Download - Páncreas exocrino y glándulas salivales

Sebastián Lavanderos B. 2do. Medicina UDP
1
PÁNCREAS EXOCRINO Y GLÁNDULAS SALIVALES
VISIÓN GENERAL DE LA FISIOLOGÍA DE LAS
GLÁNDULAS EXOCRINAS
EL PÁNCREAS Y LAS GLÁNDULAS SALIVALES PRINCIPALES
SON GLÁNDULAS COMPUESTAS EXOCRINAS
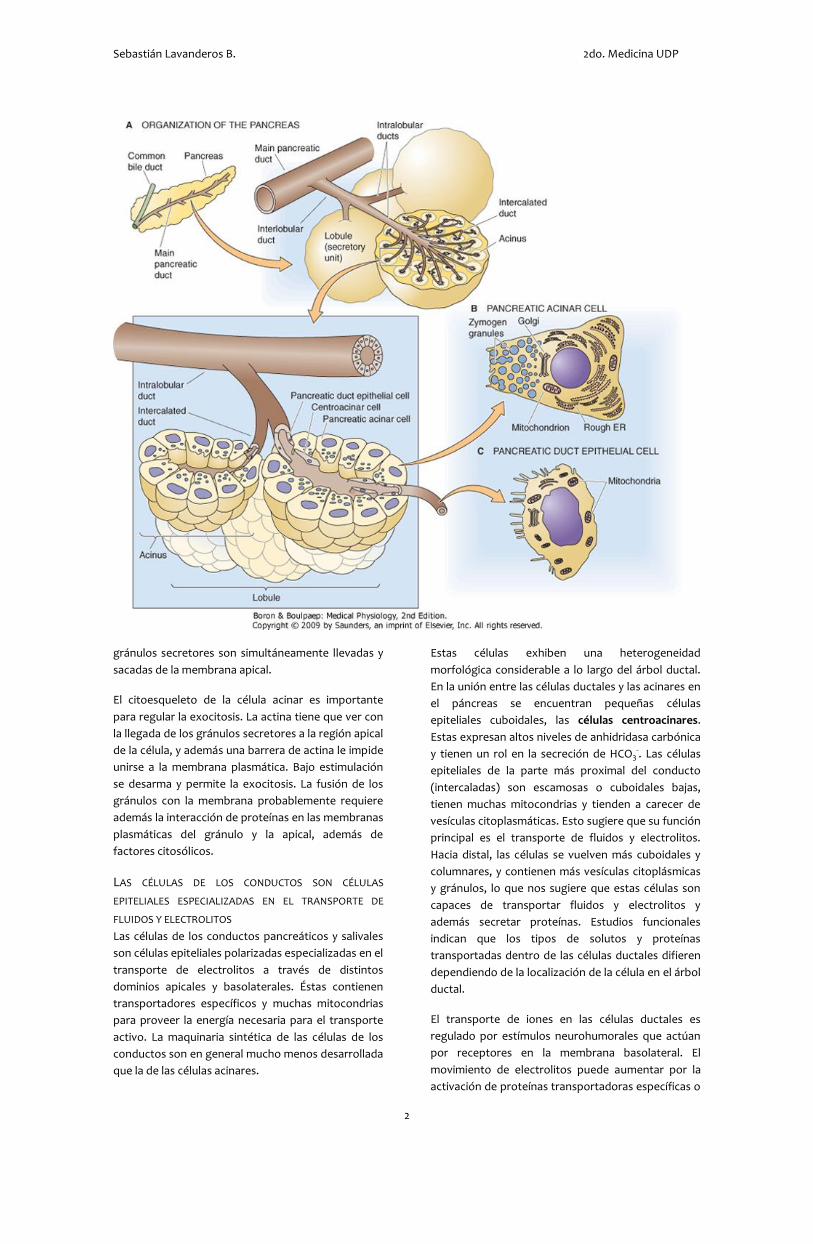
El páncreas exocrino y las glándulas salivales
principales son glándulas exocrinas compuestas –
órganos secretores especializados que tienen un
sistema de conductos ramificado a través del cual
liberan sus productos de secreción. La función
principal de estas glándulas exocrinas es ayudar en la
digestión de la comida. La saliva producida por las
glándulas salivales lubrica la comida ingerida e inician
la digestión del almidón. El jugo pancreático, rico en
HCO3- y enzimas digestivas neutraliza los contenidos
ácidos gástricos que entran al intestino delgado, y
completa la digestión intraluminal de los
carbohidratos, proteínas y grasas ingeridas. Cada una
de estas glándulas está bajo control neural y humoral
que genera una respuesta secretora coordinada
cuando nos alimentamos.
Morfológicamente, el páncreas y las glándulas
salivales se dividen en lóbulos, cada uno de los cuales
es una división del parénquima drenado por un
conducto intralobular singular. Grupos de lóbulos
separados por tejido conectivo son drenados por
conductos interlobulares más grandes, que vacían
hacia un conducto principal, que conecta la glándula
entera con el lumen del tracto GI.
Dentro de los lóbulos están las estructuras
funcionales microscópicas de la glándula. Cada
unidad secretora está compuesta de un acino y un
pequeño conducto intercalar. El acino es una
agrupación de 15 a 100 células acinares que sintetizan
y secretan proteínas al lumen de la estructura
epitelial. En el páncreas, estas secretan ~20
cimógenos (precursores enzimáticos inactivos) y
enzimas distintas. En las glándulas salivales, los
productos proteicos principales de las células
acinares son la α-amilasa, mucinas y proteínas ricas en
prolina. Las células acinares tanto del páncreas y
glándulas salivales secretan un fluido parecido al
plasma que acompaña a las proteínas secretoras. La
secreción acinar completa final se conoce como
secreción primaria.
Cada lumen acinar está conectado con el final
proximal de un conducto intercalado. A distal, estos
se van uniendo para formar conductos cada vez más
grandes que al final forman el conducto intralobular
que drena al lóbulo completo. Estos ductos proveen
la vía de salida para la secreción primaria, pero esta
es modificada por las células epiteliales que limitan
los conductos, que cambian la composición de fluidos
y electrolitos de la secreción primaria. De esta
manera, la secreción exocrina final representa el
producto combinado de dos poblaciones celulares
distintas, la célula acinar y la del conducto.
Además de los conductos y acinos, las glándulas
exocrinas están ricamente inervadas e irrigadas.
Fibras postgangliónicas simpáticas y parasimpáticas
contribuyen a la inervación autónoma que regula la
secreción a través de la liberación de NTs
colinérgicos, adrenérgicos y peptídicos, que se unen
a receptores en las células acinares y del conducto.
Vías centrales y reflejos participan en la regulación
neural de la secreción exocrina. Los nervios
autónomos también llevan fibras del dolor aferentes
que son activadas por la inflamación glandular y
trauma. La vasculatura no solo lleva oxígeno y
nutrientes, sino también lleva hormonas que regulan
la secreción.
LAS CÉLULAS ACINARES SON CÉLULAS ESPECIALIZADAS
QUE SINTETIZAN PROTEÍNAS
Las células acinares, como las del páncreas y
glándulas salivares, son células epiteliales polarizadas
que se especializan en la producción y exportación de
grandes cantidades de proteínas. Entonces, tiene un
retículo endoplasmático (RE) muy extenso. Sin
embargo, su característica más significativa es la
abundancia de gránulos de secreción electrodensos
en el polo apical de la célula. Estos son pools de
almacenamiento de las proteínas de secreción, listos
para liberar sus contenidos después de la
estimulación de la célula por agentes
neurohumorales. Los gránulos de secreción de las
células acinares pancreáticas contienen una mezcla
de cimógenos y enzimas requeridos para la digestión.
Los gránulos secretores de las células acinares
salivales tienen α-amilasa (en la parótida) o mucinas
(en las sublinguales). Los gránulos secretores en el
páncreas se ven uniformes, mientras que en las
glándulas salivales tienen condensaciones focales
conocidas como esférulas.
La exocitosis, proceso mediante el cual los gránulos
secretores liberan sus contenidos, es una serie
compleja de eventos que involucran el movimiento
de los gránulos a la membrana apical, fusión de los
gránulos con la membrana, y liberación de sus
contenidos al lumen acinar. La secreción es gatillada
por hormonas o actividad neural. Al inicio de esta, el
área de la membrana plasmática apical aumenta unas
30 veces. Después, la activación de una vía endocítica
lleva a la recuperación de las membranas de los
gránulos secretores para reciclarlas, lo que hace que
el área de la membrana apical disminuya y vuelva a su
valor normal. Entonces, durante el estado
estacionario de la secreción, las membranas de
Sebastián Lavanderos B. 2do. Medicina UDP
2
gránulos secretores son simultáneamente llevadas y
sacadas de la membrana apical.
El citoesqueleto de la célula acinar es importante
para regular la exocitosis. La actina tiene que ver con
la llegada de los gránulos secretores a la región apical
de la célula, y además una barrera de actina le impide
unirse a la membrana plasmática. Bajo estimulación
se desarma y permite la exocitosis. La fusión de los
gránulos con la membrana probablemente requiere
además la interacción de proteínas en las membranas
plasmáticas del gránulo y la apical, además de
factores citosólicos.
LAS CÉLULAS DE LOS CONDUCTOS SON CÉLULAS
EPITELIALES ESPECIALIZADAS EN EL TRANSPORTE DE
FLUIDOS Y ELECTROLITOS
Las células de los conductos pancreáticos y salivales
son células epiteliales polarizadas especializadas en el
transporte de electrolitos a través de distintos
dominios apicales y basolaterales. Éstas contienen
transportadores específicos y muchas mitocondrias
para proveer la energía necesaria para el transporte
activo. La maquinaria sintética de las células de los
conductos son en general mucho menos desarrollada
que la de las células acinares.
Estas células exhiben una heterogeneidad
morfológica considerable a lo largo del árbol ductal.
En la unión entre las células ductales y las acinares en
el páncreas se encuentran pequeñas células
epiteliales cuboidales, las células centroacinares.
Estas expresan altos niveles de anhidridasa carbónica
y tienen un rol en la secreción de HCO3-. Las células
epiteliales de la parte más proximal del conducto
(intercaladas) son escamosas o cuboidales bajas,
tienen muchas mitocondrias y tienden a carecer de
vesículas citoplasmáticas. Esto sugiere que su función
principal es el transporte de fluidos y electrolitos.
Hacia distal, las células se vuelven más cuboidales y
columnares, y contienen más vesículas citoplásmicas
y gránulos, lo que nos sugiere que estas células son
capaces de transportar fluidos y electrolitos y
además secretar proteínas. Estudios funcionales
indican que los tipos de solutos y proteínas
transportadas dentro de las células ductales difieren
dependiendo de la localización de la célula en el árbol
ductal.
El transporte de iones en las células ductales es
regulado por estímulos neurohumorales que actúan
por receptores en la membrana basolateral. El
movimiento de electrolitos puede aumentar por la
activación de proteínas transportadoras específicas o

Sebastián Lavanderos B. 2do. Medicina UDP
3
vía aumento del número de transportadores en la
membrana plasmática.
LAS CÉLULAS CALICIFORMES PRODUCEN MUCINA EN LAS
GLÁNDULAS EXOCRINAS
Además de las células acinares y ductales, las
glándulas exocrinas contienen un número variable de
células caliciformes, que secretan glicoproteínas de
alto peso molecular conocidas como mucinas.
Cuando se hidratan, estas forman moco. El moco
tiene varias funciones, entre las cuales destacan la
lubricación, hidratación y la protección mecánica de
células epiteliales de superficie. También tienen un
rol inmune, al unirse a patógenos e interactuando
con células inmunes competentes. Esto ayuda a
prevenir infecciones. En el páncreas, las células
caliciformes secretoras de mucina se encuentran
entre las células epiteliales que limitan los conductos
grandes y distales. Estas pueden llegar a ser el 25% de
las células epiteliales del conducto pancreático
principal en algunas especies En las glándulas
salivales, estas se ven en los conductos grandes
distales, aunque en menos cantidad que en el
páncreas. Sin embargo, en las glándulas salivales, la
mucina es secretada también por las células acinares.
LA CÉLULA ACINAR DEL PÁNCREAS
LA CÉLULA ACUNAR SECRETA PROTEÍNAS DIGESTIVAS EN
RESPUESTA A LA ESTIMULACIÓN
La amilasa es secretada en su forma completamente
activa, por lo que se usa como marcador de secreción
de las células acinares pancreáticas cuando se estudia
su secreción a nivel celular.
Cuando las células acinares se encuentran sin
estimulación, secretan bajos niveles de proteínas
digestivas a través de una vía de secreción
constitutiva. Las células acinares estimuladas por
agentes neurohumorales secretan proteínas a través
de una vía regulada. La secreción regulada de los
acinos y lóbulos in vitro se detecta a los 5 minutos de
estimulación y es dependiente de energía. Durante
un periodo de estimulación de 30 a 60 minutos las
células acinares secretan de 5 a 10 veces más amilasa
que vía liberación constitutiva. Sin embargo, secretan
sólo de un 10 a 20% de sus reservas en gránulos.
Aumentan la síntesis proteica para llenarlas luego.
La célula acinar tiene 2 patrones de secreción
regulada: monofásica y bifásica. Un agonista que
genere una relación dosis-respuesta monofásica
(como el GRP) causa una secreción que alcanza un
nivel máximo que no baja con concentraciones más
altas del agente. Al revés, un secretagogo que genere
una relación dosis-respuesta bifásica (como la CCK y
el carbacol) hace que la secreción alcance un nivel
máximo que disminuye subsecuentemente con
concentraciones más altas del agente. Esta respuesta
bifásica podría reflejar la presencia de receptores de
alta y baja afinidad y se relaciona con la patogénesis
de la pancreatitis aguda.
LA SECRECIÓN REGULADA DE PROTEÍNAS POR LAS
CÉLULAS ACINARES PANCREÁTICAS ES MEDIADA A TRAVÉS
DE LA COLECISTOQUININA Y RECEPTORES MUSCARÍNICOS
Aunque se han encontrado al menos 12 receptores
distintos en la membrana de la célula acinar
pancreática, los más importantes en la regulación de
la secreción de proteínas son los receptores de CCK y
los muscarínicos de ACh. Estos dos son muy similares.
Ambos están unidos a proteína Gαq, y usan la vía de
transducción de señal de PLC/Ca+2, y ambos
aumentan la secreción de enzimas en la célula acinar.
Existen 2 receptores de CCK muy ligados entre sí, que
se distinguen por su estructura, afinidad a ligandos y
distribución en tejidos. Ambos son activados por CCK
o gastrina, pero a distinta afinidad. El receptor CCKA
tiene mayor afinidad por la CCK que por la gastrina, y
el CCKB tiene más menos la misma afinidad por las 2.
Los receptores de CCK pueden existir en estados de
alta y baja afinidad. Bajas concentraciones de CCK
(picomolar) activan las formas de alta afinidad de los
receptores de CCK y estimulan la secreción.
Concentraciones suprafisiológicas (10 a 100 veces
mayores) de CCK activan las formas de baja afinidad
del receptor e inhiben la secreción. Estos distintos
estados de afinidad usan patrones de señalización
distintos. Es probable que en condiciones fisiológicas
sólo los estados de alta afinidad de los receptores de
CCK o muscarínicos se encuentren activados. La
estimulación de los estados de baja afinidad por
concentraciones suprafisiológicas de CCK o ACh no
sólo inhibe la secreción enzimática sino también
podría dañar a la célula acinar (Pancreatitis Aguda).
El receptor muscarínico de la célula acinar es
probablemente del tipo M3. Se localiza en la
membrana basolateral de la célula. También se
encuentran muchos otros receptores en la célula
acinar, como para GRP, SS y VIP, CGRP, insulina y
secretina. Aunque podrían tener un papel en la
regulación de la secreción, sus roles no han sido
identificados con claridad.
La activación de receptores que estimulan vías de
transducción de señal distintas podría llevar a una
respuesta secretora estimulada. La estimulación
simultánea del receptor CCK de alta afinidad (que
actúa vía [Ca+2]i) y del receptor VIP (que actúa vía
cAMP) genera un efecto cooperativo en la secreción.
De manera alternativa, las células acinares
previamente estimuladas podrían entrar a un periodo
refractario temporal debido a la estimulación
subsecuente, fenómeno que se conoce como
desensibilización.

Sebastián Lavanderos B. 2do. Medicina UDP
4
EL CA+2
ES EL SEGUNDO MENSAJERO PRINCIPAL PARA LA
SECRECIÓN DE PROTEÍNAS POR LAS CÉLULAS ACINARES
PANCREÁTICAS
CA+2
En estado de descanso, la [Ca+2]i oscila lentamente.
En presencia de concentraciones máximas
estimulatorias de CCK o ACh, la frecuencia de las
oscilaciones aumenta, pero se notan pequeños
cambios en su amplitud. Se requiere este aumento en
la frecuencia de las oscilaciones de la [Ca+2]i para que
se secreten proteínas. Concentraciones supra-
máximas (hiperestimulantes) de CCK o ACh generan
un gran spike (espiga) súbito de [Ca+2]i y eliminan las
oscilaciones adicionales. Este spike es el que se asocia
a la inhibición de la secreción que parece ser mediada
por la ruptura de los componentes citoesqueléticos
que se requieren para la secreción.
CGMP
La estimulación fisiológica de la célula acinar por CCK
o ACh genera un aumento rápido y prominente en los
niveles de [cGMP]i. Este aumento se ha asociado al
metabolismo del NO. La inhibición de la NO sintasa
bloquea el aumento en la [cGMP]i después de la
estimulación por secretagogos. Hay evidencia que
sugiere que el cGMP tiene que ver con la regulación
de la entrada de Ca+2 y su almacenamiento en la
célula acinar.
CAMP
La secretina, VIP y CCK aumentan la producción de
cAMP y por tanto activan a la PKA en las células
acinares pancreáticas. Bajas concentraciones de CCK
causan estimulaciones transitorias de la PKA,
mientras que concentraciones suprafisiológicas de
CCK causan un aumento en la [cAMP]i mucho más
prominente y prolongado que hace que aumente
más la PKA. La ACh, sin embargo, tiene un efecto
muy pequeño (si es que tiene) en la vía de
señalización del cAMP.
EFECTORES
Los efectores más importantes de los segundos
mensajeros intracelulares son las proteína quinasas.
La estimulación de los receptores de CCK y
muscarínicos en las células acinares llevan a la
generación de señales similares de Ca+2 y la activación
de proteína quinasas dependientes de calmodulina y
miembros de la familia de PKCs. La activación de los
receptores de secretina o VIP hacen que aumente la
[cAMP]i y por tanto activan a la PKA. Estos segundos
mensajeros probablemente activan proteína
fosfatasas, así como otras quinasas que no salen en la
figura a continuación. Los blancos de las quinasas y
fosfatasas activadas en la célula pancreática acinar
son desconocidos. Algunos regulan la secreción,
otros median la síntesis proteica, crecimiento,
transformación y muerte celular.
ADEMÁS DE LAS PROTEÍNAS, LA CÉLULA PANCREÁTICA
TAMBIÉN SECRETA UN FLUIDO SIMILAR AL PLASMA
Además de las proteínas, las células acinares
pancreáticas secretan un fluido isotónico similar al
plasma, rico en NaCl, que hidrata el material denso
rico en proteínas que secretan las células acinares. El
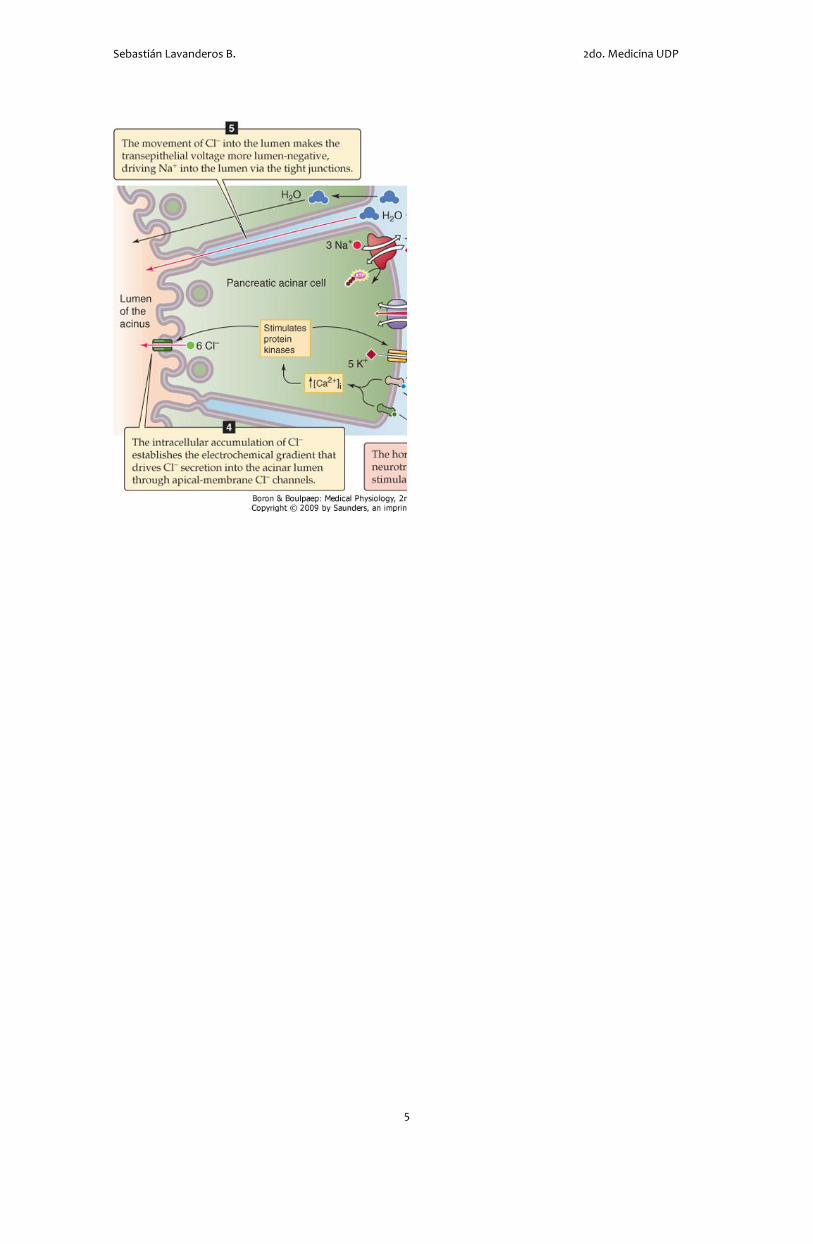
proceso fundamental de transporte para esto es la
secreción de Cl- por la membrana apical. Para que
ocurra el movimiento transcelular (del plasma al
lumen) de Cl-. Este se debe mover hacia la célula
atravesando la membrana basolateral. Esto ocurre
por un cotransportador Na/K/Cl. La bomba Na-K
genera el gradiente de Na+ que energiza el
cotransportador Na/K/Cl. El K+ que entra a través de
la bomba Na-K y del cotransportador Na/K/Cl sale a
través de canales de K+, que hay en la membrana
basolateral. Entonces, necesitamos la bomba, el
cotransportador y los canales para sostener la
absorción basolateral de Cl- a la célula acinar.
El aumento en la [Cl-]i producido por la absorción
basolateral de este ion conduce la secreción de Cl-
bajo su gradiente electroquímico a través de canales
en la membrana apical. A medida que el voltaje
transepitelial se va haciendo más negativo en el
lumen, el Na+ se mueve a través de la vía paracelular
selectiva para cationes (i.e. tight junctions) para
unirse al Cl- que ha sido secretado hacia el lumen.
También se mueve agua por esta vía paracelular,
Sebastián Lavanderos B. 2do. Medicina UDP
5

Sebastián Lavanderos B. 2do. Medicina UDP
6
además de las AQP en las membranas tanto
basolateral como apical. De esta manera, el efecto
neto de estos procesos de transporte acinar es la
producción de un fluido isotónico rico en NaCl que es
el ~25% de la secreción total de fluido pancreático.
De la misma manera que la secreción de proteína por
las células acinares, la secreción de fluido y
electrolitos es estimulada por secretagogos que
aumentan la [Ca+2]i.
LA CÉLULA DEL CONDUCTO PANCREÁTICO
LA CÉLULA DEL CONDUCTO PANCREÁTICO SECRETA
NAHCO3 ISOTÓNICO
La función fisiológica principal de las células del
conducto pancreático es secretar un fluido rico en
HCO3- que alcaliniza e hidrata las secreciones
primarias ricas en proteína de la célula acinar. En paso
apical de la secreción transepitelial de HCO3- es
mediado en parte por un intercambiador Cl-HCO3,
que manda HCO3- intracelular al lumen del conducto.
Debe haber Cl- en el lumen para que este transporte
pueda ocurrir. Aunque hay un poco de Cl- en las
secreciones primarias de la célula acinar, canales
aniónicos en la membrana apical de la célula del
conducto proveen el Cl- adicional para el lumen en un
proceso llamado reciclaje de Cl-. El más importante de
estos es el CFTR (Transregulador de la conductancia
de membrana de Fibrosis Quística), un canal de Cl-
activado por cAMP presente en las membranas
apicales de las células de los conductos pancreáticos.
Los canales apicales de Cl- también pueden servir
directamente como conductos para el movimiento de
HCO3- desde la célula ductal hacia el lumen.
El HCO3- intracelular que sale de la célula del
conducto a través de la membrana apical viene de 2
vías. La primera es la absorción directa de HCO3- a
través de un cotransportador electrogénico Na/HCO3
(NBCe1). El segundo mecanismo es la generación
intracelular de HCO3- a partir de CO2 y OH-, catalizado
por la anhidridasa carbónica. El OH- de esta reacción
viene junto a H+ del H2O. De esta manera, se acumula
H+ que debe ser sacado de la célula por la membrana
basolateral. Esto ocurre por intercambio Na-H o una
bomba de H+ ATP-dependiente. Las células de los
conductos pancreáticos tienen vesículas ácidas
intracelulares (que presumiblemente contienen
bombas vacuolares de H+) que se movilizan a la
membrana basolateral después de la estimulación
por secretina, que es un potente secretagogo. De
hecho, las bombas de H+ se encuentran en su mayor
actividad en condiciones de estimulación
neurohumoral. Entonces, 3 transportadores
basolaterales proveen directa o indirectamente el
HCO3- intracelular que las células de los conductos
pancreáticos requieren para secreción: (1) el
cotransportador electrogénico Na/HCO3, (2) el
intercambiador Na-H y (3) la bomba de H+. La célula
del conducto pancreático secreta un ~75% del total de
secreción de fluido pancreática.
LA SECRETINA (A TRAVÉS DE CAMP) Y ACH (A TRAVÉS
DE CA+2) ESTIMULAN LA SECRECIÓN DE HCO3
- POR EL
CONDUCTO PANCREÁTICO
Cuando son estimularas, las células epiteliales del
conducto pancreático secretan una solución
isotónica de NaHCO3. Estas células tienen receptores
para secretina, ACh, GRP (que estimulan la secreción
de HCO3-) y sustancia P (que la inhibe). Hay evidencia
de actividad moduladora de la CCK sobre la
secreción, pero no se han identificado receptores.
La secretina es el regulador humoral más importante
de la secreción ductal de HCO3-. La activación de su
receptor estimula a la adenil ciclasa, lo que aumenta
la [cAMP]i, que activa la PKA. Se ha observado que
bajas concentraciones de secretina que no aumentan
mediblemente la [cAMP]i pueden estimular la
secreción de HCO3-. Esto sugiere que la respuesta a
secretina podría ser mediada por (1) pequeños
aumentos imposibles de medir en el cAMP total en la
célula, (2) aumentos de cAMP localizados en
pequeños compartimentos intracelulares, o (3)
activación de vías de segundos mensajeros
alternativas. La secretina actúa estimulando el canal
de Cl- CFTR apical y el cotransportador basolateral
Na/HCO3, sin afectar al intercambiador Na-H.
La secreción de HCO3- también es regulada por el
sistema parasimpático, a través de ACh, que aumenta
la [Ca+2]i y activa proteína quinasas dependientes de
Ca+2 (como la PKC y quinasas dependientes de
calmodulina) en la célula del conducto pancreático.
La ACh es inhibida por atropina, lo que nos sugiere
que este NT está actuando a través de receptores
muscarínicos en la célula del conducto pancreático.
Aunque la secreción ductular también es estimulada
por GRP, no se sabe cómo, pero se sabe que no es
por [Ca+2]i ni [cAMP]i.
En ratas, la secreción ductular basal y estimulada de
HCO3- es inhibida por sustancia P. El segundo
mensajero que media esto es desconocido, y logra
inhibir la secreción sin importar qué secretagogo la
estimule, por lo que se piensa que probablemente
actúa distal al sitio donde se generan los segundos
mensajeros, por ejemplo inhibiendo el
intercambiador Cl-HCO3.
LOS CANALES DE CLORURO APICALES DE MEMBRANA SON
SITIOS IMPORTANTES DE REGULACIÓN NEUROHUMORAL
Se han identificado como proteínas efectoras de las
quinasas y fosfatasas activadas por los mecanismos
neurohumorales que regulan a las células del
conducto pancreático a los canales apicales de Cl-, los
canales basolaterales de K+ y el cotransportador
Sebastián Lavanderos B. 2do. Medicina UDP
7
Na/HCO3. El canal CFTR tiene dominios de unión de
nucleótidos que controlan su apertura y cierre,
además de un dominio regulatorio con múltiples
sitios de fosforilación para la PKA y PKC. Los agentes
neurohumorales que controlan la secreción de
fluidos y electrolitos por las células del conducto
pancreático actúan aquí. De esta manera, el canal
CFTR de Cl- es regulado por ATP vía 2 mecanismos:
interacción con los dominios de unión a nucleótidos y
fosforilación de proteínas.
En células del conducto pancreático de ratas, los
canales sensibles a Ca+2 de K+ basolaterales parecen
ser los blancos de la estimulación neurohumoral.
Cosas que activen la vía del cAMP estimulan la
fosforilación por PKA, promoviendo la respuesta de
estos canales al [Ca+2]i y aumentando su probabilidad
de estar abiertos.
LAS CÉLULAS DEL CONDUCTO PANCREÁTICO TAMBIÉN
PUEDEN SECRETAR GLICOPROTEÍNAS
Aunque la función principal de las células del
conducto pancreático es secretar HCO3- y agua, estas
células pueden también sintetizar y secretar varias
glicoproteínas de alto peso molecular, que no se
acumulan en gránulos de secreción, sino que más
bien parece que se están continuamente sintetizando
y secretando de pequeñas vesículas citoplásmicas. La
secretina aumenta la secreción de glicoproteínas, a
través de la estimulación de su síntesis y no de su
transporte o exocitosis per sé. Estas proteínas
podrían proteger en contra del daño a las células
mucosales inducido por proteasas.
COMPOSICIÓN, FUNCIÓN Y CONTROL DE LA
SECRECIÓN PANCREÁTICA
EL JUGO PANCREÁTICO ES UNA SECRECIÓN ALCALINA
RICA EN PROTEÍNAS
Los humanos producen ~1,5 L/día de fluido
pancreático. El páncreas tiene las tasas de síntesis y
secreción de proteínas más altas del cuerpo. Cada
día, el páncreas manda de 15 a 100 g de proteínas
hacia el intestino delgado. El nivel de secreción
pancreática se determina por un balance entre
estimulación e inhibición de la secreción.
El páncreas humano secreta más de 20 proteínas, que
en su mayoría son cimógenos (precursores de
enzimas digestivas) o enzimas digestivas activas. Las
proteínas secretadas responsables de la digestión se
pueden clasificar de acurdo a sus sustratos en:
proteasas, que hidrolizan proteínas, amilasas, que
digieren carbohidratos, lipasas y fosfolipasas, que
rompen lípidos y nucleasas que digieren ácidos
nucleicos. La función de otras proteínas (como la
GP2, litostatina y proteína asociada a la pancreatitis)
no han sido bien definidas aún.
La GP2 es una proteína que ha sido implicada en la
regulación de la endocitosis. Bajo ciertas
circunstancias puede formar agregados proteicos en
el jugo pancreático junto a la litostatina, que pueden
obstruir el lumen de los acinos en pacientes con
fibrosis quística y pancreatitis crónica.
La proteína asociada a la pancreatitis es una proteína
que está presente en bajas concentraciones en
estado normal, sin embargo, sus niveles aumentan
cientos de veces en las fases tempranas de un daño
pancreático. Esta proteína es un agente
bacteriostático que podría ayudar a prevenir la
infección pancreática en el combate de la
pancreatitis.
El jugo pancreático es rico también en Ca+2 y HCO3-.
Las concentraciones de calcio están en el rango de
los milimolares, y podrían ser necesarias para inducir
la agregación de proteínas secretoras y dirigirlas
hacia la vía secretora. El bicarbonato secretado por
las células del conducto pancreático neutraliza las
secreciones ácidas gástricas que entran al duodeno y
le permite a las enzimas digestivas funcionar
apropiadamente. También facilita la solubilización
micelar de lípidos y el funcionamiento de las células
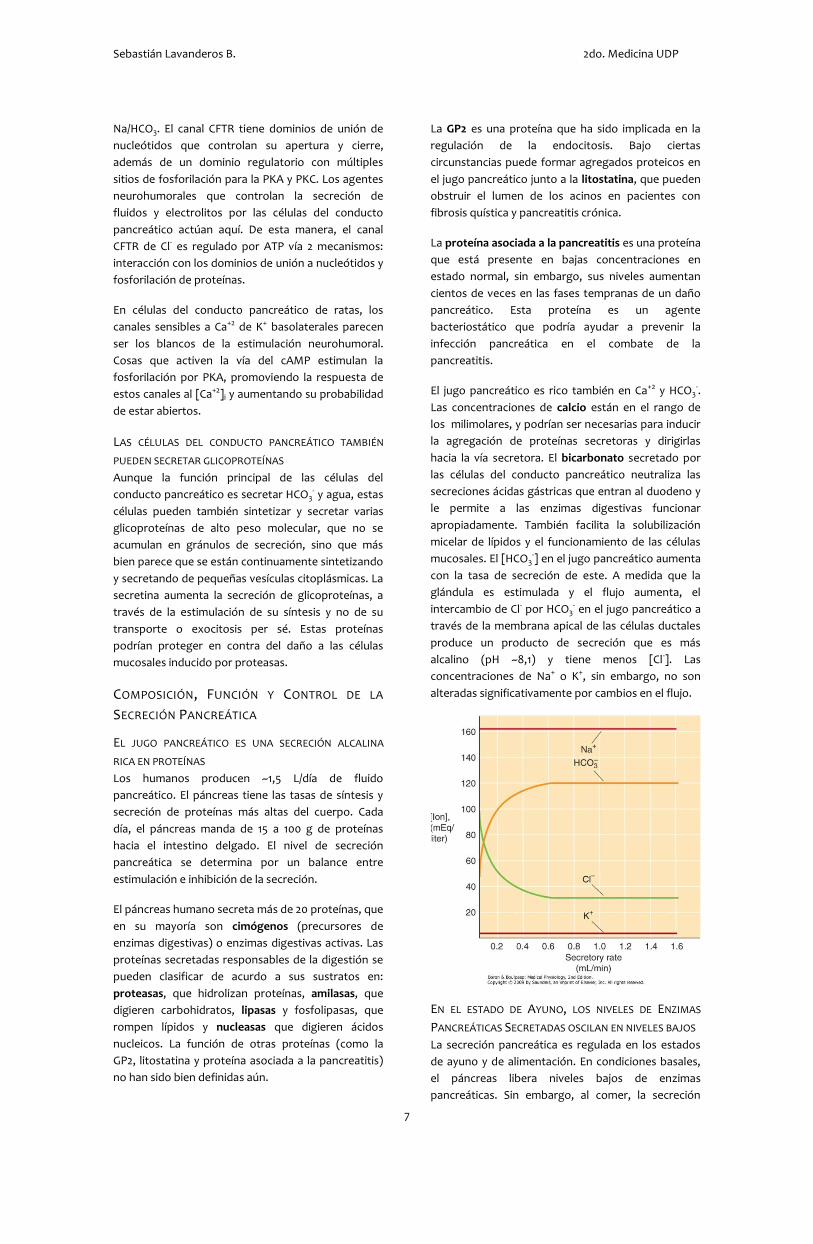
mucosales. El [HCO3-] en el jugo pancreático aumenta
con la tasa de secreción de este. A medida que la
glándula es estimulada y el flujo aumenta, el
intercambio de Cl- por HCO3- en el jugo pancreático a
través de la membrana apical de las células ductales
produce un producto de secreción que es más
alcalino (pH ~8,1) y tiene menos [Cl-]. Las
concentraciones de Na+ o K+, sin embargo, no son
alteradas significativamente por cambios en el flujo.
EN EL ESTADO DE AYUNO, LOS NIVELES DE ENZIMAS
PANCREÁTICAS SECRETADAS OSCILAN EN NIVELES BAJOS
La secreción pancreática es regulada en los estados
de ayuno y de alimentación. En condiciones basales,
el páncreas libera niveles bajos de enzimas
pancreáticas. Sin embargo, al comer, la secreción

Sebastián Lavanderos B. 2do. Medicina UDP
8
pancreática aumenta secuencialmente de 5 a 20
veces los niveles basales. Los sistemas que regulan la
secreción parecen ser redundantes.
Como otros órganos del tracto GI superior, el
páncreas tiene una tasa de secreción basal (en
reposo), aun cuando no se está comiendo o
digiriendo algo. Durante este periodo interdigestivo
(ayuno), las secreciones pancreáticas varían
cíclicamente, lo que corresponde a cambios cíclicos
en la motilidad del intestino delgado. La secreción
pancreática es mínima en la fase I de motilidad
intestinal en fase quieta, en la fase II la motilidad
duodenal aumenta, y la secreción pancreática
también. Durante el periodo interdigestivo, la
secreción enzimática es máxima cuando la motilidad
intestinal (MMCs) es máxima. Sin embargo, esta tasa
de secreción interdigestiva es sólo un 10 a 20% de la
estimulada por comidas. Las fases peak de la
actividad motora intestinal y secretora pancreática
son seguidas de un periodo de disminución (fase IV).
Las tasas de secreción de fluidos y electrolitos durante
la fase interdigestiva son usualmente menos de un 5%
que los niveles máximos.
El patrón cíclico de la secreción interdigestiva
pancreática es mediada por mecanismos intrínsecos y
extrínsecos. El mecanismo predominante de
regulación pancreática es vía el sistema
parasimpático. La CCK y las vías adrenérgicas
también tienen un rol. La CCK parece estimular la
secreción enzimática en las fases I y II. Al revés, el
tono α-adrenérgico basal parece suprimir la
secreción pancreática interdigestiva. El rol del SNA es
de regulación de la secreción basal pancreática.
LA CCK DE LAS CÉLULAS I DUODENALES ESTIMULA LA
SECRECIÓN ENZIMÁTICA POR LOS ACINOS, Y LA
SECRETINA DE LAS CÉLULAS S ESTIMULA LA SECRECIÓN
DE HCO3- Y FLUIDO POR LOS CONDUCTOS
La CCK es importante regulando la secreción
pancreática. Es liberada por las células I duodenales y
actúa en las células acinares pancreáticas
aumentando la secreción de proteínas. En respuesta
a una comida, los niveles de CCK plasmáticos
aumentan de 5 a 10 veces en 10 a 30 minutos. Los
lípidos son el secretagogo más potente de CCK.
También los productos de la digestión de proteínas,
carbohidratos y ácido, pero en menor extensión. Los
factores de liberación de CCK son péptidos liberados
por las células mucosales del duodeno o secretadas
por el páncreas que estimulan la secreción de CCK. El
nivel de estos factores refleja un balance entre las
cantidades relativas de nutrientes y enzimas
digestivas presentes en el lumen intestinal, así que el
nivel de factores refleja el medio digestivo del
duodeno. El nivel relativo de proteínas vs. proteasas
en el intestino delgado determina la cantidad de
factor de liberación de CCK disponible para dirigir la
liberación de CCK y por tanto secreción pancreática.
La CCK actúa en la célula acinar por vías directas e
indirectas. Estimula directamente la secreción
enzimática vía un receptor CCKA en la célula acinar y
podría estimular indirectamente la secreción
enzimática activando el sistema nervioso
parasimpático. La estimulación vagal lleva la
secreción pancreática a niveles cercanos al máximo.
La atropina reduce la secreción de enzimas y HCO3-
durante la fase intestinal de una comida, y también
inhibe la secreción en respuesta a estimulación por
niveles fisiológicos de CCK exógeno. Esto sugiere que
la CCK de alguna manera estimula la vía
parasimpática, que, a su vez, estimula los receptores
muscarínicos en la célula acinar.
Como la CCK, el GRP también podría ser un regulador
fisiológico de la secreción de enzimas pancreáticas.
La estimulación con GRP induce la secreción
enzimática. El GRP parece venir de las terminaciones
nerviosas vagales.
La secretina es el estimulador humoral más potente
de la secreción de fluidos y HCO3- por el páncreas. Es
liberada desde células neuroendocrinas tipo S en la
mucosa del intestino delgado en respuesta a la
acidificación duodenal (pH < 4,5) y en una extensión
menor a los ácidos biliares y lípidos. La secretina
actúa junto a CCK, ACh y otros agentes para estimular
la secreción de HCO3-.
Además de las hormonas de origen intestinal, la
insulina y otras hormonas secretadas por los islotes
de Langerhans dentro del páncreas podrían también
influenciar la secreción pancreática exocrina. El flujo
sanguíneo desde los islotes pancreáticos posibilitaría
esto, al exponer a las células pancreáticas acinares a
altísimas concentraciones de hormonas de los islotes.
Un resultado de este arreglo podría ser que la
insulina modifique la composición de las enzimas
digestivas dentro de la célula acinar y aumente los
niveles relativos de amilasa.
COMER GATILLA LAS FASES CEFÁLICA, GÁSTRICA E
INTESTINAL DE LA SECRECIÓN PANCREÁTICA, MEDIADAS
POR UNA COMPLEJA RED DE INTERACCIONES
NEUROHUMORALES
El periodo digestivo ha sido dividido en 3 fases,
basándose en el sitio donde la comida actúa para
estimular la secreción pancreática. Estas fases son
secuenciales y actúan de manera coordinada.
FASE CEFÁLICA
Durante esta fase el sentir, saborear y oler comida
usualmente genera un pequeño incremento en la
secreción de fluidos y electrolitos, pero un efecto
prominente en la secreción de enzimas ( 25-50% en

Sebastián Lavanderos B. 2do. Medicina UDP
9
relación al máximo evocado por CCK endógena). Esta
fase es corta y se disipa rápidamente al remover la
comida, y es mediada por aferencias y eferencias
vagales, esta última vía ACh que estimula receptores
muscarínicos en las células acinares.
FASE GÁSTRICA
En la fase gástrica, la presencia de comida en el
estómago modula la secreción pancreática vía: (1)
liberación de hormonas, (2) estimulación de vías
neurales, y (3) modificando el pH y disponibilidad de
nutrientes en la parte proximal del intestino delgado.
La presencia de péptidos y aminoácidos estimula la
liberación de gastrina (células G antrales y duodeno
proximal), que actúan en los receptores de
gastrina/CCKB y a menor extensión los de CCKA, que
no están presentes en algunas especies. La
importancia de la regulación por gastrina no está
clara, y aunque la presencia de comida en el
estómago afecta a la secreción pancreática, el rol
más importante del quimo en el control de la
secreción pancreática es después de que los
contenidos gástricos entran al intestino delgado.
FASE INTESTINAL
En esta fase, el quimo que entra a la región proximal
del intestino delgado estimula una gran respuesta
secretora pancreática por 3 mecanismos principales:
1. El ácido gástrico que entra al duodeno y en
menor extensión los ácidos biliares y lípidos
estimulan a las células S duodenales para
que liberen secretina, que estimula a las
células del conducto pancreático para que
liberen HCO3- y fluidos.
2. Los lípidos y péptidos y aminoácidos a
menor extensión, estimulan a las células I
duodenales para que liberen CCK, que
estimula a las células acinares para que
liberen enzimas digestivas.
3. El mismo estímulo que estimula a las células
I también activa un reflejo vagovagal
enterohepático que estimula las células
acinares.
El patrón de secreción enzimática depende de los
contenidos de la comida. Una comida líquida gatilla
una respuesta ~60% del máximo. Una sólida gatilla
una respuesta más prolongada, y una comida rica en
calorías gatilla la respuesta más potente.
La química de los nutrientes también afecta la
secreción. Los carbohidratos tienen poco efecto en la
secreción, mientras que los lípidos son potentes
estimuladores de la secreción de enzimas
pancreática. Un dato importante, los triglicéridos no
estimulan la secreción pancreática, sus productos de
hidrólisis (monoglicéridos y ácidos grasos libres) sí.
Algunos ácidos grasos también estimulan la secreción
de HCO3- pancreática, y reducen la secreción de ácido
gástrico y retrasan el vaciamiento gástrico, por lo que
podrían tener un rol importante en modular las
condiciones de pH en la parte proximal del intestino
delgado. El rompimiento de proteínas genera
intermediarios con efectos estimulantes. Los
aminoácidos no esenciales tienen poco efecto en la
secreción de proteínas, mientras que algunos
aminoácidos esenciales estimulan la secreción
(fenilalanina, valina y metionina), y los péptidos que
los contienen también.
Fase Estimulante Vía
Regulatoria
% Secreción Enzimática
Máxima
Cefálica Ver Oler Saborear Masticar
Vías Vagales 25%
Gástrica Digestión Gastrina?
Vagal-colinérgico
10-20%
Intestinal Aminoácidos Ác. Grasos H+
CCK Secretina Reflejos Entero-pancreáticos
50-80%
La potencia relativa de los distintos nutrientes en la
estimulación de la secreción es inversamente
proporcional a las reservas pancreáticas de enzimas
digestivas, de esta manera, sólo una pequeña porción
de amilasa se libera para digerir carbohidratos, pero
se liberan fracciones mayores de lipasa pancreática
para digerir eficientemente la grasa en la mayoría de
las comidas. El páncreas exocrino tiene la habilidad
de responder a cambios a largo plazo en la
composición de la dieta modulando las reservas de
enzimas pancreáticas. De esta manera, dietas altas en
carbohidratos pueden llevar a un incremento relativo
en el contenido pancreático de amilasa.
EL PÁNCREAS GRANDES RESERVAS DE ENZIMAS
DIGESTIVAS PARA LOS CARBOHIDRATOS Y PROTEÍNAS,
NO ASÍ PARA LOS LÍPIDOS
El páncreas exocrino guarda más enzimas que las
requeridas para digerir una comida. La mayor reserva
es la de enzimas requeridas para digestión de
carbohidratos y proteínas. Las reservas enzimáticas
para digestión de lípidos (especialmente para la
hidrólisis de triglicéridos) son más limitadas. Estudios
indican que empieza a ocurrir mal digestión de grasas
luego de la remoción de un 80-90% del páncreas,
observación con implicancia clínica importante que
indican que individuos pueden tolerar grandes
resecciones de páncreas por tumores sin el riesgo de
desarrollar mal digestión o diabetes posoperativa. Si
ocurre mal digestión de grasas o diabetes es un
indicador de destrucción masiva del páncreas.

Sebastián Lavanderos B. 2do. Medicina UDP
10
LA GRASA EN LA PARTE DISTAL DEL INTESTINO INHIBE LA
SECRECIÓN PANCREÁTICA
Una vez que se ha alcanzado la estimulación máxima,
la secreción pancreática comienza a caer después de
varias horas. Los sistemas de regulación devuelven
gradualmente la secreción al estado basal
(interdigestivo). Los mecanismos regulatorios de
esto no se encuentran tan bien caracterizados como
los que estimulan la secreción, pero se sabe que la
presencia de grasa en el final distal del intestino
delgado reduce la secreción pancreática en la
mayoría de los animales incluido el humano. Esta
inhibición podría estar mediada por péptido YY (PYY)
que podría suprimir la secreción pancreática al actuar
en vías neurales inhibitorias así como disminuyendo
el flujo sanguíneo pancreático. La somatostatina
(sobre todo SS-28), liberada de las células D
intestinales, y el glucagón (liberado de las células α
de los islotes pancreáticos), también podrían ser
factores que devuelvan la secreción pancreática a su
estado interdigestivo después de comer.
DISTINTOS MECANISMOS PROTEGEN AL PÁNCREAS DE SER
AUTODIGERIDO
La activación prematura de las enzimas pancreáticas
dentro de las células acinares podría llevar a la
autodigestión y tener un papel en el inicio de la
pancreatitis. Para prevenir esto, existen mecanismos
que previenen la activación enzimática prematura.
Primero, las proteínas digestivas están almacenadas
en los gránulos secretores como cimógenos, que sólo
se activan después de entrar el intestino delgado.
Aquí, la enzima intestinal enteroquinasa convierte el
tripsinógeno a tripsina, que inicia la conversión del
resto de cimógenos a sus formas activas. Segundo,

Sebastián Lavanderos B. 2do. Medicina UDP
11
las membranas de los gránulos secretores son
impermeables a proteínas. Tercero, inhibidores
enzimáticos como el inhibidor pancreático de
tripsina se encuentran empaquetados
conjuntamente en el gránulo secretor. Cuarto, la
condensación de lo cimógenos, el bajo pH y las
condiciones iónicas dentro de la vía secretora podrían
limitar la actividad enzimática al no ofrecer un
microambiente óptimo para su función. Quinto, las
enzimas que se activan prematuramente son
degradadas por otras enzimas o secretadas antes de
que causen daños.
La degradación de enzimas activadas
prematuramente podría estar mediada por otras
enzimas presentes dentro del gránulo secretor o al
mezclar los contenidos del gránulo secretor con
enzimas lisosomales que podrían degradar a las
enzimas activas. 3 mecanismos hacen que las
proteasas digestivas se mezclen con enzimas
lisosomales: (1) las enzimas lisosomales podrían estas
empaquetadas en conjunto dentro del gránulo
secretor, (2) los gránulos podrían fusionarse
selectivamente con lisosomas, o (3) los gránulos
podrían ser absorbidos por los lisosomas. La falla de
uno de estos mecanismos resulta en activación
prematura de las enzimas e inicia la pancreatitis.
LA CÉLULA ACINAR SALIVAL
DISTINTAS CÉLULAS ACINARES SALIVALES SECRETAN
DISTINTAS PROTEÍNAS
La estructura organizacional de las glándulas salivales
es similar a la del páncreas, las unidades acinares
secretoras drenan a conductos progresivamente más
grandes. No como el páncreas, la distribución celular
es más heterogénea y contiene 2 poblaciones
distintas de células acinares que sintetizan y secretan
distintos productos proteicos. Las células acinares de
la glándula parótida secreta un producto seroso con
abundancia de α-amilasa. Muchas células acinares de
las glándulas sublinguales secretan un producto
mucoso compuesto principalmente de glicoproteínas
mucina. La morfología de estas dos poblaciones
celulares difiere también. La glándula submandibular
contiene células acinares serosas y mucosas. En los
humanos, a diferencia de otras especies, se
entremezclan unidades mucosas y serosas acinares.
Aparte de la α-amilasa y las glicoproteínas mucina las
células acinares salivales secretan también proteínas
ricas en prolina, que como las mucinas están
altamente glicosiladas, y están en los gránulos
secretores acinares y son liberados por exocitosis.
VÍAS NEURALES COLINÉRGICAS Y ADRENÉRGICAS SON
LOS ACTIVADORES FISIOLÓGICOS MÁS IMPORTANTES DE
LA SECRECIÓN REGULADA POR LAS CÉLULAS ACINARES
SALIVALES
En diferencia al páncreas, en donde la estimulación
humoral es importante en estimular la secreción, las
glándulas salivales son controladas mayoritariamente
por el SNA. Los principales agonistas de la secreción
salival acinar son la ACh y la NE, liberadas de
terminaciones nerviosas simpáticas y parasimpáticas.
El receptor colinérgico en la célula salival acinar es del
tipo M3 subtipo glandular. Los adrenérgicos son del
subtipo α y β. Otros receptores que se han
identificado son los de sustancia P (NK1), VIP,
purinérgicos (P2z), neurotensina, prostaglandinas y
factores de crecimiento epidermal (EGF). Hay
algunos de estos receptores que se encuentran más
en las células del conducto que en las células
acinares. También difieren entre especies. Por esto es
difícil establecer exactamente la regulación de las
glándulas, pero es razonable decir que los NTs
colinérgicos y adrenérgicos estimulan la exocitosis.
TANTO EL CAMP COMO EL CA+2
MEDIAN LA SECRECIÓN
SALIVAL ACINAR
La secreción de proteínas por la célula acinar salival y
pancreática se asocia a aumentos en la [cAMP]i y en
la [Ca+2]i. La activación del cAMP a través del receptor
β-adrenérgico es el estimulante más potente de la
secreción de amilasa en la parótida de rata. La
activación de la vía por Ca+2 a través de los receptores
α-adrenérgicos, muscarínicos y de sustancia P
también estimula la liberación de amilasa por la
parótida. Aumentos en la [Ca+2]i causan la activación
vía proteína G de la PLC, lo que lleva a la generación
de IP3 y DAG. El IP3 hace que se libere Ca+2 desde los
reservorios internos y esto estimula proteína
quinasas dependientes de Ca+2 como la PKC y la
calmodulina quinasa, mientras que el DAG activa
directamente la PKC. El ATP liberado en conjunto con
la NE activa un receptor P2z, que es un receptor canal
que permite que entre Ca+2 y por tanto [Ca+2]i.
La secreción de fluidos y electrolitos es la segunda
función de las células acinares y salivales. La
secreción primaria es isotónica resultante de la
absorción basolateral de Cl- vía cotransportadores
Vía Autónoma Neurotransmisor Receptor Vía Respuesta (Secreción de)
Parasimpática ACh Sustancia P
Muscarínico M3 Taquiquinina NK-1
Ca+2
Ca+2 Fluido > Proteínas Fluido > Proteínas
Simpática α-Adrenérgico β-Adrenérgico
α-Adrenérgico β-Adrenérgico
Ca+2 cAMP
Fluido > Proteínas Proteínas > Fluido

Sebastián Lavanderos B. 2do. Medicina UDP
12
Na/K/Cl en conjunto con bombas Na-K y canales
basolaterales de K+. La secreción de Cl- y agua al
lumen es mediada por canales apicales de Cl- y AQP.
El Na+ y algo de agua llegan al lumen por vía
paracelular. Las células acinares de algunas especies
expresan también anhidridasa carbónica e
intercambiadores paralelos Cl-HCO3 y Na-H
basolaterales, lo que sugiere que otras vías podrían
contribuir también a la secreción primaria.
La estimulación de la secreción de fluidos y
electrolitos por las células acinares salivales es
mediada en su mayoría por estimulación colinérgica y
α-adrenérgica. La sustancia P vía su propio receptor
también produce cambios en la conductancia. Estos
efectos son mediados por aumentos en la [Ca+2]i, que
afecta a los canales apicales de Cl- y K+ vía
fosforilación por quinasas Ca+2-dep que podrían
afectar la probabilidad de que los canales estén
abiertos y de esta forma aumentar la conductancia.
LA CÉLULA DEL CONDUCTO SALIVAL
LAS CÉLULAS DEL CONDUCTO SALIVAL PRODUCEN UN
FLUIDO HIPOTÓNICO POBRE EN NACL Y RICO EN KHCO3
El conducto modifica la composición de la secreción
primaria, que es isotónica como el plasma, a través
de mecanismos de transporte activo. Su actividad se
ve reflejada en las invaginaciones de membrana y
abundantes mitocondrias, que le dan un aspecto
estriado a las células ductales. En general, se absorbe
Na+ y Cl-, y en menor extensión se secreta K+ y HCO3-.
Ya que el epitelio no es muy permeable al agua, el
lumen se vuelve hipotónico.
La reabsorción de Na+ ocurre en 2 pasos. Primero, el
canal de Na+ epitelial apical (ENaC) hace que el Na+
entre a la célula. Después, la bomba Na-K basolateral
saca este Na+.
La reabsorción de Cl- a través de la membrana apical
también ocurre en dos pasos. La entrada de Cl- ocurre
por un intercambiador Cl-HCO3. Los canales apicales
de Cl-, incluyendo el canal CFTR que reciclan este Cl-
absorbido por el intercambiador Cl-HCO3. Las células
ductales también tienen canales basolaterales de Cl-
que proveen una vía de salida para el Cl-.
La secreción de HCO3- ocurre a través del
intercambiador Cl-HCO3 apical. Esto depende de un
CFTR funcional, confirmando el acoplamiento del
CFTR al intercambiador Cl-HCO3.
La secreción de K+ ocurre a través de la absorción
basolateral de K+ a través de la bomba Na-K. El
mecanismo de la salida de K+ apical no se ha
establecido bien, pero podría ser por int. K-H.
LA ESTIMULACIÓN PARASIMPÁTICA DISMINUYE LA
ABSORCIÓN DE NA+, MIENTRAS QUE LA ALDOSTERONA
AUMENTA LA ABSORCIÓN DE NA+ POR LAS CÉLULAS
DUCTALES
La regulación de los procesos de transporte se
entiende menos en las glándulas salivales que en el
páncreas. En una salival intacta, la ACh por vía
parasimpática es la principal estimulación de la
secreción. En la célula ductal, agonistas colinérgicos
que actúan vía receptores colinérgicos aumentan la
[Ca+2]i y presumiblemente activan vías de regulación
dependientes de Ca+2. Los efectores de esta vía no se
conocen. El rol de las células ductales en la
producción incrementada de saliva se refleja en una
absorción de NaCl menor más que una secreción
aumentada de KHCO3.
Los efectos específicos de la estimulación adrenérgica
en el transporte de la célula ductal no son claros. Sin
embargo, la activación β-adr [cAMP]i y activa el
canal CFTR de Cl-.
La función de las células del conducto salival son
reguladas también por las hormonas circulantes. El
mineralocorticoide aldosterona estimula la absorción
de NaCl y secreción de K+. Si bien no se ha examinado
bien su rol en las células del conducto salival, la
aldosterona en otros epitelios absorbentes de Na+
(como el riñón y el colon) estimula el transporte de
Na+ aumentando la actividad del ENaC y la bomba Na-
K. Estas células también podrían tener receptores
para neuropéptidos como el VIP.
LAS CÉLULAS DEL CONDUCTO SALIVAL TAMBIÉN
SECRETAN Y ABSORBEN PROTEÍNAS
Las células ductales manejan proteínas de 3 maneras
distintas. Las sintetizan y secretan hacia el lumen,
sangre o las reabsorben desde el lumen.

Sebastián Lavanderos B. 2do. Medicina UDP
13
Las células ductales epiteliales intralobulares en las
submandibulares de roedores sintetizan varias
proteínas que se almacenan en gránulos
intralobulares y se secretan en respuesta a estímulos
neurohumorales. Las proteínas más abundantes
sintetizadas para secreción por estas células son el
EGF, NGF y kalicreína. Las células ductales salivales
también podrían sintetizar, guardar y secretar
algunas enzimas digestivas, como la α-amilasa y
ribonucleasas. La estimulación α-adrenérgica produce
la liberación de los gránulos, lo que nos indica que la
secreción proteica por las células ductales podría
estar regulada por la división simpática del SNA.
Aunque algunos péptidos reguladores se han
detectado en las células ductales salivales, no hay
evidencia que indique que estos se guardan en
gránulos o se secretan al lumen. Además, las células
ductales sintetizan receptores poliméricos de IgA,
responsables por la endocitosis basolateral de IgA, y
también sintetizan un componente secretor que
facilita la liberación apical de IgA.
Estas células ductales también pueden remover
sustancias orgánicas desde el lumen ductal (como la
ferritina). Además, se ha detectado que estas células
expresan receptores de transferrina, lo que indica
que podría ocurrir endocitosis regulada en estas
células.
COMPOSICIÓN, FUNCIÓN Y CONTROL DE LA
SECRECIÓN SALIVAL
DEPENDIENDO DE LA COMPOSICIÓN PROTEICA, LA
SECRECIÓN SALIVAL PUEDE SER SEROSA, SEROMUCOSA O
MUCOSA
La mayoría de la saliva (~90%) es producida por las
glándulas salivales principales, que son la parótida,
sublingual y submandibular. El 10% restante viene de
numerosas glándulas menores distribuidas en toda la
submucosa de la cavidad oral. Cada glándula salival
produce un tipo distinto de secreción, que puede ser
serosa, seromucosa o mucosa, los cuales se
diferencian por su cantidad de glicoproteínas. En los
humanos, la parótida es serosa, la sublingual y
submandibular es seromucosa y las salivales menores
producen una secreción mucosa.
Las secreciones serosas son ricas en α-amilasa, y las
mucosas en mucina. Sin embargo, las proteínas más
abundantes en la saliva de la parótida y
submandibular son proteínas ricas en prolina (1/3 del
total es prolina). Estas tienen propiedades
antimicrobiales, contribuyen a la lubricación de la
comida ingerida y estimulan la integridad de los
dientes vía interacciones con el Ca+2 y el
hidroxiapatito. La saliva también tiene cantidades
menores de lipasas, nucleasas, lisozimas,
peroxidasas, lactoferrina, IgA secretora, factores de
crecimiento, péptidos regulatorios y proteasas
vasoactivas como la kalicreína y renina.
Las funciones principales de la saliva son prevenir la
deshidratación de la mucosa oral y proveer
lubricación para la masticación y tragado de la
comida ingerida. El sentido del gusto y del olor en
menor extensión dependen de un suministro
adecuado de saliva. También es importante para
mantener una higiene oral adecuada. Y, aunque la α-
amilasa es un constituyente importante de la saliva,
esta no parece ser esencial para la digestión efectiva
de carbohidratos en la presencia de un páncreas que
está funcionando normal. Lo mismo pasa con la
lipasa lingual.
A FLUJOS BAJOS, LA SALIVA ES HIPOTÓNICA Y RICA EN K+,
MIENTRAS QUE A FLUJOS MAYORES, SU COMPOSICIÓN SE
PARECE MÁS A LA DEL PLASMA
La composición de la saliva varía de glándula en
glándula y de especie en especie. La secreción
primaria de las células acinares en reposo son
similares al plasma en su composición. La única
diferencia es que tienen más [K+]. En la mayoría de las
especies la estimulación no altera significativamente
la función de transporte celular ni la composición de
la secreción primaria. La filtración de las tight
junctions entre células acinares contribuye a la
formación de un producto de secreción similar al
plasma.
La composición de la saliva primaria es modificada
subsecuentemente por procesos de transporte en la
célula ductal. A tasas de secreción bajas (basales), Na+
y Cl- son absorbidos y K+ es secretado por las células
ductales de la mayoría de las glándulas salivales. Esto
genera una saliva rica en K+ hipotónica en reposo. A
tasas de secreción mayores, la composición del
producto de secreción final comienza a acercarse a la
secreción primaria parecida al plasma. La saliva
humana es siempre hipotónica, y la [K+] salival es
siempre mayor que la plasmática. En humanos el flujo
aumentado alcaliniza la saliva y aumenta su [HCO3-].
Esto neutraliza el ácido gástrico que normalmente
vuelve al esófago.
LA ESTIMULACIÓN PARASIMPÁTICA AUMENTA LA
SECRECIÓN DE SALIVA
Los humanos producen ~1,5 L/día de saliva. En
condiciones basales, las glándulas salivales producen
saliva a una velocidad de ~0,5 mL/min, y baja mucho
cuando se duerme. Después de estimulación, la
velocidad aumenta hasta 10 veces. Aunque las
glándulas responden a agonistas colinérgicos y
adrenérgicos, en condiciones fisiológicas es
importante la regulación parasimpática.
CONTROL PARASIMPÁTICO
Se origina en el núcleo salivatorio de la médula
oblongada. Entradas locales y centrales al núcleo

Sebastián Lavanderos B. 2do. Medicina UDP
14
salivatorio regulan las señales parasimpáticas. El
sabor y estímulos táctiles de la lengua son
transmitidos a la médula oblongada que puede
excitar a la secreción salival. Impulsos centrales
gatillados por la visión y olor de la comida también
excitan el núcleo salivatorio e inducen la salivación
antes de la ingestión de comida.
Fibras pregangliónicas parasimpáticas viajan por el PC
VII al ganglio submandibular, desde donde las fibras
postgangliónicas llegan a las glándulas sublinguales y
submandibulares. Las fibras pregangliónicas
parasimpáticas también viajan en el PC IX hacia el
ganglio ótico, desde donde fibras postgangliónicas
van hacia las parótidas. Además, algunas fibras
parasimpáticas alcanzan su destino final a través de la
rama bucal del PC V hacia las parótidas, o a través de
las ramas linguales del PC V a las sublinguales y
submandibulares. Estas estimulan directamente las
glándulas salivales con ACh. La disrupción de esta
inervación resulta en atrofia glandular.
CONTROL SIMPÁTICO
Las glándulas salivales son también inervadas por el
sistema simpático, a través de los ganglios cervicales
superiores, que viajan junto a los vvs sanguíneos
hasta las glándulas salivales. La estimulación
simpática aumenta el flujo de saliva, pero su
interrupción no tiene efectos mayores. Esta es el
estimulador principal de las células mioepiteliales,
que disminuyen la resistencia al flujo de los ductos
intercalados, y por tanto facilitan el flujo de secreción
de la saliva. También puede tener un control
indirecto modificando el flujo sanguíneo a la
glándula, aunque no es importante. Igual es el caso
con el VIP y sustancia P. Los mineralocorticoides
producen saliva con menos Na+ y más K+.







