Download - ANABOLISMO DE LOS CARBOHIDRATOS

UNIVERSIDAD DE EL SALVADOR
FACULTAD MULTIDISCIPLINARIA PARACENTRAL
DEPARTAMENTO DE CIENCIAS AGRONÓMICAS
INGENIERÍA AGROINDUSTRIAL
INGENIERÍA AGRONÓMICA
TEMA:
“ANABOLISMO DE CARBOHIDRATOS”
ASIGNATURA:
BIOQUÍMICA
DOCENTE:
LIC. RODRIGO ANTONIO MELÉNDEZ MORALES.
PRESENTAN:
EDWIN JOEL RAMOS MARTÍNEZ. RM15075
KENIA ISOLINA HENRÍQUEZ MARTÍNEZ. HM15024
GRUPO: No. 4
CICLO: I – 2016
SAN VICENTE, 09 DE JUNIO DE 2016

ii
ÍNDICE
Contenido Pág. INTRODUCCIÓN ..............................................................................................................................iii
OBJETIVOS. ..................................................................................................................................... 4
OBJETIVO GENERAL ................................................................................................................. 4
OBJETIVOS ESPECÍFICOS ...................................................................................................... 4
1. “ANABOLISMO DE LOS CARBOHIDRATOS” ................................................................... 5
1.1. GLUCONEOGÉNESIS. ................................................................................................... 5
1.3. VALOR BIOLÓGICO DE LOS CARBOHIDRATOS. ................................................... 8
1.4. IMPORTANCIA DE LA GLUCONEOGÉNESIS ........................................................... 8
2. FORMACION DE FOSFOENOLPIRUVATO A PARTIR DEL PIRUVATO ...................... 9
2.1. DIFERENCIAS ENTRE LOS PROCESOS DE GLICOLISIS Y
GLUCOGENOLISIS DEL PIRUVATO. .................................................................................... 10
2.2. FOSFOENOLPIRUVATO CARBOXICINASA ............................................................ 11
2.3. FRUCTOSA 1,6-BIFOSFATASA. ................................................................................ 11
2.4. GLUCOSA – 6 – FOSFATASA. ................................................................................... 12
3. PRECURSORES PARA LA GLUCONEOGÉNESIS. ....................................................... 13
3.1. GLUCONEOGÉNESIS A PARTIR DE AMINOÁCIDOS........................................... 14
3.2. GLICEROL. ..................................................................................................................... 15
3.3. PROPIANATO. ............................................................................................................... 15
3.4. ACETATO. ....................................................................................................................... 15
4. REGULACIÓN DE LA GLUCONEOGÉNESIS. ................................................................. 16
5. CONCLUSION. ....................................................................................................................... 17
6. BIBLIOGRAFIA. ...................................................................................................................... 18

iii
INTRODUCCIÓN
El trabajo tiene por objetivo dar a conocer el proceso de la biosíntesis de la
glucosa y de otros carbohidratos a partir de precursores sencillos.
Todas las formas de vida necesitan de energía para la síntesis de estructuras
imprescindibles para su desarrollo y formación, tanto plantas, animales y
microbios. Esta energía es utilizada en reacciones bioquímicas en su
metabolismo, para poder efectuar todos estos procesos es necesario la obtención
de nutrientes, como aminoácidos, ácidos grasos y de los destacan los
carbohidratos como compuestos altos en contenido energético. La obtención de
los carbohidratos se puede realizar de diversos modos para su asimilación
dependiendo del tipo de organismo, así pues, puede ser por obtención de materia
orgánica previamente elaborada (heterótrofos) y organismos que sintetizan
carbohidratos ya sea por compuestos no glucocídicos o reservas de energía en
forma de glucógeno o almidón en reacciones bioquímicas. Uno de los temas más
importantes en bioquímica y de los cuales depende el lograr comprender los
diferentes procesos de nutrición tanto en plantas, animales como microbios es “El
Anabolismo de los Carbohidratos”. Una de ellas consiste en las varias reacciones
mediante las cuales los productos intermedios del ciclo de los ácidos
tricarboxilicos se transforman en piruvato. Este proceso se efectúa en todos los
organismos y se denomina gluconeogénesis. El otro de los caminos principales de
alimentación estriba en las reacciones que provocan la reducción neta de CO2
para formar glucosa; esta senda no tiene efecto en los heterótrofos, pero
constituye una característica identificadora de los autótrofos.

4
OBJETIVOS.
OBJETIVO GENERAL
Indagar sobre los procesos anabólicos de los carbohidratos y la
importancia de estos como eslabones principales del desarrollo vital de
los distintos organismos que los utilizan.
OBJETIVOS ESPECÍFICOS
Estudiar los procesos de gluconeogénesis como mecanismos de
síntesis de carbohidratos.
Conocer los diferentes precursores no glucocídicos que forman
carbohidratos por las diferentes rutas anabólicas.

5
1. “ANABOLISMO DE LOS CARBOHIDRATOS”
1.1. GLUCONEOGÉNESIS.
La gluconeogénesis es el proceso de síntesis de glucosa o de glucógeno a partir
de precursores que no son carbohidratos (Harper, 2012). La biosíntesis de la
glucosa (anabolismo), y de otros carbohidratos a partir de precursores más
sencillos es el proceso biosintetico más notable que tiene lugar en la biosfera
según Lehninger (1972).
Todos los organismos tienen una ruta de biosíntesis de glucosa o
gluconeogénesis. No siempre puede estar disponible la glucosa de fuentes
externas o de reservas intracelulares. Ciertos tejidos en los mamíferos, en
especial de hígado y riñones, pueden sintetizar glucosa a partir de precursores
simples, como lactato y alanina. Bajo condiciones de ayuno, la gluconeogénesis
suministra casi toda la glucosa al organismo. Se requiere la glucosa para
metabolismo en ciertos tejidos, como en el cerebro. Durante el ejercicio, los
músculos convierten a la glucosa en piruvato y lactato, que van al hígado y son
convertidos en glucosa (H. Horton et al.2008).
En el dominio de los organismos fotosintéticos (plantas), se producen enormes
cantidades de hexosas a partir de dióxido de carbono y de agua, y las hexosas a
su vez, se convierten en almidones, celulosa y otros polisacáridos. En las células
heterotróficas también constituye un proceso central la conversión del piruvato, el
lactato, los aminoácidos y otros precursores simples en glucosa y en glucógeno
(Lehninger, 1972).
Un aporte de glucosa es necesario, en especial para el sistema nervioso y los
eritrocitos (Harper, 2012).

6
H. Horton et al. En 2008, determina que la glucosa es un sustrato básico, en
ocasiones incluso único, de muchos tejidos o vías celulares. El cerebro se nutre
principalmente de glucosa, pero en caso de extrema necesidad se puede
alimentar de otros sustratos lipídicos. Los eritrocitos se alimentan única y
exclusivamente de glucosa, por lo que es muy importante que el nivel de esta en
sangre se mantenga. Se trata por lo tanto de una reversión del proceso, pero no
de una inversión, porque existen pasos que son totalmente irreversibles, y son los
catalizados por los enzimas: HK/GK, PFK1 (fofofructoquinasa 1), PK (fofoquinasa).
Se requerirán enzimas que hagan que el proceso vaya en dirección contraria, y
estas son 4 y son:
Piruvato carboxilasa.
Fosfoenolpiruvato carboxiquinasa (PEPCK).
Fructosa – 1,6 – bisfosfatasa.
Glucosa – 6 – fosfatasa.
Utilizando esos 4 enzimas se puede revertir el proceso, aunque también
intervienen enzimas que actúan en la glucólisis, ya que las reacciones que
catalizan no son irreversibles (Espinoza L. 2012).
La síntesis de una molécula de glucosa a partir de dos de piruvato requiere cuatro
moléculas de ATP y dos de GTP (en animales), así como dos moléculas de
NADH. La ecuación neta para la gluconeogénesis es
2 Piruvato + 2 NADH + 4 ATP + 2 GTP + 6 H2O + 2 H Glucosa + 2 NAD_ + 4
ADP + 2 GDP + 6 Pi
En la etapa de hexosa de la gluconeogénesis, no se recupera energía en los
pasos que convierten la fructosa 1,6-bifosfato a glucosa, porque la fructosa 1,6-
bifosfato no es un intermedio de alta energía. La síntesis de una molécula de
glucosa en la gluconeogénesis consume un total de seis equivalentes de ATP y
7
dos moléculas de NADH. Como era de esperar, la biosíntesis de la glucosa
requiere energía, y su degradación libera energía (H. Horton et al.2008).
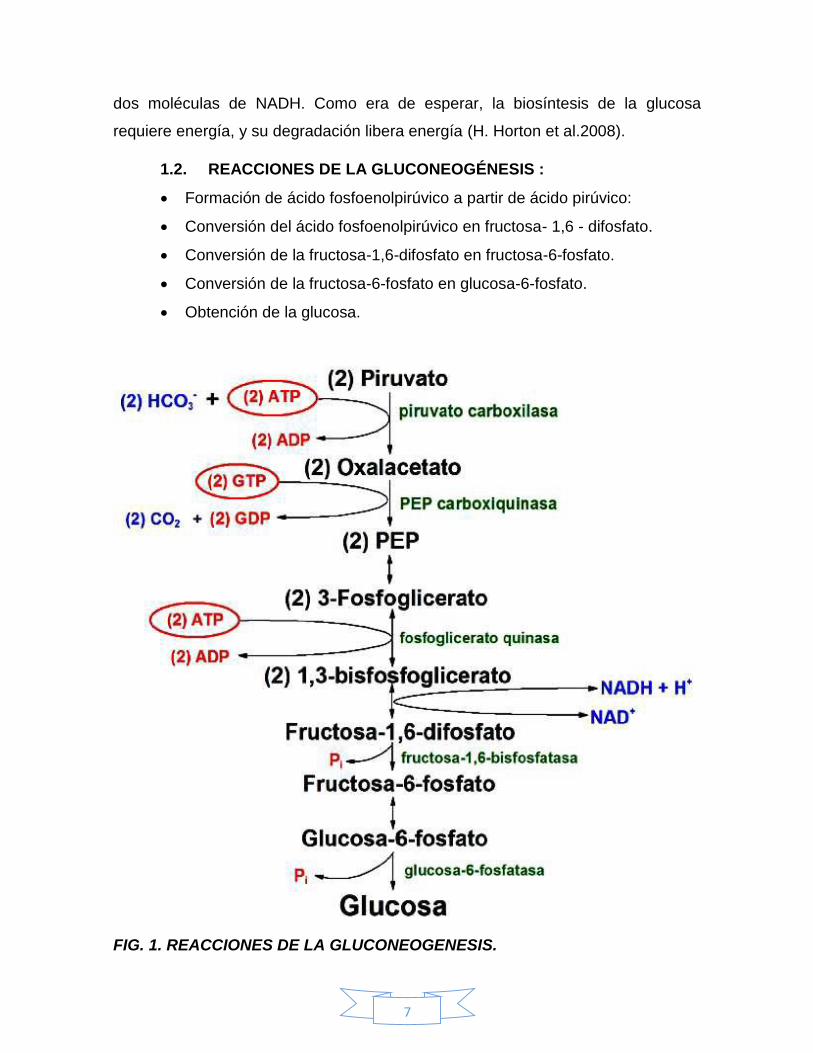
1.2. REACCIONES DE LA GLUCONEOGÉNESIS :
Formación de ácido fosfoenolpirúvico a partir de ácido pirúvico:
Conversión del ácido fosfoenolpirúvico en fructosa- 1,6 - difosfato.
Conversión de la fructosa-1,6-difosfato en fructosa-6-fosfato.
Conversión de la fructosa-6-fosfato en glucosa-6-fosfato.
Obtención de la glucosa.
FIG. 1. REACCIONES DE LA GLUCONEOGENESIS.

8
1.3. VALOR BIOLÓGICO DE LOS CARBOHIDRATOS.
Los carbohidratos constituyen la fuente principal de energía tanto en organismos
autótrofos como heterótrofos, energía metabólica que impulsa el desarrollo de los
mismos.
Espinoza L. (1998) en sus estudios sobre el “Metabolismo de los Carbohidratos”
adjudica a estos los valores biológicos siguientes:
a) La utilización de algunos de ellos como fuente de energía por los
animales y en general por todos los organismos aerobios.
b) Se utilizan algunos de ellos como reserva energética por ejemplo,
almidón en las plantas y el glucógeno en los animales superiores.
c) Participan en la síntesis de otras sustancias de gran importancia
biológica. Ejemplo: síntesis de aminoácidos, ácidos grasos, glicerina,
etc.
d) Función estructural extracelular. Ejemplo: celulosa en los tejidos
vegetales.
1.4. IMPORTANCIA DE LA GLUCONEOGÉNESIS
La glucosa obtenida mediante la gluconeogénesis se utiliza en la síntesis de otros
monosacáridos, disacáridos y polisacáridos estructurales. Por otra parte, la
gluconeogénesis que es muy activa en el hígado contribuye a la recuperación del
organismo después de un ejercicio físico prolongado, pues contribuye a eliminar
ácido láctico de la sangre (Harper, 2012).
Glucogénesis: La glucogénesis aparece por acumulación de glucógeno en los
tejidos, como consecuencia de un defecto en su metabolismo ya sea una
incapacidad para degradarlo inactividad de la enzima o porque esta no es
funcional, o secundario a la formación adecuada del glucógeno lo que impide
posteriormente su degradación. En otras palabras es el almacenamiento de
glucosa en el hígado el cual recibe el nombre de glucógeno (Espinoza L. 1998).

9
2. FORMACION DE FOSFOENOLPIRUVATO A PARTIR DEL PIRUVATO
La conversión de piruvato a fosfoenolpiruvato en la gluconeogénesis se lleva a
cabo en dos pasos. El primero de ellos es la reacción de piruvato y dióxido de
carbono para dar oxaloacetato. Este paso requiere de energía, disponible por
hidrolisis de ATP.
La enzima que cataliza esta reacción es la piruvato carboxilasa, una enzima
alosterica que se encuentra en la mitocondria. La acetil-CoA es un efecto
alosterico que activa al piruvato carboxilasa. Cuando hay altos niveles de acetil-
CoA (en otras palabras cuando hay más acetil-CoA) de los necesarios para que se
lleve a cabo el ciclo del ácido cítrico, el piruvato (un precursor del acetil-CoA) se
dirige a la gluconeogénesis (con frecuencia, el oxaloacetato del ciclo del ácido
cítrico también constituye un punto inicial para la gluconeogénesis) (Campbell y
Farrell, 2003).
La piruvato carboxilasa se estimula para dirigir al piruvato hacia el oxaloacetato, y
no la acetil-CoA. El oxaloacetato puede entrar al ciclo del ácido cítrico o servir
como precursor en la biosíntesis de la glucosa.
La reacción de piruvato carboxilasa tiene una función importante en la fijación de
dióxido de carbono en bacterias y algunos eucariotas. Sin embargo, mucho del
oxaloacetato que se produce no se usa en la gluconeogénesis. Más bien
reabastece la reserva de intermedios en el ciclo del ácido cítrico que sirven como
precursores en la biosíntesis de aminoácidos y lípidos (H. Horton et al.2008).

10
2.1. DIFERENCIAS ENTRE LOS PROCESOS DE GLICOLISIS Y
GLUCOGENOLISIS DEL PIRUVATO.
H. Horton et al. (2008) plantea que en el proceso de formación de glucosa o
gluconeogénesis (síntesis o formación) a partir de precursores no glucocídicos es
parecido a los procesos de glicolisis (ruptura o degradación), es por ello
importante hacer una comparación de las rutas que sigue el piruvato en estos
procesos. Siempre con el fin de generar glucosa ante condiciones adversas del
medio o del organismo.
FIG. 2. Comparación entre las reacciones de la glicolisis y glucogenólisis.

11
2.2. FOSFOENOLPIRUVATO CARBOXICINASA
La fosfoenolpiruvato carboxicinasa (PEPCK, phosphoenolpyruvate carboxykinase).
Cataliza la descarboxilación y fosforilación de oxaloacetato a fosfoenolpiruvato
usando GTP como el donador de fosfato. En hígado y riñones, la reacción de la
succinato tiocinasa en el ciclo del ácido cítrico produce GTP (en lugar de ATP
como en otros tejidos), y este GTP se usa para la reacción de fosfoenolpiruvato
carboxicinasa, lo que proporciona un enlace entre la actividad del ciclo del ácido
cítrico y la gluconeogénesis, con el fin de prevenir la eliminación excesiva de
oxaloacetato para gluconeogénesis, lo que alteraría la actividad del ciclo del ácido
cítrico (Harper, 2012).
La síntesis de fosfoenolpiruvato en dos pasos, a partir de piruvato, es común en la
mayor parte de los eucariotas, incluyendo al hombre. Es la razón principal de que
se muestre cuando se describe la gluconeogénesis. Sin embargo, muchas
especies de bacterias pueden convertir el piruvato directo a fosfoenolpiruvato en
una reacción dependiente de ATP y catalizada por fosfoenolpiruvato sintasa. Esta
es una ruta mucho más eficaz que la de dos pasos en los eucariotas, catalizada
por piruvato carboxilasa y PEPCK. La presencia de fosfoenolpiruvato sintasa en
células bacterianas se debe a que la gluconeogénesis eficiente es mucho más
importante en las bacterias que en los eucariotas, ya que la mayor parte de las
bacterias no se pueden basar en una fuente exógena de glucosa (H. Horton et
al.2008).
2.3. FRUCTOSA 1,6-BIFOSFATASA.
El fosfoenolpiruvato generado a partir del piruvato, gracias a las reacciones
anteriores, se convierte fácilmente en fructosa-1,6-difosfasto por inversión de las
reacciones de la glucolisis (Lehninger, 1972).
Sin embargo Campbell y Farrell (2012), describen que: se trata más bien de la
hidrolisis de la fructosa-1,6-difosfato para producir fructosa -6-fosfato e ion fosfato.

12
En esta reacción, la hidrólisis del éster fosfato se asocia a un gran cambio
negativo de energía libre de Gibbs (_G), que hace que esta reacción sea
metabólicamente irreversible. La enzima en los mamíferos presenta cinética
sigmoidal, y es inhibida de manera alostérica por AMP y por la molécula
reguladora de fructosa 2,6-bifosfato. Así, las dos enzimas que catalizan la
interconversión de la fructosa 6-fosfato y la fructosa-1,6-bifosfato son controladas
en forma recíproca por la concentración de fructosa-2,6-bifosfato.
2.4. GLUCOSA – 6 – FOSFATASA.
En esta etapa del camino hacia la glucosa la fructosa-6-fosfato se convierte, de
modo reversible en glucosa-6-fosfato por la acción de la fosfohexoisomerasa
(Lehninger, 1972). El paso final de la gluconeogénesis es la hidrólisis de la
glucosa 6-fosfato para formar glucosa. La enzima es glucosa 6-fosfatasa.
Esta es una reacción hidrolítica metabólicamente irreversible. Aunque se presenta
a la glucosa como producto final de la gluconeogénesis, eso no es cierto en todas
las especies. En la mayor parte de los casos, la ruta biosintética termina con la

13
glucosa 6-fosfato. Este producto es una forma activada de glucosa (H. Horton et
al.2008).
Sin embargo Lehninger (1972), expresa que en determinadas células como el
hígado, riñón y epitelio intestinal de los vertebrados, el glucosa-6-fosfato puede
resultar desfosforilado y liberar glucosa. Cabe destacar también que el hígado
constituye la fuente más importante de la glucosa sanguínea.
3. PRECURSORES PARA LA GLUCONEOGÉNESIS.
Los principales precursores para la gluconeogénesis en los mamíferos son lactato
y la mayor parte de los aminoácidos, en especial alanina. La glicerina, que se
produce por hidrólisis de los triacilgliceroles, también es un sustrato para la
gluconeogénesis (H. Horton et al.2008).
Según Espinoza L. (2012), El lactato es el principal sustrato gluconeogénico, ya
que casi todas las células tienen la capacidad de sintetizarlo. Este lactato es
generado por la glicólisis (en gran medida) en los músculos activos, y también así
por los glóbulos rojos de manera continua; entra al torrente sanguíneo y llega al
hígado, donde se convierte en piruvato por acción de la lactato deshidrogenasa,
constituyendo el Ciclo de Cori (H. Horton et al.2008).
FIG.3. Ciclo de Cori, o Ciclo del Ácido Láctico.

14
3.1. GLUCONEOGÉNESIS A PARTIR DE AMINOÁCIDOS.
Los esqueletos de carbono en la mayor parte de los aminoácidos son
catabolizados a piruvato o a compuestos intermedios en el ciclo del ácido cítrico.
Los productos finales de estas rutas catabólicas pueden servir directamente como
precursores para la síntesis de glucosa 6-fosfato en las células capaces de
efectuar la gluconeogénesis (H. Horton et al.2008).
Aquellos aminoácidos que pueden precursores del fosfoenolpiruvato y, por
consiguiente, de la glucosa, son aminoácidos glucogénicos. Hay un aminoácido,
la leucina, que en los vertebrados no puede contribuir a la formación neta de
glucosa, dado que todos sus átomos se convierten en acetil-CoA o en CO2
(Lehninger, 1972).
El ciclo de glucosa-alanina es un sistema parecido de transporte. El piruvato
puede aceptar un grupo amino de un -aminoácido, como glutamato, para formar
alanina con el proceso de transaminación.
La alanina va al hígado donde sufre transaminación con -cetoglutarato a fin de
volver a formar piruvato para la gluconeogénesis. Los aminoácidos se convierten
en una de las fuentes principales de carbono para gluconeogénesis durante el
ayuno, cuando se agota el suministro de glucógeno (H. Horton et al.2008).

15
3.2. GLICEROL.
El catabolismo de los triacilgliceroles produce glicerol y acetil-CoA. Como se
mencionó antes, la acetil-CoA contribuye a la formación neta de glucosa por
reacciones del ciclo del glioxilato. El ciclo del glioxilato no contribuye a la síntesis
neta de glucosa a partir de lípidos en las células de mamíferos. Sin embargo, el
glicerol se puede convertir en glucosa en una ruta que comienza con la
fosforilación a glicerol 3-fosfato, catalizada por glicerol cinasa. El glicerol 3-fosfato
entra a la gluconeogénesis después de convertirse en dihidroxiacetona fosfato. (H.
Horton et al.2008).
3.3. PROPIANATO.
La oxidación de ácidos grasos con un número impar de átomos de carbono y la
oxidación de algunos aminoácidos genera como producto final de la oxidación
propionil-CoA. La propionil-CoA se convierte en el intermediario del ciclo de Krebs,
succinil-CoA. Esta conversión se lleva a cabo por la enzima dependiente de ATP,
propionil-CoA carboxilasa, la metilmalonil-CoA epimerasa y finalmente por la
enzima que requiere vitamina B12, la metilmalonil-CoA mutasa. La utilización del
propionato en la gluconeogénesis solamente tiene una significancia cuantitativa en
los rumiantes (Lehninger 1972).
3.4. ACETATO.
Muchas especies pueden usar acetato como fuente principal de carbono. Estas
especies pueden convertir el acetato en acetil-CoA, que puede ser el precursor
para el oxalacetato. Las bacterias y los eucariotas unicelulares, como las
levaduras, usan el acetato como precursor para la gluconeogénesis. Algunas
especies de bacterias pueden sintetizar al acetato en forma directa a partir de
CO2. En ellas, la ruta de gluconeogénesis les proporciona una forma de sintetizar
glucosa a partir de sustratos inorgánicos (H. Horton et al.2008).

16
4. REGULACIÓN DE LA GLUCONEOGÉNESIS.
La gluconeogénesis se controla cuidadosamente in vivo. La glicólisis y la
gluconeogénesis son rutas opuestas, catabólicas y anabólicas, que comparten
algunos pasos enzimáticos.
Sin embargo, algunas reacciones son únicas para cada ruta. Por ejemplo, la
fosfofructocinasa-1 cataliza una reacción en la glicólisis, y la fructosa 1,6-
bifosfatasa cataliza la reacción contraria en la gluconeogénesis, y ambas
reacciones son metabólicamente irreversibles. Por lo regular, sólo una de las
reacciones se efectúa en forma importante. Bajo algunas condiciones, las enzimas
operan al mismo tiempo, y la reacción neta es la hidrólisis de ATP a ADP y Pi
(Lehninger 1972)
FIG. 4. Regulación de la gluconeogénesis.

17
5. CONCLUSION.
La gluconeogénesis es el proceso de síntesis de glucosa o glucógeno a partir de
precursores que no son carbohidratos. Tiene especial importancia cuando el
carbohidrato no está disponible a partir de la dieta. Los sustratos importantes son
aminoácidos, lactato, glicerol y propionato.
La vía de la gluconeogénesis en hígado y riñones utiliza las reacciones en la
glucólisis que son reversibles, más cuatro reacciones adicionales que evitan el
paso por las reacciones no de equilibrio irreversibles.
Dado que la glucólisis y la gluconeogénesis comparten la misma vía pero operan
en direcciones opuestas, es necesario que sus actividades se regulen de manera
recíproca. El hígado regula la glucosa en la sangre después de una comida,
porque contiene la glucocinasa que promueve el aumento de la utilización
hepática de glucosa.

18
6. BIBLIOGRAFIA.
Campbell, MK; Farrell So. 2003. Bioquímica. 4a Ed. Universidad Estatal De
Colorado. Thomson. 497-504 P. ISBN 970-686-335-4.
Harper, K. Murray, A. Bender, M. Botham, Kennelly, W. Rp. Weil, Phd 2012
Bioquímica Ilustrada, 29va Edición, Capítulo 22, McGraw-Hill. 187- 192 P. ISBN
978-607-15-0914-7
Espinoza L. F. 1968- 2012 “Metabolismo De Carbohidratos” Universidad Católica
Agropecuaria Del Trópico Seco. Consultado El Día 04/ 06/ 2016.Págs. 16-20. Guía
Técnica.
Lehninger A., 1972 Bioquímica, 4ta Edición. Ediciones Omega, S. A. Capítulo 22
Págs. 517-543. ISBN 84-282-0211-7.
H. Robert Horton, Laurence A. Morán, K. Gray Scrimgeour, Marc D. Perry, J.
David Rawn, 2008, Principios De Bioquímica Cuarta Edición, México, Editor:
Rubén Fuerte Rivera Consultado El Día 04/06/ 2016 Págs. 357-366.







