú. 2a ed. Stanfrod University Press. USA, 1064 p.
Kesarcodi-Watson A., Klumpp D. W., Lucas J. S. 2001. Comparative feeding and
physiological energetics in diploid and triploid Sydney rock oysters
(Saccostrea commercialis): II. Influences of food concentration and tissue
energy distribution. Aquaculture, 203:195-216.

53
Kiefert L., McLaurin-Moreno D., Arizmendi-Castillo E., Hänni H. A., Elen S. 2004.
Cultured pearls from the Gulf of California, Mexico. Gems Gemology, 40:26-
38.
Kirby-Smith W. W. 1970. Growth of the scallops Argopecten irradians concentricus
(Say) and Argopecten gibbus (Linne), as influenced by food and
temperatures. Ph. D. Thesis, Duke University, Durham, North Carolina 126 p.
Kobayashi S., Tobata M. 1949. Studies on culture of pearl: II. Activity of pearl oyster
in winter. Bulletin of the Japanese Society for the Science of Fish, 14:196-
199.
Kooijman S. A. L. M. 2010. Dynamic energy budget theory for metabolic organisation.
3era. Cambridge University Press, Great Britain, 72 p.
Laing I., Utting S. D. 1980. The influence of salinity on the production of two
commercially important unicellular marine algae. Aquaculture, 21:79-86.
Laing I., Utting S. D., Kilada R. W. S. 1987. Interactive effect of diet and temperature
on the growth of juvenile clams. Journal of Experimental Marine Biology and
Ecology, 113:23-38.
Lamghari M., Almeida M. J., Berland S., Huet H., Laurent A., Milet C., Lopez E. 1999.
Stimulation of bone marrow cells and bone formation by nacre: In vivo and In
vitro studies. Bone, 25:91-94.
Le Moullac G., Soyez C., Sham-Koua M., Levy P., Moriceau J., Vonau V., Maihota
M,, Cochard J. C. 2013. Feeding the pearl oyster Pinctada margaritifera
during reproductive conditioning. Aquaculture Research, 44:404-411.
Maeda-Martínez A. N., Saucedo P. E., Mazón-Suástegui M., Acosta-Salmón H.,
Romero-Meléndez Z. 2016. Scope for growth of juvenile Cortez oyster,
Crassostrea corteziensis fed isocaloric diets prepared with microalgae and
cereal flours. Latin American Journal of Aquatic Research, 44:845-849.
MacDonald B. A., Bricelj M., Shumway S. E. 2006. Physiology: Energy acquisition
and utilization. En: Shumway S. E., Parsons G. J. (Eds.) Scallops: biology,
ecology and aquaculture. Elsevier, Amsterdam, p. 417-492.

54
Martínez-Fernández E., Acosta-Salmón H., Rangel-Dávalos C. 2004. Ingestion and
digestion of 10 species of microalgae by winged pearl oyster Pteria sterna
(Gould, 1851) larvae. Aquaculture, 230:417-423.
Mazón-Suástegui J. M., Avilés-Quevedo M. A. 1988. Ensayo preliminar sobre la
alimentación de bivalvos juveniles con dietas artificiales. Revista
Latinoamericana de Acuicultura, 36:56-62.
Mondal S. K. 2006. Effect of temperature and body size on food utilization in the
marine pearl oyster Pinctada fucata (Bivalvia: Pteriidae). Indian Journal of
Marine Sciences, 35:43-49.
Monteforte M., Aldana A. 1994. Spat collection, growth and survival of pearl oyster
Pteria sterna under extensive culture conditions in Bahía de La Paz, South
Baja California, México. Abstracts, Pearls’94 International Pearl Conference,
Honolulu, USA. Journal of Shellfish Research, 13:21-34.
Monteforte M., García-Gasca A. 1994. Spat collection studies on pearl oysters
Pinctada mazatlanica and Pteria sterna (Bivalvia: Pteriidae) in Bahía de La
Paz, South Baja California, Mexico. Hydrobiologia, 291:21-34.
Monteforte M., Wright H. 1994. Ecology of pearl oyster spat collection in Bahía de La
Paz: South Baja California, México: temporal and vertical distribution,
substrate selection, associated species. Abstracts, Pearls’94 International
Pearl Conference, Honolulu, USA. Journal of Shellfish Research, 13:342-343.
Monteforte M., Kappelman-Piña E., López-Espinosa B. 1995. Spatfall of pearl oyster,
Pteria sterna (Gould), on experimental collectors at Bahía de La Paz, South
Baja California, Mexico. Aquaculture Research, 26:497-511.
Monteforte M. 2013. Instalación y operación de granjas perleras: Un manual para
Pinctada mazatlanica y Pteria sterna. Académica Española, Barcelona,
España, 184 p.
Monteforte M., Cariño M. 2013. Condiciones para el desarrollo de granjas perleras y
producción de perlas: estado del arte, potencial y perspectiva. Revista de
Biología Marina y Oceanografía, 48:1-16.
Monteforte M., Bervera H., Ramírez J. J., Saucedo P. E., López C. O. 2005. Effect of
stocking density on growth and survival of the rainbow pearl oyster Pteria

55
sterna (Gould 1852) during nursery and late culture in Bahía de La Paz, Baja
California Sur, México. Aquaculture International, 13:391-407.
Miyauchi T. 1962. Clearance rate of the pearl oyster Pinctada fucata martensii. II.
Effects of temperature and salinity. Suisan Zoshoku, 10:73.
McAnally-Salas L., Valenzuela-Espinoza E. 1990. Crecimiento y supervivencia de las
larvas de la ostra concha nácar Pteria sterna en condiciones de laboratorio.
Ciencias Marinas, 16:29-41.
McLaurin D., Arizmendi E., Farell S., Nava M. 1999. Pearls and pearl oysters from the
Gulf of California, México, an update. The Australian Gemmologist, 20:239-
245.
Navarro J. M., Thompson R. J. 1996. Physiological energetics of the horse mussel
Modiolus modiolus in a cold ocean environment. Marine Ecology Progress
Series, 138:135-148.
Navarro J. M., Widdows J. 1997. Feeding physiology of Cerastoderma edule in
response to a wide range of seston concentrations. Marine Ecology Progress
Series, 152:175-186.
Navarro J. M., Leiva G. E., Martínez G., Aguilera C. 2000. Interactive effects of diet
and temperature on the scope for growth of the scallop Argopecten
purpuratus during reproductive conditioning. Journal of Experimental Marine
Biology and Ecology, 247:67-83.
Newell R. C. 1979. Biology of intertidal animals. Marine Ecology Surveys Ltd.,
Faveresham, England, 791 p.
Newell R. C., Bayne B. L. 1973. A review on temperature and metabolic acclimation
in intertidal marine invertebrates. Netherlands Journal of Sea Research,
7:421-433.
Newell R. C., Branch G. M. 1980. The effects of temperature on the maintenance of
metabolic energy balance in marine invertebrates. Advances in Marine
Biology, 17:329-396.
Newell R. I. E. 2004. Ecosystem influence of natural and cultivated populations of
suspension-feeding bivalve molluscs: a review. Journal of Sellfish Research,
23:51-61.

56
Numaguchi K., Tanaka Y. 1986. Effects of temperature on mortality and growth of the
spat of the pearl oyster Pinctada fucata martensii. Bulletin of National
Research Institute of Aquaculture, 9:35-39.
Numaguchi K. 1994. Effect of water temperature on the filtration rate of Japanese
pearl oyster Pinctada fucata martensii. Suisan Zoshoku, 41:1-6.
Peck L. S. 2002. Ecophysiology of Antarctic marine ectotherms: limits to life. Polar
Biology, 25:31:40.
Pereira-Mouriès L., Almeida M. J., Ribeiro C., Peduzzi J., Barthélemy M., Milet C.,
Lopez E. 2002. Soluble silk-like organic matrix in the nacreous layer of the
bivalve Pinctada maxima. European Journal of Biochemistry, 269:4994-5003.
Pérez-Estrada C. J. 2000. Descripción anatómica e histológica de la concha nácar
Pteria sterna Mollusca: Pteriidae. Tesis de Licenciatura. Universidad
Autónoma de Baja California Sur. La Paz, Baja California Sur, México 80 p.
Pérez-Camacho A., Labarta U., Navarro E. 2000. Energy balance of mussels Mytilus
gallopronvicialis the effect of length and age. Marine Ecology Progress
Series, 199:149-158.
Poutiers J. M. 1995. Bivalvos (Acephala, Lamellibranchia, Pelecypoda) En: Flescher
W., Krupp F., Schneider W., Sommer C., Carpenter K. E., Niem V. H. (Eds)
Guía FAO para la identificación de especies para los fines de la pesca.
Pacífico Centro-Oriental. Vol. I. Plantas e Invertebrados. Roma, p 99-222.
Pouvreau S., Bodoy A., Buestel D. 2000. In situ suspension feeding behavior of the
pearl oyster, Pinctada margaritifera: combined effects of body size and
weather-related seston composition. Aquaculture, 181:91-113.
Pouvreau S., Bourles Y., Lefebvre S., Gangnery A., Alunno-Bruscia M. 2006.
Application of a dynamic energy budget model to the Pacific oyster,
Crassostrea gigas, reared under various environmental conditions. Journal of
Sea Research, 56:156-167.
Precht H., Laudien H., Havsteen B. 1973. The normal temperature range. En: Precht
H., Cristophersen J., Hensel H., Larcher W. (Eds.) Temperature and life,
Springer, New York, NY, USA p. 302-325.

57
Prosser C. L. 1973. Comparative animal physiology. 3era. W. B. Saunders Co.,
Philadelphia, London, Toronto, 966 p.
Rajesh K. V., Mohamed K. S., Kripa V. 2001. Influence of algal cell concentration,
salinity and body size on the filtration and ingestion rate of cultivable Indian
bivalves. Indian Journal of Marine Sciences, 30:87-92.
Rangel C., Chávez J. 1994. Desarrollo científico y tecnológico del cultivo de la madre
perla y concha nácar. Secretaría de Pesca, Dirección General de
Acuacultura. Convenio SEPESCA/UABCS, México. 104 p.
Rangel-Dávalos C., Cáceres-Martínez C. 1994. Pearl oyster culture in México. En:
Abstracts of the International Pearl Conference and Exposition, Pearls’94.
Journal of the Shellfish Research, 13:47-348.
Riisgard H. U. 2001. Physiological regulation versus autonomous filtration in filter-
feeding bivalves: Starting points for progress. Ophelia, 54:193-209.
Riisgard H. U., Larsen P. S. 2000. Comparative ecophysiology of active zoobenthic
filter feeding essence of current knowledge. Journal of Sea Research,
44:169-193.
Rueda J. L., Smaal A. C. 2004. Variation of the physiological energetics of the bivalve
Spisula subtruncata (da Costa, 1778) within an annual cycle. Journal of
Experimental Marine Biology and Ecology, 301:141-157.
Ruiz-Rubio H., Acosta-Salmón H., Olivera A., Southgate P. C., Rangel-Dávalos C.
2006. The influence of culture method and culture period on quality of half-
pearls (‘mabe’) from the winged pearl oyster Pteria sterna, Gould, 1851.
Aquaculture, 254: 269-274.
Sadok S., Uglow R. F., Haswell S. J., 1999. Some aspects of nitrogen metabolism in
Mytilus edulis: effects of aerial exposure. Marine Biology, 135: 297-305.
Sarà G., Pusceddu A. 2008. Scope for growth of Mytilus galloprovincialis (Lmk.,
1819) in oligotrophic coastal waters (Southern Tryrrhenian Sea, Italy). Marine
Biology, 156:117-126.
Sarà G., Romano C., Widdows J., Staff F. J. 2008. Effect of salinity and temperature
on feeding physiology and scope for growth of an invasive species

58
(Brachidontes pharaonis - MOLLUSCA: BIVALVIA) within the Mediterranean
sea. Journal of Experimental Marine Biology and Ecology, 363:130-136.
Sarà G., Kearney M., Helmuth B. 2011. Combining heat-transfer and energy budget
models to predict thermal stress in Mediterranean intertidal mussels.
Chemistry and Ecology, 27:135-145.
Saucedo P. E. 1991. Ensayo sobre repoblamiento de bancos naturales de concha
nácar Pteria sterna y madre perla Pinctada mazatlanica (Bivalvia:Pteriidae)
en El Merito, Bahía de La Paz, Sudcalifornia, México. Tesis de Licenciatura,
Universidad Autónoma de México, 75 p.
Saucedo P. E. 1995. Crecimiento, relaciones alométricas y reproducción de las
ostras perleras Pinctada mazatlanica y Pteria sterna (Bivalvia: Pteriidae) bajo
condiciones de repoblamiento en el Merito, Bahía de la Paz, Baja California
Sur, México. Tesis de Maestría. Centro Interdisciplinario de Ciencias Marinas
de IPN, La Paz, Baja California Sur, México 100 p.
Saucedo P. E., Monteforte M. 1997a. Breeding cycle of pearl oysters Pinctada
mazatlanica and Pteria sterna in Bahia La Paz, South Baja California,
Mexico. Journal of Shellfish Research, 16:348-349.
Saucedo P. E., Monteforte M. 1997b. In situ growth of pearl oyster Pinctada
mazatlanica (Hanley 1856) and Pteria sterna (Gould 1851) under
repopulation conditions at Bahía de La Paz, Baja California Sur, Mexico.
Aquaculture Research. 28:367-368.
Saucedo P. E., Ocampo-Victoria L., Bervera-León H., Monteforte M. 2004. Effect of
temperature on oxygen consumption and ammonia excretion in the Calaifa
mother-of-pearl oyster Pinctada mazatlanica (Hanley, 1856). Aquaculture,
229: 377-387.
Saucedo P. E., Southgate P. C. 2008. Reproduction, development, and growth. En:
Southgate P. C., Lucas J. S. (Eds.) The pearl oyster: biology and culture.
Elsevier Science, Oxford, United Kingdom, p. 129-184.
Saucedo P. E., Castillo-Domínguez A., Melgar-Valdés C. 2015. Aprovechamiento de
almejas dulceacuícolas en Tabasco para formación de perlas y artesanías.

59
Colección José N. Rovirosa, Universidad Juárez Autónoma de Tabasco,
Villahermosa, México, 133 p.
Saucedo P. E. 2016. Advances in hatchery production of the winged pearl oyster
Pteria sterna. World Aquaculture Magazine, (en prensa).
Segal E. 1961. Acclimation in molluscs. American Zoologist, 235-244.
Serrano-Guzmán S. J., Salina-Ordáz D. 1993. Cultivo de larvas y producción de
semilla de Pteria sterna (Mollusca:Bivalvia) en un criadero comercial. Revista
de Investigación Científica, 4:81-90.
Seki M. 1972. Studies on natural factors affecting the growth and pearl quality of
Japanese pearl oyster Pinctada fucata in culture environments. Bulletin Mie
Pref Fish Res, 32-149.
Sevilla M. L. 1969. Contribución al conocimiento de la madre perla Pinctada
mazatlanica (Hanley 1845). Revista de la Sociedad Mexicana de Historia
Natural, 30:223-262.
Sicard-González M. T. 1999. Temperatura letal superior y temperatura optima en una
población de almeja catarina (Argopecten ventricosus Sowerby II, 1842).
Tesis de Maestría. Centro Interdisciplinario de Ciencias Marinas, La Paz,
Baja California Sur, México 94 p.
Singh C. J., Bojórquez V. G., Meza D. J. M. 1982. Resultados finales de las
actividades de estudios de ostras perleras en la Bahía de La Paz, B. C. S.
durante los años 1981-1982. Delegación Federal de Pesca en el Estado de
Baja California Sur. Secretaría de Pesca 50 p.
Somero G. N. 2002. Thermal physiology and vertical zonation of intertidal animals:
optima, limits, and costs of living. Integrative Comparative Biology, 42:780-
789.
Soria G., Merino G., Von Brand E. 2007. Effect of increasing salinity on physiological
response in juvenile scallops Argopecten purpuratus at two rearing
temperatures. Aquaculture, 270:451-463.
Sokolova I. M., Frederich M., Bagwe R., Lanning G., Sukhotin A. A. 2012. Energy
homoeostasis as an integrative tool for assessing limits of environmental

60
stress tolerance in aquatic invertebrates. Marine Environmental Research,
79:1-15.
Sugiyama A., Tomori A. 1988. Oxygen consumption of black-lip pearl oyster. Suisan
Zoshoku, 36:121-125.
Sukhotin A. A., Lajus D. L., Lesin P. A. 2003. Influence of age and size on pumping
activity and stress resistance in the marine bivalve Mytilus edulis L. Journal of
Experimental Marine Biology and Ecology, 284:129-144.
Scarlet M. P. J., Halldórsson H. P., Granmo A. 2015. Scope for growth and condition
index in the clam Meretrix meretrix (L.) as biomarkers of pollution in Espírito
Santo Estuary, Mozambique. Regional Studies in Marine Science, 63-71.
Schoener T. W. 1986. Mechanistic approaches to community ecology: a new
reductionism? American Zoologist, 26:81-106.
Schulte E. H. 1975. Influence of algal concentration and temperature on the filtration
rate of Mytilus edulis. Marine Biology, 30:331-341.Thompson R. J.,
MacDonald B. A. 2006. Physiological integration and energy partitioning. En:
Shumway E., Parsons G. J. Scallops: Biology, Ecology and Aquaculture.
Volume 35, Developments in Aquaculture and Fisheries, p. 493-520.
Shirai S., Sano Y. 1972. Reporte de una investigación sobre los recursos perleros del
Golfo de California. Reporte Interno. SEPESCA. México 55 p.
Shumway S. E. 1982. Oxygen consumption in brachiopods and the possible role of
punctae. Journal of Experimental Marine Biology and Ecology, 58:207-220.
Spicer J. I. 2014. What can an ecophysiological approach tell us about the
physiological responses of marine invertebrates to hypoxia? Journal of
Experimental Biology, 217:46-56.
Torres-Zepeda M. G., Morales-Salvador J., Peralta-Montes L. 2002. Evaluación de la
captación estacional y batimétrica de semilla de la madreperla Pinctada
mazatlanica (Hanley 1856) y la concha nácar Pteria sterna (Gould 1851) en
colectores artificiales en la Bahía de Acapulco (Guerrero, México). En:
Memorias del Congreso Iberoamericano Virtual de Acuicultura
(CIVA,http://www.revistaaquatic.com/civa2002), pp. 388-408.

61
Uemoto H., 1968. Relationship between oxygen consumption by the pearl oyster and
its environmental temperature. Bull Natl Pearl Res Lab, 13:1617-1623.
Velasco L. A., Navarro J. M. 2002. Feeding physiology of infaunal (Mulinia edulis) and
epifaunal (Mytilus chilensis) bivalves under a wide range of concentrations
and qualities of seston. Marine Ecology Progress Series, 240:143-155.
Velasco L. A., Navarro J. M. 2003. Energetic balance of infaunal (Mulina edulis King,
1831) and epifaunal (Mytilus chilensis Hupé, 1854) bivalves in response to
wide variations in concentrations and quality of seston. Journal of
Experimental Marine Biology and Ecology, 296:79-92.
Velasco L. A. 2007. Energetic physiology of the Caribbean scallops Argopecten
nucleus and Nodipecten nodosus fed with different microalgal diets.
Aquaculture, 270:299-311.
Vite-García N., Saucedo P. E. 2008. Energy storage and allocation during of Pacific
winged pearl oyster Pteria sterna at Bahía de la Paz, Baja California Sur,
México. Journal of Shellfish Research, 27:375-383.
Ward J. E., Levinton J. S., Shumway S. E. 2003. Influence of diet on pre-ingestive
particle processing in bivalves. I: Transport velocities on the ctenidium.
Journal of Experimental Marine Biology and Ecology, 129-149.
Ward J. E., Shumway S. E. 2004. Separating the grain from the chaff: particle
selection in suspension-and deposit-feeding bivalves. Journal of Experimental
Marine Biology and Ecology, 300: 83-130.
Warren C. E., Davis G. E. 1967. Laboratory studies on the feeding bioenergetics and
growth of fishes. En: Gerking S. D. (Ed.) The biological basis of -freshwater
fish production, Blackwell, Oxford, p. 175-214.
Widdows J., Bayne B. L. 1971.Temperature acclimation of Mytilus edulis with
reference to its energy budget. Journal of the Marine Biological Association of
the United Kingdom, 51:827-843.
Widdows J. 1976. Physiological adaptation of Mytilus edulis to cyclic temperatures.
Journal of Comparative Physiology, 105:115-128.

62
Widdows J. P., Fieth P., Worall C. M. 1979. Relationships between seston, available
food and feeding activity in the common mussel Mytilus edulis. Marine
Biology, 50:195207.
Widdows J., Donkin P., Worrall C. M. 1988. Interpretation of relationship between
growth and concentration of aromatic hydrocarbons in the tissue of Mytilus
edulis: mechanisms of toxicity and ecological consequences. Marine
Environmental Research, 24:254.
Widdows J., Page D. 1993. Effects of tributyltin and dibutyltin on the physiological
energetic of the mussel Mytilus edulis. Marine Environmental Research,
35:233-249.
Widdows J., Johnson D. 1988. Physiological energetics of Mytilus edulis: scope for
growth. Marine Ecology Progress Series, 46:113-121.
Widdows J., Burns K. A., Menon N. R., Page D. S., Soria S. 1990. Measurement of
physiological energetics (scope for growth) and chemical contaminants in
mussels (Arca zebra) transplanted along a contaminant gradient in Bermuda.
Journal of Experimental Marine Biology and Ecology, 138:99117.
Widdows J., Donkin P., Brinsley M. D., Evans S. V., Salked P. N., Franklin A., Law R.
J., Waldeock M. J. 1995a. Scope for growth and contaminant levels in North
Sea mussels Mytilus edulis. Marine Ecology Progress Series, 27:131-148.
Widdows J., Donkin P., Evans S. V., Page D. S., Salked P. N. 1995b. Sublethal
biological effects and chemical monitoring of Sullom Voe (Shetland) using
mussels (Mytilus edulis). Proceedings of the Royal Society of Edinburgh
Section B. 103:99-112.
Widdows J., Nasci C., Fossato V. U. 1997. Effects of pollution on the scope for
growth of mussels (Mytilus galloprovincialis) from the Venice Lagoon Italy.
Marine Environmental Research, 43:69-79.
Widdows J., Brown S., Salkeld P., Elliot M. 2000. Temporal changes in intertidal
sediment erodibility: influence of biological and climatic factors. Continental
Shelf Research, 20:1275-1289.
Widdows J., Donkin P., Staff F. J., Matthiessen P., Law R. J., Allen Y. T., Thain J. E.,
Allchin C. R., Jones B. R. 2002. Measurement of stress effects (scope for

63
growth) and contaminant levels in mussels (Mytilus edulis) collected from the
Irish Sea. Marine Environmental Research, 53:327-356.
Wilbur A. E., Hilbish T. J. 1989. Physiological energetics of the ribbed mussel
Geukensia demissa (Dillwyn) in response to increased temperature. Journal
of Experimental Marine Biology and Ecology, 131:161-170.
Winter J. E. 1969. Über den Einfluß der Nahrungskonzentration und anderer Faktoren
auf Filtrierleistung und Nahrungsausnutzung der Muscheln Arctica islandica
und Modiolus modiolus. Marine Biology, 4:87-135.
Winter J. E. 1978. A review on the knowledge of suspension-feeding in
lamellibranchiate bivalves, with special reference to artificial aquaculture
systems. Aquaculture, 13:1-33.
Wong W. H., Cheung S. G. 2003. Seasonal variation in the feeding physiology and
scope for growth of green mussels, Perna viridis in estuarine Ma Wan, Hong
Kong. Journal of the Marine Biological Association of the United Kingdom,
83:543-552.
Wright H., Monteforte M. 1995. Ecology of pearl oyster spat collection in Bahía de La
Paz, South Baja California, México: Temporal and vertical distribution,
substrate selection, associated species. Resume 535. En: Abstracts of
International Congress Aquaculture’95, San Diego, California.
Wright-López. H. 1997. Ecología de la captación de semilla de madreperla Pinctada
mazatlanica y concha nácar Pteria sterna (Bivalvia: Pteriidae), en la isla
Gaviota, Bahía de La Paz, B. C. S., México. Tesis de Maestría en Ciencias,
Centro Interdisciplinario de Ciencias Marinas de IPN, La Paz, Baja California
Sur, México 139 p.
Wright-López H., Holguin-Quiñones O., Arreguín-Sanchez F., Roque-Villada I. 2009.
Crecimiento y mortalidad de la concha nácar Pteria sterna en bancos
silvestres de Baja California Sur, México. Revista de Biología Tropical,
57:659-670.
Young T. 1999. Exploring the potential of indigenous species for aquaculture in Bahia
Magdalena: Growth rate of cultivated concha nácar (Pteria sterna).

64
Swarthmore College, Swarthmore, PA. School for Field Studies, Centro de
Estudios Costeros, Puerto San Carlos, B. C. S. 1-7.
Yukihira H., Klumpp D. W., Lucas J. S. 1998a. Comparative effects of microalgal
species and concentration on suspension feeding and energy budgets of the
pearl oysters Pinctada margaritifera and P. maxima. Marine Ecology
Progress Series, 171:71-84.
Yukihira H., Klumpp D. W., Lucas J. S. 1998b. Effects of body size on suspension
feeding and energy budgets of the pearl oysters Pinctada margaritifera and P.
maxima. Marine Ecology Progress Series, 170:119-130.
Yukihira H., Klumpp D. W., Lucas J. S. 1999. Feeding adaptations of the pearl
oysters Pinctada margaritifera and P. maxima to variations in natural
particles. Marine Ecology Progress Series, 182:161-173.
Yukihira H., Lucas J. S., Klumpp D. W. 2000. Comparative effects of temperature on
suspension feeding and energy budgets of the pearl oysters Pinctada
margaritifera and P. maxima. Marine Ecology Progress Series, 195:179-188.
Zhang J., Fang J. G., Hawkins A. J. S., Pascoe P. L. 2004. The effect of temperature
on clearance rate and oxygen consumption of scallops, Chlamys farreri.
Journal of Shellfish Research, 23:715-721.







